
Maricela Paramo Santa Ana Watershed Association Riverside, CA
description

1
The paramo vegetation of the volcanicRuiz- Tolima massif*
La vegetación de páramo del macizo volcánicoRuiz- Tolima**
by/por
S. Salamanca, A.M. Cleef & J.O. Rangel Ch.
Dedicated to the memory of José Cuatrecasas Arumo.
Resumen
Este capítulo presenta una clasificación, en unidades sintaxonómicas, de los tipos de vegetación del páramo,de acuerdo con el método propuesto por la Escuela Ziirich-Montpellier que fue retrabajado posteriormentepor Westhoff & Van der Maarel (1973). Cleef (1981) aplicó por primera vez este método en Colombia. Elnivel jerárquico básico del presente estudio es la asociación, definida como una unidad de vegetación queconsiste en una comunidad de composición tlorística bien definida y de fisionomía uniforme y se encuentrabajo condiciones unifonnes de habitat. La asociación se define mediante una serie de levantamientos quetienen cierto número de especies en común (especies diagnóstico). El conjunto de datos analizados constade 34 levantamientos efectuados a lo largo del transecto Parque Los Nevados (levantamientos TPN), de71 levantamientos efectuados por A.M. C1eef en 1980 en colaboración con el primer autor (levantamientosAMe) y finalmente de 164 levantamientos realizados por el primer autor en 1980 (parte en conjunto conE.H. Kloosterman y C. Kloosterman, levantamientos SS) y en 1982. Estos datos se ordenaron en tablas deconstancia (Miiller Dombois & Ellenberg 1974) correspondiendo cada una a una formación vegetal diferente. Se establecieron grupos de levantamientos teniendo en cuenta las similaridades y disimilaridades encuanto a composición de especies. Con base en los grupos definidos se elaboró una tabla síntesis (tablaromana), en la cual la constancia de una determinada especie dentro de un grupo de levantamientos seexpresa en cinco clases (cada una del 20%) las cuales se designan mediante numerales romanos. Esta tablasíntesis (Tabla 1) que se ordenó mediante el método Twinspan (HillI979) muestra las afinidades tlorísticasentre los grupos y se utilizó para definir las unidades fitosociológicas finales. Para la nomenclatura de lossintaxa se empleó la metodología propuesta en la segunda edición del Código de Nomenclatura Fitosoci-
* Tables 1,2,3,4 and 6 on CD in the back ofthis volume.** Tablas 1,2,3,4 Y 6 en CD al final de este volumen.

2 The paramo vegetation
ológica (Barkman et al., 1986). En la definición de los tipos de vegetación se tnvieron en cuenta las clasificaciones disponibles de otras vegetaciones altoandinas y de páramo. Este estndio considera principalmente los tipos de vegetación del páramo propiamente dicho y el superpáramo. También se suministra unareferencia rápida a los bosques altoandinos que bordean los páramos. La distribución espacial esperada encintnrones altitndinales se encuentra un poco distorsionada debido al carácter volcánico del área. Kloosterman (capitnlo15, este volumen) provee una descripción detallada del paisaje en relación con la distribución espacial de la vegetación de páramo en el Parque Los Nevados.
A continuación se citan los sintaxa descritos (la mayoría descritos por Salamanca, Cleef & Rangel 1992)al interior de cada una de las seis formaciones vegetales principales:
Gynoxyo-Polylepidetum sericeae, Baccharido tricuneatae-Hypericetum laricifolii, y Pentacalietum vernicosi Cleef 1981. Además se tipificaron dos comunidades, de Gynoxys sp., y de Escallonia myrtilloidesAciachne acicularis dentro de los bosques enanos de páramo y los matorrales (Tabla 2). Calandrinio acaulis-Calamagrostietum rectae, Calamagrostietum ejJitso-rectae, Festuco dolichopihyllae-CalamagrostietumejJitsae y Espeletio hartwegianae-Calamagrostietum effusae dentro de los pajonales del páramo propiamante dicho (Tabla 3). Wernerietum crasso-humilis, Floscaldasio hypsophilae-Distichietum muscoides,Carici peucophilae-Wernerietum crassae y Gentianello dasyanthae-Plantaginetum rigidae, dentro de loscojines de plantas vasculares (Tabla 4). Muhlenbergietum cleejii Cleef 1981 corr. Cleef, Aciachnetum acicularis Vareschi 1953 corro Salamanca, Cleef & Rangel, y Agrostio breviculmis-Lachemilletum orbiculatae, dentro de los prados (Tabla 5). Cerastio floccosi-Pentacalietum gelidae, Elaphoglosso mathewsiiMuehlenbecldetum vulcanicae, Racomitrio crispuli-Lachemilletum nivalis, Thamnolio vermicularisRacomitrietum crispuli, Senecioni latiflori-Calamagrostietum ligulatae, y Senecioni canescentis-Cerastietum jloccosi, dentro de las comunidades subnivales del superpáramo (Tabla 6). Lupino alopecuroidesAgrostietum araucanae, Baccharido caespitosae-Agrostietum araucanae, y Diplostephio eriophoro-Loricarietum colombianae, dentro del pastizal azul del superpáramo (Tabla 7). La tabla sinóptica 4 muestralas interrelaciones entre los 31 sintaxa definidos para la vegetación de páramo del Parque Los Nevados.
En el presente estudio se describen también por primera vez algunos sintaxa de la vegetación zonal delpáramo del Parque Los Nevados a nivel de alianza, orden y clase. Son: Espeletio-Calamagrostietea rectoeffusae cl. nov., Calamagrostietalia recto-effusae ord. nov., Cerastio subspicatae-Calamagrostion rectaeal. nov., Festuco dolichophyllae-Calamagrostion effusae al. nov., Agrostiofoliatae-Cerastionfloccosae al.nov., y Festuco breviaristatae-Agrostion araucanae al. nov.
Finalmente se propone una zonificación altitndinal de la vegetación zonal para la palie alta del transectoTPN. En el flanco oeste del volcán de Santa Rosa se encuentra la secuencia más completa de la vegetaciónzonal del páramo. En el páramo propiamente dicho, los pajonales más extendidos son aquellos pelienecientes a la asociación Calandrinio acaulis-Calamagrostietum rectae . La distribución espacial de este tipode vegetación es disimétrica, pues alcanza altitndes un poco menores en los flancos exteriores de los volcanes. El límite superior de los pajonales de páramo propiamente dicho que pertenecen a la asociaciónCalamagrostietum effuso-rectae, se encuentra a una altitud mayor en la vertiente este del volcán de SantaRosa que en la vertiente oeste. Por otra parte, en la vertiente oeste del volcán de Santa Isabel estos pajonalesasí como los que pertenecen a la asociación Espeletio hartwegianae-Calamagrostietum effusae, se encuentran restringidos a una pequeña extensión, mientras que los pajonales azonales que pertenecen a la asociación Festuco dolichophyllae-Calamagrostietum effitsae, ocupan un área mayor.
El límite altitndinal entre el superpáramo alto y bajo es difuso. De 4400 m hacia arriba dos asociacionesque pertenecen a la vegetación subnival, el Senecioni canescentis-Cerastietumfloccosi y el Senecioni canescentis-Calamagrostietum ligulatae se encuentran en el mismo rango altih¡dinal o incluso a más bajaaltitud que algunas asociaciones como el Lupino alopecuroides-Agrostietum araucanae y el Diplostephioeriophoro-Loricarietum colombianae, que pertenecen al pastizal azul del superpáramo bajo. El Diplostephio eriophoro-Loricarietum colombianae se encuentra bastante extendido en la vertiente oeste del volcánde Santa Isabel (de 4350 a 4470 m) mientras que ocupa solamente una pequeña franja a ambos lados delvolcán de Santa Rosa.
La línea arbórea se encuentra alrededor de 3800 m en las vertientes externas tanto del volcán de Santa
Rosa como del de Santa Isabel. Por otra parte en las vertientes internas de los dos volcanes, las cualesterminan en el Lago de Otún a 3900 m, se encuentra una vegetación azonal de bosques enanos de páramopertenecientes a la comunidad de Aciachne acicularis-Escallonia myrtilloides.
En cuanto a la vegetación azonal, el Gentianello dasyanthae-Plantaginetum rigidae, es el tipo de vegetación más extendido dentro de las turberas de plantas en cojín. Se encuentra entre 3700 y 4200 m en la

La vegetación de páramo 3
vertiente este del volcán de Santa Isabel y entre 4100 y 4200 m en la vertiente este del volcán de SantaRosa. El Floscaldasio hypsophilae-Distichietum muscoides, mucho más restringido en extensión, se encuentra a 4150 m en el lado oeste del volcán de Santa Rosa y a 4350 m en el lado este del volcán de SantaIsabel, cerca de la transición con el superpáramo. En la vertiente oeste del volcán de Santa Isabel a 4200m, se encuentran pequeños parches de turberas pertenecientes al Wernerietum humilis var. de Distichiamuscoides. Por otra parte el Carici peucophilae-Wernerietum crassae se registró a 3800 m en la vertienteeste del volcán de Santa Isabel.
A 4300 m se registró un bosque enano perteneciente al Gynoxyo-Polylepidetum sericeae en la vertienteoeste del volcán de Santa Isabel. Entre 3830 y 3900 m se encuentran bosques enanos del Baccharidotricuneatae-Hypericetum laricifolii en la vertiente oeste del volcán de Santa Isabel. Por otra parte el Pentacalietum vernicosi se registró a 4340 m también en la vertiente oeste del volcán de Santa Isabel y a 4350m en la vertiente este del mismo.
Los prados azonales pertenecientes a la asociación Muhlenbergietum cleefii se registraron solamente enun levantamiento en la márgen oeste del Lago de Otún a 4000 m. El Aciachnetum acicularis subass. lupinetosum microphylli, se registró en la vertiente este del volcán de Santa Rosa a 4050 m.
En la vertiente oeste del volcán de Santa Isabel los tipos de vegetación más altos pertenecen a dosasociaciones diferentes de la vegetación subnival en el superpáramo alto. El primero registrado a 4540 mes el Senecioni canescentis-Cerastietum./loccosi. Un poco más abajo, a 4400 m se encuentra la asociaciónSenecioni latiflori-Calamagrostietum ligulatae. El Diplostephio eriophoro-Loricarietum colombianae,perteneciente al pastizal azul del superpáramo bajo se encuentra a 4450 m. Entre 4350 y 4400 m se encuentra otra asociación perteneciente al pastizal azul del superpáramo bajo, el Lupino alopecuroidesAgrostietum araucanae. La transición al páramo propiamente dicho se encuentra alrededor de 4400 m.Entre esta altitud y 4150 m se encuentran los pajonales del páramo propiamente dicho pertenecientes a laasociación Calandrinio acaulis-Calamagrostietum recta e . A 4140 m se encuentran los pajonales azonalesdel Festuco dolichophyllae-Calamagrostietum effusae.
En la vertiente este del volcán de Santa Isabel el tipo de vegetación más alto (a 4475 m) pertenece alLupino alopecuroides-Agrostietum araucanae. Esta vegetación se extiende hasta 4400 m. La transición alpáramo propiamente dicho se encuentra alrededor de 4350 m. Los pajonales del Calandrinio acaulis-Calamagrostietum rectae, del páramo propiamente dicho alto se encuentran entre 4125 y 4315 m. En estemismo rango altitudinal también se encuentran algunos pajonales pertenecientes al Calamagros(ietum e{filso-rectae y al Espeletio hartwegianae-Calamagrostietum effusae.
El tipo más alto de vegetación registrado en la vertiente oeste del volcán de Santa Rosa (a 4550 m)pertenece a la asociación Lupino alopecuroides-Agrostietum araucanae. Entre 4390 y 4460 m se encuentrael pastizal azul del Diplostephio eriophoro-Loricarietum colombianae. La transición al páramo propiamente dicho se encuentra alrededor de 4400 m. Entre 4250 y 4330 m se encuentran los pajonales delCalandrinio acaulis-Calamagrostietum rectae. Entre 4050 y 4175 m se encuentran los pajonales del Calamagrostietum effilso-rectae. El pajonal azonal perteneciente a la asociación Festuco dolichophyllae-Calamagrostietum effusae, se encuentra entre 3850 y 3960 m.
En la vertiente este del volcán de Santa Rosa la vegetación más alta pertenece a la asociación Senecionicanescentis-Cerastietum./loccosi, de la vegetación subnival y se encuentra a 4400 m. A esta misma altitude incluso unos metros por encima, se encuentran el Lupino alopecuroides-Agrostietum araucanae, y elDiplostephio eriophoro-Loricarietum colombianae, ambos del pastizal azul. Los primeros pajonales delpáramo propiamente dicho, pertenecientes al Calandrinio acaulis-Calamagrostietum rectae, se encuentrantambién a 4400 m, y descienden hasta 4200 m.
Introduction and methods
The word "paramo" has been commonly used since the Spanish Conquest. For the conquerorsthose lands, high up in the mountains ofthe New World, had many similarities with the remotehigh areas in the mountains ofthe Iberic Peninsula. In northern Peru, the paramo is called "jalea".In the rest of Peru, Bolivia, northern Argentina and Chile the dry open vegetation type abovethe upper forest line is termed "puna" in the Quechua language.
Although in recent European and American literature the open vegetation above the upperforest hne is often referred to by "tropical alpine", because ofthe equatorial Andean setting, the

4 The paramo vegetation
Table 1. 8ynoptic table showing the relationship among the different paramo syntaxa (on CD inthe back of this volume).- Tabla sinóptica que muestra las relaciones entre los diferentes sintaxa del páramo (en CD alfinal de este volumen).
A 1 Lupino alopecuroides-Agrostietum araucanae ass. nov.A2 Baccharido caespitosae-Agrostietum araucanae ass. nov.A3 Diplostephio eriophori-Loricarietum colombiana e ass. nov.81a Calandrinio acaulis-Calamagrostietum recta e ass. nov. varo typicum varo nov.81 b Calandrinio acaulis-Calamagrostietum rectae ass. nov. varo of Espeletia hartwegiana
ssp. centroandina varo nov.82 Calamagrostietum effuso-rectae ass. nov.83 Festuco dolichophyllae-Calamagrostietum effusae ass. nov.84 Espeletio hartwegianae-Calamagrostietum recta e ass. nov.F1 Gynoxyo-Polylepidetum sericeae ass. nov.F2 Dwarf forests (bosques enanos) of Gynoxys sp.F3 Dwarf forests (bosques enanos) of Aciachne pulvinata and Escallonia myrtilloidesF4 Baccharido tricuneatae-Hypericetum laricifolii ass. nov.F5 Pentacalietum vernicosi Cleef 1981P 1 Muhlenbergietum cleefii Cleef 1981P2a Aciachnetum acicularis subass. typicum subass. nov.P2b Aciachnetum acicularis subass. lupinetosum microphyllae subass. nov.P3 Agrostio breviculrnis-Lachemilletum orbiculatae Cleef 198181a Cerastio f1occosi-Pentacalietum gelidae ass. nov. varo inops varo nov.81 b Cerastio f1occosi-Pentacalietum gelidae ass. nov. varo ofStereocaulon vesuvianum varo
nov.82 Elaphoglosso mathewsii-Muhlenbeckietum vulcanicae ass. nov.83 Racomitrio crispuli-Lachemilletum nivalis ass. nov.84 Thamnolio vermicularis-Racomitrietum crispuli ass. nov.85 Senecio latiflori-Calamagrostietum ligulatae ass. nov.86 Senecio canescentis-Cerastietum f1occosi ass. nov.87 8ubnival vegetation complex (complejo de vegetación subnival)T1a Wernerietum humilis ass. nov. varo of Hypochoeris sessiliflora varo nov.T1 b Wernerietum humilis ass. nov. varo of Distichia muscoides varo nov.
T2 Floscaldasio hypsophilae-Distichietum muscoides Cleef 1981T3 Carici peucophilae-Wernerietum crassae Cleef 1981 var of Cotula mexicana varo nov.T4a Gentianello dasyanthae-Plantaginetum rigidae ass. nov. subass. typicum subass. nov.T4b Gentianello dasyanthae-Plantaginetum rigidae ass. nov. subass. brometosum lanatae
subass. nov.
word "paramo" is preferred here. Moreover, the word "paramo" is gaining wider use and refersexactly to the equatorial cold and humid high mountain vegetation of South and Central America,which is largely derived from the tropical Andean stock.
The aim ofthis chapter is to provide a classification, in syntaxonomical units, ofthe vegetationtypes of the paramos in the Ruiz- Tolima volcanic massif according to the principIes and methodofthe phytosociological school ofZurich-Montpellier (Westhoff & Van der MaareI1973). Ihismethod was first applied, and detailed, in Colombia for the paramo vegetation (Cleef 1981). Thebasic hierarchicallevel used is the 'association'. Ihis is defined as a vegetation unit consistingof a community within an uniform environment with well-defined floristic composition and anuniform physiognomy. The association is defined by one or more relevés, which share a numberof character species (i. e. diagnostic species). A member of an association should be recognizablein the field by the presence of a specific group of diagnostic species.

La vegetación de páramo 5
The paramo vegetation data set analyzed consists of 34 relevés made in 1980 along the ParqueLos Nevados transect (TPN relevés; data are presented in the appendix after chapter 3), 71relevés made by A.M. Cleef(AMC relevé s), mostIy in collaboration with the first author in 1980and, finally, 164 relevé s made by the first author in 1980 in part together with E.H. Kloostermanand C. Boonen and in 1982 (SS relevés) . First of all this data set was ordered into constancytables (sensu Miiller Dombois & Ellenberg 1974), one per vegetation formation. Relevé groupswere established on the similarities in species campo sitian using TWINSP AN (Hill1979). Basedon al! the defined groups, a summary table was constructed, where the constancy of a givenspecies within a group ofrelevés is expressed in five c1asses (each 20%) designated by Romannumerals. This summary table (Table 1), which was ordered with TWINSPAN, shows the floristic affinities between the groups and was used to define the final phytosociological units. Forselecting the names of the syntaxa, the methodology proposed in the second edition of the Codeof Phytosociological Nomenc1ature by Barkman et al. (1986) was followed. With regard to thec1assification of the Ruiz- Tolima paramo vegetation types, the c1assifications already availablefrom paramo and high Andean vegetation elsewhere were taken into account.
The first text (in English) of the present study was part of a photocopied manuscript, whichwas used by the first author for the defence ofher Ph.D. thesis at the University of Amsterdam(Salamanca 1991). Later this study was translated in Spanish (Salamanca 1992). We considerthe printed version of Salamanca (1992), and especially chapter 4 (Salamanca, Cleef & Rangel1992) as the formal reference publication for the validation of the different syntaxa described.Complementary to earlier editions ofSalamanca (1991, 1992), we have updated the informationand corrected taxonomical plant names, as far as necessary, with reference to the earlier published syntaxa. In addition we have described in this study also a number of syntaxa above thelevel ofthe association (alliance, order, c1ass), especially for the zonal paramo vegetation.
This study focuses on the zonal, and most common azonal, vegetation types within thegrassparamo and the superparamo. A quick reference to the uppermost high Andean forests,bordering the paramo, is also provided. The expected spatial distribution of the vegetation inaltitudinal belts is somewhat distorted by the volcanic character of the area. Kloosterman et al.(this volume) provide a detailed description of the landscape in relation to the spatial distributionof the paramo vegetation of the Parque Los Nevados.
As to the correct spelling of the names of the plant species reference can be made to Luteyn(1999). We did not distinguish between Werneria Kunth and Xenophyllum V.A. Funk in thisstudy and maintained all species belonging to both genera under Werneria.
The paramo concept
Several papers dealing with physiognomic and floristic descriptions, as well as with phytosociological classifications, of the paramo vegetation have been published. The first modern studieswere published by Cuatrecasas (1934,1954,1958,1968,1979,1980). Cleef(e.g. 1978, 1979a,1979b, 1980a, 1980b, 1980c) studied the general physiognomy and the altitudinal sequence ofthe paramo vegetation, as well as the campo sitian and the phytogeographical position of thevascular flora of the paramos of the Colombian Cordillera Oriental. Cleef (1981) published arather complete phytosociological study on the paramo vegetation of the Colombian CordilleraOriental. In the case of the vegetation of the Parque Los Nevados Cleef et al. (1983) providedthe first approach (after Cuatrecasas 1934) on the reconaissance ofthe paramo and Andean forestvegetation types as well as information on the altitudinal zonation of the Parque Los Nevadosvegetation. In a similar study, Cleef & Rangel (1984) c1assified the paramo vegetation along theBuritaca transect, located in the Sierra Nevada de Santa Marta. Rangel & Franco (1985), Sturm& Rangel (1985), Rangel & Lozano (1986), Franco et al. (1986), Duque & Rangel (1989),Sánchez & Rangel (1990), Restrepo & Duque (1992) and Becking et al. (1997) also provided

6 The paramo vegeta tia n
phytosociological studies ofthe paramo vegetation in different localities ofthe three ColombianCordilleras. Studies concerning the structure, biomass, productivity and nutrient status ofbunchgrass paramos in the Eastern Cordillera are found in Lutz & Vader (1987) and Beekman &Verweij (1987). For the volcanic Central Cordillera more recently also in Hofstede (1995) forthe Parque Los Nevados area. Ferwerda (1987) and Van Groen (1987) discussed the influenceof agriculture and burning on natural bunchgrass paramos in the Eastern Cordillera. Verweij(1995) studied patterns ofburning and grazing for the Los Nevados National Park. A vegetationmap based on photo interpretation and field control of the Parque Los Nevados is presented inKloosterman et al. (chapter 5, this volume).
Other studies concerning different types ofparamo vegetations have been published for northern Ecuador, Peru, Venezuela, Panamá and Costa Rica. See for references Luteyn (1999).
In the present study, the concepts of zonal and azonal vegetation follow Walter (1979). Vegetation types mainly depending on the macroclimate are considered as zonal, while others, whichstrongly depend on local edaphic factors, are considered as azonal. However, it was not alwayseasy to decide whether some vegetation types should be ranked as zonal or azonal; see Cleef(1981) for further details.
Altitudinal zonation and physiognomy
Cuatrecasas (1958, 1968) described three major zones ofparamo vegetation in the ColombianAndes, which can be distinguished by means of their physiognomy and floristics. These zonesare; the subparamo, including scrub paramo, just above the upper forest line; the paramo proper,or grassparamo, and the superparamo, just below the ice cap. Furthermore, Cleef (1979a, 1980b,1981) subdivided each of these zones in a higher and a lower subzone for the Colombian EasternCordillera.
This altitudinal zonation is although valid for the Colombian Eastem Cordillera. In the RuizTolima volcanic massif of the Central Cordillera no proper shrubby subparamo vegetation ispresent, except perhaps for the area North ofthe Tolima Volcano. In this area Cuatrecasas (1934)described two types of shrubby vegetation referred to as 'Vaccinietum floribundi' (3500-3600m) and 'Espeletietum hartwegianae-Hypericeti laricifoliosum' (3450-3600 (3800) m (see Cuatrecasas, chapter 4, this volume). Cleef et al. (1983) and Van der Hammen & Cleef (1986),concluded that the near absence of a subparamo belt in this vo1canic massif can be attributed tothe absence of a number oftaxa which characterize the subparamo zone in the Colombian EastemCordillera.
In the Parque Los Nevados, patches of high Andean dwarf forests are found between 3600 to3900 (3800) m. The grassparamo is situated from 3800 (3900) to 4400 (4300) m and the superparamo occurs from 4300 to 4800 m. The nival zone starts around 4800 m, albeit with manyexceptions due to local circumstances. See also the vegetation map ofKloosterman et al. (Chapter 5, this volume). Elsewhere in the Colombian Andes, e.g. Sierra Nevada de Santa Marta (Cleef& RangeI1984), Westem Cordillera (Cleef et al. in prep.) and near Popayán (Becking et a1.l997)subparamo scrub is patchy. This apparently results from the steep slopes and small ridges.
The Venezuelan paramos cover an altitudinal range from 3000 to 4800 m (Monasterio 1980).The paramo region is subdivided into two altitudinal zones; the transition being place at 4000m. The lower zone, 'Paramo Andino' broadly corresponds with the Colombian subparamo andgrassparamo. The higher zone ('Páramo desértico' and 'Desierto Periglacial') broadly correspond to the superparamo belt. Vegetation types and tife forms of the upper grassparamo andsuperparamo of the Mérida Andes have recently been studied by Berg (1998). Shrubs of Ericaceae and/or Compositae in contact with the open paramo vegetation, which could be regardedas a kind of subparamo vegetation, have also been described for Venezuela (Monasterio, f.c.;Vareschi 1980).

