
Crosstalk between neovessels and mural cells directs the...
8
1607 Research Article Introduction Angiogenesis, the growth of new vasculature from pre-existing capillaries, is an important component of normal growth, tissue repair and neoplasia (Carmeliet, 2005; Davis and Senger, 2005). In both normal and tumor stroma, type I collagen is the most abundant component of the extracellular matrix and serves not only as a key scaffolding material but also as a physical barrier for new vessel formation (Chun et al., 2004; Seandel et al., 2001; Davis and Senger, 2005; Saunders et al., 2006). As multiple matrix metalloproteinases (MMPs), including MMP-1, MMP-2, MMP-8, MMP-13 and the membrane type 1 MMP MT1-MMP (also known as MMP-14) display type I collagenolytic activities in vitro (Brinckerhoff et al., 1987; Ohuchi et al., 1997; Hotary et al., 2000; Atkinson et al., 2001), several of these enzymes have been proposed to play important roles in regulating angiogenesis in vivo (Zhou et al., 2000; Chun et al., 2004; Ling et al., 2004; Zijlstra et al., 2004). Recently, however, increasing evidence has begun to accumulate that MT1-MMP serves as the dominant pericellular collagenase during angiogenesis (Zhou et al., 2000; Koike et al., 2002; Chun et al., 2004; Saunders et al., 2006). Whereas neovessel formation is severely impaired in tissues rich in type I collagen in Mmp14-null mice (Zhou et al., 2000), little is known with regard to the factors that regulate vascular MT1- MMP expression in the in vivo setting. To this end, we have established a transgenic mouse line wherein the lacZ gene (encoding -galactosidase) was inserted into the endogenous Mmp14 gene to monitor its transcriptional regulation (MT1- MMP +/lacZ mice). In combination with an ex vivo angiogenesis model using murine tissues cultured under 3-dimensional (3D) conditions, we now demonstrate that MT1-MMP expression is largely confined to the sprouting tip of the developing vasculature, a site where cell proliferation and focal degradation of collagen proceed preferentially. Further, we find that MT1-MMP protein expression is downregulated throughout the vessel stalk by surrounding mural cell The membrane-anchored matrix metalloproteinase MT1- MMP (also known as Mmp14) plays a key role in the angiogenic process, but the mechanisms underlying its spatiotemporal regulation in the in vivo setting have not been defined. Using whole-mount immunohistochemical analysis and the lacZ gene inserted into the Mmp14 gene, we demonstrate that MT1-MMP vascular expression in vivo is confined largely to the sprouting tip of neocapillary structures where endothelial cell proliferation and collagen degradation are coordinately localized. During angiogenesis in vitro, wherein endothelial cells are stimulated to undergo neovessel formation in the presence or absence of accessory mural cells, site-specific MT1- MMP expression is shown to be controlled by crosstalk between endothelial cells and vascular smooth muscle cells (VSMC). When vessel maturation induced by VSMCs is inhibited by introducing a soluble form of the receptor tyrosine kinase Tek, MT1-MMP distribution is no longer restricted to the endothelial tip cells, but instead distributes throughout the neovessel network in vitro as well as ex vivo. Taken together, these data demonstrate that vascular maturation coordinated by endothelial cell/mural cell interactions redirects MT1-MMP expression to the neovessel tip where the protease regulates matrix remodeling at the leading edge of the developing vasculature. Supplementary material available online at http://jcs.biologists.org/cgi/content/full/120/9/1607/DC1 Key words: MT1-MMP, Angiogenesis, Endothelial cells, Mural cells, Type I collagen Summary Crosstalk between neovessels and mural cells directs the site-specific expression of MT1-MMP to endothelial tip cells Ikuo Yana 1 , Hiroshi Sagara 1 , Satoshi Takaki 2 , Kiyoshi Takatsu 2 , Kenji Nakamura 3 , Kazuki Nakao 4 , Motoya Katsuki 5 ,Shun-ichiro Taniguchi 6 ,Takanori Aoki 7 , Hiroshi Sato 8 ,Stephen J. Weiss 9 and Motoharu Seiki 1, * 1 Division of Cancer Cell Research and 2 Division of Immunology, Institute of Medical Science, University of Tokyo, 4-6-1 Shirokane-dai, Minato-ku, Tokyo 108-8639, Japan 3 Mouse Genome Technology Center, Mitsubishi Kagaku Institute of Life Sciences, Tokyo 194-8511, Japan 4 Laboratory for Animal Resources and Genetic Engineering, Center for Developmental Biology, RIKEN, Kobe 650-0047, Japan 5 National Institute for Basic Biology, Okazaki 444-8585, Japan 6 Institute on Aging and Adaptation, Shinshu University Graduate School of Medicine, Japan 7 Daiichi Fine Chemical Corporation, 530 Chokeiji, Takaoka, Toyama 933-8511, Japan 8 Department of Molecular Virology and Oncology, Cancer Research Institute, Kanazawa University, Japan 9 Division of Molecular Medicine and Genetics, Department of Internal Medicine, University of Michigan Comprehensive Cancer Center, Life Sciences Institute, University of Michigan, Ann Arbor, MI, USA *Author for correspondence (e-mail: [email protected]) Accepted 18 March 2007 Journal of Cell Science 120, 1607-1614 Published by The Company of Biologists 2007 doi:10.1242/jcs.000679 Journal of Cell Science Journal of Cell Science
Transcript of Crosstalk between neovessels and mural cells directs the...

1607Research Article
IntroductionAngiogenesis, the growth of new vasculature from pre-existingcapillaries, is an important component of normal growth, tissuerepair and neoplasia (Carmeliet, 2005; Davis and Senger,2005). In both normal and tumor stroma, type I collagen is themost abundant component of the extracellular matrix andserves not only as a key scaffolding material but also as aphysical barrier for new vessel formation (Chun et al., 2004;Seandel et al., 2001; Davis and Senger, 2005; Saunders et al.,2006). As multiple matrix metalloproteinases (MMPs),including MMP-1, MMP-2, MMP-8, MMP-13 and themembrane type 1 MMP MT1-MMP (also known as MMP-14)display type I collagenolytic activities in vitro (Brinckerhoff etal., 1987; Ohuchi et al., 1997; Hotary et al., 2000; Atkinson etal., 2001), several of these enzymes have been proposed to playimportant roles in regulating angiogenesis in vivo (Zhou et al.,2000; Chun et al., 2004; Ling et al., 2004; Zijlstra et al., 2004).Recently, however, increasing evidence has begun to
accumulate that MT1-MMP serves as the dominant pericellularcollagenase during angiogenesis (Zhou et al., 2000; Koike etal., 2002; Chun et al., 2004; Saunders et al., 2006). Whereasneovessel formation is severely impaired in tissues rich in typeI collagen in Mmp14-null mice (Zhou et al., 2000), little isknown with regard to the factors that regulate vascular MT1-MMP expression in the in vivo setting. To this end, we haveestablished a transgenic mouse line wherein the lacZ gene(encoding �-galactosidase) was inserted into the endogenousMmp14 gene to monitor its transcriptional regulation (MT1-MMP+/lacZ mice). In combination with an ex vivo angiogenesismodel using murine tissues cultured under 3-dimensional (3D)conditions, we now demonstrate that MT1-MMP expression islargely confined to the sprouting tip of the developingvasculature, a site where cell proliferation and focaldegradation of collagen proceed preferentially. Further, we findthat MT1-MMP protein expression is downregulatedthroughout the vessel stalk by surrounding mural cell
The membrane-anchored matrix metalloproteinase MT1-MMP (also known as Mmp14) plays a key role in theangiogenic process, but the mechanisms underlying itsspatiotemporal regulation in the in vivo setting have notbeen defined. Using whole-mount immunohistochemicalanalysis and the lacZ gene inserted into the Mmp14 gene,we demonstrate that MT1-MMP vascular expression invivo is confined largely to the sprouting tip of neocapillarystructures where endothelial cell proliferation andcollagen degradation are coordinately localized. Duringangiogenesis in vitro, wherein endothelial cells arestimulated to undergo neovessel formation in the presenceor absence of accessory mural cells, site-specific MT1-MMP expression is shown to be controlled by crosstalkbetween endothelial cells and vascular smooth muscle cells(VSMC). When vessel maturation induced by VSMCs is
inhibited by introducing a soluble form of the receptortyrosine kinase Tek, MT1-MMP distribution is no longerrestricted to the endothelial tip cells, but instead distributesthroughout the neovessel network in vitro as well as ex vivo.Taken together, these data demonstrate that vascularmaturation coordinated by endothelial cell/mural cellinteractions redirects MT1-MMP expression to theneovessel tip where the protease regulates matrixremodeling at the leading edge of the developingvasculature.
