
The Phosphoinositide Phosphatase Sjl2 Is Recruited to Cortical Actin Patches in the Control of
14
MOLECULAR AND CELLULAR BIOLOGY, Apr. 2005, p. 2910–2923 Vol. 25, No. 8 0270-7306/05/$08.000 doi:10.1128/MCB.25.8.2910–2923.2005 Copyright © 2005, American Society for Microbiology. All Rights Reserved. The Phosphoinositide Phosphatase Sjl2 Is Recruited to Cortical Actin Patches in the Control of Vesicle Formation and Fission during Endocytosis† Christopher J. Stefan, Steven M. Padilla,‡ Anjon Audhya,§ and Scott D. Emr* Department of Cellular and Molecular Medicine, The Howard Hughes Medical Institute, University of California, San Diego, School of Medicine, La Jolla, California Received 12 June 2004/Returned for modification 26 July 2004/Accepted 18 January 2005 The Saccharomyces cerevisiae synaptojanin-like proteins (Sjl1, Sjl2, and Sjl3) are phosphoinositide (PI) phosphatases that regulate PI metabolism in the control of actin organization and membrane trafficking. However, the primary sites of action for each of the yeast synaptojanin-like proteins remain unclear. In this study, we show that Sjl2 is localized to cortical actin patches, sites of endocytosis. Cortical recruitment of Sjl2 requires the actin patch component Abp1. Consistent with this, the SH3 domain-containing protein Abp1 physically associates with Sjl2 through its proline-rich domain. Furthermore, abp1 mutations confer defects resembling loss of SJL2; sjl1 abp1 double-mutant cells exhibit invaginated plasma membranes and impaired endocytosis, findings similar to those for sjl1 sjl2 mutant cells. Thus, Abp1 acts as an adaptor protein in the localization or concentration of Sjl2 during late stages of endocytic vesicle formation. Overexpression of the Hip1-related protein Sla2 delayed the formation of extended plasma membrane invaginations in sjl2 ts cells, indicating that Sla2 may become limiting or misregulated in cells with impaired PI phosphatase activity. Consistent with this, the cortical actin patch protein Sla2 is mislocalized in sjl1 sjl2 mutant cells. Together, our studies suggest that PI metabolism by the synaptojanin-like proteins coordinately directs actin dynamics and membrane invagination, in part by regulation of Sla2. Phosphoinositides (PI) control numerous cellular processes including cell signaling, growth, vesicular trafficking, and actin cytoskeletal arrangements (8, 43, 50, 54). In particular, phos- phatidylinositol(4,5)-bisphosphate [PI(4,5)P 2 ] provides impor- tant regulatory roles at the cell surface, as a precursor of the signaling molecules DAG and IP 3 , and through direct interac- tions with downstream effector proteins (31, 34). During en- docytic internalization, PI(4,5)P 2 recruits or activates several proteins, including the clathrin adaptor complex AP-2, AP180, epsin, and the GTPase dynamin, leading to the formation and fission of clathrin-coated vesicles (13, 15, 16, 25, 40). PI(4,5)P 2 also recruits or controls several actin-regulatory proteins (54). Actin polymerization may provide a propulsive force to facil- itate membrane invagination and fission events or to transport newly formed endocytic vesicles away from the plasma mem- brane. Thus, PI(4,5)P 2 synthesis plays several important roles in the formation and fission of clathrin-coated vesicles during endocytosis. Likewise, previous studies implicate dephosphorylation of PI(4,5)P 2 by the polyphosphoinositide phosphatase (PPIPase) synaptojanin as an important step in endocytic internalization (7, 22, 27, 41, 42, 51). Synaptojanins contain two distinct PI phosphatase domains and a C-terminal proline-rich domain (PRD). The N-terminal Sac1 domain (PPIPase domain) acts on several positions of the inositol ring, while its central phos- phatase domain specifically acts at the 5 position (5-Pase do- main) (20, 37). The PRD region functions as a targeting do- main via interactions with various SH3 domain-containing proteins, such as endophilin, amphiphysin, and CIN85 (5, 28, 42, 45, 51). Coated vesicles accumulate in cells bearing genetic disruptions of synaptojanin isoforms in the mouse, Drosophila melanogaster, and Caenorhabditis elegans, suggesting that turn- over of PI(4,5)P 2 mediates release of coat proteins from mem- branes of newly formed vesicles (7, 22, 51). Since synaptojanins regulate PI(4,5)P 2 -mediated actin polymerization, they may also control earlier vesicle formation and/or fission events, along with their role in postfission vesicle uncoating. Three synaptojanin-like proteins are present in Saccharomy- ces cerevisiae: Sjl1, Sjl2, and Sjl3 (also named Inp51, Inp52, and Inp53) (46, 49). Like mammalian synaptojanin, Sjl2 and Sjl3 exhibit Sac1 PPIPase activities, and all three possess 5-Pase activity (Fig. 1) (20). Factors that target the yeast synaptojanin- like proteins via their PRDs have yet to be demonstrated in vivo. Interestingly, the C-terminal domains of Sjl1, Sjl2, and Sjl3 do not share extensive identity (Fig. 1). Thus, while syn- aptojanins display similar activities in vitro, each may provide specialized functions in vivo and may be uniquely recruited or activated at distinct membrane sites. Previous studies have indicated that Sjl2 provides essential overlapping functions at both the plasma membrane and in- tracellular endosomal/late Golgi compartments (17, 44, 46, 48, 49). Moreover, we have shown that yeast cells expressing only a temperature-conditional SJL2 allele inappropriately accumu- * Corresponding author. Mailing address: Department of Cellular and Molecular Medicine, The Howard Hughes Medical Institute, Uni- versity of California, San Diego, School of Medicine, La Jolla, CA 92093-0068. Phone: (858) 534-6462. Fax: (858) 534-6414. E-mail: [email protected]. † Supplemental material for this article may be found at http: //mcb.asm.org/. ‡ Present address: Veterans Medical Research Foundation, San Di- ego, CA 92161. § Present address: Ludwig Institute for Cancer Research, La Jolla, CA 92093. 2910 Downloaded from https://journals.asm.org/journal/mcb on 16 November 2021 by 39.124.117.21.
Transcript of The Phosphoinositide Phosphatase Sjl2 Is Recruited to Cortical Actin Patches in the Control of

MOLECULAR AND CELLULAR BIOLOGY, Apr. 2005, p. 2910–2923 Vol. 25, No. 80270-7306/05/$08.00�0 doi:10.1128/MCB.25.8.2910–2923.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
The Phosphoinositide Phosphatase Sjl2 Is Recruited to Cortical ActinPatches in the Control of Vesicle Formation and Fission
during Endocytosis†Christopher J. Stefan, Steven M. Padilla,‡ Anjon Audhya,§ and Scott D. Emr*
Department of Cellular and Molecular Medicine, The Howard Hughes Medical Institute, University of California,San Diego, School of Medicine, La Jolla, California
Received 12 June 2004/Returned for modification 26 July 2004/Accepted 18 January 2005
The Saccharomyces cerevisiae synaptojanin-like proteins (Sjl1, Sjl2, and Sjl3) are phosphoinositide (PI)phosphatases that regulate PI metabolism in the control of actin organization and membrane trafficking.However, the primary sites of action for each of the yeast synaptojanin-like proteins remain unclear. In thisstudy, we show that Sjl2 is localized to cortical actin patches, sites of endocytosis. Cortical recruitment of Sjl2requires the actin patch component Abp1. Consistent with this, the SH3 domain-containing protein Abp1physically associates with Sjl2 through its proline-rich domain. Furthermore, abp1� mutations confer defectsresembling loss of SJL2; sjl1� abp1� double-mutant cells exhibit invaginated plasma membranes and impairedendocytosis, findings similar to those for sjl1� sjl2� mutant cells. Thus, Abp1 acts as an adaptor protein in thelocalization or concentration of Sjl2 during late stages of endocytic vesicle formation. Overexpression of theHip1-related protein Sla2 delayed the formation of extended plasma membrane invaginations in sjl2ts cells,indicating that Sla2 may become limiting or misregulated in cells with impaired PI phosphatase activity.Consistent with this, the cortical actin patch protein Sla2 is mislocalized in sjl1� sjl2� mutant cells. Together,our studies suggest that PI metabolism by the synaptojanin-like proteins coordinately directs actin dynamicsand membrane invagination, in part by regulation of Sla2.
Phosphoinositides (PI) control numerous cellular processesincluding cell signaling, growth, vesicular trafficking, and actincytoskeletal arrangements (8, 43, 50, 54). In particular, phos-phatidylinositol(4,5)-bisphosphate [PI(4,5)P2] provides impor-tant regulatory roles at the cell surface, as a precursor of thesignaling molecules DAG and IP3, and through direct interac-tions with downstream effector proteins (31, 34). During en-docytic internalization, PI(4,5)P2 recruits or activates severalproteins, including the clathrin adaptor complex AP-2, AP180,epsin, and the GTPase dynamin, leading to the formation andfission of clathrin-coated vesicles (13, 15, 16, 25, 40). PI(4,5)P2
also recruits or controls several actin-regulatory proteins (54).Actin polymerization may provide a propulsive force to facil-itate membrane invagination and fission events or to transportnewly formed endocytic vesicles away from the plasma mem-brane. Thus, PI(4,5)P2 synthesis plays several important rolesin the formation and fission of clathrin-coated vesicles duringendocytosis.
Likewise, previous studies implicate dephosphorylation ofPI(4,5)P2 by the polyphosphoinositide phosphatase (PPIPase)synaptojanin as an important step in endocytic internalization
(7, 22, 27, 41, 42, 51). Synaptojanins contain two distinct PIphosphatase domains and a C-terminal proline-rich domain(PRD). The N-terminal Sac1 domain (PPIPase domain) actson several positions of the inositol ring, while its central phos-phatase domain specifically acts at the 5� position (5-Pase do-main) (20, 37). The PRD region functions as a targeting do-main via interactions with various SH3 domain-containingproteins, such as endophilin, amphiphysin, and CIN85 (5, 28,42, 45, 51). Coated vesicles accumulate in cells bearing geneticdisruptions of synaptojanin isoforms in the mouse, Drosophilamelanogaster, and Caenorhabditis elegans, suggesting that turn-over of PI(4,5)P2 mediates release of coat proteins from mem-branes of newly formed vesicles (7, 22, 51). Since synaptojaninsregulate PI(4,5)P2-mediated actin polymerization, they mayalso control earlier vesicle formation and/or fission events,along with their role in postfission vesicle uncoating.
