
Pollen analysis of New Zealand honey - UMFpollen frequencies on the basis of a total count of pollen...
32
New Zealand Journal of Agricultural Research, 1985, Vol. 28: 39-70 0028 – 8233/85/2801-0039$2.50/0 O © Crown copyright 1985 39 Pollen analysis of New Zealand honey N. T. MOAR Botany Division, DSIR Private Bag, Christchurch, New Zealand Abstract A pollen analytical study of New Zealand honey provides a basis for identifying the origins of a honey in terms of locality and floral source. The information may be used to develop analytical standards for pollen, contributing to quality control of a product offered for export or for the home market. General principles outlined by the International Commission for Bee Botany have been used as a guide, although in practice these are considerably modified. Samples were processed by acetolysis, and absolute pollen counts were obtained by spiking with a known number of spores. Most New Zealand honey falls within the "normal" category (20 000-100 000 pollen grains in a standard 10 g sample). Clover honey is in this category. Thyme honey with a pollen content less than 20 000 grains per 10 g sample, and manuka honey where the pollen content exceeds 100 000 grains, are examples of "under-represented" and "over represented" categories respectively. The analyses confirm the importance of white clover to apiarists, they provide details of characteristic pollen spectra for New Zealand honey, and draw attention to difficulties associated with assessing honey type by organoleptic criteria. Standards developed for New Zealand honey by pollen analysis should be flexible enough to accommodate changes as more data become available. Lycopodium Keywords melissopalynology; pollen analysis; honey INTRODUCTION There has never been a systematic pollen analytical study of New Zealand honey, although nearly 70 years ago Waters (1915a, b, 1916) began a series of short papers describing pollen grains commonly found in local honey. The series was begun to "sat- isfy the increasing demand" for such information. Nevertheless, it was 30 years before the next seri- ous study was attempted. Harris & Filmer (1947) Received 27 July 1984 used pollen analysis during the search for the toxic principle in honey from Pongakawa, Bay of Plenty. It is now known that the toxin, a derivative of tutin, is incorporated into honey by bees which have worked the honeydew excreted by the naturalised passion- vine hopper, as it feeds on the sap of tutu plants (Sutherland & Palmer-Jones 1947; Connor 1977). Harris & Filmer (1948) extended their observations to include bee loads (pollen pellets) taken from hives in the same area, and their work showed that pollen analysis could contribute to a knowledge of New Zealand honey despite the difficulty of cor- relating pollen with nectar sources. They concluded that "if there were no relationship between the pro- portion of pollen and parent nectars, a more fortuitous occurrence of pollen maxima might be expected". Scolypopa australis, (Coriaria arborea) Nevertheless this optimism did not generate much response from apiarists, possibly because of a negligible export market, and home consumption favouring the production of clover honey. Pollen analysis therefore was not used as in Europe to determine geographical origins, or floral sources of honey (e.g., Crane 1975). As interest in creating export markets for characteristic New Zealand honeys increased, it became clear that customer requirements, including those of product quality, had to be met. These requirements were emphasised at the Christchurch Honeydew Seminar (1978) where matters relating to the production, utilisation, and recognition of honeydew honey were discussed. However, occasional pollen analyses have shown that hyphae of a sooty mould are characteristic of honeydew honey and that the pollen spectrum of a floral source honey is sometimes characteristic of the district in which it has been produced. The work reported in this paper expands these general observations into a more coherent pollen analytical statement of New Zealand honey. Identification of honey-type is based upon the results of pollen analysis and not upon the more traditional organoleptic characteristics of taste, colour, and odour. Beginning late in 1979, the study extended over 3 years. Besides establishing analytical criteria for pollen, it was hoped to provide apiarists with guidelines for monitoring the quality of their product as well as meeting the requirement of a microscopical examination sometimes demanded by importing countries.
Transcript of Pollen analysis of New Zealand honey - UMFpollen frequencies on the basis of a total count of pollen...

New Zealand Journal of Agricultural Research, 1985, Vol. 28: 39-700028 – 8233/85/2801-0039$2.50/0 O © Crown copyright 1985
3 9
Pollen analysis of New Zealand honey
N. T. MOARBotany Division, DSIRPrivate Bag, Christchurch, New Zealand
Abstract A pollen analytical study of New Zealandhoney provides a basis for identifying the origins ofa honey in terms of locality and floral source. Theinformation may be used to develop analyticalstandards for pollen, contributing to quality controlof a product offered for export or for the homemarket. General principles outlined by theInternational Commission for Bee Botany havebeen used as a guide, although in practice these areconsiderably modified. Samples were processed byacetolysis, and absolute pollen counts were obtainedby spiking with a known number ofspores. Most New Zealand honey falls within the"normal" category (20 000-100 000 pollen grains ina standard 10 g sample). Clover honey is in thiscategory. Thyme honey with a pollen content lessthan 20 000 grains per 10 g sample, and manukahoney where the pollen content exceeds 100 000grains, are examples of "under-represented" and"over represented" categories respectively. Theanalyses confirm the importance of white clover toapiarists, they provide details of characteristicpollen spectra for New Zealand honey, and drawattention to difficulties associated with assessinghoney type by organoleptic criteria. Standardsdeveloped for New Zealand honey by pollenanalysis should be flexible enough to accommodatechanges as more data become available.
Lycopodium
Keywords melissopalynology; pollen analysis;honey
INTRODUCTION
There has never been a systematic pollen analyticalstudy of New Zealand honey, although nearly 70years ago Waters (1915a, b, 1916) began a series ofshort papers describing pollen grains commonlyfound in local honey. The series was begun to "sat-isfy the increasing demand" for such information.Nevertheless, it was 30 years before the next seri-ous study was attempted. Harris & Filmer (1947)
Received 27 July 1984
used pollen analysis during the search for the toxicprinciple in honey from Pongakawa, Bay of Plenty. Itis now known that the toxin, a derivative of tutin, isincorporated into honey by bees which have workedthe honeydew excreted by the naturalised passion-vine hopper, as it feeds on thesap of tutu plants (Sutherland &Palmer-Jones 1947; Connor 1977). Harris & Filmer(1948) extended their observations to include beeloads (pollen pellets) taken from hives in the samearea, and their work showed that pollen analysiscould contribute to a knowledge of New Zealandhoney despite the difficulty of cor- relating pollenwith nectar sources. They concluded that "if therewere no relationship between the pro- portion ofpollen and parent nectars, a more fortuitousoccurrence of pollen maxima might be expected".
Scolypopa australis,(Coriaria arborea)
Nevertheless this optimism did not generate muchresponse from apiarists, possibly because of anegligible export market, and home consumptionfavouring the production of clover honey. Pollenanalysis therefore was not used as in Europe todetermine geographical origins, or floral sources ofhoney (e.g., Crane 1975). As interest in creatingexport markets for characteristic New Zealand honeysincreased, it became clear that customer requirements,including those of product quality, had to be met.These requirements were emphasised at theChristchurch Honeydew Seminar (1978) wherematters relating to the production, utilisation, andrecognition of honeydew honey were discussed.However, occasional pollen analyses have shown thathyphae of a sooty mould are characteristic ofhoneydew honey and that the pollen spectrum of afloral source honey is sometimes characteristic of thedistrict in which it has been produced. The workreported in this paper expands these generalobservations into a more coherent pollen analyticalstatement of New Zealand honey. Identification ofhoney-type is based upon the results of pollen analysisand not upon the more traditional organolepticcharacteristics of taste, colour, and odour. Beginninglate in 1979, the study extended over 3 years. Besidesestablishing analytical criteria for pollen, it was hopedto provide apiarists with guidelines for monitoring thequality of their product as well as meeting therequirement of a microscopical examinationsometimes demanded by importing countries.

