
Gabion Baskets specialises in the manufacture of products ...

Osmoregulation in Juvenile and Adult Lampreys1
Department c.fZoology, University oj'Guelph, Guelph, Onr. NIG 2 WI
BEAMISH, F. W. M. 1980. Osmoregulation in juvenile and adult lampreys. Can. J. Fish. Aquat. Sci. 37: 1735-1750.
Nonparasitic lampreys are well adapted for a freshwater existence but are less tolerant of salt water than representatives of the parasitic species. Anadromous feeding juvenile sea lampreys, Purromyzon marinus, are able to control serum osmotic and ionic concentrations in all salinities between 0 and 34%. Juveniles of the anadromous European river lamprey, Lamperra fluviurilis, are slightly less tolerant of salinity change. The transition of P. marinus to a landlocked existence has resulted in a reduction in the capacity for marine osmoregulation. Tolerance to salt water among landlocked P. marinus is related directly to size emphasizing the advantage aflforded through a reduction of surface area relative to body volume. Changes in serum osrnolality are primarily the result of corresponding shifts in sodium and chloride ions. Changes in chloride are not accompanied by an equivalent shift in sodium suggesting that the relationship between the two ions is not passive. Regulation of sodium by landlocked P. marinus is not precise as that exhibited by the anadromous form particularly in the higher salinities. Nontrophic upstream migrant lampreys display a reduction in their marine osmo- regulatory mechanisms and a restoration of the freshwater osmoregulatory apparatus. Fresh- water osmotic and ionic regulation deteriorates in spent adult lampreys.
Key words: blood, ionic regulation, osmotic regulation, parasitic and nonparasitic lampreys, salinity, life cycle
BEAMISH, F. W. M. 1580. Osmoregulation in juvenile and adult lampreys. Can. J. Fish. Aquat. Sci. 37: 1739-1750.
Les lamproies non parasites sont bien adaptees Zt une existence en eau douce mais ssnt moins tolerantes B I'eau salee que les representants d'espkces parasites. Les jeunes lamproies marines, Petrorqmta mclriraus, anadromes et se nourrissant peuvent regler les concentrations osmotiques et ioniques de leur &rum B toute salinite entre 0 et My&,. Les jeunes de la lamproie fluviatile europeenne anadrome, bnrmpetra j?uviarilis, sont legkrement moins tolerants ti un changement de salinite. Le passage de P. marinus B une existence confinee en eau douce entraina une diminution de capacite osmorCgulatrice en mer. Chez P. r?aur.irtus confine en eau douce, la tolerance h 19eau salke est dirwtement like % la taille, ce qui fait ressortir l'avantage d'une diminution de surface par rapport au volume du corps. Les changements d'oxmolalite serique rCsultent avant tout du glissement correspondant des ions sodium et chlorure. Les change- ments de chlorure ne sont pas accompagnes d'un glissement equivalent de sodium, suggerant que la relation entre les deux ions n'est pas passive. La regulation du sodium par P. marinus confine en eau douce n'est pas aussi precise yue celk de la forme anadrome, en particulier aux fortes salinites. Les lamproies se depla~ant vers l'amont montrent une diminution de leurs mkanismes d'osmsr6gulation en mer et une restauration de leur appareil osmoregulateur d'eau douce. La regulation osnlotique et ionique d'eau douce se detCriore cher. les lamproies adultes qui ont fray&
Received September 24, 1579 Accepted March 26, 1980
MOST vertebrates are tolerant of small changes in internal ionic composition and concentration. However, the variation compatible with life is extremely limited. Among marine teleosts and cyclostomes (Petromy- zontia) body water is lost f rom the hypoosmotic blood
T h i s paper forms part of the Proceedings of the Sea Lamprey International Symposium (SLIS) convened at Northern Michigan University. Marquette, Michigan, July 30-A~gust 8, 1979.
Printed in Canada (J5789) Imprim6 au Canada (J5789)
R e p le 24 septembre 1979 Accepte le 26 mars 1980
a t the gills and across the integument (Black 1957) . Water loss is countered by a low urine production (Morris 19721, swallowing of seawater (Krogh 19391, and the entrance of ambient water across the integu- ment (Potts 1968). Cyclsstomes, unlike teleosts, have no mineralized tissue with which t o exchange and accumulate ions of the extracellular fluid and Bdood (Urist 1963 1. Excess monovalent ions absorbed f rom the digestive tract are believed to be secreted by spe- cialized giIl epithelial o r "chloride" cells in bath the cyclostomes and teleosts (Keys 193 1; Dsy le and Gorecki 1961 ; Threadgsld and Houston 1964: Picker-

1740 CAW. S. FISH. AQUAT. SCH., VOL. 37, I980
ing and Morris 8976). Bivalent ions remain in the gut and are removed in the feces (Hoar 1966). Fresh- water homeostasis depends on the elimination of water from the hyperosmotic blood coupled with the trans- portation of ions from the ambient environment or, in the case of teleosts, from skeletal stores. The osmotic influx of water in freshwater fishes is controlled by a copious dilute urine (Parry 1966; Morris 1972). Salt loss is replaced from ingested food (Hoar 1966) and active recruitment by the ion-transporting or "chloride" cells (Peek and Youson 1979a).
Juvenile lampreys are found in both fresh and saline waters. Those species which do not feed during the juvenile interval of their life cycle are found only in freshwater. Of the parasitic species some spend the juvenile trophic portion of their life cycle in saline water before returning as adults to freshwater rivers in which they reproduce and thereafter die. Other parasitic species remain in freshwater throughout their entire life cycle. This report summarizes present information on the capacity of juvenile and adult lampreys for ionic and osmotic regulation and the mechanisms by which con- trol is achieved.
Regulatory Mechanisms
Water balance and ionic-composition and concentra- tion are regulated in lampreys by the gills, kidney, gut, and integument. In a saline environment juvenile Bam- preys maintain water and ion balance by drinking seawater. Monovalent ions are absorbed by specialized cells located in the anterior portion of the digestive tract, with any excess being excreted via the gills. Most divalent ions remain in the gut and are excreted along with the nondigestible components of the ingested food. Excess divalent ions which enter the body fluids are excreted by the kidneys in concentrated urine. Ira juveniIe lampreys the foregut is a new structure which develops during transformation by formation of a lumen in a solid epithelial chord (Hardisty et al. 1970). The timing of the opening sf the foregut appears not to be in synchrony within and among species, a factor which would be expected to contribute to variability in osmoregulatory efficiency. Thus, while the foregut of most specimens of the anadromous Eampetra Ruviatilis opens by Iate October, it may remain closed in some animals until May (Weissenberg 1927). Pre- sumably the foregut opens earlier in Betromyzon marinus than in river lampreys (Hardisty and Potter 1971). In freshwater, ion balance is maintained by the active uptake of salts at the gills, gut, and kidney to- gether with the production of dilute urine.
The gills of many teleosts and lampreys are char- acterized by specialized ion transport cells widely known as "chloride9' cells. In the marine or estuarine environment, chloride cells are believed to be involved in ion excretion whereas in freshwater they are thought to be responsible for ion absorption. Chloride cells are involved also in the elimination of nitrogenous wastes
in the form of ammonia (Read 1968; Be Renzis and Maetz 1973; Evans 1975) which may account at least in part for their active appearance in lampreys adapted to freshwater (Peek and Youson 1979a). Several types of chloride cells are present within the gill epithelium of most fishes (Doyle and Gorecki 1961; Threadgold and Houston 1964) and the anadromous P . marinus (Peek and Youson 1979a1, likely reflecting stages of cell differentiation (Conte and Lin 1967) or cells at various levels of activity (Shirai and Utida 1970). The appearance of cells is influenced also by the ambient salinity (Philpott and Copeland 1963; Shirai and Utida 1970).
