
The Intergovernmental Oceanographic Commission of UNESCO...
12
The publication of Harmful Algae News is sponsored by the Spanish Institute of Oceanography, Vigo and the Institute of Biology, University of Copenhagen. The Intergovernmental Oceanographic Commission of UNESCO HARMFUL A LGAE NEWS An IOC Newsletter on toxic algae and algal blooms No. 28 August 2005 http://ioc.unesco.org/hab/news.htm (Cont’d on p.3) SEED: A new EU-US collaboration on HABs Detection of toxic Alexandrium catenella (Whedon & Kofoid) Balech in clam production zone of North Lake and Channel, Tunisia • Tunisia Fig. 2. Alexandrium catenella: (a) microphotograph with phase contrast; (b) microphotograph with epifluorescence, ventral view; (c) dorsal view; (d) Po, 1’ and Sa; (e) sulcal plates; (f) apical plates of epitheca; (e) Hypotheca. The word SEED is not an acronym or abbreviation; it directly refers to seed as produced by plants. However, the indication ‘SEED’ refers to a new project entitled Life cycle transforma- tions among HAB species, and the en- vironmental and physiological factors that regulate them, for which we do not want an acronym. The project is funded by the Global Change and Ecosystems Programme of the European Commission and the U.S. National Science Foundation. SEED started in March 2005 and will continue for three years. SEED is endorsed by the Global Ecology and Oceanography of Harmful Algal Blooms programme of the Intergovernmental Oceano- graphic Commission of UNESCO (GEOHAB) as a Targeted Research Project. Two coordinators, a steering com- mittee, and 12 partners make up the (Cont’d on p.2) A dozen species of Alexandrium have been identified in the Bay of Tu- nis, of which two are toxic, A. tamarense and A. minutum [1]. During our monitoring of the production zones of bivalve molluscs in northern Tuni- sia, A. catenella was detected for the first time in the Tunis Channel in Au- gust 1997. This species seems to be well adapted to the Mediterranean, and may have been introduced with ballast wa- ter [2]. In addition, these are the kinds of ecosystem, estuarine and lagoonal, which are most vulnerable to invasions [2].
Transcript of The Intergovernmental Oceanographic Commission of UNESCO...
1
The publication of Harmful Algae News is sponsored by the Spanish Institute of Oceanography, Vigoand the Institute of Biology, University of Copenhagen.
The Intergovernmental Oceanographic Commission of UNESCO
HARMFUL ALGAE NEWSAn IOC Newsletter on toxic algae and algal blooms
No. 28
August 2005
http://ioc.unesco.org/hab/news.htm
(Cont’d on p.3)
SEED: A new EU-US collaboration on HABs
Detection of toxic Alexandrium catenella (Whedon &Kofoid) Balech in clam production zone of North Lakeand Channel, Tunisia
• Tunisia
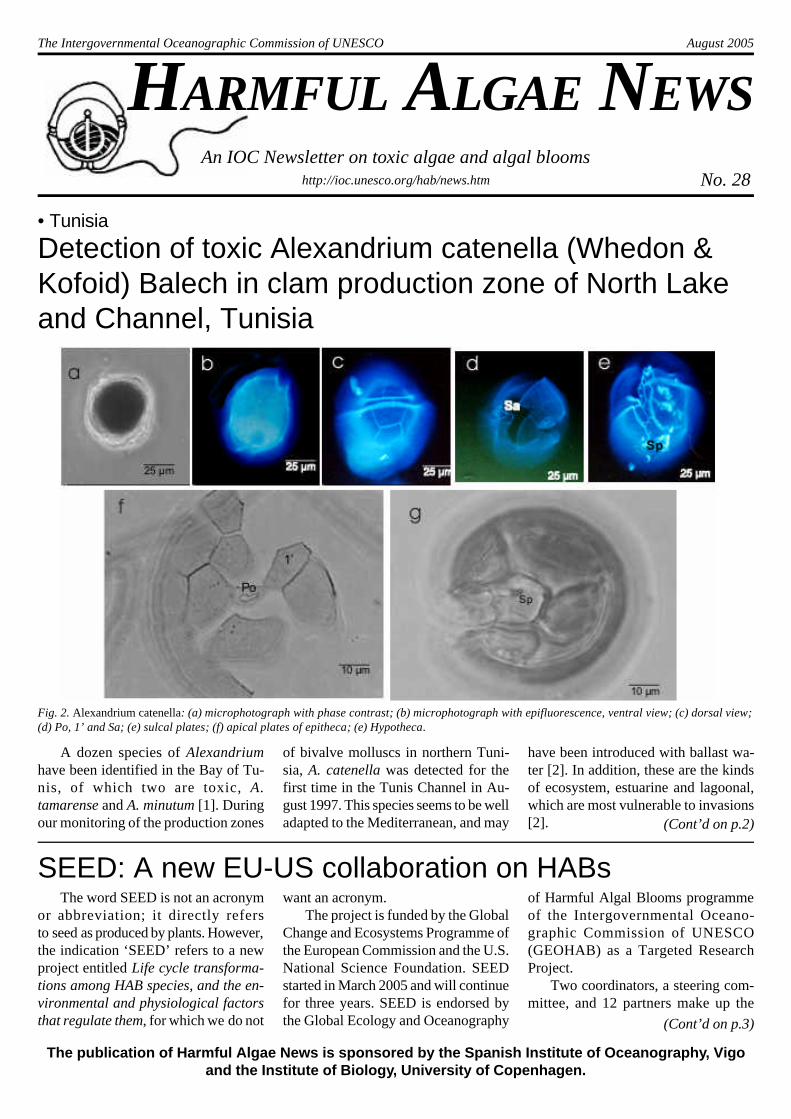
Fig. 2. Alexandrium catenella: (a) microphotograph with phase contrast; (b) microphotograph with epifluorescence, ventral view; (c) dorsal view;(d) Po, 1’ and Sa; (e) sulcal plates; (f) apical plates of epitheca; (e) Hypotheca.