Supplementary material available online athttp://jcs.biologists.org/cgi/content/full/120/9/1607/DC1
Key words: MT1-MMP, Angiogenesis, Endothelial cells, Mural cells,Type I collagen
Summary
Crosstalk between neovessels and mural cells directsthe site-specific expression of MT1-MMP toendothelial tip cellsIkuo Yana1, Hiroshi Sagara1, Satoshi Takaki2, Kiyoshi Takatsu2, Kenji Nakamura3, Kazuki Nakao4,Motoya Katsuki5,Shun-ichiro Taniguchi6,Takanori Aoki7,Hiroshi Sato8,Stephen J. Weiss9 and Motoharu Seiki1,*1Division of Cancer Cell Research and 2Division of Immunology, Institute of Medical Science, University of Tokyo, 4-6-1 Shirokane-dai, Minato-ku,Tokyo 108-8639, Japan3Mouse Genome Technology Center, Mitsubishi Kagaku Institute of Life Sciences, Tokyo 194-8511, Japan4Laboratory for Animal Resources and Genetic Engineering, Center for Developmental Biology, RIKEN, Kobe 650-0047, Japan5National Institute for Basic Biology, Okazaki 444-8585, Japan6Institute on Aging and Adaptation, Shinshu University Graduate School of Medicine, Japan7Daiichi Fine Chemical Corporation, 530 Chokeiji, Takaoka, Toyama 933-8511, Japan8Department of Molecular Virology and Oncology, Cancer Research Institute, Kanazawa University, Japan9Division of Molecular Medicine and Genetics, Department of Internal Medicine, University of Michigan Comprehensive Cancer Center, LifeSciences Institute, University of Michigan, Ann Arbor, MI, USA*Author for correspondence (e-mail: [email protected])
Accepted 18 March 2007Journal of Cell Science 120, 1607-1614 Published by The Company of Biologists 2007doi:10.1242/jcs.000679
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1608
populations that act through a network depending on thereceptor tyrosine kinase Tek, which contains an Ig and EGFhomology domain 2. Our results outline a dynamic cellularstrategy that is used by the developing vasculature to confineMT1-MMP activity to endothelial tip cells during neovesselformation.
ResultsMonitoring transcription of Mmp14 via �-galactosidaseactivityTo monitor the expression of the Mmp14 transcript, weemployed a recently developed mouse strain harboring a lacZgene under the control of the endogenous Mmp14 promoter.Homologous recombination of the murine Mmp14 gene locusencoding the pro- and catalytic domains (exons 1-5) with thetargeting vector carrying the lacZ gene disrupts expression ofMT1-MMP and, at the same time, affords the opportunity tomonitor endogenous promoter activity via nuclear lacZ activity(Fig. 1A).
�-Gal staining of the whole MT1-MMP+/lacZ mouse embryoreveals a pattern of MT1-MMP expression that parallels thatobserved previously by in situ hybridization (Apte et al., 1997).Using this system, the transcription of Mmp14 in stable, non-growing vascular beds was monitored in MT1-MMP+/lacZ mice.As shown in Fig. 1B, lacZ activity was largely negative in veinand capillary endothelium of peritoneal tissues in 14-day-oldheterozygous mice. Whereas stromal cell populations as well
as the fibroblast-like adventitial cells surrounding large arteriesstained strongly positive (see supplementary material Fig. S1),endothelial cells themselves displayed minimal �-gal stainingin both small and large vessels (Fig. 1C).
Next, to determine whether MT1-MMP expression is alteredduring angiogenesis, MT1-MMP+/lacZ mice were injected witha type I collagen gel containing vascular endothelial growthfactor (VEGF) trapped within a slow release polymer. Sevendays post-injection, the collagen gel was resected and analysedfor lacZ activity followed by immunostaining for CD31 (Fig.2A,B). Within the vasculature itself, nuclear �-gal staining waslocalized specifically to the tip of the elongated CD31-positivetubule (Fig. 2B). Notably, the tip-specific lacZ activity was onlyobserved during the first 7 days of culture in vivo, disappearingat approximately at day 14 when vascular network formationwas completed (data not shown). In an ex vivo angiogenesismodel using mouse tissue explants embedded in 3D gels of typeI collagen, the growing neovasculature likewise displayed focal�-gal expression in CD31-positive cells that was confined to thenucleus of sprouting tip cells (Fig. 2C). By contrast, only lowlevels of �-gal staining were observed in the neovessel stalk (i.e.the region of the endothelial cell tubule lying behind theadvancing tip cell), which also stained positively for thebasement membrane macromolecule type IV collagen (Fig.2C). Quantitative analyses of �-gal-positive endothelial cells inthe explant model demonstrated that more than 80% of thenuclear staining detected was confined to the tip cell population
(Fig. 2D). As described previously, explants recoveredfrom Mmp14-null mice were unable to mount anangiogenic response ex vivo (Chun et al., 2004) (and datanot shown). Taken together, these data suggest thatMmp14 is transcribed preferentially within the tip cells ofextending neovessels while only low level expression isobserved in either mature vessels or the stalk region ofgrowing neovessels.
