
Supplementary Materials for -...
52
www.sciencemag.org/cgi/content/full/science.aai8898/DC1 Supplementary Materials for Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance Yiwen Deng, Keran Zhai, Zhen Xie, Dongyong Yang, Xudong Zhu, Junzhong Liu, Xin Wang, Peng Qin, Yuanzhu Yang, Guomin Zhang, Qun Li, Jianfu Zhang, Shuangqing Wu, Joëlle Milazzo, Bizeng Mao, Ertao Wang, Huaan Xie, Didier Tharreau, Zuhua He* *Corresponding author. Email: [email protected] Published 2 February 2017 on Science First Release DOI: 10.1126/science.aai8898 This PDF file includes: Materials and Methods Figs. S1 to S17 Tables S1 to S7 References
Transcript of Supplementary Materials for -...

www.sciencemag.org/cgi/content/full/science.aai8898/DC1
Supplementary Materials for
Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance
Yiwen Deng, Keran Zhai, Zhen Xie, Dongyong Yang, Xudong Zhu, Junzhong Liu, Xin Wang, Peng Qin, Yuanzhu Yang, Guomin Zhang, Qun Li, Jianfu Zhang, Shuangqing
Wu, Joëlle Milazzo, Bizeng Mao, Ertao Wang, Huaan Xie, Didier Tharreau, Zuhua He*
*Corresponding author. Email: [email protected]
Published 2 February 2017 on Science First Release DOI: 10.1126/science.aai8898
This PDF file includes:
Materials and Methods Figs. S1 to S17 Tables S1 to S7 References

2
Materials and Methods Plant materials and pathogen inoculation
The rice parents, transgenic materials and NILs used in this study were listed in table
S6. For blast resistance evaluation in seedling rice, two-week-old rice seedlings were
spray-inoculated with blast spore suspensions (1 ×105 spores/ml) in a dew growth
chamber for 24 h in darkness at 26°C, and were subsequently kept at 12 h/12 h
(day/night), 26°C and 90% relative humidity for 7 days. Lesion types on leaves were
observed and scored from 0 (resistant) to 5 (susceptible) according to the standard scale
as described (27). Percentages of lesion areas (disease index) were scored using image
analysis with software ImageJ (http://rsbweb.nih.gov/ij/). Relative infection ratio of blast
fungus in rice cells was determined by DNA-based quantitative PCR as described
previously (28). In brief, the rice ubiquitin gene (LOC_Os03g13170) was used to
quantify the rice DNA, the Pot2 transposon sequence used to quantify the fungus, the
infection ratio (N:MgPot2/[N:OsUbq × 100]) was then calculated from the determined
number of target sequences of MgPot2 and OsUbq for each sample. Each inoculation was
assayed with 20 DNA samples.
For bacterial blight resistance assay, rice plants were inoculated with Xanthomonas
oryzae pv oryzae strain PXO99A using the leaf clip method as described (29). Lesion lengths
and bacterial growth were recorded at 14 dpi. For sheath blight resistance assay, plants were
inoculated with Rhizoctonia solani AG1-IA (isolate RH-9) with the toothpick method. In
brief, R. solani colony plugs were transferred to new PDA plates and co-incubated with short
(0.8-1.0 cm) woody toothpicks for 5 d at 28°C. Toothpicks with mycelia were then inserted
into the third leaf sheath of rice plants grown in the paddy field at booting stage. Sheath
blight symptom was recorded at 15 dpi with the 6-score system (30).
Field trial, yield evaluation and statistical analysis
Field resistance to blast was assessed in the Enshi blast nursery (Hubei province, mid-
western China) and the Daweishan blast nursery (Hunan province, southeastern China)
field in the consecutive 3 years (2012-2014), which have been long used to select blast
resistant varieties with high selection pressure of blast disease. The Pigm donor variety
GM4, NIL-Pigm lines and the recurrent parents (9311 and Maratelli) were tested in the

3
blast nurseries. Field test was also conducted for transgenic rice in our Linshui protected
blast nursery (Hainan Island, southern China). In addition, fourteen NILs containing
individual blast resistance gene Pi2, Pi9, Pi11, Pia, Pib, Pik, Pii, Pikp, Piks, Pit, Pita,
Pita2, Piz and Pizt in the genetic background of a high susceptible cultivar
Lijiangxintuanheigu, respectively (31), were also grown for resistance spectrum
comparison. Plants were grown in the nurseries with 3 replications with more than 300
plants each replicate. To ensure uniform infection of blast, a row of susceptible variety
was planted every five rows as well as on the borders. Resistance was evaluated
according to the standard evaluation system of IRRI for leaf lesion types with scores
ranging from 0 to 9 (32). Disease severity of leaf blast was scored by the percentage of
lesion area. Neck (panicle) blast severity was scored as a percentage of panicle infection
as described (33). Yield performance was evaluated in field tests of NIL-Pigm (9311 and
Maratelli) at the Daiweishan blast nursery and the neighbor experimental station (2014),
and the Enshi blast nursery under disease conditions during 2012 and 2014. NIL-Pigm
and parent lines were planted in a completely Randomized Block Design with 6
replicates in a rice paddy with an interplant spacing of 20 × 20 cm. Statistical analysis
was performed by Student’s t-test or Tukey-Kramer test for multiple comparisons.
Construction of rice BAC library and BAC screening and sequencing
The genomic DNA was isolated from the etiolated leaves of GM4 and partially
digested with HindIII and inserted into the BAC cloning vector plndigoBAC-5 to
construct a BAC library with 40,500 clones with an average insert size of 110 kb.
Construction and screening of the BAC library were performed as described (34). The
BACs covering the Pigm locus were isolated using PCR-based markers M35572 and
M80410 that co-segregate with Pigm (table S7). Two positive and overlapping clones
BAC7 and BAC30 were completely sequenced by shot-gun technique using an ABI 3730
sequencer (Applied Biosystems). The genomic sequence of the Pigm locus was annotated
by using the gene prediction program Fgenesh (www.softberry.com) and was manually
edited by a homology search against available databases on GenBank
(www.ncbi.nlm.nih.gov/genbank).

4
Plasmid construction and rice transformation
To construct the plasmids for complementation test, the BAC30 clone containing the
entire Pigm locus was partially digested with Sau3AI, the digested fragments larger than
15-kb in size were purified and ligated into the binary vector pCAMBIA1300 digested by
BamHI. The subclones were verified by PCR and sequencing using specific primers
(table S7). To generate overexpression clones, the entire coding regions (CDS) of PigmR,
PigmS and R4 were amplified using gene-specific primers from leaves or panicles of
GM4, and the PCR products were inserted into the binary vector pCAMBIA1300-
CaMV35S to generate overexpression plasmids 35S::PigmR, 35S::PigmS and 35S::R4.
The CDS of PigmS also was cloned into the binary vector pCAMBIA1300-PigmR with
the PigmR promoter to conduct the chimeric NLR gene PigmR::PigmS. All the constructs
and the empty vector were introduced into susceptible variety NIPB via Agrobacterium-
mediated transformation to generate more than 30 independent lines for each construct,
which were selected by PCR-based gene expression assays. In addition, 35S::PigmS was
also transformed into the resistant NIL-Pigm plants. To construct the clones of PigmS
driven by entire and different truncated promoters, the full CDS of PigmS was first
inserted into the binary vector pCAMBIA1300 to generate plasmid p1300-PigmS. The
different truncated promoters with removal of either MITE1, MITE2 or non-MITE region
were amplified from the PigmS promoter using specific primers and individually inserted
into pCAMBIA1300. All the constructs and empty vector were transformed into NIL-
Pigm to generate more than 30 independent lines for each construct, which were selected
by PCR-based gene expression assays. To generate double transgene plants of
PigmR/PigmS or PigmR/35S::PigmS or PigmR/PigmR::PigmS, transgenic plants PigmR
were crossed with transgenic plants PigmS, 35S::PigmS and PigmR::PigmS respectively,
stable homozygous progenies were selected by PCR-based expression assays.
To generate the PigmR RNAi constructs, the 200-bp 5’ UTR fragment of PigmR was
amplified by RT-PCR using specific primers and inserted as inverted repeats into the
conventional RNAi vector PTCK303 to generate hairpin RNAi constructs. The generate
double RNAi of PigmR and PigmS (PigmRS-RNAi), a 500-bp cDNA fragment of the
PigmR LRR region that has high similarity with the PigmS LRR domain was amplified
by RT-PCR and inserted as inverted repeats into the conventional RNAi vector PTCK303

5
to generate a hairpin RNAi construct. The 300-500bp cDNA fragments of the key genes
involved in the rice RdDM pathway, OsRDR2 (LOC_Os04g39160), OsAGO4a
(LOC_Os01g16870) (16) and OsDCL3a (LOC_Os01g68120) (17), were amplified using
gene-specific primers from the cDNAs of GM4 and individually inserted as inverted
repeats into PTCK303 to generate hairpin RNAi construct respectively. All the resulting
RNAi vectors and the empty vector were introduced into NIL-Pigm via Agrobacterium-
mediated transformation to generate more than 50 independent RNAi lines for each
construct, selected by PCR-based gene expression assays. The OsAGO4a RNAi construct
and the empty vector also were introduced into the susceptible variety NIPB. All primers
used for clone constructions are listed in table S7.
For the β-glucuronidase (GUS) reporter gene constructs, DNA fragments containing
the entire PigmS promoter (-1600 to -1) isolated from BAC30 and the different truncated
PigmS fragments as described above were inserted upstream of the GUS coding sequence
in the binary pCAMBIA1300-GUS vector to make fusion reporters (table S7). All the
resulting constructs were sequenced and introduced into the japonica variety NIPB via
Agrobacterium-mediated transformation to generate more than 20 independent lines that
showed similar expression patterns of the fusion reporters.
GUS histochemical staining
GUS staining was performed following the standard procedure (35). The different
issues including leaves, internodes, roots and young panicles were collected from at least
five independent lines for each construct, submerged in the staining solution and
incubated at 37°C up to 48 h or until blue color became visible. Then the samples were
incubated with 95% ethanol for 24 h to remove the chlorophylls. The GUS-staining
panicles and pollens were taken photos using a microscope.
RNA extraction and quantitative RT-PCR (qPCR) analysis
Total RNAs were extracted from different rice tissues using TRIzol reagent and treated
with RNase-free DNase I according to the manufacturer’s protocol (Invitrogen). The
resulting RNA was reverse-transcribed using the Superscript III RT kit (Invitrogen). For
defense and abiotic stress responses, expression of the defense–related genes, OsPR1

6
(LOC_Os07g03710), OsPR3 (LOC_Os04g41620), OsPR5 (LOC_Os12g43430), OsPBZ1
(LOC_Os12g36880), and the abiotic stress responsive genes, OsHSP70-1
(LOC_Os11g08470), OsHSP70-2 (LOC_Os11g08440) (25), OsDEF48
(LOC_Os11g47120), OsDEF70 (LOC_Os10g20550), OsDEF56 (LOC_Os08g15550)
(26), were analyzed. qRT-PCR was performed using SYBR Green (Takara) with the
Eppendorf AG 22331 cycler following the manufacturer's instructions, each qRT-PCR
assay was replicated at least three times with three independent RNA preparations, and
the rice OsActin1 gene (LOC_Os03g50885) was used as an internal control. The primers
for qRT-PCR are listed in table S7.
Yeast two-hybrid assays
The different domains of PigmR and PigmS including CC, NB-ARC, LRR and full-
length cDNAs, were amplified and inserted into the vectors pDEST22 and pDEST32
(Invitrogen) by the Gateway cloning technology, respectively, using the gene-specific
primers (table S7). The resulting constructs were transformed into yeast strain AH109.
Co-transformants were plated on synthetic medium without uracil, tryptophan, leucine
and histidine, supplemented with 3mM 3-AT and incubated at 28°C for 3 days.
Experimental procedures for screening and plasmid isolation were performed according
to the manufacturer’s user guide (Invitrogen).
Split luciferase complementation assay
For split luciferase complementation assay constructs, the full-length CDS of PigmR
and PigmS were amplified and inserted into the vector pCAMBIA-35S-NLuc and
pCAMBIA-35S-CLuc to generate the constructs CLuc-PigmR, CLuc-PigmS, PigmR-
NLuc, and PigmS-NLuc. Transient expression in Nicotiana benthamiana leaves was
performed by Agrobacterium-infiltration (36). In brief, Agrobacterium strains GV3101
containing the indicated constructs were cultured overnight in LB media. Equal amounts
of Agrobacterium cultures for CLuc and NLuc constructs were mixed to a final
concentration OD600 = 1.0 and collected and resuspended in infiltration buffer (10mM
MES, pH5.6, 10 mM MgCl2 and 150 μM acetosyringone). The mixed suspensions were
incubated at room temperature for ~3 h and then infiltrated into fully expanded young

7
leaves of N. benthamiana plants grown at 25 °C. The infiltrated plants were immediately
covered with plastic bags and placed at 23 °C for 48 h. Plants were then incubated at
28 °C with 16 h light/d for LUC activity measurement, with negative controls.
Coimmunoprecipitation (Co-IP) assays
Co-IP assays were performed according to a published procedure (37) with minor
modifications. Briefly, the CC domains of PigmR and PigmS were amplified and inserted
into the vector pCAMBIA1305-35S-FLAG and pCAMBIA1305-35S-HA to generate the
tagged constructs PigmR-CC-FLAG, PigmS-CC-FLAG, PigmR-CC-HA, and PigmS-CC-
HA. The plasmid GUS-HA was used as a control. All the plasmids were individually
transformed into Agrobacterium strain GV3101. The clones carrying different constructs
were co-infiltrated into N. benthamiana leaves. About 1g infiltrated leaf tissues were
ground into powder in liquid nitrogen and homogenized in 800 μL of extraction buffer
(50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5% Triton X-100, 5% glycerol, 2 mM EDTA,
and 1% protease inhibitor cocktail). Immunoblotting and Co-IP experiments were
performed following the Pierce HA Tag IP/Co-IP Kit instruction (Thermo).
Protein site mutagenesis and cell death assay in N. benthamiana
The conserved amino acids in the PigmS CC domain were site-mutated by PCR-based
approach as previously reported (9), and the mutated PigmS proteins were fused with
YFP (PigmS:YFP) or Myc (PigmR:Myc) to determine their expression in the N.
benthamiana. PigmR-triggered cell death in N. benthamiana was investigated as
described (38). In brief, suspensions (OD600 = 0.5) of Agrobacterium GV3101
containing the PigmR:Myc, PigmS:YFP, and mutant PigmS:YFP fusion expression
constructs respectively were infiltrated into fully expanded young leaves of 4-week-old N.
benthamiana plants grown at 25 °C. YFP was expressed as an internal control. The
infiltrated plants were immediately covered with black bags and placed at 23 °C. The
hypersensitive response cell death on infiltrated leaves was recorded and photographed at
48 hpi. Proteins were extracted from the infiltrated N. benthamiana leaves at 40 hpi
(hours post infiltration) and purified by anti-Myc or anti-GFP beads, the
immunoprecipitations were applied to analyze accumulation of the PigmR:Myc,

