
Supplementary Materials for -...
34
www.sciencemag.org/cgi/content/full/science.aad0512/DC1 Supplementary Materials for Polysialylation controls dendritic cell trafficking by regulating chemokine recognition Eva Kiermaier,*† Christine Moussion,† Christopher T. Veldkamp, Rita Gerardy-Schahn, Ingrid de Vries, Larry G. Williams, Gary R. Chaffee, Andrew J. Phillips, Friedrich Freiberger, Richard Imre, Deni Taleski, Richard J. Payne, Asolina Braun, Reinhold Förster, Karl Mechtler, Martina Mühlenhoff, Brian F. Volkman, Michael Sixt*† *Corresponding author. E-mail: [email protected] (E.K.); [email protected] (M.S.) †These authors contributed equally to this work. Published 10 December 2015 on Science Express DOI: 10.1126/science.aad0512 This PDF file includes: Materials and Methods Figs. S1 to S5 Tables S1 Full Reference List
Transcript of Supplementary Materials for -...

www.sciencemag.org/cgi/content/full/science.aad0512/DC1
Supplementary Materials for
Polysialylation controls dendritic cell trafficking by regulating chemokine recognition
Eva Kiermaier,*† Christine Moussion,† Christopher T. Veldkamp, Rita Gerardy-Schahn, Ingrid de Vries, Larry G. Williams, Gary R. Chaffee, Andrew J. Phillips, Friedrich Freiberger, Richard Imre, Deni Taleski, Richard J. Payne, Asolina Braun, Reinhold
Förster, Karl Mechtler, Martina Mühlenhoff, Brian F. Volkman, Michael Sixt*†
*Corresponding author. E-mail: [email protected] (E.K.); [email protected] (M.S.) †These authors contributed equally to this work.
Published 10 December 2015 on Science Express
DOI: 10.1126/science.aad0512
This PDF file includes:
Materials and Methods Figs. S1 to S5 Tables S1 Full Reference List

2
Materials and Methods Mice All mouse strains used in this study (C57BL/6, St8sia4-/-, Ccr7-/-, Ccl19-/-) were bred and maintained at IST Austria according to the local regulations. Antibodies and reagents
The following antibodies and conjugates were used in this study: mouse α-CCR7-PE (4B12), rat α-mouse I-A/I-E-eFluor450 (M5/114.15.2), rat α-mouse I-A/I-E-biotin (2G9), hamster α-mouse CD11c-APC (N418), hamster α-mouse CD11c-biotin (N418), rat α-mouse B220-PE-Cy7 (RA3-6B2), rat α-mouse CD3-eFluor450 (17A2), rat α-human/mouse CD44-PE-Cy7 (IM7), Streptavidin-FITC all from eBioscience; rat α-mouse CD62L (MEL-14, BD Biosciences), rat α-mouse CD11b-BrillantViolet421 (M1/70, BD Biosciences), α-mouse CD11c-PE-Cy7 (HL3, BD Biosciences), rat α-mouse CD4-V500 (RM4-5, BD Biosciences), rat α-mouse CD103 (M290, BD Biosciences), hamster α-mouse CD69 (H1.2F3, BD Biosciences), α-mouse I-A/I-E-BrillantViolet510 (M5/114, BioLegend), rat α-mouse M1/70-PE (M1/70, Biolegend), rat α-CD8α-PerCP (53-6.7; Biolegend); rat α-CD207/Langerin-e647 (929F3.01, Dendritics); rabbit α-laminin (L9393, Sigma), rabbit α-mouse neuropilin-2 (D29A5), rabbit α-Akt (C67E7)/α-phospho-Akt (S473, D9E), rabbit α-Erk1/2 (137F5)/α-phospho-Erk1/2 (Th202/Thr204, D13.14.E) all from Cell Signaling; rabbit α-GAPDH (Abcam); mouse α-GFP antibody (2B6) was kindly provided by Stefan Schüchner and used at a final concentration of 3.5μg/ml ; goat α-mouse IgG-HRP conjugate, goat α-rabbit IgG-HRP conjugate both from Bio-Rad; donkey α-rat Alexa-Fluor488, donkey α-rat Alexa-Fluor647, Streptavidin-Cy3, all from Jackson ImmunoResearch. All commercially available antibodies and conjugates were diluted and used according to the manufacturer’s recommendations.
Mouse α-polySia monoclonal antibody 735 (mab 735) and active/inactive endoneuraminidase (-GFP) were purified as described previously (24-26). Mab 735 was used at a final concentration of 2μg/ml. Expression and purification of full length CCL21 (CCL21-FL 1-111), truncated CCL21 (CCL21trunc 1-79) and the CCL19/21 chimera (CCL19 1-77-CCL21 78-111) The proteins were expressed and purified using a procedure adapted from Lu et al. (27). Codon optimized DNA coding for the desired fusion protein SMT3-CCL21 1-111 or SMT3-CCL21 1-79 were cloned into the BamHI or HindIII sites of pQE30 while SMT3-CCL19 1-77-CCL21 78-111 was cloned into the same sites in pET28a. All plasmids were sequence verified. Each expression plasmid were individually transformed into BL21 [DE3] E. coli (pET28A- SMT3-CCL19 1-77-CCL21 78-111) or BL21 [pREP4] E.coli (pQE30-SMT3-CCL21 1-111, pQE30-SMT3-CCL21 1-79). One-liter cultures were grown at 37˚C in either lysogeny broth or [U-15N] or [U-15N/13C] M9 minimal media to an

3
optical density at 600nm of 0.5-0.7 and then induced with 1mM isopropyl-β-D-thiogalactopyranoside for five hours. Cell pellets were collected by centrifugation and stored at -20˚C. Cells were resuspended in 10mL of buffer A (50mM sodium phosphate, 300mM NaCl, 10mM imidazole, pH 8.0) containing 1mM phenylmethylsulfonyl fluoride (PMSF) and 0.1% (v/v) 2-mercaptoethanol. Resuspended cells were lysed by sonication and the lysate was centrifuged at 15,000 × g for 15 minutes. All the proteins expressed insolubly as an inclusion body. The inclusion bodies were separately dissolved in 10mL of buffer AD (50mM sodium phosphate, 300mM NaCl, 10mM imidazole and 6M guanidine hydrochloride, 0.1% (v/v) 2-mercaptoethanol, pH 8.0), clarified by centrifugation at 15,000 × g for 15 minutes and batch loaded onto 2mL of His60 nickel resins for thirty minutes. The columns were washed with 40mL of buffer AD and eluted with 30mL of buffer BD (100mM sodium acetate, 300mM NaCl, 10mM imidazole, and 6M guanidine HCl, pH 4.5). Elutions for the His6-SMT3-CCL19/21, His6-SMT3-CCL21 (1-111), and His6-SMT3-CCL21 (1-79) fusions were dialyzed 4˚C with stirring against 4L of 0.3% acetic acid overnight and then against 4L of refolding buffer (20mM Tris, pH 8.0). After 15-20 hours, 400µg of His6-Ulp1 (ubiquitin like protease-1) was added and left for additional 15-20 hours.
After clarification by centrifugation at 4,000 × g for 15 minutes, the digestions were separately loaded onto SP HP HiTrap™ 1mL columns at a flow rate of 2.5mL/min. The columns were washed with 15mL of wash buffer (100mM Tris, 25mM NaCl, pH 8.0) to remove His6-SUMO and His6-Ulp1. The chemokines were eluted with 15mL of elution buffer (100mM Tris, 2M NaCl, pH 8.0). CCL19/21 (chimera), CCL21 1-111 (CCL21-FL), and CCL21 1-79 (CCL21trunc) were further purified using reverse phase HPLC (0.1% aqueous trifluoroacetic acid buffer with a CH3CN gradient from 21 to 42% (v/v) over 30 minutes). MALDI-TOF mass spectrometry was used (Applied Biosystems Voyager 6004) to confirm molecular weight. Immunohistology LNs were fixed overnight in 0.05M phosphate buffer, pH 7.4 containing 2mg/ml NaIO4, 0.1mM L-lysine and 1% paraformaldehyde (all from Sigma) and dehydrated in consecutive sucrose gradients (10, 20 and 30% in 0.05M phosphate buffer) to preserve LN structures. Organs were frozen in O.C.T and cut into 10μm cryosections. Sections were air-dried before rehydrating in TBS+1% Tween (Sigma) and blocking in 1% BSA (Sigma). Antibody staining was carried out according to the manufacturer’s dilution recommendations. Slides were analyzed with an upright Zeiss LSM700 confocal microscope equipped with a Plan-Apochromat 10x/0.45 objective (Zeiss). For analysis of Langerin signal intensities the background below twice the overall mean intensity was subtracted and the remaining intensity, above the threshold, normalized by the overall intensity. In this way only significant contributions above the mean intensity are considered while compensating for the varying size and brightness level of the image. Flow Cytometry LNs and spleens were treated with collagenase D (Roche) or II (Gibco, 0.5μg/ml) for 20

4
minutes at 37°C and digestion stopped with 10mM EDTA. LNs and spleen were mechanically crushed to isolate immune cells and erythrocytes of spleen were lysed in red blood cell (RBC) buffer containing ammonium chloride (1.7M), potassium hydrogen carbonate (100mM), and EDTA (1mM). Before staining, 1-2x106 cells were incubated for 15 min at room temperature with blocking buffer (RPMI, 10% FCS, 5mM EDTA) containing 5mg/ml α-CD16/CD32 (2.4G2, BD Biosciences). For surface staining, cells were incubated for 30 min on ice with conjugated monoclonal antibodies (mAbs) diluted at the recommended concentration in blocking buffer. iEndoN-GFP was used at a final concentration of 0.6mg/ml. Flow cytometry analysis was performed on a FACS CANTO II flow cytometer (BD Biosciences). For phenotypic analysis of Langerin+ cells, after surface staining and fixation in 1% paraformaldehyde, cells were permeabilized with 0.5% saponin and intracellular staining was performed with α-mouse Langerin antibody for 45 minutes. In vivo lymphocyte homing assay Naïve lymphocytes were isolated from LNs and spleen (after RBC lysis) of St8sia4-/- and control mice and labeled with 10μM 5-(and-6-) Carboxytetramethylrhodamine, Succinimidyl Ester (TAMRA) or 5μM Carboxyfluorescein Succinimidyl Ester (CFSE; both Molecular Probes, Life Technologies) in PBS, 0.01% BSA. Labeling was stopped by adding 5 volumes of full media (R10; RPMI 1640 supplemented with 10% FBS, L-Glutamin, β-ME and Penicillin/Streptomycin, all from Invitrogen). 15-20x106 labeled lymphocytes were injected intravenously in PBS into St8sia4-/- and control recipient mice. Three hours after injection, mice were sacrificed and lymphocytes isolated from LNs and spleen to recover transferred cells. The percentage of fluorescent cells recruited to each lymph node was determined by flow cytometry. Generation of bone marrow derived dendritic cells and footpad injections Bone marrow-derived dendritic cells (BMDCs) were generated from flushed bone marrow suspension as previously described (28). Cells were grown in R10 medium supplemented with 20% GM-CSF hybridoma supernatant for seven to nine days before stimulation with 200ng/mL LPS (Sigma-Aldrich; E.Coli 0127:B8) over night. Mature BMDCs were labeled with 10μM 5-(and-6-) Carboxytetramethylrhodamine, Succinimidyl Ester (TAMRA) or 5μM Carboxyfluorescein Succinimidyl Ester (CFSE; both Molecular Probes, Life Technologies), respectively. Labeling was stopped by adding 5 volumes of R10 media. A total amount of 5x106 cells in 50μl PBS (2.5x106 cells/color) were injected into the hind footpad of recipient mice, which were sacrificed 48 hours later and popliteal LNs analyzed by flow cytometry. For phenotypic analysis of maturated DC in vivo, 10mg of LPS diluted in 50ml of PBS were injected into one food pad. FITC painting For experiments involving the tracking of skin DC into the lymph node after skin inflammation, St8sia4-/- and control mice were anesthetized with isofluorane and each

5
side of the ear was painted with 25ml of 2% FITC solution prepared in dibutylphtalate/acetone (1/1 vol/vol; Sigma-Aldrich). Three days later, mice were sacrificed and a lymph node cell suspension was generated from the pool of draining cervical LNs. To calculate the total number of recruited skin Langerin+ DCs into the lymph nodes, the total number of lymph node cells, was multiplied by the percentage of CD11c+MHCIIhighLangerin+FITC+ cells measured by flow cytometry. Ex vivo crawl-out migration assay and immunostaining of mouse ear sheets Mouse ear sheets were prepared as previously demonstrated (17) with the following modifications: ventral ear sheets were incubated up-side-down in R10 medium for 48 hours and subsequently fixed with 1% paraformaldehyde over night. Ear sheets were permeabilized with 0.2% Triton/PBS for 15 minutes and blocked with 1% BSA (PAA Laboratories) for one hour. Staining of lymphatic vessels and mature DCs was carried out using α-LYVE-1 and α-MHCII-biotin primary antibodies diluted in 1% BSA and secondary antibodies conjugated to FITC and Streptavidin-Cy3, respectively. For end point analysis of crawl-in assays, ventral ear sheets were mounted and immunostained in custom-made chambers using LYVE-1 primary- and Alexa647-conjugated secondary antibodies as described above. A total of 1x106 labeled BMDCs was added in R10 medium on top of the ear sheets and incubated for 15 minutes at 37°C to allow entering of cells into the dermis. Ear sheets were rinsed twice with R10 medium and incubated at 37°C for two hours before fixing with 1% paraformaldehyde. Confocal images of ear explants were acquired using an upright Zeiss LSM700 confocal microscope equipped with a Plan-Apochromat 20x/0.8 objective (Zeiss). Maximum intensity projections were analyzed with Fiji software to reconstitute lymphatic vessel structures according to LYVE-1 staining. DC location was estimated manually for at least five different regions per ear sheet and at least three different mice per genotype. Intralymphatic injections C57BL/6 mice were housed and bred in a pathogen-free animal facility. Mice were anesthetized by intraperitoneal injection of 20mg/kg xylazine and 100mg/kg ketamine. After leg-depilation with cream, a skin incision was made on the dorsal side of the lower hind leg and the main lymphatic vessel draining the footpad area to the popliteal lymph node was exposed by microsurgery. A glass capillary (3 in., 1.2mm/0.68mm OD/ID; WPI) was pulled with a micropipette puller (Model P-97; Sutter instrument CO), broken and grounded with a micropipette grinder (MicroData Instrument, Inc). BMDCs were labeled for 15 min at 37°C with 10μM TAMRA in PBS, 0.01% BSA and the microcapillary was filled with 5μl cell suspension (in PBS) using to a PLI-100A microinjector (Harvard Apparatus). A micromanipulator (Kleindiek Nanotechnik) was used for the stabilization of the capillary during injection. A pressure of approximately 5kPa in 120s pulses was used for the delivery of cells, avoiding injection of air. At all times during surgery, exposed tissues were kept moist with sterile PBS and observed with Leica M125 stereomicroscope. Incisions were closed with tissue adhesive (3M Vetbond) at the end of surgery. The draining popliteal LN was harvested 10h later, fixed overnight