The word SEED is not an acronymor abbreviation; it directly refersto seed as produced by plants. However,the indication ‘SEED’ refers to a newproject entitled Life cycle transforma-tions among HAB species, and the en-vironmental and physiological factorsthat regulate them, for which we do not
want an acronym.The project is funded by the Global
Change and Ecosystems Programme ofthe European Commission and the U.S.National Science Foundation. SEEDstarted in March 2005 and will continuefor three years. SEED is endorsed bythe Global Ecology and Oceanography
of Harmful Algal Blooms programmeof the Intergovernmental Oceano-graphic Commission of UNESCO(GEOHAB) as a Targeted ResearchProject.
Two coordinators, a steering com-mittee, and 12 partners make up the
(Cont’d on p.2)
A dozen species of Alexandriumhave been identified in the Bay of Tu-nis, of which two are toxic, A.tamarense and A. minutum [1]. Duringour monitoring of the production zones
of bivalve molluscs in northern Tuni-sia, A. catenella was detected for thefirst time in the Tunis Channel in Au-gust 1997. This species seems to be welladapted to the Mediterranean, and may
have been introduced with ballast wa-ter [2]. In addition, these are the kindsof ecosystem, estuarine and lagoonal,which are most vulnerable to invasions[2].
2
In the Mediterranean, the first ap-pearance of A. catenella dates from1983 along the Catalan coast [3], andits distribution gradually extended alongthe northwest Mediterranean coast withrecurrent blooms during the period 1997to 1999 in the port of Barcelona, andwith associated production of paralytictoxins [4]. Toxic episodes due to thepresence of gonyautoxins andsaxitoxins in L’étang de Thau are attrib-uted to A. tamarense with A. catenellaalso suspected [5]. The latter species hasalso been noted in Sardinian waters(Tyrrhenian Sea) [6].
The present report follows the an-nual pattern of this toxic species in the
Fig. 1. Sampling stations in shellfish production area of North lake and Channel of Tunis.
Tunis
N
Gulf of Tunis
Channel of Tunis
North lake of Tunis
T STEG1
T2
1 km
ChekliTunis T1
T Khéreddine1
T Radès2
South lake of Tunis
North Lake and Channel of Tunisia (Fig.1) where the main resources of the clamTapes decussatus are concentrated.These two sites have been monitoredfor toxic planktonic microalgae since1996 [7].
The North Lake of Tunis was ear-lier marked by the presence of recur-rent ‘green tides’ of Ulva lactuca, andimportant ecological changes have oc-curred in the last dozen years follow-ing restoration works which were madein the period 1984 to 1988. In particu-lar, there has been a net increase inmacrophyte diversity, and marinephanerogams have appeared [8].Among the toxic species mentioned in
the clam productionzones in the NorthLake and Channel,only species produc-ing diarrhetic toxins,Dinophysis spp andProrocentrum lima,have previously beendetected [9, 10]. Thenavigation channel isstrongly perturbedby anthropogenicpollution due to theexistence of threeports, with a directimpact on thebenthic macrofaunawhich may extend asfar as the adjacentGulf of Tunis [11]. Afirst appearance ofred water due to abloom of Noctilucascintillans was re-corded in March
2003 along the southwest coast of theGulf.
