
“Ecology around us” “Ecology around us”. What science studies nature? Ecology.
Upload
truongnguyetCategory
view
219download
0
BSOVE 19( 1): 1- 86 ( 1994)
ISSN 0146-6429
Bulletin of the
SOCIETY FOR
VECTOR ECOLOGY
11 AL :
4I
IL fi' - i
Volume 19, Number 1 June, 1994

BULLETIN OF THE
SOCIETY FOR VECTOR ECOLOGYVolume 19- Number 1 June 1994
Marc J. Klowden, EditorDivision of Entomology
University of IdahoMoscow, ID 83844-2339
Phone: ( 208) 885- 7546
Fax: ( 208) 885- 7760
EDITORIAL BOARD
Mir S. Mulla, Chairman, W. J. Crans R. S. Nasci
University of California Rutgers University CDC-DVBIDRiverside, CA 92521 New Brunswick, NJ Fort Collins, CO
R. C. Axtell C. I. Dahl M. W. ServiceNorth Carolina State University University of Uppsala Liverpool School of Tropical Med.Raleigh, NC Uppsala, Sweden Liverpool, England
N. Becker F. R. Holbrook E. D. Walker
University of Heidelberg USDA-ARS Michigan State UniversityGermany Laramie, WY East Lansing, MI
D. R. Barnard R. S. Lane S. K. WikelMed. Vet. Entomol. Lab University of California Oklahoma State UniversityUSDA, Gainesville, FL Berkeley, CA Stillwater, OK
H. Briegel G. G. Marten S. C. Williams
University of Zurich New Orleans MCD San Francisco State UniversityCH-8057, Zurich, Switzerland New Orleans, LA San Francisco, CA
E. P. Catts L. S. MianWashington State University County Vector ControlPullman, WA San Bernardino, CA
Published by the Society for Vector Ecologyto disseminate pertinent information from all
facets of the field of vector ecology and related disciplines.
Communications relating to editorial matters and manuscriptsshould be addressed to the Editor. Communications relating to galley proofs,
reprints, subscriptions, SOVE membership, change of address, and other mattersshould be addressed to the Business Office.
Membership Plus Bulletin$ 45.00 PUBLICATIONS AND BUSINESS OFFICEStudent Membership$ 22.50 Society for Vector Ecology
Institutional Subscription$ 45.00 P. O. Box 87
Sustaining Members$ 100.00 Santa Ana, California 92702 USA

SOCIETY FOR VECTOR ECOLOGY
BOARD OF DIRECTORS
1994 OFFICERS
President Susan M. PalchickPresident-Elect Bruce F. EldridgeVice-President A. Ralph BarrPast-President Norbert Becker
Secretary- Treasurer Gilbert L. Challet
REGIONAL DIRECTORS
Southwestern Minoo B. MadonNorthwestern Steven V. RomneyNorth Central Thomas R. WilmotSouth Central Cluff E. HoplaNortheastern Wayne J. CransSoutheastern Jonathan F. DayEuropean Frantisek Rettich
PUBLICATIONS AND BUSINESS OFFICE
Society for Vector EcologyP. O. Box 87
Santa Ana, California 92702 USA
Membership Plus Bulletin$ 45.00Student Membership$ 22.50
Institutional Subscription$ 45.00
Sustaining Members$ 150.00

BULLETIN
OF THE
SOCIETY FOR VECTOR ECOLOGY
Volume 19 June, 1994 Number 1
CONTENTS
Guidelines for Contributors ii
Submitted Papers
Seasonality ofAnopheles Maculatus, the Main Vector of Malaria in Peninsular Malaysia Near the ThailandBorder W. A. Rahman, A. Abu Hassan, C. R. Adanan, and M. R. Rashid 1
Host-Parasite Relationships Between Fleas ( Siphonaptera) and Bats( Chiroptera) Hibernating in Ice andLimestone Caves In Slovakia M. Trpis 8
Host-Parasite Relationships Between Fleas( Siphonaptera) and Small Mammals of the Tatras Mountains in
Slovakia M. Trpis 13
Host Specificity and Ecology of Fleas( Siphonaptera) of Small Mammals in Mountains of North-CentralSlovakia M. Trpis 18
Vector Competence ofWestern European Mosquitoes for Arboviruses: A Review ofField and Experimental
Studies J. O. LundstrOm 23
A Critique ofTechniques for Detection, Identification, and Classification of Sibling Species ofMosquitoesA. F. Cockburn 37
The Impact of Water Quality on the Persistence of Bacillus SphaericusM. Ludwig, M. Beck, M. Zgomba, and N. Becker 43
Color and UV Reflectance of Canopy Traps for Collecting Horse Flies( Diptera: Tabanidae) in LouisianaL. J. Hribar and L. D. Foil 49
A Redescription of the Morphology of Mosquito( Diptera: Culicidae) Ovarioles During Vitellogenesis ..M. I. Sokolova 53
Proceedings
First International Congress of Vector Ecology, San Diego, California, 3-8 October 1993
The Dismal State of Mosquito Systematics: Perspectives of a Classical Taxonomist T. J. Zavortink 69
Ross River Virus: Disease Trends and Vector Ecology in Australia Richard C. Russell 73
Some Sandfly Food is a Leishmania Poison Y. Schlein and R. L. Jacobson 82

BULLETIN OF THE SOCIETY FOR VECTOR ECOLOGY
Guidelines for Contributors
The Bulletin of the Society for Vector Ecology is an international journal concerned withall aspects of the biology, ecology, and control of arthropod vectors and the interrelationshipsbetween the vectors and the disease agents they transmit. The journal publishes original researcharticles and research notes, as well as comprehensive reviews of vector biology based onpresentations at Society meetings. All papers will be reviewed by at least two referees who arequalified scientists and who recommend their suitability for publication. Acceptance of
manuscripts is based on scientific merit and is the final decision of the editor, but these decisions
may be appealed to the Editorial Board.
Scientific contributions should be sent to Dr. Marc J. Klowden, Editor, Division of
Entomology, University of Idaho, Moscow, Idaho 83843, U.S. A. Manuscripts must be doublespaced on a single side of bond paper with 25 mm margins. An original and two clear copies
are required. Draft mode dot matrix type should not be used. Submission of text on a 3- 1/ 2"computer diskette formatted in MS-DOS is encouraged. Microsoft Word, Word Perfect, or
Wordstar formats are acceptable, as well as unformatted text files. Please indicate the type of
format on the diskette label. Papers must be organized under the following headings, each ona separate page, in order: Title page, abstract, text, acknowledgments( ifappropriate), references
cited, tables, figure legends, and figures. The title page should contain the names of all authorsand their affiliations and the identification and address of the corresponding author. Pages
should be numbered consecutively starting with the title page. References should conform tothe style in recent volumes. Illustrations that are submitted must be clearly labeled and legibleafter reduction.
Pagecharges, which partially defray the cost ofpublication, are$ 35 per printed page.SOVE members who are unable to pay page charges may apply for a limited number ofwaivers. Reprint charges are shown in the table below.
Pages 1- 4 5- 8 9- 12 13- 16 17- 20
50 copies 60.00 105.00 150.00 195.00 240.00
or less
Each add' l 20.00 30.00 40.00 50.00 60.00
50 copies
Same order
ii

BULL. SOC. VECTOR ECOL., 19( 1): 1- 7 DECEMBER, 1993
SEASONALITY OF ANOPHELES MACULATUS, THE MAIN VECTOR OF MALARIA
IN PENINSULAR MALAYSIA NEAR THE THAILAND BORDER
W. A. Rahman 1, A. Abu Hassanl, C. R. Adanan 1, and M. R. Rashid2
ABSTRACT: Seasonal distribution of the malaria vector mosquito, Anopheles maculatus in northern Peninsular
Malaysia near the Thailand border was studied by collecting mosquitoes using a cow-baited net trap. Anophelesmaculatus is abundant just before and after rainy periods. Heavy rain may flood mosquito larvae from their breedingsites and decrease the numbers of adults during that season.
INTRODUCTION MATERIALS AND METHODS
Anopheles ( Cellia) maculatus Theobald occurs Study Areathroughout the Oriental zoogeographic region. It is Observations were conducted from January 1990
highly variable in both its morphology and ability to to December 1991 in the small village of Kampongtransmit malaria( Reid 1968), and may represent another Bongor, located about 13. 5 km from the Thailand border
complex of species having little or no morphological to the north of the area( 101° 11' E longitude, 5° 30.5'
differentiation ( Reid 1970). In West Malaysia, An. N latitude)( Fig. 2). The human inhabitants were mainlymaculatus is a widespread species and is found in rubber tappers, but because tapping rubber was notforested and foothill areas. The construction of the East- possible during the rainy season, during this time theyWest Highway linking eastern and western coastal parts were employed as forest-product gatherers. Most houses
ofWestMalaysia in the early 1980s has made new lands were built on elevated wooden platforms, about 2 m
in hilly areas available for the planting of rubber and oil high, with a few at ground level and covered with nipah
palm. It has been shown that whenever the cover of leaf or zinc roofing. Interiors and under- flooring ofjungle is cleared from hilly areas, An. maculatus, the houses were sprayed by government malaria controlmain vectorof the disease, breeds prolifically in seepages workers twice a year with 25 percent EC DDT at a rate
and streams and may thus be responsible for the high of 2 gm/m2.
malaria endemicity in those newly-opened areasSandosham 1970). Vegetation, Topography, and Meteorology
In 1988, there was a dramatic rise in malaria cases The 150 houses in the village were in or near rubber
in the district of Hulu Perak especially for villages at plantations, abandoned rice-fields, rambutan trees, or
forest fringe. Kampong Bongor is one of such villages. scrub plants comprised mainly ofAmaranthus spinosus,The number of malaria cases detected for the year were Ageratum conyzoides, Eupatorium odoratum, and
high in the village ( Fig. 1). Between 1988- 1991, 53 Melastoma malabathricum. The study site was locatedpercent of cases( 195) were ofPlasmodiumfalciparum, on a river valley with slopes gently rising from 4° from45 percent ( 167) were of Plasmodium vivax, and 2 the river to nearly 20° near the foothill, about 600-750percent ( 7) were of mixed infections. Rahman et al. feet above sea level ( Malaysia 1969). The regional
1992) studied the transmission of malaria in this geology of the area is of Silurian-Ordovision age. Thevillage and incriminated An. maculatus as the major rock consists of mainly schist, phyllite, slate, and
vector. limestone with minor intercalation of sandstone and
The present study reports on the seasonal distribution volcanics( Malaysia 1985). Daily rainfall readings wereof An. maculatus populations in an endemic area near recorded at Kuala Kenderong Meteorological Station,the Malaysia-Thailand border. Department of Irrigation and Drainage, Malaysia,
1School of Biological Sciences, Universiti Sains Malaysia, Minden 11800, Penang, Malaysia.
2Centre for Off-Campus Studies, Universiti Sains Malaysia, Minden 11800, Penang, Malaysia.

2 BULL. SOC. VECTOR ECOL. DECEMBER, 1993
prnminn
et)
XX\
alimmummummiiii
lm
ti
t
a
EZZZ
OM N N I O Vi O
saseD euulew Jo iactumN
Figure 1. Malaria cases detected in Kampong Bogor, Perak, during 1988- 1991.
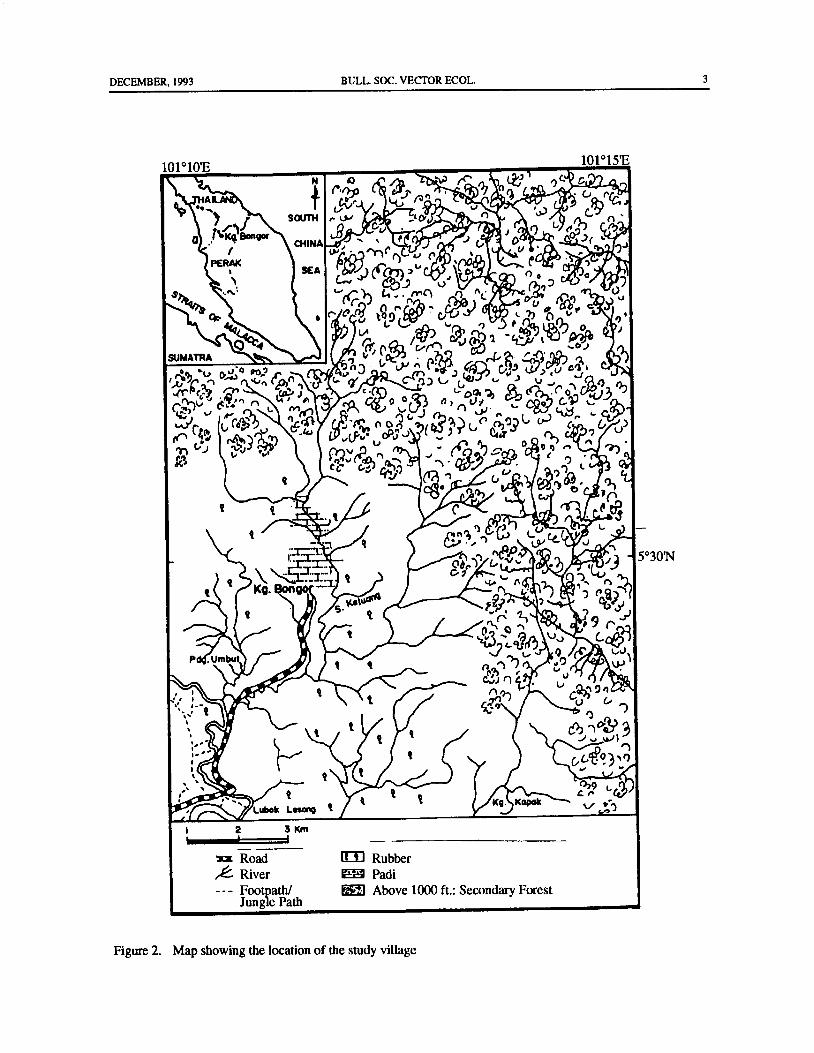
DECEMBER, 1993 BULL. SOC. VECTOR ECOL. 3
101° 10'E101° 15'E
N 4 v) , Le1 C) G .• ..-
1'), 0
q t • c,,,, cc,-.•
j Sotr H , w o- nI n
o l KaBonQorCHINA • ' ' Q G 0
e.:1•")) • .
u . ••
PERK
W ' Yl C. u n •'Dd.. P ,,',
1SEA , ) V al)
V,
A u a 47J
lU j-..,cJtra
SUMATRA VL t ^ '
A, A. - 7 n L,
b-,,,, -., 0 0,..,
Co Vfo`" f.!'.i v: no/
3G.
3j( 5 T 1.n
i0.9' c'., L`'•
1Cu
f9 Garn
Cfu n
r1 tNv/P , u+ ( fin G L ° ' c
ca y) X11
1- . D ', 0;
t
G Ly 01
y
lM1l
I t Cf
e r•1 8 _, -c,
r.+I_ tG 7 ,
mot. ('
r.
S.
V
l,JP . Umbut
t I 0 , 1'Y
u I ' wlPif r 3,...
g.);1 {,0
I
Cal ni_ 9n/
1 t cd '-)
It
I 6))..;)„"....)4
1
t79
2 t G^i,• ° ..!- t t Kg. Koh
c- ,lubok Legal; t v r..„%:;)'
I 2 3Km
Road Rubber
ee River Padi
Footpath/ Above 1000 ft.: Secondary ForestJungle Path
Figure 2. Map showing the location of the study village

4 BULL. SOC. VECTOR ECOL. DECEMBER, 1993
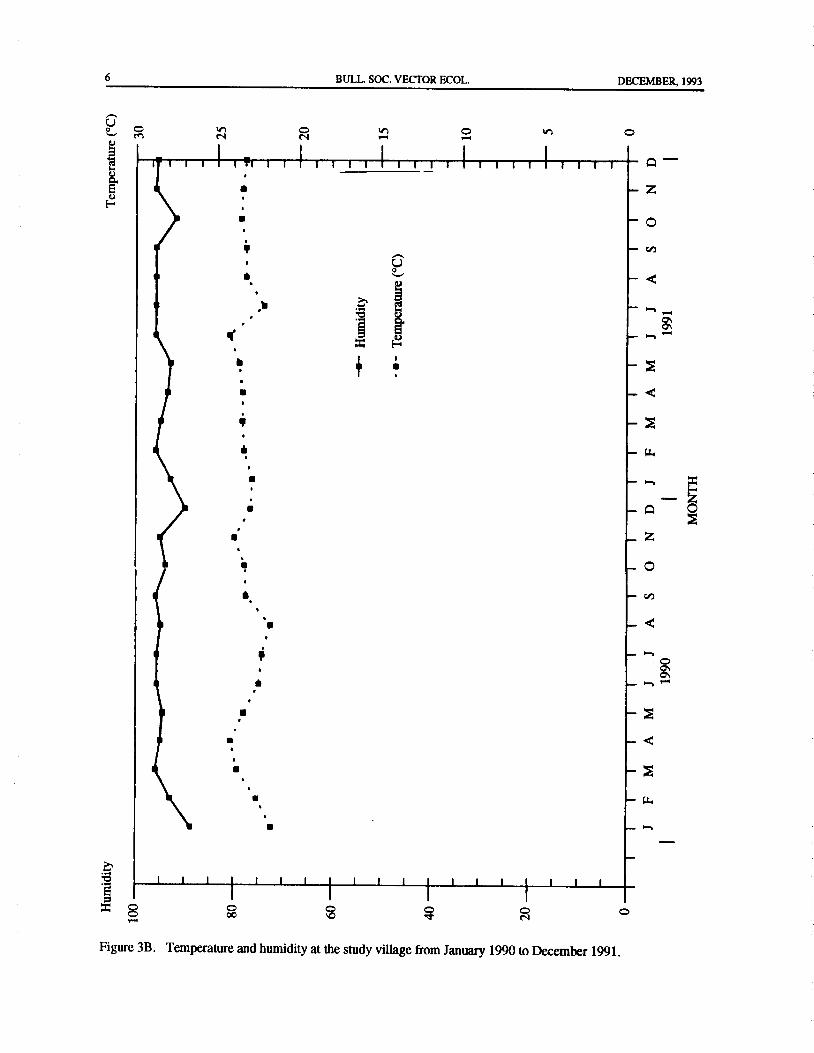
situated about 0. 5 km from the study area The mean were the driest months of the year. Other species ofannual rainfall( 1986- 1990) was 3362.4 mm, distributed Anopheles collected were An. aconitus, An. kochi, An.unevenly throughout the year, with heavy monsoon barbirostris, and An. philippinensis.rains during July-December. Monthly temperature and During the dry season, due to high catches, therehumidity readings were recorded at the study area. was a clear relationship between An. maculatus density
and drought in the sampling area; the frequency of highMosquito Collections mosquito catches was relatively more during periods of
Preliminary indoor and outdoor landing catches on low rainfall, but low during periods ofhigh rainfall( Fig.human and cow-baited net trap catches were carried 3A). However, relationships between An. maculatusout in November and December 1989. Although results densities with that of temperature and relative humidityfrom landing catches were discouraging, cow-baited were not distinct(Fig. 3B). Anopheles maculatus larvaenet trap catches were more promising. Thus, the cow- were mostly found in seepages, small pools of water,baited net trap( Service 1976) was chosen because the small streams, and drains in the cleared areas of thecatches were more promising and Anopheles mos- foothill and the paddy fields.quitoes were zoophilic with cow: man feeding ratioof 3: 1 ( Reid 1968). Anopheles mosquitoes were col- DISCUSSION
lected for one night each month from January 1990 toDecember 1991, using the cow-baited net trap method. At Kampong Bongor, An. maculatus possessesCaptures were made for 10 min.periods at the end of the characteristics of a good malaria vector because of itshour from 1900 to 0700 h by a two-man team at ground presence in high numbers, especially during the earlierlevel, 2 m outside an inhabited dwelling. However, the part of the year when many cases of malaria weresunset time was set at 1900 hours to allow for seasonal reported to the District Hospital( Annual Report of thevariation in daylight length according to local mean Malaysian Ministry of Health 1990, unpub.). It istime. Mosquitoes were identified the following morning; interesting to note that populations of An. maculatusonly An. maculatus was considered. seemed to occur in large numbers during the dry season,
with the numbers very much reduced during the wetLarval Survey season. In neighboring Thailand, Rosenberg et al.
A search for An. maculatus larvae was carried out ( 1990) also reports a highly efficient transmission ofonce a month from January 1990 to December 1991, malaria during the dry season, but due to Anophelesusing standard 350 ml dipper in seepages and streams of minimus/dirus, and not An. maculatus. If female An.the cleared areas of the foothill, paddy fields, and moist maculatus dispersed randomly from points of blooddepressions. feeding and oviposition, then there should have been no
sharp discontinuities in survival rates within the smallRESULTS study area. The village has rice paddies that are
randomly distributed, and because most AnophelesThe numbers of An. maculatus mosquitoes col- mosquitoes are capableof flights over uncovered ground
lected and the meteorological measurements for 24 of 0. 8 km ( Colless 1953), sites for oviposition aremonths are plotted in Figure 3A. The dry season started plentiful. The results of this study suggest that manyAn.in January and ended in May 1990, while the rest of the maculatus females may have been successful in layingperiod received substantial amount ofrainfall except for their eggs in seepages, paddy fields, and other suchthe month ofFebruary 1991. Temperatures at the study moist depressions that were convenient for them assite ranged between 21. 8° C and 24.3° C. The coolest adults but that were unfavorable for the larval offspringmonth occurred in August 1990, while the hottest was during the rainy season. Large numbers of these larvaeJune 1991. Relative humidity values ranged between would probably be washed away in heavy rains and fast-88.8 percent and 96.0 percent. The lowest value was flowing waters. The life expectancy of adult An.recorded in January 1990, the highest in March and maculatus is longer in the dry than in the wet seasonSeptember 1990 and from June to September 1991. Ratanatham et al. 1988), and could also bean explanation
During this study a total of 790 An. maculatus was for its high numbers during the dry season as recordedcollected. Sixty- two percent were collected in the dry in the present study.season( January-May 1990), and only 38 percent during Surprisingly, although An. maculatus is athe rest of study period( June 1990 to December 1991). widespread species in neighboring Thailand, it has beenDensities of An. maculatus were highest in February incriminated only once as a malaria vector( Upatham etand March ( 36 percent of total collections), which al. 1988). Under field conditions this species apparently

DECEMBER, 1993 BULL SOC. VECTOR ECOL. 5
4.,
5 I I I I
illE z
1 Cl)
I
I
Ii I2
E00 2
1 z
1 0
1
f4 Iu 1 0,
th,7
I_____:o
IF
E, i,
I.?.co
U
2
0 s2 .ot-si 8 oo 2 4 0
Figure 3A. Monthly rainfall and catches of Anopheles maculatus at the study village from January 1990 toDecember 1991.
6 BULL. SOC. VECTOR ECOL. DECEMBER, 1993
o O ON n
I I I I ( 1 111 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 q
H
z
o
g.El
U
r1
N
u.
q 0
o
2
2
w
1 1 1
11 1 1 ( 1 1 1 ( 1 1 1 j 1 1 1
8 N 0
Figure 3B. Temperature and humidity at the study village from January 1990 to December 1991.

DECEMBER, 1993 BULL. SOC. VECTOR ECOL. 7
does not serve as a malaria vector; but nevertheless, Malaysia. S. E. Asian J. Trop. Med. Pub. Hlth. 20:there is a possibility that it may act as potential malaria 415-420.
vector in Thailand. DDT residual house spraying hasbeen carried out as part of the vector control program in Malaysia. 1969. Topographic Sheet 19: Kerunai,
the study village since 1969, but its affect on An. National Mapping, Malaysia.maculatus populations is doubtful. There was a belief
among the public health workers responsible for Malaysia. 1985. Geological Map of Peninsular
Kampong Bongor that An. maculatus had developed Malaysia, Geological Survey ofMalaysia, Ministryresistance to DDT. Although most An. maculatus of Primary Industries, Malaysia.collected from different parts of Malaysia have yet toshow thisresistance( Loongetal. 1989), the development Rahman, W. A., A. Abu Hassan, Mohd Razha A.
of resistance in An. maculatus from Pakistan, India, Rashid, and Abdul Hamid Khalid. 1992. Malaria
Myanmar, and Thailand( Brown 1986) showed that it is transmission in a remote village located in northern
possible, especially in an environment like Kampong peninsular near Malaysia-Thailand border. Trop.Bongor where DDT has continuously been used for a Biomed. 9: 1- 7.
long period of time. Furthermore, with more landsextensively being opened up for rubber, oil palm, and Ratanatham, S., E. S. Upatham, C. Prasittisuk, W.
fruit tree plantations, the steady increase of favorable Ojanasunan, N. Theerasilp, A. Tremongkol, andV.vector habitats for the breeding of An. maculatus Viyanant. 1988. BionomicsofAnophelesminimus
mosquitoes has made it more difficult for the health and its role in malaria transmission in Thailand. S.
workers of the district to control populations of this E. Asian J. Trop. Med. Pub. Hlth. 19: 283- 289.malaria vector.
Reid, J. A. 1968. Anopheline mosquitoes of MalayaAcknowledgments and Borneo. Studies of the Institute for Medical
Research, Kuala Lumpur, Malaysia. No. 31, 520
The authors are grateful to all personnel of the pp.
Department of Health, Grik, Hulu Perak District,
Ministry of Health, Malaysia for their assistance and Reid, J. A. 1970. Systematics of malaria vectors.
collaboration. We are grateful to the following Anopheline systematics and malaria control, with
individuals for their able technical help: Mr.Burhanuddin special reference to Southeast Asia. Misc. Pub.
Saad, Mr. Yusuf Omar, Mr. Abdullah Nayan, Mr. Ent. Soc. Am. 7: 56.
Abdullah Ibrahim, and Mr. Khalid Puteh of the School
ofBiological Sciences, Universiti Sains Malaysia. This. Rosenberg, R., G. A. Richard, and L. Somchit. 1990.project was made possible with Universiti Sains Malaysia Highlyefficient transmission ofmalaria in Thailand.Research and Development Grants Nos. 122/0204/0130 Trans. R. Soc. Trop. Med. Hyg. 84: 22-28.and 123/ 3305/ 2403.
Sandosham, A. A. 1970. Malaria in rural Malaya. Med.
J. Malaya 24: 221- 226.
REFERENCES CITED
Service, M.W. 1976. Mosquito ecology- FieldsamplingBrown, A. W. A. 1986. Insecticide resistance in methods. Applied Science Publishers, London, 583
mosquitoes - a pragmatic review. J. Am. Mosq. pp.
Cont. Assoc. 2: 123- 140.Upatham, E. S., C. Prasittisuk, S. Ratanatham, C. A.
Colless, D. H. 1953. Observations on the flight range Green, W. Rojanasunan, P. Setakana, N.Theerasilp,of Anopheles. Med. J. Malaya 7: 179- 184. A. Tremongkol, V. Viyanant, S. Pantuwatana, and
R. G. Andre. 1988. Bionomics of Anopheles
Loong, K. P., G. L. Chiang, and H. H. Yap. 1989. maculatus complex and their role in malaria
Susceptibility status of Anopheles maculatus transmission in Thailand. S. E. Asian J. Trop.Theobald( Diptera: Culicidae) to DDTinPeninsular Med. Pub. Hlth 19: 259- 269.

BULL. SOC. VECTOR ECOL., 19( 1): 8- 12JUNE, 1994
HOST-PARASITE RELATIONSHIPS BETWEEN FLEAS
SIPHONAPTERA) AND BATS (CHIROPTERA) HIBERNATINGIN ICE AND LIMESTONE CAVES IN SLOVAKIA
M. Trpisl
ABSTRACT: Between 1955 and 1959, 163 specimens of fleas were collected from 2,268 bats hibernating in Slovakcaves. Seven species, including two subspecies, of fleas were found: Ischnopsyllus( I.)octactenus; lschnopsyllus1.) simplex, which occurred in two subspecies, Ischnopsyllus simplex simplex and Ischnopsyllus simplex mysticus;
Ischnopsyllus( 1.) intermedius; Ischnopsyllus( H.)hectactenus; Rhinolophopsylla unipectinata; andNycteridopsyllapectactena. Fleas infested 9 of 14 species ofbats hibernating in underground caves: Rhinolophusferrum-equinum,Rhinolophus hipposideros, Eptesicus serotinus, Eptesicus nilsoni, Plecotus auritus, Myotis myotis, Myotisoxyghnatus, Myotis mystacinus and Barbastella barbastellus. Myotis mystacinus was the species most infested withfleas. The most abundant species of fleas were I. octactenus, I. hectactenus, and R. unipectinata. Quantitativerelationships between bats and fleas is presented.
INTRODUCTION on fleas from bats in Slovakia before or during thisinvestigation in 1955- 1959 was published by Rosicky
Slovakia isknown for its numerous limestone caves. ( 1950, 1952; Hurka 1956, 1957). After 1959, additionalPermanentdeepunderground ice caves are also found in information was published by Hurka( 1963a, 1963b).central Slovakia near the town ofDobsina. These caves The flea collection was deposited at the Slovakprovide shelters for many species of bats. In summer, National Museum in Bratislava in 1968.bats occupy the entrances to the caves, sleeping duringthe day and hunting insects at night. In the late fall and MATERIALS AND METHODSwinter, bats migrate deeper into the caves for theirhibernation period. In the spring, they leave the deep From 1955 to 1959, 2,268 hibernating bats wereunderground caves and migrate closer to the entrances. examined, 163 ( 7.2%) of which were infested withDetailed studies on the biology and distribution of bats fleas. Fleas in bats' fur move very quickly. To collectin Slovakia were done by Vachold( 1956). fleas from bats, the bats were placed in containers that
The composition of bat species occupying in caves received bursts ofether every one to two minutes; fleasduring the summer monthsdiffers from that living in the were then combed out of the fur onto a piece of whitecave during the winter. In summer, we found six species cloth. The fleas were then preserved in 70 percent ethylof bats in Slovak caves: Myotis schreibersii, Myotis alcohol. Collection sites, species ofbats, and number ofmyotis, Myotis oxyghnatus, Rhinolophus euryale, fleas collected were recorded. Before microscopicRhinolophus hipposideros, and Rhinolophus ferrum- examination, the fleas were cleared in phenol-xyleneequinum. In the winters from 1955 to 1959, we found and embedded in Canada balsam on microscope slides.the following species of bats over-wintering in Slovakcaves: Rhinolophus ferrum-equinum, R. hipposideros, RESULTSEptesicus serotinus, Eptesicus nilsoni,Plecotus auritus,M. myotis, M. oxyghnatus, Myotis mystacinus, and During the five winters of our studies in CentralBarbastella barbastellus. Slovakia, 14 species ofbats were captured and identified:
Distribution and host specificity of fleas collected Rhinolophus ferrum-equinum ( Schreiber) 1775,from Europe and elsewhere were studied by Kolenati Rhinolophus hipposideros( Bechstein) 1800, Eptesicus1856), Weidner( 1937), Vermeil( 1961), Ioffand Skalon serotinus( Schreiber) 1775, Eptesicus nilsoni Keyserling1954), Hurka 1969, Peus( 1972), and others. Information et Blasius 1839, Plecotus auritus( Linne) 1758, Myotis
1The Johns Hopkins University, School of Hygiene and Public Health, Department of Immunology and InfectiousDiseases, Baltimore, 21205, USA.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 9
myotis ( Borkhausen) 1797, Myotis oxyghnatus ( TABLE 1), Ischnopsyllus octactenus and I.hexactenus
Monticelli) 1885, Myotis mystacinus ( Kuhl) 1810, occurred most often on bats( Fig. 1). The host species
Barbastellabarbastellus( Schreiber) 1775. Another five most frequented by fleas were M. mystacinus, M.
species of bats: Rhinolophus euryale Blasius, 1853; myotis, and R. ferrum-equinum( Fig. 2). In most cases,
Vespertilio murinus Linne, 1758; Vespertilio discolor a single flea specimen was found on the bats. Less often,
Matterer et Kuhl, 1819; Myotis emarginatus( Geoffroy) three to six specimens of fleas were hosted by one bat.
1806; and Miniopterus schreibersii( Kuhl) 1819, were The maximum number of fleas found on a single bat( M.
captured in caves, but they were free of fleas. mystacinus) was nine.
On hibernating bats, seven species, including twosubspecies, of fleas were found: Ischnopsyllus ( I.) DISCUSSION
intermedius ( Rothchild) 1898, lschnopsyllus ( I.)octactenus( Kolenati) 1858, Ischnopsyllus( I.)simplex Our investigations of fleas on bats was done
simplex Rothchild 1906, Ischnopsyllus ( I.) simplex exclusively in the winter period, during the bats'mysticus Jordan 1942, Ischnopsyllus ( H.) hexactenus hibernation. All hibernating bat species occurred in theKolenati) 1858, Rhynolophopsylla unipectinata form of colonies, which were found most often in large
Taschenberg) 1880, and Nycteridopsylla pentactena underground dome-like rooms. Individual bats in the
Kolenati) 1858. colony hang very close to each other, often with bodyThe most abundant flea was!. octactenus, followed contact (Fig. 3). While most bats in the colony were
by I. hexactenus, R. unipectinata, I. s. simplex, 1. motionless, hanging from the ceiling rocks, some didintermedius, I. s mysticus, and N. pectactena. Five of not fall into a deep hibernating sleep. Hibernation ofthese species of fleas occurred on four to six bat species the bat hosts may influence and possibly control
30
s 20 A,
ct Y 1ftf
CO by
8 1.
M1.: e- 1
8 10 // V — s
fY
Ai
Y. Z:
a
A i Rr}
t fiS5
y
0
Fleas
v4a
y
g 2 U H
o V 4
9VFigure 1. Quantitative occurrence of individual species of fleas on bats hibernating in caves.

10 BULL. SOC. VECTOR ECOL. JUNE, 1994
hibernation of ectoparasites. During such a quiescent bats, on one hand, is very restricted in that bat fleas areperiod the metabolism of fleas may slow down so that not found on other mammalian hosts. On the other hand,blood intake may not be required. Fleas and other the relationship is unrestricted in that most bat fleasectoparasites inhabiting the bats fur have not been found share their bat hosts.
blood fed. We noticed that during the over-winteringperiod we did not see gravid flea females. This is a rather Acknowledgmentsinteresting phenomenon that should be investigatedfurther to advance understanding of the host-parasite I thank my colleague Dr. Julius Vachold for hisrelationship during the hibernation period. It may be collaboration and friendship during the collection ofthat the flea ectoparasites survive the winter period in bats in the deep underground caves. The work reportedlower developmental stages such as larvae or pupae here was supported financially in part by the Slovakliving in droppings of guano. At the end of the hiber- Academy of Sciences, Bratislava, Slovakia, and in partnation, fleas may attach to bats passing through the cave by the Johns Hopkins University, Baltimore, Maryland,entrances. Anotherpossibility is that gravid flea females USA.
lay their eggs loosely into the host fur, from which eggsdrop onto piles of guano. Bats, instead of fleas, would REFERENCES CITED
thus be responsible for this process. Fleas may alsoover-winter as larvae and pupae in guano, and during the Hurka, K. 1956. Die Larvae des Fledermausflohesspring when bats migrate from deep underground cave Ischnopsyllus intermedius( Roths.) ( Aphaniptera:spaces back to cave entrances, newly hatched fleas may Ischnopsyllidae. Acta Soc. zool. Bohem 20: 372-hitch onto their hosts. 474.
The parasite-host relationship between fleas and Hurka, K. 1957. Prispevek systematice, faunistice a
40 - 1V
30
20 -
cu •
Y I Y G: R'.
10 - R F
0
Bats
5 h yV
hh
Z S 0 O D p,
i g OZ o Z C 0. W h
2 L.ti
k
Figure 2. Infestation of hibernating bats with fleas.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 11
JUa aO N N 00 00 ' D
dN ,0 O; NOO
Cr) DNhNMr , 0 M
laqumNmrN — 1mr M vD
snilalsvgavq v11a, svq.mg 400
snulov atu spayy M Nr, Net — 1 M
OTM
snJVuy86xo suo* N N wet • th
V
szjoCUI sliorCjtl vD N ." O et ON 00N •rs-
y
x0snjunv sn, oJald ' --. S M N
Viuosllu snolsmdq — M —
r;
044
tel0 snupojas snaLsaJdg
0
oa)
6b
g soioplsoddny snydolouyyg N — " 00 CA
ga
3 umulnba-ulniiaf snydolouNg --, in oN N vj
2a
d9. 14' H
o,4ao
aag '. u

12 BULL SOC. VECTOR ECOL JUNE, 1994-',..
4.. -
IV
4.
t
Pte' L
or
lax a*
y. 4i S','''.,7'...
oft. 4#
4 , # 4‘.,, ,
e'
er
S S ,
a
g
frex6{$
A `4, v w r
s
Figure 3. A colony of hibernating bats hanging from the cave ceiling.
bionomii netopyrich blech v CSR. Csl. parasitol. 4: Kolenati, F. A. 1856. Die Parasites der Chiropteren.145- 166. Brune.
Hurka, K. 1963x. Bat fleas ( Aphaniptera, Pens, F. 1972. Zur Kenntnis der Flo DeutschlandIschnopsyllidae) of Czechoslovakia. Contribution Insects, Siphonaptera) IV.FautlistikundOecologieto We distribution,morphology, bionomy, ecology der Saugetierflohe. Zool. Jb. Syst. 99: 408-504.and systematics. Part I. Subgenus Ischnopsyllus Rosicky, B. 1950. Predbeznykatalogblch( Aphaniptera)Westw. Acta Faun. Ent. Mus. Nat. Prague 9: 57- z uzemia Slovenska. Prirodovedny sbornik 5: 155-120. 176.
Hurka, K. 1963b. Bat fleas ( Aphaniptera, Rosicky, B. 1952. I. Dodatek k prodromu blechIschnopsyllidae) of Czechoslovakia. II. Subgenus Aphaniptera) CSR. Acta entomol. mus. national.Hectactenopsylla Oud., Genus Rhinolophopsylla Prague 28: 5- 18.Oud., Subgenus Nycteridopsylla Oud., Subgenus Vachold, J. 1956. K otazke vyskytu •a rozsireniuDinicteropsylla Ioff. Acta Univ. Carolinas- Biol. netopierov( Chiroptera) na Slovensku. Biol. prace,1963: 1- 73. 3/ 14: 1- 163.
Hurka, K. 19w. Systematic, faunal and bionomical Vermeil, C. 1961. Jordan et Rothschild 1921notes on the European and Asiatic flea species of Rhinolophopsylla unipectinata crabs Siphonapterathe family Ischnopsyllidae ( Aphaniptera). Acta nouveau pour la Tunisie. Ann. Parasit. Hum. Com.Univ. Carolinas- Biologics: 11- 26. 36: 168.
Ioff, I. G. and O. I. Skalon. 1954. Opredelitel bloc Weidner, H. 1937. Beitrag zur Kentnis derBiologie desvostocnoj Sibirii,DalnegoVostokaiprilegajuscich Fledennausflohes Jschnopsyllus hexactenus Kol.rajonov. Medgiz, Moskva, 275 pp. Z. Parasitenk. 9: 543- 548.