6
in PLP fixative buffer (0.05M phosphate buffer, 0.1M L-lysine, pH 7.4, 2 mg/ml NaIO4, 10 mg/ml paraformaldehyde), and dehydrated in consecutive sucrose gradients (10, 20, and 30% in phosphate buffer). Tissues were snap frozen in Tissue-Tek. 6μm frozen sections were stained with rabbit α-laminin Ab (Sigma- Aldrich) and secondary α-rabbit Alexa 647 Ab to visualize LN structures and localize injected BMDCs. Representative images were analyzed using IMARIS imaging software. Based on the laminin staining, surface corresponding to the outside border of the cortical sinus was selected manually. A distance transformation was performed outside this new defined surface, yielding a new (distance-) channel with intensity values directly corresponding to the distance of the nearest edge of the isosurface. TAMRA-labeled cells were represented as dots and distance-channel intensities were used for quantifying the average migratory distance from the LN’s edge. Chemotaxis assays In vitro 3D dendritic cell migration assays have been carried out as previously described (29). Briefly, 3-4x105 cells were suspended in a medium:collagen mixture (PureCol bovine collagen, (INAMED) in 1× minimum essential medium eagle (MEM, Invitrogen)) and 0.4% sodium bicarbonate (Sigma) at a volume ratio of 1:2 yielding in a final collagen concentration of 1.6mg/ml. Collagen gel mixtures were casted into custom-made migration chambers and incubated for 45 minutes at 37°C to allow polymerization of the gel. Individual chemokines were suspended in R10 medium to a final concentration of 0.33μM and placed on top of the gel. DC migration was followed by time-lapse video microscopy and cells tracked either manually by using the ‘Manuel tracking- and chemotaxis tool’ plug-in provided by Fiji or automated by an in-house generated algorithm. Automated cell tracking Average migration speed of all cells was calculated for each time point using a custom made script, determining lateral displacements that optimizes its overlap with the previous frame. Migration speed was calculated from the optimal y-displacement in pixels, divided by the time between two consecutive frames. Generation and transfection of CCR7-GFP glycosylation mutants Mouse CCR7-GFP expression plasmid (pcDNA3.1-) was a gift of Daniel Legler and the plasmid encoding full length hamster ST8Sia IV was generated as described (30). Individual putative CCR7 glycosylation sites were mutated to alanine by using the site directed mutagenesis kit (Stratagene) according to the manufacturer’s instructions. HEK293 cells were grown in DMEM (Invitrogen) supplemented with 10%FBS and transfected using Lipofectamine (Invitrogen) at 80% confluence. Protein extraction, Immunoprecipitation and Western blotting

7
To identify polysialylated proteins in mature DCs, 1x106 cells were lysed in 500μl RIPA buffer (Cell Signaling) containing 50μl/ml protease inhibitor cocktail (Roche) and subsequently incubated with inactive EndoN-coupled to MyOne® tosylactivated Dynabeads (Invitrogen) at +4°C under constant rotation. Coupling of Dynabeads was carried out according to the manufacturer’s recommendations. Proteins were eluted with 100mM Glycine, pH 2.0 at room temperature and subjected to mass spectrometric analysis. For in vitro reconstitution of CCR7 polysialylation, transfected HEK293 cells of one well of a 6-well plate were lysed in 60μl RIPA buffer containing 50μl/ml protease inhibitor cocktail. Polysialylated proteins were immunoprecipitated and eluted as described above. 6μl of cell lysate was retained as input. To remove polySia from N- and O-glycans whole cell lysates were incubated with active EndoN at a final concentration of 1.5μg/ml for two hours at 37°C. Proteins were separated by SDS-PAGE under reducing conditions and transferred to a polyvinylidene difluoride membrane (Invitrogen). Primary antibodies were used according to the recommended dilutions and detected by peroxidase-conjugated α-mouse IgG or α-rabbit IgG antibodies. Detection of proteins was performed by enhanced chemiluminescence (ECL) detection using a VersaDoc imaging system (Biorad). MS-based protein identification The nano HPLC system used was an UltiMate 3000 HPLC RSLC nano system (Thermo Fisher Scientific, Amsterdam, Netherlands) coupled to a Q Exactive mass spectrometer (Thermo Fisher Scientific, Bremen, Germany), equipped with a Proxeon nanospray source (Thermo Fisher Scientific, Odense, Denmark). Peptides were loaded onto a trap column (Thermo Fisher Scientific, Amsterdam, Netherlands, PepMap C18, 5 mm × 300 μm ID, 5 μm particles, 100 Å pore size) at a flow rate of 25μL min-1 using 0.1% TFA as mobile phase. After 10 min, the trap column was switched in line with the analytical column (Thermo Fisher Scientific, Amsterdam, Netherlands, PepMap C18, 500 mm × 75 μm ID, 2 μm, 100 Å). Peptides were eluted using a flow rate of 230 nl min-1, and a binary 2h gradient, respectively 165 min. The gradient starts with the mobile phases: 98% A (water/formic acid, 99.9/0.1, v/v) and 2%B (water/acetonitrile/formic acid, 19.92/80/0.08, v/v/v) increases to 35%B over the next 120 min, followed by a gradient in 5 min to 90%B, stays there for five min and decreases in 5 min back to the gradient 98%A and 2%B for equilibration at 30°C. The Q Exactive mass spectrometer was operated in data-dependent mode, using a full scan (m/z range 350-2000, nominal resolution of 70 000, target value 1E6) followed by MS/MS scans of the 12 most abundant ions. MS/MS spectra were acquired using normalized collision energy 30%, isolation width of two and the target value was set to 1E5. Precursor ions selected for fragmentation (charge state two and higher) were put on a dynamic exclusion list for 30 seconds. Additionally, the underfill ratio was set to 20% resulting in an intensity threshold of 5E4. The peptide match feature and the exclude isotopes feature were enabled. Data Analysis:

8
For peptide identification, the RAW-files were loaded into Proteome Discoverer (version 1.4.0.288, Thermo Scientific). All hereby created MS/MS spectra were searched using MS Amanda against the human swissprot protein sequence database (from 2014-07-06; 16678 sequences, 9357021 residues). The following search parameters were used: β-methylthiolation on cysteine was set as a fixed modification, oxidation on methionine was set as variable modification. Monoisotopic masses were searched within unrestricted protein masses for tryptic peptides. The peptide mass tolerance was set to ±10 ppm and the fragment mass tolerance to ±20 ppm. The maximal number of missed cleavages was set to 2. The result was filtered to 1% FDR using Percolator algorithm integrated in Thermo Proteome Discoverer. All non-mouse proteins were excluded from the list. Protein Areas are calculated as average of the three highest peptide areas, also integrated in Thermo Proteome Discoverer. Signaling assays Mature DCs were stimulated with 20ng/ml chemokine for the indicated time points and subsequently lysed in RIPA buffer containing 50μl/ml protease and phosphatase inhibitor cocktail (both from Roche) at +4°C. Protein lysates were separated by SDS-PAGE, followed by immunoblotting and detection of the respective (phospho)- proteins as described above. Protein NMR All NMR experiments were performed at 25˚C at the Medical College of Wisconsin NMR facility on either a Bruker 600 or 500 MHz spectrometer equipped with a triple resonance cryoprobe. All NMR samples contained the same NMR buffer (25mM deuterated MES, 10% D2O, 0.02% NaN3, pH 5.9). Spectra of [U-15N] CCL21trunc (residues 1-79) and full length [U-15N] CCL21 (residues 1-111) were collected at a protein concentration of 200µM. Titrations of [U-15N] CCL21trunc (residues1-79) and full length [U-15N] CCL21 (residues 1-111) with incremental additions of polysialic acid (DP9) were monitored by 15N-1H HSQC spectra. Backbone amide chemical shift perturbations were computed as [(5ΔδH)
2 + (ΔδN) 2]0.5
where ΔδH and ΔδN are the change in backbone amide 1H and 15N chemical shift, respectively. Dissociation constants were determined using the equation
where Δδ is the chemical shift perturbation, Δδmax is the maximum chemical shift perturbation at 100% bound CCL21, Kd is the dissociation constant, and x is the polySia (DP9) concentration. PolySia (DP9) was produced as previously described (31, 32). Statistical analysis
€
Δδ = Δδmax ×(Kd + [CCL21]+ x) − (Kd + [CCL21]+ x)2 − 4[CCL21]x
2[CCL21]

9
Results are expressed as mean ±SD with n numbers indicated for each experiment. Differences between two groups were examined by two-tailed unpaired student’s t-tests. For intralymphatic injections differences were examined by two-tailed paired student’s t-test. *P<0.05, **P<0.01 and ***P<0.001.

10
Fig. S1. PolySia on dendritic cells is required for regular lymph node homeostasis Results In order to identify polySia on different immune cells we prepared cell suspensions from LNs of St8sia4-/- and control mice and analyzed them by flow cytometry. We could not detect polySia on B and T cells (Figure S1A). Accordingly, lymphocyte trafficking was unaffected in St8sia4-deficient and control mice revealing that polySia is dispensable for T cell homing (Figure S1B). Further classification of dendritic cell subsets revealed polySia expression on the surface of migratory MHCIIhigh/Langerin+ DCs and on conventional CD8+ DCs (Figure S1C) as well as on bone marrow derived DCs (Figure S1D).

11
Figure S1
T cells B cells St8sia4+/+ St8sia4-/-
polySia
St8sia4+/+ St8sia4-/-
PLN
sple
en
A
MHCII
CD11
c
cDCs
polySia
MHCIIhigh St8sia4+/+ St8sia4V-/-
St8sia4+/+ St8sia4V-/-
polySiaCD11b
CD8
CD8+ CD11b+
St8sia4+/+ St8sia4-/-
St8sia4+/+ St8sia4-/-
cDCs
B
C
CD8+%
CD11b+%
0,0
0,4
0,8
1,2
1,6
2,0
2,4
PLN spleen
St8SiaIV+/+St8SiaIV-/-
P=0.213ns
% o
f pos
itive
cells
with
in S
LO
St8sia4+/+ recipient
St8sia4+/+ St8sia4-/-
0,0
0,4
0,8
1,2
1,6
2,0
2,4
PLN spleen
St8SiaIV+/+St8SiaIV-/-
P=0.806ns
% o
f pos
itive
cells
with
in S
LO
St8sia4-/- recipient
St8sia4+/+ St8sia4-/-
imDCs mDCs
220
120100
80
6050
40
30
20
MW[kDa]St8sia4 +/+ -/- +/+ -/-
α-polySia
mSt8sia4+/+mSt8sia4-/-
imSt8sia4+/+imSt8sia4-/-
unstained
polySia
D

12
Legend (A) Flow cytometry of lymphocytes isolated from either PLNs or spleen of St8sia4-/- and control mice and analyzed for cell surface polySia levels. B and T cells were specified according to B220+ and CD3+ staining and analyzed for cell surface polySia levels. PolySia staining has been carried out with inactive EndoN-GFP. (B) In vivo short-term homing of St8sia4-/- and control lymphocytes in wild type and polySia-deficient recipient mice. St8sia4-/- and control lymphocytes were isolated from PLNs and spleen and labeled with 10μM TAMRA or 5μM CFSE, respectively. PLNs and spleens of recipient mice were removed three hours after injection and analyzed by flow cytometry. Graphs depict average percentage of fluorescent cells within SLOs of at least 5 different mice per genotype ±SD. Differences between the two groups were examined by two-tailed unpaired student’s t-test. (C) Specification of DC subsets positive for cell surface polySia under steady state conditions. DC sub-populations were classified according to CD11b+, CD8+ and Langerin+ staining. (D) (left) Immunodetection of total polySia levels in whole cell lysates of immature CD11c+ MHCIIlow (iDCs) and mature CD11c+ MHCIIhigh (mDCs) BMDCs. (right) Cell surface polySia levels in immature and mature BMDCs.

13
Fig. S2. Non-polysialylated CCR7 shows impaired signaling towards CCL21 Results Upon chemokine binding, CCR7-mediated G protein activation triggers distinct intracellular signaling cascades involving MAP kinases as well as the PI3K/Akt pathway (33). To dissect the contribution of these signaling pathways to polySia-mediated chemotaxis we stimulated LPS-treated St8sia4-/- and control BMDCs with either CCL19 or CCL21 and compared the levels of phosphorylated effector kinases. CCL19 stimulation led to a transient phosphorylation of Akt and Erk in both cell types with strongest signals 2-5 min after stimulation (Figure S2). However, phosphorylation levels of both kinases in St8sia4-/- DCs were significantly lower after CCL21 stimulation, indicating that in the absence of polySia, CCR7 does not trigger intracellular signaling cascades upon CCL21 stimulation. These results are consistent with the observed migration defect of St8sia4-/- DCs.

14
St8sia4-/-St8sia4+/+
CCL19
0 2 5 15 30’ 0 2 5 15 30’
60
50
MW [kDa]
60
50
α!P-Akt
α-Akt
α-Erk
α!P-Erk
50
40
50
40
CCL21
0 2 5 15 30’ 0 2 5 15 30’
α-P-Akt
α!Akt
60
50
60
50
St8sia4+/+ St8sia4-/-
α-P-Erk
α-Erk
50
40
50
40
MW [kDa]
Figure S2

15
Legend BMDCs from St8sia4-/- and control mice were maturated with LPS over night and adjusted to equal cell numbers the next day. Cells were stimulated with 20ng/ml CCL19 (left panel) or CCL21 (right panel) for the indicated time points and immediately harvested and lysed on ice. Membranes were probed with specific phospho-kinase antibodies as well as total kinase levels. Shown are representative immunoblots of three independent experiments.

16
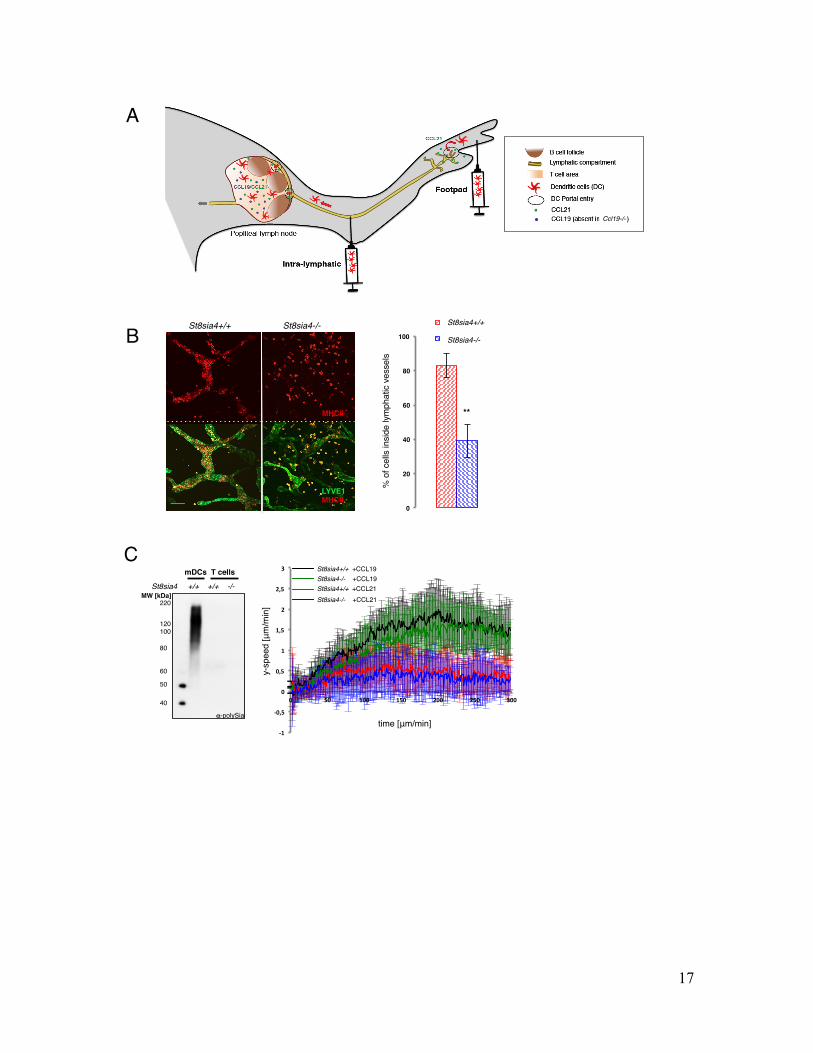
Fig. S3. Polysialylation affects CCL21 sensing in peripheral tissues
17
!1#
!0,5#
0#
0,5#
1#
1,5#
2#
2,5#
3#
0# 50# 100# 150# 200# 250# 300#
St8SiaIV+/+ +CCL19
St8SiaIV-/- +CCL19
St8SiaIV+/+ +CCL21
St8SiaIV-/- +CCL21
time [μm/min]
y-sp
eed
[μm
/min
]
St8sia4+/+ +CCL19St8sia4-/- +CCL19St8sia4+/+ +CCL21St8sia4-/- +CCL21220
120100
80
60
50
40
St8sia4 +/+ +/+ -/-
mDCs T cells
α-polySia
MW [kDa]
Figure S3
A
BSt8sia4+/+ St8sia4-/-
MHCII
LYVE1MHCII
% o
f cel
ls in
side
lym
phat
ic v
esse
ls
0
20
40
60
80
100
St8SiaIV+/+
St8SiaIV-/-
**
St8sia4+/+
St8sia4-/-
Ccl19-/-
C

18
Legend (A) Schematic representation of experimental setup and chemokine guidance of in vivo DC migration. In contrast to footpad injections, fluorescently labeled BMDCs were injected directly within the afferent lymphatic vessel of Ccl19-/- recipient mice. Draining popliteal LNs were analyzed by immunohistology 10 hours later. (B) (left) z-stack projection of split ear sheets stained against LYVE1 (green) and MHCII (red). Respective genotypes are indicated above. (right) Quantification of DCs inside lymphatic vessels. Bars represent average values ±SD of five different fields per ear of three different animals. Differences between the two groups were examined by two-tailed unpaired student’s t-test. Scale bar: 100μm. (C) (left) Immunoblotting of polySia in activated St8sia4-/- and activated control T cells in comparison to mature dendritic cells (mDCs). (right) Activated St8sia4-/- and control T cells were embedded within 3D collagen I matrices and exposed to soluble gradients of CCL19 and CCL21. Average y-directed velocities ±SD of 6 experiments with cells from 3 different mice are shown over time.