The characteristic morphology of A.catenella is shown in Fig. 2. These cellsare solitary, and rarely seen in chainsor pairs. Total length varies from 25 to32.5 µm, and the transverse diameterfrom 19 to 32 µm. The epitheca is veryfragile relative to the hypotheca, whichsometimes lacks the pore at the level ofplate Sp. This species is found at cer-tain times of the year (March 2005) ac-companied by Scripsiella trochoideaand Gonyaulax spinifera. Differentia-tion between these cells is made bymeans of epifluorescent microscopyafter staining the cells with calcofluor.
Annual variations in the abundanceof A. catenella above the critical levelof 500 cells/L have rarely been foundexcept in June in the Channel of Tunis,and in two periods, December and Feb-ruary to March, in the North Lake (Fig.3). Estimated maximum concentrationsare respectively 1300 cells/L in theChannel (T2) in March 2005 and in June2004, and 1000 cells/L in the NorthLake (T1 Chikly).
It is still too early to state that thisspecies is increasing in abundance in theclam production zones in the NorthLake and Channel of Tunis. Our futurestudies should allow us to follow theevolution of A. catenella which has beendetected since its first appearance.
References
1. Daly Yahia-Kéfi, O. et al., 2001. Oceanol. Acta29: S17- S25.
2. Wyatt, T. & J.J. Carlton, 2002. CIESMworkshop Monographs 20: 41- 46.
3. Margalef, R. & M. Estrada, 1987. Investig.Pesq. 51: 121-140.
4. Vila, M. et al., 2001. Mar. Ecol. Prog. Ser. 222:73-83.
5. Masselin, P. et al., 2001. Proc. 9th Int. Conf.Harmful Algal Blooms: 26-29.
6. Lugliè, A. et al., 2003. Bocconea, 16(2): 1045-1051.
7. Turki, S. 2004. Bull. INSTM 31 (in press).8. Trabelsi, E.B. et al., 2001. MEDCOAST 01, 2:
945-952.9. Turki, S. & A. El Abed, 1999. Bull. INSTM, 11:
165-174.10. Turki S. & A. El Abed, 2001. Harmful Algae
News 22: 10.11. Zammouri N. et al., 2001. MEDCOAST 01, 2:
641-650.
S. Turki & N. Balti, Institut Nationaldes Sciences et Technologies de laMer, 2016 la Goulette, Tunisia.Email: [email protected]
(Cont’d from p. 1)
0
200
400
600
800
1000
1200
F1 F4 M3 A1 A4 M3 J2 Jl4 S1 O4 N4 D3 J2 J'4 F3 M2
Month
T1Steg T1Chikly T1 Kherredine
0
200
400
600
800
1000
1200
1400
F1 F3 M1 M3 M4' A2 A4 M2 M4 J2 Jl2 A2 S1 O1 N1 N4
Month
T2Radès T2Canal
Fig. 3. Concentrations of A. catenella (cells/L) in the clam productionzone in the North Lake and Channel of Tunis (February 2004 - March2005).
North Lake of Tunis
Channel of Tunis
Cel
ls/L
Cel
ls/L

3
consortium of SEED and include sci-entists from 7 European countries andthe USA.
SEED aims to understand to whatextent environmental and physiologi-cal factors may influence the non-veg-etative stages of the life cycles of harm-ful algal bloom (HAB) species, therebycontributing to the increase in harmfulalgal blooms (HAB) in European ma-rine, fresh, and brackish waters. SEEDwill focus on the life histories of someof the most relevant HAB species inEurope.
The areas to be studied comprisethe regions bordering coastal sites in theWestern Mediterranean Sea, AtlanticOcean, North Sea, Baltic Sea, andSwedish lakes. All of these regions haveheavy anthropogenic influences: fish-eries, urban development, aquacultureand tourism. All are subject to the fre-quent occurrence of HABs, with a va-riety of detrimental impacts includinghuman intoxications, closure of shell-fish farms, water discoloration causinga negative impact on tourism, etc.
The approach of SEED is compara-tive, ranging from species to ecosystemlevels. Despite considerablephylogenetic distance, many morpho-logical and physiological traits of HABspecies are convergent and common.
