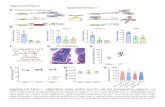
Supplementary Figure 1. Analysis of accumulation and ......Jul 08, 2013 · Supplemental Data....
17
Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118. 1 Supplementary Figure 1. Analysis of accumulation and topology of CURT1 proteins. (A) Accumulation of CURT1 proteins in different ecotypes of A. thaliana. Total protein extracts corresponding to 3 μg (CURT1A, B and C) or 10 μg (CURT1D) of Chl from three different A. thaliana ecotypes (Col-0, Ler, and No-0) matching the background of the mutant lines used in this study were fractionated by SDS-PAGE, and blots were probed with antibodies raised against CURT1A, B, C and D. The Ponceau Red (P.R.) stained protein blot served as loading control. Note that for CURT1D no signal at the expected size of 15.7 kDa was obtained; the band at >16 kDa represents an unspecific signal. (B) Overview of tagged CURT1A variants used in this study. The CURT1A cDNA was inserted into binary vectors between the 35S promoter and the sequences coding for the HA or cmyc tags and introduced into curt1a-2 plants. In the left panel, transgene expression analysis was carried out on total protein extracts from WT and T3 plants homozygous for the transgenes. CURT1A is overexpressed in all four lines tested and corresponding lines are designated in the following as “oeCURT1A-HA” or “oeCURT1A-cmyc”. In the middle panel, thylakoid proteins from T4 plants expressing CURT1A-HA were analysed by immunoblot analysis. These lines express CURT1A at lower levels than WT and are designated as “CURT1A-HA”. In the right panel, expression of oeCURT1A-cmyc in curt1a-2 plants and WT (T3 generation) is shown. (C) Schematic representation of the two possible topologies of CURT1 proteins, with the two TMs indicated by black boxes, the regions used to raise antibodies by grey boxes and the trypsin cleavage sites by asterisks. Note that the regions used as epitopes for the CURT1A- and B-specific antibodies are predicted to contain trypsin cleavage sites. (D) Immunoblot analysis of thylakoid membrane preparations from WT (Ler), oeCURT1D-HA and CURT1A-HA curt1a-2 plants using antisera that recognise N-terminal epitopes of CURT1A and B, HA or PsbO (as a control for luminal thylakoid proteins). Samples were prepared before (-Trypsin) and after (+Trypsin) treatment with trypsin. In intact thylakoids, only stroma-exposed polypeptides are accessible to the enzyme. The behaviour of LHCII,
Transcript of Supplementary Figure 1. Analysis of accumulation and ......Jul 08, 2013 · Supplemental Data....

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
1
Supplementary Figure 1. Analysis of accumulation and topology of CURT1 proteins. (A) Accumulation of CURT1 proteins in different ecotypes of A. thaliana. Total protein extracts corresponding to 3 µg (CURT1A, B and C) or 10 µg (CURT1D) of Chl from three different A. thaliana ecotypes (Col-0, Ler, and No-0) matching the background of the mutant lines used in this study were fractionated by SDS-PAGE, and blots were probed with antibodies raised against CURT1A, B, C and D. The Ponceau Red (P.R.) stained protein blot served as loading control. Note that for CURT1D no signal at the expected size of 15.7 kDa was obtained; the band at >16 kDa represents an unspecific signal. (B) Overview of tagged CURT1A variants used in this study. The CURT1A cDNA was inserted into binary vectors between the 35S promoter and the sequences coding for the HA or cmyc tags and introduced into curt1a-2 plants. In the left panel, transgene expression analysis was carried out on total protein extracts from WT and T3 plants homozygous for the transgenes. CURT1A is overexpressed in all four lines tested and corresponding lines are designated in the following as “oeCURT1A-HA” or “oeCURT1A-cmyc”. In the middle panel, thylakoid proteins from T4 plants expressing CURT1A-HA were analysed by immunoblot analysis. These lines express CURT1A at lower levels than WT and are designated as “CURT1A-HA”. In the right panel, expression of oeCURT1A-cmyc in curt1a-2 plants and WT (T3 generation) is shown. (C) Schematic representation of the two possible topologies of CURT1 proteins, with the two TMs indicated by black boxes, the regions used to raise antibodies by grey boxes and the trypsin cleavage sites by asterisks. Note that the regions used as epitopes for the CURT1A- and B-specific antibodies are predicted to contain trypsin cleavage sites. (D) Immunoblot analysis of thylakoid membrane preparations from WT (Ler), oeCURT1D-HA and CURT1A-HA curt1a-2 plants using antisera that recognise N-terminal epitopes of CURT1A and B, HA or PsbO (as a control for luminal thylakoid proteins). Samples were prepared before (-Trypsin) and after (+Trypsin) treatment with trypsin. In intact thylakoids, only stroma-exposed polypeptides are accessible to the enzyme. The behaviour of LHCII,

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
2
visualised by Ponceau Red (P.R) staining, served as control for trypsin activity. If the N and C termini of the proteins faced the stroma (Topology 1 in panel C), tryptic digestion should cleave the segments of the proteins against which the antibodies were raised. Tryptic cleavage of oppositely oriented CURT1 proteins (with the N- and C-termini facing the lumen; Topology 2 in panel B) at one site within the inter-TM region is expected to result in fragments between 6.3 and 8.6 kDa detectable by the antibodies specific for the HA-tag, CURT1A or B. As expected, PsbO on the lumenal side of the thylakoid membrane was not affected by trypsin, whereas Lhcb1 was fragmented due to cleavage of its stroma-exposed regions. The C-termini of CURT1A-HA and CURT1D-HA were completely digested, leaving almost no proteolytic fragment that could be detected by the HA antibody. Similarly, most of the CURT1B protein was digested without generating detectable fragments. CURT1A was only degraded to a minor degree, indicating that the N-terminus of the protein might be protected from trypsin through interaction with other proteins. The absence of fragments between 6.3 and 8.6 kDa clearly implies that CURT1 proteins adopt Topology 1. Moreover, the incomplete digestion of CURT1 proteins by trypsin indicates that they are protected, most likely because they form oligomers (see Figure 3). Further support for the suggested CURT1 topology is the prerequisite of a stromal orientation of the antibody-binding domains for efficient Co-immunoprecipitation (see Figure 3E) and immunogold-labelling (see Figure 7).