La vegetación de páramo 7
Harling (1979) defined three altitudinal paramo vegetation belts for Ecuador; e.g. grassparamo(3400-4000 m), shrub and cushion paramo (4000-4500 m) and the desert paramo (mainly above4500 m). Acosta-Solís (1984) established the following altitudinal zones for the Ecuadorianparamos: the low paramo from 3300 m to 4000 m; the "middle paramo" from 4000 to 4500 m,and the high paramo from 4500 m upwards. This corresponds rather well with the divisionproposed by Cuatrecasas (l.c.) for the Colombian paramos. A narrow belt (comparable with whatHarling (1979) and Ramsay (1992) described as cushion paramo for Ecuador), where the tussocks of Poaceae are intermingled with vascular cushion plants, is present around 4000-4100m on the West slope of the Ruiz Voleano. The zonal paramo vegetation of Ecuador has beenstudied by Ramsay (1992).
High andean forests
The high Andean forests are the altitudinally highest forests in the Andes. They are found alongthe upper forest line and locally as isolated stands in the paramo belt. Grubb (1977) differentiatedthese forests, consisting of small-sized trees, as subalpine rain forests (SARF), occupying a propel' altitudinal position as compared to the upper montane rain forests (UMRF). However, in ouropinion, they belong to the upper montane rain forest belt, differing only in stature, speciescomposition (fewer species), and slower nutrient/water cycling processes because oflower meanannual temperature. See for references Grubb (1977), Veneklaas (1990), Bruynzeel & Veneklaas(1998), Cavelier (1996), Marrs et al. (1988) and van del' Hammen (final chapter, this volume).
The upper forest limit, even under equatorial-montane conditions, is not constant. Also, itoften has been subject to strong human influence. As a result, the distributions of forest andparamo vegetation intermingle. For example, in the TPN transect area the altitudinal variationis between 3600 and 4000 m.
Paleoecological studies have shown that some 5000-6000 years B.P. the forest line reacheda couple ofhundred meters higher than today (Van del' Hammen & González 1960, 1965, Vandel' Hammen et al. 1973; Van del' Hammen 1974, 1979; Kuhry et al. 1983; Melief 1985; Salomons 1986). They have also shown the forest line to occur much lower, around 2000 m, duringthe Late Pleniglacial.
The present uppermost forest limit is situated at c. 3800 m in the Ruiz- Tolima massif. Paleoecological studies in the El Bosque locality (Kuhry et al. 1983; Kuhry 1988) have shown nochanges in this position during the last 3000 years. Nevertheless, the uppermost forests in thearea today show a strong human impact and have been partially destroyed (see also Verweij &Beukema 1992).
Monasterio (1980), considering the patchy distribution of the forest limit in Venezuela, surmises that some ofthe forests which are found within the borders ofthe paramo are high Andeanforest refuges. The withdrawal downslope ofthe forests is, according to her, due to competitionof diverse species of Espeletia for the same habitats.
Regarding todays presence of puna vegetation in the Peruvian and Bolivian altiplanos, Ellenberg (1979) suggests that the patchy distribution of Polylepis and Escallonia woodland abovethe present timberline, highlights the ability offorest vegetation to grow beyond their recognizedenvironmentallimit. Therefore, they are considered as relicts of more favourable past climaticconditions. Indeed Ellenberg (1979) asserts that paramo and puna grasslands cannot be regardedas climax vegetation, but are caused by deforestation, burning and grazing. Detailed discussionon this account is given by Holdridge (1947), Weberbauer (1945), Ellenberg (1959,1979), Tosi(1960), Koepcke (1961), Scott (1974), Unzueta (1975) and Beck & Ellenberg (1977). Recently,Laegaard (1992) also suggested that the lower paramo belt in Ecuador largely results from deforestation of the high Andean Polylepis forest. The paramo-like bunchgrasslands replace upperAndean forests. In OUT study area this replacement also might have occurred in some local patch-

8 The paramo vegetation
es. Near the forest line, patches ofparamo bunchgrasslands, dominated by Festuca dolichophylla, might have presumably supported a high Andean forest, in earlier days. The so-called "socaló" agrosystem, facilitating access of cattle to the forest by selective felling and by stimulatinggrawth of the herb layer, might have also contributed to influencing the altitudinal position ofthe upper forest line replacing the open patches oftrees by grazed, open vegetation (Verweij &Beukema 1992; Kok et al. 1995).
The high Andean forests, just below the paramo belt in the study area, originally classifiedas the Hesperomeletum lanuginosae (Cuatrecasas 1958) are found on the southern slope of theTolima Volcano, falllargely within the altitudinal range of 3300-3800 m (see also Chapters2 and 4 this volume). Among the trees which characterize these forests are Hesperomeles lanuginosa, Miconia salicifolia, Monnina revoluta, Solanum bogotense, Diplostephium revolutum,Myrsine dependens and Hedyosmum huilense. Intermingled with these forests, Cuatrecasas(1954,1958) also reported within the altitudinal range of3300-3400 m forests with Weinmanniatolimensis and W hirtella, as well as forests dominated by Gynoxys spp., and Pentacalia spp.There are some notable differences regarding the physiognomy as well as the floristic composition between the forests of the re1atively dry east slope and the more humid west slope of thestudy area. Cleef et al. (chapter 2, this volume) and Rangel et al. (chapter 3, this volume) dealin more detail with the phytosociology of the montane forests for this part of the Central Cordillera. The third author added transect studies on montane rain forests in the deep río Otúnvalley just south ofthe TPN transect on the west slope ofthe Central Cordillera (Rangel 1994).
Paramo dwarf forests
Cleef (1981) described dwarf and scrub forests of Aragoa abietina, Diplostephium revolutum,Hypericum spp., Pentacalia andicola, P. nitida, P. vernicosa, and many others, in the paramosof the Colombian Eastern Cordillera. Monasterio (1980) described paramo "forests" composedof Libanothamnus neriifolia, Aragoa cupressina and Alnus acuminata, in Venezuela. Valencia& Jorgensen (1992) studied the composition and structure of humid montane forests with Miconia theaezans, M. pustulata and Piper andreanum close to Quito, Ecuador.
In the Parque Los Nevados area rather open and scrubby paramo dwarf forests with Gynoxystolimensis and other Gynoxys spp., Polylepis sericea and Escallonia myrtilloides form an azonalgraup separated fram the high Andean forests since they clearly occur 'extrazonally' within theparamo belt. There they form mosaics with the lower bunchgrass paramo vegetation Those aremostly found along streams and in places where the water table is near the surface between 3600m and 3900 m. Small patches of Polylepis forests are found in sheltered place s as high as 4300m, mainly on scree slopes associated with the steep walls of the lava escarpments. Compared tothe uppermost contiguous high Andean forests extrazonal paramo dwarf forests contain fewerforest species, but also contain paramo species which are less present in the forest line stands.Occasionally, small patches of dwarf forest of Diplostephium revolutum on the west slope ofthe Cisne V olcano and of Pentacalia vaccinioides in the Quebrada Africa valley have beenobserved.
Syntaxonomy of the paramo dwarf forests
Gynoxyo-Polylepidetum sericeae Salamanca, Cleef & Rangel 1992Gynoxys-Polylepis serieea dwarfforestBosque enano altoandino eon Gynoxys y Polylepis serieeaType relevé AMC 623. Table 2; Vol. 1, Photo 28, 29; this volume Photo 1,2,3

La vegetación de páramo 9
Photo 1. Interior of the open dwarf forest of Polylepis sericea (Gynoxyo-Polylepidetum sericeae) at 4300m on the western side of the Alto La Línea (Province Risaralda). In the understorey is a low shrub of Pentacalia vernicosa. This is associated with species of Diplostephium and Gynoxys. Ground rosettes of Valeriana plantaginea are predominant together with bunchgrass. Small moss cushions of Chorisodontiummittenii are conspicuous on the trunks (relevé Cleef et al. 623, type locality for this association).
- Interior del bosque abierto de Polylepis sericea (Polylepidetum sericeae) a 4300 m en el lado occidentaldel Alto La Línea (Dpto. Risaralda). En el sotobosque hay arbustos de Pentacalia vernicosa asociados conespecies de Diplostephium y Gynoxys. Predominan las rosetas acaules de Valeriana plantaginea junto conmacoJlas de gramíneas. Las almohadillas de Chorisodontium mittenii son visibles sobre los troncos de Polylepis (Iev. Cleef et al. 623, localidad tipo de la asociación).
Physiognomy:These dwarfforests are the most common ones in the paramos ofthe Ruiz-Tolima massif. Theyconsist of;
tree 1ayer, about 5-10 m high, covering from 30% to 100%,shrub 1ayer, 1-2 m high, either rather sparse (8%) or re1ative1y dense (20%),herb 1ayer, c. 50 cm high, covering from 2% to 75%,ground 1ayer covering from 2% to 95%, with redominance ofbryophytes anci/or lichens.
Composition and syntaxonomy:C1eef et al. (1983) provided a short description of Polylepis serie ea forests ofthe Ruiz- Tolimamassif. The Gynoxyo-Polylepidetum serieeae is re1ative1y poor in vascu1ar p1ants. The tree 1ayerconsists of Polylepis serieea and Gynoxys sp., the 1atter - probab1y G. tolimensis - is rare1ypresent.

10 The paramo vegetation
Photo 2. Canopy aspect ofthe Polylepis serieea dwarfforest (Gynoxyo-Polylepidetum serieeae) ofphoto1. Dead trunks of Polylepis serie ea are still covered by epiphytes, such as Chorisodontium mittenii (Musci)and Parmeliaceae (Lichens).
- Aspecto de las copas del bosque de Polylepis serieea (Gynoxyo-Polylepidetum serieeae) de la foto 1.Además se observaron troncos muertos de Polylepis serieea todavia con epífitas de Chorisodontium mittenii (musgos) y Parmeliaceae (líquenes).
Polylepis sericea is a rosaceous tree, belonging to the section Sanguisorbeae. More than 15species have been reported (Simpson 1979). The genus is endemic ofthe tropical Andes, occurring from the northern part of Argentina to Venezuela. Although in Colombia there are twospecies, in the Ruiz-Tolima massif only Polylepis sericea dwarf forests have been found. According to Simpson (l.c.), Polylepis sericea occurs from the Mérida Andes of Venezuela in thenorth, to the Titicaca Lake (Bolivia) in the south.
The presence of Gynoxys div. spp., within Polylepis forests is well known (Cleef 1981; Arnal1983) from the Eastern Cordillera (Colombia) and the Sierra Nevada de Mérida (Venezuela).Species in common between the Polylepis forests of the Colombian Central and the EasternCordilleras are Calamagrostis recta, Festuca sublimis, Pentacalia vernicosa and of the mossesare: Campylopus spp., and Pleurozium schreberi, all occurring in the ground layer. 1t is not c1earyet if the Venezuelan Gynoxys-Polylepis sericea forests also belong to the Gynoxyo-Polylepidetum sericeae.
Ecology and distribution:Dwarfforests belonging to the Gynoxyo-Polylepidetum sericeae occur in the study area between4000 and 4200 m. These forests are found on damp soil, along paramo ravines or in shelteredplaces among boulders on scree slopes.
Azocar & Monasterio (1979) studied the mesoc1imate in Polylepis sericea forest in the Mucubají pararno in the Venezuelan Andes. They noted the large amplitude of thermic and hydricconditions characteristic of the habitats of Polylepis sericea forests, present throughout the paramo belt in Venezuela from 3600 to 4300 m (ArnaI1983).
Walter & Medina (1969) discussed the presence of Polylepis sericea forests in scree slopesin the Venezuelan Andes.

La vegetación de páramo 11
Photo 3. Detail of stem of Polylepis sericea with epiphytic Chorisodontium mittenii (Musci) of photo 1.
- Detalle del tronco de Polylepis sericea con musgos epífitos de Chorisodontium mittenii del sitio de lafoto grafía 1.
Cleef (1981) summarized phytosociological traits of Polylepis quadrijuga forests, occurringbetween 3735 and 4025 m in the Guantiva, La Rusia and El Cocuy paramos fram the ColombianEastern Cordillera. In the Paramo de Chingaza Velez et al. (1998) studied the high productivityof a stand of these rasaceous dwarf forests.
Dwarfforests of Gynoxys sp. (G. tolimensis?)Bosque enano altoandino de Gynoxys (G. tolimensis?)Table 2
Composition and syntaxonomy:These dwarf forests are relatively rich in species. The tree layer is dominated by 2-5 m tallGynoxys sp. (G. tolimensis ?). Saracha (syn.Acnistus) quitensis andHypericum laricifolium occur often. Diagnostic species include Carex jamesonii, Calceolaria pentlandii, Cortaderia sericantha and Gynoxys tolimensis, Ribes sp., Valeriana pavonii, Myrrhidendron glaucescens isassociated.

12 The parama vegetatian
These Gynoxys dwarfforests also have a number af species, e.g. Saracha (Acnistus) quitensis,Myrrhidendron glaucescens, Hypericum laricifolium and Ribes sp., in common with the dwarfforests dominated by Gynoxys div. spp. studied by Cleef (1981) and T. van der Hammen andR. Jaramillo M. (unpubl.) in the Eastern Cordillera of Colombia.
Ecology and distribution:The Gynoxys tolimensis dwarf forests are mostly found in sheltered rock outcrops (covering asurface up to 20 m2) at high altitude. The relevés which confonn the basis for this descriptionwere made between 3960 and 4200 ID in the Ruiz-Tolima study area. The highest record is from4400 m. Gynoxys sp. (G. tolimensis?) has been reported down to 3000 m in the Cordillera Centralof Colombia in the Natural reserves of Puracé and Los Nevados.
Aciachne acicularis-Escallonia myrtilloides dwarf forestsBosque enano de Escallonia myrtilloides con Aciachne acicularisTable 2; Figs. 1, 2; Photo 4
TPN 13975 m
Fig. 1. Physiognomy and structure of Aciachne acicularis-Escallonia myrtilloides dwarf forest (TPN 1,3975 m) with strong evidence of grazing and sheltering by cattle. Moraine west ofLaguna OtÚn, Risaraldaprovince. See Photo 4.
- Fisionomía y estructura del bosque enano de Aciachne acicularis-Escallonia myrtilloides (TPN 1, 3975m) con evidencia fuerte de pastoreo y pisoteo de ganado. Morrena al oeste de la Laguna Otún, Dpto. de Risaralda. Véase Foto 4.

La vegetación de páramo
TPN73930 m
1 m
13
Fig. 2. Physiognomy and structure of Aciachne acicularis-Escallonia myrtilloides dwarf forest with columnar Espeletia hartwegiana (relevé TPN 7, 3930 m) in the grass paramo on the soutwest edge ofLagunaOtún, Risaralda province.
- Fisionomía y estructura de matorral bajo de Aciachne acicularis-Escallonia myrtilloides con frailejonesaltos de Espeletia hartwegiana (lev. TPN 7, 3930 m) en el páramo propiamente dicho en la márgen suroccidente de la Laguna Otún, Dpto. de Risaralda.
Physiognomy:The description of these forests is based on two relevé s (TPN 1 and TPN 7). The tree layerattains a cover of only 60% and 75%, and is exdusively composed of Escallonia myrtilloides.Calamagrostis recta constitutes the tussock layer, with a cover of 10%. In the low, and open,herb layer, light demanding Aciachne acicularis has a cover ofbetween 4 % and 12%. The coverof terrestric bryophytes is c. 30%. In TPN 7 Espeletia hartwegiana is present with a cover of25 %. The presence of Aciachne acicularis seems related with grazing.
Composition:The vegetation sample has been based only on two relevés near the Laguna de Otún, and thereforthis vegetation type has not been ranked according to the Ziirich-Montpellier approach. Differential is the cover of Geranium sibbaldioides and Aciachne acicularis. Characteristic taxa are
also Gentianella dasyantha, Hypericum lancioides, Lachemilla holosericea and Stellaria cuspidata, among others. The influence on the vegetation of grazing is strongly induced by Aciachneacicularis, Agrostis haenkeana, Baccharis genistelloides, Eryngium humile, Lachemilla orbic~tlata, Rumex acetosella and Veronica serpyllifolia (Verweij & Budde 1992). Laegaard (1987)studied Aciachne in the tropical Andes and described most Colombian records under Aciachneacicularis Laegaard. In the past, Colombian Aciachne was referred to as A. pulvinata Bentham.

14 The paramo vegetation
Photo 4. Remnants of an Aciachne acicularis-Escallonia myrtilloides dwarf forest at 3975 m on the western edge ofthe Otún lake (Risaralda province). Heavy grazing in the past has resulted in the open herb layertoday comprised of Agrostis cf. haenkeana (relevé TPN I and relevé Cleef & Salamanca 578). See Fig. l.
- Remanentes de un bosque enano paramuno de Aciachne acicularis-Escallonia myrtilloides a 3975 msnmcerca de la orilla W de la Laguna de Otún (Dpto. de Risaralda). Como consecuencia de la influencia delpastoreo en el pasado el estrato ralo de hierbas está conformado por Agrostis cf. haenkeana (lev. TPN I andlev. Cleef & Salamanca 578). Véase Fig. 1.
Ecology and distribution:The ecology of this type of forest is not well understood yet. Escallonia scrub and dwarf forestpatches have developed on old block lava flows as well as on morainic terrain near Lake atún.Locally, Escallonia myrtilloides is associated with dense stands of stem rasettes of Espeletiahartwegiana. The aspect and composition of the ground ¡ayer, especially on top of the lateralmoraine, has been changed by animal grazing and droppings. AIso, fuel wood gathering tookplace causing an impact on Escallonia treelets. These stunted Escallonia forests were sampledat 3910 and 3930 m in the vicinity ofLake atún (see Photo 148). Escallonia myrtilloides dwarfforests have also been reported in the surroundings of Popayán (Restrepo & Duque 1992) andfram the Cordillera Oriental of Colombia (Cleef 1981).
Baccharido tricuneatae ~ Hypericetum laricifolii Salamanca, Cleef & Rangel 1992Baccharis tricuneata - Hypericum laricifolium shrubMatorral de Hypericum laricifolium con Baccharis tricuneataType: relevé AMC 635. Table 2
Physiognomy:Three layers compose the structure of these tall scrub stands;
scrub layer 2-3 m high, with a canopy cover fram 5% to 55%,medium high to high herb layer, mainly of bunchgrasses (covering fram 3 % to 30%),dense graund layer, consisting mostIy ofbryophytes (covering fram 10% to 45%).

La vegetación de páramo 15
In some forests a low herb layer is rather conspicuous, reaching covers around 25 %.Ihis community is in fact an azonal paramo scrub association for the sake of convenience
and tloristic similarity included in the paramo dwarf forest table.
Composition and syntaxonomy:For the Colombian Eastem Cordillera, Cleef (1981) described the Hypericetum laricifolii as anew association that included Hypericum laricifolium fa. laricoides as the dominant diagnosticspecies. In the association described here fa. typicum of Hypericum laricifolium is dominant(Robson 1987).
Cerastium subspicatum, Geranium sibbaldioides, and Pernettya prostrata are character species in cOllllllon with the Hypericetum larici{olii ofthe Eastern Cordillera and the shrub of Baccharido tricuneatae-Hypericetum laricifolii. Baccharis tricuneata, Conyza uliginosa, LupinushumifÚsus andMuehlenbeckia vulcanica are diagnostic for Baccharido tricuneatae-Hypericetumlaricifolii of the Central Cordillera.
Ecology and distribution:According to Robson (J 987) Hypericum laricifolium is present from central Peru to Venezuela.In the Colombian Andes, the distribution range is rather wide; Cleef (1981) reports Hypericumlaricifolium communities occuring as low as 3580 m; other species of Hypericum occurred aslow as 2500 m.
Hypericum laricifolium thickets occur from the low to the high paramo proper. Ihey are foundin the altitudinal range of 3835 to 4200 m in the study area along paramo streams, on the edgesof bogs, along erosion gullies. Moreover, Hypericum laricifolium is already present in the highAndean belt as a component of the Hesperomeles lanuginosa forests and it is also frequentlyfound near Plantago rigida and Distichia muscoides bogs near the lower superparamo border.According to our observations (e.g. in the Quebrada Africa valley and near Laguna El Mosquito),stands of Baccharido-Hypericetum laricifolii develop frequently as a kind of secondary scrubafter destruction by tire or cutting of patches of high Andean (Gynoxys) forest.
The paramo proper
Ihe physiognomy of the zonal paramo proper vegetation is mainly determined by the presenceof bunches, or tussocks, of Calamagrostis recta and C. effusa. Calamagrostis intermedia mayalso occur, intermingled with the tussocks of C. effusa, as is the case in the Puracé area (Restrepo& Duque 1992). Other Poaceae such as Agrostis haenkeana, Bromus lanatus, Festuca dolichophylla and F. sublimis are found mainly in the more or less azonal bunchgrasslands ofthe paramoproper, and the latter in heavily grazed and burned paramos.
Chusquea tessellata (synonym: Swallenochloa tessellata, Clark 1989), an important component ofthe humid paramo vegetation in the Colombian Eastern Cordillera (Cleef 1981), seemsabsent from the Ruiz- Tolima massif. However, this bamboo species has been reported from theNevado del Huila paramos in the Central Cordillera (Rangel et al. 1985) and also from the Puracéarea (Duque & Rangel 1989; Restrepo & Duque 1992).
Scattered scrubs of Baccharis macrantha, Diplostephium eriophorum, D. schultzii, Hypericumlaricifolium, H. lancioides andPentacalia vernicosa are common among the tussocks ofPoaceae.In relatively open places, many small rosettes and prostrate shrubs shape the ground layer. Iheacaulescent, or ground rosettes, of Eryngium humile, Gnaphalium antennarioides, Hieracium avilae, Hypochoeris sessiliflora, Oreomyrrhis andicola Oritrophium peruvianum and Senecio repensfrequently occur. Baccharis caespitosa, Disterigma empetrifolium, Pernettya prostrata and Vaccinium floribundum are good examples of prostrate paramo dwarf shrubs. Extensive areas ofbogs,mainly consisting of vascular cushion plants like Plantago rigida, Distichia muscoides, Werneria

16 The paramo vegetation
crassa and W humilis are an important component of the landscape around 4000 m. Scatteredpatches of scrubs, mainly associated with slope debris, are composed of Diplostephium schultzii,D. violaceum, Hypericum lancioides. and Pentacalia vernicosa. Although they never form a continuous belt, they occur more frequendy towards the transition to the superparamo. In places witha very high grazing pressure and burning, the bunchgrassparamo is replaced by meadows or pastures (this subject is further discussed by Verweij & Budde 1992; see also Hofstede 1995 andVerweij 1995; for Ecuador by Ramsay & Oxley 1996). In this chapter, those meadows of Aciachne acicularis, Lachemilla orbiculata and Muhlenbergia cleefii will also be described. In largeextensions of paramo bunchgrasslands, around the Otún lake, Calamagrostis bunches have beenreplaced by loose tufts of Agrostis haenkeana (Verweij & Budde 1992).
In contrast with the Colombian paramos, the occurrence of bunchgrass paramos seems to berather limited in Venezuela. Monasterio (1980) reports extensive areas exclusively dominatedby the stemrosettes of Espeletia spp., but without the tussocks ofPoaceae. From the Venezuelanparamos, Poaceae are represented by Calamagrostis pittieri, Chusquea spencei, Stipa sp., Danthonia secundiflora, Poa sp., and Festuca fragilis have been documented (Berg 1998). Bergrecendy reported rather different upper grassparamo communities in the high Cordillera of Mérida. These communities are dominated by Calamagr:ostis cf. heterophylla, C. pittieri, C. cf.planifolia, and Festuca tolucensis. In Ecuador, as well as in the northern part ofPeru, the bunchgrassparamos have a wide extension. Acosta-Solís (1984) lists Calamagrostis bogotensis, C.ejjitsa, C. heterophylla, C. humboldtiana, C. ligulata, C. macrophylla, C. planifolia, C. recta,C. rigida, Festuca andicola, F. do lichophylla, F. sublimis and Stipa ichu as the most commonPoaceae from the Ecuadorian paramos. In the Cordillera of Talamanca, Costa Rica, Chaverri &Cleef (1996) have recorded patches ofbunchgrasslands of Stipa hans-meyeri and Festuca dolichophylla in a matrix of Chusquea (Swallenochloa) subtessellata bamboo-paramo. Calamagrostis ntermedia, C. pittieri and Agrostis tolucensis also commonly occur there. In Ecuador, Ramsay(1992) provided an overview of bunchgrass paramos using stratified sampling tecnique thatcovers the Ecuadorian Andes.
Zonal bunchgrassparamo
Calamagrostis recta and C. effusa, either single or mixed, form the larger part of the zonalbunchgrassparamo. The environmental parameters responsible for the spatial distribution ofthose two different species of Poaceae are not yet well known. Calamagrostis recta seems toprefer deep sandy soils, and occurs optimally under drier and colder conditions, especially inthe volcanic cordilleras of Colombia and Ecuador. Calamagrostis effusa in Colombia shows thetendency to dominate the low and middle, more humid, bunchgrassparamos; whi1e in Venezuela,this species also occurs in the more dry paramos (Monasterio 1980). On the other hand, in theCo1ombian Eastern Cordillera, Calamagrostis effusa is the most common bunchgrass.
Espeletia hartwegiana ssp. centroandina is often present in paramo bunchgrass1ands. In thestudy area, the present extension of this stemrosette species is strongly influenced by human activities. Although little is known about the ecology of Espeletia hartwegiana in the ColombianCentral Cordillera, at 1east during its early stages of growing, it seems to be quite sensitive torepetitive practices ofburning and grazing (Kloosterman et al., this vo1ume; Verweij & Kok 1992).
Syntaxonomy of the bunchgrasslands
Cuatrecasas (1934) presented a first synsystematic ordering ofthe bunchgrassparamo vegetationin the Central Cordillera. He defined the "Espeletietum hartwegianae Calamagrostiosum" and

La vegetación de páramo 17
the "Espeletietum hartwegianae-Hypericetum laricifoli" on the Iolima Volcano, as "associations" including all the Calamagrostis recta-e. effitsa bunchgrassparamos. In fact Cuatrecasas'associations share some species with the associations defined in this study. According to Cleef& Rangel (1984), most ofthe north Andean bunchgrass paramos be long to the class provisionallydefined as Espeletio-Calamagrostietea. Ihis c]assification of the bunchgrass paramo is introduced here again, but is now based on many more re]evés than in the case ofthe Sierra Nevadade Santa Marta. It is envisaged for the near future, that we can present a more definitive classification of all the zonal paramos of the northem Andes. Here we will provide the forma] description of the phytosocio]ogica] alliance, the order and the class of the bunchgrass paramosvalid not only for the Parque Los Nevados, but also for the Central Cordillera and for the northemAndes.
Espeletio-Calamagrostietea recto-effitsae C]eef, Range] & Salamanca cl. nov.Class of paramo bunch grassland of Espeletia-Calamagrostis recta/C. effusaClase de pajonal de páramo de Espeletia-Calamagrostis recta/e. effusaIype: Calamagrostietalia recto-effusae (this study).
Physiognomy:Equatorial open zona] paramo bunchgrass vegetation (varying in height from (10-)20 cm up to]00 cm) with 01' without the presence of stemrosettes.
Composition and syntaxonomy:Diagnostic bunchgrass species include: Calamagrostis recta, e. effusa, e. bogotensis (tuft) andFestuca sublimis. Stemrosettes: Espeletia spp., but also other members ofthe Espeletiinae (Heliantheae, Compositae). Also diagnostic are species of Altensteinia, Baccharis tricuneata, Bartsia,Castelleja, Halenia, Lachemilla, and Geranium sibbaldioides, Gnaphalium antennarioides,Grammitis moniliformis, Hypochoeris sessiliflora, Luzula racemosa, Oritrophium peruvianum,Pernettya prostrata, among others. The core flora is tropica]-Andean with temperate influencesCleef 1979; Van del' Hammen & C]eef 1986; Chaverri & Cleef 1996.). Bunchgrass species IikeFestuca dolichophylla (Mexico to Argentina), Calamagrostis macrophylla (Puracé to CentralPeru), and e. intermedia (Costa Rica to Bo]ivia) are a1so diagnostico However to a lesser extentsince they also occur elsewhere in the tropical American mountains.
In the Parque Los Nevados study area, the order Calamagrostietalia recto-effusae (ord. nov,this study) is included. Gn a later occasion we hope to be able to define the syntaxonomic classification for paramo bunchgrasslands e]sewhere in the Colombian Andes.
Dwarf bamboo paramo vegetation dominated by Chusquea tessellata (northem Andes) andChusquea subtessellata (Centra] America) may also be included in this c]ass. It is supposed thatthe bamboo paramos belong to a separate order Espeletio-Chusqueetalia subtessellato-tessellatae (prov.).
Synecology and distribution:Ihe mesic to dry bunch- 01' tussock grasslands are present at high altitudes in the northem Andes(Ecuador, Colombia, Venezuela) and apparently a]so in mountains in Panama and Costa Ricaon sloping as well as on ]evel dry to humid ground with shallow to deep soi]s with a considerablecontent of blackish humic material. Daily temperatures fluctuate considerably according to thediuroal climate type defined earlier by Iroll (1968): low temperatures during the night, highertemperatures during daytime. Annual precipitation may vary between about 800 mm and morethan 3000 mm.