40 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Fig. 1 Localities from which honey samples were obtained. In many instancesthe location is a generalised one involving several neighbouring districts whichare indicated below. Details of site locations are available from the author onrequest. North Island: 1 Whangarei; 2 Dargaville; 3 South Head KaiparaHarbour; 4 Rangitoto Island; 5 Auckland; 6 Waerenga; Te Kauwhata,Rotongaro; 7 Whangamata, Paeroa; 8 Gordonton, Morrinsville, Hamilton,Eureka, Matamata, Taetoaroa, Ohaupo; 9 Broadlands, Manawaru, KaimaiRange, Paeroa, Papamoa- Tauranga; 10 Otorohanga, Waitomo, Te Kuiti; 11Tokoroa; 12 Taneatua; 13 Mangakino, Whakamaru, Taupo, Broadlands; 14Ruatoria, Hiruharama; 15 Rakauroa, Otoko, Tolaga Bay, Gisborne, Poverty Bay;16 Taupo Road, Raupanga, Wairoa; 17 Opunake; 18 Waverley; 19 Raetihi; 20Kimbolton, Aokautere, Linton; 21 Greytown. South Island: l Nelson; 2Blenheim; 3 Molesworth; 4 Greymouth/Westport area; 5 Blackball, Lady Lake,Haupiri; 6 Taramakau River; 7 Hanmer; 8 Kaikoura; 9 Fox Creek, HorsfordDowns, Okuku Pass, Mt. Thomas, Ashley, Oxford, Swannanoa, Kimberley; 10Hororata, Darfield; 11 Rakaia Gorge, Pudding Hill, Staveley; 12 Franz Josef; 13Chertsey, Seafield, Laghmor; 14 Benmore, Otematata, Aviemore; 15 Glenavy,Pukeuri, Ngapara, 16 Tarras; 17 Poolburn, Ranfurly; 18 Alexandra, Fruitlands,Roxborough, Dumbarton; 19 Milford Sound/Te Anau high- way; 20Mandeville; 21 Tahakopa Valley.
The International Commission for Bee Botany, acommission of the International Union of Bio-logical Sciences, has published recommendationsrelating to the palynology of honey based on manyyears of experience in Europe (Louveaux et al.1978). The objectives are to assist in thedevelopment of quality control by pollen analysis, todiscover geographical origins, and to determine thenectar source of any particular honey. The principalmethods involve qualitative and quantitative pollenanalysis. The first permits calculation of relativepollen frequencies on the basis of a total count ofpollen and other microscopical particles, and therecognition of a pollen spectrum characteristic of ahoney from a particular area. The
second (quantitative analysis) involves calculationof the absolute number of pollen grains in a honeysample, which can be used as a basis fordetermining the floral source of any honey, as wellas assessing claims made by an apiarist as to thepurity of the product. This report discusses theresults of pollen analysis of 119 samples from manyparts of New Zealand, although not every region isrepresented (Fig. 1). The number of specialisthoneys received, in contrast to mixed source andclover honey, was not great, and some of them wererejected as such after pollen analysis. Despite theselimitations the results should provide a basis for abetter understanding of New Zealand honey, and atechnique which can

Moar – Pollen analysis
be modified or improved in the future to extend andto test the observations reported here.
METHODS
Because of the need to deal with many samplesquickly, involving the counting of thousands ofpollen grains, simple and rapid laboratory procedureswere required. The procedure adopted is based upona 10 g honey sample, standard acetolysis (Erdtman1943), and "spiking" – adding tab lets containing aknown number of spores to the sample(Stockmarr 1971). Two batches of tablets were usedcontaining 12 500 500 and the second 10 850 200spores per tablet respectively. Two tablets from thesame batch were added to each sample.
The number of pollen grains counted for qualitativeanalysis varies according to the information required,and the principles outlined by Louveaux et al. (1978)are adopted in this paper. The simplest, intended toestablish the general character of the honey, involvesthe identification of only the most numerous, orcharacteristic pollen grains, and is defined as anorienting analysis. More detailed counts, referred toas complete analyses, are used to determine thegeographical origins of a honey, and involveidentification of every pollen-type noted andcalculation of their relative frequencies in counts ofup to 300 pollen grains. The pollen- types aredistributed among frequency classes, eachrepresenting a defined range as follows: predominantpollen, 45% or more; secondary pollen, 16- 44%;important minor pollen, 3-15%; minor pollen, <
Lycopodium
" "
"
3%.Louveaux et al. (1978) state that counts of 1200 ormore pollen grains per sample are necessary to applyreliable percentage frequencies to individual pollen-types, expressed with an accuracy of 1%.
For the present study complete analyses werealways made and, for many samples, invo1vedcounts of 1000 or more pollen grains. However,consistent results were obtained with lower counts(Table 1) and as a general rule a total of 500 pollengrains was considered adequate, although thestandard error at 2% was greater. The pollen sumexcludes pollen of anemophilous (wind-pollinated)plants, nectarless plants, and any other microscopicparticles which may be present. Quantitative data arebased on results of complete analyses, and theabsolute pollen content of the 10 g sample is derivedfrom the ratio of the total pollen counted to thenumber of spores counted during thepollen analysis. Details of processing and countingare presented in Appendix l.
"
Lycopodium
41
honey depends upon the purity of the sample. Mostsamples received for pollen analysis were named inthe field on the basis of their organolepticcharacteristics and were as pure as the collectorcould obtain. Some however were known to beblends, or to have been taken from commercialpacks. These were processed and examined, buttheir results were not used when assessing thecharacteristics of a unifloral honey. A unifloralhoney is derived mainly from one species, but notexclusively so (Louveaux et al. 1978) and isextracted by standard methods of centrifugation.Most unifloral honey, including white clover,contains 20 000-100 000 pollen grains in a standard10 g sample, and in this respect is regarded as"normal" (Crane 1975) – the principal pollen type ispredominant and therefore present in frequencies of45% or more. The term "single species honey" isreserved for honey produced by bees working onespecies under controlled conditions. In this context a"normal" honey derived from various floral sourceswithout any one predominating is a "mixed sourcehoney". Since pollen production varies greatlybetween species, some unifloral honey may containless than 20 000 pollen grains, and some more than100 000 pollen grains. Honey in the lower range is"under-represented" in terms of "normal" absolutepollen content, and that in the higher range is "over-represented". Because the quality of New Zealandthyme honey is sometimes questioned by animporting country it was necessary to collect thepurest honey possible for pollen analysis. Ministryof Agriculture and Fisheries officers attempted to dothis by placing hives in 2 areas dominated by thymeat the beginning of its flowering period andremoving them when flowering had passed its peak.The honey produced during this time was extractedby hand centrifuge. Having established that thepollen content of a sample is under- or over-represented in relation to normal honey, the nextstep is to calculate the minimum percentagefrequency that defines it as a unifloral honey. Forthis only the purest samples are used, and as notedalready, samples received as blends, commercialpacks, or without collecting details are discarded.Average values for absolute pollen content andrelative frequencies for particular pollen types arecalculated and corrected for the minimum "normal"value of 45%. This value is then expressed as apercentage of the sum of the average absolute pollencontent of the pollen-type under review and theaverage absolute pollen content of clover-typepollen in "normal" clover honey. The followingcalculations, using thyme honey as an example,illustrate this.

