
The bovine placenta before and after birth: placental ...€¦ · Animal Reproduction Science...
16
Ž . Animal Reproduction Science 60–61 2000 145–160 www.elsevier.comrlocateranireprosci The bovine placenta before and after birth: placental development and function in health and disease D.H. Schlafer ) , P.J. Fisher, C.J. Davies Department of Biomedical Sciences, College of Veterinary Medicine, Cornell UniÕersity, T6-020, VRT, Ithaca, NY 14853 USA Abstract Ž . This paper reviews bovine placental development, anatomy microscopic and gross , nomencla- ture and classification. The paper focuses on the biology of those specialized cells that arise from the outermost layer of very early embryos, the trophoblast cells, and on placental macrophages, cells that play a key role in fetalrplacental defense. Data is presented from an immunohistochemi- cal quantitative study that characterizes the ontogeny of placental macrophages using placental Ž . tissues from 21 cows sampled from 4 months of pregnancy through the post partum period . Understanding of bovine placental development is essential for veterinarians, pathologists, diag- nosticians and researchers. Lesions of diagnostic significance can be recognized for many economically important infectious abortifacient diseases, and there is growing evidence that pregnancy failure of cloned calves is due in part to unexplained placental failure. Placentology and placental pathology are becoming of increasing importance. q 2000 Elsevier Science B.V. All rights reserved. Keywords: Bovine; Placenta; Development; Anatomy; Histology; Macrophage; Disease; Review 1. Introduction The cow is one of the ‘‘eutherian’’ species, i.e. those that have placentas. The word ‘‘placenta’’ is taken from latin meaning ‘‘flat cake’’ from the apparent gross similarity of the human discoid placenta to round flat loaves of unleavened bread commonly made in ancient times. The gross appearance of placentas varies greatly between species. ) Corresponding author. Tel.: q 1-607-253-3352; fax: q 1-607-253-3317. Ž . E-mail address: [email protected] D.H. Schlafer . 0378-4320r00r$ - see front matter q 2000 Elsevier Science B.V. All rights reserved. Ž . PII: S0378-4320 00 00132-9
Transcript of The bovine placenta before and after birth: placental ...€¦ · Animal Reproduction Science...

Ž .Animal Reproduction Science 60–61 2000 145–160www.elsevier.comrlocateranireprosci
The bovine placenta before and after birth: placentaldevelopment and function in health and disease
D.H. Schlafer ), P.J. Fisher, C.J. DaviesDepartment of Biomedical Sciences, College of Veterinary Medicine, Cornell UniÕersity, T6-020, VRT,
Ithaca, NY 14853 USA
Abstract
Ž .This paper reviews bovine placental development, anatomy microscopic and gross , nomencla-ture and classification. The paper focuses on the biology of those specialized cells that arise fromthe outermost layer of very early embryos, the trophoblast cells, and on placental macrophages,cells that play a key role in fetalrplacental defense. Data is presented from an immunohistochemi-cal quantitative study that characterizes the ontogeny of placental macrophages using placental
Ž .tissues from 21 cows sampled from 4 months of pregnancy through the post partum period .Understanding of bovine placental development is essential for veterinarians, pathologists, diag-nosticians and researchers. Lesions of diagnostic significance can be recognized for manyeconomically important infectious abortifacient diseases, and there is growing evidence thatpregnancy failure of cloned calves is due in part to unexplained placental failure. Placentology andplacental pathology are becoming of increasing importance. q 2000 Elsevier Science B.V. Allrights reserved.
Keywords: Bovine; Placenta; Development; Anatomy; Histology; Macrophage; Disease; Review
1. Introduction
The cow is one of the ‘‘eutherian’’ species, i.e. those that have placentas. The word‘‘placenta’’ is taken from latin meaning ‘‘flat cake’’ from the apparent gross similarityof the human discoid placenta to round flat loaves of unleavened bread commonly madein ancient times. The gross appearance of placentas varies greatly between species.
) Corresponding author. Tel.: q1-607-253-3352; fax: q1-607-253-3317.Ž .E-mail address: [email protected] D.H. Schlafer .