8
PigmS:YFP and the mutated PigmS:YFP fusion proteins with anti-Myc or anti-GFP
antibody.
Protein competition experiment
For competition of PigmS with PigmR in dimerization, the fusion protein constructs,
PigmR-CC-GST, PigmR-CC-His, and PigmS-CC-MBP, were generated and expressed in
E. coli (DE3) and purified by antibody-conjugated beads, and protein reaction samples
were prepared (0.75 µg/µl), respectively. Competition assay was performed as described
(39). In brief, PigmR-CC-GST and PigmR-CC-His (20 µl) were co-incubated in the anti-
GST beads. Then different amount of PigmS-CC-MBP (0, 5, 15 and 20 µl) was added in
the same mix to compete the homodimerization of PigmR-CC-GST and PigmR-CC-His.
Interacting proteins were pulled down using anti-GST beads and immunoblotted with
different antibodies to determine protein levels. .
Bisulfite sequencing
Genomic DNAs were prepared from leaves and pollens of NIL-Pigm using CTAB
method and DNeasy plant kit according to the manufacturer’s instructions (Qiagene).
Genomic DNA (2 μg) was digested using HindIII, followed by phenol/chloroform
extraction and ethanol precipitation. DNA was treated using the Epitect bisulfite kit
according to the manufacturer’s instructions (Qiagen). The bisulphited DNA was used for
PCR amplification of the PigmS promoter. PCR primers were designed with Methyl
Primer Express software (Applied Biosystems). Amplified PCR fragments were cloned
into pGEM-T easy vector (Promega) and sequenced. Sequencing data was analyzed using
Web-based Kismeth software (40). Primers used for bisulfite sequencing are listed in
table S7.
Small RNA sequencing
Small RNAs were isolated from total RNAs from the leaves of two-week-old NIL-Pigm
and NIPB seedlings. Small RNA libraries were constructed and deep-sequenced with an
Illumina Hiseq 2000 according to the manufacturer’s instructions (Illumina) by BGI.
Small RNAs (21 and 24-nt siRNAs) were mapped to the PigmS promoter sequence using

9
software Bowtie-0.12.7 (41)
.
Small RNA gel blot assay
Small RNA gel blot experiments were performed as previously described (42). Total
RNAs prepared from leaves of NIL-Pigm and the transgenic lines OsAGO4aRNAi,
OsRDR2RNAi and OsDCL3aRNAi were loaded to detect the siRNAs derived from the
MITE1 and MITE2 regions of the PigmS promoter. The probe amplified with specific
primers (table. S7) was labeled with digoxigenin (DIG) using the PCR DIG Probe
Synthesis Kit (Roche). U6 was served as an internal control.
Molecular breeding of Pigm
The cultivar GM4 (Pigm) was crossed with varieties 9311(indica), Maratelli (japonica)
and Nipponbare (japonica), and backcrossed four times with the recurrent parents to
generate the BC4F1 plants, which self-crossed three times to generate the BC4F4 lines.
Total of 192 SSR markers covering all 12 rice chromosomes were used to detect the
backgrounds of the selected NILs. The NIL-Pigm lines carrying a single ~500-kb
fragment with the Pigm locus from the donor GM4 were selected in the 9311, Maratelli
and Nipponbare genomes. The NIL-Pigm lines (9311 and Maratelli) were cultivated at
the two different blast nurseries located in Enshi (Hubei province) and Daweishan
(Hunan province) for field evaluation (see above).
Database deposition
The full genomic sequence of the Pigm locus including PigmR and PigmS can be found
in GenBank (accession number KU904633). . The entire small RNA sequencing data can
be obtained from the NCBI Gene Expression Omnibus (GEO) database under series
accession number GSE83522.

10
Fig. S1. Pigm confers durable blast resistance and does not confer resistance to other
rice diseases.
(A) Field resistance phenotype of EnD3 (Pigm) exhibiting high resistance to M. oryzae in
the Enshi blast nursery, compared with the susceptible variety V14B. (B) Field resistance
score records from 1984 to 2015 indicate that EnD3 confers durable resistance to M.
oryzae in the blast nursery for more than 30 years. (C, D) PigmR transgenic plants did
not change susceptibility to bacterial blight (Xanthomonas oryzae pv oryzae). (E, F)
PigmR transgenic plants did not change susceptibility to fungal sheath blight
(Rhizoctonia solani).

11
Fig. S2. Gene structure of the three NLR members of the Pigm locus.
The full-length open reading frame (ORF) was determined for the expressed members,
R4, R6 (named PigmR) and R8 (named PigmS), by RT-PCR and sequencing using
specific primers listed on table S7. Exons are indicated by black rectangle. Introns are
indicated by lines. The 5’-UTR (untranslated region) and 3’-UTR are indicated by white
rectangle. The initiation (ATG) and termination (TGA) codons are also indicated.
12
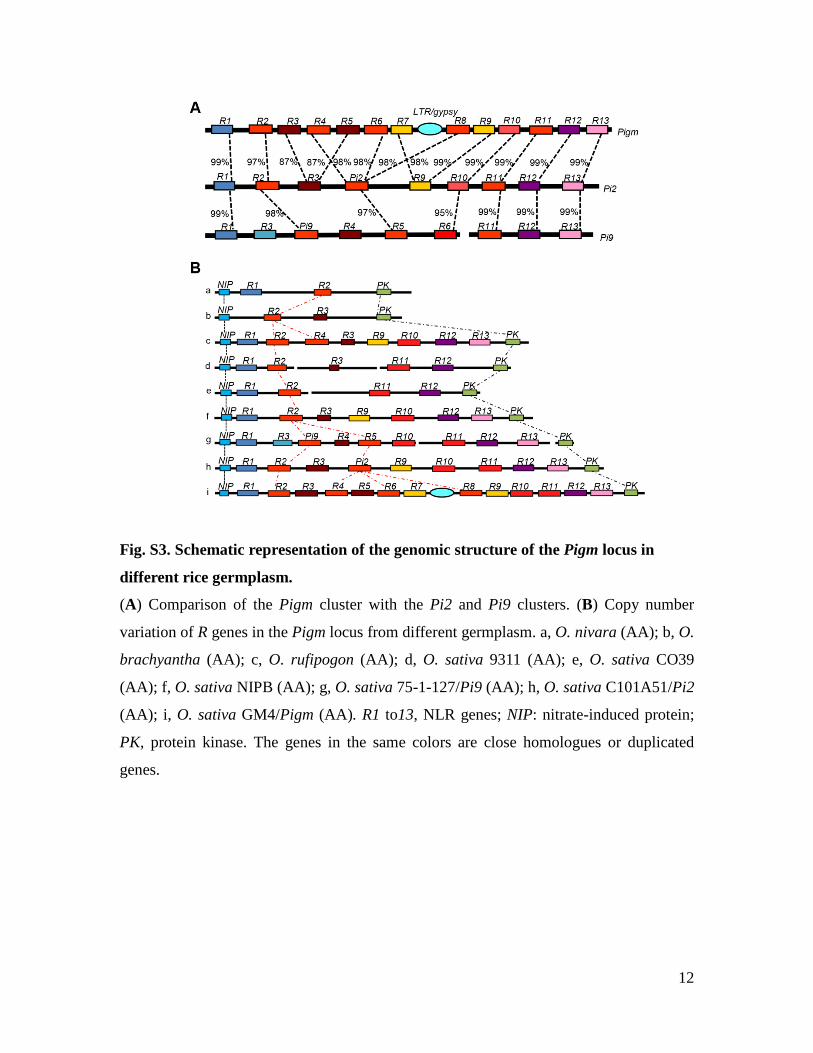
Fig. S3. Schematic representation of the genomic structure of the Pigm locus in
different rice germplasm.
(A) Comparison of the Pigm cluster with the Pi2 and Pi9 clusters. (B) Copy number
variation of R genes in the Pigm locus from different germplasm. a, O. nivara (AA); b, O.
brachyantha (AA); c, O. rufipogon (AA); d, O. sativa 9311 (AA); e, O. sativa CO39
(AA); f, O. sativa NIPB (AA); g, O. sativa 75-1-127/Pi9 (AA); h, O. sativa C101A51/Pi2
(AA); i, O. sativa GM4/Pigm (AA). R1 to13, NLR genes; NIP: nitrate-induced protein;
PK, protein kinase. The genes in the same colors are close homologues or duplicated
genes.
13
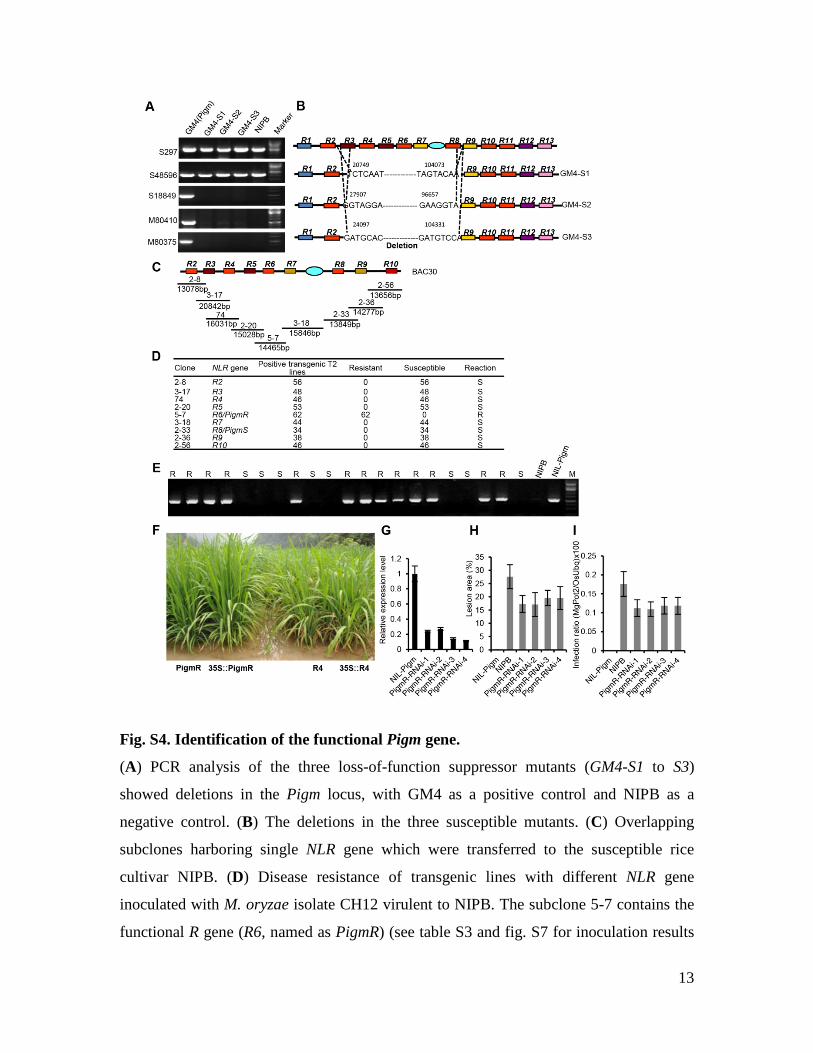
Fig. S4. Identification of the functional Pigm gene.
(A) PCR analysis of the three loss-of-function suppressor mutants (GM4-S1 to S3)
showed deletions in the Pigm locus, with GM4 as a positive control and NIPB as a
negative control. (B) The deletions in the three susceptible mutants. (C) Overlapping
subclones harboring single NLR gene which were transferred to the susceptible rice
cultivar NIPB. (D) Disease resistance of transgenic lines with different NLR gene
inoculated with M. oryzae isolate CH12 virulent to NIPB. The subclone 5-7 contains the
functional R gene (R6, named as PigmR) (see table S3 and fig. S7 for inoculation results

14
with more isolates). (E) PCR analysis of 21 plants from the T2 generation of a PigmR
line indicated that all the transgene positive plants were blast resistant, with NIL-Pigm as
the positive and NIPB as negative controls. R, resistance; S, susceptible (D, E). (F)
Disease resistance of transgenic lines with PigmR, driven by its native promoter (PigmR)
or the 35S promoter (35S::PigmR) in the blast nursery, with the transgenic R4 lines (R4
and 35S::R4) as controls that do not confer blast resistance. Note that the PigmR and
35S::PigmR transgenic plants exhibit complete resistance against M. oryzae. (G) qRT-
PCR detection of PigmR transcript levels in the 4 representative PigmR-RNAi lines in
NIL-Pigm background, the rice OsActin1 was used as a control to normalize expression
levels. (H) Lesion areas of the PigmR-RNAi lines at 7 dpi with CH12. (I) Infection ratio
of the PigmR-RNAi lines at 7 dpi with CH12.
15
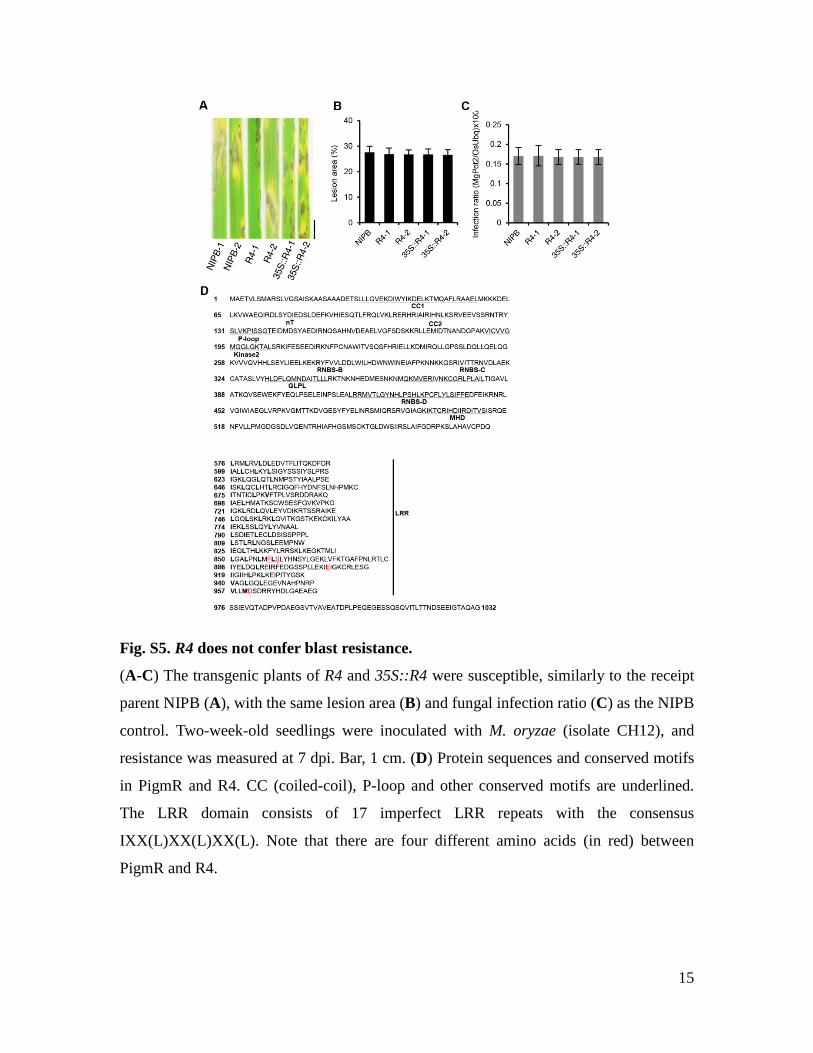
Fig. S5. R4 does not confer blast resistance.
(A-C) The transgenic plants of R4 and 35S::R4 were susceptible, similarly to the receipt
parent NIPB (A), with the same lesion area (B) and fungal infection ratio (C) as the NIPB
control. Two-week-old seedlings were inoculated with M. oryzae (isolate CH12), and
resistance was measured at 7 dpi. Bar, 1 cm. (D) Protein sequences and conserved motifs
in PigmR and R4. CC (coiled-coil), P-loop and other conserved motifs are underlined.
The LRR domain consists of 17 imperfect LRR repeats with the consensus
IXX(L)XX(L)XX(L). Note that there are four different amino acids (in red) between
PigmR and R4.