18 The paramo vegetation
Calamagrostietalia recto-effusae Cleef, Salamanca & Range] ord. nov.Order of paramo bunch grassland of Calamagrostis recta/e. effusaOrden del pajonal de paramo de Calamagrostis recta/e. effusaType: Cerastio-Calamagrostion rectae Salamanca, Cleef & Range] 1992
Physiognomy:Open zona] coarse and tall bunchgrass vegetation with or without stemrosettes. Between thebunches or tussocks, a number of herbs (scapose and prostrate species, ground rosettes) arepresent.
Composition and syntaxonomy:The bunchgrass paramos on volcanic substrates studied in the Parque Los Nevados are examplesof the vegetation inc]uded in this order. It is supposed that this type of paramo bunchgrass vegetation is present in the Central Cordillera ofColombia, the high vo]canoes in southern Colombia(Macizo Colombiano, Nudo de los Pastos), towards Central Ecuador (Ramsay 1992) and perhapsa]so in the Cordillera de Mérida, Venezuela. In the Parque Los Nevados study are a, the alliancesof Cerastio-Calamagrostion rectae (all. nov., this study) and Festuco-Calamagrostion ejjitsae(all. nov., this study) are included in this order. Diagnostic are: Calamagrostis recta, C. eIJitsa,C. intermedia, Espeletia hartwegiana, E. pycnophylla, Festuca dolichophylla, F. sublimis. Alsodiagnostic taxa are Baccharis caespitosa, Lachemilla pitosa, Bromus lanatus, Gentianella dasyantha, Lupinus microphyllus, and Satureja nubigena. This order is different from the bunchgrasslands on soils relative]y poor in nutrients on sedimentary rocks, Calamagrostietalia ejjitsaeorder nov. prov., a]ready referred to by C]eef & Rangel (1984) for the bunchgrass paramos ofthe Colombian Cordillera Oriental and the Sierra Nevada de Santa Marta.
Synecology and distribution:Paramo proper bunchgrasslands are generally found on deep, relatively nutrient rich, blackishvolcanic soils, located on volcanoes in the northern Andes (Central Ecuador to Central Colombia). These are situated above the upper forest line (at c. 3600-3800 m) and the ]ower limit ofthe superparamo (at C. 4200--4300 m). The Parque Los Nevados bunchgrass paramos are thereforconsidered as a type locality.
Cerastio subspicati-Calamagrostion rectae Cleef, Salamanca & Range], all. nov.Alliance of paramo bunch grassland of Calamagrostis recta with Cerastium subspicatumAlianza de pajonal del páramo de Calamagrostis recta con Cerastium spicatumType: Calandrinio acaulis-Calamagrostietum rectae Sa]amanca, Cleef & Rangel 1992
Physiognomy:Open zonal bunchgrassland most]y with Espeletia stemrosettes. The ground cover between thebig tussocks, or bunches, consists of a number of low herbs and bryophytes. They are affectedby dai]y frost heaving in the uppermost reaches of the bunchgrass zone.
Composition and syntaxonomy:Character species include Aphanactis jamesoniana, Calamagrostis recta, Cerastium subspicatum, Colobanthus quitensis, Erigeron chionophilus, Lachemilla holosericea, Senecio formosusandValeriana plantaginea. In the Parque Los Nevados study area, paramo bunchgrassland ofCalandrinio-Calamagrostietum rectae and Calamagrostietum effuso-rectae are included in thealliance here described. The assemblage of the diagnostic species referred to here is differentiated against the bunchgrass paramo of Festuco-Calamagrostion effusae downslope. Upslope,the superparamo blue grasslands (Festuco breviaristatae-Agrostion araucanae) are found.

La vegetación de páramo 19
Synecology and distribution:Most of these zonal bunchgrasslands are present on deep vo1canic soils. These are subjected todaily frast heaving. This affects particularly the uppermost bunchgrass communities. In the studyarea ofthe Parque Los Nevados bunchgrass vegetation ofthis alliance is present between about4000 and 4400 m al1.
Calandrinio acaulis-Calamagrostietum rectae Salamanca, Cleef & Rangel1992Paramo bunchgrassland of Calamagrostis recta with Calandrinia acaulisPajonal del páramo de Calamagrostis recta con Calandrinia acaulisType: relevé TPN 5. Table 3; Figs. 3-8; Photo 5-9;(see aIso vol. 1, Photo 12, 17; Fig. 20)
Physiognomy:The Calamagrostis recta bunchgrass vegetation, which can be highly variable in cover, encompasses those bunchgrasslands reaching fram the upper part of the paramo proper to the lower¡imit of the superparamo. Three layers can be distinguished;- high tussock (bunchgrass) layer up to 1 m, almost exclusively composed of Calamagrostis
recta,- low herb layer,- ground layer with rosette herbs and a high cover of terrestric mosses,
In the variant of Espeletia hartwegiana a low stemrosettes of Espeletia (up to c. 1 m) arealways presen1. in the bunchgrass layer.
Composition and syntaxonomy:The almost exclusive contribution of Calamagrostis recta to the bunchgrass layer is the maincriterion for the definition ofthis association. Agrostis breviculmis, Calandrinia acaulis, Drabalindenii and Montia meridensis are diagnostic taxa. The differences in the floristic composition
TPN 84550 m
1 m
Fig.3. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant typicum at 4550m on the east slope ofthe summit ofvolcano Santa Rosa (relevé TPN 8), Risaralda province.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum recta e variante typicum a 4550 m en el lado este del cumbre del Volcán de Santa Rosa (lev. TPN 8), Dpto. de Risaralda.

20 The paramo vegetation
TPN94330 m
1 m
Fig.4. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant typicum at 4330m in the Paramo of Santa Rosa, east ofthe Valle San Ramón between OtÚn and Hacienda La Sierra (relevéTPN 9), Risaralda province. See Photo 5.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum rectae variante typicum a 4330 m en el Páramo de Santa Rosa, al este del Valle de San Ramón en el camino de OtÚn a laHacienda La Sierra (lev. TPN 9), Dpto. de Risaralda. Véase Foto 5.
TPN 44350 m
Fig. 5. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant of Espeletiahartwegiana ssp. centroandina at 4350 m on the northwest slope ofNevado Santa Isabel (relevé TPN 4),Risaralda province.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum rectae variante deEspeletia hartwegiana ssp. centroandina a 4350 m en la vertiente norte-occidental del Nevado de Santa Isabel (lev. TPN 4), Dpto. de Risaralda.

La vegetación de páramo
TPN 304315m
1 m
21
Fig. 6. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant of Espeletiahartwegiana ssp. centroandina at 43 l 5 m south ofthe pass and southeast of the Nevado Santa Isabel in theheadwaters ofQuebrada Africa (relevé TPN 30), Tolima province. See Photo 9.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum rectae variante deEspeletia hartwegiana ssp. centroandina a 4315 m al sur del Paso y al sureste del Nevado de Santa Isabelen las cabeceras de la Quebrada Africa (lev. TPN 30), Dpto. del Tolima. Véase Foto 9.
TPN64150 m
Fig. 7. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant of Espeletiahartwegiana ssp. centroandina at 4150 m between the Laguna Otún and the soutwest slope ofNevado SantaIsabel (relevé TPN 6), Risaralda province. See Photo 6.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum rectae variante deEspeletia hartwegiana ssp. centroandina a 4150 m en el camino de Otún al Nevado de Santa Isabel (lev.TPN 6), Dpto. de Risaralda. Véase Foto 6.

22 The paramo vegetation
TPN 324125 m
1m
Fig. 8. Zonal upper bunch grass páramo of Calandrinio-Calamagrostietum rectae variant of Espeletiahartwegiana ssp. centroandina at 4125 m at the southern side of Quebrada Africa in the headwaters ofValledel Totarito (relevé TPN 32), Tolima province. See Photo 7.
- Pajonal zonal del páramo propiamente dicho alto del Calandrinio-Calamagrostietum rectae variante deEspeletia hartwegiana ssp. centroandina a 4125 m en la márgen derecha de la Quebrada Africa en las cabeceras del Valle del Totarito (lev. TPN 32), Dpto. del Tolima. Véase Foto 7.
Photo 5. Open bunchgrass páramo ofthe variant typicum (relevé TPN 9) of Calandrinio acaulis-Calamagrostietum rectae close to its upper Iimit at 4330 m in the Páramo de Sta. Rosa, close to the Valle de SanRamón (Risaralda province). See Fig. 4.
- Pajonal paramuno abierto de la variante typicum (lev. TPN 9) del Calandrinio acaulis-Calamagrostietumrectae a 4330 m en su límite altitudinal en el Páramo de Sta. Rosa, cerca del Valle de San Ramón (Dpto. deRisaralda). Véase Fig. 4.

La vegetación de páramo 23
Photo 6. Collecting of soil samples and plant species in relevé site TPN 6 at 4150 m in the bunchgrass páramo of the variant of Espeletia hartwegiana ssp. centroandina of Calandrinio acaulis-Calamagrostietumrectae on the southwestern slope ofNevado Sta. Isabel, about 1 km north ofthe Otún lake (Risaralda province). See Fig. 7.
- Colección de muestras de suelo y de plantas dellev. TPN 6 a 4150 m en el pajonal paramuno de la variante de Espeletia hartwegiana ssp. centroandina del Calandrinio acaulis-Calamagrostietum rectae en ellado SW del Nevado de Sta. Isabel aproximadamente l km al norte de la Laguna de Otún (Dpto. de Risaralda). Véase Fig. 7.
Photo 7. Relevé TPN 32 at 4125 m in the Totarito Valley, left side ofQuebrada Africa, northeast ofthepass (Tolima province). Semi-open páramo bunchgrassland of the variant of Espeletia hartwegiana ssp.centroandina de Calandrinio acaulis-Calamagrostietum rectae. See Fig. 8.
- Levantamiento TPN 32 a 4125 m en el Valle de Totarito, márgen derecha de la Quebrada Africa, al norteoriente del paso (Dpto. del Tolima). Pajonal paramuno semiabierto de la variante de Espeletia hartwegianassp. centroandina de Calandrinio acaulis-Calamagrostietum rectae. Véase Fig. 8.

24 The paramo vegetation
Photo 8. Flowering Senecio isabelis at 4300 m in the upper grass páramo (Calandrinio acaulis-Calamagrostietum rectae val'. of Espeletia hartwegiana ssp. centroandina) east ofthe Laguna de OtÚn (Risaraldaprovince).
- Senecio isabelis florecido en el pajonal del páramo propiamente dicho alto (Calandrinio acaulis-Calamagrostietum rectae var. de Espeletia hartwegiana ssp. centroandina) a 4300 msnm al oriente de la Laguna de OtÚn (Dpto.de Risaralda).
Photo 9. Relevé TPN 30 at 4315 m in the headwaters of the Valle Quebrada Africa, south of the pass andsoutheast ofNevado de Santa Isabel. Páramo bunchgrassland ofthe variant Espeletia hartwegiana ssp. centroandina of Calandrinio acaulis-Calamagrostietum rectae on sloping ground. Nearby are two tracks withinfluence of grazing and trampling (on more level ground). See Fig. 6.
- Levantamiento TPN 30 a 4315 m sobre un talud de derrubios al Sur del Paso (al suroriente del Nevado deSanta Isabel) en el valle alto de la Quebrada Africa (Dpto. del Tolima). Pajonal paramuno de la variante deEspeletia hartwegiana ssp. centrondina del Calandrinio acaulis-Calamagrostietum recta e en la parte inclinada. Cerca hay dos caminos de herradura con influencia de pastoreo (en la palie más plana). Véase Fig. 6.

La vegetación de páramo 25
of the two variants belonging to this syntaxon reflect extreme environmental changes in thebunchgrasslands near the altitudinallimit of the paramo proper.
Syneco]ogy and distribution:This association is found on sandy deposits fomled by o]d pyroclastic flows. Most of these arecurrently covered by incipient soils, affected by perig]acial processes in the lower border of thesuperparamo. The uppermost distribution of this syntaxon coincides well with the altitudinallimit of growth of Calamagrostis recta. Frost heaving between the bunches occurs almost dailyin bare soil patches. The association Calandrinio acaulis-Calamagrostietum rectae is newlydescribed for the Ruiz-Tolima massif. It extends between 4]25 and 4350 m on the VolcanoNevado de Santa Isabel; between 4200 and 4440 m on the Vo1cano Nevado de Santa Rosa andit was found between 4140 and 4300 m on the Volcano Nevado del Cisne.
In the Colombian Eastem Cordillera, the extension of Calamagrostis recta bunchgrasslandsis very limited. Cleef (198]) reported bunchgrasslands, with Calamagrostis recta and Jamesoniabogotensis as a characteristic species, as high as 4500 m in the uppermost bunchgrass paramoon the dry side of the Sierra Nevada del Cocuy.
Variant typicum Salamanca, Cleef & Rangel1992Variante typicumType: relevé SS 82. Table 3; Figs. 3, 4; Photo 5
Physiognomy:]n this variant of Calamagrostis recta vegetation, the Espeletia stemrosette layer is absent. However, the herb layer is rather dense, covering as much as 45%.
Composition and syntaxonomy:The absence of Espeletia hartwegiana is the differentiating criterion. Moreover, this syntaxonis relatively poor in plant species. Some species, largely restricted to the lower superparamo,like Agrostis ¡oliata, Lachemilla tanacetifolia and Thamnolia vermicularis occur in the vegetation of the variant typicum.
Syneco]ogy and distribution:The soils (mainly Vitrandepts and Cryorthents) above 4200 m are poor]y developed and affectedby the periglacia] conditions of the superparamo climate. This bunchgrass vegetation is transitional to the ]ower-most superparamo. Bunchgrass paramos be]onging to this variant are in theupper part of the distribution range (mainly between 4200 and 4400 m) of Calandrinio acaulisCalamagrostietum rectae.
Variant of Espeletia hartwegiana ssp. centroandina Salamanca, C]eef & Range] 1992Variante de Espeletia hartwegiana ssp. centroandinaType: relevé TPN 5. Tab]e 3; Figs. 5-8; Photo 6-9;(see also vol. 1, Fig. 20; Photo 12, 17)
Physiognomy:In the vegetation of this variant the tussocks of Calamagrostis recta are intermingled with thelow stemrosettes of Espeletia hartwegiana ssp. centroandina. The Espeletia ]ayer covers between ]0% and 45 %. The herb layer, covers between 4% and 20% and is somewhat more openthan in the grass]and of var. typicum. A]so shrubs are more abundant than in the var. typicum.grassland.

26 The paramo vegetation
Composition and syntaxonomy:Differentiating from other variants is the presence of Espeletia hartwegiana ssp. ceritroandinaand the overall higher species richness. Castillejajissifolia, Niphogeton dissecta, Ophioglossumcrotalophoroides and Polystichum polyphyllum are absent fram the grassland of var. typicum.of Calandrinio acaulis-Calamagrostietum rectae.
Synecology and distribution:Soils are more developed than under the bunchgrass vegetation of variant typicum. Comparedto the bunchgrass paramo of variant typicum. slightly higher temperatures cause limited frastheaving and allows for the presence of more species. Bunchgrass paramos belonging to thisvariant are mainly found between 4125 and 4300 ffi. In the study area these are foundwithin theupper part of the paramo proper belt, and are contiguous below those of the bunchgrassparamoof Calandrinio acaulis-Calamagrostietum rectae var. typicum.
Calamagrostietum ejjitso-rectae Salamanca, Cleef & Rangel 1992Paramo bunchgrassland of Calamagrostis effusa and C. rectaPajonal de páramo de Calamagrostis effusa y C. rectaType: relevé SS 2. Table 3; Fig. 9; Photo 10, 11;(see also vol. 1, Photo 15)
Physiognomy:
This syntaxon refers to dense bunchgrasslands, with an average cover of 50% of the tussocklayer. In this five layered vegetation type, the Espeletia stemrosette layer covers from 5% to25%; the scrub ¡ayer about 5%; the herb layer between 5% and 25%; and the ground ¡ayer
TPN 263900 m
1m
Fig.9. Zonal bunch grass páramo of Calamagrostietum ejJitso-rectae at 3900 m on 'lahar' at the northemside ofQuebrada Africa in the headwaters ofValle del Totarito (relevé TPN 26), Tolima province. See Photo 11.
- Pajonal zonal del páramo propiamente dicho del Calamagrostietum effitso-rectae a 3900 m sobre flujode lodo en la márgen izquierda de la Quebrada Africa en las cabeceras del Valle del Totarito (lev. TPN 26),Dpto. del Tolima. Véase Foto 11.

La vegetación de páramo 27
Photo 10. Páramo bunchgrass vegetation (relevé TPN 31) of Calamagrostietum effilso-rectae with stem rosettes of Espeletia hartwegiana ssp. centroandina at 4200 m between the pass and the sources ofTotaritoriver in the headwaters of Quebrada Africa (Tolima province).
- Pajonal paramuno (lev. TPN 31) del Calamagrostietum effitso-rectae con caulirrósulas de Espeletia hartwegiana ssp. centroandina a 4200 msnm entre el paso y el nacimiento del río Totarito en el valle alto de laQuebrada Africa (Dpto. del Tolima).
Photo 11. Stem rosette-bunchgrassland (rel. TPN 26) of Calamagrostietum ejJuso-rectae con Espeletiahartwegiana ssp. centroandina at 3900 m in the Totarito valley, left side ofthe Quebrada Africa (Tolimaprovince). See Fig. 9.
- Pajonal paramuno con caulirrósulas (lev. TPN 26) del Calamagrostietum ejJuso-rectae con Espeletiahartwegiana ssp. centroandina a 3900 m en el Valle del Totarito, márgen izquierda de la Quebrada Africa(Dpto. del Tolima). Véase Fig. 9.
from 3% to 25%. Ground cover can become rather dense in those places not shadowed by thetussocks. The cover ofthe bryophytes ranges from less than 1% to 50%. Sometimes lichens canbe abundant, covering as much as 30% of the ground layer.

28 The paramo vegetation
Composition and syntaxonomy:Diagnostic taxa for this association are Castilleja fissifolia, Calamagrostis ejjÚsa and Espeletiahartwegiana occurring together. This association has a group of species in common with Calandrinio acaulis-Calamagrostietum rectae. Ihese include, besides Calamagrostis recta andC. efjÚsa; Aphanactis jamesoniana, Cerastium subspicatum, Luzula racemosa, SenecioformosusandValeriana plantaginea. The association Calamagrostietum ejJuso-rectae also shares somespecies with the two associations described by Cuatrecasas (1934) fram Tolima Volcano (seealso chapter 4).
Synecology and distribution:Ihe bunchgrass paramo belonging to this association thrives on non-differentiated Andosols,with an A~C profile, pH values around 6 and sometimes with high contents of aluminiul11. Iheassociation Calamagrostietum ejJuso-rectae includes the bunchgrasslands of Calamagrostis recta and C. effilsa of the middle part ofthe paramo proper belt throughout the study area. It extendscOl11l11onlybetween 3900 and 4200 m.
Festuco dolichophyllae-Calamagrostion ejJusae Cleef, Salamanca & Rangel all. nov.AlIiance of paramo bunchgrassland of Festuca dolichophylla and Calamagrostis effusaAlianza de pajonal del páramo de Festuca dolichophylla and Calamagrostis effusaType: Espeletio hartwegianae-Calamagrostietum ejjÚsae Salamanca, Cleef & Rangel 1992
Physiognomy:Ihis syntaxon refers to dense bunchgrass paral110 vegetation with or without stemrosettes. Between the bunches species of low herbs and dwarf shrubs, bryophytes and lichens are found.
Composition and syntaxonomy:Diagnostic for this syntaxon are: Calamagrostis ejJusa (highest presence and cover), Carexpichinchensis, Espeletia hartwegiana ssp. centroandina (p.p.), Festuca dolichophylla (the lattertwo were recorded in half of the relevés that belong to this alliance), Hieracium avilae, Lycopodium spurium and Nertera granadensis. The alliance comprises two associations; FestucoCalamagrostietum effilsae and Espeletio-Calamagrostietum ejJusae. The alliance is separatedfrol11 Cerastio-Calamagrostion rectae by the combined presence of diagnostic species of bothassociations. The alliance is separated from Cerastio-Calamagrostion rectae by the near absenceof Baccharis caespitosa, Diplostephium schultzii, Erigeron chionophilus, Gentianella dasyantha, Jamesonia goudotii, Lachemilla nivalis and Pentacalia vernicosa among others. FestucoCalamagrostietum ejJusae is separated from Espeletio-Calamagrostietum effÚsae by the presence of Azorella multifida, Bromus lanatus, Calamagrostis recta, Cotula minuta, Lucilia kunthiana, Oreomyrrhis andicola and Ranunculus peruvianus. Espeletia is absent in Festuco-Calamagrostietum ejJusae, but diagnostic in Espeletio-Calamagrostietum ejJusae together withAgrostis tolucensis, Baccharis rupicola, B. genistelloides, Castilleja fissifolia and Halenia spp.
Synecology and distribution:In general these grassland coml11unities are less adapted to cold. These grassland cOl11munitiesare strong1y affected by buming and grazing more than in any other vegetation type. Bunchgrasslands belonging to the alliance are commonly distributed between 3850 and 4250 111in theParque Los Nevados.

La vegetación de páramo
Festuco dolichophyllae-Calamagrostietum effusae Salamanca, Cleef & Rangel1992Bunchgrass paramo ofFestuca dolichophylIa and Calamagrostis effusaPajonal de páramo de Festuca dolichophylIa and Calamagrostis efIusaType: relevé SS 85. Table 3; Fig. 10; Photo 12
29
Physiognomy:Three layers can be distinguished in this type ofbunchgrass paramo:- medium close to close tussock layer, 0.6 to 1 m high, mainly composed of Festuca dolicho
phylla and Calamagrostis ejjitsa, covering between 50% and 90%,- sparse herb layer, around 30 cm high, covering between 2% and 15%,- close ground layer, including rosette herbs, prostrate dwarf shrubs, bryophytes and lichens.
Composition and syntaxonomy:The diagnostic species are: Calamagrostis effusa and Festuca dolichophylla. The absence ofAphanactis jamesoniana, Cerastium subspicatum, Colobanthus quitensis, Luzula racemosa,Senecio formosus andValeriana plantaginea is also diagnostico Bromus lanatus, Calamagrostisrecta, Cotula minuta, Lucilia kunthiana, Oreomyrrhis andicola, Ranunculus praemorsus andTaraxacum spp. are also present within the Calandrinio acaulis-Calamagrostietum rectae.
Synecology and distribution:The grassland of Festuco dolichophyllae-Calamagrostietum ejjitsae was observed in the externalfringes of small glacier valleys, on moraines, or within those places that are occasionally flooded.Individual tussocks of Festuca dolichophylla frequently accompany Plantago rigida cushionbogs. Cleef et al. (1983) already reported bunchgrasslands, with Festuca dolichophylla as thedominant tussock grass, on the external slopes ofthe volcanic massifbetween 3800 and 4250 m.
TPN 143960 m
1 m
Fig.l0. Zonal bunch grass páramo of Festuco-Calamagrostietum effitsae at 3960 m in the valley ofQuebradaBetania, Hacienda La Sierra, Municipality of Santa Rosa de Cabal (relevé TPN 14), Risaralda province.
- Pajonal zonal del páramo propiamente dicho del Festuco-Calamagrostietum e.ffitsae a 3960 m en el vallede la Quebrada Betania, Hacienda La Sierra, Municipio de Santa Rosa de Cabal (lev. TPN 14), Dpto. deRisaralda.

30 The paramo vegetation
Photo 12. Relevé site ofTPN 35 at 3800 m in bunchgrassland of Festuco dolichophyllae-Calamagrostietum ejJusae with stem rosettes of Espeletia hartwegiana ssp. centroandina in the Totarito valley, left sideofthe Quebrada Africa (Tolima province).
- Sitio del levantamiento TPN 35 a 3800 m en el pajonal del Festuco dolichophyllae-Calamagrostietumeffilsae con caulirrósulas de Espeletia hartwegiana ssp. centroandina en el Valle de Totarito, márgen izquierda de la Quebrada Africa (Dpto. del Tolima).
Festuca dolichophylla is a common element ofthe bamboo paramos in the Eastern Cordilleraof Colombia e.g. Chusquea (communities with species of Sphagnum and Breutelia described byCleef (1981). Chusquea tesselata was previously denominated Swallenochloa tesselata by thisauthor.
Acosta-Solís (1984) and J0rgensen & Ulloa Ulloa (1994) reported Festuca dolichophylla asa common species (2700--4200 m) in the Ecuadorian paramos. Chaverri & Cleef (1996) studiedvegetation with Festuca dolichophylla and Chusquea subtessellata in the Costa Rican paramosbetween 3400 and 3600 m. They also reported Pernettya prostrata and species of Geranium,Gnaphalium and Lachemilla as companions. Festuca dolichophylla also has been recorded as aconstituant of dry alpine grassland in Ecuador (J0rgensen & Ulloa Ulloa 1994), Peru (Gutte1985) and Bolivia (Seibert & Menhofer 1992).
Espeletio hartwegianae-Calamagrostietum ejJusae Salamanca, Cleef & Range11992Paramo bunchgrassland of Calamagrostis effusa with Espeletia hartwegianaPajonal de páramo de Calamagrostis effusa con Espeletia hartwegianaType: relevé TPN 34. Table 3; Fig. 11;(see also vol. 1, Photo 3-5)
Physiognomy:These stemrosette bunchgrasslands show a distinct four layered stratification;- tussock layer, 0.6-1 m high, with a cover between 30% and 70%,- Espeletia layer, 1.5-3 m high, with the stemrosettes of Espeletia covering between 1% and 40%,- herb layer, with a height of 30 cm covering between < 1% and 20%,- ground layer, covering between 2% and 15%.

La vegetación de páramo
TPN104175 m
1 m
31
Fig.11. Zonal bunch grass páramo of Espeletio-Calamagrostietum efJitsae at 4175 m on the west slope ofvolcanoSanta Rosa (relevé TPN 10), Risaralda province.
- Pajonalzonal del páramo propiamente dicho del Espeletio-Calamagrostietum ejJusae a 4175 m en el ladooestedel Volcán de Santa Rosa (lev. TPN 10), Dpto. de Risaralda.
Composition and syntaxonomy:Diagnostic for this association is the absence of Aphanactis jamesoniana, Cerastium Sub5picatum, Luzula racemosa, Senecio formosus and Valeriana plantaginea.
In stands with a relatively open tussock layer small acaulescent rosettes, like Eryngium humile,Senecio repens and Gnaphalium antennarioides as well as terrestric bryophytes, become moreabundant and conspicuous, with cover as high as 80%.
Baccharis rupicola, Castilleja fissifolia, Espeletia hartwegiana, Lysipomia muscoides andNiphogeton dissecta form a group of species shared by Calandrinio acaulis-Calamagrostietumrectae var. Espeletia hartwegiana, Calamagrostietum ejJuso-rectae and Espeletio hartwegianaeCalamagrostietum ejJusae.
Synecology and distribution:The bunchgrassparamos of Espeletio hartwegianae-Calamagrostietum ejJusae are associated
with humic and poorly differentiated Andosols (Thouret 1983). This association is rather com1110nin the middle part ofthe paramo proper belt in the study area (between 3920 and 4250 m).Cleef et al. (1983) ha ve already reported bunchgrasslands of Calamagrostis ejJusa and Espeletiahartwegiana ssp. centroandina from the external, humid, slopes of the Ruiz- Tolima massif.
Rangel & Lozano (1986) reported Calamagrostis ejJusa-Espeletia hartwegiana communitieson the Puracé Volcano between 3300-3400 m. Stunn & Rangel (1985) report Calamagrostiseffusa bunchgrass paramos from the volcanoes Cumbal and Chiles. On the Nevado del Huila,according to the same authors, Calamagrostis ejJusa is not dominant nor does it forrn a typicalbunchgrassparamo, though it is present in the paramo belt.

32 The paramo vegetation
The bunches of Calamagrostis effitsa dominate throughout the paramo proper in the EasternCordillera and, according to Cleef (1981), they can be found in dry paramos as well as in humidbamboo paramos together with Chusquea.
Azonal paramo vegetation
Cushion bogs
Geophytes and chamaephytes are the main constituents ofthe cushion-like vegetation, currentlywell known from the high tropical Andes. The phytosociological ranking of the cushion bogvegetation proposed by Cleef (1981) for the Colombian Eastern Cordillera seems valid for allthe other paramos in Colombia within the order Oritrophio- Wernerietalia Cleef 1981 and thealliances Wernerion crassae-pygmaeae and Gentiano-Oritrophion (Cleef, l.c.). Rivas-Martinez& Tovar (1982) provided a syntaxonomical update of tropical Andean cushion bogs includingthe class level: Plantagini rigidae-Distichietea muscoides.
Wernerietum humilis Salamanca, Cleef & Rangel1992Cushion bog ofWerneria humilisTurbera de cesped de Werneria humilisType: relevé AMC 603. Table 4; Photo 13
Physiognomy:Stands include small loose cushions, with a layer of cushion chamaephytes covering between15% and 90%. Tussocks of Poaceae may be present with low cover percentages (1% to 15%).The bryophyte layer is well developed; achieving up to 60% of the total cover.
Photo 13. Cushion of Werneria humilis (Wernerietum humilis) situated in a Plantago rigida cushion bog(Gentianello dasyanthae-Plantaginetum rigidae) at about 4200 m on the northwestern side ofthe Nevadodel Ruiz (Caldas province). Noteworthy are also some ground rosettes of Gentianella dasyantha.
- Cojín de Werneria humilis (Wernerietum humilis) ubicado en una turbera de cojines de Plantago rigida(Gentianella dasyanthae-Plantaginetum rigidae) a ca. 4200 m en el lado occidental del Nevado del Ruiz(Dpto. de Caldas). Además son notorias algunas rosetas sésiles de Gentianella dasyantha.