42 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Table 1 4Percentage frequency of main pollen types in each of 7 slides, based on pollensums ranging from 100 to 1000.
(1) Average values of thyme pollen in thymehoney.
(3) Minimum frequency thyme pollen in unifloralthyme honey
5 8 0 1 100X = 20%
5801 + 23 116 1
Because present data are limited, calculated valuesare accepted as guides which may be modified as moresamples are analysed.
Correction factors can be determined experimentally(Maurizio 1975), but these demand long- term studiesand guaranteed sample purity, which were beyond thescope of the present investigation.
Pollen grains were identified by comparison withprepared reference slides, and the level ofidentification depended upon the characteristics ofeach pollen type. Thus, although white clover
is an important nectar source and itspollen is found in most New Zealand honeys, it issometimes difficult to separate from the pollen of other
(Trifolium repens)
=

Moar – Pollen analysis
clovers present as minor components in a sample.It is therefore recorded as white clover-type pollen.However, the characteristic pear-shaped pollen ofvipers bugloss allows identifica-tion to the species level.
(Echium vulgare)
RESULTS
The results of all pollen analyses are tabulatedaccording to floral source. Most samples are listedas a named unifloral honey although some aretreated as being derived from mixed sources (seeTable 12).
The samples forwarded for pollen analysis werenamed in the field on the basis of organolepticcharacteristics, i.e., flavour, odour, and colour. Itwas realised early in the survey thatcharacterisation by pollen analysis may be atvariance with this traditional method ofidentification as illustrated by the followingexamples.
Barberry buttercupling nodding
thistle and tawarihoney samples were received and
examined during the survey. In most instances thepollen frequency of the named source was so lowas to suggest mis- identification, especially asanother pollen, often white clover-type, waspredominant. A sample received as barberry honey(H24) contained
(Berberis vulgaris),(Ranunculus), (Calluna vulgaris),
(Carduus nutans), (Ixerbabrexioides)
19% barberry pollen, 82% whiteclover-type pollen, and 2% buttercup pollen.Although barberry pollen tends to fragment underacetolysis, there were very few broken grains notedduring the counts, so the predominance of whiteclover-type pollen was not fortuitous. The samplesreceived as nodding thistle honey (H46, H116,H122, H123, H124) and tawari honey (H40, H47,H186) are classed as clover honey, as pollen of thenamed plant is either absent or present in lowfrequencies.
Two samples, H45 and H192, were received aspenny royal honey. The firstcontained 59% white clover-type pollen and 6%penny royal pollen, and is regarded as a cloverhoney (see Table 3). The second was dominated bylotus pollen (65%), with penny royal (14%) beingthe next most common pollen – it is listed as lotushoney (see Table 10). Penny royal pollen wasrecorded as a minor element in another 18 sam-ples, but the flavour of penny royal nectar wasdetected in only 2 of these (H68 and H125). Bothcontained 2% penny royal pollen; the first isclassed as clover honey, and the second as a mixedsource honey (see Tables 3 and 12). Unifloralpenny royal honey is rare in Europe where thepollen is mostly recorded as a minor component inhoney (Maurizio & Louveaux 1962).
(Mentha pulegium)
43
Heather honey, whether derived from ling or Spanishheath has a characteristic flavour(Walsh 1967). Ling pollen is poorly represented (2%)in the only ling honey sample (H26) received duringthe survey, whereas white clover- type pollen andlotus spp.) pollen together accounted for 94%of the pollen sum. However, a honey from Glenledi,near Milton, (H63) of uncertain source, but "probablySpanish heath", contained secondary pollen including
(21%), gorse (25%), and manuka (31%).Maurizio & Louveaux (1964) note that ling producesan almost pure honey in parts of Europe and that thepollen is predominant or secondary in importance(see also Maurizio 1949, 1979). It is interesting that aNew Zealand ling honey containing only 1% lingpollen was acceptable in Germany as a ling-typehoney, but not as ling honey, on the basis of itsorganoleptic properties (Rope pers. comm.). Thereare insufficient data to characterise New Zealandheather honey on pollen analytical criteria with anyconfidence, and until better documentation isavailable the Glenledi sample is treated as a mixedsource honey, and the ling sample as a clover honey.
(Erica lusitanica)
(Lotus
Erica
Ability to identify honey on the basis of traditionalmethods varies between individuals who mayrecognise different sources for the same, or similar,product. Four samples (H27, H126, H130, H191)identified by flavour as manuka
buttercup, manuka blend, and manukarespectively, illustrate this point. The "manuka" (H27)and the "buttercup" honey (H126) each contained <
(Leptospermumscoparium),
1%buttercup pollen and a similar frequency of manukapollen (22 and 20% respectively). In the "manuka"blend (H130), 59%, manuka pollen, and (19%buttercup pollen was recorded in contrast to the 55%manuka pollen and 5% buttercup pollen in the"manuka" honey (H191). These 4 are classed as mixedsource honeys (see Table 12) on pollen analyticalcriteria, and emphasise the difficulties involved inidentifying nectar sources consistently and accuratelyby flavour. One hundred and twenty pollen types, and afew fern spores, were identified during the pollenanalyses. Thirteen of the pollen types are derived fromwind-pollinated plants of which dock grass(Gramineae), and plantain are the mostregularly recorded, albeit in low numbers. Beechpollen is regularly present inhoneydew honey and the rest are sporadic in theiroccurrence. All other pollen types come from nectarproducing plants, except Californian poppy
kawakawapigeon wood
and pukatea theflowers of which produce little or no nectar. Theirpresence in honey is probably
(Rumex),(Plantago)
(Nothofagus fusca-type)
(Eschscholtzia californica), (Macropiperexcelsum), Muehlenbeckia, (Hedycaryaarborea), (Laurelia novae-zelandiae)








Moar – Pollen analysis
accidental, although pukatea and Californian poppyare known to be worked by bees for pollen (Walsh1967; Maurizio & Grafl 1982).
Grass, white clover-type, and Rosaceae pollenoccurred in almost every sample, and dandelion
lotus, thistle, and willowin more than half the samples received.
Most pollen recorded occur as minor orsecondary components in honey, and only a few –kamahi manuka,matagouri rataspp.), vipers bugloss, and white clover-type – areconsistently predominant. Thyme pollen
is usually recorded as a secondary pollen(16 – 44% in thyme honeys, although it isoccasionally predominant ( > 45%). Rewarewapollen is either an importantminor (3-15%) or secondary pollen in rewarewahoneys.
The number of pollen types in any one sampleis not great (generally 9-22). The lowest number (4)was recorded in the only Chatham Island samplereceived (H29), and the highest (33) occurred in arewarewa honey (H206).
(Taraxacum-type), (Salix)
(Weinmannia racemosa),(Discaria toumatou), (Metrosideros
(Thymusvulgaris)
(Knightia excelsa)
)
Geographical origins
A New Zealand honey can usually be recognised byits pollen spectrum. However, to identify the districtfrom which a honey is derived, pollen analyticalcharacteristics need to be closely defined. Thepresent survey has gone some way in achieving thisobjective, and the results demonstrate that a localitymay sometimes be identified as much by thepresence of a particular pollen, as by the pollenspectrum.
Thus, goats rue pollen (Galega officinalis)
Elaeocarpus, (Quintinia),
occursonly in honey from the Manawatu district in thesouthern North Island, and relatively highfrequencies of thyme pollen, sometimes associatedwith pollen of Californian poppy, are characteristicofhoney from the Central Otago valleys of theClutha, Kawarau, and Manuherikia Rivers (Wilsonet al. 1979). Similarly, honey with relatively highfrequencies of vipers bugloss pollen probably comesfrom the drier inland areas of eastern South Island.Tawari pollen restricts the source area to forestedsites in central and northern North Island, andrewarewa pollen identifies any North Island districtwhere there are forests or forest remnants withrewarewa. Rewarewa extends south into the extremenorth-east of the South Island, but so far honey fromthis area has not been available for examination.Whereas rewarewa and tawari pollen identify aNorth Island honey, the combination of kamahi,
quintinia and rata pollengenerally is representative of South Island Westlandhoneys. Although clover honey is producedthroughout New Zealand, careful scrutiny of
51
pollen spectra separates a North Island from a SouthIsland honey (see especially Tables 2 and 3).
Pollen analytical characterisation of honey
The pollen content of the unifloral, mixed source,and honeydew honeys identified are reported, andcomplement the tables which record the details ofeach pollen analysis.
Unifloral honey
(1) Clover honey (Tables 2 and 3)Fifty-five samples are listed as clover honey – 27
from the South Island and 28 from the North Island.Honey from the North Island contains more pollentypes than that from the South Island and each reflectsthe different combination of species available to bees.White clover-type pollen was the dominant pollentype in North Island samples. Lotus pollen, the nextmost important pollen type in North Island samples,along with low frequencies of dandelion and grasspollen. The combination of these with a mixedrepresentation of other herbs (buttercup, thistle) andsome pollen derived from native shrubland (manuka)or forest (rewarewa, tawari) identifies a North Islandorigin.
South Island honey is clearly dominated by whiteclover-type pollen and there are low frequencies ofgorse, Rosaceae, and other herbs in most samples.This combination of pollen types is especiallycharacteristic of pastoral areas in Canterbury andNorth Otago; inland areas are often identified by thepresence of bulbinella, matagouri, and vipers buglosspollen.
Absolute pollen counts for white clover-type pollenvaried from 16,000 to 120,000, and most were in the20,000-100,000 range so that a minimum frequencyof 45% satisfies the requirement of a unifloral honey.
White clover is so widely grown that clover honeyis produced throughout New Zealand and whiteclover-type pollen can be expected in varyingproportions in most honey.
(2) Thyme honey (Table 4)Eight samples are listed as thyme honey.Thyme, matagouri, and white clover-type pollen
occur in every sample, and other frequently recordedpollen-types are dock, grass, gorse-type, vipersbugloss, and willow.
Absolute counts for thyme pollen varied from 3700to 9100 in the purest samples examined (H18, H61,H62, H162). On this basis thyme pollen is under-represented in honey and it is calculated that aminimum of 20% thyme pollen is required tocharacterise a unifloral honey; this is in accord with