16
Fig. S6. Functional identification of the four amino acids in the PigmR LRR domain.
(A) Substitution of four amino acids within the LRR domain of PigmR with ones of R4:
F858V, S860Y, E909Q and D961Y. (B) Inoculation experiment with different blast
isolates showed that the site-specific substitutions in the 4 amino acids changed resistance
spectrum. MR, middle resistance; MS, middle susceptibility.

17
Fig. S7. Blast resistance of PigmR against representative strains with
complementary avirulence effectors from different countries.
(A) Inoculation of different blast strains with different avirulence spectra indicates that
the 3 representative PigmR transgenic lines are all resistant to the world-selection strains
with diverse predicted effectors, supporting that PigmR confers broad-spectrum
resistance to M. oryzae. The susceptible variety Maratelli and the transgenic parent NIPB
were used as controls. (B) Factorial analysis of genotypic diversity of the M. oryzae
strains collected from worldwide rice-cultivated regions. Red dots are the collection
positions of the strains used in this study, and gray dots indicate positions of M. oryzae
isolates from a worldwide collection (7, 8).
18
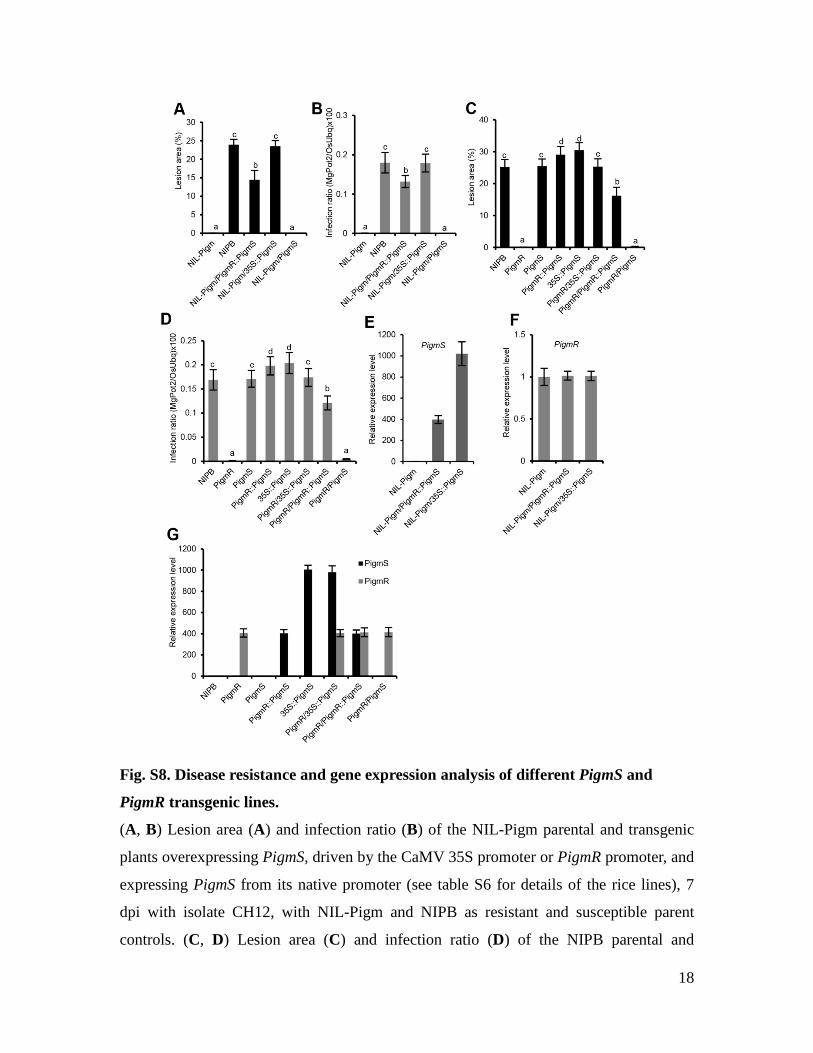
Fig. S8. Disease resistance and gene expression analysis of different PigmS and
PigmR transgenic lines.
(A, B) Lesion area (A) and infection ratio (B) of the NIL-Pigm parental and transgenic
plants overexpressing PigmS, driven by the CaMV 35S promoter or PigmR promoter, and
expressing PigmS from its native promoter (see table S6 for details of the rice lines), 7
dpi with isolate CH12, with NIL-Pigm and NIPB as resistant and susceptible parent
controls. (C, D) Lesion area (C) and infection ratio (D) of the NIPB parental and

19
transgenic plants overexpressing PigmS in NIPB and PigmR transgene background at 7
dpi. Data are shown as means ± SD (n = 30). Means labeled with different letters indicate
significant difference at 1% level via Tukey-Kramer test for multiple comparisons (A to
D). (E) PigmS transcript levels in NIL-Pigm transgenic plants ectopically expressing
PigmS by 35S or the PigmR promoter, with the wild-type NIL-Pigm as control. (F)
PigmR expression was not affected in NIL-Pigm transgenic plants overexpressing PigmS
(A). (G) Transcript levels of PigmS and PigmR in transgenic plants overexpressing
PigmS in NIPB or PigmR transgene background (C). Gene expression levels were
detected by qRT-PCR. The rice OsActin1 was used as a control to normalize expression
levels (E to G).

20
Fig. S9. Yeast two-hybrid assay to identify protein interaction.
(A, B) Yeast two-hybrid assay confirmed that the CC domains of PigmR and PigmS are required for pairwise interaction of PigmR and PigmS, confirmed by growth of yeast cells on basal media without supplement of Trp, Leu, Ade and His and supplemented with 3mM 3AT. (C, D) Yeast two-hybrid assay showed that PigmS does not interact with the full length CDS and different domains of Pish that confers race-specific resistance to M. oryzae isolate YN2 in the NIPB genome. (E) PigmS did not decrease resistance to isolate YN2 avirulent to NIPB. Two-week-old seedlings were inoculated, photographs were taken 7 dpi. Bar, 1 cm.
21
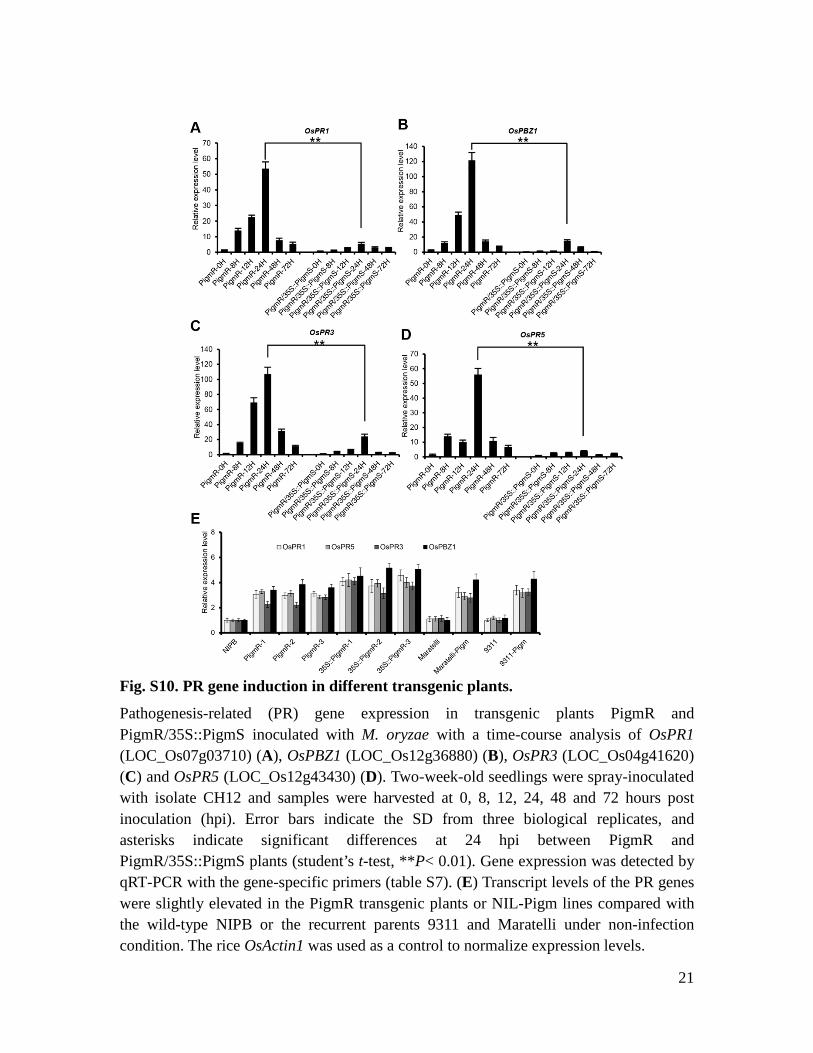
Fig. S10. PR gene induction in different transgenic plants.
Pathogenesis-related (PR) gene expression in transgenic plants PigmR and PigmR/35S::PigmS inoculated with M. oryzae with a time-course analysis of OsPR1 (LOC_Os07g03710) (A), OsPBZ1 (LOC_Os12g36880) (B), OsPR3 (LOC_Os04g41620) (C) and OsPR5 (LOC_Os12g43430) (D). Two-week-old seedlings were spray-inoculated with isolate CH12 and samples were harvested at 0, 8, 12, 24, 48 and 72 hours post inoculation (hpi). Error bars indicate the SD from three biological replicates, and asterisks indicate significant differences at 24 hpi between PigmR and PigmR/35S::PigmS plants (student’s t-test, **P< 0.01). Gene expression was detected by qRT-PCR with the gene-specific primers (table S7). (E) Transcript levels of the PR genes were slightly elevated in the PigmR transgenic plants or NIL-Pigm lines compared with the wild-type NIPB or the recurrent parents 9311 and Maratelli under non-infection condition. The rice OsActin1 was used as a control to normalize expression levels.
22
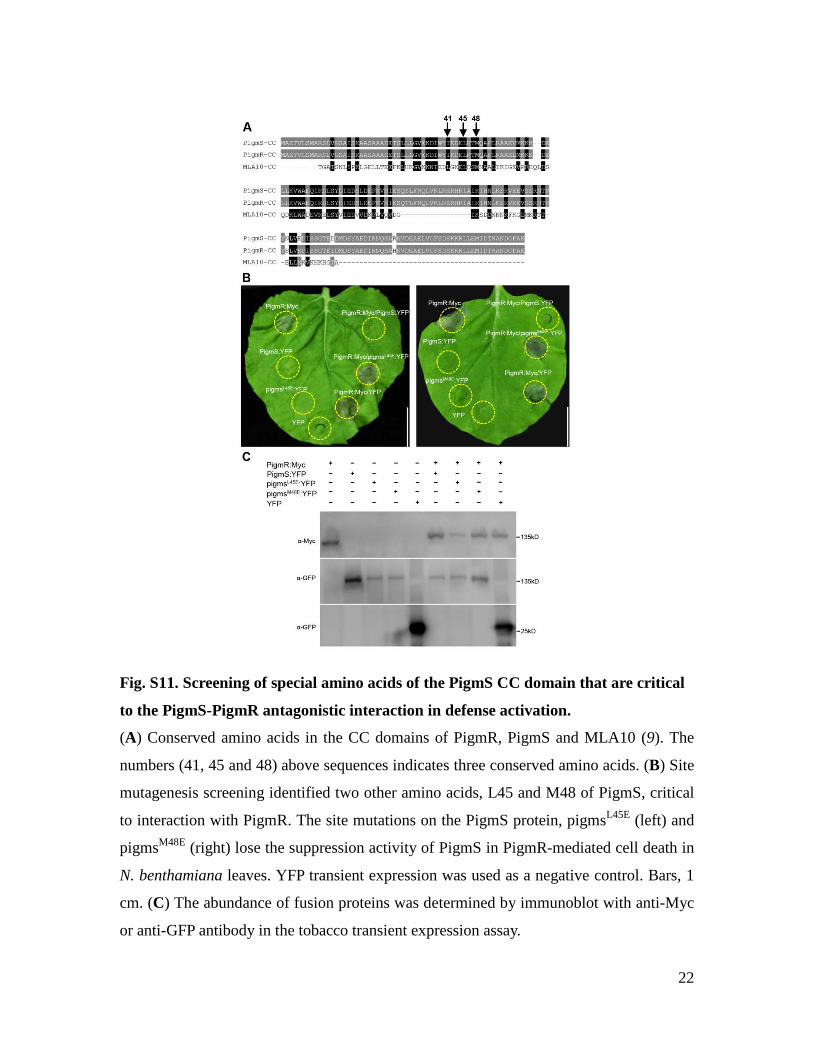
Fig. S11. Screening of special amino acids of the PigmS CC domain that are critical
to the PigmS-PigmR antagonistic interaction in defense activation.
(A) Conserved amino acids in the CC domains of PigmR, PigmS and MLA10 (9). The
numbers (41, 45 and 48) above sequences indicates three conserved amino acids. (B) Site
mutagenesis screening identified two other amino acids, L45 and M48 of PigmS, critical
to interaction with PigmR. The site mutations on the PigmS protein, pigmsL45E (left) and
pigmsM48E (right) lose the suppression activity of PigmS in PigmR-mediated cell death in
N. benthamiana leaves. YFP transient expression was used as a negative control. Bars, 1
cm. (C) The abundance of fusion proteins was determined by immunoblot with anti-Myc
or anti-GFP antibody in the tobacco transient expression assay.
23
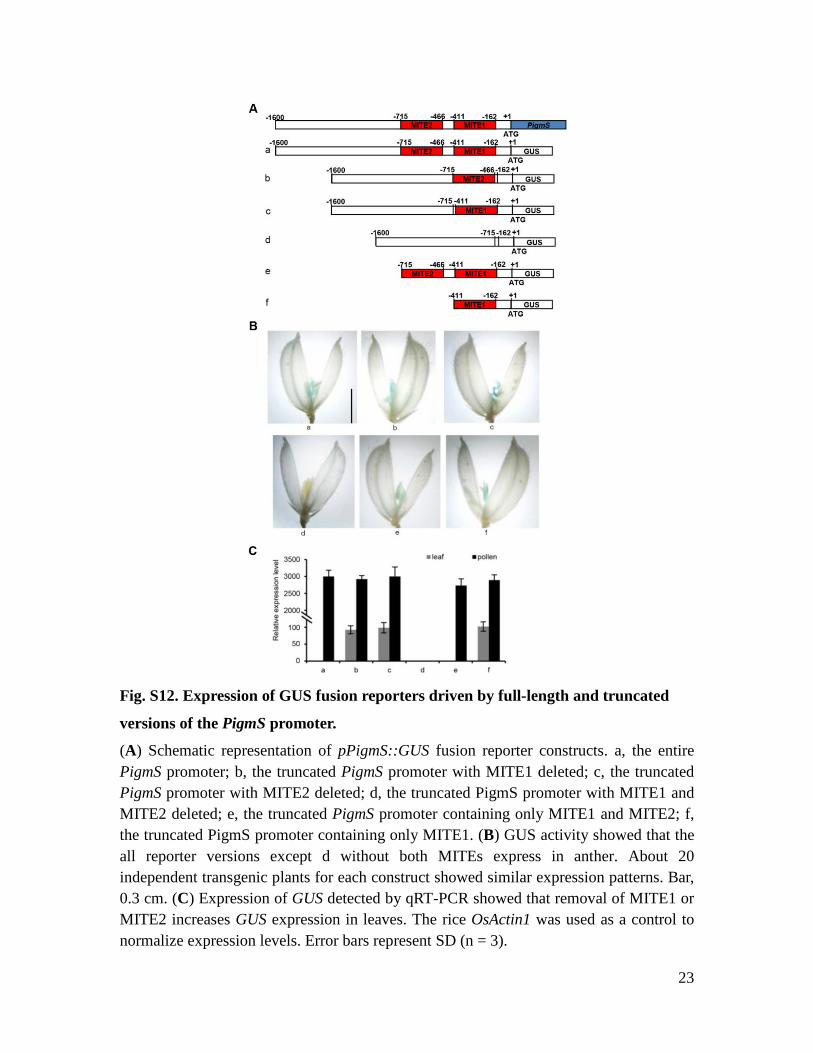
Fig. S12. Expression of GUS fusion reporters driven by full-length and truncated
versions of the PigmS promoter.
(A) Schematic representation of pPigmS::GUS fusion reporter constructs. a, the entire PigmS promoter; b, the truncated PigmS promoter with MITE1 deleted; c, the truncated PigmS promoter with MITE2 deleted; d, the truncated PigmS promoter with MITE1 and MITE2 deleted; e, the truncated PigmS promoter containing only MITE1 and MITE2; f, the truncated PigmS promoter containing only MITE1. (B) GUS activity showed that the all reporter versions except d without both MITEs express in anther. About 20 independent transgenic plants for each construct showed similar expression patterns. Bar, 0.3 cm. (C) Expression of GUS detected by qRT-PCR showed that removal of MITE1 or MITE2 increases GUS expression in leaves. The rice OsActin1 was used as a control to normalize expression levels. Error bars represent SD (n = 3).
24
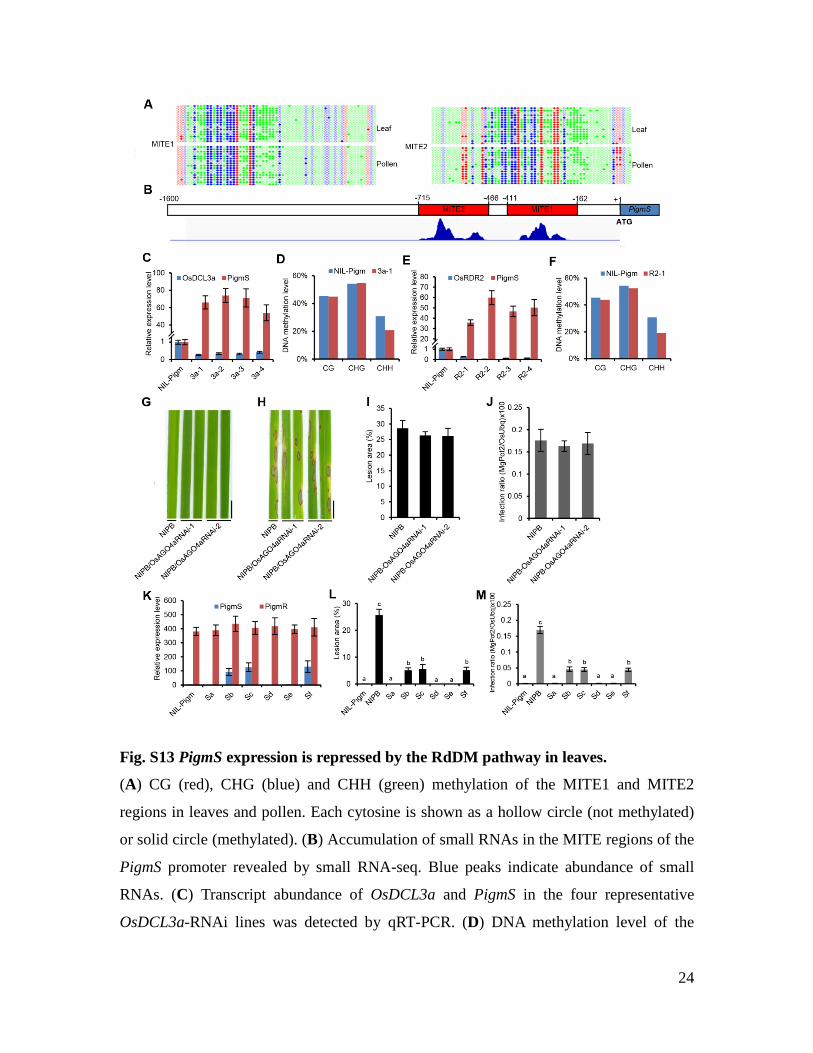
Fig. S13 PigmS expression is repressed by the RdDM pathway in leaves.
(A) CG (red), CHG (blue) and CHH (green) methylation of the MITE1 and MITE2
regions in leaves and pollen. Each cytosine is shown as a hollow circle (not methylated)
or solid circle (methylated). (B) Accumulation of small RNAs in the MITE regions of the
PigmS promoter revealed by small RNA-seq. Blue peaks indicate abundance of small
RNAs. (C) Transcript abundance of OsDCL3a and PigmS in the four representative
OsDCL3a-RNAi lines was detected by qRT-PCR. (D) DNA methylation level of the