0378-4320r00r$ - see front matter q2000 Elsevier Science B.V. All rights reserved.Ž .PII: S0378-4320 00 00132-9

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160146
There are very few similarities between the appearance of a placenta from a woman andone from a cow; but during very early embryonic development, before they enter theuterus, they appear nearly identical. Once placental development begins, dramaticdifferences become apparent.
When this great variation in anatomy of the definitive placenta from animal species toanimal species is combined with the seemingly complex medical terms applied todevelopmental and histologic features, placentology can become quickly frustrating. Thepurpose of this paper is to present bovine placental development from a simplifiedperspective, to discuss and illustrate gross and microscopic features and to presentpreliminary quantitative data on the ontogeny of bovine placental macrophages, cellsthat are important in utero-fetal defense systems.
Fig. 1. Essential steps in early placental development.The outer layer of the expanded blastocyst, the trophectoderm, is composed of trophoblast cells. This layer is
Žalso referred to as ‘‘the trophoblast’’. Cells on the inner aspect of the blastocoel cavity which becomes the.yolk sac cavity will develop into the embryo. The outer trophoblast layer, combined with the subjacent
somatic mesoderm, is called the chorion and it becomes vascularized by apposition to a bulging fluid-filledallantoic sac that develops from the embryo’s hindgut. This layer of allantois and chorion becomes thechorioallantois of the fetal membranes. The amnion is formed around the embryo by folding of the chorion toreach over the embryo and fuse, thereby, enclosing the embryo in a membrane lined by amniotic epithelialcells.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 147
2. Placental development
Following fertilization, the embryo goes through stages of division, modeling andŽ .cellular compaction morula stage . Cells that make up the early embryo are called
‘‘blastomeres’’. The ‘‘blastocyst stage’’ occurs when the embryo develops a fluid-filledcentral cavity, the ‘‘blastocoel’’, that is surrounded by a single layer of cells, the
Ž .‘‘trophectoderm’’ Fig. 1 . These cells, forming this outer wall, have assumed special-ized functions and are known as trophoblast cells. They will remain as the outermost
Fig. 2. Bovine Fetuses — Early Placentation.These bovine fetuses are contained in a clear membrane the ‘‘amnion’’ forming a round fluid-filled structure
Ž .surrounding them. The chorioallantois has been torn to better demonstrate the amniotic sacs. These threefetuses are approximately 30, 40 and 50 days of gestation. Placental vessels are evident extending from thefetus along the umbilical cord to the chorioallantois, which initially attaches to the endometrium at about 33days of gestation. Cotyledons are not yet visible.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160148
layer of cells essentially forming a single cell layer covering the outside of the placentaof all domestic animal species. This outer layer of trophoblast cells, together with thesomaticrparietal mesoderm, form the ‘‘chorion’’.
With time, more fluid is produced within the blastocoel resulting in an expansion ofthe size of the embryo, which is then called the ‘‘expanded blastocyst’’. The fluid-filled
Ž .blastocoel will become the cavity of the yolk sac Fig. 1 .During the blastocyst stage, another specialized population of cells, those that will
become the embryo proper, replicate and appear as a flat cluster of cells at one pole of
Fig. 3. Bovine fetus and placenta with cotyledons evident.The amnion has expanded and discrete cotyledons are now readily visible. Specialized areas called ‘‘cotyle-dons’’ begin to develop on the surface of the chorioallantois at about 30 days gestation and are readily visiblein this photograph of an approximately 70-day fetus. Cotyledons on that part of chorioallantois filling theuterine horn are larger than those lining the placental membranes in the other horn. Cotyledons closer to the tipof the uterine horn will also be smaller. Total numbers vary greatly from 70 to 120.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 149
Ž .the hollow blastocyst. This cluster of cells is the embryonic disc inner cell mass thatŽ .will become the embryo. Another cell layer endoderm grows from the inner cell mass
Žto line the cavity of the yolk sac, and a third layer of cells, the mesoderm mesos.middle , will then extend from the embyonic disc and form a layer between the
trophectoderm and the endoderm.The expanded blastocyst maintains its round structure, with essentially three concen-
Ž .tric layers of cells trophectoderm, mesoderm and endoderm surrounding the early yolksac cavity with the mass of cells that will differentiate into the embryo located along onepole. Following this, the embryo will elongate. The amnion will form as folds of thetrophectoblast cells, together with somatic mesoderm, rise up from around the embryo
Ž .proper to ultimately join and create a fluid-filled space around the embryo see Fig. 2 .As the embryo begins to form into a fetus, but before the ventral abdominal wall is
closed, an outpocketing of the hindgut extends from the fetus into the loose tissues ofthe splanchnic mesoderm. This sac-like stucture is the ‘‘allantois’’ and with thesplanchnic mesoderm, it carries with it the vessels that will ultimately vascularize the
Ž .chorion and amnion Fig. 1 .Only one additional major remodeling event remains. The expanding allantois
becomes directly apposed to the chorion . The allantois and the chorion fuse to form thechorioallantois, which, with the amnion, are the ‘‘extra-embryonic fetal membranes’’.Depending on the species, the chorioallantois will assume different shapes and micro-scopic forms as it apposes and, in some cases, invades the endometrium of the graviduterus.