Three synaptojanin-like proteins are present in Saccharomy-ces cerevisiae: Sjl1, Sjl2, and Sjl3 (also named Inp51, Inp52, andInp53) (46, 49). Like mammalian synaptojanin, Sjl2 and Sjl3exhibit Sac1 PPIPase activities, and all three possess 5-Paseactivity (Fig. 1) (20). Factors that target the yeast synaptojanin-like proteins via their PRDs have yet to be demonstrated invivo. Interestingly, the C-terminal domains of Sjl1, Sjl2, andSjl3 do not share extensive identity (Fig. 1). Thus, while syn-aptojanins display similar activities in vitro, each may providespecialized functions in vivo and may be uniquely recruited oractivated at distinct membrane sites.
Previous studies have indicated that Sjl2 provides essentialoverlapping functions at both the plasma membrane and in-tracellular endosomal/late Golgi compartments (17, 44, 46, 48,49). Moreover, we have shown that yeast cells expressing onlya temperature-conditional SJL2 allele inappropriately accumu-
* Corresponding author. Mailing address: Department of Cellularand Molecular Medicine, The Howard Hughes Medical Institute, Uni-versity of California, San Diego, School of Medicine, La Jolla, CA92093-0068. Phone: (858) 534-6462. Fax: (858) 534-6414. E-mail:[email protected].
† Supplemental material for this article may be found at http://mcb.asm.org/.
‡ Present address: Veterans Medical Research Foundation, San Di-ego, CA 92161.
§ Present address: Ludwig Institute for Cancer Research, La Jolla,CA 92093.
2910
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

late PI(4,5)P2 in intracellular compartments (48). These find-ings have thus led to the question of whether Sjl2 acts primarilyat the plasma membrane or at endosomes in the control ofPI(4,5)P2 steady-state distribution. To address this question,we have examined the localization of Sjl2 in vivo. Sjl2 wasrecruited to cortical actin patches in a manner dependent onfilamentous actin and the PRD of Sjl2. We further addressedhow Sjl2 is recruited to its site of action at the plasma mem-brane and found that the SH3-containing protein Abp1 wasessential for this process. Finally, we found that overexpressionof the ANTH domain-containing protein Sla2 can partiallysuppress the endocytic defects in sjl1� sjl2ts sjl3� mutant cells,suggesting that Sla2 becomes a limiting factor during endocyticinternalization in cells with impaired PI 5-Pase activity.
MATERIALS AND METHODS
Yeast strains and plasmid constructions. Standard yeast genetic methods tocreate integrated deletions, truncations, and fusions were used throughout (33).Saccharomyces cerevisiae strains used in this study are listed in Table 1. Primersused in this study are available upon request.
To generate the Sjl2�37C-green fluorescent protein (GFP) fusion used in thesestudies, the 0.6-kb EcoRI fragment (encoding the 39 C-terminal residues of Sjl2)was removed from pRS416SJL2 (48) to create pRS416SJL2�RI, which then hada unique EcoRI site. PCR was then used to amplify GFP(S65T)-encoding se-quences. The resulting product was digested with EcoRI and subcloned in frameand in the appropriate orientation into pRS416SJL2�RI, which was cleaved withEcoRI to create pRS416SJL2�37C-GFP. To generate the Sjl2�PRD-GFP fusion,the 3.7-kb SalI-XhoI fragment from pRS416SJL2 was subcloned into pRS416cleaved with XhoI to create pRS416SJL2�XI, which then had a unique XhoI siteand thus excluded 190 C-terminal residues encoding the PRD of Sjl2. PCR wasthen used to amplify GFP(S65T)-encoding sequences. The resulting product wasdigested with XhoI-EcoRI and subcloned in frame into pRS416SJL2�XI, whichwas cleaved with XhoI-EcoRI. The K1120V and P1121D mutations were intro-duced into pRS416SJL2-GFP by QuikChange mutagenesis (Stratagene, La Jolla,Calif.). In order to overexpress SLA2 from a multicopy plasmid, a 4.3-kb EcoRIfragment encompassing the SLA2 locus (53) was subcloned into pRS426, whichhad been cleaved with EcoRI.
Generation of Sjl2-specific antisera, immunofluorescence, and microscopy onintact cells. A 0.7-kb HindIII fragment of SJL2 encompassing codons 893 to 1129was subcloned into pGEX-KG, and the resulting glutathione S-transferase fusionprotein was inducibly expressed in Escherichia coli, purified from bacterial ex-tracts, and then used to immunize New Zealand White rabbits as describedpreviously (2). Resulting antibodies were further purified by acetone extracts ofsjl2� cells. Immunofluorescence microscopy procedures were carried out essen-tially as described previously (2, 17), except that cells were costained with rho-damine phalloidin according to the manufacturer’s recommendations (Molecu-lar Probes, Eugene, Oreg.).
To monitor endocytosis, cells were labeled with FM4-64 (Molecular Probes) asdescribed elsewhere (48). Briefly, cells were grown to early-log phase in yeastextract-peptone-dextrose (YPD) and shifted to the appropriate temperature for90 min. Cells (2 OD600 [optical density at 600 nm] units) were harvested bycentrifugation and labeled with 16 nM FM4-64 and 100 nM CMAC (MolecularProbes) in YPD prewarmed to the appropriate temperature, followed by a chasein YPD without the vital dyes at the appropriate temperature for 30 min. Cellswere concentrated and visualized by fluorescence microscopy.
Live cells expressing Sjl2�37C-GFP, Sjl2�PRD-GFP, Sjl2K1120V,P1121D,�37C-GFP, or Sla2-GFP were grown to mid-log phase and harvested for visualization.
FIG. 1. Yeast synaptojanin-like proteins possess unique C-terminaldomains. The Sac1 domains in Sjl1p, Sjl2p, and Sjl3p are shown byopen ovals. The asterisks in the Sac1-like domain of Sjl1 indicate thatit does not possess PPIPase activity. Dark gray bars, 5-Pase domains inSjl1, Sjl2, and Sjl3. Relevant PI metabolism activities are shown forthese PI phosphatase domains. The light gray boxes between Sjl1, Sjl2,and Sjl3 show the percentages of identity between their PPIPase do-mains and between their 5-Pase domains. The C-terminal PRDs (blackbars) of the yeast synaptojanins do not share extensive identity. CB,clathrin box motifs in Sjl2 and Sjl3. NPF, a putative EH or Sla1 domainbinding site in Sjl1.
TABLE 1. Strains used in this study
Strain Genotype Reference or source
SEY6210 MAT� leu2-3,112 ura3-52 his3-�200 trp1-�901 lys2-801 suc2-�9 39aSEY6210.1 MATa leu2-3,112 ura3-52 his3-�200 trp1-�901 lys2-801 suc2-�9 39aGOY24 Same as SEY6210.1 except pep4::LEU2 prb1::LEU2 Emr lab strain collectionJGY132 Same as SEY6210 except sjl2�::HIS3 sjl3�::TRP1 17YCS62 Same as SEY6210.1 except sjl1�::HIS3 48YCS63 Same as SEY6210.1 except sjl2�::HIS3 48YCS66 Same as SEY6210.1 except sjl1�::HIS3 sjl2�::HIS3 48YCS156 Same as JGY132 except sjl1�::hisG and harboring pRS415SJL2 (LEU2 CEN6 SJL2) 48YCS176 Same as YCS156 except harboring pRS415sjl2ts-8 (LEU2 CEN6 sjl2ts-8) 48YCS192 Same as SEY6210 except rvs167�::HISMx This studyYCS223 Same as SEY6210.1 except SJL2-13xmyc::HisMx This studyYCS279 Same as YCS156 except SNC1-GFP::URA3 This studyYCS280 Same as YCS176 except SNC1-GFP::URA3 This studyYCS284 Same as SEY6210 except SLA2-GFP::HisMx This studyYCS286 Same as GOY24 except sjl2�PRD-13xmyc::HisMx This studyYCS288 Same as SEY6210 except sla1�::kanMx This studyYCS301 Same as SEY6210.1 except abp1�::HisMx This studyYCS323 Same as SEY6210.1 except sjl1�::HIS3 abp1�::HisMx This studyYCS325 Same as GOY24 except abp1�::HisMx This studyYCS327 Same as SEY6210.1 except abp1�::HisMx sjl2�::HIS3 This studyYCS358 Same as SEY6210.1 except sla2�::HisMx This studyYCS374 Same as YCS66 except SLA2-GFP::HisMx This studyAAY1701 Same as SEY6210 except ABP1-DsRed::HisMx 1
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2911
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.
Where indicated, 200 �M latrunculin A (Calbiochem, La Jolla, Calif.) was addedto cells 15 min prior to microscopy. For FM4-64 staining of the plasma mem-brane, cells were grown to mid-log phase at the appropriate temperature, incu-bated at the restrictive temperature for 90 min when indicated, and then har-vested. Metabolic inhibitors (10 mM NaN3 and NaF) were applied, andnonviable cells were stained with FM4-64 (Molecular Probes) at 0°C, as de-scribed elsewhere (44, 48). Cells were visualized on a Zeiss Axiovert S1002TVfluorescence microscope equipped with fluorescein isothiocyanate and rhoda-mine filters, captured with a Photometrix camera, and deconvolved by usingDelta Vision software (Applied Precision, Issaquah, Wash.). Fields showinglocalization of GFP-tagged forms of Sjl2 were two-dimensional representationsof several Z-sections using Delta Vision software. Results presented were basedon observations of more than 100 cells. For live-cell two-color movies followingSjl2-GFP and Abp1-DsRed localization, cells were visualized on a spinning diskconfocal microscope (Yokogawa) equipped with a 100� NA-1.4 objective and acooled charge-coupled device camera (Hamamatsu) controlled by Metamorphsoftware (Universal Imaging). Image pairs from single optical sections scanned at488 and 568 nm were collected every 1.8 s. Throughout, representative fieldswere shown and scales were constant within each figure.
In vivo biochemical analyses. For the coisolation of Sjl2 or Sjl2-myc withGFP-Abp1, 20 OD600 equivalents of cells were harvested, washed, sphero-plasted, and lysed in phosphate-buffered saline lysis buffer, consisting of 8 g ofNaCl/liter, 0.2 g of KCl/liter, 1.44 g of Na2HPO4/liter, 0.24 g of KH2PO4/liter (pH7.2), 0.1 mM EDTA, the protease inhibitors aminoethyl-benzene sulfonyl fluo-ride (AEBSF; Calbiochem), benzamidine, and pepstatin A, and a protease in-hibitor cocktail (Complete; Roche). Lysates were solubilized with 0.5% Tween20 (Sigma) by incubation on ice for 10 min. Samples were cleared by centrifu-gation at 13,000 � g, and supernatants were processed to immunoisolate GFP orGFP-Abp1 by using anti-GFP antibodies (provided by C. Zuker) and proteinA-Sepharose beads (Pharmacia). Material was then visualized by Western blot-ting with either anti-Sjl2 or anti-myc (Genosys, The Woodlands, Tex.) antibodies.