BULL. SOC. VECTOR ECOL., 19( 1): 13- 17 JUNE, 1994
HOST-PARASITE RELATIONSHIPS BETWEEN FLEAS (SIPHONAPTERA)
AND SMALL MAMMALS OF THE TATRAS MOUNTAINS IN SLOVAKIA
M. Trpisl
ABSTRACT: Relationships between fleas and small mammals were investigated in two mountain ranges of
Slovakia, High Tatras, and Low Tatras. In the High Tatras, 195 flea specimens were collected from 272 smallmammals. Fleas collected from nine species of small mammals (Sorex araneus, Sorex alpinus, Neomys fodiens,
Neomys anomalus, Neomys a. milleri, Apodemus flavicollis, Clethrionomys glareolus, Microtus arvalis, and
Microtusnivalis) belonged to 12 species( Hystrichopsylla talpae, Ctenophthalmus bisoctodentatus, Ctenophthalmus
agyrtes, Ctenophthalmus assimilis, Doratopsylla dasycnema dasycnema, Palaeopsylla soricis rosickyi, Palaeopsylla
similis, Peromyscopsylla silvatica, Peromyscopsylla fallax, Malaraeus penicilliger, Malaraeus arvicolae, and
Megabothris turbidus). The most abundant fleas in the High Tatras were D. d. dasycnema, P. s. rosickyi, and C.agyrtes, and small mammals most often infested with different species of fleas were, N.fodiens, S. araneus, and S.
alpinus. In the Low Tatras, 98 specimens of small mammals belonging to three species ( Apodemus sylvaticus,Apodemusflavicoll is, and Neomysfodiens) were captured in spring- loaded traps. One hundred and fifteen specimensof fleas belonging to seven species( Hystrichopsylla talpae, Ctenophthalmus agyrtes, Doratopsylla d. dasycnema,Palaeopsylla soricis, Palaeopsylla similis, Palaeopsylla steini, and Nosopsyllus fasciatus) were collected. In the
Low Tatras, the small mammal infested with fleas was N.fodiens.
INTRODUCTION MATERIAL AND METHODS
Before this survey was undertaken in 1965, the flea Study Areafauna of the High Tatras had been studied by only a few The High Tatras are located in northern Slovakia in
investigators( Jordan 1932, Rosicky 1950, 1955, 1957a, the western Carpathian Mountains. The entire area is
Jurik 1955), and elsewhere in Europe by Wagner( 1936) included in the Tatra National Park. It is an area of
and Pens ( 1972). Research on the distribution and natural beauty with picturesque mountains, numerousecology of fleas of the High Tatras intensified after the mountain lakes, and streams traversing coniferous1970' s ( Mrciak and Rosicky 1970, 1975; Ryba and forests. The highest peak of the High Tatras mountain
Rosicky 1975; Cyprich et all. 1976; Ryba 1977. The range is Gerlach ( 2,633 m above sea level). Manystudies by Cyprich et al.( 1976) and Kiefer and Cyprich thousands of visitors come to the Tatra National Park
1984) concentrated on the ecology of such specific each year for recreational purposes as well as for health
hosts as Cricetus, Arvicola, Rattus, and Sorex and on reasons, mainly for the treatment of tuberculosis. Mosttheir association with flea ectoparasites. On the Polish towns in the High Tatras are located at intermediate
side of the Slovak High Tatras Mountains, fleas were elevations ( up to 1, 300 m). The park offers manyinvestigated by Bartkowska( 1973). diverse biotopes for small mammals, hosting various
The purpose of this investigation was to determine flea species. The small mammals used in this study werethe host-parasite relationships between small mammals captured in the High Tatras mainly at intermediateand fleas in the mountains of High Tatras and Low elevations up to 1, 500 m. Only a few were collected atTatras in the areas most often visited by tourists to higher elevations ( above 1, 500 m), where the onlyexamine the possible association of fleas with humans human settlements are isolated cottages constructed
and to compare the flea fauna on small mammals of primarily for use by mountain climbers and skiers.High Tatras with flea fauna of Low Tatras. In the LowTatras Mountains, the Demanova Valley
The collection of fleas included in this study has opens to the Vah River Valley near the city ofLiptovskybeen in the custody of the Slovak National Museum in Mikulas. The highestpeaks of the LowTatrasMountains
Bratislava since 1968. are Chopok( 2,025 m above sea level), Dumbier( 2,045
1The Johns Hopkins University, School of Hygiene and Public Health, Department of Immunology and InfectiousDiseases, Baltimore, Maryland, 21205, USA.

14 BULL. SOC. VECTOR ECOL. JUNE, 1994
m above sea level), and Poludnica( 1, 550 m above sea ApodemussylvaticusLinne 1759, Microtus arvalis Pallaslevel). The slopes of the Demanova Valley consist of 1778, Microtus agrestis Linne 1761, Microtus nivalislimestone and contain several limestone caves. Severe mirhanreini Schiefer 1935, Pitimys tatricus Kratochvil
winters and cool summers characterize its climate. The 1952, PitimyssubterraneusDeselys-Longchamps 1836,mean temperature remains below freezing from the and Clethrionomys glareolus Schreber 1780. Six speciesmiddle of November until the middle of March. Total ofsmall mammals captured, S.minutus, M.avelanarius,annual rainfall is between 1, 000 and 3, 000 mm, peaking A. sylvaticus,M.agrestis, P. tatricus, andP.subterraneusin June. were free of fleas, therefore werenot included in TABLE
1.
Captures ofSmall Mammals and Collections ofFleas Twelve species of fleas were caught in the HighSmall mammals were caught in spring traps baited Tatras on nine species of mammals: Hystrichopsyslla
either with bacon or with wicks impregnated with a lard talpae ( Curtis) 1826, Ctenophthalmus ( C.)and fried-onion mixture. The traps were set in vegetation bisoctodentatus Kolenati 1836, Ctenophthalmus ( C.)areas, under shrubs and trees, in the late afternoon, and agyrtes ( Heller) 1896, Ctenophthalmus ( E.) assimilis
the mammals collected the next morning. Small ( Taschenberg) 1880, Doratopsylla dasycnemamammals caught in traps were collected before sunrise dasycnema ( Rothschild) 1897, Palaeopsylla soricisto prevent the fleas from escaping from their dead hosts. rosickyi Smit 1960, Palaeopsylla similis Dampf 1910,Captured mammals were removed from the traps and Peromyscopsylla silvatica ( Meinert) 1896,placed in clear plastic bags, which were sealed. The Peromyscopsylla fallax( Rothschild) 1909, Malaraeusbags with mammals were then taken to the field station ( A.)penicilliger( Grube) 1852, Malaraeus( A.) arvicolaelocated near the entrance to the Demanova caves, where ( Ioff) 1950, and Megabothris turbidus ( Rothschild)all parasites were removed from the mammals; air was 1909. Associations between fleas and their host areblown on a specimen in order to detect and collect listed in TABLE 1. The most common flea on small
moving fleas from its fur. The fleas collected from each mammals in High Tatras appeared to be Doratopsyllamammal were placed into separate test tubes and dasycnema; the next two most common flea speciespreserved in 70 percent ethyl alcohol. At the laboratory were P. s. rosickyi and C. agyrtes. The most flea-of the Slovak Academy of Sciences in Bratislava, the infestedhosts were N.fodiens,S. araneus, and S. alpinus.fleas were cleared in phenol-xylene and mounted on It is interesting to note that C. agyrtes shared threeslides in Canada balsam. The fauna of small mammals species of small mammalian hosts, D. dasycnema, fourof the High Tatras has been previously described by species, P. soricis, four species, and M. turbidus, threeFeriancova-Masarova and Hanak( 1965). species( TABLE 1). It is also important to observe that
All collections of small mammals in the Low Tatras S. araneus harbored three species of fleas, S. alpinusMountains took place in the Demanova Valley along the three species, andN.fodiens seven species; N.anomalusDemanovka River ( 19°40' E; 49° 05' N) from 1956 to milleri, C.glareolus, and M.nivalis were each associated1959. The 1956 collections were done in August, the with two species of fleas( TABLE 1).
1957 ones in March and May, and the 1959 ones in In the Low Tatras, 115 flea specimens were collected
January. The keys by loff and Skalon ( 1954) and from 98 small mammals belonging to five species:Rosicky( 1957b) were used for the identification of fleas Sorex araneus, N.fodiens, N. a. milleri, A. flavicollis,in this study. and A. sylvaticus( TABLE 2).
The following seven species of fleas were found inRESULTS AND DISCUSSION the Low Tatras: Hystrichopsylla talpae, C.( C.) agyrtes,
D. dasycnema, P. soricis,P. similis,Palaeopsylla steiniIn the High Tatras, collections of small mammals Jordan 1932, and Nosopsyllus fasciatus (Bose) 1801.
and fleas were done from June to September in 1957 and Palaeopsylla similis appeared to be the predominantin 1958. During that period, 195 fleas were collected species. It accounted for over 78 percent of the fleafrom 272 captured small mammals. Fourteen species population of the Demanova Valley. Doratopsyllaand one subspecies of small mammals were captured dasycnema, C. agyrtes, and P. soricis accounted for 8.7and checked for flea ectoparasites in the High Tatras: percent, 7.8 percent, and 2.6 percent, respectively.Sorex araneus Linne 1758, Sorex minutus Linne 1766, Hystrichopsylla talpae, P. steini, and N. fasciatusSorex alpinus Schnitz 1837, Neomys fodiens Schreber occurred for less than 1 percent. The most flea-infested1777, Neomys anomalus Cabrera 1907, Neomys small mammal was N.fodiens.
anomalusmilleriMottaz 1907, Muscardinus avelanarius Hystrichpsylla talpae, C. agyrtes, P. soricis, andLinne 1758, Apodemus flavicollis Melchior 1834, P. similis, occurred in both the High Tatras and Low
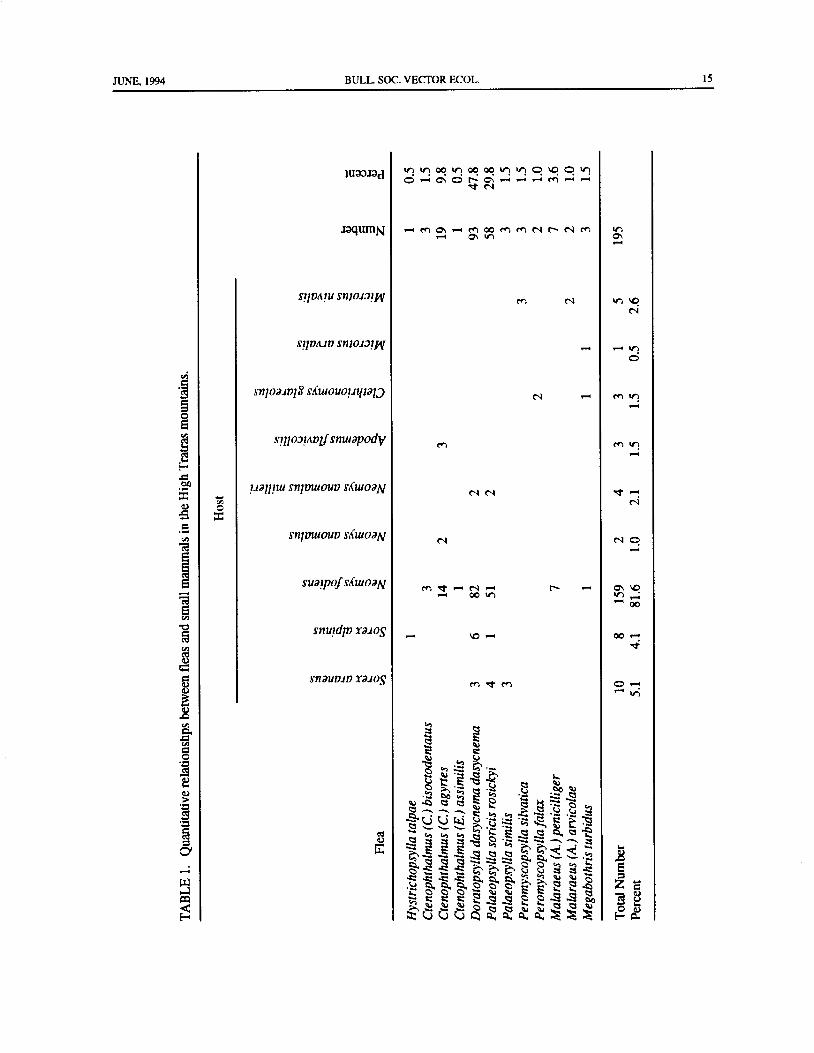
JUNE, 1994 BULL. SOC. VECTOR ECOL. 15
Juaandv, v; oov, o00ov, v! o4OOto
C5 . o oroir.., r: .-: M ,-: ,.. 1
et N
JacpunN '-. m oN . gl, oc m M N t- N M
sijnniu SnJOJOJyy r, N oN
silvtuV SnJOJdlJtlN
O
vi
isnioainjX sdwouowpao N r, r, v,
0
5
gsal/oa.z tv fsnuiapodd r, M , r,
E.
iJal/ Iui snimuoun sICuWoaN N N et r-,
h N
o.,
snimuoun sXwoaN N N o
suamof s,CuioaN Met N on Ooo in in
o0
snuzdjn raJos
a) SnaunJn xaJOs K, M o
42
azo)
oo V
v
2
ot .
y V CO 0>
t CS O yCS - - - y y 'r '•• •G V
N 3Cf
h •y 'r' —. -•,
3
4per' ^ i- 4, 1% t8i 8 ' a `A • tC y '. i o> o>
l C O O4o, Oao 0 0 0 E 2 2 '° Z o;cciQ O C! CS oa CU
CI I:" o au
E-, UUC) ga, a, a, a, E. au

16 BULL. SOC. VECTOR ECOL. JUNE, 1994
TABLE 2. Quantitative relationships between fleas and small mammals in the Low Tatras mountains.
Host
h a.
Flea lIlaHystrichpsylla talpae 1 1 0.9Ctenophthalmus agyrtes 2 1 6 9 7.8Doratopsylla dasycnema dasycnema 10 10 8. 7Palaeopsylla soricis 3 3 2.6
Palaeopsylla similis 90 90 78.2Palaeopsylla steini 1 1 0.9Nosopsyllus fasciatus 1 1 0.9
Total Number 3 1 111 115
Tatras. Doratopsylla dasycnema, P. steini, and N. This investigation was financially supported inpartfasciatus occurred only in the Low Tatras. Apodemus by the Slovak Academy of Sciences in Bratislava,sylvaticus was free of fleas in the High Tatras but was Slovakia and in part by the Johns Hopkins University inparasitized by C. agyrtes and N.fasciatus in the Low Baltimore, Maryland, USA.Tatras.
The most frequently parasitized host in bothmountains was N.fodiens with seven species of fleas in REFERENCES CITEDthe High Tatras and six species in the Low Tatras.
All areas of the High Tatras and Low Tatras Bartkowska, K. 1973. Siphonaptera Tat/ Polskich.mountains are accessible to visitors for hiking,mountain Frag. Faunistica 19: 277-283.climbing, and skiing. Public facilities such as hotels, Cyprich, D., M. Kiefer, and M. Kminiak. 1976. Blchyshops, and restaurants are located in elevations up to Siphonaptera) krysy vodnej( Arvicola terestris L.,1, 500m above sea level. Many tourists visit these places 1758) v podmienkach Slovenska. Biologia,and hike in the higher elevations. People are occasionally Bratislava 31: 573- 581.bitten by the human flea Pulex irritans Linne 1858 or by Feriancova-Masarova, Z.and V. Hanak. 1965. Cicavce.the cat and dog flea Ctenocephalides fells ( Bouche) Stavovce Slovenska, 4, Vydavatelstvo SAV,1835 or Ctenocephalides canis ( Curtis) 1826, Bratislava.
respectively, but neither of these species occurred in our Ioff,I.G. and O. I.Skalon. 1954. Akey for Identificationcollections of fleas from small mammals orhumans. So of Fleas of Eastern Siberia, Far East and Adjacentfar, no flea-borne pathogens have been reported from Territories. USSR Academy of Sciences, Medgiz,the Tatras. Moscow, pp. 275.
Jordan, K. 1932. Siphonaptera collected by HerrAcknowledgments Georg Stein in the High Tatras. Nov. Zool., 38:
261- 263.I thank Dr.Julius Vachold and Ing. Jan Mituch with Jurik, M. 1955. Prispevokk poznaniublch( Aphaniptera)
whom I had an opportunity tocollect the small mammals. CSR. Acta univ. agric. et silvicult., Brno. 2: 175-I enjoyed and greatly appreciated their friendship and 180.enthusiasm for this research. Kiefer,M. and D. Cyprich. 1984. Blchy( Siphonaptera)

JUNE, 1994 BULL. SOC. VECTOR ECOL. 17
potkana obycajneho ( Rattus norvegicus Berken- Rosicky, B. 1955. Aphanipterofauna Vysokych Tatierbout) v podmienkach Slovenska. ActaF.R.N. Univ. s poznamkami o vyskovem rozvrstveni blech v
Comen.- Formatio et Protectio. Nature 10: 39- 54. tomto horstvu. Zool. ent. listy 4: 365- 383.Mrciak, M. and B. Rosicky. 1970. Ekologicke studie o Rosicky, B. 1957a. Nektere zoogeograficky vyznamne
nidikolnej faune drobnych cicavcov vo Vysokych nalezy blech ( Aphaniptera) s drobnych ssavcu zTatrach. Zbornik Narodne parky, bohatstvo uzemi CSR. Ceskoslovenska parasitologie 4: 291-
civilizacie. Proceedings, TANAP 360- 383. 298.
Mrciak, M. and B. Rosicky. 1975. Some features of the Rosicky, B. 1957b. Blechy- Aphaniptera. Fauna CSRzoogeography of high altitude parasites of birds sv. 10, CSAV, Prague, 439 pp.and mammals on the example of High Tatra Ryba, J. 1977. Blechy drobnych savcu a jejich hnizd vmountains ( Slovakia). Biologia ( Bratislava) 30: Tatrach. Zpravy Csl. zool. spol. 10- 12: 51.589-597. Ryba, J. and B. Rosicky. 1975. Rhadinopsylla
Peus, F. 1972. Zur Kenntnis der Flohe Deutschland ACtenophthalmus) mesoides Smit, 1957
Insecta, Siphonaptera) IV.Faunistik und Oecologie S iphonaptera, Hystrichopsyllidae) in Czecho-
der Saugetierflohe. Zool. Jb. Syst, 99: 408- 504. slovak western Carpathians. Foliaparasitol.( Praha)
Rosicky, B. 1950. Predbezny katalog blch( Aphaniptera) 22: 379- 383.
z uzemia Slovenska. Prirodovedny sbornik 5: 155- Wagner, J. 1936. Flohe. Die Tierwelt Mitteleuropas,
176. Leipzig 17: 1- 24.

BULL. SOC. VECTOR ECOL., 19( 1): 18- 22 JUNE, 1994
HOST SPECIFICITY AND ECOLOGY OF FLEAS (SIPHONAPTERA)
OF SMALL MAMMALS IN MOUNTAINS OF NORTH-CENTRAL SLOVAKIA
M. Trpisl
ABSTRACT: To determine the host-specificity and ecology of individual species of fleas, small mammals, mostlyrodents, were collected in spring traps at various biotopes in the the Slovak Beskyds, Oravian Magura, and the SmallFatra mountains of north-central Slovakia. In the Orava region, 393 fleas were collected from 14 species of smallmammals. Pulex irritans was only occasionally collected from humans. The preferred hosts of fleas wereClethrionomysglareolus,Apodemusagrarius,andMicrotusarvalis. The predominant flea species was Ctenophthalmusagyrtes. In the Small Fatra Mountains, six species of small mammals were collected in the Vratna and Stefanovavalleys, where 13 species of fleas were collected. The most common flea species were Ctenophthalmus ( C.)
bisoctodentatus, Ctenophthalmus( C.) agyrtes, and Doratopsylla dasycnema. The host species most frequently andextensively infested by fleas was Neomysfodiens, but the largest number of fleas was found in mole nests.
INTRODUCTION MATERIALS AND METHODS
The host specificity, ecology, and geographical Small( 5 x 10 cm), mechanical, spring- loaded rodentdistribution of fleas have been studied by medical traps baited with bacon were used to capture smallentomologists and parasitologists in various European mammals. Traps were placed in low vegetation, undercountries ( Wagner 1930, 1936; Peus 1972; loff and shrubs and trees, along river banks, as well as in humanSkalon 1954). The host specificity of fleas in the former habitations. Traps were set in the late afternoon andCzecho-Slovakia was studied by Rosicky 1950a, 1950b, were collected before sunrise the nextmoming to prevent1955, 1957a; Jurik 1960; Hurka 1963a, 1963b; Cyprich fleas from leaving the dead animals. Each capturedand Kiefer 1975; Cyprich et al. 1976; Ryba 1977; Kiefer animal was placed in a separate plastic bag to preventand Cyprich 1984; Dudich 1987; Cyprich 1989; Trpis, mixtureorlossofectoparasites. Small mammals trapped1994a; 1994b. in the Orava region were then brought to the Slovak
Before this investigation in Small Fatra in 1955- Academy ofSciences Field Experimental Station located1956, and in the Orava region in 1958- 1959, no flea in the village of Slanicka Osada. Fleas were collectedsurveys had been carried out, although some other from the small mammals by blowing air into the fur ofregions had been occasionally explored, primarily by the captured mammals with ahair-dryer. Fleas collected
Rosicky ( 1950b, 1957a). In this study, fleas were from individual animals were recorded and preserved incollected from mole nests in Small Fatra. Additional 70 percent ethyl alcohol. O' Mahony' s methodinformation on the association of fleas with mole nests ( unpublished) was used to process and mount fleas onwas published by Rosicky( 1957a) and Junk( 1968). microscope slides in Canada balsam. Briefly, fleas
The goals of this study were to determine the preserved in 70 percent ethyl alcohol were placed intocommon species of fleas prevalent in this area, to 10 percent KOH for at least 12 hrs, washed in distilledevaluate their association with small mammals and their water for 1 min., placed into glacial acetic acid for 15possible association with humans, and to comment on min., cleared in xylene saturated with phenol for 15 min.the potential of fleas in the transmission of zoonotic to 12 hrs or longer, depending on intensity ofdiseases to humans. This study was initiated in 1958 pigmentation, and washed in xylene for 15 min. Thewhen the authorwas associated with the Slovak Academy treated fleas were placed into a few drops of Canadaof Sciences. The entire flea collection was deposited in balsam diluted with xylene and covered with a coverthe Slovak National Museum in Bratislava in 1968. slip. After the slides had dried, the fleas were examined
1The Johns Hopkins University, School of Hygiene and Public Health, Department of Immunology and InfectiousDiseases, Baltimore, Maryland, 21205, USA.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 19
under a compound microscope and identified to species Krasna Horka, Sucha Hora, Krusetnica, Podbiel, Tichausing the keys developed by Wagner( 1930, 1936), by Dolina, Zverovka, and Oraysky Podzamok.Ioff and Skalon( 1954), and by Rosicky( 1957b).
In the Small Fatra region, thirty mole nests of Talpa RESULTS
europea, were additionally dug out during the fall andwinter. In the laboratory, fleas were isolated in Berlese Host Specificity of Fleas from the Orava Regionfunnels from the nesting material and processed as In the Orava region, 393 fleas belonging to 17above. species and 11 genera were collected from 551 small
mammals( TABLE 1). The following flea species wereDescription of the Study Area found associated with various species ofsmall mammals:
The Orava is an elevated basin located in northern Ctenophthalmus agyrtes Heller 1896 ( collected fromSlovakia, between 19° 10'- 1945' E and 49°06'- 49° 38' N. nine species of small mammals and representing 58.5%The Orava River flows through the central valley of the total fleas collected), Palaeopsylla soricis rosickyiadjoining the Vah River near Kralovany city at its Smit 1960 ( collected from seven species of smalllowest point, 430 m above sea level. The Slovak mammals and representing 8.4% of the total fleaBeskyds Mountains, located in northern Slovakia,belong collection), Doratopsylla dasycnema dasycnemato the third-highest mountain range in Slovakia. Babia Rothschild 1897 ( collected from seven species ofHora Mountain is the highest peak, rising 1, 725 m above mammals and representing 8. 1% of the total fleasea level. The next highest mountain range in this area collection), Megabothris turbidus Rotschild 1909isOrayskaMagura, with the highestmountain, Kubinska ( collected from six species of small mammals andHola, rising 1, 345 m above sea level. The Skorusinske making up about 5% of the total), and LeptopsyllaMountains, divided by the Studeny Potok creek and the segnis Schonherr 1816( collected from four species ofOravica River, constitute the third highest mountain small mammals, and representing about 5% of the total
range in the area. The highest mountain in the latter fleas). Palaeopsylla similisDumpf 1910 andMalaraeusrange is Skorusina, 1, 312 m above sea level. penicilligerGrube 1852parasitized fourand three species
Small Fatra is an integral part of the mountains that of small mammal hosts, respectively, and constitutedextend northward from the Vah River, between the 3. 8 percent and 2.5 percent of the total flea collectionvillage of Strecno and the city of Vrutky. It forms the ( TABLE 1). The remaining 10 flea species occurred inouter part of Karpathian Mountains. The geologic low numbers and parasitized only a few small mammals.layers of granite alternate with flint, limestone, and Fleas were collected from 14 species of smalldolomite layers. In the limestone crust of various mammals, belonging to nine genera, and from humans.thicknesses and densities, water created valleys at Besides rodents, which constituted the largest group ofdifferent elevations. flea hosts( nine species), a small group of insectivores
In mountainous regions of Slovakia, coniferous ( three species) was studied, some of which hostedand deciduous forests alternate with mountain meadows numerous flea species. The most frequently flea-infestedand agricultural fields. Slovakia also has a rich fauna of mammal was Apodemus sylvaticus( 24.2%), followedsmall mammals that inhabit a variety of biotopes by Apodemus agrarius ( 12. 5%), Microtus arvalisFeriancova-Masarova and Hanak 1965) and support ( 12.5%), and Sorex araneus ( 9.4%), which was the
an abundant fauna of ectoparasites, among which are insectivore most often infested( TABLE 1). Fleas werefleas. collected only sporadically from the remaining mammal
The study sites were located in the mountains, hosts.
valleys, meadows, and ecotones between forests andagricultural developments, and along brooks and rivers Host Specificity of Fleas in the Small Fatra Regionas well as in villages in the Orava ( Slovak Beskyds In the Small Fatra region, 270 fleas were collectedMountains; Slovenske Beskydy) and Terchova( Small from 104 small mammals during the spring. TheFatra Mountains; Mala Fatra) regions. In the Orava following species of small mammals were captured inregion, small mammals and their fleas were collected spring- loaded traps or caught alive in the Small Fatraduring the winter, spring, and summer months in 1958 region: Talpa europea, Sorex araneus, Sorex minutus,and 1959. Small mammals were trapped at three Neomys fodiens, Apodemus flavicollis, and
elevations( mountains, foothills, and lowlands in valleys) Clethrionomys glareolus.at Polhora, Slana Voda, Rabca, Zubrohlava, Bobrov, Thirteen species of fleas were collected on sixNamestovo, Slanicka Osada, Vavrecka, Tapesovo, species of small mammals( TABLE 2). The dominantLokca, Orayska Lesna, Hrustin, Tvrdosin, Oravice, species of flea, Ctenophthalmus ( C.) bisoctodentatus

20 BULL. SOC. VECTOR ECOL. JUNE, 1994
luaalad 60N6 pNO0~6 Ooci0; 66 6 veNiit 6 ,-;.- 6un
1 O mr .- 1 N K1 V.) N N M ap o .„-, ON VD M MJag1II[Ix N
M M N -- N M O
OWOH N M v1 Ni
SlyvftJV SnJOl31111 l M N M ON WI
N
SI.l1Saa? J slalsala1 vjoapuy o o .,
v
snwoalv18auwououyp M — N M N in en LnM
snouv t Cs snuiapody n ,, N M ten N
4o sipodmvlfsnuuapody — 0 D
N —.uncn
w.0sm 1v18v snuiapody A N M N • . N — ON u•
00a.c4 snjnosnui sny/ n o` M N M
8 0x vinpanu siCtuo,GQ N N VD ,,,a)
snlrnuouv siCluoaN N try
C 0
cgsuaIpofsXu/ oaN N I
N
snuidiv xa.loS r` er 00
y swum rams I N un5 cb
1 snauvav xawg 00 00 ., O N e'
6 1 ,.4 M •CTG
it vadoina vdms N N ,
8 O
O
o 1g
yCS
V h C tn ,:: ygy
O r_ CL V
U C W W . gct zZ' v .p .t • o% %
s y y y y o •i 'v . E ts v
a cccg > ao zF C.) U0i0q0... at. ra. c4 la. .• z Q. E- 0 CL

JUNE, 1994 BULL. SOC. VECTOR ECOL. 21
Table 2. Quantitative relationship of flea species and their mammalian hosts in the Small Fatra Mountains ofSlovakia.
Host
honov 1e
V Op
o d o
Uoe
FleaI 4
Hystrichopsylla lalpae 6 6 2.2Ctenophthalmus bisoctodentatus 104 1 105 38. 9Ctenophthalmus agyrtes 2 1 3 84 2 92 34. 1Ctenophthalmus assimilis 2 2 0. 7Ctenophthalmus congener 1 1 2 0.7Doratopsylla dasycnema 2 2 17 21 7. 8Palaeopsylla soricis rosickyi 2 18 20 7.4Palaeopsylla similis 5 4 9 3. 4Palaeopsylla kohauti 2 2 0.7Palaeopsylla steini 3 2 5 1. 9Peromyscopsylla falax 1 1 0.4Peromyscopsylla bidentata 3 3 1. 1Megabothris turbidus 1 1 2 0.7
Total Number 121 3 2 48 87 9 270Percent 44.8 1. 1 0.8 17. 8 32.2 3. 3
Kolenati 1836, was most often associated with mole that live in underground burrows and roam on thenests( TABLE 2). The next most prevalent flea species ground in shrubberies may to some degree share thewas Ctenophthalmus( C.)agyrtes Heller 1896, followed same types of habitats. Similarly, one small mammalbyDoratopsylla dasycnema dasycnema Rotschild 1897, species may host several species of fleas.and Palaeopsylla soricis rosickyi Smit 1960. The When comparing flea species associated with theirremaining species occurred in lower densities ofbetween mammalian hosts in the two explored mountain ranges,3.4 percent and 0.4 percent. The largest number of it is interesting to see that those species that werefleas( 121) was collected from mole nests. The largest abundant in the Orava region may also be abundant innumber of flea species( seven) was found to be parasitic Small Fatra, e. g. C. agyrtes and D. dasycnema. On theon Neomys fodiens( TABLE 2). Six flea species were other hand, T. europea in the Orava region wasfound on both T. europea and C. glareolus. parasitized only by one flea species, while in the SmallCtenophthalmus agyrtes andD. d.dasycnema occurred Fatra Mountains several species of fleas occur on T.on five and three mammalian hosts, respectively( TABLE europea. Neomys fodiens and C. glareolus were hosts2). of many species of fleas in both areas.
A lack of host-specificity in some species of fleasDISCUSSION may increase the potential for acquisition and
interspecific transmission of pathogens among wildMost flea species were found on several different animals. Some zoophilic flea species, moreover, may
small mammals, an indication that the small mammals transmit pathogens among wild animals and become

22 BULL. SOC. VECTOR ECOL. JUNE, 1994
indirectly involved in the transmission ofcertain zoonotic Ioff,I.G. and O. I. Skalon. 1954. A key for identificationdiseases. In this respect, further studies of relationships of fleas of Fastern Siberia, Far East and adjacent
among wild animals, fleas, and human hosts would be territories. USSR Academy of Sciences, Medgiz,important. Moscow, 275 pp.
Jurik, M. 1960. Aphaniptera drobnych cicavcov
Acknowledgments Badinskeho pralesa. Biologia, Bratislava 17: 383-
387.
The friendship extended by Dr. Julius Vachold, Jurik, M. 1968. Fleas of the mole Talpa europea L. in
Ing. Jan Mituch, and Miss Ludmila Tonkovic and their Czechoslovakia ( Aphaniptera). Acta. ent.
help in collecting the small mammals are greatly bohemoslovaca 65: 67- 75.
appreciated. The work reported here was fmancially Kiefer,M.and D. Cyprich. 1984. Blchy( Siphonaptera)supported in part by the Slovak Academy of Sciences, potkana obycajneho ( Rattus norvegicus
Bratislava, Slovakia, and in part by the Johns Hopkins Berkenhout) v podmienkach Slovenska. Acta
University, Baltimore, Maryland, USA. F.R.N. Univ.Comen.- FonnatioetProtectioNature
10: 39- 54.
REFERENCES CITED Peus, F. 1972. Zur Kenntnis der Flohe Deutschland
Insecta, Siphonaptera) IV.FaunistikundOecologie
Cyprich, D. 1989. Revision and distribution of the der Saugetierflohe. Zool. Jb. Syst. 99: 408- 504.
specific fleas ( Siphonaptera) of European suslik Rosicky, B. 1950a. Biocenosy a ekologieblech stredniCitellus citellus ( L.) the Genus Citellophillus Evropy. Vest. csl. zool. spol. 14: 97- 148.Wagner. Annot. Zool. Bot., Bratislava 194: 1- 48. Rosicky, B. 1950b. Predbezny katalog blch ( Apha-
Cyprich, D. and M. Kiefer. 1975. Siphonapterofauna niptera) z uzemia Slovenska. Prirodovedny sbornikhniezd chrcka rolneho( Cricetus cricetus L., 1758) 5: 155- 176.
v obdobi jeho premnozenia na vychodoslovenskej Rosicky, B. 1955. Aphanipterofauna Vysokych Tatternizine. Biologia, Bratislava 30: 599- 604. s poznamkami o vyskovem rozvrstveni blech v
Cyprich, D., M. Kiefer, M. Kminiak. 1976. Blchy tomto horstvu. Zool. ent. listy 4: 365- 383.Siphonaptera) krysy vodnej( Arvicola terestris L., Rosicky, B. 1957a. Nektere zoogeograficky vyznamne
1758) v podmienkach Slovenska. Biologia, nalezy blech ( Aphaniptera) s drobnych ssavcu zBratislava 31: 573- 581. uzemi CSR. Ceskoslovenska parasitologie 4: 291-
Dudich, A. 1987. Synuzie blch( Siphonaptera) piskora 298.
vrchovskeho ( Sorex alpinus) Schinz, 1837 v Rosicky, B. 1957b. Blchy( Aphaniptera) CSR. FaunaZapadnych Karpatoch. Biologia ( Bratislava) 42: CSR, Praha, 439 pp.603-616. Ryba, J. 1977. Blechy drobnych savcu a jejich hnizd v
Feriancova-Masarova,Z. and V. Hanak. 1965. Cicavce. Tatrach. Zpravy Csl. zool. spol. 10- 12: 51.Stavovce Slovenska, 4, Vydavatelstvo SAV, Trpis, M. 1994a. Host-parasite relationships between
Bratislava. fleas ( Siphonaptera) and small mammals of the
Hurka, K. 1963a. Bat Fleas ( Aphaniptera, Ischno- Tatras mountains in Slovakia. Bull. Soc. Vect.
psyllidae) of Czechoslovakia. I. Subgenus Ecol. 19( 1): 13- 17.
Ischnopsyllus Westw. Acta Faun. Ent. Mus. Nat., Trpis, M. 1994b. Host-parasite relationships between
Prague 9: 57- 120. fleas ( Siphonaptera) and bats ( Chiroptera)
Hurka, K. 1963b. Bat Fleas ( Aphaniptera, hibernating in ice and limestone caves in Slovakia.Ischnopsyllidae) of Czechoslovakia. II. Subgenus Bull. Soc. Vect. Ecol. 19( 1): 8- 12.
Hexactenopsylla Oud., Rhinolophopsylla Oud. Wagner, J. 1930. Katalog der palaearktischenSubgenus Nycteridopsylla Oud., Subgenus Aphanipteren, Wien, 55 pp.Dinycteropsylla Ioff. Acta Universit. Carolinae- Wagner, J. 1936. Flohe. Die TierweltMitteleuropas 17,
Biologica 1: 1- 73. Leipzig, 24 pp.