Collagenolysis and proliferation of endothelial tipcellsGiven the preferential expression of MT1-MMP inendothelial tip cells, these results raised the possibility
Journal of Cell Science 120 (9)
Fig. 1. Establishment of MT1-MMP+/lacZ mouse strain tomonitor Mmp14 transcription. (A) Schematic representation oftargeting Mmp14. Exons 1-5 encoding the catalytic domain ofMT1-MMP were targeted and the lacZ gene, encoding �-galactosidase fused with a nuclear localization signal (NLS)was cloned in-frame with a phosphoglycerate kinase (PGK)-gpt/neo resistance gene cassette. (B) Vascular bed Mmp14expression. Peritoneal tissue whole-cell mounts with associatedsegments of the musculus rectus abdominis were isolated from3-day-old mice [MT1+/+ (left panel) and MT1+/lacZ (rightpanel)] and stained for �-gal in combination with CD31.Whereas �-gal staining was not observed in MT1-MMP+/+
tissues, lacZ-positive cells were found throughout the stromaand also in association with perivascular cells in tissues ofMT1+/lacZ mouse (see below). Bars, 0.25 mm for large panels,0.1 mm for insets. (C) Transverse sections of peritoneum andassociated musculus rectus abdominus muscle from theMT1+/lacZ mice were stained both for �-gal (red) and CD31(green). Within the vascular bed, �-gal staining was confinedto perivascular cells, with limited or no staining associated withthe CD31-positive endothelial cells. Bar, 0.25 mm.
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1609Regulation of MT1-MMP during angiogenesis
that MT1-MMP-dependent collagenolytic activity (Chun et al.,2004; Saunders et al., 2006) is similarly restricted to theleading edge of the developing neovessel. As such, murinetissue explants were cultured in native type I collagen gelsoverlaid with a second layer of type I collagen conjugated witha quenched fluorescence probe (DQ collagen; Fig. 3A andsupplementary material Fig. S2). Significantly, fluorescentsignal developed solely around the MT1-MMP-expressing tipcells in a fashion consistent with focal collagenolysis (Fig. 3A).As expected, focal collagenolytic activity could not be detectedin the presence of BB-94 which blocked in-growth ofendothelial cell sprouts into the layer of DQ collagen
(supplementary material Fig. S2). Further, as MT1-MMPcollagenolytic activity can also control cell proliferation in 3Dmatrix environments (e.g. Hotary et al., 2003),bromodeoxyuridine (BrdU) incorporation was confined to thetype IV collagen poor endothelial cells at the neovessel front(Fig. 3B; white arrow). Quantitative analyses demonstrate thatBrdU incorporation is almost completely restricted to theendothelial tip cells (Fig. 3C). Thus, collagenolysis and cellproliferation appeared to coincide with the site-specificexpression of MT1-MMP at the leading edge of the advancingneovessel.
MT1-MMP and Tek expression are invertly relatedduring angiogenesisInteractions between endothelial cells and perivascular muralcells (i.e. pericytes or smooth muscle cells) promote blood
Fig. 2. MT-MMP expression during neovessel formation in vivo andex vivo. (A) Neocapillary formation was induced in vivo in type Icollagen gel implants containing a VEGF slow-release polymer.Neovessels are indicated by arrows. Bar, 1 mm. (B) Whole-cell mountof neovessels infiltrating the collagen gel implant stained for �-galand CD31. In neovessels, nuclear �-gal staining was predominatelyassociated with the advancing tip of the blood vessels (arrows, leftpanel), which are pointed at by arrows in the accompanying tracing(right panel). Bar, 0.25 mm. (C) Muscle explants derived from MT1-MMP+/lacZ mice were embedded in 3D collagen gels and neovesselformation was induced with a growth factor cocktail. In the left panel,the advancing tip of CD31-positive neovessels (arrowheads) exhibitstrong �-gal staining (arrows). Following staining for type IVcollagen (right panel), the advancing endothelial cell tips (arrows)were largely poor of type IV collagen, whereas the neovessel stalk isstrongly positive of type IV collagen (arrowheads). Bars, 0.25 mm.(D) Neovessel number per explant was quantified at day 7 by phase-contrast microscopy and the numbers of tubes with either tip-specific(i.e. within two cells of the neovessel tip) or pan-vessel (i.e.neovessels staining with two or more endothelial stalk cells) �-galstaining was determined (*P<0.01, F-test).
Fig. 3. Characterization of neovascular tip cells. (A) In the ex vivoexplant system, neovessels invading surrounding 3D gelsimpregnated with DQ collagen (left panel, phase-contrastmicrograph), express collagenolytic activity as assessed byfluorescence microscopy (right panel). Collagen degradationsurrounding the neovascular tip (white arrows) is detected as a greensignal. Bar, 0.1 mm. (B) BrdU incorporation specific for endothelialtip cells. After a 48-hour pulse period, neovessels arising from the3D muscle explants were stained for BrdU (green) and type IVcollagen (red). Whereas the neovessel stalks stained strongly for typeIV collagen (white arrowheads, left panel), little or no BrdUincorporation was detected in these regions (yellow arrowhead, rightpanel). Instead, BrdU incorporation (white arrow) was largelyconfined to endothelial tip cells poor of the type IV collagen. Thewhite arrow shows a specific nuclear signal indicating BrdUincorporation at the neovascular tip cell. The yellow arrowheadindicates a BrdU-negative stalk cell. Bar, 0.1 mm. (C) BrdUincorporation (percent positive cells) was localized to eitherendothelial tip cells or neovessel stalks and quantified in 14 separateexperiments (P=0.00382, F-test).
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence
1610
vessel maturation and stabilization (Korff et al., 2001; Lafleuret al., 2001; Saunders et al., 2006; von Tell et al., 2006). Asneovessel outgrowths are largely enveloped by mural cells inex vivo models (Gerhardt et al., 2003; Zhu et al., 2002)(supplementary material Fig. S3), we considered the possibilitythat interactions of pericytes, vascular smooth muscle cells(VSMCs) and endothelial cells participate in regulating MT1-MMP expression. The mural-cell-derived soluble factorangiopoietin 1 (Ang-1), is a major ligand for Tek, a cell surface,endothelial-specific receptor tyrosine kinase that plays acentral role in vessel maturation (Suri et al., 1996; Carmeliet,2005; von Tell et al., 2006). To determine whether the Tek axismight impact on the spatiotemporal expression of MT1-MMP,we monitored Tek expression in endothelial cells duringangiogenesis. Using tissue explants isolated from transgenicmice that express GFP under the control of the endogenous Tekpromoter, the GFP signal was preferentially localized to theneovessel stalks (Fig. 4A,B, white arrows). By contrast, Tek
promoter expression was absent in the endothelial tip cells(Fig. 4A,B, yellow arrows), in a fashion consistent with theproposition that MT1-MMP expression is suppressed in theTek-positive endothelium.