La vegetación de páramo 33
Composition and syntaxonomy:The cushion vegetation of the association Wernerietum humilis is rather poor in species diversity.Distichia muscoides and Plantago rigida are diagnostic in the variant of Distichia muscoides;Hypochoeris sessiliflora in the other variant. Carex peucophila, a diagnostic species of the alliance, is present in most of the relevés ;"ith a high total cover.
Synecology and distribution:The cushions belonging to this association (occurring in the paramo praper between 4000 and4400 m) are found on the wet and cold glacial valley floors, where often the water table reachesthe surface. Werneria humilis cushions frequently occurr in Plantago rigida bogs. Isolated cushions of Wernerietum humilis are also found in very wet (micra )habitats ofthe lower superparamooutcrops. Here they are associated with the open grassland ofthe association Senecioni latifloriCalamagrostietum ligulatae.
Although Werneria humilis has a wide distribution in the superparamos of the Eastem andCentral Cordilleras, the true cushion vegetation of this association is not common. Rangel &Lozano (1986) also report Werneria humilis cushions from 4050 m on the Puracé Vol cano,Central Cordillera of Colombia.
Cleef et al. (1983) reported Werneria humilis among the character taxa of the associationOritrophio limnophili- Wernerietum pygmaeae found on the Santa Isabel Volcano. The samespecies also occurs in the superparamo vegetation of the Sumapaz and the Sierra Nevada delCocuy paramos in the Colombian Eastem Cordillera. Acosta-Solís (1984) reports Werneria humilis as an element of the paramo cushion vegetation, in Ecuador. Ramsay (1992) described inEcuador different zonal types of what he calls cushion paramo (4100--4400 m) with Werneriahumilis from the volcanoes El Altar, Cotacachi and the Páramo de Guamaní in Ecuador.Werneria humilis seems also present in Peru.
Variant of Hypochoeris sessiliflora Salamanca, Cleef & Rangel1992Variante de Hypochoeris sessilifloraType: relevé AMC 603. Table 4
Composition and syntaxonomy:Differential species of this variant are Breutelia sp., Hypochoeris sessiliflora, Muhlenbergiacleefii and Riccardia spp.
Synecology and distribution:Small stands of the variant of Hypochoeris sessiliflora of the association Wernerietum humilisare found in the altitudinal range of 4000 m to 4400 m, mainly in small flat areas on the slopesof the Nevado del Ruiz.
Variant of Distichia muscoides Salamanca, Cleef & Rangel1992Variante de Distichia muscoides
Type: rel. SS 113. Table 4
Composition and syntaxonomy:Distichia muscoides is the differential species. Moreover, the floristic composition at the specieslevel is less diverse, and species likeAgrostis haenkeana, Azorella aretioides, Baccharis caespitosa, Bromus lanatus and Gentiana sedifolia among others, are absent fram the vegetation ofbogs belonging to this variant. But present in the vegetation is the variant of Hypochoeris sessiliflora.

34 The paramo vegetation
Synecology and distribution:Lax cushions of the variant of Distichia muscoides of the asociation Wernerietum humilis were
found at 4200 m in flat areas, on moist soil and along the slopes of Santa Isabel Volcano.
Floscaldasio hypsophilae-Distichietum muscoides Cleef 1981Variant of Cotula minuta vaL nov. (this study)Floscaldasia hypsophila-Distichia muscoides cushion bog; Cotula minuta variantTurbera de cojines de Distichia muscoides con Floscaldasia hypsophila; Variante de Cotula minutaTable 4; Fig. 12;(see also vol. 1, Photo 27, 30)
Physiognomy:The association comprises flat to convex cushions of Distichia muscoides. The young and firmcushions are poor in companion species. However, the cover ofthe small herbs is higher on thetop of mature cushions.
Composition and syntaxonomy:The association Floscaldasio hypsophilae-Distichietum muscoides already has been describedfor the Cordillera Oriental. Cleef et al. (1983) also provided a short description ofthis vegetationtype present in the study area.
Besides Distichia muscoides and Floscaldasia hypsophila other character species are Bryumargenteum, Calamagrostis coarctata and Gymnocoleopsis multiflora. On decaying cushions,Calamagrostis coarctata may achieve high cover. Cotula minuta is differential for the CentralCordilleran Distichia cushion bogs. They belong to the variant of Cotula minuta. Different species have been reported within hollow communities around lake Otún at 3950 m the reddish
Fig. 12. Small cushion bog of Floscaldasio-Distichietum muscoides (relevé Cleef & Salamanca 582) at4300 m in contact with upper bunchgrass paramo (Calandrinio-Calamagrostietum rectae) and lower superparamo sClUb of Loricarietum colombianae (relevé Cleef & Salamanca 593) on outcrops of volcanic rockon the soutwest side ofNevado Santa Isabel, Risaralda province.
- Turbera de cojines del Floscaldasio-Distichietum muscoides (lev. Cleef & Salamanca 582) a 4300 m encontacto con pajonal del páramo propiamente dicho alto (Calandrinio-Calamagrostietum rectae) y matorralbajo del superpáramo bajo del Loricarietum colombianae (lev. Cleef & Salamanca 593) sobre afloramientosde roca volcánica en el lado suroccidente del Nevado de Santa Isabel, Dpto. de Risaralda.

La vegetación de páramo 35
coloured Calamagrostis jamesonii is particularly conspicuous. Scirpus cernuus has also beenfound common at 3950 m on top of the Distichia cushions in neighbouring Laguna Mosquito.
Synecology and distribution:In the Ruiz-Tolima massif, Distichia muscoides cushion mire is present mainly above 4000 min the high paramo proper and in the superparamo. Distichia muscoides forms soligenous cushions in concave terrains with a peaty substrate, especialIy in the depressions left by former glaciallakes.
Rangel & Lozano (1986) observed Distichia muscoides cushions at 3950 m on the PuracéVolcano. Duque & Rangel (1989) studied the same cushion type vegetation, which they rankedas Agrostio boyacensis-Distichietum muscoides.
In Ecuador Ramsay (1992) reported a Distichia muscoides-Azorella corymbosa- Werneria humidis cushion paramo at 4100 m on the El Altar Volcano, Ecuador. In addition to Gutte (1980)with the association Stylito andicolae-Distichietum muscoides, Rivas-Martínez & Tovar (1982)described the Calamagrostio jamesonii-Distichietum muscoides fram Central Peru. Seibert &Menhofer (1992) studied a "Scirpo deserticolae-Distichietum muscoides" from the Bolivian Andes. Ruthsatz (1977; 1995) studiedDistichia muscoides cushion bogs fram the Andes ofnorthernArgentina and northern Chile.
Carici peucophilae- Wernerietum crassae Cleef 1981var. Cotula mexicana Salamanca, Cleef & Rangel 1992Werneria crassa cushions with Carex cC.peucophila; Cotula mexicana variantTurbera de cesped de Werneria crassa con Carex cf. peucophila; Variante de Cotula mexicanaType: rel. SS 152. Table 4
Physiognomy:The vegetation of this association, defined by Cleef (198 1) for the paramos of the ColombianCordillera Oriental, comprises loose cushions of Werneria crassa. These cover between 40%and 80 % and are associated with a well-developed bryophyte layer, up to coverages of 25%.Sometimes tussocks of Poaceae can be found fram 5% to 50%. In some of the relevés, dwarfshrubs achieve a cover fram 2% to 60%.
Composition and syntaxonomy:Diagnostic species of this Central Cordilleran association are Cotula mexicana, Breutelia sp.,Cladina cf. rangiferina, Muhlenbergia cleefii, Plagiocheilus solivaeformis and Riccardia sp(p.).Cleef (1981) defined Carici peucophilae-Wernerietum crassae (with subass.wernerietosumcrassae) as a new syntaxon for the Eastern Cordillera. From this syntaxon Bartsia sp., Breuteliasp., Muhlenbergia cleefii, Hypericum lancioides, Oritrophium limnophilum, Plantago rigida andRiccardia spp., among others, also occur in the variant described here. Differential for the Central Cordillera are Bromus lanatus, Cotula mexicana, Gentianella dasyantha and Werneriacrassa ssp. crassa (versus Werneria crassa ssp. orientalis).
Synecology and distribution:In the Ruiz- Tolima volcanic massif, the vegetation belonging to Carici- Wernerietum crassae isfound mainly on flat glacial valley bottoms as small patches on the fringes of Plantago rigidabogs. The relevés which constitute the basis for the description of this variant were 10cated inthe altitud in al range of 3800 m to 4200 m. Relevés with an important cover of Hypericum lancioides apparently represent a successional stage on old Werneria crassa cushions parallel tosuch series of succession described on Plantago rigida and Distichia cushions in the ColombianEastern Cordillera (Cleef 1981; Bosman et al. 1993).

36 The paramo vegetation
A similar type of vegetation, but with another species of Werneria e.g. the vegetation ofOritrophio limnophili-Wernerietum pygmaeae subass. typicum, has been reported fram one 10cality in the Sierra Nevada de Santa Marta (Cleef & Rangel 1984). In Ecuador, Acosta-Solís(1984) found Werneria disticha, W graminifolia, W pumila, W pygmaea, W rigida and Wrosea, on the volcanic sandy deposits of the Chimborazo V olcano. Monasterio (1980), referredto Werneria pygmaea cushions within the Plantago rigida bogs on glacial valley bottoms in theSierra Nevada de Mérida, Venezuela.
Gentianello dasyanthae-Plantaginetum rigidae Salamanca, Cleef & Rangel1992Gentianella dasyantha-Plantago rigida cushion bogTurbera de cojines de Plantago rigida con Gentianella dasyanthaType: relevé SS 1. Table 4; Photo 13-15;(see also vol.1, Photo 26, 31)
Physiognomy:The bogs of this association include compact flat cushions, in which Plantago rigida attainsbetween 20% and 100% cover. Tussocks ofPoaceae, grawing on top ofthe cushions, with coverbetween 1% and 30%, are characteristic. Some (dwarf) shrubs may be present with cover between 2 % and 50%. In appraximately half of the relevés, bryophytes were present; coveringbetween < 1% and 60%.
Composition and syntaxonomy:Diagnostic species ofthis association are Baccharis caespitosa, Bromus lanatus, Diplostephiumeriophorum, Gentianella dasyantha, Hypericum laricifolium, Lachemilla galioides, Oreomyrrhis andicola, Pentacalia vaccinioides, P. vernicosa and Valeriana plantaginea. With the ex-
Photo 14. Cushion bog ofthe Gentianello dasyantha-Plantaginetum rigidae association in the grass páramo ofthe Parque Los Nevados.
- Turbera de cojines perteniciente al Gentianello dasyanthae-Plantaginetum rigidae en el páramo propiamente dicho del Parque Los Nevados.

La vegetación de páramo 37
Photo 15. Cushion bog of Plantago rigida associated with bunches of Festuca sublimis (Gentianello dasyanthae-Plantaginetum rigidae) at 4200 m on the northwestern slope ofthe Nevado del Ruiz near Las Brisas(Caldas province). The volcanic outcrops support low shrubs of Loricaria colombiana (Compositae) andmoss cover.
- Turbera de cojines de Plantago rigida con pajonal de Festuca sublimis (Gentianello dasyanthae-Plantaginetum rigidae) a ca. 4200 m en el lado norte occidental del Nevado del Ruiz, cerca de Las Brisas (Dpto.de Caldas). Los afloramientos de roca volcánica están cubiertos de arbustillos de Loricaria colombiana(Compositae) y musgos.
ception of some of the character species for higher rank syntaxa, there are not many species incommon with the diagnostic species given by Cleef (1981) for Hyperico lancioides-Plantaginetum rigidae of the paramos from the Eastern Cordillera of Colombia.
Two subassociations are provisionally described within the present Gentianello dasyanthaePlantaginetum rigidae, the subassociation typicum, and the subassociation brometosum lanatae.Ihe first is poorer in species. Bromus lanatus, Elaphoglossum mathewsii, Hieracium avi/ae,Niphogeton dissecta, Pentacalia vernicosa, Senecio formosus and Valeriana plantaginea aredifferential for the subassociation brometosum lanatae. The association Gentianello dasyanthaePlantaginetum rigidae subass.typicum is characterized by the presence of Oreobolus cf. obtusangulus.
Oreobolus obtusangulus, a common element of the Plantago rigida bogs in the ColombianEastern Cordillera (Cleef 1978; 1981; Bosman et al. 1993), is very rare in the Ruiz-Tolimamassif. Ihis cushion species was found only two times during the tieldwork: one record fromthe Páramo de Santa Rosa and another from the Quindío V oleano. It is supposed that the vo]canicenvironment, rich in nutrients, is limiting the presence of Oreobolus.
Syneco]ogy and distribution:Cushion mires belonging to the association Gentianello dasyanthae-Plantaginetum rigidae arefound in concave and flat glacial depressions in the paramo proper be]t.
Plantago rigida cushion bogs occur in most Co]ombian paramos. Although apparently scarcein the Sierra Nevada de Santa Marta (there is on]y one observation: Sturm & Rangel 1985), theyare wide]y distributed in the Eastern and in the Central Cordilleras and a]so occur in the WesternCordillera, the Tatamá massif and on the Farallones de Cali (Ca]derón 1993).

38 The parama vegetatian
Benoist (1935), Diels (1937), God]ey (1978), Acosta-Solís (1984) report Plantago rigidacushion bogs fram the paramos of Pichincha and Chimborazo, as well as from many other ]0calities in Ecuador. Also in Ecuador, Ramsay (1992) described aPlantago rigida cushion paramofram 4200 m at the El Altar Volcano, Ecuador. In Central Peru, Gutte (1980) described anassaciation of Carex microglochin-Plantago rigida fram 3800-4800 m. This vegetation typehas affinities with the Gentianello dasyanthae-Plantaginetum rigidae at syntaxonomica] ranksabove the association leve], with species like Gentiana sedifolia, Luzula racemosa, Oritrophiumlimnophilum, Ourisia muscosa andWerneria pygmaea. Rivas-Martinez & Tovar (1982) described the same community of Centra] Peru as Hypselo reniformis-Plantaginetum rigidae, be]onging to the alliance Hypselo-Plantaginion rigidae (Calamagrostio jamesonii-Distichietaliamuscoides MartÍnez & Tovar, l.c). Seibert & Menhofer (1992) studied a Calamagrostio ovataePlantaginetum rigidae fram the Bo]ivian Andes. Monasterio (1980) reported Plantago rigidabogs fram glacial valley bottams in the paramo belt ofthe Sierra Nevada de Mérida, Venezuela.
Sphagnum-Chorisodontium bog
Sphagnum bogs appear to be rather rare in the Parque Los Nevados study area. This could beattributed ta the relatively nutrient rich volcanic substrates and low precipitation compared toparamo elsewhere in the Colombian Andes.
Just one relevé (Table 4a), has been taken fram a species-rich Sphagnum-Chorisodontium bogwith a substantia] open aerial cover af Calamagrostis effusa at 3710 m along the upper foresthne just below Hacienda La Sierra, above Termales de Santa Rosa de Cabal, on the west slopeof the TPN transect.
Paramo scrub
Apart fram the association Baccharido-Hypericetum laricifolii described above, another scrubcommunity can be reported.
Pentacalietum vernicosi Cleef 1981 carr. Salamanca, Cleef & Rangel 1992Pentacalia vernicosa shrubMatorral de Pentacalia vernicosaTab]e 2
Physiognomy:These shrub]ands of the high paramo proper exhibit three layers:- shrub ]ayer, covering between 20 and 75%,- tussock layer, covering between < 1 and 40%,- graund ]ayer, covering between < 1 and 30%.
Sometimes bryophytes can dominate in the ground layer, reaching a cover as high as 80 %.
Composition and syntaxonomy:This association was originally described as Senecionetum vernicosae by C]eef (1981) for thesouthern paramos of the Co]ombian Eastern Cordillera. Character species for the Centra] Cordillera include Senecio isabelis and S. latiflorus. Common species of this association to Centra]and Eastern Cordilleras are Calamagrostis recta, Campylopus spp., Geranium sibbaldioides,Lachemilla nivalis, Lachemilla holosericea, Leptodontium wallisii, Oreomyrrhis andicola, Pelti-

La vegetación de páramo
Table 4a. Sphagnum-Chorisodontium bogo
39
Relevé nr.Relevé area
Slope (0)
ExposureAltitude m
cover %
6510
3115152
1<1<1<1<1<1
<1<1+<1+
51
<1<13
<11
<1<1<1<1
AMC 6086m x 6 m3SW3710
vascular species:
Calamagrostis effusaEspeletia aff. hartwegiana(yellowish broad-Ieaved)Hypericum laricifoliumOritrophium peruvianumLycopodium d. brevifoliumFestuca dolichophyllaIsoetes andinaPernettya prostrataUncinia macrolepis(U meridensis,)Lucilia kunthiana
Senecio repensHypochaeris sessilifloraCotula minutaLachemilla d. hispidulaSisyrinchium sp. (small,yellow flowering)Erigeron sp.Nertera granadensisOphioglossum crotalophoroidesCarex sp. (brownish, small)Hypericum lancioidesBartsia sp.Pedicularis incurvaWerneria humilisGeranium sibbaldioides
Ericaceae (unknown)Bromus sp.Azorella multifidaJamesonia rotundifolia.
Plagiocheilus solivaeformis
Locality: Parque Los Nevadas, RisaraldaHacienda La Sierra; upper forest line24 January 1980
microtopography: hummock-hollow
cover % bryophytes & lichenslcryptogams:
40 Chorisodontium sp.10 Sphagnum magellanicum
< 1 Pezizales (orange)35 Plagiochila sp.25 Riccardia spp.< 1 Lepidozia sp.2 Anastrophyllum sp.1 Campylopus cucullatifolius2 Adelanthus lindenbergianus
<1+ Jensenia sp.<1+ Lophocolia sp.
5 Campylopus spp.1 Herbertus sp.1 Metzgeria sp.
< 1 Lejeuniaceae (Iight-greenish)
<1+ Funaria calvescens<1+ Ascomycetes (yellowish)
1 Pleurozium schreberi< 1 Leptodontium wallisii<1 Cladonia subg. Cenomyce
< 1+ Leptoscyphus d. c1eefii.
gera spp., Satureja nubigena and Senecio formosus. Senecio niveo-aureus is found instead ofSenecio isabelis in Pentacalietum vernicosi of the Co1ombian Eastern Cordillera (C1eef 1981).
Synecology and distribution:Pentacalia vernicosa is a frequent companion of the Plantago rigida bogs. Scattered shrubs ofthis species are also characteristic for Calamagrostis recta bunchgrass1ands. Range1 & Lozano(1986) described the species as characteristic for the Calamagrostis macrophylla-C ejJusa andChusquea (Swallenochloa) tessellata-Espeletia hartwegiana ssp. centroandina communities onthe Puracé Volcano. Cleef (1981) reported the Pentacalietum vernicosi fram the southern paramos of the Co1ombian Eastern Cordillera, as a new association in the altitudinal range between3800 and 4100 m.

PRADOSHUHLEN8ERGIETUH r.STIGIAUE ACIACHNETUH ft'ULVfHATA(AOItOSTIO SREVICULHIS
lACLA CONSTANCIAtUBA", nP'ICUHIIJS .•.•• , L. I1IC:tOP'H'fL.LAELACHEHILLETUI1 OUICULATAE
l!Wtlntllm:t¡nl,o"C106 C415 C491 C257 CI60 C1501 CS65C336 C320 e 156C31 C452 C567SSI S·52544 C15J4 CI5J3 CS1e531529532S7" C!577524 C576 C624 C61 ti512C596
,\lt:tu,l(m.S.I).m.)3S30 3610 3670 3775 4OtiO 4000 321503950 4040 3960 3640 3130 3ellO4030 4070 4050 3e25 3eoo 39153950 3900 3900 4200 3960 4150 3740 3600 3t1S0 39t104\50
lOCQliúoú(l$pP'u"euoA'y'y'yG,y AoooSISIOOOO,OOASIooo
Ar<)ll11414369121221S1615152S6• 1515151541544425 4
r~n(jleMe2
-lO-24210637202102-7220 720 -520 -S62 2
EXPOSICIónNNWENE- 5WWNW5SNW
-SE5 NW-NEE S- WNW
"laxo oprox.1146461037156101616"9261S14201614621 913e1441314
.\Otros Grumlnlle6070953560100409Ses9096eseo707560709060S10"30354SlO5,130'O
~Es~nllo herboceo15,O -211<l- ,1-3,1-<1-10,1,1323243311S,1- 2131-<1<1
SESl.rotorosMteSSS- -7-,1 12<1- 3320 •2011•1062S2725470909292995021
:t[slrolo mQc(lllo~
-<1-- ---1'O 212<1- 42-S,133-2,1 3~E$troto arbusl.IYO
--- --<13 3131010-2S7----<l ---<1XMlISCI
60107"-S60 -,1 ,1-315 -33<1- ,1-S2-31--2~Llchllns
--------<1- -,1- - -- -<1------ --<1
~5lJ&lo d8snudo-- --- -- ---ss--- ------- --
lAXA11llhlenberglu fOS1l910l0
'O709535409640 ,1';!lros~lS sr.
,"• 1erelll.elH:1 ctlr~seo
'O'0"Cllre:-; DIChmch!lnsis
'O1 <1,1
JUOCllS C!..IP~roldes101
leptotJonllumlutllum,l'2
Sphuooum cuspl<lQium7Q
Brell',f::il11 oll1onll60
Gol¡umconl!'scens00
'lI~rMno PY9mtl(l~
5
GllrurL1Ulnconfl;r~umS
CllffiPY!OpIlS cO\'lfollUS
S
,.'\clochne pU¡'(lno,o95es909 •• 5.040SO607060SS
LuplnuS mlcrophyl1us,1-S4<1 ,l'2 ,1- SS <13
Ctllundnnlll QCCUllS,1,,_ <l-11" ,14 <1
H~per1cum 10nClOI(JElS3410 ,, <1<1
Geronlum slbblliólOlCI9S<1-<111 3S
/l.cuullmoll'opun:ll0tll<1<12<1 1
1!~¡Jlmcum Corllco~onum"3 32
.\coertocu1mú(1Slochyo,1,1-1
Ciodle 0!l!lregolo,1-<1- ,1
(sp916,IQ hort .•...eglQno23
Pol:¡1.rtctHlm jUnlpennum,1<1-
'\!lroHls1.r1e-hodes,1<l'
LOchum\11ogo1lo1des
,1,llochem\lloorblculQto
4,1153S4,1-605070307090'S8599SO¡,grO$tlsl'lOllnki?ono
30252<1,1SS •303515 1530eolJupoctlol;lns ssss11lfloro
,,_234,l <1-<11 2 1,1S
Vl;lronlCO Sarp~l1\foho1<1,1 1,1
Tl:lrllC':llCtlffiSP.<1-1,1<1<1
I'oóllnnurl<1106,l'
.\grOHIS ~olucen$IS242
Tnrl):lum r~pens2<16
Plllntl:l!jo l~ne'lrl s,,12
UnCln\(l ffien<l{<nSl$30
~romllS u010101(les15
Corto<JenusenconlM15
t:i!J:n!Jltl ri?pens15
Rumex llce~OSl;1l0<1-1<1<1- <1-22S<1'<1', <1-<1
l~fl:l)(jon~llIm pungans5<1315 33<1- ,<1',1'
Enn!j\llm r.urnlla<1<11 1<1 1 ,l' <1<110
'\9(0$11S brfYlculm1S20<1-<1<1-,1- SS<1
BromusloO(l\us,11"-S,13 3,
Creom!jrrh\s onólcolo<12 ,1-23 <1-<1<1
Cl)r(ll( o:"r. C. peucophlloS ,1-10 115 ,1
seneCIO (l'pens323 2 101 <1
-l::J"CD
"Oal...,al3O<CD
COCD
§:o·::J
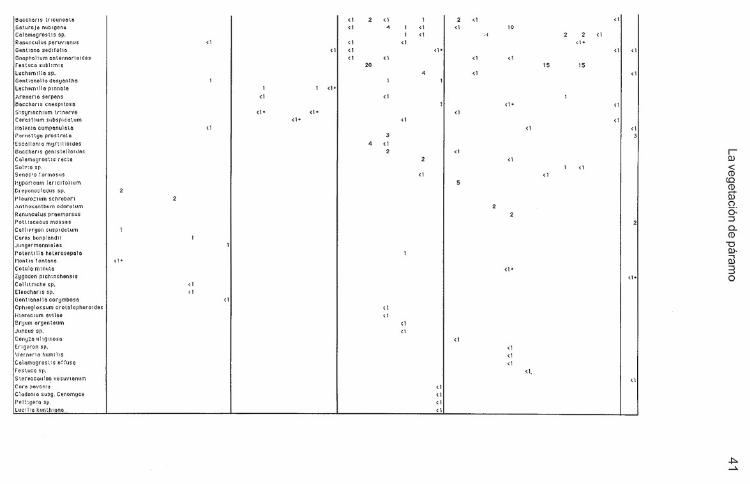
Buccrll~ns lrlcune-oto <12<1 12<1 dSoturoJ<l nt:bl!l(lM
<1•1d <1 10
C01~mO!)rOS~lS Sp.
1<1 22<1
RonunculuS peruvlonus
<1<1<1 d-Ganuono sea:lollo
<1<1 "-<'1<1
Gnophol1um onlennorloldes
<1<1 <1<1
festucoiOubllmlS20ISIS
locMmll1osp .
•<1 I<1
Genllonel1<.\(Josyonlhel
1
loch9mlll~ plnnQI~
1d-,\renono serpens
d<1
8occtoons coesp:tosl)
,,-<1
5:s!jnnchlum tnnerv6
,,-<1- <1
CeroSllllmSlIbsplCQlum<1-d <1
HOiilll10Ctlmponu)¡HO
" <1<1
J't>rn6II!JQprQstnHQ,,
(scQllon:o m!)rtllioHJeS
•<1
Bucct,ons genlslelloHJllS
2<1 rCuloffiugrOStls rec,o
2<1 Q)
So~\,;0 sp
<1<S"n<\·c'O):ormoslls
<1<1 ID
1!!Jp&ncum lorlclfollum
SCO
IDt.. ellllnocio~us SP.
2OiPleuro:wmschriltlsn
2
,\l'Ith()~Ul'lth'lmodorl;l\um
2(")
ROl'lunculusproemorsus
2O:
PotUQceous mosses
2:J
ColilPr!Jon clIspldl;llum
a.Corf;l:->bonp:lln(,lll
ID
.)unoerm~n010¡eS
1 ""O
Pote-n111ioheteroseplllo
1Q).
11ontlofonllll't0
,l' m
COlu\() mlnu~O
<l'3z~goaon p1chlnchenSl$
<1.oCoI1HnChll sp.
<1
Eleochonssp.
,1
GenllonellocorLlmboso
<1
CPhloglossum crotolophoroHJlls
<1
H1MllclUm 0\'1100<1
er!Jum orgen,eum"
.)uncus SI'.
<1
Conyzo 1l~19lr,O)so
<1
[r1glilronsp.
<1
'ilerneno hum1l1S<1
C011)ffitlgros,:s e~fuso<1
fesluco sp.<1.
S \ereOCOll~o~ \'eSUV10numI<1
Coro pOI'onl"<1
Clol:!onlo sUDg.Canoffi!)Ce<1
P('I':'.le-rtlsp.
<1
luclllll klln~hlllno<1