Ultrastructural studies on the interlamellar regions of juvenile and adult lamprey gills (Nakao 1974, 1977; Pickering and Morris 1976; Youson and Freeman 1976; Peek and Youson 1979a) suggest the presence of chloride cells with similar features to those displayed by marine teleosts (Philpott and Copeland 1963; Shirai and Utida 1970; Newstead 1971; Kikuchi (1977). Chloride cells from lampreys have akundant mito- chondria and a highly developed reticulum of smooth cytoplasmic tubules continuous with the Iateral and basal plasma membranes. The continuity of the smooth endoplasmic reticulum with the plasma membrane in young juvenile landlocked P. marinus (Youson and Freeman 1976) has been observed also in the chloride cells of E. .iapo~licn (Nakao 1974; Wakao and Uchi- nomiya 1974) and early upstream migrants of L . fleaviatilis (Morris and Pickering 1976).
Among larval lampreys, chloride cells are few in number and resemble those identified in freshwater teleosts in that their smooth endoplasmic reticulum is confined to the apical cytoplasm in the form of vesicles. Chloride cells in larval lampreys are rich in mito- chondria (Morris and Pickering 1975; Youson and Freeman 1976) and are believed to assist in ion uptake. The rapid increases in osmolality which have been ob- served for landlocked sea Iamprey during metamor- phosis (Mathers and Beamish 1974) appear to coincide with the early formation of the juvenile type of chloride cell (Peek and Youson 8979b). During the upstream spawning migration the juvenile type of chloride cells degenerate and are replaced by the larval type (Morris and Pickering 1976).
The biochemical mechanism by which lampreys transport ions across the gill epithelium has only re- cently been studied (Beamish et al. 1978). At the site of the chloride cells a sodium-potassium dependent ATPase enzyme ( (Na-K) -ATPase) has been identi- fied in both teleosts (Epstein et al. 1967; Kamiya and Utida 1968; Pfeiler and Kirschner 1972; Zaugg and McLain 1976) and anadromous sea lampreys (Beamish et al. 1978). This enzyme is believed to constitute the pump responsible for the outward transport of sodium and inward movement of potassium across the gill epithelium (Adams et aI. 1975). Gill (Na-K) -ATPase activity is absent in ammocoetes of the anadromous sea lamprey but present in young feeding juveniIes of

BEAMISH : LAMPREY OSMOREGULATIQN 1741
this species (Beamish et al. 1978). Peek and Youson ( 1979a) suggest that the site of (Na-K)-ATPase may be the membrane of the cytoplasmic tubules in the chloride cells.
Gill (Na-K)-ATPase activity declines with the age of the anadromous sea lamprey, being highest in young feeding animals in salt water and effectively absent from spawning migrants in freshwater. The function- ing of (Na-K) -ATPase in small juvenile sea lampreys presumably enhanccs the efficiency with which they are able to make the transition from fresh to salt water. Once lampreys begin feeding, thc composition of body fluids ingested from the host fish may facilitate osmo- regulation and at the same time stimulate the animal's capacity for osmoregulation making the role of the enzymc less important. Further, as lampreys increase in size the corresponding reduction of the surface area in relation to volume lowers the osmotic stress exerted by the external medium.
Lamprey kidncys are largely responsible for ex- creting osmotic water and the absorption of solutes which in freshwater assists in the preservation of thc ionic composition of body fluids (Morris 1960). The paired kidneys are ustially unequal in size, with one longer and of greater cross-sectional area (Youson et al. 1974). The structure of the larval pronephros and opisthonephros has been described in detail (Whecler 1899; Hatta 1900; Inukai 1929: Youson and McMillan 1970a, b, 1971a, b) . The larval kidney degenerates anteriorly throughout life and disappears entirely dur- ing metamorphosis at which time the opisthonephros develops (Schneider 1879; Youson 1970; Ooi and Youson 1979). The opisthonephric kidney degen- erates gradually until the cnd of the spawning migra- tion when a massive deterioration occurs (Morris 1972). Details of the structure of the opisthonephros are given in Muller ( 1875), Meyer (1876), Wheeler ( 1899), Krause ( 192 1 ) , Viilnichcnko ( 1966), Youson and McMillan 1970a, b, 1971 a, b) , and others.
Measurements of urine production and quality have been made for lampreys in freshwater (Wikgren 1953: Sawyer 1955; Morris 1956; Hardisty 1956; Bentley 1962; Bentley and Follett 1963: Bull and Morris 1967; Pickering and Morris 1970; Malvin et al. 1970). Most measurements are, however, subject to question due to handling diuresis (Morris 1972) or the effects of anesthesia (Moriartv et al. 1978). 'The most reliable measurements indicate that lampreys have a high urine production relative to that of freshwater tcleosts. Some ions are excreted in aswciation with the high produc- tion of urine. Wikgren ( I 953) estimated electrolyte excretion in the urine to represent 2.5-309; of the total ionic loss. Other estimates have been made and are discussed in detail in Morris (1972). Calculations by Morris (1972) for lamprevs in freshwater suggest that sodium, potassium. and chloride are largely re- absorbed by the kidney although absorption efficiency for potassium is not high. Further. the kidney reabsorbs approximately one half of the water produced as a
result of glomerular filtration (Morris 1972). Efficiency of ion absorption and water reabsorption varies directly with temperature (Wikgren 1953). Morris ( 1972) suggests that this is consistent with the hypothesis that water absorption is a passive process resulting from the osmotic flow of water from the urine to the blood. Thus, a reduction in ion absorption at low temperature could cause a decline in water reabsorption.
Permeability of the integument to water has been investigated for L. plnneri and L. fluvintilis in fresh- water by ligating the urinogenital papilla and measur- ing weight change (Hardisty 1956). Urine production determined in this manner coincided percisely with direct measurements made by Wikgren (1953). Hardisty (1956) suggests that the branchial epithelium plays the major role in water transport. On the basis of equal body weights the permeability of the integu- ment to water was estimated to be about twice as great in L. pluneri as in L. fluviatilis.
In salt water, juvenile lampreys are believed to pro- duce very limited quantities of urine (Morris 1972). It is important to point out that kidney function in salt water is poorly understood for lampreys. Qn the basis of glomerular filtration rates obtained by Bentley and Follett (1963) together with consideration for tubular absorption of water, Pickcring and Morris (1970) calculated that kidney tubules preferentially secrete divalent ions.
Regulation of salt and water balance in the higher vertebrates is under hormonal control. Water cxcretion among the higher vertebrates may be regulated by one or more hypophysial peptides such as arginine vasotocin (Kobertson 1974). Among the primitive cyclostomes the physiological importance of hypophysial peptides is still uncertain (Hardisty 1972; Robertson 1974). Injection of arginine vasotocin into L. fluviatilis did not cause a change in body weight nor urine volume. I t did, however, cause an elevation in urine sodium and potassium with an increased total renal loss of sodium (Bentley and Follett 1963). Presumptive adrenocortical cells exist in association with the cardinal veins of lampreys and may be the source of corti- costeroids influencing salt and watcr balance (Chester Jones 1963). An attempt to identify and quantify cor- tisol and corticosterone in the plasma of the land- locked sca lamprey has shown only doubtful traces of these compounds (Weisbai-t and Idler 1970). Injections of aldosterone caused a slight decline in urine output but significant changes in total urinary loss of sodium or potassium were not demonstrable (Bentley and Follett 1963). In saline water aldosterone reduced the total loss of sodium which caused Bentlcy and Follett (1963) to suggest that the hormone inhibits the extrarenal loss of sodium at the gills. Certainly, at present it is not possible to generalize on the hormones present in the pituitary-hypothalamus complex or those of the adrenocortical tissue, or on their function in osmotic and ionic regulation (Robertson 1974; Larsen 1980; Gorbman 1980).