19
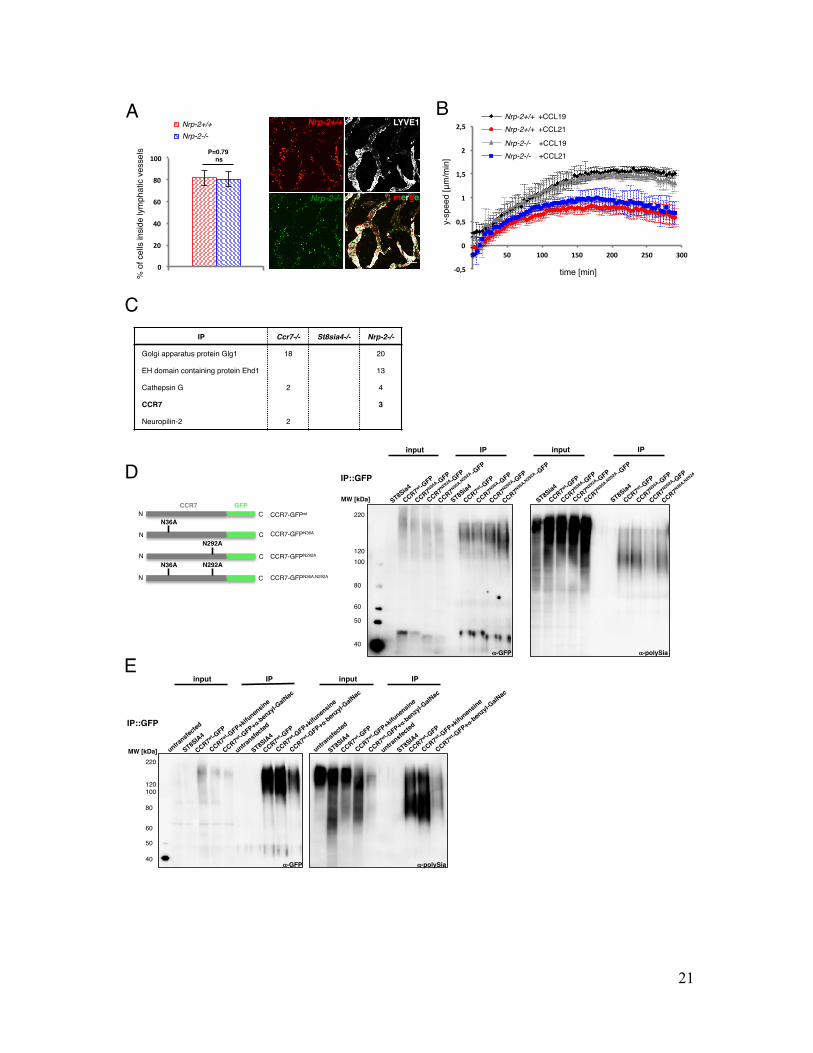
Fig. S4. Migration of Nrp2-deficient dendritic cells and polysialylation of CCR7 on mature dendritic cells Results Migration of Nrp2-deficient dendritic cells Neuropilin-2 (NRP-2) is the only known potential carrier of polySia, which is abundantly expressed on DCs and as such has been implicated in CCL21 responses (7-9, 23). To address whether loss of polysialylated NRP-2 is responsible for the DC-specific migration defect in St8sia4-/- animals we used ear explants and BMDCs from Nrp-2-deficient mice (34) and determined migration as previously described ((17); Figure S4A). BMDCs from Nrp-2-/- and control mice both entered the lymphatic vessels equally well when incubated on wildtype ear explants suggesting that NRP-2 is dispensable for DC migration in vivo. Accordingly, analysis of gradient-directed velocities, as described in the main text, did not reveal a difference between Nrp-2-/- and control BMDC migration towards CCL19 or CCL21 (Figure S4B). These data demonstrate that polysialylated NRP-2 is dispensable for chemokine sensing of murine DCs. Furthermore, so far unidentified polySia carrier must be responsible for the observed migration defect in St8sia4-deficient mice. Polysialylation of CCR7 on mature dendritic cells To identify additional polySia carriers, proteins from whole cell lysates of Nrp-2-/- BMDCs were immunoprecipitated using inactive EndoN covalently coupled to magnetic beads. The protein fraction was analyzed by mass spectrometry leading to four so far unknown polysialylated proteins with CCR7 as the sole cell surface protein (Figure S4C). To confirm modification of CCR7, we reconstituted CCR7 polysialylation in HEK293 cells. Due to the lack of specific antibodies, we expressed CCR7-green fluorescent protein (GFP) fusion protein together with the polysialyltransferase ST8SiaIV in HEK293 cells. Immunoprecipitation of CCR7-GFP and subsequent separation of whole cell lysates by SDS-PAGE followed by immunoblotting revealed a dispersed signal between 220 and 120kDa (Figure 3A). In addition to glycosylation, CCR7 was shown to be constitutively ubiquitylated on several lysine residues in HEK293 cells (35) explaining the highly dispersed signal of CCR7-GFP at high molecular weight. Glycans can be attached to the free amide nitrogen of an asparagine residue placed in the sequence Asn-X-Ser/Thr (N-linked glycosylation) or to the hydroxyl oxygen of serine and threonine residues (O-linked glycosylation). Since polySia is attached to the extracellular part of the receptor, we searched for putative glycosylation sites within the N-terminus and the three extracellular loops of CCR7, a heptahelical transmembrane GPCR, and found two matching consensus sequences allowing N-glycosylation (Asn36 and Asn292). To confirm polysialylation of N-glycans at these sites, Asn36 and Asn292 were mutated to alanine by site-specific mutagenesis leading to three different glycosylation-mutants (Figure S4D, left panel). Whole cell lysates and immunoprecipitates from N-glycosylation deficient mutants of CCR7-GFP showed a shift in mobility towards lower molecular weight indicating that CCR7 is indeed N-glycosylated (Figure S4D, right panel). Yet, pull-down of CCR7-GFP and

20
immunodetection with mAb 735 revealed polysialylation of CCR7 even for the double mutant (CCR7N36A,N292A) indicating additional attachment sites of polySia chains presumably to O-linked glycans. To distinguish between N- and O-linked polysialylation of CCR7, we treated HEK293 cells with the glycosylation inhibitors kifunensine and α-benzyl GalNAc. Kifunensine inhibits N-glycosylation by preventing α-mannosidase I from trimming mannose residues from precursor glycoproteins. α-benzyl GalNAc is a structural analogue of GalNAc-Ser/Thr and blocks O-glycan synthesis as it is used as surrogate substrate or decoy by glycosyltransferases involved in the extension of GalNAc-Ser/Thr sites. Treatment of HEK cells with either of the two inhibitors did not interfere with cell growth and cell surface CCR7-GFP expression (data not shown). However, treatment with both inhibitors strongly interfered with cell growth and led to dettachment of cells. Consequently transfection rates with CCR7-GFP were very poor (data not shown). PolySia levels from whole cell lysates were strongly reduced after inhibition of O-linked glycosylation (Figure S4E, right panel, input). Immunoprecipitation of CCR7-GFP and subsequent immunodetection of polySia still showed polysialylation of CCR7 after inhibition of either N- or O-linked glycosylation (Figure S4E, right panel, IP) verifying polysialylation of CCR7 on N- and O-glycosylation sites. Alltogether these results establish CCR7 as a highly glycosylated protein with terminal polySia residues attached to N- and O-linked glycans.
21
Figure S4
A B
merge
LYVE1
Nrp-2-/-
Nrp-2+/+
0"
20"
40"
60"
80"
100"
1"
NRP-2 +/+NRP-2 -/-
% o
f cel
ls in
side
lym
phat
ic v
esse
ls P=0.79ns
Nrp-2+/+ Nrp-2-/-
(0,5"
0"
0,5"
1"
1,5"
2"
2,5"
0" 50" 100" 150" 200" 250" 300"
NRP-2+/+ +CCL19NRP-2+/+ +CCL21NRP-2-/- +CCL19NRP-2-/- +CCL21
y-sp
eed
[μm
/min
]
time [min]
Nrp-2+/+ +CCL19Nrp-2+/+ +CCL21
Nrp-2-/- +CCL19Nrp-2-/- +CCL21
C
220
120100
80
60
50
40α-GFP α-polySia
MW [kDa]
IP::GFP
input IP input IP
D
CCR7-GFPwt
CCR7-GFPN36A
CCR7-GFPN292A
CCR7-GFPN36A,N292A
N C
N C
N C
N C
N36A
N36A
N292A
N292A
CCR7 GFP
IP::GFP
α-polySia
220
120100
80
60
50
40
MW [kDa]
α-GFP
input IP input IP
IP Ccr7-/- St8sia4-/- Nrp-2-/-
Golgi apparatus protein Glg1 18 20
EH domain containing protein Ehd1 13
Cathepsin G 2 4
CCR7 3
Neuropilin-2 2
E

22
Legend (A) (right) z-stack projection of wild type split ear sheets incubated with eFluor630-labeled Nrp-2-/- (green) and TAMRA-labeled control (red) BMDCs. Ears were stained against LYVE1 (Alexa488; grey) to visualize lymphatic vessels. (left) Quantification of cells inside lymphatic vessels. Bars indicate average values ±SD of five different fields per view of three independent experiments. Scale bar: 100μm. (B) Mature Nrp-2 and control BMDCs were embedded within 3D collagen I matrices and exposed to soluble gradients of CCL19 and CCL21. Average y-directed velocities ±SD of 6 experiments with cells from 3 different mice are shown over time. (C) List of proteins containing peptides identified by mass spectrometry after pull-down of polySia with iEndoN-coupled magnetic beads in Nrp-2-/- BMDCs. Numbers indicate different peptides of the respective protein identified. Only the most abundant proteins with at least two peptides matching the detected sequences with >99% probability were considered. (D) (left panel) Ccr7-gfp constructs. Due to the lack of specific antibodies, CCR7 was expressed as GFP-fusion protein. Putative N-glycosylation sites were mutated to alanine by site-directed mutagenesis (N36A, N292A). All Ccr7-gfp constructs were co-expressed together with the polysialyltransferase ST8Sia IV. (right panel) Pull-down of CCR7-GFP constructs from HEK293 cells using α-GFP-coupled magnetic beads. CCR7-GFP and polySia were detected using α-GFP and monoclonal antibody 735, respectively. (E) Inhibition of N- and O-glycosylation in HEK293 cells co-transfected with Ccr7-gfp and St8sia4 plasmids. Cells were treated with kifunensine and α-benzyl GalNAc for 4 days before pull-down of CCR7-GFP using α-GFP-coupled magnetic beads. CCR7-GFP and polySia were detected as described above.

23
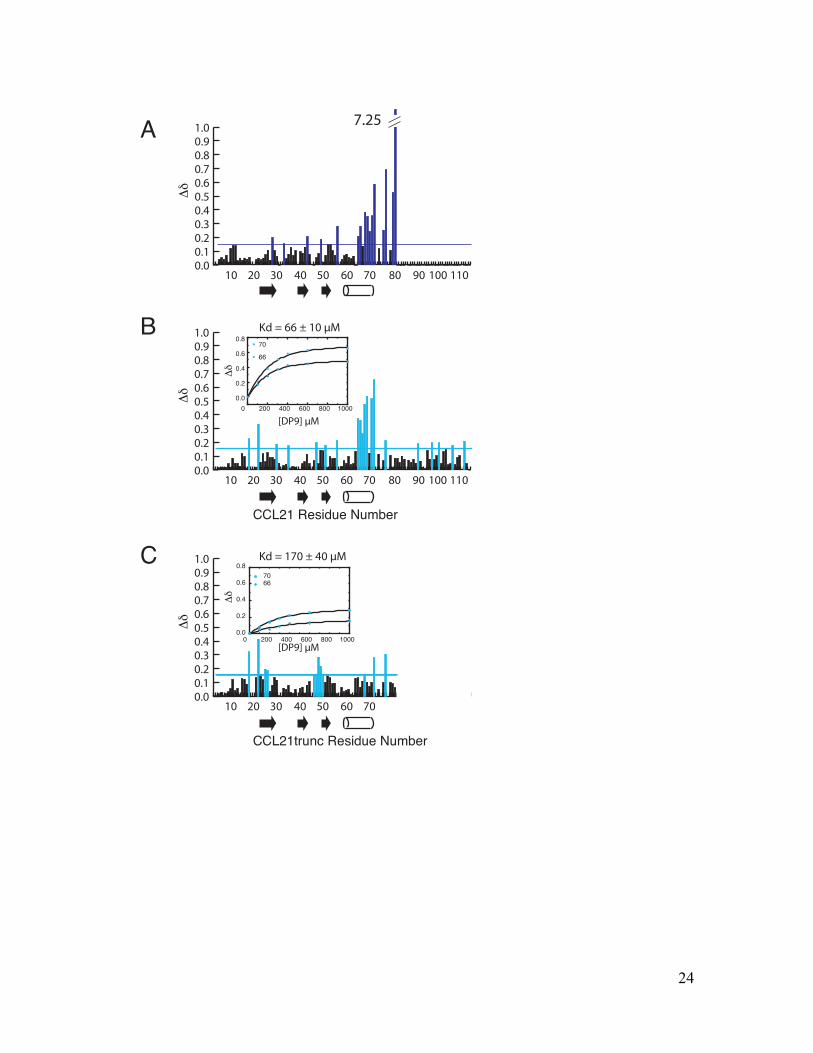
Fig. S5. Identification of an autoinhibitory interaction site within CCL21
24
0.00.10.20.30.40.50.60.70.80.91.0 7.25
10 20 30 40 50 60 70 80 90 100 110
Δδ
A
B
C
0.00.10.20.30.40.50.60.70.80.91.0
10 20 30 40 50 60 70 80 90 100 110
Δδ
[DP9] µM
Δδ
[DP9] µM
Δδ
0 200 400 600 800 10000.0
0.2
0.4
0.6
0.870
66
Kd = 66 ± 10 µM
Kd = 170 ± 40 µM
0.00.10.20.30.40.50.60.70.80.91.0
10 20 30 40 50 60 70 80 90 100 110
Δδ
CCL21 Residue Number
CCL21trunc Residue Number
0 200 400 600 800 10000.0
0.2
0.4
0.6
0.8
6670
Figure S5
24

25
Legend
(A) Chemical shift differences (Dd) between 200 µM CCL21trunc 1-79 and CCL21-FL 1-111 plotted versus CCL21 residue number to identify CCL21 residues involved in autoinhibition. (B) CCL21-FL 1-111 chemical shift perturbations induced by a 5-fold molar excess of polySia (DP9) plotted versus CCL21 residue number. The insert shows a plot of CCL21 chemical shift perturbations for residue 66 (dots) and 70 (diamonds) upon titration with DP9. The Kd was determined using a model that takes into account ligand depletion and non-linear regression. (C) CCL21trunc 1-79 chemical shift perturbations induced by a 5-fold molar excess of polySia (DP9) plotted versus CCL21trunc residue number. The insert shows a plot of CCL21trunc chemical shift perturbations upon titration with DP9. The Kd was determined using a model that takes into account ligand depletion and non-linear regression.

