
Vector Ecology folder/journal/sovejournal74-2000/SOVE 1999, VO… · The Journal of Vector Ecology...
130
Journal of Vector Ecology Journal of the Society for Vector Ecology i1 Volume 24, No. 2 DECEMBER 1999 JVE 24( 2): 111- 232 ISSN 1081- 1710 Printed on Acid- Free Paper
-
Upload
vuongkhuong -
Category
Documents
-
view
222 -
download
0
Transcript of Vector Ecology folder/journal/sovejournal74-2000/SOVE 1999, VO… · The Journal of Vector Ecology...

Journal
of
Vector EcologyJournal of the Society for Vector Ecology
i1
Volume 24, No. 2 DECEMBER 1999JVE 24( 2): 111- 232 ISSN 1081- 1710
Printed on Acid-Free Paper

Journal of Vector Ecology
Volume 24- Number 2- December1999
Published by the Society for Vector Ecology
Marc J. Klowden, EditorDivision of Entomology
University of IdahoMoscow, ID 83844- 2339
Phone: ( 208) 885- 7546
Fax: ( 208) 885- 7760
E- mail: mklowden @uidaho.edu
EDITORIAL BOARD
M. S. Mulla, Chair( 2000) L. S. Mian( 2000)
University of California California State UniversityRiverside, CA, USA San Bernardino, CA, USA
N. Becker( 2001) R. S. Nasci ( 2000)
KABS Centers for Disease Control
Waldsee, Germany and Prevention
Ft. Collins, CO, USA
H. Briegel( 1999)
University of Zurich J. W. Beehler ( 2001)
Zurich, Switzerland Northwest Mosquito and
Vector Control District,
C. I. Dahl( 2001) Corona, CA , USA
University of UppsalaUppsala, Sweden E. D. Walker( 1999)
Michigan State UniversityR. S. Lane( 1999) East Lansing, MI, USAUniversity of CaliforniaBerkeley, CA, USA
The Journal of Vector Ecology is published biannually in June and December. Authors agree to transfer thecopyright for their article to the publisher when the article is accepted for publication. Authorization to photocopyarticles is granted by the Society for Vector Ecology provided the indicated fee is sent to the Copyright ClearanceCenter, Inc., 222 Rosewood Drive, Danvers, MA 01923, USA. Individuals may make single copies of articleswithout charge. Communications relating to editorial matters and manuscripts should be addressed to the Editor.Communications relating to galley proofs, reprints, subscriptions, SOVE membership, change of address, andother matters should be addressed to the Business Office.
Publications and Business Office: Society for Vector Ecology, 1966 Compton Avenue, Corona, CA 92881- 3318USA, (909) 340-9792; ( 909) 340- 2515 ( Fax); E-Mail: soveoffice @pe.net
Subscription Rates: Membership, including the Journal ofVector Ecology,$ 50.00, Student membership$ 25. 00,Institutional subscription $ 50.00, Sustaining membership $ 100.00

SOCIETY FOR VECTOR ECOLOGY
1999 BOARD OF DIRECTORS
OFFICERS
President David A. Dame
President-Elect John D. Edman
Vice-President Marc J. Klowden
Past- President Rex E. Thomas
Secretary- Treasurer Major S. Dhillon
REGIONAL DIRECTORS
Southwestern Minoo B. Madon
Northwestern Sammie Dickson
North Central Thomas R. Wilmot
South Central Cluff E. Hopla
Northeastern Wayne J. Crans
Southeastern Jonathan F. DayEuropean Romeo Bellini
SOVE Journal Editor Marc J. Klowden
SOVE Newsletter Editor Cluff E. Hopla

JOURNAL OF VECTOR ECOLOGY
Volume 24 DECEMBER, 1999 Number 2
CONTENTS
Board of Directors iiGuidelines for Contributors iiiEditorial Message v
Hilton B. Munns Memoriamxi
Submitted Papers
Effects of Partial Blood Engorgement and Pretest Carbohydrate Availability on the Repellency of Deet toAedes albopictus Rui-De Xue and Donald R. Barnard 111
A World Checklist of Genera, Subgenera, and Species of Ticks( Acari: Ixodida) Published from 1973- 1997James E. Keirans and Richard G. Robbins 115
Larval Habitats of Anopheline Mosquitoes in the Upper Orinoco, VenezuelaE. Rejmankova, Y. Rubio- Palis, and L. Villegas 130
Mosquito Control and Bacterial Flora in Water Enriched with Organic Matter and Treated with Bacillus
thuringiensis subsp. israelensis and Bacillus sphaericus FormulationsTram T. H. Nguyen, Tianyun Su, and Mir S. Mulla 138
Field Efficacy of Fipronil 3G, Lambda-cyhalothrin 10% CS, and Sumithion 50EC Against the DengueVector Aedes albopictus in Discarded Tires
S. Sulaiman, Z. A. Pawanchee, A. Wahab, J. Jamal, and A. R. Sohadi 154
Reproductive Biology of Lutzomyia shannoni ( Dyar) ( Diptera: Psychodidae) Under Experimental
Conditions
Estrella Cardenas, Cristina Ferro, Dario Corredor, Orlando Martinez, and Leonard E. Munstermann 158
The Fleas ( Siphonaptera) of South Carolina with an Assessment of Their Vectorial ImportanceLance A. Durden, William Wills, and Kerry L. Clark 171
Experimental Studies of Interactions Between Wild Turkeys and Black-Legged Ticks
Richard S. Ostfeld and David N. Lewis 182
Susceptibility of the Malaria Vector Anopheles culicifacies ( Diptera: Culicidae) to DDT, Dieldrin,Malathion, and Lambda-Cyhalothrin R. S. Sharma 187
Bacteria and Mosquito Abundance in Microcosms Enriched with Organic Matter and Treated with a
Bacillus thuringiensis subsp. israelensis FormulationTram T. H. Nguyen, Tianyun Su, and Mir S. Mulla 191
Effects of Neem Products Containing Azadirachtin on Blood Feeding, Fecundity, and Survivorship ofCulex tarsalis and Culex quinquefasciatus( Diptera: Culicidae) Tianyun Su and Mir S. Mulla 202
Compatibility of Bacillus thuringiensis serovar israelensis and Chemical Insecticides for the Control ofAedes Mosquitoes P. Seleena, H. L. Lee, and Y. F. Chiang 216
Ticks of South Carolina( Acari: Ixodoidea) .... D. C. Williams, W. Wills, L. A. Durden, and E. W. Gray 224
JOURNAL OF VECTOR ECOLOGY
Guidelines for Contributors
The Journal of Vector Ecology is an international journal published by the Society forVector Ecology. It is concerned with all aspects of the biology, ecology, and control of arthropodvectors and the interrelationships between the vectors and the disease agents they transmit. Thejournal publishes original research articles and research notes, as well as comprehensive reviews
of vector biology based on presentations at Society meetings. All papers are reviewed by at leasttwo referees who are qualified scientists and who recommend their suitability for publication.Acceptance of manuscripts is based on their scientific merit and is the final decision of the editor,
but these decisions may be appealed to the editorial board.Manuscripts intended for publication should be sent to Dr. Marc J. Klowden, Editor,
Division of Entomology, University of Idaho, Moscow, Idaho 83844- 2339, U. S. A.
mklowden @uidaho.edu>. Manuscripts must be double-spaced on a single side of bond paper
with 25 mm margins. An original and two clear copies are required. Submission of text on a 3-
1/ 2" computer diskette formatted in MS-DOS is encouraged. Any word processing format isacceptable, but please indicate the program that was used on the diskette label. Papers must be
organized under the following headings, each on a separate page, in order: title page, abstract,text, acknowledgments( if appropriate), references cited, tables, figure legends, and figures. The
title page should contain the names of all authors and their affiliations as well as the corresponding
author's mailing address, e- mail, and fax number. It should also include a keyword index
containing no more than five words that best describe the paper. Pages should be numbered
consecutively starting with the title page. References should conform to the style in recent
volumes. Illustrations that are submitted must be of high quality and remain legible afterreduction.
Page charges, which partially defray the cost of publication, are $ 35 per printed page.
SOVE members who are unable to pay page charges may apply for a limited number of waivers.Reprint charges are shown in the table below.
Pages 1- 4 5- 8 9- 12 13- 16 17- 20
50 copies 70.00 115. 00 160.00 205. 00 255.00
or less
Each add' l 30.00 48.00 66.00 84.00 102.00
50 copies
Same order
Communications relating to editorial matters and manuscripts should be addressed to theEditor. Communications concerning galley proofs reprints, subscriptions, SOVEmembership, and change of address should be addressed to the Business Office.
PUBLICATIONS AND BUSINESS OFFICE
Society for Vector Ecology1966 Compton Avenue
Corona, CA 92881- 3318
Phone: ( 909) 340- 9792; Fax: ( 909) 340-2515; E-Mail: soveoffice @pe.net

Journal of Vector EcologyVolume 24- Number 2- December1999
Published by the Society for Vector Ecology
Editorial Message
The editor would like to express his appreciation to the many reviewers who spent considerabletime evaluating manuscripts for publication in the Journal of Vector Ecology during the past two years.
Charles S. Apperson Roger Eritja Barry R. MillerRichard C. Axtell Woodbridge A. Foster Carl J. MitchellNorbert Becker J. Howard Frank Dennis MooreJeffrey Beehler Marilyn Geary Mir S. MullaJohn C. Beier Christopher Geden Roger S. NasciM.F. Bowen John E. Gimnig Dara NunnWilliam E. Bradshaw Paul R. Grimstad George F. O'MearaMichael Brown Robert Hancock Susan M. PalchickDavid A. Carlson Jerome A. Hogsette Sally L. PaulsonDave D. Chadee Cluff E. Hopla Michael J. PerichJames E. Cilek Murray B. Isman William K. ReisenCarlo Costantini Lawrence A. Lacey Thomas W. ScottDavid A. Dame Robert S. Lane Michael W. ServiceJonathan F. Day Cynthia Lord Daniel E. SonenshinePeter DeChant L.P. Lounibos David StillerGail Chambers DeSantis Farida Mahmood Willem TakkenTeun Dekker Thomas N. Mather Rex E. ThomasLance Durden Janet C. McAllister Edward D. WalkerStephen Doggett Roger W. Meola Thomas WaltonJohn D. Edman Richard W. Merritt James P. WebbSanford Eigenbrode Richard P. Meyer Tom R. WilmotBruce F. Eldridge Lal S. Mian Mark L. Wilson
Jocelyn G. Millar
ADVERTISING
Commercial advertising space is available in the Journal; full page ( blackand white) at $ 150.00 per issue, half page ( black and white) at $ 90.00 perissue. Inquiries may be addressed to Major S. Dhillon, Ph.D., SOVE
Business Office, 1966 Compton Avenue, Corona, CA 92881- 3318 USA,909) 340-9792; ( 909) 340- 2515 ( Fax); E-Mail: soveoffice @pe.net
The publisher reserves the right to approve or refuse any advertisement.The publisher is not responsible for any claims, litigations, or expensesresulting from the advertiser' s unauthorized use of any name, photograph,sketch, or words protected by registered trademark or copyright.

VOLUME 24 DECEMBER, 1999 NUMBER 2
IN MEMORIAM
HILTON B. MUNNS
1929- 1999
f
l*
d
4 1
Aar 4.
of
f
a
JI P 4 /
d
ti
Si{ f
tom
ff it
i i -}," e
t `1
i 1
y f° YY
At
iliZI4
4

NUMBER 2 DECEMBER, 1999 VOLUME 24
A unique first name. Munzy' s genuine friendship and his great affection forA unique last name. the SOVE that earned him lifelong friendships withMunns was popularly known as " MUNZY," a many of us here and abroad, plus the honor ofbeing one
unique nickname! of the most beloved members of the Society. MunzyIt seems that the common denominator is unique, was presented with the Distinguished Service Award
in eulogizing this remarkable human being, a kind, to the Society in 1992.compassionate gentleman, and a genuine friend. I first met Munzy sometime in 1980 or soon
Munzy was born on 26 February 1929, in thereafter, our friendship was instant. We traveled allFennimore, Wisconsin. On his final day, 9 September through the USA and Europe many times together. But1999, Munzy was once again in Fennimore. He was 70 it never really mattered how long one had knownyears old. His hometown paper, the Fennimore Times Munzy— whether it was decades, years, months, orcharacterized Munzy for living"... an exemplary life even a day or two, one often experienced the feelingwith that quiet dignity that identified him as a man that the friendship with him would be a long- lastingamong men." one.
Following a postwar stint in the army and college, Warm, gentle, kind, compassionate, genuine, and
Munzy began his career as a " cheese- maker" in most of all, giving— Munzy always gave, not becauseFennimore. He came to California in 1963, first working he felt he had to— he gave for the sake of giving, neverin the dairy section of the food processing chemicals really expecting anything in return. He made it a habitdepartment for Wyandotte Chemicals( now a subsidiary of usually being there to extend a helping hand to thoseof the giant BASF Chemical Company). He then that needed it. He acquired and fostered the friendshipworked forRalston— Purina until he formed Fennimore of many of us ever since he first became a member ofChemicals in 1971. As the hometown paper the SOVE in the early 1980's.characterized Munzy: " He loved Fennimore with a Although many sponsors from the private industrypassion and took advantage of every opportunity to graciously contributed, Munzy always " walked thatbring honor to his native home." And, until the very extra mile" to assure that the hosted socials(" attitude
end, many years after leaving Fennimore, his Fennimore adjustment hours") were enjoyable, and these became
friends always received calendars which proudly an integral part of the SOVE meetings. He left thisincluded the name of their town in his company name. legacy behind, therefore, we must strive to keep it
A good friend of Munzy' s, Dr. Maria Zgomba alive. Munzy was also a golf enthusiast. I am sure thatfrom far away Yugoslavia), expressed her, including his golfing buddies will surely " feel his presence" at
other colleagues' sentiments as follows: ". . It is the" 19th hole" whenever they happen to be there.unbearable to think that there won' t be any more Munzy is survived by his wife, Jessica; his threeof his vivid, charming personality to meet and enjoy daughters, Meridyth, Andrea, and Melanie and their
Munzy' s discreet, nonintrusion generosity wherever mother LuAnn; Jessica' s three children David, Glenn,
he appears. . ." We too, share the same sentiments. and LoriAnne; 12 grandchildren; and 4 sisters.
When his daughters eulogized their father, they Munzy is not amongst us anymore, but we willquoted Horace Mann, who said: ` Be ashamed to die always remember his warm and friendly smile, hisuntil you have won some great victory for humanity." genuine friendship, and now we must bid him fare-His family, as well as all of his friends share the same well.
feeling; Munzy passed on unashamed.Most of the sentiments expressed above, including Minoo B. Madon.
many more by all those who knew Munzy, were Greater Los Angeles Countymirrored in my hurriedly prepared eulogy I delivered Vector Control District
at the XIIth. Euro- SOVE Conference in Wageningen, 12545 Florence Avenue
Holland, on Saturday 11 September 1999. It was Santa Fe Springs, California 90760
xii

Journal of Vector Ecology 24(2): 111- 114
Effects of Partial Blood Engorgement and Pretest Carbohydrate
Availability on the Repellency of Deet to Aedes albopictus
Rui-De Xue and Donald R. Barnard
Center for Medical, Agricultural, and Veterinary EntomologyUSDA, ARS, P. 0. Box 14565, Gainesville, Florida 32604 U.S.A.
Received 21 September 1998; Accepted 16 April 1999
ABSTRACT: The pretest availability of 10% sucrose solution and/ or partial blood engorgement in Aedes
albopictus Skuse significantly influenced mosquito attack rates and the time of repellent protection inlaboratory bioassays. In 46 cm L x 38 cm W x 37 cm H cages used in USDA repellent tests, non-blood-fed and partially blood- fed mosquitoes attempted to bite at similar rates. In small cages( 5 cm dia. x 4 cmH), holding individual females, mean mosquito attack rates were reduced when females were partiallyblood fed, compared with those not blood fed. The protection period from bites by Ae. albopictus using25% ethanolic deet( N,Ndiethyl- 3- methylbenzamide) increased significantly in small and USDA standardcages when females had pretest access to sucrose solution, compared with females starved for 12 h. Partial
blood engorgement in mosquitoes affected repellent protection time in USDA standard test cages but not
in small cages.
Keyword Index: Mosquito, sucrose, carbohydrate, blood, feeding, repellents.
INTRODUCTION MATERIALS AND METHODS
Two pretest conditions that can affect mosquito Immature and adult Ae. albopictus (' F25) were
host- seeking behavior and repellency in repellent maintained in the laboratory at 27° C and 80% relative
bioassays are carbohydrate availability and partial blood humidity in a 14: 10 ( L:D) photoperiod. Larvae wereengorgement( Scott et al. 1993). In Aedes aegypti L., reared in groups of 200 using 30 cm long x 19 cm widerepellent efficacy is correlated with the pretest availability x 5 cm high trays containing 1000 ml of well water.ofsugar solution ( Khan et al. 1975); repellents are most Adults emerged in screened cages and were provided
effective against this species when the mosquitoes have continuous access to 10% sucrose/ water solution. Blood
access to sugar water before testing, and are least meals were obtained from restrained 5- 7 week-oldchicks.
effective when females have been starved. Mosquito responses to deet were evaluated in two
The effects of pretest sugar availability and partial different test arenas: ( 1) a screened cage( the" USDA
blood engorgement on repellent activity against Aedes standard" cage), 46 cm long x 38 cm wide x 37 cm highalbopictus Skuse are unknown. In the study presented ( volume: 64, 676 cm3),
containing 200 femalehere, we determined if repellent protection time against mosquitoes, and( 2) small cages ( 5 cm dia. x 4 cm H;
this species was affected by either factor when deet( N,N volume: 78. 4 cm3), each containing a single female anddiethyl- 3- methylbenzamide) was applied to human skin with fine mesh( 1. 7/ mm) cloth over the open end. The
and tested against caged populations of mosquitoes. latter test arena approximated the space and mosquito
Our findings will be used to standardize repellent density conditions defined by the above cited ASTMbioassays against Ae. albopictus and may be useful for standard; specifically, individual small cages were usedrevision of standard E951- 94 ( American Society for to eliminate the positional bias resulting from preferenceTesting and Materials 1994). This standard presumes by landing and probing mosquitoes for the outmostthe use of nulliparous mosquitoes but does not specify ports of the five feeding port, box- shaped apparatuscarbohydrate availability during the pretest period. specified in the standard.

112 Journal of Vector Ecology December, 1999
Mosquito Attack Rates and Deet Repellency in USDA Mosquito Attack Rates and Deet Repellency in SmallStandard Cages Cages
Four treatments( 10% sugar water, no sugar water, Four treatments( 10% sugar water, no sugar water,
partial blood meal, no blood meal) were used in this partial blood meal, no blood meal) were used in thisexperiment, one treatment per cage, with each treatment experiment with each treatment replicated three times.
replicated repeated three times. Each replicate required Small cages, as described above, were used to cage100 female mosquitoes, which were partially blood fed individual female mosquitoes.
using the following technique: the feathers on an adult One hundred mosquitoes were used in each replicate;chicken were clipped from one side of the abdomen and 50 of these were not blood fed and 50 were partiallyfrom adjacent areas on the wing. A small cage( 5 cm dia. blood fed immediately before testing. Cages werex 4 cm H), with the one open end covered by 1. 7- mm separated into four groups of 25 cages per group withmesh cloth, was used to hold a single female mosquito. each group placed into one of four plastic trays( 56 cmThe screened end of the small cage was placed against L x 43 cm W x 8 cm H) according to pretest sugarthe chicken' s skin while the chicken was restrained. availability and blood- feeding status of the female. AllOnce the mosquito began to feed, engorgement was trays were lined with paper towels. In two of the traysallowed until the mosquito gut appeared one- quarter to ( with 25 blood- fed females; 25 non- blood- fed females),one- third full of blood. Feeding was interrupted by the paper towels were soaked to runoff with a 10%
lifting the small cage containing the mosquito from the sucrose solution. Towels in the remaining two trays( 25chicken' s skin. The first group of 50 females partially blood- fed females; 25 non- blood-fed females) wereblood fed in this manner was transferred to a test cage soaked with water only.and provided cotton saturated with 10% sucrose solution Mosquito Attack Rates. To test mosquito attackfor 12 hours before testing. A second group of 50 rates, the screened end of each cage was individuallypartially blood- fed females was transferred to a test cage pressed against the skin on the forearm of a humanand provided cotton saturated with water only for 12 volunteer. If the mosquito landed and probed the skinhours before testing. The remaining 100 non- blood-fed surface, the observation was recorded as an attempted
mosquitoes were divided equally into the third and bite. If the mosquito did not probe the skin within the 1fourth cages; those in the third cage were provided min. test period, a non- bite was recorded.sucrose solution for 12 hours before testing and those in Repellent Test. One ml of25% ethanolic deet wasthe fourth cage were provided water only for 12 hours applied to the forearm skin ofa human volunteer betweenbefore testing. the wrist and the elbow( approximately 650 cm2). The
Mosquito Attack Rates. To determine mosquito screened end of each cage was held against the treatedattack rates, the arm of a human volunteer was presented skin for 3 min. and the mosquito observed for probing.to the mosquitoes in each cage for 1 min. Mosquitoes Observations were repeated at 30-min. intervals. Thethat probed the skin exposed in a 9. 8 x 4. 8 cm, 1. 7 mm repellency test for each tray ended when a cumulativemesh- covered window in a vinyl glove, that otherwise total of three female mosquitoes from both groups of theprotected the forearm and hand from mosquito bite, 25 cages in the tray had probed the skin. Protection timewere categorized as attempting to feed. The mean attack was calculated as above.
rate was calculated for each treatment as the average Design and Data Analysis. USDA standard cagepercentage of mosquitoes that probed the skin in three and small cage experiments each were made as a 2 x 2replicates. factorial using a split plot design ( Steel and Torrie
Repellent Test. One ml of 25% deet in ethanol 1980). Factor one was sugar availability( 10% sucrose
solution was applied to the forearm ofa human volunteer solution; water only), factor two was blood engorgementbetween the wrist and the elbow( the hand was protected status ( non blood fed, partially blood fed). Meanfrom mosquito bite by covering it with a latex glove). percentage attack rate and repellent protection timeMosquitoes inside each cage were allowed access to the were analyzed separately and by cage type using analysistreated forearm for 3 min. ( Xue et al. 1995) with of variance procedures( Gustafson, 1989).
observations repeated every 30 min. The test for a cagewas stopped when a cumulative total of three mosquitoes RESULTS
from the cage attempted to feed. Repellent protectiontime( hours) was calculated for each cage as that elapsed Mosquito Attack Rates and Deet Repellency in USDAbetween deet application and the end of the test. The Standard Cagesmean protection time from mosquito bite was calculated Mosquito Attack Rates. Carbohydrate availabilityfor each treatment as the average for three replicates. significantly influenced attack rates by Ae. albopictus

December, 1999 Journal of Vector Ecology 113
F18= 65. 19, P< 0. 0001)( TABLE 1) with mean rates Repellent Test. The protection time of deet againsthigher in the water only group compared with the Ae. albopictus was affected by sugar availability( F18=sucrose group. Interaction between sugar availability 73.51, P < 0.0001) ( TABLE 2) and was longest in
and blood engorgement status was not significant. mosquitoes with access to sugar, compared with those
Repellent Test. The protection time ofdeet against without access to sugar, regardless ofblood-engorgementAe. albopictus was affected by sugar availability( F1,8= status. Interaction between sugar availability and blood-55. 13, P< 0.001) and blood engorgement status( F1,8=
engorgement status was not significant.
6. 13, P< 0.05) ( TABLE 1). Mosquitoes with pretest
access to sucrose were repelled by deet longer than DISCUSSION
females provided water only, regardless of bloodengorgement status, whereas, repellent protection time If the attack rate in a population of mosquitoes usedwas longer against partially blooded females than against in a repellent bioassay is low, the protection time will benon-blood- fed females, regardless of sugar availability. overestimated( Kalmus and Hocking 1960). Thus, it is
There was no significant interaction between sugar important to know the hunger status of the mosquitoes
availability and blood-engorgement status. before a repellent test is made. From other studies( Xue
et al. 1995, Xue and Barnard 1996) we know that bloodMosquito Attack Rates and Deet Repellency in Small feeding in Ae. albopictus is affected by mosquito bodyTest Cages size and age, parity, and by the time of day when
Mosquito Attack Rates. In small cages( TABLE observations are made. In the present study, we have2), sugar availability ( F18 = 57. 31, P < 0.0001) and shown that attack rates in Ae. albopictus are affected byblood engorgement status ( F18= 111. 33, P< 0. 0001) the pretest availability ofcarbohydrates. In addition, weinfluenced mosquito attack rates. The rates were highest showed that partial blood feeding in mosquitoes affectsin sugar- starved females, regardless of blood- attack rates in small cage tests but not in tests with
engorgement status, and lowest in partially blood- fed USDA standard cages. One reason for this differencefemales, regardless of sugar availability. There was no may be the forced proximity of mosquitoes to the skinsignificant interaction between sugar availability and surface in small cages, which facilitates orientation to ablood-engorgement status. feeding site.
TABLE 1. Mean mosquito attack rates by, and repellent protection times against, partially blood- fed and non-blood- fed female Aedes albopictus provided water or sucrose solution for 12 hours before testing inUSDA cages.
Partially Blood-Fed Non-Blood-Fed
Water Sucrose Water Sucrose
Mosquito attack rate(%± SE) 25. 0± 3. 1 11. 0± 2. 3 24. 7± 1. 2 14. 0± 3. 5
Repellent protection time( hour± SE) 7. 0± 0.5 8. 5± 0.5 6. 2± 0.3 8. 2± 0.3
TABLE 2. Mean mosquito attack rates by, and repellent protection times against, partially blood fed and non-blood fed female Aedes albopictus provided water or sucrose solution for 12 hours before testing insmall cages.
Partially Blood-Fed Non-Blood-Fed
Water Sucrose Water Sucrose
Mosquito attack rate(%± SE) 60.0± 10. 1 23. 0± 7. 0 100.0± 0. 1 72. 0± 3. 5
Repellent protection time( hour± SE) 8. 3± 0.3 10.0± 0.2 7. 8± 0.3 9.7± 0.6

114 Journal of Vector Ecology December, 1999
Pretest sucrose availability affected the protection commercial mosquito repellent formulations on
time of deet against Ae. albopictus. Although a the skin. American Society for Testing andcomparison of repellent protection time according to Materials. E951- 94, P. 1- 6. Philadelphia, PA,6 pp.cage type was not an objective of this study, we did Davis, E. E. 1985. Insect repellents: concepts of theirnote that blood-engorgement status affected protection mode of action relative to potential sensorytime in USDA cages, but not small cages; and that mechanisms in mosquitoes( Diptera: Culicidae). J.protection time was longest in the small cages. These Med. Entomol. 22: 237- 243.disparate responses among cage types could be explained Gustafson, T. L. 1989. True Epistat Manual, 3rd ed.,on the basis ofneuronal inhibition in mosquitoes( Davis Richardson, TX, 229 pp.1985) in small cages resulting from the small space and Kalmus, H. and B. Hocking. 1960. Behavior of Aedespoor air exchange and the inability of the mosquito to mosquitoes in relation to blood- feeding andmove away from the deet treated skin surface. repellents. Ent. Exp. Appl. 3: 1- 26.
We conclude that sucrose availability to mosquitoes Khan, A. A., H. I. Maibach, and D. L. Skidmore. 1975.prior to a repellent test influences the period of repellent Insect repellents: Effect of mosquito and repellent-effectiveness. We also conclude that mosquitoes that related factors on protection time. J. Econ. Entomol.have access to sugar solution and to blood before a 68: 43- 45.
repellent test is made are repelled longer, by the same Scott, T. W., E. Chow, D. Strickman, P. Kittayapong, R.dose of deet, than mosquitoes receiving neither sugar A. Wirtz, L. H. Lorenz, and J. D. Edman. 1993.nor blood. Blood- feeding patterns of Aedes aegypti( Diptera:
The results of this study are important because they Culicidae) collected in a rural Thai village. J. Med.underscore the need to control pretest conditions for Entomol. 30: 922- 927.
mosquitoes ifthe resultsoflaboratory repellentbioassays Steel, R. G. D. and J. H. Torrie. 1980. Principles andare to be considered reliable. They also suggest the need Procedures of Statistics, 2nd ed. McGraw-Hill
to amend the ASTM standard with regard to carbohydrate Book Company, 633 pp.availability to mosquitoes before a repellent test is made Xue, R. D. and D.R. Barnard. 1996. Human host aviditybecause this factor affects mosquito responses to a in Aedes albopictus: influence of mosquito bodyrepellent. size, age, parity, and time of day. J. Am. Mosq.
Contr. Assoc. 12: 58- 63.
REFERENCES CITED Xue, R. D., D. R. Barnard, and C. E. Schreck. 1995.
Influence of body size and age in Aedes albopictusAmerican Society for Testing and Materials. 1994. on human host attack rates and the repellency of
Standard test methods for laboratory testing of non- deet. J. Am. Mosq. Contr. Assoc. 11: 50- 53.

Journal of Vector Ecology 24( 2): 115- 129
A World Checklist of Genera, Subgenera, and Species of
Ticks (Acari: Ixodida) Published from 1973- 1997
James E. Keirans' and Richard G. Robbins2
U.S. National Tick Collection, Institute ofArthropodology and Parasitology,Georgia Southern University, Statesboro, Georgia 30460-8056, USA.
Armed Forces Pest Management Board, Walter Reed Army Medical Center,Washington, D.C. 20307-5001, USA.
Received 20 November 1998; Accepted 8 March 1999
ABSTRACT: Researchers on ticks and tickborne diseases have been extremely fortunate in having at theirfingertips the tick bibliographies produced by Harry Hoogstraal and his coworkers at the U.S. Naval facilityat Cairo, Egypt, and by Mildred Doss and her colleagues at the U.S. Department of Agriculture laboratoryat Beltsville, Maryland, USA. The Doss checklist of tick families, genera, species, and subspecies is now
25 years out of date, and the following checklist of one new genus, nine new subgenera, and 110 new speciesof Ixodida brings together the nomenclature on ticks produced during the last quarter century.
Keyword Index: Tick taxa( Ixodida) since 1973.
INTRODUCTION referenced their entries to two other acarological
bibliographies: the Index-Catalogue itself ( Authors,
During the 1970s, a team of bibliographers, headed Parts 1- 18, and Supplements 1- 17) and the Bibliographyby the late Mildred Doss( September 2, 1903- December of Ticks and Tickborne Diseases by the late Harry21, 1993), Animal Parasitology Institute ( now Hoogstraal ( 1917- 1986). In their checklist, the Doss
Biosystematics and National Parasite Collection Unit team cite most references to new tick taxa published upBNPCU), Livestock and Poultry Sciences Institute), to 1972( and some subsequently), but through usage we
Agricultural Research Service, U.S. Department of have determined that the year 1973 marks the point atAgriculture, and by George Anastos, Department of which the Doss list ceases to be an all- inclusive work.
Zoology, University of Maryland, compiled a series of With the passage of a quarter century, we believe thatsubject indices on the genera and species of ticks, their the time has come to augment our predecessors'hosts, and distributions. These works were issued as compilation.
Special Publications of the USDA' s Index-Catalogue of Since 1973, descriptions of 1 new tick genus, 9 newMedical and Veterinary Zoology. After collating subgenera, and 110 new species have appeared in 96
references to tick genera and species( Part I, 3 volumes, scientific papers, worldwide. Of the new species, 34
January 1974) and tick hosts ( Part II, 3 volumes, July ( 31%) are argasids, described in 5 genera, while 761974), Doss and colleagues produced a checklist of tick ( 69%) are ixodids, described in 10 genera. Ornithodorosfamilies, genera, species, and subspecies( Part III, May antiquus is known only from Dominican amber; the1977), based largely on the contents of the preceding remaining 109 new species represent all zoogeographicparts. Their fourth and final installment on tick regions, as follows: Afrotropical 31, Australian 10,distribution appeared in 1978. Nearctic 4, Neotropical 21, Oriental 15, and Palearctic
Users of the" Doss bibliographies" generally agree 28. In our checklist, tick taxa and hosts are listedthat they are invaluable. Despite the enormity of their alphabetically, without regard to phylogenetic propin-task, the USDA-University ofMaryland team succeeded quity. However, depositories appear in the sequencein listing by category much of the world literature on given in the original descriptions, using the followingticks and tickborne diseases. As well, they cross- acronyms for collections cited more than once:

116 Journal of Vector Ecology December, 1999
ANIC, CSIRO: Australian National Insect Argasidae
Collection, Commonwealth Scientific and
Industrial Research Organisation, Canberra.
BMNH: The Natural History Museum, London; Alectorobius camicasi Sylla, Cornet and Marchand,
formerly British Museum( Natural History). 1997. Acarologia 38: 239- 254, figs. 1- 19,
CASP: Institute of Parasitology, Czech( formerly tabs. 1- 2.
Czechoslovak) Academy of Sciences, HOSTS: Rousettus aegyptiacus ( E. Geoffroy), R.Prague. angolensis( Bocage).
FMNH: Field Museum of Natural History, DISTRIBUTION: Senegal, Bandia( 14° 35' N, 17°
Chicago, USA. 01' W), Saboya ( 13° 36' N, 16° 26' W), and
FVZ: Department of Parasitology, Veterinary Ebarak, Salemata( 12° 35' N, 12° 50' W).
Faculty at Zaragoza, Spain. DEPOSITORIES: USNTC; IFAN, Dakar, Senegal;
IES: Zoological Collection of the Institute of Museum national d' Histoire naturelle, Paris;
Ecology and Systematics, Havana, Cuba. authors' collections.
IFAN: L' Institut Fondamental d' Afrique Noire. NOTE: The original description cited the host
IICT: Centro de Zoologia do Instituto de subspecies Rousettus aegyptiacus occidentalis
Investigacao Cientffica Tropical, Lisbon, and R. angolensis smithi. According to WilsonPortugal. and Reeder, eds. 1993. Mammal Species ofthe
IZAC: Institute de Zoologia, Academia de World, 2nd ed., Smithsonian Institution Press,
Ciencias de Cuba, Havana. Washington, xviii+ 1206 pp., the subspecificIZAS: Institute of Zoology, Academia Sinica, epithets occidentalis and smithi are synonyms
Beijing, People' s Republic of China. of R. aegyptiacus and R. angolensis,
MBB: Museum Bogoriense, Bogor, Java, respectively.
Indonesia.
MBR: Museo Argentino de Ciencias Naturales Antricola armasi de la Cruz and Estrada-Pena, 1995.
Bernardino Rivadavia," Buenos Aires. Acarologia 36: 277- 286, figs. 14- 19.
MNHM: Museum of Natural History, Maputo, HOST: Collected on bat guano.
Mozambique. DISTRIBUTION: Cuba, Pinar del Rio Province,
NAMRU-3: United States Naval Medical Guanahacacibes, Cueva de la Ventana.
Research Unit Number Three, Cairo, Egypt. DEPOSITORIES: USNTC; FVZ; BMNH; IES.
NTMD: Northern Territory Museum, Darwin,Australia. Antricola centralis de la Cruz and Estrada- Pena, 1995.
ORSTOM: Office de la Recherche Scientifique Acarologia 36: 277- 286, figs. 1- 7.
et Technique Outre-Mer. HOST: Collected in a bat cave.
OVI: Veterinary Research Institute, DISTRIBUTION: Cuba, Las Villas Province,
Onderstepoort, South Africa. Remedios, Buenaventura, Cueva del Maja.
USNTC: United States National Tick Collection, DEPOSITORIES: USNTC; IES; BMNH; FVZ.
Statesboro, Georgia.
VRLH: Veterinary Research Laboratory, Harare, Antricola cernyi de la Cruz, 1978. Poeyana( 184): 1-
Zimbabwe. 17, fig. 1 c.WAMP: Western Australia Museum, Perth. HOST: Collected in a bat cave.
ZISP: Zoological Institute, St. Petersburg, DISTRIBUTION: Cuba, Las Villas, Rodas, Cueva
Russia. de Castellanos.
DEPOSITORY: IZAC.
We take pleasure in dedicating this update to the Antricola gran"asi de la Cruz, 1973. Simposium XXX
memory of Mildred Doss, whose conscientiousness and Aniversario de la Sociedad Espeleologica decharm greatly facilitated the always arduous task of Cuba. Serie Espeleologica y Carsologica( 44):bibliography. Our thanks also to J. Ralph Lichtenfels, 3- 13, fig. 1, fig. 2a, d, g, j, figs. 3, 4.Research Leader and Supervisory Zoologist, BNPCU, HOST: Collected in a bat cave.
who provided important background information on DISTRIBUTION: Cuba, Las Villas, Yaguajay,Ms. Doss and her career, and to Renjie Hu for obtaining Punta Judas, Cueva del Abono.
and translating various Chinese publications. DEPOSITORIES: IZAC; CASP.

December, 1999 Journal of Vector Ecology 117
Antricola habanensis de la Cruz, 1976. Poeyana( 151): Ukazzi Hill, and Rift Valley Province, Kajiado.1- 8, figs. la, d. DEPOSITORIES: USNTC; OVI; South African
HOST: Collected in a bat cave. Institute for Medical Research, Johannesburg,DISTRIBUTION: Cuba, La Habana Province, South Africa; BMNH; ZISP.
Catalina de Gaines, Cueva del Mudo.
DEPOSITORY: IZAC. Argas assimilis Teng and Song, 1983. Acta
Zootaxonomica Sinica 8: 153- 156, figs. 1- 5.Antricola hummelincki de la Cruz and Estrada-Pena, In English: NAMRU-3- T1774).
1995. Acarologia 36: 277- 286, figs. 8- 13. HOST: Hirundo daurica japonica Temminck andHOST: Mormoops megalophylla( Peters). Schlegel.DISTRIBUTION: Venezuela, Curacao, Hato, DISTRIBUTION: People' s Republic of China,
Cueva di Rato. Jiangxi Province, Tonggu County.DEPOSITORY: USNTC. DEPOSITORIES: IZAS; Department of
Parasitology, Jiangxi Medical College.Antricola martelorum de la Cruz, 1978. Poeyana
184): 1- 17, fig. lf. Argas beijingensis Teng, 1983. Acta ZootaxonomicaHOST: Collected in a bat cave. Sinica 8: 255- 261, figs. 1- 9. ( In English:DISTRIBUTION: Cuba, La Habana, Santa Cruz NAMRU-3- T1775).
del Norte, Finca Galera, Cueva de los HOST: Collected from roosts of Columba liviaMurcielagos. Gmelin.
DEPOSITORY: IZAC. DISTRIBUTION: People' s Republic of China,
Shijing mountain region of Beijing.Antricola naomiae de la Cruz, 1978. Poeyana( 184): 1- DEPOSITORY: Not stated but probably IZAS.
17, fig. ld.HOST: Collected in a bat cave. Argas dalei Clifford, Keirans, Hoogstraal, and Corwin,DISTRIBUTION: Cuba, Matanzas, Camarioca, 1976. Annals ofthe Entomological Society of
Cueva de Santa Catalina. America 69: 917- 925, figs. 1- 16.DEPOSITORY: IZAC. HOST: Collected from roosts of Speotyto
cunicularia nanodes Berlepsch and Stolzmann.Antricola occidentalis de la Cruz, 1978. Poeyana DISTRIBUTION: Peru, Lima,LaMolina( 12° 05' S,
184): 1- 17, fig. la. 76° 57' W).
HOST: Collected in a bat cave. DEPOSITORIES: USNTC; Museo de Entomo-DISTRIBUTION: Cuba, Pinar del Rio, San Andres logia, Universidad Nacional Agraria, Lima,
de Caiguanabo, Galalon, Cueva de los Majaes. Peru.
DEPOSITORY: IZAC.
Argas dewae Kaiser and Hoogstraal, 1974. Annals ofAntricola siboneyi de la Cruz and Estrada-Pena, 1995. the Entomological Society ofAmerica 67: 231-
Acarologia 36: 277- 286, figs. 20- 26. 237, figs. 1- 28.HOST: Collected on bat guano. HOSTS: Chalinolobus dwyeri Ryan, C. gouldiiDISTRIBUTION: Cuba, Santiago de Cuba, Gray, Eptesicus pumilus Gray, Nyctophilus
Siboney, Cueva de los Majaes. geoffroyi Leach, Pipistrellus tasmaniensisDEPOSITORIES: USNTC; IES; FVZ; BMNH. Gould), Rhinolophus megaphyllus Gray.
DISTRIBUTION: Australia, New South Wales,Argasafricolumbae Hoogstraal, Kaiser, Walker, Ledger, Victoria, Tasmania.
Converse, and Rice, 1975. Journal ofMedical DEPOSITORIES: ANIC, CSIRO; Queen Victoria
Entomology 12: 194- 201, figs. 1- 23. Museum, Launceston, Tasmania; USNTC.HOSTS: Collected from nests of Columba guinea
phaeonota Gray, Geronticus calvus( Boddaert), Argasfalco Kaiser and Hoogstraal, 1974. Annals oftheand Ptyonoprogne fuligula rufigula ( Fischer Entomological Society of America 67: 5- 10,and Reichenow). figs. 1- 21.
DISTRIBUTION: Republic of South Africa, HOST: Falco cenchroides Vigors and Horsfield.Transvaal ( now Gauteng) Province, Pretoria DISTRIBUTION: Australia, Western Australia,and Pietersburg; Kenya, Eastern Province, Cue Shire, Remington Breakaway( 26° 28' S,

118 Journal of Vector Ecology December, 1999
117° 35' E). DEPOSITORY: USNTC.
DEPOSITORIES: ANIC, CSIRO; USNTC;
BMNH. Argas polonicus Siuda, Hoogstraal, Clifford, and
Wassef, 1979. Journal of Parasitology 65:Argas gilcolladoiEstrada-Pena, Lucientes, and Sanchez, 170- 181, figs. 1- 30.
1987. Journal of Parasitology 73: 824- 828, HOST: Columba livia Gmelin.
figs. 1- 7. DISTRIBUTION: Poland, Krakow( 50° 03' N, 19°
HOST: Gyps fulvus L. 58' E).
DISTRIBUTION: Spain, Zaragoza Province, 15 DEPOSITORIES: USNTC; Polish Academy ofkm NNW of Tabuenca( 41° 45' N, 01° 32' W). Sciences, Institute of Systematic and
DEPOSITORIES: Unidad de Parasitologia, Experimental Zoology, Krakow; DepartmentFacultad de Veterinaria, Zaragoza, Spain; of Animal Morphology, Institute of Biology,USNTC. Adam Mickiewicz University, Poznan; senior
author' s collection; ZISP; BMNH; FMNH.
Argas lowryae Kaiser and Hoogstraal, 1975. Annals ofthe Entomological Society ofAmerica 68: 585- Argas ricei Hoogstraal, Kaiser, Clifford, and Keirans,
590, figs. 1- 21, tab. 1. 1975. Annals of the Entomological Society ofHOST: Falco cenchroides Vigors and Horsfield. America 68: 873- 881, figs. 1- 21.
DISTRIBUTION: Australia, Western Australia, HOSTS: Cathartes aura septentrionalis Wied,
Nullarbor Plain, Horseshoe Cave ( 31° 39' S, Coragyps a. atratus( Bechstein).
127° 26' E). DISTRIBUTION: U.S. A., Texas, Zavala County,DEPOSITORIES: ANIC, CSIRO; BMNH; 3. 5 mi.( 5. 6 km) E of La Pryor( 28° 55' N, 99°
USNTC. 47' W) on Highway 57. Also from Bastrop,Bexar, Kimble, Menard, and Travis Counties.
Argas macrodermae Hoogstraal, Moorhouse, Wolf, DEPOSITORIES: USNTC; Entomologyand Wassef, 1977. Annals ofthe Entomological Department, Texas A& M University, CollegeSociety ofAmerica 70: 861- 870, figs. 1- 27. Station; FMNH.
HOST: Macroderma gigas Dobson.
DISTRIBUTION: Australia, Queensland, Argas sinensis Jeu and Zhu, 1982. Acta Entomologica
Limestone Ridge, Johannsen' s Cave( 23° 10' S, Sinica 25: 328- 331, figs. 1- 6.
150° 27' E) near Rockhampton. HOST: Pipistrellus abramus( Temminck)[ now P.
DEPOSITORIES: Queensland Museum, Brisbane; javanicus( Gray)].Department of Parasitology, University of DISTRIBUTION: People' s Republic of China,
Queensland, Brisbane; BMNH; USNTC. Sichuan Province.
DEPOSITORY: Department of Parasitology,Argas monolakensis Schwan, Corwin, and Brown, Chongqing Medical College, Chongqing,
1992. Journal ofMedical Entomology 29: 78- Sichuan.
97, figs. 1- 36, 41, 43, tabs. 1- 10.
HOST: Collected under and around nests of Larus Carios hadiae Klompen, Keirans, and Durden, 1995.
californicus Lawrence. Acarologia 36: 25- 40, fig. 7.DISTRIBUTION: U. S. A., California, Mono HOSTS: Cynopterusbrachyotis( Miiller),Dobsonia
County, islands in Mono Lake. viridis ( Heude), Rousettus amplexicaudatus
DEPOSITORY: USNTC. E. Geoffroy).DISTRIBUTION: Indonesia, Halmahera Island,
Argas moreli Keirans, Hoogstraal, and Clifford, 1979. Jailolo District, Kampung Pasir Putih; LombokJournal ofMedical Entomology 15: 246- 252, Island, Bilekedit; Java.
figs. 1- 11. DEPOSITORY: USNTC.
HOSTS: Unknown; found on the wall of a house
and from unspecified locales. Wild birds and Carios multisetosus Klompen, Keirans, and Durden,
domestic chickens are probable hosts. 1995. Acarologia 36: 25- 40, fig. 6.DISTRIBUTION: Peru, Junin Province, Quebrada HOSTS: Dobsonia moluccensis ( Quoy and
de Gagaracca( 10° 57' S, 76° 01' W). Also from Gaimard), D. viridis( Heude), Dobsonia sp.Arequipa Province. DISTRIBUTION: Papua New Guinea, Madang

December, 1999 Journal of Vector Ecology 119
District, 15 km N of Madang; Indonesia, Irian Agraria, La Molina, Lima, Peru; Florida StateJaya, Owi Island, Biak Island, Sarwon and Collection of Arthropods, Bureau ofSorido; Indonesia, Seram Island, Manusela Entomology, Gainesville, Florida, USA.National Park; Indonesia, Halmahera Island,Jailolo District, Kampung Pasir Putih. Ornithodoros vansomereni Keirans, Hoogstraal, and
DEPOSITORY: USNTC. Clifford, 1977. Annals of the EntomologicalSociety ofAmerica 70: 221- 228, figs. 1- 14.
Cariospapuensis Klompen, Keirans, and Durden, 1995. HOSTS: Collected in nests of Hirundo abyssinicaAcarologia 36: 25- 40, figs. 1- 5. unitatis Sclater and Mackworth-Praed, H.
HOSTS: Dobsonia moluccensis ( Quoy and daurica emini Reichenow, and MyrmecocichlaGaimard), D. viridis( Heude). cinnamomeiventris subrufipennis Reichenow.
DISTRIBUTION: Indonesia, Halmahera Island, DISTRIBUTION: Kenya,Eastern Province, incave,Jailolo District, Kampung Pasir Putih; Papua Ngomeni Rock ( 04° 08' S, 38° 35' E), and inNew Guinea, Central District, Kairuku, Kukuba cave, peak of Ukazzi Hill( 00° 50' S, 38° 35' E).Cave. DEPOSITORIES: USNTC; Kenya National
DEPOSITORY: USNTC. Museum, Nairobi, Kenya.
Ornithodoros antiquus Poinar, 1995. Experientia 51: Urnithodorosyunkeri Keirans, Clifford, and Hoogstraal,384- 387, figs. 1- 5. 1984. Journal of Medical Entomology 21:
HOST: Found in two pieces of amber. 344- 350, figs. 1- 18.
DISTRIBUTION: Dominican Republic, Cordillera HOST: Collected in marine bird nesting site,Septentrional mountain range, La Toca mine. probably of Sula nebouxii Milne-Edwards.
DEPOSITORY: In the private collection of Jim DISTRIBUTION: Ecuador, Galapagos Islands,Work, Ashland, Oregon, U.S. A. Isabella ( Albemarle) Island, Punta Vicente
Roca( 00° 03' S, 91° 33' W). Also on the islandsOrnithodoros collocaliae Hoogstraal, Kadarsan, Kaiser, ofCulpepper( Darwin), Plaza Sur, Santa Cruz,
and Van Peenen, 1974. Annals of the Espanola, Daphne Major, Fernandina, andEntomological Society of America 67: 224- Seymour Norte.
230, figs. 1- 23. DEPOSITORIES: USNTC; BMNH.
HOST: Collected in and near nests of Collocalia NOTE: Also on and around nesting areas ofesculenta linchi Horsfield and Moore. Diomedeairrorata Salvin, Nannopterum( now
DISTRIBUTION: Indonesia, East Java, Baluran, more commonly placed in the genusWonoredjo (07° 56' S, 114° 22' E), and West Phalacrocorax) harrisi Rothschild,SpheniscusJava, Palabuhanratu. mendiculus Sundevall, Sterna lunata Peale,
DEPOSITORIES: MBB; USNTC; BMNH. and Sula dactylatra Lesson. Collections fromAmblyrhynchus cristatus Bell and Zalophus
Ornithodoros cyclurae De la Cruz, 1984. Poeyana wollebaeki Sivertsen ( now Z. californianus227): 1- 6, figs. 1- 2d. Lesson)) are probably accidental.
HOST: Collected in nasal cavity of Cyclura nubilaGray). Proknekalia Keirans, Hoogstraal, and Clifford, 1977.
DISTRIBUTION: Cuba, Granma Province, Cabo Annals ofthe Entomological Society ofAmericaCruz. 70: 221- 228. [ Subgenus of Ornithodoros.
DEPOSITORY: IZAC. Type- O. ( P.) vansomereni]
Ornithodoros spheniscus Hoogstraal, Wassef, Hays,
and Keirans, 1985. Journal of Parasitology Ixodidae
71: 635- 644, figs. 1- 18, tab. 1.
HOST: Spheniscus humboldti Meyen. Africaniella Santos Dias, 1974. Revista de CienciasDISTRIBUTION: Peru, Ica, Punta San Juan ( 15° Veterindrias Serie A, 7: 73- 107. [ Subgenus of
21' S, 75° 11' W), Punta Blanca( 05° 49' S, 81° Aponomma. Type - A. ( A.) transversale
05' W) and Punta San Fernando( 15° 08' S, 70° Lucas)].
21' W).
DEPOSITORIES: USNTC; Universidad Nacional Amblyommaarianae Keirans and Garris, 1986. Journal

120 Journal of Vector Ecology December, 1999
ofMedical Entomology 23: 622- 625, figs. 1- 8. Adelaide, Australia; USNTC.
HOST: Alsophis portoricensis Reinhardt and NOTE: Coordinates for several other collection
Liitken. localities are detailed in Table 1 of the original
DISTRIBUTION: Puerto Rico, Barrio Puynado article.
Adentro, Municipio of Vega Baja.
DEPOSITORY: USNTC. Anomalohimalaya cricetuli Teng and Huang, 1981.NOTE: A junior synonym of Amblyomma Acta Entomologica Sinica 24: 99- 102, figs. 1-
quadricavum ( Schulze, 1941) - see Keirans 15. ( In Chinese; English summary).and Klompen. 1996. Proceedings of the HOST: Cricetulus migratorius( Pallas).
Entomological Society ofWashington 98: 164- DISTRIBUTION: People' s Republic of China,
165. Xinjiang Uygur Autonomous Region, HashiDistrict.
Amblyommaglauerti Keirans, King, and Sharrad, 1994. DEPOSITORY: IZAS.
Journal ofMedical Entomology 31: 132- 147,figs. 7- 12, tab. 2. Anomalohimalaya lotozkyi Filippova and Panova, 1978.
HOSTS: Varanus glauerti Mertens, V. glebopalma Parazitologiya 12: 391- 399, figs. 9- 28. ( In
Mitchell. Russian; English summary).DISTRIBUTION: Australia, Western Australia, HOST: Alticola argentatus( Severtzov).
Buccaneer Archipelago, Lachlan Island ( 16° DISTRIBUTION: Tajikistan, Peter the First Ridge.
37' S, 123° 31' E). DEPOSITORY: ZISP.
DEPOSITORIES: WAMP; NTMD; USNTC;
BMNH. Aponomma glebopalma Keirans, King, and Shan-ad,NOTE: Coordinates for numerous other localities 1994. Journal of Medical Entomology 31:
are detailed in Table 2 of the original article. 132- 147, figs. 1- 6, tab. 1.
HOSTS: Varanus glauerti Mertens, V.glebopalma
Amblyomma hainanense Teng, 1981. Acta Mitchell.
Zootaxonomica Sinica 6: 399- 401, figs. 1- 6. DISTRIBUTION: Australia, Western Australia,
In English: NAMRU-3 - T1570). Prince Regent River Reserve( 15° 32' S, 125°
HOST: Undetermined species of snake. 19' E).
DISTRIBUTION: People' s Republic of China, DEPOSITORIES: WAMP; NTMD; USNTC;
Guangdong Province, and on Hainan Island. BMNH.
DEPOSITORY: IZAS. NOTE: Coordinates for numerous other localities
NOTE: Hainan Island is also a Province of the are detailed in Table 1 of the original article.
P.R.C.
Aponomma inopinatum Santos Dias, 1989. Boletim da
Amblyomma pseudoparvum Guglielmone, Mangold, Sociedade Portuguesa de Entomologia 4: 29-
and Keirans, 1990. Acarologia 31: 143- 159, 48, figs. 1- 3a.
figs. 15- 29. HOST: Varanus exanthematicus( Bosc).
HOSTS: Dolichotis salinicola Burmeister. Also DISTRIBUTION: Democratic Republic ofCongo,
found on cattle. Monga.
DISTRIBUTION: Argentina, Salta Province, DEPOSITORY: Musee Royal de l' Afrique
Department ofRivadavia,Rivadavia( 24° 11' S, Centrale, Tervuren, Belgium.
62° 53' W). Also in Department of Anta.
DEPOSITORIES: USNTC; MBR. Aponommaorlovi Kolonin, 1992. Folia Parasitologica
39: 93- 94, fig. 1.Amblyomma vikirri Keirans, Bull, and Duffield, 1996. HOST: Python molurus bivittatus Kuhl.
Systematic Parasitology 34: 1- 9, figs. 1- 18, DISTRIBUTION: Vietnam, Shonla( 21° 20' N, 103°
tabs. 1- 2. 50' E).
HOST: Egernia stokesii( Gray). DEPOSITORY: ZISP.
DISTRIBUTION: Australia, South Australia,
Flinders Ranges near Hawker, Warru- Asiacentor Filippova and Panova, 1974.
warldunha Range( 31° 54' S, 138° 25' E). Entomologicheskoe Obozrenie 53: 470- 476.
DEPOSITORIES: South Australia Museum, In English: NAMRU-3- T1083). [ Subgenus

December, 1999 Journal of Vector Ecology 121
of Dermacentor; type - D. ( A.) pavlovskyi H. (G.) calvus Nuttall and Warburton].Olenev].
Haemaphysalis anomaloceraea Teng, 1984. ActaBoophilus florae Santos Dias, 1987. Garcia de Orta, Zootaxonomica Sinica 9: 37-40, figs. 1- 12. ( In
Serie de Zoologia 14: 17- 26, figs. 1- 4. English: NAMRU-3 - TI776).HOST: Aepyceros melampus( Lichtenstein). HOST: Not stated, probably collected on vegetation.DISTRIBUTION: Mozambique, Gaza Province, DISTRIBUTION: People' s Republic of China,
Govuro District, Parque Nacional do Zinave. Yunnan Province, wilderness of LushuiDEPOSITORY: IICT; BMNH; Museum d' Histoire County.
Naturelle de Paris; USNTC. DEPOSITORY: IZAS.
NOTE: In all probability a junior synonym ofBoophilus decoloratus( Koch). Haemaphysalis bacthaensis Phan 1977. Va bet va con
trung ky sinh o Vietnam. Tap 1. Ve( Ixodoidea).Boophilus scheepersi Santos Dias, 1989. Garcia de Mo to va phan Iaoi. ( In Vietnamese).
Orta, Serie de Zoologia 15: 41- 44, figs. 1- 4. HOST: Chloropsis cochinchinensis( Gmelin).HOST: Petrodromus tetradactylus beirae Roberts DISTRIBUTION: Vietnam, Bacthai.
now P. tetradactylus Peters]. DEPOSITORY: Unknown.DISTRIBUTION: Mozambique, Sofala Province, NOTE: We have been unable to acquire the
Lagoa Ura. publication by Phan. Kolonin( 1992) In: V. E.DEPOSITORIES: IICT; BMNH; OVI. Sokolov, ed. Zoological Researches inNOTE: In all probability a junior synonym of Vietnam, NaukaPublishers, Moscow, considers
Boophilus decoloratus( Koch). Haemaphysalis bacthaensis to be a junior
synonym of H. ornithophila Hoogstraal andBothriocroton Keirans, King, and Sharrad, 1994. Kohls.
Journal ofMedical Entomology 31: 132- 147.Subgenus of Aponomma; type - A. ( B.) Haemaphysalis danieli Cerny and Hoogstraal, 1977.
glebopalma Keirans, King and Sharrad]. Journal of Parasitology 63: 567- 574, figs. 1-27.
Dermacentonomma Santos Dias, 1978. Ciencias HOSTS: Holotype 9 from vegetation; tentativelyBiologica Biologia Molecular e Celular 4: 49- associated nymphs and larvae from Alticola52. [ New genus for Aponomma bourreti roylei Gray, Apodemus flavicollis( Melchior),Toumanoff]. Cricetulus migratorius ( Pallas), Marmota
caudata( Geoffroy), Ochotona roylei( Ogilby).Dermacentor montanus Filippova and Panova, 1974. DISTRIBUTION: Pakistan, Swat, Tirich-Mir
Entomologicheskoe Obozrenie 53: 470-476, Valley( 36° 15' N, 71° 51' E). Collections alsofigs. 1- 5. ( In English: NAMRU-3- T1083). from Wakhan area of Badakhashan Province,
HOST: Ochotona rutila( Severtzov). Afghanistan.
DISTRIBUTION: Tajikistan, southern slopes of DEPOSITORIES: CASP; USNTC.
Peter the First Ridge near the middle course ofthe Obikhingou River. Haemaphysalis demidovae Emel' yanova, 1978. In: O.
DEPOSITORY: ZISP. M. Kozhova, ed. Natural conditions and
resources ofPrikhubsugul( Mongolian People' sDermacentor ushakovae Filippova and Panova, 1987. Republic). Proceedings of Soviet-Mongolian
Parazitologiya 21: 450-458, pls. 1- 5, tabs. 1- 2. Complex Khubsugul Expedition, Ministry ofHOST: Collected on vegetation. Higher and Special Secondary Education ofDISTRIBUTION: Kazakstan, valley of the Chilik the A. A. Janova State University, Irkutsk,
River. Also from Kyrgyzstan and Turk- RSFSR ( 6): 162- 171, pl. 2, figs. 1- 10; pl. 3,menistan. figs. 1- 9.( In Russian). ( In English: NAMRU-
DEPOSITORY: ZISP. 3- T1738).
HOSTS: Citellus [ now Spermophilus] dauricusGarnhamphysalis Hoogstraal and Wassef, 1981. Society Brandt. Also on Ochotona pallasi ( Gray),
of Protozoologists ( Special Publication 1): Pamir high-mountain vole," and " Siberian117- 124. [ Subgenus ofHaemaphysalis; type- souslik."

122 Journal of Vector Ecology December, 1999
DISTRIBUTION: Mongolia, southern Khangai, Temminck, are hosts for this tick species.
Khure-Marl. Also in the Tarbagatai and Taishiri
regions. Haemaphysalis menglaensis Pang, Chen, and Xiang,DEPOSITORY: Not stated. 1982. Zoological Research 3( Suppl.): 45- 51,
figs. 1- 13.
Haemaphysalis filippovae Bolotin, 1979. HOST: Cervus sp.Zoologicheskiy Zhurnal 58: 267- 269, figs. 1- DISTRIBUTION: People' s Republic of
5. ( In Russian; English summary).( In English: China,Yunnan Province, Meng La.NAMRU-3- T1438). DEPOSITORY: Institute of Microbiology and
HOST: Not stated. Epidemiology, Academy of Military MedicalDISTRIBUTION: Russia, Primor' ye, Nadezh- Science, Beijing.
dinsky region, on the northern slope of thelower Anan' evka River valley( right tributary Haemaphysalis moschisuga Teng, 1980. Acta
of the Razdnol' naya River). Zootaxonomica Sinica 5: 144- 149, figs. 15- 27.
DEPOSITORY: ZISP. HOST: Moschus berezovskii Flerov.
DISTRIBUTION: People' s Republic of China,
Haemaphysalis grochovskajae Kolonin, 1992. In: V. Qinghai Province, and Qusum, XizangE. Sokolov, ed. Zoological Researches in Autonomous Region.
Vietnam, Nauka Publishers, Moscow ( in DEPOSITORY: IZAS.
Russian), pp. 242- 276, figs. 1- 5.HOST: Cattle. Haemaphysalis norvali Hoogstraal and Wassef, 1983.
DISTRIBUTION: Vietnam, Laocai. Onderstepoort Journal ofVete rinary ResearchDEPOSITORY: ZISP. 50: 183- 189, figs. 1- 35.
HOST: Erinaceus frontalis Smith [ now Atelerix
Haemaphysalis kadarsani Hoogstraal and Wassef, frontalis( Smith)].
1977. Journal ofParasitology 63: 1103- 1109, DISTRIBUTION: Zimbabwe, Matabeleland, Bula-
figs. 1- 28. wayo District. Also from Matopos National
HOSTS: Rattus dominator Thomas [ now Park, Victoria Falls Road, and Kumalo.
Paruromys dominator( Thomas)], Echiothrix DEPOSITORIES: USNTC; VRLH; OVI;
leucura Gray. ORSTOM, Bondy, France.DISTRIBUTION: Indonesia, Central Sulawesi.
DEPOSITORIES: MBB; USNTC; BMNH. Haemaphysalis paraleachi Camicas, Hoogstraal, and
El Kammah, 1983. Journal of ParasitologyHaemaphysalis lobachovi Kolonin, 1995. Folia 69: 400-404, figs. 1- 20.
Parasitologica 42: 239, figs. 1- 5. HOST: Viverra civetta( Schreber).
HOST: Hystrix cristata L. DISTRIBUTION: Cameroon, Nanga-Eboko ( 04°
DISTRIBUTION: Ethiopia, near Awash National 41' N, 12° 22' E).
Park( 08° 50' N, 39° 50' E). DEPOSITORIES: USNTC; J.- L. Camicas Col-
DEPOSITORIES: Zoological Museum ofMoscow lection( ORSTOM); P.- C. Morel Collection.
University, Moscow, Russia; author' scollection. Haemaphysalisphasiana Saito, Hoogstraal, and Wassef,
1974. Journal of Parasitology 60: 198- 208,Haemaphysalis mageshimaensis Saito and Hoogstraal, figs. 1- 35, tab. 1.
1973. Journal of Parasitology 59: 569- 578, HOST: Phasianus versicolor Vieillot.
figs. 1- 35. DISTRIBUTION: Japan, Niigata Prefecture, Sado
HOST: Collected by flagging vegetation. Island, Kanai( 38° 03' N, 138° 22' E).
DISTRIBUTION: Japan, Kagoshima Prefecture, DEPOSITORY: USNTC.
Mage Shima( Mage Island), Kumage- gun( 30° NOTE: Russia, China and Korea are listed along45' N, 130° 51' E). with other hosts for tick specimens that are
DEPOSITORIES: Medical Zoology Department, tentatively regarded as H. phasiana.Niigata University School of Medicine;USNTC. Haemaphysalis primitiva Teng, 1982. Acta
NOTE: It is presumed that Sika deer, Cervus nippon Zootaxonomica Sinica 7: 46- 48, figs. 1- 8.