Fig. 4. Placentome from a term pregnancy with umbilical arteries injected with colored latex. Note the intricatevascular pattern of branches of the umbilical arteries and veins as they penetrate or leave the placentome.

()
D.H
.Schlaferet
al.rA
nimalR
eproductionScience
60–
612000
145–
160150
Fig. 5. Drawing of two bovine placentome and fetal membranes attached to the endometrium.The relationship between the chorioallantois forming cotyledonary villi and the endometrium is demonstrated in this drawing. Branching cotyledonary villi penetrate
Ž Ž .the crypts of the endometrium at specialized sites called caruncles. Together, they form the placentome. Reprinted from Davies et al. 2000 by permission of the.publisher W-B Saunders .

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 151
3. Gross anatomy
In the cow, the chorioallantois attaches at about 4 weeks of gestation to the lining ofthe uterus. This becomes more intimate over specialized areas of the endometrium called
Žcaruncles approximately 100 are present, evenly distributed throughout the en-.dometrium . These sites can be seen grossly even in uteri of fetal calves. As the extra
embryonic fetal membranes grow and expand within the lumen of the uterus, the flatsurface of the chorioallantois will begin to become irregular over these caruncles. Theremodeling of these areas, now recognized grossly as cotyledons, progresses to form
Žvillous projections that interdigitate with recesses in the surface of the caruncle Figs. 3,.5 and 6 . The surface of the caruncles develops crypts and the apposing chorioallantois
is modeled to form finger-like villous projections that enter into them, thus, enhancingŽ .the contact surface area Figs. 4–6 . The combined cotyledonary and caruncular tissues
together are called ‘‘placentomes’’. As gestation progresses, these become domed ovoidŽ .shaped structures ranging from 10 to 12 cm long and 2–3 cm thick Figs. 4 and 5 .
Significant variation in size normally occurs. Placentomes are arranged in a very orderlyŽ .manner in four rows that run lengthwise along both uterine horns Fig. 7 . They are
largest in the horn in which the fetus develops and tend to decrease in size towards theapex of both horns. The surface area of contact between the cow and fetal calf is greatly
Fig. 6. Bovine placentome with the cotyledonary villi partially separated from caruncular crypts.This photograph taken with the partially separated placentome under water demonstrates the relationshipbetween the fetal villi and the maternal crypts.
