
Aplikasi Formula Campuran Rizobakteri untuk Pengendalian...
36
Volume 10, Nomor 2, April 2014 Halaman 37–44 DOI: 10.14692/jfi.10.2.37 Aplikasi Formula Campuran Rizobakteri untuk Pengendalian Penyakit Busuk Akar Rhizoctonia dan Peningkatan Hasil Kedelai di Tanah Ultisol Application of Mixture Formulation of Rhizobacteria to Control Rhizoctonia Root Rot Disease and Enhance of Soybean Yield in Ultisol Soil Andi Khaeruni*, Asniah, Muhammad Taufik, Gusti Ayu Kade Sutariati Universitas Halu Oleo, Kendari 93232 ABSTRAK Penyakit busuk akar yang disebabkan oleh Rhizoctonia solani merupakan salah satu penyakit penting pada tanaman kedelai di lahan ultisol di Sulawesi Tenggara. Penggunaan rizobakteri merupakan salah satu teknik pengendalian yang dapat dipertimbangkan untuk mengendalikan patogen tersebut. Penelitian ini bertujuan menentukan efektivitas formula campuran rizobakteri untuk mengendalikan penyakit busuk akar rhizoctonia, meningkatkan pertumbuhan dan hasil tanaman kedelai di tanah ultisol. Penelitian disusun dalam rancangan acak lengkap dengan tujuh perlakuan, yaitu aplikasi formulasi rizobakteri pada benih, formulasi rizobakteri pada benih dan 2 minggu setelah tanam, formulasi rizobakteri pada benih, 2 dan 4 minggu setelah tanam, formulasi rizobakteri pada benih dan aplikasi fungisida 2 minggu setelah tanam, fungisida pada benih dan 2 minggu setelah tanam, fungisida pada benih, 2 dan 4 minggu setelah tanam, dan kontrol (tanpa rizobakteri dan fungisida). Semua perlakuan diinokulasi dengan Rhizoctonia solani. Hasil penelitian menunjukkan aplikasi rizobakteri pada benih 2 dan 4 minggu setelah tanam atau aplikasi pada benih yang dikombinasikan fungisida pada umur 2 minggu setelah tanam sangat efektif mengendalikan penyakit busuk akar rhizoctonia pada tingkat insidensi penyakit 0%, meningkatkan pertumbuhan tinggi tanaman dan jumlah daun sebesar 119% dan 170%, dan bobot biji bernas sebesar 1870% tanaman kedelai di tanah ultisol dibandingkan dengan tanaman kontrol. Kata kunci: agens hayati, insidensi penyakit, pemacu pertumbuhan tanaman, Rhizoctonia solani ABSTRACT Rhizoctonia root rot disease caused by Rhizoctonia solani is one of the most important disease in soybean area, including at ultisol land in Southeast Sulawesi. Rhizobacteria application is one alternative method to control this pathogen. The aim of this experiment was to study of rhizobacteria indigenous formulation to control of Rhizoctonia root rot disease and increase soybean yield in Ultisol soil. A complete randomized design with seven treatments was used *Alamat penulis korespondensi: Jurusan Agroteknologi, Fakultas Pertanian Universitas Halu Oleo, Jalan HEA Mokodompit, Kendari 93232 Tel: 0401-3193596, Faks: 0401-3193596, Surel: [email protected] 37 ISSN: 2339-2479
Transcript of Aplikasi Formula Campuran Rizobakteri untuk Pengendalian...

Volume 10, Nomor 2, April 2014Halaman 37–44
DOI: 10.14692/jfi.10.2.37
Aplikasi Formula Campuran Rizobakteri untuk Pengendalian Penyakit Busuk Akar Rhizoctonia dan
Peningkatan Hasil Kedelai di Tanah Ultisol
Application of Mixture Formulation of Rhizobacteria to Control Rhizoctonia Root Rot Disease and Enhance of Soybean Yield in Ultisol Soil
Andi Khaeruni*, Asniah, Muhammad Taufik, Gusti Ayu Kade SutariatiUniversitas Halu Oleo, Kendari 93232
ABSTRAK
Penyakit busuk akar yang disebabkan oleh Rhizoctonia solani merupakan salah satu penyakit penting pada tanaman kedelai di lahan ultisol di Sulawesi Tenggara. Penggunaan rizobakteri merupakan salah satu teknik pengendalian yang dapat dipertimbangkan untuk mengendalikan patogen tersebut. Penelitian ini bertujuan menentukan efektivitas formula campuran rizobakteri untuk mengendalikan penyakit busuk akar rhizoctonia, meningkatkan pertumbuhan dan hasil tanaman kedelai di tanah ultisol. Penelitian disusun dalam rancangan acak lengkap dengan tujuh perlakuan, yaitu aplikasi formulasi rizobakteri pada benih, formulasi rizobakteri pada benih dan 2 minggu setelah tanam, formulasi rizobakteri pada benih, 2 dan 4 minggu setelah tanam, formulasi rizobakteri pada benih dan aplikasi fungisida 2 minggu setelah tanam, fungisida pada benih dan 2 minggu setelah tanam, fungisida pada benih, 2 dan 4 minggu setelah tanam, dan kontrol (tanpa rizobakteri dan fungisida). Semua perlakuan diinokulasi dengan Rhizoctonia solani. Hasil penelitian menunjukkan aplikasi rizobakteri pada benih 2 dan 4 minggu setelah tanam atau aplikasi pada benih yang dikombinasikan fungisida pada umur 2 minggu setelah tanam sangat efektif mengendalikan penyakit busuk akar rhizoctonia pada tingkat insidensi penyakit 0%, meningkatkan pertumbuhan tinggi tanaman dan jumlah daun sebesar 119% dan 170%, dan bobot biji bernas sebesar 1870% tanaman kedelai di tanah ultisol dibandingkan dengan tanaman kontrol.
Kata kunci: agens hayati, insidensi penyakit, pemacu pertumbuhan tanaman, Rhizoctonia solani
ABSTRACT
Rhizoctonia root rot disease caused by Rhizoctonia solani is one of the most important disease in soybean area, including at ultisol land in Southeast Sulawesi. Rhizobacteria application is one alternative method to control this pathogen. The aim of this experiment was to study of rhizobacteria indigenous formulation to control of Rhizoctonia root rot disease and increase soybean yield in Ultisol soil. A complete randomized design with seven treatments was used
*Alamat penulis korespondensi: Jurusan Agroteknologi, Fakultas Pertanian Universitas Halu Oleo, Jalan HEA Mokodompit, Kendari 93232Tel: 0401-3193596, Faks: 0401-3193596, Surel: [email protected]
37
ISSN: 2339-2479

J Fitopatol Indones Khaeruni et al.
in this experiment. The treatments were (A) rhizobacteria formulation for seed treatment, (B) rhizobacteria formulation for seed treatment and repeated at 2 weeks after planting, (C) rhizobacteria formulation for seed treatment, repeated at 2 and 4 weeks after planting, (D) rhizobacteria formulation for seed treatment and fungicide applied at 2 weeks after planting, (E) fungicide seed treatment, and repeated at 2 weeks after planting, (F) fungicide seed treatment, and repeated at 2 and 4 weeks after planting, and control (without rhizobacteria and fungicides). All treatments were inoculated by R. solani and replicated three times. The results showed that rhizobacteria seed treatment and repeated at 2 and 4 weeks after planting was the most effective treatment to control Rhizoctonia root rot disease, and increase plant height and leaf number up to 119% and 170%, respectively, and increased the yield of soybean up to 1870% in ultisol soil compared to plant with control treatment.
Key words: biocontrol agents, disease insidence, plant growth promoting, Rhizoctonia solani
PENDAHULUAN
Rhizoctonia solani penyebab penyakit busuk akar merupakan satu patogen penting pada tanaman kedelai di lahan ultisol di Sulawesi Tenggara. Patogen ini dapat bertahan hidup di dalam tanah dalam bentuk sklerotium atau miselium dorman dan dapat pula bertahan hidup pada biji. Infeksi tanaman terjadi pada fase pratumbuh, saat benih tumbuh, maupun fase pascatumbuh yang mengakibatkan tanaman berwarna kuning, kerdil, layu, dan mati.
Di Indonesia fungisida berbahan aktif mankozeb digunakan petani untuk mengendalikan penyakit busuk akar rhizoctonia. Keefektifan fungisida berbahan aktif mankozeb dapat mengendalikan penyakit busuk akar rhizoctonia antara 52.5% dan 76.6% (Dorrance et al. 2003). Pestisida sintetik harus digunakan secara bijaksana karena dapat mempengaruhi karakteristik fisik dan biologi tanah dan meninggalkan residu yang membahayakan lingkungan dan makhluk hidup lainnya. Oleh karena itu, pengembangan bioteknologi menggunakan mikroorganisme agens pemacu pertumbuhan tanaman (PPT) sekaligus sebagai agens pengendali hayati terhadap patogen diperlukan.
Fatima et al. (2009) melaporkan bahwa penggunaan rizobakteri mampu meningkatkan ketahanan tanaman gandum terhadap R. solani. Khaeruni et al. (2011) mengemukakan bahwa pencampuran tiga isolat rizobakteri indigenos Bacillus subtilis ST21b, B. cereus ST21e, dan
Serratia galur SS29a yang diformulasikan dalam bahan pembawa gambut dan lempung mampu menekan perkembangan penyakit layu fusarium pada tanaman tomat sampai 60%. Penelitian lain menunjukkan bahwa aplikasi rizobakteri pada benih yang disusul dengan aplikasinya saat 2 dan 4 minggu setelah tanam di sekitar perakaran tanaman, selain meningkatkan ketahanan tanaman terhadap penyakit layu fusarium juga meningkatkan hasil panen tomat di tanah ultisol (Khaeruni et al. 2013). Sebelum formula rizobakteri tersebut diperkenalkan kepada masyarakat luas masih perlu diteliti kemampuan dan efektivitasnya dalam mengendalikan patogen tular tanah lainnya, seperti Rhizoctania solani pada tanaman kedelai.
Oleh karena itu, penelitian ini bertujuan menentukan efektivitas formula campuran rizobakteri Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a untuk mengendalikan penyakit busuk akar rhizoctonia dan meningkatkan pertumbuhan dan hasil tanaman kedelai di tanah ultisol.
BAHAN DAN METODE
Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a, merupakan rizobakteri hasil isolasi dari tanah ultisol di Sulawesi Tenggara (Khaeruni et. al. 2010) dan R. solani merupakan koleksi Laboratorium Ilmu Hama dan Penyakit Tumbuhan, Universitas Halu Oleo.
38

J Fitopatol Indones Khaeruni et al.
Penelitian disusun dalam rancangan acak lengkap yang terdiri atas tujuh perlakuan (Tabel 1). Semua perlakuan diinokulasi dengan R. solani dengan tiga ulangan. Setiap unit percobaan terdiri atas 7 tanaman sehingga secara keseluruhan terdapat 147 tanaman.
Persiapan Medium TanamMedium tanam yang digunakan ialah tanah
ultisol yang dicampur dengan pupuk kandang dengan perbandingan 9:1 (v:v) yang disterilkan dengan sterilisasi uap dan dimasukkan ke dalam kantong plastik berukuran 20 cm x 30 cm sebanyak 147 buah.
Penyiapan Inokulum dan Inokulasi PatogenSebanyak empat potong miselium R. solani
dengan ukuran 1 cm x 1 cm yang berumur 7 hari pada medium agar-agar desktrosa kentang (ADK) dibiakkan dalam 200 mL Medium cair dekstrosa kentang dan biakan ini dikocok dengan kecepatan 150 rpm pada suhu ruang 27 °C selama 14 hari. Massa miselium dipanen dengan menyaringnya menggunakan kain kasa steril berlapis kertas tisu steril. Selanjutnya kerak miselium dicuci dengan air steril 2 kali dan selanjutnya disuspensikan dengan 500 mL air steril. Suspensi miselium
R. solani dicampur dengan pasir steril dengan perbandingan 1:5 (v:b) yang dijadikan sebagai sumber inokulum.
Pasir steril yang mengandung miselium R. solani diinokulasikan dengan cara menaburkannya ke dalam lubang pada medium tanam lalu ditutup dengan tanah. Inokulasi dilakukan dua kali, pertama dilakukan sebanyak 10 g per pot saat 2 minggu sebelum tanam dan kedua sebanyak 5 g per pot saat 4 minggu setelah tanam.
Pembuatan Suspensi dan Formula Rizobakteri
Suspensi rizobakteri dibuat dengan mengambil masing-masing 5 ose isolat rizobakteri B. subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a umur 2 hari pada medium trypticase soy agar (TSA) yang dilarutkan secara homogen dalam 50 mL akuades steril dalam botol Schoot secara terpisah. Larutan diinkubasi di atas alat pengocok dengan kecepatan 150 rpm pada suhu ruang selama 12 jam. Suspensi rizobakteri pada kerapatan 109–1010 cfu mL-1 dicampur dengan sekam halus (1:100 v:b) dan digunakan sebagai inokulum untuk dicampurkan pada benih kedelai varietas Anjosmoro dengan perbandingan 1:1 (v:v) dan diinkubasi pada suhu ruang selama 4 jam sebelum ditanam.
Formula rizobakteri dibuat dengan cara mencampurkan 100 mL suspensi rizobakteri B. subtilis ST21b, B. cereus ST21e, Serratia galur SS29a masing-masing dengan kerapatan sel 109–1010 cfu mL-1 dalam 1 kg bahan formulasi gambut dan lempung halus yang telah disterilkan dengan perbandingan 3:2. Formulasi rizobakteri tersebut diaplikasikan pada medium tanam pada perlakuan B, C, dan D (Tabel 1) dengan cara menaburkan 10 g per tanaman pada lubang di sekeliling perakaran tanaman.
Aplikasi Fungisida Sintetik pada Benih dan Tanaman
Aplikasi fungisida sintetik pada benih dilakukan dengan cara merendam benih dalam larutan fungisida berbahan aktif
39
Tabel 1 Perlakuan yang diujikan untuk aplikasi pengendalian hayati terhadap penyakit busuk akar rhizoctonia
Kode Perlakuan
A formula rizobakteri pada benih
Bformula rizobakteri pada benih dan 2 minggu setelah tanam (MST)
C formula rizobakteri pada benih, 2 dan 4 MST
D formula rizobakteri pada benih dan fungisida 2 MST
E fungisida pada benih dan 2 MST
F fungisida pada benih, 2 dan 4 MST
K tanpa perlakuan rizobakteri dan fungisida

J Fitopatol Indones Khaeruni et al.
mankozeb 0.1% selama 1 jam, lalu benih dikeringanginkan sebelum ditanam. Aplikasi fungisida pada tanaman dilakukan dengan menyiram larutan fungisida sebanyak 10 mL dengan konsentrasi yang sama di sekitar akar tanaman sesaui dengan waktu aplikasi pada perlakuan D, E, dan F (Tabel 1).
Penanaman dan Pemeliharaan TanamanBenih ditanam dalam pot plastik yang
telah disiapkan sebanyak 2 benih per pot dengan cara membuat lubang kecil sedalam kurang lebih 2 cm dari permukaan tanah. Dua minggu setelah tanam dipilih satu tanaman yang dibiarkan hidup sebagai tanaman uji. Penyiraman dilakukan dua kali dalam satu hari, pagi dan sore, sedangkan pengendalian hama dan gulma dilakukan secara manual jika ditemukan pada tanaman uji.
Pengamatan Pengamatan tinggi tanaman dan jumlah
daun trifoliat dilakukan setiap minggu pada umur 2 sampai 6 minggu setelah tanam pada lima tanaman sampel yang dipilih secara acak. Tinggi tanaman yang diukur mulai dari pangkal batang di atas permukaan tanah sampai daun pucuk paling atas, sedangkan jumlah daun trifoliat dihitung mulai dari daun bawah sampai daun yang tertinggi yang telah terbentuk sempurna.
Hasil panen yang diamati meliputi: jumlah polong, bobot polong, jumlah dan bobot kering biji bernas. Jumlah dan bobot polong diamati pada 5 tanaman sampel pada saat panen ketika polong telah memasuki masak fisiologi, sedangkan pengamatan jumlah dan bobot kering panen biji bernas dilakukan pada saat satu minggu setelah panen dari tanaman sampel yang sama.
Insidensi penyakit diamati pada jumlah tanaman dengan gejala layu yang ditandai oleh menguningnya daun dan atau merunduknya tangkai daun. Pengamatan dilakukan pada saat tanaman berumur 3, 4, dan 5 minggu setelah tanam. Insidensi penyakit dihitung dengan menggunakan rumus sebagai berikut:
IP, insidensi penyakit; n, jumlah tanaman yang bergejala; N, total tanaman yang diamati.
Data hasil pengamatan dianalisis meng-gunakan metode sidik ragam dan dilanjutkan dengan Uji Jarak Berganda Duncan pada α 5%.
HASIL
Pertumbuhan Tanaman dan Hasil PanenFormula rizobakteri indigenos berpengaruh
nyata terhadap tinggi tanaman (Tabel 2). Tanaman tertinggi pada pengamatan minggu ke-2 hingga ke-6 setelah tanam terdapat pada perlakuan aplikasi rizobakteri pada benih, 2 dan 4 minggu setelah tanam (perlakuan C) dengan tinggi tanaman pada umur 6 minggu setelah tanam mencapai 104.2 cm. Tinggi tanaman pada perlakuan C tersebut berbeda nyata dengan perlakuan yang menggunakan fungisida sintetik (perlakuan E dan F) dan kontrol (K), dengan tinggi tanaman masing-masing 45.9 cm, 39.8 cm, dan 47.4 cm sehingga terjadi peningkatan pertumbuhan tinggi tanaman 119% dibandingkan dengan tanaman kontrol.
Rata-rata jumlah daun tanaman kedelai yang diberi perlakuan rizobakteri selalu lebih banyak dan berbeda nyata dengan perlakuan fungisida sintetik dan kontrol (Tabel 3). Daun terbanyak pada setiap waktu pengamatan terdapat pada perlakuan aplikasi rizobakteri pada benih, 2 dan 4 minggu setelah tanaman (perlakuan C) dengan jumlah daun pada minggu ke-6 sebanyak 26.8 helai. Jumlah daun pada perlakuan C tersebut berbeda nyata dengan perlakuan yang menggunakan fungisida sintetik (perlakuan E dan F) dan kontrol (K) dengan jumlah daun masing-masing 9.6 helai, 8.9 helai, dan 9.9 helai sehingga terjadi peningkatan jumlah daun sebesar 170% dibandingkan dengan tanaman kontrol.
Hasil panen menunjukkan bahwa aplikasi formula rizobakteri indigenos berpengaruh nyata terhadap hasil panen kedelai yang diinokulasi R. solani (Tabel 4). Pengamatan hasil panen berupa berupa jumlah polong bobot kering polong, jumlah biji, bernas, dan
40
IP= nN
x 100% , dengan