1742 CAN. J. FISH. AQUAT. SCP., VOL. 37, 1980
Regulation and Salinity Tolerance During Metamorphosis
Subsequent to the freshwater sedentary larva1 stage, lampreys undergo a nontrophic period of metamor- phosis during which a number of morphological and physiological changes occur (Manion and Stauffer 1970; Hardisty and Potter 1971; Potter et al. 1978). Among the parasitic lampreys, these changes include the machinery by which internal ionic and osmotic concentration can be regulated over a greater- range of ambient salinities than can be tolerated by the larvae. Ammocoetes are well adapted for life in freshwater; however, in saline water they exhibit a rather ineffective control of their internal osmotic and ionic concentra- tion (Hardisty 1956; Morris 1972; Mathers and Beamish 1974; Beamish et al. 1978). Accordingly larval lampreys have a relatively low tolerance for salt water. Mortalities of larval anadromous sea lamprey were recorded in a salinity as low as 10% with the 24-h LC50 being 14.8%c (Beamish et al. 1975). Larvae of the nonparasitic species appear equally sensitive to salinity. Thus, Hardisty (1956) found mortaIities among larval L. pluirc~ri in solutions of NaCl equivalent to about 280 mosmol . kg-I or 1 1 %.
Osmotic regulation during nletamorphosis has been measured only for the landlocked sea lamprey (Mathers and Beamish 1974). During the early stages of rneta- morphosis serum osmolality of the landlocked sea lamprey in freshwater increases rapidIy reaching a plateau by the early macrophthalmia stage (Fig. 1 , Mathers and Beamish 1974). Serum osmolality of lar- val sea lampreys in freshwater is -260 mocmol- kg-I. By the early macropkthalmia stage osmolality increases to 304 mosmole kg-' and remains at a similar level to the completion of metamorphosis. Not until the macrophthalmia stage is reached are sea lampreys abIe to osmoregulate in a saline solution equivalent to 1 0 % ~ . After 1 d exposure to a salinity of logc, osmolality of ammocoetes and metamorphosing stages III and IV (Manion and Stauffer 1970) are -344, 333, and 335 mosmol kg-I, respectively. Osmolality of early macrophthalmia in 1 O$ is about 300 mosmol kg-', similar to that in freshwater.
Salinity Tolerance in Juveniles and Adults
On completion of metanlorphosis, parasitic juvenile lampreys migrate downstream to either a lake or ocean where they begin the second trophic interval of their life cycle. Completion of the downstream migration may be delayed by as much as 6 mo during which time lampreys remain within the river without feeding (Applegate 1950: Potter and Beamish 1977). Of those species which spend their juvenile interval in freshwater only the landlocked P. marinus has been examined with respect to salinity tolerance and osmotic performance (Mathers and Beamish 1974). Among the marine lampreys, osmotic relationships have been determined for P. marinus, L. fluviatilis, and Larnpetra tridentata
(Fontaine 1930; Galloway 1933; Hardisty 1956; Morris 1956, 1958; Pickering and Morris 1970; Potter and Huggins 1973: Beamish et al. 1978; J. Richards, Bep. Fish. Oceans, Nanaimo, B.C., personal communica- tion).
Adjustment to seawater soon after metamorphosis has been demonstrated for P. marinus (Beamish et al. 1978), I;. JEuvifitilis (Hardisty 1956: Potter and Hug- gins 1973), and I;. tridentata (J . Richards personal communication). Direct transfer of young anadromous juvenile P. marinus and I;. tridrntuta to full seawater did not result in any mortalities within a 2-wk experi- mental period. Indeed, the transfer of L, tridentatn t(9
full seawater appears to encourage the initiation of Geeding. Young juvenile L. fluvkatilis are slightly better able to withstand direct transfer from fresh to full sea- water than metamorphosing individuals of the same species (Potter and Huggins 1973). However, for both stages some mortalities were recorded over the 3-wk experimental period. Mortality of downstream migrant individuals captured in March was 25$, when exposed to full scawater compared with 20%( among animals taken in Maylearly June. About 30% of the metamor- phosing animals when transferred directly to full sea- water died within the experimental 3-wk period. Sur- vival of j~lveniIes was almost complete if the transfer to full seawater was made by exposing animals to increments of one third seawater each for 4 d.
Tolerance to salt water among the landlocked P. nzarinus is related to size of the feeding animals (Mathers and Beamish 1974). Over one half of the landlocked feeding juveniles of 127-188 mm died within 10 d following direct transfer to 26%. Only when landlocked juveniles reached almost 280 rnm did they survive direct transfer to full seawater for periods of -2 wk.
Coincident with the completion of the juvenile inter- val and the migration into freshwater rivers prior to reproduction is a marked reduction in the marine osmoregulatory mechanism. Galloway (1933) and ( 1956) found that adult migrant anadron~ous P. marinrrs and L. fliiviatilis immersed in one half sea- water died within 24 h. Beamish et al. (1978) deter- mined the salinity at which 50% of the anadromous early upstream migrant P. marinus died within 48 h to be -15.2%C which is in general accord with that found by Morris ( 1956).
Nonparasitic lampreys appear to be less tolerant of salinity than representatives of the parasitic species. Hardisty (1956) found some mortalities among juvenile L. planeri exposed to a solution of 10% NaCl. hlor- tality was complete at 22 and 34%( NaCl within 6 and 3 h, respectively. More recently J. Richards (personal communication) found that gradual exposure of juve- nile I;. richrrrdsoni to saline water equivalent to 15% rcsulted in total mortality within 4 or 5 d.
Osmotic and Ionic ReguIation The regulatory mechanisms in marine feeding lam-

BEAMISH: LAMPREY OSMOREGULATION
Landlocked P. marinus
July Sept. Oct. Nor. Dee* Jan. Maxup Dec. JunxIy Aug.
Transforming
I II 8 1 1 IV Macropthalmia Feeding Juvenile Near ly
Mature
Anadromous
P. marinus L . f luviat i l is
FIG. 1. QsmoIality, chloride, and sodium concentration of the life stages of the Imd- locked and anadromous Petromyzon nzurinus (Mathers and Beamish 1974; Beamish et al. 1978) and Lunlpetra fZuviatiJis (Bekhuyzen 1904; Galloway 1933; Hardisty 1956; Picker- ing and Morris 1970) in freshwater. Values for P. mariws (10 + 1°C) are based on blood serum, those for L. fluviutilis on plasma. Mean length (nun) is given within or beside the bar and the sample size is provided on top of each bar.
preys are well developed but unablc to provide for environment maintain their blood hyperosmotic to complete blood homoiosmoticity between fresh and full freshwater and hypoosmotic to seawater. seawater. Juveniles of the landlockcd forms and the In freshwater, feeding landlocked sea lampreys ex- nonparasitic species are unable to maintain as tight a hibit osmotic independence (Urist and Van der Putte control over their blood concentration in saline water 1967; Mathers and Beamish 1974). There is, however, as that exhibited by the anadromsus species. Feeding a significant increase in osmotic concentration between individuals of all species that are tolerant of the saline the larval and juvenile stage which signals a change in

1744 C.4N. J. FISH. .4QUA4T. SCF.. VQL. 37, 1980
the regulatory mechanisms during metamorphosis (Mathers and Beainish 1974). Serum osrnolality de- clines precipitousIy during the period of reproduction to a level comparable to that for ammocoebes (Fig. I ) . This decrease undoubtedly reflects an increase in skin permeability (Hardisty 1956; Morris 1958), and a degeneration o l the kidney (Youson 1970), and the gut (Pickering and Morris 1970; Pickering and Dockray 1972).