25
MITE sequences in the leaves of the OsDCL3a-RNAi line, detected by bisulfite
sequencing. (E) Transcript abundance of OsRDR2 and PigmS in the four representative
OsRDR2-RNAi lines was detected by qRT-PCR. (F) DNA methylation level of the MITE
sequences in the leaves of the OsRDR2-RNAi line, detected by bisulfite sequencing. Note
that CHH methylation of the MITE region in leaves is decreased in the RNAi lines
compared with the wild-type (NIL-Pigm) (D, F). (G, H) Disease phenotypes of the two
different OsAGO4a-RNAi lines in the NIPB background infected with isolates YN2 (G)
and CH12 (H). Bar, 1 cm. (I, J) Quantitative analysis of lesion area (I) and M. oryzae
growth (J) in the OsAGO4a-RNAi lines infected with isolate CH12 at 7 dpi. The results
indicate that the OsAGO4a does not affect blast resistance in NIPB that does not have the
Pigm locus or PigmS gene. (K) Expression levels of PigmS and PigmR in NIL-Pigm
transgenic lines expressing PigmS from different truncated promoters in leaves. Sa, the
full length PigmS promoter; Sb, the truncated PigmS promoter with MITE1 deleted; c,
the truncated PigmS promoter with MITE2 deleted; d, the truncated PigmS promoter with
MITE1 and MITE2 deleted; e, the truncated PigmS promoter containing only MITE1 and
MITE2; Sf, the truncated PigmS promoter containing only MITE1 (see fig. S12A for
details). Note that removal of the MITE sequence increases PigmS expression in leaves,
similar to the GUS reporter expression (fig. S12, B and C), but does not affect PigmR
expression. Consequently, PigmR-mediated resistance is compromised in the PigmS
transgenic lines (Sb, Sc and Sf). (L, M) Quantitative analysis of lesion area (L) and M.
oryzae growth (M) in the in NIL-Pigm transgenic lines expressing PigmS from different
truncated promoters infected with isolate CH12 at 7 dpi. The rice OsActin1 was used as a
control to normalize expression levels (C, E, K). Error bars represent SD (n = 3) (C, E,
K), (n = 30) (I, J, L, M).

26
Fig. S14 Effect of PigmR and PigmS on grain yield components in transgenic lines
and NIL-Pigm.
(A-C) Comparison of grain yield per plant (A), 1000-grain weight (B) and seed setting
ratio (C) between the NILs (9311 vs 9311-Pigm, Maratelli vs Maratelli-Pigm) under no-
disease conditions. (D) Transcript levels of PigmR in leaves and PigmS in anthers of the
RNAi lines that synchronously knocked down PigmR and PigmS (PigmRS-RNAi), which
were normalized as 1 with OsActin1 in the wild-type NIL-Pigm leaves and anthers,
respectively. Data are shown as means ± SD (n = 3). (E-G) Comparison of seed setting
ratio (E), 1000-grain weight (F) and grain yield per plant (G) between the PigmR RNAi,
PigmRS RNAi transgenic lines and the NIL-Pigm control (NIPB background). (H, I)