J Fitopatol Indones Khaeruni et al.
bobot biji bernas pada perlakuan rizobakteri pada benih dan aplikasi fungisida sintetik 2 minggu setelah tanam (perlakuan D) selalu lebih tinggi: jumlah polong sebanyak 147 biji, bobot polong 69.1 g per tanaman, jumlah biji bernas sebanyak 231.1 biji dengan bobot 53.2 g per tanaman, walaupun tidak berbeda nyata dengan perlakuan rizobakteri lainnya
(perlakuan A, B, dan C), namun berbeda nyata dengan perlakuan tanpa rizobakteri (perlakuan E, F, dan K). Terjadi peningkatan bobot biji bernas pada perlakuan D sebesar 1870% dibandingkan dengan bobot biji bernas pada tanaman kontrol dengan bobot biji bernas 2.7 g per tanaman .
41
Tabel 2 Pengaruh aplikasi formula campuran rizobakteri Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a terhadap tinggi tanaman tanaman kedelai yang diinokulasi Rhizoctonia solani
PerlakuanRata-rata tinggi tanaman* minggu ke-
(cm)2 3 4 5 6
A 20.8 a 35.0 a 53.8 b 68.9 a 95.5 aB 20.7 a 32.5 a 56.3 ab 77.1 a 101.0 aC 19.8 ab 35.7 a 65.2 a 80.2 a 104.2 aD 18.3 bc 34.3 a 55.5 ab 72.1 a 94.9 aE 16.6 c 23.6 b 27.3 c 33.7 b 45.9 bF 15.6 c 20.9 b 25.8 c 37.7 b 39.8 bK 17.3 c 23.1 b 27.8 c 35.7 b 47.4 b
A, formulasi rizobakteri pada benih; B, formulasi rizobakteri pada benih dan 2 minggu setelah tanam (MST); C, formulasi rizobakteri pada benih, 2 dan 4 MST; D, formulasi rizobakteri pada benih dan fungisida 2 MST; E, fungisida pada benih dan 2 MST; F, fungisida pada benih, 2 dan 4 MST; dan K, tanpa perlakuan rizobakteri dan fungisida. *Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf kepercayaan 95%.
Tabel 3 Pengaruh aplikasi formula campuran rizobakteri Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a terhadap jumlah daun tanaman kedelai yang diinokulasi Rhizoctonia solani
PerlakuanRata-rata jumlah daun trifoliat/tanaman* pada minggu ke-
2 3 4 5 6A 4.0 a 6.4 b 8.1 a 12.5 a 19.2 bB 4.0 a 6,7 a 9,1 a 12.9 a 23.7 aC 4.1 a 6.7 ab 9.1 a 15.9 a 26.8 aD 4.0 a 6.7 a 8.9 a 12.2 a 24.9 aE 3.6 b 4.7 c 4.1 c 6.1 b 9.6 cF 3.5 b 4.8 c 5.2 b 6.7 b 8.9 cK 3.4 b 4.8 c 4.9 bc 6.3 b 9.9 c
A, formulasi rizobakteri pada benih; B, formulasi rizobakteri pada benih dan 2 minggu setelah tanam (MST); C, formulasi rizobakteri pada benih, 2 dan 4 MST; D, formulasi rizobakteri pada benih dan fungisida 2 MST; E, fungisida pada benih dan 2 MST; F, fungisida pada benih, 2 dan 4 MST; dan K, tanpa perlakuan rizobakteri dan fungisida. *Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf kepercayaan 95%.

J Fitopatol Indones Khaeruni et al.
Insidensi Penyakit Busuk Batang Rhizoctonia
Insidensi penyakit pada pengamatan minggu ke-3, ke-4, dan ke-5 setelah tanam perlakuan rizobakteri pada benih yang diikuti aplikasi pada tanaman umur 2 dan 4 minggu setelah tanam (perlakuan C) dan aplikasi rizobakteri pada benih dan fungisida 2 minggu setelah tanam (perlakuan D) mampu mencegah terjadinya penyakit busuk batang rhizoctonia hingga akhir pengamatan. Sementara pada waktu yang sama perlakuan fungisida sintetik menyebabkan (perlakuan E dan F) insidensi penyakit berkisar 33.3%, dan kontrol mencapai 66.6% (Tabel 5).
PEMBAHASAN
Rizobakteri merupakan kelompok bakteri yang hidup di rizosfer tanaman dan berinteraksi secara intensif dengan akar tanaman maupun tanah dan berperan sebagai agens PPT dalam ekosistem tanah. Hal ini dibuktikan bahwa aplikasi rizobakteri inidigens dalam penelitan ini meningkatkan pertumbuhan tanaman, hasil panen, serta ketahanan tanaman kedelai terhadap R. solani.
Pada akhir pengamatan saat tanaman berumur 6 minggu setelah tanam, tanaman tertinggi (104.2 cm) dan daun terbanyak (26.8
helai) diperoleh pada perlakuan inokulasi rizobakteri pada benih, 2 dan 4 minggu setelah tanam (perlakuan C), walaupun nilai-nilai tersebut tidak berbeda nyata dengan perlakuan rizobakteri lainnya, termasuk perlakuan campuran rizobakteri dan fungisida sintetik (perlakuan D). Sedangkan perlakuan fungisida pada benih, 2 dan 4 minggu setelah tanam (perlakuan F) menunjukkan pertumbuhan tanaman yang kurang baik.
Pertumbuhan terbaik pada tanaman yang diaplikasi dengan rizobakteri karena rizobakteri yang digunakan menghasilkan hormon PPT dan apabila semakin sering diaplikasikan maka pertumbuhan juga memberikan hasil yang lebih baik. Hasil penelitian sebelumnya menyatakan bahwa isolat B. subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a mampu memproduksi indol asam asetat (IAA) masing-masing 33.00 ppm, 59.44 ppm, dan 28.87 ppm (Khaeruni et al. 2010). Lebih lanjut dinyatakan bahwa ketiga isolat rizobakteri tersebut juga memiliki kemampuan sebagai pelarut fosfat dan menfiksasi nitrogen nonsimbiotik. Kemampuan rizobakteri sebagai PPT ditunjukkan dengan kemampuan dalam menyediakan dan memobilisasi penyerapan berbagai unsur hara dalam tanah serta mensintesis dan mengubah konsentrasi
42
Tabel 4 Pengaruh aplikasi formula campuran rizobakteri Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a terhadap hasil panen tanaman kedelai yang diinokulasi Rhizoctonia solani
Perlakuan JP*(biji/tan)
BKP*(g/tan)
JBB*(Biji/tan)
BBB*(g/tan)
A 132.7 a 52.1 b 170.2 a 23.8 cB 139.9 a 54.3 b 181.4 a 29.0 cC 145.7 a 59.7 ab 217.1 a 39.1 bD 147.0 a 69.1 a 231.1 a 53.2 a E 67.1 b 21.4 c 92.4 b 10.2 dF 49.0 bc 16.0 c 47.4 bc 5.7 deK 38.3 c 13.0 c 29.7 c 2.7 e
JPT, jumlah polong; BKP, berat kering polong; JBB, jumlah biji bernas; BBB, bobot biji bernas. A, formulasi rizobakteri pada benih; B, formulasi rizobakteri pada benih dan 2 minggu setelah tanam (MST); C, formulasi rizobakteri pada benih, 2 dan 4 MST; D, formulasi rizobakteri pada benih & fungisida 2 MST; E, fungisida pada benih dan 2 MST; F, fungisida pada benih, 2 & 4 MST; dan K, tanpa perlakuan rizobakteri dan fungisida.*Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf kepercayaan 95%.

J Fitopatol Indones Khaeruni et al.
43
berbagai fitohormon (Thakuria et al. 2004; Ahmad et al. 2005; Compant et al. 2005).
Insidensi penyakit pada tanaman kedelai dapat diketahui melalui pengamatan gejala eksternal yang ditampakkan. Gejala ini diawali dengan menguningnya daun mulai dari daun bawah dan seterusnya bergerak ke atas, tanaman juga menjadi kerdil. Gejala yang tampak pada pangkal batang ditandai dengan timbulnya bercak-bercak cokelat, akar menjadi busuk, mengering dan berwarna cokelat. Menguningnya daun dan membusuknya akar tanaman kedelai pada penelitian ini diduga karena adanya serangan R. solani pada jaringan pengangkut dari akar ke seluruh bagian tanaman sehingga tanaman tidak mendapatkan suplai air, mineral, dan unsur hara dari dalam tanah.
Hasil pengamatan insidensi penyakit dalam penelitian ini menunjukkan bahwa aplikasi formula rizobakteri indigenos yang digunakan efektif menekan penyakit busuk akar rhizoctonia pada tanaman kedelai karena tidak timbulnya penyakit walaupun tanaman tersebut diinokulasi R. solani. Sementara tanaman yang tidak mendapat perlakuan rizobakteri maupun tanaman yang hanya diaplikasi dengan fungisida, penyakit
busuk batang rhizoctonia timbul dengan insidensi penyakit berkisar 33.3%-66.7%. Hal ini mengindikasikan bahwa rizobakteri lebih unggul mengatasi serangan R. solani dibandingkan dengan fungisida sintetik mankozeb. Rendahnya insidensi penyakit atau ketidakmunculan gejala pada perlakuan rizobakteri diduga karena bakteri tersebut memiliki kemampuan untuk menghasilkan atau memproduksi senyawa metabolik sekunder, seperti antibiotik yang mampu menekan perkembangan patogen dan enzim kitinase yang mampu mendegradasi kitin yang merupakan salah satu komponen penyusun dinding sel R. solani sehingga perkembangan cendawan tersebut terhambat akibat adanya aktivitas rizobakteri di sekitar perakaran tanaman. Efektivitas B. subtilis ST21b dan Serratia galur SS29a dalam mendegredasi kitin sangat kuat sehingga mampu memiliki aktivitas antagonis yang cukup kuat secara in vitro dengan potensi penghambatan terhadap patogen R. solani, yaitu 18.5% dan 48.2% (Khaeruni et al. 2010). Hasil ini menguatkan penelitian sebelumnya yang menyatakan bahwa kelompok rizobakteri yang dapat menekan pertumbuhan cendawan patogen tanaman di antaranya berkaitan dengan
Tabel 5 Pengaruh aplikasi formula campuran rizobakteri Bacillus subtilis ST21b, B. cereus ST21e, dan Serratia galur SS29a terhadap kejadian penyakit tanaman kedelai yang diinokulasi Rhizoctonia solani
PerlakuanRata-rata insidensi penyakit* pada minggu ke-
(%) 3 4 5
A 0.0 c 9.5 c 14.8 cB 9.5 b 14.3 b 14.3 cC 0.0 d 0,0 d 0.0 dD 0.0 d 0.0 d 0.0 dE 4.8 c 9.5 c 33.3 bF 4,8 b 14.3 b 33.3 bK 52.3 a 52.4 a 66.7 a
A, formulasi rizobakteri pada benih; B, formulasi rizobakteri pada benih dan 2 minggu setelah tanam (MST); C, formulasi rizobakteri pada benih, 2 dan 4 MST; D, formulasi rizobakteri pada benih dan fungisida 2 MST; E, fungisida pada benih dan 2 MST; F, fungisida pada benih, 2 dan 4 MST; dan K, tanpa perlakuan rizobakteri dan fungisida. *Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf kepercayaan 95%.

J Fitopatol Indones Khaeruni et al.
44
kemampuannya menghasilkan enzim kitinase dan antibiotik (Idriss 2002; dan Sutariati et. al. 2005).
Peningkatan pertumbuhan dan kesehatan tanaman berimplikasi pada peningkatan hasil panen. Perlakuan yang menunjukkan peningkatan pertumbuhan dan kesehatan tanaman yang lebih baik juga berkorelasi positif dengan peningkatan hasil panen. Perlakuan formulasi rizobakteri pada benih yang dikombinasi dengan aplikasi fungsida sintetik pada umur 2 minggu setelah tanam memberikan hasil panen yang lebih baik dibandingkan dengan perlakuan lain, namun tidak berbeda nyata dengan perlakuan rizobakteri pada benih, 2 dan 4 minggu setelah tanam. Hasil penelitian ini menunjukkan bahwa aplikasi rizobakteri dapat diterapkan dalam sistem pengendalian penyakit terpadu karena dapat diintegrasikan dengan teknik pengendalian lainnya termasuk dikombinasikan dengan penggunaan fungisida sintetik sehingga bersinergi dalam meningkatkan kesehatan, memacu pertumbuhan, dan meningkatkan hasil panen tanaman.
UCAPAN TERIMA KASIH
Tulisan ini merupakan sebagian dari hasil penelitian Insentif Riset Dasar (RD-2010-0080) yang didanai oleh KNRT Tahun Anggaran 2010. Penulis mengucapkan terima kasih kepada Mariana Anwar untuk bantuan teknis penelitian di laboratorium dan rumah kasa.
DAFTAR PUSTAKA
Ahmad F, Ahmad I, Khan MS. 2005. Indole acetic acid production by the indigenous isolat of Azotobacter and fluorescent Pseudomonas in the presence and absence of trypthofan. Turk J Biol. 29:29–34.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Mini review: use of plant growth-promoting rhizobacteria for biocontrol of plant diseases: principles, mechanism of action and future prospect. Appl Environ Microbiol. 71:4951–4959.
DOI: http://dx.doi.org/10.1128/AEM.71.9. 4951-4959.2005.
Dorrance AE, Kleinhenz MD, Mcclure SA, Tuttle NT. 2003. Temperature, moisture, and seed treatment effects on Rhizoctonia solani root rot of soybean. Plant Dis. 87: 533–538. DOI: http://dx.doi.org/10.1094/PDIS.2003.87.5.533.
Fatima Z, Saleemi M, Zia M, Sultan T, Aslam M, Rehman RU, Chaudhary MF. 2009. Antifungal activity of plant growth-promoting rhizobacteria isolates against Rhizoctonia solani in wheat. African J of Biotechnol. 8(2):219-225.
Idriss E. 2002. Extracellular phytase activity of Bacillus amyloliquefaciens F2B45 contributs to its plant growth promoting effect. Microbiology. 148:2097–2109.
Khaeruni A, Sutariati GAK, Wahyuni S. 2010. Karakterisasi dan uji aktivitas bakteri rizosfer lahan ultisol sebagai pemacu pertumbuhan dan agensia hayati cendawan patogen tular tanah secara in-vitro. J Hama Penyakit Tumbuhan Trop. 10(2):123–30.
Khaeruni A, Syair, Sarmiza. 2011. The effectiveness of rhizobacteria mixture to control fusarium wilt disease and stimulate tomato plant growth in ultisol soil. Di dalam: Proceeding of International Seminar of Indonesian Phytopathology Sociaty; 2011 Des 3–5. Solo (ID): Perhimpunan Fitopatologi Indonesia.
Khaeruni A, Wahab A, Taufik M, Sutariati GAK. 2013. Keefektifan waktu aplikasi formulasi rizobakteri indigenos untuk mengendalikan layu fusarium dan meningkatkan hasil tanaman tomat di tanah ultisol. J Hort. 23(4):365–371.
Sutariati GAK, Widodo, Sudarsono, Ilyas S. 2005. Isolasi bakteri rizosfer dan karakteristik kemampuannya untuk peng-hambat pertumbuhan koloni cendawan patogen. Agriplus. 15:272–281.
Thakuria DNC, Talukdar C, Goswami S, Hazarika RC, Boro, Khan MR. 2004. Characterization and screening of bacteria from rhizosphore of rice grown in acidis soils of assam. Current Sci. 86(7):978–985.