52 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Table 4 Pollen analysis of thyme honey, Central Otago, South Island.(Thymus vulgaris)
Locality
European experience (Maurizio 1946, 1975). Thymehoney is a product of Central Otago (South IslandLocation 16, Fig. 1) which is the only district in NewZealand where thyme occurs as a dominant in thenaturalised vegetation.
An attempt to produce an especially pure thymehoney failed. The unexpectedly high frequencies of
Umbelliferae pollen in samples H155and H156 reflected the presence of
(Aciphylla-type)Aciphylla plants
which were subsequently discovered in large numbersnear the thyme communities in which the hives werelocated. Sample H155 is accepted as a thyme honey, butH156 is treated separately as Umbelliferae honey (seeTable 10).

Moar – Pollen analysis Pollen analysis of manuka honey,North Island.
Table 5 (Leptospermum)
Locality
Pollen source
LeptospermumLotusSalixTrifolium repens-type
Cirsium-typeRosaceae
MetrosiderosTaraxacum-type
LoniceraRanunculusEucalyptusAcacia
ElaeocarpusHedycaryaKnightia
LigustrumMenthaMida
PhormiumPseudopanaxQuintinia ThymusTilia
Weinmannia
Caryophyllaceae
Labiatae
Onagraceae
Ulex-type
Gramineae (W)
(W)Cyperaceae(W)
(W)(W)
(W)
CoriariaCoprosma
PlantagoRumexCyathea
Pollensum APF
53
(3) Manuka honey (Table 5)Six samples are listed as manuka honey.
Manuka, grass, lotus, white clover-type, andwillow pollen occur in every sample, andRosaceae and thistle pollen are present in 5 ofthem. Manuka pollen is clearly predominant, andother pollen is almost always present in the"minor" category. Absolute pollen content is inexcess of 100 000, and in this respect manukapollen is over-represented in comparison with"normal" honey, and minimum frequencies of 70%manuka pollen are
necessary to classify a sample as unifloral manukahoney. Sample H2 has an absolute pollen count ofover 1 000 000; it cannot be compared with othersamples because the honey was extracted by"crushing and straining" (Simpson pers.comm.) andnot by centrifugation. Manuka honey has a strongflavour and is produced wherever there are suitableshrublands, often in areas of regenerating forest. Twotaxa – manuka andkanuka produce identicalpollen
(Leptospermum scoparium)(Leptospermum ericoides) –

54 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Table 6 Pollen analysis of rata honey. Sample H 1 was received as pohutukawahoney from North Island Locality 4. The locality for H145 is not precisely known,but it originated in Westland.
(Metrosideros)(M. excelsa)
Locality
Pollen source
MetrosiderosLotusTaraxacum-typeTrifolium repens-type
ElaeocarpusQuintiniaWeinmannia
LeptospermumPennantiaRosaceae
AristoteliaCirsium-typeRanunculus
CordylineUlex-type
cf. LigustrurnNothofagus fusca-typeRipogonumRubus
Ericaceae
Acaena
HoheriaKnightia
LoniceraMedicagoPhormiumPseudopanaxCiriseliniaSalixSchefflera
CistaceaeCompositae
Labiatae
Gramineae (W)(W)
(W)(W)
Coprosma DacrydiumcupressinumPlantago
Pollen sumAPF
grains. Both produce honey of similar flavour, butWalsh (1967) comments that kanuka is anunreliable source of nectar although a honeysurplus is sometimes produced. In view of thiscomment the samples examined are accepted asmanuka honey.
(4) Rata honey (Table 6)Six samples are listed as unifloral rata honey, but
only 3 (H95, H97, H145) were used to determineabsolute pollen content. The others were receivedas commercial packs and not used for that reason.

Moar – Pollen analysis
Table 7 Pollen analysis of kamahi honey. The precise locality ofSample H146 is not known.
(Weinmannia)
Locality
Pollen source
Weinmannia
Elaeocarpus
Trifolium repens-type
Rosaceae
LeptospermumLotusMetrosiderosTaraxacum-typeUlex-type
AristoteliaHedycaryaLigustrumQuintinia
Cirsium-type
Knightia
Laurelia
Muehlenbeckia
Pennantia
Phormium
Pittosporum
Ranunculus
Ripogonum
Rubus
Salix
Tetrapathaea
Trifolium pratense
Plantago
Coprosma
Nothofagus fusca-type
Gramineae (W)(W)(W)
Cyperaceae (W)(W)
Unidentified
Pollen sumAPF
55
Besides rata, characteristic pollen types are:dandelion; kamahi; lotus; quintinia;and white clover-type. Kaikamako
and Rosaceae are each represented in5 samples, and only 11 out of the 37 recorded arecommon. Absolute pollen content varies withinthe "normal" range although it tends towards theupper limit of 100,000 pollen grains. The honey istreated as "normal" so that a minimum frequencyof 45% rata pollen identifies a rata honey.
Elaeocarpus;(Pennantia
corymbosa)
A sample (Hl) received from North Island Locality5 as pohutukawa honeycontains few forest pollen and reflects the pastoralcharacter of the locality. Its North Island originsare confirmed by the presence of rewarewa pollen.Pohutukawa and rata pollen are similar inappearance, and size differences are so slight thatseparation is not easy. The term rata may involvenorthern rata or southern rata (M.
and since their ranges overlap thenectar
(Metrosideros excelsa)
(M. robusta)umbellata),