December, 1999 Journal of Vector Ecology 123
HOST: Not stated but probably collected on Haemaphysalis subterra Hoogstraal, El Kammah, andvegetation. Camicas, 1992. International Journal of
DISTRIBUTION: People' s Republic of China, Acarology 18: 213- 220, figs. 1- 35.Sichuan Province, wilderness ofPeng County. HOSTS: For adults, Atilaxpaludinosus( G. Cuvier),
DEPOSITORY: Not stated but probably IZAS. Herpestes sanguineus ( Ruppell) [ now
Galerella sanguinea ( Ruppell)], GenettaHaemaphysalis quadriaculeata Kolonin, 1992. In: V. maculata ( Gray), Herpestes ichneumon ( L.),
E. Sokolov, ed. Zoological Researches in Ichneumia albicauda ( G. Cuvier), MungosVietnam, Nauka Publishers, Moscow ( in mungo ( Gmelin); for immatures, AethomysRussian), pp. 242- 276, figs. 1- 6. chrysophilus( de Winton), Praomys natalensis
HOST: Dogs. Smith) [ now Mastomys natalensis ( Smith)],DISTRIBUTION: Vietnam, Bacthai. Tachyoryctes splendens Ruppell.DEPOSITORY: ZISP. DISTRIBUTION: Kenya, Rift Valley, 25 miles
SW Nairobi on Magadi Road ( 01° 22' S, 36°Haemaphysalis qinghaiensis Teng, 1980. Acta 39' E). Also Democratic Republic of Congo
Zootaxonomica Sinica 5: 144- 149, figs. 1- 14. Zaire), Ethiopia, Republic of South Africa,HOST: Goats. Tanzania, Zambia, Zimbabwe.DISTRIBUTION: People' s Republic of China, DEPOSITORIES: USNTC; H. Hoogstraal
Qinghai Province, Huangyuan. Collection at NAMRU-3, Cairo, Egypt; J.- L.DEPOSITORY: IZAS. Camicas Collection at ORSTOM.
NOTE: One nymph collected from GalerellaHaemaphysalis simplicima Hoogstraal and Wassef, sanguinea( Ruppell).
1979. In: Uilenberg, Hoogstraal and Klein.1979. Archives de l' Institut Pasteur de Haemaphysalis suntzovi Kolonin, 1993. Journal ofMadagascar, Numero Special: 34- 39, plate Medical Entomology 30: 966- 968, figs. 1- 14.35, figs. 1- 4; plate 36, figs. 5- 20; plate 37, figs. HOSTS: Hystrix brachyura L., Sus scrofa L.21- 28. DISTRIBUTION: Vietnam, Daclac, Dac- Genh( 12°
HOSTS: Echinops telfairi Martin, Setifer setosus 30' N, 107° 50' E). Also from Gailai Contum,Schreber). Buon-Loi( 14° 10' N, 108° 30' E); Quangninh,
DISTRIBUTION: Madagascar, Tulear. Island Bamun ( 21° 05' N, 107° 40' E).DEPOSITORIES: USNTC; Institut Pasteur, DEPOSITORIES: ZISP; BMNH; Museum of
Tananarive, Madagascar; BMNH. Comparative Zoology, Harvard University,Cambridge, Massachusetts, USA; USNTC.
Haemaphysalis sindensis Bilqees and Masood, 1973.
Sind University Research Journal ( Science Haemaphysalis xinjiangensis Teng, 1980. Acta
Series) 7: 41- 56, figs. 1- 7. Entomologica Sinica 23: 86- 89, figs. 1- 12.HOST: Varanus monitor ( L.) [ now Varanus HOST: Collected in an alpine pasture and offa wild
bengalensis( Daudin)]. goat.
DISTRIBUTION: Pakistan, near Karachi. DISTRIBUTION: People' s Republic of China,DEPOSITORY: Sind University, School of southern part of Xinjiang Uygur Autonomous
Parasitology, Department of Zoology. Region.
NOTE: A junior synonym of Aponomma gervaisi DEPOSITORY: IZAS.Lucas).
Hyalomma arabica Pegram, Hoogstraal, and Wassef,Haemaphysalis sinensis Zhang, 1981. Acta Veterinaria 1982. Journal of Parasitology 68: 150- 156,
etZootechnicaSinica 12: 169- 173, figs. 1- 15, figs. 1- 17, tabs. 1- 2.tab. 1. HOSTS: Goats and sheep.
HOSTS: Cattle and goats. DISTRIBUTION: Yemen Arab Republic, Ta' izzDISTRIBUTION: Fang County, Hubei Province, Province, southern tihama( lowland) foothills,
People' s Republic of China. Al Hamilee( Al Muroa)( 13° 20' N, 43° 35' E),DEPOSITORY: Institute of Animal Husbandry Misgab as Seloo ( 13° 20' N, 44° 20' E), and
and Veterinary Science, Yang- xin County, Saudi Arabia, Mecca.Hubei Province, People' s Republic of China. DEPOSITORIES: USNTC; BMNH; Department

124 Journal of Vector Ecology December, 1999
of Biology, Faculty of Science, King Abd el Ixodes columnae Takada and Fujita, 1992. Journal ofAziz University, Jidda, Saudi Arabia. the Acarological Society of Japan 1: 37- 44,
NOTE: It is postulated that the original host for this fig. 1 a- c, fig. 2a-c, fig. 3a-d.tick species was the Nubian ibex, Capra HOSTS: Collected by flagging vegetation and fromnubiana F. Cuvier. Apodemusargenteus( Temminck),A. speciosus
Temminck), Clethrionomys[ now Phaulomys]
Hyalomma hystricis Dhanda and Raja, 1974. Oriental andersoni( Thomas), Sciurus[ now Petaurista]
Insects 8: 531- 536, figs. 1- 12. leucogenys ( Temminck), Sciurus lis
HOST: Hystrix indica Kerr. Temminck, and Phasianus colchicus L.
DISTRIBUTION: India, Tamil Nadu, Salem DISTRIBUTION: Japan, Hokkaido; Honshu: Akita,
District. Also in Madurai District, Andipatti Aomori, Fukushima, Gumma, Saitama, and
forest area of Tamil Nadu. Yamagata Prefectures.
DEPOSITORIES: Virus Research Centre, Poona, DEPOSITORY: Fukui Medical School, Fukui;
India; Zoological Society of India, Calcutta; Laboratory ofOhara General Hospital, Omachi,collection of Dr. E. Ebenezer Raja; BMNH. Fukushima, Japan.
NOTE: Specific collecting localities in theIxodes bivari Santos Dias, 1990. Boletim da Sociedade various prefectures are given in Table 1 of the
Portuguesa de Entomologia 4: 153- 170, 5 original description.
unnumbered figs.
HOST: Oryctolagus cuniculus( L.). Ixodes copei Wilson, 1980. International Journal ofDISTRIBUTION: Portugal, Leiria District, Porto Acarology 6: 157- 162, figs. 1- 8.
de Mos. HOST: Mimus polyglottos( L.).
DEPOSITORIES: IICT; BMNH. DISTRIBUTION: Jamaica, St. Thomas Parish, Blue
Mountains, Penlyne, Whitfield Hall.
Ixodes brewsterae Keirans, Clifford, and Walker, 1982. DEPOSITORY: USNTC.
Journal ofMedical Entomology 19: 309- 329,figs 1- 12. Ixodes corwini Keirans, Clifford, and Walker, 1982.
HOSTS: A tilaxpaludinosus( G. Cuvier), Herpestes Journal ofMedical Entomology 19: 309- 329,ichneumon( L.). figs. 35- 50.
DISTRIBUTION: Uganda, Buganda, Namulonge HOSTS: Aonyx capensis ( Schinz). Also on
00° 32' N, 32° 37' E). Also Democratic Galerellapulverulenta( Wagner), G. sanguinea
Republic of Congo and Congo. Riippell), Genetta genetta( L.), and G. tigrina
DEPOSITORIES: USNTC; BMNH; OVI. Schreber).
DISTRIBUTION: Republic of South Africa,
Ixodes calcarhebes Arthur and Zulu, 1980. Systematic Tsitsikama National Park( 33° 58' S, 23° 45' E).
Parasitology 1: 241- 244, text figs. 1- 2; p1. 1, DEPOSITORIES: OVI; USNTC; BMNH.
figs. 1- 5, pl. 2.
HOST: Praomys natalensis Smith[ now Mastomys Ixodes dammini Spielman, Clifford, Piesman, and
natalensis( Smith)]. Corwin, 1979. Journal ofMedical EntomologyDISTRIBUTION: Zambia, found in the tick 15: 218- 234, figs. 1- 30, tabs. 1- 3.
collection of the Pest Research Unit Labora- HOST: Collected by flagging vegetation.tories of the National Council for Scientific DISTRIBUTION: USA, Massachusetts, Nantucket
Research, Chilanga, Lusaka. Island( 41° 20' N, 70° 02' W).
DEPOSITORY: Same as for distribution. DEPOSITORY: USNTC.
NOTE: Numerous hosts and localities are listed in
Ixodes catherinei Keirans, Clifford, and Walker, 1982. Tables 1 and 2 of the original description. A
Journal ofMedical Entomology 19: 309- 329, junior synonym of Ixodes scapularis Say.figs. 51- 56.
HOST: Lepus saxatilis F. Cuvier. Ixodes donarthuri Santos Dias, 1980. Publicacoes do
DISTRIBUTION: Republic of South Africa, East Instituto de Zoologia " Dr. Augusto Nobre"
Cape Province, Clark' s Siding( 31° 25' S, 27° 151): 1- 11, fig. la-e.08' E). HOSTS: Redunca arundinum ( Boddaert),
DEPOSITORIES: OVI; USNTC. Sylvicapra grimmia( L.).

December, 1999 Journal of Vector Ecology 125
DISTRIBUTION: Mozambique, Nampula maculata( Gray)]. Also found on CephalophusProvince, Mutuali. sylvicultor( Afzelius).
DEPOSITORIES: MNHM; BMNH. DISTRIBUTION: Uganda, Kigezi District, KabaleNOTE: Probably ajunior synonym ofIxodes neitzi. 01° 15' S, 29° 59' E). Also Democratic Republic
of Congo, Haut-Congo, Yangambi( 00° 47' N,Ixodes drakensbergensis Clifford, Theiler, and Baker, 24° 28' E).
1975. Onderstepoort Journal of Veterinary DEPOSITORIES: USNTC; OVI; BMNH.Research 42: 33- 40, figs. 1- 13, tab. 1.
HOSTS: Collected by flagging vegetation. Also Ixodes matopi Spickett, Keirans, Norval, and Clifford,on goats, cattle and Taurotragus oryx 1981. Onderstepoort Journal of VeterinaryPallas). Research 48: 23- 30, figs. 1- 27a.
DISTRIBUTION: Republic of South Africa, Natal HOST: Oreotragus oreotragus( Zimmermann).now Kwa-Zulu Natal), Giant' s Castle Nature DISTRIBUTION: Zimbabwe, Matabeleland South
Reserve( 29° 16' S, 29° 30' E), and Tank Area Province, Rhodes Matopos ( now Matopos)118( 29° 35' S, 29° 50' E). National Park, Maleme Dam ( 20° 33' S, 28°
DEPOSITORIES: OVI; USNTC; BMNH. 30' E).
DEPOSITORIES: OVI; USNTC; BMNH.Ixodes eastoni Keirans and Clifford, 1983. Journal of NOTE: Ticks congregate on twigs containing pre-
Medical Entomology 20: 90-98, figs. 1- 28. orbital gland secretions of this host.HOSTS: Clethrionomysgapperi( Vigors), Microtus
longicaudus ( Merriam), M. pennsylvanicus Ixodes moscharius Teng, 1982. Acarina: Ixodidae. In:Ord), Neotoma cinerea( Ord), Sorex cinereus Insects of Xizang, Vol. II, pp. 449- 461, figs.
Kerr, Tamias minimus Bachman, Zapus 18- 26. Institute ofZoology, Academia Sinica.hudsonius( Zimmermann). HOST: Moschus berezovskii Flerov.
DISTRIBUTION: USA, South Dakota, Lawrence DISTRIBUTION: Xizang, Zham and Nylam.County, Spearfish Canyon ( 44° 25' N, 103° DEPOSITORY: IZAS.
52' W), also Pennington Co., Black Hills
National Forest; Wyoming, Weston Co. Ixodes myospalacis Teng, 1986. Acta ZootaxonomicaDEPOSITORY: USNTC. Sinica 11: 46- 53, figs 1- 10.
HOST: Myospalax fontanierii( Milne-Edwards).Ixodes galapagoensis Clifford and Hoogstraal, 1980. DISTRIBUTION: People' s Republic of China,
Proceedings of the Entomological Society of Gansu Province, Pingliang County( 35° 30' N,Washington 82: 378- 383, figs. 1- 14. 106° 40' E). Also in Ningxia Huizu Autonomous
HOST: Oryzomys galapagoensis bauri( Allen). Region and Shanxi Province.DISTRIBUTION: Ecuador, Galapagos Islands, DEPOSITORY: IZAS.
Santa Fe ( Barrington) Island ( 00° 49' N, 90°04' W). Ixodes neitzi Clifford, Walker, and Keirans, 1977.
DEPOSITORY: USNTC. OnderstepoortJournalofVeterinary Research44: 143- 149, figs. 1- 16.
Ixodes ghilarovi Filippova and Panova, 1988. La HOST: Redunca fulvorufula( Afzelius).Systematique des Insectes et Acarines ( Aca- DISTRIBUTION: Republic of South Africa,demie des Sciences de L' URSS Horae Transvaal ( now Gauteng) Province, LoskopSosietatis Entomologicae Unionis Soveticae): Dam Nature Reserve( 25° 24' S, 29° 21' E).212-217, figs. 1- 6, tab. 1, map. DEPOSITORIES: OVI; USNTC; BMNH.
HOST: Chionomys gud( Satunin).
DISTRIBUTION: Russia, Caucasus Mountains, Ixodes nicolasi Santos Dias, 1981. Anais da FaculdadeSouth Daghestan. de Ciencias do Porto 63: 119- 123, fig. la-f.
DEPOSITORY: ZISP. HOST: Sylvicapra grimmia( L.)
DISTRIBUTION: Mozambique, InhambaneIxodes macfarlanei Keirans, Clifford, and Walker, Province, Mambone.
1982. Journal of Medical Entomology 19: DEPOSITORY: MNHM.309- 329, figs. 23- 34.
HOSTS: Genetta tigrina rubiginosa[ now Genetta Ixodes ochotonarius Teng, 1973. Acta Entomologica

126 Journal of Vector Ecology December, 1999
Sinica 16: 73- 81, figs. 20- 33. HOSTS: Cattle. Also on goats.
HOST: Ochotona thibetana( Milne-Edwards). DISTRIBUTION: People' s Republic of China,
DISTRIBUTION: People' s Republic of China, Fujian and Anhui Provinces.
Sichuan Province, Maerkang. DEPOSITORIES: IZAS.
DEPOSITORY: IZAS.
NOTE: Although we have not seen specimens of Ixodes zairensis Keirans, Clifford, and Walker, 1982.
this taxon, from the description and figures we Journal ofMedical Entomology 19: 309- 329,believe it to be a junior synonym of Ixodes figs. 13- 22.
hyatti Clifford, Hoogstraal, and Kohls. HOSTS: Crocidura flavescens ( I. Geoffroy) andCrocidura sp.
Ixodes pararicinus Keirans and Clifford, 1985. In: DISTRIBUTION: Democratic Republic of Congo,
Keirans, Clifford, Guglielmone and Mangold, Gemena Zone, Tandala( 02° 58' N, 19° 21' E).
1985. Journal of Medical Entomology 22: DEPOSITORIES: USNTC; BMNH; OVI.
401- 407, figs. 1- 14.
HOSTS: Cattle. Also on horses. Keiransiella Santos Dias, 1992. Garcia de Orta, SerieDISTRIBUTION: Argentina, Salta Province, Zoologia 19: 11- 19. [ SubgenusofAmblyomma;
Capital Department, Lesser ( 24° 40' S, 65° type- A. ( K.) albopictum Neumann].
34' W). Also in Tucamen Province, Argentina
and in Uruguay, Departments ofFlorida,Rocha Koloninum Santos Dias, 1992. Garcia de Orta, Serie
and Maldonado. Zoologia 19: 11- 19. [ Subgenus ofAmblyomma;DEPOSITORIES: USNTC; MBR; BMNH. type- A.( K.) bibroni( Gervais)= Amblyomma
dissimile Koch].
Ixodes rangtangensis Teng, 1973. Acta EntomologicaSinica 16: 73- 81, figs. 13- 19. Mammalixodes Emel' yanova, 1979. In: M. V. Efimov
HOST: Moschus berezovskii Flerov. and N. M. Pronin, eds. Zooparasitology ofDISTRIBUTION: People' s Republic of China, Lake Baikal Basin. Akademia Nauk SSSR,
Sichuan Province, Rangtang County. Sibirskoe Otdelenie Buryatskii Filial OtdelDEPOSITORY: IZAS. Biologii, Ulan- Ude, pp. 5- 27. ( In Russian).
Subgenus of Pholeoixodes; type - P. ( M.)
Ixodes siamensis Kitaoka and Suzuki, 1983. Tropical prokop' yevi Emel' yanova]. ( In English:
Medicine 25: 205- 219, figs. 1- 5, tab. 2. NAMRU-3- T1739).
HOSTS: Anourosorex squamipes Milne-Edwards,
Mus pahari Thomas. Ornithixodes Emel' yanova, 1979. In: M. V. EfimovDISTRIBUTION: Thailand, Doi Inthanon. and N. M. Pronin, eds. Zooparasitology ofDEPOSITORIES: National Science Museum, Lake Baikal Basin. Akademia Nauk SSSR,
Natural History Institute, Shinjuku, Tokyo; Sibirskoe Otdelenie Buryatskii Filial Otdel
National Institute of Animal Health, Tsukuba, Biologii, Ulan- Ude, pp. 5- 27. ( In Russian).
Ibaraki, Japan. Subgenus of Pholeoixodes; type - P. ( 0.)
arboricola Schulze and Schlottke].( In English:
Ixodes sigelos Keirans, Clifford, and Corwin, 1976. NAMRU-3- T1739).
Acarologia 18: 217- 225, figs. 1- 22.
HOSTS: Abrocoma bennetti Waterhouse, Pholeoixodes prokop' yevi Emel' yanova, 1979. In: M.Aconaemys fuscus ( Waterhouse), Octodon V. Efimov and N. M. Pronin, eds.
degus ( Molina), Phyllotis sp., probably P. Zooparasitology of Lake Baikal Basin.darwini( Waterhouse). Akademia Nauk SSSR, Sibirskoe Otdelenie
DISTRIBUTION: Chile, Malleco Province, Parque Buryatskii Filial Otdel Biologii, Ulan- Ude,Nahuelbuta ( 37° 50' S, 72° 57' W), also in pp. 5- 27, figs. 46- 79.( In Russian). ( In English:
Santiago and Maule Provinces. NAMRU-3- T1739).
DEPOSITORIES: USNTC; Texas Tech University, HOST: Erinaceus dauuricus [ now MesechinusLubbock, Texas, USA. dauuricus( Sundevall)].
DISTRIBUTION: Mongolia, northeastern steppes.Ixodes sinensis Teng, 1977. Acta Entomologica Sinica Also found in the southeastern Transbaikal.
20: 342- 344, figs. 1- 11. DEPOSITORY: Not stated.

December, 1999 Journal of Vector Ecology 127
Rhipicephalus aquatilis Walker, Keirans, and Pegram, Keirans. 1995. Onderstepoort Journal of1993. Onderstepoort Journal of Veterinary Veterinary Research 62: 89- 95, fig. la-b, 2a- f,Research 60: 205- 210, figs. 1- 9, tab. 1. 3, tab. 1.
HOSTS: Tragelaphus spekii Sclater, occasionally HOST: Cattle.
from Panthera leo ( L.), P. pardus ( L.) and DISTRIBUTION: Uganda, Masaka District,
cattle. Kawoko- Masaka ( 00° 30' S, 31° 35' E);
DISTRIBUTION: Uganda, Tanzania, and Zambia Tanzania, Igula Village, Ihimbu Gunguli( 07°
in semi- aquatic habitats. 50' S, 35° 47' E); Zambia, Lutale, Mumbwa
DEPOSITORIES: BMNH; OVI; USNTC. 15° 16' S, 26° 50' E).
DEPOSITORIES: USNTC; BMNH; OVI.
Rhipicephalus arakeri Hiregoudar, 1975. Mysore
Journal of Agricultural Science 9: 473- 479, RhipicephaluslounsburyiWalker, 1990. Onderstepoort
figs. 1- 10. Journal of Veterinary Research 57: 57- 75,HOST: Rattus rattus( L.). figs.-1- 25, tab. 1.
DISTRIBUTION: India, Gujarat, Anand. HOSTS: Sheep. Also on various antelope species.DEPOSITORY: Zoological Survey of India, DISTRIBUTION: Republic of South Africa,
Calcutta. Eastern Cape Province, Dordrecht( 31° 22' S,
NOTE: A junior synonym of Rhipicephalus 27° 02' E).
ramachandrai Dhanda. DEPOSITORIES: OVI; USNTC.
NOTE: Ticks are found on the feet of these animals,
Rhipicephalus bergeoni Morel and Balis, 1976. Revue usually between the toes and on the heels.d' Elevage etde Medecine Veterinaire des Pays Numerous other collecting localities in SouthTropicaux 29: 141- 148, figs. la-h, 2a- i. Africa are cited on pg. 64 of the original
HOST: cattle. description.
DISTRIBUTION: Ethiopia, Harrar, Hubeta. Also
from Bagemder, Bale, Gemu Gofa, Gojam, RhipicephalusneumanniWalker, 1990. Onderstepoort
and Sidamo. Journal of Veterinary Research 57: 57- 75,DEPOSITORY: Not stated. figs. 26- 49, 52- 54, 58, tab. 2.
HOSTS: Sheep. Also on goats to a lesser extent.Rhipicephalus camicasi Morel, Mouchet, and Rodhain, DISTRIBUTION: Namibia, Bethanien District,
1976. farm" Soutdoringvlei"( c. 26° 05' S, 17° 10' E).
Revue d' Elevage et de Medecine Veterinaire Also in the Northern, Western, and Eastern
des Pays Tropicaux 29: 337- 340, fig. 1. Cape Provinces, Republic of South Africa.
HOSTS: Sheep. Also on goats and Lepus capensis DEPOSITORIES: OVI; USNTC; BMNH.
L. NOTE: Ticks are found on the feet of these animals,
DISTRIBUTION: Afars and Issas( now Djibouti), usually between the toes. Numerous other
Randa. Also Ethiopia, Harrar, and Shoa. collecting localities are cited on pp. 72 and 74DEPOSITORY: Not stated. of the original description.
Rhipicephalus exophthalmos Keirans and Walker, Rhipicephalus tetracornus Kitaoka and Suzuki, 1983.
1993. In: Keirans, Walker, Horak, and Heyne, Tropical Medicine 25: 205- 219, figs. 6- 23,
1993. Onderstepoort Journal of Veterinary tab. 2.
Research 60: 229-246, figs. 1- 17, tabs. 1- 4. HOSTS: AnourosorexsquamipesMilne- Edwards,
HOSTS: Cattle, sheep, occasionally goats; wild Maxomys surifer ( Miller), Rattus nitidus
artiodactyls such as Antidorcas marsupialis Hodgson).
Zimmermann), Oryx gazella ( L.), and DISTRIBUTION: Thailand, Doi Inthanon.
Tragelaphus strepsiceros ( Pallas); leporids DEPOSITORY: National Science Museum, Natural
such as Lepus saxatilis F. Cuvier. History Institute, Shinjuku, Tokyo, Japan.DISTRIBUTION: Namibia, Botswana, South
Africa, and Angola. Rhipicephalus zambeziensis Walker, Norval, and
DEPOSITORIES: BMNH; OVI; USNTC. Corwin, 1981. Onderstepoort Journal ofVeterinary Research 48: 87- 104, figs. 1- 56,
Rhipicephalus interventus Walker, Pegram, and tabs. 1- 2.

128 Journal of Vector Ecology December, 1999
HOST: Cattle. R. zambeziensis.DISTRIBUTION: Zimbabwe, Gwanda District, PAKISTAN: Haemaphysalis danieli, H. sindensis.
West Nicholson, Drihoek Ranch ( 21° 25' S, PAPUA NEW GUINEA: Carios multisetosus.29° 46' E). This species is also found in other PEOPLE' S REPUBLIC OF CHINA: Argas assimilis,areas of Zimbabwe, Zambia, Namibia, A. beijingensis, A. sinensis, AmblyommaBotswana, and in northern and western hainanense, Anomalohimalaya cricetuli,
Transvaal( now Gauteng) Province, Republic Haemaphysalis anomaloceraea, H. mengaaensis,of South Africa. H. moschisuga, H. primitiva, H. qinghaiensis, H.
DEPOSITORIES: OVI; BMNH; USNTC; VRLH. sinensis, H. xinjiangensis, Ixodes myospalacis, 1.
ochotonarius, I. rangtangensis, I. sinensis.
PERU: Argas dalei,A. moreli, Ornithodoros spheniscus.Countries Where These Tick Species are Found POLAND: Argas polonicus.
PORTUGAL: Ixodes bivari.AFGHANISTAN: Haemaphysalis danieli. PUERTO RICO: Amblyomma arianae.ANGOLA: Rhipicephalus exophthalmos. REPUBLIC OF SOUTH AFRICA:Argasafricolumbae,ARGENTINA: Amblyomma pseudoparvum, Ixodes Haemaphysalis subterra, Ixodes catherinei, I.
pararicinus. corwini, I.drakensbergensis, I.neitzi,RhipicephalusAUSTRALIA: Argas dewae, A. falco, A. lowryae, A. exophthalmos, R. lounsburyi, R. neumanni, R.
macrodermae, Amblyomma glauerti, A. vikirri, zambeziensis.
Aponomma glebopalma. RUSSIA: Haemaphysalis filippovae, Ixodes ghilarovi.BOTSWANA: Rhipicephalus exophthalmos, R. SAUDI ARABIA: Hyalomma arabica.
zambeziensis. SENEGAL: Alectorobius camicasi.CAMEROON: Haemaphysalis paraleachi. SPAIN: Argas gilcolladoi.CHILE: Ixodes sigelos. TAJIKISTAN: Anomalohimalaya lotozkyi, Derma-CONGO: Ixodes brewsterae. centor montanus.
CUBA: Antricola armasi, A. centralis, A. cernyi, A. TANZANIA: Haemaphysalis subterra, Rhipicephalusgranasi,A. habanensis, A. martelorum,A. naomiae, aquatilis, R. interventus.A. occidentalis,A. siboneyi, Ornithodoros cyclurae. TIBET( XIZANG) Ixodes moscharius.
DEMOCRATIC REPUBLIC OF CONGO: Aponomma THAILAND: Ixodes siamensis, Rhipicephalusinopinatum, Haemaphysalis subterra, Ixodes tetracornus.
brewsterae, I. macfarlanei, I.zairensis. TURKMENISTAN: Dermacentor ushakovae.DJIBOUTI: Rhipicephalus camicasi. UGANDA: Ixodes brewsterae, I. macfarlanei,DOMINICAN REPUBLIC: Ornithodoros antiquus. Rhipicephalus aquatilis, R. interventus.ECUADOR: Ornithodoros yunkeri, Ixodes URUGUAY: Ixodes pararicinus.
galapagoensis. USA: Argas monolakensis, A. ricei, Ixodes dammini, I.ETHIOPIA: Haemaphysalis lobachovi, H. subterra, eastoni.
Rhipicephalus bergeoni, R. camicasi. VENEZUELA: Antricola hummelincki.INDIA: Hyalomma hystricis, Rhipicephalus arakeri. VIETNAM: Aponomma orlovi, HaemaphysalisINDONESIA: Carios hadiae, C. multisetosus, C. bacthaensis, H.grochovskajae, H.quadriaculeata,
papuensis, Ornithodoros collocaliae, Haema- H. suntzovi.physalis kadarsani. YEMEN ARAB REPUBLIC: Hyalomma arabica.
JAPAN: Haemaphysalis mageshimaensis, H.phasiana, ZAMBIA:Haemaphysalis subterra, Ixodes calcarhebes,Ixodes columnae. Rhipicephalus aquatilis, R. interventus, R.
KAZAKSTAN: Dermacentor ushakovae. zambeziensis.
KENYA: Ornithodoros vansomereni, Haemaphysalis ZIMBABWE: Haemaphysalis norvali, H. subterra,subterra. Ixodes matopi, Rhipicephalus zambeziensis.
KYRGYZSTAN: Dermacentor ushakovae.MADAGASCAR: Haemaphysalis simplicima. REFERENCESMONGOLIA: Haemaphysalis demidovae, Pholeoixodes
prokop' yevi. Doss, M. A., M. A. Fan, K. F. Roach, and G. Anastos.MOZAMBIQUE: Boophilus florae, B. scheepersi, 1974. Ticks and tickborne diseases. I. Genera and
Ixodes donarthuri, 1. nicolasi. species of ticks. Part 1. Genera A-G. Index-NAMIBIA:Rhipicephalus exophthalmos, R. neumanni, Catalogue of Medical and Veterinary Zoology.

December, 1999 Journal of Vector Ecology 129
Special Publication No. 3. United States Department Egypt, U.A.R., v+ 498 pp.of Agriculture, Washington, DC, 429 pp. Hoogstraal, H. 1970. Bibliography ofticks and tickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311974. Ticks and tickborne diseases. I. Genera and December 1969. Volume 2. Special publicationspecies of ticks. Part 2. Genera H-N. Index- December 1970, United States Naval MedicalCatalogue of Medical and Veterinary Zoology. Research Unit Number Three( NAMRU-3), Cairo,Special Publication No. 3. United States Department Egypt, U.A.R., 495 pp.of Agriculture, Washington, DC, 593 pp. Hoogstraal, H. 1971. Bibliography of ticksandtickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311974. Ticks and tickborne diseases. I. Genera and December 1969. Volume 3. Special publicationspecies of ticks. Part 3. Genera O-X. Index- June 1971, United States Naval Medical ResearchCatalogue of Medical and Veterinary Zoology. Unit Number Three ( NAMRU-3), Cairo, Egypt,Special Publication No. 3. United States Department U.A.R., 435 pp.of Agriculture, Washington, DC, 329 pp. Hoogstraal, H. 1972. Bibliography of ticks and tickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311974. Ticks and tickborne diseases. II. Hosts. Part December 1969. Volume 4. Special publication1. A-F. Index-Catalogue ofMedical and Veterinary June 1972, United States Naval Medical ResearchZoology. Special Publication No. 3. United States Unit Number Three ( NAMRU-3), Cairo, Egypt,Department of Agriculture, Washington, DC, 489 U.A.R., i+ 355 pp.pp. Hoogstraal, H. 1974. Bibliography of ticks and tickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311974. Ticks and tickborne diseases. II. Hosts. Part December 1973. Volume 5, Part I. Special2. G-P. Index-Catalogue ofMedical and Veterinary publication August 1974, United States NavalZoology. Special Publication No. 3. United States Medical Research Unit Number Three( NAMRU-Department of Agriculture, Washington, DC, iv+ 3), Cairo, Egypt, U.A.R., vii+ 492 pp.489- 976. Hoogstraal, H. 1978. Bibliography ofticks and tickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311974. Ticks and tickborne diseases. II. Hosts. Part December 1976. Volume 5, Part II. Special3. Q- Z. Index-Catalogue ofMedical and Veterinary publication April 1978, United States Naval Medical
Zoology. Special Publication No. 3. United States Research Unit Number Three( NAMRU-3), Cairo,Department of Agriculture, Washington, DC, vi+ Egypt, U.A.R., iii+ 455 pp.977- 1268. Hoogstraal, H. 1981. Bibliography ofticks and tickborne
Doss, M. A. and G. Anastos. 1977. Ticks and tickborne diseases from Homer ( about 800 B. C.) to 31diseases. III. Checklist of families, genera, species, December 1979. Volume 6. Special publicationand subspecies of ticks. Index-Catalogue ofMedical July 1981, United States Naval Medical Researchand Veterinary Zoology. Special Publication No. Unit Number Three ( NAMRU-3), Cairo, Egypt,3. United States Department of Agriculture, U.A.R., iii+ 407 pp.Washington, DC, 97 pp. Hoogstraal, H. 1982. Bibliography of ticks and tickborne
Doss, M. A., M. A. Farr, K. F. Roach, and G. Anastos. diseases from Homer ( about 800 B. C.) to 311978. Ticks and tickborne diseases. IV. December 1981. Volume 7. Special publicationGeographical distribution ofticks. Index-Catalogue May 1982, United States Naval Medical Researchof Medical and Veterinary Zoology. Special Unit Number Three ( NAMRU-3), Cairo, Egypt,Publication No. 3. United States Department of U.A.R., iii+ 219 pp.Agriculture, Washington, DC, 648 pp. Hoogstraal, H. 1988. Bibliography ofticks and tickborne
Hoogstraal, H. 1970. Bibliography of ticks and tickborne diseases from Homer ( about 800 B. C.) to 31diseases from Homer ( about 800 B. C.) to 31 December 1984. Volume 8. Special publicationDecember 1969. Volume 1. Special publication January 1988, United States Naval MedicalOctober 1970, United States Naval Medical Research Unit Number Three( NAMRU-3), Cairo,Research Unit Number Three( NAMRU-3), Cairo, Egypt, U.A.R., x+ 327 pp.

Journal of Vector Ecology 24(2): 130- 137
Larval Habitats of Anopheline Mosquitoes
in the Upper Orinoco, Venezuela
E. Rejmankova', Y. Rubio- Palis2, and L. Villegas3'4
Department ofEnvironmental Science and Policy,University of California, Davis, CA 95616 USA
2Division de Investigaciones, Escuela de Malariologia y SaneamientoAmbiental, Apartado 2073, Maracay 2101- A, VENEZUELA
3 Distrito Sanitario 4, Direccion Regional de Salud, Amazonas, VENEZUELA
4Present Address: Shoklo Malaria Research Unit, THAILAND
Received 22 December 1998; Accepted 8 March 1999
ABSTRACT: Survey of larval habitats of anopheline mosquitoes was conducted in Ocamo in the Stateof Amazonas, southern Venezuela. The sampled habitats belonged to three different hydrological types:
lagoons( 26 habitats), forest pools including flooded forest( 16 habitats), and forest streams( 4 habitats).Out of 46 habitats surveyed, 31 contained anopheline larvae. Six species were found: Anopheles darlingi,
Anopheles triannulatus,Anopheles oswaldoi,Anophelesperyassui,Anophelespunctimacula, andAnopheles
mediopunctatus. Anopheles triannulatus was the most abundant species. Significantly higher numbers ofanopheline larvae, in general, and of An. triannulatus specifically were found in lagoons with submersedmacrophytes and sparse emergent graminoids than in forest pools with detritus.
Keyword Index: Larval habitats, Anopheles darlingi, An. triannulatus, An. oswaldoi, An. peryassui, An.
punctimacula, An. mediopunctatus, Upper Orinoco, Ocamo.
INTRODUCTION documented from the Ocamo area by Rubio-Palis( 1995)and Rubio-Palis et al. ( 1997), but the larval habitats
An ecological approach that relates ( in predictive have not been described. In this paper we report results
models) vector distribution, environmental charac- of a systematic survey of anopheline larval habitatsteristics, and human activities to malaria transmission conducted in Ocamo in July, 1997. The results of thishas recently been advocated for malaria control programs survey are supported by additional data collected overRoberts and Rodriguez 1994, Rubio- Palis and several trips to the region by one of the authors( YRP).
Zimmerman 1997). Defining environmental conditions This study provides the only available data on larvalthat characterize larval habitats of malaria vectors is an ecology of malaria vectors from this region and shouldintegral part of this approach. While we know a lot be viewed as a potential starting point for more detailedabout distribution and behavior of adult mosquitoes, investigations of larval habitats in the Upper Orinoco.
larval habitat studies have often been neglected.
An increase in numbers ofmalaria cases has occurred METHODS
in many countries where malaria has been previouslyunder control( PAHO 1998). One of the regions that has Site Description
experienced recent increase in the incidence of malaria The survey oflarval habitats was carried out in July,is the Upper Orinoco, specifically the Yanomami 1997 along the Ocamo and Orinoco Rivers( Fig. 1) in thesettlements in the Biospheric Reserve Alto Orinoco- Alto Orinoco-Casiquiare Biospheric Reserve, southernCasiquiare in the State ofAmazonas, southern Venezuela. Venezuela. No high resolution maps/ remote sensingAdult seasonal abundance, behavior, and entomological images are available for the region; the most detailedinoculation rate of the principal malaria vector of this map is on a scale of 1: 500,000. We created our own mapregion, Anopheles darlingi, have been recently by plotting positions of the rivers, villages, and sampling
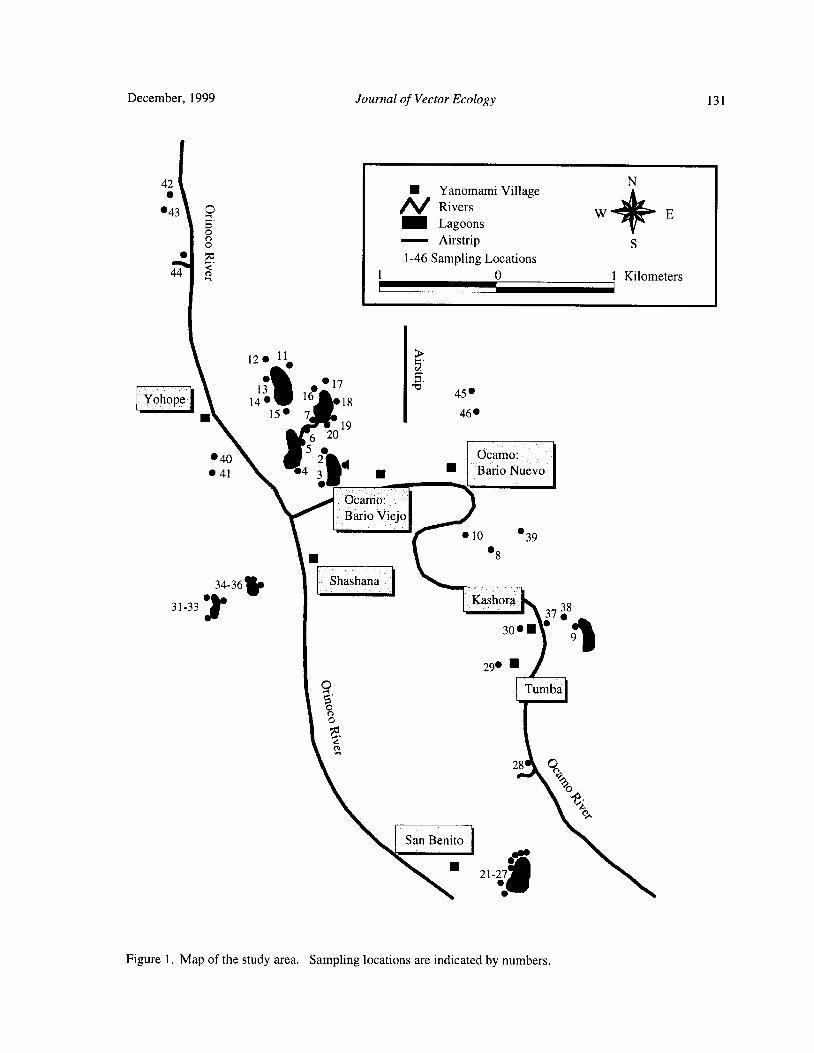
December, 1999 Journal of Vector Ecology 131
42 NYanomami Village
O N Rivers43 W ;`, E
o'Lagoons
o Airstrip S
a? 1- 46 Sampling Locations1 0 1 Kilometers
12• 11 Y
1317 b'•
Yohope I 14• 1°
1845
15• 7 46•
19
6 20
405
2 Ocamo:
41 4 3 Bario Nuevo
Ocamo:
Bario Viejo
0 • 39
8
34-36kShashana
ex31- 33Kask ora'
3•837 7
30• • 129• ll
O Tumba 1o00
GCDH28 9,
O
San Benito I M
21-
27a
Figure 1. Map of the study area. Sampling locations are indicated by numbers.

132 Journal of Vector Ecology December, 1999
locations as we recorded them with the GPS ( Trimble sampled. Thirty dips for mosquito larvae were takenNavigation). The area includes several villages, Santa from each habitat with a standard mosquito dipper.
Maria de los Guaicas - Ocamo, located close to the Larvae were reared to adults and some were preserved
confluence of Ocamo and Orinoco rivers; Clavotheri, in 80% ethanol. Identified voucher specimens are
Carl itos, Kashora, and Tumba along Ocamo river; and deposited in the collection of the Entomology Laboratory,Shashana, San Benito, Lechoza, and Yohope along the Research Division, School of Malariology in Maracay,Orinoco river. Venezuela, and at the Museum Support Center,
Rio Ocamo, one of the major tributaries of the Smithsonian Institution.
Upper Orinoco river is draining the central and southwestslopes of Sierra Parima in east- central Amazonas. Both Environmental Factors
the Upper Orinoco and Ocamo rivers can be characterized The environmental data were recorded for each site
as clear water rivers with yellow-brown color, relatively ( habitat) either as nominal values ( hydrology type,low suspended solids, low nutrients, and circumneutral surrounding environment, degree of shade), estimatedpH. These conditions reflect low mineral content of percentages( tree cover, shrub cover, detritus cover), or
parent rocks( igneous metamorphic basement of granite measured water characteristics ( pH, conductivity, ionoverlain with layers of sand). Geomorphologically the concentrations). Standard methods of water analyses
study area is part of the Upper Orinoco lowlands( mean were used for determining the concentrations of cationselevation about 135 m above see level) typified by a ( APHA 1985).
mosaic of periodically flooded forests including manypalm species ( Steyermark et al. 1995). Climate is Field Data Analysis
macrothermic ombrophilous( hot and rainy) with mean The individual habitats, defined by dominant plantannual T> 24° C and mean annual precipitation> 2200 species/ forms and hydrology/ water chemistry, weremm( Ministerio del Ambiente station in Ocamo, 1997). categorized into higher units referred to as habitat types
The area is sparsely inhabited by Amerindians of ( Rejmankova et al. 1992). Cluster analysis ( average
Yanomami ethnic group practicing a shifting agriculture. linkage, chord Euclidean distance, Podani 1996) was
Forests in the vicinity of villages are interspersed with conducted. The environmental variables measured as
small fields, " conucos." Numerous lagoons are part of percentages were subjected to the angular transformation.
the fringing river floodplain. The floodplain is seasonally The two-tailed t-test was used to compare the means of
inundated by flow from the main river channels. When environmental variables for sites with or without larvae.
the floodwater recedes, the lagoons are isolated from the Null hypothesis of independence of larval occurrence
river until the following inundation. During the isolation on types of habitats was tested with chi- square criterion
period, water level is primarily controlled by rainfall, ( Sokal and Rolf 1995).
evapotranspiration, and groundwater seepage( Hamilton
and Lewis 1990). In addition to lagoons, flooded forest RESULTS
usually a transient habitat formed after a rain), smallforest pools, and forest streams represent potential larval In July, 1997, we sampled 46 potential larvalhabitats. The lagoons support diverse aquatic vegetation, habitats in the Ocamo area ( see the map, Fig. 1 forincluding emergentmacrophytes( graminoids, Ludwigia sampling locations). These habitats belonged to three
spp., Montrichardia arborescens); submersed plant different hydrological types: lagoons ( 26 habitats),
communities represented by genera Elodea, Mayaca, forest pools including flooded forest( 16 habitats), andUtricularia, and Cabomba; and occasional floating forest streams ( 4 habitats). The environmental
macrophytes( Lemna, Salvinia). Forest pools and flooded differences among these three types of habitats areforests provide habitats dominated by detritus, i. e., an summarized in TABLE 1. Lagoon habitats were
accumulation of predominantly organic debris on the significantly deeper than the other two types and theywater surface. had water mineral content ( specific conductivity and
potassium, calcium and magnesium cations)
Larval Sampling comparable to streams but lower than forest pools.
Surveys of larval habitats were conducted at the Lagoons were also significantly less shaded than forestbeginning of the wet season( July, 1997). Sampling was pools and streams. They usually contained diverseconcentrated in about 1 km buffer zones along the rivers communities of submersed macrophytes, which were
see Fig. 1). All potential habitats encountered when absent in forest pools and streams.
walking around the villages and through the forest with Anopheline larvae were present in 31 out of 46
a Yanomami guide, well familiar with the area, were sampled habitats. Six species were identified: Anopheles

December, 1999 Journal of Vector Ecology 133
TABLE 1. Means and standard deviations ofenvironmental variables measured in habitats belonging to the threehydrological types. Means sharing the same letter are not significantly different( Scheffe' s test).
Variable Lagoons Forest Pools Forest Streamsn 26 16 4
Water Depth[ cm] 57.0 ( 23. 6) a 16. 0 ( 1. 0) b 30. 0 ( 18. 5) bpH 5. 7 ( 0.5) a 5. 7 ( 0.3) a 5. 2 ( 0.4) bSpecial Conditions [µ S cm-
I] 14.6 ( 3. 5) a 20. 7 ( 12. 3) b 9.0 ( 1. 3) a
K+[ppm] 0.8 ( 0.7) a 1. 7 ( 1. 5) b 0.5 ( 0.2) a
Ca++ [ ppm] 1. 5 ( 0.9) a 2.6 ( 2. 1) b 1. 2 ( 1. 1) a
Mg++ [ ppm] 0.2 ( 0. 1) a 0.4 ( 0. 3) b 0. 1 ( 0.08) a
Tree Cover[%] 20. 6 ( 28. 6) a 67. 8 ( 19. 6) b 57.5 ( 18. 9) bShrub Cover[%] 2. 1 ( 3. 9) a 11. 4 ( 21. 3) b 3. 8 ( 2. 5) a
Submersed[%] 33. 3 ( 39.0) 0.0 0.0Detritus [%] 13. 9 ( 23. 4) a 40.0 ( 16. 3) b 56.3 ( 30.9) b
Nyssorhynchus) darlingi Root, An. ( Nyssorhynchus) cover, low water depth, and frequent presence ofdetritus.triannulatus ( Neiva and Pinto), An. ( Nyssorhynchus) Cluster A closely overlaps with the hydrological typeoswaldoi ( Peryass), An. (Anopheles) peryassui Dryar " lagoon." Correspondingly, the frequency analysis ofand Knab, An. ( Anopheles) punctimacula Dryar and larval occurrence in the two main clusters showed thatKnab, andAn.( Anopheles) mediopunctatus( Theobald). significantly more An. triannulatus, as well as more ofIn addition, one of the authors collected An. total anopheline larvae were found in cluster ANyssorhynchus) marajoara Galvao and Damasceno ( TABLE 2). Frequency analysis testing the deviation
and An. ( Nyssorhynchus) argyritarsis Robineau- from randomness of larval distribution among all fourDesvoidy in the lagoons ( locations 1- 3, Fig. 1) on a clusters ( clusters 1, 2, 3, and cluster B) showed theprevious date( Yasmin Rubio- Palis, unpublished data). heterogeneity ofdistribution ofAn. triannulatus amongNot all the larvae were identified to species level since individual clusters to be only marginally significantmany collections consisted only of first or second larval ( chi- square 7. 18; DF= 3; p= 0. 06).instars. The abundances of other identified species, TABLE 3 shows the variables that significantlyexcept for An. triannulatus, were too low to allow for varied between habitats with An. triannulatus presenttesting of associations between individual mosquito versus absent. Tree cover and consequently a degree ofspecies and habitat types Consequently, only associa- shade, cover of broad- leaved macrophytes, total covertions for An. triannulatus and for total anophelines are ofsubmersed macrophytes, as well as cover of individual
presented. The frequency analysis of larval occurrence submersed species Utricularia spp. and Mayaca spp.,in the three types of hydrologically defined habitats cover of detritus, and cover of filamentous algae wereTABLE 2) showed that larvae ofAn. triannulatus were significantly different between habitats with and without
significantly more frequent in lagoons than in forest An. triannulatus larvae. Differences in water depth andpools or streams. The same analysis using data for all pH were not quite significant ( p= 0.08), and specific
anopheline larvae showed similar trend but the conductivity as well as individual cations did not differassociation was only marginally significant( chi-square between positive and negative habitats.4. 99; DF= 2; p= 0.08).
The cluster analysis of habitats based on DISCUSSION
environmental variables revealed two main clusters( A
and B) differing mainly in the tree cover and water depth There are several reasons why lagoons wouldFig. 2). Cluster A can be further divided into three provide favorable habitats for immature stages of
distinct smaller clusters dominated by submersed mosquitoes. Vegetation is usually well developed inmacrophytes( cluster# 1), grasses( cluster# 2), and sedges lagoons, since they are more permanent than ephemeralcluster# 3). Cluster B is characterized by a high tree forest pools and have a higher light availability. Higher
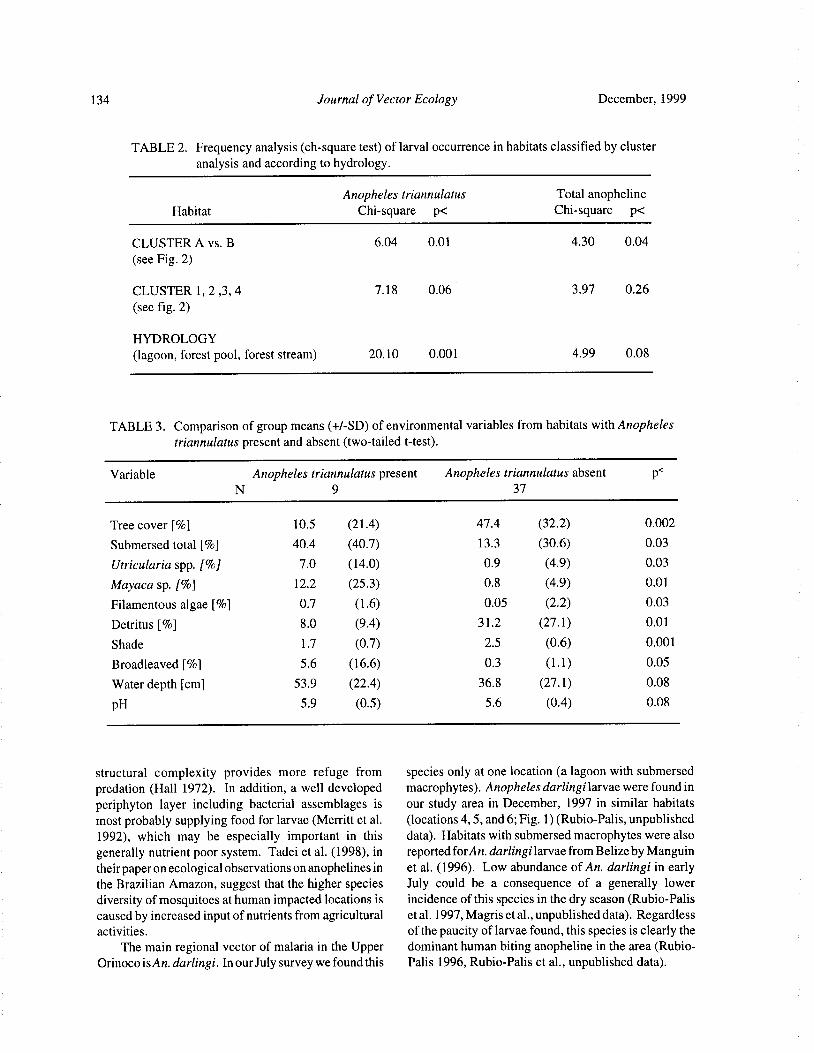
134 Journal of Vector Ecology December, 1999
TABLE 2. Frequency analysis( ch- square test) of larval occurrence in habitats classified by clusteranalysis and according to hydrology.
Anopheles triannulatus Total anopheline
Habitat Chi- square p< Chi-square p<
CLUSTER A vs. B 6.04 0.01 4.30 0.04
see Fig. 2)
CLUSTER 1, 2, 3, 4 7. 18 0.06 3. 97 0.26
see fig. 2)
HYDROLOGY
lagoon, forest pool, forest stream) 20. 10 0. 001 4.99 0.08
TABLE 3. Comparison of group means(+/- SD) of environmental variables from habitats with Anopheles
triannulatus present and absent( two- tailed t-test).
Variable Anopheles triannulatus present Anopheles triannulatus absent p<
N 9 37
Tree cover[%] 10.5 ( 21. 4) 47. 4 32.2) 0.002
Submersed total [%] 40.4 ( 40.7) 13. 3 30.6) 0.03
Utricularia spp. [%] 7.0 ( 14. 0) 0.9 4.9) 0.03
Mayaca sp. [%] 12. 2 ( 25. 3) 0.8 4.9) 0.01
Filamentous algae [%] 0. 7 1. 6) 0.05 2.2) 0. 03
Detritus [%] 8. 0 9. 4) 31. 2 27. 1) 0. 01
Shade 1. 7 0.7) 2. 5 0.6) 0.001
Broadleaved[ To] 5. 6 ( 16.6) 0. 3 1. 1) 0.05
Water depth[ cm] 53.9 ( 22.4) 36. 8 27. 1) 0.08
pH 5. 9 0.5) 5. 6 0.4) 0.08
structural complexity provides more refuge from species only at one location( a lagoon with submersedpredation ( Hall 1972). In addition, a well developed macrophytes). Anopheles darlingi larvae were found in
periphyton layer including bacterial assemblages is our study area in December, 1997 in similar habitatsmost probably supplying food for larvae( Merritt et al. ( locations 4, 5, and 6; Fig. 1)( Rubio-Palis, unpublished1992), which may be especially important in this data). Habitats with submersed macrophytes were also
generally nutrient poor system. Tadei et al. ( 1998), in reported forAn. darlingi larvae from Belize by Manguintheir paper on ecological observations on anophelines in et al. ( 1996). Low abundance of An. darlingi in earlythe Brazilian Amazon, suggest that the higher species July could be a consequence of a generally lowerdiversity of mosquitoes at human impacted locations is incidence of this species in the dry season( Rubio-Paliscaused by increased input of nutrients from agricultural et al. 1997, Magris et al., unpublished data). Regardless
activities. of the paucity of larvae found, this species is clearly theThe main regional vector of malaria in the Upper dominant human biting anopheline in the area( Rubio-
Orinoco is An. darlingi. In our July survey we found this Palis 1996, Rubio-Palis et al., unpublished data).
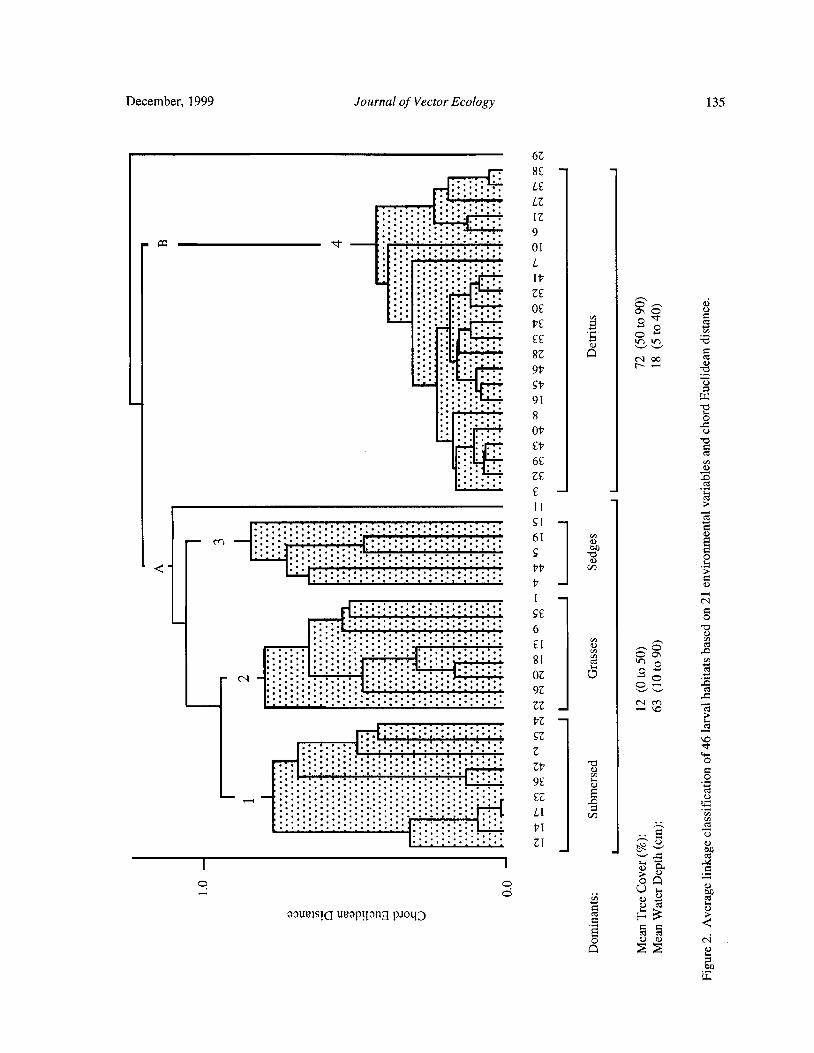
December, 1999 Journal of Vector Ecology 135
6Z
r 8£L
L£
LZ
9Z
Aa 01•ii - i:..i L
117
Z£
r Lb£ Y o o
CA
bo
9b b1!Sb
9i b8 0Ob
b ao
1EZ£ az
I[
Si
il3
cr1 --- S oWO
Q bb
bZ
N
S£ o6 0
I i o8[ m tt
Cz
ocv OZ e5 o
9Z
bZ
SZ VD
Zb aF I. 9£
o
az
bim
ZT _ to
0 0
p U F.
N aon
aaueasrQ uuapL[ ang RIND m H 3Q
on
w

136 Journal of Vector Ecology December, 1999
The most frequent larval species in the study area University of California, Davis, conducted the cationwas An. triannulatus. This species is regarded as a analysis. Remi was a great guide through the Ocamo
secondary vector of malaria by many authors from region. Yasmin Rubio-Palis and Leopoldo Villegas were
similar regions in Brazil( Tadei et al. 1983, Arruda et al. funded by Proyecto Control de Enfermedades1986, Deane et al. 1988, Oliveira-Ferreira et al. 1990, Endemicas/ Malariologia.
Povoa 1993) and Peru( Hayes et al. 1987). Nevertheless,
it has not been collected during intensive human landing REFERENCES CITED
and CDC trap catches inside human dwellings in thestudy area( Rubio- Palis et al., unpublished data), which American Public Health Association ( APHA). 1985.
documents its exophilic behavior. As shown in TABLE Standard methods for the examination ofwater and
3 and the frequency analyses ( TABLE 2), this species wastewater, 16th ed. APHA, Washington, DC.,
was mostly associated with submersed macrophytes 1, 193 pp.and sparse growth of graminoids in lagoons. This is in Arruda, M., M. B. Carvalho, R. Nussenzweig, M.agreement with Conn ( 1991) who reports collecting Maracic, W. Ferreira, and A. H. Cochrane. 1986.
larvae ofAn. triannulatus from ponds with floating and Potential vectors of malaria and different
emergent macrophytes. Tadei et al. ( 1983) report An. susceptibility to Plasmodium falciparum andtriannulatus to be among the most frequently occurring Plasmodium vivax in northern Brazil identified byspecies in their study area( northern Amazon Basin). immunoassay. Am. J. Trop. Med. Hyg. 35: 873-
Anopheles mediopunctatus was found only twice 881.
during the survey, each time in a shaded forest pool in Conn, J. 1991. Cytogenetic analysis of the polytene
detritus. Deane et al.( 1988) reported this species from chromosomes of Anopheles triannulatus( Diptera:
similar habitats, i.e., shaded forest pools and streams Culicidae) from western Venezuela. Genome 34:
with clear water. Anophelespunctimacula was collected 267- 272.
from lagoons with submersed macrophytes and detritus, Deane, L. M., C. D. Ribeiro, R. Lourenco de Oliveira, J.
which is in agreement with Gorham et al ( 1967) and A.Oliveira-Ferreira, and E. Guimaraes. 1988. StudyRejmankova et al.( 1998). Each, An. oswaldoi and An. on the natural history of malaria in areas of theperyassui were only found once in this survey, associated Rondonia State—Brazil and problems related to its
with submersed macrophytes and detritus, respectively. control. Rev. Inst. Med. Trop. Sao Paulo 30: 153-Rubio-Palis ( 1991) collected these species in ground 156.
pools with floating vegetation in shaded areas. Gorham, J. R., C. J. Stojanovich, and H. G. Scott. 1967.
Habitat description, as provided here and in other Clave ilustrada para los mosquitos anofelinos de
papers( Rejmankova et al. 1993, 1998), is based primarily sudamerica oriental. U.S. Department of Health,
on hydrology and aquatic vegetation present. One of the Education and Welfare, Public Health Service,
advantages of using the aquatic vegetation for Atlanta, Georgia. 64 pp.characterization of larval habitats is its integrating and Hall, T. F. 1972. The influence of plants on anopheline
therefore more permanent indication value. For example, mosquito breeding. Am. J. Trop. Med. Hyg. 21:stands of submersed Mayaca require certain water depth, 787- 794.
nutrients, salinity, light, and time to get established. By Hamilton, S. K. and W. M. Lewis, Jr. 1990. Basin
finding certain anopheline species repeatedly in habitats morphology in relation to chemical and ecologicaldefined by this submersed macrophytes, we can infer characteristics of lakes on the Orinoco River
specific chemical and physical characteristics of the floodplain, Venezuela. Arch. Hydrobiol. 119: 393-
habitat and its relative permanence. This is important 425.
especially in situations when time or other constrains Hayes, J., G. Calderon, R. Falcon, and V. Zambrano.
do not allow us to do repeated surveys of the same 1987. Newly incriminated anopheline vectors ofhabitats. human malaria parasites in Junin Department, Peru.
J. Am. Mosq. Contr. Assoc. 3: 418- 422.Acknowledgments Manguin, S., D. R. Roberts, R. G. Andre, E. Rejmankova,
and S. Hakre. 1996. Characterization ofAnopheles
We thank Mr. James Pecor, Walter Reed darlingi ( Diptera: Culicidae) larval habitats inBiosystematics Unit, for verifying the identifications of Belize, Central America. J. Med. Entomol. 33:
early stage larvae. Professor Justiniano Velasquez, 205- 211.
Central University, Venezuela, kindly assisted with the Merritt, R. W., R. H. Dadd, and E. D. Walker. 1992.identification of submersed macrophytes. Jae Kim, Feeding behavior, natural food, and nutritional

December, 1999 Journal of Vector Ecology 137
relationships of larval mosquitoes. Annu. Rev. Ann. New York Acad. Sci. 740: 396- 402.
Entomol. 37: 349- 376. Rubio-Palis, Y. 1991. Vector biology and malariaOliveira-Ferreira, J., R. Lourenco de Oliveira, A. Teva, transmission in western Venezuela. Ph.D. Thesis.
L. M. Deane, and C. T. D. Ribeiro. 1990. Natural University of London, 261 pp.malaria infections in the anophelines in Rondonia Rubio-Palis, Y. 1995. Observaciones sobre el patron de
state, Brazilian Amazon. Am. J. Trop. Med. Hyg. actividad hematofagica del vector de malaria
43: 6- 10. Anopheles darlingi en las poblaciones del sur de
Pan American Health Organization. 1998. Status of Venezuela. Bol. Dir. Malariol. San. Amb. 35: 66-
malaria programs in the Americas. XLVI Report. 70.
Washington DC; PAHO, 26 pp. Rubio- Palis, Y. 1996. Observations on the biting activityPodani, J. 1996. SYN-TAX 5. 02.Mac. Computer of different populations of the malaria vector
programs for multivariate data analysis on the Anopheles darlingi in southern Venezuela. J. Am.
Macintosh system. User' s Guide, Scientia Mosq. Contr. Assoc. 12: 461.Publishing, Budapest., 91 pp. Rubio- Palis, Y. and R. H. Zimmerman. 1997.
P6voa, M. 1993. Studies on malaria in Sierra do Navio Ecoregional classification of malaria vectors in the
region, Amapa State, Brazil. Ph. D. Thesis, Neotropics. J. Med. Entomol. 34: 499- 510.
University of London, 259 p. Rubio-Palis, Y.,M.Magris, L.Villegras, and C. Menares.
Rejmankova, E., H. Savage, M. Rodriguez, and D. 1997. Seasonal variation, biting activity andRoberts. 1992. Aquatic vegetation as a basis for entomological inoculation rate of Anopheles
classification ofAnopheles albimanus Wiedemann darlingi in the upper Orinoco River, Venezuela. J.
Diptera: Culicidae) larval habitats. Environ. Am. Mosq. Contr. Assoc. 13: 117.Entomol. 21: 598- 603. Sokal, R. R. and F. J. Rohlf. 1995. Biometry, 3rd ed. W.
Rejmankova, E., D. R. Roberts, R. E. Harbach, J. Pecor, H. Freeman, New York, 887 pp.E. L. Peyton, S. Manguin, R. Krieg, J. Polanco, and Steyermark, J. A., P. E. Berry, and B. K. Hoist. 1995.L. Legters. 1993. Environmental and regional Flora of the Venezuelan Guyana. Volume 1. Timber
determinants of Anopheles larval distribution in Press, Portland, 320 pp.northern Belize. Environ. Entomol. 22: 978- 992. Tadei, W. P., B. M. Mascarenhas, and M. G. Podesta.
Rejmankova, E., K. O. Pope, D. R. Roberts, M. G. Lege, 1983. Biologia de anofelinos amazonicos. VIII.
R. Andre, J. Greico, and Y. Alonzo. 1998. Conhecimentos sobre a distribuicao de especies de
Chatacterization and detection of Anopheles Anopheles na regiao de Tucurui- Maraba ( Para).
vestitipennis andAnopheles punctimacula( Diptera: Acta Amazonica 13: 103- 140.
Culicidae) larval habitats in Belize with field survey Tadei, W. P., B. D. Thatcher, J. M. M. Santos, V. M.
and SPOT satellite imagery. J. Vector Ecol. 23: 74- Scarpassa, I. B. Rodrigues, and M. S. Rafael. 1998.
88. Ecologic observations on anopheline vectors of
Roberts, D. R. and M. H. Rodriguez. 1994. The malaria in the Brazilian Amazon. Am. J. Trop.environment, remote sensing and malaria control. Med. Hyg. 59: 325- 335.