42 The paramo vegetation
The association Pentacalietum vernicosi occurs in the study area between 4000 and 4400 m,either on the foot slopes of scree debris, near the boundary of the superparamo, or on the wetsoils along small ravines in the high paramo proper.
Paramo meadows
A few types of azonal meadows will be described in the present study. Most of them be longto the alliance Lachemillion orbiculatae Cuatrecasas 1934, with the exception of the Muhlenbergietum jastigiatae Cleef 1981 (now Muhlenbergietum cleejii corro Salamanca et al., thisstudy).
The alliance Lachemillion orbiculatae Cuatrecasas 1934 includes most of the meadows with
Lachemilla orbiculata as a prominent species. This taxon replaces the natural vegetation in thenorthem Andes mostJy by repeated buming and intensive grazing at the altitudinal limit of theAndean forest in the northem Andes.
Muhlenbergietum cleefii Cleef 1981 corro Salamanca, Cleef & Rangel (this study).Muhlenbergia c1eefiimeadowPrado de Muhlenbergia c1eefiiTable 5
This association has been described by Cleef (1981) as Muhlenbergietum jastigiatae from theparamos of the Colombian Eastem Cordillera. These humid meadows composed of small cushions of Poaceae were observed in the damp soils transitional to Plantago rigida cushion bogsnear the Otún lake at 3900 m. However, these have also been observed elsewhere in similarhabitats.
Diagnostic taxa of this association are Breutelia spp., Carex pichinchensis and Galium canescens. According to Laegaard (1995), the common Muhlenbergia cushiongrass species growing on damp soil in the Colombian paramos belongs to the newly described M. cleefii Laegaard,and not to M. jastigiata (Pres!) Henr., which is a misidentification.
In the Parque Los Nevados study area also transitions have been noted from this associationto Caricetum pichinchensis resulting in a Breutelia spp. facies and a Sphagnum cuspidatumfacies of this species-poor vegetation type.
Aciachnetum acicularis Vareschi 1953 em. Cleef 1981; corro Salamanca, Cleef & Rangel, (thisstudy).Aciachne acicularis meadowPrado de Aciachne acicularisTable 5
Composition and syntaxonomy:The vegetation of Aciachnetum acicularis is dominated by prickly cushion grasses, and wasoriginally described as Aciachnetum pulvinatae from the Sierra Nevada de Mérida in Venezuela(Vareschi 1953). Laegaard (1987) studied the genus Aciachne, which was earlier believed to bemonotypic with A. pulvinata. However, two new species were recognized by Laegaard (J.c.): A.acicularis and A. flagellifera. Aciachne acicularis has a wide distribution from Venezuela toBolivia, and was also matched for the Los Nevados study area. The association Aciachnetumacicularis can be divided in two subassociations: typicum Cleef 1981 and lupinetosum microphylli Salamanca, Cleef & Rangel 1992 (type relevé AMC 634). The first subassociation is

La vegetación de páramo 43
restricted to the Aciachne acicularis meadows of the Colombian Eastem Cordillera, while thesubassociation lupinetosum microphylli has been identified for the Ruiz- Tolima volcanic massif.Agrostis cf. haenkeana, Hypochoeris sessiliflora, Lachemilla orbiculata and Lupinus microphyl[usare mainly found in the subassociation lupinetosum microphylli. A diagnostic species for theRujz area is Hypericum lancioides, while Calandrinia acaulis and Geranium sibbaldioides arefound in the same association in the Eastem Cordillera (Cleef 1981).
Synecology and distribution:In the Ruiz-Tolima massif, relatively big patches ofthe vegetation belonging to this associationwere observed on the dry and heavily grazed eastem slope ofthe Santa Rosa Volcano between3800 and 4100 m, and in the headwaters of the Quebrada Africa on top of old lake sedimentsat 3800 m.
Similarly Aciachne acicularis vegetation is reported for the paramos ofVenezuela (Vareschi1953; 1970; Monasterio 1980; Berg 1998) and also for Costa Rica (Chaverri & Cleef 1996). InPeru (Gutte 1985) and Bolivia (Seibert & Menhofer 1991; 1992) Aciachne pulvinata ("Gnaphalio-Aciachnetum pulvinatae") vegetation is natural to the dry puna landscape. However, Laegaard (1987) indicated that Aciachne pulvinata and A. acicularis seem almost sympatric in Peruand Bolivia.
Agrostio breviculmis-Lachemilletum orbiculatae Cleef 1981Agrostis breviculmis-Lachemilla orbiculata herb fieldPrado de Lachemilla orbiculata with Agrostis breviculmisTable 5
Physiognomy:These low herbaceous meadows are dominated by ground rosettes of Lachemilla orbiculata. Insome cases small shrubs and/ or bryophytes can attain a high percentage of the total cover. Theassociation was described for the Eastem Cordillera of Colombia (Cleef 1981).
Composition and syntaxonomy:Poa annua, Taraxacum sp. andTrifolium repens are diagnostic taxa for the study area. In theAgrostio breviculmis-Lachemilletum orbiculatae meadows of the Central Cordillera, Agrostiscf. haenkeana is far more common than A. breviculmis.
Synecology and distribution:The meadows of Agrostio breviculmis-Lachemilletum orbiculatae are found in the study area indeforested areas near the altitudinal Iimit of the high Andean forest, 01' as a replacement oftussock grasses in heavily grazed bunchgrassparamos. They occupy an altitude range from 3800m to 4200 m and are widely distributed in the northem Andes.
Aquatic vegetation
During fieldwork in the Otún area of the Parque Los Nevados, a number of quick relevés andobservations were made on the aquatic vegetation (submerged and wet shore communities) ofparamo lakes. This work included observations on the hydroseral sequence (Fig. 13).
The highest lake (Fig. 13e) on the southwest slope ofNevado de Santa Isabel contained submerged vegetation (at 50 cm depth) of Isoetetum karstenii (Cleef 1981). Isoetes karstenii (identification confirmed by DI'. H.P. Fuchs 1980) covered 20% ofthe sandy lake bottom (rel. AMC584). The association Isoetetum karstenii is here for the first time documented outside the Cor-
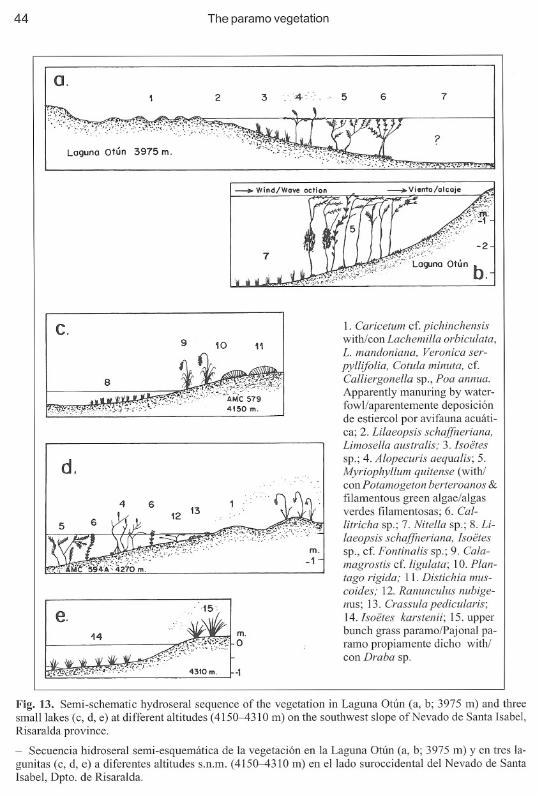
44 The paramo vegetation
a.
1. Caricetum cf. pichinchensiswith/con Lachemilla orbiculata,L. mandoniana, Veronica ser
pyllifolia, Cotula minuta, cf.Calliergonella sp., Poa annua.Apparently manuring by waterfowl/aparentemente deposiciónde estiercol por avifauna acuática; 2. Lilaeopsis schaffneriana,Limosella australis; 3. Isoetessp.; 4. Alopecuris aequalis; 5.Myriophyllum quitense (with/conPotamogeton berteroanos &filamentous green algae/algasverdes filamentosas; 6. Callitricha sp.; 7. Nitella sp.; 8. Lilaeopsis schaffneriana, Isoetessp., cf. Fontinalis sp.; 9. Calamagrostis cf. ¡¡gulata; 10. Plantago rigida; 11. Distichia muscoides; 12. Ranunculus nubigenus; 13. Crassula pedicularis;14. Isoetes karstenii; 15. upperbunch grass paramo/Pajonal paramo propiamente dicho withlcon Draba sp.
11109
-15'
~
e.m.
14 .' .~~.•..;.,>:",:•...• " ..•. O":'-,.~,~";. :,.::.::-
4310m. -1
d.
c.
Fig. 13. Semi-schematic hydroseral sequence of the vegetation in Laguna atún (a, b; 3975 m) and threesmalllakes (e, d, e) at different altitudes (4150--4310 m) on the southwest slope ofNevado de Santa Isabel,Risaralda province.
- Secuencia hidroseral semi-esquemática de la vegetación en la Laguna atún (a, b; 3975 m) y en tres lagunitas (e, d, e) a diferentes altitudes s.n.m. (4150--4310 m) en el lado suroccidental del Nevado de SantaIsabel, Dpto. de Risaralda.

La vegetación de páramo 45
dillera Oriental of Colombia. The species has been reported in Colombia (between about 3500and 4425 m) and also for the Mérida Andes in Venezuela.
In small lakes at 4260 m and 4150 m on the same slope (Fig. 13c, d) Isoetes karstenii wasstill present with low (l 0-15 %) cover.
In these lakes it was found in association with Crassuletum peduncularis Cleef 1981 corroCleef (this study). In the Colombian Eastern Cordillera this syntaxon has been earlier describedas Tillaeetum paludosae Cleef 1981 (based on Tillaea paludosa Schltdl., now a synonym ofCrassula peduncularis (Sm.) Meigen) with two subassociations: isoetetosum and typicum (Cleef1981). For the time being we believe that the six Central Cordilleran AMC-relevés (4100-4170m) belong to the same syntaxon as in the Eastern Cordillera of Colombia: Crassuletum peduncularis Cleef 1981 corr. Cleef (Table 5a).
lA. Steyermark has later described a second species, Crassula (Tillaea) venezuelensis witha distribution area in the high Andes from Venezuela to Pem. We are not convinced that Crassulavenezuelensis (Steyerm.) M. Bywater & Wickens is a 'good species', and maintain for the moment Crassula peduncularis for the study area. Hence, the community should be termed Crassuletum peduncularis, as the only syntaxon in this amphibious habitat on sandy-silty paramo lakeshores.
In Laguna Otún (Fig. 13a,b) a hydroseral sequence was studied from the swampy shores withmire and reedswamp vegetation (different communities of order Marchantio-Epilobietalia Cleef1981) to submerged communities of Hydrocotylo ranunculoides-Myriophylletum quitensis Cleef1981 corro Cleef and deep lake bottom stands belonging to Nitella sp.(Charetea). Patches ofAlopecuris aequalis with filamentous algae and floating Azolla cf. filiculoides, Calamagrostisligulata, Carex bonplandii, Caricetum pichinchensis Cleef 1981, Juncus sp. with Epilobium sp.,Hydrocotyle ranunculoides, Lilaeopsis schajJneriana and Limosella australis have been recorded partly on mud, partly floating, on or close to the unsafe shores of Laguna Otún (3975 m).
Table 5a. Crassu/etum pedicu/aris Cleef 1981 corro Cleef
relevé nr. AMC
580622581594615616
relevé size (m2)
420.5 0.5 22altitud e m
416041704160427041704170water depth cm
2550- 102020
cover % Crassu/a pedicu/aris803570659035
(RJM 6266) Oitrichum sp. (GVR 1226)
2 1565Callitriche sp.
< 1 31/soétes karstenii
101515E/atine sp.
< 12530
Ranuncu/us nubigenus
90Myriophyllum quitense
2
Li/aeopsis schaffneriana1
Lachemi//a mandoniana20
Orepanocladus sp.1< 1
Ca/amagrostis Iigu/ata Uuv.)3
/soétes sp. (RJM 6265)
8Algae
100
Localities:Parque Los Nevados; Nevado S. Isabel, SW slope (AMC 580,581,594), NW slope El Portón (AMC
615,616), Laguna de Silencio, East of Otún lake (AMC 622).

46 The paramo vegetation
Rheophytic vegetation bordering streams at 3730 m in the Quebrada Africa sector includedsmall stands of Montia fontana (AMC 629) and Limosella australis (AMC 628). Limosella australis was further recorded (AMC 647) near the shore of Laguna del Silencio (4150 m) aboveLaguna Otún. In shallow running water of Quebrada Totarito (3320 m) (AMC 628) and in Quebrada Las Damas (3030 m) (AMC 629) bryophyte growth of Rhynchostegium riparioides wasnoted on boulders with 80 and 30% cover respectively. The same species (cover 80 %) was alsodocumented in Quebrada La Sierra near Hacienda La Sierra (3675 m) in the upper forest line(AMC 607). The species seems very common in this rheophytic habitat at the East slope (Tolima)and West slope (Risaralda) of the Parque Los Nevados. This rheophytic vegetation type wasearlier reported from the lower paramos of the Eastern Cordillera of Colombia (Cleef 1981) asDendrocryphaeo latifoliae-Platyhypnidietum riparioides Cleef & Gradstein 1981. At present thecorrect name for this association is Dendrocryphaeo latifoliae-Rhynchostegietum riparioidesCleef & Gradstein 1981 corro Cleef.
The superparamo
From C. 4300 m upwards the vegetation cover starts to be more sparse; the absence of Calamagrostis recta, C. eJJusa and Espeletia hartwegiana is a noticeable feature of the superparamo.Similarly as in the Eastern Cordillera of Colombia (Cleef 1981) the superparamo zone of theCentral Cordilleran study area can be subdivided into an upper (subnival) and a lower superparamo. Small tufts of Festuca breviaristata and Agrostis araucana that form the so-called bluegrasslands (Cleef et al. 1983; Salamanca et al. 1992), are characteristic for the lower superparamo. Replacing the low Pentacalia vernicosa shrubs of the bunchgrassparamo, Loricaria columbiana is the most common shmb in the lower superparamo. Near the lower limit ofthe nivalbelt, the vegetation cover becomes increasingly scattered and only small patches of subnivalvegetation are found. Calandrinia acaulis, Cerastium jloccosum, Draba spp., Erigeronchionophilus, Lachemilla nivalis, Pentacalia gelida and Senecio canescens are almost exclusivetaxa to the superparamo. Distichia muscoides dominated cushion bogs are common in the lowersuperparamo.
Subnival communities
The subnival communities are present at the altitudinallimit of the upper superparamo vegetation. They exhibit a pioneer character, are of variable t10ristic composition, and show conspicuous adaptations to the extreme environmental conditions (Monasterio 1980; Pfitsch 1988). Thevolcanic origin of the substrate, in combination with the diurnal freezing and thawing, haveshaped unique habitats. The ongoing retreat of the glaciers is followed by colonization of thehighly specialized vegetation. The sparse subnival vegetation is found on the coarse, and/or,fine-grained volcanic sandy deposits, where well-differentiated soils are not yet developed.
Syntaxonomy of the subnival communities
Agrostio foliatae-Cerastion floccosae Cleef, Salamanca & Rangel all. nov.AlIiance of Agrostis foliata and Cerastium floccosumAlianza de Agrostis foliata y Cerastium floccosum
Type: Cerastio floccosi-Pentacalietum gelidae Salamanca, Cleef & Rangel 1992Table 6

La vegetación de páramo 47
Physiognomy:The vegetation type ofthis al1ianee is dominated by searee, patehy, open vegetation ofthe lowerand uppermost superparamo. This varies from dominanee of smalJ shrubs, whitish ground rosettes, loose grass tufts, sma11tussoeks and aeroearpous bryophytes and liehens. Bryophytes andliehens l11ayfreely move over the substrate as a eonsequenee of night1y frost heaving.
Composition and syntaxonomy:Diagnostie taxa of the assemblage are Agrostis foliata, Baccharis caespitosa, Bromus lanatus,Cerastiumfloccosum, Draba hallii, D. hazenii, D. pachythyrsa, Erigeron chionophilus, Grammitis moniliformis, Hypochaeris sessiliflora, Luzula racemosa, Lycopodium crassum, Poa trachyphylla, Senecio canescens, Stereocaulon vesuvianum and Valeriana plantaginea.
AIso the eharaeter species ofthe six associations, referred to below, have been included here.This classification is based on relevés from the study area; for a robust c1assification at the orderand c1ass level, it is necessary to compare with superparamo elsewhere in the northern Andes.The assoeiation Racomitrio-Lachemilletum is the vegetation type with highest speeies density.In faet, this vegetation type is 10ca11ypresent on the lowermost border (4150--4250 m) of thesuperparamo. Racomitrium crispulum and Thamnolia vermicularis are present in both the associations Racomitrio-Lachemilletum nivalis and Thamnolio-Racomitrietum nivalis. Those mayconstitute either a suba]]iance, or final1y merge into one association (Thamnolio-Racomitrietumprov.), when more relevés become available. Cuatrecasas (1934) described 'Culcitietum rufescentis-Agrostiosum' from the Tolima Volcano. In this syntaxon, he integrated a11zonal superparamo vegetation types from the Loricaria colombiana scrub at 4320 m up to the last standsat 4500 m (see Cuatrecasas, this volume).
Syneeology and distribution:This type of superparamo is representative for the Parque Los Nevados area. As detailed for theMérida Andes by Pfitsch (1988) daily frost heaving and low temperatures, combined with highradiation levels, are the main abiotic factors. On the Puracé V olcano, some of the associationsbelonging to this a11iance seem also to be present (Rangel & Lozano 1986). In the study area,coml11unitiesofthis alliance extend between about 4150 m and 4600 m.
Cerastiofloccosi-Pentacalietum gelidae Salamanca, Cleef & Rangel1992Subnival Pentacalia gelida dwarf shrubMatorral bajo subnival de Pentacalia gelidaType: relevé SS 63. Table 6; Photo 16
Physiognomy:This open, dwarf shrub-like, vegetation consists only of one sparse layer, about 30 cm high,mainly cOl11posedof endemic Pentacalia gelida, with sma]] eoriaceous leaves, covering between1% and 70 %.
Composition and syntaxonomy:Within this assoeiation two different variants can be recognized: one speeies poor, almost entirely consisting of Pentacalia gelida (variant inops, type relevé SS 63), and another relativelyspecies rich (variant of Stereocaulon vesuvianum, type relevé SS 102).
Synecology and distribution:This vegetation is found on pyroclastic flow deposits affected by diurnal freezing and thawing.The environment is permanent1y wet and cold, and the sandy deposits move according to erionival proeesses. The dwarfshrub of Cerastio-Pentacalietum gelidae was found in the Ruiz-

48 The paramo vegetation
Photo 16. Subnival dwarfshrub patches of Pentacalia gelida (Cerastiofloccosi-Pentacalietum gelidae) inthe upper superpáramo ofthe Nevado del Ruiz, 'arenal' ofthe La Olleta volcano at 4650 m (Caldas province). The dwarfshrubs are characterized by dark green, coriaceous leaves and a well developed root systemo The substrate consists of moving ash deposits with stripes caused by solifluction and frost heave (relevé Cleef & Salamanca 610).
- Vegetación subnival en islotes de arbustillos de Pentacalia gelida (Cerastio floccosi-Pentacalietum gelidae) en el superpáramo alto del Nevado del Ruiz, arenal de La Olleta, a 4650 m (Dpto. de Caldas). Losarbustillos se caracterizan por sus hojas coriáceas verdes y un sistema radicular bien desarollado. En el substrato de cenizas móbiles se oberservan las huellas del efecto combinado de la soliflucción y la gravedad(lev. Cleef & Salamanca 610).
Tolima massifbetween 4400 and 4650 m. The variant inops belongs to the uppermost vegetationlimit, while the variant of Stereocaulon vesuvianum can be found at somewhat lower altitudes.
No similar association has been described for other studied superparamos of Colombia, suchas the Eastern Cordillera or the Sierra Nevada de Santa Marta. Nevertheless, some ofthe specieslike cf. Oreoweisia bogotensis, Senecio canescens and Thamnolia vermicularis, occasional inCerastio-Pentacalietum gelidae, are reported to be common in the superparamo communitiesdescribed by Cleef (1981) for the Colombian Eastern Cordillera. It is interesting to note thatdwarshrub of Pentacalia (Senecio) guicanensis, a vicariant community occurs undel' similarenvironmental conditions in the Sierra Nevada del Cocuy.
Senecioni latiflori-Calamagrostietum ligulatae Salamanca, Cleef & Rangel 1992Subnival Senecio latiflorus-Calamagrostis ligulata vegetationVegetación subnival de Senecio latiflorus y Calamagrostis ligulataType: relevé AMC 590. Table 6; Photo 17
Physiognomy:This vegetation is composed of scattered ground rosettes of Senecio latiflorus together withsmall cushions of Werneria humilis and tussocks of Calamagrostis ligulata.

La vegetación de páramo 49
Two ]ayers are present;- tussock ]ayer, with cover of Calamagrostis ligulata ranging between 5 and 80%,- ground layer, main]y composed ofthe cushions of Werneria humilis, with cover between 3%
and ]5%. Bryophytes may cover as much as 90% in the ground ]ayer.
Composition and syntaxonomy:Diagnostic species of this association are Calamagrostis ligulata, Lucilia kunthiana, Seneciolatiflorus and Werneria humilis ..
The association of Senecioni latiflori-Calamagrostietum ligulatae has t10ristic affinities withsome of the communities with Calamagrostis ligulata described by C]eef (1981) for the highparamos ofthe Colombian Eastern Cordillera, viz. the Sierra Nevada del Cocuy and the Nevadode Sumapaz. A]so, site conditions on wet 01'damp ground are are indicative to azonal vegetation.For practica] reasons, the association of Senecioni-Calamagrostietum ligulatae is being provisionally included under the alliance Agrostio-Cerastion jloccosae.
Note: The formal name of this association was erroneously omitted at the base of p. 57 inSalamanca (1992). The name of this association however was correctly quoted on p. 58 under'Composición y sintaxonomía'.
Synecology and distribution:Vegetation belonging to this association is present on humid coarse-grained volcanic sands, indepressions in small ravines in the high superparamo, between 4400 and 4500 m, in the ParqueLos Nevados. In fact, this vegetation type is rather azona] than zonal.
Photo 17. Vegetation of Calamagrostis liguiata and ground cover of Bryum sp. (Cleef 10432) (Senecionilatiflori-Calamagrostietum ligulatae) at 4470 m in the upper superpáramo ofthe western slope ofthe NevadodelRuiz (Caldas province). The small valley bottom between both moraines is permanently wet as aresultof seepage fram the surrounding slopes (relevé Cleef & Salamanca 612).
- Vegetaciónde Calamagrostis ligulata y tapetes de Bryum sp. (Cleef 10432) (Senecioni latiflori-Calamagrostietum ligulatae) a 4470 m en el superpáramo alto dellado occidental del Nevado del Ruiz (Dpto.deCaldas).En el vallecito entre las dos morrenas hay escolTentíapermanente (lev. Cleef & Salamanca 612).

50 The paramo vegetation
Senecioni canescentis-Cerastietum floccosi Salamanca, Cleef & Rangel 1992Senecio canescens-Cerastium floccosum superparamo vegetationVegetación subnival de Senecio canescens y Cerastium floccosum
Type: relevé SS 139. Table 6; Photo 18-23; (see also vol. 1, Photo 8, 9)
Physiognomy:This association consists of very sparse superparamo vegetation at the lower limit of the snowline. The total cover ranges fram 5% to 20%.
Composition and syntaxonomy:Cerastium jloccosum, Poa trachyphylla andSenecio canescens are diagnostic species. This syntaxon is poor in species, with an average of only 5 vascular species. The Senecioni-Cerastietumis differentiated against all other here studied subnival associations by the absence of their respective diagnostic species.
Distribution and synecology:Subnival vegetation of Senecioni canescentis-Cerastietum jloccosi occurs in the loose, not yetstabilized, sandy volcanic deposits, but also covers superparamo morraines (TPN 2). This vegetation was found between 4400 and 4550 m; mainly on the slopes ofthe Ruiz and Santa Isabelvolcanoes. Superparamo vegetation, with Senecio canescens and Poa paucijlora, is also described by Rangel & Lozano (1986) fram Puracé Vo1cano.
The next syntaxa do not properly belong to the subnival belt, but are characteristic of thetransition subnival - lower superparamo blue grasslands. There they occur on the lava escarpments, in contact with the blue grasslands.
Photo 18. Whitish ground rosettes of Senecio canescens (Senecioni canescentis-Cerastietum jloccosi) at4570 m in the upper superpáramo ofNevado del Ruiz, 'arenal' ofLa Olleta, constituting the almost upperlimit of vascular plant growth (Caldas province). The substrate consists of unstabilized volcanic sand andash deposits that are subjected to daily solifluction. The snow patches persist only few hours in the shadowofthe rosettes (relevé Cleef & Salamanca 611).
- Rosetas blancas acaules de Senecio canescens (Senecioni canescentis-Cerastietumfloccosi) en el límitealtitudinal de la vegetación vascular en el superpáramo alto del Nevado del Ruiz, arenal de La Olleta, a 4570m (Dpto. de Caldas). El substrato consiste de depósitos de arenas volcánicas sueltas, todavía no estabilizados, debido a la acción diaria de la soliflucción. Los parches de nieve persisten unas horas en la sombra delas rosetas (lev. Cleef & Salamanca 611).

La vegetación de páramo 51
Photo 19. Vegetation aspect of the upper superpáramo with ground rosettes of Senecio canescens (Senecioni canescentis-Cerastietumjloccosi) at the high pass (4550 m) between the voJcanic cones ofNevadodel Ruiz and La Olleta (Caldas province).
- Aspecto de la vegetación del superpáramo alto con rosetas esparcidas de Senecio canescens (Senecionicanescentis-Cerastietumjloccosi) en el paso alto (4550 msnm) entre el edificio principal del volcán del Ruizy su cráter adventicio La Olleta (Dpto. de Caldas).
Photo 20. Upper limit of the vascular vegetation at about 4600 m on La Olleta (Nevado de Ruiz, Caldasprovince). Whitish ground rosettes of Senecio canescens (Senecioni canescentis-Cerastietum jloccosi) aregrowing together with dark, greenish, dwarfshrub of Pentacalia gelida on the unstable 'arenales' ofthe upper superpáramo.
- Limite altitudinal de la vegetación vascular a ca. 4600 m en La Olleta (Nevado de Ruiz, Dpto. de Caldas).Observan rosetas blancas acaules de Senecio canescens (Senecioni canescentis-Cerastietum floccosi)creciendo junto con arbustillos verde oscuro de Pentacalia gelida en los arenales mobiles del superpáramoalto.

52 The paramo vegetation
Photo 21. Draba pennell-hazenii shown as tiny stem rosettes colonizing the unstable 'arenales' ofthe superpáramo at 4350 m on the northwestern side ofthe Nevado de Ruiz (Caldas province).
- Draba pennell-hazenii en forma de caulirrósula pequeña que coloniza los arenales del superpáramo a4300 m en el flanco norte occidental del Nevado Ruiz (Dpto. de Caldas).
Photo 22. Branched stem rosette up to 20 cm height ofyellow-flowered Draba pennell-hazenii at 4350 min the shelter ofvolcanic outcrops in the superpáramo ofthe northwestern side ofNevado de Ruiz (Caldasprovince).
- Caulirrósula ramificada de Draba pennell-hazenii (hasta 20 cm) en un abrigo de roca volcánica en el superpáramo a 4350 m, flanco norte occidental del Nevado de Ruiz (Dpto. de Caldas).