The anadroinous sea lamprey displays much greater variability in osmotic concentration in freshwater throughout the juvenile interval of its life cycle than the landlocked form (Fig. I ) . Despite this variability osmotic concentration remains consistently below that for the landlocked form. The decline in osmolality with size of feeding anadromous individuals is likely a reflection of the adjustment of the regulatory mecha- nisms for a marine existence during this period of the life cycle.
Upstream migrant adults exhibit an osmolality in freshwater which is above that of feeding juveniles. This rise apparently signals the restoration of the fresh- water osmoregulatory apparatus. Freshwater osmoregu- lation deteriorates in spent adults indicating the tissues responsible for maintaining hyperosmoticity have ceased functioning efficiently, perhaps coupled with a leakage of water across the integument. Serum osmolality in migrant sea lamprey appears to be inde- pendent of body length (Fig. 2b suggesting that regula- tion of ionic constituents depends to a lesser extent on integumcnt permeability than on other mechanisms.
Plasma osmotic concentration has been measured in freshwater for the nontrophic upstream migrant L. flznviatilis (Fig. 1 ) . Unfortunately measurements have not been made on spent individuals. However, L. fluviatilis demonstrates greater osmotic independence than the anadromous sea lamprey during this phase of the life cycle.
Changes in osmotic concentration in freshwater are mainly attributable to fluctuations in chloride and sodium concentrations (Fig. 1) . Pickering and Morris (1970) suggest the low chloride values for early migrant L. fluvintilis, and L. tridentata (Urist 1963) indicates that the freshwater regulatory mechanism is incom- pletely established when individuals first enter fresh- water. However, in the anadromous sea lamprevs both sodium and chloride were higher in early migrants than in feeding animals and decreased with the ap- proach of sexual maturity.
Relatively few measurements have been made of other blood ions (Table 1 ) . Consistent and low levels of serum calcium and phosphate are prevalent in the blood of the landlocked sea lamprey (Urist and Van der Putte 1967). Urist (1 963) has shown that the lamprey is a typical freshwater vertebrate in its serum calcium and phosphate levels. About 28 and 11% of calcium and phosphate is present as a complex with protein. Early migrant female lampreys display higher calcium levels than males which Pickering and Bockray (1972)
0
0
ondloc ked
0
0
0
0
Nearly M o t u r e
Length rnm
FIG. 2. Osmolality of migrant Pc.tromyacan n~urirzus in relation to length. Measurements were made in freshwater at 10 + 1°C (Mathers and Beamish 1974; Bean~ish et al. 1978).
suggest is related to the process of producing calcium- rich vitellin by the maturing female. Serum potassiun~ in a normally functioning animal is regulated by sodium and throughout the juvenile stages of feeding and early migration remains at a similar concentration. In mature and spent adult lampreys potassium levels are approxi- mately double those in juvcnile individuals, probably a result of leaching from cells which deteriorate during this terminal period of the life cycle.
Feeding anadromous P. rnurirzus are able to osmo- regulate in all salinities between 0 and 34% (Beamish et al. 1978). Qsmolality of small juveniles s f -138 mm in length remains essentially unchanged at 236 inosmol- kg-' between 0 and l6$0 (Table I , Fig. 3) . In higher salinities osmolality increases but only by 10% in full seawater. Small juveniles are capable of acclimating to daily increments of 2% exhibiting no significant change in osmolality over exposure periods of 1-8 d following a given increase in salinity. Serum osmolality of large feeding juveniles, ~ 2 8 2 mm in length increases continuously between 0 and 26%, the highest salinity in which the species has been examined (Fig. 3 ) . Howcver, for identical salinities significant differences in osmolality betwecn the two size-classes of feeding individuals are demonstrable only in fresh- water. Both size-classes maintained hyperosmotic blood serum in 0 and 8Yc and hypoosmotic in higher salinities.
Small feeding juvenile landlocked sea lampreys main-

TA
BL
E
1.
Blo
od p
lasm
a or
ser
um o
smot
ic a
nd io
nic
conc
entr
atio
ns.
Na'
K
- C
a++
M
g++
C
I-
S04
--
HC
03
- P-
- S
alin
ity
Ser
um
/ O
smol
a8ity
P
rote
in
Sta
ge
%
plas
ma
mO
s.kg
-"
mm
ol. L
-I
mg.
mL
-'
Ref
eren
ce
Spe
cies
P. m
arin
us (
land
lock
ed)
Met
amor
phos
ing
Sta
ge 1
S
tage
I1
Sta
ge 1
11
Sta
ge 1
V
Mac
ropk
thal
mia
S
tage
I11
Sta
ge I
V
Mac
roph
thal
mia
M
etam
orph
osin
g
Ser
um
Ser
um
, .
Ser
um
$3
,*a
Pla
sma
Pla
sma
Ser
um
Ser
um
Ser
um
Pla
sma
Pla
sma
Pla
sma
Pla
sma
Mat
hers
an
d B
eam
ish
(197
4)
7.
..
I,
1.
.. 9
.
.. 7
. 9
,
., 3
1
,$
7s
..
.. 7
, 7
.
71
99
7
. 11
1.30
32
.0
Uri
st a
nd
van
der
Put
te (
1967
)
Juve
nile
F
eedi
ng
1.40
35
.0
Uri
st a
nd
van
der
Put
te (
1967
) M
athe
rs a
nd
Bea
mis
h (1
974)
1
$
,. $
1
$3
3
, 7
7
I,
.. 7
.
,I
11
1,
Adu
lt
P.
mar
inus
(an
adro
mou
s) J
uven
ile
Mig
rant
M
igra
nt
Spa
wni
ng
1.40
39
.0
Uri
st (
1963
) 1.
31
Uri
st (
1963
)
Fee
ding
(sm
all)
-
Bea
mis
R e
t al
. (8
978)
a
Fee
ding
(la
rge)
F
eedi
ng (
larg
e)
Fee
ding
(la
rge)
F
eedi
ng
8 R
ober
tson
, in
Mor
ris
(197
2)
c B
eam
ish
et a
l. (
1978
) F
2.5
Pic
keri
ng a
nd
Mor
ris
(897
0)
5 1.
8 F
onta
ine
(193
0, 1
932)
5
Bea
mis
h et
al.
(19
78)
Z
Adu
lt
Ear
ly m
igra
nt
Ear
ly m
igra
nt
Ear
ly m
igra
nt
Nea
rly
mat
ure
Spe
nt
Adu
lt
Adu
lt
Adu
lt
Ear
ly m
igra
nt
Har
dist
y (1
956)
Dek
huyz
en (
1904
) G
allo
way
(18
33)
Rob
erts
on (
1954
) H
ardi
sty
(195
6)
Pic
keri
ng a
nd
Mor
ris
(197
0)
Har
dist
y (1
956)
P
icke
ring
an
d M
orri
s (1
970)
P
icke
ring
an
d M
orri
s (1
970)
Ear
ly m
igra
nt
Nea
rly
mat
ure
Mat
ure
"Con
fide
nce
lim
its
95 %
. "e
amis
h (u
npub
lish
ed d
ata)
. 'H
igh
valu
es d
ue t
o so
me
kem
olys
is o
f th
e sa
mpl
e.