Journal of Vector Ecology 24( 2): 138- 153
Mosquito Control and Bacterial Flora in Water Enriched with
Organic Matter and Treated with Bacillus thuringiensis
subsp. israelensis and Bacillus sphaericus Formulations
Tram T. H. Nguyen, Tianyun Su, and Mir S. Mulla
Department ofEntomology, University ofCalifornia, Riverside, CA 92521- 0314, USA
Received 1 February 1999; Accepted I1 May 1999
ABSTRACT: Three tests were conducted during July 17 to October 30, 1998 to study the impact of twomosquitocidal microbial agents on mosquito larvae and their contribution to bacterial flora in aquatic
microcosms. Formulations ofBacillus thuringiensis subsp. israelensis( Bti) and Bacillus sphaericus strain2362 ( Bsph) were applied at various rates to outdoor tubs enriched with rabbit pellets and filled with
irrigation water from a reservoir. Mosquito larvae were effectively controlled by all treatments; themagnitude of initial and persistent control depended on materials and dosages applied. Bacterial flora were
assessed in the irrigation water as well as water in the enriched tubs before and after treatment with the
microbial agents. The irrigation water contained 800- 1000 total bacterial cells/ ml. The populations of total
bacteria and spore formers peaked on day 3 after enriching and filling the tubs, then declined progressivelyto the low levels at the end of the tests. After treatment, the numbers of Bti and Bsph spores in treated tubs
prevailed at a dosage-dependent manner, their populations peaked at three hours after treatment, and
declined progressively thereafter. The contribution of Bti and Bsph spores to the total bacterial flora was
negligible but significant to the counts of spore- forming bacteria. The gram- negative bacteria made upmore than 80% of the total bacterial flora during the test periods; and, of these, gram- negative rodsconstituted the greatest proportion, gradually increasing from the time of flooding to the end of the tests.Gram- negative cocci also occurred in relatively great proportion, but showed a reverse trend as comparedwith the gram- negative rods, declining gradually from pretreatment to the end of the tests. Gram-positiverods( spore formers), including Bti and Bsph, occurred in low numbers in all the tests but increased slightlyin treated tubs due to the addition of Bti and Bsph spores. Gram-positive cocci occurred occasionally insome water samples.
Keyword Index: Bacillus thuringiensis subsp. israelensis, Bacillus sphaericus, bacterial flora, spore-forming bacteria, Culex mosquitoes.
INTRODUCTION Karch et al. 1991, Mulla et al. 1999). The most
studied mosquitocidal strains of Bsph are strainsBacillus thuringiensis subsp. israelensis de Barjac 1593, 2297, and 2362 ( Singer 1990). These strains
Bti) and Bacillus sphericus Neide ( Bsph) are two have high activity against larvae of Culex mosquitoes.entomopathogenic gram-positive, aerobic, endospore- Despite its more limited host range, Bsph providesforming bacteria that produce parasporal proteinaceous more persistent larvicidal activity than Bti undercrystals toxic to mosquito larvae. Bti has been evaluated polluted water conditions ( Yap 1990; Mulla 1991;for mosquito and black fly control in many countries Mulla et al. 1997, 1999). Bacillus sphaericus strainde Barjac and Sutherland 1990, Mulla 1990). It was 2362 has been more extensively studied than the other
registered in 1980 by the US- EPA for use against strains and was registered by the US- EPA in 1991 formosquitoes and black flies and has been found to be mosquito control. It has been successfully tested andeffective against these two groups of insects in many used in mosquito control programs in Spain ( Arandatypes of habitats ( de Barjac and Sutherland 1990, and Eritja 1992), Thailand ( Mulla et al. 1997, 1999),

December, 1999 Journal of Vector Ecology 139
United States ( Mulla 1991), West Africa ( Karch et MATERIALS AND METHODSal. 1991, Skovmand and Bauduin 1997), and other
countries. Test Facilities
Mosquito larvae breed in a variety ofaquatic habitats Three tests were conducted during the summer andand their food consists of microorganisms, detritus, fall seasons, from the middle of July to the end ofalgae, protozoans, bits of leaves, and small living and October 1998. In total, 24 fiberglass tubs measuring 1. 0dead invertebrates( Clements 1992, Merritt et al. 1996). x 1. 0 x 0. 4 m deep, placed in an open sunlit area at theBacteria are dominant microorganisms in the aquatic Aquatic and Vector Control Research Facilitynatural habitats of mosquito larvae. Bacterial densities ( Midgeville, University of California, Riverside) werehave been estimated by direct microscopic count used. The tubs were enriched with rabbit pelletsmethod( DMC), and have been reported to range from ( Brookhurst®, Brookhurst Mill, Riverside, CA, crude
18. 8- 65. 4 x106
cells/ m1 in surface microlayer of water protein 17%) at the rate of 100 grams per tub( 0.04%) to
in marsh habitats where anopheline larvae were found. facilitate continuous and sustained oviposition by wildIn subsurface water, counts have ranged from 3. 8- gravid mosquitoes. The tubs were then filled to a depth
14. 0 x106
cells/ml ( Walker and Merritt 1993). In of 30 cm( 236 liters) ofwater from an irrigation reservoir.
another study, two different methods of bacterial A constant water level was maintained in each tub byassessment yielded different cell counts. Smith et al. float valves. Within a day or two after flooding in1998) collected surface water samples from a fresh warmer months, wild populations of Culexstigmatosoma
water marsh habitat supporting anopheline larvae and Dyar, Culex quinquefasciatus Say, and Culex tarsalismade bacterial counts using the DMC and culturing on Coquillett oviposited nightly in these tubs. During thetrypticase soy agar medium. The culture method course of the tests, the tubs also supported the production
yielded bacterial density in the range of 1. 0- 1. 5 x105
of algae and some aquatic macroinvertebrates, such as
cells/ml, while the DMC method gave bacterial counts ephydrid flies, chironomid midges, mayfly naiads,from 9. 7 x
105- 1. 3 x 10' cells/ ml. Bacteria constitute a dragonfly naiads, and others. The tubs were treated withmajor source of food of mosquito larvae. Using acridine Bti formulation on day 7 postflooding in tests 1 and 2 ororange and epifluorescence microscopy, Nilsson( 1987) both Bti and Bsph formulations on day 10 postfloodingfound that the number of bacteria per gut in some Aedes, in test 3, when 3rd- 4th- instar mosquito larvae were
Culex, Culiseta, and Anopheles species larvae ranged present in large numbers. The check and treatment tubs
from 6. 68 x106- 2. 18 x
107. In field-collected Aedes were assigned randomly before flooding using six( teststriseriatus Say and Anopheles quadrimaculatus Say 1 and 2) or four( test 3) replicates. After termination of
4th- instar larvae, the number ofbacteria averaged 2. 2 x a given test, the tubs were cleaned and kept dry for three106
and 2. 0 x106
cells/gut, respectively ( Walker et al. to seven days before starting a new test.1988). In another study, four food types consisting ofbacteria, detritus, euglenoid protozoans, and algae were Bacterial Insecticides and Application
found in the gut of 4th- instar larvae of Coquillettidia Three preparations of Bti and Bsph ( provided byperturbans Walker. Bacteria were found in highest Abbott Laboratories, North Chicago, IL) were used in
numbers averaging 7. 07 x105
cells/ gut. Next in numbers the tests. The water dispersible granules( WDG) of Bti
found were detrital particles, euglenoid protozoans, and ( VectoBac WDG, ABG-6490, Lot# 30- 067- BR, 4,000
algae( Merritt et al. 1990). ITU/mg, received on 7/ 10/ 1997) were applied at theNatural populations of bacteria play an important rates of 0. 27, 0. 53, and 1. 1 lb/ ac, equaling 25, 50, and
role in the breakdown of organic matter and 100 mg per tub, respectively. The Bsph WDGeutrophication of aquatic habitats and constitute a major formulation ( VectoLex WDG, ABG 6491, Lot # 32-
source of food for mosquito larvae. We initiated the 094- BR, 600 ITU/mg, received 8/ 5/ 1998) was appliedpresent studies to determine: 1) the relationship between at the rates of 0.05 and 0. 1 lb/ac, equaling 5 and 10 mgnatural microbial flora and mosquito breeding; and 2) per tub, respectively. The commercial product VectoLexthe impact of various mosquitocidal formulations of Bti CG( corn grit)( Lot# 29- 853- N8, 50 ITU/mg, receivedVectoBac) andBsph( VectoLex) onbacterialabundance 8/ 4/ 1998) was applied at the rates of 1. 4 and 2. 7 lb/ ac,
in microcosms sustaining heavy breeding of natural equaling 125 and 250 mg per tub, respectively.populations of mosquitoes. The VectoBac and VectoLex The WDG formulations were brownish fine sized
formulations were applied to outdoor tubs enriched with granules with a loose appearance that readily dispersedorganic matter, and the population trends of mosquito in water by gentle stirring or shaking. Before application,larvae, total bacterial flora, spore- forming bacteria, as the WDG formulations were suspended in distilled
well as Bti and Bsph spores were studied over time. water at 1% for VectoBac WDG or 0. 1% for VectoLex

140 Journal of Vector Ecology December, 1999
WDG. The required aqueous aliquots based on the surface( not surface film) and about 0. 5- 1.0 cm abovetreatment rates were slowly applied to the corners and the bottom of the tub. From each tub, five samples wereedges of the tubs using a 5 ml pipette. The required collected at the four corners and middle for the top andamounts of the VectoLex CG granules were broadcast bottom of each tub. The samples from the top or theby hand to the corners and edges of the tubs getting good bottom of all replicates were composited in 250 mlcoverage. sterile flasks, each composite sample amounting to 150
ml( tests 1 and 2) or 100 ml( test 3) of water from six orAbiotic Water Parameters four replicates of each treatment as well as check. The
Maximum-minimum water temperatures were water samples were kept in a refrigerator at 5° C andmeasured during each test period by leaving a minimum- processed a few hours after collection.maximum thermometer submerged in the water located Media for Bacterial Isolation: To estimate thein the center of the tub arrangement. In every tub, bacterial densities in water samples, nutrient agar mediumdissolved oxygen ( DO, ppm), electrical conductivity and selective media containing antibiotics were usedEC, µ S), and salinity ( ppt) were measured by YSI ( Jones 1970, Yousten et al. 1985). Nutrient agar medium
Model 85 Handheld Water Quality Meter on every ( NA medium: 2. 3% Difco nutrient agar) was used forsampling day. The pH values of the tub water were identifying diversity of total bacteria, including spore-determined by pH test strips. forming bacteria. For recovery of Bti spores, we used
selective medium ( NYPC medium) containing 2. 3%Mosquito Control nutrient agar, 0.05% yeast extract, 0.01% polymixin B
In VectoBac WDG tests ( 1 and 2), immature sulfate, 0.0001% chloramphenicol. Selective nutrient-mosquitoes( larvae and pupae) were sampled just before yeast medium with streptomycin( NYST medium: 2.3%treatment( day 0 or day 7 postflooding) and on days 3, nutrient agar, 0. 05% yeast extract, 0.01% streptomycin)
7, and 14 posttreatment. In the test of VectoBac and was used for recovery of Bsph spores. The media wereVectoLex( test 3), immature mosquitoes were sampled prepared, autoclaved, and poured into Petri dishes threejust before treatment( day 0 or day 10 postflooding) and days before plating out water samples onto the media.on days 3, 7, 14, 21, and 28 posttreatment. Five dip This time was necessary for drying the medium surfacesamples were taken from each tub; one dip from each and detecting contamination of Petri dishes if any.corner and one from elsewhere where mosquito larvae Dishes showing any contaminations were discarded.were noted in large numbers. The mosquito immatures Bacterial culture and count: Prior to plating thewere counted and divided into three categories as early water samples onto microbiological media, a 1. 0-mlinstars ( 1st and 2nd), late instars ( 3rd and 4th), and aliquot from each water sample was serially diluted in 9pupae. ml autoclaved water. For detection of bacterial cells,
dilutions ranged from 10- 104 times. Three Petri dishesBacterial Assessment containing 20 ml NA medium were inoculated with 0. 1
Water Samples: To study bacterial density in tests ml aliquot of each serial dilution, and incubated at 28-1 and 2, both top and bottom water from assigned check 30° C for 48- 72 hours. Autoclaved water for dilutionand treated tubs was collected on day 4 pretreatment was also plated on NA as a control. After the incubationday 3 postflooding), just before treatment ( day 0 period, colony- forming units ( CFUs) were counted
posttreatment or day 7 postflooding), and on days 3, 7, visually and characterized by color, form, elevation,and 14 posttreatment. Additionally, in test 3 we took edge, and diameter. Cultivable cell densities weresamples of water flowing from the float valves to averaged from the CFUs and adjusted for dilutions anddetermine bacterial density in the water coming from inoculum volume and were expressed as CFUs/ml. Cellthe reservoir. Water from assigned check and treated morphology and gram- stained cells were examinedtubs was collected on day 10 pretreatment( 3 h after start under light microscopy at 400x and under oil immersionof filling), day 7 pretreatment( day 3 postflooding), just at 1, 000x. Bacteria were categorized as rods prevailingbefore treatment( day 0 or day 10 postflooding), and on singly or in chains and cocci consisting of monococci,days 3, 7, 14, 21, and 28 posttreatment for study of diplococci, and tetracocci.bacterial density. For determination of Bti and Bsph For determination of the densities of spore- formingspores soon after treatment with VectoB ac and VectoLex bacteria including Bti and Bsph, water samples wereformulations, additional water samples from check and pasteurized at 65° C for 30 min to kill vegetative andtreated tubs were also collected at three hours after non- spore- forming bacteria before diluting and plating.treatment. Using sterile pipettes fitted with a pro-pipette, Pasteurized water samples were spread on NA medium5 ml of water was collected at 0.5- 1. 0 cm below the for recovery of total spore- forming bacteria as well as on

December, 1999 Journal of Vector Ecology 141
selective media NYPC and NYST for recovery of Bti that demonstrated equal efficacy. The larval densities inand Bsph spores. Colonies were counted visually and the checks declined naturally on day 14 posttreatment,characterized 48- 72 hours after incubation at 28- 30° C. and no control due to the treatment was indicated on thisCell morphology and gram- stained cells were observed sampling day.microscopically. In a few cases where the identificationof cells and spores was uncertain, isolated bacterial Bacterial Assessmentcolonies were removed for culturing on agar slants at Total bacteria: Since the bacterial flora and trends30° C for 24 hours, and then were kept at 5° C to reconfirm were essentially the same in tests 1 and 2, we haveidentification by microscopic examination. combined the data of the two experiments and report
their averages. The average numbers of total bacteriaData Analysis
CFUs/ m1) in top water cultivable on NA mediumAverage mosquito larval densities( larvae/dip) and peaked on day 4 pretreatment( day 3 postflooding), and
average bacterial densities( CFU/ml) in the check and declined sharply by 91- 93% on day 0 pretreatment( daytreated tubs were analyzed for significance by one factor 7 postflooding). The total bacterial counts declinedANOVA ( Scheffe F test) for repeated measurements gradually afterward to the end of the test periods( day 14Abacus Concepts, Inc. 1987). For mosquito control, posttreatment) when the reductions were 97-98% asthe reduction in the population of immature mosquitoes compared with the peak populations on day 4was calculated using Mulla' s formula ( Mulla et al. pretreatment( Fig. 1B). The changing pattern ofbacterial1971):(% R)= 100- [( C 1/ T1) x( T2/C2)] x 100, where numbers in bottom water was similar to that in top waterCl= mean number of larvae in control pretreatment, T1 ( Fig. 1B).
mean number of larvae in treated pretreatment, T2= As to the impact of microbial treatment on bacterialmean number of larvae in treated posttreatment, and C2 flora, the total bacterial counts in the check tubs were
mean number of larvae in control posttreatment. slightly but significantly lower than in treated tubs withthe highest rate of VectoBac WDG( 1. 1 lb/ac) on day 3
RESULTS posttreatment in top water. The same situation held trueon day 7 posttreatment. No treatment-related differences
VectoBac WDG Tests( Test 1 and 2) occurred on day 14 posttreatment. In bottom water,treatment- related differences in total bacterial counts
Abiotic Water Parameters were only noted on day 7 posttreatment, where the tubsDuring the course of these tests, maximum water treated with the middle rate ofVectoBac WDG( 0.53 lb/
temperatures ranged from 32. 2- 37. 8° C and minimum ac) contained more spore formers than check and otherranged from 22. 1- 23. 3° C. No significant differences treatments( Fig. 1B).were detected in abiotic water parameters( pH, DO, EC, Sporeforming bacteria: In water samples collectedand salinity) among various treatment regimens on on day 4 pretreatment( day 3 postflooding), we recoveredevery sampling day( data omitted). But it was noted that some spore- forming bacteria, which produced bigoxygen tension due to microbial degradation of the elongated ovoid spores. These bacteria occurred singlyorganic matter reached an almost anoxic level on day 3 or in long chains. The average numbers of all spore-postflooding. Oxygen tension increased after this initial forming bacteria in top water increased gradually fromperiod when the organic matter decomposition had day 4 pretreatment, peaked on day 3 posttreatment, andpassed the peak period. declined thereafter( Fig. 2A). The changing pattern in
populations of total spore formers in bottom water wasMosquito Control similar to that in top water( Fig. 2A).
The population trends of mosquito larvae in test 1 With regard to the impact of VectoBac WDGand 2 were essentially the same, and the data of the two treatment on total spore- former counting after treatment,experiments are combined and the averages are presented the spore formers in treated tubs with the highest rate ofFig. 1A). Immediately prior to treatment, larval densities 1. 1 lb/ac were significantly higher than check and other
in all the assigned check and treatments were statistically treatments in top water on days 3 and 14 posttreatment.the same. On days 3 and 7 posttreatment, all three rates No treatment-related differences were indicated on day0.27, 0.53, and 1. 1 lb/ac) of VectoBac WDG yielded 7 posttreatment ( Fig. 2A). In bottom water, the same
good control. The number of larvae was reduced by impact was only seen on day 3 posttreatment. No94. 3, 94.9, and 72. 7%( day 3) and 93. 6, 91. 3, and 68. 7% treatment- related differences were indicated on theday 7), respectively. It was clear that the lower rate following sampling days( Fig. 2A).0.27 lb/ ac) was less effective than the two higher rates Bti spores: After treatment with VectoBac WDG,
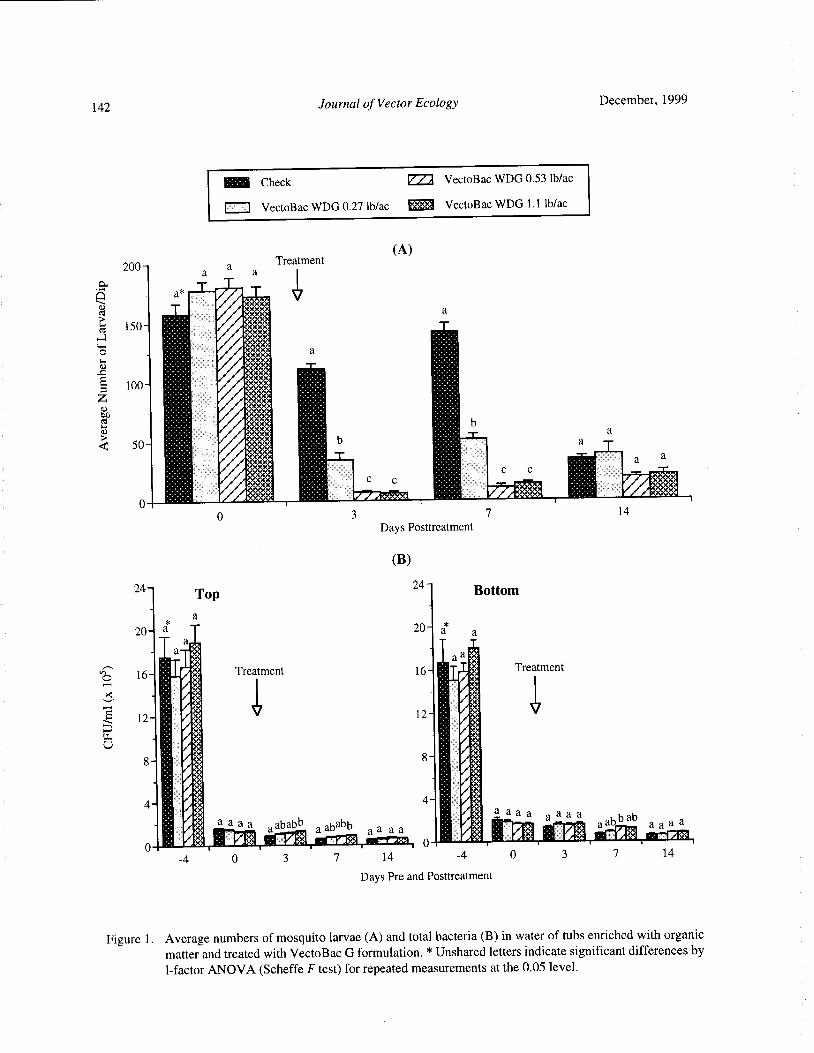
142 Journal of Vector Ecology December, 1999
Check VectoBac WDG 0. 53 lb/ ac
1 VectoBac WDG 0.27 lb/ac +'+'+ t1 VectoBac WDG 1. 1 lb/ ac
200- a
aa
TreatmentA)
vs
150- F}++«++#+'
1 I •++#+
Ci
G 100— F}+++4+++i
y i
I
0 3 7 14
Days Posttreatment
B)
24Top
24Bottom
a
20 a 20 a a
a,
a I4 aa; 4;a i
0 16 /:; Treatment 16 4 Treatment,....
t.,< r
iNI/
12 I F++, 12 f+ih* i
II$+i i
8
i
4 I'4 I+ a a asa b aa a a ababb
aababb a a a
a s+ aa
s +
a+ a ab
aba a a
fa, I44 P44 ,## i+I++i 4 I .0
a
n • . i#i . a. • '«. F1.. i i!a!•
4 0 3 7 14 4 0 3 7 14
Days Pre and Posttreatment
Figure 1. Average numbers of mosquito larvae( A) and total bacteria( B) in water of tubs enriched with organicmatter and treated with VectoBac G formulation. * Unshared letters indicate significant differences by1- factor ANOVA( Scheffe F test) for repeated measurements at the 0.05 level.

December, 1999 Journal of Vector Ecology 143
Check VectoBac WDG 0.53 lb/ ac
VectoBac WDG 0. 27 lb/ ac 5•1::;. VectoBac WDG 1. 1 lb/ ac
A)
5Top
6Bottom b
A 4Treatment 5- Treatment «:«.
k*Mi 4i
a..A 4- ji, a t'
3 a : t aF «. a
a +#*i d a I*a
abb 3- bd +. /.;*,
M++i f•
24* : 1110. 4.4+'
abc + 4; a a
b a a *: ia
ab
c.*: a a a
F *+*+ a a F+ i b i*+i a a•
C aIV Ill
IIIf#*
1 a a
faa i fia f4 4/
M*#* f#*
i i
01* Ou. aY*+i a+
0i
7 14 4 0 3 7 14
k Days Pre- and Posttreatment
E
E B)
U
2. 5 Top2.5
Bottom
c
2 2
iiig
1. 5 b 1. 5b +++.
r+*+++•
1F+*+*+•
ba ++;+;
0.5ab«++.
a +++ a +«« a '++ i a ++; a s *+*+*i
3 7 14 3 7 14
Days Posttreatment
Figure 2. Average numbers of spore- forming bacteria including Bti( A) and Bti spores alone( B) present in waterenriched with organic matter and treated with VectoBac WDG formulation( tubs flooded 7 days before
treatment, day 0 indicates just before treatment). * Unshared letters indicate significant differences by1- factor ANOVA( Scheffe F test) for repeated measurements at the 0.05 level.

144 Journal of Vector Ecology December, 1999
we isolated Bti spores on NA as well as on selective were statistically the same. Significant mosquito larvalNYPC medium in all treated tubs. Overall, Bti spore control was achieved on days 3( 75.7- 97. 4% reductions)counts in treatment samples gradually decreased from and 7 ( 50.0- 65. 0% reductions) posttreatment in all
day 3 posttreatment to the end of the tests in both top and treatments. On day 14 posttreatment, only VectoLexbottom water( Fig. 2B). In top water of treated tubs, the WDG gave partial but significant control( 50.0- 59. 0%average counts ofBti spores showed adosage- dependent reductions) at both rates. Other treatments ( VectoBacmanner on every sampling day posttreatment. The high WDG and VectoLex CG) did not show any more control.rate of 1. 1 lb/ ac contributed significantly more Bti On days 21 and 28 posttreatment, no more control wasspores than the middle rate of0.53 lb/ ac and the low rate indicated in all treatments( Fig. 4).of 0. 27 lb/ ac. No differences were indicated betweenthe middle and low rates. Similar trends were noted in Bacterial Florabottom water. On day 3 posttreatment, significant Total bacteria: In the third test, in addition todifferences in Bti spore counting were detected among assessing the bacterial population in the water enrichedthree treatment rates. On day 7 posttreatment, significant with rabbit pellets, we also took samples of water
difference only showed between the lowest and the flowing from the float valves, which contained very fewhighest rates. The situation on the last sampling day was bacteria averaging 0.8- 1. 0 x 103 cells/ml.the same as that in top water on this sampling day, i. e., In top water of assigned check and treated tubs, thethe significantly higher count of Bti spore was average numbers of total bacteria increased from veryencountered in the highest rate of 1. 1 lb/ ac ( Fig. 2B). low counts on day 10 pretreatment( at 3 h after start of
Gram-stained bacteria: As to the bacterial types in filling) to the peak populations on day 7 pretreatment.top and bottom water, it was noted that gram-negative On day 0( before treatment), bacterial densities decreasedGr-ve) bacteria constituted 90-99% of the populations sharply by 81- 83% as compared with day 7 pretreatment.
on different sampling days during the course of the two From day 3 posttreatment to the end of the test, thetests. In general, while Gr-ve rods increased from 34- number of total bacteria continued to decrease slightly.52% on day 4 pretreatment to 81- 93% on day 14 On day 28 posttreatment, when the test was concluded,posttreatment. The Gr-ve cocci proportion decreased the reductions in total bacterial populations were 96-steadily from 48- 66% to 5- 16% during the same period 97% as compared with peak populations on day 7of time. Gram-positive( Gr+ve) rods determined in our pretreatment ( Fig. 5). The overall changing trends oftests were spore- forming bacteria, to which Bti belongs, total bacteria in bottom water were essentially the sameoccurred in very low density and never exceeded 4% of as those in top water( Fig. 5).the total bacterial flora. Gr+ve cocci occurred only After treatment with VectoBac and VectoLexoccasionally in some water samples, constituting 4- 9% formulations, differences in bacterial population declinesof total bacterial flora( Fig. 3). were noted between check and treatments. In the top
water on days 3, 7, 14, and 21 posttreatment, the totalVectoBac and VectoLex Test( Test 3) bacterial counts in check tubs were slightly but
significantly lower than in treated tubs. On the lastAbiotic Water Parameter sampling day ( day 28 posttreatment), the tubs treated
Water temperatures during the 39 days ofevaluation with VectoLex WDG at 0. 1 lb/ac contained more totalwere considerably cooler than the previous two tests in bacteria than the check and other treatments( Fig. 5). Asthe summer, ranging from 12. 2- 17. 8° C and 25. 6- 27. 8° C to the impact of microbial treatments on total bacterialfor the minimum and the maximum, respectively. As in counts in bottom water, some slight but significantthe previous tests, no significant differences were differences were indicated on some occasions. On daydetected in abiotic water parameters( pH, DO, EC, and 3 posttreatment, the tubs treated with VectoBac WDGsalinity) among various treatment regimens on every showed higher bacterial counts than the check andsampling day( data omitted). But as noted in test 1 and VectoLex WDG at the low rate( 0.05 lb/ ac). On day 72, the oxygen tension reached to an almost anoxic level posttreatment, however, all treatments showed higheron day 3 postflooding, then increased after this initial total bacterial counts than the check. On day 14period upon the completion ofdecomposition ofenriched posttreatment, the tubs treated with high rates oforganic materials. VectoLex products( both WDG and CG) showed higher
total bacterial counts than the check and other treatments.Mosquito Control The treatment of VectoLex WDG at the high rate still
In this test, the numbers of total larvae on day 0 showed higher counts of total bacteria than check andbefore treatment) in assigned check and treatments other treatments on days 21 posttreatment. The situation

December, 1999 Journal of Vector Ecology 145
Gr-ve rods El Gr-ve cocci Gr+ve rods Gr+ve cocci
100—Top Bottom•
11111 . II180
A) -
1I .1 1
0 \ ` \ \ \ ii . ' '100-
80-
B) I
ii . ' 'x100-
111180— I I I I
0 \ \ \ k. ., k \100-
1III :JrIi' . i— 77 14 4 0 3 7 14
Day Pre and Posttreatment
Figure 3. Proportion of gram-negative( Gr- ve) and gram-positive( Gr+ve) bacteria present in water from untreatedA) and treated tubs with VectoBacWDG formulation at 0.27 lb/ac( B), 0. 53 lb/ac( C), and 1. 1 lb/ac( D)tubs flooded 7 days before treatment, day 0 indicates just before treatment).
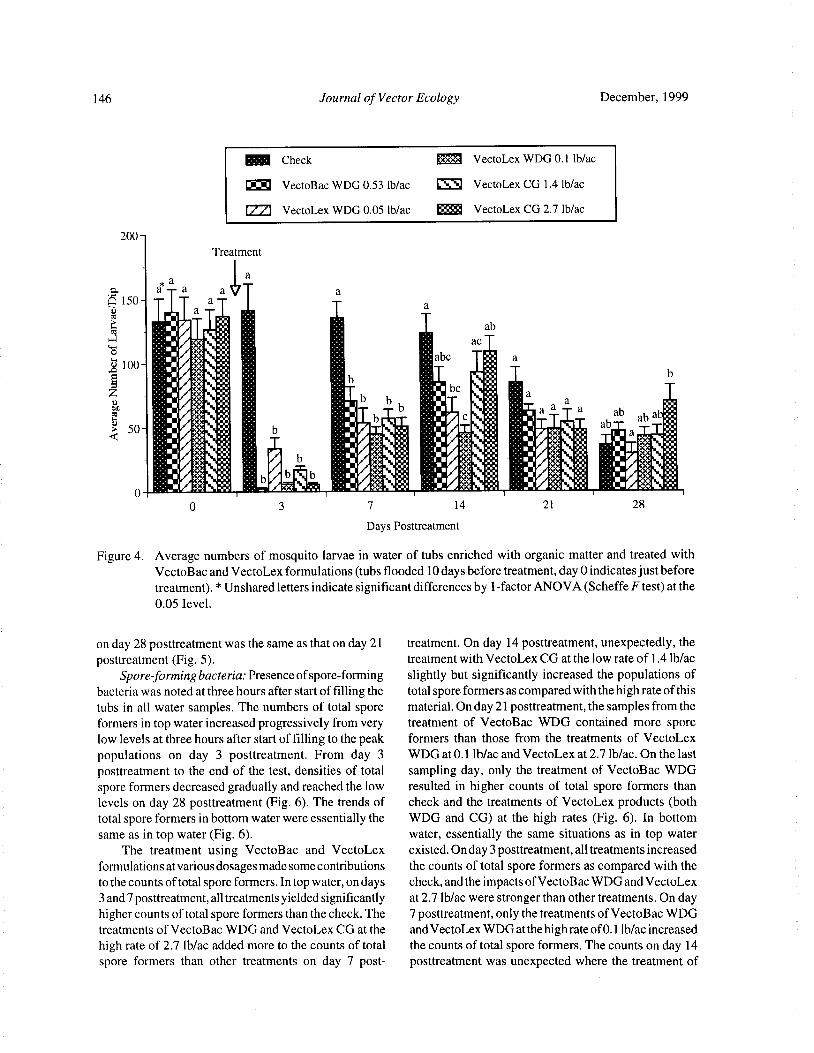
146 Journal of Vector Ecology December, 1999
Check VectoLex WDG 0. 1 lb/ ac
VectoBac WDG 0. 53 lb/ ac CK-9 VectoLex CG 1. 4 lb/ac
VectoLex WDG 0.05 lb/ ac ? ;: VectoLex CG 2. 7 lb/ac
200-
Treatment
a
p a a aJ1! a
150- — aa
ma1110 11..
ab
100- /;; .•
I V.: i be :
o iI//+* •'• i b ::: i• a•':
I ;: ':} b i c :•' :: 1
a aab
aba.:•:
ab
i+ b11.+., ::.
14* fir :.+:4,* ::
1 a ''••:
0 3 7 14 21 28
Days Posttreatment
Figure 4. Average numbers of mosquito larvae in water of tubs enriched with organic matter and treated with
VectoBac and VectoLex formulations( tubs flooded 10 days before treatment, day 0 indicates just beforetreatment).* Unshared letters indicate significant differences by 1- factor ANOVA( Scheffe F test) at the0.05 Ievel.
on day 28 posttreatment was the same as that on day 21 treatment. On day 14 posttreatment, unexpectedly, theposttreatment( Fig. 5). treatment with VectoLex CG at the low rate of 1. 4 lb/ ac
Spore-forming bacteria: Presence ofspore- forming slightly but significantly increased the populations ofbacteria was noted at three hours after start of filling the total spore formers as compared with the high rate of this
tubs in all water samples. The numbers of total spore material. On day 21 posttreatment, the samples from theformers in top water increased progressively from very treatment of VectoBac WDG contained more spore
low levels at three hours after start of filling to the peak formers than those from the treatments of VectoLex
populations on day 3 posttreatment. From day 3 WDG at 0. 1 lb/ac and VectoLex at 2.7 lb/ ac. On the last
posttreatment to the end of the test, densities of total sampling day, only the treatment of VectoBac WDGspore formers decreased gradually and reached the low resulted in higher counts of total spore formers than
levels on day 28 posttreatment( Fig. 6). The trends of check and the treatments of VectoLex products ( both
total spore formers in bottom water were essentially the WDG and CG) at the high rates ( Fig. 6). In bottom
same as in top water( Fig. 6). water, essentially the same situations as in top waterThe treatment using VectoBac and VectoLex existed. On day 3 posttreatment, all treatments increased
formulations at various dosages made some contributions the counts of total spore formers as compared with the
to the counts of total spore formers. In top water, on days check, and the impacts ofVectoBac WDG and VectoLex
3 and 7 posttreatment, all treatments yielded significantly at 2. 7 lb/ ac were stronger than other treatments. On dayhigher counts of total spore formers than the check. The 7 posttreatment, only the treatments of VectoBac WDGtreatments of VectoBac WDG and VectoLex CG at the and VectoLex WDG at the high rate of0. 1 lb/ ac increased
high rate of 2.7 lb/ac added more to the counts of total the counts of total spore formers. The counts on day 14spore formers than other treatments on day 7 post- posttreatment was unexpected where the treatment of

December, 1999 Journal of Vector Ecology 147
Check 1:0:41 VectoLex WDG 0. 1 lb/ac
VectoBac WDG 0.53 lb/ac ® VectoLex CG 1. 4 lb/ ac
VectoLex WDG 0.05 lb/ac VectoLex CG 2.7 lb/ ac
14- Top Treatment
A
12-a a
a
aaab
10-
8 111iro.i
4- ICI l * 11,0I. ot4+i
11I:;; ; a aaaaa bbb bb bccch 2
1 **+ a bbe c c d cdCD t.4 '' .:-./:;.; I': ' + •
a. , , be -..„ be be b
aa a a a a :
r+/«## •' /t4*i 1 $ 1._: eon::
b.!!}# a s
aft a
U• 14-
Bottom
12-a a Treatment
a
10-
vL,vi
8- h#i
6- 0;+ o
r ••. '• aaaaaa
a baabaab bcbcc
2- r
L1F M+ II0 +•.1 +
4:4 :•I!#L ._ L -. L/ 4#•. .' mil t+t;._ .' N f+i._ .'-, iA. 1+# L,.
10 7 0 3 7 14 21 28
Days Pre and Posttreatment
Figure 5. Average numbers of total bacteria present in water of tubs enriched with organic matter and treated withVectoBac and VectoLex formulations( tubs flooded 10 days before treatment, day 0 indicates just beforetreatment).* Unshared letters indicate significant differences by 1- factor ANOVA( Scheffe F test) at the0.05 Ievel.
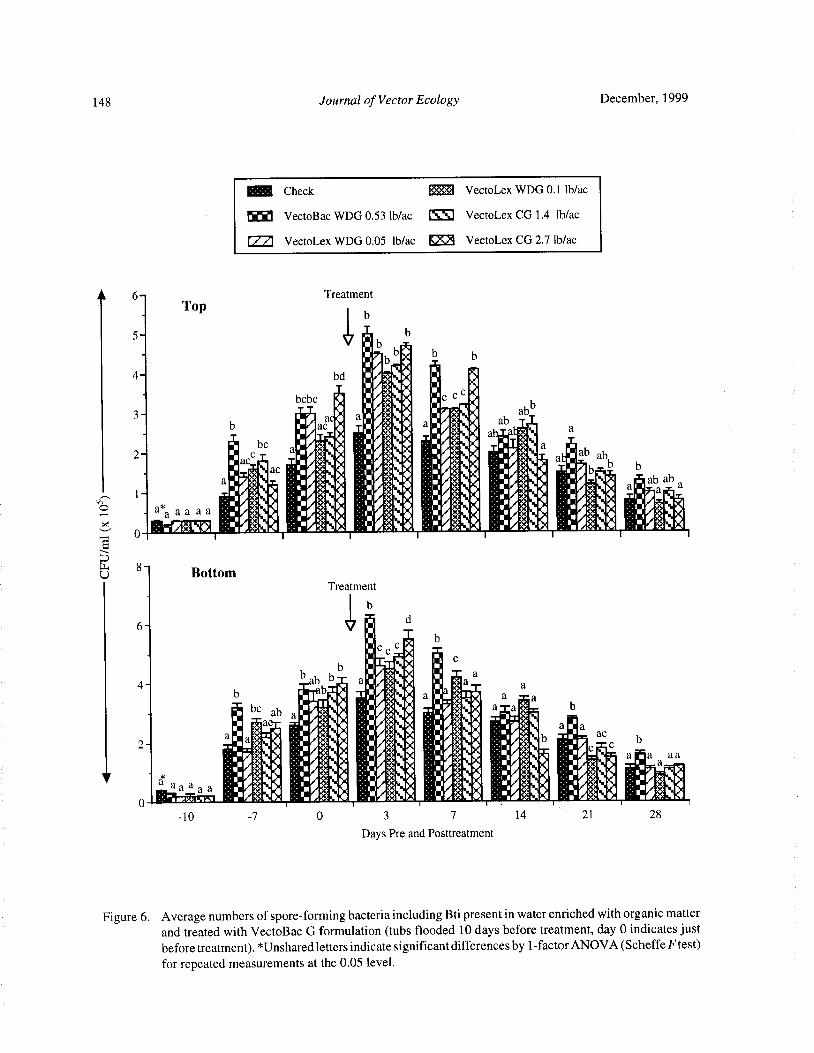
148 Journal of Vector Ecology December, 1999
Check 4 VectoLex WDG 0. 1 lb/ ac
VectoBac WDG 0.53 lb/ ac ® VectoLex CG 1. 4 lb/ ac
VectoLex WDG 0.05 lb/ac ® VectoLex CG 2.7 lb/ac
A 6— Treatment
Topb
5- b1. b
bbWb b
4- bd ;/;+; s' 0 .'h.A . ; 1
bcbc * 11/+'+ NA i'c cC.3- 1#
aim' :. • ' 1..+± o.1
b 1 ac r, 1: AV a' 1 : , 1 ab , a
1 bea
1 , k1' •:. 1 ++ i
r+i, *1 : t+`
2- 1 c1: :
410 ..• *. 8*1 ...; + t+L1 a1 ab
ac 1 1 +, :.:1; «+± • AM , a. 1
ab
i+: o • «
ti 1F : 1r«+N O 1 '..' ++ i _1' 7' 1)
b b
F+tiaC :
ice+ ' i '. I 1« '
iJEIFJiiJtIi :. « a '.'
I 1« alb a
III' 1 , «+ *1 1
F+t 1
5
u8-
BottomTreatment
b
6- ind
1lccc _ b
1 11b I•
I. c
b b i ;+*:
ill' a
4*i a4- a . a il t 1i +++ a
b I'b Iii . +;: a
I. :;IN a
beab a 1 4): ::rii• ',
APAil
i .:-.i*
mm Ma
a +i7 ., «+ I 11,++ ::: . 0441, aC
2- i •. i1 *;+ 4 . :* rb
aa a a
a a -:-
e Nee -IPAID :.:101ei ': j1F irAll '. III«++1
P :•-11 , W # .. 10t
10 7 0 3 7 14 21 28
Days Pre and Posttreatment
Figure 6. Average numbers of spore- forming bacteria including Bti present in water enriched with organic matterand treated with VectoBac G formulation( tubs flooded 10 days before treatment, day 0 indicates justbefore treatment).* Unshared letters indicate significant differences by 1- factor ANOVA( Scheffe F test)for repeated measurements at the 0.05 level.

December, 1999 Journal of Vector Ecology 149
VectoLex CG at the high rate of 2.71b/ ac showed lowest density in the check declined drastically due to naturaltotal spore former counts. On days 21 and 28 phenomena, and larval abundance in the treated was notposttreatment, the treatment of VectoBac WDG still markedly different from that in the check. Because ofincreased the counts of total spore formers as compared the natural decline, it cannot be concluded with certaintywith check and other treatments( Fig. 6). whether the low prevalence of larvae in treatments was
Bti and Bsph spores: In this test, as expected, no Bti due to Bti or natural decline ( lack of oviposition) orand Bsph spores were found in any samples collected both.
before treatment, and the samples from check after In another test, low dosages of the VectoLex WDGtreatment. However, after treatment with VectoBac formulation produced a persistent control of larvae up toWDG and VectoLex formulations, the spores were 14 days posttreatment. But in this test, as before, therecovered on selective media. In top water samples, the larval abundance from day 21 posttreatment decreasedspore counts of Bti and Bsph were the highest at three substantially in the check approximating that of larvaehours after treatment, then declined progressively to in the treatments. This declining trend of larvae as avery low levels at the end of the test( Fig. 7). The similar result of reduced oviposition is partly responsible for thepopulation trends ofBti and Bsph spores were indicated low larval abundance in both the treated and untreatedin bottom water( Fig. 7). regimens.
With respect to the spore counts of Bti and Bsph Embarking on the present studies, we hypothesizedin the check and various treatments, in top water, the that mosquito ovipositional activity and, in turn, larvaltubs treated with VectoLex WDG at the low rate of0.05 abundance are a function ofthe decomposition of organiclb/ ac contained fewer spores than those treated with matter and abundance of bacterial flora. Since weVectoBac WDG at three hours posttreatment. On the enriched the tubs with rabbit pellets prior to filling, wefollowing sampling days, highest spore counts were hypothesized that drastic changes in bacterial flora willencountered in the treatment of VectoBac WDG( Fig. ensue following flooding. Extensive bacteriological7). In bottom water, at three hours posttreatment, assessment lent support to this hypothesis. In the firstVectoBac WDG treatment produced higher spore counts place, the irrigation water from the reservoir used to fillthan other treatments except VectoLex WDG at the high the tubs had low bacterial counts, averaging 800- 1000rate of 0. 1 lb/ac. From day 3 through 28 posttreatment, cells/ ml. However, soon after the start of filling( 3 h), amore spore counts were encountered in the treatment of large number of bacteria were detected in the top andVectoBac WDG than in other treatments( Fig. 7). bottom water in the enriched tubs ( 0.3- 0.4 x 105 cells/
Gram stained bacteria: The data obtained with ml). This increase in bacterial numbers is very likely theregard to the changes in bacterial types in top and result of bacteria present before filling, and due tobottom water in the third test showed that Gr-ve bacteria bacteria present on or in the rabbit pellets added to theas in previous tests, accounted for 80-99% of the total tubs. The bacterial flora propagated rapidly and reachedbacteria on different sampling days during the course of peak populations on day 4 pretreatment ( day 3the test. Gr-ve rods increased from 40- 60% at three postflooding). Bacterial abundance was positivelyhours after start of filling the tubs and day 7 pretreatment con-elated with ovipositional activity of mosquitoesto 97- 98% on day 28 posttreatment( Fig. 8). Gr-ve cocci ( data omitted). After reaching a peak, the bacterial floraincreased from 20- 24% at three hours after start of followed a natural decline, reaching the low levels at thefilling the tubs to 55- 60% on day 7 pretreatment, then end of the tests. This natural decline pattern was similardecreased to 4- 16% on day 21 posttreatment. On day 28 in both the check and treatment regimens.posttreatment, no Gr-ve cocci were recovered in all In tests 1 and 2 in the summer season, treatmentswater samples. As in the first and second tests, Gr+ve with VectoBac WDG formulation contributed little ifrods found in the third test were spore- forming bacteria, any to the total bacterial flora. This was because of thetheir numbers made up no more than 4% of the total application ofrelatively small quantities of the productsbacteria. Gr+ve cocci were found in low numbers most necessary for the control ofmosquito larvae. The sporesduring the earlier phase of the test period( Fig. 8). of Bti were not detected in the check or other tubs before
treatment, but they were found in all treated tubs afterDISCUSSION treatment. On day 3 posttreatment, a higher number of
Bti spores was found in both top and bottom water. It isInitially, the Bti WDG formulation yielded a high possible that the peak was reached soon after treatment.
level of mosquito control at the two higher rates( 0.53 Concerning the dynamic changes in the abundanceand 1. 1 lb/ac). The extent of control was still high at 7 of bacterial types, it was noted that gram-negative( Gr-days posttreatment. At 14 days posttreatment the larval ve) rods and Gr-ve cocci constituted most of the total
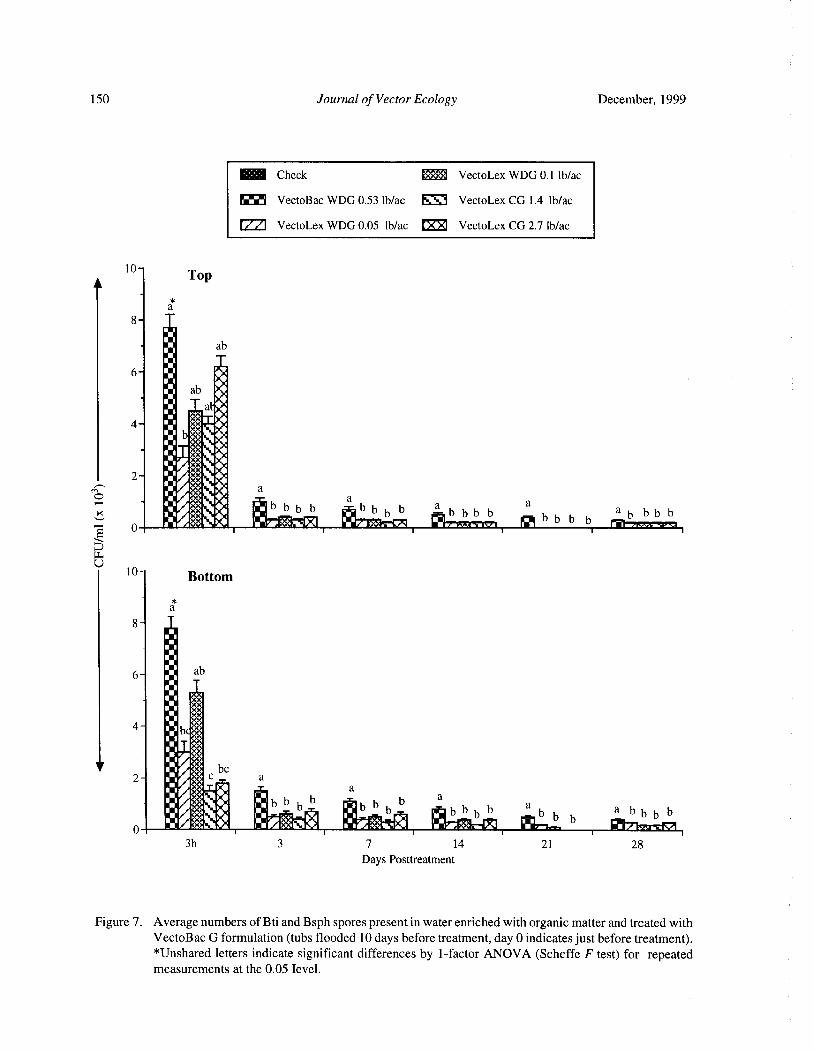
150 Journal of Vector Ecology December, 1999
Check ti. i: VectoLex WDG 0. 1 lb/ac
VectoBac WDG 0.53 lb/ ac ® VectoLex CG 1. 4 lb/ ac
VectoLex WDG 0.05 lb/ac ® VectoLex CG 2.7 lb/ ac
10Top
a
8
tiab
6 1
1 ab •,1
alb#1 v+v
1F
4 : +++{
hrior 4
1 *++{
r1 %+
ti 1# a
r+t{c
b b b b : b b b b a b b b ba
b b a b b b b0 1 1 :+:*; L IEf+ (. • F •'.•: IR. • a:*IT arm a 1
b ba.
Li
10 Bottom
a
8
6 :: ab
umml
4 ; b ++
V ti ::*: be2 :+:+: c a
1 *{ f _ at+{
I
i1 +• ;:a 60 : bbbb
rlbbbb
a
b b aai1 eb b b bb b
1 t*+*{ IF iii k K.; P I 4- 1— b b
3h 3 7 14 21 28
Days Posttreatment
Figure 7. Average numbers of Bti and Bsph spores present in water enriched with organic matter and treated with
VectoBac G formulation( tubs flooded 10 days before treatment, day 0 indicates just before treatment).Unshared letters indicate significant differences by 1- factor ANOVA ( Scheffe F test) for repeated
measurements at the 0.05 Ievel.

December, 1999 Journal of Vector Ecology 151
Gr-ve rods EJ Gr-ve cocci Gr+ve rods 56 Gr+ve cocci
100— Top Bottom
80- "
0-: 1 14I A) -
0 \ iiiiiI 100-
i:
80—
40—
C0 II
100-
80-
N c _ 1
0
100— 0 1 gliiiiiN
80—
60- ' N 1 \ \ ` ( D) - 1 1
11iiiui0 \ \ \ \ \
10 - 7 0 3 7 14 21 28 10 - 7 0 3 7 14 21 28
Day Pre and Posttreatment
Figure 8. Proportion of gram- negative( Gr-ve) and gram- positive( Gr+ve) bacteria present in water from untreatedA) and treated tubs with VectoBacWDG at 0.53 lb/ ac( B), VectoLex WDG 0. 1 lb/ ac( C), and VectoLex
CG 1. 4 lb/ac( D)( tubs flooded 10 days before treatment, day 0 indicates just before treatment).

152 Journal of Vector Ecology December, 1999
bacterial flora during the experimental period. Gr-ve large number of total existing natural populations of
rods increased gradually in their abundance over time bacteria, especially at the peak population level.while the Gr-ve cocci showed the reverse trend declining However, if one considers the spore- forming bacteriain abundance at the same time. Gr-positive( Gr+ve) rods alone, then the treatments made with larvicidal dosages
spore forming to which Bti and Bsph belong) occurred can increase the number of spores in the top and bottomin very low densities. In general, the numbers of total water of the treated habitats. This increase, however,
spore- forming bacteria in treated tubs increased slightly could be influenced by biotic and edaphic factors inafter treatment with Bti formulation. Gr+ve cocci were mosquito breeding habitats.
noted occasionally, and no obvious trend was noted intheir populations during the course of the tests.
AcknowledgmentsIn the experiment conducted in the cooler season
when both Bti and Bsph formulations were applied, theWe wish to thank John D. Chaney( Department of
bacterial populations also peaked on day 7 pretreatment
Entomology, University of California, Riverside) forday 3 postflooding). Just before treatment ( 10 days
assistance in field and laboratory during the course ofpostflooding), the total bacterial flora had declined
this study.sharply and further slight declines continued onsubsequent sampling days as in the previous experiments.Spore- forming bacteria reached a peak in all treatments REFERENCES CITED
and the check on day 3 posttreatment, then declinedsteadily as did the total bacteria on subsequent sampling Abacus Concepts, Inc. 1987. StatView SE+ Graphics.
days. However, the numbers of total spore formers in Abacus Concepts, Inc., Berkeley, CA, 234 pp.Bti and Bsph treatment regimens were higher than those Aranda, C. and R. Eritija. 1992. Laboratory and fieldof the check on most sampling days. Soon after treatment efficacy of Bacillus sphaericus 2362 against Culex3 h), high numbers of Bti and Bsph spores were found pipiens in the river Llobregat delta ( Barcelona,
in the top and bottom water of treated tubs. None of their Spain). Bull. Soc. Vector Ecol. 17: 41- 44.
spores were detected in the check. The decline ofBti and Clements, A. N. 1992. The Biology of Mosquitoes,Bsph spores in both top and bottom water was very Volume 1. Development, Nutrition, and
pronounced on day 3 after treatment. Bti and Bsph spore Reproduction. Chapman and Hall. London,
counts declined further on subsequent sampling days. It Glasgow, New York, Tokyo, Melbourne, Madras
was also noted that the populations of Gr-ve and Gr+ve 1: 73- 80.
bacteria followed the similar trends to those found in the de Barjac H. and D. J. Sutherland( eds). 1990. Bacterial
previous experiments ( tests 1 and 2), but the Gr+ve Control of Mosquitoes and Black Flies:
cocci were found most during the earlier phase of the Biochemistry, Genetics and Application ofBacillustest. thuringiensis and Bacillus sphaericus. Rutgers
In all three tests, total bacterial flora was higher in University Press, New Brunswick, NJ. 349 pp.the treated regimens than in the checks in most of the Jones, J. G. 1970. Studies on fresh water bacteria: effect
samples, especially in top water in test 3. This difference of medium composition and method on estimates
was very likely due to the elimination of mosquito of bacterial population. J. Appl. Bacteriol. 33: 679-
larvae in the treated tubs. Abundant larval populations 686.
in checks were responsible for ingesting bacterial cells, Karch, S., Z. A. Manzambi, and J. J. Salaun. 1991. Field
resulting in reduced numbers of bacteria. trials with VectoLex ( Bacillus sphaericus) and
From these studies, it is clear that bacterial flora VectoBac ( Bacillus thuringiensis H14) against
play an important role in the oviposition activity of Anopheles gambiae and Culex quinquefasciatus
gravid mosquitoes as indicated by larval production and breeding in Zaire. J. Am. Mosq. Contr. Assoc. 8:as a source of food for larvae. Bacterial decomposition 376- 385.
of organic matter produces transspecific behavior Merritt, R. W., D. A. Craig, R. S. Wotton, and E. D.
modifying substances that strongly influence mosquito Walker. 1996. Feeding behavior of aquatic insects:oviposition. Bacterial flora also serves as a source of case studies on black fly and mosquito larvae.food for mosquito larvae. Oviposition cues provided by Invertebr. Biol. 115: 206- 217.
the decomposition of organic matter also are signaling Merritt, R. W., E. J. Olds, and E. D. Walker. 1990.
abundant supply of food for larvae. These studies also Natural food and feeding behavior of Coquillettidiashowed that the addition of spores by treatments with perturbans larvae. J. Am. Mosq. Contr. Assoc. 6:Bti and Bsph formulations add very little to the already 35- 42.

December, 1999 Journal of Vector Ecology 153
Mulla, M. S. 1990. Activity, field efficacy, and use of Bacillus thuringiensis and Bacillus sphaericus.( H.Bacillus thuringiensis israelensis against de Barjac and D. J. Sutherland, eds.). Rutgers Univ.
mosquitoes. Pp. 134- 160 in Bacterial Control of Press, New Brunswick, NJ, 349 pp.Mosquitoes and Black Flies: Biochemistry, Genetics Skovmand, 0.and S. Bauduin. 1997. Efficacy ofgranularand Application of Bacillus thuringiensis and formulation of B. sphaericus against Cx.Bacillus sphaericus. ( H. de Barjac and D. J. quinquefasciatus and An. gambiae in West African
Sutherland, eds.). Rutgers Univ. Press, New countries. J. Vector Ecol. 22: 43- 51.
Brunswick, NJ, 349 pp. Smith, T. W., E. D. Walker, and M. G. Kaufman. 1998.
Mulla, M. S. 1991. Biological control of mosquitoes Bacterial density and survey of cultivablewith entomopathogenic bacteria. Chin. J. Entomol. heterotrophs in the surface water of a freshwater
Spec. Publ. 6: 93- 104. marsh habitat ofAnopheles quadrimaculatus l arvae
Mulla, M. S., R. L. Norland, D. M. Fanara, H. A. Diptera: Culicidae). J. Am. Mosq. Contr. Assoc.Darwazeh, and D. W. McKean. 1971. Control of 14: 72- 77.
chironomids midges in recreational lakes. J. Econ. Walker, E. D., E. J. Olds, and R. W. Merritt. 1988. Gut
Entomol. 64: 300- 307. content analysis of mosquito larvae ( Diptera:
Mulla, M. S., J. Rodcharoen, W. Ngamsuk, A. Tawatsin, Culicidae) using DAPI stain and epifluorescenceP. Pan- Urai, and U. Thavara. 1997. Field trials with microscopy. J. Med. Entomol. 25: 551- 554.Bacillus sphaericus formulations against polluted Walker, E. D. and R. W. Merritt. 1993. Bacterial
water mosquitoes in a suburban area of Bangkok, enrichment in the surface microlayer of an
Thailand. J. Am. Mosq. Contr. Assoc. 13: 297- 304. Anopheles quadrimaculatus ( Diptera: Culicidae)
Mulla, M. S., T. Su, U. Thavara, A. Tawatsinand, W. larval habitat. J. Med. Entomol. 30: 1050- 1052.
Ngamsuk, and P. Pan- Urai. 1999. Efficacy of new Yap, H. H. 1990. Field trials of Bacillus sphaericus forformulations of the microbial control agent Bacillus mosquito control. Pp. 307- 320 in Bacterial Controlsphaericus against polluted water mosquitoes in of Mosquitoes and Black Flies: Biochemistry,Thailand. J. Vector Ecol. 24: 99- 110. Genetics and Application ofBacillus thuringiensis
Nilsson, C. 1987. Feeding and food utilization by and Bacillus sphaericus. ( H. de Barjac and D. J.
mosquito larvae. Ph.D. Dissertation, Univ. of Sutherland, eds.). Rutgers Univ. Press, New
Uppsala, Dept. of Zoology, Sweden, 269 pp. Brunswick, NJ, 349 pp.Singer, S. 1990. Introduction to the study of Bacillus Yousten, A. A., S. B. Fretz, and S. A. Jelly. 1985.
sphaericus as a mosquito control agent. Pp. 221- Selective medium for mosquito-pathogenic strains236 in Bacterial Control of Mosquitoes and Black of Bacillus sphaericus. Appl. Environ. Microbiol.
Flies: Biochemistry, Genetics and Application of 49: 1532- 1533.

Journal of Vector Ecology 24( 2): 154- 157
Field Efficacy of Fipronil 3G, Lambda-cyhalothrin 10% CS,
and Sumithion 50EC Against the Dengue Vector
Aedes albopictus in Discarded Tires
S. Sulaiman', Z. A. Pawanchee2, A. Wahab2, J. Jamal2, and A. R. Sohadi3
Department ofBiomedical Science, Faculty ofAllied Health Sciences,Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia.
2Vector Control Unit, Department ofHealth, Municipality of Kuala Lumpur, Malaysia.Department of Parasitology and Medical Entomology, Faculty ofMedicine,
Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia.
Received 18 February 1999; Accepted 29 July 1999
ABSTRACT: The efficacy of three insecticides, fipronil 3G, lambda-cyhalothrin 10% CS, and sumithion50EC were evaluated against the dengue vector Aedes albopictus in discarded tires in Kuala Lumpur,Malaysia. The dosage given for each insecticide was 0.01 g of active ingredient/m2. Fipronil 3G was themost effective larvicide with a residual activity of up to two weeks, causing 88% mortality in Aedesalbopictus. Lambda-cyhalothrin 10% CS was effective for one week causing 92% larval mortality and twoweeks with 63% larval mortality. Sumithion 50EC had a residual efficacy of one week with 79% larval
mortality.
Keyword Index: Fipronil 3G, lambda-cyhalothrin 10% CS, sumithion 50EC, Aedes albopictus, discardedtires
INTRODUCTION The objective of this study was to compare theefficacy of fipronil 3G, lambda-cyhalothrin 10% CS,
In Southeast Asia, Aedes albopictus ( Skuse) has and sumithion 50EC against the dengue vector Ae.been incriminated as a secondary vector ofdengue fever albopictus in discarded tires in a dengue endemic area inand Aedes aegypti( Linnaeus) as the principal vector of Kuala Lumpur.
dengue viruses ( Russell et al. 1969, Chan et al. 1971,Jumali et al. 1979, Harinasuta 1984). Human-made MATERIALS AND METHODS
habitats, such as discarded tires, are becoming importantsources for the prolific breeding and dispersal of Ae. The experiment was conducted on flat groundalbopictus into new geographical areas ( Reiter and under shade. About 280 discarded tires were placedSprenger 1987, Laird et al. 1994). horizontally on the ground in each of four plots selected
Vythilingam ( 1988) used fenitrothion in fogging for the study. Each plot measured 5x 10m( 50m2) and thetrials in the field against Ae. aegypti adults and reported distance between plots was 5m. Three plots were assigned100% mortality. With the same insecticide it caused 89 to receive one control agent each, namely, fipronil 3Gto 100% mortality in Ae. aegypti larvae. Lam and Tham ( Regent®) granular formulation ( manufactured by
1988) conducted a field trial with ULV application of Rhone-Poulenc Sector Agro Rue Pierre Biazet, BPmalathion 96% technical grade and sumithion L-40S 9163 Lyon, France), lambda-cyhalothrin 10% CSagainst Ae. aegypti and Ae. albopictus at residential ( IconCS®) ( manufactured by Zeneca Agrochemicals,areas in Ipoh municipality, Perak, Malaysia. Malathion United Kingdom), and Sumithion 50EC( manufactured96% TG showed little larvicidal activity, whereas, by Sumitomo Chemical Company, Ltd., Japan). Bothsumithion L-40S appears to have good larvicidal and lambda-cyhalothrin 10% CS and sumithion 50EC wereadulticidal effects both indoors and outdoors. diluted with water and applied as a residual spray inside

December, 1999 Journal of Vector Ecology 155
each tire at a concentration of 0.01 gm a. i/ m2
and a immature stages), with lambda-cyhalothrin 10% CSvolume of 5 liters per plot. The fipronil G granules were mortality was 92%( 45 died out of 49 immature stages),placed inside each tire by hand at a rate of 0.01 g a. i/ m2.
sumithion 50EC produced 79% mortality( 65 died out ofThe dosage was based on the area of the plot. 82 immature stages), and controls had 2% mortality( 5
Prior to spraying, the tires in each plot were left died out of 331 immature stages). There were again noundisturbed in the open for about three weeks for Ae. significant differences ( P> 0. 05) between the threealbopictus breeding to take place. Between 2 and 3 liters insecticides, but all were significantly different from theof rainwater collected in each tire. No other mosquito controls( P< 0.0005). Two weeks after spraying, fipronilspecies was identified in this study. All the tires in each 3G yielded the highest mortality of 88%( 28 died out ofplot were sampled for immatures before spraying, 24 32 immature stages), lambda- cyhalothrin 10% CS ofhours after spraying, and weekly thereafter. Sampling 63% ( 15 died out of 24 immature stages), sumithionwas conducted by taking only one dip with a ladle from 50EC of42%( 19 died out of 45 immature stages), whileeach tire for every plot, and the numbers ofdead and live the controls had no mortality( 354 alive). There was noimmatures were recorded from each dip. Live immatures significant difference between the three insecticideswere replaced into their respective tires. The percentage fipronil 3G, lambda-cyhalothrin 10% CS, and sumithionmortality was based on dead immature stages sampled 50EC ( P> 0.05). However, all the insecticides were
by the ladle from each tire sampled. The above significantly different from the controls( P< 0.05).experiment was conducted three times at different time Three weeks after spraying, fipronil 3G showed theperiods. Statistical analysis was performed using the highest mortality among all the insecticides evaluatedANOVA and LSD test( Choi, 1978). with 51% mortality( 43 died out of 85 immature stages),
lambda-cyhalothrin 10% CS killed 48%( 19 died out of
RESULTS AND DISCUSSION 40 immature stages), sumithion 50EC of 13%( 13 died
out of98 immature stages), and controls had no mortalityFipronil 3G produced the highest mortality of Ae. ( 370 alive). There was no significant difference between
albopictus of 95% ( 290 died out of 304 immature fipronil 3G and lambda-cyhalothrin 10% CS ( P> 0. 05),stages), lambda-cyhalothrin 10% CS of 90%( 247 died but fipronil 3G was significantly different from sumithionout of 275 immature stages), sumithion 50EC killed 50EC( P< 0.05). Similarly, lambda-cyhalothrin 10% CS92%( 244 died out of264 immature stages), and controls was significantly different from sumithion 50ECwere without any mortality( 381 alive, no mortality) 24 ( P< 0.05) three weeks after spraying. However, all thehours after treatment, respectively ( TABLE 1). There insecticides were significantly different from the controlswere no significant differences ( P> 0.05) among the ( P> 0.05).
activities of the three insecticides, but they were Four weeks after spraying, fipronil 3G yielded thesignificantly different from the controls( P< 0.0005) 24 highest mortality of 41% ( 11 died out of 27 immaturehours after treatment. A week after spraying fipronil 3G stages), lambda-cyhalothrin 10% CS of 3% ( 3 died outthe highest mortality of 92% resulted( 48 died out of 52 of 115 immature stages), sumithion 50EC of 5%( 7 died
TABLE 1. The percentage mortality of immature stages of Aedes albopictus larvae intires after treatment with fipronil 3G, lambda-cyhalothrin 10% CS, andsumithion 50EC.
Mortality of Immature StagesLambda-
Weeks After Fipronil cyhalothrin Sumithion
Treatment 3G 10% CS 50EC Control
24 hours 95 90 92 0
1 week 92 92 79 2
2 weeks 88 63 42 0
3 weeks 51 48 13 0
4 weeks 41 3 5 0
5 weeks 2 0 0 0
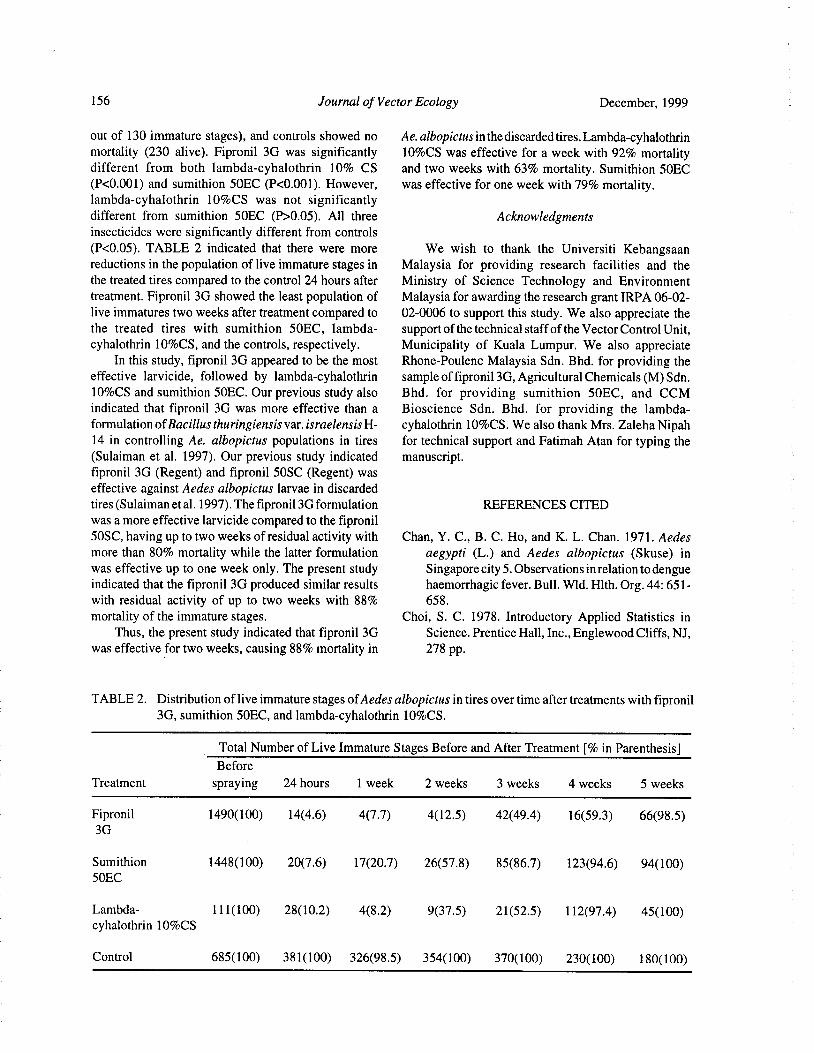
156 Journal of Vector Ecology December, 1999
out of 130 immature stages), and controls showed no Ae. albopictus in the discarded tires. Lambda-cyhalothrin
mortality ( 230 alive). Fipronil 3G was significantly 10% CS was effective for a week with 92% mortalitydifferent from both lambda-cyhalothrin 10% CS and two weeks with 63% mortality. Sumithion 50ECP< 0.001) and sumithion 50EC ( P< 0. 001). However, was effective for one week with 79% mortality.
lambda- cyhalothrin 10% CS was not significantlydifferent from sumithion 50EC ( P> 0.05). All three Acknowledgments
insecticides were significantly different from controlsP< 0.05). TABLE 2 indicated that there were more We wish to thank the Universiti Kebangsaan
reductions in the population of live immature stages in Malaysia for providing research facilities and thethe treated tires compared to the control 24 hours after Ministry of Science Technology and Environmenttreatment. Fipronil 3G showed the least population of Malaysia for awarding the research grant IRPA 06- 02-live immatures two weeks after treatment compared to 02-0006 to support this study. We also appreciate thethe treated tires with sumithion 50EC, lambda- support of the technical staff of the Vector Control Unit,
cyhalothrin 10% CS, and the controls, respectively. Municipality of Kuala Lumpur. We also appreciateIn this study, fipronil 3G appeared to be the most Rhone- Poulenc Malaysia Sdn. Bhd. for providing the
effective larvicide, followed by lambda-cyhalothrin sample of fipronil 3G, Agricultural Chemicals( M) Sdn.10% CS and sumithion 50EC. Our previous study also Bhd. for providing sumithion 50EC, and CCMindicated that fipronil 3G was more effective than a Bioscience Sdn. Bhd. for providing the lambda-formulation ofBacillus thuringiensis var. israelensis H- cyhalothrin 10% CS. We also thank Mrs. Zaleha Nipah14 in controlling Ae. albopictus populations in tires for technical support and Fatimah Atan for typing theSulaiman et al. 1997). Our previous study indicated manuscript.
fipronil 3G ( Regent) and fipronil 50SC ( Regent) was
effective against Aedes albopictus larvae in discarded
tires( Sulaiman et al. 1997). The fipronil 3G formulation REFERENCES CITED
was a more effective larvicide compared to the fipronil
50SC, having up to two weeks of residual activity with Chan, Y. C., B. C. Ho, and K. L. Chan. 1971. Aedes
more than 80% mortality while the latter formulation aegypti ( L.) and Aedes albopictus ( Skuse) in
was effective up to one week only. The present study Singapore city 5. Observations in relation to dengueindicated that the fipronil 3G produced similar results haemorrhagic fever. Bull. Wld. Hlth. Org. 44: 651-with residual activity of up to two weeks with 88% 658.
mortality of the immature stages. Choi, S. C. 1978. Introductory Applied Statistics inThus, the present study indicated that fipronil 3G Science. Prentice Hall, Inc., Englewood Cliffs, NJ,
was effective for two weeks, causing 88% mortality in 278 pp.
TABLE 2. Distribution of live immature stages ofAedes albopictus in tires over time after treatments with fipronil3G, sumithion 50EC, and lambda-cyhalothrin 10% CS.
Total Number of Live Immature Stages Before and After Treatment[% in Parenthesis]
Before
Treatment spraying 24 hours 1 week 2 weeks 3 weeks 4 weeks 5 weeks
Fipronil 1490( 100) 14( 4.6) 4( 7. 7) 4( 12. 5) 42( 49.4) 16( 59. 3) 66(98. 5)
3G
Sumithion 1448( 100) 20( 7. 6) 17( 20.7) 26( 57. 8) 85( 86.7) 123( 94.6) 94( 100)
50EC
Lambda- 111( 100) 28( 10.2) 4( 8. 2) 9( 37. 5) 21( 52. 5) 112( 97. 4) 45( 100)
cyhalothrin 10% CS
Control 685( 100) 381( 100) 326( 98. 5) 354( 100) 370( 100) 230( 100) 180( 100)

December, 1999 Journal of Vector Ecology 157
Harinasuta, C. 1984. Mosquito-borne diseases in mechanism for the worldwide dispersal ofcontainerSoutheast Asia. Mosq. Borne Dis. Bull. 1: 1- 11. breeding mosquitoes. J. Am. Mosq. Contr. Assoc.
Jumali, Sunarto, D. J. Gubler, S. Nalim, S. Eram, and J. 3: 494- 501.
S. Saroso. 1979. Epidemic dengue haemorrhagic Russell, P. K., D. J. Gould, T. M. Yuill, A. Nisalak, andfever in rural Indonesia III.Entomological studies. P. E. Winter. 1969. Recovery of dengue- 4 virusesAm. J. Trop. Med. Hyg. 28: 717- 724. from mosquito vectors and patients during an
Laird, M., L. Calder, R. C. Thornton, R. Syme, P. W. epidemic of dengue haemorrhagic fever. Am. J.
Golder, and M. Mogi. 1994. Japanese Aedes Trop. Med. Hyg. 18: 580- 583.albopictus among four mosquito species reaching Sulaiman, S., Z. A. Pawanchee, A. Wahab, J. Jamal, A.
New Zealand in used tires. J. Am. Mosq. Contr. R. Sohadi, and A. R. A. Rahman. 1997. Field
Assoc. 10: 14- 23. evaluation of Fipronil 3G, Fipronil 50SC and
Lam, W. K. and A. S. Tham. 1988. A field evaluation of Bacillus thuringiensis H- 14 against the dengue
the effectiveness of ULV application of malathion vector Aedes albopictus in tires. Med. Entomol.
96% technical grade and sumithion L-40S against Zool. 48: 351- 354.
Aedes aegypti ( Linnaeus) and Aedes albopictus Vythilingam, I. 1988. A field trial of the comparative
Skuse) in Ipoh municipality, Perak, Malaysia. effectiveness of fenitrothion and Resigen ofAedes
Trop. Biomed. 5: 81- 88. aegypti by thermal fog application. Mosq. BorneReiter, P. and D. Sprenger. 1987. The used tire trade: a Dis. Bull. 4: 99- 102.