27
Other agronomic traits including tiller number (H) and plant height (I) are not affected by
PigmR and PigmS in transgenic plants. Data are shown as means ± SD (n = 60) (A to C,
E to I). Means labeled with different letters indicate significant difference at 5% level via
Tukey-Kramer test for multiple comparisons (E to G), and asterisks represent
significance difference determined by the Student’s t-test at P < 0.05 (A to C).
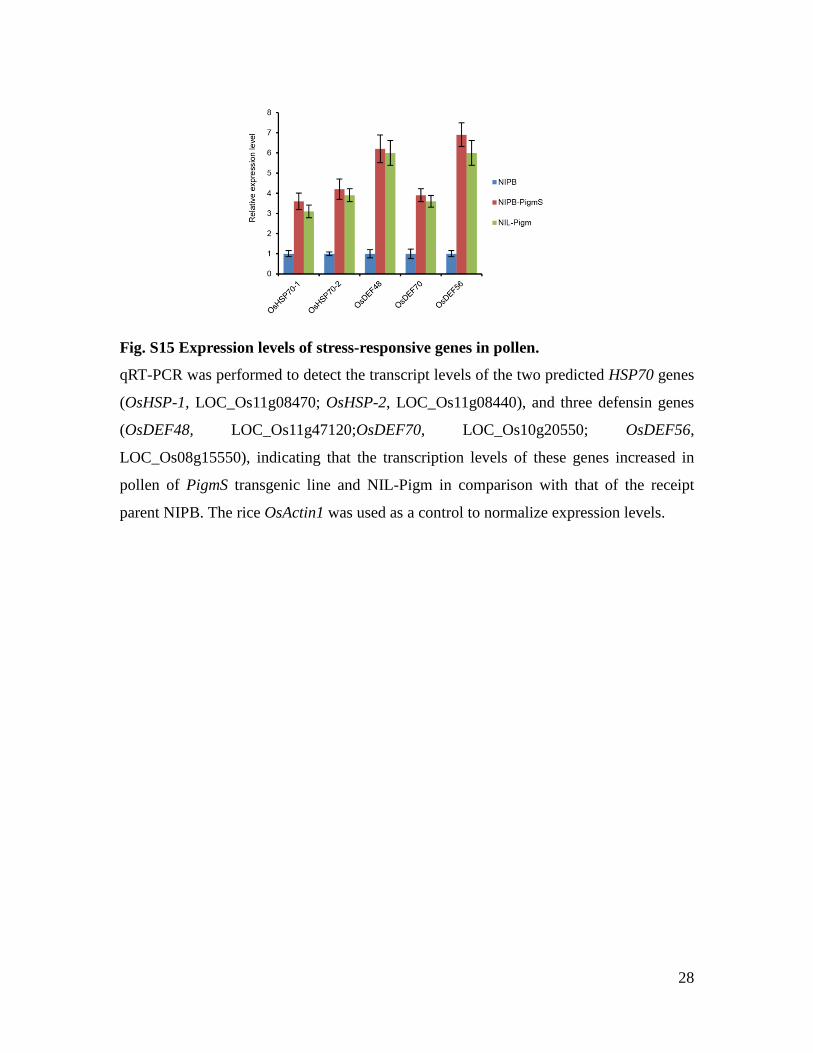
28
Fig. S15 Expression levels of stress-responsive genes in pollen.
qRT-PCR was performed to detect the transcript levels of the two predicted HSP70 genes
(OsHSP-1, LOC_Os11g08470; OsHSP-2, LOC_Os11g08440), and three defensin genes
(OsDEF48, LOC_Os11g47120;OsDEF70, LOC_Os10g20550; OsDEF56,
LOC_Os08g15550), indicating that the transcription levels of these genes increased in
pollen of PigmS transgenic line and NIL-Pigm in comparison with that of the receipt
parent NIPB. The rice OsActin1 was used as a control to normalize expression levels.
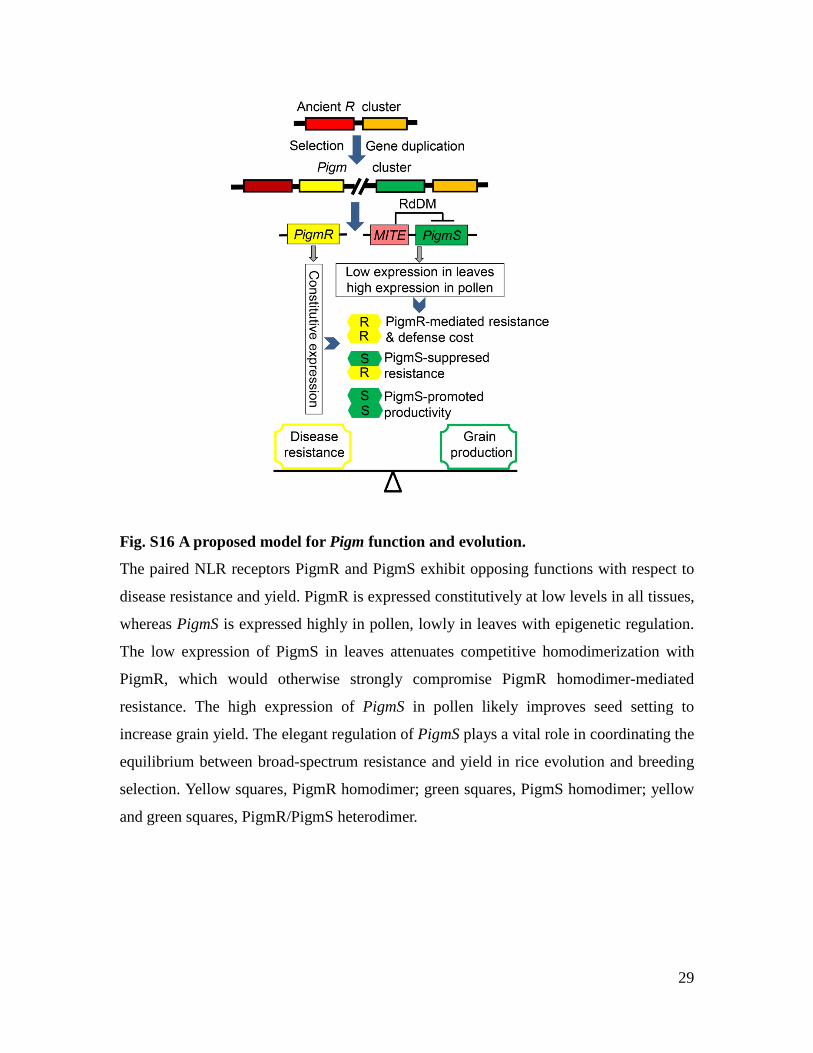
29
Fig. S16 A proposed model for Pigm function and evolution.
The paired NLR receptors PigmR and PigmS exhibit opposing functions with respect to
disease resistance and yield. PigmR is expressed constitutively at low levels in all tissues,
whereas PigmS is expressed highly in pollen, lowly in leaves with epigenetic regulation.
The low expression of PigmS in leaves attenuates competitive homodimerization with
PigmR, which would otherwise strongly compromise PigmR homodimer-mediated
resistance. The high expression of PigmS in pollen likely improves seed setting to
increase grain yield. The elegant regulation of PigmS plays a vital role in coordinating the
equilibrium between broad-spectrum resistance and yield in rice evolution and breeding
selection. Yellow squares, PigmR homodimer; green squares, PigmS homodimer; yellow
and green squares, PigmR/PigmS heterodimer.
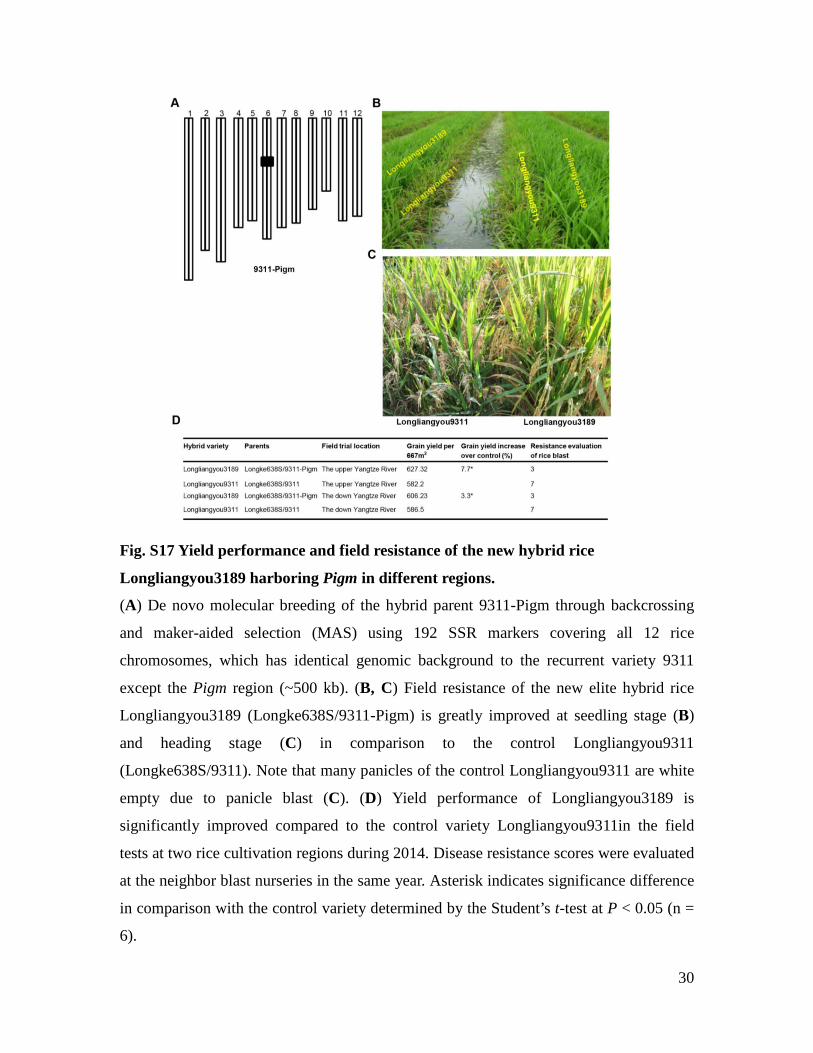
30
Fig. S17 Yield performance and field resistance of the new hybrid rice
Longliangyou3189 harboring Pigm in different regions.
(A) De novo molecular breeding of the hybrid parent 9311-Pigm through backcrossing
and maker-aided selection (MAS) using 192 SSR markers covering all 12 rice
chromosomes, which has identical genomic background to the recurrent variety 9311
except the Pigm region (~500 kb). (B, C) Field resistance of the new elite hybrid rice
Longliangyou3189 (Longke638S/9311-Pigm) is greatly improved at seedling stage (B)
and heading stage (C) in comparison to the control Longliangyou9311
(Longke638S/9311). Note that many panicles of the control Longliangyou9311 are white
empty due to panicle blast (C). (D) Yield performance of Longliangyou3189 is
significantly improved compared to the control variety Longliangyou9311in the field
tests at two rice cultivation regions during 2014. Disease resistance scores were evaluated
at the neighbor blast nurseries in the same year. Asterisk indicates significance difference
in comparison with the control variety determined by the Student’s t-test at P < 0.05 (n =
6).
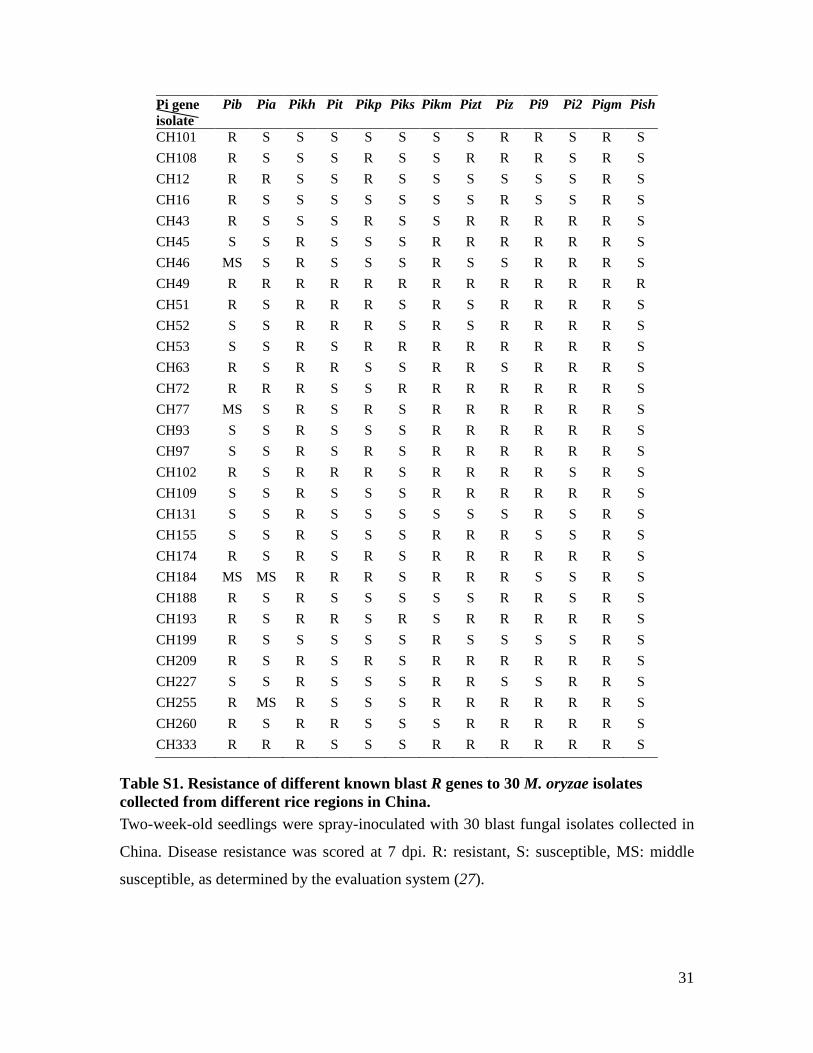
31
Pi gene isolate
Pib Pia Pikh Pit Pikp Piks Pikm Pizt Piz Pi9 Pi2 Pigm Pish
CH101 R S S S S S S S R R S R S CH108 R S S S R S S R R R S R S CH12 R R S S R S S S S S S R S CH16 R S S S S S S S R S S R S CH43 R S S S R S S R R R R R S CH45 S S R S S S R R R R R R S CH46 MS S R S S S R S S R R R S CH49 R R R R R R R R R R R R R CH51 R S R R R S R S R R R R S CH52 S S R R R S R S R R R R S CH53 S S R S R R R R R R R R S CH63 R S R R S S R R S R R R S CH72 R R R S S R R R R R R R S CH77 MS S R S R S R R R R R R S CH93 S S R S S S R R R R R R S CH97 S S R S R S R R R R R R S CH102 R S R R R S R R R R S R S CH109 S S R S S S R R R R R R S CH131 S S R S S S S S S R S R S CH155 S S R S S S R R R S S R S CH174 R S R S R S R R R R R R S CH184 MS MS R R R S R R R S S R S CH188 R S R S S S S S R R S R S CH193 R S R R S R S R R R R R S CH199 R S S S S S R S S S S R S CH209 R S R S R S R R R R R R S CH227 S S R S S S R R S S R R S CH255 R MS R S S S R R R R R R S CH260 R S R R S S S R R R R R S CH333 R R R S S S R R R R R R S
Table S1. Resistance of different known blast R genes to 30 M. oryzae isolates collected from different rice regions in China. Two-week-old seedlings were spray-inoculated with 30 blast fungal isolates collected in
China. Disease resistance was scored at 7 dpi. R: resistant, S: susceptible, MS: middle
susceptible, as determined by the evaluation system (27).

32
Variety Origin Release year GuanfengA Fujian 2002 GufengA Fujian 2002 ChuanguA Sichuan 2005 Bing1A Hunan 2009 FuyiA Fujian 1995 AnfengA Fujian 2011 Fuyou58 Hubei 1990 Gunong13 Guangxi ND Digu Fujian 1992 XiafengA Fujian 2002 Endao3 Hubei 1994 Shuangchao25 Guangdong 1985 ZhongguA Zhejiang 2010 Peiyou2 Chongqing 2008 Guangkang13A Fujian 2003 Guangyou2643 Fujian 2013 Chuanguyou538 Sichuan 2012 Guyou428 Fujian 2014 Guangyou66 Sichuan 2014 Guangyou673 Sichuan 2014 Guyou527 Fujian 2007 Quanyou527 Fujian 2010 Quanyou77 Fujian 2007 Quanyou94 Fujian 2007 Leyou94 Fujian 2007 ChangfengA Fujian 2011 LianfengA Fujian 2002 LefengA Fujian 2005 FufengA Fujian 2005 JiefengA Fujian 2005 ChengfengA Fujian 2010 Shenyou9734 Guangdong 2012 Shenyou9752 Guangdong 2012 Shen97A Guangdong 2007 N7A Sichuan 2003 Jun1A Hubei 2010 N5B Sichuan 2009 Fuhui9801 Chongqing 2001 803B Chongqing 2002 Fuhui9802 Chongqing 2010 Lingyou2 Chongqing 2008
Table S2. Commercial blast resistant rice varieties harboring Pigm.
A nationwide collection of commercial rice varieties with high blast resistance was

33
screened for the Pigm locus with Pigm-specific primers (table S7). These varieties
containing Pigm are mostly indica, some have been long grown. ND, not determined.
34
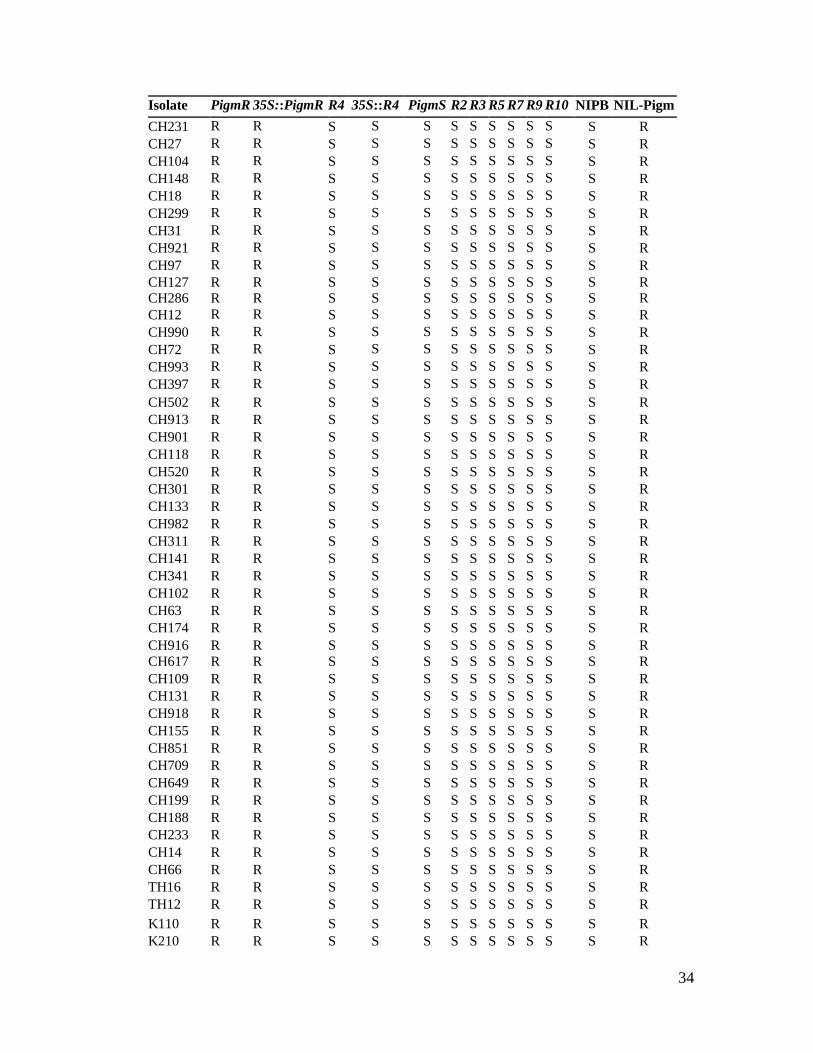
Isolate PigmR 35S::PigmR R4 35S::R4 PigmS R2 R3 R5 R7 R9 R10 NIPB NIL-Pigm CH231 R R S S S S S S S S S S R CH27 R R S S S S S S S S S S R CH104 R R S S S S S S S S S S R CH148 R R S S S S S S S S S S R CH18 R R S S S S S S S S S S R CH299 R R S S S S S S S S S S R CH31 R R S S S S S S S S S S R CH921 R R S S S S S S S S S S R CH97 R R S S S S S S S S S S R CH127 CH286
R R
R R
S S
S S
S S
S S
S S
S S
S S
S S
S S
S S
R R
CH12 R R S S S S S S S S S S R CH990 R R S S S S S S S S S S R CH72 R R S S S S S S S S S S R CH993 R R S S S S S S S S S S R CH397 R R S S S S S S S S S S R CH502 R R S S S S S S S S S S R CH913 R R S S S S S S S S S S R CH901 R R S S S S S S S S S S R CH118 R R S S S S S S S S S S R CH520 R R S S S S S S S S S S R CH301 R R S S S S S S S S S S R CH133 R R S S S S S S S S S S R CH982 R R S S S S S S S S S S R CH311 R R S S S S S S S S S S R CH141 R R S S S S S S S S S S R CH341 R R S S S S S S S S S S R CH102 R R S S S S S S S S S S R CH63 R R S S S S S S S S S S R CH174 R R S S S S S S S S S S R CH916 R R S S S S S S S S S S R CH617 R R S S S S S S S S S S R CH109 R R S S S S S S S S S S R CH131 R R S S S S S S S S S S R CH918 R R S S S S S S S S S S R CH155 R R S S S S S S S S S S R CH851 R R S S S S S S S S S S R CH709 R R S S S S S S S S S S R CH649 R R S S S S S S S S S S R CH199 R R S S S S S S S S S S R CH188 CH233
R R
R R
S S
S S
S S
S S
S S
S S
S S
S S
S S
S S
R R
CH14 R R S S S S S S S S S S R CH66 R R S S S S S S S S S S R TH16 R R S S S S S S S S S S R TH12 R R S S S S S S S S S S R K110 R R S S S S S S S S S S R K210 R R S S S S S S S S S S R