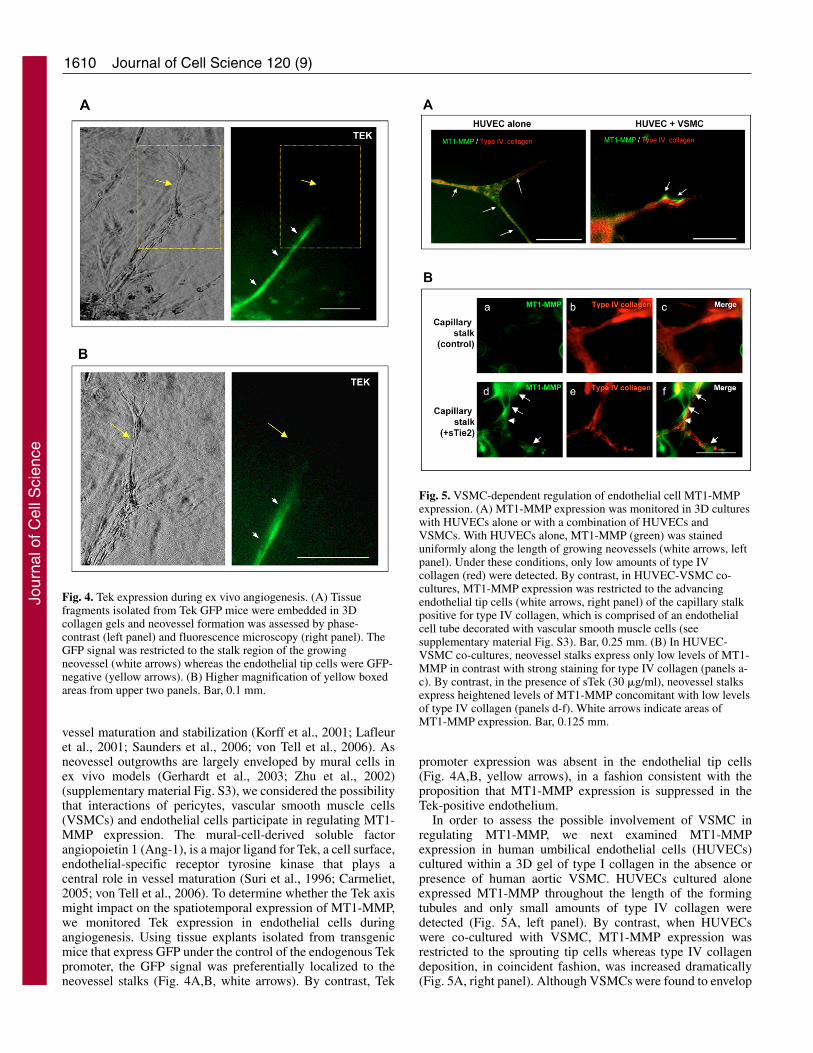
In order to assess the possible involvement of VSMC inregulating MT1-MMP, we next examined MT1-MMPexpression in human umbilical endothelial cells (HUVECs)cultured within a 3D gel of type I collagen in the absence orpresence of human aortic VSMC. HUVECs cultured aloneexpressed MT1-MMP throughout the length of the formingtubules and only small amounts of type IV collagen weredetected (Fig. 5A, left panel). By contrast, when HUVECswere co-cultured with VSMC, MT1-MMP expression wasrestricted to the sprouting tip cells whereas type IV collagendeposition, in coincident fashion, was increased dramatically(Fig. 5A, right panel). Although VSMCs were found to envelop
Journal of Cell Science 120 (9)
Fig. 4. Tek expression during ex vivo angiogenesis. (A) Tissuefragments isolated from Tek GFP mice were embedded in 3Dcollagen gels and neovessel formation was assessed by phase-contrast (left panel) and fluorescence microscopy (right panel). TheGFP signal was restricted to the stalk region of the growingneovessel (white arrows) whereas the endothelial tip cells were GFP-negative (yellow arrows). (B) Higher magnification of yellow boxedareas from upper two panels. Bar, 0.1 mm.
Fig. 5. VSMC-dependent regulation of endothelial cell MT1-MMPexpression. (A) MT1-MMP expression was monitored in 3D cultureswith HUVECs alone or with a combination of HUVECs andVSMCs. With HUVECs alone, MT1-MMP (green) was staineduniformly along the length of growing neovessels (white arrows, leftpanel). Under these conditions, only low amounts of type IVcollagen (red) were detected. By contrast, in HUVEC-VSMC co-cultures, MT1-MMP expression was restricted to the advancingendothelial tip cells (white arrows, right panel) of the capillary stalkpositive for type IV collagen, which is comprised of an endothelialcell tube decorated with vascular smooth muscle cells (seesupplementary material Fig. S3). Bar, 0.25 mm. (B) In HUVEC-VSMC co-cultures, neovessel stalks express only low levels of MT1-MMP in contrast with strong staining for type IV collagen (panels a-c). By contrast, in the presence of sTek (30 �g/ml), neovessel stalksexpress heightened levels of MT1-MMP concomitant with low levelsof type IV collagen (panels d-f). White arrows indicate areas ofMT1-MMP expression. Bar, 0.125 mm.
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1611Regulation of MT1-MMP during angiogenesis
the endothelial cell tubules only at intervals (supplementarymaterial Fig. S4), expression of MT1-MMP was suppressedsignificantly along the entire length of the neovessel stalks(Fig. 5A). Thus, interactions occurring between theendothelial cells and surrounding VSMC in the neovascularstalk provide a potential cellular basis for the selectiveexpression of MT1-MMP.
To examine directly the possible involvement of Teksignals in mural cell-mediated MT1-MMP suppression,HUVEC-VSMC co-cultures were established in thepresence of a recombinant, soluble form of Tek (sTek).Interestingly, when neovessel growth was allowed toproceed in the presence of sTek, the area of the endothelialcell tubules expressing MT1-MMP was significantlyextended along the entire length of the vessel stalk (i.e. thetotal area of MT1-MMP-positive staining increased from6±4% to 75±21%, n=5), whereas type IV collagendeposition was depressed (Fig. 5B). Likewise, in explantsof MT1+/lacZ mouse tissue cultured in the presence of sTek,�-gal was redistributed throughout the length of the tubules(Fig. 6A). Whereas the total tube number was unaffectedby sTek, the number of neovessels expressing endothelialtip cell-specific �-gal staining was decreased by ~75% (Fig.6B). Finally, sTek also relaxes MT1-MMP suppression inthe explant cultures, such that the protease is more globallyexpressed across the neovessel surface in concert with amarked diminution in type IV collagen deposition (Fig.6C). Taken together, these results support a model whereinTek signals negatively regulate the transcription of Mmp14in the stalk region and act to confine MT1-MMP expressionto the leading edge of the neovasculature.
DiscussionDespite increasing evidence supporting a criticalrequirement for MT1-MMP in angiogenic responses(Hiraoka et al., 1998; Zhou et al., 2000; Lafleur et al., 2002;Chun et al., 2004; Saunders et al., 2006), little is knownwith regard to the factors controlling the endothelialcellspecific, spatiotemporal expression of MT1-MMP invivo. Given the low levels of MT1-MMP detected in theendothelium of mature vessels, it seemed plausible topropose that vascular MT1-MMP expression are restrictedto neovessel formation. Indeed, we now demonstrate thatMT1-MMP is specifically expressed in advancingendothelial tip cells and is subsequently downregulated asthe vessel structure stabilizes and matures. As angiogenesis isimpaired in the absence of MT1-MMP (Zhou et al., 2000;Chun et al., 2004; Saunders et al., 2006), the site-specificexpression of the protease in endothelial tip cells probablyplays a crucial role in guiding new blood vessels through thestroma rich in type I collagen. Whereas seemingly intactvascular networks can be formed in Mmp14-null mice duringearly development, the content of type I collagen of multiplemouse tissues is maintained at low levels in utero and increasesmost dramatically in the postnatal state (Caulfield and Borg,1979; Mays et al., 1988; Sabeh et al., 2004).