For sizing analysis of Sjl2 complexes in wild-type and abp1� cells, 100 OD600
equivalents of cells were harvested, washed, and lysed in phosphate-bufferedsaline lysis buffer plus 1% Triton X-100. Solubilized lysates were cleared bycentrifugation at 13,000 � g and subsequently loaded onto a linear 10 to 40%glycerol gradient equilibrated in lysis buffer plus 1% Triton X-100. Samplescontaining cleared lysates or molecular weight standards (aldolase, ferritin, andthyroglobulin) were then subjected to centrifugation at 100,000 � g for 4 h in anSW41 rotor. Fractions were then collected from the resulting gradients andanalyzed by Western blotting with anti-Sjl2 antibodies. Sizing standards weredetected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (adolase),OD280 absorbance (adolase and thyroglobulin), and OD420 absorbance (ferritin).The ESCRT-I 350-kDa complex was detected by using anti-Vps23 antibodies (2).
RESULTS
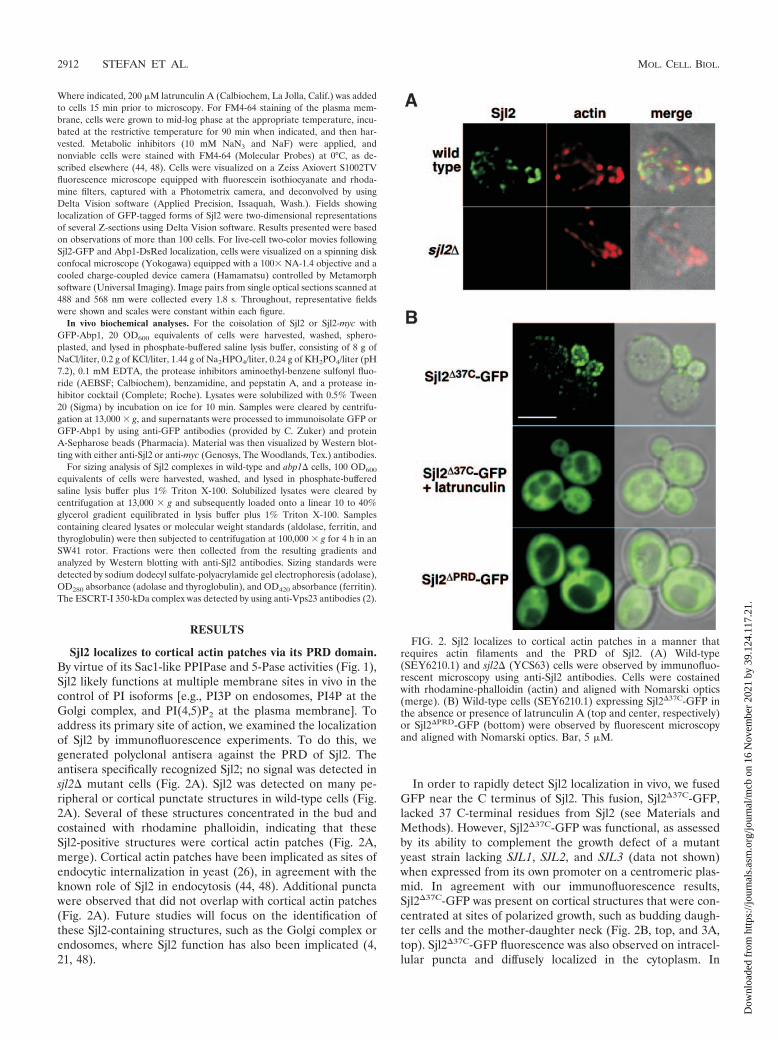
Sjl2 localizes to cortical actin patches via its PRD domain.By virtue of its Sac1-like PPIPase and 5-Pase activities (Fig. 1),Sjl2 likely functions at multiple membrane sites in vivo in thecontrol of PI isoforms [e.g., PI3P on endosomes, PI4P at theGolgi complex, and PI(4,5)P2 at the plasma membrane]. Toaddress its primary site of action, we examined the localizationof Sjl2 by immunofluorescence experiments. To do this, wegenerated polyclonal antisera against the PRD of Sjl2. Theantisera specifically recognized Sjl2; no signal was detected insjl2� mutant cells (Fig. 2A). Sjl2 was detected on many pe-ripheral or cortical punctate structures in wild-type cells (Fig.2A). Several of these structures concentrated in the bud andcostained with rhodamine phalloidin, indicating that theseSjl2-positive structures were cortical actin patches (Fig. 2A,merge). Cortical actin patches have been implicated as sites ofendocytic internalization in yeast (26), in agreement with theknown role of Sjl2 in endocytosis (44, 48). Additional punctawere observed that did not overlap with cortical actin patches(Fig. 2A). Future studies will focus on the identification ofthese Sjl2-containing structures, such as the Golgi complex orendosomes, where Sjl2 function has also been implicated (4,21, 48).
In order to rapidly detect Sjl2 localization in vivo, we fusedGFP near the C terminus of Sjl2. This fusion, Sjl2�37C-GFP,lacked 37 C-terminal residues from Sjl2 (see Materials andMethods). However, Sjl2�37C-GFP was functional, as assessedby its ability to complement the growth defect of a mutantyeast strain lacking SJL1, SJL2, and SJL3 (data not shown)when expressed from its own promoter on a centromeric plas-mid. In agreement with our immunofluorescence results,Sjl2�37C-GFP was present on cortical structures that were con-centrated at sites of polarized growth, such as budding daugh-ter cells and the mother-daughter neck (Fig. 2B, top, and 3A,top). Sjl2�37C-GFP fluorescence was also observed on intracel-lular puncta and diffusely localized in the cytoplasm. In
FIG. 2. Sjl2 localizes to cortical actin patches in a manner thatrequires actin filaments and the PRD of Sjl2. (A) Wild-type(SEY6210.1) and sjl2� (YCS63) cells were observed by immunofluo-rescent microscopy using anti-Sjl2 antibodies. Cells were costainedwith rhodamine-phalloidin (actin) and aligned with Nomarski optics(merge). (B) Wild-type cells (SEY6210.1) expressing Sjl2�37C-GFP inthe absence or presence of latrunculin A (top and center, respectively)or Sjl2�PRD-GFP (bottom) were observed by fluorescent microscopyand aligned with Nomarski optics. Bar, 5 �M.
2912 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

Sjl2�37C-GFP-expressing cells treated with the actin-depoly-merizing agent latrunculin A, Sjl2�37C-GFP fluorescence wasredistributed throughout the cytoplasm (Fig. 2B), indicatingthat Sjl2�37C-GFP localization or recruitment was dependenton filamentous actin.
A previous study has shown that the PRD of Sjl2 is sufficientfor recruitment to nonpolarized actin filaments under hyper-osmotic stress conditions (39). We examined whether this re-gion of Sjl2 was necessary for the recruitment of Sjl2 to corticalactin patches at the plasma membrane. We fused GFP to atruncated form of Sjl2 lacking 190 C-terminal residues(Sjl2�PRD-GFP). This mutant form of Sjl2 was expressed atwild-type levels and at the predicted molecular weight, as de-termined by immunoblotting experiments (data not shown).However, Sjl2�PRD-GFP localized diffusely throughout the cy-toplasm rather than on punctate structures (Fig. 2B). Takentogether, our results indicated that Sjl2 is recruited to corticalpunctate structures and that this process requires filamentousactin and the PRD of Sjl2.
Abp1 is necessary for Sjl2 recruitment in vivo. Next, wesearched for trans-acting factors that concentrate Sjl2 at corti-cal patches. Since the PRD was necessary for Sjl2 recruitmentand SH3 domains bind proline-rich motifs, we examined rolesfor SH3-containing proteins in the localization of Sjl2 at cor-tical actin patches. We identified the actin-binding proteinAbp1 as necessary for Sjl2�37C-GFP recruitment to corticalstructures. In abp1� mutant cells, Sjl2�37C-GFP localized dif-fusely throughout the cytoplasm (Fig. 3A), a pattern similar tothat of Sjl2�PRD-GFP expressed in wild-type cells. In contrast,the SH3-containing proteins Sla1 and Rvs167, which have re-cently been shown to interact biochemically with Sjl2 (47),were not essential for Sjl2�37C-GFP cortical recruitment (Fig.3A). However, cytoplasmic Sjl2�37C-GFP fluorescence ap-peared higher in rvs167� and sla1� cells than in wild-type cells.Thus, while multiple factors may be involved in targeting Sjl2,our results have suggested an important role for Abp1.
A previous in vitro study used phage display to identify aproline-rich peptide (PPVVKKP) in the PRD of Sjl2 (residues1115 to 1121) as a possible binding site for the Abp1 SH3 domain(14). To address whether these residues were involved in Sjl2localization in vivo, we constructed a mutant form of Sjl2�37C-GFP that bore substitutions in the 1115PPVVKKP1121 motif (Fig.3B). This mutant form of Sjl2 (Sjl2K1120V,P1121D,�37C-GFP) wasmislocalized to the cytoplasm of wild-type cells (Fig. 3B), a pat-tern similar to the diffuse localization of wild-type Sjl2�37C-GFPin abp1� cells. Altogether, our results suggested that Abp1 acts asa recruitment factor for Sjl2 and identified specific residues withinthe Sjl2 PRD important for Sjl2 localization, likely by direct as-sociation with the Abp1 SH3 domain.
Sjl2 and Abp1 colocalize and physically interact in vivo. Toexamine whether Sjl2 colocalizes with Abp1 and to monitor thetemporal relationship of these actin patch proteins, we coex-pressed Sjl2�37C-GFP and a functional Abp1-DsRed fusionprotein (1) and imaged cells in real time by spinning diskconfocal microscopy. Due to differences in the relative inten-sities of Sjl2�37C-GFP and Abp1-DsRed fluorescence,Sjl2�37C-GFP was expressed from a high-copy-number plasmidin these experiments. In agreement with our results indicatingthat Abp1 was necessary for Sjl2 cortical recruitment, signifi-cant colocalization was observed between Sjl2�37C-GFP and
Abp1-DsRed at cortical patches (in four of seven Sjl2�37C-GFP patches at the initial time point [Fig. 4A]). Closer exam-ination indicated that two additional Sjl2�37C-GFP patchesoverlapped with faint Abp1-DsRed fluorescence (Fig. 4A).More importantly, this analysis revealed an ordered sequencefor the recruitment and disappearance of Sjl2 and Abp1 atcortical patches. Based on observations of more than 30patches from several cells, Abp1-DsRed appeared at patchesprior to Sjl2�37C-GFP, as Sjl2�37C-GFP consistently concen-trated at preexisting Abp1-DsRed patches. To demonstrate anexample of this progression, a specific region in Fig. 4A wasmagnified (by 120%) and shown throughout the course of thisexperiment in Fig. 4B. Interestingly, the two proteins coexistedon patches only transiently, with Abp1-DsRed disappearingfrom the patch rapidly after Sjl2�37C-GFP recruitment (Fig.4B). In addition, Sjl2�37C-GFP remained in patches followingthe disappearance of Abp1-DsRed. For example, each of theSjl2�37C-GFP patches that initially colocalized with Abp1-DsRed (Fig. 4A) persisted following the disappearance ofAbp1-DsRed. This process may explain the observation ofSjl2�37C-GFP patches that did not appear to display significantoverlap with Abp1-DsRed at the start of this experiment. Al-ternatively, Sjl2 may exist in a separate complex that transientlyinteracts with Abp1 patches.