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
3
Supplemental Figure 2. Mutations of the CURT1 genes. (A) Insertion lines. Both curt1a alleles (curt1a-1, SGT_3_4785; curt1a-2, GT_5_19630) are in the Landsberg erecta (Ler) background, carry insertions of the Dissociation (Ds) transposon and were obtained from the IMA (Parinov et al., 1999) and Exotic (Sundaresan et al., 1995) collections, respectively. The curt1b-1 allele (15-2546-1) is from the RIKEN Ds collection (Ito et al., 2002) (http://rarge.psc.riken.jp/dsmutant/index.pl) and was induced in the Noessen (No-0) background, whereas curt1b-2 (FLAG_218A11) originates from the FLAGdb/FST T-DNA collection (Samson et al., 2002) and is in the genetic background of the ecotype Wassilewskija (WS). In curt1c-1 (SALK_052057) from the SALK T-DNA collection (Alonso et al., 2003) (http://signal.salk.edu; genetic background Col-0), the gene is disrupted by insertion of the ROK2 T-DNA. The curt1d-1 mutant (SAIL_1240_C05) carries an insertion of the pCSA110 T-DNA and originates from the Syngenta SAIL collection (Sessions et al., 2002) (Col-0 background). The translated parts of exons (boxes), as well as intron sequences (lines), are depicted. For each insertion mutant used in this study, the site and the orientation of the insertion are indicated. The T-DNA and transposon insertions are not drawn to scale. The presence of gene insertions was confirmed by PCR with gene- and insertion-specific primers (Supplemental Table 4 online) and sequencing of the PCR products. Loss of expression of the gene products was confirmed by RT-PCR or Northern analysis, as well as by immunoblot analysis (see Figure 3). (B) For CURT1A, C and D, RT-PCR analysis was performed on transcripts from WT and mutant plants with gene-specific primers located 5’ (1) or 3’ (2) to the insertion and ACTIN1 primers as positive control. Because no suitable primer pair could be designed for one of the two insertions in CURT1B, Northern analysis was performed. Aliquots (30 µg) of total leaf RNA were fractionated on a denaturing agarose gel, transferred to a positively charged nylon membrane and hybridised with a CURT1B cDNA probe. Ribosomal RNA was stained with Methylene Blue (M.B.) as a loading control.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
4
Supplemental Figure 3. Analysis of CURT1 protein abundance and oligomerisation. (A) Relative abundance of CURT1 proteins in curt1 mutants as determined in Figure 3A and Supplemental Figures 3B-D. Protein abundance relative to WT is indicated on the grey scale,

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
5
ranging from 0% (white) to 100% (black). CURT1D abundance is set to 0% in mutants containing the curt1d-1 mutation and to 100% in WT. (B-D) Total protein extracts from Col-0 (1x corresponds to 3 µg of Chl) or curt1 mutant plants were fractionated by SDS-PAGE, and blots were probed with antibodies raised against CURT1A, B and C. The Ponceau Red (P.R.)-stained protein blot served as loading control. Note that Ler and No-0 behaved like Col-0 (see Supplemental Figure 1A online). (E) Quantification of MBP fusions of PetC and PsaD as controls for Figure 3B. The calculated concentration of the respective protein in thylakoid preparations is given below each panel. (F) Blue-Native (BN) polyacryamide gel electrophoresis (PAGE) analysis of thylakoid multiprotein complexes. Left: WT (Ler) and mutant (curt1a-1) thylakoids were solubilised with 1.6 % (w/v) digitonin and then fractionated by BN-PAGE. The bands were assigned to the PSI-LHCI-LHCII supercomplex (PSI-LHCII), PSI-LHCI monomers (PSImono) and PSII dimers (PSIIdi), dimeric Cytochrome b6/f complexes (Cyt b6/fdi) trimeric (LHCIItri) and monomeric (LHCIImono) LHCII (Pesaresi et al., 2009; Armbruster et al., 2010). Right: After immunoblotting, the blots were immunodecorated with CURT1A-specific antibodies. The circled Arabic numbers indicate the different CURT1A-containing complexes. The strong signal between bands 5 and 6 derives from chemiluminescence of the hemes contained in the Cyt b6/f complex. (G) Immunoblots of thylakoid proteins from curt1a-1, curt1b-1 and curt1c-1 separated by 2D gel electrophoresis were analysed as in Figure 3C. The positions of the major thylakoid multi-protein-pigment complexes are indicated by Roman numerals as in Figure 3C. (H) As a loading control for Figure 3D, blots with crosslinked products were stained with Ponceau Red (P.R.). (I) Identification of interactors of CURT1B by CoIP. Thylakoid membranes from WT (No-0) and curt1b-1 mutants were co-incubated with a CURT1B-specific antibody. Complexes were solubilised with 1.6% (w/v) digitonin and precipitated with Protein-A agarose before centrifugation. The pellet was repeatedly washed and bound proteins were eluted by adding SDS-PAGE loading dye. Thylakoid membrane (T) and immunoprecipitated (P) proteins were analysed by SDS-PAGE and Western analysis employing antibodies specific for CURT1B, CURT1A and PsaF. (J) CoIP analysis of CURT1B was performed as in Figure 3E, except that the immunoprecipitated proteins were also analysed with a CURT1C-specific antibody. (K) CoIP analysis was carried out with antibodies against cmyc on thylakoid proteins from WT (Ler) or plants overexpressing CURT1A-cmyc either in the curt1a-2 or the WT background (see Supplemental Figure 1 online). Immunoprecipitated proteins were detected with an antibody raised against CURT1A.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
6
Supplemental Figure 4. CURT1 proteins are not constitutive components of the major thylakoid photosynthesis complexes and their lack affects photosynthesis pleiotropically. (A) Blue Native (BN) polyacryamide gel electrophoresis (PAGE) analysis of thylakoid multiprotein complexes. Thylakoids corresponding to 30 µg Chl from WT (Ler) and curt1 mutant plants were solubilised with 1.0 % (w/v) dodecyl--D-maltoside (-DM). The extracts

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
7
were then fractionated by BN-PAGE. The bands detected were identified as specific protein complexes in accordance with previously published profiles (Granvogl et al., 2006; Schwenkert et al., 2006; Peng et al., 2008; Armbruster et al., 2010): PSI-NDH supercomplex (PSI-NDH), PSII supercomplexes (PSIIsuper), PSI monomers and PSII dimers (PSImono and PSIIdi), PSII monomers and dimeric Cyt b6/f (PSIImono and Cyt b6/fdi), PSII monomers w/o CP43 (CP43-PSII), multimeric (LHCIImult), trimeric (LHCIItri) and monomeric (LHCIImono) LHCII. (B) CURT1B is not a subunit of PSI. Immunoblot analysis of PSI preparations with a CURT1B-specific antibody. PSI was isolated from WT thylakoids by sucrose-gradient centrifugation (DalCorso et al., 2008). Thylakoid and PSI proteins corresponding to 10 µg Chl were separated by SDS-PAGE and either stained with Coomassie Blue (lower panel) or subjected to immunoblot analysis. Positions of PSI subunits previously identified by immunodetection are indicated. (C) Immunoblot analysis of thylakoid proteins (5 µg Chl) from WT (No-0) and mutant (psal-1, psao-1, curt1a-1 and curt1b-1) plants. Antibodies directed against CURT1A, CURT1B, PsaL, PsaO or Lhca3, as well as an antibody (“-PsaP”) previously raised against a peptide from CURT1B (previously denoted TMP14 and PsaP)(Khrouchtchova et al., 2005), were used. Clearly, the “-PsaP” antibody does not recognise CURT1B but crossreacts with PsaL even though the two proteins have no apparent similarity. This led to the erroneous conclusion that CURT1B was a PSI subunit (PsaP). (D-G) Average values ± SD (bars) for three to five different plants are indicated by filled (WT) or open (curt1abcd) circles. (D) Dependence of the relative electron transport rate (ETR) on light intensity. ETR is expressed relative to the maximum ETR in the WT (100%). (E) Kinetics of re-reduction of P700
+ following a short saturating light pulse (7000 mol photons m-2s-1) that led to the complete oxidation of P700.