In vivo, endothelial cells at the advancing tips of developingneovessels have been reported to display properties distinctfrom those observed in the vessel stalk (e.g. Gerhardt et al.,2003). Likewise, in our studies, MT1-MMP expression waslargely restricted to CD31-positive endothelial tip cells that
expressed collagenolytic activity, proliferated, expressed lowlevels of Tek and remained free of a mural cell coat. Cautionshould, however, be ascribed to characterizing tip cells asstrictly endothelial in terms of their structural or functionalphenotype. During vasculogenesis as well as angiogenesis,endothelial cells can undergo an endothelial-mesenchymaltransformation wherein tissue-invasive cells adopt VSMC-likecharacteristics, including the expression of �-smooth muscleactin (DeRuiter et al., 1997; Frid et al., 2002; Ishisaki et al.,2003; Liebner et al., 2004; Timmerman et al., 2004; Paruchuriet al., 2006). Hence, because endothelial cells engagetranscriptional programs necessary to support invasive activity,the leading cell population might be predicted to assume aplastic phenotype more consistent with the uniquerequirements of the tip cell population. Further, we have foundthat, whereas the vessel stalk is ensheathed by pericellular
Fig. 6. Tek-dependent regulation of MT1-MMP expression during exvivo angiogenesis. (A) MT1-MMP transcription in MT1-MMP+/lacZ
explants (as monitored by lacZ activity) was assessed in the absence orpresence of sTek. Wheras lacZ activity is confined to endothelial tipcells in the absence of sTek (white arrow, left panel), addition of sTekinduced widespread MT1-MMP expression in growing neovessels(white arrows, right panel). Mural cells and fibroblasts stain positive forsmooth muscle actin (SMA). Bar, 0.25 mm. (B) Total numberneovessels (blue bars) and of neovessels displaying endothelial tip-cell-specific MT1-MMP expression (red bars) was determined in explantcultures in the absence or presence of sTek after a 7-day culture period.Whereas the total number of neovessels was unaffected by sTek, thenumber of capillary structures exhibiting tip-specific MT1-MMPinduction was significantly reduced in the eight experiments performed(P=0.0045, F-test). (C) In murine explants, treatment with sTek (rightpanel) induced widespread expression of MT1-MMP (green, indicatedby yellow arrows) and the concomitant decrease in type IV collagenstaining (red) compared with control explants (left panel). Bar, 0.25 mm.
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1612
matrix positive for type IV collagen, the advancing front of theneovessel is almost completely devoid of a type IV collagenmatrix. Interestingly, a similar inverse pattern of MT1-MMPexpression and type IV collagen deposition could berecapitulated by culturing HUVECs under 3D conditions in theabsence or presence of VSMCs. Together, these data suggestthat crosstalk between endothelial and mural cells may beresponsible for regulating both the spatiotemporal expressionof MT1-MMP and the maintenance of a stable perivascularbasement membrane in vivo.
Mural-cell-derived growth factors such as Ang-1 have beenproposed to play central roles in regulating vascular maturationby initiating a Tek-dependent signaling cascade in associatedendothelial cells (Carmeliet, 2005; von Tell et al., 2006). In ourstudy, we found that Tek-mediated signals induce thetranscriptional suppression of MT1-MMP in the vessel stalk.These results support a model whereby endothelial tip cell-specific expression of MT1-MMP is accentuated as aconsequence of the mural-cell-dependent silencing of MT1-MMP expression within the neovessel stalk (see Fig. 7).Interestingly, recent studies have proposed that mural-cell-derived MMP inhibitors similarly act to suppress endothelialcell MT1-MMP proteolytic activity (Lafleur et al., 2001;Saunders et al., 2006). Whereas these reports demonstrate thatpericytes or smooth muscle cells can regulate MT1-MMPactivity at the post-translational level, our data support anadditional – but upstream – role for the mural-cell-dependentcontrol of MT1-MMP transcription itself. As MT1-MMP iscapable of degrading multiple basement-membranecomponents, including type IV collagen, laminin and
fibronectin (d’Ortho et al., 1997; Koshikawa et al., 2000;Hotary et al., 2006), quenching both MT1-MMP expressionand also MT1-MMP enzymatic activity in the vessel stalk maycontribute to vascular stability by preventing the unregulatedproteolysis of the perivascular matrix.
By restricting MT1-MMP activity to the leading edge ofthe neovasculature, the growing vessel would be in an optimalposition to advance itself into the surrounding matrix.Interestingly, MT1-MMP expression by endothelial tip cellsalso correlated with BrdU incorporation. This findingcontrasts with a previous study in which BrdU wasincorporated preferentially into the stalk of retinal neovessels(Gerhardt et al., 2003). Although we have confirmed theseobservations in the retina (data not shown), a crucialdifference between the retinal environment, the peritoneumand also our ex vivo culture conditions is the content of typeI collagen of the respective tissues. Indeed, we find that theretinal environment is poor of type I collagen compared withthe tissues studied here (supplementary material Fig. S5). Inaddition, type I collagen can itself induce MT1-MMPexpression (Ellerbroek et al., 2001; Lafleur et al., 2006) andit seems plausible that the surrounding collagenousenvironment acts to both modulate protease expression andcell function at the neovascular tip. Consequently, endothelialcell invasion and proliferation may take place in aconcomitant fashion as MT1-MMP-dependent changes in cellshape, cytoskeletal tension and migration impact on theproliferative response (Hotary et al., 2003; Chun et al., 2004;Chun et al., 2006). Finally, although our studies have focusedon endothelial-cell-derived MT1-MMP, VSMC and also
pericytes are additional sources of MT1-MMP activity (Shofuda et al., 1997;Filippov et al., 2005; Lehti et al., 2005).However, the expression level of MT1-MMP in the mural cells that cover theneovascular stalk appears to be low asevaluated by �-gal staining. It remains tobe determined whether the expression ofMT1-MMP in migrating VSMC is alsoregulated following recruitment to thevessel wall via endothelial-cell crosstalk.
Recent studies have suggested thatendothelial cell MT1-MMP serves as apotential target for inhibiting tumorangiogenesis (Haas et al., 1998; Zhou et al.,2000; Sounni et al., 2002; Seiki and Yana,2003; Chun et al., 2004; Plaisier et al.,2004). Our findings, however, predict anarrow window of opportunity for targetingMT1-MMP in the neovasculature as the
Journal of Cell Science 120 (9)
Tip cells
VascularMaturation
・・MT1-MMP induction
・Cell proliferation
・Basement membrane
deposition
collagenolysis
Sprouting direction
SMC BMEC
VascularMaturation
・MT1-MMP induction
・Cell proliferation
・Basement membrane
deposition
Stalk
Fig. 7. Model for site-specific MT1-MMPexpression during neovessel formation. MT1-MMP-expressing endothelial tip cells aremaintained in an immature state as they driveinvasion and proliferation. By contrast,endothelial stalk cells assume a quiescent stateas perivascular mural cells signal neighboringendothelial cells to suppress MT1-MMPexpression and increase type IV collagendeposition.