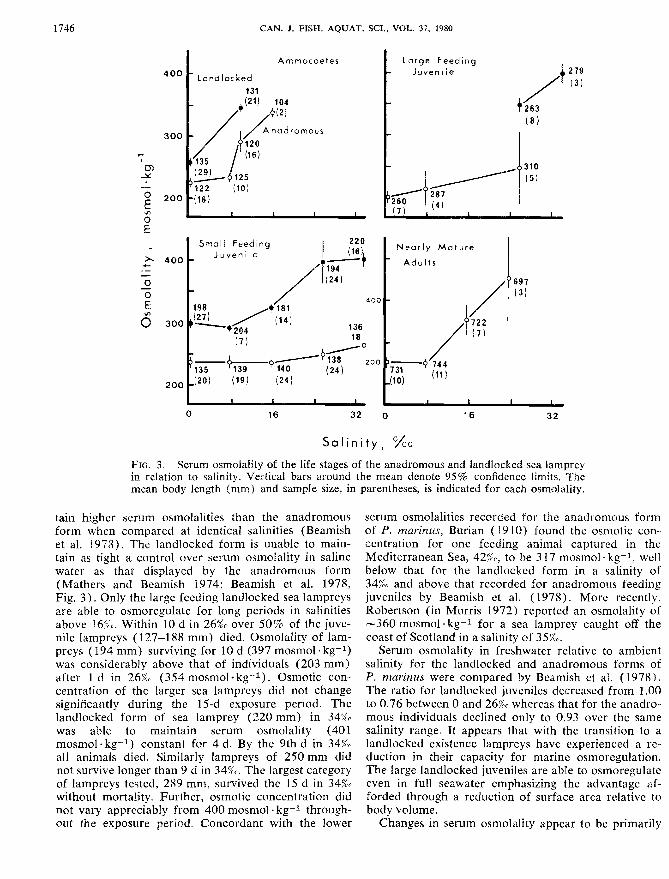
CAN. J . FISH. AQUAT. SCI.. VOL. 37, 1980
A m m o c o e
104 O(2)
nad romous
tes L a r g e F e e d i n g
J u v e n i l e /I ::: 263
N e a r l y M a t u r e
I
FIG. 3. Serum osmolality of the life stages of the anadromous and landlocked sea lamprey in relation to salinity. Vertical bars around the mean denote 95% confidence limits. The mean body length (mm) and sample size, in parentheses, is indicated for each osmolality.
tain higher serum osmolalities than the anadromous form when compared at identical salinities (Beamish et al. 19'78). The landlocked form is unable to main- tain as tight a control over serum osmolality in saline water as that displayed by the cmadromous form (Mathers and Bearnish 1974; Beamish et al. 1978, Fig. 3 ) . Only the large feeding landlocked sea lampreys are able to osmoregulate for long periods in salinities above 16rcc. Within 20 d in 26% over 50% of the juve- nile lampreys (127-188 mm) died. Osmolality of lam- preys ( 194 mm) surviving for 18 d (397 mosmol - kg-l) was considerably above that of individuaIs (203 mm) after 1 d in 26% (354 mosmol k g 1 ) . Osmotic con- centration of the larger sea lampreys did not change significantly during the 15-d exposure period. The landlocked form of sea lamprey (220 mm) in 34g0 was able to maintain serum osmolality (401 mosmol kg-] ) constant for 4 d. By the 9th d in 34%( all animals died. Similarly lampreys of 250 mm did not survive longer than 9 d in 34%(. The largest category of lampreys tested, 289 mm, survived thc 15 d in 34%0 without mortality. Further, osmotic concentration did not vary appreciably from 400 mosmol . kg-f through- out the exposure period. Concordant with the lower
serum osmolalities recorded for the anndromous form of P. mnrinus, Burian (19 18) found the osmotic con- centration for one feeding animal captured in the Mediterranean Sea, 42%, to he 3 17 mosmol kg-', well below that for the landlocked form in a salinity of 34% and above that recorded for anadromous feeding juveniles by Beamish et al. (1978). More recently, Robertson (in Morris 1972) reported an osmolality of -360 mosmol kg-1 for a sea lamprey caught off the coast of Scotland in a salinity of 35%.
Serum osrnolality in freshwater relative to ambient salinity for the Iandlocked and anadromous forms of P. marinus were compared by Beamish et al. ( 1978). The ratio for landlocked juveniles decreased from 1.00 to 0.76 between 0 and 26% whereas that for the anadro- mous individuals declined only to 0.93 over the same salinity range. It appears that with the transition to a landlocked existence lampreys have experienced a re- duction in their capacity for marine osmoregulation. The large landlocked juveniles are abIe to osmoregulatc even in full seawater emphasizing the advantage af- forded through a reduction of surface area relative to body volume.
Changes in serum osmolality appear to be primarily

BEAMISH: LAMPREY OShlOREGULATION 1747
2 2 0
Landlocked
1 8 0
1 4 0 Anadronious
1 0 0
Ch lo r i de , mrnol. L-'
FIG. 4 ( A ) . Serum sodium and chloride concentrations of the anadromous sea lamprey. Vertical bars around the mean denote 95% confidence limits. ( B ) Relationship of serum sodium to chloride concentrations. Redrawn from Beamish et al. (1978) .
the result of corresponding shifts in sodium and chloride ions (Table 1, Fig. 4A) (Urist and van Jer Putte 1967; Mathers and Beamish 1974; Beamish et al. 1978). Organic molecules such as glucose, glycerol, and amino acids along with some inorganic ions ac- count for the difference between osmolality and sodium plus chloride (Potts and Parry 1964). In both land- locked and anadromous feeding sea lampreys, changes in chloride ion are not accompanied by an equivalent
shift in sodium, suggesting that the relationship be- tween the two ions is not passive (Fig. 4B). For a given change in chloride, that by sodium was less; however, this was not as pronounccd in the higher salinitics (Fig. 4A) . The close control held over sodium has been demonstrated also for several teleosts (Houston et al. 1968; Byrne et al. 1972: Lutz 1972). Lutz ( 1972) suggests that the physiological significance of sodium is in the uptake of metabolitcs by the cell as well as in the regulation of potassium. Eandlockcd P. marinrts are not able to regulate sodium as pre- cisely as anadromous sea lampreys, particularly in the higher salinities, which may contribute to the difficulties they experience in salt water.
Early migrants of the anadromous L. flimiafi7is fail to regulate their internal osmotic concentration in salinities above - 18%( (Hardisty 1956). Between fresh- water and the isosmotic salinity the hyperosmotic blood serum increases from a value between -250 and 300 (Dekhuyzcn 1904; Galloway 1933; Hardisty 1956; Pickering and Morris 1970) to 450m0smol .kg-~ (Hardisty 1956). well above the range of concentra- tions found for the feeding stages of both the anadro- mous and landlocked forms of P. rncrrinus (Mathers and Beamish 1974; Beamish et al. 8 978).
Osmoregulatory performancc among the nonparasitic Iampreys has been examined in rclation to ambient salinity only for L. plnneri (Hardisty 1956). Animals maintain a hyperosmotic blood serum to a salinity of about 12% at which regulation s f the internal con- ccntration ceases and some mortalities occur.