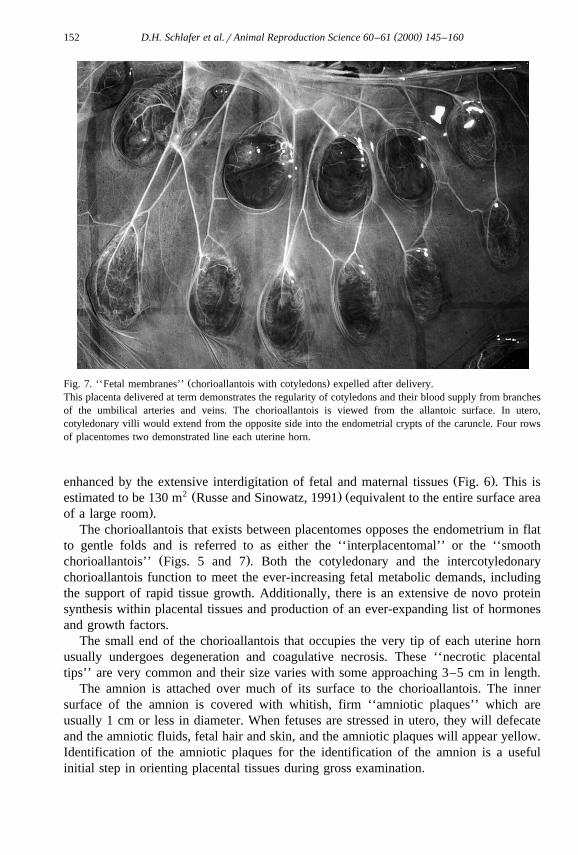
( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160152
Ž .Fig. 7. ‘‘Fetal membranes’’ chorioallantois with cotyledons expelled after delivery.This placenta delivered at term demonstrates the regularity of cotyledons and their blood supply from branchesof the umbilical arteries and veins. The chorioallantois is viewed from the allantoic surface. In utero,cotyledonary villi would extend from the opposite side into the endometrial crypts of the caruncle. Four rowsof placentomes two demonstrated line each uterine horn.
Ž .enhanced by the extensive interdigitation of fetal and maternal tissues Fig. 6 . This is2 Ž . Žestimated to be 130 m Russe and Sinowatz, 1991 equivalent to the entire surface area
.of a large room .The chorioallantois that exists between placentomes opposes the endometrium in flat
to gentle folds and is referred to as either the ‘‘interplacentomal’’ or the ‘‘smoothŽ .chorioallantois’’ Figs. 5 and 7 . Both the cotyledonary and the intercotyledonary
chorioallantois function to meet the ever-increasing fetal metabolic demands, includingthe support of rapid tissue growth. Additionally, there is an extensive de novo proteinsynthesis within placental tissues and production of an ever-expanding list of hormonesand growth factors.
The small end of the chorioallantois that occupies the very tip of each uterine hornusually undergoes degeneration and coagulative necrosis. These ‘‘necrotic placentaltips’’ are very common and their size varies with some approaching 3–5 cm in length.
The amnion is attached over much of its surface to the chorioallantois. The innersurface of the amnion is covered with whitish, firm ‘‘amniotic plaques’’ which areusually 1 cm or less in diameter. When fetuses are stressed in utero, they will defecateand the amniotic fluids, fetal hair and skin, and the amniotic plaques will appear yellow.Identification of the amniotic plaques for the identification of the amnion is a usefulinitial step in orienting placental tissues during gross examination.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 153
Amniotic fluid is produced by several means which include fluxes of fluid across theamnionic epithelium and unkeratinized fetal skin, production by the lungs, and, in laterpregnancy, by urine. During most of gestation, urine produced by the fetus is expelledinto the bladder and through the urachus within the umbilical cord into the allantoiccavity. Near term, however, it has been shown that the composition of amniotic fluidchanges to become more like urine. Fetuses older than 240 days gestation urinate
Ž .through the urethra into the amniotic cavity Reeves et al., 1972 . The amount ofamniotic and allantoic fluid is approximately equal in the 3-month gestation bovine
Ž .fetus, but by term, the volume of allantoic fluid 10–15 l is greater than the volume ofŽ . Ž .amniotic fluid 2.5–7.5 l Russe and Sinowatz, 1991 .
4. Histology
As noted, trophoblast cells form a continuous epithelial layer of the chorion, over theentire surface of the chorioallantois. Subpopulations of the trophoblast cells, however,are present, and have different functions.