Journal of Vector Ecology 24(2): 158- 170
Reproductive Biology of Lutzomyia shannoni (Dyar)Diptera: Psychodidae) Under Experimental Conditions
Estrella Cardenas', Cristina Ferro', Dario Corredor2,
Orlando Martinez2, and Leonard E. Munstermann3
Instituto Nacional de Salud, Laboratorio de Entomologic, Santa Fe de Bogota, Colombia,2 Universidad Nacional de Colombia, Facultad de Agronomic, Santa Fe de Bogota, Colombia.
3 Department ofEpidemiology and Public Health, YaleUniversity School ofMedicine, New Haven, CT, USA.
Received 5 March 1998; Accepted 3 August 1999
ABSTRACT. Baseline biological growth data of Lutzomyia shannoni( Dyar) were compared under two
experimental conditions within insulated styrofoam chests and in standard laboratory incubators. Thedevelopmental time from egg to adult was 67 and 52 days, respectively. Based on cohorts of 100 femalesin each experiment, horizontal life tables were constructed. The following predictive parameters wereobtained under each of the two conditions: net rate of reproduction ( 23. 5 and 18. 0 females per cohort
female), generation time( 11. 4 and 9. 4 weeks), intrinsic rate of population increase( 0.27 and 0. 30), andfinite rate of population increment( 1. 31 and 1. 36). The reproductive value for each class age of the cohortfemales was calculated. The observed parameters were obtained under each experimental condition: netrate of reproduction ( 1. 9 and 2. 5 females per cohort female), generation time ( 11. 7 and 9. 6 weeks),
intrinsic rate of population increase( 0.05 and 0.09), and finite rate of population increment( 1. 06 and1. 10). Vertical life tables were elaborated and mortality was described for every generation in each cohort.In addition, for two successive generations, additive variance and heritability for fecundity wereestimated.
Keyword Index: Lutzomyia shannoni, phlebotomine sand flies, life cycle, fecundity, intrinsic rate ofincrease, oviposition, heritability.
INTRODUCTION can support the development of at least three species ofLeishmania: Le. mexicana, Le. panamensis and Le.
Lutzomyia shannoni is a zoophilic species with a chagasi( Ferro et al. 1998).broad geographical distribution in the Western The reproductive cycles of vector arthropods playHemisphere, extending from the southeastern United a fundamental role in the epidemiology of the diseaseStates to northern Argentina( Young and Duncan 1994). they transmit. The parameters affecting transmission ofLocal distribution limits are dependent on factors of disease include fecundity rate, mortality rate, density,rainfall, temperature, physical barriers to migration, distribution by ages, migration rate, and geneticforest habitat, and abundance of appropriate vertebrate variation of the vector arthropods ( Black and Moorehosts( Young and Arias 1991). In the United States, it 1996, Tabachnick and Black 1996). However, none ofis a vector ofvesicular stomatitis virus( VSV), a disease this information is available for Lu. shannoni. To betteraffecting cattle, horses, and swine( Comer et al. 1990, understand baseline parameters of the life cycle,1991). In Colombia, Lu. shannoni isa species ofmedical growth under two experimental conditions wasand veterinary importance and occurs from sea level up compared. Horizontal life tables were constructedto approximately 1, 300 m( Young 1979). The role of based on the developmental time of each instar inLu. shannoni in VSV epidemiology in Colombia is cohorts of 100 females, observed and predictiveunknown, but the Instituto Colombiano Agropecuario population parameters were calculated. Stage specificICA) has recorded cattle infected with the New Jersey mortality data were collected, and, for two successive
serotype ofVSV in nearly 100 communities( ICA 1999). generations, the additive variance and heritability ofIn addition, under laboratory conditions, Lu. shannoni fecundity were estimated.

December, 1999 Journal of Vector Ecology 159
MATERIAL AND METHODS shown in the Figures. 1, 2.
Maintenance of Cohorts in the Laboratory Calculation of Predictive Population Parameters
The specimens ofLu. shannoni came from a colony Based on Horizontal Life Tables
maintained since 1992 in the Entomology Laboratory, In groups of organisms with intervals of discrete
Instituto Nacional de Salud( INS), Bogota, Colombia. ages, the net reproductive rate( R0) was calculated withA paired comparison was made under two experimental the following equation:conditions, in a styrofoam chest and in incubator.
The females were fed on hamster blood two days Ro = I Ix mxafter pupal emergence. Females were blood fed as x = o
follows: first, the adults were transferred from the
rearing chambers to cloth cages( 20 cm x 20 cm), then where lx is the survival proportion if the cohort females,a hamster was anesthetized( sodium pentothal, 10 mg/ and mx is the average of the cohort females. Thekg animal weight) and introduced into the cloth cages generation time( Tc) was calculated with the followingfor one hour. After blood feeding, a 60% glucose equation:
solution was provided daily on cotton swabs placed onthe muslin covering of each rearing chamber. The I x lx mxrearing and feeding ofadults and larvae ofLu. shannoni x= o
followed the methods of Modi and Tesh( 1983), Endris Tc =et al. ( 1982), and modifications of Ferro et al. ( 1998). Ro
Each day, data were collected on the number offemales remaining alive, date of emergence of the where x is the age class of the cohort female, lx is thepupae of the each female, date of blood feeding of each survival proportion if the cohort females, mx is thefemale, date of the oviposition of each female, number average of the cohort females, and Ro is the netof oviposited eggs for each female, number of retained reproductive rate. The intrinsic rate of population
eggs per female, number of non- embryonated eggs per increase( rc) was calculated from the following equation:female, number of embryonated eggs per female, and
number of days that each female survived in the adult In Rostage. The life cycle parameters were estimated by rc =
tracking the development of each cohort female Tcprogeny and averaging these data. Fecundity was
The maximum possible value of the parameter ( r) forestimated by summing eggs oviposited and those
species in given biotic and physical environmentsretained by the female. Retained eggs were counted by was denoted by ( rm). This value was calculated fromdissection after the females died.
the ( re) as the r initial ( ri) and then, with an iterativeIn Experiment 1, the cohort of 100 females was
method, each ri was proved simultaneously until thetaken from the 22nd generation of the colony. After
left side of the Euler and Lotka equation was equaledblood feeding, individual females were separated into
to I. At this point, the maximum value of the intrinsicrearing chambers( 5. 5 cm high and 6. 6 cm in diameter).
natural rate of increase rm was obtained ( for itsThe rearing chambers were placed in a styrofoam chest.
calculation, see Rabinovich 1980).
Inside, relative humidity and temperature were con-tinuously monitored with a hygrometer and maximum- Euler-Lotka equation:
minimum thermometer. In Experiment 2, the cohort
consisted of 100 couples ( one female and one male)
Ix mx e- rmx__taken from the 27th generation of the colony.
Immediately after blood feeding the females, eachx= 0
couple was separated into a rearing chamber ( 5. 5 cm
where( e) is the base of the natural logarithm, and lx mxhigh and 6. 6 cm in diameter). The rearing chambersis obtained from the horizontal life table of the cohort
were placed in plastic boxes with lids, and temperaturefemales. The finite rate of increase was calculated with
and humidity were monitored within each box. Theplastic boxes were maintained inside a temperature-
the equation:
controlled incubator. Changes in relative humidity and X= e rmtemperature in each experiment are summed in TABLE
1, and the daily oscillations of these parameters are where( e) is the base of natural logarithms and rm is the

160 Journal of Vector Ecology December, 1999
TABLE 1. Variation, standard deviation, average relative humidity, and temperature in two experimentalconditions for rearing Lutzomyia shannoni.
Experimental ConditionsParameters Styrofoam Chest Incubator
Range SD Average Range SD Average
Relative humidity 80-96% 3. 4% 88. 1% 90- 100% 2. 3% 94.4%Minimum temperature 21- 27° C 1. 2° C 25. 1° C 22- 27° C 0.9° C 25. 5° CMaximum temperature 27- 30° C 0.84° C 28° C 26-28° C 0.5° C 26. 8° C
CF
100 100
RH
80- 80 2c.
b E
0 60- 60
oyn bV V
40- 40 xa MT
cat
20- m 20 rx
P
1 6 11 16 21 26 31 36 41 46 51 56 61 66 71 76 81
Time, in days
Figure 1. Longevity of the cohort females and developmental time of each instar in one generation of Lutzomiyashannoni maintained in a styrofoam chest environment in relation to relative humidity and the temperature.CF, cohort females; L 1, Larvae I; L2, larvae II; L3, larvae III; L4, larvae IV; P, pupae; A, adults. RH, relativehumidity; MT, maximum temperature; mT, minimum temperature.
maximum rate of increase. According to Fisher( cited where ( e) is the base of natural logarithm, ( rm) is theby Pianka 1988), the reproductive value ( Vx) of the maximum natural rate of increase,( x) is the age of thecohort females at the age( x) was formerly expressed as cohort females, and lx mx is the product of thethe relative reproductive value at the moment of the survivorship of the each cohort female by its fecunditybirth V°( in practice equals 1). Vx was calculated by the at an age( x).
following equation: The construction of horizontal life tables and the
V ermx calculation of the predictive population parameters
x were based on methods of Pianka( 1988), RabinovichE ermx
lx mx 1980), and Southwood( 1978). This was assisted withV° lx xa small computer program ( E. Cardenas and M.
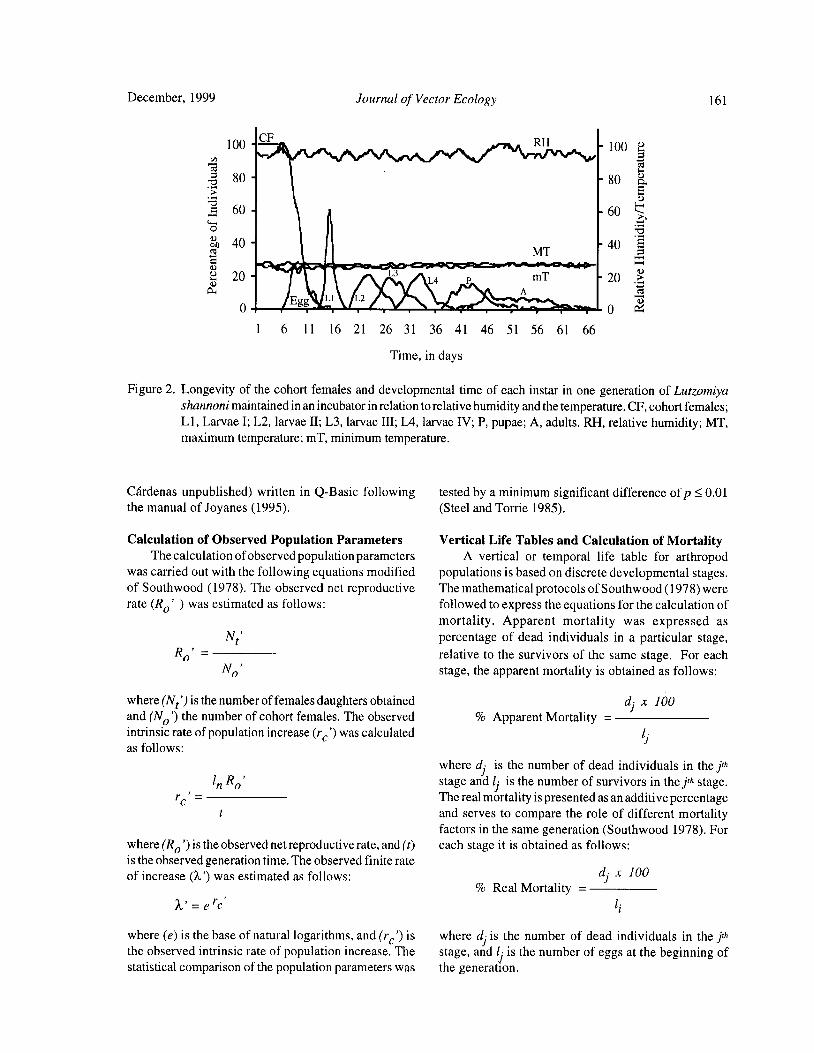
December, 1999 Journal of Vector Ecology 161
100CF RH 100
80
I80 a.
c 60 60 5
ta
oY
40MT
40
y 20 L3L4 P mT 20
AI_ .. 0 { Yi
1 6 11 16 21 26 31 36 41 46 51 56 61 66
Time, in days
Figure 2. Longevity of the cohort females and developmental time of each instar in one generation of Lutzomiyashannoni maintained in an incubator in relation to relative humidity and the temperature. CF, cohort females;L1, Larvae I; L2, larvae II; L3, larvae III; L4, larvae IV; P, pupae; A, adults. RH, relative humidity; MT,maximum temperature; mT, minimum temperature.
Cardenas unpublished) written in Q- Basic following tested by a minimum significant difference of p<_0. 01
the manual of Joyanes ( 1995). Steel and Torrie 1985).
Calculation of Observed Population Parameters Vertical Life Tables and Calculation of MortalityThe calculation of observed population parameters A vertical or temporal life table for arthropod
was carried out with the following equations modified populations is based on discrete developmental stages.
of Southwood ( 1978). The observed net reproductive The mathematical protocols ofSouthwood( 1978) were
rate( Ro' ) was estimated as follows: followed to express the equations for the calculation of
mortality. Apparent mortality was expressed asNt' percentage of dead individuals in a particular stage,
Ro' _ relative to the survivors of the same stage. For each
No' stage, the apparent mortality is obtained as follows:
where( Nt') is the number of females daughters obtained d• x 100
and( No') the number of cohort females. The observed Apparent Mortality =intrinsic rate ofpopulation increase( rc') was calculated ljas follows:
where d• is the number of dead individuals in the jth
In Ro' stage and li is the number of survivors in the jth stage.rc' = The real mortality is presented as an additive percentage
and serves to compare the role of different mortalityfactors in the same generation( Southwood 1978). For
where( R0') is the observed net reproductive rate, and( t) each stage it is obtained as follows:
is the observed generation time. The observed finite rate
of increase( X') was estimated as follows: d) x 100
Real Mortality =a, = e rc li
where( e) is the base of natural logarithms, and( rc') is where di is the number of dead individuals in the jththe observed intrinsic rate of population increase. The stage, and l• is the number of eggs at the beginning ofstatistical comparison of the population parameters was the generation.

162 Journal of Vector Ecology December, 1999
The irreplaceable or indispensable mortality is that Horizontal Life Tables and Predictive Populationpart of the generation mortality that will not occur if the Parameters
mortality factor in question were removed from the life Under the assumption that the cohort parentalsystem. Allowance is made for the action of subsequent females were reared in styrofoam chests or an incubator,mortality factors( Southwood 1978). The irreplaceable their pre- reproductive ages will approximate those of
mortality is calculated as follows: their respective progeny ( see Pianka 1988). The pre-
reproductive ages of cohort females were estimated
Ak x 100 based on the average pre- reproductive ages of theirIrreplaceable Mortality = respective progeny- 10.7 weeks for styrofoam chests
li and 8. 6 weeks for the incubator. With ,these data,
together with the average survivor data ofcohort femaleswhere Ak is the number of adults over the observed if (2 weeks for both), the class ages column ( x) of themortality had not occurred at the kth stage, and li is the horizontal life tables was constructed. The pre-number of eggs at the beginning of the generation. reproductive data provided an estimate of reproductive
value ( Vx) ( TABLES 3, 4). The mx' column wasCalculation of Genetic Parameters formed according to Rabinovich( 1972). The predictive
The genetic parameters additive variance( 62A) and population parameters were estimated in absence of
heritability( h2) for the fecundity of Lu. shannoni of the mortality according to Southwood( 1978).laboratory colony were estimated, following the method From TABLE 3, the following values were obtainedfather-descending Linear Simple Regression( Falconer for predictive population parameters: net reproduc-1970, Strickberger 1978). For this purpose, the number tive rate ( R0) = 23. 50 daughter females per cohortof oviposited eggs, retained eggs, and total eggs was female, generation time( Ta)= 11. 44 weeks, the naturalrecorded for 30 parent females; these data were averaged rate of increase( rm)= 0.27 daughter females for eachfor three daughters of each line. cohort female per week, and the finite rate of increase
for each cohort was?= 1. 31 individuals per female perRESULTS week.
From TABLE 4, the following values were obtainedLife Cycle for predictive population parameters: net reproductive
The oscillations of the relative humidity and rate ( R0) = 18 daughter females per cohort female,temperature were not drastic in the two experiments generation time( Tc)= 9. 42 weeks, the natural rate ofFig. 1, 2); however, these oscillations were reduced in increase( rm)= 0.30 daughter females for each cohort
the incubator where the average life cycle time was female per week, and the finite rate of increase for eachshorter by two weeks( Fig. 2). The longevity of cohort cohort was A.= 1. 36 individuals per female per week.females never extended beyond two weeks in either The last columns of TABLES 3 and 4 showedexperiment. The curves labeled" Egg" in Figures 1 and estimates of the reproductive values per age class2 showed the distribution percentage of oviposited of the cohort females. For cohort maintained in aeggs in each cohort. In both, the maximum peak of styrofoam chest, the increase in reproductive valueoviposition occurred at the ninth day of the female adult was exponential ( despite a fecundity of zero betweenlife. Oviposition pattern and emergence times of the 1st 0.5 to 9.5 weeks) until the females reached an age ofand 2nd- instar larvae were similar in the two micro- 11. 5 weeks. For the incubator cohort, the increase inenvironments. The 3rd, 4th larval instars, and pupae in reproductive value was also exponential ( note zerothe styrofoam chests required an average of one fecundity between 0.5 to 7. 5 weeks) until the femalesadditional day each over those in the incubator; reached an age of9. 5 weeks( Figs. 3, 4). These maximummoreover, these three stages demonstrated greater peaks of Vxcoincided with the week in which ovipositionvariance in time within stage( Figs. 1, 2). was initiated, confirming the theoretical expectation.
In TABLE 2, the life cycle averages forLu. shannoni Vx then abruptly decreased in the following week whenobtained in an incubator were compared with those the majority of the cohort females died. This populationobtained by Ferro et al.( 1998) in styrofoam chests. No parameter provided a clear estimate of the increasesignificant differences were obtained of the average potential of Lu. shannoni laboratory colony.duration in the following stages: eggs, 1 st- instar larvae,and adults; however, significant differences were Observed Population Parametersobtained for 2nd, 3rd, and 4th larval instars, and pupae In the styrofoam chest, 192 adult daughter femalesbetween the two experiments. were obtained from the 100 cohort; therefore, the net
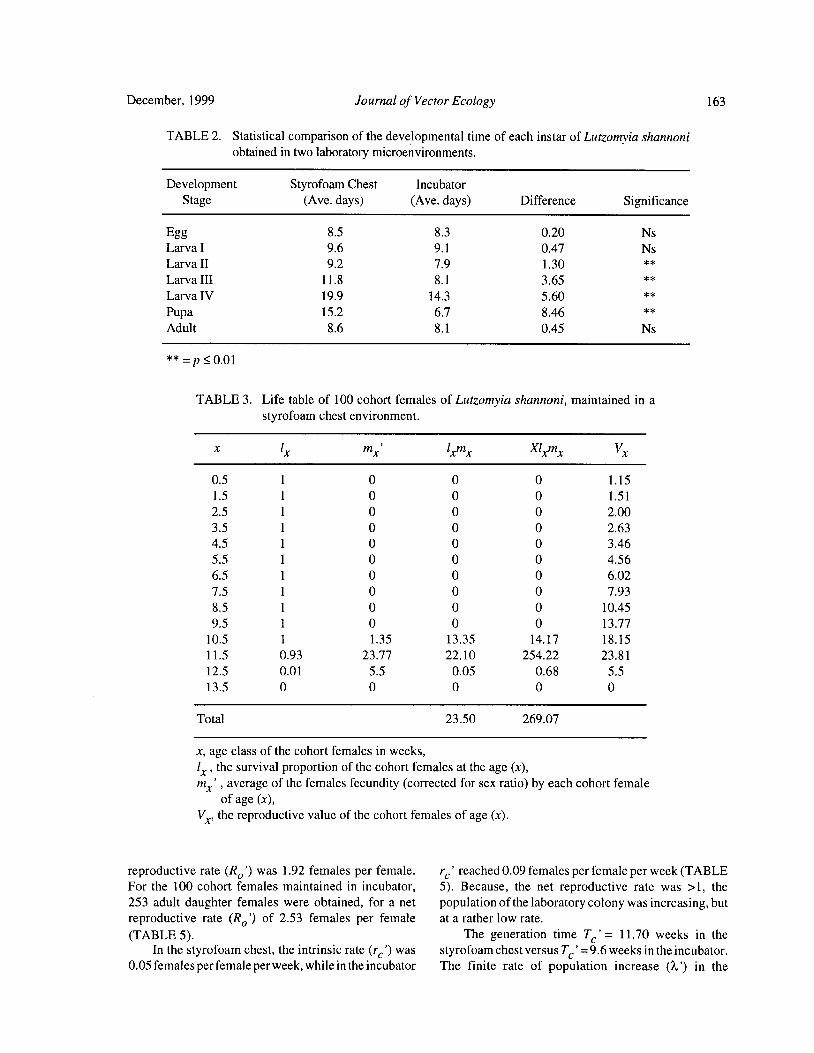
December, 1999 Journal of Vector Ecology 163
TABLE 2. Statistical comparison of the developmental time of each instar of Lutzomyia shannoni
obtained in two laboratory microenvironments.
Development Styrofoam Chest Incubator
Stage Ave. days) Ave. days) Difference Significance
Egg 8. 5 8. 3 0.20 Ns
Larva I 9. 6 9. 1 0.47 Ns
Larva II 9. 2 7. 9 1. 30
Larva III 11. 8 8. 1 3. 65
Larva IV 19. 9 14. 3 5. 60
Pupa 15. 2 6. 7 8. 46
Adult 8. 6 8. 1 0.45 Ns
p 5 0.01
TABLE 3. Life table of 100 cohort females of Lutzomyia shannoni, maintained in a
styrofoam chest environment.
x lx nix' lxmx Xlxmx Vx
0. 5 1 0 0 0 1. 15
1. 5 1 0 0 0 1. 51
2. 5 1 0 0 0 2.00
3. 5 1 0 0 0 2. 63
4. 5 1 0 0 0 3.46
5. 5 1 0 0 0 4. 56
6. 5 1 0 0 0 6. 02
7. 5 1 0 0 0 7. 93
8. 5 1 0 0 0 10.45
9. 5 1 0 0 0 13. 77
10. 5 1 1. 35 13. 35 14. 17 18. 15
11. 5 0.93 23. 77 22. 10 254.22 23. 81
12. 5 0.01 5. 5 0.05 0. 68 5. 5
13. 5 0 0 0 0 0
Total 23. 50 269.07
x, age class of the cohort females in weeks,
lx, the survival proportion of the cohort females at the age( x),mx' , average of the females fecundity ( corrected for sex ratio) by each cohort female
of age( x),
Vx, the reproductive value of the cohort females of age( x).
reproductive rate ( R0') was 1. 92 females per female. rc' reached 0. 09 females per female per week( TABLEFor the 100 cohort females maintained in incubator, 5). Because, the net reproductive rate was > 1, the
253 adult daughter females were obtained, for a net population of the laboratory colony was increasing, butreproductive rate ( R0') of 2. 53 females per female at a rather low rate.
TABLE 5). The generation time Tc'= 11. 70 weeks in the
In the styrofoam chest, the intrinsic rate( re') was styrofoam chest versus Tc'= 9. 6 weeks in the incubator.
0.05 females per female per week, while in the incubator The finite rate of population increase ( X') in the
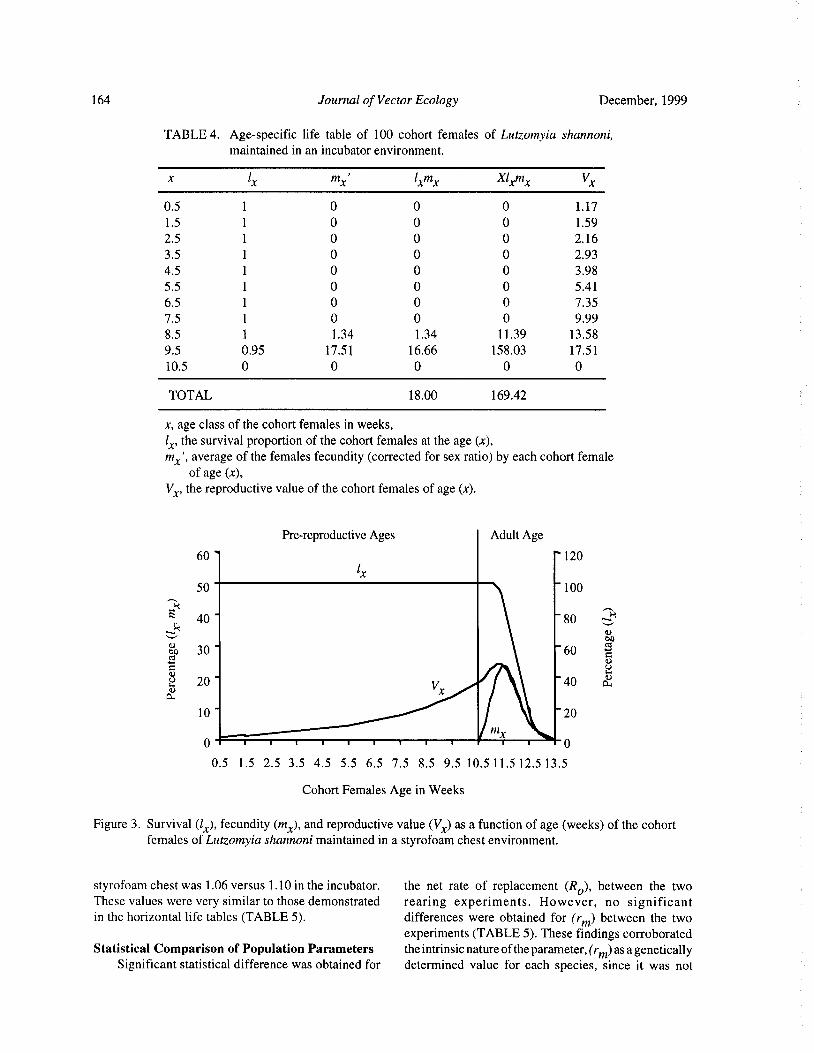
164 Journal of Vector Ecology December, 1999
TABLE 4. Age-specific life table of 100 cohort females of Lutzomyia shannoni,
maintained in an incubator environment.
x lx mx' lxmx Xlxmx V
0. 5 1 0 0 0 1. 17
1. 5 1 0 0 0 1. 59
2. 5 1 0 0 0 2. 16
3. 5 1 0 0 0 2. 93
4. 5 1 0 0 0 3. 98
5. 5 1 0 0 0 5. 41
6.5 1 0 0 0 7. 35
7. 5 1 0 0 0 9.99
8. 5 1 1. 34 1. 34 11. 39 13. 58
9. 5 0.95 17. 51 16.66 158. 03 17. 51
10.5 0 0 0 0 0
TOTAL 18. 00 169.42
x, age class of the cohort females in weeks,
lx, the survival proportion of the cohort females at the age( x),mx', average of the females fecundity( corrected for sex ratio) by each cohort female
of age( x),
Vx, the reproductive value of the cohort females of age( x).
Pre-reproductive Ages Adult Age
60 " 120
lx50 100
E 40 " 80ti
yon
e 30 - 60
0
2 20 " V 40 a:
a:x
10 " 20
0 0
0. 5 1. 5 2. 5 3. 5 4. 5 5. 5 6. 5 7. 5 8. 5 9. 5 10. 5 11. 5 12. 5 13. 5
Cohort Females Age in Weeks
Figure 3. Survival( 1x), fecundity( mx), and reproductive value( Vx) as a function of age( weeks) of the cohortfemales of Lutzomyia shannoni maintained in a styrofoam chest environment.
styrofoam chest was 1. 06 versus 1. 10 in the incubator. the net rate of replacement ( R0), between the two
These values were very similar to those demonstrated rearing experiments. However, no significantin the horizontal life tables ( TABLE 5). differences were obtained for ( rm) between the two
experiments( TABLE 5). These findings corroborated
Statistical Comparison of Population Parameters the intrinsic nature of the parameter,( rm) as a geneticallySignificant statistical difference was obtained for determined value for each species, since it was not
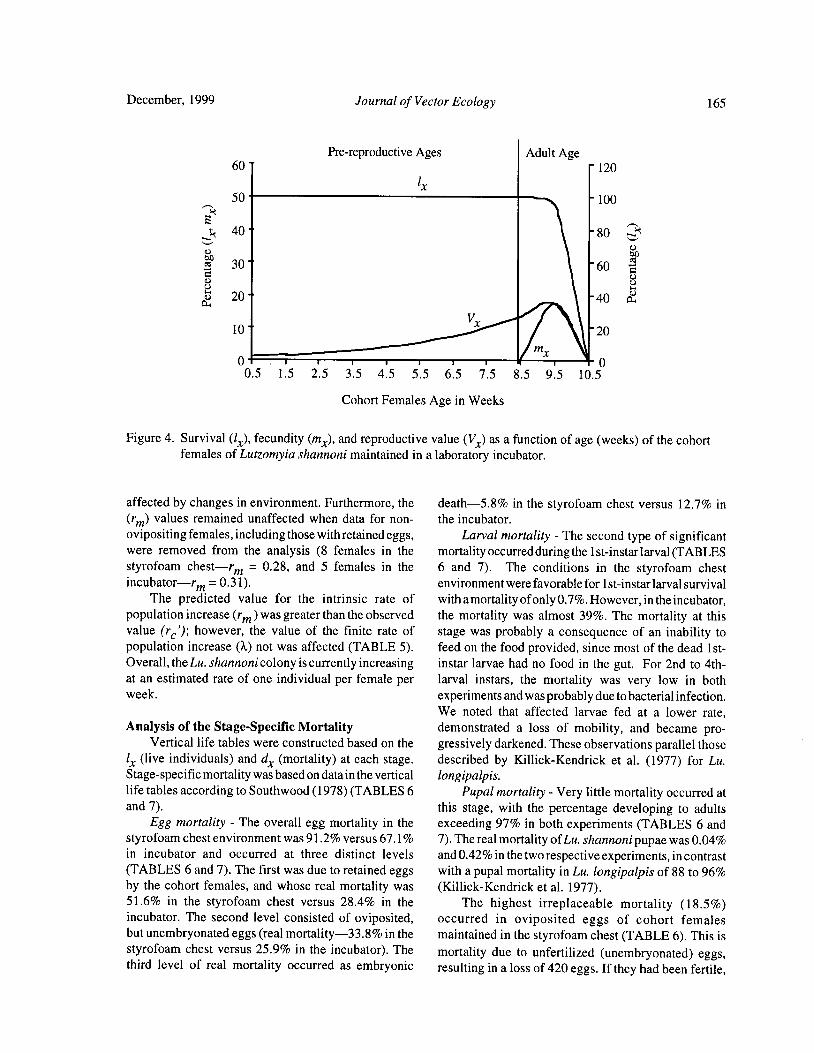
December, 1999 Journal of Vector Ecology 165
Pre-reproductive Ages Adult Age60' 120
lx50- 100
40- 80
on
c 30 60
s.20- 40 a:
V10'
x20
0. 5 1. 5 2. 5 3. 5 4. 5 5. 5 6. 5 7. 5 8. 5 9. 5 10.5
Cohort Females Age in Weeks
Figure 4. Survival( 1x), fecundity( mx), and reproductive value( Vx) as a function of age( weeks) of the cohortfemales of Lutzomyia shannoni maintained in a laboratory incubator.
affected by changes in environment. Furthermore, the death- 5. 8% in the styrofoam chest versus 12. 7% in
rm) values remained unaffected when data for non- the incubator.
ovipositing females, including those with retained eggs, Larval mortality - The second type of significantwere removed from the analysis ( 8 females in the mortality occurred during the ist-instarlarval( TABLESstyrofoam chest— rm = 0. 28, and 5 females in the 6 and 7). The conditions in the styrofoam chestincubator— rm= 0. 31). environment were favorable for 1st- instar larval survival
The predicted value for the intrinsic rate of with a mortality ofonly 0.7%. However, in the incubator,population increase( rm) was greater than the observed the mortality was almost 39%. The mortality at thisvalue ( rc'); however, the value of the finite rate of stage was probably a consequence of an inability topopulation increase ( X) not was affected( TABLE 5). feed on the food provided, since most of the dead 1st-Overall, the Lu. shannoni colony is currently increasing instar larvae had no food in the gut. For 2nd to 4th-at an estimated rate of one individual per female per larval instars, the mortality was very low in bothweek. experiments and was probably due to bacterial infection.
We noted that affected larvae fed at a lower rate,Analysis of the Stage-Specific Mortality demonstrated a loss of mobility, and became pro-
Vertical life tables were constructed based on the gressively darkened. These observations parallel thoselx ( live individuals) and dx ( mortality) at each stage. described by Killick-Kendrick et al. ( 1977) for Lu.
Stage- specific mortality was based on data in the vertical longipalpis.
life tables according to Southwood( 1978)( TABLES 6 Pupal mortality- Very little mortality occurred atand 7). this stage, with the percentage developing to adults
Egg mortality - The overall egg mortality in the exceeding 97% in both experiments ( TABLES 6 andstyrofoam chest environment was 91. 2% versus 67. 1% 7). The real mortality ofLu. shannoni pupae was 0.04%in incubator and occurred at three distinct levels and 0.42% in the two respective experiments, in contrastTABLES 6 and 7). The first was due to retained eggs with a pupal mortality in Lu. longipalpis of 88 to 96%
by the cohort females, and whose real mortality was ( Killick-Kendrick et al. 1977).51. 6% in the styrofoam chest versus 28.4% in the The highest irreplaceable mortality ( 18. 5%)incubator. The second level consisted of oviposited, occurred in oviposited eggs of cohort femalesbut unembryonated eggs( real mortality- 33. 8% in the maintained in the styrofoam chest( TABLE 6). This isstyrofoam chest versus 25.9% in the incubator). The mortality due to unfertilized ( unembryonated) eggs,third level of real mortality occurred as embryonic resulting in a loss of 420 eggs. If they had been fertile,
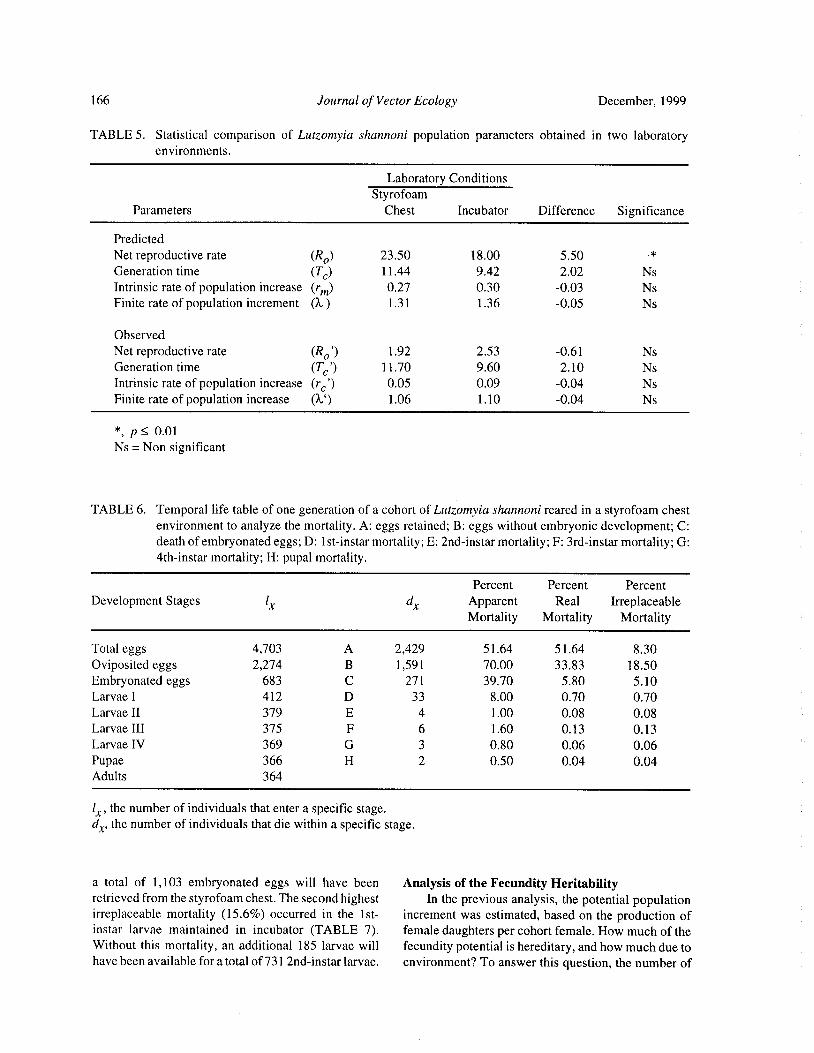
166 Journal of Vector Ecology December, 1999
TABLE 5. Statistical comparison of Lutzomyia shannoni population parameters obtained in two laboratoryenvironments.
Laboratory ConditionsStyrofoam
Parameters Chest Incubator Difference Significance
Predicted
Net reproductive rate R0) 23. 50 18.00 5. 50
Generation time Tc) 11. 44 9.42 2.02 Ns
Intrinsic rate of population increase ( rm) 0.27 0.30 0.03 Ns
Finite rate of population increment ( 2 ) 1. 31 1. 36 0.05 Ns
Observed
Net reproductive rate R0') 1. 92 2.53 0.61 Ns
Generation time Ta') 11. 70 9. 60 2. 10 Ns
Intrinsic rate of population increase ( rc') 0.05 0.09 0.04 Ns
Finite rate of population increase ( X') 1. 06 1. 10 0.04 Ns
p<_ 0.01
Ns= Non significant
TABLE 6. Temporal life table of one generation of a cohort of Lutzomyia shannoni reared in a styrofoam chest
environment to analyze the mortality. A: eggs retained; B: eggs without embryonic development; C:death of embryonated eggs; D: lst-instar mortality; E: 2nd- instar mortality; F: 3rd- instar mortality; G:4th-instar mortality; H: pupal mortality.
Percent Percent Percent
Development Stages lX dX Apparent Real Irreplaceable
Mortality Mortality Mortality
Total eggs 4, 703 A 2,429 51. 64 51. 64 8. 30
Oviposited eggs 2,274 B 1, 591 70.00 33. 83 18. 50
Embryonated eggs 683 C 271 39. 70 5. 80 5. 10
Larvae I 412 D 33 8. 00 0.70 0. 70
Larvae II 379 E 4 1. 00 0.08 0.08
Larvae III 375 F 6 1. 60 0. 13 0. 13
Larvae IV 369 G 3 0.80 0.06 0.06
Pupae 366 H 2 0.50 0.04 0.04
Adults 364
lx, the number of individuals that enter a specific stage.dX, the number of individuals that die within a specific stage.
a total of 1, 103 embryonated eggs will have been Analysis of the Fecundity Heritabilityretrieved from the styrofoam chest. The second highest In the previous analysis, the potential populationirreplaceable mortality ( 15. 6%) occurred in the 1st- increment was estimated, based on the production ofinstar larvae maintained in incubator ( TABLE 7). female daughters per cohort female. How much of theWithout this mortality, an additional 185 larvae will fecundity potential is hereditary, and how much due tohave been available for a total of 731 2nd- instar larvae. environment? To answer this question, the number of
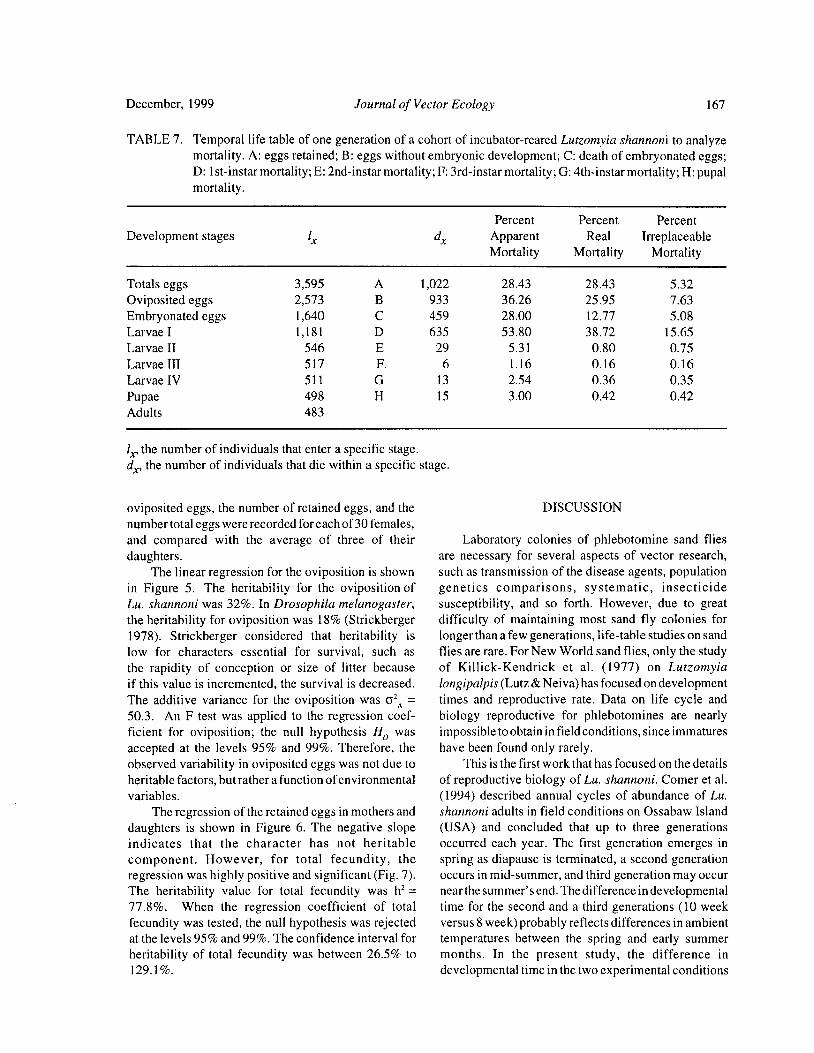
December, 1999 Journal of Vector Ecology 167
TABLE 7. Temporal life table of one generation of a cohort of incubator- reared Lutzomyia shannoni to analyze
mortality. A: eggs retained; B: eggs without embryonic development; C: death of embryonated eggs;D: 1st- instar mortality; E: 2nd- instar mortality; F: 3rd- instar mortality; G: 4th- instar mortality; H: pupalmortality.
Percent Percent Percent
Development stages lx dx Apparent Real Irreplaceable
Mortality Mortality Mortality
Totals eggs 3, 595 A 1, 022 28.43 28. 43 5. 32
Oviposited eggs 2,573 B 933 36.26 25. 95 7. 63
Embryonated eggs 1, 640 C 459 28.00 12.77 5. 08
Larvae I 1, 181 D 635 53. 80 38. 72 15. 65
Larvae II 546 E 29 5. 31 0. 80 0. 75
Larvae III 517 F. 6 1. 16 0. 16 0. 16
Larvae IV 511 G 13 2.54 0.36 0. 35
Pupae 498 H 15 3. 00 0.42 0. 42
Adults 483
lx, the number of individuals that enter a specific stage.dx, the number of individuals that die within a specific stage.
oviposited eggs, the number of retained eggs, and the DISCUSSION
number total eggs were recorded for each of 30 females,
and compared with the average of three of their Laboratory colonies of phlebotomine sand fliesdaughters. are necessary for several aspects of vector research,
The linear regression for the oviposition is shown such as transmission of the disease agents, population
in Figure 5. The heritability for the oviposition of genetics comparisons, systematic, insecticide
Lu. shannoni was 32%. In Drosophila melanogaster, susceptibility, and so forth. However, due to greatthe heritability for oviposition was 18%( Strickberger difficulty of maintaining most sand fly colonies for1978). Strickberger considered that heritability is longer than a few generations, life- table studies on sand
low for characters essential for survival, such as flies are rare. For New World sand flies, only the studythe rapidity of conception or size of litter because of Killick-Kendrick et al. ( 1977) on Lutzomyia
if this value is incremented, the survival is decreased. longipalpis( Lutz& Neiva) has focused on development
The additive variance for the oviposition was 62A = times and reproductive rate. Data on life cycle and
50.3. An F test was applied to the regression coef- biology reproductive for phlebotomines are nearlyficient for oviposition; the null hypothesis Ho was impossible to obtain in field conditions, since immatures
accepted at the levels 95% and 99%. Therefore, the have been found only rarely.observed variability in oviposited eggs was not due to This is the first work that has focused on the details
heritable factors, but rather a function ofenvironmental of reproductive biology of Lu. shannoni. Comer et al.variables. 1994) described annual cycles of abundance of Lu.
The regression of the retained eggs in mothers and shannoni adults in field conditions on Ossabaw Island
daughters is shown in Figure 6. The negative slope ( USA) and concluded that up to three generationsindicates that the character has not heritable occurred each year. The first generation emerges in
component. However, for total fecundity, the spring as diapause is terminated, a second generationregression was highly positive and significant( Fig. 7). occurs in mid- summer, and third generation may occurThe heritability value for total fecundity was
h2= near the summer' s end. The difference in developmental
77. 8%. When the regression coefficient of total time for the second and a third generations ( 10 week
fecundity was tested, the null hypothesis was rejected versus 8 week) probably reflects differences in ambientat the levels 95% and 99%. The confidence interval for temperatures between the spring and early summerheritability of total fecundity was between 26.5% to months. In the present study, the difference in129. 1%. developmental time in the two experimental conditions
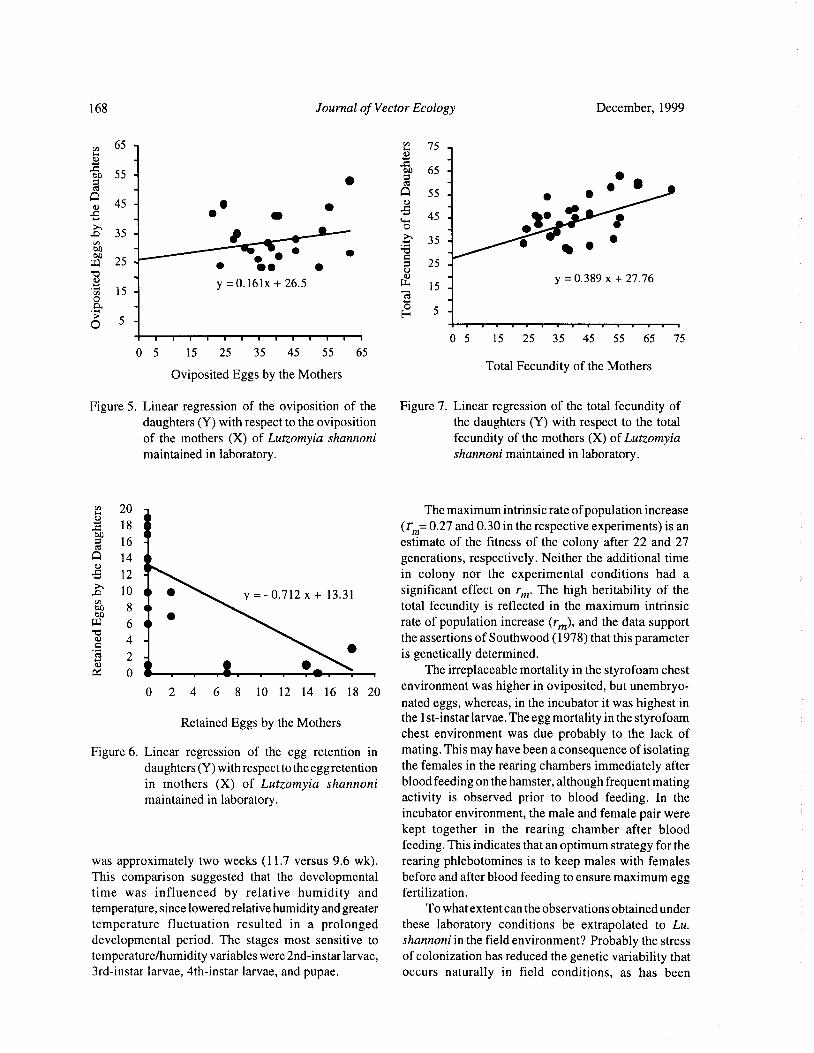
168 Journal of Vector Ecology December, 1999
65 - 41 75 -o
O65 -55
etoQ
Q 55 - 45 - a)
5 _ 45 _ l•o0 35 - j 35 -
W 25 o 25 -
o - Ny = 0. 389 x+ 27. 76
15 -y= 0. 161x+ 26. 5 w 15 -
o. - 05 -
O 5 -H
I 0 5 15 25 35 45 55 65 75
0 5 15 25 35 45 55 65
Oviposited Eggs by the MothersTotal Fecundity of the Mothers
Figure 5. Linear regression of the oviposition of the Figure 7. Linear regression of the total fecundity ofdaughters( Y) with respect to the oviposition the daughters ( Y) with respect to the total
of the mothers ( X) of Lutzomyia shannoni fecundity of the mothers( X) of Lutzomyiamaintained in laboratory. shannoni maintained in laboratory.
20 The maximum intrinsic rate ofpopulation increase
18 r. 0.27 and 0.30 in the respective experiments) is an
0 16 estimate of the fitness of the colony after 22 and 27al
Q 14 generations, respectively. Neither the additional time12 in colony nor the experimental conditions had a10 •
y =- 0. 712 x + 13. 31 significant effect on rm. The high heritability of theo8 total fecundity is reflected in the maximum intrinsicw 6 • rate of population increase ( rm), and the data supportc 4 the assertions of Southwood( 1978) that this parameter
2 is genetically determined.0 The irreplaceable mortality in the styrofoam chest
0 2 4 6 8 10 12 14 16 18 20environment was higher in oviposited, but unembryo-
nated eggs, whereas, in the incubator it was highest in
Retained Eggs by the Mothers the l st- instar larvae. The egg mortality in the Styrofoamchest environment was due probably to the lack of
Figure 6. Linear regression of the egg retention in mating. This may have been a consequence of isolatingdaughters( Y) with respect to the egg retention the females in the rearing chambers immediately afterin mothers ( X) of Lutzomyia shannoni blood feeding on the hamster, although frequent matingmaintained in laboratory. activity is observed prior to blood feeding. In the
incubator environment, the male and female pair were
kept together in the rearing chamber after bloodfeeding. This indicates that an optimum strategy for the
was approximately two weeks ( 11. 7 versus 9.6 wk). rearing phlebotomines is to keep males with femalesThis comparison suggested that the developmental before and after blood feeding to ensure maximum eggtime was influenced by relative humidity and fertilization.
temperature, since lowered relative humidity and greater To what extent can the observations obtained under
temperature fluctuation resulted in a prolonged these laboratory conditions be extrapolated to Lu.developmental period. The stages most sensitive to shannoni in the field environment? Probably the stresstemperature/humidity variables were 2nd- instar larvae, of colonization has reduced the genetic variability that3rd- instar larvae, 4th- instar larvae, and pupae. occurs naturally in field conditions, as has been

December, 1999 Journal of Vector Ecology 169
demonstrated for several colonies of Lu. longipalpis J. Trop. Med. Hyg. 42: 483- 490.Mukhopadhyay et al. 1997, Morrison et al. 1995) and Corner, J. A., D. E. Stallknecht, J. L. Corn, and V. F.
several mosquito species ( Munstermann 1994). The Nettles. 1991. Lutzomyia shannoni ( Diptera:
colony environment cannot duplicate conditions of Psychodidae): a biological vector of the New Jerseylarval nutrition, temperature, humidity, and host serotype of vesicular stomatitis virus on Ossabaw
availability; and, consequently, will strongly affect Island, Georgia. Parassitologia 33( Suppl. 1): 151-
adult behavior, egg mortality, larval growth period, and 158.
adult mortality. Furthermore, female longevity in the Comer, J. A., D. M. Kavanaugh, D. E. Stallknecht, and
field is undoubtedly greater than that observed in the J. L. Corn. 1994. Population dynamics ofLutzomyia
laboratory, with an associated low egg mortality. This shannoni( Diptera: Psychodidae) in relation to the
conclusion is based on the necessary chronology of epizootiology of vesicular stomatitis virus on
disease transmission by these insect vectors. The field Ossabaw Island, Georgia. J. Med. Entomol. 31:
females must ingest blood at least twice— first to obtain 850- 854.
the parasite, then ( after a defined parasite incubation Endris, R. G., P. V. Perkins, D. G. Young, and R. N.and sand fly oviposition period) to transmit the disease Johnson. 1982. Techniques for laboratory rearingagent to healthy hosts. Nonetheless, the laboratory data of sand flies ( Diptera: Psychodidae). Mosquito
have provided essential baseline information concerning News 42: 400- 407.
the reproductive potential for Lu. shannoni. For Falconer, D. S. 1970. Introduccion a la Genetica
laboratory colonies, it suggests more efficient rearing Cuantitativa. Compaflia Editorial Continental S. A.
practices and at which points in the life cycle are more 430 pp.critical to colony maintenance. For studies of Ferro, C., E. Cardenas, D. Corredor, A. Morales, and
phlebotomine population biology in the field, these L.E. Munstermann. 1998. Life cycle and fecundityobservations provide guidelines for approximating the analysis of Lutzomyia shannoni ( Dyar) ( Diptera:
boundaries of expected biological behaviors. Finally Psychodidae). Mem. Inst. Oswaldo Cruz 93: 195-
these data provide clues to how and where this species 199.
might survive in the field and, eventually, documenting ICA. 1999. Boletin de sanidad animal 1996- 1997.
its complete life cycle in the natural habitat. Grupo de epidemiologia veterinaria, Div. Sanidad
Animal, Inst. Colomb. Agropec., Bogota, Colombia
Acknowledgments In Press).
Joyanes, L. 1995. Programacion en QuickBasic/QBasic.
We thank the anonymous reviewers whose Segunda edicion, McGraw-Hill. Impreso en
suggestions greatly improved the presentation. Marco Espana, 641 pp.Fidel Suarez is acknowledged for his continuous Killick-Kendrick, R., A. J. Leaney, and P. D. Ready.maintenance of the laboratory colony. We also want to 1977. The establishment, maintenance and
thank Margarita Cardenas for her collaboration in the productivity of a laboratory colony of Lutzomyiaelaboration of a small computer program. This research longipalpis ( Diptera: Psychodidae). J. Med.
was supported by the United States National Institutes Entomol. 13: 429- 440.
of Health ( AI-34521 to L.E.M) and the Colombian Modi, G. B. and R. B. Tesh. 1983. A simple technique
Instituto Nacional de Salud ( Project No. 3100302 to for mass rearing Lutzomyia longipalpis andC. F. of the Entomology Laboratory). Phlebotomus papatasi ( Diptera: Psychodidae) in
the laboratory. J. Med. Entomol. 20: 568- 569.Morrison, A. C., C. Ferro, R. Pardo, M. Torres, and L.
REFERENCES CITED E. Munstermann. 1995. An ecological and genetic
study of Lutzomyia longipalpis in a centralBlack IV, W.C. and C. G. Moore. 1996. Population Colombian focus of visceral leishmaniasis. Bol.
biology as a tool for studying vector-borne diseases. Dir. Malariol. Saniamiento Ambiental 35 ( Suppl.
Pp. 292- 416 in The Biology of Disease Vectors( B. 1): 235- 248.
J. Beaty, and W. C. Marquardt, eds). University Mukhopadhyay, J., E. F. Rangel, K. Ghosh, and L. E.Press of Colorado, Niwot, CO, 632 pp. Munstermann. 1997. Patterns ofgenetic variability
Corner, J. A., R. B. Tesh, G. B. Modi, J. L. Corn, and V. in colonized strains of Lutzomyia longipalpis
F. Nettles. 1990. Vesicular stomatitis virus, New Diptera: Psychodidae) and its consequences. Am.
Jersey serotype: replication in and transmission by J. Trop. Med. Hyg. 57: 216- 221.Lutzomyia shannoni( Diptera: Psychodidae). Am. Munstermann, L. E. 1994. Unexpected genetic

170 Journal of Vector Ecology December, 1999
consequences of colonization and inbreeding: Ediciones Omega, S. A. Barcelona, 937 pp.allozyme tracking in Culicidae ( Diptera). Ann. Tabachnick, W. J. and W.C. Black, IV. 1996. PopulationEntomol. Soc. Am. 87: 157- 164. genetics in vector biology. Pp. 417-437 in The
Pianka, E. R. 1988. Evolutionary Ecology. Harper and Biology of Disease Vectors( B. J. Beaty, and W. C.Row, New York, 468 pp. Marquardt, eds). University Press of Colorado,
Rabinovich, J. E. 1980. Introduccion a la Ecologia de Niwot, CO, 632 pp.Poblaciones Animales. CompaniaEd. Continental, Young, D. G. 1979. A Review of the BloodsuckingMexico City, Mexico, 313 pp. Psychodid Flies of Colombia ( Diptera:
Rabinovich, J. E. 1972. Vital statistics of Triatominae Phlebotominae and Sycoracinae). Tech. Bull. 806,Hemiptera: Reduviidae) under laboratory Agric. Exp. Station, IFAS, Univ. Florida,
conditions. I. Triatoma infestans Klug. J. Med. Gainesville, 226 pp.Entomol. 9: 351- 370. Young, D. G. and J. A. Arias. 1991. Flebotomos:
Southwood, T. R. E. 1978. Ecological Methods, with Vectores de Leishmaniasis en las Americas.Particular Reference to the Study of Insect Panamerican Hlth Org. Tech. Bull. No. 33, 28 pp.Populations. Chapman and Hall, London, 524 pp. Young, D. G. and M. A. Duncan. 1994. Guide to the
Steel, R. G. D. and J. H. Torrie. 1985. Bioestadistica Identification and Geographic Distribution ofPrincipios y Procedimientos. Spanish Ed., Lutzomyia Sand Flies in Mexico, the West Indies,McGraw-Hill, New York, 622 pp. Central and South America( Diptera: Psychodidae).
Strickberger, M. W. 1978. Genetica. Segunda edicion. Am. Entomol. Inst. Gainesville, FL, 881 pp.