(F) P700 oxidation and re-reduction kinetics during a 250-ms saturating light pulse. (G) Time courses of induction and relaxation of NPQ monitored during dark-to-light (80 mol photons m-2s-1, white bar) transitions. The 4-min light period (white bar) was followed by a 2-min dark period (black bar).

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
8
Supplemental Figure 5. Quantification of thylakoid phosphorylation in curt1 mutants. (A) Thylakoid proteins were extracted from WT (Col-0) and curt1 mutant (curt1a-2 and curt1abcd) plants that had been kept for 16 h in the dark (D), subsequently exposed to low light (LL) for 2 h, and then to high light (HL) for 2 h. After fractionation by SDS-PAGE, phosphorylation of thylakoid proteins was detected by immunoblot analysis with a phosphothreonine-specific antibody. pCP43, phosphorylated CP43; pD1/D2, phosphorylated D1/D2; pLHCII, phosphorylated LHCII. The lower panels show portions of the Ponceau S-stained PA gels as loading control; the position of LHCII is indicated. (B) Total signals for pCP43, pD1+pD2 and pLHCII and all four phosphoproteins together in panel c and two repetitions of the experiment were quantified. Average values ± SD are provided. The total of the signals measured from the three genotypes for a given phosphoprotein and light condition was set to 100%.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
9
....|....| ....|....| ....|....| ....|....| ....|....| ....|....| 5 15 25 35 45 55 Nostoc1 ------ISRQ VSQFLEKLPD YLGSFFQDYK --QPLITVAL ILAAIVTAKL VLALLDAIND Anabeana2 ----QQITNQ VTDFLAELPE YLGSFYQQYS --RSILTVLL ILAALVTLKV VLALLGAING Nodularia1 -----RVARR ITEFLEQLPA YVSGFFQKYQ --RSLINVAL ILTAIITVKV VAAILGAINY Raphidiops ------YGQE VSNFLGTLPE YLVGLFDKYK --QPLLTLGL IVTAGVTVKV ILAVLDSLND Anabaena1 ----RKYGEQ VSDFLATLPD YVGNFFNQYK --QPLVSVGL IVASIVAVKV LLAVLDSLND Arthrospir -------KEK VVVFLSELPG YVSKFFGEYQ --KPIVTILL IVVAVVTIKV LFAVVDALND Oscillator -----QIKDQ VFSVLSELPA YVSNFFQEYQ --KALITIGL IAAGGITIKV TLAVLDSLND Synechoco1 ------AVKP ITDFLSELPE ELGKFFADYK --QPLITVGL IVAALITVKL TFALIGAIND Cyanothece -------FDY VMQILAKVPE YLGEFFSEYK --QPLTIVGL SLLAIITVKI IVAVLGAVDD Microcysti ---------- VMSYLANVPD YIGSFFSGYK --KPLIVLGA ILASFVTVKV TLAVLDAIDD Synecho1 ----QEWLQP VWEVLGKIPQ MTGEFFEDNK --QPLISLGI ILLGIISVKI LIAVLDAIND Populus1 ------SEES -PVDANEIFT DLKEKWDAVE --NKSTVIIY GGGAVVAVWL SSILIGAVNS Populus2 ------SEES -SVDANEVFT DLKEKWDAVE --NKSTVILY GGGAIVFVWL SSIVIGAINS At4g01150 ----ASSEET SSIDTNELIT DLKEKWDGLE --NKSTVLIY GGGAIVAVWL SSIVVGAINS Vitis2 ----ASSDES GPVDAGELFS DLMERWDALE --NKSTVFIY GGGAIVAVWL SSIIVSAINS Vitis1 ----ASSSEE TSLNTEELFT DLKAKWDALE --NKSTVFLY GGGAIVAVWL SSVVVGAVNS Zea3 ----ASSDDS SAASGDELIA DLKAKWDAVE --NKSTVLTY AGGAVVALWL TSVIVGAINS Oryza3 ----SDDTSS SAATGDELIE DLKAKWDAVE --NKSTVLTY AGGAIIALWL SSVIVGAVNS Zea2 --KAASEDTS ASG--DELIE DLKAKWDAVE --DKPTVLLY GGGAVVALWL TSVVVGAINA Zea1 ---AASEDTS ASGG-DELIE DLKAKWDAVE --DKPTVLLY GGGAVVALWL TSVVVGAINA Oryza2 ----ASDDTS TSASGDELVA DLKAKWEAIE --DKPTFLLY SGGAVVALWL TTVVVGAINS Physco3 ----TSDSSD SSAQIQEFVE DLKVKWDQTE --NKTTVAIY AGGALVTLWL SSIIVGAINS Physco2 ----TSDSSD SSAQIQEFVE DLKVKWDQTE --NKTTVAIY AGGALVTLWL SSIIVGAINS Physco1 ----ATAEGD SSAQIQEFID DLKAKWDQTE --NKSTVVIY AGGALVALWF SSTIVGAINS Picea1 ----EDTSTT SSVQTEEVLT NLKEKWDAVE --NKPTVLIY GGGALLALWI SSIIVAAINS Picea2 ----EDTSDS VSKDFDEVVG DLKEKWDSVE --NKSTLLVY GGGALAALWL SATVVSAINS Chlamy1 ----SSSTET TSFDSEKVLK DLQEKWDAVD --NKGAVAAY AAGAVVALWL SSTIVNAINA Volvox1 ----ETTTSA PSFDSEKVLK DLQEKWDAVD --NKGAVAAY AAGAVVALWL SSTIVNAINA Chlorella1 -------TEG SQVDVDALVK DLQEKWDKVE --NKTSVIVY GAGGIVVLWL ASTVVGALNS Micromon1 ----TMAVAD SKINADEVLK TISDKWEDTE --NKSTVITY VAGAAAVVWL SGTVVGAINS Oryza1 ----RSSEES SSDEDDEILS ELKEKWDAIE --NKSSVLFY GGGAIIAVWL SSIVVKAVDS Picea3 ----EESSSD DSQQFDDMLS NLKQKFDSIE --NKSTPLIY GSAALVALWV SATVVDAIDS Oryza5 --DGEVQVQ- GESTSWNVLN QIGVELDSEN --S-YTALVY GTSALVTIWI SSIVVSALDS Zea6 --EEEKEVQ- GEPASWGVLN QIGVELDSDK --S-YSALIY GTSALVAIWI SSIVVSALDS Vitis3 --IEDSQVE- EQTVAFEFLD NLNIKFDSED --P-YSIFLY GTGALTALWF ASAIVGAIDS PPP4L -NSEAPQAED EETQALEFLN --DIKLDSDK --T-YSILLY GSGAIVALYL TSAIVSSLEA Zea5 ELVPSEASSS P--DDGGLDE ILS-KLNIEV --T-PTLILT GFGAFIALWI LSSVVAAVDS Zea4 --VSSEASPS PSTDDGGLDE ILS-KLNIEV --T-PILILT GSGAFVALWI LSSVVSAVDS Oryza4 ---SLVAAVA DSVEDDALSQ ALSSKLDFKE --T-STFVMY GSGAFIAGWI LSAVVSAIDS Chlamy4 ------PSVK LQKQAEEVLK QVQGKWESTD DSEKPAAIAI IVGVIVAQIA IGATIDAVDK Volvox3 ------VAQK LQAQATEALK FVQGKWDSTD NSEKPAALAI ILGVVVAQIA IGATIDAVDR Chlamy3 ------ALEK IQSTANEAWT WVKDKWATTE DSEKPAVVGI IAGVIVAQIA IGATIDVVDR Populus4 ------GESS ESSAPLGIVK SVKNIWDDSE --DRLALVGL GFAALVAIWT SAKLILAIDK Populus5 ------GESS ESSTSLSIVK SVQNIWDDSE --DRLPLVGL GFAALVAVWA SANVITAVDK TMP14L ----ASGESS DSSTDLDVVS TIQNVWDKSE --DRLGLIGL GFAGIVALWA SLNLITAIDK Vitis5 -VAKATGESS ESSTTLSIVK SVQNVWGDPE --DRFALFGL GFAAIVAVWA SANLITAIDN Oryza10 ---KAAQDSS EPS-SGSVVK YVQSSFSSPE --DLFALAGI GFAGIAALWA SINLVEVIDK Oryza9 ---KAAQDSS EPS-SGSVVK YVQSSFSSPE --DLFALAGI GFAGIAALWA SINLVEVIDK Zea9 ----AAQDGS ESSGTGSIVK YVTSSFSTAE --DIFALAGI GFAAVAALWA SVNLIEIIDK Physco6 ----NSTVND SSAAIEDALK SVQEAWEKTD --DKVAIAGL GLAGLVAIWA AAGLINAIDK Physco5 ----ESTVDD TSAAFEDALK SVQEAWEKTD --DKVAIAGL GLAGLVAIWS AAGLINAVDK Physco4 ----EAAVDE TSAAAEDFLQ SLTEAWEKSD --DKPAIVGL GFAGLIGLWA TNGLINAIDK Picea4 ----GETSTD PVKPLTDALK PVQEAWEKTD --DKLAVGGL GFAAVIVLWA STGLIAAIDK Picea5 ----GETSTD PVKPLTDALK PVQEAWEKTD --DKLAVGGL GFAAVIVLWA STGLIAAIDK Ostroco1 ----DDLTDA ARDKFDEVTT TLSEYWEDSD --EKPALVTL GVYGIVGLVA ANGTLRAVDG Micromon2 ----RADSDD LNAKLKEVTD TVSEKWEDTD --DKPAVVTL GVYGLVGLVA ANGVLKSIDG Populus6 -------VAV ASTDLPEIVK TVQEAWDKVE --DKYAVSSV VVAGGVALWG SAGFISAIER Vitisvinif ----EAPAEV GTTEVPEIIK TVQEAWDKVE --DKYAVSSL AAAGFVGLWV STGMVSAIDK TMP14 ----ATTTEA ETTELPEIVK TAQEAWEKVD --DKYAIGSL AFAGVVALWG SAGMISAIDR Zea8 ----TSGGEG ATEEVPEIVK AAQDAWDKVE --DKYAVATI GVAAIVALWT VVGAIKAIDK Oryza8 ----AADGTG SETEVPEVVK AAQDAWAKVE --DKYAVTAI GVAALVGLWT AIGAIKAIDR Oryza7 ----PAAPLA ANAEITEFIN ALKQEWDRIE --DKYAVTTL AVAASLGMWS AGGVVSAIDR Oryza6 ----PAAPLA ANAEITEFIN ALKQEWDRIE --DKYAVTTL AVAASLGMWS AGGVVSAIDR Zea7 ----PAAPQA ANEELTEFVD ALKKEWDRIE --DKYAVTTL AVAATLGMWS AGGVVSAIDR Chlorella2 -------TTE LQDKATEVVE WAQAKWEESD --NKPGLVAT GGAAFFGLYL ISGLVNTVDR ....|....| ....|....| ....|....| ....|....| ....|....| .. 65 75 85 95 105 Nostoc1 IPLLSPLFEL VGIGYASWFI FRYLLKASTR QELANEIQSL KNQFVGE--- -- Anabeana2 IPLLAPFFEL IGLSYSTWFT FRYLIKSSTR QELTAEIRTL KSQVF----- -- Nodularia1 IPLLSPSFEL IGIGYFTWFC LRYLTKSETR QELTEKIRLF KQEIVGE--- -- Raphidiops IPLVAPTFEL IGIGYSGWFV YRYLLKASTR EELTSEIDTL KSQVFGQD-- -- Anabaena1 IPLVAPTFEL IGIGYSAWFV YRYLLKASTR QELTHEITTL KSQVV----- -- Arthrospir IPLVAPTFEL IGMGYAAWFV YRYLLKASTR QELVQEVNAL RKQVFGNKS- -- Oscillator IPLLSTTFEL VGMGYTGWFV YRYLLRASNR QELSAEIESF KEQVLGK--- --