56 New Zealand Journal of Agricultural Research, 1985, Vol. 28
source may be difficult to determine. However,most samples have originated in areas of westernSouth Island outside the limits ofwhere is often a distinctive memberof lowland or montane forests.
M. robustaM. umbellata
Table 8 Pollen analysis of matagourihoney, South Island.
(Discaria)
Locality
(5) Kamahi pollen (Table 7)Five samples, 2 from the North Island (H44,
H52) and 3 from the South Island (H146, H178,H193) are listed as kamahi honey.
Characteristic pollen types are kamahi, pokaka,Rosaceae, and white clover-type – dandelion,gorse, lotus, manuka and rata each occur in 4 outof the 5 samples.
The pollen spectra in North and South Islandsamples are similar, although the records ofpukatea and rewarewa pollen in Sample H44 areevidence of its North Island origin. Kohia
extends to about latitude44S in the South Island, so its presence in thesample does not positively identify a North Islandhoney.
Absolute pollen content is less than 100,000grains in the North Island samples and greater than100,000 in the South Island samples. The averagevalue is about 140,000 grains per sample. On thisbasis, pollen in kamahi honey is "over-represented", although an accurate assessment ofthe required frequency for a pure honey is notpossible from available data, but probably liesbetween 60 and 70%.
Kamahi honey is most frequently produced in thewestern South Island although some comes fromSouthland (South Island Locality 21) and from theNorth Island.
(Tetrapathaea tetrandra)
(6) Matagouri honey (Table 8)Three samples are treated as matagouri although
none were received under that name. One (H59)came from a commercial pack of vipers buglosshoney, a second (H78) was from an unknownsource, and the third (H16) was labelled as thymehoney.
The pollen spectra are variable and reflect thecharacteristics of the districts in which the honeywas produced. Thus the relatively high frequencyof thyme pollen identifies Central Otago (South1sland Locality 18) as the source of H16, and thefrequency of vipers bugloss pollen confirms aninland area of eastern South Island as the sourcefor H59. Absolute pollen content is high,suggesting that pollen is over-represented, butsince the collection details of one sample are notknown, and another is from a commercial pack,quantitative data are treated with reserve, and untilmore information is available no attempt will bemade to determine minimum pollen frequencies.
DiscariaRosaceae
F.chiumMedicagoSalixTaraxacum-typeTrifolium repens-typeUnidentified
Bulbinella
Cordyline
Cyathodes fraseriEucalyptusLeptospermumLotusMetrosideros
ParsonsiaPhormiumRanunculus
Thymus
Campanulaceae
Cruciferae
Papilionaceae
Thymelaceaceae
Ulex-type
Gramineae (W)(W)
(W)(W)
PlantagoNothofagus fusca-
typeRumex
Pollen sumAPF
(7) Vipers bugloss honey (Table 9) Five samplesare listed as vipers bugloss honey. Characteristicpollen types are vipers bugloss, gorse- type,matagouri, white clover-type, and Rosaceae.Absolute pollen counts, based on only a fewsamples, fall into the "normal" category, and onthis basis a minimum of 45% vipers bugloss pollenis required to identify a unifloral honey. Vipersbugloss occurs in the North Island, but it isabundant only in eastern districts of central andnorthern South Island where it occurs along road-sides and in poor, gravelly soils.
(8) Citrus honey (Table 10)Only one sample (H129) was studied. According
to Vorwohl (1973) and Maurizio (197S) citruspollen is grossly under-represented(Citrus) and a
frequency of between 10 and 20% citrus pollen

Moar – Pollen analysis Pollen analysis of vipers buglosshoney, South Island.
Table 9 (Echium vulgare)
Locality
Pollen source
EchiumDiscaria
Rosaceae
Ulex-typeTrifolium repens-type
Cirsium-typeSalix
EucalyptusEscholtziaLeptospermumTaraxacum-typeTrifolium pratensePapilionaceae
CaryophyllaceaeCruciferaeEricaceaeLigustrumLotusMedicagoMenthaMuehlenbeckiaPhormiumRanunculus
Gramineae (W)(W)Plantago
Pollen sumAPF
57
identifies a unifloral honey. The single samplemeets this criterion – the absolute pollen content is11,545 of which citrus pollen accounts for only1,288, and the relative frequency of citrus pollen is11% The pollen spectrum is characterised byrelatively high frequencies of citrus, kiwifruit
.
(Artinidia chinensis, ), ,18% willow (13%) andbuttercup (13%) pollen, all of which are usuallypoorly rep- resented in honey. According to Walsh(1967) there is not much citrus honey producedalthough it provides a good pollen source innorthern North Island.
(9) Lotus honey (Table 10) Three samples arelisted as lotus honey. The first (H29) was receivedas clover honey from the Chatham Islands, thesecond (H149) was labelled "pure bush honey",and the third (H192) was received as penny royalhoney. Their diverse ori- gins are not clearlyreflected by the pollen spectra,
and there is no pollen which characterises theChatham Islands as the source for sample H29.Since no samples were received as "pure" lotushoney no attempt has been made to determineabsolute pollen values for a unifloral honey – the 3samples are treated as lying within the "normal"range of honeys.
(10) Rewarewa honey (Table 11) Four samples(H41, H134, H207a, H207b) are listed as unifloralrewarewa honey. In these samples rewarewa,dandelion, kamahi, lotus, whiteclover-type, and willow pollen are always present,and several forest pollen types, including manuka,pigeon wood, rata, tawari, and wineberry occur inmost of them. Absolute pollen counts varyconsiderably and offer no firm basis fordetermining the characteristics of a unifloralhoney, although on present evidence a minimumfrequency of 10% rewarewa pollen is probable,
Genios- toma,

58 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Table 10 Pollen analysis of (N2, N14, Chatham Island),(N15) and (S18) honey.
Lotus CitrusAciphylla
Locality
Pollen source
Trifolium repens-typeLotusActinidiaCitrusDiscariaMenthaRanunculus
SalixThymusAciphyllaAcacia
Rosaceae
AesculusAmaranthusAristoteliaCirsium
CordylineCotula
ElaeocarpusEucalyptusHedycaryaIxerbaLeptospermumMetrosideros
PseudopanaxTaraxacum-typeUlexViciaWeinmanniaCoprosmaCoriaria
Nothofagus fusca-type
Compositae
Cruciferae
Papilionaceae
(W)(W)
Gramineae (W)
Pollensum APF
Seven samples were received as rewarewahoney. Rewarewa pollen frequencies varied from12 to 31% in the 4 listed unifloral honey samplesand from 2 to 4% in the other 3 (H44, H188,H206). Because of these differences, 3 samplesreceived as buttercup honey (H28, H117, H126), 2samples received as tawari honey (H40, H47), and1 sample received as ling honey (H26), allcontaining low frequencies of rewarewa pollen(1-8%) are included in Table 11 for comparison.The pollen analyses show that all those sampleswith low frequencies of rewarewa
,
pollen have similar spectra, and on this basis theyare classed as clover honey (H26, H28, H40, H47),kamahi honey (H44), or mixed source honey(H117, H126, H188, H206). The only unifloralrewarewa honey samples recognised are the 4cited at the beginning. Rewarewa honey isproduced in lowland areas of the North Islandwhere areas of secondary forest occur. The treegrows in the Marlborough Sounds (north-eastSouth Island), but honey from this locality has notbeen investigated.