Journal of Vector Ecology 24( 2): 171- 181
The Fleas (Siphonaptera) of South Carolina with an
Assessment of Their Vectorial Importance
Lance A. Durdent, William Wills2, and Kerry L. Clark3
Institute ofArthropodology& Parasitology, Georgia SouthernUniversity, P.O. Box 8056, Statesboro, GA 30460, USA.
15 Chandler Court, Columbia, SC 29210, USA.
Department ofHealth Sciences, University ofNorth Florida,4567 St. John' s Bluff Road South, Jacksonville, FL 32224, USA.
Received 8 March 1999; Accepted 19 April 1999
ABSTRACT: We document 25 species of fleas from South Carolina including new state records for twospecies, the ctenophthalmids Epitedia cavernicola and Rhadinopsylla orama. Host and other collection
data, by county, are provided, including many new records amassed from 1992- 1998 and some olderrecords gleaned from the Clemson University Arthropod Collection. One flea species, the rhopalopsyllidPolygenis gwyni, is especially common and widespread in South Carolina, particularly on the cotton ratSigmodon hispidus) in coastal plain habitats. The largest number of flea species( 8) from a single host
species was recorded from the cotton mouse, Peromyscus gossypinus. Several flea species of potential
medical or veterinary importance were recorded, some of which are potential vectors of pathogens, suchas the agents of murine typhus, murine typhus- like disease, sylvatic epidemic typhus, cat scratch disease,
and rodent bartonellosis. A host- flea list for South Carolina is included.
Keyword Index: Fleas, Siphonaptera, South Carolina, faunal analysis, vectorial importance.
INTRODUCTION MATERIALS AND METHODS
The flea fauna ofSouth Carolina has not previously In order to survey South Carolina' s flea fauna, webeen documented in any detail. Benton( 1980) depicted first made detailed literature searches for previouslycollection localities for 18 species of fleas from South documented flea records from the state. Next, we
Carolina in his flea atlas of the eastern United States. initiated a statewide mammal live- trapping programAdditionally, a few miscellaneous flea records for South from 1994- 1998 to survey fleas associated with variousCarolina have been documented, particularly by Fox species ofmammals. We also asked field mammalogists
1940), Trembley and Bishopp ( 1940), Johnson and working in the state to collect fleas for this study. Mist-Traub ( 1954), Pratt and Good ( 1954), Whitaker et al. netted birds were also examined for fleas( Durden et al.1994), and Durden et al. ( 1997a). Because South 1997b). Lastly, we accessed the Clemson University
Carolina encompasses several physiographic realms Arthropod Collection and recorded collection data for
ranging from coastal plain to montane ( Appalachian) any flea specimens deposited there, some of which arezones, a relatively rich fauna of eastern North American historically noteworthy.fleas could feasibly inhabit this state. Numbers of flea In the lists that follow, flea species are presented
species documented from adjacent states include 19 for alphabetically followed in parentheses by theGeorgia, 17 for North Carolina( Benton 1980), and 33 Entomological Society of America-approved commonfor Tennessee( Durden and Kollars 1997). name( Bosik 1997) if one has been assigned, and then by
Except for murine typhus, which was a widespread the family designation. Collection data are then presentedzoonosis in South Carolina until the early 1950s( Pratt by county( in upper case); South Carolina counties are1958), the status of flea-borne diseases in the state is shown in Figure 1. Literature citations for South Carolina
poorly documented. Knowledge of the flea species flea records mostly provide the author and date ofoccurring in South Carolina will permit an analysis of publication but also include the number and sex of fleathe flea- borne pathogens that maybe present in the state. specimens, the host, collection date, and name of
172 Journal of Vector Ecology December, 1999
collector, if available. The same data are presented for = Lance A. Durden ( Georgia Southern University,the new collections including the slide-mounted material Statesboro); LR = Laurie Reid; MB = Mary Bunchheld at Clemson University. In a few cases, especially ( South Carolina Dept. ofNatural Resources, Clemson);
for older material, the host species, name of the collector, SL = Susan Loeb ( U. S. Dept. of Forest Resources,
or full collection date were unavailable. Collection data Clemson, SC); RBC= R. B. Casey; REW= R. E. Ware;from a single host individual are separated by SCH= Schroeder( other initials unknown); SM= Stanleybackslashes, whereas, data from different host Miller ( Campbell Museum of Natural Resources,individuals are separated by a hyphen. Clemson University, Clemson, SC); TC = T. Currin;
In the collection lists, initials of collectors are given WED= W. E. Dove; WW= William Wills( Columbia,
according to the following key: AD= Amy Dye( South SC); WWB= W. Wilson Baker.
Carolina Dept. of Natural Resources, Clemson), ARB= Throughout this work, we follow the flea
Amy R. Banks ( Georgia Southern University, classification of Lewis ( 1993) and the mammal
Statesboro); CFR= C. F. Rainwater; CWB= Craig W. classification in Wilson and Reeder ( 1993). Flea
Banks( Georgia Southern University, Statesboro); DD specimens from this survey are deposited in theD. Dunavan; DCW= Dwight C. Williams( Cypress Clemson University Arthropod Collection, Clemson,
Gardens, Moncks Corner SC); DV = David Vaughan SC, or in the Institute ofArthropodology and ParasitologySouth Carolina Dept. of Health & Environmental at Georgia Southern University, Statesboro, GA. All
Control, Columbia); EVH= E. V. Horton; GRW= G. R. of the flea species recorded here from South CarolinaWilkie III; JCH= J. C. Hartzell, Jr.; KLC = Kerry L. can be identified using the key prepared by BentonClark( University of North Florida, Jacksonville); LAD ( 1983).
CherokeeGreen-
ville Spartan- York
Pickensburg
OconeeUnion
Lancaster Chesterfield Marl-
boroAnderson
Laurens
Fairfield KershawDarlington
Dillon
NewberryAbbeville Marion
Green-
wood Florence
Mc- Saluda Richland
HorryLexington Sumter
ry
Edgefield
Calhoun
Clarendon WilliamsburgAiken George-
town
Orangeburg
BarnwellBerkeleyDor- Y.
chester
Allendale
Colleton
HamptonCharleston
Beau-
fort
100 Kilometers
Figure 1. Map of South Carolina showing county locations.

December, 1999 Journal of Vector Ecology 173
FLEA RECORDS FOR SOUTH CAROLINA
Cediopsylkisimplex( Baker, 1895)( rabbit flea)- Family cosmopolitan ectoparasite ( Lewis 1972) of veterinaryPulicidae. importance, is common in South Carolina. We recorded
This rabbit flea is widespread in eastern North it from the Virginia opossum, a fox, domestic dogs, andAmerica( Lewis 1972). It is sometimes also recovered from inside a house in South Carolina.from predators of rabbits such as foxes, coyotes, and COLLECTION RECORDS:
dogs. AIKEN CO.: 19 ex domestic dog\7 Oct. 1979.COLLECTION RECORDS: ANDERSON CO.: 29 ex " fox"\ RBC\Dec. 1939 ( FoxAIKEN CO.: ex gray fox, Urocyon cinereoargenteus 1940); Benton( 1980).
WWB\ 23 Nov. 1962( Wilson 1966). BEAUFORT CO.: 2d' ex domestic dog\ 29 May 1945-BARNWELL CO.: 2d',19 ex eastern cottontail rabbit, 19\31 May 1945( Carpenter et al. 1945).
Sylvilagus floridanus\WW & SL\21 Jan. 1998 - BERKELEY CO.: 29 in house\DCW\ 12 Aug. 1998.3d,39 ex S. floridanus\WW& SL\21 Jan. 1998 - CHARLESTON CO.: Trembley and Bishopp( 1940).19 ex S. floridanus\WW& SL\ 30 Mar. 1998. HORRY CO.: Benton( 1980).
DILLONCO.: ld'ex domestic dog, Canis lupus\EVH\ 14 PICKENS CO.:2d1,
99 ex Virginia opossum, DidelphisApr. 1965. virginiana\MB\ 28 Oct. 1998 - 19 ex domestic
MCCORMICK CO.: ld' ex marsh rabbit, Sylvilagus dog\ 1924.palustris( Bachman)\ GRW\ 20 Feb. 1999. RICHLAND CO.: 3d',69 ex domestic dog\WW\ 13
OCONEE CO.: 39 ex New England cottontail rabbit, May 1995.Sylvilagus transitionalis\AD& MB\ 23 Feb. 1999. SUMTER CO.: Trembley and Bishopp( 1940).
PICKENS CO.: Benton( 1980).
UNSPECIFIED County/ Counties: ex S. floridanus Ctenophthalmus pseudagyrtes Baker, 1904 - FamilyAndrews et al., 1980); ex red foxes, Vulpes vulpes Ctenophthalmidae.
7 of 28 animals examined were infested) that had This flea is widely distributed in eastern Northbeen illegally translocated to South Carolina from America where it parasitizes many species of smallmidwestern states for hunting purposes( Davidson mammals, particularly shrews, moles, voles, chipmunks,
et al. 1992). and native mice ( Lewis 1974a). Durden and Kollars
1997) reported that it was abundant on these hosts in
Conorhinopsylla stanfordi Stewart, 1930 - Family Tennessee.
Ctenophthalmidae. COLLECTION RECORDS:
This is a flea ofsquirrels, especially flying squirrels BARNWELL CO.: 1 d ex cotton mouse, Peromyscus
Glaucomys spp.) in eastern North American ( Lewis gossypinus\WW& SL\22 Jan. 1998.
1974a). There are few records of it from the southern CHARLESTON CO.: Benton( 1980).
United States( Benton 1980). OCONEE CO.: ld' ex" mole"\ REW\ 1950- ld',19 exCOLLECTION RECORD: pine vole, Microtus pinetorum\WW\7 Aug. 1996.PICKENS CO.: Benton( 1980). PICKENS CO.: Benton( 1980).
Ctenocephalidescanis( Curtis, 1826)( dog flea)- Family Doratopsylla blarinae C. Fox, 1914 - FamilyPulicidae. Ctenophthalmidae.
The dog flea, a cosmopolitan ectoparasite ( Lewis This flea is a widespread ectoparasite of Blarina
1972) of veterinary significance, does not appear to be spp. short- tailed shrews in eastern North Americacommon in South Carolina especially on domestic dogs. ( Durden and Kollars 1997). However, there are few
However, it may parasitize foxes and coyotes more records of it from the southeastern United States( Benton
frequently. 1980).
COLLECTION RECORDS: COLLECTION RECORDS:
CHARLESTON CO.: Trembley and Bishopp( 1940). AIKEN CO.: 12 specimens from 9 of 50 individualKERSHAW CO.: Trembley and Bishop( 1940). southeastern short- tailed shrews, Blarina
carolinensis, examined( Whitaker et al. 1994).
Ctenocephalidesfelis( Bouche, 1835)( cat flea)- FamilyPulicidae. Echidnophagagallinacea( Westwood, 1875)( sticktight
Unlike the previous species, this flea, another flea)- Family Pulicidae.

174 Journal of Vector Ecology December, 1999
This small flea is globally widespread in temperate unusual but Durden and Kollars ( 1997) recorded thisand subtropical climates( Lewis 1972). It parasitizes a flea from eastern cottontails in western Tennessee.wide variety of bird and mammal species. Evidently, it Further collecting should clarify the status of this flea inis widespread in South Carolina. South Carolina.
COLLECTION RECORDS: COLLECTION RECORDS:AIKEN, ANDERSON, BEAUFORT, CALHOUN, UNSPECIFIED County/ Counties.: ex S. floridanus
CHARLESTON, COLLETON, DARLINGTON, Andrews et al. 1980).
FLORENCE, GEORGETOWN, HAMPTON,
HORRY, JASPER, KERSHAW, LEXINGTON, Leptopsylla segnis( Schonherr, 1911)( European mouse
MARLBORO, ORANGEBURG, RICHLAND, flea) Family Leptopsyllidae.
and SUMTER Counties: Trembley and Bishopp This cosmopolitan flea mainly parasitizes the1940), Carpenter et al. ( 1945), Pratt and Good
house mouse, Mus musculus, but it is sometimes1954), Benton( 1980).
recovered from other small rodents ( Lewis 1974b). ItDILLON CO.: 19 ex feral kitten, Felis sylvestris\WW\ 1 1
appears to be widespread in South Carolina.Jul. 1996.
COLLECTION RECORDS:FLORENCE CO.: 19 ex " jaybird" \ CFR\ 1894 ( Fox
AIKEN, ALLENDALE, ANDERSON, BAMBERG,1940).
BARNWELL, BEAUFORT, BERKELEY,ORANGEBURG CO.: 99 ex horses, Equus caballus
CHARLESTON, COLLETON, DARLINGTON,UCH\Nov. 1894( Fox 1940, Hopkins and Rothschild
DORCHESTER, FAIRFIELD, GEORGETOWN,1953).
HAMPTON, HORRY, MARLBORO, ORANGE-UNSPECIFIED County/ Counties: Trembley and
BURG, and UNION Counties: Pratt and GoodBishopp ( 1940); ex blue jay, Cyanocitta cristata
1954), Benton( 1980).Boyd et al. 1956) ex horses, mules, and asses
CHARLESTON CO.: 2d' ex Mus sp.\ 2 Jan. 1938( FoxBecklund 1964). 1940)- 2d',69 ex" rat"\ 2 Feb. 1944( Carpenter et al.1945).
Epitedia cavernicola Traub, 1957 - Family Cteno BEAUFORT CO.: ld ex" rat"\ 1945 29 ex" rat"\ 1945phthalmidae. ld',39 ex" rat"\ 1945.
This is a host-specific flea of the eastern woodrat, PICKENS CO.: 19 ex house mouse, Mus mus
Neotoma floridana. Because of its nidicolous habits culus\ DD\ 18 May 1933.
and apparent winter phenology( Benton 1980) there arerelatively few records of it. The single specimen we Nosopsyllusfasciatus( Bosc, 1800)( northern rat flea)-report here was taken at fairly high altitude. Family Ceratophyllidae.COLLECTION RECORD: This flea is a cosmopolitan ectoparasite of OldPICKENS CO.: 19 ex N. floridana\MB\ 6 Nov. World domestic rats but it is typically more prevalent in
1998\ 1, 067 m elevation( NEW STATE RECORD). temperate or cooler regions ( Traub et al. 1983). It
appears to be widely distributed in South Carolina andprobably occurs wherever commensal rats are foundEpitedia wenmanni testor( Rothschild, 1915)- Family within the state.
Ctenophthalmidae. COLLECTION RECORDS:
This is a widespread North American flea thatABBEVILLE, AIKEN, BAMBERG, BARNWELL,
mainly parasitizes Peromyscus spp. mice ( LewisBEAUFORT, CALHOUN, CHARLESTON,
1974a). CLARENDON, DORCHESTER, GEORGE-
COLLECTION RECORDS: TOWN, HORRY, MARION, ORANGEBURG,
ANDERSON CO.: Benton( 1980).and RICHLAND Counties: Pratt and Good( 1954),
BARNWELL CO.: Both collections ex P. gossypinusBenton ( 1980).
by WW& SL on 25 Jan. 1998 1d, 49 4d',29. CHARLESTON CO.: 3(3, 19\ 21 Nov. 1945.UNSPECIFIED Counties: Traub et al. ( 1983).
Euhoplopsyllus glacialis affinis( Baker, 1904)- FamilyPulicidae. Odontopsyllus multispinosus ( Baker, 1898) - Family
This flea is mainly an ectoparasite of native rabbits Leptopsyllidae.and hares in the Rocky Mountain region( Lewis 1972). This rabbit flea is widespread in the eastern UnitedIts occurrence in South Carolina must be considered States ( Lewis 1974b, Benton 1980). It appears to be

December, 1999 Journal of Vector Ecology 175
fairly widely distributed in South Carolina. only recently recorded from South Carolina( Durden etCOLLECTION RECORDS: al. 1997a). Nevertheless, it appears to be quite widespreadAIKEN CO.: 2d' ex bobcat, Lynx rufus\25 Feb. 1980 on N. floridana within the state. Some of the South
specimens in coll. of R. E. Lewis). Carolina specimens have been examined by Robert E.BARNWELL CO.: 2d',39 ex S. floridanus\WW & Lewis ( Ames, IA) who has noted some morpho-
SL\21 Jan. 1998. logical variation in them( R. E. Lewis, pers. com.) butMCCORMICK CO.: Id ex S. palustris\GRW\ 20 Feb. advocates retaining them as O. s. pennsylvanicus until a
1999. systemtic revision of the genus Orchopeas has been
OCONEE CO.: 19 ex S. transitionalis\AD & MD\23 completed.
Feb. 1999. COLLECTION RECORDS:
ORANGEBURG CO.: Ex S.floridanus\WED& SCH\ 8 BARNWELL CO.: both collected from N.floridana byFeb. 1931 ( Fox 1940, Benton 1980). WW& DV as follows- ld'\25 Jun. 1996- 19\26
UNSPECIFIED County/ Counties: ex S. floridanus Jun. 1996.
Andrews et al. 1980). CHARLESTON CO.: ex N. floridana ( Durden et al.
1997a); ld ex N. floridana\WW & DV\ 18 Jun.
Orchopeas howardi ( Baker, 1895) - Family Cerato- 1996 1d ex cotton rat, Sigmodon hispidus\WW&
phyllidae. DV\ 19 Jun. 1996 - remainder collected from
This flea parasitizes tree squirrels throughout much individual N.floridana by KLC as follows- ld',39\6
of North America and is especially common in theJul. 1994 1\ 7 Sep. 1994 4d',2\ 22 Sep. 1994
central and eastern United States( Lewis 1975) on the19\ 3 Oct. 1994 9d, 12\ 19 Feb. 1995 1d, 29\25
eastern gray squirrel, Sciurus carolinensis, and foxJun. 1995.
squirrel, Sciurus niger, as discussed by Traub et al. GEORGETOWN CO.: ex N. floridana (Durden et al.
1983). It is probably widespread on these hosts and on 1997a); remainder collected from N.floridana by
the southern flying squirrel in South Carolina. KLC as follows ld\ 17 Jul. 1995- 29\20 Jul. 1995
COLLECTION RECORDS: 19\ 10 Nov. 1995 - 6d',89\29 Nov. 1995 - 19\ 1
BARNWELL CO.: 5d',89 ex southern flying squirrel, Dec. 1995 24, 19\2 Dec. 1995 7d',69\3 Dec.
Glaucomys volans\SL\ 15 Jan. 1999 - remainder1995.
collected from individual G. volans by WW& SLJASPER CO.: ld',19 ex N.floridana\ARB& CWB\ 21
as follows 1\ 30Mar. 1998 1d, 1Q\ 20May 1998Jul. 1997- remainder collected from individual N.
1d, 19\ 12 Nov. 1998 ld',19\ 12 Nov. 1998 - floridana by ARB, CWB & LAD as follows -
1d, 2\ 12 Nov. 1998. ld',19\28 Nov. 1996 - 2d,29\28 Nov. 1996
CHARLESTON CO.: Benton ( 1980). 3d',39\ 28 Nov. 1996- 19\ 29 Nov. 1996- 19\ 30
PICKENS CO.: 19 ex " squirrel"\ RBC\ 1 Nov. 1929 Nov. 1998 9d,69\ 30 Nov. 1996.
Fox 1940)- ld ex" squirrel"\ DD\ 10 Jan. 1939- PICKENS CO.: 19 ex N. floridana\MB\ 6 Nov.
19 ex" squirrel"\ TC\ 15 Nov. 1964. 1998\ 1, 067 m elevation.
Peromyscopsylla hesperomys ( Baker, 1904) - FamilyOrchopeas leucopus ( Baker, 1904) - Family Cerato- Leptopsyllidae.phyllidae. This is mainly a parasite of Peromyscus spp. mice
This flea is distributed across North America but it that is widspread in North America ( Lewis 1974b).is especially common in the eastern half of the United Nevertheless, there are few records of this flea from theStates ( Lewis 1975) as a parasite of Peromyscus spp. southeastern United States and the localities reportedmice( Durden and Kollars 1997). Surprisingly, it does here represent the most southern records documented tonot appear to be common in South Carolina. date( Benton 1980).COLLECTION RECORDS: COLLECTION RECORDS:ALLENDALE CO.: Benton ( 1980). GEORGETOWN CO.: ex P. gossypinus( Johnson andBARNWELL CO.: 19 ex P. gossypinus\WW& SL\ 23 Traub 1954- host originally recorded as Peromyscus
Jan. 1998. leucopus by these authors); Benton( 1980).SUMTER CO.: 1 d ex P. gossypinus\ KLC\ 16 Jan. 1995.
Orchopeas sexdentatuspennsylvanicus( Jordan, 1928)
Family Ceratophyllidae. Peromyscopsylla scotti I. Fox, 1939 - Family Lepto-This eastern woodrat flea is widespread in eastern psyllidae.
North America( Benton 1980, Lewis 1975), but it was Like the previous species, this flea is also mainly a

176 Journal of Vector Ecology December, 1999
parasite of Peromyscus spp. mice( Johnson and Traub hispidus\KLC\9 Sep. 1995- 19 ex S. hispidus\WW1954). It occurs throughout most of the eastern United DV\25 Sep. 1996- 6d',69 ex S. hispidus\WW&States with several records in southern states ( Benton DV\7 May 1997 - 4d',69 ex S. hispidus\WW &1980). DV\7 May 1997 - 4c5,29 ex S. hispidus\WW &COLLECTION RECORDS: DV\8 May 1997 - ld',29 ex S. hispidus\WW &BARNWELL CO.: ld',29 ex eastern harvest mouse, DV\9 May 1997- 29 ex S. hispidus\WW& DV\ 8
Reithrodontomys humulis\WW& SL\23 Jan. 1998 Aug. 1997- 19 ex S. hispidus\ WW& DV\ 8 Aug.ld ex golden mouse, Ochrotomys nuttallASL\ 15 1997- ld ex rice rat, Oryzomys palustris\KLC\22
Jan. 1999- 19 ex O. nuttalli\SL\ 15 Jan. 1999- 29 Apr. 1994- ld ex Norway rat, Rattus norvegicus\ex P. gossypinus\SL\ 13 Jan. 1999 - 19 ex P. WW& DV\7 Aug. 1997- remainder collected fromgossypinus\ SL\ 13 Jan. 1999- remainder collected individual S. hispidus by WW & DV on 7 Aug.from individual P. gossypinus by WW & SL as 1997 as follows- ld'- ld - 19- 19- 2d',19- 39.follows- 29\21 Jan. 1998- 1 d\22 Jan. 1998- 19\22 CHESTER CO.: all collected from individual S. hispidusJan. 1998 - 19\22 Jan. 1998 - ld\ 23 Jan. 1998 - by KLC on 15 Oct. 1994- ld-1c3123 Jan. 1998- 29\23 Jan. 1998- 8d', 119\ 23 Jan. CLARENDON CO.: all collected from individual S.
1998 - 29\25 Jan. 1998 - ld',29\25 Jan. 1998 - hispidus by WW & DV as follows - 19\23 Oct.ld\25 Jan. 1998- 2d',19\26 Jan. 1998. 1996- 19\23 Oct. 1996- 29\23 Oct. 1996- 1 d',19\23
GEORGETOWN CO.: ex Peromyscus sp. ( Johnson Oct. 1996- ld',29\23 Oct. 1996- ld\24 Oct. 1996and Traub 1954); Benton( 1980). ld125 Oct. 1996.
SUMTER CO.: all collected from individual P. COLLETON CO.: all collected from individual S.
gossypinus by KLC as follows - 1 d',19\ 16 Jan. hispidus by WW& DV as follows- 2d',19\25 May1995- 29\ 12 Apr. 1995. 1997 - 4d',19\ 25 May 1997 - 19\28 May 1997 -
ld',29\28 May 1997 - ld129 May 1997 - 19\29
May 1997- 19\ 29 May 1997- 19\ 29 May 1997-Polygenis gwyni ( C. Fox, 1914) - Family Rhopa- 29\29 May 1997.lopsyllidae. DILLON CO.: all collected from individual S. hispidus
This flea is mainly a parasite of the cotton rat, S. by WW & DV as follows - ld\ 16 Jul. 1996 -
hispidus, although it is also frequently recovered from 2d',29\ 18 Jul. 1996.the ecological associates of this rodent and on Virginia GEORGETOWN CO.: ex N. floridana ( Durden et al.
opossums ( Lewis and Lewis 1994a) throughout much 1997a); ld' ex N.floridana\KLC\ 20 Jul. 1995- 1 d'of the southern United States. It was the most abundant ex N. floridana\KLC\31 Aug. 1995 - 19 ex S.flea during our survey being collected in all months hispidus\WW& DV\ 12 Sep. 1996 - 2d',39 ex S.except November and December and from several rodent hispidus\WW& DV\ 12 Sep. 1996.species, but particularly from S. hispidus. HAMPTON CO.: ld' ex P. gossypinus\WW& DV\ 25
COLLECTION RECORDS: Apr. 1997- remainder collected from individual S.BEAUFORT CO.: Benton( 1980); remainder collected hispidus by WW & DV as follows - 4d\ 22 Apr.
from individual S. hispidus by WW & DV as 1997- 2d',39\22 Apr. 1997- 8d',49\22 Apr. 1997follows- 5d',29\ 1 Oct. 1996- ld',19\ 1 Oct. 1996- 29\23 Apr. 1997- ld',39\23 Apr. 1997- 2d',39\ 232d',19\ 1 Oct. 1996- ld',19\ 2 Oct. 1996- 29\2 Oct. Apr. 1997- 6d',19\ 23 Apr. 1997- 5d',59\23 Apr.1996- 1 d\2 Oct. 1996- 19\ 2 Oct. 1996- 3d13 Oct. 1997- 7d',59\23 Apr. 1997- 1( 1, 19\25 Apr. 19971996. 3d',29\ 25 Apr. 1997 - 2d',39\26 Apr. 1997 -
BARNWELL CO.: 39 ex S. hispidus\ SL\ 15 Jan. 1999. 2d',39\26 Apr. 1997 - 1 ld',59\26 Apr. 1997 -BERKELEY CO.: all collected from individual S. 7d',49\ 25 May 1997 - 2d',29\ 17 Jun. 1997 -
hispidus by WW & DV as follows - ld\ 15 Oct. 3d',29\ 17 Jun. 1997- 19\ 19 Jun. 1997- 19\ 19 Jun.1996- 19\ 16 Oct. 1996- 2d\ 16 Oct. 1996. 1997 - 29\ 19 Jun. 1997 - ld',19\ 19 Jun. 1997 -
CHARLESTON CO.: ex N. floridana ( Durden et al. 39\ 19Jun. 1997- 5d',39\ 19Jun. 1997- 13d',129\ 191997a); ld'exN.floridana\KLC\ 16Sep. 1994- 19 Jun. 1997 - ld',29\20 Jun. 1997 - 5d',79\20 Jun.ex N. floridana\KLC\23 Jan. 1995 - 19 ex N. 1997- 7d',89\20 Jun. 1997.floridana\KLC\ 19 Feb. 1995- ld'ex N.floridana\ JASPER CO.: ld',19 ex N. floridana\ARB, CWB &KLC\ 18 Mar. 1995- Id' ex N.floridana\ KLC\20 LAD\28 Nov. 1996.
Jun. 1995- 29 ex S. hispidus\ KLC\21 Apr. 1994- McCORMICK CO.: all collected from individual S.
19 ex S. hispidus\KLC\7 Sep. 1995 - ld ex S. hispidus by WW& DV on 31 Jul. 1996 as followshispidus\ KLC\ 7 Sep. 1995 - ld',19 ex S. ld',39- 4d',19- 4d',19- 10d',109.

December, 1999 Journal of Vector Ecology 177
NEWBERRY CO.: 19 ex S. hispidus\WW\23 Jan. rodents, especially Peromyscus spp. and Microtus spp.,1999. in eastern North America( Lewis 1974a). It appears toORANGEBURG CO.: all collected from individual S. be common on P. gossypinus in South Carolina especiallyhispidus by KLC as follows - 11112 Sep. 1994 - during the cooler months.
3d',39\ 13 Sep. 1994. COLLECTION RECORDS:BARNWELL CO.: Id' ex P. gossypinus\ SL\ 13 Jan.
Pulex irritans L., 1758( human flea)- Family Pulicidae. 1999 - remainder collected from individual P.Prior to 1958, P. irritans and P. simulans were not gossypinus by WW & SL as follows - Id',29\21
recognized as separate species( Smit 1958); instead, all Jan. 1998 - ld\23 Jan. 1998 - 19\23 Jan. 1998 -specimens were assigned to P. irritans. Therefore, it 2d123 Jan. 1998- 39\23 Jan. 1998- ld',29\23 Jan.cannot be definitively stated that the records cited below 1998- 49\23 Jan. 1998- 21125 Jan. 1998- 39\25are attributable to P. irritans. Nevertheless, given its Jan. 1998- 19\ 21 Nov. 1998.almost cosmpolitan distribution ( Lewis 1972), it is CHESTER CO.: 1d ex O. palustris\ KLC\ 10 Nov. 1994likely that P. irritans occurs in South Carolina. remainder collected from individual S. hispidusCOLLECTION RECORDS: on 11 Nov. 1994 by KLC as follows- 1 d'- ld'- 19.CHARLESTON CO.: Trembley and Bishopp( 1940). EDGEFIELD CO.: Benton( 1980).FLORENCE CO.: Trembley and Bishop( 1940). PICKENS CO.: ld',19\20 Sep. 1976.
RICHLAND CO.: 1 in pitfall trap\ LR\Feb. 1999.Pulex simulans Baker, 1895 - Family Pulicidae. SUMTER CO.: 1d' ex P. gossypinus\ KLC\ 16 Jan.
This New World flea parasitizes a wide spectrum of 1995.
mammalian hosts, especially carnivores, deer, and largerodents( Hopla 1980). It appears to be widely distributed,especially on carnivores, in the southern United States Sternopsylla distincta texana ( Rothschild, 1903) -Durden and Kollars 1997). Family Ischnopsyllidae.
COLLECTION RECORDS: This is the only species of bat flea recorded fromAIKEN CO.: Id ex U. cinereoargenteus\ WWB\ 23
South Carolina ( Benton 1980). Its range appears to
Nov. 1962( Wilson 1966); Benton( 1980), include much of the southern United States( Lewis andOCONEE CO.: 4d,29 ex coyote, Canis latrans\SM\ 13 Lewis 1994b) in tandem with that of its principal host,
Mar. 1998. the Brazilian free- tailed bat, Tadarida brasiliensis.PICKENS CO.: ld' ex domestic dog\ DD\7 Feb. 1932.
COLLECTION RECORD:
UNSPECIFIED County/Counties: ex U. cinereo CHARLESTON CO.: Benton ( 1980); same collectionargenteus( 5 of 18 animals examined were infested) cited by Lewis and Lewis( 1994b).
that had been illegally translocated to South Carolinafrom midwestern states for hunting purposes Xenopsylla cheopis ( Rothschild, 1903) ( Oriental ratDavidson et al. 1992). flea)- Family Pulicidae.
The Oriental rat flea, virtually cosmopolitan inRhadinopsylla orama Smit, 1957 - Family Cteno- distribution, is especially common in warmer climatesphthalmidae. Lewis 1972). It is principally an ectoparasite of Old
This eastern North American vole flea ( Lewis World domestic rats, with many records from these1974a) was collected at the Savannah River Site in rodents in sea ports such as Charleston, South Carolina.South Carolina close to the Georgia border. This COLLECTION RECORDS:represents the most southern record of this flea AIKEN, ALLENDALE, BAMBERG, BARN-documented to date and is several hundred kilometers WELL, BEAUFORT, BERKELEY, CALHOUN,south of the next closest record in northeastern Tennessee CHARLESTON, DORCHESTER, FLORENCE,Benton 1980, Durden and Kollars 1997). GEORGETOWN, HAMPTON, HORRY,
COLLECTION RECORD: JASPER, KERSHAW, MARION, NEWBERRY,BARNWELL CO.: 19 ex P. gossypinus\WW& SL\ 22 ORANGEBURG, and RICHLAND Counties:
Jan. 1998( NEW STATE RECORD). Carpenter et al. ( 1945), Pratt and Good ( 1954),Benton ( 1980).
Stenoponia americana( Baker, 1899)- Family Cteno- BEAUFORT CO.: 2d ex " rat"\ 1945 - 29 ex " rat"\phthalmidae. 1945.
This large flea is relatively common on small UNSPECIFIED County: 19\ 22 Mar. 1960.

178 Journal of Vector Ecology December, 1999
HOST-FLEA LIST FOR Eastern cottontail rabbit, Sylvilagus floridanus:
SOUTH CAROLINACediopsylla simplex, Euhoplopsyllus
glacialis affinis, Odontopsyllus
multispinosus.
Blue jay, Cyanocitta cristata: Echidnophaga Marsh rabbit, Sylvilagus palustris: Cediopsylla
gallinacea. simplex, Odontopsyllus multispinosus.
Virginia oposuum, Didelphis virginiana: New England cottontail rabbit, Sylvilagus
Ctenocephalides felis. transitionalis: Cediopsylla simplex,
Bat": Sternopsylla distincta texana. Odontopsyllus multispinosus.
Southeastern short- tailed shrew, Blarina carolinensis: Unspecified hosts: Ctenocephalides canis, Pulex
Doratopsylla blarinae. irritans.
Mole": Ctenophthalmus pseudagyrtes.
Domestic cat, Felis sylvestris: Echidnophaga
gallinacea. DISCUSSION
Domestic dog, Canis lupus: Cediopsylla simplex,Ctenocephalides felis, Pulex simulans. The Flea Fauna of South Carolina
Bobcat, Lynx rufus: Odontopsyllus multispinosus. We have documented a relatively rich fauna of 25Fox": Ctenocephalides felis. species of fleas from South Carolina. This compares
Gray fox, Urocyon cinereoargenteus: Cediopsylla favorably with the 19 recorded flea species for adjoiningsimplex, Pulex simulans. Georgia and 17 species for North Carolina ( Benton
Red fox, Vulpes vulpes: Cediopsylla simplex. 1980), but less impressively with the 33 flea speciesCoyote, Canis latrans: Pulex simulans. known from adjacent Tennessee( Durden and Kollars,
Horse, Equus caballus, mule and ass: Echidnophaga 1997). We hypothesize that approximately 5 additionalgallinacea. flea species are denizens of South Carolina' s higher
Norway rat, Rattus norvegicus: Polygenis gwyni. elevations in the western part of the state. Further
Rat": Leptopsylla segnis, Nospsyllus fasciatus, collecting in Appalachian South Carolina will provide aXenopsylla cheopis. test of this assumption.
House mouse, Mus musculus: Leptopsylla segnis. Whereas some of the fleas we recorded are
Cotton rat, Sigmodon hispidus: Orchopeas widespread in South Carolina, others appear to be
sexdentatus pennsylvanicus, Polygenis gwyni, characteristic of coastal plain or montane zones. South
Stenoponia americana. Carolina' s widespread flea taxa appear to include C.
Eastern woodrat, Neotoma floridana: Epitedia simplex, C.fells, E.gallinacea, X.cheopis, S. americana,
cavernicola, Orchopeas sexdentatus L. segnis, N.fasciatus, O. sexdentatus pennsylvanicus,
pennsylvanicus, Polygenis gwyni. and perhaps also C. pseudagyrtes, O. multispinosus, P.
Rice rat, Oryzomys palustris: Polygenis gwyni, scotti, and O. howardi. Montane flea species that we
Stenoponia americana. recorded in South Carolina are C. stanfordi and E.
Cotton mouse, Peromyscus gossypinus: cavernicola. One flea species, P. gwyni, was distinctlyCtenophthalmus pseudagyrtes, Epitedia more common in coastal plain habitats, although it was
wenmanni testor, Orchopeas leucopus, also recorded from other physiographic zones.
Peromyscopsylla hesperomys, Peromyscopsylla Polygenis gwyni was clearly the most abundant fleascotti, Polygenis gwyni, Rhadinopsylla orama, in our survey and it was mainly associated with theStenoponia americana. cotton rat. This species, and the cat flea, C.felis, appear
Peromyscus sp.": Peromyscopsylla scotti. to be the 2 most common fleas in South Carolina.
Golden mouse, Ochrotomys nuttalli: Multiple flea species infestations were recorded for
Peromyscopsylla scotti. several mammalian hosts, most notably for the cottonEastern harvest mouse, Reithrodontomys humulis: mouse which hosted 8 flea species, but also for the
Peromyscopsylla scotti. domestic dog, cotton rat, eastern woodrat, and easternPine vole, Microtus pinetorum: Ctenophthalmus cottontail, each of which was host to 3 flea species.
pseudagyrtes. Also, Old World domestic rats( Rattus spp.) appear to beSquirrel": Conorhinopsylla stanfordi, Orchopeas parasitized by 4 flea species in South Carolina but host
howardi. data are not sufficiently definitive to be certain.Southern flying squirrel, Glaucomys volans: Some of the flea species we report from South
Orchopeas howardi. Carolina have medical and/ or veterinary importance.

December, 1999 Journal of Vector Ecology 179
Several of the species, such as C. canis, C. fells, E. Several members of the apicomplexan protozoangallinacea, and P. irritans will feed on pets, livestock genus Hepatozoon have been detected in fleas and someanimals, or humans, and may reach pest status. are known to be transmitted by fleas( Smith 1996). Of
relevance to flea species and hosts recorded in SouthSouth Carolina Fleas as Potential Vectors Carolina, is the squirrel parasite, Hepatozoon sciuri,
Some of the flea species we recorded are proven or which has been found in 0. howardi. Hepatozoonsuspected vectors of zoonotic pathogens. One of these erhardovae can be transmitted by 5 species of fleas topathogens, Rickettsia typhi, the etiologic agent ofmurine European bank voles( Krampitz and Wongchari 1980)typhus, is usually transmitted by X.cheopis, but C.fells, so it is likely that additional species of Hepatozoon areL. segnis, N.fasciatus, and P. gwyni, all of which were transmitted to other mammals by various species ofrecorded in this survey, can also transmit this pathogen fleas.Azad 1990). Murine typhus was formerly a widespread Some trypanosomes are similarly known to be
and relatively common infectious disease in most transmitted to rodents or lagomorphs by fleas. Molyneuxsouthern states including South Carolina, but human ( 1970) discussed 11 species of trypanosomes known tocases declined significantly after the 1940s largely as a be transmitted by fleas to hosts belonging to these tworesult ofrat control and rat-flea control programs( Mohr Orders. Of relevance to the flea species and hosts weet al. 1953, Pratt 1958, Traub et al. 1978). Although recorded for South Carolina, he listed N.fasciatus andPolygenis gwyni has sometimes been perceived as an X.cheopis as vectors ofTrypanosoma lewisi to domesticimportant historical vector of R. typhi in the southern rats, N.fasciatus as a vector of both T. microti and T.United States, its role as a vector of this pathogen in evotomys to voles, and C. simplex as a vector of annature may have been overestimated according to Lewis unnamed trypanosome to the eastern cottontail.and Lewis( 1994a). Murine typhus probably still persists Other pathogens detected in fleas in the southernin enzootic mammal and flea foci in parts of South United States, such as Borrelia burgdorferi, the etiologicCarolina and it could still represent a zoonotic threat agent of Lyme disease, are not thought to be transmittedwithin the state.
by fleas but may instead reflect a recent bloodmeal byAnother flea-borne rickettsia] agent, Rickettsiafelis, these fleas from, for example, a spirochetemic hostwhich causes murine typhus- like disease, is also almost ( Teltow et al. 1991). Similarly, although plague is notcertainly present in South Carolina. It has been detected present in South Carolina, potential flea vectors,in cat fleas recovered from opossums, bobcats, and especially X. cheopis, are evidently common in thedomestic dogs in neighboring Georgia and is also known state. Overall, we advocate that fleas have a persistentfrom cat fleas collected in other southern states( Azad et but underestimated role as vectors of pathogens toal. 1997).
humans, pets, livestock, and wild mammals in SouthSome species of Bartonella may be transmitted by Carolina.
fleas in South Carolina. One of these is Bartonellahenselae, the causative agent of cat scratch disease, Acknowledgmentswhich can be transmitted by cat fleas in the laboratoryChomel et al. 1996). Several other species ofBartonella We thank all of the flea collectors listed in the
are known to infect rodents in the southeastern United Materials and Methods section who supplied specimensStates( Kosoy et al. 1997) and other species of fleas may for this survey. We are grateful to M. Bunch ( Southbe vectors of these agents. For example, 0. leucopus is Carolina Department of Natural Resources, Clemson),known to be a laboratory vector ofBartonella peromysci S. Loeb, E. Ridley, and G. Chapin( U. S. Forest Service,to mice( Katavolos and Telford 1998). Clemson), and D. Vaughan( South Carolina Department
The strain of Rickettsia prowazekii that causes of Health and Environmental Control, Columbia) forsylvatic epidemic typhus in humans and asymptomatic collecting fleas for us during their mammal trappingtransient infections in its enzootic southern flying squirrel projects. We also thank John Morse, Curator of thereservoir host( Bozeman et al. 1975) is almost certainly Clemson University Arthropod Collection, for allow-present in South Carolina. McDade ( 1987) recorded ing us to examine the flea holdings, Allen Bentonhuman infections by this rickettsia] agent from several ( Fredonia, NY) for providing additional information onstates including adjacent Georgia, North Carolina, and flea collection localities depicted in his flea atlas, J.Tennessee. In the laboratory, the squirrel flea, 0. Ciegler and V. Winn for allowing us to reproducehowardi, which we found to be widespread in South their map of South Carolina( Fig. 1), and G. Bowers andCarolina, is a competent vector of this pathogen( McDade R. Bowers for allowing WW to trap mammals on their1987).
property. We are also grateful to Robert E. Lewis

180 Journal of Vector Ecology December, 1999
Ames, IA) for examining representative specimens Dis. 28: 581- 589.
of Orchopeas sexdentatus, allowing us to document Durden, L. A., C. W. Banks, K. L. Clark, B. V. Belbey,his unpublished flea records from a South Carolina and J. H. Oliver, Jr. 1997a. Ectoparasite fauna of
bobcat, and commenting on an earlier draft of this the eastern woodrat, Neotoma floridana:
paper. composition, origin, and comparison with
ectoparasite faunas of western woodrat species. J.
Parasitol. 83: 374-381.
REFERENCES CITED Durden, L. A. and T. M. Kollars, Jr. 1997. The fleas
Siphonaptera) of Tennessee. J. Vector Ecol.
Andrews, C. L., R. R. Gerrish, and V. F. Nettles. 1980. 22( 1): 13- 22.
Ectoparasites collected from eastern cottontails in Durden, L. A., R. G. McLean, J. H. Oliver, Jr., S. R.
the southeastern United States. J. Med. Entomol. Ubico, and A. M. James. 1997b. Ticks, Lyme
17: 479- 480. disease spirochetes, trypanosomes, and antibody to
Azad, A. F. 1990. Epidemiology of murine typhus. encephalitis viruses in wild birds from coastal
Annu. Rev. Entomol. 35: 553- 569. Georgia and South Carolina. J. Parasitol. 83: 1178-
Azad, A. F., S. Radulovic, J. A. Higgins, B. H. Noden, 1182.
and J. M. Troyer. 1997. Flea-borne rickettsioses: Fox, I. 1940. Fleas of eastern United States. Iowa State
ecologic considerations. Emerging Inf. Dis. 3: 319- College Press, Ames, 191 pp.327. Hopkins, G. H. E. and M. Rothschild. 1953. An
Becklund, W. W. 1964. Revised checklist of internal illustrated catalogue of the Rothschild Collection
and external parasites of domestic animals in the of Fleas ( Siphonaptera) in the British Museum
United States and Possessions and in Canada. Am. Natural History) with keys and short descrip-J. Vet. Res. 25: 1380- 1416. tions for the identification of families, genera,
Benton, A. H. 1980. An atlas of the fleas of the eastern species and subspecies. Vol. I. Tungidae and
United States. Marginal Media, Fredonia, NY, 177 Pulicidae. Trustees of the British Museum, London,
pp. 361 pp.Benton, A. H. 1983. An illustrated key to the fleas of Hopla, C. E. 1980. A study of the host associations and
the eastern United States. Bioguide No. 3. Marginal zoogeography of Pulex, Pp. 185- 207 in Fleas.Media, Fredonia, NY, 34 pp. Proceedings of the International Conference on
Bosik, J. J. 1997. Common names of insects and related Fleas( R.Traub and H. Starcke, eds.). A.A. Balkema.
organisms. Entomological Society of America, Rotterdam, 420 pp.Lanham, MD, 200 pp. Johnson, P. T. and R. Traub. 1954. Revision of the flea
Boyd, E. M., R. L. Diminno, and C. Nesslinger. 1956. genus Peromyscopsylla. Smithson. Misc. Coll.
Metazoan parasites of the blue jay, Cyanocitta 123( 4): 1- 68.
cristata. J. Parasitol. 42: 332- 346. Katavolos, P. and S. R. Telford. 1998. Flea transmission
Bozeman, F. M., S. A. Masiello, M. S. Williams, and B. ofBartonella( Grahamella) peromysciTyzzer. Am.
L. Elisberg. 1975. Epidemic typhus rickettsiae J. Trop. Med, Hyg.( Suppl.). 59: 334- 335( Abstract).isolated from flying squirrels. Nature, Lond. 255: Kosoy, M. L., R. L. Regnery, T. Tzianabos, E. L.545- 547. Marston, D. C. Jones, D. Green, G. O. Maupin, J. G.
Carpenter, S. J., R. W. Chamberlain, and R. Baker. Olson, and J. E. Childs. 1997. Distribution,
1945. Flea collections at Army Installations in the diversity, and host specificity of Bartonella inFourth Service Command. J. Econ. Entomol. 38: rodents from the southeastern United States. Am. J.
600- 602. Trop. Med. Hyg. 57: 578- 588.Chomel„ B. B., R. W. Kasten, K. A. Floyd-Hawkins, B. Krampitz, H. E. and V. Wonchari. 1980. The
Chi, K. Yamamoto, J. Roberts- Wilson, A. N. development of Hepatozoon erhardovae in
Gurfield, R. C. Abbot, N. D. Pederson, and J. E. experimental mammalian and arthropod hosts. 1.
Koehler. 1996. Experimental transmission of The evaluation of suitable arthropod vectors, Pp.Bartonella henselae by the cat flea. J. Clin. 349- 358 in Fleas. Proceedings of the International
Microbiol. 23: 1952- 1956. Conference on Fleas ( R. Traub and H. Starcke,
Davidson, W. R., M. J. Appel, G. L. Doster, O. E. Baker, eds.). A. A. Balkema, Rotterdam, 420 pp.and J. F. Brown. 1992. Diseases and parasites of Lewis, R. E. 1972. Notes on the geographical distribution
red foxes, gray foxes, and coyotes from commercial and host preferences in the Order Siphonaptera.
sources selling to fox-chasing enclosures. J. Wildl. Part 1. Pulicidae. J. Med. Entomol. 9: 511- 520.

December, 1999 Journal of Vector Ecology 181
Lewis, R. E. 1974a. Notes on the geographical Sci. 70: 516- 527.distribution and host preferences in the Order Pratt, H. D. and N. E. Good. 1954. Distribution ofsomeSiphonaptera. Part 3. Hystrichopsyllidae. J. Med. common domestic rat ectoparasites in the UnitedEntomol. 11: 147- 167. States. J. Parasitol. 40: 113- 129.
Lewis, R. E. 1974b. Notes on the geographical Smit, F. G. A. M. 1958. A preliminary note on thedistribution and host preferences in the Order occurrence ofPulex irritans L. and Pulex simulansSiphonaptera. Part 5. Ancistropsyllidae, Baker in North America. J. Parasitol. 44: 523- 526.Chimaeropsyllidae, Ischnopsyllidae, Leptop- Smith, T. G. 1996. The genus Hepatozoon ( Api-syllidae and Macropsyllidae. J. Med. Entomol. 11: complexa: Adeleina). J. Parasitol. 82: 565- 585.525- 540.
Teltow, G. J., P. V. Fournier, and J. A. Rawlings. 1991.Lewis, R.E. 1975. Notes on the geographical distribution Isolation of Borrelia burgdorferi from arthropods
and host preferences in the Order Siphonaptera. collected in Texas. Am. J. Trop. Med. Hyg. 44:Part 6. Ceratophyllidae. J. Med. Entomol. 11: 658- 469-474.676.
Traub, R., M. Rothschild, and J. Haddow. 1983. TheLewis, R. E. 1993. Notes on the geographical distribution Rothschild Collection of Fleas. The Cerato-
and host preferences in the Order Siphonaptera. phyllidae: key to the genera and host relationshipsPart 8. New taxa described between 1984 and 1990,
with notes on their evolution, zoogeography andwith a current classification of the Order. J. Med. medical importance. Academic Press, London, 288Entomol. 30: 239-256.
pp.Lewis, R. E. and J. H. Lewis. 1994a. Siphonaptera of Traub, R., C. L. Wisseman, Jr., and A. F. Azad. 1978.
North America north of Mexico: Vermipsyllidae The ecology of murine typhus - a critical review.and Rhopalopsyllidae. J. Med. Entomol. 31: 82- 98. Trop. Dis. Bull. 75: 237- 317.
Lewis, R. E. and J. H. Lewis. 1994b. Siphonaptera of Trembley, H. L. and F. C. Bishopp. 1940. DistributionNorth America north of Mexico: Ischnopsyllidae. and hosts of some fleas of economic importance. J.J. Med. Entomol. 31: 348- 368. Econ. Entomol. 33: 701- 703.
McDade, J. E. 1987. Flying squirrels and their Whitaker, J. 0.,Jr., G. D. Hartman, and R. Hein. 1994.ectoparasites: disseminators of epidemic typhus. Food and ectoparasites of the southern short-Parasitol. Today 3: 85- 87.
tailed shrew, Blarina carolinensis ( Mammalia:Mohr, C. 0., N. E. Good, and J. H. Schubert. 1953. Soricidae), from South Carolina. Brimleyana 21:
Status of murine typhus infection in domestic rats 97- 105.in the United States, 1952, and relation to infestation Wilson, D. E. and D. M. Reeder ( editors). 1993.by Oriental rat fleas. Am. J. Publ. Hlth. 43: 1514- Mammal species of the world. A taxonomic and1522.
geographic reference. 2nd. edn. Smithsonian Inst.Molyneux, D. H. 1970. Developmental patterns in Press, Washington DC, 1, 206 pp.
trypanosomes of the subgenus Herpetosoma. Ann. Wilson, N. 1966. A new host and range extension forSoc. Belge Med. Trop. 50: 229- 238. Pulex simulans Baker with a summary ofpublishedPratt, H. D. 1958. The changing picture of murine records( Siphonaptera: Pulicidae). Am. Midl. Nat.typhus in the United States. Ann. New York Acad. 75: 245- 248.

Journal of Vector Ecology 24( 2): 182- 186
Experimental Studies of Interactions
Between Wild Turkeys and Black-Legged Ticks
Richard S. Ostfeld and David N. Lewis
Institute of Ecosystem Studies, Box AB, Route 44AMillbrook, NY 12545 U.S.A.
Received 14 April 1999; Accepted 21 June 1999
ABSTRACT: Wild turkeys are increasing in abundance and distribution in eastern North America, buttheir potential role as hosts for ticks, or as predators on ticks, is unknown. We performed two experiments,one to determine whether juvenile black- legged ticks( Ixodes scapularis) feed successfully on turkeys, andthe other to determine if turkeys depredate adult black- legged ticks in forest habitats. Of 550 larval ticksplaced directly on 5 captive wild turkeys, none engorged and only 7( 1. 3%) were recovered; the remainder
apparently were consumed during preening. Of 165 nymphal ticks placed on the turkeys, 5 engorged and8 unengorged ticks were collected; 152( 93. 3%) were apparently consumed. Of 250 adult ticks introducedinto forest enclosures exposed to turkey foraging, 89. 5% were recaptured, which was not significantly
different from the 92. 2% recaptured in control enclosures from which turkeys were excluded. We concludethat wild turkeys are unlikely to host juvenile black- legged ticks in nature, and that turkey foraging isunlikely to reduce local density of adult ticks.
Keyword Index: Host, Lyme disease, parasite- host interactions, vector.
INTRODUCTION in the emergence of Lyme disease, a zoonotic diseasetransmitted by tick vectors ( Ixodes spp.)( Barbour and
Historically, the wild turkey( Meleagris gallopavo) Fish 1993, Ostfeld 1997). Forest floor vertebrates play
was an abundant member of vertebrate communities key roles in the Lyme-disease epizootic both as hosts forwithin forests and grasslands of eastern and central ticks and as sources from which feeding ticks obtain theNorth America (Healy 1992). However, this species bacterium( Borreliaburgdo rferi) that causes the disease
was decimated throughout its historical range during the ( Lane et al. 1991, Piesman and Gray 1994). Because
18th through early 20th centuries as a result of hunting wild turkeys are abundant, large, and active on the forest
and habitat destruction( Kennamer et al. 1992). Regrowth floor where ticks seek hosts, this species may interactof forests following deforestation, combined with strongly with Ixodes ticks. If turkeys serve as hosts forreintroductions, transplantations, and subsequent ticks or as reservoirs of Lyme-disease bacteria, they
population growth and dispersal of the birds, has resulted may enhance the density and infection prevalence of thein rapid increases in both density and range of this ticks, thus increasing risk of human exposure to Lymespecies. Between about 1930 and 1990, the turkey disease. Alternatively, if turkeys consume the ticks theypopulation in the eastern United States and southeastern encounter while foraging, they may reduce tick densityCanada increased from extreme scarcity to more than and hence disease risk. We are not aware of any studies2.5 x
106 individuals, which may equal or exceed the on trophic interactions between turkeys and Ixodes
abundance they maintained in precolonial times ticks. To evaluate the possibility that turkeys mayKennamer et al. 1992). influence the abundance or infection prevalence of
Regrowth of eastern forests and the consequent ticks, we undertook a study to determine experimentallyexpansion of populations of other vertebrates, most whether and how these species interact.
notably white-tailed deer ( Odocoileus virgianus) and In summer and autumn 1998, we conducted two
white- footed mice( Peromyscus leucopus), have resulted experiments on trophic interactions between turkeys

December, 1999 Journal of Vector Ecology 183
and black-legged ticks( Ixodes scapularis— formerly I. to reduce the possibility that turkeys would immediatelydammini), the primary vector ofLyme disease in eastern remove ticks by preening. The turkeys were then heldand central North America. In the first experiment, we individually for five days ( 125 hours) in wire-meshsought to determine whether turkeys are suitable hosts cages( 40 cm wide by 56 cm long by 80 cm high) thatfor juvenile ticks. If so, our intention was to determine were suspended over tubs of water. Cages were covered
whether turkeys are a competent reservoir for Borrelia by plastic tarps for protection and shading. The turkeysburgdorferi. In the second experiment, we asked whether had ad lib access to food and water through a 10 cm byturkeys depredate unfed adult ticks and consequently 15 cm opening in the front of each cage. The tubs ofreduce their abundance within local areas. water beneath cages were examined for fed or unfed
ticks once each day for five days. We used a five-dayMATERIALS AND METHODS criterion because juvenile ticks may require up to a day
to attach and typically remain attached to hosts for aboutExperimental Animals three days( Lane et al. 1991, Ostfeld 1997). After a two-
Five yearling female eastern wild turkeys were week resting period, we placed 1101arval and 6 nymphalobtained in June 1998 from Quattro' s Game Birds in I. scapularis on each turkey ( again, on the nape of thePleasant Valley, NY. The breeding stock consisted of neck), returned the turkeys to their individual cages, and
10th generation turkeys originating from wild-caught repeated the daily collection procedure for seven daysbirds native to Wisconsin, and was bred periodically ( 168 hours).
with wild stock captured in Massachusetts. These birds
were hatched in incubators and raised in large groups. Experiment II- Turkeys as Predators on Ticks
We housed the five birds together in a 2m wide by 5m The purpose of this experiment was to evaluate the
long by 2m high coop made of welded wire on a wood effectiveness ofturkeys as predators on adult/. scapularis
frame and containing a wooden perch and rain shelter. in a natural forest habitat. We used adult ticks because
The coop was placed in a native old- field at the Institute this largest life stage( ca 2 mm long in I. scapularis) isof Ecosystem Studies in Milibrook, NY, - 500m from the most likely to be visible and attractive to turkeys asthe experimental forest plots( see below). Turkeys were food. To this end, we established six 10 m by 10 msupplied with commercial turkey feed, scratch grain, enclosures in a mature oak forest site about 500 m from
and water ad lib. the turkey coop. The oak forest sites are described
Host- seeking ticks ( Ixodes scapularis) were thoroughly in Ostfeld et al. ( 1995, 1996a). Enclosures
obtained for both experiments by dragging a 1 m2
piece were arranged as three blocks with 30- 100 m between
of white corduroy cloth in a forested site- 3 km from the blocks and 15 m between the two enclosures within each
experimental site. Ticks were placed in glass vials with block. Enclosures were constructed of plastic snow-
moistened plaster of paris, and were used within five fencing 2.5 m high that was supported at the corners bydays ofcollection. The same collection and maintenance stapling the fence to tree trunks. For each pair of
procedure in prior experiments resulted in high feeding enclosures within a block, we used a coin toss to
success of larval ticks on two species of Peromyscus determine which would be the experimental ( with
mice( Hazier and Ostfeld 1995). turkeys) and control( no turkeys) unit.
We performed a pilot experiment in October 1998,
Experiment I- Turkeys as Hosts for Ticks in which we introduced 50 adult ticks( 1: 1 sex ratio) into
The primary purpose of this experiment was to the center of one of the experimental enclosures and
determine whether juvenile( larval and nymphal) black- allowed them to acclimate for 15 hours. We then
legged ticks can feed successfully on turkeys. In the introduced two turkeys into the enclosure and left them
initial phase, we used nymphal ticks known from our undisturbed for eight hours. The turkeys were then
prior studies at this site to have a 30-40% infection placed individually in cages suspended over water forprevalence with Lyme-disease bacteria( R. S. Ostfeld, 48 hours( see methods for Experiment 1) to determine
unpublished data). If nymphal ticks fed successfully, whether any of the ticks had parasitized the turkeys. Weour secondary purpose was to determine using then estimated the number of ticks that remained in the
xenodiagnosis the probability that uninfected larval forest plots using a standard drag- sampling techniqueticks feeding two weeks later would acquire the bacteria. ( e. g., Falco and Fish 1992). We exhaustively sampled
In August 1998, we placed 27 nymphal I.scapularis the forest floor of both experimental and control
ticks on each of the five turkeys. While the birds were enclosures by dragging a 1 m2
corduroy cloth inrestrained, ticks were placed on the nape of the turkeys' concentric circles, checking the cloth for ticks every 10necks with a fine brush. The nape of the neck was used paces. Because recovery rates were low, we followed

184 Journal of Vector Ecology December, 1999
the pilot experiment with an additional experiment nymphs placed on each of the five turkeys ( 135 ticks
using carbon dioxide-baited tick traps( Falco and Fish altogether), only six( 4.4%) were recovered. On average,
1992) in addition to drag- sampling. 1. 20( 0.74( SE; range 0- 4) ticks per bird were recoveredWe performed the full experiment in early to mid after 125 hours. Of the six nymphs that were recovered,
November, 1998, which is the time of peak activity of four were engorged and two were unfed; the fouradult ticks at our sites ( Ostfeld et al. 1996a,b). We engorged nymphs all came from one of the birds.
placed 100 adult ticks( 1: 1 sex ratio) in the center ofeach Of the 110 larvae and six nymphs placed on eachof the six enclosures and allowed them to acclimate for turkey in phase two( 580 ticks altogether), only 15 ticks15 hours. We then released two turkeys into each of the ( 2.6%) were recovered after 168 hours. None of the 550three experimental enclosures and left them undisturbed larvae engorged; all seven of the larvae recovered werefor 8 hours per day for the following two days( 16 hours unfed( mean= 1. 40± 1. 17 larvae per bird). Of the eighttotal). Immediately following the 16 hours of foraging, nymphs that were recovered, only one was engorgedthe birds were placed in individual cages suspended ( TABLE 1). A thorough visual examination of one ofover water for 48 hours to collect any ticks having the turkeys immediately following the seven days overparasitized the turkeys. The morning after turkey water pans revealed no ticks. Because of the low
removal, three CO2- baited tick traps were placed 2.5 m success rate in feeding nymphs to repletion on the birds,from the center of each enclosure, with at least 4.5 m and because no engorged larvae were recovered, webetween adjacent traps. Each trap was charged with- 1 could not conduct a xenodiagnosis to test for the reservoir
kg of dry ice and was checked after seven hours. competence of wild turkeys.
Immediately after the traps were checked, we drag-sampled the entire plots to collect any remaining ticks. Experiment II- Turkeys as Predators on TicksWe also drag- sampled the outer perimeter of the Turkeys were ineffective predators on adult I.enclosures to determine whether any ticks had escaped. scapularis ticks. On average, 179 adult ticks wereThe following day the tick traps were recharged and the recovered from the three forest enclosures in which
trapping and dragging procedures were repeated. We turkeys foraged, compared to 184 ticks in the controlrepeated this entire process in all six enclosures one enclosures( paired t-test, t=- 0.23, P= 0.84; Fig. 1). The
week later. To be conservative in our analysis, we did proportion of introduced ticks recovered after 16 hoursnot consider the repetitions of the tick introductions to of access by turkeys was 89. 5% in experimental
be experimental replicates. Instead, we used a paired t- enclosures( with turkeys) and 92.2% in the turkey- freetest with the presence or absence of turkeys as the controls. No ticks were recovered from the turkeys
independent variable, the experimental and control during the 48 hours they were held over waterenclosure within a block comprising the pairs ( N= 3 immediately following the experiments.pairs), and the total number of ticks recovered in the two
introductions as the dependent variable. DISCUSSION
RESULTS Results of Experiment I suggest that wild turkeys
are a poor host for juvenile Ixodes scapularis ticks, andExperiment I- Turkeys as Hosts for Ticks are likely to be parasitized only rarely in nature. In the
Very few larval or nymphal Ixodes scapularis ticks first phase, only four( 3. 0%) of 135 nymphs placed onwere recovered from the turkeys. Of the initial 27 turkeys fed to repletion, and in the second phase, only
TABLE 1. Recovery of juvenile Ixodes scapularis ticks placed on five captive wild turkeysheld individually over collecting pans of water.
Number Number(%) of Number(%) of Total Number
of Ticks Engorged Ticks Unengorged (%) of Ticks
Tick Life Stage Introduced Recovered Ticks Recovered Recovered
Larval 550 0( 0) 7( 1. 3) 7( 1. 3)
Nymphal 30 1 ( 3. 3) 7( 23. 3) 8( 26.7)

December, 1999 Journal of Vector Ecology 185
one( 3. 3%) of 30 nymphs and none of 550 larvae fed to in the same forest enclosures revealed that the turkeysrepletion. The low recovery rates for ticks in either an spent much of their time in the enclosures feeding,engorged or unengorged state strongly suggest that the and that they consumed substantial numbers of treeticks we placed on turkeys were preened off and seeds experimentally introduced into the enclosuresswallowed by the hosts. The ticks had no other route of ( F. Keesing, E. Brownold, and R. Ostfeld, unpub-escape. Casual observations revealed that the turkeys lished data). In both prior and current studies, turkeyscommonly autopreened and allopreened while housed began pecking and scratching at the forest floor withinin the coop; however, observations of the birds while several minutes of being introduced into enclosuresthey were held in individual cages were prevented by ( D. Lewis and R. Ostfeld, pers. obs.). Therefore, theprotective tarps surrounding the cages. Grooming lack of removal of adult ticks was not caused by abehavior is known or suspected to be responsible for lack of foraging effort. We tentatively conclude thatreductions in infestation rates by ixodid ticks on other natural populations of turkeys are unlikely to reducehosts( Ostfeld et al. 1993, Sonenshine 1993). population size of questing adult I. scapularis through
Ixodes scapularis ticks are known to parasitize predation. Because both turkeys and ticks occurdozens of species of vertebrate hosts, including several naturally in aggregations ( Ostfeld et al. 1996a, Lewisspecies of ground- dwelling songbirds ( reviewed by and Ostfeld, pers. obs.), it remains possible that turkeyLane et al. 1991, Fish 1993, Talleklint 1996). However, flocks may encounter and depredate clumps of ticks.we are not aware of any studies reporting I. scapularis Natural enemies of ticks appear to be commonparasitizing M. gallopavo. Davidson and Wentworth ( Carroll 1995, Hu et al. 1993, Samish and Rehacek1992) reviewed studies ofectoparasites on turkeys, and 1999, Zhioua et al. 1995), although their impacts on
reported the occurrence of three species of other ixodid tick populations are poorly understood. Duffy et al.ticks and one species of argasid on wild turkeys in the ( 1992) found that helmeted guineafowl ( Numidasoutheastern United States. The poor performance of meleagris) reduced abundance of adult I. scapularisthese ticks on turkeys, and the likelihood that they were ticks on grass lawns. However, this species of tick isconsumed during autopreening, suggests that continued substantially more abundant in forests than on lawnsrange expansion and population growth ofwild turkeys ( Adler et al. 1992, Maupin et al. 1991, Ostfeld et al.will not increase the number of feeding opportunities for 1995). Although> 40 species of birds are reported toquesting I. scapularis. feed on ticks, evidence suggesting regulation of tick
Captive wild turkeys in our forest enclosures did populations by birds is scarce ( Samish and Rehaceknot reduce abundance of adult I.scapularis, nor did the 1999). Further studies of the regulatory capacity ofticks parasitize the turkeys during their foraging birds and other predators and pathogens on ticks arebouts. Prior studies with the same individual turkeys warranted.
200
150U
r
F-9 100O
N
E 50
z
0
Present Absent
Turkeys
Figure 1. Number (+ 1 SE) of adult Ixodes scapularis ticks recovered using CO2- baited traps and cloth drag-sampling inside 10m by 10m forest enclosures. Three replicates ( experimental enclosures) had turkeyspresent for 16 hours over two days, and three( controls) had no turkeys.