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
10
Synechoco1 IPLLAPTFEL IGISYTAWFV YRYLLKASNR DELLAEFNSL KSQVLGKK-- -- Cyanothece IPLLAPLLEM VGLGYSAWFV WRYLWKASNR KELLAEFDAI KNQMFG--DN A- Microcysti IPLLAPILQL VGIVYTGWFV WRYLLKDENR RELLAEIDAL KAQVFGSHNN TL Synecho1 IPLLAPTLQL IGMGYTAWFI WRYLWKAEKR QELASEFGAL KEQIF----- -- Populus1 VPLLPKILEL VGLGYTGWFV YRYLLFKSSR KELATDIESL KKKIAGTE-- -- Populus2 VPLLPKIMEL VGLGYTGWFV YRYLLFKSSR KELATDIEVL KKKIAGTE-- -- At4g01150 VPLLPKVMEL VGLGYTGWFV YRYLLFKSSR KELAEDIESL KKKIA----- -- Vitis2 VPLLPKVMEL VGLGYTGWFV YRYLLFKSSR KELATDIEAL KKKIS----- -- Vitis1 VPLLPKIMEL VGLGYTAWFV YRYILFKSNR KELATDIEAL KKKVT----- -- Zea3 VPLLPKIMEL VGLGYTGWFV YRYLLFKESR KELADDIESL KKKIA----- -- Oryza3 VPLLPKFMEL VGLGYTGWFV YRYLLFKESR KELADDVDSL KKRIA----- -- Zea2 VPLLPKILEL VGLGYTGWFV YRYLLFKESR KELAADIETL KKKIA----- -- Zea1 VPLLPKILEL VGLGYTGWFV YRYLLFKQSR KELVDDIETL KKKIA----- -- Oryza2 VPLLPKILEL VGLGYTGWFV YRYLLFKESR KELATDIETL KKKIA----- -- Physco3 VPLLPKVMEL IGLGYTGWFV YRYLLFKSSR KELVDDIEEL KGKIT----- -- Physco2 VPLLPKVMEL IGLGYTGWFV YRYLLFKSSR KELVDDIEEL KGKIT----- -- Physco1 VPLLPKVMEL IGLGYTGWFV YRYLLFKSSR KELVEDIEEL KGKIT----- -- Picea1 VPLLPKLMEL IGLGYTGWFV YRYLLFKSSR KELATDVEEL KKKIT----- -- Picea2 IPLLPKFMEL IGLGYALWFT YSYLLFKESR KQLAEDVEEL KQKIT----- -- Chlamy1 VPLLPKLMEL VGLGYSAWFT YRYLLFKSSR EELMKDIGDL SKKIS----- -- Volvox1 VPLLPKLMEL VGLGYSAWFT YRYLLFKSSR EELLKDIGEL SKKVT----- -- Chlorella1 IPLLPKAFEL VGLGYSAWFT YRYLLFKSSR EELVEDIESL KKKISGEQ-- -- Micromon1 IPILPKVMEL VGLGYSSWFV YRYVLYKDSR KELLEQFDAL KDKVS----- -- Oryza1 VPVLPNILEL VGLGYSGWFV YRYLLFKENR EELANGFDAL KKRIT----- -- Picea3 VPLLPKVLEF IGFGYTVWFV YRYLLFKGSR EELATAIEKL KQEIM----- -- Oryza5 VPLVPQVMEV VGLGFTVWFT SRYLIFKENR DELITRIGSI KKQIL----- -- Zea6 VPLVPQVMEV VGLGFTIWFT SRYLIFKENR DELFTRVSSI KKQIL----- -- Vitis3 IPIFPKLMEI VGLGYTLWFS ARYLIFKQNR DELAAKIEEL KQQVL----- -- PPP4L IPLFPKLMEV VGLGYTLWFT TRYLLFKRNR EELKTKVSEI KKQVL----- -- Zea5 VPLLPKLLEL IGTAYTIWFI ARYLLFKESR DELFAKFEDL KQRII----- -- Zea4 VPLLPKLLEL VGTGYSIWFT ARHLLFKESR DGMFAKFKDI KERII----- -- Oryza4 IPLFPKILQI VGLGYTIWFS TRYLLFKENR DELFVKVDDL KRKIT----- -- Chlamy4 IPIVNKGLQL IGVAVTGLFT YRYFTDPAER ESVKKSIDAF VKSVT----- -- Volvox3 IPVINKGLQL IGVAVTALFF YRYFTDPSER ESVKKSVDAF VKSVT----- -- Chlamy3 IPIVNKLLQL VGLAVTAVFI YKITTDPEER TTVKSSITGF LKEVT----- -- Populus4 LPVVPSVLEL IGILFSSWFI YRYLLFKPNR EELFQIIKKS VANILGQ--- -- Populus5 LPVVPSALEF VGILYSSWFV YRYLLLKPNR EELFQIIKKS VGDILGQ--- -- TMP14L LPVISSGFEL VGILFSTWFT YRYLLFKPDR QELSKIVKKS VADIL----- -- Vitis5 LPLIPGVFEF IGILYS-WG- -RAVRNHQQV CIGSTGPVKS AARXS----- -- Oryza10 LPVLPLLFEL IGILVAWLFI YQNLLFKPDR EKFLNNIKSS VSRVL----- -- Oryza9 LPVLPLLFEL IGILVAWLFI YQNLLFKPDR EKFLNNIKSS VSRVL----- -- Zea9 LPVLPLLFEL VGILVAWLFI YNNLLFKPKR KEFLENIKSS GSRIL----- -- Physco6 LPLIPDVFEL VGIVFSGWFV YRYLLFKPDR EELLKIIDET KSKIT----- -- Physco5 LPLIPDFFEF VGILFSGWFV YRYLLFKPDR EVLFKFIDET MTKIT----- -- Physco4 LPIIPDLFEI IGILFSGWFI YRYLLFKPDR EELLKLIDEQ KAKIT----- -- Picea4 LPLIPSAFEF VGILFSGWFI YRNLLFKPDR EELWTKIQNA ISEIT----- -- Picea5 LPLIPSAFEF VGILFSGWFI YRNLLFKPDR EELWTKIQNA ISEIT----- -- Ostroco1 LPLIPDFLEL VGILFSGFFV YQNLLYKPDR AALRETISKI YNKIL----- -- Micromon2 LPLVPDLLEI VGIGFSAFYV YQNLLFKPDR AALKESISKA LDEIL----- -- Populus6 LPLIPGVLEL AGIGYTGWFA YKNLVFKPDR EALIAKIKDT