Volume 10, Nomor 2, April 2014Halaman 45–52
DOI: 10.14692/jfi.10.2.45
Karakteristik Morfologi dan Fisiologi Isolat Pantoea stewartii subsp. stewartii pada Jagung
Morphological dan Physiological Characterization of
Pantoea stewartii subsp. stewartii from Maize
Yulfi Desi1*, Trimurti Habazar2, Agustian2, Ujang Khairul2, Syamsuwirman2, Prima Novia2
1Universitas Ekasakti, Padang 251132Universitas Andalas, Padang 25163
ABSTRAK
Pantoea stewartii subsp. stewartii adalah patogen penyebab penyakit layu stewart pada tanaman jagung yang baru diketahui keberadaannya di Indonesia. Keberadaan patogen ini telah dilaporkan terdapat pada beberapa daerah, namun belum banyak laporan penelitian terkait masalah ini. Penelitian bertujuan mencirikan isolat P. stewartii subsp. stewartii dari beberapa sentra pertanaman jagung di Sumatra Barat. Isolat-isolat P. stewartii subsp. stewartii berasal dari tanaman jagung yang menunjukkan penyakit layu stewart di 6 kabupaten di Sumatera Barat. Daun tanaman jagung yang mempunyai gejala garis kuning sepanjang tulang daun digerus dan disuspensikan, selanjutnya bakterinya diisolasi pada medium nutrient glucose agar (NGA) dan diinkubasi selama 3 × 24 jam pada suhu kamar. Karakter yang diuji ialah morfologi dan fisiologi bakteri, hipersensitivitas, patogenisitas, dan uji polymorfism chain reaction. Berdasarkan karakter morfologi dan fisiologi didapatkan 7 isolat yang menunjukkan karakter Pantoea. Hasil uji PCR membuktikan bahwa isolat LA homolog dengan Pantoea stewartii galur R1-104, isolat PR1 homolog dengan Pantoea stewartii galur R1-132, dan isolat PR2 homolog dengan Pantoea stewartii galur ATCC 29923.
Kata kunci: identifikasi, karakterisasi, layu stewart
ABSTRACT
Pantoea stewartii subsp. stewartii is the pathogen causing stewart’s wilt disease in maize. The occurrence of the disease was recently reported in Indonesia with limited information. Research was conducted to characterize several isolates of P. stewartii subsp. stewartii isolated from maize plants collected from 6 districts in West Sumatra. Leaves showing yellow stripes symptoms along the vein was ground and suspeneded then the bacteria was isolated and incubated for 3 × 24 h at room temperature in nutrient glucose agar (NGA). Pure bacterial colonies gained from this medium was used for characterization. Assessment of bacterial characters was conducted based on morphological assay, physiological assay, biological assay (hypersensitivity, and pathogenicity) followed by polymerase chain reaction (PCR)-based identification. Unique characters of the family Enterobacteriaceae was observed on 7 isolates based on morphological and physiological characters, and only 4 isolates (PR1, PP, ST1, LA) gave positive reaction on hypersensitivity and pathogenicity test. Further molecular identification confirmed 3 out of 7 isolates had high homology to Pantoea stewartii, i.e. Pantoea stewartii strain R1-104, R1-132, and ATCC 29923 for isolates LA, PR1, and PR2, respectively.
Key words: characterization, identification, stewart’s wilt
*Alamat penulis korespondensi: Fakultas Pertanian, Universitas Ekasakti Padang, Jalan Veteran Dalam 26 B, Padang 25113Tel: 0751-28859/26770, Faks: 0751-32694, Surel: [email protected]
45
ISSN: 2339-2479

J Fitopatol Indones Desi et al.
PENDAHULUAN
Penyakit layu stewart pada tanaman jagung disebabkan oleh bakteri Pantoea stewartii subsp. stewartii. Sampai saat ini informasi mengenai keberadaan penyakit layu ini di Indonesia masih terbatas. Beberapa penelitian terkait P. stewartii subsp. stewartii telah dilakukan oleh Trisnawati (2010) dan Rahma (2013).
Spesies dari genus Pantoea dicirikan berdasarkan morfologi koloni, uji fisiologi, biokimia, analisis asam lemak, dan komposisi quinon. Bodman dan Farrand (1995)melakukan uji ß-galactosidase, biosintesis EPS, CAT assay, dan produksi acyl homoserine lactone (AHL). TAS (2008) menetapkan beberapa karakter fisiologi antara lain: pergerakan bakteri, sifat Gram, metabolisme glukosa, oksidasi, katalase, hidrolisis esculin, produksi acetion, produksi indol, reduksi nitrat, pertumbuhan pada cis-anonitate, produksi asam (maltose, arbutin, raffinose, cellubiose, dan arabitol).
Penelitian bertujuan mencirikan isolat bakteri P. stewartii subsp. stewartii dari beberapa sentra pertanaman jagung di Sumatra Barat.
BAHAN DAN METODE
Sumber Tanaman SakitBahan tanaman sakit berasal dari beberapa
sentra pertanaman jagung di Sumatra Barat, yaitu Kabupaten Agam (Gadut dan Lubuk Basung), Kabupaten Pasaman Barat (Padang Tujuh dan Padang Rajo), Kabupaten Lima Puluh Kota (Simalanggang dan Tanjung Pati), Kabupaten Tanah Datar (Kumanggo dan Sumaniak), Kabupaten Padang Pariaman (Lubuk Alung dan Ketaping), dan Kota Padang (Pauh dan Kuranji). Daun tanaman jagung yang bergejala garis kuning sepanjang tulang daun dipotong dengan ukuran 1.5 cm x 7 cm, kemudian dibilas dengan akuades steril, alkohol 70%, dan akuades steril kembali, selanjutnya digerus dengan 10 mL akuades steril menggunakan lumpang steril. Suspensi bakteri diencerkan sampai 10-6 dan sebanyak
100 µL dituangkan pada medium nutrient glucose agar (NGA), selanjutnya diinkubasi selama 3 × 24 jam pada suhu kamar. Komposisi NGA ialah 28 g agar-agar nutrien ditambah 10 g glukosa, dan 1000 mL akuades. Koloni bakteri yang menunjukkan karakter P. stewartii subsp. stewartii: berwarna kuning, berkilat, berlendir, datar atau cembung. Isolat murni dimasukkan ke dalam tabung berisi akuades steril, divorteks dan disimpan di dalam lemari pendingin.
Uji MorfologiUji morfologi koloni bakteri yang diamati
mengikuti Cappucino dan Sherman (2002) terhadap warna koloni (kuning, kuning pucat), ukuran koloni (kecil, sedang, besar), bentuk koloni (bulat, tidak beraturan, tersebar), keadaan koloni (berlendir, tidak berlendir), dan bentuk pinggiran koloni (rata, bergelombang).
Uji GramUji Gram dilakukan untuk membedakan
kelompok bakteri berdasarkan ketebalan dinding sel. Sebanyak 10 µL KOH 3% diteteskan pada kaca objek, sejumlah sel dari koloni bakteri dipindahkan dengan jarum ose, diaduk sampai sempurna kira-kira pada diameter 1.5 cm. Jika ujung jarum ose diangkat ke atas dan suspensi menjadi berlendir dalam waktu 5-60 detik maka isolat bakteri tersebut ialah Gram negatif. Sebaliknya, jika tidak terbentuk lendir maka isolat tersebut merupakan bakteri Gram positif.
Uji KatalaseUji katalase dilakukan untuk mem-
bantu identifikasi bakteri tergolong Enterobacteriaceae (Taylor dan Anhanzar 1972). Setetes H2O2 ditempatkan pada gelas objek, isolat uji dioleskan secara merata menggunakan tusuk gigi steril, ditetesi kembali dengan 10 tetes H2O2, apabila muncul gelembung gas berarti menunjukkan reaksi positif (+), sebaliknya apabila tidak terdapat gelembung gas berarti menunjukkan reaksi negatif (-). Bakteri yang tergolong Enterobacteriaceae menunjukkan reaksi positif (+).
46

J Fitopatol Indones Desi et al.
Uji OksidasiUji oksidasi adalah pengujian yang
digunakan untuk menentukan apakah bakteri menghasilkan sitokrom-C oksidase tertentu. Uji oksidasi dilakukan menggunakan strip oxidase test. Isolat uji digoreskan pada strip oxidase test, munculnya warna biru setelah beberapa detik menunjukkan terjadinya reduksi atau bereaksi negatif (-), apabila tidak menimbulkan warna biru berarti terjadi oksidasi atau bereaksi positif (+).
Uji Metabolisme GlukosaSalah satu medium yang dapat digunakan
untuk identifikasi bakteri genus Pantoea ialah medium hugh-leifson (HL). Komposisi medium HL terdiri atas larutan utama dan larutan glukosa. Larutan utama terdiri atas20 g peptone, 5 g NaCl, 0.3 g KH2PO4, 3 g agar-agar, 3 mL bromotimol biru 1%. Larutan glukosa terdiri atas 10 g glukosa dan 60 mL dH2O. Medium ini digunakan untuk mengetahui isolat yang mampu menghasilkan glukosa secara anaerob. Pelaksanaan metabolisme glukosa dilakukan mengikuti prosedur sebagai berikut. Medium HL steril dimasukkan ke dalam tabung, kira-kira 2/3 bagian, kemudian isolat uji diinokulasikan pada 2 tabung (A dan B). Permukaan medium ditutup dengan menuangkan 1-2 mL vaselin steril. Munculnya warna kuning pada tabung dalam waktu 24 jam pada suhu 27 °C menunjukkan reaksi positif (+) dan warna biru menunjukkan reaksi negatif (-).
Deteksi Produksi Acyl Homoserine Lactone Deteksi produksi acyl homocerine lactone
(AHL) didasarkan pada metode Bodman dan Farrand (1995). Medium yang digunakan ialah medium minimal (MM) mengandung 40 µg X-Gal (5-bromo-4chloro-3indolyl-β-d-galactocidase) mL-1 dan galur indikator yang digunakan adalah Agrobacterium tumefaciens A4404 yang diperoleh dari Laboratorium Bioteknologi dan Pemuliaan Tanaman, Fakultas Pertanian, Universitas Andalas. Isolat uji digoreskan, kemudian galur indikator digoreskan lagi membentuk dua garis sejajar. Setelah biakan diinkubasi pada suhu 28 °C akan terbentuk zone warna biru.
Deteksi Exopolisaccharide Substance Stewartan
Deteksi exopolisaccharide substance stewartan (EPS) didasarkan pada metode Bodman dan Farrand (1995). Isolat uji ditumbuhkan pada medium casamino acid peptone glucose (CPG) dengan komposisi glukosa 1%, tripton 1%, casamino acid 0.1%, dan agar-agar 1.5%. Sel bakteri dari biakan diambil dan dimasukkan ke dalam 10 mL larutan NaCl 0.9%. Campuran larutan tadi dipindahkan ke dalam gelas piala dan diputar dengan cepat selama 1 menit menggunakan stirer, seterusnya disentrifugasi selama 30 menit. Supernatan dipisahkan dan dipindahkan ke dalam gelas piala lalu ditambahkan 60 mL etil alkohol, kemudian ditempatkan di dalam ruang pendingin selama 1 malam. Sisa pelet dicuci dengan 1 mL NaCl 0.9% per tabung. Semua ekstrak dipindahkan ke gelas piala lalu ditambahkan 60 mL etil alkohol dan disimpan selama 24 jam di dalam ruang pendingin. Ekstrak EPS diambil dan dicuci dengan air steril, seterusnya disimpan di dalamlemari pembeku sampai waktu yang diinginkan.
Uji AdhesiUji adhesi dilakukan berdasarkan pada
metode O’Toole dan Kolter (Koutsoudis et al. 2006). Isolat uji ditumbuhkan pada medium luria bertani (LB) yang mengandung glukosa 0.2%. Pengujian adhesi menggunakan kotak plastik polivinil klorida yang mempunyai 5 sumur. Isolat uji dicuci berulang kali dan diwarnai dengan kristal violet 1% selama 15 menit pada suhu kamar. Sisa zat warna dipisahkan melalui pencucian dengan akuades mengalir, sisa kristal violet dilarutkan dalam 150 µL etanol 95% yang ditambahkan pada setiap sumur. Pengujian ini ditujukan untuk mengetahui tegangan permukaan dari sel bakteri. Pada kesempatan ini hasil uji tidak dapat dilanjutkan karena tidak tersedianya tensiometer.
Uji PatogenisitasUji patogenisitas dilakukan pada bibit
jagung varietas Sweet Boy umur 8 hari. Isolat yang terdeteksi sebagai Gram negatif (-) dengan kerapatan 106 sel mL-1 diinokulasikan
47

J Fitopatol Indones Desi et al.
menggunakan jarum steril pada batang bibit kira-kira 5 cm dari permukaan tanah. Gejala kebasahan akan muncul setelah 3 hari dan bibit menunjukkan gejala layu antara 5-8 hari setelah inokulasi.
Uji Hipersensitivitas Uji hipersensitifitas dilakukan untuk
mengetahui kemampuan isolat dalam menimbulkan gejala respons hipersensitif. Isolat yang terdeteksi sebagai bakteri Gram negatif dengan kerapatan 108 sel mL-1 diinfiltrasikan menggunakan jarum steril pada daun tembakau, kemudian tanaman disungkup dengan kantong plastik. Gejala akan muncul dalam waktu 24 jam setelah infiltrasi.
Uji Polymerase Chain ReactionIsolat yang akan diuji ditumbuhkan pada
medium yeast dextrose carbonate (YDC) selama 48 jam pada suhu 23 °C, 5 µL suspensi sel bakteri di dalam akuades steril ditambah dengan 50 µL NaOH 0.05 M dan selnya dilisis pada penangas air yang mendidih selama 15 menit. Sebanyak 5 µL hasil lisis tersebut ditambahkan ke 45 µL campuran reaktan yang terdiri atas 1x bufer reksi, MgCb 1.5 mM, dNTP’s masing-masing 200 µM (Boehringer Mannheim), primer masing-masing 1 µM dan 2.50 unit AmpliTaq DNA polymerase. Amplifikasi DNA dilakukan menggunakan pasangan primer ES16: 5’-GCG AAC TTG GCA GAG AT-3’ dan ESIG2c: 5’-GCG CTT G TGT TAT GAG-3’ didahului dengan denaturasi awal selama 5 menit pada 94 °C, dilanjutkan sebanyak 35 siklus meliputi denaturasi selama 1 menit pada 94 °C, penempelan primer selama 1 menit pada 55 °C sintesis selama 2 menit pada 72 °C, pada tahapan terakhir ditambah 10 menit pada 72 °C. Hasil amplifikasi dielektroforesis menggunakan gel agaros 1% dalam bufer tris boric EDTA (TBE 0.5 x) dan divisualisasi menggunakan UV. Pola pita DNA masing-masing isolat diamati untuk mengetahui keragaman isolat. Selanjutnya dilakukan sikuensing terhadap isolat tersebut dan perunutan DNA menggunakan program NCBI BLAST (http://www.ncbi.nlm.nih.gov/BLAST).
HASIL
Gejala Penyakit di LapanganGejala penyakit yang ditemui pada
beberapa pertanaman jagung di Sumatra Barat sangat beragam, di antaranya ialah terdapat garis-garis tegas berwarna kuning sepanjang pertulangan daun, pinggiran daun bergelombang, tanaman kerdil, dan daun mengering lebih cepat.
Morfologi Isolat Bakteri Isolasi bakteri dari daun bergejala garis
kuning sepanjang tulang daun didapatkan 22 isolat murni. Pada medium NGA, bakteri mempunyai ciri bentuk koloni bulat, warna koloni kuning terang dan kuning, ukuran koloni berkisar antara 1 dan 4 mm, elevasi cembung, keadaan koloni berlendir, dan pinggiran koloni bervariasi (Tabel 1).
Uji FisiologiHasil uji Gram yang dilakukan terhadap
22 isolat murni diperoleh 10 isolat merupakan bakteri Gram negatif dan 10 isolat bakteri Gram positif, sementara dua isolat tidak diuji karena tidak menghasilkan pigmen kuning pada media NGA. Sebanyak 7 isolat bereaksi positif (+) pada uji katalase dan uji metabolisme glukosa tetapi bereaksi negatif (-) pada uji oksidasi. Oleh karena itu, uji lanjut hanya dilakukan terhadap 7 isolat ini (Tabel 2).
Uji Hipersensitivitas dan Patogenisitas Uji hipersensitivitas (HS) pada tanaman
tembakau, menunjukkan reaksi positif hanya pada 4 isolat, yaitu PR1, PP, ST1, dan LA, sedangkan 6 isolat lainnya menunjukkan reaksi negatif.
Uji patogenisitas yang dilakukan terhadap bibit jagung varietas Sweet Boy menghasilkan variasi gejala. Isolat KP1, KP3, dan PR2 menunjukkan gejala daun menguning mulai dari bawah, kemudian berwarna cokelat dan akhirnya tanaman layu dan mati. Isolat PT1, PR1, PP, LA, dan PT3 menunjukkan gejala water soaking dan pada hari ke-4 tanaman layu, sedangkan isolat ST1 dan ST3 menunjukkan
48
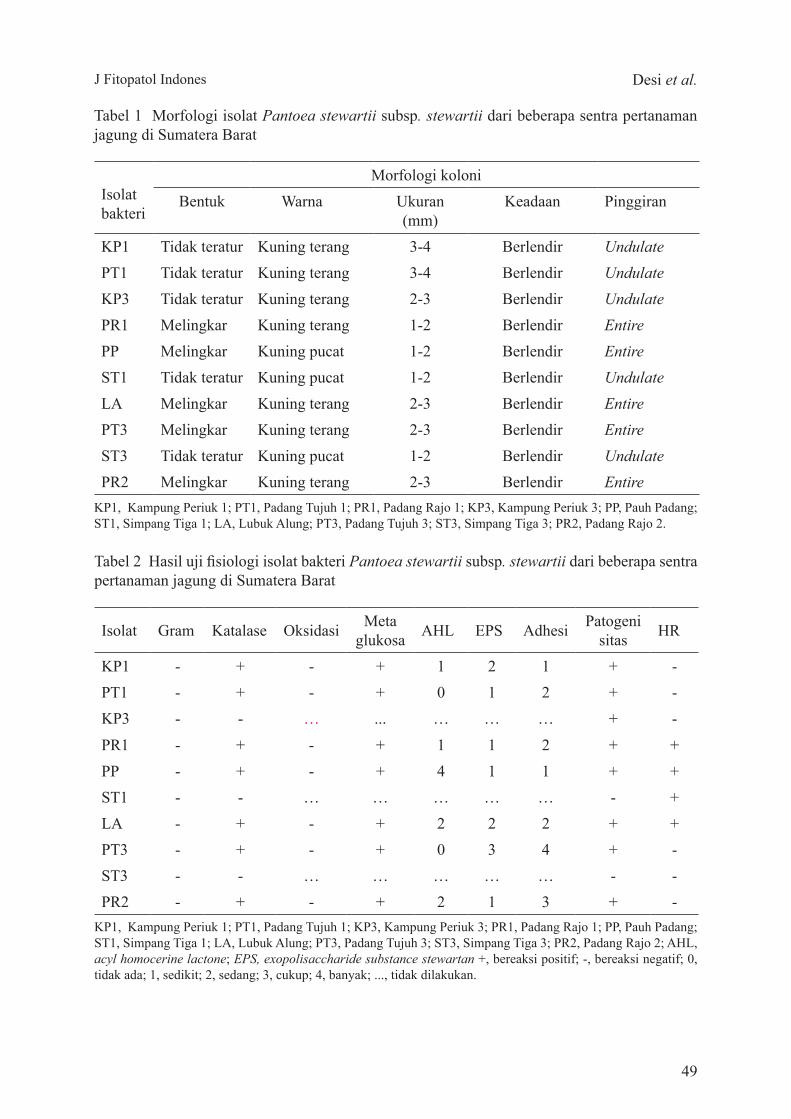
J Fitopatol Indones Desi et al.
49
Isolat bakteri
Morfologi koloniBentuk Warna Ukuran
(mm)Keadaan Pinggiran
KP1 Tidak teratur Kuning terang 3-4 Berlendir UndulatePT1 Tidak teratur Kuning terang 3-4 Berlendir UndulateKP3 Tidak teratur Kuning terang 2-3 Berlendir UndulatePR1 Melingkar Kuning terang 1-2 Berlendir EntirePP Melingkar Kuning pucat 1-2 Berlendir EntireST1 Tidak teratur Kuning pucat 1-2 Berlendir UndulateLA Melingkar Kuning terang 2-3 Berlendir EntirePT3 Melingkar Kuning terang 2-3 Berlendir EntireST3 Tidak teratur Kuning pucat 1-2 Berlendir UndulatePR2 Melingkar Kuning terang 2-3 Berlendir Entire
Tabel 1 Morfologi isolat Pantoea stewartii subsp. stewartii dari beberapa sentra pertanaman jagung di Sumatera Barat
KP1, Kampung Periuk 1; PT1, Padang Tujuh 1; PR1, Padang Rajo 1; KP3, Kampung Periuk 3; PP, Pauh Padang; ST1, Simpang Tiga 1; LA, Lubuk Alung; PT3, Padang Tujuh 3; ST3, Simpang Tiga 3; PR2, Padang Rajo 2.
Isolat Gram Katalase Oksidasi Meta glukosa AHL EPS Adhesi Patogeni
sitas HR
KP1 - + - + 1 2 1 + -PT1 - + - + 0 1 2 + -KP3 - - … ... … … … + -
PR1 - + - + 1 1 2 + +PP - + - + 4 1 1 + +ST1 - - … … … … … - +LA - + - + 2 2 2 + +PT3 - + - + 0 3 4 + -ST3 - - … … … … … - -PR2 - + - + 2 1 3 + -
Tabel 2 Hasil uji fisiologi isolat bakteri Pantoea stewartii subsp. stewartii dari beberapa sentra pertanaman jagung di Sumatera Barat
KP1, Kampung Periuk 1; PT1, Padang Tujuh 1; KP3, Kampung Periuk 3; PR1, Padang Rajo 1; PP, Pauh Padang; ST1, Simpang Tiga 1; LA, Lubuk Alung; PT3, Padang Tujuh 3; ST3, Simpang Tiga 3; PR2, Padang Rajo 2; AHL, acyl homocerine lactone; EPS, exopolisaccharide substance stewartan +, bereaksi positif; -, bereaksi negatif; 0, tidak ada; 1, sedikit; 2, sedang; 3, cukup; 4, banyak; ..., tidak dilakukan.