Two populations of mononuclear trophoblast cells have phagocytic phenotypes.ŽThese are the trophoblast cells lining the arcade area located towards the fetal side of. Ž .the placentome between the bases of cotyledonary villi Fig. 5 . These cells line the
‘‘hematophagus organs’’ where maternal erythrocytes accumulate between the surfacesof endometrial and chorioallantois tissues and are phagocytized by the trophoblast cells.A second area of specialized phagocytic trophoblast cells is found in the interpla-centomal chorioallantois overlying the openings of endometrial glands. Microscopicaccumulations of uterine milk secreted by endometrial glands raise the overlying
Ž .chorioallantois to form small microscopic dome-shaped spaces referred to as ‘‘areolae’’.Trophoblast cells over these chambers are tall and contain phagocytized uterine milk.While areolae are not visible grossly, careful examination of the surface of freshplacentomes reveals small uniformly distributed black areas that are the hematophagusorgans.
Approximately one fifth of bovine trophoblast cells are binucleated. The cellularbiology of these cells is amazing. Although they may have a similar microscopicappearance, lectin cytochemistry clearly demonstrates the existence of many subpopula-
Ž .tions Munson et al., 1989; Jones et al., 1994 .Trophoblast cells produce a number of hormones and growth factors including
Ž . Ž . Žprogesterone Reimers et al., 1985 , bovine placental lactogen bPL Duello et al.,. Ž . Ž1986 , pregnancy associated glycoproteins bPAG-1, bPAG-2, bPAG-3 Roberts et al.,. Ž .1995 , and transforming growth factor beta Munson et al., 1996 . These hormones are
associated with fetal growth and development and the maintenance of pregnancy.Progesterone receptors are present within nuclei of many different tissues of the
Žplacentome, including trophoblast cells, suggesting paracrine function Shuler et al.,.1999 .
In addition to the production of a remarkable array of products, trophoblast cells alsoundergo intriguing migration into the maternal epithelium. Ultrastructural studies usingphosphotungstic acid to stain cytoplasmic granules of binucleate cells clearly demon-strated the translocation of binucleate trophoblast cells from the chorionic layer of the

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160154
cotyledonary villi across to penetrate the single epithelial cell layer lining the caruncularŽ .crypts see Wooding et al., 1980; Wooding, 1982, 1992 . Similar migration occurs in the
interplacentomal areas. As the binucleate cells migrate, they routinely fuse with a singleendometrial epithelial cell and discharge their cytoplasmic granules. The fusion of a fetalcell, the binucleate trophoblast with a maternal cell temporarily forms a hybrid cell withthree nuclei. This hybrid represents the union of two separate individuals into a singlecell!
Figs. 8 and 9 show the dramatic difference in immunohistochemical staining usingŽmonoclonal antibody SBU-3 SBU-3 is the designation and antigen prepared from sheep
.trophoblast cells; Lee et al., 1986 stained sections of placentomal tissues comparing late
Fig. 8. Photomicrography of an SBU-3 immunohistochemically stained section of bovine placentome from theplacenta of a cow in the 8th month of gestation.Cross-sections of villi are surrounded by crypt walls of the caruncle. The larger cells that appear darker in thisphotomicrograph are binucleate trophoblast cells that are specifically stained with the SBU-3 antibody. Thereis slightly linear staining along the basement membrane of crypt epithelium. Compare this discrete cellularpattern with that in Fig. 9.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 155
Fig. 9. Photomicrography of an SBU-3 immunohistochemically stained section of bovine placentome from theplacenta collected during cesarean section.Note the deposition of stain along the area of the basement membrane of the caruncular crypts. Binucleatetrophoblast cells migrate from the surface of the villi to embed within and fuse with an endometrial epithelialcell followed by dramatic degranulation. They are then lost. Several stained trophoblast cells remain in thisphotomicrograph. The exact role binucleate cell migration, fusion and degranualtion play in parturition androrplacental release is not known.
Žgestation with the degranulated binucleate cells found immediately before delivery Fig..9 . The monoclonal antibody, originally ‘‘raised’’ against ovine placental tissues,
specifically labels binucleate trophoblast cells in ruminant placentas. It is probable thatthe antigen labeled is a bPAG. The bPAGs have been localized to binucleate cells, andperipheral systemic levels have been shown to rise over the last 10 days of pregnancyŽ . ŽPatel et al., 1997 . This is when marked binucleate cell degranulation occurs Figs. 8
.and 9 . It has been suggested that bPAG-1 is a proteinase and may function as anŽ .enzyme Roberts et al., 1995 . Its specific role in placental release is not known.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160156
Another important population of cells found in the bovine placenta are the fetalmacrophages. These cells are referred to in human medicine as ‘‘Hofbauer Cells’’Ž .Benirschke and Kaufmann, 1995 . Fetal placental macrophages may originate eitherfrom chorionic mesenchyme early in gestation, or from fetal bone marrow-derivedmonocytes. These cells produce pro-inflammatory cytokines and function in antigenpresentation suggesting that they can function as sentinel cells, are important in fetaldefense and may contribute to transplacental transmission of microbial agents.