To further demonstrate a role for Abp1 in the recruitment ofSjl2 to cortical actin patches, we examined whether Abp1 andSjl2 physically associate in vivo. Wild-type cells expressing GFPalone or functional GFP-Abp1 (9) were lysed. Next, GFP orGFP-Abp1 was immunoisolated from lysates solubilized undernative conditions by using anti-GFP antibodies. Bound mate-rial was then probed for the presence of Sjl2 by using Sjl2-specific antisera. Sjl2 copurified with GFP-Abp1 but not withGFP alone (Fig. 5A). To address whether this interaction wasmediated through the PRD of Sjl2, GFP-Abp1 was immu-noisolated from cells expressing full-length Sjl2-myc or a mu-tant form that lacked the PRD, Sjl2�PRD-myc. These experi-ments employed myc-tagged fusions, because the anti-Sjl2antisera were directed against the PRD of Sjl2. Sjl2-myc copu-rified with GFP-Abp1 but not with GFP alone (Fig. 5B), resultssimilar to those obtained using native Sjl2. In contrast,Sjl2�PRD-myc did not copurify with GFP-Abp1. Taken to-gether, these data indicated that Sjl2 and Abp1 associate invivo and that this interaction occurs through the PRD of Sjl2,in agreement with our findings that the PRD of Sjl2 and Abp1direct the recruitment of Sjl2 to cortical patches in vivo.
We further tested whether Abp1 and Sjl2 physically associ-ate in vivo by determining the relative molecular size of Sjl2 inextracts from wild-type and abp1� cells by using velocity gra-dient sedimentation analysis. In wild-type cell extracts, Sjl2 waspresent in fractions corresponding to a range of 440 to 669 kDa(Fig. 6). In extracts from abp1� cells, the profile of Sjl2 wasshifted to a lower-molecular-size range (approximately 350 to440 kDa [Fig. 6]), consistent with our earlier result that Abp1and Sjl2 interact. However, Sjl2 remained in a complex largerthan its monomeric size (approximately 120 kDa) in abp1�cells, suggesting that additional factors associate with Sjl2. Insupport of this idea, dual-color imaging of Sjl2 and Abp1fusions indicated that Sjl2-GFP persisted in cortical patchesfollowing the rapid movement or disappearance of Abp1-DsRed from these patches (Fig. 4). Further studies will be
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2913
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

necessary to define the precise composition of the Sjl2 com-plex(es). As a control, the size distribution of the 350-kDaESCRT-I complex, consisting of Vps23, Vps28, and Vps37 (2),remained constant in extracts prepared from wild-type andabp1� cells (Fig. 6).
Abp1 and Sjl2 share similar functions in vivo. To furtherexamine the role of the Abp1-Sjl2 interaction, we investigatedwhether Abp1 was involved in Sjl2 function in vivo. Thus, weexamined whether abp1� mutant cells displayed phenotypes
FIG. 3. Recruitment of Sjl2�37C-GFP to cortical patches is depen-dent on Abp1. (A) Localization of Sjl2�37C-GFP in living wild-type(SEY6210.1), abp1� (YCS301), rvs167� (YCS192), and sla1�(YCS288) cells was observed by fluorescence (Sjl2�37C-GFP) and No-marski (merge) microscopy. (B) Wild-type cells (SEY6210.1) express-ing Sjl2K1120V,P1121D,�37C-GFP (Sjl2KV,PD,�37C-GFP) were observed byfluorescence and Nomarski microscopy. The diagram shows substitu-tions in the Sjl2 PRD used in this experiment. Bars, 5 �M.
FIG. 4. Sjl2�37C-GFP and Abp1-DsRed colocalize in an ordered,transient manner. (A) Sjl2�37C-GFP was coexpressed in wild-type cellsexpressing Abp1-DsRed (strain AAY1701). Frames from a single focalplane movie visualizing Abp1-DsRed (left) and Sjl2�37C-GFP (center)are shown along with merged images (right). The time to acquire oneimage pair was 1.8 s; every other time point has been shown. Arrow-heads and asterisks indicate significant and faint colocalization ofSjl2�37C-GFP and Abp1-DsRed at cortical patches, respectively.(B) Time series showing Abp1-Sjl2 composition of a single patch(boxed region in panel A). Individual images showing Abp1-DsRed(top) and Sjl2�37C-GFP (middle) are aligned with the merged imagepairs (bottom). The dashed line indicates the cell cortex.
2914 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

similar to those of cells lacking Sjl2. Because abp1� and sjl2�single-mutant cells display no obvious phenotypes, we com-pared phenotypes exhibited by sjl1� sjl2� double-mutant cellswith those of sjl1� abp1� double-mutant cells and found thatthey displayed similar properties. Cells lacking both SJL1 and
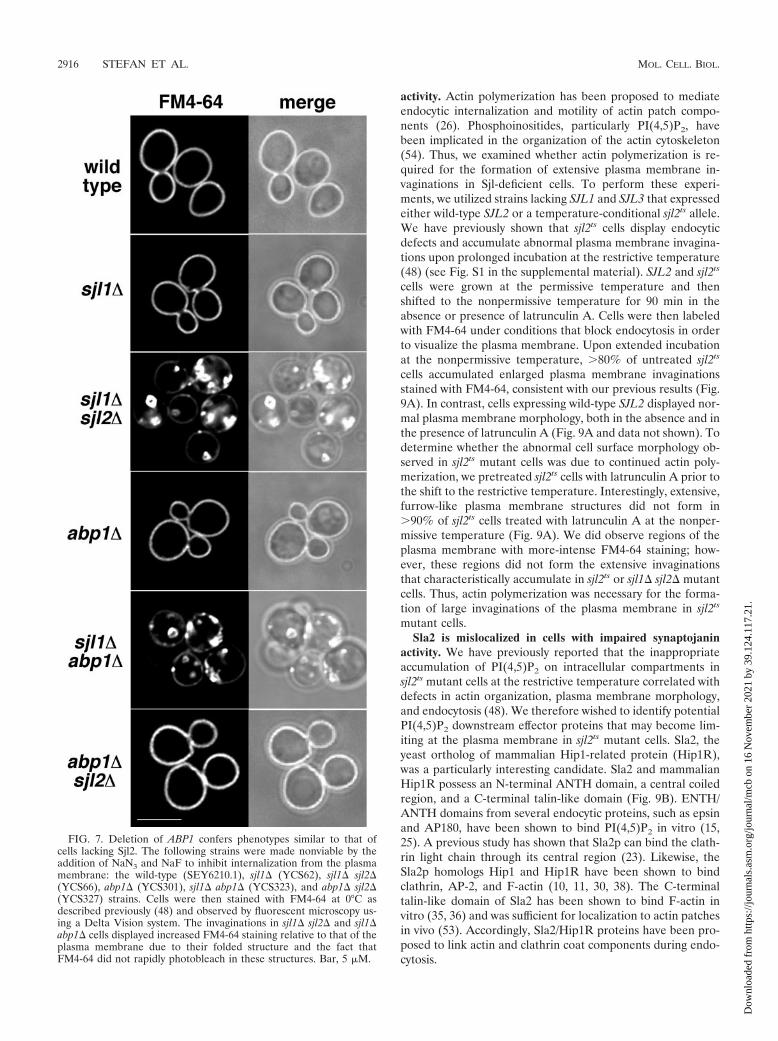
SJL2 have been shown to accumulate abnormal, enlargedplasma membrane invaginations that fail to undergo fissionand often fold around themselves to enclose portions of cyto-plasm (44, 46, 48, 49). These structures can be readily visual-ized by staining the membrane surfaces of sjl1� sjl2� cells withthe lipophilic dye FM4-64 under conditions that block endo-cytosis (0°C and metabolic inhibitors) (44). As expected,�90% of sjl1� sjl2� double-mutant cells displayed massive,furrow-like invaginations stained with FM4-64 that are contin-uous with the plasma membrane (Fig. 7). These invaginationsdisplayed increased FM4-64 staining relative to the plasmamembrane due to their double membrane structure and be-cause FM4-64 did not rapidly photobleach in these structures.Addition of FM4-64 to wild-type, sjl1�, and sjl2� cells underthese conditions did not reveal the presence of any such cellsurface structures (Fig. 7 and data not shown). Likewise, abp1�mutant cells did not accumulate invaginations of the plasmamembrane like those that form in sjl1� sjl2� double-mutantcells. Interestingly, deletion of SJL1 and ABP1 in combinationresulted in the formation of deep, furrow-like invaginations ofthe plasma membrane that were stably labeled with FM4-64 in�50% of cells observed in these experiments (Fig. 7). Thus,sjl1� sjl2� double-mutant cells and sjl1� abp1� double-mutantcells displayed similar phenotypes. Notably, abp1� sjl2� mu-tant cells did not accumulate the furrow-like plasma mem-brane structures that are present in sjl1� sjl2� double-mutantcells (Fig. 7), suggesting that Sjl1 is recruited by a differentmechanism.
Previous studies have shown that sjl1� sjl2� double mutantsdisplay defects in endocytic internalization, while sjl1�, sjl2�,or abp1� single-mutant cells do not (24, 44). Thus, we exam-ined whether sjl1� abp1� double-mutant cells displayed endo-cytic defects similar to those of sjl1� sjl2� double-mutant cells.To do this, we monitored the internalization and transport ofthe lipophilic dye FM4-64 to the vacuole. Cells were pulse-labeled with FM4-64 and CMAC (a vital dye that accumulatesin the vacuole lumen), washed, and chased in a medium thatdid not contain FM4-64 or CMAC. As expected, wild-type andabp1� mutant cells displayed FM4-64 fluorescence only invacuole membranes or late endosomal compartments adjacentto vacuoles following a 30-minute chase (Fig. 8). Likewise,�80% of sjl1� sjl2� double-mutant cells expressing Sjl2�37C-GFP from a plasmid delivered FM4-64 to vacuole membranesor compartments adjacent to vacuoles within the chase period(Fig. 8, sjl1� sjl2�37C-GFP). In contrast, sjl1� sjl2� double-mutant cells displayed defects in delivery of FM4-64 to thevacuole. FM4-64 fluorescence was observed in structures thatwere distinct from vacuoles, possibly corresponding to cellsurface invaginations (Fig. 8). Interestingly, sjl1� abp1�double-mutant cells displayed a delay in endocytic traffick-ing to vacuoles; FM4-64 was found in peripheral structures in�50% of these cells (Fig. 8). We also expressedSjl2K1120V,P1121D,�37C-GFP in cells lacking SJL1 and SJL2 andfound FM4-64 in peripheral structures in �40% of these cells(Fig. 8, sjl1� sjl2KP,VD,�37C-GFP). Together, these results fur-ther implicate Abp1 in endocytosis and suggest that the pre-dicted Abp1 binding site in Sjl2 is necessary for full Sjl2 func-tion in the endocytic pathway.