J Fitopatol Indones Desi et al.
gejala daun menguning yang dimulai dari bagian ujung, kemudian berwarna cokelat dan tanaman tidak layu.
Uji Polymerase Chain ReactionDari uji morfologi, fisiologi, dan
patogenisitas diperoleh tujuh isolat menunjukkan karakter Pantoea. Selanjutnya, identifikasi isolat tersebut menggunakan polymerase chain reaction (PCR) dengan pasangan primer ES16 dan ESIG2C (Gambar 3).
Hasil analisis urutan basa nukleotida menunjukkan bahwa tidak semua isolat tergolong genus Pantoea, hanya empat isolat diantaranya sebagai genus Pantoea dan tiga isolat sebagai spesies Pantoea stewartii yakni: isolat dari Padang Rajo 2, Lubuk Alung dan Pdang Rajo 1 memiliki kemiripan 98% asesi no. FJ118651, JQ6595401 dan JQ6595451 (Tabel 3).
PEMBAHASAN
Gejala penyakit layu stewart yang ditemukan pada beberapa sentra pertanaman jagung di Sumatera Barat sangat beragam. Gejala penyakit sesuai dengan yang dikemukakan oleh EPPO (2006), yaitu daun-daun bagian bawah awalnya memiliki garis-garis berwarna kuning. Penyakit ini berbeda dengan penyakit hawar yang umumnya menyebabkan warna daun-daunnya berkembang menjadi hijau pucat sampai kuning, terdapat garis longitudinal, dengan pinggiran tidak teratur atau bergelombang, dan dapat memanjang di sepanjang helaian daun, garis-garis ini mengering kemudian bewarna cokelat.
Uji katalase menunjukkan bahwa tujuh isolat menghasilkan gelembung atau bereaksi positif (+) dan tiga isolat bereaksi negatif (-).
50
Gambar 3 Hasil amplikasi beberapa isolat bakteri menggunakan primer 16S rNRA/ITS. M, 1 kb penanda DNA; PR2, Padang Rajo 2; LA, Lubuk Alung; PR1, Padang Rajo 1.
M PR2 M PR1L
1500 pb
Isolat Panjang sekuen DNA(pb)
Homologi sekuen 16S RNAPantoea stewartii
Homologi%
Kode aksesi GenBank
PR2 1408 galur ATCC 29923 98 FJ6118651LA 902 galur R1-104 96 JQ6595401PR1 775 galur R1-132 94 JQ6595451
Tabel 3 Analisis homologi sekuen DNA dari 3 isolat bakteri dari daun jagung bergejala penyakit layu stewart di sentra pertanaman jagung Sumatera Barat
PR2, Padang Rajo 2; LA, Lubuk Alung; PR1, Padang Rajo 1.

J Fitopatol Indones Desi et al.
51
Salah satu medium yang dapat digunakan untuk identifikasi bakteri genus Pantoea adalah medium Hugh-Leifson (HL), medium ini digunakan untuk mengetahui isolat yang mampu menghasilkan glukosa secara anaerobik (fermentasi). Pada tabung yang diberi vaseline, goresan isolat berwarna kuning artinya isolat bereaksi positif (+).
Bakteri Gram negatif tertentu menghasilkan signal dalam bentuk substitusi acyl homoserine lactone (acyl-HSLs) yang dikenal dengan autoinduser. Pesan-pesan ini memungkinkan populasi bakteri memonitor kepadatannya sendiri (querum sensing) untuk menggerakkan, mengkoordinasikan, dan menyatukan respons ketika ada kesempatan memarasit inang (Bodman dan Farrand 1995). Dalam pengujian deteksi AHL, hanya isolat PT1 dan PT3 yang tidak menghasilkan AHL. Dalam hal ini kemungkinan kedua isolat ini tidak bersifat patogen meskipun keduanya bersifat Gram negatif.
Deteksi EPS stewartan dilakukan untuk mengetahui kemampuan isolat dalam menghasilkan stewartan. Jumlah stewartan yang dihasilkan berkaitan dengan kemampuan isolat dalam menimbulkan gejala water soaking pada inang. Virulensi dari mutan P. stewartii mampu menyebabkan water soaking pada daun, juga mampu menyebabkan gejala layu sehingga diduga EPS merupakan faktor virulensi dari mutan ini. Demikian juga Minoque et al. (2005) menyatakan bahwa ekspresi EPS bergantung pada kepadatan sel bakteri yang akan menentukan perkembangan layu stewart. Koloni yang menghasilkan EPS bersifat kental, seperti lumpur, fluidal, dan permukaannya berlendir, sedangkan koloni yang kurang menghasilkan EPS bersifat tidak berlendir dan halus seperti krim (Bodman dan Farrand 1995).
Uji adhesi dilakukan untuk mengetahui lendir yang dihasilkan oleh bakteri. Semakin banyak lendir yang dihasilkan berarti semakin tinggi kemampuan bakteri untuk menempel pada inang. Bakteri uji ini menghasilkan mukus, yang menyumbat pembuluh vaskuler tanaman jagung sehingga akhirnya tanaman layu dan mati. Adanya adhesi pada bakteri
merupakan langkah penting dalam menentukan apakah bakteri bersifat parasit atau simbiotik (Koutsoudis et al. 2006).
Berdasarkan semua uji yang dilakukan, hanya 7 isolat, yaitu KP1, PT1, PR1, PP, LA, PT3, dan PR2 yang tergolong pada famili Enterobacteriaceae. Namun setelah dilakukan uji PCR, ternyata isolat PT1, PP, dan PT3 tidak tergolong ke dalam genus Pantoea, hal ini kemungkinan terjadi kesalahan dalam pengujian metabolisme glukosa.
UCAPAN TERIMA KASIH
Penelitian ini merupakan sebagian dari penelitian Hibah Bersaing yang dibiayai oleh DIPA Kopertis Wilayah X dengan Kontrak No. 0666/023-04.2.01/03/2012 tanggal 9 Desember 2011.
DAFTAR PUSTAKA
Bodman SB, Farrand SK. 1995. Capsular polysaccharide biosynthesis and pathogenicity In Erwinia stewartii require induction by An N-acylhomoserine lactone autoinducer. J Bacteriol. 177:5000–5008.
EPPO Buletin. 2006. Diagnostics antoea stewartii subsp. stewartii. Eur Med Plant Protect Org. 36(1):111–115.
Koutsoudis MD, Tsaltas D, Minoque TD, Suzanne VB. 2006. Quorum sensing regulation governs bacterial adhesion, biofilm development, and host colonization in Pantoea stewartii subsp. stewartii. Proc Natl Acad Sci. 103(15):5983–5988. DOI: http://dx.doi.org/10.1073/pnas.0509860 103.
Minoque TD, Carlier AL, Koutsoudis MD, Suzanne VB. 2005. The cell density-dependent expression of stewartan exopolysaccharide in Pantoea stewartii ssp. stewartii is a function of EsaR-mediated repression of the rcsA gene. Molecular Microbiology. 56(1):189–203. DOI: http://dx.doi.org/10.1111/j.1365-2958. 2004.04529.x
Rahma H. 2013. Kajian penyakit layu stewart pada jagung (Pantoea stewartii subsp.

J Fitopatol Indones Desi et al.
stewartii dan upaya pengendaliannya [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Taylor WI, Anhanzar D. 1972. Catalase test as an aid to identification of Enterobacteriaceae. App Microb. 24(1):58–61.
[TAS] Thai Agricultural Standard. 2008. Diagnostic Protocol for Pantoea stewartii subsp. stewartii Bacterial Wilt of Maize. Bangkok (TH): National Bureau of
Agricultural Commodity and Food Standards Ministry of Agriculture and Cooperatives.
Trisnawati F. 2010. Tingkat serangan Pantoea stewartii subsp. stewartii pada berbagai fase hawar dan layu stewartii pada jagung (Zea mays L.) di Kabupaten Pasaman Barat [skripsi]. Padang (ID): Universitas Andalas.
52

Volume 10, Nomor 2, April 2014Halaman 53–60
DOI: 10.14692/jfi.10.2.53
Analisis Keragaman Genetik Isolat Bakteri Xanthomonas oryzae pv. oryzae dari Jawa Barat dan Jawa Tengah Berdasarkan
Analisis ARDRA Gen 16SrRNA
Genetic Diversity Analysis of Bacterial Isolates Xanthomonas oryzae pv. oryzae from West Java and Central Java
Using ARDRA 16SrRNA Gene Analysis
Yadi Suryadi 1*, Dwi N. Susilowati1, Puji Lestari1, Sutoro1, Manzila Ifa1, Trini Suryani Kadir2
Sheryn Sunni Albani3, I Made Artika3
1Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor 16111 2Balai Besar Penelitian Tanaman Padi, Subang 41256
3Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Keragaman genetik lima belas isolat Xanthomonas oryzae pv. oryzae yang berasal dari beberapa daerah di Jawa Barat dan Jawa Tengah telah dianalisis menggunakan amplified ribosomal DNA restriction analysis (ARDRA) gen 16SrRNA. Pola ARDRA yang terbentuk dari hasil pemotongan dengan enzim restriksi RsaI menghasilkan sebanyak 13 pola yang menunjukkan keragaman genetika isolat X. oryzae pv. oryzae yang diuji cukup tinggi. Berdasarkan pohon filogenetika yang terbentuk, beberapa isolat memiliki kekerabatan yang cukup dekat. Isolat X. oryzae pv. oryzae 1/96 pml dan 61pml secara genetik cukup dekat dengan isolat 29d, 59pml, dan 60pml meskipun berasal dari daerah yang berbeda-beda. Isolat lain yang memiliki hubungan kekerabatan cukup dekat ialah isolat X. oryzae pv. oryzae 5mgl dengan isolat 23d, 28d, dan dan 10sbg, 8myd, serta isolat 6klt dengan isolat 3ind dan 2kr. Hasil keragaman genetika ini dapat dijadikan acuan dalam seleksi galur/isolat X. oryzae pv. oryzae untuk pengembangan tanaman padi tahan hawar daun bakteri di Indonesia
Kata kunci: filogenetika, hawar daun bakteri, keragaman genetika, padi
ABSTRACT
Fifteen Xanthomonas oryzae pv. oryzae isolates from several regions in West Java and Central Java have been studied for their genetic diversity using amplified ribosomal DNA restriction analysis (ARDRA) 16SrRNA gene.Total of 13 ARDRA patterns from digestion of DNA with RsaI restriction enzyme was obtained, indicated higher genetic diversity of X. oryzae pv. oryzae. Phylogenetic analysis revealed a close genetic relationships among isolates. Isolates X. oryzae pv. oryzae 1/96pml and 61pml are closely related with isolates 29d, 59pml, and 60pml although they are isolated from different locations. The other closely related isolates were between X. oryzae pv. oryzae 5mgl with 23d, 38d, 10sbg, 8myd; while 6klt was close to 3ind and 2kr. The result of X. oryzae pv. oryzae genetic diversity can be used as a reference for strain or/isolates selection for development ofbacterial leaf blight resistance in Indonesia
Key words: bacterial leaf blight, phylogenetic, restriction enzyme, rice
*Alamat penulis korespondensi: Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32, Indralaya, Ogan Ilir 30662 Tel: 0711-580663, Faks: 0711-580059, Surel: [email protected]
53
ISSN: 2339-2479

J Fitopatol Indones Suryadi et al.
PENDAHULUAN
Bakteri Xanthomonas oryzae pv. oryzae merupakan patogen penyebab penyakit hawar daun bakteri (HDB) pada padi. Serangan penyakit HDB menyebabkan turunnya produksi padi dan dapat menyebabkan kehilangan hasil panen. Tahun 2010 luas lahan yang terserang penyakit HDB sebesar 54 796 ha dan serangan ini meningkat pada masa tanam 2010-2011 menjadi sebesar 64 123 ha (Direktorat Perlindungan Tanaman Pangan 2011). Pengendalian penyakit HDB melalui pengembangan varietas tahan HDB merupakan salah satu cara yang efektif, murah, dan mudah diterapkan pada petani. Keragaman genetika yang luas pada bakteri X. oryzae pv. oryzae di berbagai daerah di Indonesia memerlukan pengembangan varietas padi yang tahan dan harus disesuaikan dengan penyebaran galur atau patotipenya.
Sampai saat ini paling tidak telah dilaporkan adanya 12 patotipe X. oryzae pv. oryzae di Indonesia yang berbeda variabilitas patogenisitasnya antar lokasi maupun musim sehingga menyulitkan upaya pengendalian X. oryzae pv. oryzae secara efektif. Dominasi populasi patotipe erat kaitannya dengan musim dan jenis varietas yang ditanam di lapangan (Sudir et al. 2009). Upaya pergiliran gen ketahanan (gene deployment) sangat bermanfaat untuk mengendalikan patogen X. oryzae pv. oryzae secara spesifik lokasi. Pengetahuan tentang penyebaran patotipe beserta variabilitasnya sangat penting dikaji untuk dijadikan pijakan perlunya dilakukan analisis keragaman genetik pada daerah yang diteliti.
Analisis keragaman genetik untuk karakterisasi patogen X. oryzae pv. oryzae dapat dilakukan menggunakan teknik molekuler karena cepat dan sensitif. Gen 16SrRNA dapat dilakukan dengan metode restriction fragment length polymorphism (RFLP) berdasarkan analisis pengukuran perbedaan panjang fragmen DNA yang dihasilkan oleh pemotongan enzim restriksi yang disebut amplified rDNA restriction analysis (ARDRA) (Schlegel et al. 2003).
Berdasarkan pola pita yang dihasilkan dari pemotongan enzim restriksi, pohon filogenetika dapat direkonstruksi dan dapat mengelompokkan bakteri berdasarkan jarak genetikanya.
Identifikasi keragaman genetika isolat bakteri X. oryzae pv. oryzae dari berbagai daerah di Indonesia, diharapkan dapat membantu pengembangan varietas padi tahan penyakit HDB. Tujuan penelitian ialah mengarakterisasi keragaman genetik 15 isolat X. oryzae pv. oryzae yang diperoleh dari beberapa daerah di Jawa Barat dan Jawa Tengah menggunakan analisis ARDRA.
BAHAN DAN METODE
Isolat Xanthomonas oryzae pv. oryzae dan Isolasi DNA
Sebanyak 15 isolat bakteri X. oryzae pv. oryzae yang berasal dari beberapa daerah di Jawa Barat dan Jawa Tengah (Tabel 1) diisolasi pada medium agar-agar Wakimoto (AW) (larutan sari kentang, 0.125 g Ca(NO3)2. 4H2O, 0.5 g Na2HPO4. 12H2O, 3.75 g sukrosa, dan 1.25 g pepton dengan pH 7-8). Isolat murni bakteri diremajakan pada agar-agar miring medium AW dan diinkubasi selama kurang 3-5 hari sampai muncul koloni berwarna kuning dan berlendir.
Isolasi DNA genom bakteri dilakukan menggunakan metode Lazo et al. (1987) yang dimodifikasi. Sebanyak 5 mL biakan sel bakteri dalam nutrient broth (NB) disentrifugasi pada kecepatan 8000 x g selama 10 menit pada suhu 4 °C . Pelet dicuci dan disuspensi kembali dengan 1 mL bufer STE, kemudian disentrifugasi kembali pada kecepatan 8000 x g selama 10 menit pada suhu 4 °C . Pelet disuspensi kembali dalam 200 µL bufer STE dan ditambahkan 40 µL larutan sodium dodecyl sulphate (SDS) 10%. Suspensi diinkubasi dalam oven pada suhu 65 °C selama 1.5 jam lalu didinginkan pada suhu ruang. Suspensi ditambah 4 µL Proteinase-K 10 mg mL-1, lalu diinkubasi dalam oven pada suhu 37 °C selama 4 jam. Suspensi ditambahi 200–400 µL bufer STE, selanjutnya ditambah larutan fenol dan
54