5. Disease states
In twinning during bovine pregnancy, placental vessels nearly always fuse. Theestablishment of shared vasculatures results in fetal chimarism. In male–female twinpairs, this causes characteristic, marked changes in development of the gonads, andinternal tubular and external genitalia of the female fetus and is recognized as ‘‘free-martinism’’.
Placentation is important in understanding the pathogenesis of some congenitaldiseases, mechanisms of transplacental transmission of teratogenic microbial agents, and
Ž .failure of pregnancy of cloned calves Hill et al., 1999 . It is beyond the scope of thisreview to address all aspects of in utero infection. Those infectious diseases that producegrossly evident lesions are limited to chronic fungal and bacterial infections. Mycoticinfections, most commonly, Aspergillus sp., cause chronic placentitis with severe
Ž .necrosis and destruction of cotyledonary villi. Brucellosis and yeast infection Candidaalso result in chronic infection with ‘‘functional amputation’’ of cotyledonary tissuesleading to either in utero growth retardation, severe fetal distress, and commonly,
Ž .abortion Anderson et al., 1986; Foley and Schlafer, 1987 . Transplacental transmissionof most viral agents occurs with no grossly evident tissue destruction.
The most common disease of the bovine placenta is placental retention. The precisemechanisms that are triggered, and lead to placental release are not understood. Failure
Ž .of placental release is common values cited range from 3% to 12% and is higher inindividual animals that abort or calve prematurely, in cases of twinning, following
Ž .induced deliveries, etc. see review by Laven and Peters, 1996 . During normalmaturation prior to delivery, binucleate trophoblast numbers decrease dramatically but
Žthis change does not occur in placentas that are retained Gross et al., 1991; Williams et.al., 1987 . The mechanisms for this are not known.
Field studies have demonstrated an association between the major histocompatibilityŽ .complex MHC Class I compatibility of a dam and her fetus with placental retention
Ž .Joosten et al., 1991; Joosten and Hensen, 1992 . In cases where dam and fetus shareMHC Class I antigens, placental retention is more likely to occur. This suggests acentral role for histocompatibility in triggering an immune reaction that contributes toplacental release. Class I antigens have been detected on trophoblast cells in the
Ž . Ž .intercotyledonary area Low et al., 1990 . Davies et al. 2000 have recently expandedthis observation by showing that during normal pregnancy, MHC Class I is expressed bytrophoblast cells in the intercotyledonary areas and in the arcade area of the placentomefrom the 6th month of gestation.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 157
6. Placental macrophages: ontogeny during bovine pregnancy
Placentomes were collected at Taylor Packing Plant, Wyalusing, PA. Control tissuesŽincluded fetal gut, thymus, lymph node, and maternal mesenteric lymph node 6th month
. Ž .gestation . Gestation age was estimated from fetal crownrrump length Harris, 1983 .
Fig. 10. Change in numbers of fetal macrophages throughout gestation.This graph shows the number of fetal macrophages present in cotyledonary villi at 4, 6 and 8 months ofgestation, in placentas collected at term by cesarean section, and in placentas shortly after natural vaginaldelivery. Numbers of fetal macrophages, called Hofbauer Cells in human placentas, increase dramatically inthe villi during the last trimester of bovine pregnancy.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160158
Ž . Ž . Ž .Placentas were sampled from cattle at approximately 4 ns3 , 6 ns2 , and 8 ns3months gestation.