La vegetación de páramo
':;:':~~::~:~::~'.:'~':
,- u_· ::,'2':'~';%;~;~f\::'~·i:
53
Photo 23. Superpáramo vegetation at 4350 m in a landscape ofmobile 'arenales' (caused by daily solifluction) and volcanic outcrops on the nOlihwestern side ofthe Nevado de Ruiz (Caldas province). Vegetationcomprises low shrub of Loricaria coLombiana, bunches of Festuca sp., reddish Lycopodium (Huperzia)crassum and grayish patches of StereocauLon vesuvianum.
- Vegetación de superpáramo a 4350 m en paisaje de arenales móbiles (con soliflucción diaria) y afloramientos de roca volcánica en el lado norte occidental del Nevado de Ruiz (Dpto. de Caldas). Se encuentrantambien arbustillos de Loricaria coLombiana, macollas solitarias de Festuca sp., tallos rojizos de Lycopodium (Huperzia) crassum y parches grisáceos de Stereocaulon vesuvianum.
Elaphoglosso mathewsii-Muehlenbeckietum vulcanicae Salamanca, Cleef & Rangel 1992Elaphoglossum matthewsii-Muehlenbeckia vulcanica vegetationVegetación de Elaphoglossum matthewsii and Muehlenbeckia vulcanicaType: relevé AMC 586. Table 6; Photo 24
Physiognomy:This vegetation consists oftwo layers: the first one, 15 cm high, is mainly composed ofthe fernElaphoglossum mathewsii with cover between 2 and 20%. Predominant in the ground layer arethe lichen Stereocaulon vesuvianum and the small creeping Muehlenbeckia vulcanica, with coverages for both species from 15 to 40%.
Composition and syntaxonomy:Muehlenbeckia vulcanica is the exclusive character species for this association. Elaphoglossummathewsii is also highly diagnostic fOl'this association, which is rather poor in species. Groundrosettes of Senecio canescens and Hypochaeris sessiliflora are absent
Synecology and distribution:The asociation of Elaphoglosso mathewsii-Muehlenbeckietum vulcanicae is found within threedifferent habitats: on the outcrops of volcanic andesites, on 10w stony ground moraines, and incoarse-grained pyroclastic flow deposits. In the Ruiz- Tolima volcanic massif, vegetation patchesofthis association were observed fringing the 10wermost superparamo between 4250 and 4380 m.

54 The paramo vegetation
Photo 24. Fems of Elaphoglossum mathewsii (Elaphoglosso mathewsii-Muehlenbeckietum vulcanicae)growing at 4375 m between volcanic roche moutonnée in the transition between the 'blue grassland' ofthelower superpáramo and the subnival vegetation at the southwestern side ofvolcano Sta. Isabel (type relevéCleef & Salamanca 586; Risaralda province).
- Helechos de Elaphoglossum mathewsii (Elaphoglosso mathewsii-Muehlenbeckietum vulcanicae) a 4375m en los afloramientos de roca volcánica pulida por acción del hielo en la transición entre los pastizales azules del superpáramo bajo y la vegetación subnival del superpáramo alto en el lado SW del Volcán de Sta.Isabel (lev. tipo Cleef & Salamanca 586; Dpto. de Risaralda).
Racomitrio crispuli-Lachemilletum nivalis Salamanca, Cleef & Rangel 1992Racomitrium crispulum-Lachemilla nivalis vegetationVegetación de Racomitrium crispulum y Lachemilla nivalisType: relevé AMC 642. Table 6. (see also Cleef 1981, Fig. 29)
Physiognomy:This association is composed ofhanging carpets ofmosses and low (up to about 30 cm) grayishground rosettes on outcrops ofvoIcanic rocks exposed to frequent fog and atmospheric humidity.This results from the uppermost condensation zone at this altitude.
Composition and syntaxonomy:Diagnostic species are Aa (Altensteinia) cf. colombiana, Castilleja jissifolia, Disterigma empetrifolium, Hieracium avilae, Lachemilla nivalis, Ourisia chamaedrifolia, Polystichum polyphyllumand Racomitrium crispulum (generally cover from 40 to 70%). Also diagnostic is the absence ofCerastium floccosum and Senecio canescens. Racomitrium lanuginosum and Rhacocarpus purpuracens reported by Cleef(1981) as character species for a similar Racomitrium crispulum vegetation type in the Colombian Eastem Cordillera, are found only in the plot of the type relevé(Racomitrium lanuginosum 30% cover). This association shows affinities with that of Loricarietum complanatae racomitrietosum crispuli Cleef 1981 ofthe Eastem Cordillera.

La vegetación de páramo 55
Synecology and distribution:The bryophyte vegetation belonging to Racomitrio crispuli-Lachemilletum nivalis was studiedat the humid, exposed northwest side ofthe Ruiz Volcano within a narrow altitudinal range from4150 to 4250 m. Patches have also been observed on the west slopes of the Santa Rosa and theSanta Isabel voleanoes. Rangel & Lozano (1986) reported patches of Racomitrium crispulum at4250 m on the Puracé Voleano. Another stand belonging to this association was recorded at4100 m on the Nevado de Sumapaz, Eastern Cordillera, as was referred to by Cleef(l98l) under'Other zonallower superparamo communities'.
Thamnolio vermicularis-Racomitrietum crispuli Salamanca, Cleef & Rangel1992Thamnolia vermicularis-Racomitrium crispulum pioneer vegetationVegetación pionera de Thamnolia vermicularis y Racomitrium crispulumType: relevé SS 104. Table 6; Photo 25
Physiognomy:The physiognomy of this vegetation is similar to that of the ground layer of the Racomitrietumcrispuli-Lachemilletum nivalis described above. The conspicuous moss layer covers between 40and 95%. A low herb layer, with cover from 2 to 30%, is also present.
Composition and syntaxonomy:The poorness in species diversity is a diagnostic character when compared to the previouslydescribed Racomitrietum crispuli-Lachemilletum nivalis. Thamnolia vermicularis is a free moving lichen species common to both associations with Racomitrium crispulum, described here.Thamnolio vermicularis-Racomitrietum crispuli has affinities with the community of Thamnoliavermicularis-Alectoria cf. ochroleuca described by Cleef (1981) from the paramos ofthe EasternCordillera, although the latter one is richer in lichens. Both vegetation types have Thamnoliavermicularis and Draba spp. in common.
Photo 25. Detail (ofPhoto 23) showing patch of Stereocaulon vesuvianum (lichens) associated with Racomitrium crispulum (dark mosses), grasses and a small dwarfshrub of Loricaria columbiana.
- Detalle de la foto 23. Parche en el cual predomina Stereocaulon vesuvianum (líquenes), asociado con Racomitrium crispulum (musgos oscuros), gramíneas, y un arbustillo de Loricaria columbiana.

56 The paramo vegetation
Synecology and distribution:The association of Thamnolio vermicularis-Racomitrietum crispuli is found on andesite outcropsofthe lower superparamo. The vegetation starts growing in small depressions in the rocks, wherelitter, as wel! as fine-grained ashes, are accumulated. When the vegetation is well developed, largecarpets of mosses hang over the vertical rocky wal!s. These are kept permanently wet by seepagefrom ground water. In the study area this association was found between 4200 and 4450 m.
Rangel & Lozano (1986) reported the occurrence of Racomitrium crispulum in the superparamo of the Puracé V olcano from 3960 to 4250 m. But, according to these authors, these vegetation types are very scarce and were mainly found on volcanic sand deposits. Similar mossvegetation associations (between 3400-3800 m), with Grammitis moniliforme, Racomitriumcrispulum, R. lanuginosum and Stereocaulon vesuvianum have been studied by Chaverri & Cleef(1996) from paramo outcrops in Costa Rica. AIso, patchy vegetation with Racomitrium lanuginosum var. pruinosum have been reported growing in the volcanic rock crevices from the summit of Mt. Mauna Loa (Hawaii) at 4150 m (Fosberg, 1959). Villagrán (1980) studied extenseRacomitrium lanuginosum vegetation on the Osomo Volcano in Chile.
Superparamo blue grasslands
This plant formation contains al! the grasslands of the lower superparamo belt. In contrast withthe subnival vegetation, they always occur on the so-called fine-grained volcanic ashes (Kloosterman et a!., chap. 5, this vo!.).
Syntaxonomy of the superparamo blue grasslands
Festuco breviaristatae-Agrostion araucanae Cleef, Salamanca & Rangel al!. nov.Alliance of Festuca breviaristata and Agrostis araucanaAlianza de Festuca breviaristata y Agrostis araucanaType: Lupino alopecuroides-Agrostietum araucanae Salamanca, Cleef & Rangel 1992Table 7
Physiognomy:The aspect of the vegetation of this alliance is an open grassland, consisting of low tufts with abluish hue. The open ground 1ayer consists of ground rosettes, small herbs and prostrate dwarfshrubs. Conspicuous are whitish inflorescences of Lupinus alopecuroides and Senecio canescenso Reddish clumps of Lycopodium (Huperzia) crassum locally reach high densities.
Composition and syntaxonomy:Agrostis araucana and Festuca breviaristata are diagnostic species ofthis al!iance, characterizedby high presence and cover values. The same applies for Bromus lanatus, Cerastium jloccosum,Erigeron chionophilus, Hypochaeris sessiliflora, Lachemilla nivalis, Luzula racemosa, Lycopodium (Huperzia) crassum, Senecio canescens, Stereocaulon vesuvianum and Valeriana plantaginea. The new alliance includes Lupino-Agrostietum araucanae, Baccharido-Agrostietumaraucanae and provisionally Diplostephio-Loricarietum columbianae.
Synecology and distribution:The so-called 'blue grasslands' are present in the lowermost part ofthe superparamo belt, wherethe vegetation cover may be close to 100%. Frost heaving is an important daily phenomenonhere, but also pedogenetic processes develop under the closed tussock layer. The first species

La vegetación de páramo 57
indicative of grassparamo indicate, that this grass vegetation is transitional between the superparamo being dominated by large patches ofbare ground, and the bunchgrassparamo proper. Inthe study area, small tussock grasslands of Festuco-Agrostion araucanae are found between c.4300 and 4470 m. Observations from Acosta-Solis (1984) indicate that this vegetation type maya1sobe present in the Ecuadorian Andes.
Cleef et al. (1983) studied a superparamo blue grassland of Agrostis araucana and Festucabreviaristata with Lupinus alopecuroides, Lycopodium crassum and Senecio canescens which,though it was not ranked at the time, also belongs to this alliance.
No similar superparamo blue grasslands have been described from the other two ColombianCordilleras, nor from the Sierra Nevada de Santa Marta. Agrostis araucana has been reported(according to herbarium records) from Colombia to Chile and Festuca breviaristata as far southas Bolivia (Jorgensen & Ulloa Ulloa 1994).
Lupino alopecuroides-Agrostietum araucanae Salamanca, Cleef & Rangel1992Agrostis araucana blue grassland with Lupinus alopecuroidesPastizal azul de Agrostis araucana con Lupinus alopecuroidesType: relevé TPN 22. Table 7; Figs. 14, 15; Photo 26-33; (see also vol. 1, Photo 10, 11)
Physiognomy:This vegetation consists of two layers;- tussock layer, mainly composed of Agrostis araucana tufts, and some ground rosettes. The
cover ranges from 20 to 70%. Conspicuous are the woolly columnar inflorescences of Lupinusalopecuroides,
- ground layer of small herbs, achieving maximally 15% of the total cover.
Composition and syntaxonomy:Diagnostic species of this association are Draba hallii, Gentiana sedifolia, Lucilia kunthiana,Lupinus alopecuroides, Poa trachyphylla, Senecio isabelis, S. latiflorus and S. repens. Furtheralso: Bartsia sp., Draba lindenii, Halenia campanulata, Lachemilla holosericea, Lysipomia museoides, Senecio summus, Valeriana plantaginea and Werneria humilis. The association has claseaffinities to Baccharido-Agrostietum araucanae.
Synecology and distribution:The association Lupino alopecuroides-Agrostietum araucanae was studied on the fine-grainedsandy deposits ofthe lower superparamo on the Santa Isabel Volcano, between 4350 and 4500111. This vegetation type is very common in the Parque Los Nevados.
Rangel & Lozano (1986) and Duque & Rangel (1989) reported lower superparamo grasslands(3950 to 4380 m) withLupinus alopecuroides and Valeriana microphylla on the Puracé Volcano.However, Agrostis araucana was very sparse and Festuca breviaristata was absent.
Baccharido caespitosae-Agrostietum araucanae Salamanca, Cleef & Rangel 1992Agrostisaraucana blue grassland with Baccharis caespitos aPastizal azul de Agrostis araucana con Baccharis caespitosaType: relevé SS 3. Table 7

PASTl2AL AZULLUPINOAI..OPECUROlDES·BACCHARlDO CAESPITOSAE·OIPLOSTEPHIO ERIOPHORI-
TABLA CONSTANCIAAGROSTIEIl.JMARAUCAHAEACiROSTIElUM ARAUCAJiAElORICAAltTUM COLOMDIANAE
NÚr.lCIO levantamiento5117 SI09 e60S $126 S112 T22 T28 SI25 T29 S108 5119 S121 SI20S'72 S'73 S67 S70535141 S142 S65 5665144S38 SUS C609 C593 C606 5101515
Altitud (m,s.n.m.)4380435044604390434044204470435044004360440044704400438043$043804380422044004400 4380438043704390 4430 4280 4275 4400 4280 4450
Loe;¡lidadesSI SI SR C $1 SR SI SI $1 SI SI SI $1RRRRRRRRRRSI1RROSl1RC
Arca25 25 24 25 25 50 SO 25 SO 25 25 2S 2525 18 25 25 6 40 25 25 25 2525 25 25 25 30 25 25
Pendiente55551054420525 1527225 54452221720251065'0155
Exposición""">.E>.EB\E SSW $SE SNSSESSS< """E>E>.EEy..,wv 'r/INN W&N W5W NgN SES INSW WSW SIiNV'MNI>.EN>.EN"'""'">.EN
i Taxa aprox17,.17.01720241529"2Z2027,.2Z29•2t179816'820Z526196152t
%E~\fato mileollas203765"Z9706555SO47,.422t2t"56,169Z867'3111810911101510
'Y.oEwato helbáeeo65447214770464314152Z151Z35",11•". S119S'0 •181 S430
%ESUoltO faSante10,.254'0 4". ·115"8,."•43 94.o3,5321423 .4
''/"Musei65708 15 55'5 ,3' 70"706070"d. 20406040 5625907525
%liehens9<12 .o,. 76• O15· ,.413 5 607l.23Z93 2,1
%Ew<tto afbustivo,12 4•2 ,..,1 19155191739060J8
%Arcn;¡s dcsnud<1S,120505 ,13<137•5·55 •30 20 'O
%Roc.-tS
5"·3. 15715 82520 506040330 5TAXA Lupinus alopecUfoidcs
<1,.•10Z5,<1,1,1.oSenccio laliHorus
<140 15ISd,.11Scnccio repens
d3 d•1,2 ,1Gelltl<tIli:l sedilolia
.o,..d. ,1,1,1Lueili;¡ kunltlíana
d<1,1 d ,1PO,¡ trilehyphylla
5530 dAgfostis foliata
dISIS 2Scnccio isabclis
5,1,.dC.lI<lmagrostis ¡lgulclta
,2 ,1dOraba hallii
d". ,..Baccharis caespltosa
5•2354d ,32552
Pcnlaealia gelida2•2 ,l.d 2,1,11
lupjl1u.s míerophyllus1" 4 ,1d
Gc¡anium sibbaldioidcs33,1 d i
Aumcx acetosella3
d,l.EringlUnl humile
<1,1R¡munculus praemorsus
d,1Hicr;¡cium avilae
ddLoricalia colombiana
2,.50.070'0,O30
DiploSlcphium eriopholumd15,1,•2 ,1
Pemenya proslrata1.o 12
Oisterigma empetritolium261
POltiilceOI,lS mossesa 80
C,tlam3groslis ellusao10
POlysllchum polyphyllum2.
Dfil'oa pcnnell·hazeniid,.
Jamcsoniella rubricaulis50
Racomitrium erisputum50
Orcowcisia bogolensisZ5
Afcytophyllum eL arislalumIS
Vaccinium lIoribundum5
Tortula arldic-ola5
Aíeecudía sp.5
Agroslis araucana15755 344025d6Z530Z5zo101Z535657237•1 ,..
Bromus lanarus,110'5d•20 17,1,1d126,l. 4·1510410 2d1 d
Hypochoeris st!$$ili~<lra3125425 1<1,11242215•d. ,1 223 2 d
Erigeron ehionophilus313,.. 304d. •22 44114,.. ·,l. 215 45
luzula raccmosa,.1.,d. d.,. d25 7,..,1,d35d2<1. d,1
FC.sluC<1 brcviariSl3\a5Z5 152030 5Z515515 ,o10255dZO50 7
lycopodium erassum330210433,.10",l. ,. 102 220
SIN{!OCaulon yesuvianum,1,1 ,.,.10157 22125 205 151 1
lachemilJa nivalis" ,.,.27¡3 ,1d. 33,552•
Scnccio canescr'ns240 50Z515 10 ,.,.85 d2,1<1 d
CNastium f1oeeosum,11 d7d ".,1d,. ,.,. 2,d d
Valclíana plantaginea4.,l. 527 .5 1d 13510 d·
B:msía el. pedicularioides,l.d,112,1,..o ,lo<1 <1,
Cerastium subspicawm,1•,1,1 5,.. ,1,.<1<1,1,.. <1
Oreomyrrhis andieola2,.,. ,l.d,.·,1 ,.<1,.. 2 .o
PClligcra sp.,1,.. ,. ·,l·2,. .o2 eZ5
Wcrncli¡¡ humilis7,..<1"•5 2 1
GI¡Jmmites moniJilormisd,1,l.,1 ,13 ,.<1
Arcn¡¡ria sClpcnsd.d.,l. <1.<1,. d,. I -1 (J1
Q)ex:>
"Ut:i
OliD~ ¡;¡':-.J
9!..OJOleN CDe :- co
-103
Ol
(j)CY
(j)
Ol
¡¡¡o..
::J
o..CD ~() OO::J
O(j)
::J¡¡j
(j)::J
¡¡j()
::Jw'
()'< -1::J ¡¡j :::¡-o..
CD
()
CY
"OOlro
Ol::J ::J03
o.. Oo..
:3¡¡¡
o'O
~<() ::JCD
O coco
:3CD
"Og:
§:OCD
~.:::!l
o'()
~ ::J
O; ::J~- ()
Q ()~
O:3()
"OOl
O'<
(j);::;:¡¡¡ o'
CD
::J
~Ol
2::J
o..() g:e03
CD
~e()ero

Aren,ido !;crpens d.d·< •• .1..<.<1
Gnaphalium 3n1ennarioidcs<.< • <1 <l.<1.1.
Gcrllianclla das)'antha2 7,lO
Orilropnium peru.,.¡anum< •.5I4 d
laehemilla holosericeaddd<l. d
AltcnSleinia paludosa<. <.d·<1d
Halc·nia campanulataI<.d <l.
Tharllnolia \lctmicularisdd <1
C<llamagroslis recta5 I 515
Campylopus sp<11010
CI:rdonia subg. Cenomyced·< • .<1
lysipomia museoides< •.1 d<1
Colobanthus quitensisd<1dd
EI¡¡phoglossum mattlewsii<.d· 10
Hypericum lancioides<.<.
Azololla sp. Senecto summusClaoia ag;;¡rcgalaCalox alt. C. peucophilad
Oiplostephium schultzij4<1d r
COla paOJoniad.<. O)
Calamagrostis sp.<1'0<Pcnt,1co11ia vernicosa eo
Oraba lindenii<1 (Q
Jamesonia goudolii<1.eo
MClzgcria sp.<1. <.¡¡)
Calandrinia acaulis.<.d ()Monlia meridense
d<1 O:Oraba pachythyrsa<1<1 ::lTaraxacum sp.
d.1o..Racomitrium lanuginosum 25
Niphogcton dissecta, eo
Poaannua
3 'OZygoo'on píchinchensis
2 0)-Espcletia hartwegiana
...,O)CaSfillcia lissitolia :3Pohlia sp LcprOCiSulon congesfum
OOiplostcphium el. rosma¡initolium
Fcstuca sublimis<l.
Lcptodonlium sp
I
<l.I-{clbcrtus sp.
, <l.BrachiolcjcunciS sp.
< ••Kurzi¡l sp
<l.Wcmcria Cltlssa Agloslis haenkcan3Anaslrophyllum spAphanaclis jamcsonianaOphio910s~um clotalopholoida!:Senado sp. nov. flol rosada
<1Baccharis macranrha
<'Sisyrillchium lJinerve
dAnlhoxanttlum ocloralum
dOiploschistes sp
<1BrYl1m algenleum
dPolytrichum juniperinurTl I
<1Plaqiochila sp.
<1
(J1
<!)

60 The parama vegetatian
TPN 284475 m
1 m
Fig. 14. Zonallower superpáramo ofblue grassland of Lupino-Agrostietwn araucanae at 4475 m northeastofthe pass on the southeast slope ofNevado de Santa Isabel, Quebrada Africa (relevé TPN 28), Tolima province. See Photo 30 and 31.
- Pastizal azúl zonal del superpáramo bajo del Lupino-Agrostietum araucanae a 4475 !TI al nOlieoccidentedel paso al suroriente del Nevado de Santa Isabel, Quebrada Africa (lev. TPN 28), Dpto. del Tolima. VéaseFoto 30 y 31.
TPN 1184460 m
Fig. 15. Semi-schematic cross-section of lower superparamo vegetation of low scrub (Diplostephio eriophori-Loricarietum colombianae) in contact with 'arenales' and different facies of blue grassland (LupinoAgrostietum araucanae) in the summit area at 4460 m at the northwest side ofSanta Rosa volcano, Risaralda province.
- Corte semi-esquemático de la vegetación del superpáramo bajo con matorral bajo (Diplostephio eriophori-Loricarietum colombianae) en contacto con arenales y diferentes facies del pastizal azul (Lupino-Agrostietum araucanae) en la zona del cumbre a 4460 m en el flanco norteoccidental del Volcán de Santa Rosa,Dpto. de Risaralda.
Physiognomy:This vegetatian is mainly stratified into two layers;- tussock layer covering between 10 and 70%,- ground layer, covering from 3 to 25%, consisting of ground rosettes and prostrate dwarf shrubs.Sometimes scattered shrubs of Pentacalia gelida can be observed.

La vegetación de páramo 61
Photo 26. Blue grassland belonging to the Lupino alopecuroides-Agrostietum araucanae association inthe lower superpáramo ofLagunares de Santa Isabel at about 4300 m. (Tolima province).
- Pastizal azul perteneciente al Lupino alopecuroides-Agrostietum araucanae en el superpáramo bajo deLagunares de Sta. Isabel a ea. 4300 m (Dpto. del Tolima).
Photo 27. Zonal and azonal superpáramo vegetation at 4300 m in the area ofLagunares de Sta. Isabel (Tolima province). Blue grasslands of Lupino alopecuroides-Agrostietum araucanae comprised Agrostis araucana, Festuca breviaristata and Senecio canescens. Lycopodium (Huperzia) crassum is present on the lowmorraines and on a cushion bog of Distichia muscoides (Floscaldasio hypsophilae-Distichietum muscoides) in the glaciallake.
- Vegetación zonal y azonal del superpáramo a c. 4300 m en Lagunares de Sta. Isabel (Dpto. del Tolima).Pastizales azules de Lupino alopecuroides-Agrostietum araucanae con Agrostis araucana, Festuca breviaristata y Senecio canescens. Lycopodium (Huperzia) crassum sobre morrenas bajas y por encima de cojines de Distichia muscoides (Floscaldasio hypsophilae-Distichietum muscoides) en la laguna glaciar.

62 The paramo vegetation
Photo 28. Uppermost fringe of the blue grassland (Lupino alopecuroides-Agrostietum araucanae) of thelower superpáramo at 4450 m on a recent moraine on the northwestern slope ofNevado Sta. Isabel (relevéTPN 3; Risaralda province).
- Franja superior del pastizal azul (Lupino alopecuroides-Agrostietum araucanae) del superpáramo bajosobre una morrena reciente a 4450 m en el lado norte occidental del Nevado de Sta. Isabel (lev. TPN 3;Dpto. de Risaralda).
Photo 29. Blue grassland (Lupino alopecuroides-Agrostietum araucanae) of the lower superpáramo at4550 m on the eastern side ofthe surnmit area ofvolcano Sta. Rosa (Risaralda province). Conspicuous arethe whitish ground rosettes of Senecio canescens and reddish plants of Lycopodium (Huperzia) crassum (relevé TNP 8).
- Pastizal azul (Lupino alopecuroides-Agrostietum araucanae) del superpáramo bajo a 4550 In en la zonaoriental de cumbre del Volcán de Sta. Rosa (Dpto. de Risaralda). Son notorias las rosetas blancas acaulesde Senecio canescens y los tallos rojizos de Lycopodium (Huperzia) crassum (lev. TPN 8).

La vegetación de páramo 63
Photo 30 and 31. Blue grassland (Lupino alopecuroides-Agrostietum araucanae) ofthe lower superpára1110 al 4475 m in the headwaters of Quebrada Africa, northeast of the pass located southeast ofNevado Sta.Isabel (relevé TPN 28; Tolima province). See Fig. 14.
- Pastizal azul (Lupino alopecuroides-Agrostietum araucanae) del superpáramo bajo a 4475 m al norteoriente del Paso, al suroriente del Nevado de Santa Isabel en las cabeceras de la Quebrada Africa (lev. TPN28; Opto. del Tolima). Véase Fig. 14.

64 The paramo vegetation
Photo 32. Blue grassland (Lupino alopecuroides-Agrostietum araucanae) ofthe lower superpáramo closeto the pass between the valleys of Qtún and Quebrada Africa, southeast of Nevado Sta. Isabel at 4380 111
(Risaralda province). A species poor variant of the Lupino-Agrostietum araucanae association covers theflat sandy bottom ofthe former glaciallake (4335 m) surrounded by moraines on the Qtún side ofthe pass.
- Pastizal azul (Lupino alopecuroides-Agrostietum araucanae) del superpáramo bajo cerca del Paso a4380 m al suroriente del Nevado de Santa Isabel, entre los valles de Qtún y Quebrada Africa (Dpto. de Risaralda). Se encuentra una variante más pobre en especies de la asociación Lupino-Agrostietum araucaneaeen el fondo plano y arenoso de una antigua laguna glaciar (4335 msnm) rodeada por morrenas en el ladooccidental del Paso.
Composition and syntaxonomy:Baccharis caespitosa val'. alpina, Pentacalia gelida, Lupinus microphyllus, Geranium sibbaldioides, Rumex acetosella, Hieracium avilae, Eryngium humile and Ranunculus praemorsus arediagnostic species. The association also includes a Gentianella dasyantha variant (val'. nov.;type rel. S 67) with most ofthe diagnostic species for the association and a species pOOl'variant,variant typicum (val'. nov.; type rel. S 65).
The association Baccharido-Agrostietum araucanae shows close affinities with LupinoAgrostietum araucanae, but also with Diplostephio eriophori-Loricarietum colombianae. Baccharis caespitosa, Geranium sibbaldioides, Lupinus microphyllus and Pentacalia gelida occurin both vegetation units.
Synecology and distribution:The association Baccharido caespitosiae-Agrostietum araucanae was studied on the sandy volcanic deposits affected by periglacial processes. The association reported here is on the lowersuperparamo ofthe Ruiz, the Santa Isabel and the Santa Rosa volcanoes, between 4200 and 4400 lll.