YKEVIGSS-- -- Vitisvinif LPLVPGVLEI VGIGYSGWFA YKNLIFKPDR EALIQKIKDT YKEII----- -- TMP14 LPLVPGVLEL VGIGYTGWFT YKNLVFKPDR EALFEKVKST YKDIL----- -- Zea8 LPLLPGVLEI VGIGYTGWFT YRNLVFQPDR EALIGKIKST YKEIT----- -- Oryza8 LPLLPGVLEL VGIGYTGWFT YRNLIFQPDR EALVSKIKST YNEIT----- -- Oryza7 LPIVPGLMEA VGIGYSGWFA YRNLLFKPDR EAFFAKVREV YEDII----- -- Oryza6 LPIVPGLMEA VGIGYSGWFA YRNLLFKPDR EAFFAKVREV YEDII----- -- Zea7 LPVVPGLMQA VGIGYSGWFA YRNLLFKPDR DAFFAKVREI YEDII----- -- Chlorella2 LPLIHTGFEL LGLSVTAWFA YRWFFVAGEK EAITSKVTSI TKDIGLDL-- --
Supplemental Figure 6. Alignment of orthologous CURT1 protein sequences for phylogenetic analyses. For the phylogenetic analysis of CURT1 sequences, the C-terminal 99 amino acids of each protein were aligned using ClustalW (v2.0.12; http://mobyle.pasteur.fr/http://mobyle.pasteur.fr/; default settings, gap extension: 1).

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
11
Supplemental Table 1. Proteomics-based data on subcellular localisation and phosphorylation of CURT1 proteins.
total chloroplast thylakoids envelope phosphorylated
R Z K P Fri Fro Fe PhosPhAt 3.0
CURT1A + + + + + + + +
CURT1B + + + + + + + +
CURT1C + + + + + +
CURT1D +
Data on the subcellular localisation of proteins were derived from proteomics studies
indicated in the table: R, (Rutschow et al., 2008); Z, (Zybailov et al., 2008), K,
(Kleffmann et al., 2004); P, (Peltier et al., 2004); Fri, (Friso et al., 2004); Fro,
(Froehlich et al., 2003); Fe, (Ferro et al., 2010). Protein phosphorylation data are
from the PhosPhAt 3.0 database (http://phosphat.mpimp-golm.mpg.de/). Note that
mature CURT1A and B have phosphorylation sites N-terminal to the two TMs and
sites C-terminal to the two TMs are found in CURT1C. Because the vast majority of
thylakoid proteins are phosphorylated in their stroma-exposed regions (Pesaresi et
al., 2011), this argues in favour of a topology of CURT1 proteins in which both N- and
C-termini face the stroma (see Supplemental Figure 1 online).

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
12
Supplemental Table 2. Chlorophyll fluorescence parameters measured in wild-type
and curt1 mutant leaves.
Fv/Fm ΦII NPQ 1-qL
Ler 0.83 ± 0.01 0.74 ± 0.01 0.27 ± 0.01 0.26 ± 0.01
No-0 0.83 ± 0.01 0.74 ± 0.02 0.23 ± 0.04 0.25 ± 0.04
Col-0 0.84 ± 0.01 0.74 ± 0.01 0.24 ± 0.02 0.26 ± 0.02
curt1a-1 0.83 ± 0.01 0.70 ± 0.03 0.19 ± 0.03 0.43 ± 0.03
curt1a-2 0.84 ± 0.01 0.71 ± 0.03 0.19 ± 0.03 0.40 ± 0.03
curt1b-1 0.84 ± 0.01 0.75 ± 0.02 0.23 ± 0.02 0.22 ± 0.02
curt1c-1 0.83 ± 0.01 0.73 ± 0.02 0.25 ± 0.04 0.26 ± 0.04
curt1d-1 0.83 ± 0.01 0.73 ± 0.01 0.23 ± 0.02 0.24 ± 0.02
curt1ab 0.84 ± 0.01 0.65 ± 0.04 0.15 ± 0.04 0.57 ± 0.04
curt1ac 0.83 ± 0.01 0.65 ± 0.05 0.15 ± 0.02 0.48 ± 0.02
curt1abc 0.83 ± 0.01 0.66 ± 0.03 0.16 ± 0.04 0.55 ± 0.05
curt1abd 0.82 ± 0.01 0.65 ± 0.04 0.12 ± 0.03 0.48 ± 0.03
curt1abcd 0.83 ± 0.02 0.64 ± 0.02 0.15 ± 0.01 0.55 ± 0.01
Mean values for five plants ( SD) are shown. Fifth to seventh true leaves were
chosen for measurements. To determine FV/FM, 0.8 s pulses of white light (5000
mol photons m-2 s-1) were used. A 10 min illumination with actinic light (40 mol
photons m-2 s-1) was used to drive electron transport between PSII and PSI before
measuring II, NPQ and 1-qL.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
13
Supplemental Table 3. Fatty acid composition of Arabidopsis leaf lipids.