35
K102 GUY11
R R
R R
S S
S S
S S
S S
S S
S S
S S
S S
S S
S S
R R
Table S3. Blast resistance evaluation of transgenic lines with different R members of
the Pigm cluster to 50 M. oryzae isolates.
Two-week-old seedlings of the stable transgenic lines were spray-inoculated with a set of
blast collection (50 isolates). Disease resistance was scored at 7 dpi. NIPB is the wild-
type control, and NIL-Pigm is the donor control. Note that only PigmR expressed with its
native promoter (PigmR) or the 35S promoter (35S::PigmR) conferred resistance to all
isolates tested as the donor NIL-Pigm.
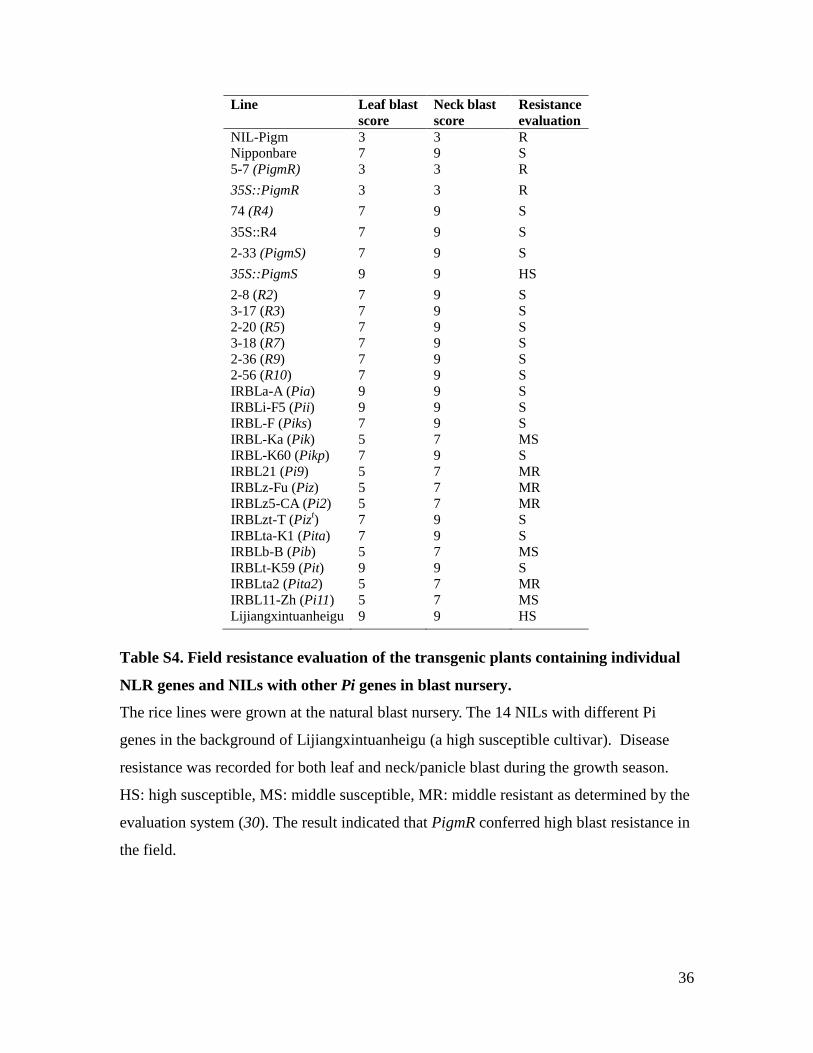
36
Line Leaf blast score
Neck blast score
Resistance evaluation
NIL-Pigm Nipponbare
3 7
3 9
R S
5-7 (PigmR) 3 3 R 35S::PigmR 3 3 R 74 (R4) 7 9 S 35S::R4 7 9 S 2-33 (PigmS) 7 9 S 35S::PigmS 9 9 HS 2-8 (R2) 3-17 (R3) 2-20 (R5) 3-18 (R7) 2-36 (R9) 2-56 (R10)
7 7 7 7 7 7
9 9 9 9 9 9
S S S S S S
IRBLa-A (Pia) IRBLi-F5 (Pii) IRBL-F (Piks) IRBL-Ka (Pik) IRBL-K60 (Pikp) IRBL21 (Pi9) IRBLz-Fu (Piz) IRBLz5-CA (Pi2) IRBLzt-T (Pizt) IRBLta-K1 (Pita) IRBLb-B (Pib) IRBLt-K59 (Pit) IRBLta2 (Pita2) IRBL11-Zh (Pi11)
9 9 7 5 7 5 5 5 7 7 5 9 5 5
9 9 9 7 9 7 7 7 9 9 7 9 7 7
S S S MS S MR MR MR S S MS S MR MS
Lijiangxintuanheigu 9 9 HS
Table S4. Field resistance evaluation of the transgenic plants containing individual
NLR genes and NILs with other Pi genes in blast nursery.
The rice lines were grown at the natural blast nursery. The 14 NILs with different Pi
genes in the background of Lijiangxintuanheigu (a high susceptible cultivar). Disease
resistance was recorded for both leaf and neck/panicle blast during the growth season.
HS: high susceptible, MS: middle susceptible, MR: middle resistant as determined by the
evaluation system (30). The result indicated that PigmR conferred high blast resistance in
the field.
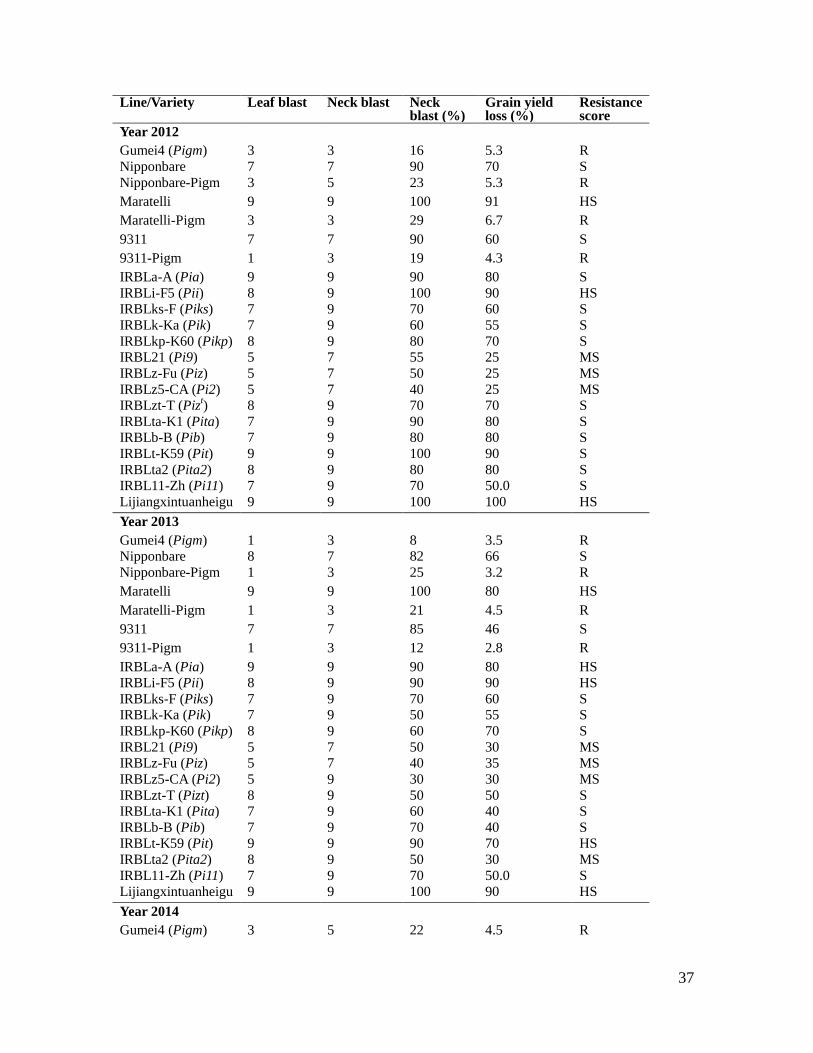
37
Line/Variety Leaf blast
Neck blast Neck blast (%)
Grain yield loss (%)
Resistance score
Year 2012 Gumei4 (Pigm) Nipponbare
3 7
3 7
16 90
5.3 70
R S
Nipponbare-Pigm 3 5 23 5.3 R Maratelli 9 9 100 91 HS Maratelli-Pigm 3 3 29 6.7 R 9311 7 7 90 60 S 9311-Pigm 1 3 19 4.3 R IRBLa-A (Pia) IRBLi-F5 (Pii) IRBLks-F (Piks) IRBLk-Ka (Pik) IRBLkp-K60 (Pikp) IRBL21 (Pi9) IRBLz-Fu (Piz) IRBLz5-CA (Pi2) IRBLzt-T (Pizt) IRBLta-K1 (Pita) IRBLb-B (Pib) IRBLt-K59 (Pit) IRBLta2 (Pita2) IRBL11-Zh (Pi11)
9 8 7 7 8 5 5 5 8 7 7 9 8 7
9 9 9 9 9 7 7 7 9 9 9 9 9 9
90 100 70 60 80 55 50 40 70 90 80 100 80 70
80 90 60 55 70 25 25 25 70 80 80 90 80 50.0
S HS S S S MS MS MS S S S S S S
Lijiangxintuanheigu 9 9 100 100 HS Year 2013 Gumei4 (Pigm) Nipponbare
1 8
3 7
8 82
3.5 66
R S
Nipponbare-Pigm 1 3 25 3.2 R Maratelli 9 9 100 80 HS Maratelli-Pigm 1 3 21 4.5 R 9311 7 7 85 46 S 9311-Pigm 1 3 12 2.8 R IRBLa-A (Pia) IRBLi-F5 (Pii) IRBLks-F (Piks) IRBLk-Ka (Pik) IRBLkp-K60 (Pikp) IRBL21 (Pi9) IRBLz-Fu (Piz) IRBLz5-CA (Pi2) IRBLzt-T (Pizt) IRBLta-K1 (Pita) IRBLb-B (Pib) IRBLt-K59 (Pit) IRBLta2 (Pita2) IRBL11-Zh (Pi11)
9 8 7 7 8 5 5 5 8 7 7 9 8 7
9 9 9 9 9 7 7 9 9 9 9 9 9 9
90 90 70 50 60 50 40 30 50 60 70 90 50 70
80 90 60 55 70 30 35 30 50 40 40 70 30 50.0
HS HS S S S MS MS MS S S S HS MS S
Lijiangxintuanheigu 9 9 100 90 HS Year 2014 Gumei4 (Pigm) 3 5 22 4.5 R
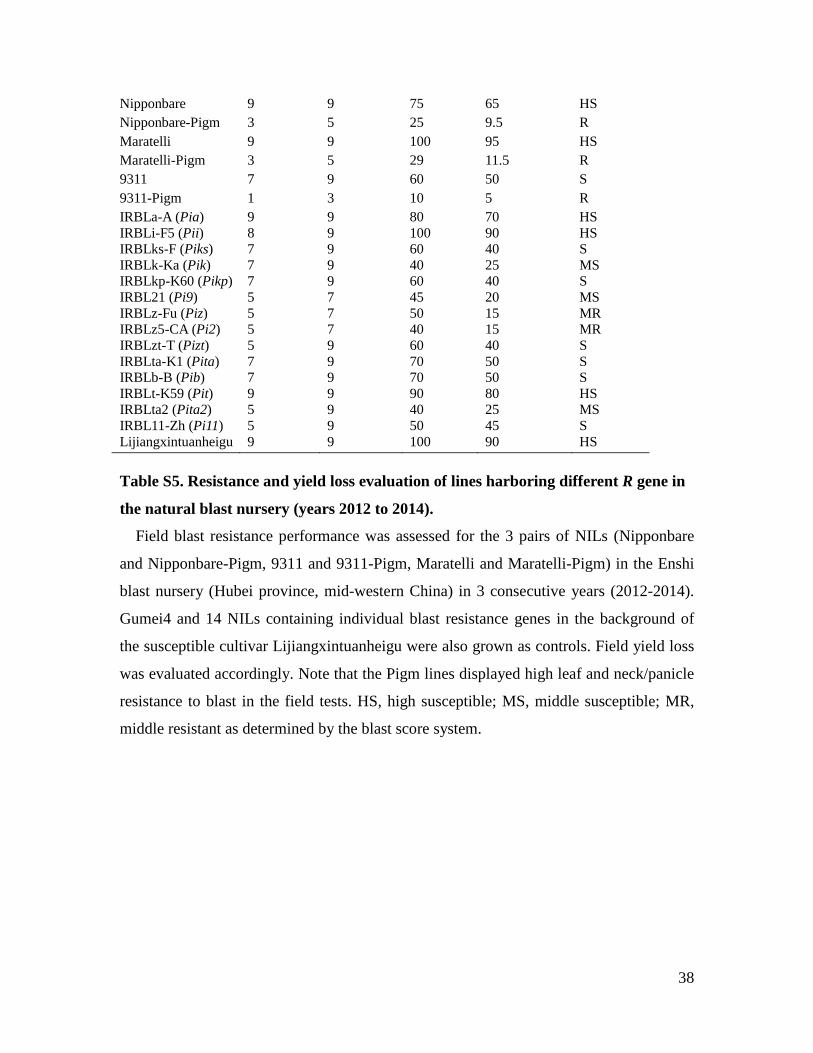
38
Nipponbare 9 9 75 65 HS Nipponbare-Pigm 3 5 25 9.5 R Maratelli 9 9 100 95 HS Maratelli-Pigm 3 5 29 11.5 R 9311 7 9 60 50 S 9311-Pigm 1 3 10 5 R IRBLa-A (Pia) IRBLi-F5 (Pii) IRBLks-F (Piks) IRBLk-Ka (Pik) IRBLkp-K60 (Pikp) IRBL21 (Pi9) IRBLz-Fu (Piz) IRBLz5-CA (Pi2) IRBLzt-T (Pizt) IRBLta-K1 (Pita) IRBLb-B (Pib) IRBLt-K59 (Pit) IRBLta2 (Pita2) IRBL11-Zh (Pi11)
9 8 7 7 7 5 5 5 5 7 7 9 5 5
9 9 9 9 9 7 7 7 9 9 9 9 9 9
80 100 60 40 60 45 50 40 60 70 70 90 40 50
70 90 40 25 40 20 15 15 40 50 50 80 25 45
HS HS S MS S MS MR MR S S S HS MS S
Lijiangxintuanheigu 9 9 100 90 HS Table S5. Resistance and yield loss evaluation of lines harboring different R gene in
the natural blast nursery (years 2012 to 2014).
Field blast resistance performance was assessed for the 3 pairs of NILs (Nipponbare
and Nipponbare-Pigm, 9311 and 9311-Pigm, Maratelli and Maratelli-Pigm) in the Enshi
blast nursery (Hubei province, mid-western China) in 3 consecutive years (2012-2014).
Gumei4 and 14 NILs containing individual blast resistance genes in the background of
the susceptible cultivar Lijiangxintuanheigu were also grown as controls. Field yield loss
was evaluated accordingly. Note that the Pigm lines displayed high leaf and neck/panicle
resistance to blast in the field tests. HS, high susceptible; MS, middle susceptible; MR,
middle resistant as determined by the blast score system.