Nearly mature migrant anadromous sea lampreys are able to regulate their osmotic concentration in dilute saline solutions to about 8Y0 (Beamish et al. 1978). Exposure of nearly mature sea lampreys to 8% for 4 d did not alter significantly the osmolality found after 1 d in this salinity. In 16% serum osmolality rose pre- cipitously to 358 mosmol-kg-l, a significant increase over that found at the lower salinities. The increase in osmolality continued to 24ftC at which it was 466 mosmol-kg-l. Loss of the swallowing mechanism and a reduced capacity to absorb ions and water across the gut wall were suggested by Pickering and Morris (1970) as factors contributing to the breakdown of the marine osmoregmlatory ability in upstream migrat- ing sea lampreys and L. fluvic~tili,~. Freshwater osmo- regulation deteriorated in spent adults. Osmolality and the concentration of sodium and chloride dropped below those of any other stage in freshwater indicating that the tissues responsible for maintaining hyper- osmoticity had ceased to function efficiently, perhaps coupled with a leakage of ions or water across the integument. An increase in integument permeability to water was reported by Morris (1958) and Pickering and Morris (1978) in L. flul-iatilis and P. marinus on the spawning migration. Hardisty ( 1956) found in- creased body watcr in L. planeri and L. fluviatili,~ as each approached sexual maturity. Regulation is restricted further by a massive degencration of the kidneys which

1748 CAN. J . FISH. AQUAT. SCI., VOL. 37, 1980
occurs at the time of spawning (Youson 1970). "The habitat of the earliest agnathans is uncertain.
Robertson (1954, 1957) has argued in favor of a marine environment on the basis of paleontological and physiological evidence, the former being supported more recently by the findings of Denison (1956), White (1958). and Tarlo (1962). In contrast, Marshall and Smith (1930) proposed a freshwater origin and based their views on the presence of a glomerular kid- ney. Robertson ( 1957) and Morris (1965, 1972) have suggested the vertebrate glomerulus may have developed in seawater where it served primarily as an organ of ion balance. I n support of this, Youson et al. (1974) were unable to detect significant differences in the cross- sectional area of the renal corpuscles of freshwater and marine sea lampreys. Certainly a glomerular kid- ney would have facilitated the entry of cyclostomes into freshwater. Volume regulation and conservation of ions by the production of a hypoosmotie urine to- gether with the function of ion regulation would permit animals to maintain a hyperosmotic blood in an ion- deficient environment (Robertson 1957, 1974; Munz and McFarland 1964). O n balance, Morris (1972) concluded the earliest agnathans were marine animals equipped with a primitive osmoregulatory mechanism similar to that of the present-day myxinoids. H e sug- gests that cyclostomes more recently entered freshwater and adapted their metabolic processes to function in a concentration of blood ions beIow that which is pre- valent in marine forms, thereby reducing the osmotic stress between the internal and external environment. The subsequcnt reentry into the marine habitat may have been facilitated by the development of a swallow- ing extrarenal excretory mechanism.
The present-day landlocked forms of parasitic lam- preys as well as the nonparasitic species are believed to have evolved from marine parasitic species (Hubbs 1925, Hubbs and Potter 1971). Hardisty (1956) sug- gested that this evolution involved selection for in- dividuaIs of smaller size and reduced osmoregulatory performance which is cntirely consistent with recent observations o n salinity tolerance, and osmotic and ionic regulation.
Acknowledgments Technical assistance was provided by Ms S. Sadler and
Mrs E. Thomas.
ADAMS, B. L.. W. S. ZAUGG, AND L. R. MGLAIN. 1995. Inhibi- tion of ~ a l t water survival and Na-K ATPase elevation in steelhead trout (Salmo gairdneri) by moderate watet temperatures. Trans. Am. Fish. Soc. 104: 766-769.
APPLEGATE, V. C. 1950. Natural history of the sea lamprey (Petrornyzan marinus) in Michigan. U.S. Fish. Wildl. Serv. Spec. Sci. Rep. Fish. Serv. 55: 237.
BEAMISH, F. W. H., P. D. STRACHAN, AND E. THOMAS. 1978. Osmotic and ionic performance of the anadromous sea lamprey, ~ c . r r o n ~ ~ z o n mttrinrrs. Cornp. Biochem. Physiol. 60A : 435-443. HARDISTY, M. W., AND I. C. POTTER. 1971. The general biology
BENTLEY, P. F. 1962. Permeability of the skin of the cyclo- stome Lampetra fluviutilis to water and electrolytes. Comp. Biochem. Physiol. 6: 95-97.
BENTLEY, P. J., AND B. K. FOLLETT. 1963. Kidney function in a primative vertebrate, the cyclostome Lampetra Jluviutilis. J. Physiol. (Lond.), 169: 902-918.
BLACK, V. S. 1957. Excretion and osmoregulation, p. 163-205. In ha. E. Brown [ed.] Thc physiology of fishes. Vol. 1. Academic Press, New York.
BULL, J. M., AND R. MORRIS. 1967. Studies on freshwater osmoregulatiorl in the ammocoete larva or Lamptra plamv-i (Bloch). 1. Ionic constituents fluid compartments, ionic compartments and water balance. J. Exp. Biol. 47: 485-494.
BYRNE, J. M., F. W. H. BEAMISH, AND R. L. SAUNDEKS. 1972. Inf uence of salinity temperature, and exercise on plasma osmolarity and ionic concentration in Atlantic salmon (Salmo salar). J. Fish. Res. Board Can. 29: 1217-1220.
BURIAN, R. 1910. Funktion der nierenglomeruli und ultra- filtration, Pfluegers Arch. Gesamten Physiol. Menschen Tiere 136: 741-760.
CHESTER JONES, 1. 1963. Adrenocorticostero~ds, p. 488-502. In A. Brodal and R. Fange [ed.] The biology of myxine. Universitetsforlaget, Oslo, Norway.
CONTE, F. P., AND D. L ~ N . 1967. Kinetics of cellular morpho- genesis in gill epithelium during seawater adaptation of Oncorhynchus (Walb.). Comp. Biochem. Physiol. 23: 945-957.
DE RENZIS, G., AND J. MAETZ. 1973. Studies on the mech- anisms of chloride absorption by the goldfish gill : relation with acid-base regulation. J. Exp. Biol. 59: 339-358.
BEKHUYZEN. M. C. 1904. Ergebnisse von osmotischen studien, namentlich bei knochenfischen, an der biologischen sta- tion des bergenser museums. Bergens Mus. Arbok Naturvitensk. Rekke 8 : 3-7.
DENISON, R. H. 1956. A review of the habitat of the earliest vertebrates. Chicago Nat. Hist. Mus. Fieldiana Geol. 11 : 3 59-457.
DOYLE, W. L., AND D. GORECKI. 1961. The so-called chloride cell of the fish gill. Physiol. Zool. 34: 81-85.
EPS~EIN, F. H., A. I. KATZ, AND G. E. PICKFORD. 1967. Sodium and potassium-activated adenosine triphosphatase of gills: role in adaptation of teleostr to salt water. Science 156: 1245-1247.
EVANS, D. H. 1975. Ton exchange mechanisms in fish gills. Cornp. Biochem. Physiol. 51A: 491-495.
FONTAINE, M. 1930. Recherches sur le milieu interieur de la lamproi marine (Petrontyzon marinus). Se? variations en fonction de celles du milieu exterieur. C . R. Acad. Sci. Paris 191 : 680-682.
1932. Sur la relation existant chez %es poissons marins et potamotoques entre le teneur en phosphore inorganique du serum et l'ossification du squellete. C . R. Acad. Sci. Paris 194: 395-396.
GALLOWAY, T. MCL. 1933. The osmotic pressure and saline content of the blood of Perromyza/z jhviatilis. J. Exp. Biol. 10: 313-316.
GOKBMAN, A. 1980. Evolution of the brain-pituitary relation- ship: evidence from the Agnatha. Can. J. Fish. Aquat. Sci. 37: 168S1686.