186 Journal of Vector Ecology December, 1999
Acknowledgements wasp parasitoid of Ixodes dammini. J. Med.
Entomol. 30: 277- 280.
We are grateful to Melissa Moyer for help in the Kennamer, J. E., M. Kennemer, and R. Brenneman.
field, and to Dr. Felicia Keesing for insightful comments 1992. History. Pp. 6- 17 in The Wild Turkey:on a draft of the manuscript. Support was provided by Biology and Management ( J. G. Dickson, ed.).the National Science Foundation ( DEB 9615414) and Stackpole Books, Mechanicsburg, PA, 463 pp.the National Institutes of Health( RO1 AI40076). Lane, R. S., J. Piesman, and W. Burgdorfer. 1991. Lyme
borreliosis: relation of its causative agents to its
REFERENCES CITED vectors and hosts in North America and Europe.
Ann. Rev. Entomol. 36: 587- 609.
Adler, G. H., S. R. Telford, III, M. L. Wilson, and A. Maupin, G., D. Fish, J. Zultowsky, E. G. Campos, andSpielman. 1992. Vegetation structure influences J. Piesman. 1991. Landscape ecology of Lymethe burden of immature Ixodes dammini on its main disease in a residential area ofWestchester County,host, Peromyscus leucopus. Parasitol. 105: 105- New York. Am. J. Epidemiol. 133: 1105- 1113.
110. Ostfeld, R. S. 1997. The ecology ofLyme-disease risk.Barbour, A. G. and D. Fish. 1993. The biological and Am. Sci. 85: 338- 346.
social phenomenon of Lyme disease. Science 260: Ostfeld, R. S., O. M. Cepeda, K. R. Hazler, and M. C.
1610- 1616. Miller. 1995. Ecology of Lyme disease: habitatCarroll, J. F. 1995. Laboratory evaluation of predatory associations of ticks( Ixodes scapularis) in a rural
capabilities of a common wolf spider ( Araneae, landscape. Ecol. Applic. 5: 33- 361.
Lycosidae) against two species of ticks ( Acari, Ostfeld, R. S., K. R. Hazler, and O. M. Cepeda. 1996a.
Ixodidae). Proc. Entomol. Soc. Wash. 97: 746- 749. Temporal and spatial dynamics oflxodesscapularis
Davidson, W. R. and E. J. Wentworth, E. J. 1992. Acari: Ixodidae) in a rural landscape. J. Med.
Population influences: diseases and parasites. Pp. Entomol. 33: 90-95.
101- 118 in The Wild Turkey: Biology and Ostfeld, R. S., C. G. Jones, and J. O. Wolff. 1996b. Of
Management( J. G. Dickson, ed.). Stackpole Books, mice and mast: ecological connections in eastern
Mechanicsburg, PA, 463 pp. deciduous forests. BioSci. 46: 323- 330.
Duffy, D. C., R. Downer, and C. Brinkley. 1992. The Ostfeld, R. S., M. C. Miller, and J. Schnurr. 1993. Ear
effectiveness ofhelmeted guineafowl in the control tagging increases tick( Ixodes dammini) infestationof the deer tick, the vector of Lyme disease. Wilson rates of white- footed mice( Peromyscus leucopus).
Bull. 104: 342- 345. J. Mamm. 74: 651- 655.
Falco, R. C. and D. Fish. 1992. A comparison of Piesman, J. and J. S. Gray. 1994. Lyme disease/ Lymemethods for sampling the deer tick, Ixodes dammini, borreliosis. Pp. 327- 350 in Ecological Dynamics ofin a Lyme disease endemic area. Exp. Appl. Acarol. Tick-Borne Zoonoses.( D. E. Sonenshine and T. N.
14: 165- 173. Mather, eds.). Oxford University Press, NY, 447Fish, D. 1993. Population ecology of Ixodes dammini. pp.
Pp. 25- 42 in Ecology and Management of Lyme Samish, M. and J. Rehacek. 1999. Pathogens and
Disease ( H. Ginsberg, ed.). Rutgers Univ. Press, predators of ticks and their potential in biological
New Brunswick, NJ, 224 pp. control. Ann. Rev. Entomol. 44: 159- 182.
Hazler, K. R. and R. S. Ostfeld. 1995. Larval density Sonenshine, D. E. 1993. Biology of ticks. Volume 2.and feeding success of Ixodes scapularis on two Oxford Univ. Press, NY, 465 pp.species of Peromyscus. J. Parasitol. 81: 870- 875. Talleklint, T. 1996. Transmission of Lyme borreliosis
Healy, W.M. 1992. Population influences: environment. spirochetes at the tick vector- mammal reservoir
Pp. 129- 143 in The Wild Turkey: Biology and interface. Ph.D. thesis, Uppsala University.Management( J. G. Dickson, ed.). Stackpole Books, Zhioua, E., R. A. Lebrun, H. S. Ginsberg, and A.Mechanicsburg, PA, 463 pp. Aeschlimann. 1995. Pathogenicity ofSteinernema
Hu, R. J., K. E. Hyland, and T. N. Mather. 1993. carpocapsae and S. glaseri ( Nematoda: Steiner-
Occurrence and distribution in Rhode Island of nematidae) to Ixodes scapularis( Acari, Ixodidae).
Hunterellus hookeri( Hymenoptera: Encyrtidae), a J. Med. Entomol. 32: 900-905.

Journal of Vector Ecology 24( 2): 187- 190
Susceptibility of the Malaria Vector Anopheles culicifacies (Diptera:
Culicidae) to DDT, Dieldrin, Malathion, and Lambda-Cyhalothrin
R. S. Sharma
National Anti Malaria Programme
22-Shamnath Marg, Delhi 110054, INDIA
Received 18 February 1999; Accepted 10 September 1999
ABSTRACT: The susceptibility of the malaria vector Anopheles culicifacies to DDT, dieldrin, malathion,and lambda-cyhalothrin was determined in Karnal, Yamunanagar, and Ambala districts of Haryana State,
India. The vector population showed a high degree of resistance to DDT and dieldrin. The test mortalityto DDT and dieldrin ranged from 25% to 28% and 18% to 20%, respectively, in Nadasahib of Ambaladistricts. The mortality of An. culicifacies to malathion ranged between 65% and 68%. All the tests withlambda-cyhalothrin resulted in 100% mortality of An. culicifacies. DDT and dieldrin resistance did notconfer cross- resistance to lambda-cyhalothrin in An. culicifacies.
Keyword Index: DDT, Malathion, Anopheles culicifacies.
INTRODUCTION widespread in Gujarat and Maharastra ( Vittal andDeshpande 1983, Das et al. 1986).
Insecticides continue to be the mainstay of vector- Synthetic pyrethroids have been found to be effectiveborne disease control programs in India. The extensive for the control of mosquitoes ( Chester et al. 1992).and intensive use of residual insecticides reduced the During the 1996 malaria epidemic in the Gurgaonvector population, and malaria control was achieved to district of Haryana State, HCH and malathion werea great extent in India by the mid-sixties. Afterwards, replaced by synthetic pyrethroids, which controlled An.however, malaria incidence started rising to a peak level culicifacies( Sharma et al. 1997). The response of multi-of 6. 4 million by 1976. Vector resistance to DDT, BHC, resistant An. culicifacies to lambda-cyhalothrin is notand malathion was considered to be one of the known in Haryana State. The present field study on thecontributory factors for the resurgence of malaria in susceptibility of An. culicifacies populations using theIndia. The steady development of resistance to an diagnostic dosage of 4% DDT, 0.4% dieldrin, 0. 1%
increasing number of insecticides by an increasing lambda-cyhalothrin, and 5% malathion was undertaken
number of vector species over wider geographical areas in different districts of Haryana State.has impeded malaria control programs in India.Anopheles culicifacies was highly susceptible to MATERIALS AND METHODSmalathion in Haryana State ( Sharma 1990, Sharmaet.al. 1996), where malathion was introduced in 1981 The study was carried out in Ambala, Karnal, andfor regular indoor residual spraying for DDT and BHC Yamunanagar districts ofHaryana State. The diagnosticresistant populations ofAn. culicifacies. Over the years, concentration for the detection ofresistance in Anophelesthe An. culicifacies populations have reacted differently culicifacies was 5% malathion. The susceptibility testsunder different ecological and environmental conditions were carried out according to the WHO standard testranging from the development of resistant populations ( WHO 1975), in which the mosquitoes were exposed toto the selection of refractory behavior, affecting malaria impregnated papers with an oil solution containing thetransmission to varying degrees( Bang 1985). In India, diagnostic concentration of the test insecticide. Thethe first report ofmalathionresistancecamefromGujarat impregnated papers were supplied by WHO. Thestate( Rajagopal 1977). The development of resistance exposure period was one hour in all the tests. Fully fedin An.culicifacies to malathion was found to be An. culicifacies collected from the field were used for
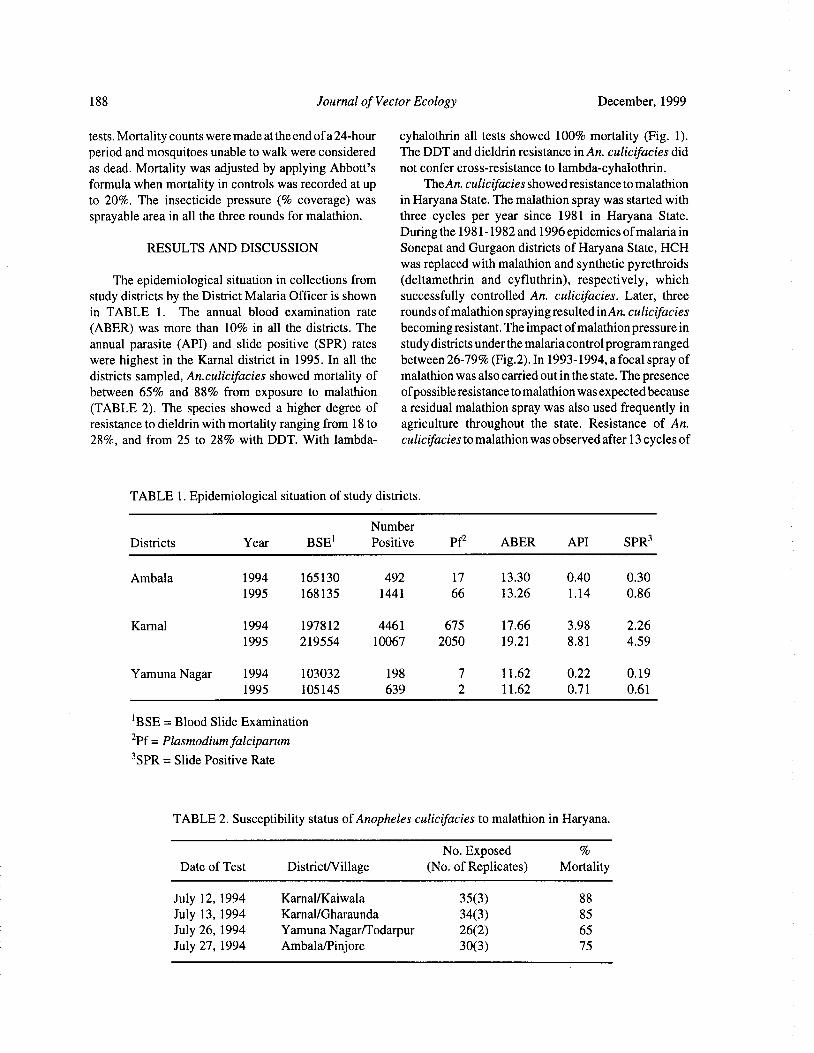
188 Journal of Vector Ecology December, 1999
tests. Mortality counts were made at the end ofa 24-hour cyhalothrin all tests showed 100% mortality ( Fig. 1).period and mosquitoes unable to walk were considered The DDT and dieldrin resistance in An. culicifacies did
as dead. Mortality was adjusted by applying Abbott' s not confer cross- resistance to lambda-cyhalothrin.
formula when mortality in controls was recorded at up TheAn. culicifacies showed resistance to malathion
to 20%. The insecticide pressure (% coverage) was in Haryana State. The malathion spray was started withsprayable area in all the three rounds for malathion. three cycles per year since 1981 in Haryana State.
During the 1981- 1982 and 1996 epidemics ofmalaria inRESULTS AND DISCUSSION Sonepat and Gurgaon districts of Haryana State, HCH
was replaced with malathion and synthetic pyrethroids
The epidemiological situation in collections from ( deltamethrin and cyfluthrin), respectively, which
study districts by the District Malaria Officer is shown successfully controlled An. culicifacies. Later, threein TABLE 1. The annual blood examination rate rounds ofmalathion spraying resulted in An. culicifaciesABER) was more than 10% in all the districts. The becoming resistant. The impact ofmalathion pressure in
annual parasite ( API) and slide positive ( SPR) rates study districts under the malaria control program rangedwere highest in the Karnal district in 1995. In all the between 26-79%( Fig.2). In 1993- 1994, a focal spray ofdistricts sampled, An.culicifacies showed mortality of malathion was also carried out in the state. The presence
between 65% and 88% from exposure to malathion ofpossible resistance to malathion was expected because
TABLE 2). The species showed a higher degree of a residual malathion spray was also used frequently inresistance to dieldrin with mortality ranging from 18 to agriculture throughout the state. Resistance of An.
28%, and from 25 to 28% with DDT. With lambda- culicifacies to malathion was observed after 13 cycles of
TABLE 1. Epidemiological situation of study districts.
Number
Districts Year BSE' Positive Pf2 ABER API SPR3
Ambala 1994 165130 492 17 13. 30 0.40 0. 30
1995 168135 1441 66 13. 26 1. 14 0. 86
Karnal 1994 197812 4461 675 17. 66 3. 98 2. 26
1995 219554 10067 2050 19. 21 8. 81 4. 59
Yamuna Nagar 1994 103032 198 7 11. 62 0.22 0. 19
1995 105145 639 2 11. 62 0.71 0.61
BSE= Blood Slide Examination
2Pf= Plasmodium falciparum
3SPR= Slide Positive Rate
TABLE 2. Susceptibility status of Anopheles culicifacies to malathion in Haryana.
No. Exposed
Date of Test District/Village No. of Replicates) Mortality
July 12, 1994 Karnal/ Kaiwala 35( 3) 88
July 13, 1994 Karnal/ Gharaunda 34( 3) 85
July 26, 1994 Yamuna Nagar/Todarpur 26( 2) 65
July 27, 1994 Ambala/Pinjore 30( 3) 75
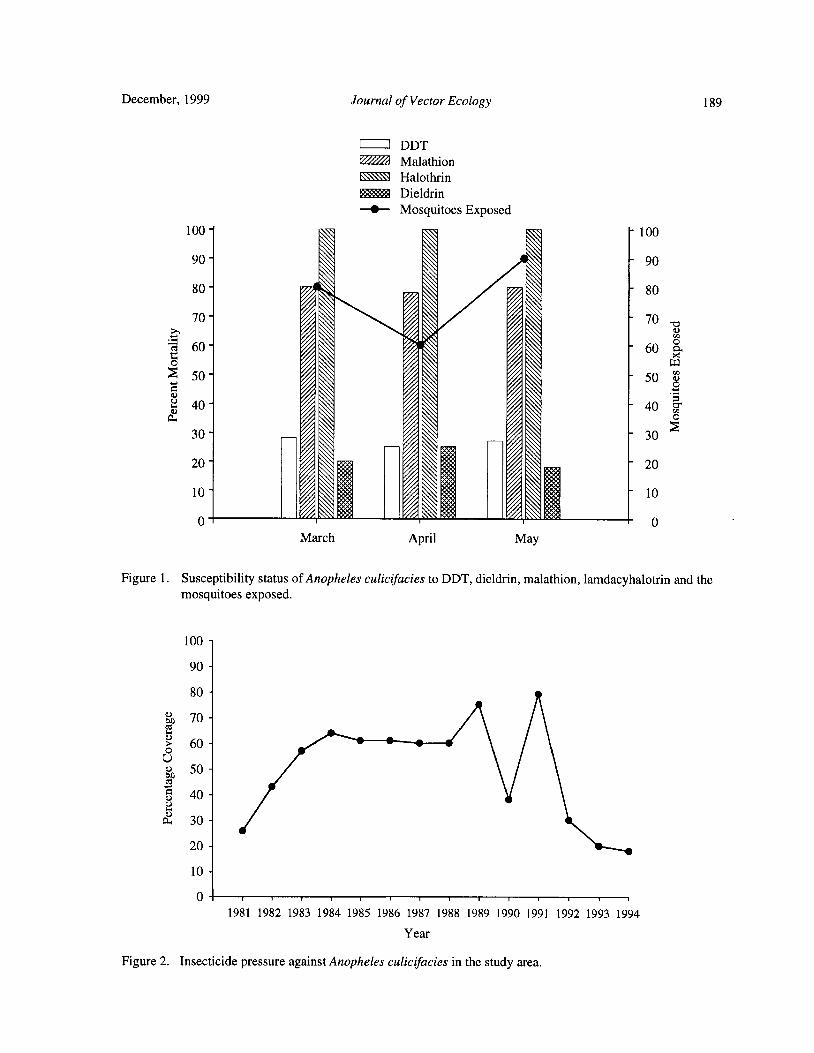
December, 1999 Journal of Vector Ecology 189
1 1 DDT
Malathion
Halothrin
Dieldrin
S Mosquitoes Exposed
100= 190
80 V V 80
50' 50
a40 40 ,
30' 30
20' 20
10' 10
March April May
Figure 1. Susceptibility status of Anopheles culicifacies to DDT, dieldrin, malathion, lamdacyhalotrin and themosquitoes exposed.
1 00 -
1 9 8 1 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994
Year
Figure 2. Insecticide pressure against Anopheles culicifacies in the study area.

190 Journal of Vector Ecology December, 1999
spray in the Thane district of Maharashtra ( Vittal and Deshpande. 1986. Susceptibility of DDT, dieldrinDespande 1983). The development of resistance in An. and malathion resistant to Anopheles culcifacies
culicifacies to malathion in Haryana State is due to populations to deltamethrin. J. Am. Mosq. Cont.prolonged selection pressure maintained by the Control Assoc. 2: 553- 555.
Program and the predominant indoor resting and biting Rajagopal, R. 1977. Malathion resistance in Anopheles
habits of the vector species. culicifacies in Gujarat Ind. J. Med. Res. 66: 27- 28.
Sharma, R. S. 1990. Susceptibility status of malariaAcknowledgements vectors Anopheles culicifacies and Anopheles
stephensi ( Diptera: Culicidae) to malathion.
The authors are thankful to Shri P. K. Jena and Shri Entomol. 15: 253- 256.
Y. S. Parihar for their dedicated assistance in the field. Sharma R. S, S. N. Sharma, and R. S. Sharma. 1996.
The authors are also thankful to Shri Jarnail Singh and Susceptibility of DDT, Dieldrin and the
Mr. S. K. Katyal for secretarial assistance. development ofmalathion resistance in Anopheles
culicifacies ( Diptera: Culicidae) in Ambala
REFERENCES CITED District of Haryana. J. Vect. Ecol. 21: 14- 16.
Sharma R. S., L. Shiv, S. N. Sharma, R. D. Joshi, and G.
Bang, Y. H. 1985. Implementations in the control of P. S. Dhillon. 1997. Malaria outbreak in Mewat
malaria vectors with insecticides in tropical region of Gurgaon district of Haryana State. J.
countries of South East Asia. Part- 1. Insecticide Commun. Dis. 29: 307- 308.
Resistance. J. Commun. Dis. 17: 199- 218. Vittal, M. and L. B. Deshpande. 1983. Development of
Chester, G., N. N. Satapathy, and B. H. Woolpen 1992. malathion resistance in DDT, HCH resistance to
Exposure and health assessment during application Anopheles culicifacies population in Thane district
of lambda-cyhalothrin for malaria vector control in Maharashtra) J. Commun. Dis. 15: 144- 145.
Pakistan. Bull. World Hlth. Org. 70: 615- 629. World Health Organization 1975. Manual on Practical
Das, M., S. P. Srinivasan, J. S. Khamre, and L. H. Entomology- Part- II, 190 pp.

Journal of Vector Ecology 24( 2): 191- 201
Bacteria and Mosquito Abundance in MicrocosmsEnriched with Organic Matter and Treated with a
Bacillus thuringiensis subsp. israelensis Formulation
Tram T. H. Nguyen, Tianyun Su, and Mir S. Mulla
Department ofEntomology, University of California,Riverside, CA 92521- 0314, USA
Submitted 30 April 1999; Accepted 31 August 1999
ABSTRACT: Bacteria and mosquito abundance were studied in outdoor tubs unenriched and enriched with0.04% rabbit pellets during the winter, 1999. The irrigation water used to fill the tubs contained a totalbacterial count of 1. 15- 1. 35 x 10' cells/ml. Adding rabbit pellets for enrichment yielded a total bacterialcount of 5. 50- 7. 63 x 105 cells/ g. Bacterial densities in unenriched water were significantly lower than inenriched tubs on every sampling day. When bacterial densities in both enriched and unenriched regimensreached peak populations on day 3 post- flooding, their numbers in enriched water were 25- fold higher thanthe unenriched water. Under cool weather conditions, mosquito oviposition activity was low and larvaldevelopment was very slow. Egg raft counts and larval densities in enriched water were nevertheless higherthan those in unenriched water. After reaching peak populations on day 3 post- flooding, the natural declinein bacterial densities in the top portion of enriched water without mosquito larvae was lower compared withthat in water with larvae. In water with larval present, the decline of bacterial levels in top water was greaterthan in bottom water on day 7 post- flooding. VectoBac G, a granular formulation ofBacillus thuringiensissubsp. israelensis caused a reduction in larval numbers of 80, 93, 73% at the rate of 5. 5 lb/ac and a reductionof 94, 93, 86% at the rate of 10. 6 lb/ ac on days 1, 3, 7 posttreatment, respectively. After treatment, thereductions of bacterial densities in untreated tubs were greater than treated tubs. These results indicate that
mosquito larvae play an important role in the decline of bacterial populations by their feeding activity.
Keyword Index: Organic matter, enriched water, oviposition, Bacillus thuringiensis subsp. israelensis,mosquitoes, bacteria.
INTRODUCTION aeruginosa from fatty acid substrates, which producesovipositional attractants for both Cx. pipiens molestus
Organic matter has been used in field experiments Forskal andAe. aegypti. In addition, bacteria also servedas ovipositional attractants and stimulants to obtain as an important component of food sources ofmosquitosustained production of mosquito larvae ( Beehler and larvae( Nilsson 1987, Walker et al. 1988, Merrit et al.Mulla 1995, Rodcharoen et al. 1997). Supplemental 1990, 1996).
enrichment of mosquito breeding sources with chicken We recently studied the bacterial flora and thelay mash, alfalfa pellets, and rabbit pellets produces contribution of the microbialmosquitocidalformulations
large numbers of Culex mosquitoes ( Culex quinque- of Bacillus thuringiensis subsp. israelensis de Barjacfasciatus Say, Culex tarsalis Coquillette, Culex ( B. t. i.) and Bacillus sphericus Neide strain 2362 tostigmatosoma Dyar) for field evaluation of microbial bacterial abundance in microcosms sustaining heavyagents and mosquitofish as larval predators( Walton and natural populations of mosquito larvae in the summerMulla 1991, Su and Mulla 1999, Nguyen et al. 1999). and fall seasons ( Nguyen et al. 1999). In the present
Bacterial activity can modify the ovipositional study, we investigated the bacterial abundance in waterbehavior of gravid mosquitoes. The bacterium in microcosms unenriched and enriched with organicAerobacter aerogenes isolated from hay infusion matter to address the relationship between organicproduced chemicals attractive to ovipositing Cx. quinque- pollution and bacterial abundance. To determine thefasciatus and Aedes aegypti L. ( Hazard et al. 1967). effect ofmosquito activity on bacterial densities, bacterialIkeshoji et al.( 1975) isolated the bacterium Pseudomonas abundance was assessed in enriched water in the presence

192 Journal of Vector Ecology December, 1999
or absence of mosquito larvae, and tubs treated with the parameters were measured forall tubs on every samplingmicrobial agent B. t. i, where mosquito larval densities day. The mean values for dissolved oxygen ( DO) andwere reduced, were also examined. The experiments corrected electrical conductivity at 25°C ( CEC) werewere carried out in the winter season of 1999, when determined with a YSI Model 85 Handheld Water
water temperatures and mosquito larval breeding were Quality Meter. Water temperature was monitored withlower than in the summer and fall seasons. a minimum-maximum thermometer submerged in a tub
located in the center of the tub arrangement, and
MATERIALS AND METHODS temperatures were recorded each time the tubs were
sampled for mosquito larvae and/ or bacterial density.Test Facilities and Material
Two tests were carried out in the winter season of Bacterial Assessment
1999 in outdoor fiber-glass tubs measuring 1. 0 x 1. 0 x In order to assess the background bacterial
0.4 m deep at the Aquatic and Vector Control Research populations in the water used for tub filling, three waterFacility at University of California, Riverside. In test 1, samples, each of which consisted of 120- 150 ml, were
because of lack of adequate mosquito production due to directly collected from the float valves of threecool weather, only bacterial abundance in water randomized tubs during the period of filling the tubs inunenriched and enriched with organic matter was both test 1 and 2.
assessed. In total, 24 tubs were used in this experiment. To determine the background bacterial density inTwelve of them were selected at random to be enriched the dry rabbit pellets used for tub enrichment in test 1with rabbit pellets ( Brookhurst®, Brookhurst Mill, and 2, about 2 kg of pellets were placed in a plasticRiverside, CA, crude protein 17%) at the rate of 0.04%, bucket of 10 quarts capacity before enriching the tubs.equaling 100 grams per tub. The other 12 tubs were left Three 10 g samples were collected from the peripheryunenriched. The tubs were filled with water from an and center of the top layer of the pellets in plastic bucketirrigation reservoir and water depth was kept constant at and put in separate sterile plastic Petri dishes. Rabbit
30 cm( 236 liters) using float valves. In test 2, 16 tubs pellets were then ground to powder in the laboratorywere enriched with rabbit pellets at the same rate as in with a porcelain mortar and pestle and processed for
test 1. The purpose of this experiment was to determine isolation and determination ofbacterial numbers present
bacterial abundance in enriched water with or without in the pellets.
mosquito larvae and in microbial treatments using B. t.i. To determine bacterial abundance in the tubs after
granular formulation( VectoBac G). Eight randomized filling in test 1, water from both the top and bottom oftubs were treated with two rates( four tubs for each rate) enriched and unenriched tubs was collected at three
of VectoBac G, while the other eight tubs were left hours after filling the tubs and on days 3, 8, 15, and 22untreated. Among the untreated tubs, four tubs were post- flooding. No treatments were made with B.t.i. inassigned as check 1 with mosquito larvae present, while this test. In test 2, water samples were collected at threethe other four tubs were used as check 2 where mosquito hours after enriching and filling the tubs and on days 3,egg rafts were removed every other day during the test 7, and 13 post- flooding. After treatment with VectoBacperiod to prevent larval breeding. Wild populations of G, water samples were collected at three hours and on
Culex stigmatosoma, Cx. quinquefasciatus, and Cx. days 1, 3, and 7 posttreatment. Sterile pipetts ( 5m1)
tarsalis oviposited nightly in these filled tubs. fitted with a pro-pipette were used to collect five samples,
The B. t.i. granular formulation( VectoB ac G, corn one at each corner and one at the middle ofthe undisturbed
cob granules, Lot# 25- 801- N8, 200 ITU/mg, Abbott water at 0.3- 0. 5 cm below the surface and about 0.5- 1. 0Laboratories, North Chicago, IL, received 7/ 10/ 1997) cm above the bottom of each tub. The top and bottomwas applied to the eight tubs in test 2, 13 days after water samples of all replicates were composited
enriching and filling the tubs, when 3rd-4th- instar separately in 250 ml sterile flasks, each compositemosquito larvae were present in large numbers. The sample amounting to 100 ml of water from the fourformulation was applied at two rates of 5. 3 and 10.6 lb/ replicates.
ac, equaling 0.5 and 1. 0 g per tub, respectively. The To estimate the bacterial densities in water samplesrequired amounts of VectoBac G for each rate were and in rabbit pellets, the surface spread plated method
gently broadcast by hand on the water surface at the on nutrient agar was used( Jones 1970). For quantitative
corners and middle of each tub. determination of bacterial cells, 1 ml from each water
sample was serially diluted in 9 ml of autoclaved waterWater Quality Determination and dilutions ranged from 10° to 10' times to facilitate
During the course of the tests, water quality colony counting. One gram of powdered rabbit pellets

December, 1999 Journal of Vector Ecology 193
was weighed and suspended in 99 ml sterile water. After Statistical Analyses
shaking for 5 min, 1 ml of this aliquot was serially Water quality parameters( DO and CEC), averagediluted in test tubes containing 9 ml sterile water. Three bacterial densities( CFU/ml), and average egg raft countsPetri dishes containing 20 ml NA medium( 2. 3% Difco ( egg rafts/tub) as well as average larval densities( larvae/nutrient agar) were inoculated with 0. 1 ml aliquot of dip) in the check and treated tubs were analyzed foreach serial dilution, and incubated at 30°C for 48 hours. significance by one factor ANOVA( Scheffe F test) forAutoclaved water for dilution was also plated on NA as repeated measurements.
a control. After an incubation period of 48 hours, the
colony- forming units ( CFUs) were counted visually RESULTS
and adjusted for dilutions and inoculum volume, and
were expressed as mean number of CFUs/ ml of water TEST 1
sample or as mean number of CFUs/ gram of rabbit Water Qualitypellets. Test 1 was carried out from January 11 to February
To determine densities of spore- forming bacteria 2, 1999. The water temperatures during the test periodincluding B. t.i., water samples were heated to 65° C for were low, minimum, and maximum ranged 6. 8- 10.2° C
30 minutes to kill vegetative and non- spore- forming and 15. 7- 20.2° C, respectively ( Fig. 1A). Marked
bacteria before diluting and plating. These water samples differences in dissolved oxygen( DO) values were noted
were diluted from 10° to102
times and spread on NA between unenriched and enriched water. In unenriched
medium. After incubation at 30° C for 48 hours, colonies water, the DO was 6.7 ppm at three hours after filling thewere counted visually and expressed as CFUs/ml. tubs and progressively increased to 14. 1- 14. 2 ppm on
days 15- 22 post-flooding( Fig. 1B). The DO in enrichedMosquito Production and Control water, however, was similar to that in the unenriched
Mosquito oviposition was assessed by counting the water at three hours after enriching and filling the tubstotal number of egg rafts( presented as mean number of ( 6. 5 ppm), but the readings sharply decreased to 0.04,egg raft per tub) on every sampling day in test 1 and 2 0. 11. and 0.49 ppm DO on days 3, 8, and 15 post-
except check 2) in each tub and placing them in plastic flooding, respectively, then increased to 9. 8 ppm on daysentinel cups, which were partially submerged in the 22 post- flooding. On days 3, 8, 15, and 22 post- flooding,water of the tubs using floatation collars. The bottom of DO readings in enriched water were significantly lowerthese cups was removed previously to allow the hatched than in unenriched water ( Fig. 1B). The CEC in
larvae to swim freely out into the water. This technique unenriched water increased from 528 mS at three hours
allowed us to count the newly laid egg rafts at each after start of filling the tubs to 545. 7 mS on day 8 post-sampling interval. In test 2, in order to investigate the flooding, then rapidly decreased to 420.6 mS on day 22influence of mosquito larvae on microbial abundance, post- flooding. In enriched water, the CEC increasedthe egg rafts were removed and discarded every other from 557. 8 mS at three hours after enriching and fillingday from four enriched tubs, which were assigned as the tubs to 590 mS on day 8 post- flooding, then declinedcheck 2 to determine bacterial abundance without to 552.6 mS on day 22 post- flooding. Significantmosquito larvae. These tubs were devoid of mosquito difference in CEC was indicated between unenriched
larvae throughout the experiment. and enriched water on days 15 and 22 post-floodingMosquito larvae were assessed by taking five-dip ( Fig. 1C).
samples per tub, one dip from each corner and one fromwhere larvae were noted in large numbers on every Bacterial Abundance
sampling day. In order to minimize the dipping When the background bacterial population in
disturbance to the water surface, during a dipping cycle reservoir water used to fill the tubs was monitored, the
for a treatment assignment, one dip was taken from each bacterial densities in water flowing from the float valvestub, so there was a 3- 5 minute period between each averaged 1. 15 x 103 total bacterial cells/ml. Rabbit
dipping in a given tub. Larval populations were pellets used for enrichment contained a large number of
categorized into early ( 1st and 2nd) and late ( 3rd and bacteria, averaging 5. 50 x 105 total bacterial cells/ g.4th) instars, then counted and recorded as mean number After filling the tubs, the bacterial density in water of theof larvae per dip. The pupae, ifany, were excluded from unenriched regimen was significantly lower than that incounting because no feeding activity was occurring at the enriched regimen on every sampling day during thepupal stage. The percentage reduction(% R) ofmosquito test period( Fig. 2A). In top water of unenriched tubs atlarvae due to B.t.i. treatments was calculated using the three hours after filling the tubs, total bacterial numbersformula given by Mulla et al. ( 1971). were 1. 80 x
103cells/ ml( equal to the incoming water),
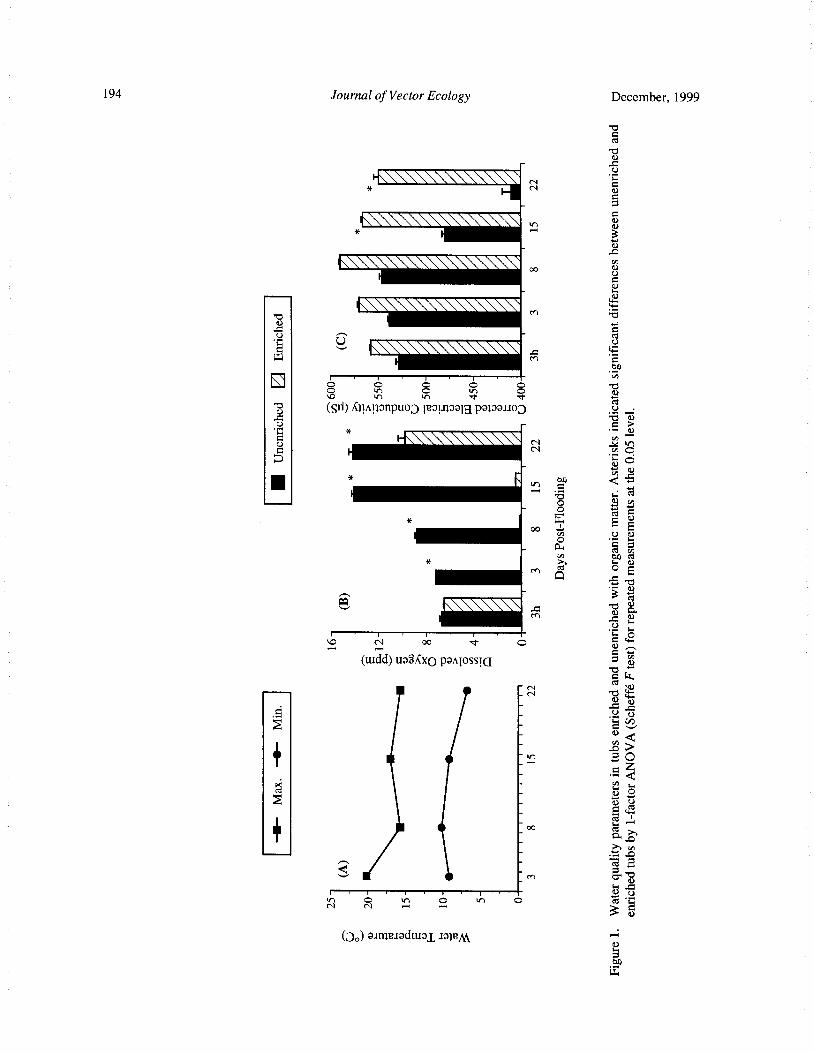
194 Journal of Vector Ecology December, 1999
bacbN
U
N9E N
y0p0
kn
ykn
3n
11= ii6iiii oo 0cnVQ
w
CEi66iiiw
bc+ b
U 0
W m 0on
A
pa)tn
p o 8V
sri), Clini; anpuo3 lRai.1aalg papauo3 oU
IC NO
t.•
oo II* oo
E a—. a: cd vl
Q oM
b
3 :b a)
iiii61 M
U F.
N o0 O w
urdd) uaRXxo panlossLQ ovim'
CI
N 0
0 Qa
Y
C
t"2 i
0_ N0 w
yIII
Do) arn;eaadural, J IuMa>
aw"

December, 1999 Journal of Vector Ecology 195
which peaked at 6. 22 x104
cells/ml on day 3 post- Mosquito Productionflooding, then progressively declined to 1. 10 x 104cells/ Because ofcool temperatures during test 1, mosquitoml at the end of the test on day 22 post- flooding. oviposition levels and larval numbers were low, theseHowever, in top water of tubs enriched with rabbit data are not presented in figure form. Despite this, onpellets, the total bacterial numbers were 2.20 x
104cells/ day 8 post- flooding, the mean number of egg rafts in
ml at three hours after enriching and filling the tubs, unenriched water ( 4. 9 ± 1. 4 egg rafts/ tub) waswhich sharply increased and peaked at 1. 53 x
106cells/ significantly lower than in the enriched( 17. 8± 2. 6 egg
ml on days 3 and 8 post- flooding, maintaining a high rafts/ tub). Similarly, the number of larvae in unenrichedcount of 1. 10 x
106cells/ml on day 15 and finally water was significantly lower than in the enriched tubs
declining to 1. 45 x105
cells/ml on day 22 post- flooding amounting to 6. 1± 2.7 and 19. 6± 5. 8 larvae/dip on dayFig. 2A).
22 post- flooding, respectively. No species identificationTotal bacterial densities in the bottom water of of the mosquito larvae was conducted because of low
unenriched as well as enriched tubs were slightly higher larval densities in this test.than those in top water on every sampling day, howeverthese data were not included in figures. In bottom water TEST 2of unenriched tubs, bacterial numbers increased from Water Quality2. 30 x 10' cells/ml at three hours after filling the tubs to Test 2 was conducted from February 23 to Marchthe peak population of 6.55 x
104cells/ml on day 3 post- 15, 1999. Water temperatures fluctuated from aminimum
flooding, then gradually declined to 1. 10 x104
cell s/ ml of 8. 5- 10.8° C to amaximum of 16. 8- 25° C. No differenceat the end of the test. Bacterial numbers in bottom water in water quality among the various treatments wasofenriched tubs sharply increased from 2. 40 x
104cells/ detected. DO readings were 3. 6 ppm at three hours after
ml at three hours after enriching and filling the tubs to enriching and filling the tubs, decreasing to almostthe peak population of 1. 66 x
106cells/ ml on day 3 post- anoxic conditions( 0.04 and 0.06 ppm) on days 3 and 7
flooding. High numbers continued to prevail with 1. 64 post- flooding, then gradually increasing to 15. 9 ppm atx
106cells/ ml on day 8 post- flooding, then declined to the end of the test on day 7 posttreatment( day 20 post-
1. 21 x106
cells/ml on day 15 and 1. 48 x105
cells/ ml on flooding). CEC values increased progressively fromday 22 post- flooding. The growth and decline trends of 549. 1 mS at three hours after enriching and filling thetotal bacterial abundance over time were essentially the tubs to 614. 8 mS on day 1 posttreatment( day 14 post-same in the top and bottom waters. flooding), then decreased slightly to 564.9 mS on day 7
As with the total bacteria, densities ofspore- formers posttreatment. These data are not presented in figurein unenriched water samples were significantly lower form.
than those in enriched water on every sampling day. Inunenriched water, spore- formers densities increased Bacterial Abundanceslightly from 0.76 x
102cells/ml at three hours after In test 2, irrigation water contained an average of
filling the tubs to 0. 80 x102
cells/ ml on day 3 post- 1. 35 x 10' total bacterial cells/ml and rabbit pelletsflooding, then progressively declined to 0. 30 x
102cells/ contained 7. 63 x 105 total bacterial cells/ g. After filling
ml on day 22 post- flooding ( Fig. 2B). In top water of the tubs, a marked decline in total bacterial populationsenriched tubs, the average count of spore- formers was were noted in all treatment regimens ( tubs without1. 20 x
102cells/ml at three hours after enriching and mosquito larvae, tubs with larvae and tubs treated with
filling the tubs, increasing to 1. 75 x102
cells/ml on day B.t. i. formulation). In top water, the bacterial densities3 post- flooding, then gradually declining to 0.75 x
102
increased from 1. 5- 2. 30 x 104 cells/ml at three hourscells/ml on day 22 post- flooding( Fig. 2B). after enriching and filling the tubs to the peak populations
Spore- forming bacterial densities in bottom water of 1. 94- 2.26 x 106 cells/ ml on day 3 post-flooding( Fig.of unenriched and enriched tubs were essentially the 3), then gradually declining to 1. 60 x 104- 1. 11 x 105same as or slightly higher than in densities in top water, cells/ml on day 7 posttreatment( day 20 post- flooding),depending on sampling days. The spore- formers when the test was terminated. Bacterial densities in thereached peak populations on day 3 post- flooding( 0.80 bottom water were higher than those in the top water onx
102and 2.05 x
102cells/ml in unenriched and enriched every sampling day. Their numbers increased from
water, respectively), gradually declining to the low 1. 90-4. 20 x 104 cells/ ml at three hours after enrichinglevels at the end of the test on day 22 post- flooding. The and filling the tubs to peak populations of 1. 99- 2.32 xoverall growth and decline patterns of spore- forming 106 cells/ml on day 3 post- flooding, then decliningbacteria over time were similar in top and bottom progressively to 2. 50 x 104- 1. 27 x 105 cells/ml on day 7waters.
posttreatment.
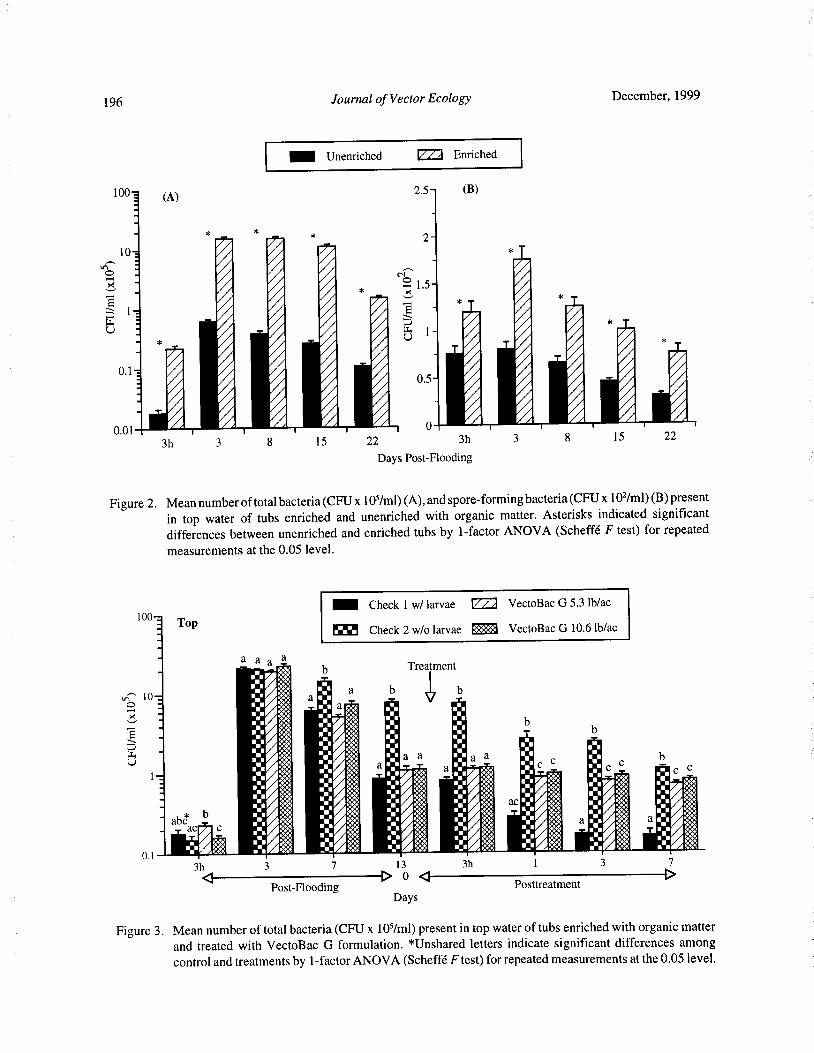
196 Journal of Vector Ecology December, 1999
Unenriched ® Enriched
100! ( A) 2.5— ( B)
2-
10—
O _ N
i l
1. 5'
u1 _ E 1_
mUi*oorr 010. 1
0.5-
1 i0.01 I 1 1 1
0 1 I 1
3h 3 8 15 22 3h 3 8 15 22
Days Post- Flooding
Figure 2. Mean number of total bacteria( CFU x 105/ ml)( A), and spore- forming bacteria( CFU x 102/ ml)( B) presentin top water of tubs enriched and unenriched with organic matter. Asterisks indicated significantdifferences between unenriched and enriched tubs by 1- factor ANOVA ( Scheffe F test) for repeatedmeasurements at the 0.05 level.
INE Check 1 w/ larvae ® VectoBac G 5. 3 lb/ ac
100-7Top
Check 2 w/o larvae .:::: VectoBac G 10.6 lb/ ac
a a a ab Treatment
Fes::
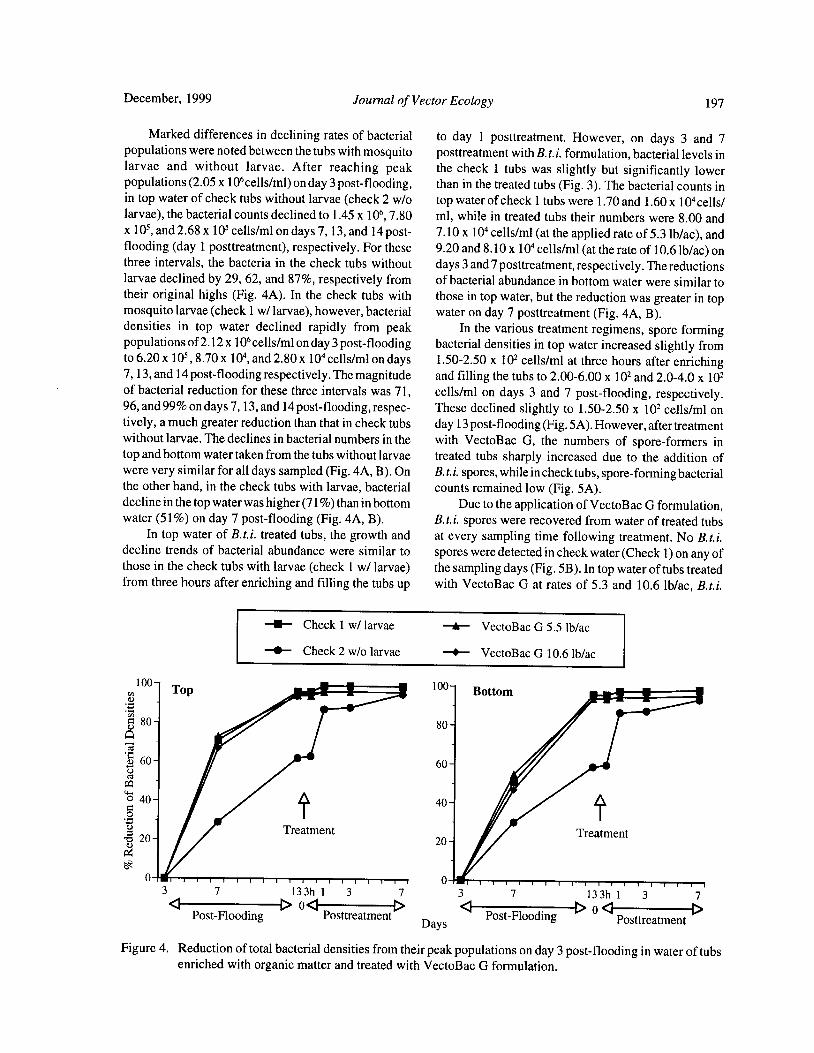
December, 1999 Journal of Vector Ecology 197
Marked differences in declining rates of bacterial to day 1 posttreatment. However, on days 3 and 7populations were noted between the tubs with mosquito posttreatment with B.t.i. formulation, bacterial levels inlarvae and without larvae. After reaching peak the check 1 tubs was slightly but significantly lowerpopulations( 2. 05 x
106cells/ml) on day 3 post- flooding, than in the treated tubs( Fig. 3). The bacterial counts in
in top water of check tubs without larvae( check 2 w/o top water of check 1 tubs were 1. 70 and 1. 60 x 104cells/larvae), the bacterial counts declined to 1. 45 x
106, 7. 80 ml, while in treated tubs their numbers were 8. 00 andx
105,and 2. 68 x
105cells/ml on days 7, 13, and 14 post- 7. 10 x 104 cells/ ml( at the applied rate of 5. 3 lb/ ac), and
flooding( day 1 posttreatment), respectively. For these 9.20 and 8. 10 x 104 cells/ ml( at the rate of 10.6 lb/ ac) onthree intervals, the bacteria in the check tubs without days 3 and 7 posttreatment, respectively. The reductionslarvae declined by 29, 62, and 87%, respectively from of bacterial abundance in bottom water were similar totheir original highs ( Fig. 4A). In the check tubs with those in top water, but the reduction was greater in topmosquito larvae( check 1 w/ larvae), however, bacterial water on day 7 posttreatment( Fig. 4A, B).densities in top water declined rapidly from peak In the various treatment regimens, spore formingpopulations of2. 12 x
106cells/ml on day 3 post- flooding bacterial densities in top water increased slightly from
to 6.20 x105, 8. 70 x
104,and 2. 80 x
104cells/ml on days 1. 50- 2. 50 x 102 cells/ ml at three hours after enriching
7, 13, and 14 post- flooding respectively. The magnitude and filling the tubs to 2. 00- 6.00 x 102 and 2.0- 4.0 x 102of bacterial reduction for these three intervals was 71, cells/ ml on days 3 and 7 post- flooding, respectively.96, and 99% on days 7, 13, and 14 post- flooding, respec- These declined slightly to 1. 50- 2.50 x 102 cells/ml ontively, a much greater reduction than that in check tubs day 13 post- flooding( Fig. 5A). However, after treatmentwithout larvae. The declines in bacterial numbers in the with VectoBac G, the numbers of spore- formers intop and bottom water taken from the tubs without larvae treated tubs sharply increased due to the addition ofwere very similar for all days sampled( Fig. 4A, B). On B.t. i. spores, while in check tubs, spore- forming bacterialthe other hand, in the check tubs with larvae, bacterial counts remained low( Fig. 5A).decline in the top water was higher( 71%) than in bottom Due to the application ofVectoBac G formulation,water( 51%) on day 7 post- flooding( Fig. 4A, B). B.t.i. spores were recovered from water of treated tubs
In top water of B. t.i. treated tubs, the growth and at every sampling time following treatment. No B.t.i.decline trends of bacterial abundance were similar to spores were detected in check water( Check 1) on any ofthose in the check tubs with larvae( check 1 w/ larvae) the sampling days( Fig. 5B). In top water of tubs treatedfrom three hours after enriching and filling the tubs up with VectoBac G at rates of 5. 3 and 10. 6 lb/ac, B. t.i.
U Check 1 w/ larvae A- VectoBac G 5. 5 lb/ac
Check 2 w/o larvae VectoBac G 10.6 lb/ac
100Top a. 100
Bottom
A80
tiV181W.P-
elS'atment
80
o
60 60
edpa
40 40
o
b 20 20Treatment
a)
g
0 03 7 13 3h 1 3 7 3 7 13 3h 1 3 7a D 0a D a
Post-Flooding PosttreatmentDays
Post-FloodingD 0 a
PosttreatmentD
Figure 4. Reduction of total bacterial densities from their peak populations on day 3 post- flooding in water of tubsenriched with organic matter and treated with VectoBac G formulation.

198 Journal of Vector Ecology December, 1999
spores were detected in relatively high numbers, reaching DISCUSSION
5. 3 and 10.00 x 103 spores/ ml, respectively, three hoursafter treatment, then rapidly declining to 4.00 and 5. 50 The results of test 1 showed a natural pattern of
x102
spores/ml, respectively, on day 7 posttreatment. In growth and decline of bacterial populations in outdoor
tubs treated with VectoBac G at the high rate of 10. 6 tubs both unenriched and enriched with organic matter
lb/ac, B. t. i. spore counts in top water were signifi- in the winter season. The irrigation water contained a
cantly higher than in water treated at the low rate very low number of total bacterial cells per milliliter,of 5. 3 lb/ ac at three hours and day 3 posttreatment( Fig. while the rabbit pellets used for enrichment of the tubs
5B). contained a large number of bacteria per gram. The
As in the top water, B. t.i. spores in bottom water bacterial populations in enriched water at three hours
were found in high numbers of 7. 75 and 10.0 x103
after enriching and filling the tubs were 11- fold higherspores/ ml at three hours after treatment with VectoBac than those in unenriched water. The bacteria preexistingG at the rates of 5. 3 and 10.6 lb/ ac, respectively, then in rabbit pellets constituted an important source of
gradually declining to 6. 30 x102
and 1. 36 x103
spores/ bacterial abundance in the enriched regimen. These
ml, respectively, on day 7 posttreatment. The treatment bacteria, along with those coming from irrigation water,using the high rate showed significantly higher B.t.i. reproduced in much higher densities in the enriched tubs
spore counts than the low rate at three hours, and three when the rabbit pellets were present. The peak
and seven days posttreatment( Fig. 5B). populations of bacteria on day 3 post- flooding in theenriched regimen were 25- fold higher than in unenriched
Mosquito Production and Control with B.t.i. water. The organic matter required for bacterial growth
Mosquito oviposition activity was noted on day 1 and propagation was in low quantity in unenrichedafter enriching and filling the tubs and reached a peak on water, while in enriched water, the bacteria decomposed
day 7 post- flooding ( Fig. 6A), then progressively and used the ample supply ofrabbit pellets for metabolicdeclined to a low level on day 3 posttreatment. However, activity, growth, and reproduction. After reaching theon day 7 posttreatment, a minor peak of oviposition peak populations, bacterial densities in unenriched and
activity occurred in both check and treated tubs. enriched regimens gradually declined by 83 and 91% at
Early instar( 1st and 2nd) larvae were noted on day the end of the test on day 22 post- flooding, respectively.7 post- flooding in assigned check and treated tubs. Due to lack of adequate mosquito production in cool
Larval densities reached averages of 105. 5- 135. 0 total weather in test 1, these bacterial declines were natural
larvae/dip (Fig. 6B) before treatment on day 13 post- when the source of required nutrients in the environment
flooding( treatment day). The application of VectoBac were gradually exhausted.
Gat the two rates of5. 3 and 10.61b/ ac resulted in equally Analysis of the relationship between bacterialgood control up to seven days( Fig. 6B). The reduction abundance and dissolved oxygen values indicates that
in larval population was 80, 93, and 73% for the low rate the high bacterial counts were directly related to the lowof 5. 3 lb/ac and 94, 93, and 86% for the high rate of 10.6 oxygen concentration in the enriched regimen. In the
lb/ac on days 1, 3, and 7 posttreatment, respectively. unenriched water, the DO values were significantlyThe densities of late instar( 3rd and 4th) larvae were higher than in the enriched water soon after flooding.
39- 49 larvae/dip at the time of treatment in check and The DO in the enriched water decreased to a very lowtreated tubs. The two rates of VectoBac G yielded level which inversely coincided with peak bacterialequally good control up to seven days( Fig. 6C). On days populations. The great reduction in oxygen concentration
1, 3, and 7 posttreatment, VectoBac G reduced the at the peak time of bacterial population in enriched
populations of late instar larvae by 67, 95, and 91% at water indicates strong biological demand for oxygen5. 3 lb/ ac and 91, 95, and 99% at 10. 6 lb/ac, respectively. ( BOD) by the bacteria during the logarithmic division
As to the changes in species composition in check phase. The occurrence of low DO and high bacterial
tubs during sampling period as indicated by identification populations concomitantly was also true in test 2.of late instar larvae, Culex stigmatosoma kept a relatively The results of test 2 clearly showed the influence ofstable proportion of 45. 3- 40. 8%, while Cx. mosquito larval presence in high numbers on bacterial
quinquefasciatusdecreasedprogressivelyposttreatment abundance in water enriched with organic matter.from 33. 8% to 15. 8%. Culex tarsalis, however, increased Bacterial densities in enriched water devoid ofmosquitofrom 14. 9% on treatment day to 35. 0% on day 7 larvae as well as with larvae and treated with B. t.i.posttreatment. Culiseta incidens and Cs. inonata were granular formulation progressively declined to lowfound in low proportions ( 2.0- 5. 8%) during the test numbers at the end of the test. However, the natural
period. decline ofbacterial abundance from peak populations in
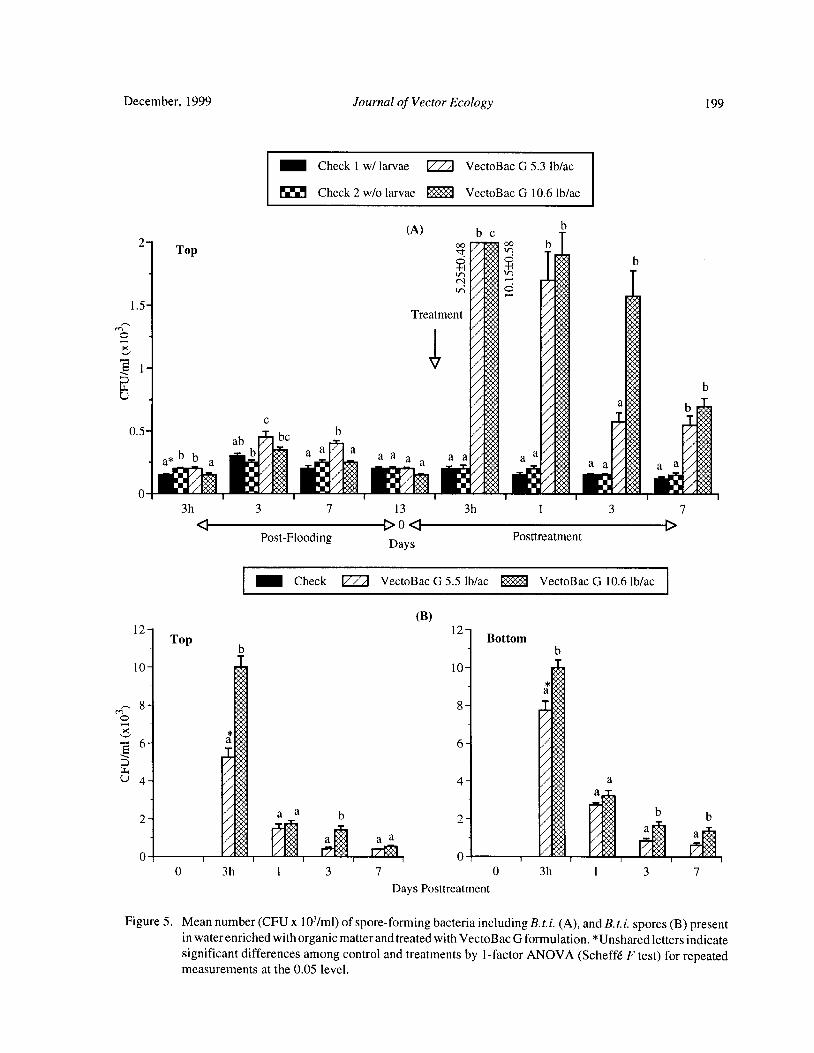
December, 1999 Journal of Vector Ecology 199
Check 1 w/ larvae ® VectoBac G 5. 3 lb/ac
Check 2 w/o larvae R* Sf VectoBac G 10.6 lb/ac
A) b cb
2- Top v PIq:
b
v b
1. 5- Ji AN
ow
Treatment ioo• oe K4
ow
moow ow
mg ow
o iel1vo.
0 0•. mei,VA
t:, 0V: 5:, b ,•;•
too,4owow0. 5-
beb
øri
ab
b a ab 0 a a a p. d o. a a a s a O- a*
ba O•, 4:$a
0 fi 00• _ / Oi% ' P•• A COQ /'•
4 / Oii' i /'• /0•'1
3h 3 7 13 3h 1 3 7
4 D0 4 DPost-Flooding Days
Posttreatment
IIIIII Check ® VectoBac G 5. 5 lb/ ac ZZ VectoBac G 10.6 lb/ ac
B)
12T
12Top Bottom
b b
10 10
O ii i
E r''•''U 4 t:S
IFLt. 4 a
2a s b b
2
a aM a :4 a A a IJ!;'iij! oo
0 3h 1 3 7 0 3h 1 3 7
Days Posttreatment
Figure 5. Mean number( CFU x 10'/ ml) of spore- forming bacteria including B.t.i. (A), and B. t.i. spores( B) presentin water enriched with organic matter and treated with VectoBac G formulation.* Unshared letters indicate
significant differences among control and treatments by 1- factor ANOVA( Scheffe F test) for repeatedmeasurements at the 0.05 level.

200 Journal of Vector Ecology December, 1999
Check ® VectoBac G 5. 3 lb/ ac .. VectoBac G 10. 6 lb/ ac
A)
50-
o 40- aTreatment
a a
4 Ivon 30-to
W
o aa a a aa
ao a
Z 20- a
pr.Q
10—
A•:.:•••••::•••:::::..::•::.::
a por ;;:::::::::
3 7 13 1 3 7
4 D04 DPost-Flooding
Da sPosttreatment
Y
B) C)
160- Treatment 70- Treatmenta
b. Va
a
aba
a 60a
e - a
e 120- aa
cd 50- a
O 7 - a •.•..
40-
w 80- y
o 30-
z - o -N C
o 40^ b20-
b
bb
b ¢ 10-
b• b lb b
p p
0 1 3 7 0 1 3 7
Days Posttreatment
Figure 6. Mosquito oviposition( A), total larvae( B), and late instar larvae( C) developing in water of tubs enrichedwith organic matter and treated with VectoBac G formulation. * Unshared letters indicate significant
differences among control and treatments by 1- factorANOVA( Scheffe F test) for repeated measurementsat the 0.05 level.

December, 1999 Journal of Vector Ecology 201
enriched water withoutmosquito larvae was much lower enrichment on temporal distribution and abundance
than in water with larvae, which was brought about by of culicine egg rafts. J. Am. Mosq. Contr. Assoc.the feeding activity of mosquito larvae. The marked 11: 167- 171.
decline in bacterial numbers in the top and bottom water Hazard, E. I., M. S. Mayer, and K. E. Savage. 1967.with larvae was noted on day 7 post- flooding, with the Attraction and oviposition stimulation of gravidmagnitude of decline higher in the top water than in the female mosquitoes by bacteria isolated from haybottom water. This difference in bacterial declines is infusion. Mosq. News 27: 133- 136.probably due in part to more larval feeding activity in Ikeshoji, T., K. Saito, and A. Yano. 1975. Bacterialsurface water than in the bottom water. The influence of production of the ovipositional attractant of
mosquito abundance on bacterial populations was further mosquitoes on fatty acid substrates. Appl. Entomol.evidenced by the greater reduction in bacterial densities Zool. 10: 239- 242.
in untreated tubs as compared with the treated tubs, Jones, J. G. 1970. Studies on fresh water bacteria: effectwhere B. t.i. treatments caused significant reductions in of medium composition and method on estimates
larval numbers. of bacterial population. J. Appl. Bacteriol. 33: 679-The treatment with the B.t.i. formulation contributed 686.
very little to the total bacterial population. This was Merritt, R. W., E. J. Olds, and E. D. Walker. 1990.
because of the application of relatively small quantities Natural food and feeding behavior of Coquillettidiaof the product necessary for the control of mosquito perturbans larvae. J. Am. Mosq. Contr. Assoc. 6:larvae. However, B.t.i. spores contributed significantly 35- 42.
after treatment to the population of spore- forming Merritt, R. W., D. A. Craig, R. S. Wotton, and E. D.bacteria, which prevailed in very low numbers as Walker. 1996. Feeding behavior of aquatic insects:compared with the total bacterial population. The rapid case studies on black fly and mosquito larvae.decline in B. t.i. spore numbers in treated tubs after Invertebr. Biol. 115: 206- 217.
treatment could be the result of larval feeding activity Mulla, M. S., R. L. Norland, D. M. Fanara, H. A.
and settling-out of bacterial spores. Darwazeh, and D. W. McKean. 1971. Control of
There probably are other factors which influence chironomid midges in recreational lakes. J. Econ.
the population trends ofbacteria in microcosms enriched Entomol. 64: 300- 307.
with organic matter. Bacterial densities and growth Nguyen, T. T. H., T. Su, and M. S. Mulla. 1999.
could be affected by weather, background bacterial Mosquito control and bacterial flora in water en-
populations in incoming water and enrichment substrates, riched with organic matter and treated withBacillus
and availability of organic matter. These factors could thuringiensis subsp. israelensis and Bacillus sphae-likewise influence mosquito larval abundance and ricus formulations. J. Vector Ecol. 24: 138- 153.
survival. We have noted that the decline in bacterial Nilsson, C. 1987. Feeding and food utilization bynumbers was much faster and greater in the presence of mosquito larvae. Ph.D. Dissertation, Univ. of
larvae than the natural bacterial decline in the tubs Uppsala, Dept. of Zoology, Sweden. 269 pp.without larval breeding. In the absence of larvae, the Rodcharoen, J., M. S. Mulla, and J. D. Chaney. 1997.declines in bacterial numbers in the top and bottom Organic enrichment of breeding sources forwater were quite similar. However, the decline in sustained productivity of mosquitoes ( Diptera:bacterial numbers in the top water layer was greater than Culicidae). J. Vector Ecol. 22: 30- 35.
that in the bottom water in the presence of larvae. Su, T. and M. S. Mulla. 1999. Field evaluation of newTherefore, it is evident that mosquito larvae play an water dispersible granular formulation of Bacillusimportant role in the decline of bacterial populations by thuringiensis spp. israelensis and Bacillustheir feeding activity. sphaericus against Culex mosquitoes in micro-
cosms. J. Am. Mosq. Contr. Assoc. 15: 356- 365.Acknowledgment
Walker, E. D., E. J. Olds, and R. W. Merritt. 1988. Gut
We wish to thank John D. Chaney at Department content analysis of mosquito larvae ( Diptera:
of Entomology, University of California, Riverside, for Culicidae) using DAPI stain and epifluorescencehis assistance during the course of this study and critical microscopy. J. Med. Entomol. 25: 551- 554.
reading of this manuscript. Walton, W. E. and M. S. Mulla. 1991. Integrated control
of Culex tarsalis larvae using Bacillus sphaericusREFERENCES CITED and Gambusia affinis: effects on mosquitoes and
nontarget organisms in field mesocosms. Bull. Soc.Beehler, J. W. and M. S. Mulla. 1995. Effect of organic Vector Ecol. 16: 203- 221.