Fatty Acid Col-0 Ler No-0 curt1a curt1ab curt1abcd
16:0 19.6 ± 1.0 18.5 ± 1.9 17.9 ± 0.5 19.6 ± 0.8 21.5 ± 2.3 20.5 ± 1.9
16:1 3.3 ± 0.1 3.0 ± 0.2 3.0 ± 0.1 3.1 ± 0.2 3.6 ± 0.4 3.2 ± 0.5
16:3 11.8 ± 0.3 13.5 ± 1.6 12.9 ± 1.4 11.8 ± 1.8 10.4 ± 1.4 11.6 ± 1.2
18:0 9.5 ± 1.1 8.2 ± 1.1 8.2 ± 0.4 9.6 ± 0.6 11.0 ± 1.6 11.1 ± 1.9
18:1 2.9 ± 0.1 3.0 ± 0.3 2.9 ± 0.3 3.0 ± 0.3 3.1 ± 0.1 2.9 ± 0.1
18:2 12.6 ± 0.4 10.6 ± 0.3 11.4 ± 1.1 11.0 ± 1.3 11.1 ± 0.2 10.5 ± 0.5
18:3 40.3 ± 1.5 43.4 ± 1.9 43.8 ± 0.8 41.9 ± 1.4 39.4 ± 3.1 40.3 ± 2.5
Mean values (± SD) of three measurements are shown and expressed as mol%.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
14
Supplemental Table 4. Oligonucleotides used for PCR and RT-PCR.
Gene Forward primer (5´3´) Reverse primer (5´3´) experiment
CURT1A CGTCCCGGGATGGCGATATCA GCCAGATCTTTCGCTTCCTGC Fusion to
HA/cmyc
CGGAATTCATGGCTTCTTCAGAAG GCTCTAGACTATTCGCTTCCTGCG Fusion to MBP
CGCCATGGCGATATCAGTAGCAGC CGCCATGGCGCTTCCTGCGATCTTCT Fusion to RFP
AACACCACTGAGTTGGATTTC AACATACAGAGAGAAAGCTGG Genotyping
AAGAAGAAGTTCTCGGCGTG GACGAGGTCTCTTCTGAAGA RT-PCR
GGATGGTCTTGAGAAC GCTTCCTGCGATCTTCTTCT RT-PCR
CTCGAATAGTGAAGCTCCTC CACTCACTATCAGACCCAAG RT-PCR
CURT1B CGGAATTCATGGCCGCTAAAG CCTCTAGACTAGCTGCTCCC Fusion to MBP
ACTCCAAGATCGTCTCTACG CATGCATGGTTAGCTTAGCTT Genotyping
AAAGCCACTGCTTACTGTCGG GCTGCTCCCTAATATGTCTTT RNA blot
analysis
CURT1C CGGAATTCATGGCTAGTGGAGA GCTCTAGATCACTGGCCAAGTA Fusion to MBP
CGCCATGGCTTCAATTTCTGCA CGCCATGGCCTGGCCAAGTATATCC Fusion to RFP
CAACTTTGCCTTCGCCATTGT CTGGCCAAGTATATCCGCTA Genotyping
CAACTTTGCCTTCGCCATTGT GCTGCTAGGGTTTCGAGTAA RT-PCR
TGGGCATCATTGAATCTCATC CTGGCCAAGTATATCCGCTA RT-PCR
CURT1D CGCCCGGGATGGAGCTCTGCACCA GCAGATCTCTCACTATCAGACCC Fusion to
HA/cmyc
CGGAATTCATGTGTAGTAGAAAG GCTCTAGATCACTCACTATCAG Fusion to MBP
CGCCATGGAGCTCTGCACCA CGCCATGGCCTCACTATCAGACCC Fusion to RFP
TGCACCAGGTCTACCACAAT TTGAGTTTCCTCATCTTCAGC Genotyping
TGCACCAGGTCTACCACAAT GCAACAACTACTCCATCACG RT-PCR
CTCGAATAGTGAAGCTCCTC CACTCACTATCAGACCCAAG RT-PCR
PetC GCGAATTCGCGTCGAGTATTCCA GCTCTAGATTAAGACCACCATGGA Fusion to MBP
PsaD1 GCGAATTCATGGCCGAGAAAACA GCTCTAGATCACAAATCATAACTTTG Fusion to MBP
ACTIN1 TGCGACAATGGAACTGGAATG AGCATGTGGAAGTGCATACC RT-PCR

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
15
Supplemental References 1 – Supplemental References for Supplemental Figures and
Tables
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P., Stevenson,
D.K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C., Heller, C., Jeske, A.,
Koesema, E., Meyers, C.C., Parker, H., Prednis, L., Ansari, Y., Choy, N., Deen, H.,
Geralt, M., Hazari, N., Hom, E., Karnes, M., Mulholland, C., Ndubaku, R., Schmidt, I.,
Guzman, P., Aguilar-Henonin, L., Schmid, M., Weigel, D., Carter, D.E., Marchand, T.,
Risseeuw, E., Brogden, D., Zeko, A., Crosby, W.L., Berry, C.C., and Ecker, J.R. (2003).
Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657.
Armbruster, U., Zühlke, J., Rengstl, B., Kreller, R., Makarenko, E., Rühle, T.,
Schünemann, D., Jahns, P., Weisshaar, B., Nickelsen, J., and Leister, D. (2010). The
Arabidopsis thylakoid protein PAM68 is required for efficient D1 biogenesis and photosystem
II assembly. Plant Cell 22, 3439-3460.
DalCorso, G., Pesaresi, P., Masiero, S., Aseeva, E., Schünemann, D., Finazzi, G., Joliot,
P., Barbato, R., and Leister, D. (2008). A complex containing PGRL1 and PGR5 is involved
in the switch between linear and cyclic electron flow in Arabidopsis. Cell 132, 273-285.
Ferro, M., Brugiere, S., Salvi, D., Seigneurin-Berny, D., Court, M., Moyet, L., Ramus, C.,
Miras, S., Mellal, M., Le Gall, S., Kieffer-Jaquinod, S., Bruley, C., Garin, J., Joyard, J.,
Masselon, C., and Rolland, N. (2010). AT_CHLORO, a comprehensive chloroplast
proteome database with subplastidial localization and curated information on envelope
proteins. Mol Cell Proteomics 9, 1063-1084.