39
Variety/line MAS/transgene Purpose GM4 Pigm donor Gene cloning Maratelli Recurrent parent NIL development 9311 Recurrent parent NIL development Nipponbare (NIPB) Susceptible variety Transgene receipt and
NIL development NIL-Pigm Near-isogenic lines harboring Pigm in
Nipponbare Transgene receipt variety and resistance analysis
9311-Pigm Near-sogenic lines harboring Pigm in 9311
Resistance and grain yield analysis
Maratelli-Pigm Near-isogenic lines harboring Pigm in Maratelli
Resistance and grain yield analysis
PigmR PigmR transgenic lines in Nipponbare Resistance and grain yield analysis
35S::PigmR PigmR overexpressing transgenic lines in Nipponbare
Resistance and grain yield analysis
PigmS PigmS transgenic lines in Nipponbare Resistance and grain yield analysis
35S::PigmS PigmS overexpressing transgenic lines in Nipponbare
Resistance and grain yield analysis
PigmR::PigmS Chimeric PigmS transgenic lines driven by the PigmR promoter based in Nipponbare
Resistance and grain yield analysis
PigmR/PigmR::PigmS Double transgenic lines in Nipponbare developed by crossing
Resistance and grain yield analysis
PigmR/35S::PigmS Double transgenic lines in Nipponbare developed by crossing
Resistance and grain yield analysis
PigmR/PigmS NIL-Pigm/PigmR::PigmS
Double transgenic lines in Nipponbare developed by crossing Chimeric PigmS transgenic lines driven by the PigmR promoter based in NIL-Pigm
Resistance and grain yield analysis Resistance and grain yield analysis
NIL-Pigm/PigmS PigmS transgenic lines in NIL-Pigm Resistance and grain yield analysis
NIL-Pigm/35S::PigmS PigmS overexpressing transgenic lines in NIL-Pigm
Resistance and grain yield analysis
R4 R4 transgenic lines in Nipponbare Resistance analysis 35S::R4 R4 overexpression transgenic lines in
Nipponbare Resistance analysis
R2 R2 transgenic lines in Nipponbare Resistance analysis R3 R3 transgenic lines in Nipponbare Resistance analysis R5 R5 transgenic lines in Nipponbare Resistance analysis R7 R7 gDNA based on Nipponbare Resistance analysis R9 R9 transgenic lines in Nipponbare Resistance analysis R10 R10 transgenic lines in Nipponbare Resistance analysis PigmR-RNAi PigmR-RNAi lines in NIL-Pigm Resistance and grain
yield analysis PigmRS-RNAi Simultaneous PigmR and PigmS RNAi
lines in NIL-Pigm Resistance and grain yield analysis
4a-1 OsAGO4a-RNAi lines in NIL-Pigm DNA methylation and

40
resistance analysis 3a-1 OsDCL3a-RNAi lines in NIL-Pigm DNA methylation R2-1 OsRDR2-RNAi lines in NIL-Pigm DNA methylation N1 OsAGO4a-RNAi lines in Nipponbare Resistance analysis Sa PigmS CDS transgenic lines driven by
the full PigmS promoter in NIL-Pigm Resistance analysis
Sb PigmS CDS transgenic lines driven by the truncated promoter (deletion of MITE1) in NIL-Pigm
Resistance analysis
Sc PigmS CDS transgenic lines driven by the PigmS truncated promoter (deletion of MITE2) in NIL-Pigm
Resistance analysis
Sd PigmS CDS transgenic lines driven by the PigmS truncated promoter (deletion of MITE1 and MITE2) in NIL-Pigm
Resistance analysis
Se PigmS CDS transgenic lines driven by PigmS truncated promoter (only with MITE1 and MITE2) in NIL-Pigm
Resistance analysis
Sf PigmS CDS transgenic lines driven by PigmS truncated promoter (only containing MITE1) in NIL-Pigm
Resistance analysis
a GUS fusion reporter transgenic lines driven by the full PigmS promoter in Nipponbare
GUS staining analysis
b GUS fusion reporter transgenic lines driven by the PigmS truncated promoter (deletion of MITE1) in Nipponbare
GUS staining analysis
c GUS fusion reporter transgenic lines driven by the PigmS truncated promoter (deletion of MITE2) in Nipponbare
GUS staining analysis
d GUS fusion reporter transgenic lines driven by the PigmS truncated promoter deletion of MITE1 and MITE2) in Nipponbare
GUS staining analysis
e GUS fusion reporter transgenic lines driven by the PigmS truncated promoter (containing MITE1 and MITE2) in Nipponbare
GUS staining analysis
f GUS fusion reporter transgenic lines driven by the PigmS truncated promoter (containing only MITE1) in Nipponbare
GUS staining analysis
IRBLa-A (Pia) IRBLi-F5 (Pii) IRBLks-F (Piks) IRBLk-Ka (Pik) IRBLkp-K60 (Pikp) IRBL21 (Pi9) IRBLz-Fu (Piz) IRBLz5-CA (Pi2) IRBLzt-T (Pizt) IRBLta-K1 (Pita) IRBLb-B (Pib)
The near-isogenic lines with individual Pi genes in Lijiangxintuanheigu for resistance tests
Blast nursery test

41
IRBLt-K59 (Pit) IRBLta2 (Pita2) IRBL11-Zh (Pi11) Lijiangxintuanheigu Table S6. Rice materials developed and used in this study.
All transgenic plants were developed in the Nipponbare background, and the Pigm NILs were developed by backcrossing with the recurrent varieties in this study. The IRBL NILs are world-collection with Pi genes.

42
Gene/clone primer 5'-3' Purpose C5483 TTAGGCTGCTTGTCTTGGG Mapping and genotyping GGGAGGAGGAATGGTAGGAA C24 ACCTCCAGGCTCTAGTCA CCTCTGTTGTTAATCTTCG C29742 CAGTGAAACGAACGCTATG AATAGGAAGGGTTGATGTTG P18849 AACCCTTACCTTGTTACTATCCTCT AAAGATGTAACTATTGCTACTGTCCA S26205 GTTCTCCACTTCACCTCCAT TTGCTCTACCCAAACCTTTA M80375 GACGAGTAAACGAGAAGTCACG GGACAACCATATTCCCCTAAGA M80410 GGATTGTCTTGTCTCTCTCGC CAGGACTTAGGGTTTCTCTCTTT S48596 GTAGTCATCAAGGTCGTAGTCTCG GCCACTGCTTTGCGGTAC C680 TAACTAACACATTATGCCTGCC CGTTTTGAATACTAGCTTCTCC C0428 AAGGTTCTCGTGGTTTCA TCCCCATTGTTTATAGCAG M35572 TCGCTATCCGTATCCACAAC ACTTCTCCGCAAGATCAACA S2 GTTGAAGAAGTGAGTAGCAGGAA Screening the subclone AGTAGCAATGTTATGGCATCGT S3 TATACAGACAAGCAACGCAGTC CATACCGACAACACATACAACCT S4 CGCTACTCAACTGCCTGATG CACTCCAAGATGTTACTGTCCTC S5 GTTACAACTACTTACCGTCTCATCT TCCTTCTAATATCCACTACCTCCAA S6 ATGGTCTTGTTGTAGGTCTGGTA AAGGTTGTGGATACGGATAGCA S7 TCTCACCTCCATTCTGTTCTCC ACTCTTGAATTGCTCTGCTGTC S8 TGGCAATGGCAATAATCGGTTA TGAGGTTGTGGATACGGATAGC S9 TTGTCGGAGTTGCTGGTCTT GGAGAACAGAATGGAGGTGAGA S10 GGTAGCAAGCAGTGGTGGTA ACATTCTAGCGACTCCTTACATAC

43
R4 CGCTGGTCGCCTAGTGCTATCT cDNA amplification CTCTGCCCTTTCTGTTCGCTGA PigmR CGCTGGTCGCCTAGTGCTATCT CTCTGCCCTTTCTGTTCGCTGA PigmS GCTGCTTGCTGTTCTGAGCG AGCCCTGCCCTATCTGTTCG PigmR TTCCTTTCTCTTTATCATAATT qRT-PCR detection AGATTCCAACCTGCACTTGCCT PigmS GTCCTTGATCTTTATCAGAAAG TGGTTCCAACCTGCAGAATCTG OsActin1 GTCCTCTTCCAGCCTTCCTT TACCACCACTGAGAACGATGT OsAGO4a GAGGACCCTTCAATGCGACG TCAGCAAGAACACAGAGCAGAAA OsRDR2 TGGACTACACAGCAACAAGGC TTTCAGTGGCGAACGGTC OsDCL3a GTCTGGAAATCTGTCGCCTG GAGTAGCACTTATCCTGCGGAA OsPR1 GTGTCGGAGAAGCAGTGGTA CGAGTAGTTGCAGGTGATGAAG OsPR3 GTCACCGAGGCGTTCTTCA GCTTGGAGTCGTCGTTGGT OsPR5 GCAGCCAGGACTTCTACGA TGTGTCTTGGTGTTGTCTTCG OsPBZ1 CGCCGCAAGTCATGTCCTAA CCACGATGTCCTTCTCCTTCTC OsHSP70-1 GGACGCCAAGATGGACAAGA CTGGAAGAAGTCGTGGAGCAT OsHSP70-2 AACAACAACAGCAGCGACAG GGTTGCTCCAGATGCCTTCT OsDEF48 CTTGATTGTGCGACTGCTGAT GCCTTCAACTTGGATTCCTTCT OsDEF70 GCCGTTTGTTTCCTCCTAGTT CACAGGTACTTGATGCAGAACA OsDEF56 CTGAACAATGGTGCTGGTATGA GTGAGACTGGACTTTGGCTTT GUS GCGTGGTGATGTGGAGTATTGC TCGCTGATGGTATCGGTGTGAG R8PS1557 GTGTGTTAGTGTGTATAGTGATGTGAA DNA methylation CAACTCTACTCAACTAAATCTCCCTTC R8P1226 AAATAAAATGAGGTATAAATTGTAAAATT

44
ATAATTCACATCACTATACACACTAACA R8PS778 AGTAAATGGAGTAAAATAAATTTTATTA TATTAAATAAATTTTACAATTTATACCT R8pros363 TTTAGGAATTATAGAATTAAGTAAAGTATTT TTCTAAAATTAATTTTATAATATTTTTCAT R8PS525 TTTAGTAGTGGGGATGATTTGTATAGATATATAGG TCCATCTATACCCTATATCCAAATTCCAATCTCA 35S::PigmR CAATGTCGACATGGCGGAGACGGTGCTGAG Transgene constructs ATCAACTAGTTCAGCCAGCTTGAGCTGTG 35S::PigmS CAATGTCGACATGGCGGAGACGGTGCTGAG ATCAACTAGTTCAGCCAGCTTGAGCTGTG PigmR::PigmS ATGCAAGCTTTGGCGTCTTGAGGAGTCGTA ATGCCTGCAGGGACTCGTTCGACTCTCCCT CAATGTCGACATGGCGGAGACGGTGCTGAG ATCAACTAGTTCAGCCAGCTTGAGCTGTG 35S::R4 CAATGTCGACATGGGCGGAGACGGTGCTGA AATACTGCAGTCAGCCAGCTTGAGCTTGAG Sa GTCAAGCTTTTATCTGGCCCAATTTTATATTG CAGTCGACCGACTCTGCTCGACTAGATCTC Sb GTCAAGCTTTTATCTGGCCCAATTTTATATTG AGAGGATTATCATTATAAGAGCAAGTTCAATAGTA TTGAACTTGCTCTTATAATGATAATCCTCTCTGCT CAGTCGACCGACTCTGCTCGACTAGATCTC Sc GTCAAGCTTTTATCTGGCCCAATTTTATATTG GTCCACTATTGTACCTGCTCCAATATTGTCCATCTG
TAC
GTACAGATGGACAATATTGGAGCAGGTACAATAGTGGAC
CAGTCGACCGACTCTGCTCGACTAGATCTC Sd GTCAAGCTTTTATCTGGCCCAATTTTATATT CAAGCAGAGAGGATTATCACGTTAGCACTTAGCT
GGCGC
GCGCCAGCTAAGTGCTAACGTGATAATCCTCTCTGCTTG
CAGTCGACCGACTCTGCTCGACTAGATCTC Se GCTCAAGCTTTTAGAGCAGGTACAATAGCAGG CTGTCGACCGACTCTGCTCGACTAGATCTC Sf GCAAGCTTGAGCAGGTACAATAGTGGACTA CTGTCGACCGACTCTGCTCGACTAGATCTC PigmR-RNAi ATGGTACCGGACTAGTTGGAAAATAATGG
AGGAC
CTGGATCCGTGAGCTCGCAACCAATCTCACGACTG
PigmRS-RNAi TTGGTACCGGACTAGTGAGGATGTTACGG
GTCTTG
GCTGGATCCGTGAGCTCTACTACTGGTCCGCTTG
OsAGO4aRNAi ATTGGTACCGGACTAGTGAGTTCACGGTA
GTTCTGGA
CTGGATCCGTGAGCTCCAGTATTTGCTGGACTTGTT

45
OsDCL3aRNAi ATTGGTACCGGACTAGTTGTGGACTTCTTGAGTTGGATG GCTGGATCCGTGAGCTCAGTAGTGGCTGTGAGGGGTG OsRDR2RNAi ATTGGTACCGGACTAGTTCAGGGCACTTGATTTTACAC GCTGGATCCGTGAGCTCAGCAAAACATTATGGCTTCTC CLuc-PigmR ATCAACTAGTATGGCGGAGACGGTGCTGAG Luciferase CCGTGTCGACTCCGCCAGCTTGAGCTGTG complementation PigmR-NLuc ATCAACTAGTATGGCGGAGACGGTGCTGAG CCGTGTCGACTCCGCCAGCTTGAGCTGTG CLuc-PigmS ATCAACTAGTATGGCGGAGACGGTGCTGAG CCGTGTCGACTCCGCCAGCTTGAGCTGTG PigmS-NLuc ATCAACTAGTATGGCGGAGACGGTGCTGAG CCGTGTCGACTCCGCCAGCTTGAGCTGTG PigmR-CC-HA ATCAACTAGTATGGCGGAGACGGTGCTGAG Constructs for co-IP CGTGTCGACTTTGGCCGGACCATCATTAGC and pull down PigmR-CC-FLAG ATCAACTAGTATGGCGGAGACGGTGCTGAG CGTGTCGACTTTGGCCGGACCATCATTAGC PigmS-CC-HA ATCAACTAGTATGGCGGAGACGGTGCTGAG CGTGTCGACTTTGGCCGGACCATCATTAGC PigmS-CC-FLAG ATCAACTAGTATGGCGGAGACGGTGCTGAG CGTGTCGACTTTGGCCGGACCATCATTAGC PigmR-CC-His GTGGTCGACCTTGGCCGGACCATCATTAGC CGTGGTCGACATGGCGGAGACGGTGCTGAG PigmR-CC-GST ACCACTAGTCTTGGCCGGACCATCATTAGC TGGTCGACTGATGGCGGAGACGGTGCTGAG PigmS-CC-MBP GGCGGCCGCCTTGGCCGGACCATCATTAGC TGGCGGCCGCATGGCGGAGACGGTGCTGAG GUS-HA CACTAGTATGTTACGTCCTGTAGAAACC GCGTCGACTCCTTGTTTGCCTCCCTGCTG PigmR-CDS CACCATGGCGGAGACGGTGCTGAG Constructs for TCAGCCAGCTTGAGCTGTG yeast-two-hybrid PigmR-CC CACCATGGCGGAGACGGTGCTGAG TCATTTGGCCGGACCATCATTAGC PigmR-NBS CACCATGATCTGTGTTGTTGGGATG TCAGCAGTTTGTCCAGATCAATTG PigmR-LRR CACCATGAGGATGTTACGGGTCTTGGA TCAGCCAGCTTGAGCTGTG PigmS-CDS CACCATGGCGGAGACGGTGCTGAG TCAGCCAGCTTGAGCTGTG PigmS-CC CACCATGGCGGAGACGGTGCTGAG TCATTTGGCCGGACCATCATTAGC PigmS-NBS CACCATGATCTGTGTTGTTGGGATG