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1613Regulation of MT1-MMP during angiogenesis
protease is only transiently expressed within the endothelial tipcells of the growing vessels. Nevertheless, given the immaturestatus of mural-cell-deficient tumor vessels (e.g. Baluk et al.,2005), more global patterns of MT1-MMP expression may bemaintained within the tumor vasculature, thereby rendering itsusceptible to anti-MT1-MMP therapy. Additional studies willbe needed to address these issues directly but our findingssupport a growing body of evidence, suggesting that thespatiotemporal regulation of MT1-MMP expression andactivity in endothelial cells by surrounding mural cellpopulations serves as a key determinant of neovessel formationduring angiogenic states.
Materials and MethodsAnimals, cell lines, reagents and antibodiesC57BL/6 mice and Wistar rats were obtained from CLEA JAPAN (Tokyo, Japan).MT1-MMP+/lacZ mice were generated from heterozygous embryonic stem cells(derived from the E14 embryonic stem cell) line that were injected into C57BL/6blastocysts. The Institutional Review Board in the University of Tokyo approved allanimal studies. Human umbilical endothelial cells (HUVECs) and human vascularsmooth muscle cells (VSMCs) were purchased from KURABO (Neyagawa, Osaka,Japan), and cultured in modified MCDB131 medium supplemented with 5% fetalcalf serum and calf brain extract. An anti-MT1-MMP mouse monoclonal antibody(mAb) raised against the hemopexin domain of MT1-MMP (anti-HPX) wasdeveloped in collaboration with Daiichi-Fine Chemical Co., Ltd (Toyama, Japan).Anti-mouse and rat CD31 monoclonal antibodies (clones MEC13.3 and TLD-3A12,respectively) were purchased from BD Biosciences PharMingen (San Diego, CA)and a polyclonal antibody (clone AB756P) for type IV collagen was obtained fromChemicon (Temecula, CA). Anti-smooth-muscle actin (SMA; clone 1A4) waspurchased from Sigma (St Louis, MO). Recombinant murine VEGF and endothelialcell growth supplement were obtained from Peprotech (London, UK) andCalbiochem (La Jolla, CA), respectively. BB-94, a synthetic MMP inhibitor, was agift from Peter Brown (British Biotech, Oxford, UK), whereas TIMP-1 and TIMP-2 were provided by Daiichi Fine Chemical Co. (Takaoka, Japan).
Detection of �-galTissues were fixed in cold PBS containing 2% paraformaldehyde and 0.2%glutaraldehyde for 5 minutes, washed, and incubated overnight at 37°C in X-Galbuffer (1.3 mg/ml potassium ferricyanide, 1 mg/ml potassium ferricyanide, 0.2%Triton X-100, 1 mM MgCl2, 1 mg/ml Xgal in PBS pH 7.2). For ex vivo cultures,samples were immersed in a periodate-lysine-paraformaldehyde (PLP) buffer (2%paraformaldehyde, 75 mM lysine, 10 mM sodium periodate, 45 mM sodiumphosphate pH 7.4) for 40 minutes, followed by successive washes in PBS containing1% NP-40 and 1% SDS. To detect �-gal, samples were incubated with X-Galsolution for 3-24 hours as described above.
In vivo angiogenesis assayA double-layered matrix composed of an outer layer of type I collagen and an innersphere of a thermosensitive, slow-release polymer (Ikeda Rika, Tokyo, Japan)impregnated with 50 ng/ml of VEGF was implanted subcutaneously. At 7 or 14days, the gel was resected, stained for �-gal and immunostained for CD31. The totalnumber of tubules was determined in three independent tissue fragments for eachexperiment of five or more performed. The number of �-gal-positive tip cells or �-gal-positive tubes was determined in stained samples. Statistical significance wasdetermined by the F test.
ImmunohistochemistryFrozen sections were fixed in PLP buffer for 10 minutes, washed, and endogenousperoxidase activity was quenched by treating the samples with methanol containing0.3% H2O2. Samples were then blocked with 0.5% BSA in PBS followed byincubation with primary antibodies for 30 minutes at the appropriate concentration.Following multiple washes, samples were incubated with the secondary antibodyfor 30 minutes, washed and developed in a diaminobenzidine staining system(Nichirei, Japan). In ex vivo cultures, samples were fixed in PLP buffer for 4 hours,washed in PBS, immersed in methanol containing 0.3% H2O2, and subsequentlyblocked in PBS with 0.3% Triton X-100 containing 1% skimed milk. Samples werethen incubated with primary antibodies at the appropriate concentration overnight,followed by multiple washes before incubation with the secondary fluorescentantibody for 90 minutes. Fluorescent images were obtained using a CCD fluorescentmicroscope (IM70-Cool SNAP, OLYMPUS).
3D culture conditionsFragments of intracostal muscle or diaphragm were isolated from 1-day-old to 14-day-old C57BL/6 MT1-MMP+/+ mice, MT1-MMP+/lacZ mice or from 21-day-old
Wistar rats. Tissues were immersed in MCDB131 supplemented with a mixtureof antibiotics and antimycotics (Invitrogen-GIBCO) for 30 minutes. Samples wereminced (pieces of 3-5 mm) and embedded in 60-120 �l of neutralized type Icollagen at a final concentration of 2.4 mg/ml (type I-A Nitta Gelatine, Osaka,Japan) in 48-well or 96-well plates (Corning). Cultures were incubated in a CO2
(5%) air (95%) atmosphere at 37°C for 30 minutes until the gel was polymerized,and then cultured for 14 days in modified MCDB131 medium supplemented with20% FBS, VEGF (50 ng/ml), endothelial cell growth supplement (75 �g/ml), andheparin (100 �g/ml). For 3D HUVEC cultures, an aggregate of approximately5000 cells was suspended in 5 �l of medium and embedded in 70 �l of type Icollagen. In 3D co-cultures of HUVECs and VSMCs, a mixture of 3000 HUVECsand 2000 VSMCs was suspended in 5 �l of medium and embedded in collagengels as described above. Where indicated, VSMCs were labeled with thefluorescent dye, PKH26 (Sigma), according to the manufacturer’s instructions.Cultures were maintained for 7 days in the pro-angiogenic medium, which wasexchanged every 3 days. BrdU incorporation (48-hour pulse) was performedaccording to the manufacturer’s instructions (Pharmacia). The percentage ofBrdU-positive tip cells or BrdU-positive stalks was determined in threeindependent fragments for each of 14 experiments. Statistical significance wasdetermined by the F test.
Collagen degradation assayNeocapillary collagenolytic activity was evaluated with a quenched fluorescence-labeled substrate (DQ collagen, Molecular Probes, Eugene, Oregon). Briefly, tissuechunks were embedded in unlabeled collagen, and following a 7-day culture period,a mixture of neutralized DQ collagen and unlabeled type I collagen (3:100 ratio)was poured on top of the embedded rat tissue fragment in the absence or presenceof BB-94, TIMP-1 or TIMP-2. Following a 48-hour incubation period, images werecollected by fluorescence microscopy.