Friso, G., Giacomelli, L., Ytterberg, A.J., Peltier, J.B., Rudella, A., Sun, Q., and Wijk,
K.J. (2004). In-depth analysis of the thylakoid membrane proteome of Arabidopsis thaliana
chloroplasts: new proteins, new functions, and a plastid proteome database. Plant Cell 16,
478-499.
Froehlich, J.E., Wilkerson, C.G., Ray, W.K., McAndrew, R.S., Osteryoung, K.W., Gage,
D.A., and Phinney, B.S. (2003). Proteomic study of the Arabidopsis thaliana chloroplastic
envelope membrane utilizing alternatives to traditional two-dimensional electrophoresis. J
Proteome Res 2, 413-425.
Granvogl, B., Reisinger, V., and Eichacker, L.A. (2006). Mapping the proteome of
thylakoid membranes by de novo sequencing of intermembrane peptide domains.
Proteomics 6, 3681-3695.
Ito, T., Motohashi, R., Kuromori, T., Mizukado, S., Sakurai, T., Kanahara, H., Seki, M.,
and Shinozaki, K. (2002). A new resource of locally transposed Dissociation elements for

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
16
screening gene-knockout lines in silico on the Arabidopsis genome. Plant Physiol 129, 1695-
1699.
Khrouchtchova, A., Hansson, M., Paakkarinen, V., Vainonen, J.P., Zhang, S., Jensen,
P.E., Scheller, H.V., Vener, A.V., Aro, E.M., and Haldrup, A. (2005). A previously found
thylakoid membrane protein of 14kDa (TMP14) is a novel subunit of plant photosystem I and
is designated PSI-P. FEBS Lett 579, 4808-4812.
Kleffmann, T., Russenberger, D., von Zychlinski, A., Christopher, W., Sjolander, K.,
Gruissem, W., and Baginsky, S. (2004). The Arabidopsis thaliana chloroplast proteome
reveals pathway abundance and novel protein functions. Curr Biol 14, 354-362.
Munekage, Y., Hojo, M., Meurer, J., Endo, T., Tasaka, M., and Shikanai, T. (2002). PGR5
is involved in cyclic electron flow around photosystem I and is essential for photoprotection in
Arabidopsis. Cell 110, 361-371.
Parinov, S., Sevugan, M., Ye, D., Yang, W.C., Kumaran, M., and Sundaresan, V. (1999).
Analysis of flanking sequences from dissociation insertion lines: a database for reverse
genetics in Arabidopsis. Plant Cell 11, 2263-2270.
Peltier, J.B., Ytterberg, A.J., Sun, Q., and van Wijk, K.J. (2004). New functions of the
thylakoid membrane proteome of Arabidopsis thaliana revealed by a simple, fast, and
versatile fractionation strategy. J Biol Chem 279, 49367-49383.
Peng, L., Shimizu, H., and Shikanai, T. (2008). The chloroplast NAD(P)H dehydrogenase
complex interacts with photosystem I in Arabidopsis. J Biol Chem 283, 34873-34879.
Pesaresi, P., Pribil, M., Wunder, T., and Leister, D. (2011). Dynamics of reversible protein
phosphorylation in thylakoids of flowering plants: the roles of STN7, STN8 and TAP38.
Biochim Biophys Acta 1807, 887-896.
Pesaresi, P., Hertle, A., Pribil, M., Kleine, T., Wagner, R., Strissel, H., Ihnatowicz, A.,
Bonardi, V., Scharfenberg, M., Schneider, A., Pfannschmidt, T., and Leister, D. (2009).
Arabidopsis STN7 kinase provides a link between short- and long-term photosynthetic
acclimation. Plant Cell 21, 2402-2423.
Rutschow, H., Ytterberg, A.J., Friso, G., Nilsson, R., and van Wijk, K.J. (2008).
Quantitative proteomics of a chloroplast SRP54 sorting mutant and its genetic interactions
with CLPC1 in Arabidopsis. Plant Physiol 148, 156-175.
Samson, F., Brunaud, V., Balzergue, S., Dubreucq, B., Lepiniec, L., Pelletier, G.,
Caboche, M., and Lecharny, A. (2002). FLAGdb/FST: a database of mapped flanking
insertion sites (FSTs) of Arabidopsis thaliana T-DNA transformants. Nucleic Acids Res 30,
94-97.

Supplemental Data. Armbruster et al. (2013). Plant Cell 10.1105/tpc.113.113118.
17
Schwenkert, S., Umate, P., Dal Bosco, C., Volz, S., Mlcochova, L., Zoryan, M.,
Eichacker, L.A., Ohad, I., Herrmann, R.G., and Meurer, J. (2006). PsbI affects the stability,
function, and phosphorylation patterns of photosystem II assemblies in tobacco. J Biol Chem
281, 34227-34238.
Sessions, A., Burke, E., Presting, G., Aux, G., McElver, J., Patton, D., Dietrich, B., Ho,
P., Bacwaden, J., Ko, C., Clarke, J.D., Cotton, D., Bullis, D., Snell, J., Miguel, T.,
Hutchison, D., Kimmerly, B., Mitzel, T., Katagiri, F., Glazebrook, J., Law, M., and Goff,
S.A. (2002). A high-throughput Arabidopsis reverse genetics system. Plant Cell 14, 2985-
2994.
Sundaresan, V., Springer, P., Volpe, T., Haward, S., Jones, J.D., Dean, C., Ma, H., and
Martienssen, R. (1995). Patterns of gene action in plant development revealed by enhancer
trap and gene trap transposable elements. Genes Dev 9, 1797-1810.
Zybailov, B., Rutschow, H., Friso, G., Rudella, A., Emanuelsson, O., Sun, Q., and van
Wijk, K.J. (2008). Sorting signals, N-terminal modifications and abundance of the chloroplast
proteome. PLoS ONE 3, e1994.



![A B 80 [rCLB19] (%) 60 bound - · PDF fileKindgren et al. (2015). Plant Cell 10.1105/tpc.114.134189 1. Supplemental Figure 2. Relative Kd, but not absolute Kd, is reproducible between](https://static.fdocuments.in/doc/165x107/5aa41f8c7f8b9a1d728b71b5/a-b-80-rclb19-60-bound-et-al-2015-plant-cell-101105tpc114134189.jpg)