BULL. SOC. VECTOR ECOL., 19( 1): 23- 36 JUNE, 1994
VECTOR COMPETENCE OF WESTERN EUROPEAN MOSQUITOES FOR
ARBOVIRUSES: A REVIEW OF FIELD AND EXPERIMENTAL STUDIES
J. O. Lundstroml
ABSTRACT: In Western Europe, seven mosquito-borne arboviruses of the genera Alphavirus, Flavivirus, and
Bunyavirus have been described. The Tahyna subtype of California encephalitis virus, Ockelbo subtype of Sindbis
virus, and West Nile virus cause relatively mild diseases in humans, while the Inkoo subtype of Californiaencephalitis virus, Sindbis virus, Lednice virus, and Batai virus have not been associated with human disease inWestern Europe. Tahynavirus has been isolated from mosquitoes of five genera. TwoAedes and one Culiseta specieswere shown to be competent experimental vectors. Field and experimental studies indicated that Aedes vexans was
the main vector. Ockelbo virus has been isolated only in northern Europe from mosquitoes of three genera. TwoCulex and two Aedes species were shown to be competent experimental vectors. Field and experimental studies
showed that Culex torrentium was the main enzootic vector, while several Aedes species may transmit the virus fromviremic birds to humans. West Nile virus has been isolated from south European mosquitoes of three genera. Aedesand Anopheles species were found to be competent experimental vectors. However, field studies implicated Culex
modestus as the main vector. Lednice virus has only been isolated from Cx. modestus in Slovakia. This species wassusceptible to experimental infection but failed to transmit the virus. Inkoo virus has only been isolated from northEuropean mosquitoes, while Batai virus has been isolated from north and central European mosquitoes. Neither of
these two viruses have been experimentally studied in mosquitoes.
INTRODUCTION generalized malaise. Symptoms associated with Tahyna
virus infection also include cough, pharyngitis,
Clinical Importance ofMosquito-Borne Arboviruses conjunctivitis, headache, nausea, anorexia, acute arthritis,
in Europe abdominal pain, generalized weakness, and acute central
Seven mosquito-borne arboviruses or subtypes have nervous system illness. It has been shown that everybeen isolated from arthropods sampled in Western seventieth case of febrile illness and every fifth case ofEurope ( TABLE 1). The Tahyna, Ockelbo and West central nervous system illness in children living inNile viruses cause relatively mild diseases in humans Slovakia have been caused by Tahyna virus infectionBardos et al. 1980, Brummer-Korvenkontio and ( Bardos et al. 1980).
Kuusisto 1984, Espmark and Niklasson 1984, The Ockelbo subtype of Sindbis virus has been
Kolobukhina et a1. 1990, Lvov et al. 1982, Niklasson et associated with outbreaks of arthralgia and rash in
a1. 1984, 1988, Skogh and Espmark 1982), while Inkoo humans in Sweden, Finland, Russia, and Norwayvirus, Sindbis virus, Lednice virus, and Batai virus have ( Brummer-Korvenkontio and Kuusisto 1984, Espmark
not been associated with human disease in Western and Niklasson 1984, Hoddevik unpublished information,
Europe.However, arecentstudy suggested an association Lvov et al. 1982, Skogh and Espmark 1982, LundstrOm
between a malaria- like syndrome and Batai virus et al. 1991). The illness has a sudden onset with fever
infection in patients from Sudan( Nashed et al. 1993). and maculopapular exanthema distributed over the trunk
Human clinical cases caused by Tahyna virus have and limbs, and pain in the small joints of hands and feet
been reported from Slovakia( Bardos et al. 1975, 1980, and the larger joints of arms and legs ( Skogh and
Simkova and Sluka 1973) and Russia( Kolobukhina et Espmark 1982, Espmark and Niklasson 1984). The rash
al. 1990). The clinical picture of the infection is a febrile lasts for one day to three weeks, and is associated withillness with marked respiratory involvement and rare itch in 6 percent of patients. The joints affected were
central nervous system involvement( Grimstad 1988). ankle in 62 percent ofpatients, wrist in 50 percent, knee
The onset is usually sudden with fever, sore throat, and in 42 percent, hips in 26 percent and fingers in 18
Department of Clinical Virology, Karolinska Institute, S- 10521 Stockholm, Sweden, and Department of Zoology,Uppsala University, Villavagen 9, S- 75236 Uppsala, Sweden.

24 BULL. SOC. VECTOR ECOL. JUNE, 1994
percent ( Espmark and Niklasson 1984). Arthralgia examined for viremia.Evidently, Tahyna virus is capablepersisted for several years in 20 percent of the patients of producing viremia in almost all mammal speciesNiklasson et al. 1988). Sindbis virus has a wide tested ( AspOck and Hofmann 1971, Malkova 1970,
geographical distribution in African, Asian andEuropean ROdI et al. 1977, 1978, 1979, 1987, Simkova 1963,countries and in Australia, but outbreaks of human 1964, 1966a, Smetana et al. 1966). However, Tahynadisease have only been reported from northern Europe virusdidnotinduceadetectableviremiainbats( Simkovaand South Africa( Niklasson 1988). 1965), birds ( Simkova 1962, Mallcova and Marhoul
West Nile virus has been active from 1962 to 1965 1966), or amphibians and reptiles (AspOck and Kunzin the Camargue area ofsouthern France, but since 1966 1971).
the virus seems to have disappeared from this area The maximum Tahyna viremia titers were> 104.9Rodhain and Hannoun 1980). Clinical West Nile virus LD50/m1 in the rabbit, Oryctolagus cuniculus, 104.7
disease is usually mild, with a one to two week LDSO/ml in the European hare, Lepus europaeus, andconvalescence( Hayes 1988). Common symptoms are 105 LD50/ml in the hamster, Mesocricetus auratusfever, malaise, frontal headache, pain associated with ( Simkova 1963), > 104.3 LD50/m1 in the hedgehog,eye movement, and muscle pain. More prominent signs Erinaceus europaeus ( Simkova 1964, 1966a), 105. 1
include enlargementsoflymph nodes and maculopapular LD50/m1 in the European suslik, Citellus citellus, 104.8rash. Cases ofsevere meningoencephalitis associated to LD50/m1 in the fat dormouse, Clis glis( Malkova 1970),West Nile virus infection were first reported in elderly 102. 1 to 103 LD50/m1 in the red fox, Vulpes vulpespatients in Israel, but have subsequently been described ( Aspock and Hofmann 1971, Rodl et al. 1977), 103. 1
also in young patients. Less commonly associated with LDS0/m1 in the badger, Meles meles ( Aspiick andWest Nile virus infection are acute anterior myelitis, Hofmann 1971), 103 LDSO/mI in the polecat, Mustelamyocarditis, and acute pancreatis. putorius( ROdl et al. 1978), and 103 LD50/ml in the stone
This review will focus on mosquitoes as biological marten, Martesfoina( Rodl et al. 1978). Low viremiasvectors of arboviruses in Western Europe during the ( titers not reported) were detected in the common mole,summer season. The data are presented for each virus as Talpa europaea( Malkova 1970), the red squirrel, Sciurusfield evidence implicating potential vector species, vulgaris, and in the muskrat, Ondatra zibethica( ROdI etviremias in potential amplification hosts, and mosquito al. 1987).oral susceptibility and transmission capability. The main amplification hosts for Tahyna virus in
Western Europe are European hares and rabbits( BardosTAHYNA VIRUS 1975, Minar 1969, Simkova 1966b). The maximum
titers of Tahyna viremia in these two species wereField Evidence Implicating Potential Vector Species >_ 104.7 LD50/ml.
Tahyna virus was first isolated from mosquitoescollected in Slovakia( Bardos and Danielova 1959). It Mosquito Oral Susceptibility and Transmissionhas now been isolated in several countries mainly in Capabilitycentral Europe from mosquitoes of five genera( TABLE Simkova et al. ( 1960) experimentally infected1). The majority of the isolates have been obtained from Slovakian Ae. vexans with Tahynaviruseitherby feedingAedes species( 72 isolates), while two to three isolates them on a pledget containing 108.6 to 109.3 LDSWml, orper genus were obtained from Culex, Culiseta, by feeding on a suckling mice with viremias of 105.8 toAnopheles, and Coquillettidia. The largest number of > 106.8 LD50/ml. The virus multiplied and persisted inisolates per species were fromAedes vexans( 31 isolates), mosquitoes, and the virus titer leveled at 105.8 LD5olfollowed by Aedes caspius/dorsalis( 21 isolates), Aedes mosquito after seven days and remained at that level forcantans/annulipes( 10isolates), andAedescinereus( six the 21 days tested. Transmission experiments wereisolates). The high number of Tahyna virus isolates carried out after 4 to 16 days of extrinsic incubation atfrom Ae. vexans in Slovakia, in contrast to the lack of room temperature. Several mosquitoes were fed on eachisolates from this species and high number of isolates recipient mouse, and transmission occurred in 9 of 12fromAe. caspius/dorsalis in Austria, indicate that strains attempts. In a more detailed study, Danielova( 1966a)ofTahyna virus utilize different vector species in different examined Ae. vexans for Tahyna virus susceptibilityparts of its geographical distribution. and transmission capability ( TABLE 2). The Tahyna
virus infection and transmission rates increased withViremias in Potential Amplification Hosts increasing infective dose. A high virus dose (> 107.3
A large number of native vertebrate species have LD50/m1) induced an 88 percent infection rate, and a 50been experimentally infected with Tahyna virus and percent( for all refeeding mosquitoes) or 100 percent

JUNE, 1994 BULL. SOC. VECTOR ECOL. 25
TABLE 1. Isolates of mosquito-borne arboviruses from arthropods collected in the Western European
region. Isolates from mosquitoes are only from unengorged females identified at least to genera.
Genera/groupVirus/subtype Country Source( number of isolates)
Alphavirus
Sindbis virusl Italy Hyaloma marginatum( 1)
Subtotal 1 isolate
Ockelbo virus2 Sweden Culex pipiens/torrentium( 6)
Culiseta morsitans( 6)
Aedes cinereus( 2)
Culiseta spp.( 1)Norway Aedes spp. ( 1)Russia Aedes spp. ( 1)
Subtotal 17 isolates
Flavivirus
West Nile virus3 France Culex modestus( 3)
Slovakia Aedes cantans( 1)
Portugal Anopheles maculipennis( 1)
Subtotal 5 isolates
Bunyavirus/California groupTahyna virus4 Slovakia Aedes vexans( 27)
Aedes cantans( 9)
Aedes cinereus( 6)
Aedes caspius( 3)
Aedes sticticus( 1)
Culiseta annulata( 1)Culex modestus( 1)
Austria Aedes caspius/dorsalis( 16)
Aedes cantanslannulipes( 1)
Aedes caspius( 1)
Aedes flavescens( 2)
Coquillettidia richiardii( 2)Anopheles maculipennis( 3)
Germany Aedes vexans( 2)
Serbia Aedes vexans( 2)
Hungary Culiseta annulata( 1)
France Aedes caspius( 1)
Culex modestus( 1)Romania Culex pipiens( 1)
Italy Aedes caspius/vexans( 2)
Subtotal 83 isolates
Inkoo virus5 Sweden Aedes communis( 6)
Aedes punctor( 2)Finland Aedes communis/puctor( 1)
Aedes spp. ( 1)Aedes/ Culiseta( 1)
Norway Aedes communis( 5)
Aedes hexodontus( 2)
Aedes sticticus( 1)
Aedes spp. ( 1)Subtotal 20 isolates
Continued on next page.

26 BULL. SOC. VECTOR ECOL. JUNE, 1994
TABLE 1 - continued
Genera/ groupVirus/subtype Country Source( number of isolates)
BunyavirusBunyamwera groupBatai virus6 Austria Anopheles maculipennis( 71)
Coquillettidia richiardii( 1)Slovakia Anopheles maculipennis( 3)Yugoslavia Anopheles maculipennis( 1)Sweden Aedes communis( 1)
Norway Anopheles claviger( 2)
Subtotal 79 isolates
Bunyavirus/Turlock groupLednice virus? Slovakia Culex modestus( 7)
Subtotal 7 isolates
Grand total 212 isolates
1Gresikova et al. 1978.
2Niklasson et al. 1984, Lvov et al. 1984, Francy et al. 1989, Hoddevik et al. unpublished information.3Hannoun et al., 1964, Mouchet et al. 1970, Filipe 1972, Labuda et al. 1974.4Bardos and Danielova 1959, Kolman et al. 1964, Malkova et al. 1965, Hannoun et al. 1966, Aspock andKunz 1967, Balducci et al. 1968, Chippaux et al. 1970, Aspock et al. 1970, Molnar et al. 1973, Malkovaet al. 1974, Arcan et al. 1974, Gligic and Adamovic 1976, Danielova et al. 1970, 1972, 1976, 1978,Danielova and Holubova 1977, Pilaski and Mackenstein 1985.
5Brummer-Korvenkontio et al. 1973, Traavik et al. 1978, 1985, Francy et al. 1989, LundstrOm et al. 1992a.6Bardos and Cupkova 1962, Smetana et al. 1967, Aspock 1968, Aspock and Kunz 1968, Brudnjak et al.
1970, Aspock et al. 1970, Traavik et al. 1985, Francy et al. 1989.7Malkova et al. 1972, 1974.
refeeding infected mosquitoes only) transmission rate. ( Danielova et al. 1968, Danielova and Minar 1969,Danielova( 1968) showed that Tahyna virus could reach Danielova 1972, Simkova and Danielova 1969). It wasthe salivary glands ofAe. vexans in four days after oral shown that Cs. annulata was susceptible to oral infectioninfection, and RCA et al.( 1979) showed that this species with Tahyna virus( TABLE 2), with infection rates of54could transmit the virus to rabbits. percent when fed on a pledget containing 105 LDSO/ml.
Danielova ( 1966b) experimentally infected This species was also capable of transmitting the virusSlovakian mosquitoes of several species by feeding to chimpanzees( Simkova and Danielova 1969).
them on pledgets containing 103 to 106.8 LD50/m1 andfound that Ae. cantans, Ae. flavescens, Ae. sticticus,Ae. OCKELBO VIRUS
communis, and Ae. excrucians were susceptible to oralinfection with Tahyna virus( TABLE 3). Aedes sticticus Field Evidence Implicating Potential Vector Specieswas studied in more detail and it was shown that a virus Ockelbo virus was first isolated from mosquitoestiter of 106.8 LD50/m1 gave an infection rate of 52 collected in central Sweden during the 1982 outbreak ofpercent ( n= 25). Transmission to suckling mice was Ockelbo disease( Niklasson et al. 1984). This virus hasachieved by refeeding Ae. sticticus, the only species been isolated in Sweden, Russia and Norway fromtested for transmission. unengorged mosquitoes of three genera ( TABLE 1).
In several experimental studies, Slovakian Cs. The majorityofSwedish isolates were from ornithophilicannulata were infected by feeding on apledget containing species, Cs. morsitans ( 6 isolates) and Cx. pipiens/Tahyna virus, or by feeding on a viremic mouse torrentium ( six isolates), and only two isolates were

JUNE, 1994 BULL. SOC. VECTOR ECOL. 27
TABLE 2. Experimental vector competence of Slovakian Aedes vexans and Culiseta annulata forTahyna virus after Danielova( 1966a, 1972). Each mosquito fed on a pledget containingblood from a viremic hamster mixed with glucose was held at room temperature or at 25°C
for 13- 16 days, and was then given a noninfected mouse for the second blood meal.
Transmission rate in percent
Virus titer number tested)
in infective Infection rate
meal in log10 in percent All refeeding Only refeeding
SMLD50/m1 number tested) mosquitoes infected mosquitoes
Aedes vexans
3. 5- 3. 8 11 ( 27) NT1 NT
6.5 59 ( 27) 44 ( 9) 67 ( 6)
7. 3 88 ( 26) 50 ( 16) 100 ( 8)
Culiseta annulata
3. 8 ND2 ( 12) NT NT
5. 0 54 ( 26) NT 28 ( 14)
1NT= not tested.
2ND= no virus detected.
from a species( Ae. cinereus) with more general feeding longer in passeriforms than in anseriforms. The
habits ( Service 1971, Jaenson and Niklasson 1986, maximum duration in passeriforms was not determined
Francy et al. 1989). Sindbis virus has been isolated from because a high proportion of these birds were viremic
ticks in Italy( Gresikova et a1. 1978), but it has not been during the fifth and last sampling day. Experimentallyisolated from mosquitoes in Western Europe( TABLE infected Swedish bank voles( Cletrionomys glareolus)
1). produced Ockelbo viremias of 1027 to 107.2 PFU/mlOckelbo virus was only isolated from mosquitoes ( adults, n= 4) or 107.5 to 109.2 PFU/ml( age 2 days, n= 4)
collected during July and August, with the highest ( Lundstrdm, unpublished information).
monthly field infection rates( FIR) in August( Francy et The main amplification hosts for Ockelbo virus in
al. 1989). The monthly FIR for Cx. pipiens/torrentium Sweden are several passeriform species of the genera
5.0/ 1, 000 in July, and 12.6/ 1, 000 in August) was Turdus and Fringilla( Francy et al. 1989, Lundstrdm etconsistently higher than in Cs. morsitans( 2.0/ 1, 000 in al. 1992b, 1992c, 1993). Peak mean daily viremia titersJuly, and 4. 1/ 1, 000 in August). Aedes cinereus had a in these bird species were 105.8 to 107.5 PFU/mlFIR of 0.3/ 1, 000 in August. Lundstrdm et al. 1993).
Virus isolations from field collected mosquitoes
indicate that a Culex species( pipiens or torrentium) is Mosquito Oral Susceptibility and Transmissionthe main enzootic vector among birds, and that Ae. Capabilitycinereus is the main vector for transmission of Ockelbo The vector competence for Ockelbo virus was
virus from viremic birds to humans( Francy et al. 1989). studied in Swedish Cx. pipiens, Cx. torrentium, Ae.
cinereus, Ae. communis, and Ae. excrucians( TABLE
Viremias in Potential Amplification Hosts 5). Mosquitoes were engorged on Ockelbo virus infected
Ockelbo virus could produce viremia in all 14 viremic chickens, were held at 17°C for 14 to 28 days,
indigenous bird species of three taxonomic orders and the ability to transmit virus was tested by lettingexamined( TABLE 4). For birds older than two weeks, each mosquito refeed on a susceptible ( noninfected)
the mean daily viremia titers expressed as plaque- chicken ( Lundstrom et al. 1990a, 1990b, Turell et al.
forming units ( PFU) per ml blood were significantly 1990). Viremias in infected chickens, at the time of
higher in passeriforms( 105.8 to 107.5 PFU/ml) than in mosquito feedings, ranged from 103.5 to 108.5 PFU/ml.anseriforms( 103.7 to 104.5 PFU/ml)( Lundstrdm et al. All Aedes species were highly susceptible ( infection1993). In addition, the duration of the viremia was rates? 9• percent, n= 116), and 50 percent( n= 6) of the

28 BULL. SOC. VECTOR ECOL. JUNE, 1994
TABLE 3. Susceptibility ofSlovakian mosquitoes for oral infection with Tahyna virusafter Danielova( 1966b). Each mosquito fed on a pledget containing bloodfrom a viremic hamster mixed with glucose, and was held at unknown
temperature for a varying number of days.
Virus titer
in infective Number of
meal in loglo Days extrinsic mosquitoes
Mosquito Species SMLDso/ ml incubation tested'
Virus detected
Aedes cantans 3. 9 11 21
6.8 16 3
Aedes communis 3. 9 10 3
Aedes excrucians 3.9 10 56.8 16 1
Aedes flavescens 3. 9 10 2
Aedes sticticus2 4.5- 4.9 8- 12 24
Culex pipiens 5. 5- 5. 9 14- 15 18
Virus not detected
Aedes cinereus 3. 0 8- 23 10
4.7 10 5
Aedes punctor 4.9 13 3
Culex pipiens 3. 5 14 254.7-4.9 10- 14 24
Culiseta annulata 5. 2 10 1
Anopheles maculipennis 5. 2- 5. 5 8- 14 9
Mosquitoes were tested for virus content in pools of maximum 25 specimens each.2The only species tested for ability to transmit virus and it transmitted Tahyna virus to
a rabbit after 15 to 16 days of extrinsic incubation.
refeeding Aedes mosquitoes transmitted virus to being the most efficient vector( LundstrOm et at. 1990a,chickens. Infection rates in Cx. torrentium were z88 1990b, Turell et al. 1990).
percent ( n= 86) and all refeeding mosquitoes ( n= 11)transmitted virus. In Cx pipiens both infection and WEST NILE VIRUStransmission rates were less than 45 percent ( n= 483,
respectively n= 142). Field Evidence Implicating Potential Vector SpeciesThus, both Culex and Aedes mosquitoes were West Nile virus has been isolated in Portugal,
competent laboratory vectors of Ockelbo virus at the Slovakia, and France from mosquitoes of three generaenvironmental temperatures prevailing in Ockelbo ( TABLE 1). Virus isolations from field collectedendemic areas during summer, with Cx. torrentium specimens implicated An. maculipennis, Ae. cantans

JUNE, 1994 BULL. SOC. VECTOR ECOL. 29
TABLE 4. Proportion of birds viremic and peak mean daily viremia titers per species and age- group forindigenous Swedish birds after inoculation ofapproximately 102.7 plaqueforming units( PFU)of Ockelbo virus after Lundstrom et al. 1993.
Order Age at the time of infection
Species 1 week 2 weeks
Anseriformes
Canada goose Branta canadensis 25 ( 4) 1 2.02 50 ( 2) 3. 7
Bean goose Anserfabalis 100 ( 6) 6.2 ND3 ( 1)
Mallard Anas plathyrhynchos 50 ( 8) 5.0 58 ( 12) 4.5
Goldeneye Bucephala clangula 100 ( 2) 6. 3 83 ( 6) 4.5
Passeriformes
Blackcap Sylvia atricapilla 100 ( 4) 6.2 NT4
Redwing Turdus iliacus NT 100 ( 4) 6.2
Song trush Turdus philomelos NT 100 ( 1) 6.6
Fieldfare Turdus pilaris 100 ( 2) 5.0 57 ( 7) 6.1
Great tit Parus major 100 ( 3) 4. 8 ND ( 3)
House sparrow Passer domesticus NT 67 ( 3) 5. 8
Chaffinch Fringilla coelebs NT 83 ( 6) 6.9
Greefinch Carduelis chloris NT 83 ( 12) 7. 5
Yellowhammer Emberiza citrinella NT 44 ( 16) 6. 3
Galliformes
Capercaillie Tetrao urogallus 100 ( 2) 8. 2 100 ( 2) 6.4
Percent viremic( number tested).
2Peak mean daily viremia in log10 PFU/ ml blood.3ND= virus not detected.
4NT= not tested.
and Cm modestus as vectors( Hannoun et al. 1964, Filipe LEDNICE VIRUS
1972, Labuda et al. 1974).
Field Evidence Implicating Potential Vector SpeciesMosquito Oral Susceptibility and Transmission Lednice virus has only been isolated in SlovakiaCapability and only from Cx. modestus( TABLE 1). Several other
An experimental study showed that French Ae. mosquito species mainlyAedes spp., have been examinedgeniculatus, Ae. punctor, and An. plumbeus were ( Malkova et al. 1972, 1974).
competent vectors for an African strain of West Nile
virus( Vermeil et al. 1960). Mosquitoes were infected Viremias in Potential Amplification Hosts
by feeding on a viremic chicken inoculated with West Bird species including the duck, Anas sp., theNile virus 24 to 48 hours previously. After six days of goose, Anser sp., the pheasant, Phasianus colchius, theextrinsic incubation at 25°C or 28°C the mosquitoes black-headed gull, Larus ridibundus, and the coot,
were allowed to refeed on mice. Detection oftransmission Fulica atra, have been studied for experimental viremia
was by observation ofmice for signs ofparalysis. A total with Lednice virus ( Danielova and Malkova 1976,
of 10 specimens of each mosquito species were tested, Malkova and Danielova 1977, Malkova et al. 1979).
but the mosquitoes were not assayed singly for infection Ducklings, gosling, and young pheasants produced aand transmission. Thus, neither infection nor low titer viremia after experimental infection with
transmission rates could be determined. Lednice virus, but the viremia titers were not reported
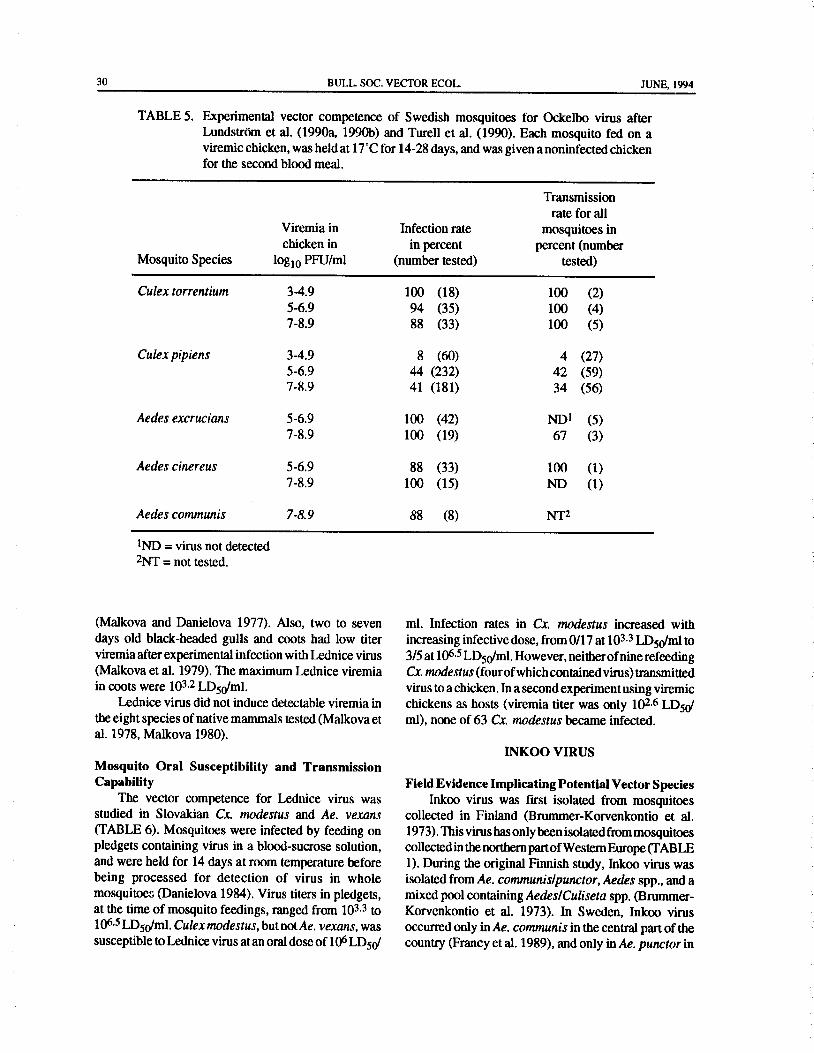
30 BULL. SOC. VECTOR ECOL. JUNE, 1994
TABLE 5. Experimental vector competence of Swedish mosquitoes for Ockelbo virus afterLundstrOm et al. ( 1990a, 1990b) and Turell et al. ( 1990). Each mosquito fed on a
viremic chicken, was held at 17°C for 14- 28 days, and was given anoninfected chickenfor the second blood meal.
Transmission
rate for all
Viremia in Infection rate mosquitoes in
chicken in in percent percent( number
Mosquito Species log10 PFU/ml number tested) tested)
Culex torrentium 3- 4.9 100 ( 18) 100 ( 2)
5- 6.9 94 ( 35) 100 ( 4)
7- 8.9 88 ( 33) 100 ( 5)
Culex pipiens 3- 4.9 8 ( 60) 4 ( 27)
5- 6.9 44 ( 232) 42 ( 59)7- 8.9 41 ( 181) 34 ( 56)
Aedes excrucians 5- 6.9 100 ( 42) ND' ( 5)
7- 8. 9 100 ( 19) 67 ( 3)
Aedes cinereus 5- 6.9 88 ( 33) 100 ( 1)
7- 8.9 100 ( 15) ND ( 1)
Aedes communis 7-8.9 88 ( 8) NT2
1ND= virus not detected
2NT= not tested.
Malkova and Danielova 1977). Also, two to seven ml. Infection rates in Cx modestus increased withdays old black-headed gulls and coots had low titer increasing infective dose, from 0/17 at 103. 3 LD50/m1 toviremia after experimental infection with Lednice virus 3/ 5 at 106.5 LD50/ml. However, neither ofnine refeedingMalkova et al. 1979). The maximum Lednice viremia Cx. modestus( fourofwhichcontainedvirus) transmitted
in coots were 103.2 LD50/ml. virus to a chicken. In a second experiment using viremicLednice virus did not induce detectable viremia in chickens as hosts ( viremia titer was only 102.6 LD501
the eight species of native mammals tested( Malkova et ml), none of 63 Cx. modestus became infected.al. 1978, Malkova 1980).
INKOO VIRUS
Mosquito Oral Susceptibility and TransmissionCapability Field Evidence Implicating Potential Vector Species
The vector competence for Lednice virus was Inkoo virus was first isolated from mosquitoesstudied in Slovakian Cx. modestus and Ae. vexans collected in Finland ( Brummer-Korvenkontio et al.TABLE 6). Mosquitoes were infected by feeding on 1973). This virus has only been isolated frommosquitoes
pledgets containing virus in a blood-sucrose solution, collected in the northern partofWestern Europe( TABLEand were held for 14 days at room temperature before 1). During the original Finnish study, Inkoo virus wasbeing processed for detection of virus in whole isolated from Ae. communis/punctor, Aedes spp., and amosquitoes( Danielova 1984). Virus titers in pledgets, mixed pool containing Aedes/ Culiseta spp.( Brummer-at the time of mosquito feedings, ranged from 103.3 to Korvenkontio et al. 1973). In Sweden, Inkoo virus106.5 LD50/ml. Culex modestus, but not Ae. vexans, was occurred only in Ae. communis in the central part of thesusceptible toLednice virus at an oral dose of 106 LDSO/ country( Franey et al. 1989), and only in Ae. punctor in

JUNE, 1994 BULL. SOC. VECTOR ECOL. 31
TABLE 6. Susceptibility of Slovakian mosquitoes for oral infection with Lednicevirus after Danielova( 1984). Each mosquito fed on a pledget containinga mixture of viremic mouse brain, blood and glucose, and was held atroom temperature for 14 days.
Virus titer in Infection rate
infective meal in percent
Mosquito Species in log 10LD50/m1 number tested)
Culex modestus 6.0 POS1 ( 21)
Aedes vexans 6.0 ND2 ( 17)
Culex modestus3 3. 3 ND ( 17)
4.5 12 ( 16)
5. 5 30 ( 20)
6.5 60 ( 5)
1POS= virus detected but infection rate not determined.
2ND= virus not detected.
3None of the nine refeeding( four of which contained virus) Cx. modestus transmittedvirus to a chicken.
the northern part( Lundstrom et al. 1992a). California Experimental Infection of Vertebrates and
group virus( probably Inkoo virus) have been isolated Mosquitoes
from mosquitoes collected in Norway, most often from An experimentally infected red fox produced aAe. communis,butalso from otherAedes species( Traavik Batai viremia of 1018 LD50/ml, while no viremia waset al. 1978, 1985). detected in abadgerMeles meles( Aspock and Hofmann
Thus, field studies indicate thatAe. communis is the 1971).
main vector for Inkoo virus, and that Ae. punctor may No studies have been published with experimental
replace it as vector in the far north( Traavik et al. 1985, Batai virus infection in mosquitoes after ingesting virus.
Francy et al. 1989, LundstrOm et al. 1992a). However, Batai virus will replicate in Austrian An.
Neither the viremia titers in potential vetebrate maculipennis after intrathoracial inoculation ( Aspock
hostsnor the experimental vector competence ofpotential and Kunz 1970).
vector species have been published.CONCLUSIONS
BATAI VIRUS
The main components of vector competence are
Field Evidence Implicating Potential Vector Species vector exposure to the pathogen, vector susceptibility toBunyamwera group viruses have been isolated in the pathogen, multiplication of the pathogen in the
the northern and central parts ofWestern Europe( TABLE vector, and transmission of the pathogen ( Grimstad
1). These virus isolates have been published as Batai 1983, Hardy et al. 1983). These main components havevirus ( Francy et al. 1989), Calovo virus ( Bardos and been given different priorities by Western EuropeanCupkova 1962, Smetana et al. 1967, Aspock 1968, scientists. The majority of studies have focused onBrudjak et al. 1970, Aspock et al. 1970), or Bunyamvera identifying the mosquito species naturally infected withgroup virus ( Traavik et al. 1985). Calovo virus is arboviruses, their oral susceptibility, and a few studiesconsidered a strain of Batai virus ( Karabatsos 1985), that actually examined the ability of Europeanwhile the Norwegian strains have not been further mosquitoes to transmit the viruses upon refeeding.
classified. Extensive field studies in several Western European
The very large number of Batai virus isolates from countries indicate thatAe. vexans is the main vector for
An. maculipennis in central Europe makes it the prime Tahynavirus( TABLE 1). However, furthercomparative
candidate for further studies( TABLE 1). experimental studies with local populations ofAe.vexans,

32 BULL. SOC. VECTOR ECOL. JUNE, 1994
Ae. caspius, Ae. dorsalais, Ae. cantons, Ae. annulipes, mosquito-borne arboviruses, experimental studies areand Ae. cinereus assayed for infection and transmission warranted to define the vector species.
rates may shed more light on the relative importance ofeach species in each geographical region. Acknowledgments
Extensive studies on the vectors of Ockelbo virushave only been performed in Sweden ( TABLE 1). I am grateful to Dr. Ake Lundkvist, Department ofResults from field studies indicate that either Cx.pipiens Defence Microbiology, Swedish Institute for Infectiousor Cx. torrentium is the main enzootic vector, with Cs. Disease Control, Sweden, and professor Christine Dahl,morsitans as a third potential enzootic vector( Francy et Department of Zoology, Uppsala University, Sweden,al. 1989). Vector competence experiments showed that for valuable comments and suggestions on theCx. torrentium was an efficient laboratory vector for manuscript. Drs. Gunnar M. Hoddevik, Reidar Mehl,Ockelbo virus, and that Cx. pipiens was relatively and Bjorn P. Berdahl, National Institute ofPublic Health,refractory and apoor vector( TABLE4). The conclusion Oslo, Norway, are acknowledged for providingis that Cx. torrentium is the main enzootic vector for unpublished information.
Ockelbo virus, while Ae. cinereus and perhaps otherAedes species may transmit virus from viremic birds tohumans. The ornitophilic species Cs. morsitans may REFERENCES CITEDalso function as an enzootic vector( Jaenson et al. 1986,Jaenson and Niklasson 1986, Franey et al. 1989), but the Arcan, P., V. Topciu, N. Rosiu, and N. Csaly. 1974.experimental vector competence of this species for Isolation of Tahyna virus from Culex pipiensOckelbo virus has to be examined before this question mosquitoes in Romania. Acta Virol. (Praha) 18:will be resolved. 175.
A relatively low number ofWest Nile virus isolates Asptick H. 1968. Weitere Untersuchungen uber dieindicate that Cx. modestus could be the main vector for durch Stechmucken ubertragenen Arbovirentransmission of virus to humans( TABLE 1). However, Osterreichs. Zentralbl. Bakteriol.( A) 208: 69-80.other possible enzootic vectors, such as Cx.pipienshave Asptick H. and H. Hofmann. 1971. Der Verlauf dernot been investigated( Mouchet et al. 1970). This study experimentellen Infection von Fuchs and Dachsshowed two Aedes and one Anopheles species as mit Tahyna- and Calovo-Virus. Zentralbl.competent laboratory vectors forWest Nile virus. Further Bakteriol.( A) 217: 148- 151.field and experimental studies are warranted, especially Aspock H. and C. Kunz. 1967. Untersuschungen uberwith ornitophilic Culexmosquitoes, to define the vectors die Okologie des Tahyna- Virus. Zentralbl.for West Nile virus in Europe. Bakteriol.( A) 203: 1- 24.
Lednice virus has only been isolated from Cx. Asptick, H. and C. Kunz. 1968. Isolierung des Calovo-modestus in Slovakia( TABLE 1), and this species was Batai, = Chitor) Virus aus Stechmucken inalso susceptible to experimental infection with the virus Osterreich. Wien. Med. Wschr. 118: 497-498.TABLE 5). However, further field studies, especially Asptick H. and C. Kunz. 1970. Uberwinterung des
of ornithophilic mosquito species, and comparative Calovo- Virus in experimentell infiziertenvector competence experiments with mosquitoes infected Anopheles maculipennis messeae. Zentralblby feeding on viremic birds and transmitting to recipient Bakteriol.( A) 213: 429-433.birds, are needed to define the enzootic vectors for Asptick H. and C. Kunz. 1971. Untersuchungen uberLednice virus. die Uberwinterung von Tahyna- and Calovo-Virus
Field evidence indicates that Ae. communis is the in Amphibien and Reptilien. Zentralbl. Bakteriol.main vector for Inkoo virus in Northern Europe, but it A) 216: 1- 8.maybe replaced as the main vector by Ae. punctor in the Asptick, H., C. Kunz, and G. Pretzmann. 1970.northernmost parts of the Inkoo virus geographical Phanologie and Abundanz der Stechmuchen desdistribution ( TABLE 1). This question could only be ostlichen Neusiedlersee Gebiets( Ost-Osterreich)resolved by comparative infection and transmission in ihrer Beziehung zum Auftreten der durchexperiments. Stechmucken ubertragen Arboviren. Zentralbl.
The field data strongly indicate that An. Bakteriol.( A) 214: 160- 173.maculipennis is the main vector for Batai virus in some Balducci, M., P. Verani, M. C. Lopes, G. Sacca, and B.areas in Western Europe( TABLE 1). No information is Gregorig. 1968. Isolation of Tahyna virus fromavailable on the vector competence of European Aedes mosquitoes in northern Italy ( Goriziamosquitoes for Batai virus. As for most other European province). Acta Virol.( Praha) 12: 457-459.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 33
Bardos, V. 1975. The role ofmammals in the circulation Lednice virus. Folia Parasitol. ( Praha) 31: 379-
of Tahyna virus. Folia Parasitol. ( Praha) 22: 257- 382.
261. Danielova, V., Z. Hajkova, J. Minar, and J. Ryba. 1972.
Bardos, V. and E. Cupkova. 1962. The Calovo virus- Virological investigations ofmosquitoes in different
the second virus isolated from mosquitoes in seasons of the year at the natural focus of the
Czechoslovakia. J. Hyg. Epidemiol. Microbiol. Tahyna virus in southern Moravia. Folia Parasitol.
Inununol. 6: 186- 192. Praha) 19: 25- 31.
Bardos, V. and V. L. Danielova. 1959. The Tahyna Danielova, V. and J. Holubova. 1977. Two more
virus - a virus isolated from mosquitoes in mosquito species proved as vectors of Tahyna
Czechoslovakia. J. Hyg. Epidemiol. Microbiol. virus in Czechoslovakia. Folia Parasitol. ( Praha)
Immunol. 3: 264-276. 24: 187- 189.
Bardos, V., M.Medek, V. Kania, and Z. Hubalek. 1975. Danielova, V. and D. Malkova. 1976. Studies on
Isolation of Tahyna virus from the blood of sick viremia and antibody formation in ducklings andchildern. Acta Virol.( Praha) 19: 447. goslings after experimental infection with Lednice
Bardos, V., M. Medek, V. Kania, Z. Hubalek, and Z. Yaba 1) virus. Folia Parasitol. ( Praha) 23: 367-
Juricova. 1980. Das Klinische Bild der Tahyna- 372.
Virus ( California-Gruppe) - Infectionen bei Danielova, V., D. Malkova, J. Minar, B. Rehse-Kupper,
Kindern. Padriatr. Grenzgeb. 19: 11- 23. Z. Hajkova, J. Halgos, and L. Jedlicka. 1978.
Brudnjak, Z., V. Danielova, J. Ryba, and J. Vesenjak- Arbovirus isolations from mosquitoes in South
Hirjan. 1970. Isolation of Calovo virus from Slovakia. Folia Parasitol.( Praha) 25: 187- 191.
Anopheles maculipennis sl mosquitoes in Danielova, V., D. Malkova, J. Minar, and J. Ryba.
Yugoslavia. Folia Parasitol.( Praha) 17: 323- 324. 1976. Dynamics of the natural focus of Tahyna
Brummer-Korvenkontio, M. and P. Kuusisto. 1984. virus in southern Moravia and species succession
Pogosta disease. P. 23 in Proceedings of the Soviet- of its vectors, the mosquitoes of the genus Aedes.
Finnish symposium on etiology, epidemiology, Folia Parasitol.( Praha) 23: 243- 249.
diagnosisandprophylaxisofKarelinfever- Pogosta Danielova, V. and J. Minar. 1969. Experimental
disease, ( T. M. Skvortsova, G. A. Sidorova, L. K. overwintering of the virus Tahyna in mosquitoesBerezina, B. Fomina, V. I. Yakolev, and T. K. Culiseta annulata ( Schrk.) ( Diptera, Culicidae).
Bobrovskij, eds.). Academy of Medical Sciences, Folia Parasitol. ( Praha) 16: 285- 287.
Perozavodsk, USSR, 28 pp. Danielova, V., J. Minar, and B. Rosicky. 1968.
Brummer-Korvenkontio, M., P. Saikku, P. Korhonen, I. Experimental survival of the virus Tahyna in
Ulmanen, T. Reunala, and J. Karvonen. 1973. hibernating mosquitoes Theobaldia annulataArbovirusesinFinland. IV. AFinnishrepresentative Schrk.). Folia Parasitol.( Praha) 15: 183- 187.
of the California group. Am. J. Trop. Med. Hyg. 22: Danielova, V., J. Minar, and J. Ryba. 1970. Isolation of
404-413. Tahyna virus from mosquitoes Culiseta annulata
Chippaux, A., J. Rageau, and J. Mouchet. 1970. Schrk. 1776). Folia Parasitol. ( Praha) 17: 281-
Hibernation de l' arbovirus Tahyna chez Culex 284.
modestus Fic. en France. C. R. Acad. Sci. ( D) Espmark, A. and B. Niklasson. 1984. Ockelbo diseaseParis) 270: 1648- 1650. in Sweden: epidemiological, clinical, and
Danielova, V. 1966a. Quantitative relationships of virological data from the 1982 outbreak. Am. J.
Tahyna virus and the mosquito Aedes vexans. Acta Trop. Med. Hyg. 33: 1203- 1211.Virol.( Praha) 10: 62-65. Filipe, A. R. 1972. Isolation in Portugal of West Nile
Danielova, V. 1966b. The relation of the virus Tahyna virus from Anopheles maculipennis mosquitoes.
to some species of mosquitoes of the genera Aedes, Acta Virol. (Praha) 16: 361.
Culex and Anopheles. Folia Parasitol. ( Praha) 13: Francy, D. B., T. G. T. Jaenson, J. 0. LundstrOm, E. B.97- 102. Schildt, A. Espmark, B. Henriksson, and B.
Danielova, V. 1968. Penetration of the Tahyna virus to Niklasson. 1989. Ecologic studies of mosquitoes
various organs of theAedes vexans mosquito. Folia and birds as hosts of Ockelbo virus in Sweden and
Parasitol.( Praha) 15: 87- 91. isolation of Inkoo and Batai viruses from
Danielova, V. 1972. The vector efficiency of Culiseta mosquitoes. Am. J. Trop. Med. Hyg. 41: 355- 363.annulata mosquito in relation to Tahyna virus. Gligic, A. and Z. R. Adamovic. 1976. Isolation of
Folia Parasitol.( Praha) 19: 259- 262. Tahyna virus from Aedes vexans mosquitoes in
Danielova, V. 1984. To the problem of the vector of Serbia. Mikrobiologija 13: 119- 129.