46
TCAGCAGTTTGTCCAGATCAATTG PigmS-LRR CACCATGAGGATGTTACGGGTCTTGGA TCAGCCAGCTTGAGCTGTG Pish-CDS CACCATGGCGGAGGTGGTGTTGGCTGGC TCATCTGAATTCCTTCCAGCGG Pish-CC CACCATGGCGGAGGTGGTGTTGGCTGGC TCAGGGACCCTCAAGACTATTACC Pish-NBS CACCATGAGTGTACCAACAATTGTTGTTC TCATAGCTGATTAAAAATATCATCTC Pish-LRR CACCATGTTGAAGAATCTGAAGAAGCTAC TCATCTGAATTCCTTCCAGCGG a (full PigmS GCAAGCTTTTATCTGGCCCAATTTTATATTG Constructs for promoter) CATGGATCCCGACTCTGCTCGACTAGATCTC GUS reporters b GTCAAGCTTTTATCTGGCCCAATTTTATATTG AGAGGATTATCATTATAAGAGCAAGTTCAATAGTA TTGAACTTGCTCTTATAATGATAATCCTCTCTGCT CAGGATCCCGACTCTGCTCGACTAGATCTC c GTCAAGCTTTTATCTGGCCCAATTTTATATTG TCCACTATTGTACCTGCTCCAATATTGTCCATCTGTAC GTACAGATGGACAATATTGGAGCAGGTACAATAGTGGA CAGGATCCCGACTCTGCTCGACTAGATCTC d GTCAAGCTTTTATCTGGCCCAATTTTATATT AAGCAGAGAGGATTATCACGTTAGCACTTAGCTGGCGC GCGCCAGCTAAGTGCTAACGTGATAATCCTCTCTGCTT CAGGATCCCGACTCTGCTCGACTAGATCTC e GCTCAAGCTTTTAGAGCAGGTACAATAGCAGG CTGGATCCCGACTCTGCTCGACTAGATCTC f GTCAAGCTTGAGCAGGTACAATAGTGGACT CATGGATCCCGACTCTGCTCGACTAGATCTC PigmR:Myc ACGCGTGGTCGACATGGCGGAGACGGTGCTGAG Constructs for TCCAGACCACTAGTTCCGCCAGCTTGAGCTGTGCC cell death assay PigmS:YFP ACGCGTGGTCGACATGGCGGAGACGGTGCTGAG TCCAGACCACTAGTTCCGCCAGCTTGAGCTGTGCC pigmsI41E:YFP GAAAGACATCTGGTATGAGAAAGATGAGCTAAAAAC GTTTTTAGCTCATCTTTCTCATACCAGATGTCTTTC pigmsL45E:YFP GTATATCAAAGATGAGGAGAAAACGATGCAAGCATT AATGCTTGCATCGTTTTCTCCTCATCTTTGATATAC pigmsM48E:YFP GATGAGCTAAAAACGGAGCAAGCATTCCTTAGAGC GCTCTAAGGAATGCTTGCTCCGTTTTTAGCTCATC MgPot2 ACGACCCGTCTTTACTTATTTGG Analysis of blast AAGTAGCGTTGGTTTTGTTGGAT infection

47
OsUbq GACGGACGCACCCTGGCTGACTAC TGCCAATTACCATATACCACGAC M12 GCCAGCTAAGTGCTAACGTTAG MITE sRNA gel CGACTCTGCTCGACTAGATCTC blot assay
Table S7. Primers used for plasmid construction , PCR detection and site mutation.

48
Author Contribution
Y.D., Y.Y., E.W., H.X., D.T., and Z.H. designed experiments; Y.D., K.Z., Z.X., X.D.,
D.Y, J.L., X.W., P.Q., G. Z., Q.L., J.Z., B.M. D.T., Z.H. performed experiments and data
analysis. Y.D., E.W., and Z.H. wrote the manuscript. All authors have read, edited and
approved the content of the manuscript.

References and Notes
1. J. L. Dangl, D. M. Horvath, B. J. Staskawicz, Pivoting the plant immune system from dissection to deployment. Science 341, 746–751 (2013). doi:10.1126/science.1236011 Medline
2. S. Fukuoka, N. Saka, H. Koga, K. Ono, T. Shimizu, K. Ebana, N. Hayashi, A. Takahashi, H. Hirochika, K. Okuno, M. Yano, Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 325, 998–1001 (2009). doi:10.1126/science.1175550 Medline
3. N. Zhang, J. Luo, A. Y. Rossman, T. Aoki, I. Chuma, P. W. Crous, R. Dean, R. P. de Vries, N. Donofrio, K. D. Hyde, M.-H. Lebrun, N. J. Talbot, D. Tharreau, Y. Tosa, B. Valent, Z. Wang, J.-R. Xu, Generic names in Magnaporthales. IMA Fungus 7, 155–159 (2016). doi:10.5598/imafungus.2016.07.01.09 Medline
4. Y. Deng, X. Zhu, Y. Shen, Z. He, Genetic characterization and fine mapping of the blast resistance locus Pigm(t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety. Theor. Appl. Genet. 113, 705–713 (2006). doi:10.1007/s00122-006-0338-7 Medline
5. B. Zhou, S. Qu, G. Liu, M. Dolan, H. Sakai, G. Lu, M. Bellizzi, G.-L. Wang, The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol. Plant Microbe Interact. 19, 1216–1228 (2006). doi:10.1094/MPMI-19-1216 Medline
6. S. Qu, G. Liu, B. Zhou, M. Bellizzi, L. Zeng, L. Dai, B. Han, G. L. Wang, The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site–leucine-rich repeat protein and is a member of a multigene family in rice. Genetics 172, 1901–1914 (2006). doi:10.1534/genetics.105.044891 Medline
7. D. Saleh, J. Milazzo, H. Adreit, E. Fournier, D. Tharreau, South-East Asia is the center of origin, diversity and dispersion of the rice blast fungus, Magnaporthe oryzae. New Phytol. 201, 1440–1456 (2014). doi:10.1111/nph.12627 Medline
8. R. Gallet, C. Fontaine, F. Bonnot, J. Milazzo, C. Tertois, H. Adreit, V. Ravigné, E. Fournier, D. Tharreau, Evolution of compatibility range in the rice-Magnaporthe oryzae system: An uneven distribution of R genes between rice subspecies. Phytopathology 106, 348–354 (2016). doi:10.1094/PHYTO-07-15-0169-R Medline
9. T. Maekawa, W. Cheng, L. N. Spiridon, A. Töller, E. Lukasik, Y. Saijo, P. Liu, Q.-H. Shen, M. A. Micluta, I. E. Somssich, F. L. W. Takken, A.-J. Petrescu, J. Chai, P. Schulze-Lefert, Coiled-coil domain-dependent homodimerization of intracellular barley immune receptors defines a minimal functional module for triggering cell death. Cell Host Microbe 9, 187–199 (2011). doi:10.1016/j.chom.2011.02.008 Medline

10. M. Bernoux, T. Ve, S. Williams, C. Warren, D. Hatters, E. Valkov, X. Zhang, J. G. Ellis, B. Kobe, P. N. Dodds, Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation. Cell Host Microbe 9, 200–211 (2011). doi:10.1016/j.chom.2011.02.009 Medline
11. A. Takahashi, N. Hayashi, A. Miyao, H. Hirochika, Unique features of the rice blast resistance Pish locus revealed by large scale retrotransposon-tagging. BMC Plant Biol. 10, 175 (2010). doi:10.1186/1471-2229-10-175 Medline
12. K. Kikuchi, K. Terauchi, M. Wada, H. Y. Hirano, The plant MITE mPing is mobilized in anther culture. Nature 421, 167–170 (2003). doi:10.1038/nature01218 Medline
13. R. K. Slotkin, M. Vaughn, F. Borges, M. Tanurdzić, J. D. Becker, J. A. Feijó, R. A. Martienssen, Epigenetic reprogramming and small RNA silencing of transposable elements in pollen. Cell 136, 461–472 (2009). doi:10.1016/j.cell.2008.12.038 Medline
14. J. P. Calarco, F. Borges, M. T. A. Donoghue, F. Van Ex, P. E. Jullien, T. Lopes, R. Gardner, F. Berger, J. A. Feijó, J. D. Becker, R. A. Martienssen, Reprogramming of DNA methylation in pollen guides epigenetic inheritance via small RNA. Cell 151, 194–205 (2012). doi:10.1016/j.cell.2012.09.001 Medline
15. X. Cui, X. Cao, Epigenetic regulation and functional exaptation of transposable elements in higher plants. Curr. Opin. Plant Biol. 21, 83–88 (2014). doi:10.1016/j.pbi.2014.07.001 Medline
16. L. Wu, H. Zhou, Q. Zhang, J. Zhang, F. Ni, C. Liu, Y. Qi, DNA methylation mediated by a microRNA pathway. Mol. Cell 38, 465–475 (2010). doi:10.1016/j.molcel.2010.03.008 Medline
17. L. Wei, L. Gu, X. Song, X. Cui, Z. Lu, M. Zhou, L. Wang, F. Hu, J. Zhai, B. C. Meyers, X. Cao, Dicer-like 3 produces transposable element-associated 24-nt siRNAs that control agricultural traits in rice. Proc. Natl. Acad. Sci. U.S.A. 111, 3877–3882 (2014). doi:10.1073/pnas.1318131111 Medline
18. R. Büschges, K. Hollricher, R. Panstruga, G. Simons, M. Wolter, A. Frijters, R. van Daelen, T. van der Lee, P. Diergaarde, J. Groenendijk, S. Töpsch, P. Vos, F. Salamini, P. Schulze-Lefert, The barley Mlo gene: A novel control element of plant pathogen resistance. Cell 88, 695–705 (1997). doi:10.1016/S0092-8674(00)81912-1 Medline
19. J. K. Brown, A cost of disease resistance: Paradigm or peculiarity? Trends Genet. 19, 667–671 (2003). doi:10.1016/j.tig.2003.10.008 Medline
20. D. Tian, M. B. Traw, J. Q. Chen, M. Kreitman, J. Bergelson, Fitness costs of R-gene-mediated resistance in Arabidopsis thaliana. Nature 423, 74–77 (2003). doi:10.1038/nature01588 Medline

21. N. Denancé, A. Sánchez-Vallet, D. Goffner, A. Molina, Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Front. Plant Sci. 4, 155 (2013). doi:10.3389/fpls.2013.00155 Medline
22. R. H. Dowen, M. Pelizzola, R. J. Schmitz, R. Lister, J. M. Dowen, J. R. Nery, J. E. Dixon, J. R. Ecker, Widespread dynamic DNA methylation in response to biotic stress. Proc. Natl. Acad. Sci. U.S.A. 109, E2183–E2191 (2012). doi:10.1073/pnas.1209329109 Medline
23. H. Ito, H. Gaubert, E. Bucher, M. Mirouze, I. Vaillant, J. Paszkowski, An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature 472, 115–119 (2011). doi:10.1038/nature09861 Medline
24. L. Q. Wei, W. Y. Xu, Z. Y. Deng, Z. Su, Y. Xue, T. Wang, Genome-scale analysis and comparison of gene expression profiles in developing and germinated pollen in Oryza sativa. BMC Genomics 11, 338 (2010). doi:10.1186/1471-2164-11-338 Medline
25. S. V. K. Jagadish, R. Muthurajan, R. Oane, T. R. Wheeler, S. Heuer, J. Bennett, P. Q. Craufurd, Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J. Exp. Bot. 61, 143–156 (2010). doi:10.1093/jxb/erp289 Medline
26. S. Amien, I. Kliwer, M. L. Márton, T. Debener, D. Geiger, D. Becker, T. Dresselhaus, Defensin-like ZmES4 mediates pollen tube burst in maize via opening of the potassium channel KZM1. PLOS Biol. 8, e1000388 (2010). doi:10.1371/journal.pbio.1000388 Medline
27. J. M. Bonman, T. I. V. Dedios, M. M. Khin, Physiologic specialization of Pyricularia oryzae in the Philippines. Plant Dis. 70, 767–769 (1986). doi:10.1094/PD-70-767
28. Y. Kawano, A. Akamatsu, K. Hayashi, Y. Housen, J. Okuda, A. Yao, A. Nakashima, H. Takahashi, H. Yoshida, H. L. Wong, T. Kawasaki, K. Shimamoto, Activation of a Rac GTPase by the NLR family disease resistance protein Pit plays a critical role in rice innate immunity. Cell Host Microbe 7, 362–375 (2010). doi:10.1016/j.chom.2010.04.010 Medline
29. D. L. Yang, Q. Li, Y.-W. Deng, Y.-G. Lou, M.-Y. Wang, G.-X. Zhou, Y.-Y. Zhang, Z.-H. He, Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance. Mol. Plant 1, 528–537 (2008). doi:10.1093/mp/ssn021 Medline
30. D. Park, R. J. Sayler, Y. Hong, M. Nam, Y. Yang, A method for inoculation and evaluation of rice sheath blight disease. Plant Dis. 92, 25–29 (2008). doi:10.1094/PDIS-92-1-0025
31. H. Tsunematsu, M. J. T. Yanoria, L. A. Ebron, N. Hayashi, I. Ando, H. Kato, T. Imbe, G. S. Khush, Development of monogenic lines of rice for blast resistance. Breed. Sci. 50, 229–234 (2000). doi:10.1270/jsbbs.50.229

32. International Rice Research Institute (IRRI), Standard Evaluation System for Rice (SES) (IRRI, 2002), pp. 17–18.
33. K. Asaga, A procedure for evaluating field resistance to blast in rice varieties. J. Cent. Agric. Exp. Stn. 35, 51–138 (1981).
34. K. Farrar, I. S. Donnison, Construction and screening of BAC libraries made from Brachypodium genomic DNA. Nat. Protoc. 2, 1661–1674 (2007). doi:10.1038/nprot.2007.204 Medline
35. R. A. Jefferson, The GUS reporter gene system. Nature 342, 837–838 (1989). doi:10.1038/342837a0 Medline
36. H. Chen, Y. Zou, Y. Shang, H. Lin, Y. Wang, R. Cai, X. Tang, J.-M. Zhou, Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 146, 368–376 (2008). doi:10.1104/pp.107.111740 Medline
37. L. Liu, Y. Zhang, S. Tang, Q. Zhao, Z. Zhang, H. Zhang, L. Dong, H. Guo, Q. Xie, An efficient system to detect protein ubiquitination by agroinfiltration in Nicotiana benthamiana. Plant J. 61, 893–903 (2010). doi:10.1111/j.1365-313X.2009.04109.x Medline
38. S. Bai, J. Liu, C. Chang, L. Zhang, T. Maekawa, Q. Wang, W. Xiao, Y. Liu, J. Chai, F. L. W. Takken, P. Schulze-Lefert, Q.-H. Shen, Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLOS Pathog. 8, e1002752 (2012). doi:10.1371/journal.ppat.1002752 Medline
39. L. Pan, S. Wang, T. Lu, C. Weng, X. Song, J. K. Park, J. Sun, Z. H. Yang, J. Yu, H. Tang, D. M. McKearin, D. A. Chamovitz, J. Ni, T. Xie, Protein competition switches the function of COP9 from self-renewal to differentiation. Nature 514, 233–236 (2014). Medline
40. E. Gruntman, Y. Qi, R. K. Slotkin, T. Roeder, R. A. Martienssen, R. Sachidanandam, Kismeth: Analyzer of plant methylation states through bisulfite sequencing. BMC Bioinformatics 9, 371 (2008). doi:10.1186/1471-2105-9-371 Medline
41. B. Langmead, C. Trapnell, M. Pop, S. L. Salzberg, Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, R25 (2009). doi:10.1186/gb-2009-10-3-r25 Medline
42. S. H. Zhong, J.-Z. Liu, H. Jin, L. Lin, Q. Li, Y. Chen, Y.-X. Yuan, Z.-Y. Wang, H. Huang, Y.-J. Qi, X.-Y. Chen, H. Vaucheret, J. Chory, J. Li, Z.-H. He, Warm temperatures induce transgenerational epigenetic release of RNA silencing by inhibiting siRNA biogenesis in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 110, 9171–9176 (2013). doi:10.1073/pnas.1219655110 Medline


