Journal of Vector Ecology 24( 2): 202- 215
Effects of Neem Products Containing Azadirachtin on Blood Feeding,Fecundity, and Survivorship of Culex tarsalis and
Culex quinquefasciatus ( Diptera: Culicidae)
Tianyun Su and Mir S. Mulla
Department of Entomology, University of California,Riverside, CA 92521- 0314, U.S.A.
Received 30 April 1999; Accepted 12 July 1999
ABSTRACT: When late 3rd or early 4th- instar larvae of Culex tarsalis Coquillett and Culex quinquefasciatus
Say mosquitoes were treated with sublethal dosages of neem insecticide until pupation, the blood- feedingactivity of the resulting adults was essentially the same as that of untreated controls. In contrast, blood-
feeding activity was suppressed when newly emerged adults were fed continuously on 10 parts per millionppm) or 50 ppm azadirachtin( AZ) in 10% sucrose solution for seven days. Fecundity was also reduced
by the various neem treatments. When late 3rd or early 4th- instar larvae were treated with 0.010 ppm AZto pupation, the resultant females had a lower rate of oviposition than did the untreated controls after a fullblood meal. When late instar larvae were treated at 0.005 ppm and 0.010 ppm AZ, the resultant females
produced smaller egg rafts after a full blood meal, as compared to the controls, but egg viability was notaffected. In newly emerged adults feeding continuously on 10 ppm and 50 ppm AZ in 10% sucrose for sevendays( before blood feeding), the oviposition rate, size of egg raft, and hatching rate of the eggs after a fullblood meal were all reduced. When newly blood-fed adults were fed continuously on 10 ppm and 50 ppmAZ in 10% sucrose for five days, their oviposition rate was lower than controls in most cases, but the eggraft size and viability of eggs were not affected. In freshly blood- fed females topically treated with AZ with1 or 5 pig/ female, the oviposition rate and size of egg rafts were generally reduced. The females receivingtopical treatment laid eggs and their hatching was not affected. The longevity of adult females feedingcontinuously on 10 ppm and 50 ppm AZ in 10% sucrose solution after emergence was reduced, whereas,
the longevity of males was only affected at the higher concentration.
Keyword Index: Neem, azadirachtin, Culex tarsalis, Culex quinquefasciatus, blood feeding, fecundity,survivorship, mosquito control.
INTRODUCTION Mordue and Blackwell 1993, Mulla and Su 1999). With
regard to mosquitoes, it has been known that at least 344Due to environmental concerns and the development species of plants including the neem tree are known to
of resistance to synthetic chemical insecticides, contain bioactive materials that show someresearchers have focused attention on the development mosquitocidal activity ( Sukumar et al. 1991). Neemofalternative pesticides, such as biopesticides possessing products serve primarily as larvicides against mosquitoesa wider margin ofsafety to the environment and nontarget ( Su and Mulla I998a, Mulla and Su 1999), even thoughbiota. As renewable resources, botanical components technical AZ and experimental formulations ofAZ havepossessing pesticidal properties constitute a major also shown ovicidal capability ( Su and Mulla 1998b),category of biopesticides. Among these entities, the and crude preparations of neem seed or technical AZbioactive preparations from the neem tree( Azadirachta exerted effects on reproductive events of adult mos-indica A. Juss) containing the tetranotriterpenoid quitoes ( Dhar et al. 1996, Ludlum and Sieber 1988).azadirachtin( AZ) have received much attention. Neem Because neem products adversely affect differentextracts exhibitmultiple effects on various insect species, systems of test insects at certain dosages( Schmutterersuch as feeding deterrence, growth regulation, fecundity 1990, Ascher 1993, Mordue and Blackwell 1993), it issuppression, and even blockage of development of likely that adult mosquitoes might be equally affectedvector-borne pathogens( Schmutterer 1990, Ascher 1993, by exposure to neem products. As part of our ongoing

December, 1999 Journal of Vector Ecology 203
research on the activity and biological effects of neem Effects on Blood Feedingproducts against mosquitoes, the current study was Delayed effects from larval treatment: In order to
initiated to elucidate the impact of experimental neem elucidate any delayed effects of the treatment with neemformulations on blood feeding, fecundity, and formulation applied to the larval stage on blood feedingsurvivorship of adult Culex tarsalis Coquillett and Cx. of the resultant adults, about 1, 000 late 3rd or early 4th-quinquefasciatus Say. We examined delayed biological instar larvae from the colonies of two species were
effects of neem products on adults resulting from transferred to distilled water containing the sublethaltreatment of larvae at sublethal concentrations as well as dosages of 0.005 ppm and 0.010 ppm AZ of both
adults treated directly with neem products either per os formulations, which were made by adding 1 ml and 2 mlor via topical applications. of the 10 ppm stock suspension to 2 liters water in an
enamel pan, respectively. The larvae were providedMATERIALS AND METHODS with 5 g rabbit pellets as food in each pan, and exposed
to the treatment until pupation. Controls consisted of
Mosquito Culture late 3rd or early 4th- instar larvae from the same stocksThe test species were Cx. tarsalis and Cx. quin- as treated larvae in both test species were transferred to
quefasciatus. Larvae were reared in two separate culture the same amount of distilled water in the same size pans,
rooms at 26± 1° C, 35- 45% RH and a 16: 8( L:D) hour and fed on the same amount ofrabbit pellets as in treated
photoperiod with 1 hour dawn and 1 hour dusk. To larvae. The pupae from treated and control pans were
maintain the colony, 4 to 5 egg rafts were placed in an collected as needed. About 200- 300 pupae were
enamel pan( 40 x 24 x 6 cm) containing 3 liters distilled transferred to each of two 240- m1 waxed paper cups
water. Rabbit pellets( Brookhurst Mill, Riverside, CA., containing 200 ml distilled water and placed in a screenedcrude protein>_17%) were added as larval food to the cage. Newly emerged adults of both treatments andrearing pans at the rate of 5 g/pan when the egg rafts control were provided continuously with 10% sucrose
were set up and then again when the larvae reached solution. On day 5 postemergence, the adults werethe early 3rd- instar. Water was added to the pans deprived of sugar feeding for 12 hours and offered a
every other day to replenish loss due to evaporation. one- week- old chick overnight for blood. The followingPupae were removed from the pans and placed in morning, the total number of blood fed and unfed240-ml waxed paper cups each containing 200 ml of females was counted and blood- feeding rate (%) was
distilled water and placed in screened cages( 23 x 23 x calculated as number of blood fed/ total number of
32 cm) where the adults emerged. Adults were females multiplied by 100. Three cages of adults wereprovided continuously with 10% sucrose solution in used for each formulation at each AZ concentration
a jar with a cotton wick. On day 5 postemergence, against each test species. The differences in blood-
the adults were deprived of sugar feeding for 12 hours, feeding rate between control and treatment werethen provided with restrained 1- week- old chicks determined by a Chi square test.overnight for blood feeding( Animal Use Protocol No. Adult treatment: About 200-300 pupae from the
A- S 9901002- 1, University of California, Riverside). regular colonies of the two test species were put in a
The late 3rd or early 4th- instar larvae, newly emerged screened cage. Newly emerged adults were allowed toadults or newly blood- fed females were used for different feed continuously on a 10% sucrose solution containing
experimental purposes. 10 ppm and 50 ppm AZ, which were made by adding 1ml and 5 ml of 1, 000 ppm AZ stock suspension to 99 ml
Neem Products and Presuspensions and 95 ml distilled water held in a 200-m1 plastic jar with
The test neem formulations were Azatin WP4.5 a cotton wick. After seven days continuous feeding onWP) and Azatin EC4.5 ( EC) ( Thermotrilogy Corp., the neem- treated sugar solution, the adults were used for
Columbia, MD, USA). Both formulations contained blood- feeding tests. The control adults were from the4.5% of the active ingredient( AZ). Stock suspensions same batch of pupae, but fed on 10% sucrose solution
of the formulated products were prepared in distilled after emergence for the same period ( 7 days) without
water at the concentrations of 10 ppm and 1, 000 ppm AZ. The adults of control and treatment were deprived
AZ, which were used for larval treatment and adult offeeding on sugar solution or sugar solution containingtreatment, respectively. For each test, stock suspensions neem for 12 hours before offered the host overnight for
were freshly prepared. All the tests were conducted in an blood feeding. The same methods were used as in theinsectary maintained at 26± 1° C, 35- 45% RH and a 16: 8 previous larval treatment tests to determine blood-
L:D) hour photoperiod with 1 hour dawn and 1 hour feeding rates. Three replicates were made in each testdusk. and a Chi square test was employed to determine

204 Journal of Vector Ecology December, 1999
significant differences in blood- feeding rates between control and treatment groups were deprived of feedingcontrol and treatments. on sugar in controls or sugar and neem in treatments for
12 hours before blood feeding. After overnight bloodEffects on Fecundity feeding, 20 fully engorged females were removed to
In order to explore the effects of various types of other cages and 10% sucrose solution was providedexposures in the larval stage or direct treatment ofadults continuously. Five cages were setup for each control oreither orally or topically by neem products on fecundity treatment. The handling procedures for obtainingof adult mosquitoes, such as oviposition rate, size ofegg oviposition, egg raft, and individual egg counting andrafts, and viability of eggs, the mosquitoes were treated hatching, as well as the statistical methods used were theas follows. same as those in the previous test of" larval treatment."
Delayed effects from larval treatment: Approxi- Adult/oral treatment after blood meal: About 200-mately 1, 000 late 3rd or early 4th-instar larvae were 300 pupae from the colonies ofeach species were placedexposed to sublethal dosages of neem at 0.005 ppm and in a screened cage and the newly emerged adults were0.010 ppm AZ until pupation. The resulting pupae were allowed to feed continuously on 10% sucrose solution
collected, 200-300 of which were placed in a screened for five days. After starvation for 12 hours by removingcage. Adults emerging from these pupae were fed the sugar cup from the cage, the adults were blood fedcontinuously on 10% sucrose solution for five days. with a chick host overnight. After blood feeding, 20They were deprived of sugar feeding for 12 hours, then fully blood-engorged females were aspirated into eachgiven a chick overnight for blood feeding. The adults cage, using five cages for each treatment and control.from the colonies of both species reared without The blood-fed females were fed on 10% sucrose solutionexposures to neem were used as controls and handled in for five days as controls, and the other group fed on 10%an identical manner. The fully blood-engorged females sucrose solution containing 10 ppm or 50 ppm AZ forfrom controls and treatments were removed with an five days as treatments. On day 5 after blood feeding,aspirator, where 20 females were placed in each of five gravid females were provided with oviposition cups.cages and provided with 10% sucrose solution for five The handling procedures for oviposition setting, eggdays. The gravid females were then provided with an raft, and egg counting and hatching test, as well asoviposition cup containing 100 ml distilled water. The statistical methods were the same as those in the previousnumber ofegg rafts and number ofeggs in each raft were test of" larval treatment."counted the following morning. The oviposition rate Adult/topical treatment after blood meal: About
was calculated from the total egg rafts collected 200- 300 pupae from each colony were placed in a cage,from five cages( total of 100 females) and subjected to and newly emerged adults were fed continuously ona Chi square test to explore the impact ofneem treatments. 10% sucrose solution for five days. Before a host wasThe average number of eggs in an egg raft was also provided for an overnight blood meal, the adults werecalculated and subjected to a t test to determine if there deprived of sugar feeding for 12 hours. The followingwere any effects of neem treatment on the size of egg morning, the fully blood fed ones were aspirated andrafts. To ascertain hatching rates of eggs, five egg rafts immobilized with CO2 and treated topically on theselected at random from the egg raft collection were abdomen with 1 or 5 µ g AZ/female using a 5- 1lplaced individually in 120-m1 waxed paper cups with micropipette, in the form of 1µ l/ female and 5 . tl/ female100 ml distilled water. The hatching was assessed three ofthe 1, 000 ppm AZ suspension ofthe test formulations.days later and the larvae from each egg raft were After treatment, 20 females were transferred to each ofcounted. Hatching rate was calculated as the total number five cages and provided with 10% sucrose solution forof larvae hatched/ total number of eggs in five egg rafts five days. Control adults were handled in the sameobserved, multiplied by 100. The hatching rates of manner but treated topically with equal volumes ofcontrols and treatments were subjected to a Chi square distilled water. Five cages were set up for control andtest to evaluate the possible impact of neem treatments treatment in each case. On day 5 after blood feeding,on egg viability. gravid females were provided with oviposition cups.
Adult/oral treatment postemergence( before blood The handling procedures for oviposition setting, eggfeeding): About 200-300 pupae from the colonies of raft, and individual egg counting and hatching test, aseach species were placed in a screened cage and newly well as statistical procedures were the same as those inemerged adults were allowed to feed continuously for the previous test of" larval treatment."seven days either on a 10% sucrose solution( controls),
or on a 10% sucrose solution containing 10 ppm or 50 Effects on Adult Survivorshipppm of AZ of each test formulation. The adults of About 350- 400 pupae from each colony of the test

December, 1999 Journal of Vector Ecology 205
species were collected in two 240-ml waxed paper cups, males) were fed continuously on 10 ppm AZ in 10%each containing 200 ml distilled water, and introduced sucrose solution for seven days postemergence( before
into a screened cage for emergence. The newly emerged blood feeding), the oviposition rate in treated femalesadults were allowed to feed continuously either on 10% was significantly reduced after afull blood meal. Anothersucrose solution ( controls) or on sugar solution group ofadults was treatedper os after ingestion of a fullcontaining 10 ppm or 50 ppm AZ of the WP or the EC blood meal with 10 ppm AZ in 10% sucrose solution for
formulations ( treatments). To insure the continuous five days. In these tests, only Cx. quinquefasciatusavailability of sugar solution or the sugar solution exhibited lower oviposition rate than the controls. Topical
containing AZ, a quantity of 150 ml of the feeding application of AZ at 1 . tg/female on the abdomen ofsolutions was placed in a 200-ml plastic cup with lid, newly blood-engorged females also significantly reducedthrough which a cotton wick was placed to absorb the oviposition rate with both neem formulations in both
liquid for feeding access of the adult mosquitoes. The species( Fig. 2).feeding solution and cotton wick were replaced every In further tests, a doubled AZ concentration( 0.010
three days with fresh preparations. The dead female and ppm) was applied in larval treatment. In adult feedingmale mosquitoes were separately collected and counted tests, either postemergence for seven days or after blood
every three days. The test was conducted until the last feeding for five days, five times the AZ concentrationsadult mosquito died. Three replicates were made in both ( 50 ppm) were added to 10% sucrose solution. In topical
control and each treatment. The cumulative mortality application tests, the dosage was increased to 5 µ g/
and lethal time for 50% mortality ( LT50) with 95% female. In these tests using high doses, the ovipositionconfidential limits were determined and analyzed using rate was significantly reduced in all tests, indicating aaprobitregression line( Finney 1971, Abacus Concepts, dose- dependent action( Fig. 2).Inc. 1987). Average size ofegg rafts: The number of eggs in all
the egg rafts collected from the above oviposition testsRESULTS was counted and the averages calculated and analyzed.
In the tests using low AZ concentration, the fullyEffects on Blood Feeding engorged females resulting from larval treatment using
The neem exposure tests were carried out by 0.005 ppm AZ produced significantly smaller egg rafts
exposing both larval and adult stages. When the late 3rd than the controls. In adult treatment tests, when the
or early 4th- instar larvae of the test species were treated cohorts were allowed to feed on 10 ppm AZ for seven
with sublethal dosages ofAZ( 0.005 and 0.010 ppm AZ days postemergence before blood feeding, fully engorgedin WP and EC formulations), the blood- feeding activity females also laid significantly smaller egg rafts than didof the adults was not affected, except in the case of Cx. controls. The treatment of females by feeding on AZtarsalis treated with the EC, where the blood- feeding after blood feeding, however, did not affect egg raft size.rate in the treatment group was significantly lower than As to the post-blood feeding treatment by topicalthat of the control( Fig. 1). In another experiment where application at 1 . tg/female on the abdomen, only thethe newly emerged adults were fed on 10 ppm and 50 treatments with the EC formulation caused a reduction
ppm AZ for seven days, the blood- feeding activity in in egg raft size in both test species( Fig. 3).resulting females was adversely affected, showing a In further tests, a doubled dosage of AZ ( 0.010
significant reduction in feeding rate compared with the ppm) was used for larval treatment and the high
controls, regardless of species and formulations. concentration of 50 ppm of AZ was used for adult
Suppression of blood- feeding activity was dose- treatment by feeding( postemergence or after ingestiondependent in both species, with greater suppression of blood meal) and 5 pg/ female for topical treatment.occurring in the higher dose ( 50 ppm AZ) treatment All these treatments, except AZ feeding after ingestionFig. 1). ofblood meal, reduced the average egg raft size( Fig. 3).
Hatching ofegg rafts: Five egg rafts were selectedEffects on Fecundity at random from the egg raft collection in the tests for
Oviposition rate: In the initial tests, low dosages of oviposition rate, and transferred individually to 100 mlAZ were applied in larval and adult treatments. When distilled water in a 120- ml waxed paper cup for hatching.the late 3rd or early 4th- instar larvae were treated by In the low dosage treatments, i.e., 0.005 ppm AZ for
0.005 ppm AZ of two neem formulations, the oviposi- larval treatment, 10 ppm AZ for adult feedingtion of resulting adults of both test species after a full postemergence or after ingestion of blood meal, and 1
blood meal was comparable to the controls. In adults gg/ female for topical application, only the egg rafts laidtreated orally, when the cohorts ( both females and by the females fed on AZ for seven days after emergence
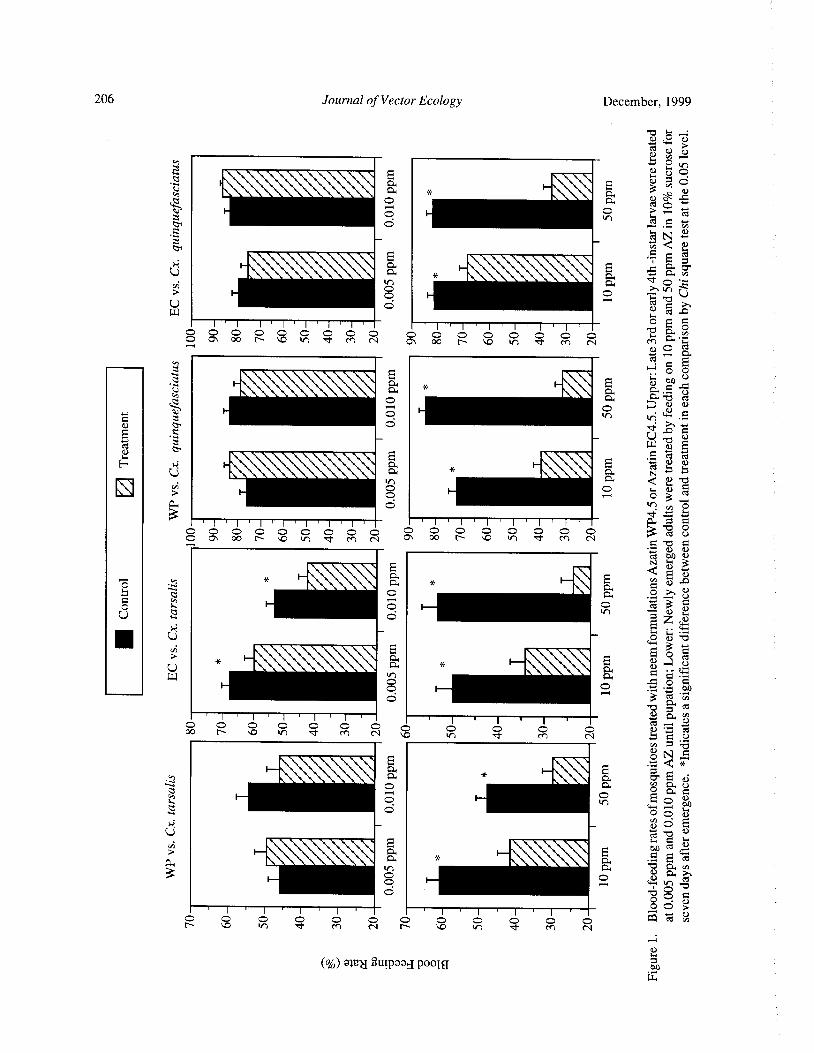
206 Journal of Vector Ecology December, 1999
cu
a 3cs O a 0o N--,
a OIn
Cr
00 0 n UW
d co gt.
I I 1 I I 1 I I ' I ' I ' I ' I ' I ' b G. a0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0n G,,.O Oh 00 N 0 V' 1 ' cF M N p\ 00 [ 0 In d M N M
a
a 0
ci
E *
a a. tb o
a a. '-elN N
co OO w C
ci
ay
tn a dg cu cri
kn ,,
I I I I I I I 1 ' 1 ' 1 ' 1 ' 1 ' 1 ` 71: 5 00 O 0 O `°
oO 0 O O 0 C ° C C C O C 3 a OV
a— a
rt. bA CU
IN E
O p, O > o
a o o 3
w
S o -E 0 ao
0
3 a.7not) a yI I I I I
00 NN C 0
In0O o o 0 o a 2
i ° 'd
off *:
11. 116aa
0
o 0 o a0 0p
oE
c:1 I— a gI
iiiii v a - o a v
O O w O. dOO O N
OC
O O O O O 0 O0 O O
InN
c6.mull 2utpood poolg
g
w

December, 1999 Journal of Vector Ecology 207
00 Ocal
U
O a' b ti
I61. 111111Xi_iiii p4 c G CbA
z obbly vi
UO
o Ucl a
A Ilk t a
U a)a)
I I ' I I ' 17 11 ' I ' a) w
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >
cA 00 N . O ' n 7r M N '--, O\ 00 r` . 0 M N ti ow y ,,..,$.
4 _
tn
zt
u. ` °b
Z, c t
jmmlla, g,*
c °"o .O ] ° w
E 5 aoo o
F w
ci N O ro ~a .
w ^0 chi o
1 I 1 I 1 1 1 I I 1 1 1 I 1 Io a a. o
0 0 0 0 0 0 0 0 0 o o o 0 0 00 oN 0 0 0 a 2 . 0
00 N D V1 !' M N p„ a)
per,O
1—. M. Mi ;Li
pp,
N d‘c~d
C p. Q > O rnp O cd - 5 ,V 0 C
cad ,Z
v w gc' s > >, v
01-- o
U w o ,-4
o o•
Cn 64
t4
Hiiiiiiiii64
aa• a °' ° Cl.
bUA al bA o1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' 1 ' a-i ' a U
O 0 O OCO O O O O c o op O o 0 0 a
2 o L2 ad
414. 1iii6i6 1. 111Mliiii*
LI.,
oo °w • 5 84 Vs
a,
Si I— Iiiiiiiiiiii:mmiliiigi
O o T'
U wo o u ott
vi>
O • o cpo
o b
4iiiiiiiii61- 4, iiiiii a • v, C w a. yo o . b a. a)
0 0 3
I ' I I 1 1 ` 1 I ' I ' 1 ' 1 ' I ' a,° N
0 co O O o 0 0 0 o o o 0 0 0 o O o.,715 an
vO v1 M NN
a)F.
O1J UOWSOdtnp Cw"
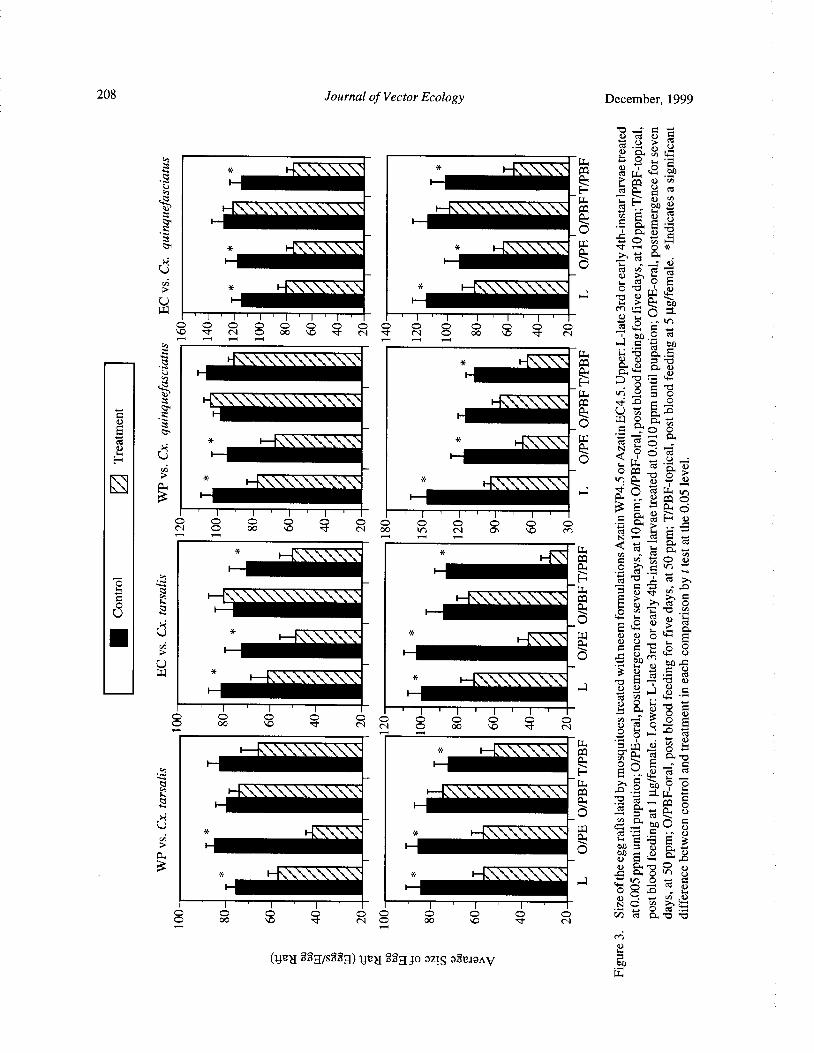
208 Journal of Vector Ecology December, 1999
z
iimiBlik Ni r4
cla) w
1— iiiiiiiii
0 .
l,"- 112
U0 >,,-
u bti7wm .. 0
in0 0 0 0 0 0 0 0 0 0 p 0 0 0 0 0w •D ' f N 0 00 \ D N N O 00 VD d• N 0 «ti ,- I ,- r .-' a i bbA
N 7 b
1ml. gc, ,...
0w
H ` a •. 4"
Izs
P4 U o a,Zk,. l W•
a 0 ..z Let o
44 V O Q 9 ° •
Pli.. li, 4,:
ii3 . „ o
o O 0 0 0 0 0 0 0 0 0 0 a > H . cN O 00 VD ' t N 00 N O") VD m
d a,
H • To a a
L 616. maccl,
U o I, -w au
1\ o s.
o 5to M
by b w ta
w • F.,
O 0N N
0 00 0N cu ct o
o o a
o\ le Lu
o ano a.) C.OA
Bilk IN Ilk Ille
2.,
o 8 z wN o ,,, w
0000
0 NN 0000
co Cn O b bN
c* i
ijuN S8gis229) 1} rg Bag Jo azis 32riand to
w

December, 1999 Journal of Vector Ecology 209
before blood feeding) exhibited a significantly lower ( Mordue and Blackwell 1993, Ascher 1993, Remboldhatching rate than the control, regardless of species and 1995). If this is also the case in mosquitoes, it mayformulations. All other treatments failed to reduce the account for the negative impact of AZ in the initiationhatching of the resultant eggs laid by treated females, process of blood feeding in test species.with the exception of Cx. tarsalis treated topically with As an extended effect of larval treatment, thethe EC( Fig. 4). resulting adults exhibited a lower oviposition rate after
In further tests, the high doses of AZ, i.e., 0.010 a complete blood meal when using 0.010 ppm AZ forppm AZ for larval treatment, and 50 ppm AZ for adult larval treatment. The resultant females laid smaller eggfeeding ( postemergence or after ingestion of blood rafts at both 0.005 ppm and 0.010 ppm AZ in larvalmeal) and 5} tg/ female for topical application were used. treatments. Treatment of larvae with neem usuallyThe treated females feeding on neem postemergence for yielded less fit adults with small body size observedseven days laid eggs with lowered viability. The eggs qualitatively, which showed a lower reproductionlaid by the females treated topically by AZafter ingestion capacity( Takken et al. 1998).of ablood meal also showed lower viability than controls, The fecundity of females was reduced by variousexcept in the case of Cx. tarsalis treated with the WP. AZ treatment regimens applied directly to the adultOther treatment regimens did not have any adverse stage. In particular, continuous feeding on 10 ppm andeffects on egg viability( Fig. 4). 50 ppm AZ in neem products in 10% sucrose for seven
days after emergence ( before blood feeding),Effects on Survivorship significantly reduced oviposition rate and size of egg
Cumulative mortality: In each case, the mortality of rafts after a full blood meal. The viability of eggsmales was higher than that of females. Feeding on both resulting from this treatment was also reduced. Thisthe WP and EC formulations in 10% sucrose solutions phenomenon was seen for both neem formulations andincreased the mortality ofboth females and males. This test species. From the previous studies, as mentionedeffect was more obvious when they were fed on 50 ppm earlier, the dominant mode of action of AZ againstAZ than on 10 ppm AZ( Figs. 5, 6). target insects is to reduce hemolymph JH titers( Mordue
Lethal time for 50% mortality (LT50): The probit and Blackwell 1993, Ascher 1993, Rembold 1995). Itanalysis of the mortality data of the females and males has been well known that in mosquitoes, JH stimulatesfeeding on neem formulations indicated that 10 ppm AZ previtellogenic development of the follicles( Meola andalone reduced the longevity of females, while 50 ppm Readio 1988, Klowden 1997). We suggested thatAZ was detrimental to the survivorship of both females ingestion of AZ after emergence prior to blood feedingand males( Fig. 7). inhibits the previtellogenic development of the ovaries
by reducing JH titers, which could be one of the reasonsDISCUSSION for reduced fecundity. The vitellogenic stage of follicle
development( see below) may also be affected by theIn the current study, the commercial formulations residual effect of ingested AZ before blood feeding. As
developed for agricultural pest control were employed to the reduced viability of the eggs laid by the femalesto investigate effects on blood feeding, fecundity, and feeding on AZ, three factors could be involved. First, JHsurvivorship of adult mosquitoes, Cx. tarsalis and Cx. mediates the development of female receptivity to malesquinquefasciatus. In most cases, larval treatment using by acting on the terminal abdominal ganglion thatsublethal doses of AZ did not have any delayed effects regulates mating behavior( Klowden 1997). If it is alsoon blood feeding of the adult females. The blood- the case in our test species, ingested AZ might reduce
feeding activity of both test species, however, was female receptivity to males through suppressed JHimpaired when the newly emerged cohorts were allowed production. Secondly, from the available evidence into feed continuously on 10 ppm and 50 ppm AZ of both other insect species, response to sexual pheromonesneem formulations. It is well known that the initiation of ( Dorn et al. 1987), spermatogenesis in males( Shimizublood- feeding behavior in mosquitoes is hormonally 1988), and transmission of spermatozoa to the femalesmodulated. Juvenile hormone( JH) III secreted by corpora ( Schmutterer and Rembold 1995) can all be affected byallata in Culexmosquitoes is responsible for the initiation the AZ treatment. We assume that continuous ingestionof blood- feeding behavior after emergence or after of AZ after emergence until blood feeding could haveoviposition ( Meola and Readio 1988). In other insect some subtle effects on mating and the inseminationspecies, one of the modes of action of AZ is to reduce process. Finally, the low hatching rate of the eggs laid byhemolymph JH titer by inhibiting the release of treated females was partially related to the interferenceallatotropins from the brain-corpus cardiacum complex ofapplied AZ with both previtellogenic and vitellogenic
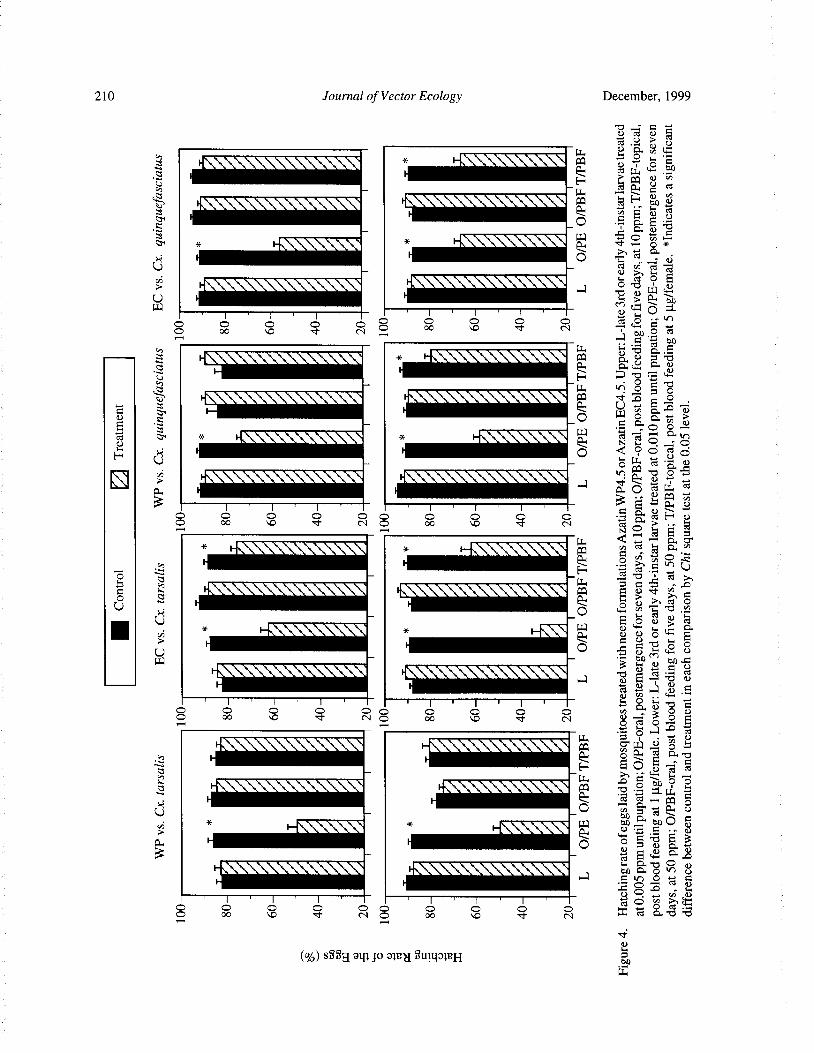
210 Journal of Vector Ecology December, 1999
ct .V > U
ct
O flu
O 0 o oo 0co N Oo0
I so. w a a0i
Q U p .O y
F" V O
fli$1° O °
ElCi
0 0 0 0 0 0 0 0 0 0 a 8 E a)0 00 \ O ' t N o 00 N 174 >
Slii. 11 SI. 1111 f:
4 IIIHU O 0 b •0
w > c3
I.
cA Pik 111i ILIWel.)
UO '
5• a) m00
a4- •
0 000 NN ° o 000 qD ' t
No0•
Ratu
p
IL NC IX N.' CZcn
E hillo
fir 400, tb=)l) t
E 3
h9Io
0 o ° 0 oa0c bo 00 d o 00
vcu
Mg am Jo awl' 2UTya; eH 0bow"
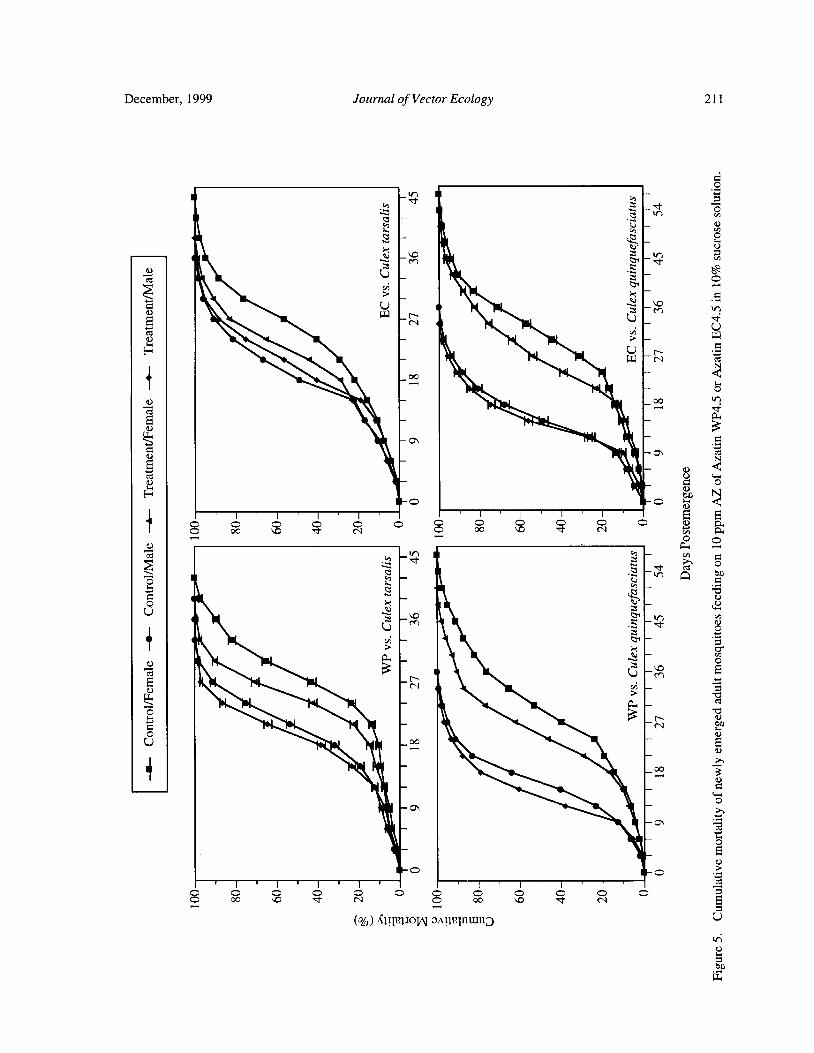
December, 1999 Journal of Vector Ecology 211
o
r •
a _©
jKt _$
ratI1
y • E M
QF't
N
fo
a 00 r
w
E
a
a \a a 0w
7I , I , I . 1 , , , 1 ,
0 0 0 0 0 0 0 0 0 o c oE \
a
7 00 ' N 2 N
oa
2 a
enCr -
1o
Ea 7
af
4 00
ta
C
a
i
c 2
1 . , . , . i , i , 1
o 2 2 9 N ° C 2 2 \ N ° E
c )%! A w a mmn» Uw
bw
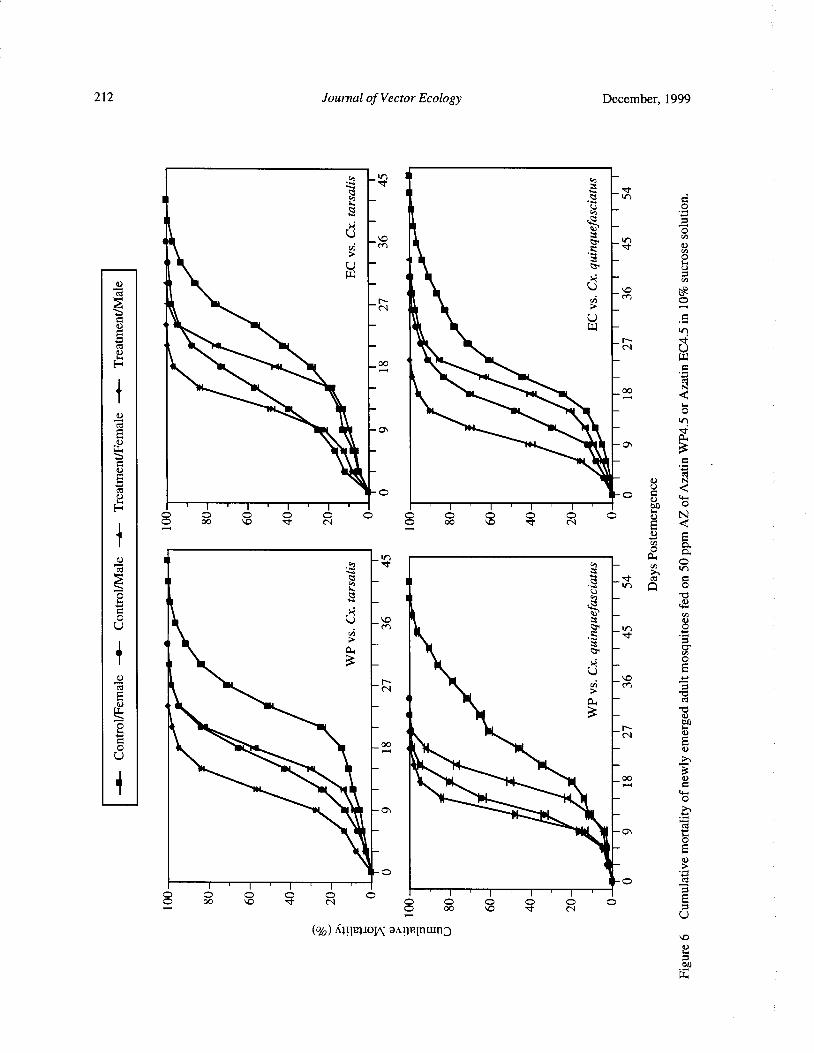
212 Journal of Vector Ecology December, 1999
h — in y —E.
4 —. 1-
U 0
IUW _ K
1U
N IM
o
Er
W —
0
b 48 N U
Et:1 00
IIIIW
r
1 N
NO
ct O C
I I I ' I I
O ° O pp ° 4 O O y
cc N co
OC3'O.
t4---, ti
i i t IL
0
to
N N
a-' 4.
0401411144404•4 1\
oo , EU
111)11%
141 _ wO
a,
O Yo
U
iil>lnioN anpnlnwnJ
a)
0bnw"
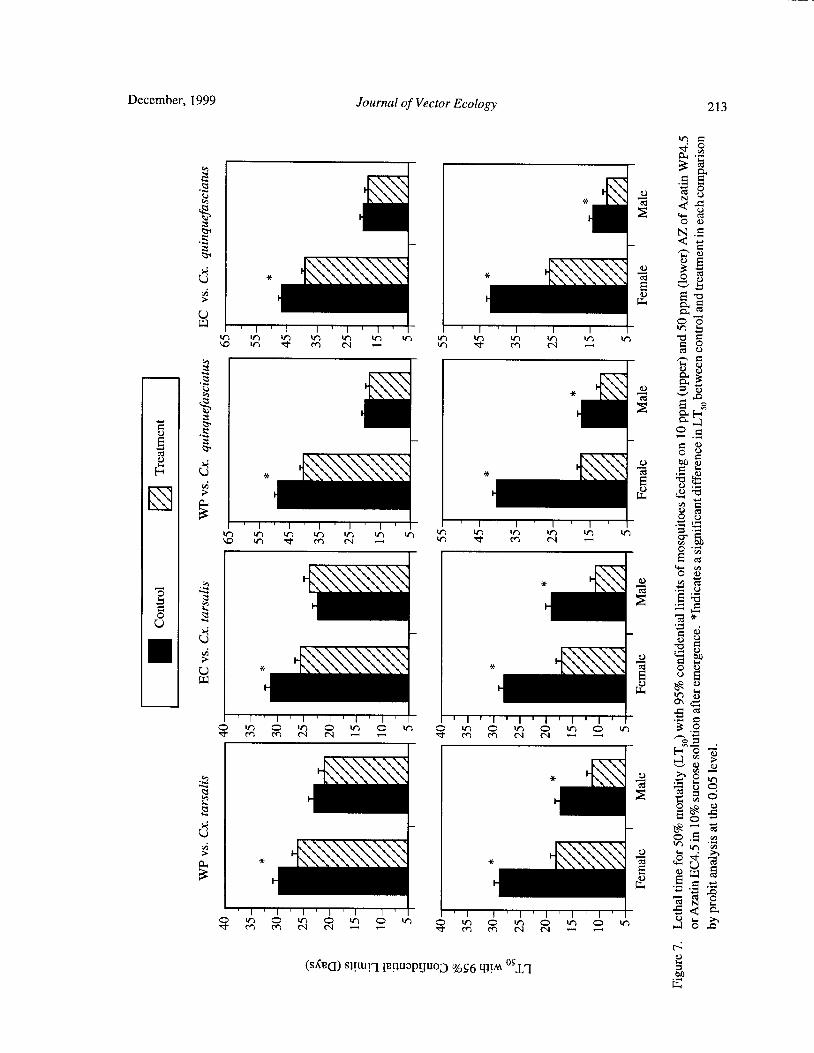
December, 1999 Journal of VcrEcology gg
2
3 §
k '' \y 6)
Ilk, ii
E
m
RE79`
7
L>\ q \1 , 1 , 1 , 1 , , , i i I
aG ? % N
u
k1 t
n z
o 2 awQ
2 §
2 0 814/
125 • Jk
ct
i ' i ' i ' r ' i ' i i i i
N N © © m
n4.
k . \k /
0 2 g *U
0•
t §
w
k 0,
1 ' i 1 , 1 , 1 , i . , . , , i . , . i . 29 m \ N N — — © 9 % % N N
0 In— —
g
u ) /
kU k /
116h.
iiiiiia ®
U
a
1 , 1 , 1 , i , 1 , 1 , 1 , 1 , 1 , , , , , 1 ,
7 N N & 2 © 9 % / N N - — ® Q # z
w
@AQ S ] i*ateo%S 11I± 1
E

214 Journal of Vector Ecology December, 1999
ovary development, yielding eggs with low viability blood meal) decapitated 10 hours after a blood meal,
see below). whereas, 96% ofdecapitated control females contained
In most cases ofAZ exposures after blood feeding, maturing oocytes( Ludlum and Sieber 1988). The lattereither orally for five days or topically at 1 and 5 µ g/ studies suggested that AZ delayed the release of one or
female application, the parameters of oviposition rate more factors from the brain that regulates oogenesis.
and average size of egg raft were affected. Hatching rate Our findings and interpretations in the current
of the eggs was reduced in some cases of topical investigation agree with this study. For explanation of
treatment. In these treatments, the females have the observed negative impact of various neem exposures
undergone the previtellogenic stage in ovarian on blood feeding and fecundity in this study, generaldevelopment before blood feeding. Therefore, lower metabolic disturbance by applied AZ could be anotherfecundities were the results of partially impaired reason, in addition to assumed endocrine disturbance.
vitellogenic process due to AZ. In anautogenous AZ has detrimental effects on a variety of tissues
mosquitoes, vitellogenesisisinitiatedafterbloodfeeding and cells, especially those with rapid mitosis, e. g.,
under the control of endocrine system, where ecdysone epidermal cells, midgut epithelial cells, ovary and testis
secreted by ovaries stimulates the fat body to synthesize etc., which results in an overall toxic syndrome after
vitellogenin using the nutrients from blood meal digestion exposure. Biological fitness of many insect species isKlowden 1997). Some studies indicated that JH may substantially reduced even at neem concentrations and
also participate in regulation of vitellogenin synthesis in doses below those interfering with the molting process.conjunction with ecdysone ( Kelly et al. 1987). JH Changes in biological fitness in test insects can be
increases the competence of follicles to ecdy- manifested as reduced lifespan ( Wilps 1989), high
steroidogenic hormone( OEH) to synthesize ecdysone mortality( Dorn et al. 1987), loss offlying ability( WilpsShapiro and Hagedorn 1982), and competence of the 1989), low absorption of nutrients ( Wilps 1989),
fatbody to ecdysone to synthesize vitellogenin( Flanagan immunodepression( Azambuj a et al. 1991), and enzymeand Hagedorn 1977). Reduced fecundity in females inhibition( Nagvi 1987). In the current study, continuous
feeding on or treated topically with AZ after blood AZ feeding after emergence increased mortality andfeeding could be partially attributable to the negative reduced longevity, which implied an extensive impacteffects of AZ on vitellogenesis through interference of AZ treatment on the target species. It seemed that
with hormone system. Circumstantial evidence is females were more vulnerable than males to AZ
available in other insects, where the mode of action of treatment.
AZ includes a reduction of hemolymph JH and When neem products are used as larvicides against
ecdysteroid hormone titers by inhibiting the release of mosquitoes, delayed effects of larval treatment on
allatotropins and prothoracicotropic hormone( PTTH) reproductive capacity in resultant adults may occur. Tofrom the brain- corpus cordiacum complex( Mordue and date a number ofneem products have been manufactured,
Blackwell 1993, Ascher 1993, Rembold 1995). In most commercialized, and registered to control pests of
cases of topical treatment using 5 . tg/female after a ornamentals, landscape plants, and food crops( Immaraju
blood meal, the eggs laid by treated females exhibited 1998). Natural mosquitopopulations will have the chance
lower hatching than the controls, which implied that to ingest and contact neem constituents during nectarpoor viability of the eggs was related to the detrimental feeding or resting on treated foliage. This exposureeffect of AZ on follicle development. Because no neem could have marked effects on blood feeding, fecundity,exposures occurred before blood feeding, the supposed and survivorship ofthe adult mosquitoes. Neem productsinterference of AZ with mating and/ or insemination also have the potential to be used as larvicides against
could be excluded. mosquitoes. As established in our current study, variousIn some comparable studies conducted on insects modes of contact and exposure induce a variety of
of medical and veterinary importance, uptake of AZ behavioral and biological effects in mosquitoes.
through the cuticle or via ingestion negatively affectedreproductive events of the adults( Mulla and Su 1999).
Acknowledgments
In mosquitoes, the gonotrophic events of femaleThese studies were supported by University of
Anopheles stephensi and An. culicifacies were impairedCalifornia Special Funds for Mosquito Control Research.
by neem exposures ( Dhar et al. 1996). In Ae. aegypti,significant transient retardation of oocyte growth was REFERENCES CITED
observed for up to 72 hours after feeding on 100 ng AZ/l blood meal. In a decapitation test, immature oocytes Abacus Concepts, Inc. 1987. StatView SE+ Graphics.
were observed in 86% of AZ-fed females ( 100 ng/µ l Abacus Concepts, Inc., Berkeley, CA, 234 pp.

December, 1999 Journal of Vector Ecology 215
Ascher, K. R. S., 1993. Nonconventional insecticidal neem extracts and some neem components, witheffects of pesticides available from the neem tree, reference to abnormalities and esterase activity inAzadirachta indica. Arch. Insect Biochem. Physiol. insect. Pp. 315- 330 in Proc. 3rd Int. Neem Conf.22: 433- 449. Nairobi, Kenya) ( H. Schmutterer and K. R. S.
Azambuja, P., E. S. Garcia, N. A. Ratcliffe, and J. D. Ascher, eds.). GTZ, Eschborn, Germany, 703 pp.Warthen Jr. 1991. Immune- depression in Rhodnius Rembold, H. 1995. Growth and metamorphosis. Pp.prolixus induced by the growth inhibitor, 174- 194 in The Neem Tree Azadirachta indica A.azadirachtin. J. Insect Physiol. 37: 771- 778. Juss and Other Meliaceous Plants: Sources of
Dhar, R., H. Dawar, S. Garg, S. F. Basir, and G. P. Unique Natural Products for Integrated PestTalwar. 1996. Effects of volatiles from neem and Management, Medicine, Industry and OtherPurposeother natural products on gonotrophic cycle and H. Schmutterer, ed.). VCH, Weinheim, New York,oviposition of Anopheles stephensi and An. Basel, Cambridge, Tokyo, 696 pp.culicifacies. J. Med. Entomol. 33: 195- 201. Schmutterer, H. 1990. Properties and potential of
Dorn, A., J. M. Rademacher, and E. Sehn. 1987. Effects natural pesticides from the neem tree, Azadirachtaof azadirachtin on reproductive organs and fertility indica. Ann. Rev. Entomol. 35: 271- 297.in the large milkweed bug, Oncopeltus fasciatus. Schmutterer, H. and H. Rembold. 1995. Reproduction.Pp. 273- 288 in Proc. 3rd Int. Neem Conf.( Nairobi, Pp. 195- 204 in The Neem Tree Azadirachta indicaKenya)( H. Schmutterer and K. R. S. Ascher, eds.). A. Juss and Other Meliaceous Plants: Sources ofGTZ, Eschborn, Germany, 703 pp. Unique Natural Products for Integrated Pest
Finney, D. J. 1971. Probit Analysis( Third Edition). Pp. Management, Medicine, Industry and Other Purpose20-49. Cambridge University Press, Cambridge, H. Schmutterer, ed.), VCH, Weinheim, New York,London, New York, New Rochelle, Melbourne, Basel, Cambridge, Tokyo, 696 pp.Sydney, 333 pp. Shapiro, J. P. and H. H. Hagedorn. 1982. Juvenile
Flanagan, T. R. and H. H. Hagedorn. 1977. Vitellogenin hormone and the development of ovariansynthesis in the mosquito: the role of juvenile responsiveness to a brain hormone in the mosquito,hormone in the development of responsiveness to Aedes aegypti. Gen. Comp. Endocrinol. 46: 176-ecdysone. Physiol. Entomol. 2: 173- 178. 183.
Immaraju, J. 1998. The commercial use of azadirachtin Shimizu, T. 1988. Suppressive effects of azadirachtinand its integration into viable pest control on spermiogenesis of the diapausing cabbageprogrammes. Pestic. Sci. 54: 285- 289. armyworm, Mamestra brassicae, in vitro. Entomol.
Kelly, T. J., T. S. Adams, M. B. Schwartz, M. J. Exp. Appl. 46: 197- 199.Birnbaum, E. C. Rubenstein, and R. B. Imberski. Sukumar, K., M. J. Perich, and L. R. Boombar. 1991.1987. Juvenile hormone and ovarian maturation in Botanical derivatives in mosquito control: areview.the Diptera: a review of recent results. Insect J. Am. Mosq. Contr. Assoc. 7: 210- 237.Biochem. 17: 1089- 1093. Su, T. and M. S. Mulla. 1998a. Antifeedancy of neem
Klowden, M. J. 1997. Endocrine aspects of mosquito products ( azadirachtin) against the mosquitoesreproduction. Arch. Insect Biochem. Physiol. 35: CulextarsalisandCulexquinquefasciatus( Diptera:491- 512. Culicidae). J. Vector Ecol. 23: 114- 122.
Ludlum, C. T. and K. P. Sieber, 1988. Effects of Su, T.and M. S. Mulla. 1998b. Ovicidal activity ofneemazadirachtin on oogenesis in Aedes aegypti. Physiol. products( azadirachtin) on Culex tarsalis and CulexEntomol. 13: 177- 184.
quinquefasciatus( Diptera: Culicidae). J. Am. Mosq.Meola, R. and J. Readio. 1988. Juvenile hormone Contr. Assoc. 14: 204- 209.
regulation of biting behavior and egg development Takken, W., M. J. Klowden, and G. M. Chambers. 1998.in mosquitoes. Pp. 1- 24 in Adv. Disease Vector Effect of body size on host seeking and blood mealRes. Vol. 5 ( K. F. Harris, ed.). Springer-Verlag, utilization in Anopheles gambiae sensu strictoNew York, 300 pp. Diptera: Culicidae): The disadvantage of being
Mordue, A. J. and A. Blackwell. 1993. Azadirachtin: an small. J. Med. Entomol. 35: 639- 645.update. J. Insect Physiol. 39: 903- 924. Wilps, H. 1989. The influence of neem seed kernel
Mulla M. S. and T. Su. 1999. Activity and biological extracts ( NSKE) from the neem tree Azadirachtaeffects of neem products against insect of medical indica on flight activity, food ingestion, reproductiveand veterinary importance. J. Am. Mosq. Contr. rate and carbohydrate metabolism in the DipteraAssoc. 15: 133- 152. Phormia terraenovae ( Diptera, Muscidae). Zool.
Naqvi, S. N. H. 1987. Biological evaluation of fresh Jb. Physiol. 93: 271- 282.

Journal of Vector Ecology 24( 2): 216- 223
Compatibility of Bacillus thuringiensis serovar israelensisand Chemical Insecticides for the Control ofAedes Mosquitoes
P. Seleena, H. L. Lee, and Y. F. Chiang
Division of Medical Entomology, Institute for Medical Research,Jalan Pahang, 50588 Kuala Lumpur, MALAYSIA
Received 13 April 1999; Accepted 28 September 1999
ABSTRACT: The compatibility of the commercial aqueous Bacillus thuringiensis serovar israelensisB. t.i.) formulation, Vectobac 12AS®, with the chemical insecticides Actellic 50EC®, Aqua Resigen®,
Resigen®, and Fendona SC®, for the simultaneous control of Aedes larvae and adults was studied by
dispersing nine different formulations using a portable mist blower, in single story half-brick houses. Theeffectiveness of the treatment was evaluated by measuring the larval mortality, adult mortality, and dropletanalysis at varying distances from the sprayer. Persistence of the larvicidal activity of the chemicalinsecticides and B. t. i was also determined by measuring the larval mortality in the test samples 7 daysposttreatment. The sprayed particles in all the trials were 50- 60 pm in size, indicating that the particles werethose of mist spray. Test samples placed within 3 m from the sprayer gave the maximum larval and adultmortality. Chemical insecticides exhibited maximum larval mortality in the 1 h posttreatment test samplesand it was comparable to the larvicidal activity of B. t.i. The larvicidal toxins of B.t. i were more stable andwere able to affect sufficient larval mortality for 7 days posttreatment. The larvicidal activity of themixtures, i.e., chemical insecticides with B.t.i, in the 1 h posttreatment test samples was not significantly
different from the larvicidal activity of the chemical insecticides and it was comparable to the larvicidalactivity of B.t.i alone. However, the larvicidal activity of the mixtures was significantly more than thechemical insecticides alone in the 7 days posttreatment test samples except for the Actellic 50EC® and
Vectobac 12AS®mixture. In all the trials, with or without B. t.i, there was no significant difference in adult
mortality, indicating that this B. t.i formulation, Vectobac 12AS®, was not antagonistic to the adulticidal
activity of the chemical insecticides. From this study, it can be concluded that chemical insecticides canbe used effectively for both adult and larval control, but the chemical insecticides do not exhibit residuallarvicidal activity. Hence, for an effective control of both Aedes larvae and adults, it is advisable to addB. t. i to the chemical insecticides, as B. t. i is specifically larvicidal and is also able to effect extended residuallarvicidal activity.
Keyword Index: Aedes, control, Bacillus thuringiensis serovar israelensis, chemical insecticides.
INTRODUCTION The chemical is dispersed by either ulta-low- volumeULV) or thermal fogging, and repeat spraying is carried
In Malaysia, dengue fever ( DF) and dengue out 7- 10 days after the first treatment. Residents in the
haemorrhagic fever ( DHF) continue to pose a serious affected areas are also encouraged to apply Abate®public health problem. The two common vector species ( temephos) in all water- storing containers as larvicidesareAedes aegypti( Linn.) and Aedes albopictus( Skuse), ( Tham 1997).
which are largely indoor and outdoor container breeders, There has been an increase in the number ofreported
respectively. Attempts to control dengue currently dengue cases since 1989, with 24,681 reported cases in
include anti- larval and anti- adult activities together 1998 ( Tham 1998). The dengue problem has been
with health education and community participation. perpetuated due to inadequate public compliance towards
Operations against adult mosquitoes involve treatment prevention and control of Aedes breeding sites and thewith malathion of outbreak areas within a radius of lack of manpower to conduct a complete inspection of
1 km from where a suspected dengue case is reported. the breeding sites.

December, 1999 Journal of Vector Ecology 217
An effective and efficient control of Aedes Spraying Trialsmosquitoes can be achieved if the health authorities are A portable mist- blower, Mist Blower MD 300,able to implement a system that can control both adults MaruyamaMfg. Co., Inc.TM with ULV attachments wasand larvae, simultaneously in the same operation. used to disperse the insecticides. The discharge rateBacillus thuringiensis serovar israelensis ( B. t.i) is an of the insecticides was maintained at 60 ml per minuteeffective microbial control agent against mosquito larvae by setting the volume switch dial at 3. The throttle leverthat is highly specific and toxic to mosquito larvae on was maintained at 4 during spraying operations.ingestion. An effective usage of B. t.i in a mosquito The spraying trials were conducted on nine differentvector control system would be in combination with days, dispersing nine different formulations. Thechemical insecticides that are being predominantly used insecticides were mixed at the field site prior to theiras adulticides. This paper reports on the compatibility application. The operator of the sprayer stood at theof dispersing B. t. i simultaneously with chemical front door of the house, pointing and rotating the nozzleinsecticides for the control of both Aedes larvae and towards its interior. The effectiveness of each trialadults.
was evaluated by measuring three different parameters:larval mortality, adult mortality, and ULV droplet
MATERIALS AND METHODS analysis.
The field studies were conducted in three unoccupied Larval Mortalitysingle story half-brick houses. Each house was 4. 5 m x In trials 1- 9 the larval mortality was measured by10.6 m x 3. 0 m( width x length x height) in size. The area placing two sets of cups, containing 200 ml distilledand volume of each house was 47.7 m2
and 143. 1 m3,
water, in each section of the house. Both sets wererespectively. To determine the effectiveness of the collected an hour after spraying. To one set of the cups,treatment in relation to the distance from the sprayer, 20 laboratory bred Ae. aegypti larvae ( L3/ L4) wereeach house was divided into three sections: 3 m, 6 m, and added per cup. The larval mortality was scored 249 m from the sprayer. h and 48 h after exposure. The second set of the
cups was left at ambient temperature ( 28- 30°C) andInsecticides relative humidity of about 85% for 7 days to determine
The trials were conducted using the following the persistency of the insecticides in the test waterinsecticides: a commercial aqueous suspension of B. t.i 7 days posttreatment. On the 7® day, 20 laboratory bredformulation, Vectobac 12AS® ( Abbott Laboratories) Ae. aegypti larvae ( L3/L4) were added into the cup.containing 1200 ITU/mg against Ae. aegypti larvae; The larval mortality was scored 24 h and 48 h afterActellic 50EC° ( Zeneca, UK) containing pirimiphos
exposure. Two additional cups per test containingmethyl 48. 1%( w/w); Aqua Resigen°( AgrEvo Environ distilled water were used in the laboratory as controlmental Health) containing s- bioallethrin 0. 14%( w/v),
replicates. The statistical significance of the larvalt-permethrin 10.26%( w/v), and piperonyl butoxide 9. 79% mortality among trials was analyzed using a Student t
w/ v); Resigen® ( AgrEvo Environmental Health) test.
containing s- bioallethrin 0. 8%( w/ w), permethrin 18. 7% Adult Mortalityw/w), and
SC®
butoxide 16. 8% ( w/ w); and
In trial Nos. 2- 9, the adult mortality was determinedFendona SC ( Cyanamid) containing alphacyper- by placing cages of 25 sucrose- fed, < 7 days old,
methrin 1. 47%( w/ w). The dosages of the insecticideslaboratory reared adult females of Ae. aegypti in each
used in the trials were based on the manufacturers' section of the house. For trial Nos. 8 and 9, an hour after
recommendations for ULV indoor application, i.e., spraying the treated mosquitoes were transferred into
Vectobac 12AS®- 500 ml/ha; Aqua Resigen®- 1: 10paper cups and fed with 10% sucrose solution in a
mixture, 100 ml mixture/2000 m3; Resigen°- 1: 10 cotton pad. Adult mortality was recorded 24 hmixture, 500 ml mixture/2000 m3; Fendona SC® 1: 10 posttreatment. Resigen®, Aqua Resigen®, and Fendonamixture, 1000 ml mixture/2000 m3;
and Actellic 50EC° SC®arepyrethroids known to cause rapid knockdown of500 ml/ ha. The chemical tank of the sprayer requires mosquitoes ( Elliott et al. 1978). Therefore, for trial
aminimum volume of2000 ml, so all formulations were Nos. 2- 7, knockdown ofadult mosquitoes was recordedprepared to a total volume of 2000 or 2200 ml. In each 5 min, 1 h, and 3 h post fogging. All the mosquitoes weretrial the spraying time( seconds/ house) was determined, then transferred into paper cups and were fed 10%ensuring that the recommended dosage of the insecticides sucrose solution in a cotton pad. Adult mortality waswas achieved in each house. recorded 24 h post spraying.