Periparturient placentomal samples were collected from seven first calf, holsteinŽheifers at C-section performed following a drop in progesterone -2 ngrml ProgestAs-
.say, Synbiotics, san Diego, CA . Placentomes were sampled from the mid-region of theuterine horn that contained the fetus. Post partum placentomes were collected from sixcases, obtained per vagina within 30 min of normal vaginal delivery.
During sampling, slabs 5 mm in thickness were carefully sectioned from the centralarea of each placentome perpendicular to the allantoic surface. Samples were frozen inchilled isopentane, cryosectioned at 8 mm, and fixed in cold acetone for 15 min. Afterblocking with normal serum, sections were incubated with primary monoclonal antibody
Ž . ŽSBU-3 Ctr. An. Biotec., Melbourne, Australia or CD68 EBM 11 Dako, Carpenteria,.CA , an antibody that labels macrophages, for 2 h at 378C in a humid chamber. Sections
Žwere then incubated with a biotinylated anti-mouse secondary antibody Vector Labora-. Žtories, Burlingame, CA followed by streptavidin peroxidase conjugate Zymed Labora-. Ž .tories, San Francisco, CA and visualized with AEC Zymed and hematoxylin. Non-im-
Ž .mune mouse ascites Zymed was used as a negative control.Analysis of the number of CD68 EBM 11 positive cells per unit area of tissue was
Ž .determined using Image-Pro Pluse Media Cybernetics image processing software. AŽ .total of 15 fields per placentome two per case were digitized. For each field, fetal villi
Ž .were traced and measured using a freeform ‘‘area of interest’’ AOI mechanism. Foreach AOI, macrophages were selected using the ‘‘color segmentation’’ function outlin-ing positively stained cells. Macrophage numbers were determined automatically, andcalculated per unit area of AOI.
Results of this study are presented in Fig. 10. Relatively few fetal macrophages werepresent in cotyledonary villi from fetuses younger than 6 months, but by 8 months,macrophages had increased by over 10-fold. Samples collected at cesarean section andfrom placentas sampled after vaginal delivery contained similar numbers. In cases ofmycotic, yeast, and bacterial placentitis, fetal macrophages become prominent within theallantoic mesenchyme in areas of inflammation. The role these cells play in fetalplacental defense and their likely role in transporting microbial agents from sites ofinflammation within the placenta to the fetus via umbilical veins needs further elucida-tion.
7. Conclusion
This paper has reviewed the basic elements of placental development, illustratedprominent gross anatomic features of bovine placentation, and discussed the cellularbiology of two of the most important cell populations, trophoblast and fetal macrophages.Original quantitative data has been presented that profiles the ontogeny of fetalmacrophages with cotyledonary villi, cells that play import functional role in in uterofetal defense, and possibly contribute, under some conditions, to transplacental transmis-sion of microbial agents from the placenta to the fetus.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160 159
Acknowledgements
Funds in support of this work have been provided by the Harold WetterbergFoundation and the USDA Animal Health and Diseases Research Program.
References
Anderson, T.D., Meador, V.P., Cheville, N.F., 1986. Pathogenesis of placentititis in the goat inoculated withBrucella abortus: Part I. Gross and histologic lesions. Vet. Pathol. 23, 219–226.
Benirschke, K., Kaufmann, P., 1995. In: Pathology of the Human Placenta. 3rd edn. Springer Verlag, NewYork, NY, pp. 84–92.
Davies, C.J., Fisher, P.J., Schlafer, D.H., 2000. Temporal and regional regulation of major histocompatabilitycomplex class I expression at the bovine uterinerplacental interface. Placenta 21, 194–202.
Duello, T.M., Byatt, J.C., Bremel, R.B., 1986. Immunohistochemical localization of placental lactogen inbinucleate cells of bovine placentomes. Endocrinology 119, 1351–1355.
Foley, G.L., Schlafer, D.H., 1987. Candida abortion in cattle. Vet. Pathol. 24, 532–536.Gross, T.S., Williams, W.F., Russek-Cohen, E., 1991. Cellular changes in the peripartum bovine fetal placenta
related to placental separation. Placenta 12, 27–35.Hill, J.R., Roussel, A.J., Cibelli, J.B., Edwards, J.F., Hooper, N.L., Miller, M.W., Thompson, J.A., Looney,
C.R., Westhusin, M.E., Robl, J.M., Stice, S.L., 1999. Clinical and pathological features of clonedŽ .transgenic calves and fetusus 13 case studies . Theriogenology 51, 1451–1465.