Moar – Pollen analysis
Mixed source honey (Table 12)
Seventeen samples are listed in this category inwhich the main pollen types recorded aresecondary or minor elements. Of these, 10 werereceived as named honey, 4 (H54, H126, H131,H132) as mixed source honey, and 1 (H117) wasof unknown origin. Those named were kamahi(H67), manuka (H27, H113, H115, H130, H191),rata (H119), rewarewa (H188, H206), and vipersbugloss (H158). Some were received incommercial packs, and none met the criteria forunifloral honeys. Twelve samples came fromvarious North Island localities, and the records ofrewarewa, tawari, and other forest pollen types insome of them, usually as minor components,reflect their origins. Similarly, the relatively highfrequencies for rata pollen in (H119) and vipersbugloss pollen in (H158) are evidence of the SouthIsland origin of these 2 samples. As usual, theNorth Island honey samples contain more pollentypes, including buttercup, grass, lotus, manuka,Rosaceae, willow, and white clover-type. OneNorth Island sample (H54) contained a few fungalspores. Absolute pollen content varies from about20,000 to 150,000 pollen grains, and most liewithin the "normal" range of20,000-100,000/10 g sample, a result similar tothat obtained by Maurizio (1949) in her survey ofpollen in mixed source Swiss honeys.
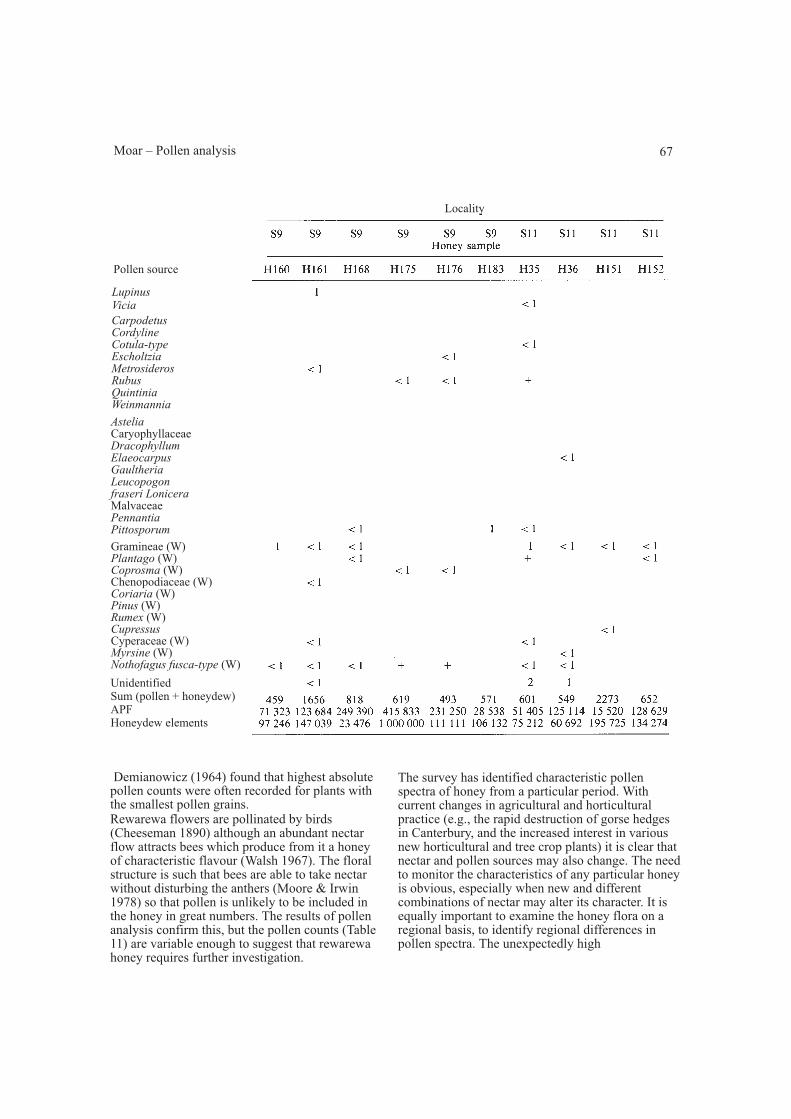
Honeydew honey (Table 13)Of the 19 samples examined, only one (H96) wascollected outside Canterbury. This sample, fromLady Lake, near Lake Brunner, contained highrelative frequencies of pollen from kamahi,quintinia, and rata – trees absent from Canterburybeech forests. Every sample contained the sporesand hyphae of a sooty mould (Hughes 1972)together with dandelion, gorse-type, and whiteclover-type pollen. Beechgrass, manuka, and willow pollen were recorded inmost samples. The honeydew elements wereincluded in the sum although their counts werevariable, a result which may be influenced by theacetolysis technique used to process the samples.There are pollen grains of 9 wind-pollinated plantsrepresented in the pollen spectra, more than in anyother honey studied so far, but of these, beech andgrass are the only regularly occurring types.
(Nothofagus fusca-type),
DISCUSSION
The results demonstrate that pollen analysis can beapplied successfully to the study of New Zealandhoney. The acetolysis procedure adopted is asuseful a technique in melissopalynology (see Lieux
59
1980) as it is in other pollen analyticalinvestigations. Materials are simple and easy toobtain, and identification of pollen with clearedexines is relatively easy. Acetolysis proceduresresult in the destruction of algal cells whichsometimes characterise a honeydew honey, and thetechnique is limited in this respect. However, thehoneydew honey samples examined all containedrelatively high numbers of fungal spores andfragments of hyphae, most contained beech pollen,and the overall number of pollen types derivedfrom wind- pollinated plants is greater than infloral source honey. This combination ofcharacters is unique for New Zealand honeyderived from beech honeydew, and identification isassured.
Quantitative pollen analysis is used to determinethe characteristics of a unifloral honey. This isachieved by spiking with spores asalready described, but some workers prefer to usetheir own prepared suspensions of pollen grains.Once the characteristics of a unifloral honey aresatisfactorily established, spiking is not strictlynecessary, but since the procedure is so simple, itsroutine use provides the opportunity for quantitativecounts if the need arises. Assessment of a honey doesnot always confirm judgements based onorganoleptic criteria which lack the quantitativecontrol of pollen analysis. The advantages of a pollenanalysis is that all, or most, nectar sources involvedare recognised, and if quantitative data are available,their relationship to each component, or to thedominant source, can be determined. Pollen analysistherefore offers a useful objective method ofclassifying honey, and certainly complementstraditional methods of doing so.
Lycopodium
In this work a judgement is more soundly based iffloral structure, nectar secretion, pollen production,and pollination of the plants involved have beendetermined. However, this information is not alwaysavailable. White clover flowers are well adapted tocross pollination by bees. When the bee alights onthe lateral petals (the alae) they are depressed by thebees weight, which forces the stigma and the stamensto protrude from their enclosing petals. The stigmaand the stamens then come into contact with the beesbody and pollen transfer is effected as the beereaches down the short calyx for nectar.
Willow pollen occurs in many honey samples, but itis never recorded as more than a minor component.The simplest explanation may be that since willowsflower early in the season a honey surplus is never, orrarely, produced. However, recent experiments withwillows suggest that honey bees exhibita degree of faithfulness to either the male or femaleplants of dioecious species
(Salix caprea)