We thank Bob Whittier and Naohiko Koshikawa (IMS Universityof Tokyo) for helpful discussions, Kunika Nishibashi, Akiko Saka,Akira Matsuda, and Tomoko Nakajima-Andou for excellent technicalassistance, and Akiko Okada (Mie University, Japan) for her initialwork to establish the MT1-MMP-targeted mouse strain. This workwas supported by the Specific Coordination Fund for PromotingScience and Technology, a Grant-in-Aid for Scientific Research fromthe Ministry of Education, Culture, Sports, Science and Technologyof Japan, and NIH grant R01 CA88308.
ReferencesApte, S. S., Fukai, N., Beier, D. R. and Olsen, B. R. (1997). The matrix
metalloproteinase-14 (MMP-14) gene is structurally distinct from other MMP genesand is co-expressed with the TIMP-2 gene during mouse embryogenesis. J. Biol. Chem.272, 25511-25517.
Atkinson, S. J., Patterson, M. L., Butler, M. J. and Murphy, G. (2001). Membranetype 1 matrix metalloproteinase and gelatinase A synergistically degrade type 1collagen in a cell model. FEBS Lett. 491, 222-226.
Baluk, P., Hashizume, H. and McDonald, D. M. (2005). Cellular abnormalities of bloodvessels as targets in cancer. Curr. Opin. Genet. Dev. 15, 102-111.
Brinckerhoff, C. E., Ruby, P. L., Austin, S. D., Fini, M. E. and White, H. D. (1987).Molecular cloning of human synovial cell collagenase and selection of a single genefrom genomic DNA. J. Clin. Invest. 79, 542-546.
Carmeliet, P. (2005). Angiogenesis in life, disease and medicine. Nature 438, 932-936.Caulfield, J. B. and Borg, T. K. (1979). The collagen network of the heart. Lab. Invest.
40, 364-372.Chun, T. H., Sabeh, F., Ota, I., Murphy, H., McDonagh, K. T., Holmbeck, K.,
Birkedal-Hansen, H., Allen, E. D. and Weiss, S. J. (2004). MT1-MMP-dependentneovessel formation within the confines of the three-dimensional extracellular matrix.J. Cell Biol. 167, 757-767.
Chun, T. H., Hotary, K. B., Sabeh, F., Saltiel, A. R., Birkedal-Hansen, H., Allen, E.D. and Weiss, S. J. (2006). A pericellular collagenase directs the 3-dimensionaldevelopment of white adipose tissue. Cell 125, 577-591.
Davis, G. E. and Senger, D. R. (2005). Endothelial extracellular matrix: biosynthesis,remodeling, and functions during vascular morphogenesis and neovessel stabilization.Circ. Res. 25, 1093-1107.
DeRuiter, M. C., Poelmann, R. E., VanMunsteren, J. C., Mironov, V., Markwald, R.R. and Gittenberger-de Groot, A. C. (1997). Embryonic endothelial cellstransdifferentiate into mesenchymal cells expressing smooth muscle actins in vivo andin vitro. Circ. Res. 80, 444-451.
d’Ortho, M. P., Will, H., Atkinson, S., Butler, G., Messent, A., Gavrilovic, J., Smith,B., Timpl, R., Zardi, L. and Murphy, G. (1997). Membrane-type matrixmetalloproteinases 1 and 2 exhibit broad-spectrum proteolytic capacities comparableto many matrix metalloproteinases. Eur. J. Biochem. 250, 751-757.
Ellerbroek, S. M., Wu, Y. I., Overall, C. M. and Stack, M. S. (2001). Functionalinterplay between type I collagen and cell surface matrix metalloproteinase activity. J.Biol. Chem. 276, 24833-24842.
Filippov, S., Koenig, G. C., Chun, T. H., Hotary, K. B., Ota, I., Bugge, T. H., Roberts,
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence

1614
J. D., Fay, W. P., Birkedal-Hansen, H., Holmbeck, K. et al. (2005). MT1-matrixmetalloproteinase directs arterial wall invasion and neointima formation by vascularsmooth muscle cells. J. Exp. Med. 202, 663-671.
Frid, M. G., Kale, V. A. and Stenmark, K. R. (2002). Mature vascular endothelium cangive rise to smooth muscle cells via endothelial-mesenchymal transdifferentiation; invitro analysis. Circ. Res. 90, 1189-1196.
Gerhardt, H., Golding, M., Fruttiger, M., Ruhrberg, C., Lundkvist, A., Abramsson,A., Jeltsch, M., Mitchell, C., Alitalo, K., Shima, D. et al. (2003). VEGF guidesangiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 161, 1163-1177.
Haas, T. L., Davis, S. J. and Madri, J. A. (1998). Three-dimensional type I collagenlattices induce coordinate expression of matrix metalloproteinases MT1-MMP andMMP-2 in microvascular endothelial cells. J. Biol. Chem. 273, 3604-3610.
Hiraoka, N., Allen, E., Apel, I. J., Gyetko, M. R. and Weiss, S. J. (1998). Matrixmetalloproteinases regulate neovascularization by acting as pericellular fibrinolysins.Cell 95, 365-377.
Hotary, K., Allen, E., Punturieri, A., Yana, I. and Weiss, S. J. (2000). Regulation ofcell invasion and morphogenesis in a three-dimensional type I collagen matrix bymembrane-type matrix metalloproteinases 1, 2, and 3. J. Cell Biol. 149, 1309-1323.
Hotary, K. B., Allen, E. D., Brooks, P. C., Datta, N. S., Long, M. W. and Weiss, S. J.(2003). Membrane type I matrix metalloproteinase usurps tumor growth controlimposed by the threedimensional extracellular matrix. Cell 114, 33-45.
Hotary, K., Li , X., Allen, E., Stevens, S. L. and Weiss, S. J. (2006). A cancer cellmetalloprotease triad regulates the basement membrane transmigration program.Genes Dev. 20, 2673-2686.
Ishisaki, A., Hayashi, H., Li, A. J. and Imamura, T. (2003). Human umbilical veinendotheliumderived cells retain potential to differentiate into smooth muscle-like cells.J. Biol. Chem. 278, 1303-1309.
Koike, T., Vernon, R. B., Hamner, M. A., Sadoun, E. and Reed, M. J. (2002). MT1-MMP, but not secreted MMPs, influences the migration of human microvascularendothelial cells in 3-dimensional collagen gels. J. Cell. Biochem. 86, 748-758.
Korff, T., Kimmina, S., Martiny-Baron, G. and Augustin, H. G. (2001). Blood vesselmaturation in a 3-dimensional spheroidal coculture model: direct contact with smoothmuscle cells regulates endothelial cell quiescence and abrogates VEGF responsiveness.FASEB J. 15, 447-457.
Koshikawa, N., Giannelli, G., Cirulli, V., Miyazaki, K. and Quaranta, V. (2000). Roleof cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5.J. Cell Biol. 148, 615-624.
Lafleur, M. A., Forsyth, P. A., Atkinson, S. J., Murphy, G. and Edwards, D. R. (2001).Perivascular cells regulate endothelial membrane type-1 matrix metalloproteinaseactivity. Biochem. Biophys. Res. Commun. 282, 463-473.