218 Journal of Vector Ecology December, 1999
Droplet Analysis m from the sprayer, thus, indicating the breakdown ofThe distribution and size of sprayed particles were sprayed Aqua Resigen® within 7 days of fogging. In
monitored through the use of magnesium oxide( MgO) comparison to trial 1, there is no significant difference
coated slides. A slide was placed amidst the cups holding in the larval mortality for 1 h posttreatment test samplesthe larvae. Droplet diameter was measured for an average ( p> 0. 05), but the larval mortality was significantly lessof 30 droplets for each MgO coated slide using a in the 7 days posttreatment test samples( p< 0.05). In
calibrated micrometer. The data were analyzed using trial 2, complete adult mortality was observed in all thethe ULV droplet analysis program of Sofield and Kent cages within 3 h post spraying.1984). In any spray the droplets are divided into two
equal parts by volume. Droplets with large volume areTrial 3: Vectobac 12AS®and Aqua Resigen®Mixture
represented by volume median diameter ( vmd) whileThe vmd and nmd of the sprayed particles was 62. 8
the small volume droplets are represented by number ± 7. 0 pm and 48. 2± 8. 5 pm, respectively, with a ratio of
median diameter ( nmd). A uniform size of droplets, 1. 3. Vectobac 12AS®, and Aqua Resigen® mixture
when the ratio of vmd to nmd is near to 1, is preferred for provided 100% larval mortality in 1 h post spray for testan efficient spray( Mount 1985). samples placed within 3 m from the sprayer( 24 and 48
h exposure). In the test samples placed beyond 3 m, a
RESULTS larval mortality of 48. 3- 70. 0% was obtained on 24 h
exposure and this was further increased to 66. 7- 86.7%
The droplet analysis and larval and adult mortality on 48 h exposure. In this trial the knockdown and
for each trial are expressed as the arithmetic mean of the recovery of larvae was not observed in the 1 hsamples from the treated sites at each distance. The posttreatment samples as in trial 2. This could be due to
larval and adult mortality results are shown together the presence ofB. t.i. toxins that are specifically larvicidal.
with the standard deviation for the mean in TABLES 1 As for the 7 days posttreatment samples, larval mortality
and 2, respectively. was observed in all the test samples, 25. 0- 98. 0% and
30. 0- 100.0% mortality on 24 and 48 h exposure,Trial 1: Commercial Aqueous B.t.i Formulation respectively. In comparison to trial 1, there is no
Vectobac 12AS®) significant difference in the larval mortality for 1 h
The vmd and nmd of the sprayed particles was 57.9 posttreatment test samples ( p > 0.05) but it was
4. 1 pm and 45. 6± 2. 1 pm, respectively, with a ratio of significantly less in the 7 days posttreatment test samples
1. 3. Vectobac 12AS® achieved a 86. 7- 95. 6% and 95. 6- ( p< 0.05). In comparison to trial 2, there is no significant100% mortality for Ae. aegypti larvae in the 1 h post difference in the larval mortality for 1 h posttreatment
treatment samples for 24 h and 48 h exposure, test samples( p> 0. 05), but it was significantly more in
respectively. A larval mortality of 100% was achieved the 7 days posttreatment test samples( p< 0. 05).
in the 7 days posttreatment samples ( 24 h and 48 h In trial 3, complete adult mortality was observed in
exposure). all the cages within 3 h of spraying, thus, indicating thatVectobac 12AS® does not have any adverse effects on
Trial 2: Aqua Resigen® the adulticidal activity of Aqua Resigen®.The vmd and nmd of the sprayed particles was 51. 1
5. 4 pm and 32.2± 5. 1 pm, respectively, with a ratio of Trial 4: Resigen®
1. 6. Aqua Resigen® achieved 100% larval mortality in The vmd and nmd of the sprayed particles was
1 h posttreatment test samples placed within 3 m from 58. 1± 3. 8 pm and 42.6± 3. 5 pm, respectively, with a
the sprayer( 24 and 48 h exposure). In the test samples ratio of 1. 4. Resigen® achieved 100% larval mortality
placed beyond 3 m, 55- 60% larval mortality was obtained in 1 h posttreatment test samples placed within 3 m from
on 24 h exposure and this was further reduced to 41. 7- the sprayer ( 24 h and 48 h exposure). In test samples
46.7% on 48 h exposure. This could be because Aqua placed beyond 3 m, 41. 7- 70.0% larval mortality was
Resigen®, as a pyrethroid, initially caused a rapid larval obtained for 24 h exposure and this was further reduced
knockdown during the first 24 h exposure but wasto 40.0- 65. 0% on 48 h exposure. This could be for the
insufficient to cause larval mortality. Thus, the larvae same reasons as for the pyrethroid Aqua Resigen®
recovered within the next 24 h, reducing the larval formulation. As for the 7 days posttreatment, a very low
mortality( 48 h exposure). In the 7 days posttreatment larval mortality was observed in samples placed withintest samples, larval mortality was only observed in 3 m from the sprayer. Larval mortality was not observedsamples placed within 3 m from the sprayer. Larval in samples placed beyond 3m from the sprayer, indicating
mortality was not observed in samples placed beyond 3 the breakdown of sprayed Resigen® within 7 days of

December, 1999 Journal of Vector Ecology 219
TABLE 1. Percent larval mortality expressed as the arithmetic mean of the samplesfrom the treated sites at each distance.
Distance From 1 Hour Post Spray 7 Days Post SpraySprayer ( m) 24 Hours 48 Hours 24 Hours 48 Hours
Trial 1: Insecticide Formulation: Vectobac 12AS®
3 95. 6± 7. 7 100± 0 100± 0 100± 06 86.7± 17. 6 95. 6± 7.7 100± 0 100± 09 95. 6± 7. 7 100± 0 100± 0 100± 0
Trial 2: Insecticide Formulation: Aqua Resigen®
3 100± 0 100± 0 43. 3± 40.4 46.7± 41. 66 60.0± 52.9 41. 7± 52.0 0 09 55.0± 50.8 46. 7± 50.3 0 0
Trial 3: Insecticide Formulation: Aqua Resigen®+ Vectobac 12AS®
3 100± 0 100± 0 98. 3± 2.9 100± 06 48. 3± 20. 2 66.7± 28. 9 25. 0± 21. 8 30.0± 17. 39 70.0± 5. 0 86.7± 15. 3 70.0± 21. 8 98. 3± 2. 9
Trial 4: Insecticide Formulation: Resigen®
3 100± 0 100± 0 15. 0± 15. 0 15. 0 ± 15. 06 41. 7± 28. 9 40.0± 26.0 0 0
9 70.0± 18. 0 65.0 ± 13. 2 0 0
Trial 5: Insecticide Formulation: Resigen® + Vectobac 12AS®
3 100± 0 100± 0 95.0± 8. 7 100± 0
6 100± 0 100± 0 67.7± 41. 6 88. 3± 20. 29 100± 0 100± 0 73. 3± 46. 2 88. 3± 20. 2
Trial 6: Insecticide Formulation: Fendona SC®
3 93. 3± 11. 6 96.7± 5. 8 73. 3± 5. 8 75. 0± 8. 7
6 80.0± 17. 3 90.0± 10. 0 48. 3± 7. 6 58. 3± 12. 6
9 91. 7± 7. 6 91. 7± 14. 4 33. 3± 7.6 61. 7± 7. 6
Trial 7: Insecticide Formulation: Fendona SC® + Vectobac 12AS®
3 100± 0 100± 0 96. 7± 5. 8 96.7± 5. 8
6 63. 3± 12. 6 83. 3± 2.9 86. 7± 11. 6 95.0± 5. 0
9 70.0± 18. 0 83. 3± 20. 8 87. 5± 10. 6 90.0± 14. 1
Trial 8: Insecticide Formulation: Actellic 50EC®
3 100± 0 100± 0 96. 7± 5. 8 100 ± 0
6 48. 3± 41. 9 61. 7± 53. 9 15. 0± 15. 0 23. 3± 7. 6
9 21. 7± 18. 9 30.0± 30.0 6. 7± 11. 6 23. 3± 5. 8
Trial 9: Insecticide Formulation: Actellic 50EC® + Vectobac 12AS®
3 100± 0 100± 0 100± 0 100± 0
6 68. 3± 7. 6 91. 7± 7. 6 10.0± 17. 3 31. 7± 7. 6
9 56. 7± 15. 3 90.0± 17. 3 0± 0 6. 7± 11. 6
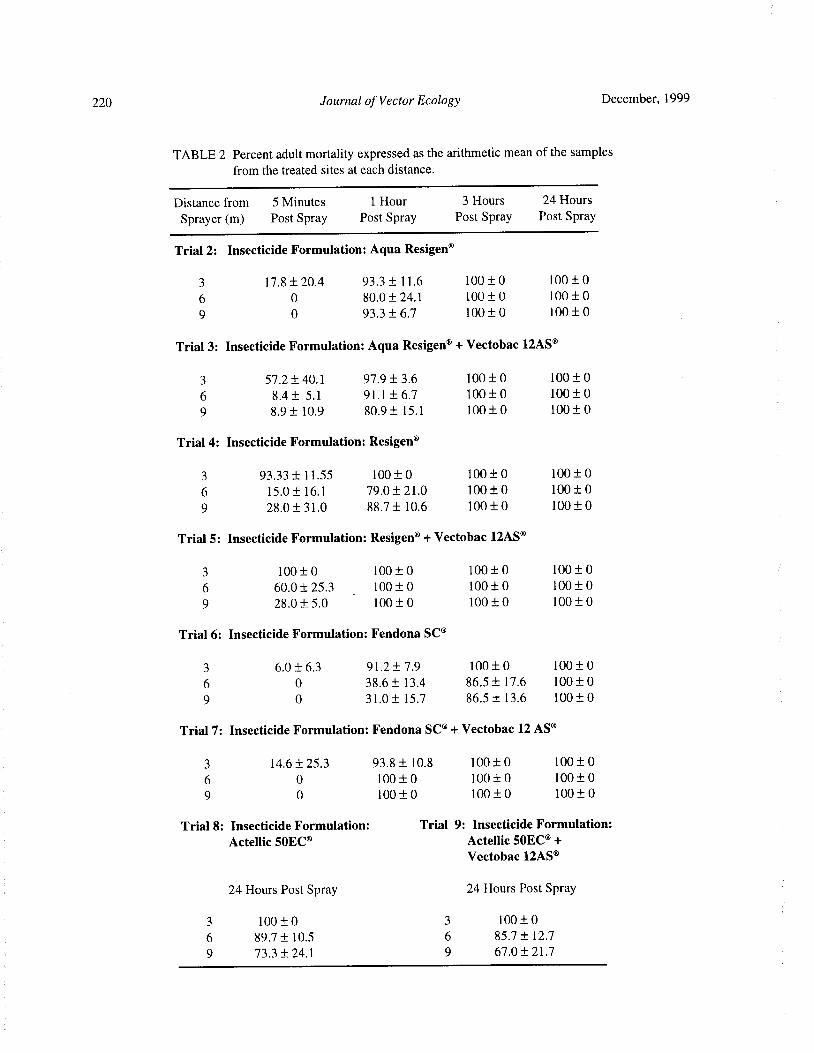
220 Journal of Vector Ecology December, 1999
TABLE 2 Percent adult mortality expressed as the arithmetic mean of the samplesfrom the treated sites at each distance.
Distance from 5 Minutes 1 Hour 3 Hours 24 Hours
Sprayer( m) Post Spray Post Spray Post Spray Post Spray
Trial 2: Insecticide Formulation: Aqua Resigen®
3 17. 8± 20.4 93. 3± 11. 6 100± 0 100± 0
6 0 80.0± 24. 1 100± 0 100± 0
9 0 93. 3± 6. 7 100± 0 100± 0
Trial 3: Insecticide Formulation: Aqua Resigen°+ Vectobac 12AS°
3 57. 2± 40. 1 97. 9± 3. 6 100± 0 100± 0
6 8. 4± 5. 1 91. 1 ± 6.7 100± 0 100± 0
9 8. 9± 10. 9 80.9± 15. 1 100± 0 100± 0
Trial 4: Insecticide Formulation: Resigen°
3 93. 33± 11. 55 100± 0 100± 0 100± 0
6 15. 0± 16. 1 79. 0± 21. 0 100± 0 100± 0
9 28. 0± 31. 0 88. 7± 10.6 100± 0 100± 0
Trial 5: Insecticide Formulation: Resigen°+ Vectobac 12AS®
3 100± 0 100± 0 100± 0 100± 0
6 60.0± 25. 3 100± 0 100± 0 100± 0
9 28. 0± 5. 0 100± 0 100± 0 100± 0
Trial 6: Insecticide Formulation: Fendona SC°
3 6. 0± 6.3 91. 2± 7. 9 100± 0 100± 0
6 0 38. 6± 13. 4 86. 5± 17. 6 100± 0
9 0 31. 0± 15. 7 86. 5± 13. 6 100± 0
Trial 7: Insecticide Formulation: Fendona SC°+ Vectobac 12 AS°
3 14. 6± 25. 3 93. 8± 10. 8 100± 0 100± 0
6 0 100± 0 100± 0 100± 0
9 0 100± 0 100± 0 100± 0
Trial 8: Insecticide Formulation: Trial 9: Insecticide Formulation:
Actellic 50EC® Actellic 50EC®+
Vectobac 12AS®
24 Hours Post Spray 24 Hours Post Spray
3 100± 0 3 100± 0
6 89. 7± 10. 5 6 85. 7± 12. 7
9 73. 3± 24. 1 9 67. 0± 21. 7

December, 1999 Journal of Vector Ecology 221
spraying. In comparison to trial l, there was no significant of 1. 4. Vectobac 12AS® and Fendona SC® mixturedifference in the larval mortality for 1 h posttreatment achieved 100% larval mortality in the 1 h post treat-test samples ( p > 0.05), but the larval mortality was ment test samples placed within 3 m from the sprayer
significantly less in the 7 days posttreatment test samples ( 24 and 48 h exposure). In the samples placed
p < 0. 05). In this trial 100% adult mortality was beyond 3 m a larval mortality of 63. 3- 70. 0% and 83. 3%observed in all the cages within 3 h of spraying. was achieved for 24 h and 48 h exposure, respectively.
In the 7 days posttreatment test samples, a larvalTrial 5: Vectobac 12AS® and Resigen® Mixture mortality of 86.7- 96.7% and 90.0- 96.7% was observed
The vmd and nmd of the sprayed particles was 58. 3 for 24 and 48 h exposure, respectively. In comparison12. 2 pm and 36.2± 17. 1 pm, respectively, with a ratio to trial 1, there is no significant difference in the
of 1. 6. Vectobac 12AS®and Resigen®mixture produced larval mortality in 1 h and 7 days posttreatment ( p >100% larval mortality in 1 h posttreatment test samples 0. 05). In comparison to trial 6, there is no significantplaced within 9 m from the sprayer ( 24 h and 48 h difference in the larval mortality at 1 h posttreatment( pexposure). In this trial the knockdown and recovery of > 0.05), but it was significantly more than at 7 dayslarvae was not observed in the 1 h posttreatment samples posttreatment( p< 0. 05).
as in trial 4, and this could be due to the presence ofB. t.i. In this trial a 100% adult mortality was observed intoxins, which are specifically larvicidal. As for the 7 all the cages within 3 h of spraying, indicating thatdays posttreatment test samples, larval mortality was Vectobac 12AS® does not have any adverse effects onobserved in all the test samples, 67.7- 95. 0% and 88. 3- the adulticidal activity of Fendona SC®.100% mortality on 24 and 48 h exposure, respectively.In comparison to trial 1, there was no significant Trial 8: Actellic 50EC®
difference in the larval mortality at 1 h and 7 days The vmd and nmd of the sprayed particles was 54. 3posttreatment test samples( p > 0.05). In comparison to ± 6.4 pm and 42. 5± 3. 1 pm, respectively, with a ratio oftrial 4, the larval mortality was significantly greater in 1. 3. In the 1 h posttreatment test samples, Actellicthe 1 h and 7 days posttreatment test samples ( p< 0.05). 50EC® produced a 100% larval mortality in samples
In trial 5, 100% adult mortality was observed in all placed 3 m from the sprayer( 24 h and 48 h exposure).the cages within 1 h of posttreatment, thus, indicating However, in samples placed beyond 3 m it only achievedthat Vectobac 12AS® does not have any adverse effects a 21. 7- 48. 3% and 30.0- 61. 7% larval mortality on 24 andon the adulticidal activity of Resigen®. 48 h exposure, respectively. This larval mortality was
further reduced in the 7 days posttreatment test samples,
Trial 6: Fendona SC° thus indicating the breakdown ofActellic 50EC° withinThe vmd and nmd of the sprayed particles was 56. 6 7 days of spraying. In comparison to trial 1, there is no
1. 2 pm and 43. 8± 4.4 pm, respectively, with a ratio of significant difference in the larval mortality for 1 h1. 3. In the 1 h posttreatment test samples, Fendona SC° posttreatment test samples ( p > 0.05), but it was
produced 80.0- 93. 3% and 90.0-96.7% larval mortality significantly less in the 7 days posttreatment test sampleson 24 and 48 h exposure, respectively. This larval ( p< 0.05).
mortality was reduced in the 7 days posttreatment In trial 8, complete adult mortality was observedsamples, thus indicating the breakdown of sprayed within 24 h of posttreatment in the cages placed 3 m
Fendona SC° within 7 days of spraying. In comparison from the sprayer. In cages placed beyond 3 m, 73. 3-
to trial 1, there was no significant difference in the larval 89.7% adult mortality was observed.
mortality for 1 h posttreatment test samples( p> 0.05),
but it was significantly less in the 7 days posttreatment Trial9: Vectobac 12AS° and Actellic 50EC° Mixturetest samples ( p< 0.05). The vmd and nmd of the sprayed particles was 52. 5
In trial 6, complete adult mortality was observed, ± 5. 2 pm and 32. 3± 4. 3 pm, respectively, with a ratio ofwithin 3 h ofposttreatment, in the cages placed 3 m from 1. 6. Vectobac 12AS® and Actellic 50EC® mixturethe spraying machine. In cages placed beyond 3 m
produced 100% larval mortality in 1 h posttreatmentcomplete adult mortality was only observed within 24 h samples placed within 3 m from the sprayer( 24 and 48of posttreatment. h exposure). In the samples placed beyond 3 m, a larval
mortality of 56.7- 68. 3% and 90. 0- 91. 7% was achieved
Trial 7: Vectobac 12AS® and Fendona SC®Mixture for 24 h and 48 h exposure, respectively. In the 7 daysThe vmd and nmd of the sprayed particles was 55. 4 posttreatment samples, complete mortality was only
5. 2 pm and 40.5± 1. 1 pm, respectively, with a ratio observed in samples placed 3 m from the sprayer( 24 h

222 Journal of Vector Ecology December, 1999
and 48 h exposure). In the 7 days posttreatment samples samples except for the Actellic 50EC° and Vectobacthat were placed beyond 3 m from the sprayer, the larval 12AS® mixture( p< 0.05). The significant difference inmortality was reduced by more than 50% in comparison the larvicidal activity for the 7 days posttreatment samplesto 1 h posttreatment samples. In comparison to trial could be due to the larvicidal activity of the chemical1, there is no significant difference in the larval insecticides degrading over time, thus, effecting a farmortality for 1 h posttreatment samples( p> 0. 05), but lesser larval mortality than the formulations mixed withit was significantly less in the 7 days posttreatment Vectobac 12AS®. This also indicates that the larvicidalsamples ( p< 0. 05). In comparison to trial 8 there is no
activity of the mixtures in the 7 days posttreatmentsignificant difference in the larval mortality for 1 h and
samples was mainly contributed by Vectobac I2AS°.7 days posttreatment test samples( p> 0.05).
In the Actellic 50EC° and Vectobac 12AS° mixture, theIn trial 9, complete adult mortality was observed
within 24 h of posttreatment in the cages placed 3 mlarvicidal activity of the Vectobac 12AS® was not
from the sprayer. In cages placed beyond 3 m, 67. 0exhibited as in other mixtures and this could be due to
85. 7% adult mortality was observed. the antagonism of Actellic 50EC° to the larvicidal
toxins of Vectobac 12AS®.
DISCUSSION The larvicidal activity of the mixtures in the 1 hposttreatment samples was not significantly different
The portable mist- blower, equipped with the ULVfrom the larvicidal activity of Vectobac 12AS®( p >
attachment, successfully dispersed the nine different0.05). However, it was significantly less in the 7 days
formulations that were tested. All formulations flowedposttreatment samples of the Aqua Resigen® and
smoothly through the sprayer nozzle. The volume
median diameter( vmd) of the sprayed particles for allVectobac 12A5® mixture and Actellic 5OEC® and
the trials was 50- 60 pm. This indicated that the particlesVectobac 12AS® mixture ( p < 0.05), suggesting that
were not of ULV spray but of mist spray ( Matthews Aqua Resigen®and Actellic SOEC®might be antagonistic
1985). The number of sprayed particles decreased with to the larvicidal toxins of Vectobac 12AS® in the
increasing distance from the sprayer. Therefore, test mixtures. On the other hand, the larvicidal activity ofsamples placed within 3 m from the sprayer gave the Resigen® and Vectobac 12AS® mixture and Fendona
maximum larval and adult mortality. SC® and Vectobac 12AS® mixture was comparable to
The chemical insecticides tested in this study are the larvicidal toxins of Vectobac 12AS®in the 1 h and
generally used as adulticides. However, this study has 7 days posttreatment test samples.shown that these insecticides used at the manu-
Overall, the Resigen® and Vectobac 12AS® mix-facturer' s recommended dosage also exhibit mosquito
ture showed significantly better larvicidal activitylarvicidal activity. The larvicidal activity of the
than Aqua Resigen® and Vectobac 12AS®, Actellicinsecticides in the 1 h posttreatment samples is
50EC® and Vectobac I2AS®, and Fendona SC® andcomparable to the larvicidal activity of the commercialaqueous B. t. i. formulation, Vectobac 12AS®, as no
Vectobac 12AS° mixtures in the 1 h posttreatment test
significant difference was observed in the larval mortalitysamples( p< 0.05) . This could be because the amount
p > 0. 05). However, the larvicidal activity of theof active ingredient in Resigen® was 2. 6- 9. 1 times
chemical insecticides was significantly less than greater than in Aqua Resigen®, Actellic 50EC°, and
Vectobac I 2A5®in the 7 days posttreatment test samplesFendona SC®.
p < 0.05). This could be due to the degradation of In all the trials, with and without Vectobac 12AS®,
larvicidal activity in the chemical insecticides. The there was no significant difference in adult mortality( plarvicidal toxins ofVectobac 12AS° are more stable and > 0.05). Thus, it can be concluded that Vectobac 12AS®
are able to effect sufficient larval mortality for a duration is not antagonistic to the adulticidal activity of chemicalof 7 days posttreatment. insecticides. From this study it can be concluded that
All the chemical insecticides mixed homogenously chemical insecticides Actellic 50EC°, Aqua Resigen®,
with Vectobac I2AS®, thus flowing smoothly without Resigen®, and Fendona SC®can be used effectively forclogging the nozzle. The larvicidal activity of the both adult and larval control. However, these chemicalmixtures in the 1 h posttreatment samples was not insecticides do not have significant residual larvicidal
significantly different from the larvicidal activity of the activity and are thus a disadvantage in Aedes control, aschemical insecticides( p> 0.05). However, the larvicidal the second application in dengue endemic areas is
activity of the mixtures was significantly greater from carried out 7- 10 days after the first spraying ( Thamthe chemical insecticides in the 7 days post spray test 1997). Within this period, Aedes larvae can emerge and

December, 1999 Journal of Vector Ecology 223
transmit dengue virus. Therefore, for simultaneous ofpyrethroids in insect control. Ann Rev. Entomol.effective control of Aedes larvae and adults, it is thus 23: 443- 469.
advisable to add Vectobac 12AS© to the chemical Matthews, G. A. 1985. Pesticide application method.insecticides, as it is specifically larvicidal and also able ELBS ed., The Bath Press, UK, pp 57- 74.to give residual larvicidal activity for 7 days after Mount, G. A. 1985. Ultra-low-volume application ofspraying. insecticides for vector control. WHO Monograph.
Acknowledgments WHONBC/ 85. 919, 30 pages.
Sofield, R. K. and R. Kent. 1984. A basic program forWe thank the Director, Institute for Medical the analysis of ULV insecticide droplets. Mosq.
Research, Malaysia, for permission to publish. We are News 44: 73- 75.
grateful to Abbott Laboratories( M) Sdn Bhd, AgrEvo Tham, A. S. 1997. Status report of dengue fever and
Environmental Health( M) Sdn Bhd, CCM Biosciences dengue haemorrhagic fever in Malaysia, presentedM) Sdn Bhd, and Cyanamid Agricultural Products( M) at the SEAMIC workshop on dengue fever in
Sdn Bhd for providing the insecticides. Manila, Philippines, 9- 15 November.
Tham, A. S. 1998. A status report on " Dengue In
REFERENCES CITED Malaysia: Current issues/ limitations and new
initiatives/ approaches" presented at the DengueElliot, M., N. F. Janes, and C. Potter. 1978. The future Control Meeting, Sarawak, 3- 5 December 1998.

Journal of Vector Ecology 24( 2): 224-232
Ticks of South Carolina (Acari: Ixodoidea)
D. C. Williams', W. Wills2, L. A. Durden3, and E. W. Gray4
Cypress Gardens, 3030 Cypress Gardens Rd., Moncks Corner, SC 29461 USA.
215 Chandler Court, Columbia, SC 29210 USA.
Institute ofArthropodology and Parasitology, Georgia Southern University, Statesboro, GA 30460 USA.
4Department ofEntomology, University of Georgia, Athens, GA 30602 USA.
Received 30 April 1999; Accepted 17 September 1999
ABSTRACT: County and host records are reported for 19 species of ticks from South Carolina:Amblyomma americanum, Amblyomma maculatum, Amblyomma tuberculatum, Aponomma latum,
Boophilus annulatus, Boophilus microplus, Dermacentoralbipictus, Dermacentorvariablis, Haemaphysalis
leporispalustris, Ixodes affinis, Ixodes brunneus, Ixodes cookei, Ixodes marxi, Ixodes minor, Ixodes
scapularis, Ixodes texanus, Ixodes woodi, Rhipicephalus sanguineus, and Ornithodoros capensis. Ixodes
woodi is recorded from South Carolina for the first time. Boophilus annulatus and Boophilus microplus
probably no longer exist in South Carolina, and Aponomma latum is an exotic species that is not establishedin South Carolina. Brief notes follow each species.
Keyword Index: Ticks, South Carolina.
INTRODUCTION 3) The Sandhills are a narrow northeast to southwest
band of hilly sand topography situated between theAlthough several tick-borne diseases are endemic Piedmont and the Coastal Plain. The Sandhills are
to South Carolina, the tick fauna of this state is not well thought to be either beaches left over from the Cretaceousdocumented. Rocky Mountain spotted fever( RMSF), period some 130 million years ago or due to erosion ofcaused by Rickettsia rickettsii, has been an important the Appalachian Chain. 4) The Coastal Plain extendshuman infection in the state for many years. Cases of almost 200 km from the Sandhills to the Coastal Zone.RMSF actually increased in the 1970s( Burgdorfer et al. 5) The Coastal Zone is a narrow band approximately 161975, Loving et al. 1978). More recently Lyme disease km in width in which the saline and brackish water fromLD) and ehrlichiosis ( Eh) have been reported from the Atlantic Ocean influence the vegetation types. This
South Carolina( Schuman and Caldwell 1989, Rumpel diverse area provides a variety of soil types, climate,and Jones 1991, Hawkins 1995). Until recently, little vegetation, and habitats for animals. South Carolina isattention has been given to systematic study of the southern-most limit for some northern animals anddistribution, hosts, and ecology of these important the northern- most limit for some southern animalsarthropods in the state. Despite some recent work ( Thompson 1982).
Barton et al. 1992, Keirans et al. 1992, Tedders 1994, Tick records were obtained from veterinarians andClark et al 1996, Durden et al. 1997, Clark et al. 1998) the Orangeburg County Society for the Prevention ofthere remains a lacuna in the bionomics and distribution Cruelty to Animals ( SPCA) ( mostly in Orangeburgof ticks within the state. With the exception of Barton County), our own collections, published literature, andet al. ( 1992) all of the above mentioned work was collection records from the U.S. National Tick Collection
conducted in the coastal and piedmont areas of South housed at Georgia Southern University' s Institute ofCarolina ( Fig. 1). Much more data are required for ArthropodologyandParasitologyinStatesboro, Georgia.surveillance and management of tick- borne diseases. Tick species are listed alphabetically, followed by the
South Carolina is divided into five physiographic county or counties in which they occurred, the hostregions ( Fig. 1). 1) The Mountains are part of the species, and the accession numbers of South Carolina
Appalachian system and are only in the extreme specimens from the U.S. National Tick Collectionnorthwest corner. 2) The Piedmont extends in a band ( appended by the last 2 digits of the year of collection,161 km wide between the Mountains and the Sandhills. all made during the
20th
century). Finally, a summary of
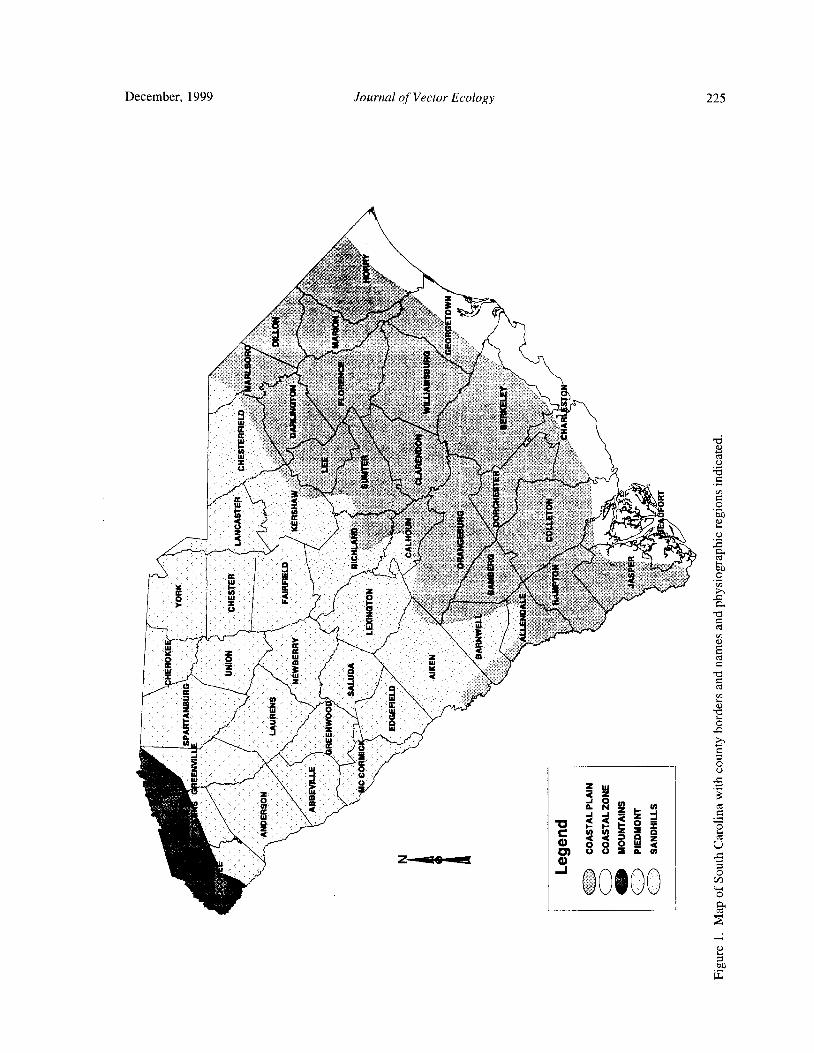
December, 1999 Journal of Vector Ecology 225
ae C
17xx i'l e
S%
QS{
r
or vbf y S' ii.3 F 3a 6 x
ff
Y3
4
l 6k f - h£
rte ;s
0 u : s. z'rs.
tbt
R
QW
k fix, R r
rL X '. Xb fl 0 T
Q Y pS f . i' 5) f
I. 5<• b E
X ' yz t tax 3"
z's
zz . ' O
0y f $
x y aW
q + qa .
5Ls
xr
6 bC
bhu z
a
F s3
tY 2
ifitiot
uiwhxRJ`.- 2, a .
y W aCd
W 2 m
o
Ro
Z Y
9 '• i.
Ii Y4 O—
O::
2C
W
c 0 ,' e 34n CI
W
Oi
d < I. o °
S
2
m GOIOO OS].ro
4.
bq
w

226 Journal of Vector Ecology December, 1999
published and personal observations about the species February, March, and December. Clark et al. ( 1998)and, where appropriate, a mention ofmedical importance collected nymphs from March through October andis included. larvae in March and from August through November.
Wilson and Baker ( 1972) listed 25 tick species Their collections were also concentrated in coastalfrom Georgia; however, 4 of these were recorded early counties.
and are not considered residents at this time. Durden Amblyomma americanum uses a variety ofwild andand Kollars ( 1992) listed 17 species from Tennessee. domestic animals including birds as hosts. AlthoughAlthough we have no list of ticks from North Carolina, this is certainly true in South Carolina as well; surveysLevine et al. ( 1989), while doing Lyme disease have concentrated on wild hosts and little information isinvestigations, reported 6 species from the state. available about infestations of domestic animals.
Here we report 18 species of ixodid ticks and 1 Numerous adult and immature A. americanum haveargasid tick from South Carolina as well as the first been observed on feral pigs by one of the authorsrecord of Ixodes woodi from this state and the first ( DW) at Hobcaw Barony, Georgetown County.record of Amblyomma maculatum taken from a coyote, Bishopp and Trembley( 1945) report that wild turkeysCanis latrans, in the southeast US. on Bull' s Island, Charleston County were observed to
be heavily infested with larvae and nymphs of thisAmblyomma americanum ( L.) . species.
COUNTY RECORDS: Beaufort, Charleston, All stages will attack humans ( Felz et al. 1996).Florence, Georgetown, Hampton, Horry, Orangeburg. Tedders ( 1994) indicated A. americanum as the tick
HOST RECORDS: Marsupialia - Didelphis most likely to be encountered by sportsmen, campers,virginiana. Carnivora - Canis lupus, Procyon lotor, and outdoor workers in the summer months in SC. TheMustela sp., Urocyon cinereoargentus. Primates - longmouthpartsofAmblyommaspp. makethisadifficultHomo sapiens. Artiodactyla- Odocoileus virginianus, tick to remove once embedded. Even without breakingBos spp. Rodentia- Sciurus niger. Avian spp.- Meleagris the mouth parts, a bite often results in a localgallopavo, Quiraca caerulea, Corvus brachyrhynchos, infection or irritation. Bolte et al. ( 1970) stated" ticksPorzana carolina. in the genera Amblyomma, Hyalomma, and Aponomma
ACCESSION NUMBERS: 122345- 94, 122308- penetrate host tissue during feeding to a greater depth94, 122307- 94, 122306- 94, 122242- 91, 060710- 31, than those of other genera. Sanders and Oliver( 1995)060714- 31, 060717, 060719- 31, 060720-36, 06229- 32, evaluated A. americanum, Dermacentor variablis, and057399- 31, 057401- 31, 057402- 34, 057404- 39, 057405- Ixodes scapularis from Georgia as vectors of a Florida39, 057275- 29, 057270- 31, 057277- 31, 057133- 29, strain ofBorrelia burgdorferi, the agent ofLyme disease.057341- 33, 057352- 33, 057398- 31, 057244- 31, 057295- They found A. americanum and D. variablis did not31, 057246- 31, 057247- 31, 057248- 31, 057255- 39, transmit this strain. On the other hand, I.scapularis was057256- 39, 057241- 29, 056836- 38, 021027- 44. an efficient vector. In Georgia, Lockhart et al. ( 1995)
REFERENCES: Barton et al. 1992, Felz et al. noticed a temporal association between A. americanum1996, Durden et al. 1997, Clark et al. 1996, Clark et al. and Ehrlichiachaffeensis, the agent ofhumanmonocytic1998, Tedders 1994. ehrlichiosis. Loving et al.( 1978) detected R. rickettsii
REMARKS: The lone star tick is abundant along in this tick in South Carolina. Burgdorfer et al.( 1975)the coastal areas of South Carolina and extends at least showed a higher R. rickettsii infection rate in thisas far west as Orangeburg County. It has also been taken species in South Carolina( 16. 6%) as compared to D.from deer in Edgefield and McCormick County( Barton variablis ( 4.9%). Amblyomma americanum also is aet al. 1992). The adults usually appear in April and competentvectorofFrancisellatularensis, the causativedisappear by July ( Clark et al. 1996, Tedders 1994), agent of tularemia( Hopla and Hopla 1994).
although Clark et al. 1998 reports collection of adultsfrom March to September with 1 male collected in Amblyomma maculatum Koch.
January, 1995. One adult was taken from a deer in COUNTY RECORDS: Allendale, Charleston,Georgetown County in November( Barton et al. 1992). Chester, Georgetown, Jasper, Newberry, Orangeburg,These records are similar to observations of this tick in Sumter.
Georgia where adults appeared in March and disappeared HOST RECORDS: Marsupialia - Didelphus
by August ( Davidson et al. 1994). Tedders collected virginiana. Carnivora - Canis lupus, Canis latrans,nymphal A. americanum during every month of the year Procyon lotor, Urocyon cinereoargenteus. Primate -from Georgetown and Charleston Counties, both coastal Homo sapiens. Artiodactyla- Odocoileus virginianus.counties. He collected larvae in all months except Rodentia- Sigmodon hispidus. Avian spp. - Agelaius

December, 1999 Journal of Vector Ecology 227
phoeniceus, Pipilo sp., Thryothorus ludovicianus. ACCESSION NUMBER: 115851- 11.ACCESSION NUMBERS: 122509- 47, 122346- REMARKS: The cattle fever tick has been
94, 122304-95, 122303- 95, 122305- 95, 057696- 33, eradicated from the U. S. James and Harwood ( 1970)057695- 31, 057654- 31, 057666- 33, 057650- 31, indicate that this species was the most economically057277- 31, 057248- 31, 057246- 31, 054244- 31, 031887- important species in the southern U.S. We have only49, 057632- 33, 057678- 36, 122785- 98. one record of this species from the U.S. National Tick
REFERENCES: Barton et al. 1992, Durden et al. Collection collected in Georgetown County.1997, Clark et al. 1996, Clark et al. 1998, Felz et al.1996, Tedders 1994. Boophilus microplus ( Canestrini)
REMARKS: South Carolina is probably the COUNTY RECORD: Georgetown.northern-most extension of the range of the Gulf Coast HOST RECORD: Artiodactyla- Cattle.tick. The adults appear in May and peak in June with a ACCESSION NUMBER: 057164- 11.few specimens appearing until mid-October ( Clark et REMARKS: We again have only one record of thisal. 1998). This species appears throughout the coastal tick from cattle in Georgetown County. Becklund
region of South Carolina and was the second most ( 1968) reported that this tick is frequently imported intocommon species collected in a study in Orangeburg the U. S. on horses from Cuba and Guatemala as well as
County ( Clark et al. 1996). Larvae and nymphs on refrigerated beef from Costa Rica and Honduras
commonly infest birds ( Bishopp and Trembley 1945). coming into Florida and on beef from Mexico comingThe adults are common on dogs and will readily attack into Ft. Worth, Texas. This species was also collectedman. Loving et al.( 1978) reported R. rickettsii from this from cattle hides from Cuba. Boophilus microplus is aspecies in South Carolina. problem in Australia where it is known, under severe
drought, to reproduce by parthenogenesis( Stone 1963).Amblyomma tuberculatum Marx. The South Carolina record is from a collection made in
COUNTY RECORD: Jasper. 1911.
HOST RECORD: Testudines - Gopherus
polyphemus. Dermacenter albipictus ( Packard).
ACCESSION NUMBER: 056913- 33. COUNTY RECORD: Abbeville, Beaufort,
REMARKS: The gopher tortoise tick probably Charleston, Chester, Chesterfield,, Edgefield, Fairfield,exists throughout the range of its host. In Georgia the Florence, Georgetown, Greenwood, Laurens,
larvae are found more frequently on mammals and birds McCormick, Newberry, Saluda, Spartanburg, Union.than reptiles ( Wilson and Baker 1972). Bishopp and HOST RECORD: Artiodactyla — Odocoileus
Trembley ( 1945) suggest that adults occur on the host virginianus.
throughout the year. We have one record from the U.S. ACCESSION NUMBER: 060254- 33, 057333- 29,
National Tick Collection ofA. tuberculatum taken from 060253- 33, 060192- 33.
a gopher tortoise in South Carolina. REFERENCE: Barton et al. 1992.
REMARKS: The winter tick is a one host tick. TheAponomma latum ( Koch). species is widespread throughout the U.S. Bishopp and
COUNTY RECORD: Florence. Trembley ( 1945) list horses, cattle, moose, and elk asHOST RECORD: Squamata( pet python). hosts. Although it probably occurs on white- tailed deerACCESSION NUMBER: 120941- 93. all over the state, Barton et al. ( 1992) did not collect it
REMARKS: The python tick is an exotic species on deer in the mountains of South Carolina. This could
occurring on African snakes and apparently is imported have been due to the time of year the deer were examined
frequently into the U.S. Durden and Kollars ( 1992) at deer check stations. Ixodes scapularis was collectedreported this species from a single specimen in Tennessee frequently with D. albipictus at check stations.and reported records from the U.S. National Tick
Collection from Connecticut, Illinois, Indiana, New Dermacenter variablis ( Say).Hampshire, New York, Pennsylvania, and Texas. One COUNTY RECORDS: Allendale, Barnwell,
female and I nymph were identified in this South Charleston, Chester, Georgetown, Lexington, Oconee,Carolina record from a pet snake kept in Lake City, SC. Orangeburg, Richland, Saluda, Spartanburg, Sumter.
HOST RECORDS: Marsupialia - Didelphus
Boophilus annulatus( Say). virginiana. Carnivora - Canis lupus, Procyon lotor,
COUNTY RECORD: Georgetown. Urocyon cinereoargenteus. Primate - Homo sapiens.HOST RECORD: Artiodactyla- Cattle. Rodentia- Rattus rattus, Sigmodon hispidus, Neotoma

228 Journal of Vector Ecology December, 1999
floridana, Oryzomyspalustris, Peromyscus gossypinus, erythrophthalmus, Cardinalis cardinalis, CatharusPeromyscus leucopus. ustulatus, Zonotrichia albicollis, Dendroica coronata.
ACCESSION NUMBERS: 122347- 94, 122311- ACCESSION NUMBERS: 056962- 33, 056959-
95, 122309- 95, 122310- 95, 120050-91, 119984-90, 31, 116916- 31.
064061- 33, 063287- 39, 063286-39, 063285- 39, 063284- REFERENCE: Durden et al. 1997.39, 063273- 39, 060449- 34, 059637- 15, 059634- 32, REMARKS: This species' common name is the059614- 14, 059431- 11, 057653- 31, 057650-31, 057256- rabbit tick. Adults are usually only found on rabbits,39, 057246- 31, 031888- 49. while immatures parasitize birds or rabbits. Durden et
REFERENCES: Clark et al. 1996 and 1998, Felz al. ( 1997) collected nymphs and larvae from severalet al. 1996, Loving et al. 1978, Tedders 1994. species of birds in coastal South Carolina. Although
REMARKS: The American dog tick is considered immatures will also parasitize small mammals, we havethe most important vector of R. rickettsii in the eastern no records of this in South Carolina. Strickland et al.U. S. ( Bishopp and Trembley 1945). Loving et al. ( 1977) suggested this species played an important role1978) studied RMSF in South Carolina. Their studies in maintaining RMSF, Q fever, and tularemia in wild
indicated D. variablis was the predominate tick found in mammal populations. It is probably more widespread inthe Piedmont region, 98% D. variablis to 2% of A. South Carolina than records indicate.
americanum. However, in the coastal region, collections
ofA. americanum were higher than those forD. variablis Ixodes affinis Neumann.
62% vs. 38%). Collections from the sandhills were COUNTY RECORDS: Berkeley, Charleston,about even for both species. Collections in Orangeburg Dorchester, Georgetown, Orangeburg, Sumter.County ( Clark et al. 1996) consisted of 259 adult D. HOST RECORDS: Carnivora - Canis lupus.
variablis collected from dogs and 8 from humans, Artiodactyla - Odocoileus virginianus. Rodentia -
compared to 14 A. americanum during the same time. Sigmodon hispidus.
Amblyomma americanum nymphs were the only ACCESSION NUMBERS: 039996-64, 122316-
immatures taken during the study. Clark et al. 1998 94, 122317- 94, 122315- 94, 122348- 94.found that Dermacentorvariablis adults usually appeared REFERENCES: Clark et al. 1998, Clark et al.
in early April and were almost all gone by September 1. 1996, Gerrish and Ossorio 1965.
Bishopp and Trembley ( 1945) list a large number REMARKS: This species was originally reportedof hosts for this tick. Lavender and Oliver( 1996) list 12 from Central and South America. It has been, so far,
host species from Bullock County, Georgia, and found restricted to the southeastern U.S. Gerrish and Ossorio
year round activity. Durden and Kollars( 1992) list 29 ( 1965) originally reported it from South Carolina onhosts from Tennessee. We have collected larvae of this deer in Dorchester County. Clark et al.( 1998) isolatedspecies in January from a cotton mouse, Peromyscus spirochetes that resembled Borrelia burgdorferi from!.gossypinus, in Barnwell County affinis collected in the sandhills and coastal zone of
One ofus( WW) observedamaledogataveterinary South Carolina. They believe that I. affinis is wellclinic in Orangeburg, South Carolina, that was suffering established in the coastal plain of South Carolina. Clarkfrom tick paralysis due to D. variablis. Forty-seven D. et al.( 1996) collected 1 male from a dog in Orangeburgvariablis were removed from the animal. It was given County. Oliver et al.( 1987) list 11 hosts for I.affinis isIV fluids and recovered after 2 days. Sanders and Oliver Georgia.
1995) showed D. variablis was unable to transmit a
Florida strain of B. burgdorferi. McLean et al. 1985 Ixodes brunneus Koch.
isolated St. Louis encephalitis virus from adult D. COUNTY RECORDS: Charleston, Cherokee,
variablis. This tick is the principal vector in the eastern Dorchester, Georgetown.
United States of R. rickettsii and is closely linked to HOST RECORDS: Avian- Zonotrichiaalbicollis,RMSF epidemiology in this region ( Felz and Durden sparrow( not identified to species), Pipilo sp., Toxostoma1998). rufum, Turdus migratorius, Spizella passerina,
Dumetella carolinensis, Hylocichla guttata, LaniusHaemaphysalis leporispalustris ( Packard). ludovicianus, Thryothorusludovicianus. Primate- Homo
COUNTY RECORDS: Charleston, Georgetown, sapiens.
McCormick. ACCESSION NUMBERS: 122351- 94, 120212-HOST RECORDS: Lagomorpha - Sylvilagus 91, 116901- 29, 116900- 29, 116899- 29, 058188- 34,
aquaticus, Sylvilaguspalustris. Avian- Colinus Virginia- 058186- 34, 058185- 33, 058183- 31, 058180- 31, 058179-nus, Parus bicolor, Thryothorus ludovicianus, Pipilo 31, 058178- 31, 058177- 31, 058176-30, 058175- 30,

December, 1999 Journal of Vector Ecology 229
058174- 30, 058172-31, 058171- 31, 058170- 24, 058165- HOST RECORDS: Rodentia - Rattus rattus,30, 057696- 33, 057695- 31. Sigmodon hispidus, Neotoma floridana, Peromyscus
REMARKS: This tick is distributed throughout gossypinus. Passeriformes - Cyanocitta cristata,most of the U.S. and is confined to birds as hosts. There Thryothorous ludovicianus, Pipilo sp., Pipilowill usually only be 1 tick attached to the host, although erythrophthalmus.
if the tick is an adult female this is sufficient to cause the ACCESSION NUMBERS: 119984- 90, 057696-death of the host( Bishopp and Trembley 1945). One of 33, 057632-33, 120050-91, 120004- 90, 122313- 95,the authors ( DW) captured a nearly flightless white- 122312- 95, 121717- 94, 121641- 94.throated sparrow, Zonotrichia albicollis, with an REFERENCES: Tedders et al. 1992, Teddersengorged female I. Brunneus below one eye. The bird 1994, Durden et al. 1997.
recovered its ability to fly overnight after the tick was REMARKS: Georgia was thought to be theremoved the previous afternoon. Rickettsia rickettsii northern- most limit for 1. minor. However, Tedders ethas been recorded from this tick in nature( Clifford et al. al. ( 1992) reported this species from coastal South1969) raising the possibility of the spread of this pathogen Carolina on roof rats. Ixodes minor was not thought toover long distances via the bird host. be a species of public health importance. Recently
Durden et al.( 1997) isolated B. burgdorferi from a largeIxodes cookei Packard number of this species suggesting it could be an important
COUNTY RECORDS: Charleston, Oconee, enzootic vector of the agent of Lyme disease since itPickens, York. feeds on rodents and birds. Clark et al.( 1998) collected
HOST RECORDS: Marsupialia - Didelphis 1. minor from cotton rats, cotton mice, and eastern woodvirginiana. Carnivora- Procyon lotor,Spilogaleputoris, rats from July through December in Georgetown County,Mustella vison. South Carolina. We have one record from a blue jay in
ACCESSION NUMBERS: 120580-91, 064950- Oconee County; because I. minor is a coastal tick, this33, 057237- 38, 122784- 98. specimen presumably was taken to this inland county by
REFERENCE: Tedders 1994. the bird after attaching to it in a coastal area.REMARKS: South Carolina records for I. cookei
occur from the coast and the mountains and, therefore, Ixodes scapularis Sayit probably also occurs in the areas in between. Medium- COUNTY RECORDS: Abbeville, Aiken,
sized mammals appear to be the main hosts for this Allendale, Anderson, Barnwell, Calhoun, Charleston,species. Tedders( 1994) found this tick to be relatively Chester, Chesterfield, Dorchester, Edgefield, Fairfield,rare with 2 females and 16 nymphs collected from Florence, Georgetown, Greenville, Greenwood,raccoons and 1 nymph from an opossum out of a total of Hampton, Laurens, McCormick, Newberry, Pickens,3, 797 ticks collected during his study. Powasson virus, Richland, Saluda, Spartanburg, Union.a serious human pathogen, has been isolated from this HOST RECORDS: Marsupialia - Didelphis
species in Canada and the northeast U. S.( Berge 1975). virginiana. Carnivora - Canis lupus, Procyon lotor,
This tick deserves a closer examination as to its Urocyon, cinereoargenteus, Vulpesvulpes,. Artiodactyladistribution and abundance in the southeast U.S. Odocoileusvirginianus, cattle. Primate- Homo sapiens.
Rodentia- Peromyscus gossypinus, Rattus norvegicus,Ixodes marxi Banks Oryzomys palustris, Mus musculus, Neotomafloridiana,
COUNTY RECORD: Marion. Sigmodon hispidus. Chiroptera, Passeriformes- SeiurusHOST RESORD: Rodentia- Sciurus carolinensis. aurocapillus. Sqaumata- Eumeces laticeps.
ACCESSION NUMBERS: 018713- 29, 066128- ACCESSION NUMBERS: 122349- 94, 122321-
24. 95, 122320-94, 122319- 95, 120168- 91, 120000-90,REMARKS: Two South Carolina records exists 122318- 95, 057333- 33, 013724- 00, 058649- 30, 121106-
for this species and both were taken from gray squirrels. 39, 121103- 39, 121086- 11, 121085- 11, 121080- 30,
Squirrels appear to be a common host for 1. marxi. It 121077- 39, 121070- 33, 121071- 30, 121069- 33, 120890-
probably has a wide distribution in South Carolina. 65, 119022- 94, 121112- 38, 121108- 33, 031887- 49,However, gray squirrels, although a game animal, are 019439- 39, 057174- 11, 057171- 11, 057167- 31, 057164-
rarely trapped and examined by biologists here. 11, 060229- 32, 058678- 31, 058677- 31, 058657- 36.
REFERENCES: Tedders et al. 1994, Barton et al.Ixodes minor Neumann 1992, Clark et al. 1996, Clark et al. 1998, Felz et al.
COUNTY RECORDS: Charleston, Georgetown, 1996, Durden et al. 1997.
Oconee. REMARKS: The black- legged tick is the most

230 Journal of Vector Ecology December, 1999
widely distributed tick species in SC ( Tedders 1994, because of the rather low infection rate he found in I.
Barton et al. 1992, Clark et al. 1996, 1998, Felz et al. scapularis.
1996, Durden et al. 1997). It has a wide range of hosts
including humans. It is a frequent parasite of deer in Ixodes texanus Banks
South Carolina( Barton et al. 1992) as well as in Alabama COUNTY RECORDS: Charleston, Georgetown.
Durden et al. 1991). Immature stages are common on HOST RECORD: Carnivora- Procyon lotor.
various reptiles, small mammals, and birds ( Bishopp ACCESSION NUMBERS: 120244- 91, 120243-
and Trembley 1945, Durden et al. 1997, Clark et al 91, 121110- 39.
1998). Tedders ( 1994) collecting ticks mainly from REFERENCE: Tedders 1994.
opossums and raccoons found that I. scapularis adults REMARKS: This species has been collected onlywere most abundant on opossums. He also collected from raccoons in South Carolina( Tedders 1994). It is
adults from gray foxes. Very few immature/. scapularis probably widespread throughout the state. Durden andwere collected in this study, however, he did record Kollars( 1992) collected this species from opossum and
nymphs and larvae on raccoons, opossums, and cotton red fox in Tennessee. It has also been collected from
mice and larvae on house mice and Florida wood rats. groundhogs and rabbits in Maryland ( Clifford et al.
Clark et al. ( 1998) found that immature I. scapularis 1961). In the western U.S. it has been taken from grayrepresented only about 3%( n= 52) of all ticks collected squirrels, martens, skunks, and chipmunks( Furman and
from 8 species of rodents. No adults were recovered Loomis 1984).
from rodents. However, among host-seeking ticks, I.scapularis was the most abundant species representing Ixodes woodi Bishopp46% of all ticks collected. Studies of infestations of I. COUNTY RECORD: Pickens
scapularis on domestic animals in South Carolina are HOST RECORD: Rodentia- Neotoma floridana
lacking. haematoreia.
In South Carolina adult I. scapularis appear first ACCESSION NUMBER: RML122818- 99.
around the middle of September and are usually gone by REMARKS: Three female specimens of I. woodi
the middle of May( Clark et al. 1998). Hence, it could recovered from an eastern woodrat trapped on Salem
be called a winter tick with respect to the adult stage. We Top( ca. 1000 m) near McKinney Mountain in Pickenshave examined specimens from white- tailed deer before Co. ( 34° 56' 04.4" N, 82°52' 39.0" W) represent the first
they were prepared for consumption and found many records of this tick in South Carolina. It is the eastern-
ticks to be in copula. They commonly attach to dogs. most record for this tick in North America although it is
One of us ( WW) walked his black Labrador retrievers only ca. 30 km east of a recent North Carolina recordduring temperatures in the teens and 20s. During these ( McCay and Durden 1996). This tick typically inhabitswalks 1. scapularis crawled onto the dogs but sometimes damp montane habitats where it mainly parasitizesthey did not attach for a week to ten days. It appears that woodrats ( Robbins and Keirans 1992). Although it is
the black-legged tick is almost certainly a vector of the relatively widely distributed in western states, it hasagent of Lyme disease( Lane et al. 1991) and probably only been recorded from Indiana, Alabama, Northis one of the main vectors in South Carolina( Durden et Carolina, and, now, South Carolina in the eastern Unitedal. 1997, Clark et al 1998). However, given its wide States( McCay and Durden 1996).distribution and relative abundance within the state,
very few reports of nymphal ticks removed from humans Rhipicephalus sanguineus ( Latreille)
exist. Felz et al.( 1996) found that in Georgia and South COUNTY RECORDS: Greenwood, Richland,
Carolina 3. 9% of all tick bites involved 1. scapularis. Orangeburg.Of the 36 I.scapularis recovered in their study, all were HOST RECORDS: Carnivora- Canis lupus.
adult ticks except for one nymph. They suggested that ACCESSION NUMBERS: 119332- 88, 119331-
immatures may be feeding on other animals including 88, 122350- 94.
lizards or that immatures went undetected because of REFERENCE: Clark et al. 1996.
their small size. Although Clark ( 1996) found B. REMARKS: The brown dog tick is a cosmopolitanhurgdorferi infection levels in cotton mice, cotton rats, species that has traveled with man and his dogs. In theand woodrats from the coastal zone of South Carolina U.S., both adults and larvae are found almost exclusively50.0 to 88. 8%) to be comparable to those of white- on dogs ( Bishopp and Trembly 1945, Clifford et al.
footed mice in the northeastern U.S., he suspected that 1961). However, one of us ( WW) collected 2 femalesthe enzootic cycle in that area may have been maintained from a bird nest in Saudi Arabia. This species is found
by I. affinis and I. minor rather than by I. scapularis commonly in animal shelters and kennels and is often

December, 1999 Journal of Vector Ecology 231
carried by dogs into the house where it may become arborviruses ( 2"`' Ed.). DHEW pub. #( CDC) 75-
established. They are rarely found outside. Control 8301.
operators sometimes fail to control this tick since they Bishopp, F. C. and H. L. Trembley. 1945. Distributionclimb walls in houses, shelters, or kennels. Although we and hosts ofcertain North American ticks. J. Parasit.have found only 3 county records, this tick is probably 31: 1- 54.
found over much of the state. Bolte, J. R., J. A. Hair, and J. Fletcher. 1970. White-
tailed deer mortality following tissue destructionOrnithodoros capensis Neumann induced by lone star ticks. J. Wildl. Mgmt. 34: 546-
COUNTY RECORD: Allendale, Charleston. 552.
HOST RECORD: Pelecaniformes - Pelecanus Burgdorfer, W., T. R. Adkins, and L. E. Priester. 1975.occidentalis. Strigiformes- Strix varia. Rocky Mountain spotted fever( tick-borne typhus)
ACCESSION NUMBERS: 118625- 87, 118572- in South Carolina: An educational program and87. tick/ rickettsial survey in 1973 and 1974. Am. J.
REFERENCE: Keirans et al. 1992. Trop. Med. Hyg. 24: 866- 872.REMARKS: This species was first reported from Clark, K. L. 1996. Prevalence of Borrelia burgdorferi
South Carolina by Keirans et al.( 1992) from Bird Key in rodents and host- seeking ticks( Acari: Ixodidae)Stono, a small island bird sanctuary in Charleston from localities in South Carolina. Ph.D. Dissertation.
County. Brown pelicans had abandoned their nests due University of South Carolina, Columbia, 132 pp.to heavy tick infestations. Yunker et al.( 1979) isolated Clark, K. L., J. H. Oliver, D. B. McKechnie, and D. C.
3 strains of Aransas Bay virus from this species. This is Williams. 1998. Distribution, abundance and
the only argasid tick we have recorded from South seasonal activities of ticks collected from rodents
Carolina. Because O. capensis is a seabird tick, we were and vegetation in South Carolina. J. Vect. Ecol. 23:
surprised to recover a larval specimen from a barred owl 89- 105.
in Allendale County. Clark, K. L., W. Wills, S. H. Tedders, and D. C.
Williams. 1996. Ticks removed from dogs and
Acknowledgments animal care personnel in Orangeburg County, SouthCarolina. J. Agromed. 3: 45- 55.
We thank the following for assistance in collecting: Clifford, C. M., G. Anastos, and A. Elbl. 1961. The
Dr. Stan Miller, Campbell Museum of Natural larval ixodid ticks of the Eastern United States
Sciences, Clemson University, Clemson, South Acarina- Ixodidae). Misc. Pub. Entomol. Soc. Am.
Carolina; Dr. John Morse, Department of Entomology, 215- 237.
Clemson University; Dr. Susan Loeb, U. S. Forest Clifford, C. M., D. E. Sonenshine, E. L. Atwood, C. S.
Service, Clemson, South Carolina; Ms. Mary Bunch, Robbins, and L. E. Hughes. 1969. Tests on ticks
South Carolina Department of Natural Resources, from wild birds collected in the Eastern United
Clemson, South Carolina; Mr. David Vaughan, South States for rickettsiae and viruses. Am. J. Trop.Carolina Department of Health and Environmental Med. Hyg. 18: 1057- 1061.Control, Bureau of Laboratories, Columbia, South Davidson, W. R., D. A. Siefken, and L. H.Creekmore.
Carolina; Mr. Ron Bowers for use of his farm as a 1994. Seasonal and annual abundance ofcollection site; Mr. James Ronan for assistance in Amblyomma americanum ( Acari: Ixodidae) in
preparing the manuscript; and Ms. Janet Ciegler and Central Georgia. J. Med. Entomol. 31( 1): 67- 71.
Ms. Virginia Winn for assistance in preparing the South Durden, L. A. and T. M. Kollars, Jr. 1992. An annotatedCarolina map. list of the ticks( Acari: Ixodidae) of Tennessee, with
records of four exotic species for the United States.
REFERENCES CITED Bull. Soc. Vector Ecol. 17: 125- 131.
Durden, L. A., R. G. Mclean, J. H. Oliver, Jr., S. R.
Barton, W. E., E. W. Gray, and D. Shipes. 1992. An Ubico, and A. M. James. 1997. Ticks, Lyme
investigation of the status of Borrelia burgdorferi disease spirochetes, trypanosomes, and antibody toand its suspected primary vector, Ixodes scapularis, encephalitis viruses in wild birds from coastal
in South Carolina. J. S. C. Med. Assoc. 88: 5- 8. Georgia and South Carolina. J. Parasitol. 83: 1 178-
Becklund, W. W. 1968. Ticks of significance found on 1182.
imports in the United States. J. Parasit. 54: 622- Durden, L. A, S. Lockhart, G. R. Mullen, and S. Smith.
628. 1991. Tick infestation of white- tailed deer in
Berge, T. O. ( Ed.) 1975. International catalogue of Alabama. J. Wildl. Dis. 27: 606- 614.

232 Journal of Vector Ecology December, 1999
Felz, M. W. and L. A. Durden. 1998. Identifying ticks: Louis encephalitis virus from adult Dermacentora pictorial guide. Patient Care 32: 172- 187. variabilis ( Acari: Ixodidae). J. Med. Entomol. 22:
Felz, M. W., L. A. Durden, and J. H. Oliver, Jr. 1996. 232- 233.
Ticks parasitizing humans in Georgia and South Oliver, J. H., Jr., J. E. Keirans, D. R. Lavender, and H.Carolina. J. Parasitol. 82: 505- 508. J. Hutcheson. 1987. Ixodes affinis Neumann
Furman, D. P. and E. C. Loomis. 1984. The ticks of Acari: Ixodidae): New host and distribution records,California. Bul. U.CA. Pub. 239 pp. description of immatures, seasonal activities in
Gerrish, R. R. and J. M. Ossorio. 1965. New record of Georgia, and laboratory rearing. J. Parasitol. 73:the tick Ixodes affinis in the United States. J. Econ. 646- 652.
Entomol. 33: 69. Robbins, R. G. and J. E. Keirans. 1992. Systematics andHawkins, M. M. 1995. Human ehrlichiosis: a case ecology of the subgenus Ixodiopsis ( Acari:
report from the South Carolina lowcountry. J. S. C. Ixodidae: Ixodes). Thomas Say Found. Vol. XIV,Med. Assoc. May: 228- 229. Entomological Society of America, Lanham, MD,
Hopla, C. E. and A. K. Hopla. 1994. Tularemia. 159 pp.Pp. 113- 126 in Handbook of zoonoses, Section A: Rumpel, C. and J. L. Jones. 1991. Lyme disease inBacterial, Rickettsial, Chlamydial, and Mycotic, South Carolina. J. S. C. Med. Assoc. Aug.: 420-2nd
ed.( G.W. Beran, ed.), CRC Press, Boca Raton, 423.
538 pp. Sanders, F. H. and J. H. Oliver, Jr. 1995. Evaluation ofJames, M. T. and R. F.Harwood. 1970. Herm' s Medical Ixodes scapularis, Amblyomma americanum, and
Entomology. The MacMillan Co., Collier- Dermacentor variabilis ( Acari: Ixodidae) fromMacMillan Limited, London, 484 pp. Georgia as vectors of a Florida strain of the Lyme
Keirans, J. E., H. J. Hutcheson, and J. H. Oliver, Jr. disease spirochete, Borrelia burgdorferi. J. Med.1992. Ornithodoros ( Alectorobius) capensis Entomol. 32: 402-406.
Neumann( Acari: Ixodoidea: Argasidae), aparasite Schuman, S. H. and S. T. Caldwell. 1989. Lyme andof seabirds, established along the Southeastern other tick-borne diseases acquired in South Carolinaseacoast of the United States. J. Med. Entomol. 29: in 1988: A survey of 1, 331 physicians. J. S. C. Med.371- 373. Assoc. 85: 311- 314.
Lane, R. S., J. Piesman, and W. Burgdorfer. 1991. Stone, B. F. 1963. Parthenogenesis in the cattle tick,Lyme Borreliosis: Relation of its causitive agent to Boophilus microplus. Nature. 200: 1233.its vectors and hosts in North America and Europe. Strickland, R. K., R. R. Gerrish, J. L. Hourigan, and G.Annu. Rev. Entomol. 36: 587- 609. O. Schubert. 1977. Ticks of veterinary importance.
Lavender, D. R. and J. H. Oliver, Jr. 1996. Ticks Animal and plant inspection Service- U.S. Dept.Arcari: Ixodidae) in Bulloch County, Georgia. J. Agr., Agr. Hndbk. 485.
Med. Entomol. 33: 224-231. Tedders, S. H. 1994. Distribution of the ticks( Ixodidae)Levine, J. F., C. S. Apperson, and W. L. Nicholson. of coastal South Carolina and control of the lone
1989. The occurrence of spirochetes in ixodid ticks star tick,Amblyomma americanum, using cyfluthrin,in North Carolina. J. Entomol. Sci. 24: 594- 602. a synthetic pyrethoid. P. h. D. Dissertation.
Lockhart, J. M., W. R. Davidson, J. E. Dawson, and D. University of South Carolina, Columbia, 146 pp.E. Stallknecht. 1995. Temporal association of Tedders, S. H., J. E. Keirans, D. C. Williams, and T. A.Amblyomma americanum with the presence of Gwinn. 1992. First report of Ixodes( Ixodes) minorEhrlichia chaffeensis reactive antibodies in white- Neumann ( Acari: Ixodidae) from South Carolina.tailed deer. J. Wildl. Dis. 31: 119- 124. J. Med. Entomol. 29: 282- 283.
Loving, S. W., A. B. Smith, A. F. DiSalvo, and W. Thompson, E. F., Jr. 1982. A guide to the amphibians,Burdorfer. 1978. Distribution and prevalence of reptiles, and mammals of South Carolina. The Statespotted fever group rickettsiae in ticks from South Printing Company, Inc. Columbia, SC, 134 pp.Carolina with an epidemiological survey of persons Wilson, N. and W. W. Baker. 1972. Ticks of Georgiabitten by infected ticks. Am. J. Trop. Med. Hyg. Acarina: Metastigmata). Bull. Tall Timbers Res.27: 1255- 1260. Stn. 10: 1- 29.
McCay, T. S. and L. A. Durden. 1996. Ticks and fleas Yunker, C. E., C. M. Clifford, J. E. Keirans, L. A.of shrews in Appalachian Georgia and North Thomas, and R. C. A. Rice. 1979. Aransas bayCarolina. J. Parasitol. 82: 666- 667. virus, a new arbovirus of the Upolu serogroup from
McLean, R. G., D. B. Franey, T. P. Monath, C. H. Ornithodoros capensis( Acari: Argasidae) in coastalCalisher, and D. W. Trent. 1985 Isolation of St. Texas. J. Med. Entomol. 16: 453- 460.


