66 New Zealand Journal of Agricultural Research, 1985, Vol. 28
Table 13 (and opposite) continued.
Locality
Pollen source
LupinusVicia
CarpodetusCordylineCotula-typeEscholtziaMetrosiderosRubusQuintiniaWeinmannia
Astelia
DracophyllumElaeocarpusGaultheriaLeucopogonfraseri Lonicera
PennantiaPittosporum
Caryophyllaceae
Malvaceae
Gramineae (W)(W)(W)
Chenopodiaceae (W)(W)
(W)(W)
(W)Cyperaceae (W)
(W)(W)
PlantagoCoprosma
CoriariaPinusRumexCupressus
MyrsineNothofagus fusca-type
Unidentified
Sum (pollen + honeydew)APFHoneydew elements
(van der Werf 1983), and the 2 commonest willowspecies in New Zealand, the crack willow
and the weeping willow (S.are dioecious. The situation is probablycomplicated by the fact that in New Zealand, crackwillows are derived from a male clone andweeping willows are derived from a female clone.This apparent preference of bees for either male orfemale plants of dioecious willow species may berelevant for other species as well. Thyme pollen isclearly under-represented in thyme honey, andfloral structure is a major factor in determiningthis. Thyme is a gynodioecious plant (i.e.,hermaphrodite flowers are produced by some
(Salixfragilis) babylonica)
plants, female flowers by other plants) and there isevidence that environmental factors play animportant and complex role in determining sexualexpression in thyme (Dommee et al. 1978). Bothtypes of flower produce abundant nectar and areequally available to bees, although pollen isavailable in only a proportion of flowers. On theother hand, manuka bears male and hermaphroditeflowers on the same plant (andromonoecious), andnectar is produced in small quantities by both typesof flower (Primack & Lloyd 1980). The smallpollen grains are therefore available whenever beeswork the flowers, and on present evidence they areclearly over-represented.
Moar – Pollen analysis 67
Locality
Pollen source
LupinusVicia
CarpodetusCordylineCotula-typeEscholtziaMetrosiderosRubusQuintiniaWeinmannia
Astelia
DracophyllumElaeocarpusGaultheriaLeucopogonfraseri Lonicera
PennantiaPittosporum
Caryophyllaceae
Malvaceae
Gramineae (W)(W)(W)
Chenopodiaceae (W)(W)
(W)(W)
Cyperaceae (W)(W)
(W)
PlantagoCoprosma
CoriariaPinusRumexCupressus
MyrsineNothofagus fusca-type
UnidentifiedSum (pollen + honeydew)APFHoneydew elements
Demianowicz (1964) found that highest absolutepollen counts were often recorded for plants withthe smallest pollen grains.Rewarewa flowers are pollinated by birds(Cheeseman 1890) although an abundant nectarflow attracts bees which produce from it a honeyof characteristic flavour (Walsh 1967). The floralstructure is such that bees are able to take nectarwithout disturbing the anthers (Moore & Irwin1978) so that pollen is unlikely to be included inthe honey in great numbers. The results of pollenanalysis confirm this, but the pollen counts (Table11) are variable enough to suggest that rewarewahoney requires further investigation.
The survey has identified characteristic pollenspectra of honey from a particular period. Withcurrent changes in agricultural and horticulturalpractice (e.g., the rapid destruction of gorse hedgesin Canterbury, and the increased interest in variousnew horticultural and tree crop plants) it is clear thatnectar and pollen sources may also change. The needto monitor the characteristics of any particular honeyis obvious, especially when new and differentcombinations of nectar may alter its character. It isequally important to examine the honey flora on aregional basis, to identify regional differences inpollen spectra. The unexpectedly high

68 New Zealand Journal of Agricultural Research, 1985, Vol. 28
values for pollen in thyme honey fromCentral Otago, and the occasions when a pollentype eludes identification further emphasise thisneed. Unless the flora of a district is welldocumented it is sometimes difficult to identify theunknown. Less than 30 years ago, honeydewhoney was regarded as an unacceptable product(Seal 1957) and attitudes in this respect have onlyrecently changed (Cook 1978, 1981). Honeydewhoney is now a prized export product (Crozier1981) which should be "produced wholly ormainly from secretions of, or found on, living partsof plants other than blossoms" (Honey exportcertification regulations 1980). The analysessuggest that most samples fit the above definitionalthough some contain relatively high values ofgorse and other pollen. Whether these qualify aspure honeydew honey is uncertain, but there is nodoubt that every sample contains honeydew. Theregular occurrence of hyphae and spores of sootymould, and pollen grains of beech, clover, willow,and gorse, provide spectra which identify thisproduct. The fluctuating values for honeydewelements may result from the preparation techniqueused or they may reflect differences in infestationof the beech trees by the sooty mould. This clearlyneeds further investigation, but the relatively highvalues for gorse, willow, and white clover-typepollen in some samples suggests that care isnecessary in selecting sites for hives which may belocated at the forest edge close to both honeydewand floral sources of honey.
Aciphylla
CONCLUSIONS
This pollen analytical survey has provided insightsinto our knowledge of New Zealand honeys, andextends the early observations made by HarrisFilmer (1948). On the basis of the data obtainedfrom the present study the following points can bemade.(1) The number of pollen types recorded duringthe survey is well over 100 but the number in anyparticular sample rarely exceeds 22. In this respectNew Zealand honey is not especially rich in pollentypes and compares with many honeyscharacteristic of north-west Europe.(2) Wind-blown pollen grains are regularly foundin New Zealand honey, as elsewhere, although theirnumbers in any one sample are insignificant. Thereare generally more pollen-types derived from wind-pollinated plants in honeydew honey than in floralhoney, which probably reflects the manner in whichthe honeydew is collected by the bees moving overthe sticky mass of sooty mould on the beech trunks.
&
(3)
(4)
(5)
(6)
(7)
The presence of these pollen types isconsidered accidental, especially as numbersare always low. However, some wind-pollinated plants, e.g., grass, plantain(Maurizio & Grafl 1982), and areworked by bees for pollen.White clover is the most important nectarsource available to bees in New Zealand, andits pollen occurs in honeys as a dominant,secondary, or minor type. This frequencyreflects the role white clover plays in NewZealand's pastoral economy, and consequentlyits value in honey production (Godley 1979).The district from which a honey originates canbe determined within limits by pollen analysis.Such deductions depend upon careful pollenidentifications together with a soundknowledge of the flora and vegetation in NewZealand. In some instances it is possible torecognise a relatively small area as the source(e.g., goats rue in the Manawatu) whereas inothers a much larger area may be involved, upto and including the North or the SouthIslands.It is clear that an analytical characterisation ofpollen content may be at variance withtraditional methods of assessing the origin of ahoney. This is especially true for mixed sourcehoneys where personal judgement is oftencritical. Pollen analysis thus provides anobjective means of assessing a honey, andfurther, can be used to monitor standards andpractices on a consistent basis. Routineexamination can be conducted rapidly bymeans of orienting or complete analysesprovided that details based on absolute pollencounts and percentage frequencies for thevarious honey types are available.Determination of absolute pollen content isnecessary when establishing the floral sourceof a honey. Spiking with tabletsprovides a relatively simple method producingcomaparable results to those based on methodsused in Europe. Thus clover and mixed sourcehoney, as in Europe and elsewhere, is a normalhoney whereas thyme and manuka areincluded in the under-represented and over-represented categories respectively. However,more samples should be examined beforeminimum frequencies are finally accepted ascharacteristic of a unifloral honey. This isespecially true for those under-representedhoneys (citrus and rewarewa) and over-represented honeys (kamahi and manuka) ofwhich only a few samples were available forstudy.
Coprosma,
Lycopodium
The samples must be processed in a uniformmanner, but the samples themselves need to becollected in the field in as uniform a way