The specific objectives are:1. To characterize and quantify the
different stages of the life cycle in abloom development of selected HABspecies in comparative environments.
2. To characterize the physiologi-cal mechanisms and tolerances under-lying dormancy stages/cysts, spores andakinetes, their formationand germination for key HAB species.
3. To explore the mating interac-tions of non-toxic and toxic HAB spe-cies to better understand the nature ofspecies invasions or dispersions and toevaluate the use of non-toxic strains asa possible mitigation strategy in HABs.
4. To quantify the flux of cells ofkey HAB species in different areas fromand to the sediments.
5. To investigate the role of sedi-mentary dynamics in the preservation,accumulation and dispersion of dor-mant stages.
6. To explore methods to identify
specific life cycle stages of key HABspecies through molecular or immuno-logical techniques.
7. To use the newly acquired knowl-edge in simple numerical models toverify the relevance of life history fea-tures and to develop or refine predic-tive and conceptual models of bloomdynamics for selected HAB species, tak-ing into account life-history.
8. To determine the role of dor-mancy stages of HAB species in differ-ent environments in relationship to theexpansion/increase of HABs. To formu-late principles that explain similaritiesbetween ecosystems.
To facilitate the development ofconceptual and numerical models ofHAB dynamics, it is imperative to rec-ognize common patterns of responseamong species. In recognition of thisconvergence and to carry out cost-ef-fective research, SEED will take advan-tage of existing model species whilekeeping the main focus on an array oftarget HAB species, which range frommarine to brackish to fresh water organ-isms, and cover a broad range ofphylogenetic types (dinoflagellates, dia-toms, raphidophytes, cyanobacteria).
New results will be used to eluci-date and evaluate the role of life cycleprocesses, especially dormancy stages,in bloom dynamics. Furthermore, oneaim of SEED is to integrate the acquiredknowledge into the development of newmodel formulations. This improved
modelling will enhance prediction andsupport the development of new man-agement and mitigation strategies forthese species.
For further information on theproject, visit our website:www.icm.csic.es/bio/seed/
E. Garcés, EU -Coordinator,,Departament de Biologia Marina iOceanografia, Institut de Ciències delMar, CMIMA, Passeig Marítim de laBarceloneta, 37-49, E08003Barcelona, Spain
D.M. Anderson,US -Coordinator, Woods HoleOceanographic Institution, BiologyDepartment, Mail Stop 32, Redfield332, Woods Hole MA 02543-1049USA
(Cont’d from p. 1)
From bottom to top and from left to right: Toni Jordi, Gotzon Basterretxea, Esther Garcés, KarinRengefors, Donald Anderson, Mercedes Masó, Robin Raine, Jordi Camp, Sílvia Anglès, JordiSolé, Kees van Lenning, Rosa Isabel Figueroa, Antonella Penna, Marina Montresor, Magda Vila,Kalle Olli, Nicolas Touzet, Deana Erdner, Santiago Fraga, Sanna Suikkanen, Isabel Bravo, AnkeKremp, Jane Lewis, Hartmut Barth, Marta Estrada, Mariagrazia Giacobbe, Elena Bertozzini,Cecilia Satta, Antonella Lugliè, Maria Laamanen, Carmen Hormigo, Dolors Blasco.
Harmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae Newsonly exists if the Editor getsinput from YOU!
Write the Editor NOW with newson your work, HAB events inyour country or region, or anyother matter you wish to sharewith HAB scientists and manag-ers worldwide. Harmful AlgaeNews has more than 2000subscribers.

4
Ostreopsis siamensis (Dinophyceae) a newtychoplanktonic record from Isabel Island NationalPark, Pacific Mexico
Fig. 1. Study area and sampling locations.
Fig. 2. Thecal plates of Ostreopsis siamensis: a) Epitheca, b) Hypotheca, c) Ventral region, d)Small and large pores.
• Mexico
An early summer sampling pro-gramme was carried out from 18th Mayto 26th June 2004, around Isabel IslandNational Park (Fig. 1). Isabel Island isa major Pacific nesting site for seabirds,principally frigate birds, blue footed andyellow footed boobies, pelicans, swal-lows, sea gulls and brown footed boo-bies. Its origin is volcanic, dating backapproximately 3.5 million years; it hasbeen used as a temporary camp for fish-ermen in the area. The climate is tropi-cal, with a long wet summer season(May to October) and a short dry win-ter season (November to April). Therainy season coincides with hurricanesand tropical storms in the zone. Diverseand abundant coral reef communitiescover 4.93% of the island [1]. The studywas done to describe the generalphytoplankton community, and to docu-ment the distribution of toxicdinoflagellates. Until now, harmful al-gae have never been investigated in thisisland. We report for the first time theoccurrence of the potentially toxicdinoflagellate Ostreopsis siamensisSchmidt 1902, and it’s geographical dis-tribution in the Mexican Pacific area.