Actin polymerization drives growth of exaggerated plasmamembrane invaginations in cells with impaired synaptojanin
FIG. 5. Sjl2 and Abp1 interact in vivo in a manner dependent onthe Sjl2 PRD. (A) Wild-type cells (SEY6210.1) or sjl2� cells (YCS63)expressing GFP or GFP-Abp1 were lysed and adjusted to 0.5% Tween20. GFP or GFP-Abp1 was immunoisolated under native conditions.Isolated material was probed with anti-Sjl2 antibodies to detect Sjl2.Input lanes show 10% of the total Sjl2 present in the lysates. (B) Cellsexpressing Sjl2-myc (YCS223) or Sjl2�PRD-myc (YCS286) and GFP orGFP-Abp1 were lysed and solubilized. GFP or GFP-Abp1 was immu-noisolated under native conditions as before. Bound material wasanalyzed by Western blotting using antisera to the myc epitope. Inputlanes show 10% of the total Sjl2-myc and Sjl2�PRD-myc in the lysates.IP, immunoprecipitation.
FIG. 6. Sjl2 is present in a high-molecular-weight complex thatcontains Abp1. Wild-type (GOY24) or abp1� cells (YCS325) werelysed, solubilized in 1% Triton X-100, and cleared by a 13,0000 � gcentrifugation step. Samples containing either cleared lysates or mo-lecular weight standards were loaded onto a linear 10 to 40% glycerolgradient equilibrated in lysis buffer plus 1% Triton X-100 and weresubjected to centrifugation at 100,000 � g. Fractions were collectedfrom the resulting gradients and analyzed by Western blotting withanti-Sjl2 antibodies (upper panels). Fractions containing sizing stan-dards are indicated. The size of the Vps23-containing ESCRT-1 com-plex was determined as a control (lower panels).
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2915
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.
activity. Actin polymerization has been proposed to mediateendocytic internalization and motility of actin patch compo-nents (26). Phosphoinositides, particularly PI(4,5)P2, havebeen implicated in the organization of the actin cytoskeleton(54). Thus, we examined whether actin polymerization is re-quired for the formation of extensive plasma membrane in-vaginations in Sjl-deficient cells. To perform these experi-ments, we utilized strains lacking SJL1 and SJL3 that expressedeither wild-type SJL2 or a temperature-conditional sjl2ts allele.We have previously shown that sjl2ts cells display endocyticdefects and accumulate abnormal plasma membrane invagina-tions upon prolonged incubation at the restrictive temperature(48) (see Fig. S1 in the supplemental material). SJL2 and sjl2ts
cells were grown at the permissive temperature and thenshifted to the nonpermissive temperature for 90 min in theabsence or presence of latrunculin A. Cells were then labeledwith FM4-64 under conditions that block endocytosis in orderto visualize the plasma membrane. Upon extended incubationat the nonpermissive temperature, �80% of untreated sjl2ts
cells accumulated enlarged plasma membrane invaginationsstained with FM4-64, consistent with our previous results (Fig.9A). In contrast, cells expressing wild-type SJL2 displayed nor-mal plasma membrane morphology, both in the absence and inthe presence of latrunculin A (Fig. 9A and data not shown). Todetermine whether the abnormal cell surface morphology ob-served in sjl2ts mutant cells was due to continued actin poly-merization, we pretreated sjl2ts cells with latrunculin A prior tothe shift to the restrictive temperature. Interestingly, extensive,furrow-like plasma membrane structures did not form in�90% of sjl2ts cells treated with latrunculin A at the nonper-missive temperature (Fig. 9A). We did observe regions of theplasma membrane with more-intense FM4-64 staining; how-ever, these regions did not form the extensive invaginationsthat characteristically accumulate in sjl2ts or sjl1� sjl2� mutantcells. Thus, actin polymerization was necessary for the forma-tion of large invaginations of the plasma membrane in sjl2ts
mutant cells.Sla2 is mislocalized in cells with impaired synaptojanin
activity. We have previously reported that the inappropriateaccumulation of PI(4,5)P2 on intracellular compartments insjl2ts mutant cells at the restrictive temperature correlated withdefects in actin organization, plasma membrane morphology,and endocytosis (48). We therefore wished to identify potentialPI(4,5)P2 downstream effector proteins that may become lim-iting at the plasma membrane in sjl2ts mutant cells. Sla2, theyeast ortholog of mammalian Hip1-related protein (Hip1R),was a particularly interesting candidate. Sla2 and mammalianHip1R possess an N-terminal ANTH domain, a central coiledregion, and a C-terminal talin-like domain (Fig. 9B). ENTH/ANTH domains from several endocytic proteins, such as epsinand AP180, have been shown to bind PI(4,5)P2 in vitro (15,25). A previous study has shown that Sla2p can bind the clath-rin light chain through its central region (23). Likewise, theSla2p homologs Hip1 and Hip1R have been shown to bindclathrin, AP-2, and F-actin (10, 11, 30, 38). The C-terminaltalin-like domain of Sla2 has been shown to bind F-actin invitro (35, 36) and was sufficient for localization to actin patchesin vivo (53). Accordingly, Sla2/Hip1R proteins have been pro-posed to link actin and clathrin coat components during endo-cytosis.
FIG. 7. Deletion of ABP1 confers phenotypes similar to that ofcells lacking Sjl2. The following strains were made nonviable by theaddition of NaN3 and NaF to inhibit internalization from the plasmamembrane: the wild-type (SEY6210.1), sjl1� (YCS62), sjl1� sjl2�(YCS66), abp1� (YCS301), sjl1� abp1� (YCS323), and abp1� sjl2�(YCS327) strains. Cells were then stained with FM4-64 at 0°C asdescribed previously (48) and observed by fluorescent microscopy us-ing a Delta Vision system. The invaginations in sjl1� sjl2� and sjl1�abp1� cells displayed increased FM4-64 staining relative to that of theplasma membrane due to their folded structure and the fact thatFM4-64 did not rapidly photobleach in these structures. Bar, 5 �M.
2916 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

FIG. 8. Deletion of ABP1 and SJL1 results in synthetic endocytic defects, similar to those caused by sjl1 and sjl2 mutations. Wild-type(SEY6210.1), sjl1� sjl2� (YCS66), abp1� (YCS301), and sjl1� abp1� (YCS323) cells, or strain YCS66 expressing sjl2�37-GFP orSjl2K1120V,P1121D,�37C-GFP (Sjl2KV,PD,�37C-GFP) from a plasmid, were labeled with the vital dyes FM4-64 and CMAC for 15 min. After labeling,cells were chased for 30 min. (Left panels) Cells under fluorescent illumination in the rhodamine channel (FM4-64). (Center panels) Cells asobserved by CMAC fluorescence to visualize the lumens of vacuoles. (Right panels) Fluorescent images were merged with corresponding DICimages (merge). Arrowheads mark examples of delayed endocytic intermediates containing FM4-64. Bar, 5 �M.
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2917
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

First, we addressed whether overexpression of Sla2 mightrestore normal plasma membrane morphology in sjl2ts cells.Thus, SLA2 was expressed from a high-copy-number plasmidin SJL2 and sjl2ts cells grown at the permissive temperature.Cells were then shifted to the nonpermissive temperature for90 min and subsequently labeled with FM4-64 under condi-tions that block endocytosis in order to visualize the plasma
membrane. Expression of SLA2 from a high-copy-numberplasmid had no effect on plasma membrane morphology inSJL2 cells (data not shown). As expected, �80% of sjl2ts cellscarrying vector alone accumulated enlarged plasma membraneinvaginations stained with FM4-64 upon extended incubationat the nonpermissive temperature (Fig. 9B). Notably, ex-tended, furrow-like plasma membrane structures did not formin �70% of sjl2ts cells overexpressing SLA2 (Fig. 9B). We didobserve punctate structures at the plasma membrane withFM4-64 staining; however, these regions did not develop intothe deep invaginations that form in sjl2ts cells.
Next, we addressed whether overexpression of Sla2 mightrescue endocytic trafficking defects in sjl2ts cells. To do this, wemonitored the transport of FM4-64 to the vacuole in SJL2 andsjl2ts cells expressing SLA2 from a high-copy-number plasmidfollowing incubation at the restrictive temperature. After a30-min chase, FM4-64 fluorescence was efficiently delivered tothe vacuole membrane in SJL2 cells carrying either vectoralone or the SLA2 plasmid (Fig. 10). As expected, sjl2ts cellsdisplayed defects in delivery of FM4-64 to the vacuole, sinceFM4-64 fluorescence was observed in punctate structures thatwere distinct from vacuoles (Fig. 10). Interestingly, expressionof SLA2 from a high-copy-number plasmid increased the rateof endocytic trafficking to the vacuole in sjl2ts cells, as FM4-64efficiently labeled the vacuole membrane in �75% of thesecells (Fig. 10). Together, these results suggested that Sla2 func-tion was limiting or misregulated in sjl2ts cells at the nonper-missive temperature.
Thus, we examined the localization of a functional Sla2-GFPfusion in wild-type and sjl1� sjl2� double-mutant cells. Cellslacking SJL1 and SJL2 have been reported to display defects inactin organization and endocytic internalization (44, 46). Inwild-type cells, Sla2-GFP was observed at cortical patches con-centrated in the bud, a pattern similar to previously publishedpatterns of Sla2 localization at cortical actin patches (53) (Fig.11A). However, in sjl1� sjl2� mutant cells, Sla2-GFP displayedrandom distribution in both mother and daughter cells and wasno longer strictly cortical (Fig. 11A). We also addressedwhether Sla2 localized to the plasma membrane furrows thataccumulate in sjl1� sjl2� cells. To do this, we labeled theplasma membranes of sjl1� sjl2� double-mutant cells express-ing Sla2-GFP with FM4-64 under conditions that block endo-cytosis (0°C and metabolic inhibitors). We did not observesignificant overlap between FM4-64 and Sla2-GFP fluores-cence in these cells (see Fig. S2 in the supplemental material),consistent with the idea that Sla2 might be limiting at sites ofendocytic internalization in sjl1� sjl2� mutant cells.