J Fitopatol Indones Suryadi et al.
kloroform, masing-masing sebanyak 120 µL. Tahap ini dilakukan secara perlahan sampai terbentuk emulsi. Suspensi yang telah teremulsi lalu disentrifugasi pada kecepatan 10 000 x g selama 15 menit. Supernatan dipindahkan ke tabung mikro steril dan dipresipitasi dengan menambahkan EtOH 95% dingin sebanyak 2 x volume supernatan. Suspensi selanjutnya diinkubasi pada suhu -20 °C selama 30 menit. Suspensi disentrifugasi pada kecepatan 8000 x g selama 2 menit lalu supernatannya dibuang. Pelet dikeringudarakan selama kurang lebih 2 jam dan disuspensikan kembali dalam 20 µL bufer TE yang mengandung 10 µg mL-1 RNase. Suspensi yang mengandung DNA ini disimpan pada suhu -20 °C untuk digunakan pada tahap selanjutnya.
Amplifikasi Gen 16S-rRNA dan Pemotongan dengan Enzim Restriksi RsaI
Sebanyak 2 µL DNA (20 ng/µL) ditambah dengan 10 µL nuclease free water. Volume total reaksi PCR sebanyak 20 µL. Campuran bahan untuk PCR terdiri atas 2 µL DNA, 12.5 µL GoTaq master mix, masing-masing 1 µL primer 63F (5’-CAG GCC TAA CAC ATG CAA GTC-3’) dan 1387R(5’-GGG CGG WGT GTA CAA GGC-3’) (Marchessi et al.1998) dan 8.5 µL ddH2O. Amplifikasi dilakukan pada kondisi suhu denaturasi awal 94 °C selama 5 menit dengan 1 siklus, denaturasi 94 °C selama 1 menit 30 detik, penempelan
55 °C selama 45 detik, dan pemanjangan 72 °C selama 1 menit dengan total 30 siklus, pemanjangan akhir 72 °C selama 1 menit dengan 1 siklus. Amplikon dielektroferesis pada gel agarosa 1.8% dalam bufer TAE 0.5x untuk memastikan adanya amplifikasi sebelum dilakukan pemotongan dengan enzim restriksi. Hasil elektroforesis direndam dengan etidium bromida dan divisualisasi menggunakan UV transiluminator.
Hasil amplifikasi gen 16SrRNA dari masing-masing sampel dipotong dengan enzim restriksi RsaI yang mempunyai situs pemotongan (5’-GTAC). Reaksi pemotongan terdiri atas 10 µL produk PCR, 18 µL ddH2O, 2 µL 10x bufer Tango, dan 1 µL enzim restriksi. Campuran tersebut diinkubasi pada suhu 37 °C selama ± 20 jam. Hasil pemotongan dielektroforesis pada gel agarosa dengan metode yang sama untuk visualisasi amplikon PCR. Hasil visualisasi dijadikan data biner terhadap pola pita yang terbentuk dan dibuat pohon filogenetikanya menginkuti program MVSP 3.2 dengan metode unweighted pair group method arithmetic mean (UPGMA).
HASIL
Amplikon Gen 16SrRNA Sebanyak 15 isolat X. oryzae pv. oryzae,
menghasilkan pita-pita DNA total yang dapat digunakan untuk amplifikasi PCR.
55
Tabel 1 Isolat Xanthomonas oryzae pv. oryzae asal tanaman padi dari beberapa wilayah di Jawa Barat dan Jawa Tengah
Isolat Asal varietas padi Asal lokasi1kr Inpari 18 Jatisari, Karawang2kr Inpari 18 Cikalong wetan, Karawang3ind C4 Gantar, Indramayu4mgl Mentikwangi Pabelan, Magelang5mgl Lokal Kaliduren, Doroyudan6klt Ciherang Kepanjen, Klaten8myd Ciherang Mayudan, Klaten10sbg Ciherang Sukamandi Jaya, Subang23d Inpari10 Pusakaratu, Subang28d Hipa6 Sukamandi,Subang29d Ciherang Ciasem, Subang59pml Si denok Comal, Pemalang60pml Ciherang Comal, Pemalang61pml Ciherang Pekuden, Pemalang1/96pml Ciherang Pemalang
J Fitopatol Indones Suryadi et al.
56
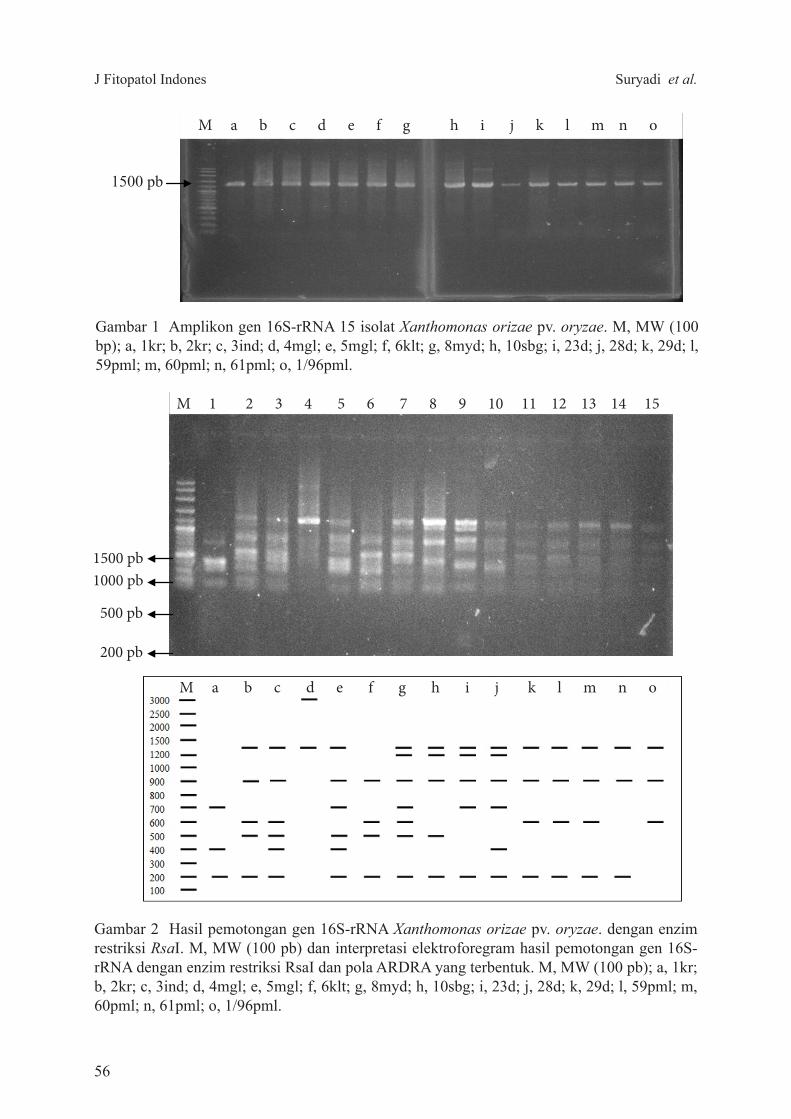
Gambar 1 Amplikon gen 16S-rRNA 15 isolat Xanthomonas orizae pv. oryzae. M, MW (100 bp); a, 1kr; b, 2kr; c, 3ind; d, 4mgl; e, 5mgl; f, 6klt; g, 8myd; h, 10sbg; i, 23d; j, 28d; k, 29d; l, 59pml; m, 60pml; n, 61pml; o, 1/96pml.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
M a b c d e f g h i j k l m n o
Gambar 2 Hasil pemotongan gen 16S-rRNA Xanthomonas orizae pv. oryzae. dengan enzim restriksi RsaI. M, MW (100 pb) dan interpretasi elektroforegram hasil pemotongan gen 16S-rRNA dengan enzim restriksi RsaI dan pola ARDRA yang terbentuk. M, MW (100 pb); a, 1kr; b, 2kr; c, 3ind; d, 4mgl; e, 5mgl; f, 6klt; g, 8myd; h, 10sbg; i, 23d; j, 28d; k, 29d; l, 59pml; m, 60pml; n, 61pml; o, 1/96pml.
1500 pb
M a b c d e f g h i j k l m n o
1500 pb
1000 pb
500 pb
200 pb

J Fitopatol Indones Suryadi et al.
DNA genomik yang dihasilkan mempunyai nilai nisbah kisaran kemurnian DNA 1.8– 2 pada panjang gelombang 260/280 yang menunjukkan kualitasnya cukup baik untuk digunakan pada proses PCR. Visualisasi pita DNA menunjukkan kemurnian DNA hasil isolasi yang diperoleh konsentrasinya cukup apabila diamplifikasi.
Gen 16SrRNA pada 15 isolat X. oryzae pv. oryzae berhasil diamplifikasi dengan PCR
sehingga menghasilkan produk PCR berupa pita tunggal yang berada pada kisaran 1400–1500 pb (Gambar 1).
Amplified rDNA Restriction Analysis (ARDRA)
Fragmen DNA hasil amplifikasi X. oryzae pv. oryzae yang dipotong dengan enzim restriksi RsaI menghasilkan 13 pola pita ARDRA yang ditandai dengan huruf a sampai m dan interpretasi elektroforegram hasil pemotongan dengan enzim restriksi RsaI (Gambar 2). Penghitungan ukuran pita hasil pemotongan dengan enzim restriksi dilihat berdasarkan marka yang digunakan. Ukuran pita terbesar 3000 pb terdapat pada kode isolat Xoo 4mgl, sedangkan pita terkecil 200 pb terdapat pada hampir setiap kode isolat (Tabel 2). Sebanyak 13 pola ARDRA yang terbentuk dari hasil pemotongan dengan enzim restriksi dijadikan data biner sebagai input data untuk pembentukan pohon filogenetika. Pohon filogenetika berdasarkan pola pita ARDRA yang terbentuk pada cut off 0.3 kb menunjukkan dua klaster utama dari 15 isolat X. oryzae pv. oryzae yang diuji. Ada 6 isolat X. oryzae pv. oryzae yang lebih dekat secara genetika dalam klaster pertama (I), yaitu 61pml, 4mgl, 59pml, 1/96pml, 60pml, dan 29d. Sebagian besar isolat teridentifikasi dalam klaster II,
57
Tabel 2 Pengelompokan ukuran pola pita hasil pemotongan dengan enzim restriksi RsaI
Pola pita
Ukuran basa (pb)
a 200, 700, 400b 200, 500, 600, 900, 1300-1400c 200, 400, 500, 600, 900, 1300-1400d 1300-1400, 3000e 200, 400, 500, 700, 900, 1300-1400f 200, 500, 600, 900g 200, 500, 600, 700, 900, 1200, 1300-1400h 200, 500, 900, 1200, 1300-1400i 200, 700, 900, 1200, 1300-1400j 200, 400, 700, 900, 1200, 1300-1400k 200, 600, 900, 1300-1400l 200, 900, 1300-1400m 600, 900, 1300-1400
Gambar 3 Pohon filogenetika Xanthomonas orizae pv. oryzae berdasarkan pola pita DNA hasil pemotongan dengan enzim restriksi RsaI.
0.04 0.2 0.36 0.52 0.68 0.84 1
Koefisien Jaccard’s

J Fitopatol Indones Suryadi et al.
yaitu sebanyak 9 isolat meliputi 1kr, 2kr, 23d, 3ind, 10sbg, 28d, 5mgl, 8myd, 6klt. Ada 3 isolat X. oryzae pv. oryzae, yaitu 29d, 60pml dan 1/96pml, memiliki kesamaan pola pita ARDRA (Gambar 3) seperti ditunjukkan oleh kedekatan genetika dalam satu klaster.
PEMBAHASAN
Salah satu teknik yang akurat untuk identifikasi molekuler pada mikroorganisme prokariot khususnya bakteri ialah identifikasi terhadap gen penyandi 16SrRNA yang dikenal juga dengan sebutan ribotyping. Identifikasi tersebut didasarkan pada tingkat kesamaan dalam urutan gen 16SrRNA sebagai sidik jari genetik bakteri atau disebut urutan sidik jari. Beberapa keuntungan dalam analisis terhadap gen penyandi 16SrRNA ini ialah RNA secara umum dimiliki semua bakteri dan merupakan unit yang konstan (Woese et al. 1983). Jika terdapat kemiripan atau sedikit perbedaan basa pada urutan nukleotida gen 16SrRNA dari dua jenis organisme maka kedua organisme tersebut memiliki hubungan kekerabatan yang dekat ditinjau dari kedekatan secara evolusinya (Pangastuti 2006).
Sampai saat ini analisis keragaman X. oryzae pv. oryzae asal Indonesia telah diobservasi menggunakan beberapa pendekatan molekuler (Tasliah et al. 2013). Mengingat kelebihan teknik ARDRA untuk karakterisasi molekuler berbagai bakteri (Susilowati et al. 2010) maka identifikasi molekuler dalam menganalisis keragaman genetik X. oryzae pv. oryzae pada studi ini menggunakan identifikasi terhadap gen penyandi 16SrRNA yang dilanjutkan dengan pemotongan enzim restriksi. Sebanyak 15 isolat X. oryzae pv. oryzae dari beberapa sentra daerah padi di Jawa Barat dan Jawa Tengah, berhasil diamplifikasi gen penyandi 16SrRNAnya menggunakan primer 63F dan 1387R. Proses amplifikasi ini menghasilkan produk pada kisaran ukuran 1400–1500 pb (Gambar 1). Hampir setiap isolat menghasilkan pita tunggal yang cukup tebal, kecuali isolat X. oryzae pv. oryzae 1/96pml. Pita yang tebal tersebut menunjukkan bahwa gen 16SrRNA isolat teramplifikasi dengan baik. Hal ini
dapat disebabkan oleh beberapa faktor yang mempengaruhi PCR, optimasi amplifikasi PCR dari DNA tiap isolat dan kesesuaian primer yang digunakan. Primer yang digunakan adalah primer spesifik 63F dan 1387R. Primer 63F memiliki panjang urutan basa 21 nukleotida dan 1387R sebanyak 18 nukleotida. Primer ini tidak membentuk struktur dupleks dengan ujung 5’ yang dapat dikenali enzim eksonuklease dan tidak terdapat nukleotida yang terpotong pada ujung 5’. Hal ini dapat mempengaruhi suhu penempelan primer (Marchessi et al. 1998) sehingga dapat membantu proses amplifikasi DNA.
ARDRA merupakan teknik RFLP yang digunakan untuk mengetahui perbedaan jenis bakteri, misalnya berdasarkan gen ribosomal DNA seperti 16SrRNA. Teknik ini menggunakan enzim restriksi untuk melihat polimorfisme dalam genom organisme. Situs enzim restriksi dari genom suatu kelompok organisme yang kemudian berubah karena mutasi atau berpindah karena genetic rearrangement dapat menyebabkan situs tersebut tidak lagi dikenali oleh enzim atau enzim restriksi akan memotong daerah lain yang berbeda. Proses ini menyebabkan terbentuknya fragmen-fragmen DNA yang berbeda ukurannya antara satu organisme dengan organisme lainnya. Polimorfisme ini selanjutnya digunakan sebagai dasar untuk membuat pohon filogenetika atau dendogram kekerabatan kelompok (Sasaki et al. 2004).
Enzim restriksi yang digunakan untuk memotong produk PCR 16SrRNA dari 15 isolat X. oryzae pv. oryzae, yaitu RsaI. Enzim RsaI merupakan enzim endonuklease restriksi tetramerik atau memiliki sisi pengenalan empat basa sehingga bila penyebaran nukleotida terjadi secara acak pada organisme dengan G+C sekitar 50% maka sekitar 256 pb dapat diharapkan memiliki satu sisi pengenalan. Enzim restriksi tersebut menghasilkan sejumlah besar fragmen yang bervariasi ukurannya (Susilowati et al. 2010).
Enzim restriksi RsaI memotong dengan sifat blunt end pada situs 5’...GT↓AC...3’ dan 3’...CA↑TG...5’. Pemotongan dengan RsaI pada isolat X. oryzae pv. oryzae yang
58