La vegetación de páramo 65
Photo 33. Blue grassland (Lupino alopecuroides-Agrostietum araucanae) of the lower superpáramo onsandy ridges on the glacier valley bottom at 4335 m directly west ofthe pass between the Otún valley andQuebrada Africa, southeast ofNevado de Santa Isabel (Risaralda province). Very conspicuous are the reddish Lycopodium (Huperzia) crassum plants between the low tussocks. In the background, Polylepis sericeadwarfforest is present (details photos I and 2).
- Pastizal azul (Lupino alopecuroides-Agrostietum araucanae) del superpáramo bajo sobre depósitosarenosos en el fondo del valle glaciar a 4335 m, inmediatamente al W del Paso entre los valles de Otún yde la Quebrada Africa, al suroriente del Nevado de Sta. Isabel (Dpto. de Risaralda). Son muy notorias lasplantas rojizas de Lycopodium (Huperzia) crassum en el pajonal ralo y bajo. En el fondo del valle hay unbosque de Polylepis sericea , el cual se muestra en detalle en las fotos 1 y 2.
Diplostephio eriophori-Loricarietum columbianae Salamanca, C1eef & Rangel1992Loricaria columbiana shrub with Diplostephium eriophorumMatorral bajo de Loricaria columbiana con Diplostephium eriophorumType: relevé AMC 593. Table 7; Figs. 12,15; Photo 34-36
Physiognomy:This open scrub is composed of two layers;- shmb layer of greenish-brawn strong1y imbricate leaves, c. 50 cm high, covering between 15
and 90%,- ground layer, with a cover up to 30%.
Composition and syntaxonomy:Loricaria columbiana, Arcytophyllum cf. aristatum, Calamagrostis ejfusa, Diplostephium eriophorum, Jamesoniella rubricaulis, cf. Oreoweisia bogotensis, Pernettya prostrata, Polystichumpolyphyllum, pottiaceous mosses, Racomitrium crispulum, Riccardia spp., Tortula andicola andVacciniumfloribundum are diagnostic taxa of this association. This syntaxon shows strong floristic affinities with some ofthe syntaxa grauped under the subnival vegetation; especially withThamnolio vermicularis-Racomitrietum crispuli and with Racomitrio-Lachemilletum nivalis.This is indicated by the presence of Disterigma empetrifolium, Lachemilla nivalis, cf. Oreoweisia bogotensis, Polystichum polyphyllum. and Racomitrium crispulum. Among the species incommon between Loricarietum complanatae described by Cleef (1981) fram the Co10mbian

66 The paramo vegetation
Photo 34. Landscape of the lower superpáramo with outcrops of volcanic rock and 'arenales' (unstablesandy and ash deposits) at about 4350 m on the northwestem side ofthe Nevado del Ruiz (Caldas province).
- Paisaje de afloramientos de roca volcánica y arenales en el lado norteoccidental del Nevado del Ruiz(Dpto. de Caldas) a aproximadamente 4350 m con representación de diferentes comunidades vegetales delsuperpáramo bajo.
Photo 35. Low shrub of Loricaria columbiana associated with bunchgrasses. This is present in the shelterofvolcanic rock at 4350 m in the lower superpáramo ofthe northwestern slope ofNevado del Ruíz (Caldasprovince).
- Matorral bajo de Loricaria columbiana, asociado con gramíneas en un abrigo de roca volcánica del superpáramo bajo a 4350 m en el flanco norteoccidental del Nevado del Ruiz (Dpto. de Caldas).

La vegetación de páramo 67
Photo 36. Detail of a branch with imbricated xeromorphic leaves and inflorescences of Loricaria columbiana (Compositae)ofphoto 35.
- Detalle de lIna rama con hojas imbricadas xeromórficas e inflorescencias de Loricaria columbiana(Col11positae)de la foto 35.
Eastern Cordillera, and the present association fram the Central Cordillera, areCalamagrostisejjÚsa,lamesoniella rubricaulis, Pernettya prostrata, Racomitrium crispulum, Riccardia spp.and Vacciniumjloribundum. Duque & Rangel (1989) described Lachemillo pectinatae-Loricarietum complanatae fram Puracé Volcano (3800 m) with a rather different floristic composition.
A different type of Loricaria columbiana vegetation was observed in contact with the highbunchgrassparamo. This has relatively high cover of Calamagrostis eIJusa and Festuca sublimis.Nevertheless, in the absence of sufficient relevés, it is not described here.
Synecology and distribution:The association of Diplostephio eriophori-Loricarietum columbianae reported here, is found onthe escarpments of lava frants at the upper Iimit of the superparamo blue grasslands, between4300 and 4450 m and common in the Parque Los Nevados.
Relationship among the paramo syntaxa
Table 1 shows the result of a Twinspan ana]ysis (Rill 1979) on the synoptic table including allsyntaxa described here. The importance of altitude and humidity as determining factors for explaining the distribution ofthe paramo vegetation types in the study area is immediately apparent.In the first Twinspan analysis the higher cover c]asses (III, IV and V) were given three timesthe weight of the smaller cover c]asses (1, II, III). In the second Twinspan ana]ysis, no weightswere given to the pseudospecies levels. The weighted Twinspan analysis gave a more satisfactory result than the unweighted one, indicating the relative importance ofthe higher cover classes. This final Twinspan table was not reordered further.
There is a major division in the tab]e, which separates the superparamo vegetation fram theother vegetation types. Appraximately half of the total number of species are absent fram thesuperparamo vegetation. The two main superparamo formations were not c]early subdivided.Vegetation types belonging to the higher superparamo interdigitate with the ones be]onging to

68 The paramo vegetation
the lower superparamo. Species like Agrostis araucana, Altensteinia (Aa) paludosa, Andreaearupestris, Cerastiumfloccosum, Draba spp., Lupinus alopecuroides, Montia meridensis, Pentacalia gelida, Poa trachyphylla, Senecio canescens, Stereocaulon vesuvianum and Thamnoliavermicularis occur exclusively in the superparamo. On the other hand, Arenaria serpens, Baccharis caespitosa, Bromus lanatus, Elaphoglossum mathewsii, Erigeron chionophilus, Grammitis moniliformis, Hypochaeris sessiliflora and Luzula racemosa are present in the majority ofthe vegetation types (with the exception of the meadows), but clearly prefer the superparamoenvironment.
A group of species, consisting of Azorella multifida, Cortaderia sericantha, Festuca sublimis,Hypericum laricifolium, Rumex tolimensis, Senecio formosus and Uncinia macrolepis (U. meridensis) are found both in the forests and in the cushion bogs. Another group of species; Baccharistricuneata, Carex pichinchensis, Conyza uliginosa, Espeletia hartwegiana, Lachemilla galioides, Ranunculus peruvianus and Satureja nubigena are shared by vegetation types belongingto the cushion bogs, the bunchgrasslands and the meadows. Castilleja fissifolia, Lachemilla holosericea, 1. tanacetifolia, Pentacalia vernicosa and Ranunculus praemorsus a\though presentin few vegetation types of the superparamo, mainly belong to the dwarf forests, the bunchgrasslands and the cushion bogs. Apart from a11the trees, some other species like Bomarea linifolia,Calceolaria pentlandii, Geranium cucullatifolium, Hydrocotyle gunnerifolia, Lappula echinata,Oxalis spp., Peperomia hartwegiana, Rubus sp., Stellaria cuspidata and Urtica ballotaefolia areexclusive to the dwarfforests. Distichia muscoides, Floscaldasia hypsophila, Geranium confertum, Myrteola oxycoccoides, Oritrophium limnophilum and Ourisia muscosa are exclusive tothe cushion bogs. The meadows form almost a completely isolated group, showing only a weakrelation with the bunchgrasslands. Species like Acaena cylindristachya, Bromus unioloides,Elytrigia repens, Plantago linearis and Salvia sp. mainly belong to the meadows. On the otherhand, the bunchgrasslands show hardly any exclusive species.
Altitudinal zonation of the paramo vegetation
In order to depict the altitudinal zonation ofthe paramo vegetation in the Ruiz-Tolima volcanicmassif the longest altitudinal sequences available were chosen. These were located along theeast and west sides, respective1y of the Santa Isabel and the Santa Rosa volcanoes, lake OtÚnlake fa11ingin between these two volcanoes. These sequences comprised thirty- four relevés alongthe Parque Los Nevados transect (TPN relevé s), and a number of relevés made by the first authortogether with E.H. and C. Kloosterman (SS relevé s) and by the second author (AMe relevés).The structural configuration ofparamo vegetation at the a11iance level ofthe Parque Los Nevadosis represented in Fig. 16.
On the west slope of the Santa Rosa volcano, the most complete sequence of the zonal vegetation is present. The most extended paramo proper bunchgrasslands belong to the associationof Calandrinio acaulis-Calamagrostietum rectae. The spatial distribution of this type of vegetation is not symmetric since it occurs at somewhat higher altitudes, at the outer slopes of bothvolcanoes. The upper limit ofthe paramo proper bunchgrasslands, belonging to the associationCalamagrostietum effuso-rectae, is found at a higher altitude on the eastern slope of the SantaRosa volcano compared to the distribution on the western slope. On the west side of the SantaIsabel volcano these bunchgrasslands, together with those of the association Espeletio hartwegianae-Calamagrostietum effusae, are restricted to a sma11belt. Bunchgrasslands belonging tothe association Festuco dolichophyllae-Calamagrostietum ejJÚsae dominate this area.
The altitudinallimit between the higher and lower superparamo is diffuse. Above 4400 m thevegetation of the associations Senecioni canescentis-Cerastietum floccosi and Senecioni canescentis-Calamagrostietum ligulatae are present. Both associations belong to the subnival vegetation of the upper superparamo. Sometimes they occur together at the same altitude with or
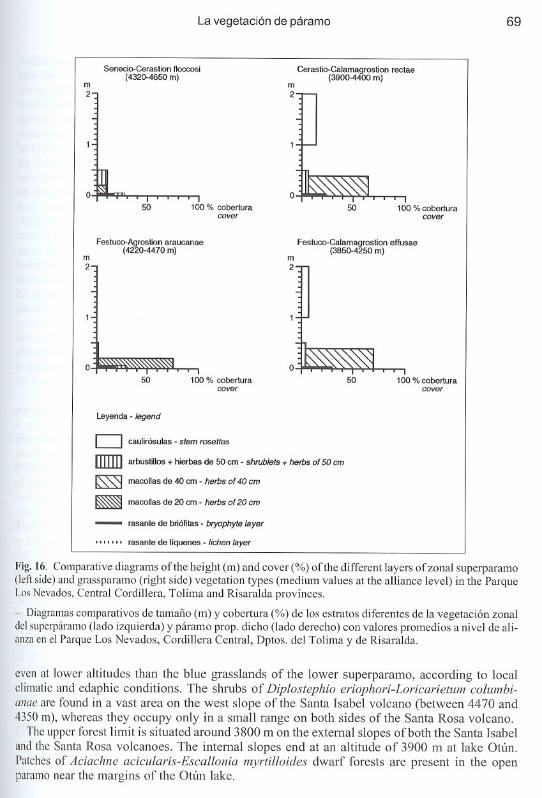
La vegetación de páramo 69
Senecio-Ceraslion f1occosi
(4320-4650 m)Cerastio-Calamagrostion rectae
(3900-4400 m)m
2
o
50 100 % coberturacaver
50 100 % coberturacaver
Festuco-Agroslion araucanae(4220-4470 m)
Festuco-Calamagrostion effusae(3850-4250 m)
100 % coberturacaver
50
o
m
2
100 % coberturacaver
50
o
m2
Leyenda - legend
D caulirósulas - stem roseNes
lIIIlll arbustillos + hierbas de 50 cm - shrublets + herbs 0150 cm
[SS3 macollas de 40 cm - herbs 0140 cm
~ macollas de 20 cm - herbs 0120 cm
rasante de briófitas - bryophyte layer
rasante de líquenes - lichen layer
Fig.16. Comparative diagrams ofthe height (m) and cover (%) ofthe different layers ofzonal superparamo(left side) and grassparamo (right side) vegetation types (medium values at the alliance level) in the ParqueLos Nevados, Central Cordillera, Tolima and Risaralda provinces.
- Diagramas comparativos de tamaño (m) y cobertura (%) de los estratos diferentes de la vegetación zonaldel superpáramo (lado izquierda) y páramo prop. dicho (lado derecho) con valores promedios a nivel de alianza en el Parque Los Nevados, Cordillera Central, Dptos. del Tolima y de Risaralda.
even at lower altitudes than the blue grasslands of the lower superparamo, according to localclimatic and edaphic conditions. The shrubs of Diplostephio eriophori-Loricarietum columbianae are found in a vast area on the west slope of the Santa Isabel volcano (between 4470 and4350 m), whereas they occupy only in a small range on both sides of the Santa Rosa volcano.
The upper forest limit is situated around 3800 m on the external slopes ofboth the Santa Isabeland the Santa Rosa volcanoes. The internal slopes end at an altitude of 3900 m at lake Otún.Patches of Aciachne acicularis-Escallonia myrtilloides dwarf forests are present in the openparamo near the margins of the Otún lake.

70 The paramo vegetation
From the azonal vegetation types, the association of Gentianello dasyanthae-Plantaginetumrigidae is by far the most extended type of cushion bogo It is encountered between 3700 and4200 m on the eastern slope of the Santa Isabel Volcano, and from 4100 to 4200 m on the easternslope of the Santa Rosa volcano. The cushion bogs belonging to the association Floscaldasiohypsophilae-Distichietum muscoides, have a more restricted distribution and are found mainlyaround 4150 m on the western slope of the Santa Rosa volcano and around 4350 m on the easternslope of the Santa Isabel volcano, near the transition with the superparamo. Small patches ofcushion bogs belonging to the association Wernerietum humilis are found on the western slopeof the Santa Isabel volcano at an altitude of around 4200 m. Those of the association Carici
peucophilae-Wernerietum crassae were recorded at about 3800 m on the eastern slope of theSanta Isabel volcano (AMC 625, 637).
Dwarfforests ofthe association Gynoxyo-Polylepidetum sericeae are recorded in relevé AMC623 (4300 m) on the western slope of the Santa Isabel volcano. However, they are also presentelsewhere. Between 3830 and 3850 m scrubs belonging to the association Baccharido tricuneatae-Hypericetum laricifolii are recorded in relevés AMC 621 (western slope ofthe Santa Isabelvolcano), AMC 635 and AMC 636 on the eastern slope of the Santa Isabel volcano (3830 to3900 m). The greyish scrubs ofthe association Pentacalietum vernicosi are found in relevés SS110 and SS 111 (both at 4340 m) on the western slope ofthe Santa Isabel volcano and in relevésSS 123 and SS 124 (4350 m) on the eastern slope of the Santa Isabel volcano.
Azonal meadows belonging to the association Muhlenbergietum cleefii are recorded in onlyone relevé at the western margin of lake Otún, around 4000 m in relevé AMC 60 l. Pricklycushiongrass of Aciachnetum acicularis subassociation lupinetosum microphylli on the easternslope ofthe Santa Rosa volcano is recorded in relevés SS 44 (4050 m) SS 51 (4030 m) and SS52 (4070 m).
The highest vegetation types on the western slope of the Santa Isabel volcano belong to twodifferent associations of the subnival vegetation types of the upper superparamo. The first isrecorded in TPN 2 (4540 m) and belongs to the association Senecioni canescentis-Cerastietumjloccosi. It is replaced at lower altitudes by the association Senecioni latiflori-Calamagrostietumligulatae, recorded in re1evés SS 106, SS 107 and SS 118 (al! three at 4400 m). Around 4450m, in TPN 3, the low scrubs of Diplostephio eriophori-Loricarietum columbianae, be10nging tothe blue grasslands of the lower superparamo, is found. Between 4400 and 4350 m anotherassociation belonging to the lower superparamo blue grasslands; Lupino alopecuroides-Agrostietum araucanae, is recorded in relevés SS 108, SS 109, SS 112, SS 117, SS 119, SS 120, SS121 and SS 125 between 4350 and 4470 m. The transition to the paramo proper is located ataround 4400 m. The first relevé with a paramo proper vegetation is TPN 4 at 4350 m; thiscomprises a bunchgrassland belonging to the association Calandrinio acaulis-Calamagrostietumrectae var. of Espeletia hartwegiana. The same vegetation type is present in TPN 5 (at 4250 m)and TPN 6 (at 4150 m). Bunchgrassland of Calandrinio acaulis-Calamagrostietum rectae varotypicum is recorded at 4200 m in relevé SS 80. Bunchgrasslands belonging to the association ofFestuco dolichophyllae-Calamagrostietum effusae are found in relevé SS 85 at 4140 m.
On the eastern side of the Santa Isabel volcano, the highest vegetation type belongs to theassociation Lupino alopecuroides-Agrostietum araucanae (recorded in TPN 28 at an altitude of4475 m). The same vegetation type is present in TPN 29 (4400 m). The transition to the bunchgrassparamo is situated at 4360 m. The bunchgrassland of Calandrinio acaulis-Calamagrostietum rectae var. of Espeletia hartwegiana is recorded in three relevés: TPN 30 (at 4315 m), TPN32 and 33 (both at 4125 m). In between these three relevés, TPN 31 (at 4200 m) records abunchgrassland belonging to the association Calamagrostietum effuso-rectae. TPN 34 (at 4015m) represents the association Espeletio hartwegianae-Calamagrostietum effusae. In TPN 26 (at3900 m) bunchgrassland of Calamagrostietum effuso-rectae is again present. The forest limiton the eastern slope ofthe Santa Isabel volcano is situated around 3800 m. In TPN 27 (at 3750m) a high Andean forest was recorded.

La vegetación de páramo 71
The highest relevé on the westem side of the Santa Rosa volcano is TPN 8 at 4550 m; itbelongs to the association Lupino alopecuroides-Agrostietum araucanae. From 4460 m, in TPNII to 4390 m, in SS 38, blue grasslands belonging to the association Diplostephio eriophoriLoricarietum columbianae are present. The transition to the paramo proper is located around4400 m and the first association of the paramo proper bunchgrasslands is recorded in TPN 9 at4330 m. It belongs to the bunchgrassland of Calandrinio acaulis-Calamagrostietum rectae var.of Espeletia hartwegiana. The same vegetation type is present in TPN 12 at 4250 m. In TPN10 (at 4175 m) and in TPN 15 (4050 m) the association Calamagrostietum effuso-rectae isrecorded. The bunchgrassland of Festuco dolichophyllae-Calamagrostietum eJJÚsae is presentin TPN 14 (3960 m) and TPN 16 (3850 m). The forest limit is located around 3800 m; the firsthigh Andean forest is recorded in TPN 17 (at 3755 m).
On the eastem slope of the Santa Rosa volcano the highest occurring vegetation type is recorded in relevé SS 40 at 4400 m. It belongs to the association of Senecioni canescentis-Cerastietwn floccosi, in the higher superparamo. In TPN 22 (4425 m) the association Lupino alopecuroides-Agrostietum araucanae of the lower superparamo is found. Another relevé (AMC605 at 4460 m) also has a vegetation type belonging to that association, while in the relevés SS38 (4390 m) and AMC 606 (4400 m) low scrub of Diplostephio eriophori-Loricarietum columbianae is present. The transition to the paramo proper is situated around 4400 m. The first bunchgrasslands are recorded in TPN 23 (4400 m), TPN 24 (4300 m), TPN 25 (4200 m), SS 41 (4400m) and SS 42 (4400 m). All of these records belong to Calandrinio acaulis-Calamagrostietumrectae val'.Espeletia hartwegiana. A dwarfforest of Aciachne acicularis-Escallonia myrtilloidesis present in TPN I (3975 m) and TPN 7 (3930 m) at the west margin of lake Otún.
Acknowledgements
Theauthorsacknowledgethe support ofDrs. Roberto Jaramillo Mejía and Santiago Díaz Píedrahita for collectionandidentificationof paramo plant species. We are grateful to profesora Eugenia Brieva for the finelinedrawingsbased on the field sketches of the second author and to Guido van Reenen for the final arrangementof these drawings.
References
Acosta Solís, M. 1984. Los páramos andinos del Ecuador. Publicaciones Científicas M.AS.,Editorial ENA, Quito, 222 pp.
Almeida, 1. A.M. Cleef, A Herrera, A. Velázquez & 1. Luna 1994. El zacatonal alpino delVolcán Popocatépetl, Mexico, y su posición en las montañas tropicales de America. Phytocoenologia 22(3): 391-436.
Ama!, D.H. 1983. Estudio ecológico del bosque alti-andino de Polylepis sericea Webb en laCordillera de Mérida. Thesis, Universidad Central de Venezuela, Caracas.
Azócar, A. & M. Monasterio 1979. Variabilidad ambiental en el páramo de Mucubají. In: M.L.Salgado-Labouriau (ed.), El Medio Ambiente Páramo, pp. 149-159. Instituto Venezolano deInvestigaciones Científicas, Caracas.
Barkman, 1.1., 1. Moravec & S. Rauschert 1986. Code of Phytosociological nomenclature, 2ndedition. Vegetatio 76: 145-195.
Beck, S. & H. Ellenberg 1977. Possibilities of Development of the Andean Highlands from theEcological Point of View. Studie im Auftrag des Bundesministeriums fUr wirtschaftlicheZusammenarbeit, Géittingen, 147 pp.

72 The paramo vegetation
Becking, M.L., A.l Negret & R.G. Hofstede 1997. Caracterización preliminar de un 'Paramillo'en la cordillera occidental del Departamento del Cauca. Novedades Colombianes (Nueva Epoca) 7: 33-44.
Beekman, A.M. & P.A. Verweij 1987. Structure and nutrient status of a paramo bunchgrassvegetation in relation to soil and climate. Internal report No. 233. Hugo de Vries Laboratory,University of Amsterdam.
Benoist, R. 1935. Le Plantago rigida H.B.K., sa structure, sa biologie. Bull. Soco Bot. France82: 462-466.
Berg, A.L. 1998. Pflanzengesellschaften und Lebensformen des Superpáramo des Parque Nacional Sierra Nevada de Mérida in Venezuela. Phytocoenologia 28(2): 157-203.
Bosman, A.F., P.e. Van der Molen, R. Y oung, & A.M. Cleef 1993. Ecology of a paramo cushionmire. Journal of Vegetation Science 4: 633-640.
Bruynzeel L.A. & E.l Veneklaas 1998. Climatic conditions and tropical montane forest productivity: the fog has not lifted yet. Ecology 79(1): 3-9.
Calderón Sánchez, E. 1993. A biogeographical approach to the vascular plant flora from thehigh mountain ofthe Farallones de Cali (W Cordillera, Colombia) In: H. Balslev (ed.), Neotropical montane forests: biodiversity and conservation. Abstracts from a symposium at theNew York Botanical Garden, June 21-26 1993. AAU Reports 31: 25
Chaverri Polini, A. & A.M. Cleef 1996. Las comunidades vegetacionales en los páramos de losmacizos del Chirripó y Buenavista, Cordillera de Talamanca, Costa Rica. Revista ForestalCentroamericana 17: 44-49.
Clark, L.G. 1989. Systematics of Chusquea sect. Swallenochloa, sect. Verticillatae, sect. Serpentes and sect. Longifoliae (Poaceae, Bambusoideae). Syst. Bot. Monogr. 27: 127 pp.
Cleef, A.M. 1978. Characteristics of neotropical paramo vegetation and its subantarctic relations.In: e. Troll & W. Lauer (eds), Geoecological relations between the southern temperate zoneand the tropical mountains. Erdwiss. Forsch. 11: 365-390. Wiesbaden.
Cleef, A.M. 1979a. Secuencia altitudinal de la vegetación de los páramos de la Cordillera Oriental, Colombia. Actas 4° Simp. Int. Ecol. Trop. 1: 282-297. Panamá.
Cleef, A.M. 1979b. The phytogeographical position of the neotropical vascular páramo florawith special reference to the Colombian Cordillera Oriental. In: K. Larsen & L.B. HolmNielsen (eds), Tropical Botany, pp. 175-184. Academic Press, London.
Cleef, A.M. 1980a. Vegetación del páramo neotropical y sus lazos australo-antárticos. ColombiaGeográfica 7(2): 7-49.
Cleef, A.M. 1980b. Secuencia altitudinal de la vegetación de los páramos de la cordillera Oriental de Colombia. Colombia Geográfica 7(2): 50-67.
Cleef, A.M. 1980c. La posición fitogeográfica de la flora vascular del páramo neotropical. Colombia Geográfica 7(2): 68-86.
Cleef, A.M. 1981. The vegetation of the páramos of the Colombian Cordillera Oriental. Dissertationes Botanicae 61, 320 pp. Cramer, Vaduz. [Also: The Quaternary of Colombia 9. Amsterdam.)
Cleef, A.M. & lO. Rangel Ch. 1984. La vegetación del páramo del Noroeste de la Sierra Nevadade Santa Marta. In: T. van der Hammen & P.M. Ruiz (eds), La Sierra Nevada de Santa Marta(Colombia), Transecto Buritaca-La Cumbre. Studies on Tropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 2: 203-266. Cramer (Borntraeger), Berlin-Stuttgart.
Cleef, A.M., lO. Rangel Ch., & S. Salamanca 1983. Reconocimiento de la vegetación de laparte alta del transecto Parque Los Nevados. In: T. van der Hammen, A. Pérez P. & P. PintoE. (eds), La Cordillera Central Colombiana, Transecto Parque Los Nevados (Introducción ydatos iniciales). Studies on Tropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 1: 150-174. Cramer, Vaduz.
Cleef, A.M., J.O. Rangel Ch. & S. Salamanca 1995. Los bosques andinos del transecto ParqueLos Nevados. In: T. van der Hammen & A.G. dos Santos (eds). Studies on Tropical Andean

La vegetación de páramo 73
Ecosystems/Estudios de Ecosistemas Tropandinos 5 (this volume/este volumen). Cramer(Borntraeger), Berlin-Stuttgart.
Cleef, A.M., lO. Rangel Ch., S. Salamanca, C. Ariza N. & G.B.A. van Reenen (in prep.). Lavegetación del páramo del Macizo del Tatamá, Cordillera Occidental, Colombia. Studies onTropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 6.
Cuatrecasas, 1 1934. Observaciones geobotánicas en Colombia. Trab. Mus. Nac. Cienc. Nat.,Ser. Bot. 27: 1-144. Paris.
Cuatrecasas, 1 1954. Outline of vegetation types in Colombia. Rapports Comm. 8eme Congr.lnt. Bot. VII: 77-78.
Cuatrecasas, 1 1958. Aspectos de la vegetación natural. Rev. Ac. Col. Cienc. Ex. Fís. Nat.10(40): 221-264.
Cuatrecasas, 1 1968. Páramo vegetation and its life forms. Coll. Geogr. 9: 163-186.Cuatrecasas, 1 1979. Comparación fitogeográfica de páramos entre varias cordilleras. In: M.L.
Salgado-Labouriau (ed.), El medio ambiente páramo, pp. 89-99. Ediciones Centro de EstudiosAvanzados, Caracas.
Cuatrecasas, 1 1980. Miscellaneous notes on neotropical flora XI. Phytologia 45(1): 17-29.Cuatrecasas, 1 2003. Observaciones geobotánicas en el Nevado del Tolima, cordillera Central,
en 1932. In: T. van del' Hammen & A.G. dos Santos (eds). Studies on Tropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 5 (this volume/este volumen). Cramer (Borntraeger), Ber!in-Stuttgart.
Die!s,1. 1937. Beitrage zur Kenntnis del' Vegetation und Flora von Ecuador. Bibl. Bot. 116: 55pp. Stuttgart.
DuqueN., A. & lO. Rangel Ch. 1989. Análisis fitosociológico de la vegetación paramuna delParque natural del Puracé. In: L.F. Herrera, R.D. Drennan & c. Uribe (eds): Cacicazgos prehispánicos en el Valle de la Plata. 1. El contexto medio-ambiental de la ocupación humana.University of Pittsburgh Memoirs in Latin American Archaeology 2: 70-95.
Ellenberg, H. 1958. Wald oder Steppe? Die naturliche Pflanzendecke del' Anden Perus, 1. DieUmschau in Wissenschaft und Technik 21: 645-648.
Ellenberg, H. 1958. Wald oder Steppe? Die naturliche Pflanzendecke del' Anden Perus, 11.DieUmschau in Wissenschaft und Technik 22: 670-681.
Ellenberg, H. 1959. Typen Tropischer Urwalder in Perú. Schweiz. Zeitschr. fUI' Forstwesen 110:169-187.
ElIenberg,H. 1979. Man' s influence on tropical mountain ecosystems in South America. 1 Ecol.67: 401-416.
Ferwerda, W. 1987. The influence ofpotato cultivation on the natural bunch grass páramo inthe Colombian Cordillera Oriental. Internal report No. 220. Hugo de Vries Laboratory, University of Amsterdam.
Fosberg, F.R. 1959. Upper limits ofvegetation on Mauna Loa, Hawaii. Eco!, 40: 144-146.Franco,R.P., lO. Rangel Ch. & G. Lozano-c., 1986. Estudios ecológicos de la cordillera Ori
entallI. Las comunidades vegetales de los alrededores de la laguna de Chingaza (Cundinamarca). Ca!dasia 15(71-75): 219-248.
Godley, E.G. 1978. Cushion bogs. In: C. Troll & W. Lauer (eds), Geoecological relations between the southern temperate zone and the tropical mountains. Erdw. Forsch. 11: 141-148.
Grllbb, P.J. 1977. Control of forest growth and distribution on wet tropical mountains: withspecial reference to mineral nutrition. Annual Review of Ecology and Systematics 8: 83-107.
Gutte, P. 1980. Beitrag zur Kenntnis zentralperuanischer Pflanzengesellschaften 11. Diehochandinen Moore und ihre Kontaktgesellschaften. Feddes Repertorium 91(5-6): 327-336.
Glltte,P. 1985. Beitrag zur Kenntnis zentralperuanischer Pflanzengesellschaften IV. Die grasreiche Vegetation del' alpinen Stufe. Wiss. Z. Karl-Marx-Univ. Leipzig, Math.-Naturwiss. R.34(4): 357-401.