Yeast cells that lack Sla2 and mammalian cells depleted ofHip1R have been shown to accumulate comet-like actin tails atsites of endocytic internalization (12, 26). We therefore exam-ined whether sla2� mutant cells form plasma membrane in-vaginations similar to those of sjl2ts or sjl1� sjl2� mutant cells.Thus, wild-type and sla2� cells were stained with FM4-64 un-der conditions that block endocytosis, as before. As expected,wild-type cells displayed normal plasma membrane morphol-ogy (Fig. 11B). Interestingly, �75% of sla2� cells displayedmultiple punctate plasma membrane structures stained withFM4-64 (Fig. 11B). While these structures did not appear to beas extensive as the invaginations that form in sjl2ts or sjl1� sjl2�cells, these results suggested that sla2� mutant cells accumu-
FIG. 9. (A) Actin polymerization is required for continued forma-tion of exaggerated plasma membrane invaginations in sjl2ts cells. SJL2(YCS157) or sjl2ts (YCS176) cells grown to mid-log phase at the per-missive temperature were then incubated at the restrictive tempera-ture for 90 min in the absence or presence of latrunculin A as indi-cated. Metabolic inhibitors (10 mM NaN3 and NaF) were added, andcells were labeled with FM4-64 at 0°C to visualize the plasma mem-brane. (B) SLA2 overexpression attenuates the formation of exagger-ated plasma membrane invaginations in sjl2ts cells. The diagram showsSla2 domain structure. YCS176 cells (sjl2ts) carrying vector alone or ahigh-copy-number SLA2 expression plasmid were grown to mid-logphase at the permissive temperature and then incubated at the restric-tive temperature for 90 min. Metabolic inhibitors (10 mM NaN3 andNaF) were added, and cells were labeled with FM4-64 at 0°C tovisualize the plasma membrane. Bars, 5 �M.
2918 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

late plasma membrane invaginations. Taken together, our re-sults have suggested that Sla2 becomes misregulated or limit-ing at the plasma membrane in cells with impaired SJL-encoded PI Pase activity, which may subsequently result ininefficient membrane fission or scission during endocytic inter-nalization.
DISCUSSION
We have shown that Sjl2 localizes to cortical actin patchesand that this localization depended on the PRD of Sjl2, actinfilaments, and the actin patch component Abp1. Previous stud-ies have indicated that elevated PI(4,5)P2 levels in cells lackingSJL-encoded PI 5-Pase activity cause defects in actin filamentorganization (39, 48). Here, we have shown that actin filamentswere necessary for the formation of extensive plasma mem-
brane invaginations in sjl2ts mutant cells, suggesting that mis-regulated actin polymerization may lead to defects in properfission and release of endocytic vesicle intermediates. Finally,our studies have indicated that overexpression of the actinpatch protein Sla2 partially rescued the endocytic and plasmamembrane morphology defects observed in sjl2ts mutant cells.
Sjl2 colocalizes with Abp1. Several lines of evidence indicatethat Sjl2 and Abp1 function at the same site in the control ofendocytosis. Both Abp1 and Sjl2�37C-GFP are recruited toactin patches in a latrunculin-sensitive manner (Fig. 2B) (26).In abp1� mutant cells, Sjl2�37C-GFP localizes diffuselythroughout the cytoplasm (Fig. 3A). The PRD of Sjl2 is nec-essary for the recruitment of Sjl2 to cortical actin patches andfor physical association with Abp1 (Fig. 2B and 5). Thus, ourresults provide in vivo evidence that Abp1 acts as an adaptorfor Sjl2 at actin patches via the PRD of Sjl2. Deletion of SJL2
FIG. 10. SLA2 overexpression increases endocytic rates in sjl2ts cells. SJL2 (YCS157) or sjl2ts (YCS176) cells carrying vector alone or ahigh-copy-number SLA2 expression plasmid were grown to mid-log phase at the permissive temperature and then incubated at the restrictivetemperature for 90 min. Cells were then labeled with the vital dyes FM4-64 and CMAC for 15 min. After labeling, cells were chased for 30 minat the nonpermissive temperature. (Left panels) Cells under fluorescent illumination in the rhodamine channel (FM4-64). (Center panels) Cellsas observed by CMAC fluorescence to visualize the lumens of vacuoles. (Right panels) Cells as observed by fluorescence and Nomarski optics(merge). Bar, 5 �M.
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2919
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

and deletion of ABP1 have similar effects; sjl1� abp1� double-mutant cells display furrow-like plasma membrane invagina-tions and endocytic defects similar to those observed in sjl1�sjl2� double-mutant cells (Fig. 7). Notably, abp1� sjl2� mutantcells do not display defects in plasma membrane morphologyor actin cytoskeletal organization (Fig. 7 and our unpublisheddata), unlike sjl1� sjl2� or sjl2� sjl3� mutant cells (44, 46).Thus, Abp1 appears to specifically recruit Sjl2 at cortical actin
patches, but not Sjl1 or Sjl3. Further studies are needed toidentify Sjl1- and Sjl3-specific targeting factors.
Abp1 is recruited to actin patches at a late step duringendocytic internalization and then rapidly moves away fromthe plasma membrane (26). Consistent with a role for Sjl2during late stages of vesicle fission and uncoating, Sjl2�37C-GFP concentrates at preexisting Abp1 cortical patches, andAbp1 disappears from patches shortly following Sjl2 recruit-ment (Fig. 4). Previous studies indicate that Abp1 also inter-acts with several other components of actin patches, includingthe Arp2/3 complex, the yeast amphiphysin ortholog Rvs167(see below), the type I myosin Myo3, and the kinases Ark1/Prk1 (6, 14, 19, 32). It is not likely that Abp1 binds to thesefactors simultaneously. So then, how are multiple Abp1 inter-actions selectively coordinated at actin patches? One possibleexplanation comes from previous studies indicating that mam-malian synaptojanin is a member of the dephosphin family ofendocytic proteins, which also includes dynamin, AP180, en-dophilin, and amphiphysin; these proteins associate with eachother only following Ca2�-stimulated dephosphorylation (29,45). Thus, a complex series of phosphorylation and dephos-phorylation events may regulate the recognition by Abp1 ofvarious targets during late events in endocytic internalization.Sjl2 might be dephosphorylated temporally later than otherAbp1 targets, thus avoiding premature termination ofPI(4,5)P2-mediated vesicle formation events. Further studiesare needed to examine whether Sjl2 is regulated by phosphor-ylation-dephosphorylation and, if so, to identify the relevantkinases and phosphatases.
Our studies also indicated that multiple Sjl2-interacting fac-tors exist, analogous to the complex interactions that controlthe localization and activity of mammalian synaptojanins. First,we found that Sjl2 �37C-GFP persisted in cortical patches fol-lowing the disappearance of Abp1-DsRed (Fig. 4). In addition,our sizing analysis indicated that Sjl2 remained in a complexlarger than its predicted monomeric size in lysates from abp1�cells (Fig. 6). Recent studies have shown that Sjl2 physicallyassociates with two other SH3 domain-containing proteins,Rvs167 and Sla1, and another protein, Bsp1 (47, 52). Rvs167 isthe yeast amphiphysin ortholog; thus, the association of Sjl2with Rvs167 is consistent with the known interaction betweenamphiphysin and mammalian synaptojanin (5, 45). Likewise, arecent study has indicated that the mammalian homolog ofSla1, CIN85, physically associates with synaptojanin 2 (28).Bsp1 was identified as a cortical actin patch protein that inter-acts with the Sac1 domains of Sjl2 and Sjl3 (47). Sla1, Rvs167,and Bsp1 may act as additional recruitment or specificity fac-tors for Sjl2 localization or regulation (see Fig. 12), consistentwith our genetic results indicating that sjl1� abp1� mutantcells did not fully phenocopy sjl1� sjl2� mutant cells. Whileour initial Sjl2�37C-GFP localization experiments suggestedthat Sla1, Rvs167, and Bsp1 were not essential for Sjl2 local-ization at cortical patches (Fig. 3 and data not shown), theseproteins may influence the stability or lifetime of Sjl2 at cor-tical patches. Our study has not excluded the possibility thatresidues deleted in Sjl2�37C-GFP provide important interac-tions with Sla1 or Rvs167, for example, that direct Sjl2 local-ization along with Abp1. Alternatively, Sjl2 may exist in astable subcomplex that transiently interacts with Abp1 patches.
FIG. 11. (A) Proper localization of Sla2 at cortical patches is de-pendent on Sjl1 and Sjl2. Localization of Sla2-GFP in living wild-type(YCS284) and sjl1� sjl2� (YCS374) cells was observed by fluorescence(Sla2-GFP) and Normarski (merge) microscopy. (B) Deletion of SLA2confers abnormal plasma membrane dynamics. Wild-type(SEY6210.1) and sla2� (YCS358) cells were made nonviable by theaddition of NaN3 and NaF to inhibit internalization from the plasmamembrane. Cells were then stained with FM4-64 at 0°C as describedpreviously (48) and observed by fluorescent microscopy (FM4-64) andNomarski optics (merge). Bars, 5 �M.
2920 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

In cells lacking Abp1, this complex may become destabilized.Thus, further experiments will be necessary to examine thetemporal and spatial recruitment of Sjl2 relative to additionalactin patch components such as Sla1, Rvs167, and Bsp1.
PI(4,5)P2 turnover regulates the timing of plasma mem-brane invagination and fission events during endocytic inter-nalization. Previous studies have shown that synaptojanins arenecessary for the postfission uncoating of adaptor proteinsfrom newly formed endocytic vesicles (7, 22, 51). Consistentwith this, sjl2ts cells have been shown to accumulate PI(4,5)P2
in intracellular compartments at the restrictive temperature(48). However, if the single function for synaptojanins duringendocytosis is to recycle PI(4,5)P2-regulated vesicle formationfactors from endosomes, subsequent rounds of internalizationmay not proceed in synaptojanin-deficient cells. Strikingly, ourstudies and previous studies have indicated that sjl2ts and sjl1�sjl2� cells accumulate numerous, extensive plasma membraneinvaginations (44, 48). At the ultrastructural level, the cyto-plasmic face of these invaginations appeared to be lined by anelectron-dense “coat”-like material. In addition, a clearing orelectron-transparent region was often observed at the tips ofthe invaginations (see Fig. S1B in the supplemental material).Further experiments are needed to determine whether theseare components of clathrin coats and the actin polymerizationmachinery. Regardless, these examinations have suggested
that vesicle formation factors may not become limiting in Sjl-deficient cells and that multiple internalization events can ini-tiate. Rather, these findings have indicated that reduced SJL-encoded PI Pase function leads to increased growth of plasmamembrane invaginations. Thus, factors important for promot-ing vesicle scission may be limiting or impaired in cells defi-cient in SJL-encoded PI Pase activity.