HARDISTY, M. W. 1956. Some aspects of osmotic regulation in lampreys. J. Exp. Biol. 33: 431-447.
1972. The interrenal, p. 171-192. In M. W. Hardisty and I. C . Potter led.] The biology of lampreys. Vol. 1. Academic Press, London.

BEAMISH: LAMPREY OSMOREGULATION 1749
of adult lampreys, p. 126-207. 11t M. W. Hardisty and I. C. Potter led.] The biology of lampreys. Vol. 1. Aca- demic Press, London.
HAKDISTY, M. W., I. C. POTTER, AND R. STURGE. 1970. A comparison of the metamorphosing and macrophthalmia stages of the lampreys Lumpetra puviatilis and L. pluneri. J. Zool. 162: 383-400.
HATTA, S. 1900. Contributions to the nlorphology of Cyclo- stomata. 11. On the development of pronephros and segmental duct in Petromyzon. J. Coll. Sci. Imp. Univ. Tokyo 13 : 31 1-426.
HOAR, W. S. 1966. General and comparative physiology. 2nd ed. Prentice-Hall, Inc., Englewood Cliffs, NJ. 848 p.
HOUSTON, A. W., R. S. REAVFS, J. A. MADDEN, AND M. A. DE WILDE. 1968. Environmental temperature and the body fluid system of the freshwater teleost. I. Ionic regulation in thermally acclimated rainbow trout (Sulnto guirdneri). Comp. Biochem. Physiol. 25 : 563-582.
Hums, C. L. 1925. The life cycle and growth of lampreys. Mich. Acad. Sci. Arts Lett. Pap. 4: 587-603.
Hums, C. L., AND I. C. POTTER. 1971. Distribution, phylogeny and taxonomy, p. 1-65. In M. W. Hardisty and I. C . Potter [ed.] The biology of lampreys. Vol. 1. Academic Press, London.
INUKAI, T. 1929. Die Vorniere (Pronephros) auf ihrer hochsten Entwick lungsstufe bei Neunaugen (Lunzpetra (Petro- myzorr) fiuviutiiis und pluneri). Z. Mikrosk. Anat. Forsch. 19: 139-162.
KAMIYA, M., AND S. UTIDA. 1968. Changes in activity of sodium-potassium activated adenosine triphosphatase in gills during adaptation of the Japanese eel to sea water. Comp. Biochem. Physiol. 26: 675-685.
KEYS, A. 1931. Chloride and water secretion and absorption by the gills of the eel. Z. Vergl. Physiol. 15: 364.
KIKUCHI, S. 1977. Mitochondria-rich (chloride) cells in the gill epithelia from four species of stenohaline freshwater teleosts. Cell Tissue Res. 180: 87-98.
KRAUSE, R. 1921. Mikroskopische anatomie der wirbeltiere, p. 746-858. In Teleostier, Plagiostomen, Zyklostomen und Leptokardier. Vol. 4. Walter De Gruyter 6i Co., Berlin.
KROGH, A. 1939. Osmotic regulation in aquatic animals. Cambridge University Press, NY. 242 p.
LARSEN. L. 0. 1980. Physiology of adult lampreys with special regard to natural starvation, reproduction, and death after spawning. Can. J. Fish. Aquat. Sci. 37: 1762-1779.
LUTZ, P. L. 1972. Ionic patterna in the teleost. Comp. Biochem. Physiol. 42(3A) : 719-733.
MALVIN. R. L., E. CARLSON. S. LEGAN, AND P. CHURCHILL. 1970. Creatinine reabsorption and renal function in the freshwater lamprey. Am. J. Physiol. 218 : 1506-1 509.
MANION, P. J., AND T. M. STAUFFER. 1970. Metamorphosis 0i the landlocked sea lamprey, Pefronayzon murinus. J. Fish. Res. Board Can. 27: 1735--1746.
MARSHALL, E. K., AND H. W. SMITH. 1930. The glomerular development of the vertebrate kidney. Biol. Bull. Mar Biol. Lab. (Wood, Hale, Mas,.) 59: 135-153.
MATHERS, J. S., AND F. W. H. BLAMISH. 1974. Changes in serum osmotic and ionic concentration in landlocked Pctrornyzon murinus. Comp. Biochem. Physiol. 49A: 677-688.
MEYER, F. 1876. Ueber due Nieren der Flussne~maugen, Petrornyzort fluvhatilis. Ztrbl. Med. Wiss. 14: 20-2 1.
MULLER, W. 1875. Ueber das Urinogenitalysystem des Am- phioxus und der cyclostomen. Jena. Z. Naturwiss. 9: 94-129.
MORIARTY, R. J., A. G. LOGAN, AND J. C. RANKIN. 1978.
Measurement of single nephron filtration rate in the kidney of the river lamprey, Lcrrnpetrafluvi~ltilis L. J. Exp. Biol. 77: 57-69.
MORRIS, R. 1956. The osmoregulatory ability of the lampern (Lmpctru fluviatilis L.) in sea water during the course of its spawning migration. J. Exp. Biol. 33: 235-248.
1958. The mechanism of marine osmoregulation in the lampern (Lumpetr~~ fluvidlis L.) and the causes of its breakdown during the spawning migration. J. Exp. Biol. 35: 649-665,
1960. General problems of osmoregulation with special reference to cyclostomes. Symp. Zool. Soc. London 1 : 1-1 6.
1965. Studies on salt and water balance in Myxine glutinosu (L.). J. Exp. Biol. 42: 359-371.
1972. Osmoregulation, p. 193-239.112 M. W. Hardisty and I. C. Potter [ed.] The biology of lampreys. Vol. 2. Academic Press, London.
MORRIS, R., AND A. D. PICKERING. 1975. Ultrastructure of the presumed ion-transporting cells in the gills of ammocoete lampreys, Lumnpetru Puviatilis (L.) and Lampetra pluneri (Bloch.). Cell Tissue Res. 163 : 327-341.
1976. Changes in the ultrastructure of the gills of the river lamprey, Lampetru jluviutilis (L.), during the ana- dromous spawning migration. Cell Tissue Res. 173 : 271-277.
MUNZ, F. W., AND W. N. MCFARLAND. 1964. Regulatory function of a primative vertebrate kidney. Comp. Bio- chem. Physiol. 13: 381-400.
NAKAO, T. 1974. Fine structure of the agranular cytoplasmic tubules in the lamprey chloride cells. Anat. Rec. 178: 49-62.
1977. Electron microscopic studies of coated mem- branes in two types of gill epithelial cells of lampreys. Cell Tissue Res. 178 : 385-396.
NAKAO, T., AND K. UCHINOMIYA. 1974. Intracisternal tubules of lamprey chloride cells. J. Electronmicrosc. 23: 51-55.
NEWSTEAD, J. D. 1971. Observations on the relationship between "chloride-type" and "pseudobranch-type" cells in the gills of a fish, Oligocottus muculo.sus. Z. Zellforsch. Mikrosk. Anat. 116: 1-6.
001, E. C., AND J. H . YOUSON. 1979. Regression of the larval opisthonephros during metamorphosis of the sea lamprey, Petromyzon mclrims L. Am. J. Anat. 154: 57-79.
PARRY, G. 1966. Osmotic adaptation in fishes. Biol. Rev. Cambridge Philss. Soc. 41 : 392-444.
PEEK, W. D., AND J. H. YOUSON. 1979a. Ultrastructure of chloride cells in young adults of the anadromous sea lamprey, Petrornyzon murinus L., in fresh water and during adaptation to sea water. J . Morphol. 160: 143-164.