Jones, C.J.P., Koob, B., Stoddart, R.W., Hoffmann, B., Leiser, R., 1994. Lectin-histochemical analysis ofglycans in ovine and bovine near-term placental binucleate cells. Cell Tissue Res. 278, 601–610.
Joosten, I., Hensen, E.J., 1992. Retained placenta: an immunological approach. Anim. Reprod. Sci. 28,451–461.
Joosten, I., Sanders, M.F., Hensen, E.J., 1991. Involvement of major histocompatibility complex class Icompatibility between dam and calf in the aetiology of bovine retained placenta. Anim. Genet. 22,455–463.
Laven, R.A., Peters, A.R., 1996. Bovine retained placenta: aetioilogy, pathogenesis, and economic loss. Vet.Rec. 139, 465–471.
Lee, C.S., Gogolin-Ewens, K., Brandon, M.R., 1986. Comparative studies on the distribution of binucleatecells in the bovine placenta of the deer and cow using monoclonal SBU-3. J. Anat. 147, 163–179.
Low, B.G., Hansen, P.J., Drost, M., Gogolin-Ewens, K.J., 1990. Expression of major histocompatibilitycomplex antigens on the bovine placenta. J. Reprod. Fertil. 90, 235–243.
Munson, L., Kao, J.J., Schlafer, D.H., 1989. Characterization of glycoconjugates in the bovine endometriumand chorion by lectin histochemistry. J. Reprod. Fertil. 87, 509–517.
Munson, L., Wilhite, A., Boltz, V.F., Wilkinson, J.E., 1996. Transforming growth factor beta in bovineplacentas. Biol. Reprod. 55, 748–755.
Patel, O.V., Sulon, J., Beckers, J.F., Takahashi, T., Hirako, M., Sasaki, N., Domeki, I., 1997. Plasma bovinepregnancy-associated glycoprotein concentrations throughout gestation in relationship to fetal number inthe cow. Eur. J. Endocr. 137, 428–432.
Reeves, J.T., Daoud, F.S., Gentry, M., Eastin, D., 1972. Changes in urinary flow in bovine fetuses during lategestation: composition of amniotic and fetal body fluids. Am. J. Vet. Res. 33, 2159–2167.
Reimers, T.J., Ullman, M.B., Hansel, W., 1985. Progesterone and prostanoid production by bovine binucleatetrophoblast cells. Biol. Reprod. 33, 1227–1236.
Roberts, R.M., Xie, S., Nagel, R.J., Low, B., Green, J., Beckers, J.F., 1995. Glycoproteins of the aspartylproteinase gene family secreted by the developing placenta. Adv. Exp. Med. Biol. 362, 231–240.
Russe, I., Sinowatz, F., 1991. In: Embryologie der Haustiere., Imogen Russe. Paul Parey, Berlin, p. 164.Shuler, G., Wirth, C.K.K., Pfarrer, C., Leiser, R., Hoffmann, B., 1999. Immunolocalization of progesterone
receptors in bovine placentomes throughout mid and late gestation and at parturition. Biol. Reprod. 61,797–801.

( )D.H. Schlafer et al.rAnimal Reproduction Science 60–61 2000 145–160160
Williams, W.F., Margolis, M.J., Manspeaker, J., Douglass, L.W., Davidson, J.P., 1987. Peripartum changes inthe bovine placenta related to fetal membrane retention. Theriogenology 28, 213–223.
Wooding, F.B.P., 1982. The role of the binucleate cell in ruminant placental structure. J. Reprod. Fertil.,Suppl. 31, 30–31.
Wooding, F.B.P., 1992. Current topic: the synepitheliochorial placenta of ruminants: binucleate cell fusionsand hormone production. Placenta 13, 101–113.
Wooding, F.B.P., Wathes, D.C., 1980. Binucleate cell migration in the bovine placentome. J. Reprod. Fertil.59, 425–430.


