74 The paramo vegetation
Harling, G. 1979. The vegetation types ofEcuador - a briefsurvey. In: K. Larsen & L.B. HolmNie1sen (eds), Tropical Botany, pp. 165-174. Academic Press, London.
Hill, M.O. 1979. TWINSPAN - A FORTRAN program for arranging multivariate data in anordered two-way table by classification of individual s and attributes. Cornell UniversityIthaca, New York, 90 pp.
Hofstede, R.G.M. 1995. Effects of burning and grazing on a Colombian páramo ecosystem.Thesis, University of Amsterdam. 199 pp.
Holdridge, L.R., 1947. Determination ofworld plant formations from simple climatic data. Science 105: 367-368.
J0rgensen, P.M. & e. Ulloa Ulloa, 1994. Seed plants ofthe High Andes ofEcuador - a checklist.AAU Reports 34, 443 pp. Department of Systematic Botany, University of Aarhus.
Koepcke, H.W., 1961. Syn6kologische Studien an der Westseite der peruanischen Anden. Bonner Geogr. Abhand. 29: 1-318.
Kok, K., P.A. Verweij & H. Beukema (1995). Effects of cutting and grazing on Andean tree1ine vegetation. In: S.P. Churchill et al. (eds), Biodiversity and conservation of neotropicalmontane forest, pp. 527-539. The New York Botanica1 Garden. New York.
Kuhry, P. 1988. A pa1eobotanical and pa1yno10gica1 study of Holocene peat from El Bosquemire, 10cated in a vo1canic area of the Cordillera Central of Colombia. Rev. Palaeobot. Palyno!. 55: 19-72.
Kuhry, P., 1.B. Salomons, P.A. Riezebos & T. van der Hammen 1983. Paleoecología de losúltimos 6000 años en el área de la Laguna de Otún - El Bosque. In: T. van der Hammen, A.Pérez P. & P. Pinto E. (eds), La Cordillera Central Colombiana, Transecto Parque Los Nevados (Introducción y datos iniciales). Studies on Tropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 1, pp. 227-261. Cramer, Vaduz.
Laegaard, S. 1987. The genus Aciachne (Poaceae). Nordic Journa1 of Botany 7: 667-672.Laegaard, S. 1992. Influence of fire in the grass paramo vegetation of Ecuador. In: H. Balslev
& 1.L. Luteyn (eds), Paramo - An Andean ecosystem under human influence, pp. 151-170.Laegaard, S. 1995. Muhlenbergia cleefii sp.nov., a new grass from the high Andes ofColombia.
Caldasia 17(82-85), 409-412. (See also Nota breve by S. Laegaard (1996) in Caldasia 18 (3), 355,correcting spelling of the epithet).
Lutz, R.T. & P.N.e. Vader 1987. Biomass, productivity and nutrient status in a Co10mbianbunchgrass páramo. Internal report No. 229. Hugo de Vries Laboratory, University of Amsterdam.
Luteyn, J.L. 1999. Páramos. A checklist of p1ant diversity, geographical distribution and botanical literature. Memoirs of the New York Botanica1 Garden, 84: 278 pp.
Marrs, R.H., 1.Proctor, A.Henney & M.D. Mountford 1988. Changes in soi1 nitrogen-mineralization and nitrification along an altitudina1 transect in tropical rain forest in Costa Rica. Journa1 of Eco10gy 76: 466-482.
Melief, A.B.M. 1985. Late Quaternary pa1eoecology ofthe Parque Nacional Natural los Nevados(Cordillera Central), and Sumapaz (Cordillera Oriental) areas, Colombia. Thesis, Universityof Amsterdam, 162 pp. [A1so: The Quaternary of Colombia 12. Amsterdam.]
Monasterio, M. 1980. Las formaciones vegetales de los páramos de Venezuela. In: M. Monasterio (ed.), Estudios eco1ógicos en los páramos andinos, pp. 94-158. Ediciones de la Universidad de los Andes, Mérida.
Miiller Dombois, D. & H. Ellenberg 1974. Aims and methods ofvegetation ecology. John Wileyand Sons, New York, 547 pp.
Pfitsch, W.A. 1988. Microenvironment and the distribution of two species of Draba (Brassicaceae) in a Venezue1an paramo. Arctic and Alpine Research 20(3): 333-341.
Pipo1y III, 1.1. 1992. Estudios en el género Myrsine (Myrsinaceae) en Colombia. Caldasia 17(1):3-10.

La vegetación de páramo 75
Ramsay,P.M. 1992. The paramo vegetation ofEcuador: the community ecology, dynamics andproductivity oftropical grasslands in the Andes. Thesis, University ofWales, Bangor, 274 pp.
Ramsay, P.M. & E.R.E. Oxley 1996. Fire temperatures and postfire plant community dynamicsin Ecuadorian grass páramo. Vegetatio 124: 129~144.
Rangel Ch., lO. & P. Franco R. 1985. Observaciones fitoecológicas en varias regiones de vidade la Cordillera Central de Colombia. Caldasia 14(67): 211~249.
RangelCh., lO. & G. Lozano C. 1986. Un perfil de vegetación entre La Plata (Huila) y el volcándel Puracé. Caldasia 14(68-70): 503~547.
RangelCh., lO. 1994. La vegetación del Parque Regional Natural Ucumarí. In: lO. Rangel Ch.(ed.), Ucumarí, un caso de la diversidad biótica andina, pp. 59-81. CARDER-UniversidadNacional de Colombia, Pereira.
Rangel Ch., lO., A.M. Cleef & S. Salamanca 2003. Los bosques ecuatoriales y subandinos deltransecto Parque Los Nevados. In: T. van der Hammen & A.G. dos Santos (eds). Studies onTropical Andean Ecosystems/Estudios de Ecosistemas Tropandinos 5 (this volume/este volumen). Cramer (Borntraeger), Berlin-Stuttgart.
Restrepo, C. & A. Duque 1992. Tipos de vegetación del Llano de Paletará. Cordillera Central,Colombia. Caldasia 17(1): 21-34.
Rivas Martínez, S. & O. Tovar 1982. Vegetatio Andinae, I. Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Perú. Lazaroa 4: 167-187.
Robson, N.K.B. 1987. Studies in the genus Hypericum 1. (Gutttiferae) 7. Section 29. Brathys(part 1). Bulletin ofthe British Museum (Natural History) Botany series (Jan. 29) 16(1), 106pp.
Ruthsatz, B. 1977. Pflanzengesellschaften und ihre Lebensbedingungen in den andinen Halbwiisten Nordwest-Argentiniens. Dissertationes Botanicae 39, 168 pp. 1Cramer, Vaduz.
Ruthsatz, B. 1995. Vegetation und Oekologie tropischer Hochgebirgsmoore in den Anden NordChiles. Phytocoenologia 25(2): 185-234.
Salamanca Villegas, S. 1991. The vegetation of the paramo and its dynamics in the volcanicmassifRuiz-Tolima (Cordillera Central, Colombia). Thesis, University of Amsterdam, 122 p.(photocopied manuscript).
Salamanca Villegas, S. 1992. La vegetación del páramo y su dinámica en el macizo volcánicoRuíz-Tolima (Cordillera Central, Colombia). Análisis Geográficos 21: 155 pp. IGAC, Santaféde Bogotá.
Salamanca, S., A.M. Cleef & lO. Range11992. La vegetación de páramo. In: La vegetación delpáramo y su dinámica en el Macizo volcánico Ruíz-Tolima (Cordillera Central, Colombia),S. Salamanca, ed. Análisis Geográficos 21: 38~66. IGAC, Bogotá.
Salomons,lB. 1986. Paleoecology of volcanic soils in the Colombian Central Cordillera (ParqueNacional Natural de los Nevados). Dissertationes Botanicae 95: 212 pp. Berlin-Stuttgart, J.Cramer. [Also: T. van der Hammen, S. Diaz-P. & VJ. Alvarez (eds), 1989. La CordilleraCentral Colombiana, Transecto Parque Los Nevados (Segunda parte). Studies on TropicalAndean Ecosystems/Estudios de Ecosistemas Tropandinos 3, pp. 15~215. Berlin-Stuttgartand The Quaternary of Colombia 13. Amsterdam.]
Sánchez,R. & lO. Rangel Ch. 1990. Estudios ecológicos en la Cordillera Oriental ColombianaV. Análisis fitosociológico de la vegetación de los depósitos turbosos paramunos de los alrededores de Bogotá. Caldasia 16(77): 155-169.
Scott, G.AJ. 1974. Grassland creation in a montane tropical rainforest and its effects on soilvegetation nutrient pools and nutrient cycles: A case study in the Gran Pajonal of EastemPem. Thesis, University of Hawaii.
Seibert, P. & X. Menhofer 1991. Die Vegetation des Wohngebietes der Kallawaya und desHochlandes von Ulla-Ulla in den bolivianischen Anden. Phytocoenologia 20(2): 145~276.
Seibert, P. & X. Menhofer 1992. Die Vegetation des Wohngebietes der Kallawaya und desHochlandes von Ulla-Ulla in den bolivianischen Anden. Phytocoenologia 20(3): 289--438.

76 The paramo vegetation
Simpson, B.B. 1979. A revision ofthe genus Polylepis (Rosaceae: Sanguisorbae). SmithsonianContr. Bot. 43: 1-62.
Sturm, H. & J.O. Rangel Ch. 1985. Eco10gía de los páramos andinos: Una visión preliminarintegrada. In: P.M. Ruiz & S. Díaz-Piedrahita, Biblioteca José Jerónimo Triana 9, 292 pp.Editora Guadalupe Ltda, Bogotá.
Thouret, J.c. 1983. Observaciones geopedológicas a lo largo del Transecto del Parque Los Nevados. In: T. van der Hammen, A. Pérez P. & P. Pinto E. (eds.), La Cordillera Central Colombiana, Transecto Parque Los Nevados (Introducción y datos iniciales). Studies on TropicalAndean Ecosystems/Estudios de Ecosistemas Tropandinos 1, pp. 113-141. Cramer, Vaduz.
Tosi Jr., J.A. 1960. Zonas de vida natural en e] Perú. Memoria explicativa sobre el mapa ecológico del Perú. IICA, Zona Andina. Boletín Técnico 5: 1-127.
Troll, C. 1968. The Cordilleras ofthe Tropical Americas. Aspects of climatic, phytogeographicaland agrarian ecology. In: C. Troll (ed.), Geo-ecology ofthe montainous regions ofthe TropicalAmericas, pp. 15-56.
Unzueta, O. 1975. Mapa Eco]ógico de Bolivia. Ministerio de Asuntos Campesinos y Agropecuarios, División de Suelos, Riegos e Ingeniería, La Paz, 312 pp.
Valencia, R. & P.M. Jorgensen 1992. Composition and stucture of a humid montane forest onthe Pasochoa volcano, Ecuador. Nordic Journal of Botany 12(2): 239-243.
Van der Hammen, T. 1974. The Pleistocene changes ofvegetation and climate in tropical SouthAmerica. J. Biogeogr. 1: 3-26.
Van der Hammen, T. 1979. History of the flora, vegetation and climate in the Colombian Cordillera Orienta] during the last five million years. In: K. Larsen & L.B. Holm-Nielsen (eds),Tropical Botany, pp. 25-34. Academic Press, London.
Van der Hammen, T. & E. González 1960. Upper Pleistocene and Holocene climate and vegetation ofthe "Sabana de Bogotá" (Colombia, South America). Leidse Geo!. Meded. 25: 125315.
Van derHammen, T. & Gonzá]ez, E. 1965. A pollen diagram from "Laguna La Herrera" (Sabanade Bogotá). Leidse Geol. Meded. 32: 183-191.
Van der Hammen, T., 1.H. Werner & H. van Dommelen 1973. Pa]ynologica] record ofthe upheaval ofthe Northem Andes: a study ofthe Pliocene and Lower Quaternary ofthe ColombianEastem Cordillera and the early evolution ofits High-Andean biota. Rev. Palaeobot. Palyno!.16: 1-122. [Also: The Quatemary ofCo]ombia 2. Amsterdam.]
Van der Hammen, T. & A.M. Cleef 1986. Development ofthe high Andean páramo flora andvegetation. In: F. Vuilleumier & M. Monasterio (eds), High altitude tropical biogeography,pp. 153-201. Oxford University Press.
Van Groen, F. 1987. Effects of fire on structure, composition and nutrient status in two bunchgrass páramos near Bogotá (Co]ombia). Internal report No. 236, 60 pp. Hugo de Vries Laboratory, University of Amsterdam.
Vareschi, V. 1953. Sobre las superficies de asimilación de sociedades vegetales de cordillerastropicales y extratropicales. Bo]etín de ]a Sociedad Venezolana de Ciencias Natura]es 14:121-173.
Vareschi, V. 1970. Flora de los páramos de Venezuela. Univ. de los Andes, Ediciones del Rectorado, Mérida, 425 pp.
Vareschi, V. 1980. Vegetationsokologie der Tropen. Ulmer, Stuttgart, 287 pp.Vargas R., O. & D. Rivera O. 1991. Comunidades vegetales del Parque Nacional Natural Chin
gaza: Sector 1 Río La Playa-Río Guatiquía. Cuadernos Divu]g. Universidad Javeriana 23: 174.
Velez, v., 1. Cave]ier & B. Devia 1998. Ecologica] traits ofthe tropical treeline species Polylepisquadrijaga (Rosaceae) in the Andes of Colombia. Joumal of Tropical Ecology 15: 771-787.

La vegetación de páramo 77
Verweij, P.A. 1995. Spatial and temporal modelling ofvegetation patterns. Burning and grazingin the paramo of Los Nevados National Park, Colombia. Thesis, University of Amsterdam,233 pp. (Also: !TC publication 30).
Verweij, P.A. & H. Beukema 1992. Aspects of human influence on upper Andean forest linevegetation. In: H. Balslev and J.L. Luteyn, (Eds), Paramo - An Andean ecosystem underhuman influence, pp. 171-175. Academic Press, London.
Verweij, P.A. & P.E. Budde 1992. Burning and grazing gradients in paramo vegetation. Initialordination analyses. In: H. Balslev and J.L. Luteyn, (Eds), Paramo - An Andean ecosystemunder human influence, pp. 177-195. Academic Press, London.
Verweij, P.A. & K. Kok 1992. Effects offire and grazing on Espeletia hartwegiana populations.In: H. Balslev and J.L. Luteyn, (Editors), Paramo - An Andean ecosystem under hurnan influence, pp. 215-229. Academic Press, London.
Villagrán M., C. 1980. Vegetationsgeschichtliche und pflanzensoziologische Untersuchungenim Vicente Pérez Rosales Nationalpark (Chile). Dissertationes Botanicae 54: 165 pp. 1. Cramer, Vaduz.
Walter, H. 1979. Vegetation ofthe earth and ecological systems ofthe Geo-biosphere. SpringerVerlag, New York, Heidelberg, Berlin, 274 pp.
Walter, H. & E. Medina 1969. La temperatura del suelo como factor determinante para la caracterización de los pisos subalpino y alpino en los Andes de Venezuela. Boletín Venezolanade Ciencias Naturales 115/116: 201-210.
Weberbauer, A. 1945. El mundo vegetal de los Andes Peruanos. Ministerio de Agricultura, Lima,776 pp.
Westhoff, V. & E. van del' Maarel1973. The Braun-Blanquet approach. In: R.H. Whittaker (ed.),Handbook of vegetation science part 5: Ordination and classification of communities, pp.617-726. Junk, The Hague.
Post Scriptum
Ihis manuscript has been finished mid 200 l. The third edition of the International Code ofPhytosociological Nomenclature (J. Veg. Sc. 11: 739-768.2000) has not been applied. In volume VII we wil! publish an overview of al! used syntaxonomical names and their formal corrections following the third edition of the Codeo
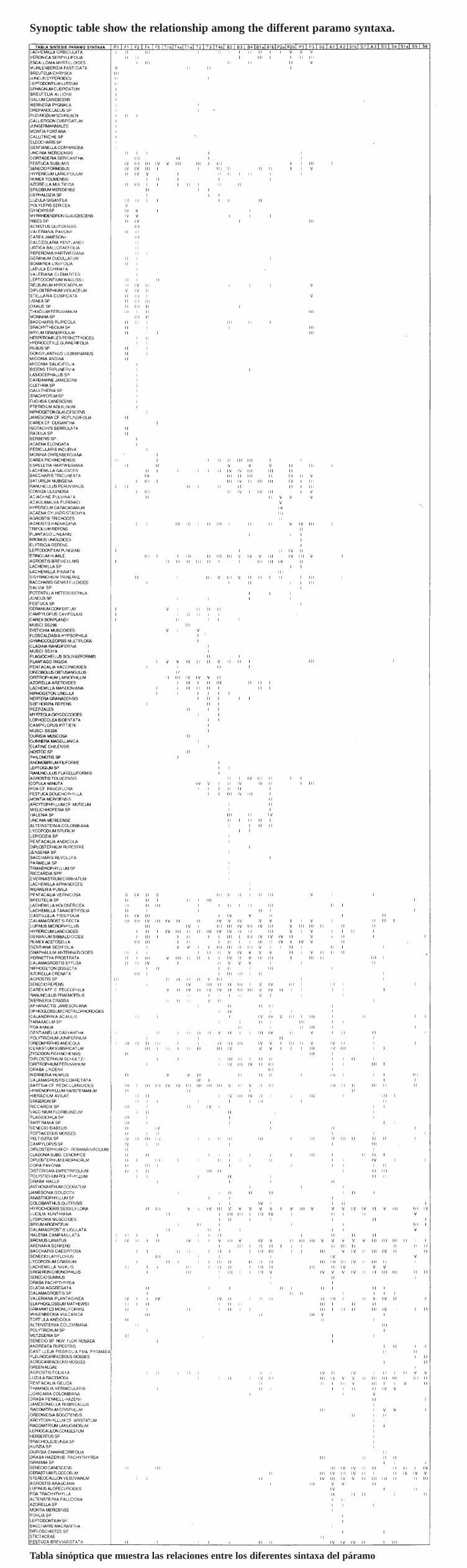
Synoptic table show the relationship among the different paramo syntaxa.
Tabla sinóptica que muestra las relaciones entre los diferentes sintaxa del páramo

Paramo dwarf forests and shrubs. Constancy table indicating the floristic compositionand the structure.
Bosques enanon de páramo y matorrales. Tabla de constancia indicando la composición florística yla estructura.
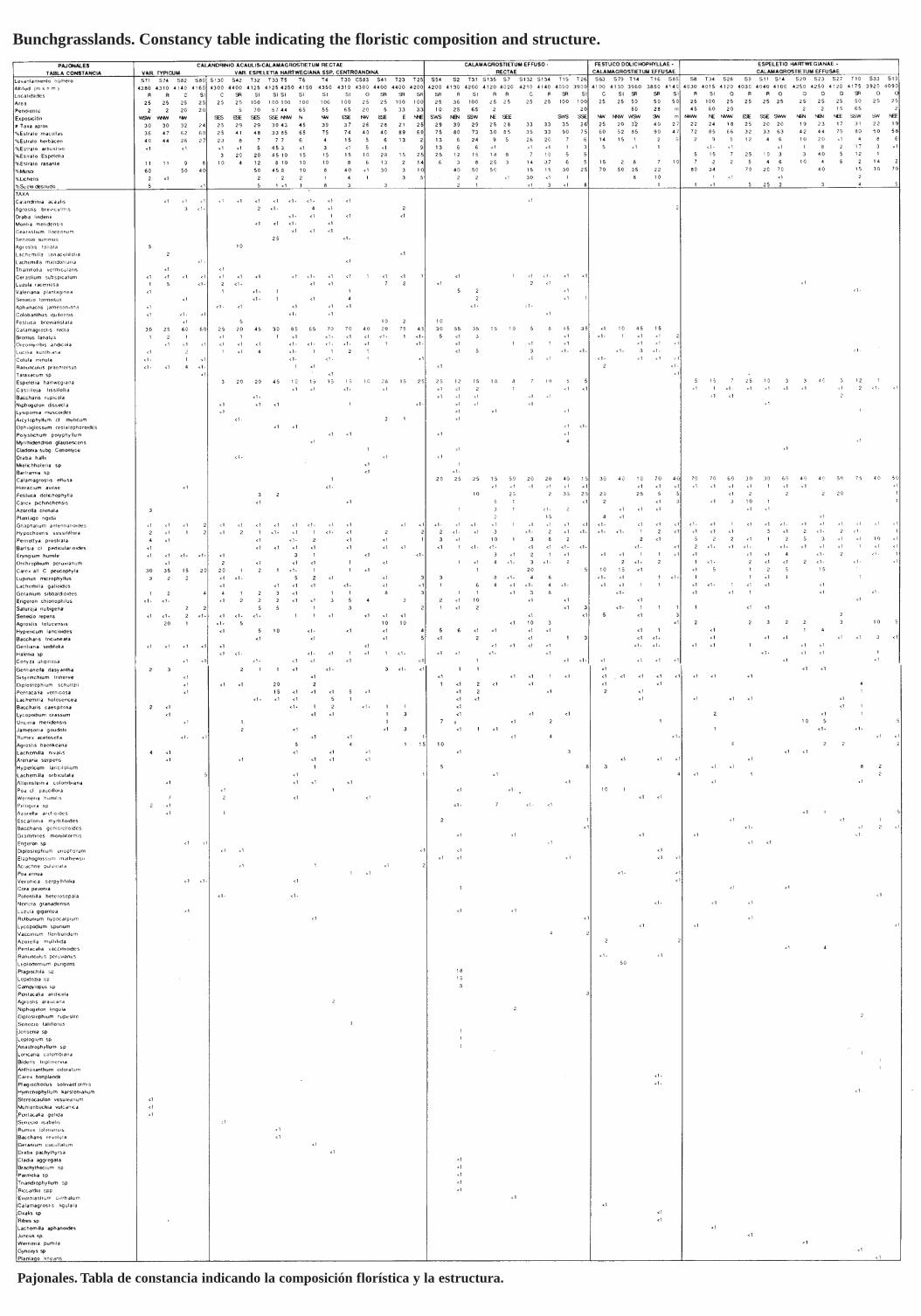
Bunchgrasslands. Constancy table indicating the floristic composition and structure.
Pajonales. Tabla de constancia indicando la composición florística y la estructura.

Cushion bogs. Constancy table indicating the floristic composition and structure.
Turberas de cojines. Tabla de constancia indicando la composición florística y la estructura.
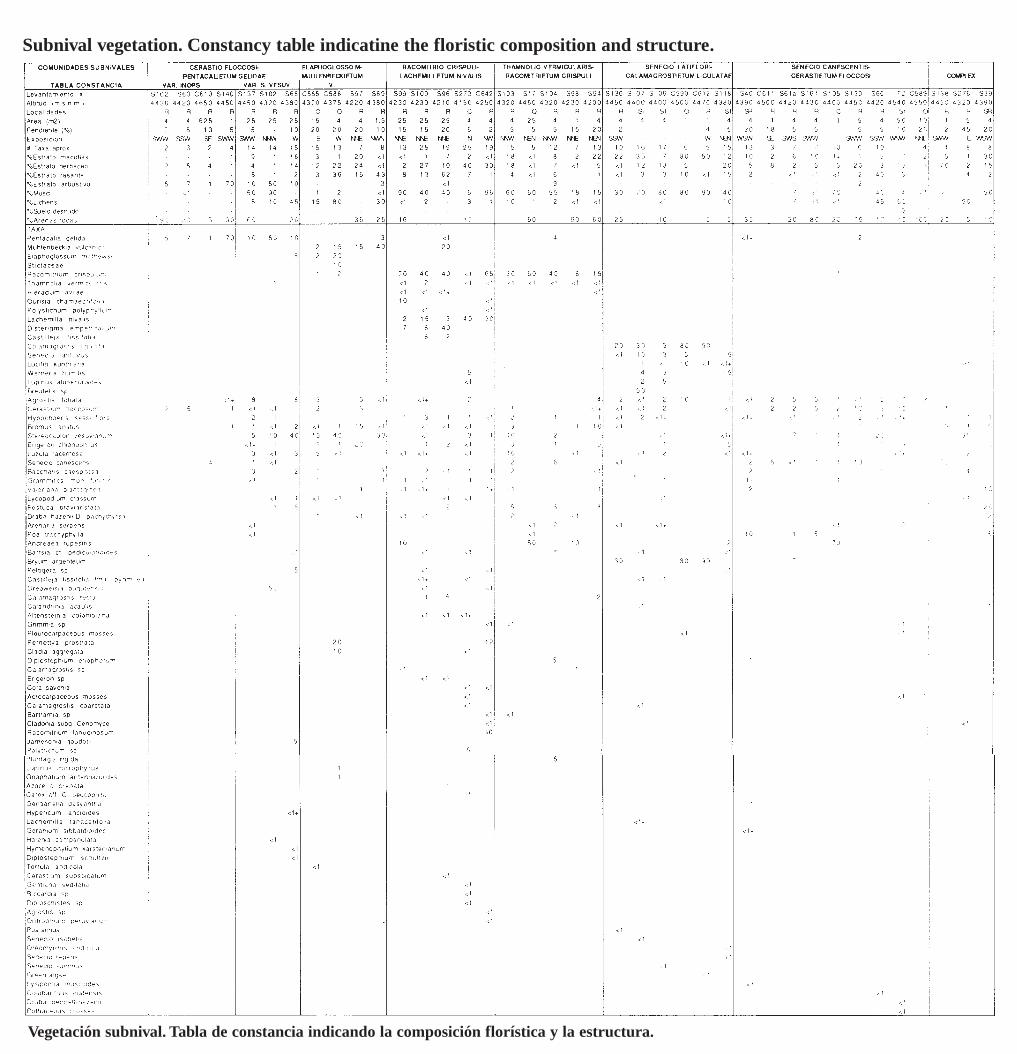
Subnival vegetation. Constancy table indicatine the floristic composition and structure.
Vegetación subnival. Tabla de constancia indicando la composición florística y la estructura.


