34 BULL. SOC. VECTOR ECOL. JUNE, 1994
Gresikova, M., M. Sekeyova, G. Tempera, S. Lundstrdm, J. 0.,A. Folke, and B. Niklasson. 1992a.Guglielmino, and A. Castro. 1978. Identification The mosquito fauna of a wetland area in northernof a Sindbis virus strain isolated from Hyaloma Sweden in relation to arbovirus transmission.marginatum ticks in Sicily. Acta Virol.( Praha) 22: Seventh European regional meeting of SOVE,231- 232. Bologna, Italy, August 1992, p. 35.
Grimstad, P. R. 1983. Mosquitoes and the incidence of Lundstrdm, J. 0., B. Niklasson, and D. B. Francy.encephalitis. Adv. Virus Res. 28: 357- 438. 1990a. Swedish Culex torrentium and Cx.pipiens
Grimstad, P. R. 1988. California group virus disease. Diptera: Culicidae) as experimental vectors ofPp. 99- 136 in The Arboviruses: Epidemiology and Ockelbo virus. J. Med. Entomol. 27: 561- 563.Ecology, Volume II ( T. P. Monath, ed.). CRC Lundstrdm, J. O., M. J. Turell, and B. Niklasson. 1990b.Press, Boca Raton, Florida, U.S. A, 280 pp. Effect of environmental temperature on the vector
Hannoun, C., R. Panthier, and B. Corniou. 1966. competence of Culex pipiens and Cx. torrentiumIsolation of Tahyna virus in the south of France. forOckelbo virus. Am. J. Trop. Med. Hyg. 43: 534-Acta Virol.( Praha) 10: 362- 364. 542.
Hannoun, C., R. Panthier, J. Mouchet, and J.- P. Eouzan. Lundstrdm, J. 0.,M. J. Turell, and B. Niklasson. 1992b.1964. Isolement en France du virus West-Nile Birds as hosts for Ockelbo virus, a mosquito-bornea' partir de malades et du vecteur Culex modestus human pathogenic virus innorthern Europe. SeventhFicalbi. C. R. Acad. Sci. ( D) ( Paris) 259: 4170- European regional meeting of SOVE, Bologna,4172. Italy, August 1992, p. 53.
Hardy, J. L., E. J. Houk, L. D. Kramer, and W. C. Lundstrdm, J. O., M. J. Turell, and B. Niklasson. 1992c.Reeves. 1983. Intrinsic factors affecting vector Antibodies to Ockelbo virus in three orders ofbirdscompetence of mosquitoes for arboviruses. Ann. Anseriformes, Galliformes and Passeriformes) inRev. Entomol. 28: 229-262. Sweden. J. Wildl. Dis. 28: 144- 147.
Hayes, C. G. 1988. West Nile fever. Pp. 59- 88 in The Lundstrdm, J. 0.,M. J. Turell, and B. Niklasson. 1993.Arboviruses: Epidemiology and Ecology ( T. P. Viremia in three orders of birds ( Anseriformes,Monath, ed.). Volume V, CRC Press, Boca Raton, Galliformes and Passeriformes) inoculated withFlorida, U.S. A, 249 pp. Ockelbo virus. J. Wildl. Dis. 29: 189- 195.
Jaenson, T. G. T., B. Niklasson, and B. Henriksson. Lundstrdm, J. O., S. Vene, A.Espmark, M.Engvall, and1986. Seasonal activity ofmosquitoes in an Ockelbo B. Niklasson. 1991. Geographical and temporaldisease endemic area in central Sweden. J. Am. distribution of Ockelbo disease in Sweden.Mosq. Contr. Assoc. 2: 18- 28. Epidemiol. Infect. 106: 567-574.
Jaenson, T. G. T. and B. Niklasson. 1986. Feeding Lvov, D. K.; T. M. Skvortsova, N. G. Kondrashina, B.patterns of mosquitoes ( Diptera: Culicidae) in V. Vershinskyu, A.L.Lesnikov, V. S. Dereviansky,relation to the transmission of Ockelbo disease in L. K. Berezina, V. L. Gromashevsky, D. P.Sweden. Bull. Entomol. Res. 76: 375- 383. Andrianova, and V. I. Yakolev. 1982. Etiology of
Karabatsos, N.( ed.). 1985. International catalogue of Karelian fever- a new arbovirus infection. Vopr.arboviruses, including certain other viruses of Virusol. 27: 690-692.vertebrates, 3rd ed. Am. Soc. Trop. Med. Hyg., Lvov, D. K, T. M. Skvortsova, L. K. Berezina, V. L.San Antonio, Texas, U.S. A., Pp. 219 and 297. Gromashevsky, B. I. Yakolev, B. V. Gushchin, V.
Kohnan, J. M., D. Malkova, A. Nemec, A. Smetana, Z. A.Aristova, G. A. Sidorova, E. L.Gushchina, S. M.Hajkova, and J. Minar. 1964. The isolation of the Klimenko, and others. 1984. Isolation of KarelianTahyna virus from the mosquito Aedes vexans in fever agent from Aedes communis mosquitoes.southern Moravia. J. Hyg. Epidemiol. Microbiol. Lancet 2: 399-400.Immunol. 8: 380-386. Malkova, D. 1970. The development of viremia and
Kolobukhina, L. V., D. K. Lvov, A. M. Butenko, M. S. neutralizing antibodies in the European suslik, fatNedyalkova, A. A. Kuznetsov, and I. V. Galkina. dormouse and common mole, experimentally1990. Signs and symptoms of infections caused by infected with the virus Tahyna. Folia Parasitol.California serogroup viruses in humans in the Praha) 17: 85-88.U.S. S. R. Arch. Virol. (Suppl. 1): 243- 247. Malkova, D. 1980. Experimental infection of
Labuda, M., O. Kozuch, and M. Gresikova. 1974. hedgehogs, hamsters, rabbits and ferrets with theIsolation of West Nile virus from Aedes cantons virus Lednice. Folia Parasitol. ( Praha) 27: 69-mosquitoes in west Slovakia. Acta Virol.( Praha) 70.18: 429-433. Malkova, D. and V. Danielova. 1977. Experimental

JUNE, 1994 BULL. SOC. VECTOR ECOL. 35
infection ofpheasants with Lednice( Yaba 1) virus. Dis. 157: 832-835.
Folia Parasitol.( Praha) 24: 382- 384. Pilaski, J. and H. Mackenstein. 1985. Nachweis des
Malkova,D., V. Danielova, J. M. Kolman, J. Minar, and Tahyna- Virus bei Stechmucken in zwei
A. Smetana. 1965. Natural focus of Tahyna virus verschiedenen europaischen Naturherden.
in South Moravia. J. Hyg. Epidemiol. Microbiol. Zentralbl. Bakteriol.( B) 180: 394-420.
Immunol. 9: 434-440. Rodhain, F. and C. Hannoun. 1980. Present status of
Malkova, D., V. Danielova, and D. Lim. 1979. arboviruses in France. Pp. 111- 116 in ArbovirusesExperimental infectionofblack-headed gulls( Larus ofthe Mediterranean Countries( J. Vesenjak-Hirjan
ridibundus L.) and coots ( Fulica atra L.) with et al., eds.). Zentrabl. Bakteriol. Suppl. 9. Gustav
Lednice( M' Poko) virus. Folia Parasitol. ( Praha) Fischer Verlag, Stuttgart and New York, 332 pp.26: 85- 88. Rodl, P., V. Bardos, Z. Hubalek, and Z. Juricova. 1977.
Malkova,D., V. Danielova, J. Minar, B. Rosicky, and J. Experimental infection of foxes with Tahyna virus.
Casals. 1972. Isolation of Yaba 1 arbovirus in Folia Parasitol.( Praha) 24: 373- 376.
Czechoslovakia. Acta Virol.( Praha) 16: 93. Rodl, P., V. Bardos, andZ.Hubalek. 1978. Experimental
Malkova, D., V. Danielova, J. Minar, and J. Ryba. infection ofPutoris eversmanni polecats andMartes
1974. Virological investigations of mosquitoes foina martens with Tahyna virus. Acta Virol.
in some biotopes of southern Moravia in summer Praha) 22: 502-505.
season 1972. Folia Parasitol. ( Praha) 21: 363- Rodl, P., V. Bardos, andZ.Hubalek. 1987. Experimental
372. infection of the squirrel( Sciurus vulgaris) and the
Malkova, D., J. M. Kolman, and Z. Hodkova. 1978. muskrat ( Ondatra zibethica) with Tahyna virus
Relationships of Lednice ( Yaba 1) arbovirus to California group, Bunyaviridae). Folia Parasitol.
some small mammals. Folia Parasitol. ( Praha) 25: Praha) 34: 189- 191.
113- 114. ROdI, P., V. Bardos, and J. Ryba. 1979. Experimental
Malkova, D. and D. Marhoul. 1966. Attempts at transmission of Tahyna virus( California group) toexperimental infection of pheasants with Tahyna wild rabbits( Oryctolaguscuniculus)bymosquitoes.
virus. Acta Virol.( Praha) 10: 375. Folia Parasitol.( Praha) 26: 61- 64.
Minar, J. 1969. Food sources of some mosquito species Service, M. W. 1971. Feeding behaviour and hostin the natural focus of Tahyna virus in southern preferences of British mosquitoes. Bull. Entomol.
Moravia. Folia Parasitol. (Praha) 16: 81- 85. Res. 60: 653-661.
Molnar, E., M. Gresikova, T. Kubaszova, L. Kubinyi, Simkova, A. 1962. Tahyna virus in birds. Acta Virol.
and J. B. Szabo. 1973. Arboviruses in Hugary. J. Praha) 6: 190.
Hyg. Epidemiol. Microbiol: Immunol. 17: 1- 10. Simkova, A. 1963. Quantitative study of experimentalMouchet, J., J. Rageau, C. Laumond, C. Hannoun, D. Tahyan virus infection in potential reservoir
Beytout, J. Oudar, B. Corniou, and A. Chippaux. animals. Acta Virol. ( Praha) 7: 414-420.
1970. Epidemiologie do virus West Nile: etude Simkova, A. 1964. Tahyna virus in hedgehogs. Acta
d' un foyer en Camargue. Ann. Inst. Pasteur( Paris) Virol.( Praha) 8: 285.
118: 839-842. Simkova, A. 1965. Tahyna virus in bats. Acta Virol.
Nashed, N. W., J. G. Olson, and A. El- Tigani. 1993. Praha) 9: 285.
Isolation ofBatai virus( Bunyaviridae: Bunyavirus) Simkova, A. 1966a. Quantitative study ofexperimentalfrom the blood of suspected malaria patients in Tahyna virus infection in hibernating hedgehogs.Sudan. Am. J. Trop. Med. Hyg. 48: 676-681. J. Hyg. Epidemiol. Microbiol. Immunol. 10: 499-
Niklasson, B. 1988. Sindbis and Sindbis- like viruses. 509.
Pp. 167- 176 in The Arboviruses: Epidemiology Simkova, A. 1966b. Tahyna virus-neutralizingand Ecology( T. P. Monath, ed.). Volume IV, CRC antibodies in naturally infected domestic rabbitsPress, Boca Raton, Florida, U.S. A, 251 pp. and hares. Cesk. Epidemiol. Mikrobiol. Immunol.
Niklasson, B., A. Espmark, J. W. LeDuc, T. P. Gargan, 15: 304- 306.
W. A. Ennis, R. B. Tesh, and A. J. Main, Jr. 1984. Simkova, A. and V. Danielova. 1969. Experimental
Association of a Sindbis- like virus with Ockelbo infection of chimpanzees with Tahyna virus bydisease in Sweden. Am. J. Trop. Med. Hyg. 33: Culiseta annulata mosquitoes. Folia Parasitol.
1212- 1217. Praha) 16: 255- 263.
Niklasson, B., A. Espmark, and J. O. Lundstrom. 1988. Simkova, A., V. Danielova, and V. Bardos. 1960.
Occurrence ofarthralgia and specific 1gM antibodies Experimental transmission of the Tahyna virus bythree to four years after Ockelbo disease. J. Inf. Aedes vexans mosquitoes. Acta Virol. (Praha) 4:

36 BULL. SOC. VECTOR ECOL. JUNE, 1994
341- 347. encephalitis group viruses isolated from mosquitoesSimkova, A. and F. Sluka. 1973. Isolation of Tahyna collected in southern and arctic Norway. Acta.
virus from the blood of a case of influenza-like Pathol. Microbiol. Scand.( B) 86: 335- 341.disease. Acta Virol.( Praha) 17: 94. Traavik, T., R. Mehl, and R. Wiger. 1985. Mosquito-
Skogh, M. and A. Espmark. 1982. Ockelbo disease: borne arboviruses in Norway: further isolationsepidemicarthritis-exanthema syndrome in Sweden and detection of antiobodies to Californiacaused by Sindbis-virus like agent. Lancet 3: 795- encephalitis viruses in human, sheep and wildlife796. sera. J. Hyg.( Camb.) 94: 111- 122.
Smetana, A.,V. Danielova, J. M. Kolman, D. Malkova, Turell, M. J., J. O. Lundstrtim, and B. Niklasson. 1990.and J. Minar. 1967. The isolation of the Calovo Transmission of Ockelbo virus by Aedes cinereus,virus from the mosquitoes of the group Anopheles Ae. communis, and Ae. excrucians ( Diptera:maculipennis in southern Moravia. J. Hyg. Culicidae) collected in an enzootic area in centralEpidemiol. Microbiol. Immunol. 11: 55- 59. Sweden. J. Med. Entomol. 27: 266-268.
Smetana, A., D. Malkova, and Z. Marhoul. 1966. Vermeil, C., J. Lavillaureix, and E. Reeb. 1960. Sur laTahyna virus in squirrels Sciurus vulgaris. J. Hyg. conservation et la transmission du virus West NileEpidemiol. Microbiol. Immunol. 10: 523- 524. par quelques arthropodes. Bull. Soc. Pathol. Exot.
Traavik, T., R. Mehl, and R. Wiger. 1978. California Filiades 57: 273- 279.

BULL. SOC. VECTOR ECOL., 19( 1): 37- 42 JUNE, 1994
A CRITIQUE OF TECHNIQUES FOR DETECTION, IDENTIFICATION,
AND CLASSIFICATION OF SIBLING SPECIES OF MOSQUITOES
A. F. Cockburn1
ABSTRACT: Although the past few years have seen an explosion of new techniques in population genetics, none
of these techniques alone is best for answering all questions. A combination of restriction fragment lengthpolymorphism analysis, isozyme electrophoresis, and polytene chromosome cytology is a good way to detect crypticspecies, DNA sequencing is the best for phylogenetics, and DNA probes are best for routine identification ofindividual insects in well-characterized cryptic species complexes. A multidisciplinary approach, includingmorphology, is essential to the study of mosquito populations.
INTRODUCTION What would be useful is the integration ofall of the
disparate available techniques into a common
Morphology has been the foundation of taxonomy framework. Our group has been collaborating in afor the 2000 years following Aristotle. If two organisms multidisciplinary effort to classifyAnopheles mosquitoeslooked alike, they were the same species. If they looked throughout the world. We have molecular biologists,
different, they were different species. Taxonomic argu- cytologists, isozyme electrophoreticists, chemists, and
ments, reduced to discussions about what" look alike" ecologists working in our laboratory; and we aremeant, were couched in the language of morphology. collaborating closely with ecologists, morphologists,Ernst Mayr introduced the biological species concept, and electron microscopists. This cooperative experience
defining a species as a group of organisms sharing a has forced our group to learn to talk the language ofcommon gene pool. In doing so he effectively kicked several disciplines and given us some insight into the
over the stable foundation of taxonomy. In its place has advantages of the various technologies.
arisen a Tower ofBabel, as cryptic species are proposed
based on specialized techniques ( Fig. 1). Cytologists SPEED VS. INFORMATION
can only talk to other cytologists, isozyme electro-phoreticists can only talk to other isozyme electro- The two most important practical considerations in
phoreticists, and neither pay attention to the few evaluating techniques are( 1) the speed with which eachremaining morphologists. The invading horde of individual insect can be processed( usually proportionalmolecular biologists, each seemingly with a unique to the cost), and( 2) the amount of information generated
technique, cannot even understand each other. for each individual. Obviously, techniques that are slowSeductive as it seems, it would not be productive to and uninformative are bad, and those that are fast and
return to the simpler days of strictly morphological highly informative are good. However, usually there istaxonomy. The description of cryptic genetic species a trade-offbetween speed and information content( Fig.has explained many puzzling problems invectorecology. 2). For example, morphological analysis is slow but
Populations ofwhatappears to be a single morphological informative, while the use of DNA probes is fast butspecies may exhibit completely different behavior in relatively uninformative. Given this situation, whichdifferent areas. The most dramatic example is the technique is best for taxonomic work?
Anopheles gambiae complex, where one species, An. We have found that the answer depends on the
gambiae, is responsible for most malaria transmission questions being asked. Different questions require
in Africa, while a morphologically identical species, different techniques. Our group is interested in threeAnopheles quadriannulatus, does not even bite humans. questions:
1USDA/ ARS, Medical and Veterinary Entomology Research Laboratory, P. O. Box 14565, Gainesville, FL 32604,USA.

38 BULL. SOC. VECTOR ECOL. JUNE, 1994
1. How many cryptic species complexes exist? for cryptic species, so speed is important. If cryptic
2. What is the phylogeny of these species? species exist, they are likely to be closely related to each3. How can individual insects best be routinely other and share most characters. We would not be
identified for studies of ecology and vector willing to describe a new species based on a singlepotential? This last question is also relevant to character; rather we want to see a constellation of
vector control programs that need to monitor characters that separate species. Therefore, it is also
populations of noxious pests. important that wecollect as much infonnation as possible
on each individual.
Each of these three questions imposes its own set of Several techniques have the necessary balancetechnical constraints. I will consider them in order. between speed and information content. They include
polytene chromosome cytology, isozyme electro-CRYPTIC SPECIES DETECTION phoresis, and DNA restriction fragment length
polymorphisms( RFLPs). Both polytene chromosome
This is the most challenging problem that we face. cytology and isozyme electrophoresis have been usedIn some cases there is evidence for a cryptic species for decades and have proven their worth for detection of
complex, forexample, when a mosquito species transmits cryptic species. RFLPs, generally of mitochondrial ormalaria in one part of its range but not elsewhere. ribosomal DNA, have been used for about ten years and
Discussions with local vector control experts are have comparable power. My own expertise is in RFLPs,invaluable for discovering such anomalies. However, so I will briefly describe its usefulness. Similarcommentsthere is no substitute for the careful examination of large can be made about the other techniques.
numbers of wild collected specimens from throughout RFLP analysis consists ofenzymatic fragmentation
the range of the morph. of DNA from an individual mosquito, electrophoresis,
We routinely screen hundreds of such specimens and detection of specific fragments with DNA probes
TOWER OF BABEL
s .. ,„„
itlt,.10,..":: ,:
sits,:.
8
175:,.r,3
i
l• • i'. 5i ®
x,
7F3 ¢
9 4. S 4,a y5„
tS -:::. ,', ,,,.; .1`;''' - Atti: o,k,t• te''‘,4,3,
ei.9
t..
5
KY,y,
Y +,, d ` 3Y1,7
5 SS
R. YS 8 3' -
44.:";1,. '''', kt';'''''''''' 1.7•-,,t, itinreD4Jt41"4,...';'.
1''--raiTt -1".'' 41Ai
l'...V''' ''4kit'I-
41'''' -' ' t t" ii3 4.t.qp" °*ti WI' 84
Cip48,..AK 1.‘` , il,1414.1-* Ailni:1114 ti ,
113B^: a
7.'''g
ill g4 :$'j2Y , f ... IF;
14
Figure 1. Tower of Babel.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 39
Cockburn and Seawright, 1988). The techniques are that show pattern Al for enzyme 1, pattern A2 for
reliable and reproducible. These are critical enzyme 2, and pattern A3 for enzyme 3 will be common,
considerations foraproject in which hundreds ofsamples as will individuals that show pattern B1 for enzyme 1,
need to be analyzed. A technique that gives erratic pattern B2 for enzyme 2, and pattern B3 for enzyme 3.
results is useless. Each mosquito DNA sample is Individuals that show a mixture of the A and B patterns
divided into several aliquots and each aliquot is digested will be rare. Intraspecific variation, on the other hand,
with a different enzyme. The end results are a series of does not give such a dichotomy. We have simplygel lanes containing a specific pattern of bands for each extended the old morphological rule of, "insects that
individual insect. Different patterns are obtained with look alike are the same species," to include the bands
each different enzyme. We routinely use eight enzymes that they produce on gels.for each mosquito. If we detect interspecific differences using one
Mostindividualsofa species have identical patterns technique( for example, RFLPs) we should be able to
with the eight enzymes. Some individuals will have one detect differences between the same individuals usingor two bands different in one of the patterns. Different the other two techniques( polytene chromosome cyto-
species usually are quite distinct, with at least three or logy and isozyme electrophoresis). The advantage of
four of the patterns different. The actual data analysis is this approach is that the limitations that afflict one
more complex, involving mapping the enzyme cut sites technique are generally unrelated to those that affect theand estimation of the nucleotide divergence( Nei and Li, others. If two or more techniques show that a genetic
1979), but the presence of cryptic species is almost split occurs in a population, we can be confident that the
always evident from casual inspection of the gels. split is real. We are implementing this in practice in twoIf two cryptic species are present, there will be two ways. First, we analyze samples of field collections
common patterns for three or four of the enzymes, and using all three techniques. While it is possible to test thethese polymorphisms will be concordant. Individuals same individuals using more than one technique by
SEQUENCEBEST
MORPHOLOGY
DNA RFLPSPOLYTENES
ISOZYMES
0ILL
PCR RAPDS
WORST PROBES
SPEED -- I' m-
Figure 2. Sequence morphology.

40 BULL. SOC. VECTOR ECOL. JUNE, 1994
dividing the specimen, it is usually more trouble than it polytene chromosome cytology, but the procedure isis worth. If no significant variation is seen between probably similar. In addition, isozyme and polyteneindividuals, we assume that the population is homo- chromosome variants are likely to be under directgeneous. If we see evidence of genetic differentiation selection, so convergent evolution( as in the wings ofwith any of these techniques, we proceed to the next birds and mosquitoes) is possible.
step.When mosquitoes are collected in the field, most of ROUTINE IDENTIFICATION
the specimens are frozen for isozyme or DNA analysis.However, egg batches are collected from a subset of For field biologists, ecologists, epidemiologists,gravid females and each family is reared separately. and mosquito control personnel, the most importantMembers ofeach family, which areof the same species, question is: what is the fastest and easiest way ofcan be analyzed with all three techniques. Ifconcordant identifying large numbers ofcryptic species? Knowledgedifferences are seen, representative families can be of the existence of cryptic species is of no practical useanalyzed with every technique available, including until we are able to determine their relative importancemorphological analysis. The morphological taxonomists as pests or vectors. This requires ecological and vectorial
will know in advance which specimens belong to each studies followed by the implementation of surveillancegenetic species, which makes their analysis much programs. These types of studies may require thesimpler. identification ofthousands of individual insects. Because
of the large numbers of insects involved, speed and costPHYLOGENY are the critical factors. A large amount of information
does not need to be obtained from each individual. InPhylogenetic analysis requires only a limited fact, a simple reliable character would be best because
number of individuals from each species, so speed is not more information may be counterproductive if it slowsan important consideration. What is critical is the identification.
amount of information generated per individual since DNA probes are clearly the method of choice forthe statistical reliability of a phylogeny is dependent routine identification. Many groups have isolated DNAupon the number of characters examined. A second probes specific for certain species of mosquitoes,important point is that the characters be homologous, including An. gambiae ( Hill et al. 1991), An. diruswhich means related by common descent. For example, ( Panyin et al. 1988), An. quadrimaculatus species A, B,birds and mosquitoes both have wings, but these are not C ( Cockburn 1990; Johnson et al. 1993, An. farautihomologous structures. species 1, 2, 3( Cooper et al. 1991), An. rangeli( Perera
DNA( or RNA orprotein) sequencing is the genetic etal. 1993), andAn. aquasalis( Perera et al. 1993). DNAtechnique best suited for phylogenetics since there is an probes are also available for many pathogens, includingessentially unlimited number of characters( a mosquito viruses, bacteria, and protozoans. Recently, DNA probesgenome has at least 100,000,000 base pairs). Homo- have also been used for identification of blood mealslogous characters can easily be obtained from each ( Sato et al. 1992). There are several highly specific andspecies by sequencing the same genes. Essentially sensitive probes available for detection ofhuman blood.
every evolutionary biologist using molecular techniques The use of DNA probes has become very simple,forphylogenetics uses DNA sequencing. The temptation and field kits are now feasible. Johnson et al. ( 1992)to use other genetic techniques to generate phylogenies describe a procedure that involves placing individualis irresistible. For example, as soon as RFLP evidence mosquitoes in the 96 wells ofa microtiterplate, squashingfor a new cryptic species is found, these patterns are them in a small amount ofbuffer, and transferring somecompared to other closely related species and aphylogeny of the liquid to a membrane filter. The filters are treatedis generated. This habit is encouraged by the availability in batches to prevent further DNA or protein bindingofpowerful computer programs for turning genetic data and then incubated for a few hours with a solution ofinto phylogenies. While this is a harmless diversion, the labelled DNA probe. Excess probe is rinsed offand theresulting phylogenies should not be taken seriously. specifically bound probe is detected. The label can beThe phylogenetic trees created from RFLP, isozyme, a radioisotope, conjugated enzyme, or fluorescentand polytene chromosome data invariably rely on a molecule. Limited technical expertise is required tosmall numberofcharacters so the Statistical reliability is interpret the result. This is important, since the traininglow. A typical RFLP or isozyme study involves 20 or 30 ( or hiring) of professionals to interpret gels, chromo-characters, only a few of which are informative. It is somes, or morphology is the major cost in the routineharder to estimate the number ofcharacters involved in identification of insects.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 41
The detection procedures forDNAprobes are similar cytology. After sorting the population into geneticallyto those used with monoclonal antibodies. Kits based on homogeneous groups, we apply as many techniques asmonoclonal antibody technology are simple and highly possible to a few representative families of each type.
reliable; there is even an over-the-counter pregnancy We also send representatives of these families to
test kit for use at home by the general public. The U.S. collaborators to try other techniques, for exampleArmy has produced a field kit for detection ofmalaria in morphological analysis. Whenever possible we also
individual mosquitoes that has been widely used( Beier perform forced matings between groups to determine
et al., 1987). DNA probes are extremely reliable and can if they are interfertile. This accumulation of databe used with mosquitoes that have been dead in a light should allow us to determine whether these are sepa-
trap for several days, with no loss of specificity. rate species, and if so, to provide a comprehensive
Specimens can be transported at room temperature or description.
processed in the field. The availability of DNA probes Next, we try to isolate DNA probes specific forforpathogens and the ability toproduce multiple identical each species. This can be helpful even for similar, but
filters from each group of insects raises the possibility morphologically distinguishable species. In some casesthatDNA( orantibody) probescouldbeusedtodetermine examination of the genetically separated specimensthe species ofeach individual, what pathogens it carries, reveals concordant morphological differences that might
and the source of the latest blood meal. A simple previously have been disregarded as " intraspecificstandardized test to determine all three of these pieces of variation." For example, we recently have found threeinformation would be enormously useful. new genetic species in the Anopheles crucians complex
near our laboratory in Gainesville. Two of these newDISCUSSION species are easily morphologically distinguishable from
the previously described species in the complex. TheseWe have put the principles outlined above into types must have been collected by dozens of entomo-
effect in our own research program( Fig. 3). When we logists, but no one had ever reported that the morpho-
begin studying a population, we screen field collections logical variants are in fact distinct species.
with DNA RFLPs, isozyme electrophoresis, and polytene Finally, the descriptions and DNA probes should
CHARACTERIZING NEW
CRYPTIC SPECIES
IDENTIFY CONFIRM STUDY
r: 0.1: 7, 4'" "ope.
to4,,,;;;.., ,&4,, lx.„ 7_,..
t, ,:ilt, tiv.::::-,
1441!ii --ON- ria„.,.,,,.1,„,„..0, 7s7,,,.„.„Ari:4:,‘,,isTE-,,
et,., ,,,,,,,,
t ,,. ,.. ,, t,,,,:,„, c,... . -,, :., ,,,,,ys`'
Y
Ilr
a , Y
iisli .,11,,. 13,..„... i .,) ...,,
Figure 3. Characterizing new cryptic species.

42 BULL. SOC. VECTOR ECOL. JUNE, 1994
provide the tools necessary for ecologists and vector of sibling species within the Anopheles gambiaebiologists. They will determine the biologically complex. Med. Vet. Entomol 5: 455- 463.
interesting pieces of the puzzle: where these species Johnson, D. W., A. F. Cockburn, and J. A. Seawright.live, what they feed on, and what diseases they transmit. 1992. Quick blots and nonradioactive detection ofThis information will allow vector control agencies to DNA probes for the identification ofmosquitoes. J.make more infonned decisions about when and where to Am. Mosq. Contr. Assn. 8: 231- 236.intervene. Johnson, D. W., A. F. Cockburn, and J. A. Seawright.
1993. Sequence of a DNA probe specific forREFERENCES Anopheles quadrimaculatus species A ( Diptera:
Culicidae). J. Med. Entomol. 30: 939-942.Beier, J. C., P. V. Perkins, R. A. Wirtz, R. E. Whitmire, Nei, M. and W. H. Li. 1979. Mathematical model for
M. Mugambi, and W. T. Hockmeyer. 1987. Field studying genetic variation in terms of restrictionevaluation of an enzyme- linked immunosorbent endonucleases. Proc. Nat.Acad. Sci.USA 76: 5269-
assay ( ELISA) for Plasmodium falciparum 5273.
sporozoite detection in anopheline mosquitoes Panyim, S., S. Yasothronsrikul, S. Tungpradubkul, V.from Kenya. Am. J. Trop. Med. Hyg. 36: 459- Baimai, R. Rosenberg, R. G. Andre, and C. A.68. Green. 1988. Identification of isomorphic malaria
Cockburn, A.F. 1990. A simple and rapid technique for vectors using a DNA probe. Am. J. Trop. Med.identification of large numbers of individual Hyg. 38: 47-49.mosquitoes using DNA hybridization. Arch. Insect Perera, 0. P., A. F. Cockburn, S. E. Mitchell, J. Conn,Biochem. Physiol. 14: 191- 199. and J. A. Seawright. 1993. Isolation of species
Cockbum, A.F. and J. A. Seawright. 1988. Techniques specific DNA probes for the South Americanfor mitochondria) and ribosomal DNA analysis of malaria vectors Anopheles aquasalisandA. rangeli.anopheline mosquitoes J. Am. Mosq. Contr.Assoc. Pp. 63- 69 in Host Regulated Developmental3: 261- 265. Mechanisms in Vector Arthropods, vol. 3 ( D.
Cooper, L., R. D. Cooper, and T. R. Burkot. 1991. The Borovsky and A. Spielman, eds.). University ofAnopheles punctilatus complex: DNA probes for Florida, 328 pp.identifying the Australian species using isotopic, Sato, C., Y. Furuya, M. Harada, and S. Suguri. 1992.chromogenic, and chemiluminescence detection Identification of human blood in mosquitoessystems. Exp. Pathol. 73: 27- 35. Diptera: Culicidae) using nonradioactive DNA
Hill, S. M., R. Urwin, T. F. Knapp, and J. M. Crampton. dot blot hybridization. J. Med. Entomol. 29: 1045-1991. Synthetic DNA probes for the identification 1048.