Moar – Pollen analysis
as possible. Comparison of results fromsamples extracted by different methods (e.g.,Centrifugation and pressing) is impossible, forabsolute pollen content may vary greatly.Similarly when attempting to establish detailsof unifloral honeys commercial packs, whichmay have been blended, should not be used,and have been discarded for this purposeduring the present survey.
ACKNOWLEDGMENTS
This work on New Zealand honeys could not have been carriedout without the co-operation of apiarists and Apiary AdvisoryOfficers of the Ministry of Agriculture and Fisheries. I amgrateful to them all for providing and for- warding samplesfrom many districts within New Zealand. The samples wereprocessed in the Botany Division laboratories by differentindividuals at various times, and I thank them for their carefultreatment of many different specimens during the 3-yearperiod.
REFERENCES
Cheeseman, T. F. 1890: Notes on the fertilisation ofReport of second meeting of the Australasian Association for
Knightia.
Connor, H. E. 1977: The poisonous plants in New Zealand.Wellington, Government Printer.
Cook, V. A. 1978: The history of the utilization of NewZealand honeydew. Advisory Services DivisionHoneydew seminar, Ministry of Agriculture and Fisheries,Christchurch, 1 – 6.
In: ed.,
1981: New Zealand honeydew from beech.62: 20 – 22.
Bee world
Crane, Eva (ed.,) 1975: Honey – a comprehensive survey.London, Heinemann.
Crozier, Lisa R. 1981: Beech honeydew: forest produce.26: 200 – 209.
NewZealand journal of forestry
Demianowicz, Z. 1964: Charakteristik der Einartighonige.273-288.Annales abeille 7:
Dommee, B.; Assouad, M. W.; Valdeyron, G. 1978: Naturalselection and gynodioecy in
l 7-28.Thymus vulgaris. Botanical
journal of the Linnean Society 77:
Erdtman, G. 1943: An introduction to pollen analysis. NewYork, Chronica botanica.
Godley, E. i. 1979: Flower biology in New Zealand. NewZealand journal of botany 17: 461 – 467.
Harris, W. F.; Filmer, D, W, 1947: A recent outbreak of honeypoisoning. Part VI. Botanical investigation of pollen and nectarHow. 29A:134 – 143.
New Zealand journal of' science and technology
1948: Pollen in honey and bee loads. New Zealand
journal of science and technology 30A: 178-187.
69
Hughes, S. J. 1972: New Zealand fungi 17. Pleomorphism inEuantennariaceae and Metacapnodiaceae, two new families ofsooty moulds. 225 – 242.New Zealand jour- nal of botany 10:
Lieux, Meredith H. 1972: A melissopalynological study of 54Louisiana (U.S.A.) honeys.
95 – 124.Review of palaeobotany and
palynology 13:
1980: Acetolysis applied to microscopical honeyanalysis. 61.Grana 19: 57 –
Louveaux, J.; Maurizio, Anna; Vorwohl, G. 1978: Methods ofmelissopalynology. 59: 139 – 157.Bee world
Maurizio, Anna 1946: Introduction au chapitre "Analyse dupollen" de la statistique des miels.
592-606.Beihefte zur
Schweizerischen Bienen-Zeitung I:
1949: Pollenanalytische untersuchungen an honig undpollenhoschen.
320-412.Beihefte zur Schwei=er- ischen Bienen-
Zeitung 2:
1951: Pollen analysis of honey. I – 5.Bee world 32:
197l: Le spectres pollinique des miels Lux-embourgeois. 221 – 238.Apidofogie 2:
1975: Microscopy of honey. Crane, Eva Acomprehensive survey of honey. London, Heinemann.pp. 240 – 257.
In: ed.,
1979: Beitrag zur kenntnis der pollenspektrumsnorwegischer honige. Apidologie 10: 359 – 394.
Maurizio, Anna; Grafl, Ina 1982: Das Trachtpflanzenbuch.Nektar und pollen-die wichtigsten nahrungsquellen derhonigbiene. (3rd ed.) Munchen, Ehrenwirth Verlag.
Maurizio, Anna; Louveaux, J. 1961: Pollens des plantesmelliferes d'Europe. II. 219 – 346. 1964:Pollens des plantes melliferes d'Europe. V. 6: 5– 45.
Pollen et spores 3:Pollen et spores
Moore, L. B.; Irwin, J. B. 1978: The Oxford book of NewZealand plants. Wellington, Oxford University Press.
Primack, Richard; Lloyd, David G. 1980: Andromonoecy inthe New Zealand montane shrub manuka,
(Myrtaceae). 67: 361 –368.
Leptospermumscoparium American journal of botany
Seal, D. W, A. 1957: Beekeeping on West Coast of SouthIsland. 167 – 168.New Zealand journal of agriculture 94:
Stockmarr, J. 1971: Tablets with spores used in absolute pollenanalysis. 615 – 621.Pollen et spores 13:
Sutherland. Maurice D.; Palmer-Jones, T. 1947: A recentoutbreak of honey poisoning. Part II. The toxic substance of thepoisonous honey.
29A: 114 – 120.New Zealand journal of science and
technology
Swallow, W. 1978: Chemical analysis of beech honeydew.Advisory Services Division Honeydew seminar. Ministryof Agriculture and Fisheries, Christchurch. pp. 65 – 70.
In:ed.,
Vorwohl, G. 1972: Das Pollenspektrum honigen aus denitalienischen alpen. Apidologie 3: 309 – 340.

70 New Zealand Journal of Agricultural Research, 1985, Vol. 28
1973: Die reprasentierung des citrus – pollens initalienischen orangehonigen. 274 – 281.Apidologie 4:
Walsh, R. S. 1967: Nectar and pollen sources. Upper Hutt,National Beekeepers Association of New Zealand.
Waters, R. 1915a: Pollen grains as source-indicators of honey.213 – 215.The journal of agriculture 11:
1915b: Pollen grains as source-indicators of honey.384 – 388.
Thejournal of agriculture 11:
1916: Pollen grains as source-indicators of honey.279-281.
Thejournal of agriculture 12:
Wilkinson, E. L.; Dann, G. M.; Smith, G. J. S. 1979; Thyme inCentral Otago. Tussock Grasslands and Mountain LandsInstitute, Lincoln College. pp. 1-30.Special publication no. 14.
APPENDIX 1
In any analytical survey of pollen in honey it isnecessary to use samples obtained in a standardmanner (e.g., standard extraction techniques) sothat results may be compared. This is especiallyimportant when determining standards forparticular honey types. Pollen analyses are basedupon the extraction of pollen grains from a 10 gsample of honey. A duplicated sample is stored as aprecaution against accident during processing or foran unforseen later requirement.Samples are processed as follows:
(l) Ten g of honey are dissolved in distilledwater in a water bath at a temperature notexceeding 45 C.(2) The sediment is concentrated by repeatedcentrifuging in a bench centrifuge at 3,000revolutions per min. If centrifuge tubes are of alimited capacity (15 ml) the sample is kept atan even temperature, and the centrifugationprocess is continued until all the sediment isincluded in one tube.(3)Two Stockmarr's tabletstablets) are added to the precipitate anddissolved in about 10 ml 10% HC1. HC1 isadded slowly and stirred until the tablets arecompletely dispersed through the solution. Thesample is then centrifuged for about 3 min at3,000 revolutions per min, and the supernatantis decanted.
(Lycopodium
(4) Acetolysis follows in the usual manner.(a) About 10 ml glacial acetic acid is added to theprecipitate, mixed thoroughly, centrifuged anddecanted.(b) About 10 ml acetolysis mixture isadded,consisting of 9 parts acetic anhydride to 1part concentrated sulphuric acid. This mixturegenerates heat and is explosive if it comes intocontact with water, so it is essential that thepreceding step using glacial acetic acid is carefullyapplied. A face mask is a useful precaution againstpossible mishap.The tubes with the acetolysis mixture are placed ina water bath at 100 C for 3 – 4 min and stirredvigorously taking care not to splash water intothem. They are then centrifuged and decanted intoa clean beaker or into a sink with fast runningwater (some reaction may occur).(c) About 12 ml glacial acetic acid is added,stirred thoroughly, centrifuged, and decanted. Theprecipitate is washed in about 12 ml distilledwater, centrifuged, and decanted.(D) About 12 ml 7% KOH is added, stirredthoroughly, centrifuged, and decanted. Then alightly coloured solution of Basic Fuchsin isadded, which stains pollen grains red.(e) One drop of well mixed precipitate is placedonto a microscope slide, mixed thoroughly withmelted glycerine jelly, and covered with a 22 X 22mm cover glass. Only enough jelly to reach theedges of the cover glass should be used – this is amatter for practice and experience – but 1-2 ml ofjelly may be enough. When the jelly is set theslide is ready for counting.
Although counting procedures are simple, amechanical stage fitted with a vernier scale is essentialto traverse the width of the cover glass. Slides arealways placed in the same relative position on themechanical stage which is moved vertically after eachtraverse by one division of the vernier to avoid overlapof the field of view. For easiest counting, a 40Xobjective is used in combination with a X10 or X12ocular, and the number of grains counted depends uponthe purpose of the analysis. An orienteering analysisinvolves no more than identification of the mostnumerous or characteristic pollen grains. To determinethe distribution among frequency classes 300 pollengrains should be counted (up to 500 for greaterprecision). spores need only be counted ifabsolute pollen content is to be calculated.
Lycopodium


