1979b. Transformation of the interlamellar epi- thelium of the gills of the anadromous sea lamprey, Petromyzort marinus L., during metamorphosis. Can. J. Zool. 57: 1318-1332.
PFEILER, E., AND L. B. KIRSCHNEW. 1972. Studies on gill ATPase of rainbow trout (Sulmo gairdncri). Biochim. Bisphys. Acta 282: 301-310.
PHILPOTT, C . W., AND D. E. COPELAND. 1963. Fine structure of chloride cells from three species of Funduh. J. Cell Biol. 18 : 389-404.
PICKERING, A. D., AND G . J. DOCKRAY. 1972. The effects of gonadectomy on osmoregulation in the migrating river lamprey, Lcrmpetru fluviarilis L. Comp. Biochem. Physiol. 4lA: 139-147.
PICKERING, A. D., AND R. MORRIS. 1970. Osmoregulation of

1750 CAN. 3. FISH. AQUAT. SCI., VOL. 37, 1980
Lumpetra fluviutilis and Petroinyzon murinus in hyper- tonic solutions. J. Exp. Biol. 53: 231-243.
1976. Fine structure of the interplatelet area in the gills of the macrophthalmia stage of the river lamprey, Lunzptra f77uekztilis (L.) Cell Tissue Res. 168: 433-443.
POTTER, I. C., AND F. W. H. BEAMIYH. 1977. The freshwater biology of adult anadromous sea lampreys Petromvzon nraritnr~. J. Zool. (London) 18 1 : 1 13-130.
POTTER, I. C,, AND R. J. HUGGINS. 1973. Observations on the n~orphology, behaviour and salinity tolerance of down- stream migrating river lampreys (Lrrmpetra fluviurilis). J. Zool. (London) 169: 365-379.
I ~ T T E R , I. C.. G. M. WRIGHT. AND J. M. YOUSON. 1978. Metamorphosis of anadrornous sea lamprey, Petroinyzotz marinus L. Can. J. Zool. 56: 561-570.
POTTS, W. T. W. 1968. Osmotic and ionic regulation. Annu. Rev. Physiol. 30: 73-184.
P o r n , W. T. W., AND G . PARRY. 1964. Osmotic and ionic regulation in animals. Pergamon Press Ltd., Oxford. 423 p.
READ, L. J. 1968. A study of ammonia and urea production and excretion in the frerhwater-adapted form of the Pacific lamprey Entosphmus tridentatus. Comp. Biochem. Physiol. 26 : 455-466.
ROBERTSON, J . D. 1954. The chemical composition of the blood of some aquatic chordates, including members of the Tunicata, Cyclostomata and Osteichthyes. J. Exp. Biol. 3 1 : 424-442.
1957. The habitat of the early vertebrates. Biol. Rev. 32: 156-187.
1974. Osmotic and ionic regulation in cyclostomes, p. 169-193. In M. Florkin and B. T. Scheer [ed.] Chemical zoology. Vol. VIII: Primitive Deuterostomians. Cyclo- stomata, Fishes. Academic Press, New York.
SAWYER, W. H. 1955. Excretion and osmoregulation. In M. E. Brown [ed.] The physiology of fishes. Vol. 1. Academic Press, New York. 194 p. (Unpublished data from V. S. Black)
SCHNEIDER, A. 1870. Beitrage zur vergleichenden Anatomie und Entwicklung-sgeschichte der Wirbelthiere. Reimer, Berlin. p. 85-92.
SHIIPAI, N., AND S. UTIDA. 1970. Development and degenera- tion of the chloride cells during seawater and freshwater adaptation of the Japanese eel, Anguillu japonicu. Z. Zellforsch. 103 : 247-264.
TARLO, L. 73. 1962. The classification and evolution of the Heterostraci. Acta Palaeontol. Polon. 8 : 249-290.
THREADGOLD, L. T.. AND A. H. HOUSTON. 1964. An electron microscope study of the "chloride ccll" of S a l r ~ o salar L Exp. Cell Res. 34: 1-23.
URIST. M. R. 1943. The regulation of calcium and other ions in the serums of hagfish and lampreys. Ann. N.Y. Acad. Sci. 109: 204-311.
URIST, M. R., AND K. A. VAN DE PUTTE. 1967. Comparative
biochemistry of the blood of fishes : identification of k h e s by the chemical composition of serum, p. 271-285. In P. W. Gilbert, R. F. Mathewson, and D. P. Rall [ed.] Sharks, skates and rays. John Hopkins Press. Baltimore.
VINNICHENKO, L. N. 1966. Electron microscope studies of lamprey kidney. In Elekromaya Mikroskopiya Kletov Zhwotnych. Nauka, Moscow.
WEISBART, M., AND D. R. IDLER. 1970. Re-examination of the presence of corticosteroids in two cyclostomes, the Atlantic hagfish (Myxine glutinosa L.) and the sea lamprey (Petronzyzon muritrus L.). J. Endocrinol. 46: 29-43.
WEISSENBERG, R. 1927. Beitrage zur Kenntnis der Biologie und Morphologie der Neunaugen. 2. Das Wefewachstum der Gonaden bei Lampetra fluviatilis und pluiteri. Z. Mikrosk. Anat. Forsch. 8 : 193-249.
WHEELER, U'. M. 1899. The development of the urinogenital organs of the lamprey. Zool. Jahrb. Abt. Anat. Ontog. Tiere 73 : 7-88
WHITE, E. I. 1958. Original environment in Craniates, p. 212-234. In T. S. West011 Led.] Studies on fossil verte- brates. Academic Press, London.
WIKGREN, B. J. 1953. Osmotic regulation in some aquatic animals with special reference to the influence of tempera- ture. Acta Zool. Fenn. 71 : 1-102.
YOUSON, J H. 1970. Observations on the opisthonephric kid- ney of the sea lamprey of the Great Lakes, Petromyzon marivtus L., at various stages during the life cycle. Can. J. ZOO^. 48 : 1313-1 316.
YOUSON, J. H., AND P. A. FREEMAN. 1976. Morphology of the gills of larval and parasitic adult sea lamprey, Petromyzon maritlus L. J . Morphol. 149 : 73-104.
YOUSON, J. H., S. J. HANSEN, AND I. M. CAMPBELL. 1974. A quantitative comparison of the kidneys of the landlocked sea lamprey of the Great Lakes and the anadromous sea lamprey of the Atlantic, Patromvzon marinus L. Can. J. Zool. 52: 1447-1455.
YOUSON, J. H., AND D. B. MCMILLAN. 1970a. The opistho- nephric kidney of the sea lamprey of the Great Lakes. Part 1 . The renal corpuscle. Am. J. Anat. 127: 287-232.
1970b. The opiqthonephric kidney of the sca lamprey of the Great Lakes. Part 2. Neck and proximal segments of the tubular nephron. Am. J. Anat. 127: 233-257.
1971a. The opisthonephric kidney of the sea lamprey of the Great Lakes. Part 3. Intermediate, distal and collecting segments of the adult. Am. J. Anat. 130: 281- 303.
197bb. The opisthonephric kidney of the sea lamprey of the Great Lakes. Part 4. Intermediate, distal and col- lecting segments of the adult. Am. J. Anat. 130: 281-303.
ZAUGG, W. S., AND L. R. MCLAIN. 1976. Influence of water temperature on gill sodium, potassium-stimulated ATPase activity in juvenile coho salmon (Oi~corl~~nclrus kisurch). Comp. Biochem. Physiol. 54A : 419-421.


