Lafleur, M. A., Handsley, M. M., Knauper, V., Murphy, G. and Edwards, D. R.(2002). Endothelial tubulogenesis within fibrin gels specifically requires the activity ofmembrane-typematrix metalloproteinases (MT-MMPs). J. Cell Sci. 115, 3427-3438.
Lafleur, M. A., Mercuri, F. A., Ruangpanit, N., Seiki, M., Sato, H. and Thompson,E. W. (2006). Type I collagen abrogates the clathrin-mediated internalization ofmembrane type I matrix metalloproteinase (MT1-MMP) via the MT1-MMPhemopexin domain. J. Biol. Chem. 281, 6826-6840.
Lehti, K., Allen, E., Birkedal-Hansen, H., Holmbeck, K., Miyake, Y., Chun, T. H.and Weiss, S. J. (2005). An MT1-MMP-PDGF receptor-� axis regulates mural cellinvestment of the microvasculature. Genes Dev. 19, 979-991.
Liebner, S., Cattelino, A., Gallini, R., Rudini, N., Iurlaro, M., Piccolo, S. and Dejana,E. (2004). �-Catenin is required for endothelial-mesenchymal transformation duringheart cushion development in the mouse. J. Cell Biol. 166, 359-367.
Ling, Q., Jacobina, A. T., Deora, A., Febbraio, M., Simantov, R., Silverstein, R. L.,Hempstead, B., Mark, W. H. and Hajjar, K. A. (2004). Annexin II regulates fibrinhomeostasis and neoangiogenesis in vivo. J. Clin. Invest. 113, 38-48.
Mays, P. K., Bishop, J. E. and Laurent, G. J. (1988). Age-related changes in theproportion of types I and III collagen. Mech. Ageing Dev. 45, 203-212.
Ohuchi, E., Imai, K., Fujii, Y., Sato, H., Seiki, M. and Okada, Y. (1997). Membranetype 1 matrix metalloproteinase digests interstitial collagens and other extracellularmatrix macromolecules. J. Biol. Chem. 272, 2446-2451.
Paruchuri, S., Yang, J. H., Aikawa, E., Melero-Martin, J. M., Khan, Z. A.,Loukogeorgakis, S., Schoen, F. J. and Bischoff, J. (2006). Human pulmonary valveprogenitor cells exhibit endothelial/mesenchymal plasticity in response to vascularendothelial growth factor-A and transforming growth factor-�2. Circ. Res. 99, 861-869.
Plaisier, M., Kapiteijn, K., Koolwijk, P., Fijten, C., Hanemaaijer, R., Grimbergen, J.M., Mulder-Stapel, A., Quax, P. H., Helmerhorst, F. M. and van Hinsbergh, V. W.(2004). Involvement of membrane-type matrix metalloproteinases (MT-MMPs) incapillary tube formation by human endometrial microvascular endothelial cells: roleof MT3-MMP. J. Clin. Endocrinol. Metab. 89, 5828-5836.
Sabeh, F., Ota, I., Holmbeck, K., Birkedal-Hansen, H., Soloway, P., Balbin, M.,Lopez-Otin, C., Shapiro, S., Inada, M., Krane, S. et al. (2004). Tumor cell trafficthrough the extracellular matrix is controlled by the membrane-anchored collagenaseMT1-MMP. J. Cell Biol. 167, 769-781.
Saunders, W. B., Bohnsack, B. L., Faske, J. B., Anthis, N. J., Bayless, K. J., Hirschi,K. K. and Davis, G. E. (2006). Coregulation of vascular tube stabilization byendothelial cell TIMP-2 and pericyte TIMP-3. J. Cell Biol. 175, 179-191.
Seandel, M., Noack-Kunnmann, K., Zhu, D., Aimes, R. T. and Quigley, J. P. (2001).Growth factorinduced angiogenesis in vivo requires specific cleavage of fibrillar typeI collagen. Blood 97, 2323-2332.
Seiki, M. and Yana, I. (2003). Roles of pericellular proteolysis by membrane type-1matrix metalloproteinase in cancer invasion and angiogenesis. Cancer Sci. 94, 569-574.
Shofuda, K., Yasumitsu, H., Nishihashi, A., Miki, K. and Miyazaki, K. (1997).Expression of three membrane-type matrix metalloproteinases (MT-MMPs) in ratvascular smooth muscle cells and characterization of MT3-MMPs with and withouttransmembrane domain. J. Biol. Chem. 272, 9749-9754.
Sounni, N. E., Devy, L., Hajitou, A., Frankenne, F., Munaut, C., Gilles, C., Deroanne,C., Thompson, E. W., Foidart, J. M. and Noel, A. (2002). MT1-MMP expressionpromotes tumor growth and angiogenesis through an up-regulation of vascularendothelial growth factor expression. FASEB J. 16, 555-564.
Suri, C., Jones, P. F., Patan, S., Bartunkova, S., Maisonpierre, P. C., Davis, S., Sato,T. N. and Yancopoulos, G. D. (1996). Requisite role of angiopoietin-1, a ligand forthe TIE2 receptor, during embryonic angiogenesis. Cell 87, 1171-1180.
Timmerman, L. A., Grego-Bessa, J., Raya, A., Bertran, E., Pérez-Pomares, J. M.,Diez, J., Aranda, S., Palomo, S., McCormick, F., Izpisúa-Belmonte, J. C. et al.(2004). Notch promotes epithelial-mesenchymal transition during cardiac developmentand oncogenic transformation. Genes Dev. 18, 99-115.
von Tell, D., Armulik, A. and Betsholtz, C. (2006). Pericytes and vascular stability. Exp.Cell Res. 312, 623-629.
Zhou, Z., Apte, S. S., Soininen, R., Cao, R., Baaklini, G. Y., Rauser, R. W., Wang, J.,Cao, Y. and Tryggvason, K. (2000). Impaired endochondral ossification andangiogenesis in mice deficient in membrane-type matrix metalloproteinase I. Proc.Natl. Acad. Sci. USA 97, 4052-4057.
Zhu, W. H., Han, J. and Nicosia, R. F. (2002). Requisite role of p38 MAPK in muralcell recruitment during angiogenesis in the rat aorta model. J. Vasc. Res. 40, 140-148.
Zijlstra, A., Aimes, R. T., Zhu, D., Regazzoni, K., Kupriyanova, T., Seandel, M.,Deryugina, E. I. and Quigley, J. P. (2004). Collagenolysis-dependent angiogenesismediated by matrix metalloproteinase-13 (collagenase-3). J. Biol. Chem. 279, 27633-27645.
Journal of Cell Science 120 (9)
Jour
nal o
f Cel
l Sci
ence
Jour
nal o
f Cel
l Sci
ence




![MT1 KS4 Edexcel GCSE World Music[1]](https://static.fdocuments.in/doc/165x107/551412be4a7959c4028b4e7a/mt1-ks4-edexcel-gcse-world-music1.jpg)


![Mt1[1] sjkt new bala edit 24 apr](https://static.fdocuments.in/doc/165x107/5589ab8bd8b42a511f8b468a/mt11-sjkt-new-bala-edit-24-apr.jpg)