26
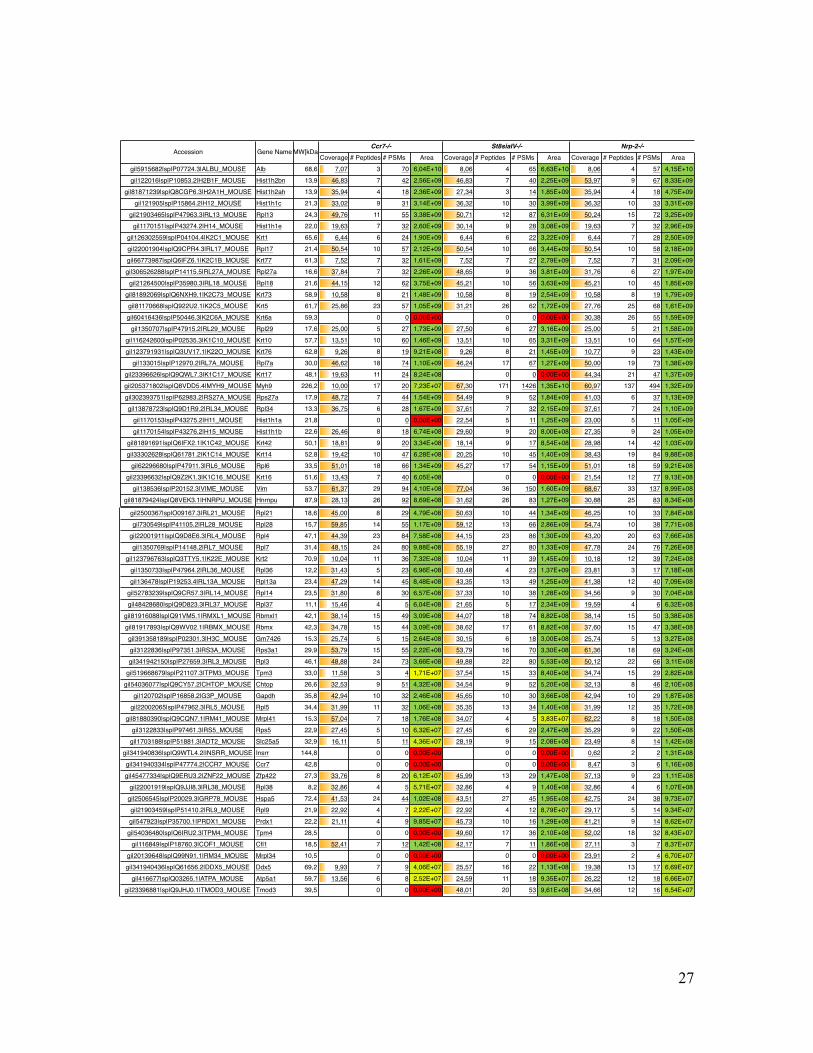
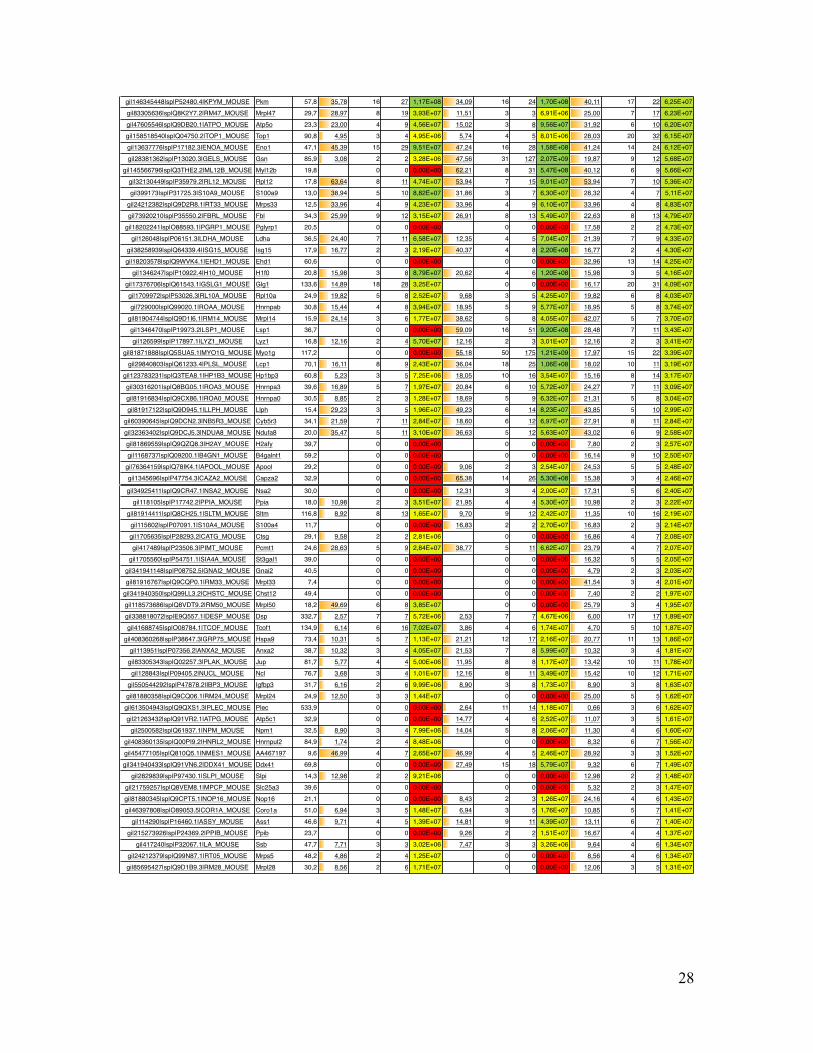
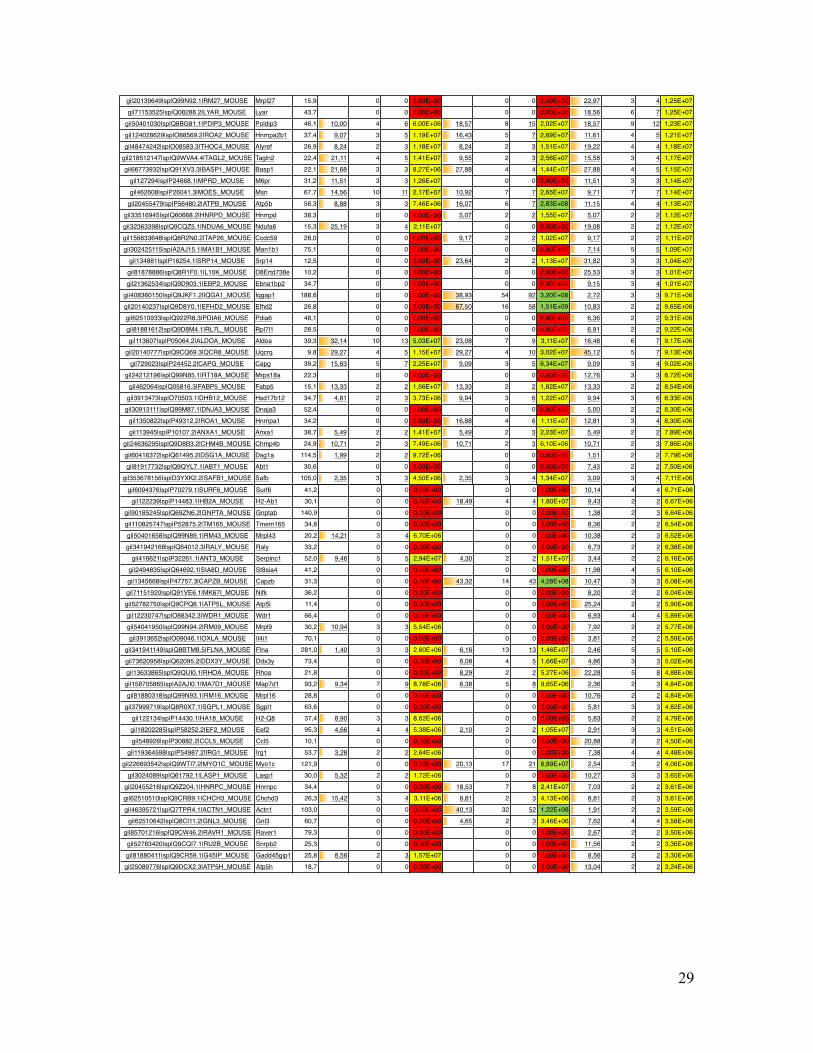
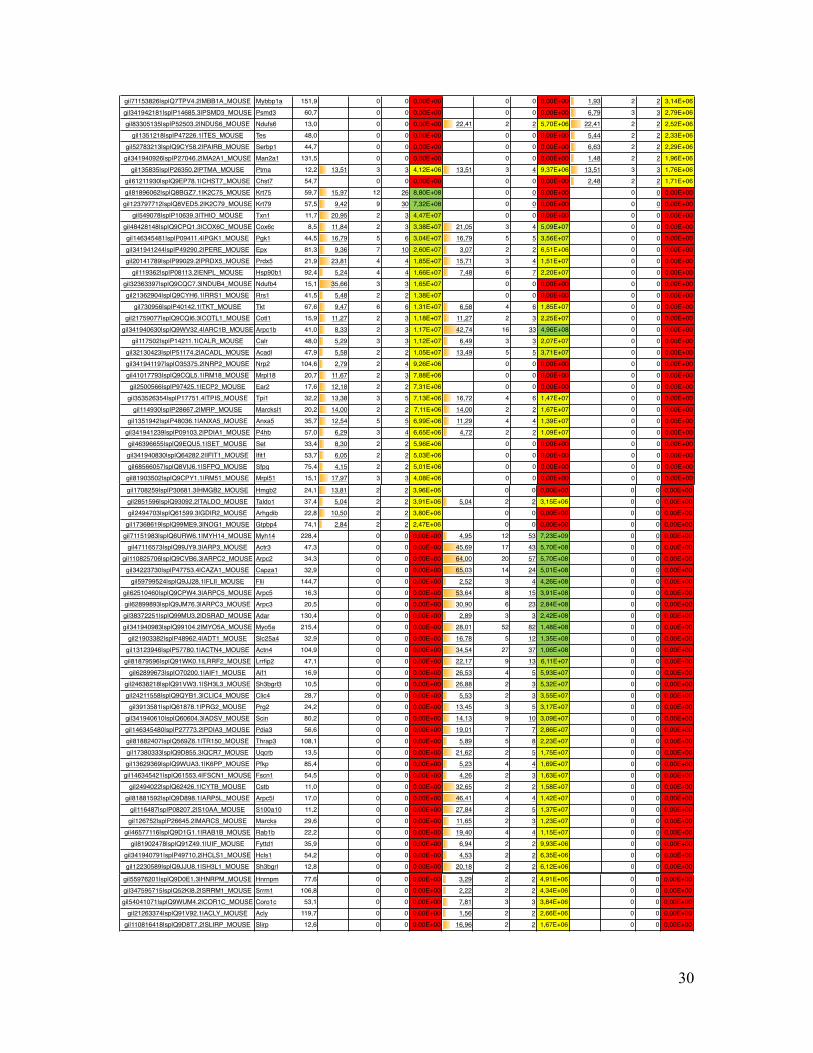
Supplementary Table Table S1. List of all identified mouse proteins by MS Amanda including their accession number, gene name, molecular weight, sequence coverage, number of peptides, number of peptide spectrum matches (PSMs) and precursor area.
27
26
Coverage # Peptides # PSMs Area Coverage # Peptides # PSMs Area Coverage # Peptides # PSMs Areagi|5915682|sp|P07724.3|ALBU_MOUSE Alb 68,6 7,07 3 70 6,04E+10 8,06 4 65 6,63E+10 8,06 4 57 4,15E+10gi|122016|sp|P10853.2|H2B1F_MOUSE Hist1h2bn 13,9 46,83 7 42 2,56E+09 46,83 7 40 2,25E+09 53,97 9 67 8,33E+09
gi|81871239|sp|Q8CGP6.3|H2A1H_MOUSE Hist1h2ah 13,9 35,94 4 18 2,36E+09 27,34 3 14 1,85E+09 35,94 4 18 4,75E+09gi|121905|sp|P15864.2|H12_MOUSE Hist1h1c 21,3 33,02 9 31 3,14E+09 36,32 10 30 3,99E+09 36,32 10 33 3,31E+09
gi|21903465|sp|P47963.3|RL13_MOUSE Rpl13 24,3 49,76 11 55 3,38E+09 50,71 12 87 6,31E+09 50,24 15 72 3,25E+09gi|1170151|sp|P43274.2|H14_MOUSE Hist1h1e 22,0 19,63 7 32 2,60E+09 30,14 9 28 3,08E+09 19,63 7 32 2,96E+09
gi|126302559|sp|P04104.4|K2C1_MOUSE Krt1 65,6 6,44 6 24 1,90E+09 6,44 6 22 3,22E+09 6,44 7 28 2,50E+09gi|22001904|sp|Q9CPR4.3|RL17_MOUSE Rpl17 21,4 50,54 10 57 2,12E+09 50,54 10 66 3,44E+09 50,54 10 58 2,18E+09gi|66773987|sp|Q6IFZ6.1|K2C1B_MOUSE Krt77 61,3 7,52 7 32 1,61E+09 7,52 7 27 2,79E+09 7,52 7 31 2,09E+09gi|306526288|sp|P14115.5|RL27A_MOUSE Rpl27a 16,6 37,84 7 32 2,26E+09 48,65 9 36 3,81E+09 31,76 6 27 1,97E+09
gi|21264500|sp|P35980.3|RL18_MOUSE Rpl18 21,6 44,15 12 62 3,75E+09 45,21 10 56 3,63E+09 45,21 10 45 1,85E+09gi|81892069|sp|Q6NXH9.1|K2C73_MOUSE Krt73 58,9 10,58 8 21 1,48E+09 10,58 8 19 2,54E+09 10,58 8 19 1,79E+09gi|81170668|sp|Q922U2.1|K2C5_MOUSE Krt5 61,7 25,86 23 57 1,05E+09 31,21 26 62 1,72E+09 27,76 25 68 1,61E+09gi|60416436|sp|P50446.3|K2C6A_MOUSE Krt6a 59,3 0 0 0,00E+00 0 0 0,00E+00 30,38 26 55 1,59E+09
gi|1350707|sp|P47915.2|RL29_MOUSE Rpl29 17,6 25,00 5 27 1,73E+09 27,50 6 27 3,16E+09 25,00 5 21 1,58E+09gi|116242600|sp|P02535.3|K1C10_MOUSE Krt10 57,7 13,51 10 60 1,46E+09 13,51 10 65 3,31E+09 13,51 10 64 1,57E+09gi|123791931|sp|Q3UV17.1|K22O_MOUSE Krt76 62,8 9,26 8 19 9,21E+08 9,26 8 21 1,45E+09 10,77 9 23 1,43E+09
gi|133015|sp|P12970.2|RL7A_MOUSE Rpl7a 30,0 46,62 18 74 1,10E+09 46,24 17 67 1,27E+09 50,00 19 73 1,38E+09gi|23396626|sp|Q9QWL7.3|K1C17_MOUSE Krt17 48,1 19,63 11 24 8,24E+08 0 0 0,00E+00 44,34 21 47 1,37E+09gi|205371802|sp|Q8VDD5.4|MYH9_MOUSE Myh9 226,2 10,00 17 20 7,23E+07 67,30 171 1426 1,35E+10 60,97 137 494 1,32E+09gi|302393751|sp|P62983.2|RS27A_MOUSE Rps27a 17,9 48,72 7 44 1,54E+09 54,49 9 52 1,84E+09 41,03 6 37 1,13E+09gi|13878723|sp|Q9D1R9.2|RL34_MOUSE Rpl34 13,3 36,75 6 28 1,67E+09 37,61 7 32 2,15E+09 37,61 7 24 1,10E+09
gi|1170153|sp|P43275.2|H11_MOUSE Hist1h1a 21,8 0 0 0,00E+00 22,54 5 11 1,25E+09 23,00 5 11 1,05E+09gi|1170154|sp|P43276.2|H15_MOUSE Hist1h1b 22,6 26,46 8 18 6,74E+08 29,60 9 20 8,00E+08 27,35 9 24 1,05E+09
gi|81891691|sp|Q6IFX2.1|K1C42_MOUSE Krt42 50,1 18,81 9 20 3,34E+08 18,14 9 17 8,54E+08 28,98 14 42 1,03E+09gi|33302628|sp|Q61781.2|K1C14_MOUSE Krt14 52,8 19,42 10 47 6,28E+08 20,25 10 45 1,40E+09 38,43 19 84 9,88E+08
gi|62296680|sp|P47911.3|RL6_MOUSE Rpl6 33,5 51,01 18 66 1,34E+09 45,27 17 54 1,15E+09 51,01 18 59 9,21E+08gi|23396632|sp|Q9Z2K1.3|K1C16_MOUSE Krt16 51,6 13,43 7 40 6,05E+08 0 0 0,00E+00 21,54 12 77 9,13E+08
gi|138536|sp|P20152.3|VIME_MOUSE Vim 53,7 61,37 29 94 4,10E+08 77,04 36 150 1,60E+09 68,67 33 137 8,99E+08gi|81879424|sp|Q8VEK3.1|HNRPU_MOUSE Hnrnpu 87,9 28,13 26 92 8,69E+08 31,62 26 83 1,27E+09 30,88 25 83 8,34E+08
Table S1Ccr7-/- St8siaIV-/- Nrp-2-/-
Accession Gene Name MW[kDa]
26
gi|2500367|sp|O09167.3|RL21_MOUSE Rpl21 18,6 45,00 8 29 4,79E+08 50,63 10 44 1,34E+09 46,25 10 33 7,84E+08gi|730549|sp|P41105.2|RL28_MOUSE Rpl28 15,7 59,85 14 55 1,17E+09 59,12 13 66 2,86E+09 54,74 10 38 7,71E+08gi|22001911|sp|Q9D8E6.3|RL4_MOUSE Rpl4 47,1 44,39 23 84 7,58E+08 44,15 23 86 1,30E+09 43,20 20 63 7,66E+08gi|1350769|sp|P14148.2|RL7_MOUSE Rpl7 31,4 48,15 24 80 9,88E+08 55,19 27 80 1,33E+09 47,78 24 76 7,26E+08
gi|123796763|sp|Q3TTY5.1|K22E_MOUSE Krt2 70,9 10,04 11 36 7,32E+08 10,04 11 39 1,45E+09 10,18 12 39 7,24E+08gi|1350733|sp|P47964.2|RL36_MOUSE Rpl36 12,2 31,43 5 23 6,96E+08 30,48 4 23 1,37E+09 23,81 3 17 7,18E+08gi|136478|sp|P19253.4|RL13A_MOUSE Rpl13a 23,4 47,29 14 45 8,48E+08 43,35 13 49 1,25E+09 41,38 12 40 7,09E+08gi|52783239|sp|Q9CR57.3|RL14_MOUSE Rpl14 23,5 31,80 8 30 6,57E+08 37,33 10 38 1,28E+09 34,56 9 30 7,04E+08gi|48428680|sp|Q9D823.3|RL37_MOUSE Rpl37 11,1 15,46 4 5 6,04E+08 21,65 5 17 2,34E+09 19,59 4 6 6,32E+08gi|81916088|sp|Q91VM5.1|RMXL1_MOUSE Rbmxl1 42,1 38,14 15 49 3,09E+08 44,07 18 74 8,82E+08 38,14 15 50 3,38E+08gi|81917893|sp|Q9WV02.1|RBMX_MOUSE Rbmx 42,3 34,78 15 44 3,09E+08 38,62 17 61 8,82E+08 37,60 15 47 3,38E+08gi|391358189|sp|P02301.3|H3C_MOUSE Gm7426 15,3 25,74 5 15 2,64E+08 30,15 6 18 3,00E+08 25,74 5 13 3,27E+08gi|3122836|sp|P97351.3|RS3A_MOUSE Rps3a1 29,9 53,79 15 55 2,22E+08 53,79 16 70 3,30E+08 61,36 18 69 3,24E+08gi|341942150|sp|P27659.3|RL3_MOUSE Rpl3 46,1 48,88 24 73 3,66E+08 49,88 22 80 5,53E+08 50,12 22 66 3,11E+08gi|519668679|sp|P21107.3|TPM3_MOUSE Tpm3 33,0 11,58 3 4 1,71E+07 37,54 15 33 8,40E+08 34,74 15 29 2,82E+08gi|54036077|sp|Q9CY57.2|CHTOP_MOUSE Chtop 26,6 32,53 9 51 4,32E+08 34,54 9 52 5,20E+08 32,13 8 46 2,10E+08
gi|120702|sp|P16858.2|G3P_MOUSE Gapdh 35,8 42,94 10 32 2,46E+08 45,65 10 30 3,66E+08 42,94 10 29 1,87E+08gi|22002065|sp|P47962.3|RL5_MOUSE Rpl5 34,4 31,99 11 32 1,06E+08 35,35 13 34 1,40E+08 31,99 12 35 1,72E+08
gi|81880390|sp|Q9CQN7.1|RM41_MOUSE Mrpl41 15,3 57,04 7 18 1,76E+08 34,07 4 5 3,83E+07 62,22 8 18 1,50E+08gi|3122833|sp|P97461.3|RS5_MOUSE Rps5 22,9 27,45 5 10 6,32E+07 27,45 6 29 2,47E+08 35,29 9 22 1,50E+08gi|1703188|sp|P51881.3|ADT2_MOUSE Slc25a5 32,9 16,11 5 11 4,36E+07 28,19 9 15 2,08E+08 23,49 8 14 1,42E+08
gi|341940836|sp|Q9WTL4.2|INSRR_MOUSE Insrr 144,8 0 0 0,00E+00 0 0 0,00E+00 0,62 2 2 1,31E+08gi|341940334|sp|P47774.2|CCR7_MOUSE Ccr7 42,8 0 0 0,00E+00 0 0 0,00E+00 8,47 3 6 1,16E+08gi|45477334|sp|Q9ERU3.2|ZNF22_MOUSE Zfp422 27,3 33,76 8 20 6,12E+07 45,99 13 29 1,47E+08 37,13 9 23 1,11E+08gi|22001919|sp|Q9JJI8.3|RL38_MOUSE Rpl38 8,2 32,86 4 5 5,71E+07 32,86 4 9 1,40E+08 32,86 4 6 1,07E+08gi|2506545|sp|P20029.3|GRP78_MOUSE Hspa5 72,4 41,53 24 44 1,02E+08 43,51 27 45 1,95E+08 42,75 24 38 9,73E+07gi|21903459|sp|P51410.2|RL9_MOUSE Rpl9 21,9 22,92 4 7 2,22E+07 22,92 4 12 8,79E+07 29,17 5 14 9,34E+07gi|547923|sp|P35700.1|PRDX1_MOUSE Prdx1 22,2 21,11 4 9 9,85E+07 45,73 10 16 1,29E+08 41,21 9 14 8,62E+07gi|54036480|sp|Q6IRU2.3|TPM4_MOUSE Tpm4 28,5 0 0 0,00E+00 49,60 17 36 2,10E+08 52,02 18 32 8,43E+07gi|116849|sp|P18760.3|COF1_MOUSE Cfl1 18,5 52,41 7 12 1,42E+08 42,17 7 11 1,86E+08 27,11 3 7 8,37E+07
gi|20139648|sp|Q99N91.1|RM34_MOUSE Mrpl34 10,5 0 0 0,00E+00 0 0 0,00E+00 23,91 2 4 6,70E+07gi|341940436|sp|Q61656.2|DDX5_MOUSE Ddx5 69,2 9,93 7 9 4,06E+07 25,57 16 22 1,13E+08 19,38 13 17 6,69E+07gi|416677|sp|Q03265.1|ATPA_MOUSE Atp5a1 59,7 13,56 6 8 2,52E+07 24,59 11 18 9,35E+07 26,22 12 18 6,66E+07
gi|23396881|sp|Q9JHJ0.1|TMOD3_MOUSE Tmod3 39,5 0 0 0,00E+00 48,01 20 53 9,61E+08 34,66 12 16 6,54E+07
28
26
gi|146345448|sp|P52480.4|KPYM_MOUSE Pkm 57,8 35,78 16 27 1,17E+08 34,09 16 24 1,70E+08 40,11 17 22 6,25E+07gi|83305636|sp|Q8K2Y7.2|RM47_MOUSE Mrpl47 29,7 28,97 8 19 3,93E+07 11,51 3 3 6,91E+06 25,00 7 17 6,23E+07gi|47605546|sp|Q9DB20.1|ATPO_MOUSE Atp5o 23,3 23,00 4 9 4,56E+07 15,02 3 8 9,56E+07 31,92 6 10 6,20E+07gi|158518540|sp|Q04750.2|TOP1_MOUSE Top1 90,8 4,95 3 4 4,95E+06 5,74 4 5 8,01E+06 28,03 20 32 6,15E+07gi|13637776|sp|P17182.3|ENOA_MOUSE Eno1 47,1 45,39 15 29 9,51E+07 47,24 16 28 1,58E+08 41,24 14 24 6,12E+07gi|28381362|sp|P13020.3|GELS_MOUSE Gsn 85,9 3,08 2 2 3,28E+06 47,56 31 127 2,07E+09 19,87 9 12 5,68E+07
gi|145566796|sp|Q3THE2.2|ML12B_MOUSE Myl12b 19,8 0 0 0,00E+00 62,21 8 31 5,47E+08 40,12 6 9 5,66E+07gi|32130449|sp|P35979.2|RL12_MOUSE Rpl12 17,8 63,64 8 11 4,74E+07 53,94 7 15 9,01E+07 53,94 7 10 5,36E+07gi|399173|sp|P31725.3|S10A9_MOUSE S100a9 13,0 38,94 5 10 8,82E+07 31,86 3 7 6,30E+07 28,32 4 7 5,11E+07gi|24212382|sp|Q9D2R8.1|RT33_MOUSE Mrps33 12,5 33,96 4 9 4,23E+07 33,96 4 9 6,10E+07 33,96 4 8 4,83E+07gi|73920210|sp|P35550.2|FBRL_MOUSE Fbl 34,3 25,99 9 12 3,15E+07 26,91 8 13 5,49E+07 22,63 8 13 4,79E+07gi|18202241|sp|O88593.1|PGRP1_MOUSE Pglyrp1 20,5 0 0 0,00E+00 0 0 0,00E+00 17,58 2 2 4,73E+07gi|126048|sp|P06151.3|LDHA_MOUSE Ldha 36,5 24,40 7 11 6,58E+07 12,35 4 5 7,04E+07 21,39 7 9 4,33E+07
gi|38258939|sp|Q64339.4|ISG15_MOUSE Isg15 17,9 16,77 2 3 2,19E+07 40,37 4 8 2,20E+08 16,77 2 4 4,30E+07gi|18203578|sp|Q9WVK4.1|EHD1_MOUSE Ehd1 60,6 0 0 0,00E+00 0 0 0,00E+00 32,96 13 14 4,25E+07gi|1346247|sp|P10922.4|H10_MOUSE H1f0 20,8 15,98 3 8 8,79E+07 20,62 4 6 1,20E+08 15,98 3 5 4,16E+07
gi|17376706|sp|Q61543.1|GSLG1_MOUSE Glg1 133,6 14,89 18 28 3,25E+07 0 0 0,00E+00 16,17 20 31 4,09E+07gi|1709972|sp|P53026.3|RL10A_MOUSE Rpl10a 24,9 19,82 5 8 2,52E+07 9,68 3 5 4,25E+07 19,82 6 8 4,03E+07gi|729000|sp|Q99020.1|ROAA_MOUSE Hnrnpab 30,8 15,44 4 8 3,94E+07 18,95 5 9 5,77E+07 18,95 5 8 3,74E+07gi|81904744|sp|Q9D1I6.1|RM14_MOUSE Mrpl14 15,9 24,14 3 6 1,77E+07 38,62 5 8 4,05E+07 42,07 5 7 3,70E+07gi|1346470|sp|P19973.2|LSP1_MOUSE Lsp1 36,7 0 0 0,00E+00 59,09 16 51 9,20E+08 28,48 7 11 3,43E+07gi|126599|sp|P17897.1|LYZ1_MOUSE Lyz1 16,8 12,16 2 4 5,70E+07 12,16 2 3 3,01E+07 12,16 2 3 3,41E+07
gi|81871888|sp|Q5SUA5.1|MYO1G_MOUSE Myo1g 117,2 0 0 0,00E+00 55,18 50 175 1,21E+09 17,97 15 22 3,39E+07gi|29840803|sp|Q61233.4|PLSL_MOUSE Lcp1 70,1 16,11 8 9 2,43E+07 36,04 18 25 1,06E+08 18,02 10 11 3,19E+07
gi|123783231|sp|Q3TEA8.1|HP1B3_MOUSE Hp1bp3 60,8 5,23 3 5 7,25E+06 18,05 10 16 3,54E+07 15,16 8 14 3,17E+07gi|30316201|sp|Q8BG05.1|ROA3_MOUSE Hnrnpa3 39,6 16,89 5 7 1,97E+07 20,84 6 10 5,72E+07 24,27 7 11 3,09E+07gi|81916834|sp|Q9CX86.1|ROA0_MOUSE Hnrnpa0 30,5 8,85 2 3 1,28E+07 18,69 5 9 6,32E+07 21,31 5 8 3,04E+07gi|81917122|sp|Q9D945.1|LLPH_MOUSE Llph 15,4 29,23 3 5 1,96E+07 49,23 6 14 8,23E+07 43,85 5 10 2,99E+07gi|60390645|sp|Q9DCN2.3|NB5R3_MOUSE Cyb5r3 34,1 21,59 7 11 2,84E+07 18,60 6 12 6,97E+07 27,91 8 11 2,84E+07gi|32363402|sp|Q9DCJ5.3|NDUA8_MOUSE Ndufa8 20,0 35,47 5 11 3,10E+07 36,63 5 12 5,63E+07 43,02 6 9 2,58E+07gi|81869559|sp|Q9QZQ8.3|H2AY_MOUSE H2afy 39,7 0 0 0,00E+00 0 0 0,00E+00 7,80 2 3 2,57E+07gi|1168737|sp|Q09200.1|B4GN1_MOUSE B4galnt1 59,2 0 0 0,00E+00 0 0 0,00E+00 16,14 9 10 2,50E+07gi|76364159|sp|Q78IK4.1|APOOL_MOUSE Apool 29,2 0 0 0,00E+00 9,06 2 3 2,54E+07 24,53 5 5 2,48E+07gi|1345696|sp|P47754.3|CAZA2_MOUSE Capza2 32,9 0 0 0,00E+00 65,38 14 26 5,30E+08 15,38 3 4 2,46E+07
26
gi|34925411|sp|Q9CR47.1|NSA2_MOUSE Nsa2 30,0 0 0 0,00E+00 12,31 3 4 2,00E+07 17,31 5 6 2,40E+07gi|118105|sp|P17742.2|PPIA_MOUSE Ppia 18,0 10,98 2 3 3,51E+07 21,95 4 4 5,30E+07 10,98 2 3 2,22E+07
gi|81914411|sp|Q8CH25.1|SLTM_MOUSE Sltm 116,8 8,92 8 13 1,65E+07 9,70 9 12 2,42E+07 11,35 10 16 2,19E+07gi|115602|sp|P07091.1|S10A4_MOUSE S100a4 11,7 0 0 0,00E+00 16,83 2 2 2,70E+07 16,83 2 3 2,14E+07gi|1705635|sp|P28293.2|CATG_MOUSE Ctsg 29,1 9,58 2 2 2,81E+06 0 0 0,00E+00 16,86 4 7 2,08E+07gi|417489|sp|P23506.3|PIMT_MOUSE Pcmt1 24,6 28,63 5 9 2,84E+07 38,77 5 11 6,62E+07 23,79 4 7 2,07E+07gi|1705560|sp|P54751.1|SIA4A_MOUSE St3gal1 39,0 0 0 0,00E+00 0 0 0,00E+00 16,32 5 5 2,05E+07gi|341941148|sp|P08752.5|GNAI2_MOUSE Gnai2 40,5 0 0 0,00E+00 0 0 0,00E+00 4,79 2 3 2,03E+07gi|81916767|sp|Q9CQP0.1|RM33_MOUSE Mrpl33 7,4 0 0 0,00E+00 0 0 0,00E+00 41,54 3 4 2,01E+07gi|341940350|sp|Q99LL3.2|CHSTC_MOUSE Chst12 49,4 0 0 0,00E+00 0 0 0,00E+00 7,40 2 2 1,97E+07gi|118573686|sp|Q8VDT9.2|RM50_MOUSE Mrpl50 18,2 49,69 6 8 3,85E+07 0 0 0,00E+00 25,79 3 4 1,95E+07gi|338818072|sp|E9Q557.1|DESP_MOUSE Dsp 332,7 2,57 7 7 5,72E+06 2,53 7 7 4,67E+06 6,00 17 17 1,89E+07gi|41688745|sp|O08784.1|TCOF_MOUSE Tcof1 134,9 6,14 6 16 7,02E+07 3,86 4 6 1,74E+07 4,70 5 10 1,87E+07gi|408360268|sp|P38647.3|GRP75_MOUSE Hspa9 73,4 10,31 5 7 1,13E+07 21,21 12 17 2,16E+07 20,77 11 13 1,86E+07gi|113951|sp|P07356.2|ANXA2_MOUSE Anxa2 38,7 10,32 3 4 4,05E+07 21,53 7 8 5,99E+07 10,32 3 4 1,81E+07gi|83305343|sp|Q02257.3|PLAK_MOUSE Jup 81,7 5,77 4 4 5,00E+06 11,95 8 8 1,17E+07 13,42 10 11 1,78E+07gi|128843|sp|P09405.2|NUCL_MOUSE Ncl 76,7 3,68 3 4 1,01E+07 12,16 8 11 3,49E+07 15,42 10 12 1,71E+07gi|550544292|sp|P47878.2|IBP3_MOUSE Igfbp3 31,7 6,16 2 6 9,99E+06 8,90 3 8 1,73E+07 8,90 3 8 1,63E+07gi|81880358|sp|Q9CQ06.1|RM24_MOUSE Mrpl24 24,9 12,50 3 3 1,44E+07 0 0 0,00E+00 25,00 5 5 1,62E+07gi|613504943|sp|Q9QXS1.3|PLEC_MOUSE Plec 533,9 0 0 0,00E+00 2,64 11 14 1,18E+07 0,66 3 6 1,62E+07gi|21263432|sp|Q91VR2.1|ATPG_MOUSE Atp5c1 32,9 0 0 0,00E+00 14,77 4 6 2,52E+07 11,07 3 5 1,61E+07gi|2500582|sp|Q61937.1|NPM_MOUSE Npm1 32,5 8,90 3 4 7,99E+06 14,04 5 8 2,06E+07 11,30 4 6 1,60E+07
gi|408360135|sp|Q00PI9.2|HNRL2_MOUSE Hnrnpul2 84,9 1,74 2 4 8,48E+06 0 0 0,00E+00 8,32 6 7 1,56E+07gi|45477105|sp|Q810Q5.1|NMES1_MOUSE AA467197 9,6 46,99 4 7 2,65E+07 46,99 4 5 2,46E+07 28,92 3 3 1,52E+07gi|341940433|sp|Q91VN6.2|DDX41_MOUSE Ddx41 69,8 0 0 0,00E+00 27,49 15 18 5,79E+07 9,32 6 7 1,49E+07
gi|2829839|sp|P97430.1|SLPI_MOUSE Slpi 14,3 12,98 2 2 9,21E+06 0 0 0,00E+00 12,98 2 2 1,48E+07gi|21759257|sp|Q8VEM8.1|MPCP_MOUSE Slc25a3 39,6 0 0 0,00E+00 0 0 0,00E+00 5,32 2 3 1,47E+07gi|81880345|sp|Q9CPT5.1|NOP16_MOUSE Nop16 21,1 0 0 0,00E+00 8,43 2 3 1,26E+07 24,16 4 6 1,43E+07gi|46397808|sp|O89053.5|COR1A_MOUSE Coro1a 51,0 6,94 3 5 1,48E+07 6,94 3 5 1,76E+07 10,85 5 7 1,41E+07gi|114290|sp|P16460.1|ASSY_MOUSE Ass1 46,6 9,71 4 5 1,39E+07 14,81 9 11 4,39E+07 13,11 6 7 1,40E+07
gi|215273926|sp|P24369.2|PPIB_MOUSE Ppib 23,7 0 0 0,00E+00 9,26 2 2 1,51E+07 16,67 4 4 1,37E+07gi|417240|sp|P32067.1|LA_MOUSE Ssb 47,7 7,71 3 3 3,02E+06 7,47 3 3 3,26E+06 9,64 4 6 1,34E+07
gi|24212379|sp|Q99N87.1|RT05_MOUSE Mrps5 48,2 4,86 2 4 1,25E+07 0 0 0,00E+00 8,56 4 6 1,34E+07gi|85695427|sp|Q9D1B9.3|RM28_MOUSE Mrpl28 30,2 8,56 2 6 1,71E+07 0 0 0,00E+00 12,06 3 5 1,31E+07
29
26
gi|20139649|sp|Q99N92.1|RM27_MOUSE Mrpl27 15,9 0 0 0,00E+00 0 0 0,00E+00 22,97 3 4 1,25E+07gi|71153525|sp|Q08288.2|LYAR_MOUSE Lyar 43,7 0 0 0,00E+00 0 0 0,00E+00 18,56 6 7 1,25E+07gi|50401030|sp|Q8BG81.1|PDIP3_MOUSE Poldip3 46,1 10,00 4 6 6,00E+06 18,57 8 15 2,02E+07 18,57 9 12 1,23E+07gi|124028629|sp|O88569.2|ROA2_MOUSE Hnrnpa2b1 37,4 9,07 3 5 1,19E+07 16,43 5 7 2,69E+07 11,61 4 5 1,21E+07gi|48474242|sp|O08583.3|THOC4_MOUSE Alyref 26,9 8,24 2 3 1,18E+07 8,24 2 3 1,51E+07 19,22 4 4 1,18E+07gi|218512147|sp|Q9WVA4.4|TAGL2_MOUSE Tagln2 22,4 21,11 4 5 1,41E+07 9,55 2 3 2,56E+07 15,58 3 4 1,17E+07gi|66773932|sp|Q91XV3.3|BASP1_MOUSE Basp1 22,1 21,68 3 3 8,27E+06 27,88 4 4 1,44E+07 27,88 4 5 1,15E+07gi|127294|sp|P24668.1|MPRD_MOUSE M6pr 31,2 11,51 3 3 1,26E+07 0 0 0,00E+00 11,51 3 3 1,14E+07gi|462608|sp|P26041.3|MOES_MOUSE Msn 67,7 14,56 10 11 2,17E+07 10,92 7 7 2,65E+07 9,71 7 7 1,14E+07gi|20455479|sp|P56480.2|ATPB_MOUSE Atp5b 56,3 8,88 3 3 7,46E+06 16,07 6 7 2,83E+08 11,15 4 4 1,13E+07gi|33516945|sp|Q60668.2|HNRPD_MOUSE Hnrnpd 38,3 0 0 0,00E+00 5,07 2 2 1,55E+07 5,07 2 2 1,12E+07gi|32363398|sp|Q9CQZ5.1|NDUA6_MOUSE Ndufa6 15,3 25,19 3 4 2,11E+07 0 0 0,00E+00 19,08 2 2 1,12E+07gi|156633648|sp|Q8R2N0.2|TAP26_MOUSE Ccdc59 28,0 0 0 0,00E+00 9,17 2 2 1,02E+07 9,17 2 2 1,11E+07gi|302425115|sp|A2AJ15.1|MA1B1_MOUSE Man1b1 75,1 0 0 0,00E+00 0 0 0,00E+00 7,14 5 5 1,09E+07gi|134881|sp|P16254.1|SRP14_MOUSE Srp14 12,5 0 0 0,00E+00 23,64 2 2 1,13E+07 31,82 3 3 1,04E+07gi|81878886|sp|Q8R1F0.1|L10K_MOUSE D8Ertd738e 10,2 0 0 0,00E+00 0 0 0,00E+00 25,53 3 3 1,01E+07gi|21362534|sp|Q9D903.1|EBP2_MOUSE Ebna1bp2 34,7 0 0 0,00E+00 0 0 0,00E+00 9,15 3 4 1,01E+07gi|408360150|sp|Q9JKF1.2|IQGA1_MOUSE Iqgap1 188,6 0 0 0,00E+00 38,93 54 92 3,20E+08 2,72 3 3 9,71E+06gi|20140237|sp|Q9D8Y0.1|EFHD2_MOUSE Efhd2 26,8 0 0 0,00E+00 67,50 16 58 1,51E+09 10,83 2 2 9,65E+06gi|62510933|sp|Q922R8.3|PDIA6_MOUSE Pdia6 48,1 0 0 0,00E+00 0 0 0,00E+00 6,36 2 2 9,31E+06gi|81881612|sp|Q9D8M4.1|RL7L_MOUSE Rpl7l1 28,5 0 0 0,00E+00 0 0 0,00E+00 6,91 2 2 9,22E+06gi|113607|sp|P05064.2|ALDOA_MOUSE Aldoa 39,3 32,14 10 13 5,03E+07 23,08 7 9 3,11E+07 16,48 6 7 9,17E+06gi|20140777|sp|Q9CQ69.3|QCR8_MOUSE Uqcrq 9,8 29,27 4 5 1,15E+07 29,27 4 10 3,02E+07 45,12 5 7 9,13E+06gi|729023|sp|P24452.2|CAPG_MOUSE Capg 39,2 15,63 5 7 2,25E+07 9,09 3 5 6,34E+07 9,09 3 4 9,02E+06
gi|24212196|sp|Q99N85.1|RT18A_MOUSE Mrps18a 22,3 0 0 0,00E+00 0 0 0,00E+00 12,76 3 3 8,72E+06gi|462064|sp|Q05816.3|FABP5_MOUSE Fabp5 15,1 13,33 2 2 1,56E+07 13,33 2 2 1,82E+07 13,33 2 2 8,54E+06gi|3913473|sp|O70503.1|DHB12_MOUSE Hsd17b12 34,7 4,81 2 3 3,73E+06 9,94 3 6 1,22E+07 9,94 3 6 8,33E+06gi|30913111|sp|Q99M87.1|DNJA3_MOUSE Dnaja3 52,4 0 0 0,00E+00 0 0 0,00E+00 5,00 2 2 8,30E+06gi|1350822|sp|P49312.2|ROA1_MOUSE Hnrnpa1 34,2 0 0 0,00E+00 16,88 4 6 1,11E+07 12,81 3 4 8,30E+06gi|113945|sp|P10107.2|ANXA1_MOUSE Anxa1 38,7 5,49 2 2 1,41E+07 5,49 2 3 2,23E+07 5,49 2 2 7,89E+06
gi|24636295|sp|Q9D8B3.2|CHM4B_MOUSE Chmp4b 24,9 10,71 2 3 7,49E+06 10,71 2 3 6,10E+06 10,71 2 3 7,86E+06gi|60416372|sp|Q61495.2|DSG1A_MOUSE Dsg1a 114,5 1,99 2 2 9,72E+06 0 0 0,00E+00 1,51 2 2 7,79E+06gi|81917732|sp|Q9QYL7.1|ABT1_MOUSE Abt1 30,6 0 0 0,00E+00 0 0 0,00E+00 7,43 2 2 7,50E+06gi|353678156|sp|D3YXK2.2|SAFB1_MOUSE Safb 105,0 2,35 3 3 4,50E+06 2,35 3 4 1,34E+07 3,09 3 4 7,11E+06
26
gi|6094376|sp|P70279.1|SURF6_MOUSE Surf6 41,2 0 0 0,00E+00 0 0 0,00E+00 10,14 4 4 6,71E+06gi|122239|sp|P14483.1|HB2A_MOUSE H2-Ab1 30,1 0 0 0,00E+00 18,49 4 4 1,80E+07 9,43 2 2 6,67E+06
gi|90185245|sp|Q69ZN6.2|GNPTA_MOUSE Gnptab 140,9 0 0 0,00E+00 0 0 0,00E+00 1,38 2 3 6,64E+06gi|110825747|sp|P52875.2|TM165_MOUSE Tmem165 34,8 0 0 0,00E+00 0 0 0,00E+00 8,36 2 2 6,54E+06gi|50401658|sp|Q99N89.1|RM43_MOUSE Mrpl43 20,2 14,21 3 4 6,70E+06 0 0 0,00E+00 10,38 2 3 6,52E+06gi|341942168|sp|Q64012.3|RALY_MOUSE Raly 33,2 0 0 0,00E+00 0 0 0,00E+00 6,73 2 2 6,38E+06gi|416621|sp|P32261.1|ANT3_MOUSE Serpinc1 52,0 9,46 5 5 2,94E+07 4,30 2 2 1,51E+07 3,44 2 2 6,16E+06gi|2494835|sp|Q64692.1|SIA8D_MOUSE St8sia4 41,2 0 0 0,00E+00 0 0 0,00E+00 11,98 4 5 6,10E+06gi|1345668|sp|P47757.3|CAPZB_MOUSE Capzb 31,3 0 0 0,00E+00 43,32 14 43 4,28E+08 10,47 3 3 6,08E+06gi|71151920|sp|Q91VE6.1|MK67I_MOUSE Nifk 36,2 0 0 0,00E+00 0 0 0,00E+00 8,20 2 2 6,04E+06gi|52782750|sp|Q9CPQ8.1|ATP5L_MOUSE Atp5l 11,4 0 0 0,00E+00 0 0 0,00E+00 25,24 2 2 5,90E+06gi|12230747|sp|O88342.3|WDR1_MOUSE Wdr1 66,4 0 0 0,00E+00 0 0 0,00E+00 6,93 4 4 5,89E+06gi|54041950|sp|Q99N94.2|RM09_MOUSE Mrpl9 30,2 10,94 3 3 5,54E+06 0 0 0,00E+00 7,92 2 2 5,77E+06gi|3913652|sp|O09046.1|OXLA_MOUSE Il4i1 70,1 0 0 0,00E+00 0 0 0,00E+00 3,81 2 2 5,59E+06
gi|341941149|sp|Q8BTM8.5|FLNA_MOUSE Flna 281,0 1,40 3 3 2,60E+06 6,16 13 13 1,46E+07 2,46 5 5 5,10E+06gi|73620958|sp|Q62095.2|DDX3Y_MOUSE Ddx3y 73,4 0 0 0,00E+00 6,08 4 5 1,66E+07 4,86 3 3 5,02E+06gi|13633865|sp|Q9QUI0.1|RHOA_MOUSE Rhoa 21,8 0 0 0,00E+00 8,29 2 2 5,27E+06 22,28 5 8 4,88E+06gi|158705865|sp|A2AJI0.1|MA7D1_MOUSE Map7d1 93,2 9,34 7 9 8,78E+06 6,38 5 8 9,65E+06 2,36 2 3 4,84E+06gi|81880318|sp|Q99N93.1|RM16_MOUSE Mrpl16 28,8 0 0 0,00E+00 0 0 0,00E+00 10,76 2 2 4,84E+06gi|37999719|sp|Q8R0X7.1|SGPL1_MOUSE Sgpl1 63,6 0 0 0,00E+00 0 0 0,00E+00 5,81 3 3 4,82E+06gi|122134|sp|P14430.1|HA18_MOUSE H2-Q8 37,4 8,90 3 3 8,62E+06 0 0 0,00E+00 5,83 2 2 4,79E+06gi|18202285|sp|P58252.2|EF2_MOUSE Eef2 95,3 4,66 4 4 5,38E+06 2,10 2 2 1,05E+07 2,91 3 3 4,51E+06gi|548926|sp|P30882.2|CCL5_MOUSE Ccl5 10,1 0 0 0,00E+00 0 0 0,00E+00 20,88 2 2 4,50E+06
gi|119364598|sp|P54987.2|IRG1_MOUSE Irg1 53,7 3,28 2 2 2,64E+06 0 0 0,00E+00 7,38 4 4 4,48E+06gi|226693542|sp|Q9WTI7.2|MYO1C_MOUSE Myo1c 121,9 0 0 0,00E+00 20,13 17 21 6,89E+07 2,54 2 2 4,06E+06gi|3024089|sp|Q61792.1|LASP1_MOUSE Lasp1 30,0 5,32 2 2 1,72E+06 0 0 0,00E+00 10,27 3 3 3,65E+06gi|20455216|sp|Q9Z204.1|HNRPC_MOUSE Hnrnpc 34,4 0 0 0,00E+00 18,53 7 8 2,41E+07 7,03 2 2 3,61E+06gi|62510510|sp|Q9CRB9.1|CHCH3_MOUSE Chchd3 26,3 15,42 3 4 3,11E+06 8,81 2 3 4,13E+06 8,81 2 3 3,61E+06gi|46395721|sp|Q7TPR4.1|ACTN1_MOUSE Actn1 103,0 0 0 0,00E+00 40,13 32 52 1,22E+08 1,91 2 2 3,59E+06gi|62510642|sp|Q8CI11.2|GNL3_MOUSE Gnl3 60,7 0 0 0,00E+00 4,65 2 3 3,46E+06 7,62 4 4 3,58E+06
gi|85701216|sp|Q9CW46.2|RAVR1_MOUSE Raver1 79,3 0 0 0,00E+00 0 0 0,00E+00 2,67 2 2 3,50E+06gi|52783420|sp|Q9CQI7.1|RU2B_MOUSE Snrpb2 25,3 0 0 0,00E+00 0 0 0,00E+00 11,56 2 2 3,36E+06gi|81880411|sp|Q9CR59.1|G45IP_MOUSE Gadd45gip1 25,8 8,56 2 3 1,57E+07 0 0 0,00E+00 8,56 2 2 3,30E+06gi|25089776|sp|Q9DCX2.3|ATP5H_MOUSE Atp5h 18,7 0 0 0,00E+00 0 0 0,00E+00 13,04 2 2 3,24E+06
30
26
gi|71153826|sp|Q7TPV4.2|MBB1A_MOUSE Mybbp1a 151,9 0 0 0,00E+00 0 0 0,00E+00 1,93 2 2 3,14E+06gi|341942181|sp|P14685.3|PSMD3_MOUSE Psmd3 60,7 0 0 0,00E+00 0 0 0,00E+00 6,79 3 3 2,79E+06gi|83305135|sp|P52503.2|NDUS6_MOUSE Ndufs6 13,0 0 0 0,00E+00 22,41 2 2 5,70E+06 22,41 2 2 2,52E+06gi|1351218|sp|P47226.1|TES_MOUSE Tes 48,0 0 0 0,00E+00 0 0 0,00E+00 5,44 2 2 2,33E+06
gi|52783213|sp|Q9CY58.2|PAIRB_MOUSE Serbp1 44,7 0 0 0,00E+00 0 0 0,00E+00 6,63 2 2 2,29E+06gi|341940926|sp|P27046.2|MA2A1_MOUSE Man2a1 131,5 0 0 0,00E+00 0 0 0,00E+00 1,48 2 2 1,96E+06gi|135835|sp|P26350.2|PTMA_MOUSE Ptma 12,2 13,51 3 3 4,12E+06 13,51 3 4 9,37E+06 13,51 3 3 1,76E+06
gi|61211930|sp|Q9EP78.1|CHST7_MOUSE Chst7 54,7 0 0 0,00E+00 0 0 0,00E+00 2,48 2 2 1,71E+06gi|81896062|sp|Q8BGZ7.1|K2C75_MOUSE Krt75 59,7 15,97 12 26 8,80E+08 0 0 0,00E+00 0 0 0,00E+00gi|123797712|sp|Q8VED5.2|K2C79_MOUSE Krt79 57,5 9,42 9 30 7,32E+08 0 0 0,00E+00 0 0 0,00E+00
gi|549078|sp|P10639.3|THIO_MOUSE Txn1 11,7 20,95 2 3 4,47E+07 0 0 0,00E+00 0 0 0,00E+00gi|48428148|sp|Q9CPQ1.3|COX6C_MOUSE Cox6c 8,5 11,84 2 3 3,38E+07 21,05 3 4 5,09E+07 0 0 0,00E+00gi|146345481|sp|P09411.4|PGK1_MOUSE Pgk1 44,5 16,79 5 6 3,04E+07 16,79 5 5 3,56E+07 0 0 0,00E+00gi|341941244|sp|P49290.2|PERE_MOUSE Epx 81,3 9,36 7 10 2,60E+07 3,07 2 2 6,51E+06 0 0 0,00E+00gi|20141789|sp|P99029.2|PRDX5_MOUSE Prdx5 21,9 23,81 4 4 1,85E+07 15,71 3 4 1,51E+07 0 0 0,00E+00gi|119362|sp|P08113.2|ENPL_MOUSE Hsp90b1 92,4 5,24 4 4 1,66E+07 7,48 6 7 2,20E+07 0 0 0,00E+00
gi|32363397|sp|Q9CQC7.3|NDUB4_MOUSE Ndufb4 15,1 35,66 3 3 1,65E+07 0 0 0,00E+00 0 0 0,00E+00gi|21362904|sp|Q9CYH6.1|RRS1_MOUSE Rrs1 41,5 5,48 2 2 1,38E+07 0 0 0,00E+00 0 0 0,00E+00gi|730956|sp|P40142.1|TKT_MOUSE Tkt 67,6 9,47 6 6 1,31E+07 6,58 4 6 1,85E+07 0 0 0,00E+00
gi|21759077|sp|Q9CQI6.3|COTL1_MOUSE Cotl1 15,9 11,27 2 3 1,18E+07 11,27 2 3 2,25E+07 0 0 0,00E+00gi|341940630|sp|Q9WV32.4|ARC1B_MOUSE Arpc1b 41,0 8,33 2 3 1,17E+07 42,74 16 33 4,96E+08 0 0 0,00E+00
gi|117502|sp|P14211.1|CALR_MOUSE Calr 48,0 5,29 3 3 1,12E+07 6,49 3 3 2,07E+07 0 0 0,00E+00gi|32130423|sp|P51174.2|ACADL_MOUSE Acadl 47,9 5,58 2 2 1,05E+07 13,49 5 5 3,71E+07 0 0 0,00E+00gi|341941197|sp|O35375.2|NRP2_MOUSE Nrp2 104,6 2,79 2 4 9,26E+06 0 0 0,00E+00 0 0 0,00E+00gi|41017793|sp|Q9CQL5.1|RM18_MOUSE Mrpl18 20,7 11,67 2 3 7,88E+06 0 0 0,00E+00 0 0 0,00E+00gi|2500566|sp|P97425.1|ECP2_MOUSE Ear2 17,6 12,18 2 2 7,31E+06 0 0 0,00E+00 0 0 0,00E+00gi|353526354|sp|P17751.4|TPIS_MOUSE Tpi1 32,2 13,38 3 5 7,13E+06 16,72 4 6 1,47E+07 0 0 0,00E+00gi|114930|sp|P28667.2|MRP_MOUSE Marcksl1 20,2 14,00 2 2 7,11E+06 14,00 2 2 1,67E+07 0 0 0,00E+00
gi|1351942|sp|P48036.1|ANXA5_MOUSE Anxa5 35,7 12,54 5 5 6,99E+06 11,29 4 4 1,39E+07 0 0 0,00E+00gi|341941239|sp|P09103.2|PDIA1_MOUSE P4hb 57,0 6,29 3 4 6,65E+06 4,72 2 2 1,09E+07 0 0 0,00E+00gi|46396655|sp|Q9EQU5.1|SET_MOUSE Set 33,4 8,30 2 2 5,96E+06 0 0 0,00E+00 0 0 0,00E+00gi|341940830|sp|Q64282.2|IFIT1_MOUSE Ifit1 53,7 6,05 2 2 5,03E+06 0 0 0,00E+00 0 0 0,00E+00gi|68566057|sp|Q8VIJ6.1|SFPQ_MOUSE Sfpq 75,4 4,15 2 2 5,01E+06 0 0 0,00E+00 0 0 0,00E+00gi|81903502|sp|Q9CPY1.1|RM51_MOUSE Mrpl51 15,1 17,97 3 3 4,08E+06 0 0 0,00E+00 0 0 0,00E+00
26
gi|1708259|sp|P30681.3|HMGB2_MOUSE Hmgb2 24,1 13,81 2 2 3,96E+06 0 0 0,00E+00 0 0 0,00E+00gi|2851596|sp|Q93092.2|TALDO_MOUSE Taldo1 37,4 5,04 2 2 3,91E+06 5,04 2 2 3,15E+06 0 0 0,00E+00gi|2494703|sp|Q61599.3|GDIR2_MOUSE Arhgdib 22,8 10,50 2 2 3,80E+06 0 0 0,00E+00 0 0 0,00E+00gi|17368619|sp|Q99ME9.3|NOG1_MOUSE Gtpbp4 74,1 2,84 2 2 2,47E+06 0 0 0,00E+00 0 0 0,00E+00gi|71151983|sp|Q6URW6.1|MYH14_MOUSE Myh14 228,4 0 0 0,00E+00 4,95 12 53 7,23E+09 0 0 0,00E+00gi|47116573|sp|Q99JY9.3|ARP3_MOUSE Actr3 47,3 0 0 0,00E+00 45,69 17 43 5,70E+08 0 0 0,00E+00
gi|110825706|sp|Q9CVB6.3|ARPC2_MOUSE Arpc2 34,3 0 0 0,00E+00 64,00 20 57 5,70E+08 0 0 0,00E+00gi|34223730|sp|P47753.4|CAZA1_MOUSE Capza1 32,9 0 0 0,00E+00 65,03 14 24 5,01E+08 0 0 0,00E+00gi|59799524|sp|Q9JJ28.1|FLII_MOUSE Flii 144,7 0 0 0,00E+00 2,52 3 4 4,26E+08 0 0 0,00E+00
gi|62510460|sp|Q9CPW4.3|ARPC5_MOUSE Arpc5 16,3 0 0 0,00E+00 53,64 8 15 3,91E+08 0 0 0,00E+00gi|62899893|sp|Q9JM76.3|ARPC3_MOUSE Arpc3 20,5 0 0 0,00E+00 30,90 6 23 2,84E+08 0 0 0,00E+00gi|38372251|sp|Q99MU3.2|DSRAD_MOUSE Adar 130,4 0 0 0,00E+00 2,89 3 3 2,42E+08 0 0 0,00E+00gi|341940983|sp|Q99104.2|MYO5A_MOUSE Myo5a 215,4 0 0 0,00E+00 28,01 52 82 1,48E+08 0 0 0,00E+00gi|21903382|sp|P48962.4|ADT1_MOUSE Slc25a4 32,9 0 0 0,00E+00 16,78 5 12 1,35E+08 0 0 0,00E+00gi|13123946|sp|P57780.1|ACTN4_MOUSE Actn4 104,9 0 0 0,00E+00 34,54 27 37 1,06E+08 0 0 0,00E+00gi|81879596|sp|Q91WK0.1|LRRF2_MOUSE Lrrfip2 47,1 0 0 0,00E+00 22,17 9 13 6,11E+07 0 0 0,00E+00gi|62899673|sp|O70200.1|AIF1_MOUSE Aif1 16,9 0 0 0,00E+00 26,53 4 5 5,93E+07 0 0 0,00E+00
gi|24638218|sp|Q91VW3.1|SH3L3_MOUSE Sh3bgrl3 10,5 0 0 0,00E+00 26,88 2 3 5,32E+07 0 0 0,00E+00gi|24211558|sp|Q9QYB1.3|CLIC4_MOUSE Clic4 28,7 0 0 0,00E+00 5,53 2 3 3,55E+07 0 0 0,00E+00gi|3913581|sp|Q61878.1|PRG2_MOUSE Prg2 24,2 0 0 0,00E+00 13,45 3 5 3,17E+07 0 0 0,00E+00gi|341940610|sp|Q60604.3|ADSV_MOUSE Scin 80,2 0 0 0,00E+00 14,13 9 10 3,09E+07 0 0 0,00E+00gi|146345480|sp|P27773.2|PDIA3_MOUSE Pdia3 56,6 0 0 0,00E+00 19,01 7 7 2,86E+07 0 0 0,00E+00gi|81882407|sp|Q569Z6.1|TR150_MOUSE Thrap3 108,1 0 0 0,00E+00 5,89 5 8 2,23E+07 0 0 0,00E+00gi|17380333|sp|Q9D855.3|QCR7_MOUSE Uqcrb 13,5 0 0 0,00E+00 21,62 2 5 1,75E+07 0 0 0,00E+00gi|13629369|sp|Q9WUA3.1|K6PP_MOUSE Pfkp 85,4 0 0 0,00E+00 5,23 4 4 1,69E+07 0 0 0,00E+00gi|146345421|sp|Q61553.4|FSCN1_MOUSE Fscn1 54,5 0 0 0,00E+00 4,26 2 3 1,63E+07 0 0 0,00E+00gi|2494022|sp|Q62426.1|CYTB_MOUSE Cstb 11,0 0 0 0,00E+00 32,65 2 2 1,58E+07 0 0 0,00E+00gi|81881592|sp|Q9D898.1|ARP5L_MOUSE Arpc5l 17,0 0 0 0,00E+00 46,41 4 4 1,42E+07 0 0 0,00E+00gi|116487|sp|P08207.2|S10AA_MOUSE S100a10 11,2 0 0 0,00E+00 27,84 2 5 1,37E+07 0 0 0,00E+00gi|126752|sp|P26645.2|MARCS_MOUSE Marcks 29,6 0 0 0,00E+00 11,65 2 3 1,23E+07 0 0 0,00E+00gi|46577116|sp|Q9D1G1.1|RAB1B_MOUSE Rab1b 22,2 0 0 0,00E+00 19,40 4 4 1,15E+07 0 0 0,00E+00gi|81902478|sp|Q91Z49.1|UIF_MOUSE Fyttd1 35,9 0 0 0,00E+00 6,94 2 2 9,93E+06 0 0 0,00E+00
gi|341940791|sp|P49710.2|HCLS1_MOUSE Hcls1 54,2 0 0 0,00E+00 4,53 2 2 6,35E+06 0 0 0,00E+00gi|12230589|sp|Q9JJU8.1|SH3L1_MOUSE Sh3bgrl 12,8 0 0 0,00E+00 20,18 2 2 6,12E+06 0 0 0,00E+00
26
gi|55976201|sp|Q9D0E1.3|HNRPM_MOUSE Hnrnpm 77,6 0 0 0,00E+00 3,29 2 2 4,91E+06 0 0 0,00E+00gi|347595715|sp|Q52KI8.2|SRRM1_MOUSE Srrm1 106,8 0 0 0,00E+00 2,22 2 2 4,34E+06 0 0 0,00E+00gi|54041071|sp|Q9WUM4.2|COR1C_MOUSE Coro1c 53,1 0 0 0,00E+00 7,81 3 3 3,84E+06 0 0 0,00E+00gi|21263374|sp|Q91V92.1|ACLY_MOUSE Acly 119,7 0 0 0,00E+00 1,56 2 2 2,66E+06 0 0 0,00E+00gi|110816418|sp|Q9D8T7.2|SLIRP_MOUSE Slirp 12,6 0 0 0,00E+00 16,96 2 2 1,67E+06 0 0 0,00E+00