J Fitopatol Indones Suryadi et al.
diuji menghasilkan 13 pola pita ARDRA yang berbeda. Pengelompokan tersebut berdasarkan kesamaan di situs pemotongan enzim restriksi RsaI pada setiap spesies. Isolat X. oryzae pv. oryzae dengan kode 29d, 59pml, dan 60pml memiliki hasil pola pita yang sama sehingga dikelompokkan dalam satu pola pita. Kesamaan pola yang dihasilkan isolat X. oryzae pv. oryzae 59pml dan 60pml tersebut dapat disebabkan oleh kesamaan daerah asal kedua isolat tersebut, yaitu Pemalang. Isolat Xoo29d berasal dari Subang, namun memiliki pola pita yang sama dengan isolat 59pml dan 60pml. Hal ini dapat diartikan bahwa ketiga isolat tersebut memiliki kekerabatan yang sangat dekat meskipun berasal dari daerah yang berbeda.
Hasil penelitian menunjukkan adanya keragaman genetika antar isolat yang diuji meskipun isolat diperoleh dari daerah yang sama atau berbeda lokasi khususnya di Jawa Barat dan Jawa Tengah. Dengan demikian, pergiliran tanaman akan semakin kompleks mengingat dominansi patotipe X. oryzae pv. oryzae akan berbeda antar lokasi di Indonesia. Keragaman genetik ini dilaporkan juga terjadi di Filipina oleh Ardales et al. (1996) yang melaporkan adanya perbedaan genetika yang cukup besar antar struktur populasi X. oryzae pv. oryzae yang diperoleh pada tingkat agroekosistem, area pengambilan contoh antar lapangan dan jenis kultivar.
Berdasarkan pohon filogenetika yang terbentuk, X. oryzae pv. oryzae isolat 1/96pml dan 61pml terdapat dalam satu percabangan dengan isolat 29d, 59pml, dan 60pml. Isolat 1/96pml dan 61pml berasal dari daerah yang sama dengan isolat 59pml dan 60pml, yaitu Pemalang. Meskipun memiliki pola pita yang berbeda, namun dapat diartikan bahwa isolat 1/96pml dan 61pml memiliki kekerabatan yang cukup dekat dengan isolat 29d, 59pml, dan 60pml. Selanjutnya, isolat X. oryzae pv. oryzae 5mgl (Doroyudan) memiliki satu percabangan dengan isolat 23d, 28d, 10sbg, dan 8myd. X. oryzae pv. oryzae 23d dan 28d berada dalam satu percabangan kecil. Kedua isolat tersebut berada dalam daerah yang berbeda, namun memiliki kekerabatan yang
cukup dekat. X. oryzae pv. oryzae 10sbg dan 8myd juga berada dalam satu percabangan kecil yang lain. Kedua isolat X. oryzae pv. oryzae ini juga berasal dari daerah yang berbeda, namun memiliki kekerabatan yang cukup dekat. Percabangan lain yang terbentuk, yaitu isolat X. oryzae pv. oryzae 6klt, 3ind, dan 2kr. Isolat 6klt berasal dari Klaten, 3ind dari Indramayu, dan 2kr dari Karawang. Ketiga isolat ini memiliki kekerabatan yang juga cukup dekat meskipun berasal dari daerah yang berbeda. Isolat Xoo1kr yang berasal dari daerah yang sama dengan isolat 2kr (Karawang) terdapat dalam percabangan yang berbeda, namun masih dalam satu klaster besar yang sama. Isolat X. oryzae pv. oryzae 4mgl (Pabelan, Magelang) berada dalam klaster yang berbeda dengan isolat lain yang diuji. Hal ini menunjukkan kekerabatan isolat 4mgl dengan yang lainnya cukup jauh. Hasil ini dapat memudahkan dalam deteksi X. oryzae pv. oryzae di lapangan dan sekaligus untuk pengendalian penyakit (Onasanya et al. 2010).
Menurut Schlegel et al. (2003) tidak terdapat keragaman pada intraspesies dan situs pemotongan yang terkonservasi pada spesies yang berdekatan menunjukkan keragaman yang rendah. Hal ini berbanding terbalik dengan hasil keragaman yang terbentuk dari X. oryzae pv. oryzae yang diuji. Hasil pohon filogenetika yang terbentuk menunjukkan keragaman yang cukup tinggi pada isolat X. oryzae pv. oryzae yang diuji meskipun berada dalam spesies yang sama. Keragaman genetik yang cukup tinggi dari X. oryzae pv. oryzae yang diuji dalam studi ini mungkin disebabkan perbedaan daerah asal setiap isolat atau telah terjadi mutasi dalam sel inang yang berbeda (Mongkolsuk et al. 2000). Berdasarkan penelitian ini terlihat pula adanya campuran patotipe yang diperoleh dari daerah yang sama yang mengimplikasikan perlunya upaya pergiliran menggunakan varietas tahan berlatar belakang gen tahan kombinasi (piramiding gen). Dalam rangka pengendalian HDB, hasil analisis keragaman genetik X. oryzae pv. oryzae ini penting artinya dan diperlukan untuk membantu mengembangkan
59

J Fitopatol Indones Suryadi et al.
varietas padi tahan HDB secara spesifik lokasi di Indonesia.
Sebanyak 15 isolat X. oryzae pv. oryzae yang diuji yang berasal dari beberapa daerah di Jawa Barat dan Jawa Tengah, secara genetika menunjukkan keragaman yang cukup tinggi. Berdasarkan analisis gen 16SrRNA, diperoleh sebanyak 13 pola pita ARDRA yang berbeda. Analisis pohon filogenetika menunjukkan bahwa antara isolat X. oryzae pv. oryzae yang berasal dari daerah yang berbeda dapat memiliki kekerabatan yang cukup dekat secara genetik.
UCAPAN TERIMA KASIH
Penulis menyampaikan ucapan terima kasih kepada APBN DIPA RPTP Dr Sutoro No Proyek.1798.009.011 yang telah membiayai penelitian ini.
DAFTAR PUSTAKA
Ardales EY, Leung H, Cruz CMV, Mew TW, Leach JE. 1996. Hierarchical analysis of spatial variation of the rice bacterial blight pathogen across agroecosystems in the Philippines. Phytopathology. 86:241–252. DOI: http://dx.doi.org/10.1094/Phyto-86-241.
[DPTP]. Direktorat Perlindungan Tanaman Pangan. 2011. Prakiraan serangan BLB pada padi di Indonesia masa tanam Tahun 2011. Jakarta (ID): Ditjen TP.
Lazo GR, Roffey R, Gabriel DW. 1987. Pathovars of Xanthomonas campestris are distinguishable by restriction fragment-length polymorphism. IJSEM. 37(3):214–221.
Marchessi JR, Sato T, Weightman AJ, Martin TA, Fry JC, Hiom SJ, Wade WG. 1998. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl Environ Microbiol. 64:795–799.
Mongkolsuk S, Wangshuk W, Tuangthong M, Loprasert S. 2000. Mutation in oxyr resulting in peroxide resistance in Xanthomonas campestris. J Bacteriol.
182:3846–3849. DOI: http://dx.doi.org/10. 1128/JB.182.13.3846-3849.2000.
Onasanya A, Basso A, Somado E, Gasore ER, Nwilene FE, Ingelbrecht I, Lamo J, Wydra K. 2010. Development of combined molecular diagnostic and DNA fingerprinting technique for rice bacteria pathogens in Africa. Biotechnol. 9:89–105. DOI: http://dx.doi.org/10.3923/biotech. 2010.89.105.
Pangastuti A. 2006. Definisi spesies prokaryota berdasarkan urutan basa gen penyandi 16S rRNA dan gen penyandi protein. Biodiversitas. 3(7):292–296.
Sasaki E, Osawa R, Nishitani Y, Whiley RA. 2004. ARDRA and RAPD analysis of human and animal isolates of Streptococcus gallolyticus. J Vet Med Sci. 66:1467–1470. DOI: http://dx.doi.org/10.1292/jvms.66. 1467.
Schlegel L, Grimont F, Grimont PAD, Bouvet A. 2003. Identification of major Streptococcal species by rrn-amplified ribosomal DNA restriction analysis. J Clin Microbiol. 41:657–666. DOI: http://dx.doi.org/10.1128/JCM.41.2.657-666.2003.
Sudir, Suprihanto, Triny S. Kadir. 2009. Identifikasi Patotipe Xanthomonas oryzae pv. oryzae, Penyebab Penyakit Hawar Daun Bakteri di Sentra Produksi Padi di Jawa. Penelitian Pertanian Tanaman Pangan. 28(3):131–138.
Susilowati DN, Nurul H, Tasliah, Mulya K. 2010. Keragaman bakteri endofitik diisolasi dari empat varietas padi dengan metode ARDRA. Berita Biologi 10(2): 241–248.
Tasliah, Mahrup, Prasetiyono J. 2013. Identifikasi molekuler hawar daun bakteri (X. oryzae pv oryzae) dan uji patogenisitasnya pada galaur-galur padi isogenik. J Agrobiogen. 9(2):49–57.
Woese CR, Guttel R, Gupta R, Woller H. 1983. Detailed analysis of the higher order structure of the16S-like ribosomal ribonucleic acids. Microbiol Rev. 47:621–669.
60

Volume 10, Nomor 2, April 2014Halaman 61–67
DOI: 10.14692/jfi.10.2.61
Keefektifan Bakteri Endofit dan Bakteri Perakaran Pemacu Pertumbuhan Tanaman dalam Menekan
Penyakit Layu Bakteri pada Tomat
The Effectiveness of Endophytic Bacteria and Plant Growth Promoting Rhizobacteria as Biocontrol
Agents of Tomato Bacterial Wilt Disease
Zhenita Vinda Tri Handini, Abdjad Asih Nawangsih* Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Penyakit layu bakteri yang disebabkan Ralstonia solanacearum merupakan penyakit penting pada tanaman tomat. Potensi bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman sebagai agens hayati untuk mengendalikan penyakit telah banyak dilaporkan. Penelitian dilakukan untuk menentukan keefektifan bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman sebagai agens hayati untuk penyakit layu bakteri pada tomat. Dua isolat bakteri perakaran pemacu pertumbuhan tanaman, Pseudomonas fluorescence RH4003, Bacillus subtilis AB89, dan 4 isolat bakteri endofit, BC10, B. amyloliquefaciens, dan Staphylococcus epidermidis BC4, diaplikasikan secara tunggal atau bersama-sama (perlakuan kombinasi) dengan metode penyiraman suspensi pada bibit tomat. Tanaman yang diberi perlakuan kombinasi bakteri P. fluorescens RH4003 dengan bakteri endofit isolat BC10 (F1*BC10) menunjukkan pertambahan tinggi yang paling besar, walaupun bobot kering tanaman antar perlakuan tidak berbeda nyata. Perlakuan S. epidermidis BC4 secara tunggal menyebabkan insidensi paling rendah dan berbeda nyata dengan perlakuan kontrol.
Kata kunci: Bacillus subtilis, B. amyloliquefaciens, pengendalian hayati, Pseudomonas fluorescens, Staphylococcus epidermidis
ABSTRACT
Bacterial wilt disease caused by Ralstonia solanacearum is an important disease on tomato. Endophytic bacteria and plant growth promoting rhizobacteria has been reported as potential biocontrol agent for many plant diseases. Research was conducted to determine the effectiveness of some isolates of endophytic bacteria (BC10, Bacillus amyloliquefaciens, dan Staphylococcus epidermidis BC4) and plant growth promoting rhizobacteria (Pseudomonas fluorescence RH4003, Bacillus subtilis AB89) as biocontrol agents for tomato wilt disease. Bacterial isolates was applied as soil drench on tomato seedling in single or combination treatments. Combination of P. fluorescens RH4003 and isolate BC10 (F1*BC10) enhanced the highest plant height, although plant dry mass among treatments was not significantly different. Single application of S. epidermidis BC4 was significantly suppressed disease incidence.
Key words: Bacillus subtilis, B. amyloliquefaciens, biological control, Pseudomonas fluorescens, Staphylococcus epidermidis
*Alamat Penulis Korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Kampus Darmaga, Jalan Kamper, Bogor 16680Tel: 0251-8629364, Faks: 0251-8629362, Surel: [email protected]
61
ISSN: 2339-2479

J Fitopatol Indones Handini dan Nawangsih
PENDAHULUAN
Penyakit layu bakteri merupakan salah satu penyakit penting pada tomat yang dapat menurunkan hasil panen. Gejala layu bakteri ialah antara lain munculnya akar adventif pada batang jika kondisi lingkungan kurang menguntungkan bagi patogen dan adanya oose bakteri yang berwarna keabu-abuan atau seperti benang-benang putih halus yang keluar dari batang yang dipotong.
Beberapa metode pengendalian penyakit layu bakteri yang telah dilakukan diantaranya ialah penanaman varietas tahan, penggunaan pestisida, rotasi tanaman, sanitasi, serta pengendalian menggunakan agens hayati yang bersifat antagonis terhadap patogen. Bakteri endofit dan bakteri perakaran yang bersifat menginduksi pertumbuhan tanaman dilaporkan memiliki potensi untuk digunakan sebagai agens hayati pada beberapa kasus penyakit tanaman (Sharma et al. 2005; Vega et al. 2005). Pengendalian penyakit layu bakteri pada tomat dengan menggabungkan bakteri endofit dan perakaran pemacu pertumbuhan tanaman belum pernah dilakukan sebelumnya. Sampai saat ini pengendalian dilakukan hanya dengan mengaplikasikan bakteri endofit atau perakaran pemacu pertumbuhan tanaman secara terpisah.
Bakteri endofit merupakan mikrob yang hidup dalam jaringan tanaman pada periode tertentu dan mampu hidup dengan membentuk koloni dalam jaringan tanaman tanpa membahayakan inangnya dan akan bersaing dengan patogen (Reiter et al. 2002). Penggunaan mikrob endofit memiliki beberapa keuntungan dibandingkan dengan penggunaan mikrob antagonis lain, di antaranya ialah mikrob endofit akan tetap ada atau bertahan selama perkembangan tanaman dan terus memberikan perlindungan bagi tanaman.
Bakteri akar perakaran pemacu pertumbuhan tanaman telah banyak diteliti untuk mengendalikan layu bakteri. Pseudomonas fluorescens dan Bacillus subtilis yang merupakan bakteri perakaran pemacu pertumbuhan tanaman dilaporkan dapat mengendalikan penyakit layu bakteri pada tomat (Nawangsih 2006).
Penelitian ini dilakukan dengan tujuan untuk menguji keefektifan kombinasi bakteri endofit dengan bakteri akar perakaran pemacu pertumbuhan tanaman dalam menekan perkembangan penyakit layu bakteri pada tomat secara in planta.
BAHAN DAN METODE
Tanaman UjiMedium tanam yang digunakan dalam
pembibitan merupakan campuran kompos dan sekam dengan perbandingan 1:1. Benih tomat disemai dan setelah berumur 3 minggu bibit tomat dipindahkan ke dalam kantong plastik berisi medium tanam untuk aplikasi di rumah kaca dan di kebun percobaan.
Medium tanam yang digunakan untuk uji bakteri endofit dan bakteri akar bakteri pemacu pertumbuhan tanaman terdiri atas tanah steril, kompos steril, dan tanah yang telah terinfestasi bakteri R. solanacearum. Medium tanam tersebut dimasukkan ke dalam kantong plastik berdiameter 20 cm dengan susunan dari bawah yang terdiri atas tanah steril dengan ketebalan ± 6 cm, tanah terinfestasi R. solanacearum dengan ketebalan ± 9 cm, lalu ditutup lagi dengan tanah steril dengan ketebalan ± 10 cm.
Inokulum Bakteri Endofit, PPT, dan R. solanacearum
Bakteri endofit (P. fluorescens RH4003 dan B. subtilis AB89), bakteri perakaran pemacu pertumbuhan tanaman (isolat BC4, BC10 dan BL10) dan bakteri patogen R. solanacearum yang digunakan adalah koleksi Laboratorium Bakteriologi Tumbuhan, Fakultas Pertanian, IPB. Bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman diremajakan pada medium agar-agar nutrien (AN) dalam cawan petri dan R. solanacearum diremajakan pada medium agar-agar King’s B Agar. Bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman yang digunakan dalam percobaan ini selanjutnya disebut sebagai bakteri agens hayati.
Suspensi bakteri yang disimpan pada suhu -20 °C didiamkan sampai mencair, kemudian diambil sebanyak 100 μL dan disebarkan pada permukaan medium AN dalam cawan petri.
62

J Fitopatol Indones Handini dan Nawangsih
Bakteri agens hayati yang tumbuh selanjutnya disiram dengan 5 mL air steril dan digosok menggunakan jarum ose hingga terbentuk suspensi. Sebanyak 1 mL suspensi tersebut diinokulasikan ke dalam 250 mL medium cair nutrien dan digoyang menggunakan alat pengocok selama 24–48 jam.
Uji Pengaruh Bakteri Pertumbuhan Tanaman
Bibit tomat berumur 3 minggu disiram akarnya dengan suspensi agens hayati sesuai perlakuan. Untuk perlakuan tunggal setiap bibit disiram dengan 50 mL suspensi agens hayati dan untuk perlakuan kombinasi disiram sebanyak 25 mL dari masing-masing agens hayati yang dikombinasikan. Untuk perlakuan kontrol tanaman sehat, bibit tomat disiram dengan 50 mL air. Perlakuan yang diberikan, yaitu K, perlakuan patogen tetapi tanpa agens hayati (Kontrol); P. fluorescens RH4003 (F1); B. subtilis AB89 (A); S. epidermidis BC4 (B); bakteri endofit BC10 (BC10); bakteri endofit BL10 (BL10); P. fluorescens RH4003 + S. epidermidis BC4 (F1* B); P. fluorescens RH4003 + bakteri endofit BC10 (F1*BC10); P. fluorescens RH4003 + bakteri endofit BL10 (F1*BL10); B. subtilis AB89 + S. epidermidis BC4 (A* B); B. subtilis AB89 + bakteri endofit BC10 (A* BC10); B. subtilis AB89 + bakteri endofit BL10 (A* BL10).
Bibit tomat yang telah diberi perlakuan selanjutnya dipindah ke dalam kantong plastik dengan medium tanam yang telah mengandung R. solanacearum. Percobaan disusun dalam rancangan acak kelompok dengan ulangan sebanyak 3 kali sebagai kelompok. Dalam satu unit perlakuan terdiri atas 5 tanaman.
Penyiraman dalam rangka pemeliharaan dilakukan sesuai kebutuhan. Pengamatan terhadap pertambahan tinggi tanaman dan penghitungan nilai area under height of plant growth curve (AUHPGC) dilakukan setiap minggu hingga tanaman berumur 5 minggu setelah tanam (MST). Nilai AUHPGC dihitung menggunakan rumus Van der Plank (1963) yang dimodifikasi, yaitu menggunakan data pertambahan tinggi tanaman.
Y, insidensi penyakit pada pengamatan minggu ke-i; t, waktu (hari).
Pengujian Pengaruh Bakteri Agens Hayati terhadap Penyakit Layu Bakteri pada Tomat di Kebun Percobaan
Bibit tomat yang telah berumur 3 minggu disiram dengan agens hayati sebanyak 100 mL untuk aplikasi tunggal dan 50 mL dari masing-masing agens hayati untuk aplikasi kombinasi. Tanaman perlakuan kontrol sehat disiram dengan 100 mL air. Setelah disiram, bibit dipindahkan ke kebun percobaan yang sudah terinfestasi R. solanacearum.
Lahan percobaan dibagi menjadi 3 blok. Dalam satu blok terdiri atas 12 bedeng per unit perlakuan yang ditanami dengan 20 bibit tomat, setiap lubang tanam terdiri atas satu bibit tomat. Setiap ulangan terdiri atas 12 plot sehingga dalam satu lahan terdiri atas 36 plot per petak.
Pengamatan dilakukan setiap 2 minggu selama 7 minggu dan peubah yang diamati ialah insidensi penyakit layu bakteri. Insiden penyakit (IP) dihitung dengan rumus yang disampaikan oleh Unterstenhofer (1963):
IP, insidensi penyakit; n, jumlah tanaman yang terserang, N, jumlah tanaman yang diamati.
Selanjutnya AUDPC dihitung dengan rumus Van der Plank (1963):
Y, insidensi penyakit; t, hari.Penghitungan terhadap indeks penekanan penyakit (keefektifan pengendalian) dilakukan menggunakan rumus:
DIc, insidensi penyakit pada perlakuan kontrol; DIb, insidensi penyakit pada perlakuan agens hayati.
Analisis data Data yang diperoleh dianalisis
menggunakan analisis ragam dengan menggunakan program SAS versi 9.1 dan dilanjutkan dengan uji Duncan pada taraf 5%.
Indeks penekanan penyakit =DIc - DIb
DIc x 100%
63
AUHPGC= ∑ ( ) n
i=1Yi +Yi+i
2(ti+1-ti)
, denganAUDPC= ∑ ( ) n 1
iYi +Yi+i
2(ti+1-ti)
n N
IP = x 100%, dengan