BULL. SOC. VECTOR ECOL., 19( 1): 43- 48 JUNE, 1994
THE IMPACT OF WATER QUALITY ON THE
PERSISTENCE OF BACILLUS SPHAERICUS
M. Ludwig', M. Beck', M. Zgomba2, and N. Becker'
ABSTRACT: The factors that influence the persistence of Bacillus sphaericus are still not completely understood.The impact ofwater quality and oxygen on the long-teen efficacy was investigated by means of bioassays and sporecounts. Bacillus sphaericus persisted in distilled water as well as in hay- infusion for at least three months. High ratesof persistence were observed for the first 14 days and a significant decrease of efficacy occurred after one month.In both test series a drop ofefficacy to 11 percent was recorded after three months. An increase in efficacy or increasein spore numbers due to recycling phenomena was not evident. Aeration had a significant influence on the
persistence ofB. sphaericus. When air was introduced continuously into both a hay- infusion and distilled water, B.sphaericus lost its efficacy after two months.
INTRODUCTION MATERIAL AND METHODS
Bacillus sphaericus has been shown to be highly To obtain different water qualities, 5 liter glass-
insecticidal to larvae of Culex spp. and some Anopheles vessels ( 10 per group) were filled with the followingspp. mosquitoes ( Mulligan et al. 1978; Mulla et al. solutions:
1984a, 1984b; Singer 1985; Mulla 1986; Lacey et al.1987). An advantage of B. sphaericus over Bacillus a) hay- infusion, nonaeratedthuringiensis var. israelensis is that it provides greater b) hay- infusion, aerated( introduction of air intoresidual larvicidal efficacy, apparently due to the longer the solution)
persistence of its toxic crystals or possible recycling c) distilled water, nonaerated
Hertlein et al. 1979, Mulligan et a1. 1980, Lacey 1984). d) distilled water, aerated
In sewer water, which represents a typical breeding sitefor Culex spp. ( O'Meara and Evans 1983, Mulla et al. For the hay- infusion, 500 grams ofdried and cut-up1987), B. sphaericus was shown to persist for a longer hay were mixed with 50 liters of spring water in 70 literperiod of time due to protection of the crystals from the plastic barrels at 25° C. After one week this suspension
damaging influence of the sunlight( Des Rochers and was filtered and the filtrate placed into the vessels.
Garcia 1984, Becker et al. 1993). However, published Three vessels of each group were treated with B.reports regarding the extent of the residual efficacy ofB. sphaericus 2362 ( Spherimos FC, unradiated, Novo
sphaericus, as well as reports concerning environmental Nordisk, Denmark) at a dosageof 10 mg/ l.The remainingfactors that may influence this phenomenon, present vessels with untreated suspensions were used for control
conflicting results( Hertlein et al. 1979, Mulligan et al. and dilution. The vessels were kept in a room with 16
1980, Nicolas et al. 1987, Hougard 1990, Lacey 1990). hours of artificial light per day at 20°C. The fate of theWith a view to more economical control of Culex active ingredient was evaluated by bioassays of
pipiens, the objective of the present study was to assess suspensions a, b, c, and d at 0, 7, 14, 30, 60, and 90 days
the influence of water quality and aeration on the post B. sphaericus treatment.
persistence of B. sphaericus. All bioassays were carried out according to WHO
German Mosquito Control Association( KABS), Ludwigstr. 99, 67165 Waldsee, GERMANY.
2University of Novi Sad, Inst. of Plant Protection, Trg. D. Obradovica 8, 21000 Novi Sad, YUGOSLAVIA
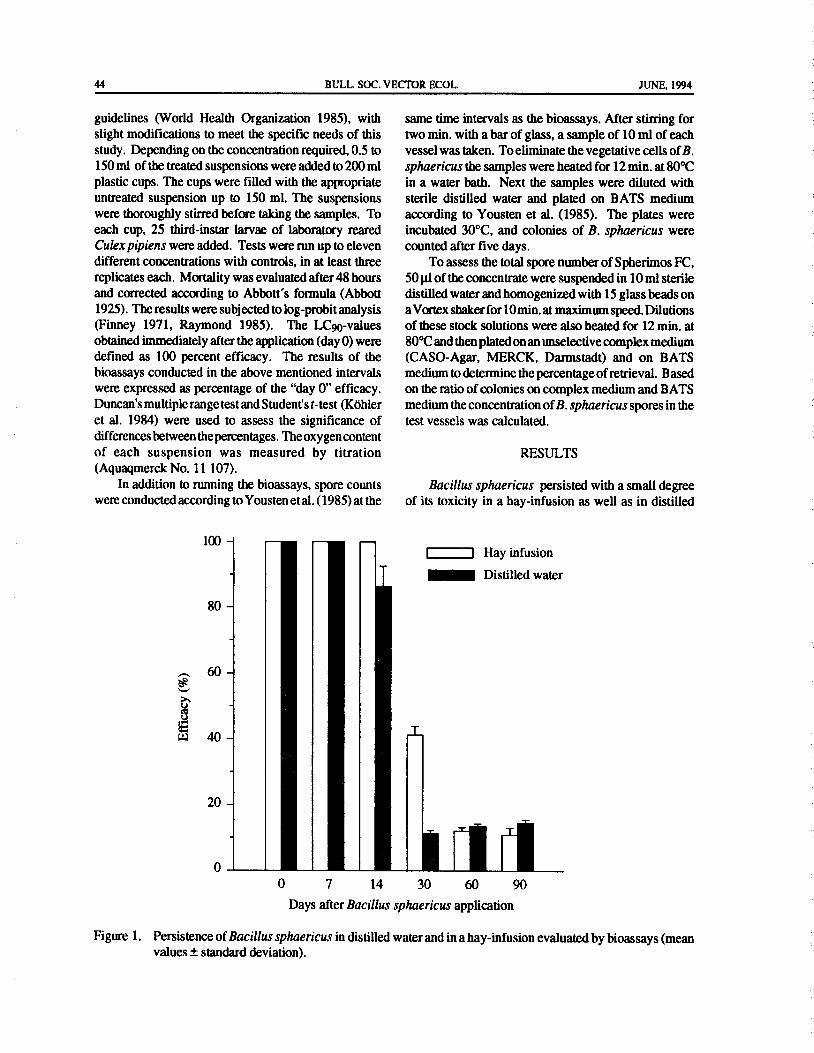
44 BULL. SOC. VECTOR ECOL. JUNE, 1994
guidelines ( World Health Organization 1985), with same time intervals as the bioassays. After stirring forslight modifications to meet the specific needs of this two min. with a bar of glass, a sample of 10 ml of each
study. Depending on the concentration required, 0.5 to vessel was taken. To eliminate the vegetative cells ofB.
150 ml of the treated suspensions were added to 200 ml sphaericus the samples were heated for 12 min. at 80°C
plastic cups. The cups were filled with the appropriate in a water bath. Next the samples were diluted with
untreated suspension up to 150 ml. The suspensions sterile distilled water and plated on BATS medium
were thoroughly stirred before taking the samples. To according to Yousten et al. ( 1985). The plates were
each cup, 25 third- instar larvae of laboratory reared incubated 30°C, and colonies of B. sphaericus wereCulex pipiens were added. Tests were run up to eleven counted after five days.
different concentrations with controls, in at least three To assess the total spore number of Spherimos FC,replicates each. Mortality was evaluated after 48 hours 5011l of the concentrate were suspended in 10 ml sterileand corrected according to Abbott' s formula( Abbott distilled water and homogenized with 15 glass beads on1925). The results were subjected to log-probit analysis a Vortex shaker for 10 min.at maximum speed. Dilutions
Finney 1971, Raymond 1985). The LC90-values of these stock solutions were also heated for 12 min. atobtained immediately after the application( day 0) were 80°C and then plated on an unselective complex mediumdefined as 100 percent efficacy. The results of the ( CASO-Agar, MERCK, Darmstadt) and on BATSbioassays conducted in the above mentioned intervals medium to determine the percentageofretrieval. Basedwere expressed as percentage of the" day 0" efficacy. on the ratio of colonies on complex medium and BATS
Duncan's multiple range test and Student's t-test( Kohler medium the concentration ofB. sphaericus spores in theet al. 1984) were used to assess the significance of test vessels was calculated.
differences between the percentages. The oxygen content
of each suspension was measured by titration RESULTS
Aquaqmerck No. 11 107).
In addition to running the bioassays, spore counts Bacillus sphaericus persisted with a small degreewere conducted according to Yousten et al.( 1985) at the of its toxicity in a hay- infusion as well as in distilled
100 —
1 I Hay infusion
Distilled water
80 —
60 -se
W 40 — L
20 —
0
0 7 14 30 60 90
Days after Bacillus sphaericus application
Figure 1. Persistence ofBacillus sphaericus in distilled water and in a hay- infusion evaluated by bioassays( meanvalues± standard deviation).

JUNE, 1994 BULL. SOC. VECTOR ECOL. 45
water for at least three months( Fig. 1). Bioassays with within the nonaerated distilled water ( 86.4 %) was
distilled waterdemonstrated that high ratesofpersistence higher than within the aerated distilled water( 69.9%).
occurred for the first 14 days, but a significant decrease After twomonths the residual effect within the nonaerated
P<_0.001) of efficacy was recorded after one month. distilled water was five-fold higher than within the
Efficacy of B. sphaericus on day 30 of the experiment aerated distilled water. After three months exposure
dropped to 11. 4 percent of the efficacy recorded on day within the aerated distilled water B. sphaericus had lost
0. Up to three months no further decrease of the efficacy all its efficacy.was observed. Spore counts support the results obtained by means
Efficacy within the hay- infusion showed high rates ofbioassays( Fig. 3). On day 0 the number ofspores wasofpersistence within the first 14 days as well( Fig. 1). A approximately 7.7 x 104 spores/ml in all four test series.significant decrease( P<_0.001) was observed after one After two weeks the spore number was reduced in all
month( 41. 3 %). After three months only 11. 0 percent test series. The reduction within the hay-infusion wasof the initial larvicidal efficacy remained. Student's t- greater than in distilled water. Aeration had a negative
test demonstrated that LC values of the test series with impact on the number of spores in hay- infusion as welldistilled water were significantly lower (P<_ 0.01) in as in distilled water. On day 60 the number of sporescomparison to the LC values determined in the hay- decreased 180- fold within the nonaerated hay- infusion,infusion( TABLE 1). Figure 2 shows the persistence of 230-fold within the aerated hay- infusion, 50- fold withinB. sphaericus within an aerated and a nonaerated hay- nonaerated distilled water and 60-fold within aerated
infusion. The content of oxygen was 0 mg/ 1 within the distilled water.
nonaerated hay- infusion, whereas the oxygen content ofthe aerated hay- infusion was 8- 9 mg/ l. DISCUSSION
The efficacy of B. sphaericus was significantlyhigher within the aerated hay infusion on day 0( TABLE Bacillus sphaericus persists and remains toxic in
1). After seven days of exposure the efficacy within the distilled water as well as in a hay- infusion for at leastaerated test series was significantly (P<_ 0.01) higher three months. This is in accordance with a report ofDes
than in the nonaerated test series. After 14 days exposure Rochers and Garcia( 1984) who found that B. sphaericus
this difference increased. After one month the efficacy persists in toxicity up to 30 days in different waterof the aerated hay- infusion dropped to 2.6 percent, qualities. Hertlein et al.( 1979) showed that B.sphaericus
whereas the efficacy in the nonaerated hay infusion was remained viable and infective nine months after
still 41. 3 percent at this time. After two months of application as a larvicidal agent of mosquitoes in a
exposure within the aerated hay- infusion B. sphaericus roadside ditch. In a field experiment by Nicolas et al.had lost all its efficacy. 1987), persistence ofsedimented spores ofB. sphaericus
Figure 2 compares the persistence of B. sphaericus in mud was shown to exceed five months. Matanmi et
within aerated and nonaerateddistilled water.The content al. ( 1990) found in bioassays of bottom water with B.
of oxygen was 2 mg/ 1 within the nonaerated and 9 mg/ sphaericus against fourth-instars of Culex quinque-
1 within the aerated test series. Results were similar to fasciatus only moderate to low larvicidal efficacy forthe test series conducted with aerated and nonaerated about 14 days. An influence of the water quality on thehay- infusion. On day 0 of the test the efficacy of B. persistence of B. sphaericus was not recorded. This is
sphaericus was nearly equal in both series( TABLE 1). in contrast to results of Hornby et al. ( 1984) who
However, after 14 days the efficacy of B. sphaericus conducted tests in sewer and freshwater ponds in Florida
TABLE 1. LC values( mean values± standard deviation) of Bacillussphaericus( Spherimos
FC) in different water qualities obtained immediately after application
Treatment LC50 LC90
hay- infusion 0.3065± 0.0314 0.9163± 0.1146
hay- infusion, aerated 0. 1621± 0.0083 0.3631± 0.0164
distilled water 0.0484± 0.0135 0.1977± 0.0211
distilled water, aerated 0.0235± 0.0033 0.2138± 0.00137
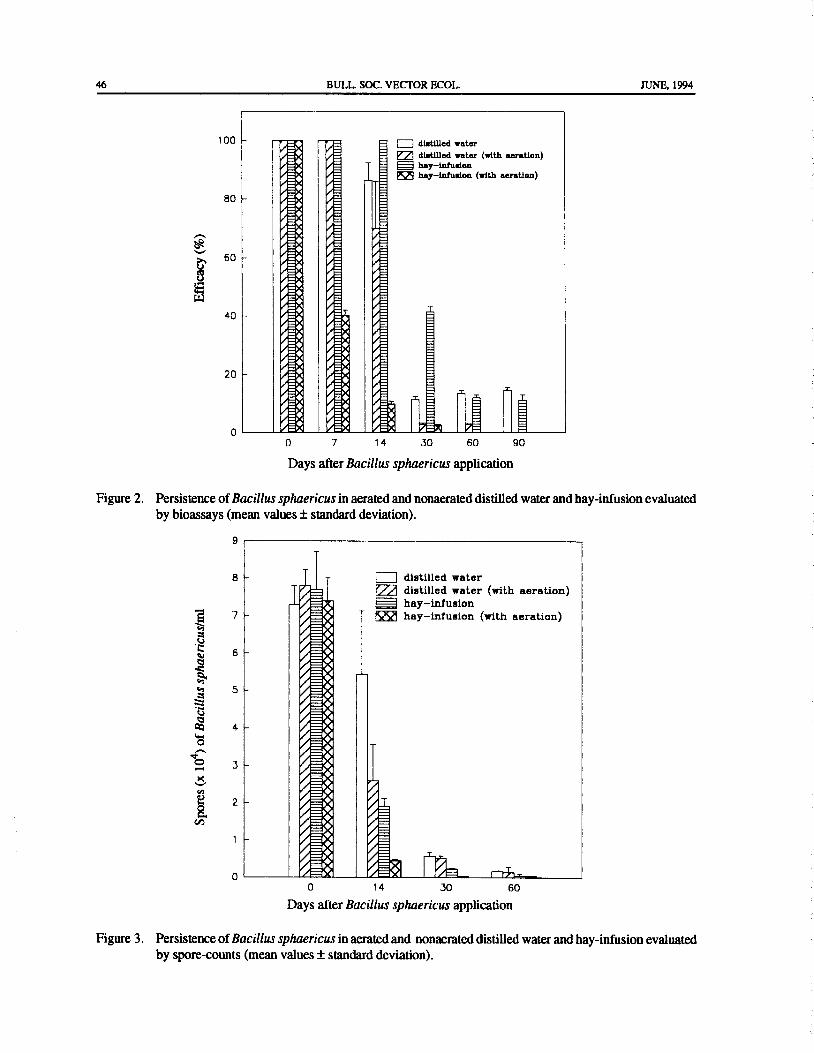
46 BULL. SOC. VECTOR ECOL. JUNE, 1994
100 - ,EA ,_ 1 I distilled water
A, /_ distilled water ( with aeration)
0/hay—infusion
A' O_ hay—infusion( with aeration)
80 - O__X/ _4
OE=
W /
EROED O_ '
0-EX OE
OEOEOE
w OE40 - OEA iiI=
l
P'A A
c' Et 0=_20 - / EP, /_/
E / E _
o0 7 14 30 60 90
Days after Bacillus sphaericus application
Figure 2. Persistence of Bacillus sphaericus in aerated and nonaerated distilled water and hay-infusion evaluatedby bioassays( mean values± standard deviation).
9 )
8 - I I distilled water
ea distilled water ( with aeration)
VII hay- infusion7 S 4 hay- infusion ( with aeration)
61
d 15 VEp4ti 5 - I_1
1•
Cri 4 - l•O Q
ao 3 - i•1
E 2 - 1
a1
01
1•
1 - 0401
1
I" 131=_00 14 30 60
Days after Bacillus sphaericus application
Figure 3. Persistence of Bacillus sphaericus in aerated and nonaerated distilled water and hay- infusion evaluatedby spore-counts( mean values± standard deviation).

JUNE, 1994 BULL. SOC. VECTOR ECOL. 47
and found that spores of B. sphaericus persisted for Control of Mosquitoes and Black Flies ( H. de
periods ranging from 30 to 90 days depending on the Barjac and D. J. Sutherland, eds.). Rutgers Univ.
quality of water. These investigators reported 100 Press, 349 pp.percent control of Cx. quinquefasciatus and Culex Kohler, W. G., G. Schachtel, and G. Voleske. 1984.
nigripalpus for 80 to 90 days in sewer water with only Biometrie. Springerverlag, Berlin, Heidelberg, Newone treatment of 2.5- 7. 5 x 104 spores of B. sphaericus York, 230 pp.ml. Similar results were obtained in experiments Lacey, L. A. 1984. Production and formulation of
conducted in fresh water but control lasted only 30 to 50 Bacillus sphaericus. Mosq. News 44: 153- 159.days. These authors speculated that the sewage habitat Lacey, L. A. 1990. Persistence and formulation of
with its high organic debris maintained at toxic levels of Bacillus sphaericus. Pp. 284-295 in BacterialB. sphaericus for a longer period of time by protecting Control of Mosquitoes and Black Flies ( H. de
the toxin from UV light. Barjac and D. J. Sutherland, eds.). Rutgers
Oxygen had a negative effect on the persistence of University Press, 349 pp.B. sphaericus. A constant supplement of oxygen seems Lacey, L. A., J. Day, and C. M. Heitzmann. 1987.
to accelerate the decomposition processes of the toxin. Longterm effects of Bacillus sphaericus on Culexquinquefasciatus. J. Invertebr. Pathol. 49: 116-
Acknowledgments 123.
Matanmi, B. A., B. A. Federici, and M. S. Mulla. 1990.
This investigation received financial support from Fate and persistence ofBacillus sphaericus used as
the UNDP/ World Bank/WHO Special Programme for a mosquito larvicide in dairy wastewater lagoons.Research and Training in Tropical Diseases. The study J. Am. Mosq. Contr. Assoc. 6: 384- 389.was also financially supported by the" Gesellschaft fur Mulla, M. S. 1986. Efficacy of the microbial agentinternationalen Erfahrungsaustausch bei der Bacillus sphaericus against mosquitoes ( Diptera:
Stechmuckenbekampfung e. V. - GiES," Waldsee, Culicidae) in southern California. Bull. Soc. Vector
GERMANY. The authors thank Dr. Mir S. Mulla, Ecol. 11: 247-254.
University of California, for his valuable comments Mulla, M. S., H. A. Darwazeh, E. W. Davidson, H. T.
concerning the manuscript. Dulmage, and S. Singer. 1984a. Larvicidal efficacyand field efficacy of Bacillus sphaericus strains
REFERENCES CITED against mosquito larvae and their safety to nontarget
organisms. Mosq. News 44: 336-342.Abbott, W. S. 1925. A method of computing the Mulla,M. S., H. A. Darwazeh, E. W. Davidson, and H.
effectiveness of an insecticide. J. Econ. Entomol. T. Dulmage. 1984b. Efficacy and persistence of18: 265- 267. the microbial agent Bacillus sphaericus against
Becker, N., M. Ludwig, M. Beck, and M. Zgomba. mosquito larvae in organically enriched habitats.1993. The impact of environmental factors on the Mosq. News. 44: 166- 173.efficacy of Bacillus sphaericus against Culex Mulla, M. S., L. S. Mian, and N. G. Gratz. 1987.
pipiens. Bull. Soc. Vector Ecol. 18( 1): 61- 66. Agricultural management practices- their impacts
Des Rochers, B. and R. Garcia. 1984. Evidence for on production of vector and pest mosquitoes. J.
persistence and recycling of Bacillus sphaericus. Agric. Entomol. 4: 97- 131.
Mosq. News 44: 160- 165. Mulligan, F. S., C. H. Schaefer, and T. Miura. 1978.
Finney, D. J. 1971. Probit analysis, 3rd edition. Laboratory and field evaluation of BacillusCambridge Univ. Press, 334 pp. sphaericus as a mosquito control agent. J. Econ.
Hertlein, B. C., R. Levy, and T. W. Miller. 1979. Entomol. 71: 774-777.
Recycling potential and selective retrieval of Mulligan, F. S., C. H. Schaefer, and W. H. Wilder.
Bacillus sphaericus from soil in a mosquito habitat. 1980. Efficacy and persistence of B. sphaericusJ. Invertebr. Pathol. 33: 217- 221. and B. thuringiensisH- 14 against mosquitoes under
Hornby, J. H., B. C. Hertlein, and T. W. Miller. 1984. laboratory and field conditions. J. Econ. Entomol.Persistent spores and larvicidal efficacy ofBacillus 73: 684-688.
sphaericus 1593 in wellwater and sewage. J. Nicolas, L.,J. Dossou- Yovo, and J. M. Hougard. 1987.
Georgia Entomol. Soc. 19: 165- 167. Persistence and recycling of Bacillus sphaericusHougard, J. M. 1990. Formulations and persistence of 2362 spores in Culex quinquefasciatus breeding
Bacillus sphaericus in Culexquinquefasciatus larval sites in West Africa. Appl. Microbiol. Biotechnol.
sites in tropical Africa. Pp. 295- 306 in Bacterial 25: 341- 345.

48 BULL. SOC. VECTOR ECOL. JUNE, 1994
O'Meara, G. F. and F. D. S. Evans. 1983. Seasonal Chapman, ed.). Am Mosq. Control Assoc. Bull. 6,patterns ofabundance among three species ofCulex 243 pp.mosquitoes in a south Florida wastewater lagoon. WorldHealth Organization. 1985. Informal consultationAnn. Entomol. Soc. Am. 48: 133- 138. on the development of Bacillus sphaericus as a
Raymond, M. 1985. Presentation d' un programme microbial larvicide. W. H. O./ TDR/BCV/d' analyse log-probit pour micro-ordinateur. Cah. SPHAERICUS/ 85: 3. 24 pp. Genf.ORSTOM Ser. Entomol. Med. Parasitol. 22: 117- Yousten, A. A., S. B. Fretz, and S. A. Jelley. 1985.121. Selective medium for mosquito-pathogenic strains
Singer, S. 1985. Bacillus sphaericus ( Bacteria). Pp. of Bacillus sphaericus. Appl. Environ. Microbiol.123- 131 in Biological control ofmosquitoes( H. C. 49: 1532- 1533.

BULL. SOC. VECTOR ECOL., 19( 1): 49-52 JUNE, 1994
COLOR AND UV REFLECTANCE OF CANOPY TRAPS FOR
COLLECTING HORSE FLIES ( DIPTERA: TABANIDAE) IN LOUISIANA
L. J. Hribar1, 2 and L. D. Foil'
ABSTRACT: The effect ofcolor and ultraviolet light( UV) reflectance on the catch ofhorse flies in carbon dioxide-
baited canopy traps was studied in south- central Louisiana. Black traps collected significantly more Tabanusamericanus Forster, Tabanus fuscicostatus Hine, Tabanus limbatinevris Macquart, Tabanus lineola group, andTabanus proximus Walker than did blue or red traps. Tabanus wilsoni Pechuman was captured in greater numbersin black traps than in red traps, whereas, there was no difference between catch of this species in black and blue trapsor in blue and red traps. There was no difference in the number of flies captured by the different colored traps forChlorotabanus crepuscularis( Bequaert), Chrysops spp., Leucotabanus annulatus( Say), Tabanus equalis Hine, andTabanus pallidescens Philip. More flies of the T. lineola group were captured in traps with increased or decreasedUV reflectance than in control traps. Traps with decreased UV reflectance captured more T. proximus than did
control traps, but there were no differences in number of T. proximus captured in control traps and traps withincreased UV reflectance, nor between traps with increased or decreased UV reflectance. Black traps are most
effective for collecting Tabanidae in southern Louisiana during midsummer. There appears to be little effect of UVreflectance on collection of most species of Tabanidae.
INTRODUCTION Area, near the town ofWashington in St. Landry Parish,Louisiana. The study site has been described previously
Canopy traps have been used for collecting by Leprince et al. ( 1991). Canopy traps ( Hribar et al.Tabanidae since Thorsteinson ( 1958) invented the 1991b) were used to collect horse flies. All traps were
Manitoba trap." Catts'( 1970) modification of this trap baited with ca. 5 kg of dry ice. The cluster design
has become a standard tool for sampling tabanid ( Hribar et al. 1991a) was chosen over other designs
populations. This trap is mainly black in color. Several ( e. g., Latin square, transects) to avoid site differencesauthors have reported that horse flies also respond to among traps. Traps within clusters were separated byother colors, especially blue ( Allan and Stoffolano only 2 to 5 meters. The carbon dioxide bait was used to1986, Browne and Bennett 1980, Neys et al. 1971, Hays attract flies to the clusters, where the differences among1968) and red( Bracken et al. 1962, Browne and Bennett traps ( color or UV reflectance) presumably would1980, Granger 1970, Hansens et al. 1971, Hays 1968, account for different numbers of flies collected by theRoth and Lindquist 1948, Snoddy 1970, Thorsteinson et traps. Due to the difficulty of separating Tabanusal. 1964). Ultraviolet light reflectance affects horse fly lineola F. and Tabanus subsimilis Bellardi, these species
host-seeking behavior( Allan& Stoffolano 1986), and were identified as the T. lineola group. Similarly, mostreducing the amount of ultraviolet light( UV) reflected deer flies were identified only to genus. Data were
from canopy traps increases numbers of horse flies analyzed by the SAS ANOVA procedure and Tukey' scaptured( Hribar et al. 1991a). This paper reports two mean separation test( SAS Institute 1987).
studies conducted to determine the effects of trap colorand UV reflectance on catch of tabanids. Color of Traps
Twelve newly-sewn canopy traps were placed inMATERIALS AND METHODS four clusters of three traps each. The skirt of the trap was
made from either red( Flare Red), blue( Blueprint), or
All experiments were conducted during June and black Tee-shot poplin fabric. Placement of traps by
July 1991 at the Thistlethwaite Wildlife Management color within clusters was assigned randomly with the
tDepartment of Entomology, Louisiana Agricultural Experiment Station, Louisiana State University AgriculturalCenter, Baton Rouge, LA 70803- 1710, USA.
2Present address: Florida Medical Entomology Laboratory, University of Florida, 200 Ninth Street S. E., VeroBeach, Florida, 32962, USA.

50 BULL. SOC. VECTOR ECOL. JUNE, 1994
exception that one trap of each color was included in Tabanus fuscicostatus Hine, Tabanus limbatinevrisevery cluster. Flies were collected daily at sunset for 12 Macquart, Tabanus lineola group, Tabanus molestusconsecutive days from 24 June- 5 July 1991. Say, Tabanus pallidescens Philip, Tabanus proximus
Walker, Tabanus trimaculatus Palisot de Beauvois,UV Reflectance Tabanus turbidus Wiedemann, Tabanus venustus Osten
Nine canopy traps were placed in three clusters of Sacken, Tabanus wilsoni Pechuman, and Whitneyomyiathree traps each. The poplin skirt of one trap in each beatifica atricorpus Philip. The most commonlycluster was sprayed with one bottle( about 532 ml) of collected species were T.fuscicostatus( 52.5%) and T.UV Killer® ( Atsko Inc., Orangeburg, S. C.), a lineola( 25.0%). Those species collected in numbers
commercially available product that reduces the amount sufficient for analysis are presented in TABLE 1.ofUV reflected from cloth fabric. Asecondtrapineach Black traps captured significantly more T.cluster was sprayed until runoff with Tinopal®( Ciba- americanus, T.fuscicostatus, T.limbatinevris, T. lineolaGeigy, Greensboro, N.C.), a brightening agent used in group, and T. proximus than did blue or red traps.laundry detergents to increase UV reflectance of cloth Tabanus wilsoni was captured in black traps more oftenfabric. The brightener was applied at a rate of 0.8 g/ l, than in red traps, but there was no difference in catchwhich is about the amount used in one washing machine between black and blue traps or between blue and redload( Silo Corp., Baton Rouge, La., pers. comm.). A traps for this species. Chlorotabanus crepuscularis,third trap in each cluster was an untreated control. Chrysops spp., L. annulatus, T. equalis, and T.Treatments were assigned randomly within clusters. pallidescens were captured equally in traps ofall colors.Flies were collected daily at sunset for 12 consecutivedays from 5 July- 17 July 1991. UV Reflectance
A total of 7, 291 horse flies in four genera and 19RESULTS species or species groups was collected: Chrysops spp.
mainly C. cursim, C. pudicus, and C. univittatus), C.Color of Traps crepuscularis, Hybomitra lasiophthalma ( Macquart),
A total of 15, 144 horse flies in five genera and 18 L. annulatus, T. americanus, T. atratus, T. calens, T.species or species groups was collected: Chrysops spp. equalis, T. fuscicostatus, T. limbatinevris, T. lineolamostly Chrysops cursim Whitney, Chrysops pudicus group, T. molestus, T. nigripes Wiedemann, T.
Osten Sacken, and Chrysops univittatus Macquart), pallidescens, T.proximus, T. trimaculatus, T.turbidus,Chlorotabanuscrepuscularis( Bequaert), Leucotabanus T. venustus, and T. wilsoni. The most commonlyannulatus( Say), Tabanus americanus Forster, Tabanus collected species were T.lineola( 33.4%), T.fuscicastarusatratus F., Tabanus calens L., Tabanus equalis Hine, ( 30.5%), and T. proximus ( 20.5%). Those species
TABLE 1. Mean numbers of horse flies captured/ trap/day in colored canopy traps, ThistlethwaiteWildlife Management Area, 1991( means within rows followed by the same letter are notsignificantly different, Tukey' s mean separation procedure).
Total Color
Species Collected Black Blue Red Probability
Chlorotabanus crepuscularis 64 0.51a 0.478 0.28a P> 0.3'Chrysops spp. 875 6.048 6.558 4.578 P> 0.2Leucotabanus annulatus 38 0.20a 0.39a 0.168 P> 0.05Tabanus americanus 690 8.29a 3. 65b 1. 59b P< 0.0001Tabanus equalis 123 0.78a 1. 068 0.578 P> 0. 1Tabanus fuscicostatus 7958 70. 148 47.43b 38.47b P< 0.0035Tabanus limbatinevris 193 2.268 0.80b 033b P< 0.0003Tabanus lineola group 3791 43. 578 18. 86b 11. 90b P< 0.0001Tabanus pallidescens 170 1. 478 1. 098 0.78a P> 0. 1Tabanus proximus 946 13.008 2. 53b 3.021' P< 0.0001Tabanus turbidus 46 0.248 0.478 0.208 P> 0.2Tabanus wilsoni 205 1. 77a 1. 53ab 0.73bc P< 0.007

JUNE, 1994 BULL. SOC. VECTOR ECOL. 51
collected in numbers sufficient for analysis are presented numbers of flies landing on that color. Granger' s( 1970)in TABLE 2. study was limited by no statistical analysis of trap catchTraps with increased or decreased UV reflectance comparisons of 16 traps, four trap colors, and a catch ofcaptured the T. lineola group in greater numbers than 1, 621 T.nigrovittatus. In our study, grouping the threedid control traps. More T.proximus were collected in colors together and baiting each trap with carbon dioxidetraps with decreased UV reflectance than were collected made the entire cluster attractive from a distance. Colorin control traps; however, there was no difference in of traps was important only after the flies arrived at the
number of T.proximus collected from traps treated with cluster. Black traps were strongly attractive to all of theUV Killer or brightener, nor between traps treated with most commonly collected species. Those species thatbrightener and control traps. were captured equally by the traps were those that werecaptured in very small numbers; these are species for
DISCUSSIONwhich the canopy trap generally is ineffective or whichare rarely collected in canopy traps. Under theseColor of Trapsexperimental conditions, black would be the mostThe collection of tabanids with traps of different effective color for canopy traps in southern Louisiana.
colors has been inconsistent as documented by differentobservers. For example, Bracken et al.( 1962), Snoddy UV Reflectance1970), Hays( 1968), and Browne and Bennett( 1980) The highest numbers of the T. lineola group and T.reported collecting equivalent numbers of tabanids proximus were collected in traps treated with UV Killer.
landing on sticky black and red surfaces. Hansens et al. Traps treated with brightener also had higher collections1971), however, found that black sticky traps were ofthe T.lineola group. There was no difference betweensuperior to red traps. Granger( 1970) found red Manning collections of T. proximus in traps treated with UVtraps to be superior to black traps for capture of T. Killer and with brightener, nor between collections ofnigrovittatus Macquart. Neys et al. ( 1971) found that traps treated with brightener and with controls.black decoys suspended from canopy traps were more The responses of horse flies to the different UVeffective than blue decoys. Allan and Stoffolano( 1986) treatments revealed that those species that were affectedfound that T.nigrovittatus landed on blue sticky panels by the treatment of canopy traps were captured inmore frequently than black and red panels. equivalent numbers both by traps treated with UV KillerAll of the aforementioned studies except that of and by traps treated with brightener. Hribar et al.Granger( 1970) involved the interaction of two different ( 1991a) found a 23- fold difference in the number of T.aspects of trap color, attractiveness from a distance, and lineola captured in canopy traps treated with UV Killer,
TABLE 2. Mean numbers of horse flies captured/ trap/day in treated canopy traps, ThistlethwaiteWildlife Management Area, 1991( means followed by the same letter are not significantlydifferent, Tukey' s mean separation procedure).
Treatment
Total UVSpecies Collected Brightener Killer Control Probability
Chlorotabanus crepuscularis 38 0.33a 0.22a 0.50a P> 0.3Chrysops spp. 317 3. 17a 2.50a 3. 14a P> 0.6Leucotabanus annulatus 28 0. 17a 0.22a 0.39a P> 0.3Tabanus americanus 62 0.67a 0.61a 044a P> 0.6Tabanus equalis 103 1. 00a 0.97a 0.89a P> 0.9Tabanus fuscicostatus 2224 22.69a 21. 86a 17.22a P> 0. 3Tabanus limbatinevris 479 4.86a 4.86a 3. 58a P> 0. 3Tabanus lineola group 2435 24.44a 27.44a 15. 75b P< 0.0016Tabanus pallidescens 22 0. 14a 0.33a 0.14a P> 0.2Tabanus proximus 1494 13. 11 ab 19.50a 8.89b P< 0.0019Tabanus turbidus 18 0. 12a 0.06a 0.28a P> 0.3Tabanus wilsoni 30 0.14a 0.19a 0.50a P> 0. 5

A
52BULL. SOC. VECTOR ECOL. JUNE, 1994
as well as a 4-fold difference in the number of T. Tabanidae: Diptera). II. The role of some visual
fuscicostatus and a 12- fold increase in the number of T. factors in the attractiveness of decoy silhouettes.
limbatinevris captured in treated traps. Why these latter Can. J. Zool. 40: 685- 695.
two species did not respond in the same manner during Browne, S. M.and G. F. Bennett. 1980. Color and shape
the present study is not known. However, the addition as mediators ofhost-seeking responses ofsimuliids
of the traps treatedwith brightener may have contributed and tabanids( Diptera) in the Tantramar marshes,
to the inconsistency between the previous study and this New Brunswick, Canada. J. Med. Entomol. 17:
one. The previous study ( Hribar et al. 1991a) was 58- 62.
conducted earlier in the year( mid-June to mid-July vs. Catts,E.P. 1970. A canopy trap for collecting Tabanidae.
early to mid-July). Three species, T. fuscicostatus, T. Mosq. News 23: 472-474.lineola group, and T.proximus, together accounted for Granger, C. A. 1970. Trap design and color as factorsover 80 percentof the flies collected in the UV experiment in trapping the salt marsh greenhead fly. J. Econ.TABLE 2), and seasonal differences in abundance may Entomol. 63: 1670- 1672.
have affected the results of this study relative to the 1991 Hansens, E. J., E. M.Bosler, and J. W. Robertson. 1971.
report. Also, differing relative numbers of T. lineola Use of traps for study and control of saltmarsh
and T. subsimilis perhaps accounts for some of the greenhead flies. J. Econ. Entomol. 64: 1481- 1486.
discrepancy between the two studies. Additionally, Hays, K. L. 1968. What attracts biting flies to their
most T. lineola group flies circle traps before entering hosts? Highlights Agr. Res. 15( 2): 6.
them, whereas many T. fuscicostatus fly straight into Hribar, L. J., D. J. Leprince, and L. D. Foil. 1991a.
traps( LDF, unpubl.) Trap interaction would therefore Increasing horse fly( Diptera: Tabandiae) catch ofbe of greater impact on the T. lineola group than on T. canopy traps by reducing ultraviolet lightfuscicostatus. Mazokhin-Porshnyakov ( 1969) has reflectance. J. Med. Entomol. 28: 874- 877.
pointed out that in nature, high UV reflectance might Hribar, L. J., D. J. Leprince, and L. D. Foil. 1991b.
signify" room to fly" to a flying insect, whereas, low UV Design for a canopy trap for collecting horse fliesreflectance may indicate" presence of an object" The Diptera: Tabanidae). J. Am. Mosq. Contr. Assoc.
differing amounts ofUV reflected from the treated traps 7: 657-659.
relative to the untreated controls may have made the Leprince,D. J., L. J. Hribar, R. T. Bessin, and L. D. Foil.
treated traps more attractive to host seeking females. 1991. Seasonal patterns ofabundance ofhorse flies
Although response to ultraviolet light is obviously an Diptera: Tabanidae) from two sites in southern
important aspect of tabanid host- seeking behavior, it Louisiana. Proc. Louisiana Acad. Sci. 54: 10- 18.
requires more thorough investigation. Mazokhin-Porshnyakov, G. A. 1969. Insect vision.
Plenum, New York, 306 pp.
Acknowledgments Neys, W. A.,R. J. Lavigne, and G. P. Roehrkasse. 1971.Attraction of Wyoming Tabanidae ( Diptera) to
We thank the staff of District 6, Louisiana decoys suspended from modified Manitoba fly
Department of Wildlife and Fisheries, and the traps. Univ. Wyoming Agr. Exp. Sta., Laramie, 11Thistlethwaite heirs for permission to work at the pp.
Thistlethwaite Wildlife Management Area. E.Chris, D. Roth, A. R. and A. W. Lindquist. 1948. Ecological
Coleman,D. Hasselschwert,D.Leprince, and A.Pecquet, notes on the deer fly at Summer Lake, Oregon. J.Louisiana State University, assisted with this study. C. Econ. Entomol. 41: 473-476.
S. Murphree, Belmont University, reviewed an earlier Snoddy, E. L. 1970. Trapping deer flies with coloredversion of the manuscript. This work was partially weather balloons( Diptera: Tabanidae). J. Georgia
supported by grant # 89- 34103- 4251 and is LAES Entomol. Soc. 5: 207- 209.
manuscript# 93- 17- 7319. SAS Institute. 1987. SAS system for elementary
statistical analysis. SAS Institute, Cary, N.C., 416
REFERENCES CITED pp.Thorsteinson, A. J. 1958. Orientation of horse flies and
Allan, S. A. and J. G. Stoffolano, Jr. 1986. The effects deer flies( Tabanidae: Diptera). I.The attractiveness
of hue and intensity on visual attraction of adult of heat to tabanids. Entomol. Exp. Appl. 1: 191-
Tabanus nigrovittatus ( Diptera: Tabanidae). J. 196.
Med. Entomol. 23: 83- 91. Thorsteinson, A. J., G. K. Bracken, and W. Hanec.
Bracken, G. K., W. Hanec, and A. J. Thorsteinson. 1964. The Manitoba horse fly trap. Can. Entomol.
1962. The orientation of horse flies and deer flies 96: 166.

BULL. SOC. VECTOR ECOL, 19( 1): 53-68 JUNE, 1994
A REDESCRIPTION OF THE MORPHOLOGY OF MOSQUITO
DIPTERA: CULICIDAE) OVARIOLES DURING VITELLOGENESIS
M. I. Sokoloval
ABSTRACT: A new model of mosquito oogenesis is suggested based on original morphological and functional
observations. Oogenesis in mosquitoes may proceed either in normal or abortive modes. Normal oogenesis consistsof previtellogenesis that is followed by pseudovitellogenesis and vitellogenesis without a previtellogenic restingstage within the ovariole. If the female has not ingested a blood meal, previtellogenesis with the subsequent
pseudovitellogenesis is initiated for two to ten days, associated with the accumulation of yolk components in theoocyte and trophocytes until the follicles degenerate. The next follicle then begins previtellogenesis andpseudovitellogenesis. Vitellogenesis interrupts previtellogenesis or pseudovitellogenesis after blood feeding,starting the mechanisms of egg maturation. Ovarioles within the ovaries of any mosquito species at any one timeof its life cycle can be segregated into two types: normal and abnormal ones, depending on their structural conditions.Normal ovarioles are characterized by progressions of vitellogenesis, whereas in abnormal ovarioles the follicle ismissing( mace- like ovarioles) or diminished in size; these latter ones could represent degenerative processes. Inaddition, non-functional germaria were found, involving a failure in cystocyte production and resorption of theiroogonial cells. All these abnormalities explain the sterility of some ovarioles, particularly in laboratory coloniesthat is recognized by reduced fecundities. Within each ovariole, normal oogenesis can be discontinued and thedegenerative processes initiated; this so-called abortive oogenesis also produces folliclular dilatations. In contrast,
after normal oogenesis is completed and ovulation occurs, the follicular epithelium becomes gradually resorbed andthe so-called egg-sac is formed. Each egg- sac will merge with the previous one and form zones of granulationfollowing a subsequent gonotrophic cycle. The reproductive age of a given female should only be deduced fromnormal ovarioles by summation ofall dilatations plus the zones ofgranulation in the egg- sac. The number ofovulatedeggs in each cycle( real fecundity) can also be defined as the sum of the zones of granulation in all ovarioles( normaland abnormal). Contrary to earlier reports, an egg never leaves the ovariole through the pedicel; this structure formsa cellular connection only between the first follicle and the calyx. After the first oviposition, the connection betweenthe remnants of the egg- sac and the second or subsequent follicles is maintained by the next connecting stalk. Theintraovarian oil injection method, together with the classification scheme of the ovariole suggested here, makes it
possible to complete a morphological description and analysis of the reproductive history of a female mosquito,including its reproductive age and potential and real fecundity.
INTRODUCTION morphological examinations of mosquito ovaries,
particularly by Christophers ( 1911) and NicholsonMosquito oogenesis is a cyclical and synchronous ( 1921). Mer ( 1936) and Macan ( 1950) have further
process, usually initiated by a blood meal in anauto- refined certain steps, and since the earlier edition of
genous mosquitoes or by internal factors in autogenous Clements ( 1963), the so-called Christophers' stagesmosquitoes during the first cycle, and is terminated by have been widely used by culicidologists. Another
the oviposition of a batch of eggs. An examination of aspect has been added to these descriptive stages, namely,the processes of oogenesis, its nutritive aspects, its the determination of the physiological age of a given
efficiency, and particularly its mechanism of endocrine female mosquito( Detinova 1962). Lehane( 1985) and
regulation, have been ofprimary interest to many groups Fox and Brust ( 1994) summarized these ovariolar
during the last two decades, and a rather compre- techniques used for many blood-sucking insects.hensive picture has emerged( Clements 1992). Many The mechanism of ovulation in insects was
physiological studies have been based on the early originally described by Brandt( 1876), but Ganin( 1866)
t Russian Medical Academy ofPostgraduate Education, Department ofTropical Diseases, Barrikadnaya 2, Moscow123836, RUSSIA. Address for correspondence: Otkritoe Shosse 28- 6- 83, Moscow 107143, RUSSIA.