Several lines of evidence suggested that Sla2, the Hip1Rhomolog shown to associate with the actin cytoskeleton andcomponents of clathrin coats, becomes limiting at the plasmamembrane in sjl mutant cells. First, Sla2-GFP was mislocalizedin sjl1� sjl2� mutant cells. More importantly, SLA2 overex-pression attenuated the formation of the large plasma mem-brane invaginations that accumulate in sjl2ts cells (Fig. 9). Wefound that overexpression of another ANTH domain-contain-ing endocytic protein, yAP180a, or the PH domain from PLC�,as a control, did not rescue the plasma membrane morphologydefects in sjl2ts cells (data not shown). Thus, suppression wasnot simply due to sequestration of excess cellular PI(4,5)P2.Furthermore, SLA2 overexpression rescued the endocytic de-fects displayed by sjl2ts cells, as assessed by monitoring of theinternalization and delivery of FM4-64 to the vacuole (Fig. 10).
Previous studies have shown that deletion of SLA2 in yeastand depletion of Hip1R in mammalian cells resulted in theformation of comet-like actin tails associated with endocytic
FIG. 12. Model for Sjl2 recruitment and function during endocytic internalization. PI(4,5)P2 synthesis by the PI(4)P 5-kinase Mss4 recruitseffector proteins that promote actin polymerization (red cables), clathrin coat formation (brown ovals), and vesicle budding (step 1). The SH3domain-containing protein Abp1 is recruited at a late stage during vesicle formation (26) and subsequently recruits Sjl2. Additional Sjl2-interactingfactors, such as the SH3 domain-containing proteins Rvs167 and Sla1 (47), may also target Sjl2 to actin patches. Following Abp1 recruitment,certain actin patch components, such as the ANTH domain-containing protein Sla2, disassociate from patches (indicated by dashed arrows) (26).Concentration of Sjl2 at patches by Abp1 may serve an important role in this process. Turnover of PI(4,5)P2 at the plasma membrane by Sjl1 andSjl2 provides an important step in coordinating actin polymerization and vesicle fission (step 2); cells lacking SJL1 and SJL2 accumulate extensiveendocytic intermediates. Synaptojanins, such as Sjl2 and Sjl3, also dephosphorylate PI(4,5)P2 on newly formed vesicles to recycle or uncoatPI(4,5)P2-binding factors, such as actin patch components and clathrin coat proteins (step 3).
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2921
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

cargo and machinery (12, 26). In addition, sla2 mutations havebeen shown to cause the formation of large, aggregated actinpatches or clumps (3, 53). These studies have indicated thatSla2 regulates actin cytoskeletal dynamics at cortical actinpatches by destabilizing actin filaments or by negatively regu-lating the Arp2/3 actin polymerization complex or its activa-tors. Thus, mislocalization of Sla2 in Sjl-deficient cells maycause increased actin polymerization and defects in productiveclathrin coat formation, ultimately leading to continued orextended growth of plasma membrane invaginations. Consis-tent with this idea, actin polymerization was necessary for theformation of extensive plasma membrane invaginations in sjl2ts
cells (Fig. 9). However, the punctate structures at the plasmamembrane in sla2� cells (Fig. 11) were not nearly as extensiveas the furrow-like invaginations that form in cells lacking SJL1and SJL2. Thus, additional PI(4,5)P2 effector proteins arelikely to be misregulated and to contribute to the dramaticeffects on plasma membrane and actin cytoskeleton dynamicsobserved in Sjl-deficient cells. Consistent with this, overexpres-sion of SLA2 was not sufficient to bypass the growth defect ofsjl2ts cells at the restrictive temperature (data not shown).
PI(4,5)P2 has been shown to stimulate actin polymerizationby binding several actin-regulatory proteins, such as WASP,cofilin, gelsolin, and capping proteins (54). Thus, increasedPI(4,5)P2 levels at the plasma membrane in Sjl1/2-deficientcells may also enhance cortical actin filament growth and sta-bility, leading in turn to additive effects on proper fission andrelease of endocytic vesicles (see Fig. 12). Consistent with thisidea, we have previously shown that increased cellularPI(4,5)P2 levels directly affect plasma membrane dynamics andactin organization, since sjl1� sjl2� mss4ts triple-mutant cells,which synthesize normal PI(4,5)P2 levels, do not display ab-normal plasma membrane furrows or depolarized actinpatches such as those seen in sjl1� sjl2� double-mutant cells(48). In yeast, at least three actin patch components, Las17 (aWASP ortholog), Pan1 (an Eps15 ortholog), and Abp1, havebeen shown to be activators of the Arp2/3 complex that medi-ates actin polymerization (18). Interestingly, Abp1 is recruitedto actin patches temporally later than Las17 and Pan1 (26).Furthermore, Las17 and Pan1 were shown to disassociate ordisappear from actin patches upon Abp1 recruitment (26).Thus, a late role for Abp1 in recruiting Sjl2 to endocytic ves-icles may be to terminate earlier PI(4,5)P2-dependent stages ofvesicle formation mediated by Las17 or Pan1. Alternatively,Sjl2 may be recruited to actin patches by Abp1 to regulateAbp1 function itself. Interestingly, disappearance or dissocia-tion of cortical Abp1 consistently occurred following Sjl2 re-cruitment (Fig. 4). Further experiments that examine the spa-tial and temporal relationships of known actin patch proteinsin cells with impaired Sjl1, Sjl2, and Sjl3 activity may providefurther insights that allow us to distinguish between these po-tential models.
Likewise, further studies are necessary to identify additionalregulatory factors for Sjl1, Sjl2, and Sjl3. Each contains uniquetargeting signals, and thus, specific recruitment and activationmechanisms for these proteins must exist. Further studies mayuncover specialized roles for these PI phosphatases within theordered processes of membrane invagination, actin cytoskel-etal dynamics, vesicle scission, and subsequent uncoatingevents that occur during endocytosis.
ACKNOWLEDGMENTS
We thank David Drubin, David Botstein, and Greg Odorizzi forproviding strains and plasmids. We are grateful to Perla Arcaira forhelpful technical assistance. We thank members of the Emr lab, espe-cially William Parrish, for useful discussions. We thank Ingrid Niesmanfor assistance with the electron microscopic analysis (ImmunoelectronMicroscopy Core, headed by M. Farquhar).
C.J.S. was supported as a fellow of the American Cancer Society bythe Holland Peck Charitable Fund and as an associate of the HowardHughes Medical Institute. A.A. is currently supported by a Helen HayWhitney postdoctoral fellowship. S.D.E. is an established investigatorof the Howard Hughes Medical Institute.
REFERENCES
1. Audhya, A., R. Loewith, A. B. Parsons, L. Gao, M. Tabuchi, H. Zhou, C.Boone, M. N. Hall, and S. D. Emr. 2004. Genome-wide lethality screenidentifies new PI4,5P2 effectors that regulate the actin cytoskeleton. EMBOJ. 23:3747–3757.
2. Babst, M., G. Odorizzi, E. J. Estepa, and S. D. Emr. 2000. Mammalian tumorsusceptibility gene 101 (TSG101) and the yeast homologue, Vps23p, bothfunction in late endosomal trafficking. Traffic 1:248–258.
3. Baggett, J. J., K. E. D’Aquino, and B. Wendland. 2003. The Sla2p talindomain plays a role in endocytosis in Saccharomyces cerevisiae. Genetics165:1661–1674.
4. Bensen, E. S., G. Costaguta, and G. S. Payne. 2000. Synthetic genetic inter-actions with temperature-sensitive clathrin in Saccharomyces cerevisiae.Roles for synaptojanin-like Inp53p and dynamin-related Vps1p in clathrin-dependent protein sorting at the trans-Golgi network. Genetics 154:83–97.
5. Cestra, G., L. Castagnoli, L. Dente, O. Minenkova, A. Petrelli, N. Migone, U.Hoffmuller, J. Schneider-Mergener, and G. Cesareni. 1999. The SH3 do-mains of endophilin and amphiphysin bind to the proline-rich region ofsynaptojanin 1 at distinct sites that display an unconventional binding spec-ificity. J. Biol. Chem. 274:32001–32007.
6. Colwill, K., D. Field, L. Moore, J. Friesen, and B. Andrews. 1999. In vivoanalysis of the domains of yeast Rvs167p suggests Rvs167p function is me-diated through multiple protein interactions. Genetics 152:881–893.
7. Cremona, O., G. Di Paolo, M. R. Wenk, A. Luthi, W. T. Kim, K. Takei, L.Daniell, Y. Nemoto, S. B. Shears, R. A. Flavell, D. A. McCormick, and P. DeCamilli. 1999. Essential role of phosphoinositide metabolism in synapticvesicle recycling. Cell 99:179–188.
8. Czech, M. P. 2003. Dynamics of phosphoinositides in membrane retrievaland insertion. Annu. Rev. Physiol. 65:791–815.
9. Doyle, T., and D. Botstein. 1996. Movement of yeast cortical actin cytoskel-eton visualized in vivo. Proc. Natl. Acad. Sci. USA 93:3886–3891.
10. Engqvist-Goldstein, A. E., M. M. Kessels, V. S. Chopra, M. R. Hayden, andD. G. Drubin. 1999. An actin-binding protein of the Sla2/Huntingtin inter-acting protein 1 family is a novel component of clathrin-coated pits andvesicles. J. Cell Biol. 147:1503–1518.
11. Engqvist-Goldstein, A. E., R. A. Warren, M. M. Kessels, J. H. Keen,J. Heuser, and D. G. Drubin. 2001. The actin-binding protein Hip1R asso-ciates with clathrin during early stages of endocytosis and promotes clathrinassembly in vitro. J. Cell Biol. 154:1209–1223.
12. Engqvist-Goldstein, A. E., C. X. Zhang, S. Carreno, C. Barroso, J. E. Heuser,and D. G. Drubin. 2004. RNAi-mediated Hip1R silencing results in stableassociation between the endocytic machinery and the actin assembly ma-chinery. Mol. Biol. Cell 15:1666–1679.
13. Evans, P. R., and D. J. Owen. 2002. Endocytosis and vesicle trafficking. Curr.Opin. Struct. Biol. 12:814–821.
14. Fazi, B., M. J. Cope, A. Douangamath, S. Ferracuti, K. Schirwitz, A. Zuc-coni, D. G. Drubin, M. Wilmanns, G. Cesareni, and L. Castagnoli. 2002.Unusual binding properties of the SH3 domain of the yeast actin-bindingprotein Abp1: structural and functional analysis. J. Biol. Chem. 277:5290–5298.
15. Ford, M. G., I. G. Mills, B. J. Peter, Y. Vallis, G. J. Praefcke, P. R. Evans, andH. T. McMahon. 2002. Curvature of clathrin-coated pits driven by epsin.Nature 419:361–366.
16. Ford, M. G., B. M. Pearse, M. K. Higgins, Y. Vallis, D. J. Owen, A. Gibson,C. R. Hopkins, P. R. Evans, and H. T. McMahon. 2001. Simultaneousbinding of PtdIns(4,5)P2 and clathrin by AP180 in the nucleation of clathrinlattices on membranes. Science 291:1051–1055.