REFERENCES AND NOTES 1. U. Rutishauser, Polysialic acid in the plasticity of the developing and adult
vertebrate nervous system. Nat. Rev. Neurosci. 9, 26–35 (2008). Medline doi:10.1038/nrn2285
2. M. Mühlenhoff, M. Rollenhagen, S. Werneburg, R. Gerardy-Schahn, H. Hildebrandt, Polysialic acid: Versatile modification of NCAM, SynCAM 1 and neuropilin-2. Neurochem. Res. 38, 1134–1143 (2013). Medline doi:10.1007/s11064-013-0979-2
3. R. L. Schnaar, R. Gerardy-Schahn, H. Hildebrandt, Sialic acids in the brain: Gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol. Rev. 94, 461–518 (2014). Medline doi:10.1152/physrev.00033.2013
4. R. A. Falconer, R. J. Errington, S. D. Shnyder, P. J. Smith, L. H. Patterson, Polysialyltransferase: A new target in metastatic cancer. Curr. Cancer Drug Targets 12, 925–939 (2012). Medline doi:10.2174/156800912803251225
5. A. Harduin-Lepers, M. A. Krzewinski-Recchi, F. Colomb, F. Foulquier, S. Groux-Degroote, P. Delannoy, Sialyltransferases functions in cancers. Front. Biosci. (Elite Ed.) 4, 499–515 (2012). Medline doi:10.2741/E396
6. Y. M. J. Al-Saraireh, M. Sutherland, B. R. Springett, F. Freiberger, G. Ribeiro Morais, P. M. Loadman, R. J. Errington, P. J. Smith, M. Fukuda, R. Gerardy-Schahn, L. H. Patterson, S. D. Shnyder, R. A. Falconer, Pharmacological inhibition of polysialyltransferase ST8SiaII modulates tumour cell migration. PLOS ONE 8, e73366 (2013). Medline doi:10.1371/journal.pone.0073366
7. S. Curreli, Z. Arany, R. Gerardy-Schahn, D. Mann, N. M. Stamatos, Polysialylated neuropilin-2 is expressed on the surface of human dendritic cells and modulates dendritic cell-T lymphocyte interactions. J. Biol. Chem. 282, 30346–30356 (2007). Medline doi:10.1074/jbc.M702965200
8. A. Rey-Gallardo, C. Escribano, C. Delgado-Martín, J. L. Rodriguez-Fernández, R. Gerardy-Schahn, U. Rutishauser, A. L. Corbi, M. A. Vega, Polysialylated neuropilin-2 enhances human dendritic cell migration through the basic C-terminal region of CCL21. Glycobiology 20, 1139–1146 (2010). Medline doi:10.1093/glycob/cwq078
9. M. Bax, S. J. van Vliet, M. Litjens, J. J. García-Vallejo, Y. van Kooyk, Interaction of polysialic acid with CCL21 regulates the migratory capacity of human dendritic cells. PLOS ONE 4, e6987 (2009). Medline doi:10.1371/journal.pone.0006987
10. P. M. Drake, J. K. Nathan, C. M. Stock, P. V. Chang, M. O. Muench, D. Nakata, J. R. Reader, P. Gip, K. P. Golden, B. Weinhold, R. Gerardy-Schahn, F. A. Troy 2nd, C. R. Bertozzi, Polysialic acid, a glycan with highly restricted expression, is found on human and murine leukocytes and modulates immune responses. J. Immunol. 181, 6850–6858 (2008). Medline doi:10.4049/jimmunol.181.10.6850
11. M. Eckhardt, O. Bukalo, G. Chazal, L. Wang, C. Goridis, M. Schachner, R. Gerardy-Schahn, H. Cremer, A. Dityatev, Mice deficient in the polysialyltransferase ST8SiaIV/PST-1 allow discrimination of the roles of