54 BULL. SOC. VECTOR ECOL. JUNE, 1994
already recognized the time at which the egg entered the paraffin oil into the calices of the ovaries( Lange et al.
calyx in Psychoda which was found to be identical to 1981).
that of Culicidae( Lehane and Laurence 1978). In the The ovaries were first removed by placing thepast several years we have reinvestigated the structures abdominal tergites in adropof0.9 percent NaCl solution
and changes within the ovarioles of various mosquito stained with 0.005 percent neutral red and gently teasedspecies and under various environmental conditions with a needle. Then, using a glass micropipette with anusing light microscopy ( Sokolova 1981, 1983, 1993; external diameter of 0.07 mm, the ovarian calices were
Sokolova and Smirnov 1985; Lange and Sokolova 1989; filled with liquid paraffm ( Merck) through the lateral
Sokolova and Lange 1989, 1993a, 1993b). A more oviducts. The ovaries were then opened with two thin
refined and precise view of the morphological details needles or microscissors by longitudinal cuts along thecan now be presented that will ultimately question some ovary. Following this, the ovaries were flattened on theof the ovarian stages and also of the female reproductive slide and are observed by light microscopy as a hangingage- grading procedures. Since all our previous reports drop preparation( Fig. 1). NaCl solutions( 0.9 percent)
were published in Russian, an English review of the that cause less osmotic damage to the tissues and thus
pertinent data is presented and recent results will be allow a longer observation time are routinely used.added. In essence, the classic Christophers' stages Ito Artifacts are easily recognized by shrinking trophocytesV are demonstrated to be too rigid and static a scheme or oocytes and other morphological irregularities in the
because there appears to be no resting stage within the follicles. Neutral red to demonstrate cell death duringovariole, no matter what the conditions are of the degenerative processes in oogenesis, a well known pro-
female. As a consequence ofall possible alterations one cedure for this purpose was used( Joly 1945, Bell 1971,may encounter in any ovariole ofa female, evidence will Bowen 1981). Necrotic zones that were dyed with
be presented that argues against using the equally classic neutral red are shown in black in all accompanyingovariolar age-grading technique, and instead a method photographs.
for retrospective analysis of female reproductive historyincluding reproductive age determination will be RESULTS
suggested.
Normal Oogenesis
MATERIALS AND METHODS A detailed description of the anatomy of thepolytrophic, meroistic mosquito ovary can be found in
The mosquitoes used in this research were strains Clements ( 1992). Presented is a semidiagrammatic
of Aedes aegypti ( L.) routinely maintained in the figure of a single ovariole to incorporate these findings
Martsinovsky Institute of Medical Parasitology and ( Fig. 2). In nulliparous females, the lumen between theTropical Medicine and at the Institute ofZoology at the common and lateral oviducts appeared to be plugged
University of Zurich, Anopheles atroparvus van Tiel., with some tissue or secretion, because the oil injected
An. sacharovi Favre, and An. stephensi List. from the into the common oviduct was unable to pass into the
Martsinovsky Institute, and field populations from calyx. There is a connecting stalk between thenorthern Karelia of Ae. dorsalis Meig., Ae. punctor germarium, where oogonial cells multiply, and the
Kirby, andAe. conununis De Geer. For comparison, An. follicles. Between the follicles and calyx there is a
gambiae strain 16c55, originating from Lagos, Nigeria similar structure, the pedicel, before the first ovulation
and obtained from the London School of Tropical has taken place, but it is best to use only one term,Medicine and Hygiene, has been included. The " connecting stalk," to refer to both of them. Connectinglaboratory mosquitoes were raised and maintained at stalks usually consist of a thread-like arrangement of27°C under long-day conditions ( 14 L: 10 D), and a single cells. The ovariole is covered by an ovariolarrelative humidity of 85± 5 percent, following standard sheath that resembles a transparent conus- like coat.
procedures. This ovariolar sheath forms connections with the
The method of dissecting the ovaries of female neighboring ovarioles with multiple muscle fibers. Duemosquitoes with forceps and needles in a salt solution to contractions of these fibers, all ovariolar structures
Detinova 1962) was found to be unsatisfactory because show motility along the longitudinal axis. At the base,only a few of the 50- 150 ovarioles in each ovary remained where the ovariolar sheath connects with the calyx,
intact, the rest being disrupted by mechanical and osmotic there are two to three concentric muscle fibers on the
stress. To improve this technique, another approach wall of the calyx( Fig. 3). The so-called" rosette cells"with a more powerful resolution of the ovarian that were described earlier by Bertram( 1962) and thatarchitecture was employed, involving the injection of surround the pedicel on the calyx wall werenotobserved.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 55
In the mosquito germarium, one oogonial stem cell at the fourth larval instar ( Fig. 4A). In the emergingdivides into one oogonial cell and a cystoblast, and the adult female, the ovarioles contain follicles with seven
latter will engage in three additional mitotic divisions, trophocytes and one oocyte, in addition to undif-
giving rise to a cluster of eight cystocytes ( seven ferentiated cystocytes in the germarial zone. Between
trophocytes plus one oocyte). The process ofcystocyte the dividing oogonial cells and eight-cell cluster theredifferentiation was reported by Fiil ( 1974) to closely are small prefollicular cells that cover the cluster and
resemble the classic schemeofDrosophila as elaborated build the connecting stalk and separate the follicle fromby King( 1970). In this connection, the question may be the germarium( Lange and Sokolova 1989). Ovariolar
raised of how many oocytes can be produced in a sheaths are also differentiated.
mosquito' s lifetime. The initial number of ovarioles is The dynamics of ovariolar differentiation after
nonnallydefinedbythenumberofappropriateoogonial eclosion varies with the species. In Ae. aegypti the
cells present in the larva or embryo. In Drosophila, each follicles were already segregated from the germarium atovariole contains up to three oogonial cells undergoing the time of eclosion, whereas in Ae. dorsalis, An.
asymmetric divisions in an irregular temporal pattern atroparvus, or An. messeae, separation of the follicle
Nothigeret al. 1978). In adultmosquitoes, each oogonial was still underway. But nevertheless, each ovariole
cell can produce one cystocyte cluster and therefore now represents a morphological entity, and the generaloocytes, several times. Clements( 1992) reports 17 to scheme is similar in most of them. Each germarium
22 gonotrophic cycles for various species. In these includes two clusters of cystocytes, one above the other
experiments up to seven were observed. Therefore, the with the lower one consisting ofeight cells and the uppergermarium appears not to limit fecundity under normal with two to six cells. At the same time, resorption of
circumstances. When certain abnormalities are oogonial cells in some germaria can be observed( Fig.encountered, however, fewerovulations will be possible, 4B).
but in such cases the oogonial information can be lost or In some species, spherical or drop-like " lumenreconstructed, depending on whether ovulation took cells" are recognized within the ovariolar sheath( Fig.place. 5). These cells are similar to those also known from
Within the ovarioles, two to eight large dividing Drosophila ( King 1970) and contain granules that arecells that produce eight-cell clusters are presentbeginning sensitive to neutral red. They are also present in the
torlpfV"
4
8 411^t O
Figure 1. General view of an ovarian hanging drop preparation after oil injection, dyed with neutral red. Aedesaegypti QUWI. 1- normal ovarioles with an egg- sac, 2-mace-like ovarioles with dilatations, 3- trachea.10x10x.

56 BULL. SOC. VECTOR ECOL. JUNE, 1994
0
laills - 1
0.440filVt.4,)
f2
4 / 001I/l *
it Irria140j 3
4:, LW 4
6
3- 9111:! • Y.vw.i- d
8 ta4
7- 4.
a
C) II I , r
2
Figure 2. Semidiagrammatic figure of a single mosquito ovariole. 1- germarium, 2- connecting stalk, 3- follicle, 4-tmphocyte, 5- oocyte, 6- follicular epithelium, 7- ovariolar sheath with seldom cells, 8- lumen cells, 9-dilatation, 10-calyx, 11- egg- sac.
pupal stage. A more detailed description of these Eclosion is also marked by the deposition of somemosquito lumen cells is provided by Sokolova and yolk material into the oocytes. This process has beenLange( 1993b). referred to as" previtellogenesis." During vitellogenesis,

JUNE, 1994 BULL. SOC. VECTOR EC • L. 57
ys,
S .
Figure 3. The calyx with concentric muscle fibers. Anopheles gambiae LOGO. 1- calyx, 2- egg-sack, 3- ovariolarsheath, 4-connecting stalk, 5- follicle. 10x40x.
fie':
4, " a
mac.,
a» # i
8 , i y
G
r
e
S"..;
Figure 4. Ovarioles in an ovary of Anopheles albimanus laryae of IV instar before pupation( A), and Aedes aegyptiUGAL female just after eclosion( B). 1- ovarioles, 2- re oogonial cells. 1Ox40x.
the dynamic process of blood- induced yolk deposition, described in other animals as well( Wyllie 1981). The
the trophocytes grow quickly but later fall behind and beginning of the accumulation of yolk in the ovariolesare completely resorbed by its end. This process maybe that are closest to the paired oviducts were observed.
referred as apoptosis or programmed cell death, and is Their maturation appears to proceed faster here than in

58 BULL. SOC. VECTOR ECOL. JUNE, 1994
aa
tst ., , ,
4i'
s
1f A
tl.'*14,.. A- Ps i N%Let
An ' * "
tlFigure 5. Lumen cells( 1) in ovarioles of Anopheles gambiae LOGO( A, 10x25x) and Culex pipiens ZOLL ( B,
10x100x). 2- normal ovariole with a dilatation and a pedicel, 3- mace- like ovariole with an egg- sac.
the apical ovarioles, showing a difference of about one When the oocyte has reached its maximal species-step in Christopher' s scale. A similar result was described specific length at the termination of secondaryby Laurence( 1964) in Ae. togoi. Accordingly, the basal vitellogenesis, the follicular epithelium secretes theoocytes reach ovulation slightly earlier than the apical chorion. When the follicular epithelium separates fromones, and the same sequence is followed during the chorion and ovulation takes place, the so-calledoviposition.
egg- sac"( Lange and Khok 1981, Lange and SokolovaThere are some controversial views on the structural 1989) is formed. Ovulation is initiated by the rupture of
changes of the pedicel and the connecting stalks during the follicular epithelium and calyx wall ( Lehane andvitellogenesis. Originally it was thought that these two Laurence 1978), accompanied byperistaltic contractionsstructures represent a channel through which oocytes of the ovariolar sheath and the calyx. Once the egg haspass ( Detinova 1962, Samarawicrema 1962). Conse- been oviposited, the follicular epithelium atrophies byquently, the pedicel would widen to accommodate the autolyticprocesses( Sokolova1983, LangeandSokolovaegg, and after ovulation a dilatation would form in its 1989). The remnants of the follicular epithelium coverplace. This led to the well-known procedure ofcounting the ruptured calyx wall. Atpresent it is not clear if theredilatations as a means of determining the number of are any regeneration processes in these calyx tissues.gonotrophic cycles. Lehane and Laurence ( 1978) Although the next connecting stalk and egg- sac form anreported that the pedicel was a solid structure without a attachment between the ovariole and calyx (Fig. 6), alumen. Histological investigations confirmed that the considerable interspecific variability in the dynamics ofconnecting stalks consist of cells( Lange and Sokolova resorption of the follicular epithelium was found1989). Moreover, the pedicels originate from ( Sokolova 1983). This apoptosis of the follicularmesodermal cells in larval ovaries and each stalk is epithelium and the corresponding reduction of itsformed by prefollicular cells in the germarium during derivative, the egg- sac, can be characterized by thefollicular segregation. Based on our observations following morphological events that have been groupedSokolova 1981, 1983; Sokolova and Smirnov 1985; into four steps ( Sokolova 1983). Immediately after
Lange and Sokolova 1989; Sokolova and Lange 1989), oviposition in all Aedes and the three Anopheles speciesthe pedicels were never rebuilt after ovulation. tested, the long egg- sacs already show large particles ofSupposedly, their only functional role is the attachment bright red color with neutral red, indicating theirof the follicle to the calyx. degenerative nature along their wall upon the tunica

JUNE, 1994 BULL. SOC. VECTOR ECOL. 59
propria ( or basement membrane that covers the observed in 97 percent of an Ae. dorsalis population, inepithelium)( Fig. 6A). During the next two steps, the 93 percent of Ae. punctor, and in 90 percent of An.size of these granules and their staining intensity are messeae, but there were always fewer than five ovariolesreduced in all mosquitoes tested together with a further per female involved( Sokolova and Lange 1989). Thesize reduction of the egg- sac, down to a short, contracted so- called pregonotrophic and agonotrophic
umbrella- like structure, as seen in Fig. 6B. In An. degenerations can also take place during the extendedgambiae the egg- sacs were smaller and contained fewer periods after eclosion in the absence of blood mealsgranules, which in this species was associated with the ( Sokolova 1981, Sokolova and Lange 1989). However,
connecting stalk ( Fig. 6C). In Cx. pipiens, these if females were withheld from water for one week orgranules were found on the apical part of the egg- sac. were given access to sucrose solutions for up to a month,When the reduction of the egg- sac is completed, an there was no significant difference in the number ofirregular circle on the calyx wall remains in Aedes, degenerating follicles at any given moment. This mightcontaining very small granules whose color has changed indicate that degeneration was not related to the age ofto greenish- red( Fig. 6D,E,F), but in other species these the female. Whenever during the 1. 5 months the femalesgranules may be nearly absent. Altogether, the duration were dissected, each had one to three degenerativeof these processes is species- specific and temperature- follicles in one to three ovarioles. At the end of onedependent. In Aedes it may take up to one week under month on a sugar diet, in 90-95 percent of the ovarioles
laboratory conditions, whereas under field conditions in the first proximate follicles had degenerated, formingNorthern Karelian territories from two weeks to one dilatations that were replaced by the next follicles. Inmonth were observed( Sokolova 1983). As a rule, in An. 5 to 15 percent of the others, these next follicles havegambiae and Cx. pipiens this reduction in the egg- sac also degenerated and replaced by the third follicles. Inoccurred the fastest of all the species examined, such ovarioles one to three dilatations, a follicle, and a
The same sequence of events occurs with each germarium were found.subsequent ovulation. Ultimately, every new egg- sac In experiments with females maintained on sugar,that is formed will merge with the previous one, thus there was a gradual accumulation of what appeared to be
forming different zones of granulation( Fig. 6G,H). A lipid droplets inside the ooplasm and trophocyte plasmapermanent autolysis that takes place in these fused egg- ( Fig. 7), termed pseudovitellogenesis, as well as an
sacs does not allow the recognition of more than three atrophy of the follicular epithelium, including thezones. The significance of this event will be apparent in thinning ofcell walls and cell swelling that preceded theconnection with the determination of female reproductive degeneration. The so-called " previtellogenic restinghistory in a following section. stage" within an ovariole, that is postulated as having no
As previously mentioned, several investigators had " detriment to the follicles"( Clements 1992, Clementsdescribed ` ovariolar dilatations" that represented and Boocock 1984), was neverobserved. Once separatedremnants of trophocytes and the follicular epithelium, from a germarium, a follicle can either degenerate if theand which were interpreted as indicating the event of female lives long enough, or may mature after a bloodone ovulation. In an analogy to mammals, these were meal or autogenously. There is no follicular restingincorrectly referred to as the corpus luteum( Detinova stage between previtellogenesis and vitellogenesis.1962). According to Lehane and Laurence( 1978), the Previtellogenesis is followed by pseudovitellogenesisdilatations are formed from the ovariolar sheath that that continues for two to ten days until one folliclesurrounds the follicle. Our observations ( Sokolova degenerates and next initiates previtellogenesis in the1981, 1983; Sokolova and Smirnov 1985; Sokolova and ovariole, and soon until the female dies or ingests blood.Lange 1989) and those ofLange and Khok( 1981) differ Moreover, starvation for 3. 5 days before death does notfrom these interpretations. The remains ofan ovulation increase the number of degenerative follicles in Ae.in an ovariole represents only the egg- sac, and since aegypti, remaining as one to three degenerative folliclesthese fuse after subsequent ovulations, they cannot be within the ovary. Thus, the reserves in the follicle arecounted reliably after three ovipositions. Dilatations do not utilized for the survival of the whole organism.indicate the degeneration of a whole follicle, as will be Follicle degeneration in females that have not ingestedshown in connection with abortive oogenesis. blood does not appear to be related to the senescence of
the mosquito or starvation, as postulated forother insectsAbortive Oogenesis Bell and Bohm 1975), but instead involves a process of
There are several ways in which the mosquito oocyte elimination rather than oosorption for nutrition.ovarioles may degenerate. Even during the first few Follicular degeneration is characterized by thehours after eclosion, follicular degeneration has been appearance of a few large granules, or lysosomes, in the
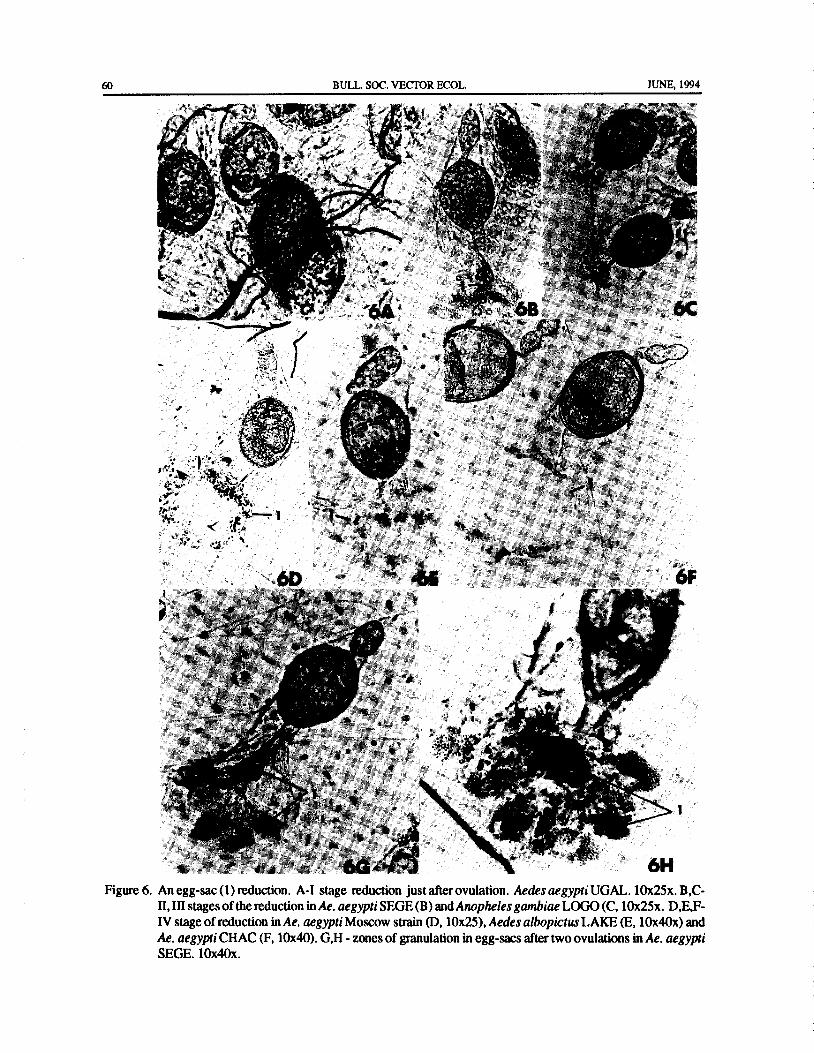
60 BULL. SOC. VECTOR ECOL. JUNE, 1994
ems. ...ty
4 I'L i,4,
4•.. ' ^. r"`
tit 'l+`
i1'
4.
r '.'
t. ';, ; y4 +. r.
Ian t .•
t l F Fak
a
r. r •
e'"' f+ ' 4* y
t. y it t' f0z' i
tl ! 1 - 0V.. ,
k, "' Ay. ". ' i 1.. i'
tit, ' a ±i 0 l‘ P. ' ,
V: , ' ,...:::'.4. •
Vii'• 6M
r
yt}•t.. "t7i
V te.': 1
if /y
J,
tt+
A'
Act
o
ice ' 44,40,-..:: tr' 4.E 1 .iro
1. - s
5, fir. gyp,; ""
aty •
a f
1,
a 1 f.Aor`
7, S
r
I tc ei ,. 4
14 I y
f'' 4 . 4''
7 6H
Figure 6. An egg- sac( 1) reduction. A-I stage reduction just after ovulation. Aedes aegypti UGAL. 10x25x. B, C-II, III stages of the reduction in Ae. aegypti SEGE( B) and Anopheles gambiae LOGO( C, 10x25x. D,E,F-
IV stage of reduction in Ae. aegypti Moscow strain( D, 10x25), Aedes albopictus LAKE( E, 10x40x) and
Ae. aegypti CHAC( F, 10x40). G,H- zones of granulation in egg- sacs after two ovulations in Ae. aegyptiSEGE. 10x40x.

JUNE, 1994 BULL, SOC. VECTOR ECOL. 61
tom, Itort
A
v. Cd#
ti
rF
ti
P"el, ..
Figure 7. Ovarioles in Aedes aegypti ELEM female after one month on a sugar diet. A-10x10x. B- 10x40x.
tss
ts
4
4
Figure 8. Degenerated follicles. A-Aedes aegypti UGAL. 10x25x. B- Aedes albopictus LAKE. 10x40x. 1-
degenerated follicle, 2-normal follicle, 3- dilatation.
follicular epithelium, that originate from spherical The follicle is later filled by very bright red granulesdroplets. They acquire a bright red color, and their ( Fig. 8A). The final step ofdegeneration is the dilatationnumber and size steadily increase as the size of the ( Fig. 8B), accompanied by a gradual disappearance offollicles decreases. The cells of the follicular epithelium the granules. Ofrly the folded tunics propria with a fewfirst increase in size and grow longer, penetrating the fine granules remains in the dilatation.
follicularcore. Trophocytes and oocyte are fragmented. Follicle degeneration within a single ovary can be

62 BULL. SOC. VECTOR ECOL. JUNE, 1994
observed at any time during oogenesis. This explains proportion. In addition, the germanium can also be in a
the presence of dilatations of different form and size. non- functional state, characterized by the absence ofThis is also consistent with the often large intra- and the large oogonial cells, sometimes containing onlyinterspecific variability in female mosquitoes where flattened mesodermal cells( Fig. 12). This seems to be indegenerating follicles can comprise up to 60 percent per an irreversible state, often foundat eclosion, and therefore
ovary. In laboratory populations, it is not uncommon permanent ovariolar sterility is assumed. One orboth offor between 30 and 95 percent of the ovarioles to mature the ovaries per female can be affected in this way( Fig.after a blood meal. In field populations after one gono- 9C), and is more frequent in laboratory colonies than introphic cycle the mean percent of ovarioles with one field collections( Sokolovaunpubl.data). This ovariolar
dilatation in Ae. dorsalis was observed to be 9.510.8 sterility may be due to the absence of oogonial cells inpercent, in Ae. punctor, 26. 110. 1 percent, and in An. the germarium, abnormalities in the speed at which the
messeae, 12. 112.4 percent. After two gonotrophic follicles separate from the germarium, which are always
cycles the percent ofovarioles with two dilatations was followed by their degeneration, follicles. thatmaycontain14.911. 6 percent in Ae. punctor, and 3. 610.7 percent in up to 32 undifferentiated cells, or follcles with only oneAn. messeae( Sokolova 1983). or two trophocytes beside the normal oocyte. The
dilatations that result may be aberrant( Sokolova andAbnormal Ovarioles Lange 1989), and may outnumber the reproductive
In addition to the follicular degeneration that can cycles by up to three times. Since the changes from atake place at any stage during their maturation period, functional mode of the germarium to any of theseother malformations can frequently be observed abnormal conditions seem to occur independently fromSokolova and Lange 1993). Thereare ovarioleswithout the gonotrophic rhythm of a given female and in any
any follicles that are called" mace- like" ovarioles as a period of its life, the discrepancy between the number ofresult of their peculiar shape( Fig. 9). The follicles that dilatations and gonotrophic cycles becomes very largeattain only halfoftheir normal size or less and that do not in these abnormal ovarioles. The frequencies of these
accumulate neutral red are referred to as " diminutive abnormalities are seldom higher than 20 percent of the
the dwarfsovarioles" ( Fig. 10), and are Detinova ovarioles of a female( except for comprehensive ovary1962). Finally, follicles may contain an atypical number sterility), but it can be found in up to 90 percent of theof cells( Fig. 11), deviating from the usual seven to one natural population. Vitellogenesis in most ovarioles
144 X4.
1. V-
4,,,;'.', N1/4.V,'744k. , .- to* Z„'''',,*44?; 41,,,,..'46.1, ."''',„ dsii .,t1- „,..,,,,-,- , 1
0 Vit. ,
t?
4t ,' ^ R 5,.
s4 '
t ..
y 9r
tY
10;,..-t,s : ' t '' '''':;,-NVI,P11',' '' o*i; '4' , :
C.;,.' '-; 5,.-yu
Figure 9. Mace-like ovarioles. A-Aedes aegypti SEGE. B- Culex pipiens ZOLL. C-Ae. aegypti BWAM sterile
ovary. 10x40x.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 63
P °.;. * IIIY
a
10 1
Figure 10. Diminutive ovariole in Anopheles gambiae LOGO. 1 normal follicle, 2- diminished follicle. 10x25x.
yt°°
a
000,,„,,,
itio.
3
0001
At4,.3a7a° )
tiatiills
k.' ff
A-'''
Ni,.7.1tr
4., *:*. 1,- -.
4v. ,‘,.
tt4,.. ,, . 1
b .__, Is . _
7., te_,,,,,;.. ,
4ki:?t, ' * '
t
Its '
At iiiiii44t1110,
itig,
i0,,i,i, Nii6 .
8v,
Figure 11. Arioles with atypical number of cells in the follicles. A-Aedes aegypti QUWI. 1Ox100x. B-Ae. aegyptiUGAL. 1Ox40x. 1- 32 undifferentiated cells, 2- 12 additive undifferentiated cells in a trophocyte area of
the follicle.

64 BULL. SOC. VECTOR ECOL. JUNE, 1994
m, ,
s a5", d s
Y
s
r 4/`
4.. i, ,,,,, , ,t,,..,„4,.,..s, i ,",,,,,, 4,,,,D. a*,
4 a»
0../-' ''
1''''''''' , 4,"nt'l(„';..:::'''''.i.'4"4::::''''‘,4:
4;1'";:,
f,..
4"Fs,Hlb\'',:
k3'4:, , ,
14i0 , - , ,'
i'',•', 5gn
i
mss»Figure 12. Functional( 1) and nonfunctional( 2) germarium in Anopheles gambiae LOGO( A, 10x25x) and Aedes
aegypti QUWI( B, 10x40x).
Germarium 9 1
91) Functional Non-functional
Follicle 1Normal Degenerate Diminutive Absent
A B C
tD E
it
FOvariolar
Stalk
Figure 13. Classificatory scheme of the morphological description of the ovariolar system of blood-suckingmosquitoes. A-pedicel, B- egg sac with two zones of granulations, C-dilatation and pedicel, D-dilatationand zone ofgranulation, E-zone ofgranulation and vestigial dilatation, F-combination of dilatations andzone of granulation.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 65
proceeds normally after a blood meal was' ingested, ditionally can be defined as normal, in which folliclesirrespective of the degeneration in neighborng follicles are separated, matured, and degenerated( gonotrophic
where yolk deposition may not occur. Thus, each dilatations) synchronously with the gonotrophic cycles,ovariole shows a certain degree ofautonomy. Each may and abnormal ones, in which follicles may be absent, ormature or degenerate according to its own fate. In An. separated and degenerated ( aberrant dilatations)
messeae, abnormal ovarioles( mace- like and diminutive asynchronously Sokolovaand Smirnov 1985; Sokolovatypes) are localized in the area where the large trachea 1993, 1994, in press). In the latter cases, follicles never
enter the ovaries( Sokolova and Lange 1993a). Because reach their definitive typical state. Normal and abortive
tracheoles are scarce at this site, it is assumed that a local oogenesis can proceed with different consequences in
deficiency in oxygen supply might be responsible. each single ovariole. Within an ovariole, previouslyWhatever the reasons for all these abnormalities formed dilatations can be observed free under the
and the extent or the temporal pattern of their appearance, ovariolar sheath ( the vestigial dilatations reported bythere was evidence. for an increase in their frequencies Sokolova 1983) in the area of the egg- sac afterovulationin laboratory colones when compared to field-collected of the next successfully matured egg ( Fig. 14). Anymaterial. The mean proportion of abnormal sterile ovariole can transform its mode of oogenesis from
ovarioles per female in natural An. messeae increased normal to abortive and to complete sterility during thefrom three percent in the first gonotrophic cycle to 11 life of the female. The mechanisms of such
percent in the third one, in Ae. punctor from four percent transformation in most cases are unknown.
in the first cycle to ten percent in the second one, while
in laboratory Ae. aegypti this proportion may vary from Analysis of Female Reproductive History15 percent in the LSTM strain to 40 percent in the OHIO Until recently, the reproductive or gonotrophic agestrain after one cycle( Sokolova unpubl. data). of female mosquitoes was determined by counting the
Thus at each given moment all ovarioles con- maximalnumberofdilatationsperovarioleinagroupof
a
4- 1, 40,
t'4
444 IV11„..:: ' h '
1 ktrc ,'' w
r
t,
t4 r a.*.„,,,
71, ,,
L *,,,,, 4 i.,4,";4. :
lc '', ',: 4,1t, le.
rt.„--
4 '',- 1,,,
w-,,,--
4....,,,:
s.:44t;I:-.' w ' ,' a t
4 , ' 7./
or. i. °,4* :.
41* . 140" *
t.,,*,,,,,f 4 4W-*.,„:,''" ''''.e If f', It
4
Ar
iti,,,,..... v,' 1,, t,.. ./-', -,
o4 ,1,tie /... tki
07., '
4.4. .-." *,..-,
i 'it'-trk *1 ..
ffi
Figure 14. Ovarioles with vestigial dilatations in egg- sacks ofAedesaegyphelesmesseaeti UGAL( A) and Anop B,
from Sokolova, 1983). 1Ox40x. 1- vestigial dilatation, 2-egg- sack in mace- like ovariole with nonfuctionalgermarium.

66 BULL. SOC. VECTOR ECOL. JUNE, 1994
females out ofa vector population( Mondet 1993). This observe nearly all the ovarioles in their intact state andpractice was initiated by the work ofPolovodova( 1949), analyze each one to determine exactly how many eggswhich was latermodified and expandedby Beklemishev were ovulated during at least three cycles( Fig. 16). Theet al. ( 1959) and Detinova ( 1962). Based on recent sum ofall the zones ofgranulation in the egg- sacs ofallresults, however, these techniques may introduce a high normal and abnormal ovarioles in both ovaries will
probability of erroneous estimates because the show the real fecundity. The detailed analysis ofindiscriminate counts of all dilatations also include ovarioles allows the correspondence of each zone of
those from ovariolar abnormalities, while egg- sacs that granulation to the exact reproductive cycle. In all
contain granules but that show no dilatations are species investigated, except for Cx. pipiens and An.excluded. Therefore it is suggested that the following gambiae, this retrospective analysis of femaletwo rules apply in order to accurately determine the reproductive history has been confirmed experimentallyreproductive age( Sokolova and Smimov1985, Sokolova ( Sokolova, unpubl. data). This method has yielded1994, in press). First,only normal ovarioles with typical valuable results in the study of natural populations offollicles should be selected for analysis, and second, Aedes andAnopheles in Russian Europe, demonstratingevery dilatation, including vestigial ones and each zone that most females had undergone from one to threeof granulation in egg- sacs, correspond to one gonotrophic cycles( Sokolova, in prep.). An analysis ofreproductive cycle in a normal ovariole( Fig. 15). Thus the female reproductive history in more detail, with thetheir sum per ovariole, which should be identical among description of some inevitable errors will be providedall ovarioles considered, isan estimate of the reproductive ( Sokolova 1994, in press).age. Among thousands of ovarioles examined fromnatural populations, only a few failed to conform to Acknowledgments
these rules, perhaps due to previous pregonotrophic
degeneration. The comparison of all normal ovarioles I thankDrs. A. B. Lange, N. A. Tamarina, and N. A.in an ovary should serve as a confirmation of age Smirnov for their assistance in early works in Moscowdetermination. State University ( Russia). Special thanks to Fr. I.
With the oil injection method, it is possible to Fluckiger for rearing mosquitoes and Fr. R. Haigis for
446k.
14fix`
is#
W°
i. & '^
rP
a
p,,,, '^
yra i.
W
ray
Figure 15. Ovarioles after three ovipositions. Aedes aegypti UGAL. 1- egg- sac, 2- dilatation, 3- connecting stalk.10x40x.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 67
First
Ovary
112 1 + 1 + 5 + 1 + 5 + 1 + 6 + 3
Second
Ovary
124 1 + 2 + 1 1 9
Figure 16. A description of the ovariolar system in Anopheles messeae after two ovipositions. The numbers under
the symbols refer to the count of actual ovarioles of each type in the ovary. The interpretation of theovariolar structures is from Figure 13. The real fecundity in the first cycle is 129 eggs in the first ovaryplus 138 eggs in the second ovary( total of 267 eggs). In the second cycle it is 112 and 125 eggs( total
of 237) in the first and second ovary, respectively.
her assistance in preparing the photographs. I am also Bell, W. J. and M. K. Bohm. 1975. Oosorption in
grateful to Drs. R. Graf and R. Stidwill, Fr. S. insects. Biol. Rev. 50: 373- 396.
Timmerman, and Hr. E. Horler for their everyday help Bertram, D. S. 1962. The ovary and ovarioles ofin carrying out the investigation in the Zoolog ical Institute mosquitoes. WHO Monogr. Ser. no. 47: 195- 214.
of the University of Zurich. The support of this work by Bowen, I. D. 1981. Techniques for demonstrating cellthe Swiss National Science Foundation is also death. Pp. 379-4442 in Cell Death in Biology andacknowledged. The author thanks Dr.M. J. Klowden for Pathology( I. D. Bowen and R. A. Lockshin, eds.).his comments and the editing of this paper. My best Chapman and Hall, London, 493 pp.thanks to Dr. H. Briegel for the permission to use his Brandt, A.F. 1876. The comparative investigations on
laboratory facilities, hishelp in preparing this manuscript, insect egg tubes and egg. Trans. Soc. Nature,and initiating the experiments with mosquito starvation. Anthropology and Ethnography Amateurs 23: 1-I am also grateful for the comments of two anonymous 150( in Russian).
reviewers. Christophers, S. R. 1911. The development of the eggfollicle in anophelines. Paludism 2: 73- 89.
REFERENCES CITED Clements, A. N. 1963. The physiology of mosquitoes.Oxford, 393 pp.
Beklemishev, V. N., T. S. Detinova, and V. P. Clements, A.N. 1992. The biology ofmosquitoes. Vol.Polovodova. 1959. Detennination ofphysiological 1. Development, nutrition and reproduction.age in Anophelines and of age distribution in Chapman and Hall, London, 509 pp.Aophelinae populations in the USSR. Bull. Clements, A. N. and M. R. Boocock. 1984. OvarianW.H.O. 21: 223-228. development in mosquitoes: stages of growth and
Bell, W. J. 1971. Starvation- induced oocyte resorption arrest, and follicular resorption. Physiol. Entomol.
and yolk protein salvage in Periplaneta americana. 9: 1- 8.
J. Insect Physiol. 17: 1099- 1111. Detinova, T. S. 1962. Age-grading methods in Diptera

68 BULL. SOC. VECTOR ECOL. JUNE, 1994
of medical importance with special reference to maculipennis, Meig. Quart. J. Microscop. Sci. 65:some vectors of malaria. WHO Monogr. Ser., No. 395- 448.
47, 216 pp. Nothiger, R., T.Schiipbach, J. Szabad, andE.Wieschaus.
Fiil, A. 1974. Structural and functional modifications 1978. Stem cells and tissue homeostasis in insect
of the nucleus during oogenesis in the mosquito development. Pp. 71- 85 in Stem Cells and TissueAedes aegypti. J. Cell Sci. 14: 51- 67 Homeostasis. The SecondSymposiumoftheBritish
Fox, A. S. and R. A. Brust. 1994. How do dilatations Society for Cell Biology( B. I. Lord, ed). 368 pp.form in mosquito ovarioles. Parasitol. Today 10: 19- Polovodova, V. P. 1949. Physiological age grading in23. Anopheles females, i.e. number of gonotrophic
Ganin, M. S. 1866. The developmental history of a fly cycles. Med. Parasitol. 18: 352-355.
egg ( Nematocera). Trans. Emperor Acad. Sci. Samarawicrema, W. A. 1962. Follicular relics in wild
suppl. 9, N5, 64 pp.( In Russian). caught Culex pipiens fatigans Wiedemann in
Joly, P. 1945. La fonction ovarienne et son controle Ceylon. Bull. W.H.O. 27: 637-640.
humoral chez les Dytiscides. Arch. Zool. Exp. Sokolova,M. I. 1981. Age alterationsandmorphological
Gen. 84: 49- 164. types of ovarioles in northern population of blood-
King, R. C. 1970. Ovarian development in Drosophila sucking mosquito females Aedes caspius dorsalismelanogaster. Acad. Press, N.Y., 227 pp. Mg. Med. Parasitol. 6: 63- 70.
Lange, A. B. and C. K. Khok. 1981. Abortive oogenesis Sokolova, M. I. 1983. Diagnostic significance of
and physiological age in blood-sucking mosquitoes ovariolar structures for age- grading in blood-Diptera: Culicidae). Med. Parasitol. 50:48- 56 sucking mosquito females ( Culicidae: Diptera).
Lange, A. B., C. K. Khok, and M. I. Sokolova. 1981. Med. Parasitol. 2: 53- 58.
Method of intraovarian oil injection and its using Sokolova, M. I. 1993. Morphofunctional changes offor the determination ofphysiological age inblood- ovarioles of blood-sucking mosquitoes ( Diptera:sucking mosquito females ( Diptera: Culicidae). Culicidae) during oogenesis. 5. Alternative betweenMed. Parasitol. 4: 51- 53. normal and abortive oogenesis in an ovariole. Med.
Lange, A. B. and M. I. Sokolova. 1989. Parasitol. 5: 41- 45.
Morphofunctional changes of ovarioles of blood- Sokolova, M. I. 1994. Reproductive history of blood-sucking mosquitoes ( Diptera: Culicidae) during sucking mosquito females ( Diptera: Culicidae).oogenesis. 1. Normal oogenesis. Med. Parasitol. 1: Med. Parasitol. 2: ( in press).
22-26. Sokolova, M. I. and A. B. Lange. 1989. Morpho-
Laurence, B. R. 1964. Autogeny in Aedes togoi Theob. functional changes of ovarioles of blood-suckingDiptera: Culicidae). J. Insect Physiol. 10: 319- 331. mosquitoes( Diptera: Culicidae) during oogenesis.
Lehane, M. J. 1985. Determining the age of an insect. 2. Abortive oogenesis. Med. Parasitol. 3: 55- 60.Parasitol. Today 1: 81- 85. Sokolova, M. I. and A. B. Lange. 1993a. Morpho-
Lehane, M. J. and B. R. Laurence. 1978. Development functional changes of ovarioles of blood-suckingof the calyx and lateral oviduct during oogenesis in mosquitoes( Diptera: Culicidae) during oogenesis.Aedes aegypti. Cell Tiss. Res. 193: 125- 137. 3. Abnormal ovarioles. Med. Parasitol. 2: 31- 38.
Macan, T. T. 1950. The Anopheline mosquitoes of Iraq Sokolova, M. I. and A. B. Lange. 1993b. Morpho-
and North Persia. Mem. Lond. Sch. Hyg. Trop. functional changes of ovarioles of blood-suckingMed. 7: 109-223. mosquitoes( Diptera: Culicidae) during oogenesis.
Mer,G.G. 1936. Experimental study on the development 4. Lumen cells in ovarioles. Med. Parasitol. 3: 30-of the ovary in A. elutus Edw.( Diptera: Culicidae). 33.
Bull. Entomol. Res. 27: 351- 359. Sokolova, M. I. and N. A. Smirnov. 1985. Analysis ofMondet, B. 1993. Application de la methode de reproductive cycles of blood-sucking mosquitoes
Polovodova a la determination de l' age Diptera: Culicidae). Biol. Sciences. 7: 32-37.
physiologique des Aedes ( Diptera: Culicidae) Wyllie, A. H. 1981. Cell death: a new classificationvecteurs de fievre jaune. Ann. Soc. Entomol. Fr. separating apoptosis from necrosis. Pp. 9-34 inN.S.) 29: 61- 76. Cell Death in Biology and Pathology( I.D. Bowen
Nicholson, A. J. 1921. The development of the ovary and R. A. Lockshin, eds.). Chapman & Hall,and ovarian egg of a mosquito, Anopheles London, 493 pp.