J Fitopatol Indones Handini dan Nawangsih
HASIL Pertambahan Tinggi Tanaman Tomat, Masa Inkubasi, dan Bobot Kering Tanaman
Tanaman yang diberi perlakuan kombinasi bakteri P. fluorescens RH4003 dengan bakteri endofit isolat BC10 (F1*BC10) menunjukkan nilai AUHPGC paling tinggi, yaitu sebesar 12.6 unit dibandingkan dengan perlakuan kontrol, maupun perlakuan lainnya (Gambar 1). Bobot kering tanaman pada semua perlakuan tidak berbeda nyata dibandingkan dengan perlakuan kontrol (Tabel 1).
Indeks Penyakit Layu Bakteri di Kebun Percobaan
Aplikasi agens hayati S. epidermidis BC4 secara tunggal (B) dan campuran antara B. subtilis AB89 dengan B. amyloliquefaciens (A*BL10) menghasilkan nilai AUDPC paling rendah dibandingkan dengan perlakuan lain termasuk kontrol (K) (Gambar 2).
Pada pengamatan 1 MST, insidensi penyakit pada tanaman yang diberi perlakuan P. fluorescens RH4003 (F1) secara tunggal dan campuran antara B. subtilis AB89 dan B. amyloliquefaciens (A*BL10) nyata lebih rendah dibandingkan dengan kontrol. Insidensi penyakit pada pengamatan 7 MST yang paling rendah terjadi pada perlakuan S. epidermidis
BC4 (B), yaitu sebesar 71.7%. Nilai tersebut nyata lebih rendah jika dibandingkan dengan insidensi penyakit pada kontrol, yaitu sebesar 90.0% (Tabel 2).
Perlakuan Bobot kering tanaman* (g)F1 15.9 ± 7.7 aB12 12.4 ± 3.1 aBC4 12.7 ± 7.0 aBC10 11.6 ± 0.8 aBL10 10.8 ± 2.1 aF1*BC4 14.9 ± 9.9 aF1*BC10 9.4 ± 3.8 aF1*BL10 11.7 ± 2.6 aB12*BC4 12.4 ± 3.0 aB12*BC10 12.6 ± 1.2 aB12*BL10 11.3 ± 2.4 aK 13.7 ± 2.8 a
Tabel 1 Rerata bobot kering tanaman tomat pada berbagai perlakuan bakteri endofit dan bakteri akar pemacu pertumbuhan secara tunggal dan kombinasi di rumah kaca
F1, Pseudomonas fluorescens RH4003; B12, Bacillus subtilis AB89; BC4, Staphylococcus epidermidis; BC10, bakteri endofit isolat BC10; BL10, B. amyloliquefaciens; K, kontrol.*Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata berdasarkan uji Duncan pada taraf 5%.
64
Gambar 1 Nilai area under height of plant growth curve (AUHPGC) pada berbagai perlakuan bakteri endofit dan bakteri akar pemacu pertumbuhan tanaman secara tunggal dan kombinasi di rumah kaca dari pengamatan 1-5 minggu setelah tanam. F1, Pseudomonas fluorescens RH4003; A, Bacilus subtilis AB89; B, Staphylococcus epidermidis BC4; BC10, bakteri endofit isolat BC10; BL10, B. amyloliquefaciens; K, kontrol.
Nila
i AU
HPG
C (u
nit)
Perlakuan agens biokontrol
15
10
5
0F1 A B
BC10BL10F1*
B
F1*BC10
F1*BL10
A*B
A*BC10
A*BL10 K
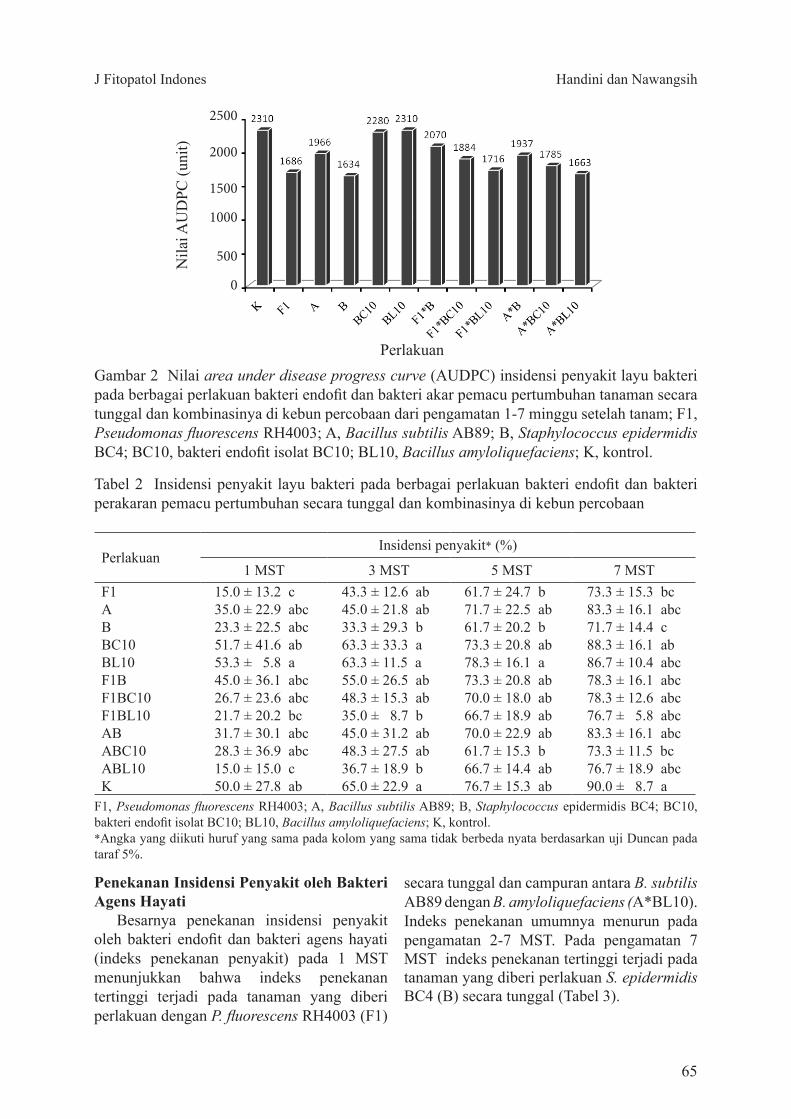
J Fitopatol Indones Handini dan Nawangsih
Penekanan Insidensi Penyakit oleh Bakteri Agens Hayati
Besarnya penekanan insidensi penyakit oleh bakteri endofit dan bakteri agens hayati (indeks penekanan penyakit) pada 1 MST menunjukkan bahwa indeks penekanan tertinggi terjadi pada tanaman yang diberi perlakuan dengan P. fluorescens RH4003 (F1)
PerlakuanInsidensi penyakit* (%)
1 MST 3 MST 5 MST 7 MSTF1 15.0 ± 13.2 c 43.3 ± 12.6 ab 61.7 ± 24.7 b 73.3 ± 15.3 bc A 35.0 ± 22.9 abc 45.0 ± 21.8 ab 71.7 ± 22.5 ab 83.3 ± 16.1 abc B 23.3 ± 22.5 abc 33.3 ± 29.3 b 61.7 ± 20.2 b 71.7 ± 14.4 c BC10 51.7 ± 41.6 ab 63.3 ± 33.3 a 73.3 ± 20.8 ab 88.3 ± 16.1 ab BL10 53.3 ± 5.8 a 63.3 ± 11.5 a 78.3 ± 16.1 a 86.7 ± 10.4 abc F1B 45.0 ± 36.1 abc 55.0 ± 26.5 ab 73.3 ± 20.8 ab 78.3 ± 16.1 abc F1BC10 26.7 ± 23.6 abc 48.3 ± 15.3 ab 70.0 ± 18.0 ab 78.3 ± 12.6 abc F1BL10 21.7 ± 20.2 bc 35.0 ± 8.7 b 66.7 ± 18.9 ab 76.7 ± 5.8 abc AB 31.7 ± 30.1 abc 45.0 ± 31.2 ab 70.0 ± 22.9 ab 83.3 ± 16.1 abc ABC10 28.3 ± 36.9 abc 48.3 ± 27.5 ab 61.7 ± 15.3 b 73.3 ± 11.5 bc ABL10 15.0 ± 15.0 c 36.7 ± 18.9 b 66.7 ± 14.4 ab 76.7 ± 18.9 abc K 50.0 ± 27.8 ab 65.0 ± 22.9 a 76.7 ± 15.3 ab 90.0 ± 8.7 a
Tabel 2 Insidensi penyakit layu bakteri pada berbagai perlakuan bakteri endofit dan bakteri perakaran pemacu pertumbuhan secara tunggal dan kombinasinya di kebun percobaan
F1, Pseudomonas fluorescens RH4003; A, Bacillus subtilis AB89; B, Staphylococcus epidermidis BC4; BC10, bakteri endofit isolat BC10; BL10, Bacillus amyloliquefaciens; K, kontrol.*Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata berdasarkan uji Duncan pada taraf 5%.
65
Gambar 2 Nilai area under disease progress curve (AUDPC) insidensi penyakit layu bakteri pada berbagai perlakuan bakteri endofit dan bakteri akar pemacu pertumbuhan tanaman secara tunggal dan kombinasinya di kebun percobaan dari pengamatan 1-7 minggu setelah tanam; F1, Pseudomonas fluorescens RH4003; A, Bacillus subtilis AB89; B, Staphylococcus epidermidis BC4; BC10, bakteri endofit isolat BC10; BL10, Bacillus amyloliquefaciens; K, kontrol.
secara tunggal dan campuran antara B. subtilis AB89 dengan B. amyloliquefaciens (A*BL10). Indeks penekanan umumnya menurun pada pengamatan 2-7 MST. Pada pengamatan 7 MST indeks penekanan tertinggi terjadi pada tanaman yang diberi perlakuan S. epidermidis BC4 (B) secara tunggal (Tabel 3).
Perlakuan Agens Biokontrol
Nila
i AU
DPC
(sat
uan)
N
ilai A
UD
PC (u
nit)
2500
2000
1500
1000
500
0
Perlakuan

J Fitopatol Indones Handini dan Nawangsih
PerlakuanIndeks penekanan insidensi penyakit (%)
1 MST 3 MST 5 MST 7 MSTF1 70.0 33.3 19.6 18.6A 30.0 30.8 6.5 7.4B 53.4 48.8 19.6 20.3BC10 0.0 2.6 4.4 1.9BL10 0.0 2.6 0.0 3.7F1B 10.0 15.4 4.4 13.0F1BC10 46.6 25.7 8.7 13.0F1BL10 56.6 46.2 13.0 14.8AB 36.6 30.8 8.7 7.4ABC10 43.4 25.7 19.6 18.6ABL10 70.0 43.5 13.0 14.8
Tabel 3 Indeks penekanan insidensi penyakit layu bakteri pada berbagai perlakuan bakteri endofit dan bakteri pemacu pertumbuhan tanaman secara tunggal dan kombinasinya di kebun percobaan
F1, Pseudomonas fluorescens RH4003; A, Bacillus subtilis AB89; B, Staphylo-coccus epidermidis BC4; BC10, bakteri endofit isolat BC10; BL10, Bacillus amylo-liquefaciens; *Relatif dibandingkan dengan kontrol; MST, minggu setelah tanam.
66
PEMBAHASAN
Indeks penekanan yang dihasilkan oleh bakteri endofit P. fluorescens yang diaplikasikan secara tunggal dan campuran antara B. subtilis AB89 dengan B. amyloliquefaciens dapat mencapai 70% tetapi hanya sampai 1 minggu setelah tanam. Pada pengamatan minggu berikutnya indeks penekanannya menurun lebih dari 30%. Hal ini menunjukkan bahwa bakteri endofit baik yang diaplikasikan secara tunggal maupun dalam campuran dengan frekuensi aplikasi pada awal tanam saja belum mampu menekan penyakit dalam jangka panjang. Untuk mengatasi hal tersebut perlu dilakukan aplikasi dengan frekuensi yang lebih banyak.
Salah satu faktor keberhasilan agens hayati dalam menekan penyakit antara lain ialah kemampuannya dalam mengolonisasi niche yang sama dengan patogen. Compant et al. (2005) mengemukakan bahwa bakteri akar pemacu pertumbuhan tanaman, sangat bergantung pada kemampuannya, baik untuk memanfaatkan lingkungan yang spesifik maupun beradaptasi terhadap perubahan kondisi lingkungannya.
Berbagai sumber bakteri endofit merupakan fokus yang sangat menarik saat
ini. Lokasinya yang berada dalam jaringan tanaman menjadikan posisi mereka yang kuat dalam mempengaruhi nutrisi tanaman, katabolisme polutan, respons terhadap tekanan atau pertahanan dan penyerangan patogen (Bent dan Chanway 2002; Melnick et al. 2008). Bakteri endofit merupakan pengoloni internal yang mempunyai keuntungan lebih sebagai agens hayati dibandingkan dengan pengoloni rizosfer (Hallmann et al. 2000). Oleh karena tanaman menyediakan tempat dan nutrisi maka bakteri dapat berkembang dalam kondisi kompetisi yang sangat kurang dan menutupi bagian dalam tanaman dari serangan patogen.
Berdasarkan hasil yang diperoleh dapat disimpulkan bahwa bakteri endofit dan bakteri perakaran pemacu pertumbuhan tanaman dan diaplikasikan secara tunggal yang memberikan penekanan relatif paling tinggi ialah S. epidermidis BC4 dan P. fluorescens RH4003. Aplikasi campuran bakteri agens hayati yang memberikan penekanan terhadap perkembangan penyakit paling tinggi adalah campuran antara B. subtilis AB89 dan B. amyloliquefaciens BL10.

J Fitopatol Indones Handini dan Nawangsih
67
UCAPAN TERIMAKASIH
Penelitian ini terselenggara dengan dana dari sebagian Program Penelitian Desentralisasi-Hibah Bersaing yang dibiayai dari DIPA IPB dengan nomor kontrak 01/I3.24.4/SPP/ PHB/2011, tanggal 28 Maret 2011.
DAFTAR PUSTAKA
Bent E, Chanway CP. 2002. Potential for misidentification of spore-forming Paenibacillus polymyxa isolat as an endophyte by using culture-based methods. Appl Environ Microbiol. 68:4650–4657. DOI: http://dx.doi.org/10.1128/AEM.68.9.4650-4652.2002.
Compant S, Duffy B, Nowak J, Clement C, Barka EA. 2005. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanism of action, and future prospect. Appl Environ Microbiol. 71(9):4951–4959. DOI: http://dx.doi.org/10.1128/AEM.71.9.4951-4959.2005.
Hallmann J, Quadt-Halmann A, Miller WG, Sikora RA, Lindow SE. 2000. Endophytic colonization of plants by the biocontrol agent Rhizobium etli G12 in relation to Meloidogyne incognita infection. Phytopathology. 91:415–422. DOI: http://dx.doi.org/10.1094/PHYTO.2001.91.4.415.
Melnick RL, Zidack NK, Bailey BA, Maximova SN, Guiltinan M, Backman PA. 2008. Bacterial endophytes: Bacillus spp. from annual crops as potential biological control agents of black pod rot of cacao.
Biol Control. 46:46–56. DOI: http://dx.doi.org/10.1016/j.biocontrol.2008.01.022.
Nawangsih AA. 2006. Seleksi dan karakterisasi bakteri biokontrol untuk mengendalikan penyakit layu bakteri (Ralstonia solanacearum) pada tomat [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Nawangsih AA, Damayanti I, Wiyono S, Kartika JG. 2010. Selection and characterization of endophytic bacteria as biocontrol agents of tomato bacterial wilt disease. Hayati. 18(2):66–70. DOI: http://dx.doi.org/10.4308/hjb.18.2.66.
Reiter B, Pfeifer U, Schwab H, Sessitsch A. 2002. Response of endophytic bacterial communities in potato plants to infection with Erwinia carotovora subsp. atroseptica. Appl Environ Microbiol. 68:2261–2268. DOI: http://dx.doi.org/10. 1128/AEM.68.5.2261-2268.2002.
Sharma PK, Sarita S, Prell J. 2005. Isolation and characterization of an endophytic bacterium related to Rhizobium/Agrobacterium from wheat (Triticum aestivum L.) roots. Current Science. 89:608–610.
Unterstenhofer G. 1963. The basic principles of crop protection field trials. Pflanzerschutz Nachtichten Bayer. 29(2):155.
Van der Plank JE. 1963. Plant Disease: Epidemics an Control. London (GB): Academic Pr.
Vega FE, Pava-Ripoll M, Posada F, Buyer JS. 2005. Endophytic bacteria in Coffea arabica L. J Basic Microbiol. 45:371–380. DOI: http://dx.doi.org/10.1002/jobm.200 410551.