neural cell adhesion molecule protein and polysialic acid in neural development and synaptic plasticity. J. Neurosci. 20, 5234–5244 (2000). Medline
12. C. Moussion, J.-P. Girard, Dendritic cells control lymphocyte entry to lymph nodes through high endothelial venules. Nature 479, 542–546 (2011). Medline doi:10.1038/nature10540
13. M. Wendland, S. Willenzon, J. Kocks, A. C. Davalos-Misslitz, S. I. Hammerschmidt, K. Schumann, E. Kremmer, M. Sixt, A. Hoffmeyer, O. Pabst, R. Förster, Lymph node T cell homeostasis relies on steady state homing of dendritic cells. Immunity 35, 945–957 (2011). Medline doi:10.1016/j.immuni.2011.10.017
14. S. E. Macatonia, S. C. Knight, A. J. Edwards, S. Griffiths, P. Fryer, Localization of antigen on lymph node dendritic cells after exposure to the contact sensitizer fluorescein isothiocyanate. Functional and morphological studies. J. Exp. Med. 166, 1654–1667 (1987). Medline doi:10.1084/jem.166.6.1654
15. R. Förster, A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Müller, E. Wolf, M. Lipp, CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell 99, 23–33 (1999). Medline doi:10.1016/S0092-8674(00)80059-8
16. A. Braun, T. Worbs, G. L. Moschovakis, S. Halle, K. Hoffmann, J. Bölter, A. Münk, R. Förster, Afferent lymph-derived T cells and DCs use different chemokine receptor CCR7-dependent routes for entry into the lymph node and intranodal migration. Nat. Immunol. 12, 879–887 (2011). Medline doi:10.1038/ni.2085
17. M. Weber, R. Hauschild, J. Schwarz, C. Moussion, I. de Vries, D. F. Legler, S. A. Luther, T. Bollenbach, M. Sixt, Interstitial dendritic cell guidance by haptotactic chemokine gradients. Science 339, 328–332 (2013). Medline doi:10.1126/science.1228456
18. A. Link, T. K. Vogt, S. Favre, M. R. Britschgi, H. Acha-Orbea, B. Hinz, J. G. Cyster, S. A. Luther, Fibroblastic reticular cells in lymph nodes regulate the homeostasis of naive T cells. Nat. Immunol. 8, 1255–1265 (2007). Medline doi:10.1038/ni1513
19. C. T. Veldkamp, E. Kiermaier, S. J. Gabel-Eissens, M. L. Gillitzer, D. R. Lippner, F. A. DiSilvio, C. J. Mueller, P. L. Wantuch, G. R. Chaffee, M. W. Famiglietti, D. M. Zgoba, A. A. Bailey, Y. Bah, S. J. Engebretson, D. R. Graupner, E. R. Lackner, V. D. LaRosa, T. Medeiros, M. L. Olson, A. J. Phillips, H. Pyles, A. M. Richard, S. J. Schoeller, B. Touzeau, L. G. Williams, M. Sixt, F. C. Peterson, Solution structure of CCL19 and identification of overlapping CCR7 and PSGL-1 binding sites. Biochemistry 54, 4163–4166 (2015). Medline
20. M. Love, J. L. Sandberg, J. J. Ziarek, K. P. Gerarden, R. R. Rode, D. R. Jensen, D. R. McCaslin, F. C. Peterson, C. T. Veldkamp, Solution structure of CCL21 and identification of a putative CCR7 binding site. Biochemistry 51, 733–735 (2012). Medline doi:10.1021/bi201601k