BULL. SOC. VECTOR ECOL., 19( 1): 69-72 JUNE, 1994
THE DISMAL STATE OF MOSQUITO SYSTEMATICS:
PERSPECTIVES OF A CLASSICAL TAXONOMIST'
T. J. Zavortink2
In biology we use the phrase" living fossil" to refer evolutionary history and distribution of the Culicidae.to an extant species with primitive characteristics that Many type specimens have not been examined inresembles species that lived long ago. It is the kind of decades, so we still face the prospect of name changes,organism you would expect to find preserved in rocks sometimes even for common, widespread specieshundreds of millions of years old that, somehow, has ( Zavortink, 1993).managed to survive. In some respects, I am a" living The first of the procedures of systematists forfossil," one of the last surviving mosquito systematists. discovering species and classifying them is to assembleThis is a sad situation for the science ofculicidology, for all available specimens in a group. The next is to sort thethere is still much work to be done in mosquito specimens to geographical source and habitat. This issystematics. Both the large number of undescribed followed by identifying the specimens as well as can bemosquito species and the absence of a sound classi- done with existing literature. Then all specimensfication and phylogeny for the family make the work of provisionally identified as the same species are examinedthose using nonmorphological taxonomic characters thoroughly, and any previously unrecognized speciesmore difficult. Statements about affinities and are sorted out. The reproductive isolation of species isillustrations ofphylogenetic trees based on comparative inferred from the distribution of morphologicalmorphology are hypotheses about evolutionary characteristics; what is looked for in particular arerelationships that can be tested with other types ofdata. populations characterized by unique sets of constantlyFurthermore, these hypotheses, like all other hypotheses, correlated features. The next step is to preparehave a heuristic quality: they suggest, help define, and preliminary keys to the species and then reexamineframe additional studies. As I noted before( Zavortink, every specimen for the key characters, sorting out any1979): " A classification . . . should stimulate further additional species that may be discerned at this stage,research that may ultimately bring about its modification and revising keys as necessary. Then all species areor demise." compared carefully and placed into groups at different
It is possible to get some idea of the state of levels on the basis of shared morphologicalmosquito taxonomy by examining the goals and characteristics and inferred genealogy. The last step isprocedures of systematics and then examining to prepare final keys, descriptions, discussions, andmosquitoes to see if these goals have been met or the illustrations for publication.procedures applied. One of the goals of systematics is Since most mosquitoes have not been studiedto discover and describe the various species of life that taxonomically in more than 50 or 60 years, theseinhabit the earth. Another is to arrange these species in procedures, particularly the application of the biologicala classification that reflects their evolutionary species concept and the development of a hierarchicalrelationships. A third is to interpretbiological diversity. classification of natural groups, obviously have notAnd, a fourth is to provide unique names for the species been applied to them. Much taxonomic work is yet to beand other taxa in the classification so that information completed.
about these taxa may be communicated. According to Mayr( 1969), in his book PrinciplesThese goals are far from being met for mosquitoes. ofSystematic Zoology, there are three stages or levels of
We have not yet found and described all the species, and taxonomic study. The first is alpha taxonomy, the levelthe classification of the family includes many groups of concerned with the characterization and naming ofquestionable monophyly, dubious rank, and unknown species. The second is beta taxonomy, the levelaffinities. Little effort has been made to interpret the concerned with the arranging of species into a natural
Paper presented at the 24th Annual Conference of the Society for Vector Ecology, November 16, 1992, SanFrancisco, California, USA.
2Department of Biology, University of San Francisco, 2130 Fulton Street, San Francisco, CA 94117- 1080, USA.

70 BULL. SOC. VECTOR ECOL. JUNE, 1994
system of lower and higher taxa. The third is gamma Over the years I have expressed the opinion that
taxonomy, the level dealing with various biological mosquito taxonomy is still by and large at the alphaaspects of taxa, ranging from the study of intraspecific taxonomic level (Zavortink, 1974, 1990). In 1990, I
populations to studies of speciation and ofevolutionary argued from the rate of description of species of Aedes
rates and trends. Mayr notes that these stages overlap and from information on the number of species of
and intergrade, but that the trend from alpha to gamma mosquitoes found in the South Pacific that we know at
is unmistakable in the taxonomic history ofevery group. best 25 to 50 percent of the species. And, on the basis
Belkin' s( 1962) Mosquitoes of the South Pacific is ofmathematical analysis of the number of species in the
still the best and, in fact, the only example of a modem family, I argued that the approximately 3, 200 speciestaxonomic study ofan entire mosquito fauna. This work we know would better be classified in 225 genera than
includes all three levels of taxonomic study mentioned the 37 genera we currently recognize.above. There is alpha taxonomy, represented by the I am not the only student of mosquitoes to havedescription of many new species, the resurrection of commented about the state of mosquito systematics.
many species from synonymy, and the description of Bates ( 1949), in his book The Natural History of
many previously unknown stages. There is beta Mosquitoes, devoted a chapter to the classification of
taxonomy, represented by the careful comparison of mosquitoes. As to the matter of the number of species,
mosquito species and their classification into tribes, he believed that most were already known. He stated:sections, genera, subgenera, groups, and complexes. " The vast majority of different kinds have probablyThis classification was done largely on the basis of already been collected, pinned, and named by someone,comparative morphology, but distribution and ecology and it seems likely that the total world fauna is on thealso were taken into consideration. Finally, there is order of magnitude of 2,000 species." As for the matter
gamma taxonomy, represented by Belkin' s attempts to of the classification of mosquitoes, Bates noted the
explain the evolution of the mosquito fauna of the South advantages of" splitting" mosquitoes into many genera,Pacific. The present fauna was explained on the basis of as was done by Theobald( 1910), who placed the 1, 050dispersals of different lineages into the South Pacific at species known to him in 149 genera( seven species per
different times and along different routes, allopatric and genus), and also the advantages of" lumping" mosquitoessympatric speciation on islands within the South Pacific, into few genera, as was done by Edwards( 1932), whoextinction ofancestral species in source areas, ecological placed the 1, 400 species known at the time into 30
competition, and hybridization between species. genera( 47 species per genus), but he did not give his
Whether or not one believes Belkin' s explanations for own preference for either system. As for the evolution
the evolution of the mosquito fauna of the South Pacific, ofmosquitoes, Bates stated simply: " The phylogeny of
his work is outstanding in attempting to explain the mosquitoes... can apparently be worked out very neatly
diversity of mosquitoes there. by the study of the male genitalia." Although Bates did
Belkin' s Mosquitoes of the South Pacific is not openly praise the state of mosquito systematics, itnoteworthy in not having a phylogenetic tree. One seems clear from his comments that he considered the
reason there isn' t one is thatBelkin was very conservative field to be in satisfactory shape.and thought it premature to attempt a complete Eldridge ( 1969) noted that there is " unmined"
reclassification ofmosquitoes on the basis of a regional descriptive ground in mosquitoes, that there are
fauna. Another reason was his belief that taxa of a geographic areas of the world that have little-studied
particular rank arose more or less contemporaneously mosquito faunas, and that some entire mosquito groups,
and formed a " phylogenetic bush" rather than a such as the sabethines in the New World tropics, are
phylogenetic tree. Yet another reason was Belkin' s very poorly known.belief that hybridization probably took place between Darsie ( 1974) noted that the study of mosquitomembers of different mosquito lineages at the time of taxonomy appears to be of interest to fewer and fewertheir differentiation, creating annectant groups that American scientists each year. He calculated from
obscure the origins of these lineages. bibliographies of papers on mosquitoes that were
Belkin' s Mosquitoes of the South Pacific set a new published regularly inMosquito News that the proportionstandard for systematic studies of mosquitoes. ofpapers dealing with taxonomy dropped from 6percentUnfortunately, no mosquito fauna elsewhere in the in the years 1951- 1952 to 2. 7 percent in the years 1971-
world has been studied and analyzed as thoroughly as 1972. This is more than a 50 percent decline in just 20that of the South Pacific. Again, the evidence suggests years. Darsie commented also on the diminishingthat much work in mosquito systematics remains to be number ofmuseums and universities that were supporting
done. research on mosquito systematics.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 71
Eldridge( 1974) noted that the taxonomy ofmosquito for systematic studies on this very important group ofvectors must be understood thoroughly before mosquito- insects [ the Culicidae]." And, a fourth provided the
borne diseases can be understood or controlled. He following statement: " The Culicidae are probably thestated that thenumberofundescribed or poorlydescribed best known family of invertebrates; an enormousmosquito species worldwide is undoubtedly literature has developed and yet phylogenetic
considerable. classification is in disarray. Many people assume thatNielsen ( 1980) stated that progress in describing all the major problems have been worked out. Nothing
mosquito species has been good, but that progress in could be farther from the case. Most work on mosquito
other aspects ofmosquito systematics is rather poor. He taxonomy was done in the period 1900- 1940. The lastmentioned ecology, distribution, behavior, and major revision was in 1932. The great centers of
phylogenetic relationships of mosquitoes as areas of taxonomic research, the U. S. National Museum and the
study in which ourknowledge is deficient. He noted that British Museum, dropped the culicidologists from their
morphological characters always have been and still staffs years ago. Yet the modern understanding ofcontinue to be the most extensively used characters in species and higher categories has not yet been applied to
mosquito taxonomy, but then went on to point out that Culicidae. Problems in disease transmission and vector
some kinds of morphological characters are little used. competence require taxonomic data that are not available.
He stated that the scanning electron microscope has not . . . As a teacher, I have often been frustrated by thebeen adequately exploited in mosquito taxonomy. primitive level of knowledge of evolution of the
Barr( 1986) traced the history of the development Culicidae. We identify species as though they wereofmorphological analysis of mosquitoes and discussed divinely created and we pay no attention to thevarious types of nonmorphological data that have been evolutionary framework."of great value in resolving complexes of sibling species It seems clear from my examination of mosquitoofmosquitoes. He noted,however, that most species we systematics, from the statements ofother scientists who
recognize still can be identified morphologically. He work with mosquitoes, and from the statements of
drew attention to the fact that numerical taxonomic reviewers of proposals for revisionary work onmethods, which allow the analysis of vast amounts of mosquitoes that the overall state of the field is rather
data, have not been widely used for the study of poor. Even more unfortunate is the fact that there are
mosquitoes, but that such techniques have the potential few opportunities to rectify this situation, for thereto help us sort out species and their relationships better simply isnot support, in the form ofpositions in museumsthan has been done in the past. and universities, for mosquito systematics.
Taken as a whole, the comments of these authors In an analysis of the state of mosquito systematics,
paintadismal picture of the state ofmosquito systematics: I believe it is useful to think in terms of historical eras.
both the number of positions for mosquito taxonomists For our purposes here, appropriate terms would be the
and the number of papers in the field have decreased; Pre- Belkin Era, the Belkin Era, and the Cladistics Era.
there are still many undescribed or poorly described If we think in these terms, then it becomes obvious that
species, poorly known groups, little-studied faunas, and our knowledge of the vast majority of the world' sunresolved phylogenetic relationships; and, newer mosquitoes is two eras behind where it could and should
instruments and methodologies are either insufficiently be. Since most groups of mosquitoes have not been
utilized or not used at all. revised in five ormore decades, our knowledge of these
I was a co-principal investigator ofan unsuccessful groups is still in the Pre-Belkin Era. And, since the
grant application for a systematic study of mosquitoes concepts and methodology of cladistics have not beenthat was submitted to the National Science Foundation applied to any mosquitoes, it is obvious that we have notseveral years ago. It is obvious from the comments of yet entered the Cladistics Era.
some of the anonymous reviewers of this proposal that Faran( 1979) called for an interdisciplinary approachthey consider the classification of mosquitoes to be to solving problems in mosquito systematics. Hebelievedsubstandard. According to one reviewer: " The status of there should be increased communication, interaction,
mosquito systematics is an irony. At the level ofspecies and cooperation among investigators. I also think therediscrimination, mosquito workers have some of the best are ample opportunities for cooperation between
andmostresolveddata, butthedataislargelyunanalyzed mosquito taxonomists and investigators usingin considering how these taxa are to be grouped." nonmorphological data. Some of the studies I have done
Another reviewer stated: " Classification of Culicidae on complexes ofmorphologically similar species wouldseems to be especially difficult. There is a need for a have been made immensely easier if I had had, forcomplete revision." A third noted: " There is a dire need example, electrophoretic data providing independent

72 BULL. SOC. VECTOR ECOL. JUNE, 1994
evidence of the reproductive isolation of populations. Eldridge, B. F. 1974. The value ofmosquito taxonomyUnfortunately for those who utilize the newer types of to the study of mosquito-borne diseases and theirdata, not only is the state ofmosquito systematics rather control. Mosq. Syst. 6: 125- 129.poor, but also there are virtually no systematists with Faran, M. E. 1979. The importance of an integratedwhom to collaborate. approach in solving a problem in mosquito
systematics. Mosq. Syst. 11: 280-288.Acknowledgments Mayr, E. 1969. Principles of Systematic Zoology.
McGraw-Hill Book Co., New York. 428 pp.I thankTruls Jensen for inviting me to participate in Nielsen, L. T. 1980. The current status of mosquito
the Mosquito Systematics and Vector Ecology systematics. Mosq. Syst. 12: 1- 6.Symposium. I also thank Mary Jane Niles and two Theobald, F. V. 1910. A Monograph of the Culicidaeanonymous reviewers for suggesting improvements to or Mosquitoes. Vol. 5. British Museum( Naturalthe manuscript. History), London. 646 pp.
Zavortink, T. J. 1974. The status of taxonomy ofREFERENCES CITED mosquitoes by the use ofmorphological characters.
Mosq. Syst. 6: 130- 133.Barr, A. R. 1986. Bases of mosquito systematics. J. Zavortink, T. J. 1979. Mosquito Studies ( Diptera,
Am. Mosq. Contr. Assoc. 2: 261- 266. Culicidae). XXXV. The new sabethine genusBates, M. 1949. The Natural History of Mosquitoes. Johnbelkinia and a preliminary reclassification of
The Macmillan Co., New York. 379 pp. the composite genus Trichoprosopon. Contrib.Belkin, J. N. 1962. The Mosquitoes of the South Am. Entomol. Inst. ( Ann Arbor). 17( 1): 1- 61.
Pacific( Diptera,Culicidae). University ofCalifornia Zavortink, T. J. 1990. Classical taxonomy of mos-Press, Berkeley and Los Angeles. 2 vols. quitoes— A memorial to John N. Belkin. J. Am.
Darsie, R. F., Jr. 1974. Introduction [ Symposium: Mosq. Contr. Assoc. 6: 593- 599.Mosquito Taxonomy— IsitNeeded?J. Mosq. Syst. Zavortink, T. J. 1993. Banksinella luteolateralis var.6: 111- 112. albothorax Theobald, 1907( currently Aedes( Neo-
Edwards, F. W. 1932. Diptera. Fam. Culicidae. In melaniconion) albothorax), B. luteolateralis var.Wytsman, P., Genera Insectorum. Fascicle 194. circumluteola Theobald, 1908 ( currently A. ( N.)Desmet-Verteneuil, Brussels. 258 pp. circumluteolus) andA.( N.)mcintoshi Huang, 1985
Eldridge, B. F. 1969. The role ofmosquito taxonomy Insecta, Diptera): proposed conservation of thein medical entomology. Mosq. Syst. News Lett. 1: specific names and designation of a neotype for A.19- 21. N.) albothorax. Bull. Zool. Nomencl. 50: 39-43.

BULL. SOC. VECTOR ECOL., 19( 1): 73- 81 JUNE, 1994
ROSS RIVER VIRUS: DISEASE TRENDS
AND VECTOR ECOLOGY IN AUSTRALIA'
R. C. Russell2
ABSTRACT: Ross River vines, a mosquito-borne alphavirus, is endemic and annually active in Australia. Althoughthe case distribution is scattered, local epidemics occur in both coastal and inland regions. Rural townships are mostaffected, but major cities have recently reported local cases. The virus is arguably the arbovirus of greatest publichealth importance in Australia. The infection is not life-threatening, but the morbidity caused by the debilitatingpolyarthritis syndrome is a serious economic concern. The epidemiology is complicated by the existence of manystrains that are biochemically distinct, with apparently different virulence and possibly different immunogenicity;some strains have a greater propensity for infection of some vector species. It is thought that the virus has mammalian
rather than avian vertebrate hosts and there is increasing evidence that the virus survives in arid areas by transovarialtransmission. The virus is quite catholic in its association with mosquito species. Virus activity appears to be drivenprimarily by local vector activity. The most important epidemic vectors are the principal rural inland ( Culexannulirostris) and coastal( Aedes vigilax and Aedes campto rhynchus) pest mosquitoes, but other species, particularlyfloodwaterAedes( e. g. Ae. normanensis), appear to be important in early season activity in inland regions, and a rangeof species in various genera have yielded virus in different situations. In recent years, outbreaks have occurred inmany parts ofAustralia and the factors influencing the activity are usually characteristic of the locality and the region.For reasons associated with mosquito and human ecology, the risk of disease is increasing for many communitiesand the virus is likely to continue as a public health problem with a significant economic impact in rural Australia.
INTRODUCTION tropical northern Australia in the Northern Territory,Queensland and the northern halfof Western Australia,
Ross River( RR) virus, amosquito-borne alphavirus, as well as in the temperate southeastern states of Newis endemic and annually active in much of Australia South Wales, Victoria and Tasmania, and also Southincluding all mainland states and the island ofTasmania Australia and the southwest ofWestern Australia. ThereMudge and Aaskov 1983). The natural vertebrate hosts are indications that humans may act as reservoir hosts in
of RR virus are thought to be non-migratory native epidemic situations and serve to distribute the virusmarsupial animals, such as kangaroos; birds are not geographically. There is typically a scattered humanconsidered to be suitable hosts. There is increasing case distribution, but local epidemics occur in bothevidence that the virus survives in arid ( and perhaps coastal and inland regions. Smaller rural townships areother) areas by transovarial transmission. most commonly affected, although local cases have
The disease caused by RR virus was first recognized occurred in all mainland capital cities ( Adelaide,in the late 1920s, although the virus and its association Brisbane, Darwin, Melbourne, Sydney, and Perth) inwith the disease were not identified until the early recent years.
1960s. It was during the 1980s that the virus and its Ross Rivervirus is arguably the arbovirus ofgreatestdisease syndrome became more familiar to laboratory public health importance in Australia. Symptoms mayworkers, clinical practitioners and health authorities, include fever, rash, myalgia, polyarthralgia, andand an appreciation of the amount of annual virus polyarthritis. While the virus infection is not life-
activity, resulting human infection, and associated threatening ( unlike the flaviviruses Murray Valleymorbidity accrued. During recent years, there has been encephalitis virus and Kunjin virus), the morbidityannual activity in each of the states including regions of caused by the debilitating polyarthritis syndrome ofRR
Paper Presented at the 1st International Congress of Vector Ecology, San Diego, California, USA, 4 October 1993.
2University of Sydney, Department of Medical Entomology, Centre for Infectious Diseases and Microbiology,Westmead Hospital, Westmead, NSW 2145, Australia

74 BULL. SOC. VECTOR ECOL. JUNE, 1994
virus infection is of considerable social and economic species)( TABLE 1).
concern for local communities, although the full Virus activity in tropical northern arid Australiaeconomic impact is not fully appreciated by health and ( e.g. northern Western Australia, the Northern Territoryother government authorities. In recent years it has and Queensland) often follows above-average seasonal
become recognized that another Australian alphavirus, rainfall sufficient to generate large populations of the
Barmah Forest ( BF) virus, causes an infection with temporary ground pool, floodwater Aedes species, suchsimilar general symptoms ( including polyarthritis) to as Ae. normanensis( Taylor). A similar situation occurs
RR virus infection, and it is likely that there has been in some temperate southern regions ( e.g. New Southsome confusion in the clinical diagnoses of the infections. Wales, Victoria, southern South Australia): in coastal
Nevertheless, RR virus is the predominant disease- salt marsh areas, spring and early summer rains provide
causing arbovirus in Australia. suitable conditions for the buildup of large summerReviews of the general epidemiology and ecology populations ofAe. camptorhynchus( Thomson) and Ae.
of the virus have been presented by Kay and Aaskov vigilax ( Skuse); in the drier inland river flood plain
1988) and Marshall and Miles( 1984), and it is beyond areas, pre-summer rains promote large populations of
the scope of this paper to cover that material in detail. floodwater Aedes species such as Ae. bancroftianus
However, it is becoming increasingly apparent that the Edwards, Ae. sagax( Skuse) and Ae. vittiger( Skuse).
ecology of the virus isnot simple; indeed itmight be said Ross River virus has been isolated from females of
that there are several ` epidemiologies' of RR virus such Aedes species shortly following rains in Newdisease in Australiadepending on the locality of infection, South Wales- from a mixed pool of Ae. bancroftianus,
the strain of virus, and the species of vector. Ae. theobaldi ( Taylor) and Ae. sagax ( Russell 1989,
The existence of many strains of RR virus that are 1992), and in Western Australia- from Ae. normanensis
biochemically distinct, with apparently different ( Broom et al. 1989). The virus has also been isolated
virulence ( at least to mice), and possibly different from male mosquitoes from inland New South Wales-
immunogenicity, has been demonstrated( see Kay and a mixed pool of Cx. annulirostris Skuse and Cx.
Aaskov 1988). More recent research on nucleotide australicusDobrotworsky& Drummond and Cq. linealiscomposition of 80 isolates, has shown that amongst ( Skuse) males( Russell 1989), and also from Western
these strains from throughout Australia there are at least Australia - from Ae. vigilax and from Ae. tremulus
four topotypes( genotype associated with a geographical ( Theobald) males( Lindsay et al. 1993a). These data( forregion), and strains with identical RNase T1 maps both male and female mosquitoes) indicate that vertical
circulate during both epidemic and non-epidemic years transmission is occurring in natural situations as
Lindsay et al. 1993b). Unpublished work in my suggested by the laboratory work of Kay ( 1982) andlaboratory has shown that some strains are more highly Vale et al.( 1992).
infective for some populations of some vector species, After the initiation of virus activity in inland areas,
indicating possible geographical associations in some Cx. annulirostris soon becomes the most important
instances( P. Wells, R. C. Russell and M. J. Cloonan, vector in mid-summer in the temperate ( and manyunpublished data). tropical) areas, although the role of Cq. linealis (and
other species) may be underestimated. Aedes vigilaxVECTORS AND VECTOR ECOLOGY and Ae. camptorhynchus appear to be the most important
coastal vectors in their respective distributions
Ross River virus activity appears to be driven throughout the infection season, but Cx. sitiens
primarily by local vector activity, and seasonality of Wiedemann may have an important involvement ininfection in human populations is related to the local some localities.
abundance of known or potential vectors. The most
important epidemic vectors are the principal rural pest RECENT VIRUS ACTIVITY AND DISEASE
species, although a range of species in various genera OUTBREAKS
have yielded virus in field studies in different situations
from many parts of Australia and may be variously Over the pastdecade, the Commonwealth( national)
important in natural enzootic or epidemic cycles. The Department of Health has conducted a routine
virus appears to be very catholic in its association with surveillance of communicable diseases in Australia
mosquito species; RR virus has been isolated from wild using a reporting system from sentinel laboratories incaught mosquitoes from the genera Aedes( 17 species), each state. This has resulted in an increased recognition
Anopheles( 2 species), Coquillettidia( 2 species), Culex of the extent and regularity ofhuman infections with RR5 species), Mansonia( 1 species), and Tripteroides( 1 virus throughout Australia, but this sentinel system

JUNE, 1994 BULL. SOC. VECTOR ECOL. 75
TABLE 1. Mosquito species and geographic region from which Ross River virus has been isolated in Australia
compiled from various published and unpublished data). Species represented in mixed pools, which
yielded virus, have not been included.
Genus Species States- regions
Aedes alternans NSW- south coast; WA- north coast
bancroftianus Vic- north inland
camptorhynchus Vic- east coast; Tas- east coast; WA- south/west
clelandi WA- sth/west
daliensis WA- nth coast
flavifrons Tas- east coast
funereus NSW- nth coast
normanensis Qld- sth inland; WA- nth inland;
NT- nth inland
notoscriptus NT- urban Darwin
phaecasiatus NT- urban Darwin
procax NSW- nth coast
ratcliffei WA- sth/west
sagax Vic- Murray valley; WA- nth inlandtheobaldi Vic- Murray valleytremulus WA- nth inland
vigilax NSW- nth& sth coast; Qld- nth& sth coast;
WA- nth coast; NT- coast, urban Darwin
undesc. species WA- nth inland, nth coast
Anopheles amictus s. l. NSW- nth inland; Qld- nth inland
annulipes s. l. NSW- Murray valley;WA- sth/west, nth inland
Coquillettidia linealis NSW- sth inland, nth& sth coast;
sp.nr. linealis WA- sth/west
Culex annulirostris NSW- nth& sth inland, nth coast;
NT- urban Darwin, nth inland, nth/east coast;
Qld- nth& sth inland; WA- nth inland, nth coast
Normanton sp.' form] WA- nth inland
australicus Vic- nth inland; WA- nth coast, sth/west
quinquefasciatus WA- nth inland, nth coast
sitiens WA- nth coast
Mansonia uniformis NSW- nth inland
Tripteroides undet. species WA- sth/west
presents arecognized under-reporting ofactual incidence each state and territory. However, this system is alsoand an incomplete picture of geographic distribution. somewhat defective inasmuch as' RR virus infection' is
More recently, a National Notifiable Diseases not separated from ' arbovirus infection' ( which maySurveillance System has been established that receives include other alpha- as well as flavivirus infections) bydata acquired by government authorities ( notified by all states ( e. g. South Australia and Tasmania) forconfirming laboratories and/or medical practitioners) in notification purposes.

76 BULL. SOC. VECTOR ECOL. JUNE, 1994
The notification system is also limited by the were notified for the six months to the end ofJune 1993,
efficiency of the reporting systems, which are still not approximately twice as many as for the same period inuniform between states. Although there have been 1992, but less than the numbers for that period of 1984.
attempts to increase specificity by moving to laboratory In recent years, better case reporting and betterreporting and recording only serologically confirmed serological techniques( and interpretation thereof) have
cases, there is substantial anecdotal evidence that at increased the quality of the data. However, the non-times when human infections are increasing, there is, uniformity of the data collecting processes makes itparadoxically, a relative decrease in notifications and impossible to compare national case numbers between
thus an increase in underreporting ofcases. This occurs years, although it is arguably not a valid comparisonwhen patients become increasingly aware of current anyway as state and regional data vary between andinfections and do not present to physicians because they within states with different vector mosquito activity,know there is no specific treatment or cure, and when and local meteorological and tidal influences.
physicians increasingly diagnose on clinical symptoms In Queensland, which is often described as havingrather than send blood samples for serological testing. the greatest endemic activity of RR virus in Australia,Thus, actual incidence of RR virus disease is difficult to the summer of 1991- 92 provided the biggest outbreak in
estimate at times of both high and low activity. some years. There was a dramatic and real increase in
Unfortunately, there has also been a lack of uniformity disease during the first six months of 1992, well abovebetween laboratories in their diagnostic techniques, the effect of increases in the reportage rate, growinginterpretation ofresults, and reporting criteria. However, medical awareness, and demand by the public fora series of clinical and serological definitions, with diagnostic tests. A total of more than 3600 cases was
guidelines for testing and reporting, was proposed reported from the first six months of the calendar year,
recently for national use by testing laboratories much above the figures for the whole of previous yearsMackenzie et al. 1993). 1, 955 cases for 1991, 1, 436 for 1990, 1, 408 for 1989,
Notwithstanding the above limitations, the total 478 for 1988, 952 for 1987, and 945 for 1986. Thenational case numbers reported through the sentinel increase in the number of cases reported in 1992 was
laboratory reporting system for the years( July 1 through most dramatic from late February to May, and theJune 30) 1988-89 to 1992- 93 were as follows: 1988- 89 majority of the 3,622 notifications were diagnosed from
2, 188 cases, 1989-90 = 728 cases, 1990-91 = 884 the southeast region of the state in contrast to the
cases, 1991- 92= 5, 287 cases, 1992-93= 5, 333 cases. previous years when the highest case rates had been
These data indicate the fluctuations between years and recorded in the north. An analysis of meteorological
the relative size of the problem, particularly for the past data from both regions showed the case rate appeared totwo years. Recent analysis of data from the correlate with the pattern of rainfall ( Phillips et al.
Commonwealth Department of Health has shown the 1992). Meteorological conditions from December 1991adjusted annual rate of notification for 1992 was 36.4 through the early months of 1992 in the densely populatedper 100,000 population, an increase from 22.9 per southeast coastal area were highly conducive to the100,000 in 1991( 1992 activity was greatest in the north: breeding of the salt marsh Ae. vigilax, and the rainfallQueensland had an annual rate of 139.6 per 100,000 and also disrupted routine vector control measures thatthe Northern Territory a rate of 134. 1 per 100,000). normally follow the high tides; vector populations wereNationally, the seasonality of infection in 1992 was thus well above normal levels. Later extensions of the
strongly marked; there was a peak( 1, 602) of cases with outbreak to the north along the coast were associatedonset in March, and a low ( 123) in July; there was a with further rainfall in late- summer.national female/male ratio of 1. 12/ 1 and there was a Despite the record numbers of cases notified, thebell-shaped age distribution of cases with the peak case rates for the heavily populated localities in theincidence of notified cases occurring in the 35- 39 years southeast outbreak in 1992 were much lower than theage group with64.5 notifications per 100,000 population 1992 and previous years' reports for more sparselyHall, 1993). populated rural areas. One small township in the west of
In 1993, following record spring and early summer the state yielded a case rate of 1, 786/ 100,000( 10 casesrainfall in South Australia, northwestern Victoria, and in population of 560) compared with 97/ 100,000 for thesouthwestern New South Wales, the largest outbreak of capital city. The overall state rate was 135/ 100,000 forRR virus disease recorded to date for the southeast of the 1992 and was 71/ 100,000 for 1991( J. Sheridan, personalcontinent occurred; in Victoria there were more than communication). The risk of disease is greater for the1, 300 notifications and in South Australia there were rural dweller in Queensland, as it appears to be also inmore than 800; in New South Walesalmost 600 infections the other states where data is available.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 77
In New South Wales, the largest recorded outbreak been revealed as probably a vector of at least secondaryof RR virus infections occurred in the summer of 1983- importance as it has yielded RR virus from a number of84 when 1, 196 serologically confirmed cases were localities in both inland and coastal areas. Although Ae.recorded( Hawkes et al. 1985). In this outbreak, the first camptorhynchus ( a vector in coastal areas of easterncases occurred in mid- late spring in western arid areas Victoria and eastern Tasmania and also in the southwestfollowing rains, which generated large numbers of of Western Australia) is present on the south coast offloodwaterAedes( Ochlerotatus) species, including Ae. New South Wales, it is only rarely abundant and hassagax,Ae. theobaldi, andAe. vittiger,before the seasonal never yielded virus in this state( Russell 1989, 1992).appearance of Cx. annulirostris. The epidemic In Victoria, during the last 10 years, the frequencysubsequently ` exploded' in the southwest of the state has been low in some calendar years and much higher induring the summer months when populations of Cx. others, ranging from a minimum of 5 cases in 1983 to aannulirostris, typically the most abundant species in the maximum of 556 in 1989, until 1993 when there hasinland during summer( and particularly more abundant been more than 1, 300 cases to the end of August( R.in areas with irrigated agriculture), rose well above Moran, personal communication). During the season ofaverage summer levels because of continuing heavy 1988- 89, more than 700 cases of RR virus disease wererainfall, which augmented the natural and irrigation- reported. The greatest proportion of cases originated inderived habitats. Cases were recorded in all inland the eastern coastal region of Gippsland and followedregions of the state, and also on the north and south three to four weeks after heavy rainfall in the salt marshcoasts where Ae. vigilax was the likely vector. areas produced unusually large populations ofadult Ae.
Following 1984, there was relatively low- level camptorhynchus; RR virus was isolated from fourannual activity until the summer of 1988- 89, when case collections of this species. The case reports peakednumbers again rose substantially and 476 serologically before the end of December 1988, and the majority ofconfirmed cases were recorded. Cases occurred from cases occurred by mid-summer. Serological surveysDecember 1988 to July 1989 with the majority of cases indicated that approximately 25 percent of the humanin January and February, the seasonal pattern being population in one locality had been previously infectedsimilar throughout the state( Hueston and Cunningham with RR virus, and the seroconversion rate from the1989). Ross River virus was isolated from mosquitoes outbreak was estimated as about 10 percent withcollected each month from January 1989 through May approximately 30 percentof these experiencing classical1989 ( Russell 1989; R. C. Russell, unpublished data). symptoms. In the following year( 1989-90) there wasIn subsequent years, 292 cases were reported to the New low rainfall throughout Victoria and only 34 casesSouth Wales Department of Health in 1990, 476 in 1991 ( mostly from the northern irrigation areas along thean average case rate of 8. 4/ 100,000, although one Murray valley where Cx. annulirostris was the most
western inland region had arate of 106/ 100,000), 345 in likely major vector) were reported( Wolstenholme 1991).1992, and 573 to the end of August 1993 ( R. Hunter, In 1990-91 there were 478 cases ( with 43% inpersonal communication). western region), and in 1991- 92 there were 135 cases.
Prior to the end of 1991, arboviral infections were Virus activity was strong again in the summer of 1992-reported by medical practitioners and it was generally 93 when there were 1, 201 cases notified( Norris 1993).accepted there was underreporting. From 1992, infections In contrast to 1988- 89, almost 80 percent were in thebecame notifiable by confirming laboratories in order to inland north and northwest and only 2 percent in theincrease efficiency and accuracy. Thus, data must be Gippsland coastal region. The species most likely totreated with caution when comparing years before and serve as the principal vector in the northern inlandafter 1992. For pre- 1992 data also, there is some concern regions in 1992-93 was the freshwater species Cx.as to comparisons between years because of increased annulirostris, associated with natural and irrigation-efficiency in data collection in some regions after the derived ground waters. The disease notifications by1984 and the 1989 outbreaks. week of onset reflected this with the greatest numbers
An arbovirus surveillance program has sampled occurring in late summer and early autumn.mosquito populations throughout the state since 1985, In Tasmania, the first serologically confirmed casesand since 1988 has processed the samples for virus iso- occurred in 1981, and since then the incidence of thelation. This has provided data that reinforces the impor- disease has varied with seasonal conditions; smalltance of Cx. annulirostris in the inland and Ae. vigilax epidemics occurred in 1982 with 21 cases, 1989 with 59on the coast as the major vectors of RR virus in New cases, and 1991 with 24 cases. A wet early autumnSouth Wales. However, amongst various other species appeared to be responsible for the activity in the 1989which have occasionally yielded virus, Cq. linealis has outbreak, when most cases were apparently infected in