17. Foti, M., A. Audhya, and S. D. Emr. 2001. Sac1 lipid phosphatase and Stt4phosphatidylinositol 4-kinase regulate a pool of phosphatidylinositol 4-phos-phate that functions in the control of the actin cytoskeleton and vacuolemorphology. Mol. Biol. Cell 12:2396–2411.
18. Goode, B. L., and A. A. Rodal. 2001. Modular complexes that regulate actinassembly in budding yeast. Curr. Opin. Microbiol. 4:703–712.
19. Goode, B. L., A. A. Rodal, G. Barnes, and D. G. Drubin. 2001. Activation ofthe Arp2/3 complex by the actin filament binding protein Abp1p. J. Cell Biol.153:627–634.
20. Guo, S., L. E. Stolz, S. M. Lemrow, and J. D. York. 1999. SAC1-like domains
2922 STEFAN ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.

of yeast SAC1, INP52, and INP53 and of human synaptojanin encode poly-phosphoinositide phosphatases. J. Biol. Chem. 274:12990–12995.
21. Ha, S. A., J. Torabinejad, D. B. DeWald, M. R. Wenk, L. Lucast, P. DeCamilli, R. A. Newitt, R. Aebersold, and S. F. Nothwehr. 2003. The synap-tojanin-like protein Inp53/Sjl3 functions with clathrin in a yeast TGN-to-endosome pathway distinct from the GGA protein-dependent pathway. Mol.Biol. Cell 14:1319–1333.
22. Harris, T. W., E. Hartwieg, H. R. Horvitz, and E. M. Jorgensen. 2000.Mutations in synaptojanin disrupt synaptic vesicle recycling. J. Cell Biol.150:589–600.
23. Henry, K. R., K. D’Hondt, J. Chang, T. Newpher, K. Huang, R. T. Hudson,H. Riezman, and S. K. Lemmon. 2002. Scd5p and clathrin function areimportant for cortical actin organization, endocytosis, and localization ofsla2p in yeast. Mol. Biol. Cell 13:2607–2625.
24. Holtzman, D. A., S. Yang, and D. G. Drubin. 1993. Synthetic-lethal interac-tions identify two novel genes, SLA1 and SLA2, that control membranecytoskeleton assembly in Saccharomyces cerevisiae. J. Cell Biol. 122:635–644.
25. Itoh, T., S. Koshiba, T. Kigawa, A. Kikuchi, S. Yokoyama, and T. Takenawa.2001. Role of the ENTH domain in phosphatidylinositol-4,5-bisphosphatebinding and endocytosis. Science 291:1047–1051.
26. Kaksonen, M., Y. Sun, and D. G. Drubin. 2003. A pathway for association ofreceptors, adaptors, and actin during endocytic internalization. Cell 115:475–487.
27. Kim, W. T., S. Chang, L. Daniell, O. Cremona, G. Di Paolo, and P. DeCamilli. 2002. Delayed reentry of recycling vesicles into the fusion-compe-tent synaptic vesicle pool in synaptojanin 1 knockout mice. Proc. Natl. Acad.Sci. USA 99:17143–17148.
28. Kowanetz, K., K. Husnjak, D. Holler, M. Kowanetz, P. Soubeyran, D. Hir-sch, M. H. Schmidt, K. Pavelic, P. De Camilli, P. A. Randazzo, and I. Dikic.2004. CIN85 associates with multiple effectors controlling intracellular traf-ficking of EGF receptors. Mol. Biol. Cell 15:3155–3166.
29. Lee, S. Y., M. R. Wenk, Y. Kim, A. C. Nairn, and P. De Camilli. 2004.Regulation of synaptojanin 1 by cyclin-dependent kinase 5 at synapses. Proc.Natl. Acad. Sci. USA 101:546–551.
30. Legendre-Guillemin, V., M. Metzler, M. Charbonneau, L. Gan, V. Chopra,J. Philie, M. R. Hayden, and P. S. McPherson. 2002. HIP1 and HIP12 displaydifferential binding to F-actin, AP2, and clathrin. Identification of a novelinteraction with clathrin light chain. J. Biol. Chem. 277:19897–19904.
31. Lemmon, M. A. 2003. Phosphoinositide recognition domains. Traffic 4:201–213.
32. Lila, T., and D. G. Drubin. 1997. Evidence for physical and functionalinteractions among two Saccharomyces cerevisiae SH3 domain proteins, anadenylyl cyclase-associated protein and the actin cytoskeleton. Mol. Biol.Cell 8:367–385.
33. Longtine, M. S., A. McKenzie III, D. J. Demarini, N. G. Shah, A. Wach, A.Brachat, P. Philippsen, and J. R. Pringle. 1998. Additional modules forversatile and economical PCR-based gene deletion and modification in Sac-charomyces cerevisiae. Yeast 14:953–961.
34. Martin, T. F. 2001. PI(4,5)P2 regulation of surface membrane traffic. Curr.Opin. Cell Biol. 13:493–499.
35. McCann, R. O., and S. W. Craig. 1999. Functional genomic analysis revealsthe utility of the I/LWEQ module as a predictor of protein:actin interaction.Biochem. Biophys. Res. Commun. 266:135–140.
36. McCann, R. O., and S. W. Craig. 1997. The I/LWEQ module: a conservedsequence that signifies F-actin binding in functionally diverse proteins fromyeast to mammals. Proc. Natl. Acad. Sci. USA 94:5679–5684.
37. McPherson, P. S., E. P. Garcia, V. I. Slepnev, C. David, X. Zhang, D. Grabs,W. S. Sossin, R. Bauerfeind, Y. Nemoto, and P. De Camilli. 1996. A presyn-aptic inositol-5-phosphatase. Nature 379:353–357.
38. Metzler, M., V. Legendre-Guillemin, L. Gan, V. Chopra, A. Kwok, P. S.McPherson, and M. R. Hayden. 2001. HIP1 functions in clathrin-mediated
endocytosis through binding to clathrin and adaptor protein 2. J. Biol. Chem.276:39271–39276.
39. Ooms, L. M., B. K. McColl, F. Wiradjaja, A. P. Wijayaratnam, P. Gleeson,M. J. Gething, J. Sambrook, and C. A. Mitchell. 2000. The yeast inositolpolyphosphate 5-phosphatases Inp52p and Inp53p translocate to actinpatches following hyperosmotic stress: mechanism for regulating phosphati-dylinositol 4,5-bisphosphate at plasma membrane invaginations. Mol. Cell.Biol. 20:9376–9390.
39a.Robinson, J. S., D. J. Klionsky, L. M. Banta, and S. D. Emr. 1988. Proteinsorting in Saccharomyces cerevisiae: isolation of mutants defective in thedelivery and processing of multiple vacuolar hydrolases. Mol. Cell. Biol.8:4936–4948.
40. Rohde, G., D. Wenzel, and V. Haucke. 2002. A phosphatidylinositol (4,5)-bisphosphate binding site within �2-adaptin regulates clathrin-mediated en-docytosis. J. Cell Biol. 158:209–214.
41. Rusk, N., P. U. Le, S. Mariggio, G. Guay, C. Iurisci, I. R. Nabi, D. Corda, andM. Symons. 2003. Synaptojanin 2 functions at an early step of clathrin-mediated endocytosis. Curr. Biol. 13:659–663. (Erratum, 13:1746.)
42. Schuske, K. R., J. E. Richmond, D. S. Matthies, W. S. Davis, S. Runz, D. A.Rube, A. M. van der Bliek, and E. M. Jorgensen. 2003. Endophilin is re-quired for synaptic vesicle endocytosis by localizing synaptojanin. Neuron40:749–762.
43. Simonsen, A., A. E. Wurmser, S. D. Emr, and H. Stenmark. 2001. The roleof phosphoinositides in membrane transport. Curr. Opin. Cell Biol. 13:485–492.
44. Singer-Kruger, B., Y. Nemoto, L. Daniell, S. Ferro-Novick, and P. De Cam-illi. 1998. Synaptojanin family members are implicated in endocytic mem-brane traffic in yeast. J. Cell Sci. 111:3347–3356.
45. Slepnev, V. I., G. C. Ochoa, M. H. Butler, D. Grabs, and P. De Camilli. 1998.Role of phosphorylation in regulation of the assembly of endocytic coatcomplexes. Science 281:821–824.
46. Srinivasan, S., M. Seaman, Y. Nemoto, L. Daniell, S. F. Suchy, S. Emr, P. DeCamilli, and R. Nussbaum. 1997. Disruption of three phosphatidylinositol-polyphosphate 5-phosphatase genes from Saccharomyces cerevisiae results inpleiotropic abnormalities of vacuole morphology, cell shape, and osmo-homeostasis. Eur. J. Cell Biol. 74:350–360.
47. Stamenova, S. D., R. Dunn, A. S. Adler, and L. Hicke. 2004. The Rsp5ubiquitin ligase binds to and ubiquitinates members of the yeast CIN85-endophilin complex, Sla1-Rvs167. J. Biol. Chem. 279:16017–16025.
48. Stefan, C. J., A. Audhya, and S. D. Emr. 2002. The yeast synaptojanin-likeproteins control the cellular distribution of phosphatidylinositol (4,5)-bisphosphate. Mol. Biol. Cell 13:542–557.
49. Stolz, L. E., C. V. Huynh, J. Thorner, and J. D. York. 1998. Identification andcharacterization of an essential family of inositol polyphosphate 5-phospha-tases (INP51, INP52 and INP53 gene products) in the yeast Saccharomycescerevisiae. Genetics 148:1715–1729.
50. Toker, A. 2002. Phosphoinositides and signal transduction. Cell. Mol. LifeSci. 59:761–779.
51. Verstreken, P., T. W. Koh, K. L. Schulze, R. G. Zhai, P. R. Hiesinger, Y.Zhou, S. Q. Mehta, Y. Cao, J. Roos, and H. J. Bellen. 2003. Synaptojanin isrecruited by endophilin to promote synaptic vesicle uncoating. Neuron 40:733–748.
52. Wicky, S., S. Frischmuth, and B. Singer-Kruger. 2003. Bsp1p/Ypr171p is anadapter that directly links some synaptojanin family members to the corticalactin cytoskeleton in yeast. FEBS Lett. 537:35–41.
53. Yang, S., M. J. Cope, and D. G. Drubin. 1999. Sla2p is associated with theyeast cortical actin cytoskeleton via redundant localization signals. Mol. Biol.Cell 10:2265–2283.
54. Yin, H. L., and P. A. Janmey. 2003. Phosphoinositide regulation of the actincytoskeleton. Annu. Rev. Physiol. 65:761–789.
VOL. 25, 2005 Abp1 RECRUITS Sjl2 TO THE PLASMA MEMBRANE 2923
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 16
Nov
embe
r 20
21 b
y 39
.124
.117
.21.


