21. J. Hirose, H. Kawashima, O. Yoshie, K. Tashiro, M. Miyasaka, Versican interacts with chemokines and modulates cellular responses. J. Biol. Chem. 276, 5228–5234 (2001). Medline doi:10.1074/jbc.M007542200
22. A. Rey-Gallardo, C. Delgado-Martín, R. Gerardy-Schahn, J. L. Rodríguez-Fernández, M. A. Vega, Polysialic acid is required for neuropilin-2a/b-mediated control of CCL21-driven chemotaxis of mature dendritic cells and for their migration in vivo. Glycobiology 21, 655–662 (2011). Medline doi:10.1093/glycob/cwq216
23. M. Rollenhagen, F. F. Buettner, M. Reismann, A. C. Jirmo, M. Grove, G. M. Behrens, R. Gerardy-Schahn, F. G. Hanisch, M. Mühlenhoff, Polysialic acid on neuropilin-2 is exclusively synthesized by the polysialyltransferase ST8SiaIV and attached to mucin-type o-glycans located between the b2 and c domain. J. Biol. Chem. 288, 22880–22892 (2013). Medline doi:10.1074/jbc.M113.463927
24. M. Frosch, I. Görgen, G. J. Boulnois, K. N. Timmis, D. Bitter-Suermann, NZB mouse system for production of monoclonal antibodies to weak bacterial antigens: Isolation of an IgG antibody to the polysaccharide capsules of Escherichia coli K1 and group B meningococci. Proc. Natl. Acad. Sci. U.S.A. 82, 1194–1198 (1985). Medline doi:10.1073/pnas.82.4.1194
25. A. Jokilammi, P. Ollikka, M. Korja, E. Jakobsson, V. Loimaranta, S. Haataja, H. Hirvonen, J. Finne, Construction of antibody mimics from a noncatalytic enzyme-detection of polysialic acid. J. Immunol. Methods 295, 149–160 (2004). Medline doi:10.1016/j.jim.2004.10.006
26. D. Schwarzer, K. Stummeyer, T. Haselhorst, F. Freiberger, B. Rode, M. Grove, T. Scheper, M. von Itzstein, M. Mühlenhoff, R. Gerardy-Schahn, Proteolytic release of the intramolecular chaperone domain confers processivity to endosialidase F. J. Biol. Chem. 284, 9465–9474 (2009). Medline doi:10.1074/jbc.M808475200
27. Q. Lu, M. C. Burns, P. J. McDevitt, T. L. Graham, A. J. Sukman, J. A. Fornwald, X. Tang, K. T. Gallagher, G. E. Hunsberger, J. J. Foley, D. B. Schmidt, J. J. Kerrigan, T. S. Lewis, R. S. Ames, K. O. Johanson, Optimized procedures for producing biologically active chemokines. Protein Expr. Purif. 65, 251–260 (2009). Medline doi:10.1016/j.pep.2009.01.017
28. M. B. Lutz, N. Kukutsch, A. L. J. Ogilvie, S. Rößner, F. Koch, N. Romani, G. Schuler, An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 223, 77–92 (1999). Medline doi:10.1016/S0022-1759(98)00204-X
29. T. Lämmermann, B. L. Bader, S. J. Monkley, T. Worbs, R. Wedlich-Söldner, K. Hirsch, M. Keller, R. Förster, D. R. Critchley, R. Fässler, M. Sixt, Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453, 51–55 (2008). Medline doi:10.1038/nature06887
30. M. Windfuhr, A. Manegold, M. Muhlenhoff, M. Eckhardt, R. Gerardy-Schahn, Molecular defects that cause loss of polysialic acid in the complementation group 2A10. J. Biol. Chem. 275, 32861–32870 (2000). Medline doi:10.1074/jbc.M003507200