78 BULL. SOC. VECTOR ECOL. JUNE, 1994
April. By contrast, in the 1991 outbreak, the majority of populations involved. The first outbreak occurred during
patients contracted the virus in mid-summer, following the spring and summer of 1988- 89. From Novemberunseasonably heavy January rain which coincided with 1988 through December 1989, more than 620 cases of
very high tides bringing inundation of coastal lagoons RR virus infection were reported from the southwest of
and marshland and generating large populations of the the state in coastal areas where very high tides inundatedvectors Ae. camptorhynchus( from which RR virus was large areas ofmarshland, and well above average winter
isolated) and Ae. flavifrons( Skuse)( which had yielded and spring rainfall in 1988 provided ideal habitatRR virus from the region in 1985) ( McManus and conditions for the salt marsh speciesAe. camptorhynchus
Marshall 1986; McManus and Russell et al. 1992). and Ae. vigilax( Condon 1991; Lindsay et al. 1992).In South Australia, the Riverland area, which is a Although Ae. vigilax was the dominant species during
continuation of the Murray River valley south from its peak summer,Ae. camptorhynchus numbers were above
New South WalesNictorian course, has historically normal and persisted well into the summer to overlap
been the area of greatest activity of RR virus( Mudge with Ae. vigilax.The early cases were probably infected1977). The largest outbreak of RR virus in the state in in mid-October when Ae. camptorhynchus was the only
recent times occurred during the 1992- 93 summer; from abundant mosquito, and there were four isolates of RR
September 1992 to the end of May 1993 there were 821 virus made from this species collected in the region in
notifications ( S. Selden, personal communication). early November.Record rainfall in 1992 across the state contributed to an Three years later, a total of 555 cases was reported
increase in mosquito numbers and several parts of the for the same southwest region between 1 September
state( including some usually semi-arid areas) reported 1991 and 30 April 1992, but this time there was a
cases for the first time, although in the final analysis the number of cases acquired within the metropolitan area
major area for infection appeared again to be the Murray of Perth. A record amount of rain fell in the southwest
valley. Serological notifications rose above expected between November 1991 and the end of February 1992levels in mid-December 1992 with a peak in the last ( more than 10 times the monthly average fell inFebruary),week of February 1993. The cases were more however, mean sea- level and tides were normal, unlike
concentrated in the drier areas to the west in the early during the 1988- 89 epidemic mentioned above. Thephase of the epidemic, with the Riverland and other unseasonable rains allowed both salt marsh and
eastern areas of the Murray valley showing peak freshwater mosquitoes to proliferate throughout the
notifications later in mid-summer. Ross River virus has summer months, and although in most years Ae.
neverbeen isolated from mosquitoes in South Australia, camptorhynchus is replaced by Ae. vigilax during theand thus the local vectors are, to that extent, unknown; hottest months, in the 1991- 92 summer Ae.
however, Cx. annulirostris is abundant in summer and camptorhynchus persisted in large numbers through
is the prime candidate in the Riverland area, Ae. late spring and summer; as had been the case duringcamptorhynchus is present and may be involved in the 1988- 89, but because of high tides and not rainfall. In
southern areas towards the coast where Ae. vigilax also the Perth metropolitan area, the rainfall resulted in large
exists in some localities, and other Aedes species may increases in populations of the freshwater vectors Cx.
have been involved in the transmission reported from annulirostris and Cq. linealis. Isolates ofRR virus werethe more and western regions in the early months of the obtained fromAe. camptorhynchus( 10) and Cq. linealis1992- 93 outbreak. 2) during the course of the outbreak. The data suggested
In Western Australia, an analysis of serological that the outbreak was initiated and maintained for the
diagnostic data from the State Health Laboratory Service first two or three months by the large late spring andfrom 1975 to 1988, showed RR virus infections had early summer population of Ae camptorhynchus, andoccurred throughout the state, with approximately 60 the virus activity spread into the Perth area as thepercent of cases in the southwest ( predominantly number of freshwater mosquitoes increased during
during summer and autumn) and 40 percent in the summer. The metropolitan activity was widespread,north ( throughout the year, and usually confined to 200 cases were reported from 104 suburbs. Epidemio-
coastal areas during the dry season but in coastal and logical and demographic data indicate that the rapid
inland areas during the wet season). Ross River virus spread of virus was due to the movement of infected
was made notifiable in 1985. In the north of the state the humans.( Lindsay et al. 1989, 1992; M.Lindsay, personalcase attack rates have been up to 10 times higher than in communication).
the southwest; however, the two most recent major Interestingly, outbreaks in the coastal areaofcentraloutbreaks in the state have occurred in the southwest ( Pilbararegion) WesternAustraliaappeartoberestricted
and differed in cause, mosquito vectors, and human to the cooler months of the year following unusually

JUNE, 1994 BULL. SOC. VECTOR ECOL. 79
heavy winter rains; Ae. vigilax is the likely vector but annulirostris is the major vector in all areas, Ae. vigilax
high tides and cyclonic rains in the hotter months of the is a vector in coastal areas in the early wet season, and
year do not lead to outbreaks ofRR virus. This contrasts Ae. normanensis is a probable vector in the inland drier
with the situation further north on the coast ( West areas ( Tai et al. 1993; P. Whelan, personal commu-
Kimberley region) where outbreaks ostensibly also due nication). In the following year, 1991- 92, there was ato Ae. vigilax have occurred in the hottest months of the concurrent outbreak of RR virus and BF virus causing a
year. It is apparent that what occurs with vectors and similar illness in a community on the Gove peninsula inviruses in one arid coastal region of Western Australia the northeast of the Arnhem Land region; 28 cases ofRR
does not occur in another. virus infection and 43 cases of BF virus infection were
In the northern( Kimberley) region, some cases of serologically indicated, and RR virus and BF virus wereRR virus infection occur each year, and have been isolated from Cx. annulirostris and Ae. vigilax,
recorded throughout the year although the greatest respectively ( A. Merianos and P. Whelan, personalactivity is associated with the wet season. Virus isolations communications).
and timing implicate Ae. normanensis and Cx.annulirostris as the major epidemic vectors. In early THE FUTURE: THE VECTORS, THE VIRUS,
1991, approximately 30 cases were recorded from towns AND THE COMMUNITY HEALTH
in the West Kimberley region following heavy rainsassociated with seasonal cyclonic activity; in early For reasons associated with mosquito and human
1993, small outbreaks again occurred in these northern ecology, the risk of RR virus disease is increasing forareas but full details are not yet available. Although RR many communities, and the virus is likely to continue asvirus has been isolated from Ae. vigilax from coastal a public health problem for the foreseeable future. Over
areas in more northern areas of Western Australia, the the past decade, an average of more than 1, 000 cases per
species appears to be not a major vector of RR virus in year nationwide has been reported from the sentinel
the southwest as it has never yielded virus in this region. laboratories alone to the national Department ofHealth;
Aedes camptorhynchus has yielded virus on a number of the actual numbers of infections are unknown but
occasions from the southwest and appears to be the variously estimated at up to 10 times reported figures.more important vector in that region. In the inland parts Although the rheumatic effects of the viral infection
of northern and central Western Australia, Cx. may last for about six months on average, they areannulirostris is generally the most important vector known to persist for at least six years ( Fraser 1986).
species for RR virus, after the first flush of activity of More recent data from New South Wales and Western
Aedes species from which virus has also been recovered Australia suggest longerperiods of incapacity associatedTABLE 1). However, Cx. annulirostris ( as with Ae. with the infection, but definitive morbidity studies are
vigilax) does not appear to be a major vector in the yet to be undertaken. The recent epidemic in South
southwest region. Australia is currently being investigated with a view toIn the Northern Territory, there appears to be an measuring morbidity and estimating economic impact.
increasing annual incidence of infection; 10 cases were The financial burden to the community, through directdocumented for 1984- 85, progressing to 84 in 1989- 90. costs associated with health care and loss ofproductivity,
Thereafter there have been 368 cases in 1990-91, 152 in particularly in agricultural areas, and indirect costs such1991- 92, and 230 in 1992- 93. Although there has been as those associated with loss of tourists and seasonal
an increase in efficiency of reporting, health authorities itinerant workers, can only be estimated. An assessmentbelieve the figures represent a real increase in incidence. of the economic significance of the outbreak in New
The largest recordedepidemic ofsymptomatic RR virus South Wales in 1983- 84, where 1, 196 cases statewide
infection occurred in the 12 months between July 1990 were serologically identified and a detailed study of aand June 1991, with 368 cases ( total population of single town showed an average incapacity of about six155,000) notified compared to an annual mean for the weeks, arrived at a conservative estimate of the overall
previous decade of 28. 8 cases. The infection season cost to have been$ 3 million( assuming the outbreak was
correlated with the wet season in this tropical region; the confined to the 1, 196 cases) or approximately$ 2,500epidemic peaked in January, andrural towns and districts per case ( Hawkes et al. 1985). In Queensland, again
were affected more than the cities. A major factor assuming that all cases were notified, the 1992 outbreakcontributing to the outbreak was thought to be the very was estimated to have cost $ 14 million ( J. Sheridan,
high rainfall for the season, the highest seasonal rainfall personal communication).
recorded in the last 10 years; the local vectors were not A recent study has revealed that antibody prevalencedetermined but previous data has indicated that Cx. rates to RR virus were substantial in various areas

80 BULL. SOC. VECTOR ECOL. JUNE, 1994
throughout inland New South Wales and northern beyond our understanding will increase the risk ofVictoria ( 25%- 72%), and were uniformly higher in infection with RR virus for human communities in1991 than they had been in 1981, with 10 year sero- Australia. Health authorities must recognize that aconversion rates of 24 percent for RR virus infection concern for future situations, where an increase inHawkes et al. 1993, Boughton et al. 1984). Whether mosquito-borne arbovirus disease has been predicted to
this is representative of other states is unknown, but follow the expected global warmingphenomenon, shouldthe outbreaks in recent years confirm there is at least be secondary to efforts to deal with an already existingsubstantial continuing ifnot increasing activity in many substantial and apparently increasing public healthareas of Australia. problem with RR virus.
If there is a real increase in virus infections inhuman communities, then this could be attributable to Acknowledgmentsincreases in virus activity in native or domestic animalvertebrate hosts, increases in vector abundance, and/or For unpublished information on human caseincreases in contact between vectors and humans numbers and virus isolates from mosquitoes I wish toresulting in transmission. It can be argued that in many thank J. Hargreaves ( Commonwealth Department ofareas there has been an increase in contact between Health), R. Hunter and W. Manning( New South Walesvector populations and human communities. Depending Department of Health), M. Lindsay ( University ofon the situation, this may have resulted from a real Western Australia), N. Marcon and R. Moran( Victorianincrease in vector populations and/or also from an Department ofHealth), I. Marshall( Australian Nationalincrease in amount of contact between humans and University), T. McManus ( Tasmanian Department ofpreviously existing vector populations. Primary Industry), S. Selden( South Australian Health
There are two general scenarios that, in at least Commission), J. Sheridan( Queensland Department ofsome areas of inland and coastal southeastern Australia, Health), and A. Merianos and P. Whelan ( Northernmight explain the apparent increase in virus/vector Territory Department of Health). Other unpublishedcontact with humans and a concomitant increase in virus isolations listed are from R. C. Russell and M. J.disease incidence/prevalence. There are data to support Cloonan.the contention that real increases in mosquito abundancehave occurred in some inland and coastal areas; in the REFERENCES CITEDformer because of provision of irrigation waters tootherwise arid areas with resultant increases in pre- Boughton, C. R., R. A. Hawkes, H. M. Nairn, J. Wild,summer populations of some Aedes species, and mid- and B. Chapman. 1984. Arbovirus infections insummer populations of Cx. annulirostris and, to a lesser humans in New South Wales: Seroepidemiology ofextent, Cq. linealis; in the latter where agricultural, the alphavirus group of togaviruses. Med. J. Aust.residential, and industrial developments in estuarine 141: 700-704.areas have resulted in changes in drainage patterns in Broom, A. K., A. E. Wright, J. S. Mackenzie, M. D.mangrove and saltmarsh habitats with resultant increases Lindsay, and D. Robinson. 1989. Isolation ofin distribution and abundance of populations of Ae. Murray Valley encephalitis and Ross River virusesvigilax and also Cx sitiens, a potential vector in more from Aedes normanensis ( Diptera: Culicidae) innorthern areas ofcoastal eastern, western, and northern Western Australia. J. Med. Entomol. 26: 100- 103.Australia. Some of these localities( inland and coastal), Condon, R. 1991. Epidemiology and acutewhich are principal examples of areas with large and symptomatology of epidemic polyarthritis inincreased vector populations, are also localities that Western Australia, 1988- 89. Comm. Dis. Intell.have a perceived higher risk ofRR virus infection, based 15: 442-447.on annualized numbers of serologically ` confirmed' Fraser, J. R. E. 1986. Epidemic polyarthritis and Rosscases.
River Virus disease. Clin. Rheu. Dis. 12: 369- 88.Increases in vector populations that follow so- Hall, R. 1993. Annual report of the National Notifiable
called development projects, or simply increases in Diseases Surveillance System, 1992- Part 1. Comm.human contact with vectors that accompany the Dis. Intell. 17: 466-87.establishment of residential developments in areas with Hawkes, R. A., C. R. Boughton, H. M. Naim, and N.mosquito problems, will inherently increase the Stallman. 1985. A major outbreak of epidemicprobability of infection with RR virus in the many polyarthritis in New South Wales during the summerapparently endemic areas. An increase in local virus of 1983- 1984. Med. J. Aust. 143: 330-333.activity that occurs in natural cycles that are as yet Hawkes, R. A., J. Pamplin, C. R. Boughton, and H. M.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 81
Naim. 1993. Arbovirus infections of humans in virus and epidemic polyarthritis. Curr.Top. Vectorhigh- risk areas of southeastern Australia: a Res. 2: 31- 56.
continuing study. Med. J. Aust. 159: 159- 162. McManus, T. J. and I. D. Marshall. 1986. The
Hueston, L. and A. L. Cunningham. 1989. Epidemic epidemiology of Ross River virus in Tasmania.polyarthritis in NSW during the summer of 1988- Arbo. Res. Aust. 4: 127- 130.
89. Arbo. Res. Aust. 5: 389-90. McManus, T. J., R. C. Russell, P. J. Wells, J. G. Clancy,
Kay, B. H. 1982. Three modes of transmission of Ross M. Fennell, and M. J. Cloonan. 1992. Further
River virus by Aedes vigilax( Skuse). Aust. J. Exp. studies on the epidemiology and effects of RossBiol. Med. Sci. 60: 339- 344. River virus in Tasmania. Arbo. Res. Aust. 6: 68-72.
Kay, B. H. and J. G. Aaskov. 1988. Ross River virus Mudge, P. R. 1977. A survey of epidemic polyarthritisepidemic polyarthritis). Pp. 93- 112 in The in the Riverland Area, 1976. Med. J. Aust. 1: 649-
Arboviruses: Epidemiology and Ecology. Volume 651.
IV( T. P. Monath, ed.) CRC Press, 243 pp. Mudge, P. R. and J. G. Aaskov. 1983. Epidemic
Lindsay, M. D. A., A. K. Broom, A. E. Wright, C. A. polyarthritis in Australia, 1980- 1981. Med. J.
Johansen, and J. S. Mackenzie. 1993a. Ross River Aust. 2: 269- 273
virus isolations from mosquitoes in arid regions of Norris, P. 1993. An outbreak of Ross River virus dis-
Western Australia: implication of vertical trans- ease in Victoria. Comm. Dis. Intell. 17: 423- 424.
mission as a means of persistence of the virus. Am. Phillips, D. A., J. Sheridan, J. Aaskov, J. Murray, andM.
J. Trop. Med. Hyg. 49: 686-696. Also printed in Wiemers. 1992. Epidemiology of arbovirus
part as: Lindsay, M. et al." Isolation of Ross River infection in Queensland, 1989- 1992. Arbo. Res.
virus from field caught male Aedes mosquitoes" Aust. 6: 245-248.
Arbo. Info. Exch., January 1993: 29- 30. Russell, R. C. 1989. New South Wales mosquito and
Lindsay, M. D. A., R. J. Coelen, and J. S. Mackenzie. arbovirus surveillance: the programme after 5 years.
1993b. Genetic heterogeneity among isolates of Arbo. Res. Aust. 5: 188- 191.
Ross River virus from different geographical Russell, R. C. 1991. Arbovirus activity in southeastern
regions. J. Virol. 67: 3576-3585. Australia. Bull. Soc. Vector Ecol. 16: 222-226.
Lindsay, M., R. Condon, J. Mackenzie, C. Johansen, M. Russell, R. C. 1992. Arbovirus surveillance in New
D' Ercole, and D. Smith. 1992. A major outbreak South Wales. Comm. Dis. Intell. 15: 229-232.
of Ross River virus infection in the southwest of Russell, R. C. 1992. The surveillance of arbovirus
Western Australia and the Perth metropolitan area. activity in N.S. W. 1989- 1992. Arbo. Res. Aust. 6:Comm. Dis. Intell. 16: 290-294. 76-80.
Lindsay, M. D., J. A. Latchford, A. E. Wright, and J. S. Tai, K. S., P. I.Whelan, M. S. Patel, and B. Currie. 1993.
Mackenzie. 1989. Studies on the ecology of Ross An outbreak of epidemic polyarthritis( Ross River
River virus in the south-west of Western Australia. virus disease) in the Northern Territory during the
Arbo. Res. Aust. 5: 28- 32. 1990- 1991 wet season. Med. J. Aust. 158: 522-
Mackenzie, J. S., A. K. Broom, C. H. Calisher, M. J. 525.
Cloonan, A.L. Cunningham, C. Gibson,L.Hueston, Vale, T. G., M. L. Dowling, and M. J. Cloonan. 1992.M. D. Lindsay, I. D. Marshall, D. A. Phillips, R. C. Infection and multiplication of Ross River virus in
Russell, J. Sheridan, D. W. Smith, T. Vitarana, and the mosquito vector Aedes vigilax( Skuse). Aust. J.
D. Worswick. 1993. Diagnosis and reporting of Zool. 40: 35- 41.
arbovirus infections in Australia. Comm. Dis. Wolstenholme, J. 1991. Ross River virus disease in
Intell. 17: 202-06. Victoria during the summers of 1988- 89 and 1989Marshall, I. D. and J. A. R. Miles. 1984. Ross River 90. Comm. Dis. Intell. 15: 137- 139.

BULL. SOC. VECTOR ECOL., 19( 1): 82- 86 JUNE, 1994
SOME SANDFLY FOOD IS A LEISHMANIA POISON'
Y. Schlein2 and R. L. Jacobson2
ABSTRACT: The studies summarized suggest that some of the diverse diets ofsandfly vectors can adversely affectthe Leishmania parasites in their gut. Phlebotomuspapatasi in the vicinity of turkey sheds were free ofLeishmaniamajor parasites, in contrast to the commonly high infection rate elsewhere in the endemic region. Experimentsdemonstrated that sandfly meals of turkey or chicken blood are lethal to L. major infections. Furthermore, such mealsreduce the potential of uninfected flies to harbor L major. Plant feeding of sandflies in nature was demonstratedby identification ofplant-tissue residues in the gut. Experimental feeding ofLeishmania-infected flies on some plantspecies caused parasite death in a high proportion of flies. The mortality of Leishmania parasites found in fieldcollected flies could therefore result from the sandflies' plant diets. Bacterial gut contaminations are harmful tosandflies and their Leishmania parasites. Their prevention is attributed to an antibacterial agent in the sandfly crop,which is a temporary store ofcontaminated sugary food, ingested from the surface ofplants. Bacterial gut infectionsofsandflies are rare in the arid Jordan Valley where contaminating food is scarce and they are probably more commonin temperate or tropical climates.
INTRODUCTION selectively on plants and honeydews in the laboratorySchlein and Warburg 1986), while Ph. ariasi has been
The cycle of Leishmania parasites in the sandfly fed on honeydew ( Killick-Kendrick and Killick-vector is limited to the gut, and the nutrients of the Kendrick 1987). The degree of feeding on honeydew insandfly provide the medium for their development. the wild was evaluated for Ph. ariasi ( Moore et al.Therefore, variations in the diet, either the sporadic 1987), Ph. perfiliewi ( MacVicker et al. 1990), andblood meals or the regular sugar meals, can influence Lutzomyia peruensis( Wallbankseta1. 1991) by detectionthe development of Leishmania infections, ofmelezitose in the gut. The ingestion ofplant tissue in
Sources of sandfly blood meals were identified for the field by Ph.papatasi was determined byobservationsamples of several vector species ( Lloyd and Napier ofcellulosic cell residues in the gut( Schlein and Jacobson1930, Vives-Sabatier 1954, George 1970, Javadian et 1994). Indirect evidence for the plant preferences ofPh.al. 1977, and other references quoted in Morrison et al. papatasi has been demonstrated by their differential1993) and the relative attraction to hosts was studied by attraction to plants in the field ( Schlein and Yuvalthe use of bait animals( Christensen and Herrer 1980; 1987).Mutinga et al. 1981, 1986). In a recent study Morrison Several of our studies were on the adverse effectset al. ( 1993) considered both the degree of feeding on on the Leishmania infections that can result from thehosts and host abundance. The pattern that emerged, sandfly vector diet. The model was L. major and Ph.which appears to be general, is a combination of papatasi from the endemic focus of cutaneousopportunistic and selective feeding. This behavior is leishmaniasis in the Jordan Valley. The sand ratpresumably dictated by the normally short range of Psammomys obesus is the rodent reservoir and theirsandfly flight that limits the choice of hosts. burrows are the main breeding and resting sites of the
It can also be assumed that a similar behavior sandflies.
guides sandflies in the intake of sugar meals. However,the specific sources ofnatural sugar meals are unknown. Effects of Avian Blood MealsSandflies were seen probing the leaves and stems of During a study in the Jordan Valley we found a highplants( Minter and Wijers 1963, Ashford 1974). Later it concentration of Ph. papatasi in the burrows of Ps.was demonstrated that Phlebotomus papatasi feeds obesus near turkey sheds. Examination of several
1 Paper Presented at the 1st International Congress of Vector Ecology, San Diego, California, USA, 7 October 1993.2Department of Parasitology, The Hebrew University- Hadassah Medical School, POB 1172, Jerusalem, ISRAEL.

JUNE, 1994 BULL. SOC. VECTOR ECOL. 83
hundreds of females ( 544) showed that they were A phenomenon similar to our initial field observationuninfected ( unpublished data). This was exceptional emerged from an epidemiological study of Rojas et al.since the infection rate of female flies from burrows in ( 1988) in Acosta County, Costa Rica. They correlatedother sites ranged from 20 percent to 50 percent in the the incidence of cutaneous leishmaniasis, caused by L.same season. The sandflies were attracted to the turkeys b. panamensis, with the housing environment andand they were also the source of most blood meals concluded that poultry in the household have the highestSchlein et al. 1982 a, 1982b). This by itself could have protective value against the disease.
been the reason for the lack of infection. However, our It is as yet unknown how the avian blood affects theexperiments showed that turkey blood is lethal to L. L. major and whether it has a similar influence on othermajor. Flies caught near the turkey sheds were Leishmania spp. Furthermore, circumstantial evidencemaintained for one week to allow for the digestion of indicates that the presence of cattle can reduce humanblood meals and then fed parasites in rabbitblood. After infections in leishmaniasis foci and this led to thesix days parasites were found in only 17. 5 percent of suggested use of cattle as a zooprophylaxis ( Addy etthese flies( TABLE 1). Similarly, only 25 percent of the al. 1983, Lainson 1989, Gosh et al. 1990, Morrison et al.laboratory reared flies that had been fed on turkey blood 1993). It would also be interesting to see in controlledand infected one week later could support the conditions whether peridomestic fowl have a similardevelopmentof the parasites. In addition, flies artificially effect.
infected with rabbit blood and parasites and givenmeals of turkey blood after 24 hrs. resulted in only 16.4 Effects of Plant Diet
percent infected flies three days later. These results In the laboratory sandflies feed on different plantswere significantly lower than the 85. 5 percent infections and honeydews in an order of preference, which isin laboratory- reared control flies that had received indicated by the rate of feeding on each diet( Schlein andparasites in rabbit blood six days earlier( Schlein et al. Warburg 1986, Schlein and Yuval 1987). Plants that1983). In another experiment no L. major parasites ranked high in the feeding preferences of Ph. papatasisurvived in the 40 flies that were fed on chicken blood were used in experiments that evaluated the effect of24 hrs. post- infection and examined three days later sugar meals on the L. major parasites in the sandfliesTABLE 1). It is noteworthy that in addition to their ( Schlein and Jacobson 1994). Only one experiment was
immediate effect, the meals of turkey blood also cause carried out with honeydew since these aphid secretionsa lasting reduction of the vector potential to harbor L. consist mainly of plant sap and amino acids ( Auclair,major. 1963; Brown, 1975), while many plants contain lectins
TABLE 1. Survival of Leishmania major promastigotes in Phlebotomus papatasi fed on turkey or chicken blood.The flies were examined six days after meals of inactivated rabbit blood with 2 X 106 promastigotes perml.
Source of flies Treatment of flies Total number No. Parasitized(%)
Turkey sheds maintained one week 63 11 ( 17. 5%)
pre- infection
Laboratory reared fed turkey blood one week 48 12 ( 25%)pre- infection
Laboratory reared fed turkey blood 24 hrs 55 9 ( 16.4%)
post- infection
Laboratory reared fed chicken blood 24 hrs 40 0 ( 0%)post- infection*
Laboratory reared infected controls 138 118 ( 85. 5%)
examined four days post- infection

84 BULL. SOC. VECTOR ECOL. JUNE, 1994
Etzler 1986) that can be harmful to Leishmania in ( 23/ 47) of the females and 10. 3 percent( 6/52) of the
cultures ( Davidowicz et al. 1975, Jacobson et al. 1982). males.
Phlebotomuspapatasi infected with L. major were fed Therefore, it appears that mortality ofL. major inon one diet for six days, following which the success of Ph. papatasi in the field could result from the plant diet
infections was evaluated by estimating the number of of the sandflies. However, specific identification of the
parasites, the degree of mortality, morphological food plants and further study of their effects on theaberrations, and cell agglutination. Infections with less harbored parasites are required for evaluation of the
than 1, 000 parasites were defined as light ( heavy role of sugarmeals in the epidemiology ofleishmaniasis.
infections are up to 1/ 2 million parasites) and those with50 percent or more disfigured and dead parasites were Bacterial Contamination of Food
considered as impaired. Feeding on the plant Malva The gut of sandflies is normally free of micro-nicaeensis and on the honeydew Icerya purchasi organisms as was observed by early investigators( Adlerproduced vital and thriving infections. Whereas, plant and Theodor 1927). The system by which sandfliesdiets of either Ricinus communis, Capparis spinosa, or evade bacterial infections is apparently as follows: TheSolanum luteum reduced the number of parasites and destination of food in the gut regions is determined byimpaired the infections in 88 percent, 55 percent, and 46 the manner of feeding. The food is indiscriminatelypercent of the flies, respectively ( TABLE 2). These entrained into the midgut when the sandfly stylets areobservations were followed by examination of field dipped through a membrane( Ready 1978), into animalcollected flies infected with L. major to see whether the tissue or a plant( Schlein and Warburg 1986). Sugars
same phenomenon occurs in nature. Parasitaemias in 94 from the surface are directed to the crop and graduallyflies were examined and 21 of them were defined as released to the midgut ( Schlein and Warburg 1985).impaired according to the above criteria. Bacterial contaminations are rare in sandfly meals of
The last stage of this study was the demonstration either blood or plants, that are taken from internal
that sandflies in the wild feed on plants and that this diet tissues while they are apparently prevalent on the plantcould be the cause of parasite mortality. We assumed surface, in the sugary honeydew, and nectar.that sandflies, while feeding on plants, also ingest The evidence that bacteria are only introduced tocellulosic cell-wall fragments. Therefore, we used the the crop suggested that this organ might contain anfluorescent dye calcofluor, which is employed for antibacterial agent. This assumption was tested bydifferential staining ofcellulose( Haigler 1991) to detect placing full crops of Ph. papatasi on agar platessuch fragments. This staining enabled the identi- inoculated with different bacterial species( Escherichia
fication of plant residues in the gut-lumen of flies that coli, Staphylococcus aureus, Shigella sonnei, Strepto-
had been fed on plants in the laboratory. The presence coccus group A, and Pseudomonas aeruginosa). The
of such residues is apparently indicative of direct presence of a bacteriostatic or bacteriocidal agent was
feeding from plants since they were not observed in the demonstrated by the clear halo that remained around theguts of either honeydew fed or unfed flies. Fly samples crops for 6 to 10 hours( Schlein et al. 1985).
that had been caught in the Jordan Valley were then This system does not afford full protection and
examined and plant residues were found in 32.8 percent bacterial infections can be acquired by feeding from
TABLE 2. Effects of plant feeding of Phlebotomus papatasi on Leishmania major in the sandfly gut.Flies were infected with meals of 104 promastigotes/ml in inactivated rabbit blood and were
examined six days later. Impaired infections are defined as containing> 50 percent dead ordeformed parasites.
Sandfly diet Flies examined Flies infected Impaired infections
Ricinus communis 62 50 44( 88%)
Capparis spinosa 33 20 11 ( 55%)
Solanum luteum 25 25 11 ( 46%)
Malva nicaeensis 31 29 0
honeydew of Icerya purchasi 52 36 0
20% sucrose solution( control) 38 32 0

JUNE, 1994 BULL. SOC. VECTOR ECOL. 85
contaminated fruits ( Killick-Kendrick 1979) or when Etzler, M. E. 1986. Distribution and function of plant
flies are artificially infected with Leishmania. In the lectins Pp. 371- 435 in The Lectins: Propertieslaboratory, and apparently in the field,bacterial infections Functions and Applications in Biology and Medicine
kill the Leishmania parasites in infected flies and cause I. E. Liener, N. Sharon, and I. J. Goldstein, eds.).
fly mortality. We have recorded four percent bacterial Academic Press, Orlando, 600 pp.
infections while examining theL. major parasites found George, G. E. 1970. Isolation of Phlebotomus fever
in Ph. papatasi from the arid Jordan Valley. Sources of virus fromPhlebotomus papatasi and determination
contamination such as honeydew, nectar, and rotting of the host ranges of sandflies ( Diptera:
fruits are more common in humid biotopes and the rate Psychodidae) in West Pakistan. J. Med. Entomol.
of bacterial infections of sandflies is higher( Schlein et 7: 670-676.
al. 1985). Such infections deserve attention since they Gosh, K. N., A. Bhattacharya, and T. N. Gosh. 1990.
may, if prevalent, decrease the transmission of Blood meal analysis of Phlebotomus argentipes in
leishmaniasis. eight districts ofWest Bengal. J. Commun. Dis. 22:
The antibacterial agent of sandflies has not been 67- 71.
studied and as far as weknow there are also no additional Haigler, C. H. 1991. Relationship between
data on the identity or prevalence of bacterial infections polymerization and crystallization in microfibrils
of sandflies. biogenesis. Pp. 99- 125 in Biosynthesis and Bio-The observations presented in this paper show that degradation of Cellulose( C. H. Haigler and P. J.
different sandfly diets can affect the transmission of Weimer, eds.). Marcel Dekker, New York, 694 pp.
leishmaniasis. Further studies can determine the Jacobson, R. L., G. M. Slutzky, C. L. Greenblatt, and L.magnitude of their effects and may lead to the F. Schnur. 1982. Surface reaction of Leishmania
development of biological methods of control. I. Lectin mediated agglutination. Ann. Trop. Med.Parasitol. 76: 45- 52.
Acknowledgments Javadian, E., R. Tesh, S. Saidi, and A. Nadim. 1977.
Studies on the epidemiology of sandfly fever inThese studies received support from NIAID-TMP- Iran III. Host-feeding patterns of Phlebotomus
AI-32076 and UNDP/World Bank/WHO Special papatasi in an endemic area of disease. Am. J.
Program for Research and Training. Trop. Med. Hyg. 26: 294-298.Killick-Kendrick, R. 1979. Biology of Leishmania in
REFERENCES CITED phlebotomine sandflies. Pp. 395-449 in Biology ofKinetoplastida Vol. 2( W. H. R Lumsden and D. A.
Addy, M., A. K. Mitra, K. K. Gosh, and A. K. Hatti. Evans, eds.). Academic Press, London, 738 pp.
1983. Host preferences ofPhlebotomus argentipes Killick-Kendrick, R. and M. Killick-Kendrick. 1987.
in different biotopes. Trop. Geogr. Med. 35: 343- Honeydew of aphids as a source of sugar for
345. Phlebotomus ariasi. Med. Vet. Entomol. 1: 297-
Adler, S. and O. Theodor. 1927. The transmission of 302.
Leishmania tropica from artificially infected sand- Lainson, R. 1989. Demographic changes and their
flies to man. Ann. Trop. Med. Parasitol. 23: 1- 18. influences on American leishmaniases. Pp. 85- 106Ashford, R.W. 1974. Sandflies( Diptera:Phlebotomidae) in Demography and Vector Borne Diseases( M. W.
from Ethiopia: taxonomic and biological notes. J. Service, ed.). CRC Press, Boca Raton FL, 402 pp.
Med. Entomol. 11: 605- 616. Lloyd, R. B. and L. E. Napier. 1930. The blood meal of
Auclair, J. L. 1963. Aphid feeding and nutrition. Ann. sandflies investigated by means of precipitinRev. Entomol. 8: 464-471. antisera. Indian J. Med. Res. 18: 347- 359.
Brown, K. S. 1975. The chemistry of aphids and scale MacVicker, J. A. K., J. S. Moore, D. H. Molyneux, and
insects. Chem. Soc. Rev. 4: 263- 288. M. Maroli. 1990. Honeydew sugars in wild caught
Christensen, H. and A. Herrer. 1980. Panamanian Italian phlebotomine sandflies ( Diptera: Psycho-
Lutzomyia( Diptera: Psychodidae) host attraction didae) as detected by high performance liquidprofiles. J. Med. Entomol. 17: 522- 528. chromatography. Bull.Entomol. Res. 80: 339- 344.
Davidowicz, R., A. G. Hernandez, R. B. Infante, and R. Minter, D. M. and D. J. B. Wijers. 1963. Studies on the
B. Convit. 1975. The surface membrane of vector of Kala-azar in Kenya IV. Experimental
Leishmania I. The effects of lectins on different evidence. Ann. Trop. Med. Parasitol. 57: 24- 31.stages of Leishmania braziliensis. J. Parasitol. 76: Moore, J. S., T. B. Kelly, R. Killick-Kendrick, M.950-953. Killick-Kendrick, K. R. Wallbanks, and D. H.

86 BULL. SOC. VECTOR ECOL. JUNE, 1994
Molyneux. 1987. Honeydew sugars in wild caught Hyg. 50: 20-27.Phlebotomus ariasi detected by high performance Schlein, Y., I.Polachek, and B. Yuval. 1985. Mycoses,liquid chromatography ( HPLC) and gas chroma- bacterial infections and antibacterial activity intography. Med. Vet. Entomol. 1: 427-434. sandflies and their possible role in the transmission
Morrison, A. C., C. Ferro, and R. B. Tesh. 1993. Host of leishmaniasis. Parasitol. 90: 57- 66.preferences of the sandfly Lutzomyia longipalpis at Schlein, Y. and A. Warburg. 1985. Feeding behavior,an endemic focus of American visceral leish- midgut distention and ovarian development inmaniasis in Colombia. Am. J. Trop. Med. Hyg. 49: Phlebotomus papatasi ( Diptera: Psychodidae). J.68- 75. Insect. Physiol. 31: 47- 51.
Mutinga, M. J., J. B. Kaddu, and L. W. Irungo. 1981. Schlein, Y. and A. Warburg. 1986. Phytophagy and theAnimal models for feeding Kenyan wild caught feeding cycle of Phlebotomus papatasi ( Diptera:phlebotomine sandflies ( Diptera:Phlebotomidae). Psychodidae) under experimental conditions. J.Insect Sci. Applic. 2: 149- 152. Med. Entomol. 23: 11- 15.
Mutinga, M. J., F. M. Kyai, C. Kamau, and D. M. Schlein, Y., A. Warburg, L. F. Schnur, and A. E.Omogo. 1986. Epidemiology of leishmaniasis in Gunders. 1982b. Leishmaniasis in the JordanKenya- III. Host preference studies using various Valley II. Sandflies and transmission in the centraltypes ofanimal baits atanimal burrows in Marigat, endemic area. Trans. Roy. Soc. Trop. Med. Hyg.Baringo District. Insect Sci. Applic. 7: 191- 197. 76: 582- 586.
Ready, P. D. 1978. The feeding habits of laboratory Schlein, Y., A. Warburg, L. F. Schnur, and J. Shlomai.bredLutzomyialongipalpis( Diptera: Psychodidae). 1983. Vector compatibility of PhlebotomusJ. Med. Entomol. 14: 545- 552. papatasi dependent on differentially induced
Rojas, J. C., R. Zeledon, J. Murillo, and A. Urbina. digestion. Acta Trop. 40: 65- 70.1988. Identification of the risk factors associated Schlein, Y. and B. Yuval. 1987. Leishmaniasis in thewith cutaneous leishmaniasis in Costa Rica. Proc. Jordan Valley IV. Attraction of PhlebotomusIDRC Int. Workshop, on the control strategies for papatasi ( Diptera: Psychodidae) to plants in theleishmaniases, Ottawa 1987: 244-249. field. J. Med. Entomol. 24: 87-90.
Schlein, Y., A. E. Gunders, and A. Warburg. 1982a. Vives-Sabatier, J. 1954. Biologia des las especies deLeishmaniasis in the Jordan valley I. Attraction of Phlebotomushallados en la Provincia de Barcelona.Phlebotomus papatasi ( Psychodidae) to turkeys. Revista Iberica Parasitol. 14: 397- 407.Ann. Trop. Med. Parasitol. 76: 517- 520. Wallbanks, K. R., J. S. Moore, L. R. Bennet, R. Soren,
Schlein, Y. and R. L. Jacobson. 1994. Mortality of D. H. Molyneux, J. M. Carlin, and J. E. Perez.Leishmania major in Phlebotomus papatasi caused 1991. Aphidderived sugars in the sandflyLutzomyiaby plant feeding of the sandflies. Am. J. Trop. Med. peruensis. Trop. Med. Parasitol. 42: 60-62.

ADVERTISING
Commercial advertising space is available in the Bulletin;full page ( black and white) at $ 150.00 per issue, half page
black and white) at $ 90.00 per issue. Inquiries may beaddressed to H. B. Munns, Advertising Manager, SOVE,P. O. Box 338, Pioneer, California 95666, telephone209) 295- 3540 or Fax ( 209) 295- 3563.
The publisher reserves the right to approve or refuse anyadvertisement. The publisher is not responsible for anyclaims, litigations, or expenses resulting from theadvertiser' s unauthorized use of any name, photograph,
sketch, or words protected by registered trademark orcopyright.


