Phytoplankton samples were col-lected during early summer at five sta-
tions in Isabel Island National Park. Thesamples containing Ostreopsissiamensis were obtained from surfacenet tows using a 62 µm pore size plank-ton net, towed by boat at a low speedfor 4 minutes, very close to coral reefin shallow waters. The specimens werefixed immediately with 4% glutaralde-hyde solution. Surface water tempera-ture (25oC), salinity (35.3‰) and dis-solved oxygen (6.15 mg.L-1) were meas-ured in-situ with an YSI model 85.Phytoplankton cells were identified us-ing the Utermöhl technique and a phase-contrast microscope. Morphologicaldetails were observed through scanningelectron microcopy (JEOL/OE jsm-6360).
The morphological characteristicsof the organisms were in accord withthe descriptions of Fukuyo [2] and Faust[3], which coincide in all plates of the
genus, the epithecal (Fig. 2a), thehypothecal (Fig. 2b), ventral zone (Fig.2c) and both kind of pores (Fig. 2d). Theshapes and dimensions of the Po plateare characteristic of each species anddifferent from the other species asshown in Fig. 3. The dorso-ventral dis-tance ranged from 88 to 129 (½ 101)µmand transdiameter from 65 to 90 (½78.4)µm. Our report confirms the pres-ence of O. siamensis as part of the natu-ral biota in this latitude. It is possiblethat Isabel Island waters generatefavorable environmental conditions forthe proliferation of benthicdinoflagellates, which are transportedlater by currents to the plankton zone(= tychoplankton). The species has of-ten been reported as benthic, epiphytic,but is most frequently associated withsand and as an epiphyte on seaweeds[4]. Further analysis will outline the

5
Fig. 3. The morphology of the Po plates: a) O. siamensis, b) O. ovata.
seasonal distribution, outbreak times,summer-winter survival and speciessuccession in this fishery location, aswell as its abundance. According toFaust [3] the ecology of O. siamensis isdiverse; the type species was identifiedfrom plankton samples collected bySchmidt from the Gulf of Thailand.Some years later, the morphology of O.siamensis was reexamined by Fukuyo[2] based on specimens from RyukyuIsland in Japan. O. siamensis is alsoknown from South Florida, FloridaKeys, Puerto Rico, U.S. Virgin Islands,Hawaii, Guam and New Zealand [5, 6].
This species is known to be toxinproducer. Nikajima et al. [7] were thefirst, followed by Yasumoto et al., [8]and Holmes et al. [9] to indicate thepresence of toxins produced by O.siamensis populations, which producesostreocin D, an analog of palytoxin, oneof the most potent marine toxins known.Palyotoxin can accumulate in fish andcrabs and is considered responsible forclupeotoxism, an illness linked to theeating of sediment-feeding fishes in Ja-pan [6]. It is also thought to be the causeof the multiplicity of symptoms in thepathology of ciguatera [10]. The toxinaffects first coral grazing fish, thenpiscivorous fish, notably groupers, andfinally humans. Other organisms af-fected are snapper, mackerel, jack, goatfish, barracuda, parrot fish, and somegastropods [5]. Until now, no clinicalcase of ciguatera has been reported inthe island by health authorities, but thehigh to moderate abundances ofOstreopsis cells observed raise the pos-sibility that ostreocin or another kindof palytoxin, are responsible for differ-ent seafood poisoning (ciguatera,clupeotoxism, etc.). Strong coastal sur-face currents have been proposed asmajor players in the transport of cellsto the island waters.
Acknowledgements
We thank the authorities of theIsabel Island National Park, Biol.Cecilia García Chavelas (Director) forher cooperation in this project, and M.C.Amílcar Cupul Magaña for their excel-lent technical assistance and supportduring the field samplings. Y. Hornelasfor her skilled assistance using SEM.This study was partially supported by
the project SEMARNAT-2002-CO1-1005.
References
1. Cupul-Magaña, A.L. & P. Medina-Rosas, 2002.Mem. XIII Congreso Nacional de Oceanografía.Puerto Vallarta, Jalisco, México. pp. 151.