31. J. H. Y. Tan, J. P. Ludeman, J. Wedderburn, M. Canals, P. Hall, S. J. Butler, D. Taleski, A. Christopoulos, M. J. Hickey, R. J. Payne, M. J. Stone, Tyrosine sulfation of chemokine receptor CCR2 enhances interactions with both monomeric and dimeric forms of the chemokine monocyte chemoattractant protein-1 (MCP-1). J. Biol. Chem. 288, 10024–10034 (2013). Medline doi:10.1074/jbc.M112.447359
32. T. G. Keys, H. L. Fuchs, J. Ehrit, J. Alves, F. Freiberger, R. Gerardy-Schahn, Engineering the product profile of a polysialyltransferase. Nat. Chem. Biol. 10, 437–442 (2014). Medline doi:10.1038/nchembio.1501
33. L. Riol-Blanco, N. Sánchez-Sánchez, A. Torres, A. Tejedor, S. Narumiya, A. L. Corbí, P. Sánchez-Mateos, J. L. Rodríguez-Fernández, The chemokine receptor CCR7 activates in dendritic cells two signaling modules that independently regulate chemotaxis and migratory speed. J. Immunol. 174, 4070–4080 (2005). Medline doi:10.4049/jimmunol.174.7.4070
34. A. Walz, I. Rodriguez, P. Mombaerts, Aberrant sensory innervation of the olfactory bulb in neuropilin-2 mutant mice. J. Neurosci. 22, 4025–4035 (2002). Medline
35. K. Schaeuble, M. A. Hauser, A. V. Rippl, R. Bruderer, C. Otero, M. Groettrup, D. F. Legler, Ubiquitylation of the chemokine receptor CCR7 enables efficient receptor recycling and cell migration. J. Cell Sci. 125, 4463–4474 (2012). Medline doi:10.1242/jcs.097519


