Volume 10, Nomor 2, April 2014Halaman 68–72
DOI: 10.14692/jfi.10.2.68
*Alamat korespondensi penulis: Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Hasanuddin Jalan Perintis Kemerdekaan Km. 10, Makassar 90245Tel: 0411-587100, Faks: 0411-587100, Surel: [email protected]
Efektivitas Isolat Bakteri dari Rizosfer dan Bahan Organik Terhadap Ralstonia solanacearum dan Fusarium oxysporum
pada Tanaman Kentang
Effectiveness of Bacterial Isolates from Several Rhizospheres and Organic Materials against Ralstonia solanacearum and
Fusarium oxysporum on Potato
Tutik Kuswinanti*, Baharuddin, Sri SukmawatiUniversitas Hasanuddin, Makasar 90245
ABSTRAK
Rizosfer merupakan daerah yang ideal untuk pertumbuhan dan perkembangan mikrob antagonis.Nutrisi yang disekresikan tanaman ke dalam rizosfer banyak dipengaruhi oleh faktor lingkungan dan selanjutnya mempengaruhi kelimpahan dan keragaman mikrob di daerah tersebut. Penelitian ini bertujuan untuk mengisolasi bakteri dari rhizosfer tanaman dan dari bahan organik di Tana Toraja, dan menguji potensi bakteri tersebut sebagai agens hayati penyakit layu kentang. Sampel tanah diambil dari rizosfer tanaman padi, bambu, kentang, dan terung belanda; serta dari bahan organik kerbau belang dan babi. Uji antagonisme isolasi, Ralstonia solanacearum dan Fusarium oxysporum dilakukan untuk mengetahui kemampuan daya hambat masing-masing isolat bakteri dengan metode biakan ganda serta tahapan identifikasi. Hasil penelitian menunjukkan bahwa dari total 74 isolat bakteri, terdapat berturut-turut 10 dn 4 isolat bakteri yang mampu menghambat pertumbuhan R. solanacearum dan F. oxysporum secara in vitro.
Kata kunci: antagonis, penghambatan, penyakit layu
ABSTRACT
Rhizosphere is an ideal area for growth and development of microbial antagonists, because it is a nutrition rich area. Nutrients that secreted in the rhizosphere are influenced by many environmental factors which in turn effect the diversity and abundance of microbial community. Based on these facts, the research were aimed to isolate bacteria from the rhizosphere and organic materials in Tana Toraja, and determine their role as biological control agents for potato wilt disease. Antagonistic test was conducted in order to determine the ability of each bacterial isolate to inhibit the growth of Ralstonia solanacearum and Fusarium oxysporum. The results showed that from totally 74 bacterial isolates, only 10 and 4 bacterial isolates were able to inhibit the growth of R. solanacearum and F. oxysporum in vitro, respectively.
Key words: antagonist, inhibition, potato wilt
68
ISSN: 2339-2479

J Fitopatol Indones Kuswinanti et al.
PENDAHULUAN
Rendahnya produktivitas kentang di Indonesia disebabkan oleh teknik budi daya yang belum optimal, kurangnya ketersediaan bibit yang bermutu dan bersertifikat, serta serangan organisme pengganggu tanaman. Salah satu penyakit pada kentang adalah penyakit layu yang disebabkan bakteri Ralstonia solanacearum dan cendawan Fusarium oxysporum. Infeksi patogen ini dilaporkan dapat menyebabkan kerugian besar pada berbagai sentra produksi dan ancaman pada daerah target pengembangan di Indonesia.
Beberapa mikrob yang menyelimuti perakaran tanaman sehat diketahui sebagai pelindung dari serangan patogen layu. Pada perakaran tanaman sehat, bakteri antagonis Bacillus subtilis, Pseudomonas fluorescens dan Streptomyces sp. dilaporkan dapat mengendalikan R. solanacearum pada kentang (Nurbaya et al. 2011).
Kabupaten Tana Toraja memiliki potensi menjadi daerah pengembangan hortikultura, termasuk kentang. Eksplorasi keragaman bakteri rizosfer sangat diperlukan untuk mengetahui potensi ketersediaan agens hayati terhadap penyakit layu kentang. Penelitian dilakukan untuk mengevaluasi efektivitas beberapa isolat rizobakteri dan bahan organik untuk menekan penyakit layu kentang.
BAHAN DAN METODE
Isolasi dan Perbanyakan Fusarium oxysporum dan Ralstonia solanacearum
Isolat Fusarium diperoleh dari koleksi Laboratorium Bioteknologi, Universitas Hasanuddin yang telah diuji sebelumnya, sedangkan bakteri R. solanacearum diisolasi dari jaringan batang tanaman kentang yang memperlihatkan gejala layu. Jaringan diambil 1 cm lalu dipotong-potong hingga berukuran 5 mm ×5 mm lalu digerus pada mortar hingga halus dan ditambahkan 1 mL akuades steril. Suspensi bakteri yang ada pada mortar diambil sebanyak 0.5 mL kemudian dimasukkan ke dalam tabung reaksi yang berisi 4.5 mL
MgSO4.7H2O. Dari larutan tersebut kemudian
diencerkan hingga mencapai pengenceran 10-6. Sebanyak 0.1 mL suspensi bakteri dari masing-masing tingkat pengenceran dipindahkan ke dalam cawan yang berisi medium spesifik untuk R. solanacearum, yaitu tetrazolium chloride agar (TTC) yang mengandung 10 g pepton, 5 g glukosa, 15 g agar, 1000 mL akuades, 8 g nutrient broth, dan 2,3,5- triphenyltetrazoliumchlorid (TTC) 1%. Suspensi disebar merata pada seluruh permukaan medium dengan menggunakan spatula. Setelah 3-5 hari dilakukan pengamatan dan seleksi koloni bakteri yang khas kemudian ditumbuhkan kembali pada medium TTC yang baru.
Uji patogenesitas dilakukan melalui uji hipersensitivitas pada daun tembakau. Isolat yang telah murni selanjutnya dipindahkan ke media nutrient growth agar (3.3 g pepton, 2.7 g nutrient broth, 2 g ekstrak khamir, gliserol 2.5% (v/v) dan 15 g agar-agar L-1) sebagai medium perbanyakan.
Bakteri AntagonisSampel untuk penelitian ini diperoleh
dari rizosfer tanaman kentang, bambu, padi, terung belanda serta bahan organik kerbau belang dan babi di Desa Minanga, Kecamatan Mengkendek, Kabupaten Tana Toraja, Provinsi Sulawesi Selatan.
Sebanyak 1 g sampel rizosfer tanaman dan bahan organik disuspensikan dalam 10 mL air steril, lalu dibuat pengenceran hingga 6 kali. Pada pengenceran 10-4, 10-5 dan 10-6, diambil sebanyak 1 mL dan ditumbuhkan pada medium King’s B. Setelah murni, isolat diperbanyak pada medium agar-agar nutrien.
Uji Kemampuan Daya Hambat Bakteri Antagonis terhadap R. solanacearum
Isolat murni bakteri dari rizosfer dan bahan organik diinokulasikan pada botol berisi medium cair nutrien (8 g nutrient broth L-1), lalu digoyang dengan kecepatan 120 rpm selama 3 hari. Setelah itu, sebanyak 1 mL medium cair dimasukkan ke tabung ependorf dan disentrifiugasi selama 10 menit dengan kecepatan 100 rpm. Supernatan selanjutnya
69

J Fitopatol Indones Kuswinanti et al.
disaring menggunakan filter 0.02 µM dan dimasukkan ke tabung ependorf berisi larutan kloroform. Selanjutnya dilakukan pengenceran isolat murni bakteri patogen dan diratakan pada medium NGA. Kertas saring steril berukuran 0.5 cm dicelupkan ke dalam larutan kloroform yang mengandung supernatan bakteri antagonis, dikeringanginkan, lalu diletakkan pada bagian tengah medium yang telah berisi bakteri patogen. Pada perlakuan kontrol, potongan kertas saring dicelupkan dalam pelarut kloroform. Kemampuan antagonisme isolat-isolat bakteri terhadap R. solanacearum diukur berdasarkan diameter zona penghambatan disekeliling kertas saring.
Uji Kemampuan Daya Hambat Bakteri Antagonis terhadap Fusarium oxysporum
Uji kemampuan bakteri menghambat F. oxysporum dilakukan dengan metode kultur ganda Fokkema (1973). Persentase penghambatan (P) dihitung dengan rumus:
Ro, jari-jari pertumbuhan patogen pada perlakuan kontrol (cm); R1 dan R2, jari-jari pertumbuhan patogen pada perlakuan (cm).
Penelitian ini disusun dalam acak lengkap dengan pengulangan sebanyak 3 kali pada setiap perlakuan. Analisis sidik ragam dilakukan, jika di antara perlakuan menunjukkan perbedaan yang nyata dengan uji Duncan pada taraf 5%.
HASIL
Isolat Bakteri dan Uji Antagonisme Terhadap R. solanacearum
Dari rhizosfer tanaman kentang, padi, bambu dan terung belanda, serta dua jenis bahan oganik berhasil diisolasi sebanyak 74 isolat bakteri, yang terdiri atas 31 isolat dari rizosfer pertanaman dan 43 isolat dari bahan organik (Tabel 1).
Dari total 74 isolat bakteri yang diperoleh, 10 isolat bakteri memiliki potensi sebagai agens antagonis karena mampu membentuk zona hambat terbesar (Tabel 2). Isolat-isolat tersebut terdiri atas 1 isolat asal rizosfer,
bambu dan 9 isolat asal bahan organik kerbau belang (Tabel 2).
Uji Kemampuan Daya Hambat Bakteri Antagonis Terhadap Fusarium oxysporum
Pengujian kemampuan daya hambat isolat-isolat bakteri terhadap F. oxysporum menunjukkan terdapat 4 isolat (BT5, KT9, KB11 dan KB25) yang memiliki kemampuan daya hambat tertinggi. Persentase daya hambat tertinggi (80.68%) diperoleh dari perlakuan dengan isolat BT5, diikuti berturut-turut oleh KB11 (71.59%), KT9 (67.41%) dan KB25 (65.91%) (Gambar 1).
PEMBAHASAN
Pengendalian hayati patogen mengguna-kan mikroorganisme yang berasosiasi dengan rizosfer dan bahan organik merupakan pendekatan yang efisien dan ramah lingkungan (Bargabus et al. 2003). Bahan organik bermanfaat sebagai nutrisi tanaman, dan mengandung sejumlah besar mikrob termasuk bakteri, cendawan dan kapang. Mikroba ini memecah residu organik menjadi senyawa yang lebih sederhana selama proses pengomposan. Peningkatan nutrisi ini meningkatkan jangkauan dan jumlah mikrob tanah, termasuk yang berpotensi sebagai agensia hayati terhadap patogen tanaman (Holguín-Castaño dan Mora-Delgado
Sumber isolat bakteri Jumlah isolat
Rizosfer kentang (KT) 12Rizosfer padi (PT) 8Rizosfer bambu (BT) 6Rizosfer terung belanda (TB) 5
Bahan organik babi (BB) 8
Bahan organik Kerbau belang (KB) 35
Tabel 1 Hasil isolasi bakteri dari empat rhizosfer pertanaman dan dua jenis bahan organik
70
P = Ro - R1+R22
x 100% , dengan

J Fitopatol Indones Kuswinanti et al.
2009). Tahapan pengembangan agens hayati meliputi isolasi, skrining in vitro, uji kemampuan antagonisme, skrining in vivo, dan pengembangan formulasi (Ahmed et al. 2007).
Hasil penelitian menunjukkan bahwa isolat bakteri lebih banyak diperoleh dari bahan organik dibandingkan dengan dari rizosfer. Hal ini diduga akibat pengaruh struktur tanah, kesuburan tanah, kelembaban tanah dan ketersediaan nutrisi. Tanah yang sehat bukan hanya subur dan banyak mengandung bahan yang menunjang kesehatan tanaman, tetapi juga mampu menyediakan lingkungan yang cocok bagi mikrob tanah, sehingga tanaman
dapat terlindungi dari patogen tanah (Nion dan Toyota 2008).
Dari total 74 isolat bakteri yang diuji hanya 10 yang memperlihatkan daya hambat terhadap R. solanacearum dan F. oxysporum, dan hambat terdapat 4 isolat yang memiliki daya hambat terbaik. Samia et al. (2013) melakukan skrining terhadap 111 galur bakteri yang berasal dari tanaman kentang dan gandum. Hanya 14 isolat bakteri dari tanaman kentang yang bersifat antagonis terhadap Phytophthora infestans, F. oxysporum f. sp. albedinis and F. solani var. coeruleum dengan persentase penghambatan mencapai 92.30%. Tehrani dan Ramezani (2003) melaporkan
Sumber isolat bakteriJumlah isolat pada masing-masing
kategori daya hambat*- + ++ +++ ++++
Rizosfer kentang (KT) 0 8 4 0 -Rizosfer padi (PT) 0 2 4 2 0Rizosfer bambu (BT) - 3 1 1 1Rizosfer terung belanda (TB) 1 4 - - -Bahan organik babi (BB) 0 7 - 1 -Bahan organik kerbau belang (KB) 0 19 4 3 9
Tabel 2 Daya hambat isolat bakteri terhadap pertumbuhan Rolstania solanacearum secara in-vitro
Kemampuan daya hambat isolat bakteri dikelompokkan berdasarkan diamater zona hambat -, 0 (tidak ada penghambatan); -, 0; A, tidak ada penghambatan; B, 0.1–1 cm; C, > 1.0–2 cm; D, > 2.0–3 cm; E, > 3.0 cm.
Gambar 1 Persentase penghambatan Fusarium oxysporum oleh 4 bakteri antagonis potensial (KB5, KB11, KT9 dan KB25) berdasarkan pengamatan pada hari ke-8 dan ke-10.
71
KB5KB5 KB11KB11 KT9KT9 KB25KB250102030405060708090
100
8 10Hari pengamatan
% P
engh
amba
tan
Pengamatan hari ke-8 10

J Fitopatol Indones Kuswinanti et al.
bahwa kelompok rizobakteri yang merupakan mikrob antagonis adalah yang memiliki daya hambat diatas 51 persen terhadap patogen tular tanah seperti F. oxysporum Schlecht. Mikroba antagonis dapat berfungsi sebagai agens pengendali patogen melalui mekanisme kompetisi, antibiosis, parasitisme atau ketahanan yang terinduksi. Hingga saat ini kelompok bakteri yang paling banyak dimanfaatkan sebagai agens pengendali hayati adalah Pseudomonas berfluoresen (Weller et al. 2002) dan beberapa strain dari Bacillus (Choudhary dan Johri 2009).
Penggunaan agens antagonis untuk meningkatkan hasil panen dan melindungi tanaman dari serangan organisme penggangu tanaman merupakan pendekatan menjanjikan dalam sistem pertanian moderen. Penelitian terhadap isolat-isolat agens antagonis ini perlu dilakukan lebih lanjut untuk mengetahui mekanisme antagonistiknya.
UCAPAN TERIMA KASIH
Penelitian ini merupakan bagian dari tesis yang dibiayai oleh proyek Insentif Ristek TA 2012, Nomor Kontrak 06/M/Kp/I/2012.
DAFTAR PUSTAKA
Ahmed IH, Labuschagne N, Korsten L. 2007. Screening rhizobacteria for biological control of Fusarium root and crown rot of sorghum in Ethiopia. Biol Control. 40(1): 97–106. DOI: http://dx.doi.org/10.1016/j.biocontrol.2006.07.017.
Bargabus RL, Zidack NK, Sherwood JE, Jacobsen BJ. 2003. Oxidative burst elicited by Bacillus mycoides isolate, Bac J, a biological control agent, occurs independently of hypersensitive cell death in sugar beet. Mol Plant–Micro Interact. 16:1145–1153. DOI: http://dx.doi.org/10. 1094/MPMI.2003.16.12.1145.
Choudhary DK, Johri BN. 2009. Interactions of Bacillus spp. and plants – With special reference to induced systemic resistance (ISR). Microbiol Res. 64(5):493–513. DOI:
http://dx.doi.org/10.1016/j.micres.2008. 08.007.
Fokkema NJ. 1973. The role of saprophytic fungi in antagonsim against Drechslera sorokiniana (Helminthosporium sativum) on agar plates and on rye leaves with pollen. Physiological Plant Pathology. 3:195–205. DOI: http://dx.doi.org/10.1016/0048-4059 (73)90082-9.
Holguin-Castaño VA, Mora-Delgado J. 2009. Dinámica microbiana en vermicompostas comerciales con y sin inoculación del hongo Trichoderma spp. Revista Luna Azul. 29:18–24.
Nion YA, Toyota K. 2008. Suppression of bacteria wilt and fusarium wilt by a Burkholderia nodosa strain isolated from Kalimantan soils, Indonesia. Microbes Environ. 23(2):134–141. DOI: http://dx.doi.org/10.1264/jsme2.23.134.
Nurbaya, Zulfikar A, Kuswinanti T, Baharuddin dan Lologau BA. 2011. Kemampuan Mikroba Antagonis dalam Mengendalikan Ralstonia solanacearum pada Sistem Budi daya Aeroponik Tanaman Kentang. J Fitomedika. 7(3):155–158.
Samia MA, Nora H, Nadia S, Abdelhadi G and Mohamed MZ. 2013. Isolation and selection of indigenous bacterial strains with suppression properties from the Rhizospheres of potato and wheat. Annual Review & Research in Biology. 3(4):405–415.
Tehrani AS. and Ramazani M. 2003. Biological control of Fusarium oxysporum, the causal agent of onion wilt by antagonistic bacteria. Comm Agr Appl Biol Sci. 68(4):543–547.
Weller DM, Raaijmakers JM, McSpadden GB, Thomashow LS. 2002. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu Rev Phytopathol. 40:309–348. DOI: http://dx.doi.org/10.1146/annurev.phyto.40. 030402.110010.
72


