2. Fukuyo, Y., 1981. Bull. Jpn. Soc. Sci. Fish. 47:967-978.
3. Faust, M.A., 1996. J. Phycol. 32: 1053-1065.4. Faust, M.A. & S.L. Morton, 1995. J. Phycol.
31: 456-463.5. Anderson, D.M., et al., 1993. Woods Hole
Oceanographic Institution Tech. Report. WHOI93-02. Woods Hole, Ma. 59 pp.
6. Rhodes, L., et al., 2002. NZ J. Mar. Freshw.Res. 36: 631-636.
7. Nakajima, I., et al., 1981. Bull. Jpn. Soc. Fish.47: 1029-1033.
8. Yasumoto, Y., et al., 1987. Biol. Bull. 172: 128-131.
9. Holmes, M.J., et al., 1988. Proc. 6th Int. CoralReef Symp. Australia, 3: 49-54.
10. Bomber, J.W., 1991. In: Ciguatera SeafoodsToxins. D. Millar (Ed.) CRC Press Inc.: 135-170.
M.C. Cortés Lara, Dpto. de Ciencias,Centro Universitario de la Costa (U.de G.), Campus Puerto Vallarta,Fax: (01-322) 2262200 and 01.Email: [email protected].
R. Cortés Altamirano, UnidadAcadémica Mazatlán (ICMyL-UNAM), Apdo. 811, Mazatlán 82040,Sinaloa, México. Fax: (01-669)9826133.Email: [email protected].
A. Sierra Beltrán, Lab. GenéticaMolecular (CIBNOR). Apdo. 128,23000 La Paz B.C.S., México.Tel: (52). 612 12 53633 ext. 3805,Fax: (52) 612 12 53625.Email: [email protected].
A. Reyes Juárez, Dpto. de Ciencias,Centro Universitario de la Costa (U.de G.), Campus Puerto Vallarta,Fax: (01-322) 2262200 and 01.Email: [email protected].

6
Toxic and harmful algal species in the Izmit Bay,Marmara Sea
• Turkey
During the last 40 years, nutrientconcentrations have obviously in-creased in many estuaries and coastalwaters in the world due to the influenceof human activities [1, 2]. This prob-lem of marine eutrophication has beenhighlighted in recent years by the oc-currence of increasingly severe toxicphytoplankton blooms in many near-shore waters worldwide. Toxicphytoplankton blooms in the sea are ofeven greater concern than in freshwa-ter ecosystems [3]. Red tides and othertoxic and/or noxious algal bloomsmainly due to progressiveeutrophication from terrestrial inputshave been observed by reporters andTurkish scientists [4]. Balkis and Aktan[5] reviewed plankton studies in theMarmara Sea (except Izmit Bay) con-tinuing since the 1990s [6-10, 11], lead-ing to the identification of 11 toxicphytoplanktonic species.
Izmit Bay, located in the north-east-ern part of the Marmara Sea is one ofthe most polluted areas in the MarmaraSea, and an important centre for Turk-ish industry. The bay has been receiv-ing more than 300 industries’ effluentstogether with untreated domestic wastewaters from the small cities, and fromIzmit which is the most populous cityof the region. Pollution problems havegradually become more serious since1960. Previous studies showed that thebay waters have become eutrophic [1,
12] and red tides due to Prorocentrumspecies, and fish mortalities, have beenobserved in some periods (unpublisheddata).
This paper focuses on the toxic and/or harmful phytoplanktonic and benthicmicroalgal species recorded in IzmitBay. In March 1999 and September2000, phytoplankton, epiphytic, andepipelic algae were collected from sam-pling sites.
Fourteen toxic and harmful species;dinophytes Ceratium furca (Ehrenberg)Claparéde & Lachmann, Dinophysisacuminata Claparéde & Lachmann,Dinophysis acuta Ehrenberg,Dinophysis caudata Saville-Kent,Dinophysis sacculus Stein,Gymnodinium sanguineum Hirasaka,Lingulodinium polyedrum (Stein)Dodge, Noctiluca scintillans(Macartney) Kofoid & Swezy,Phalacroma rotundatum (Claparéde &Lachmann) Kofid & Michener,Prorocentrum micans Ehrenberg,Prorocentrum minimum Schiller; anddiatoms Pseudo-nitzschia delicatissima(P.T. Cleve) Heiden in Heiden & Kolbe,Pseudo-nitzschia sp. (Nitzschia seriata(Hasle) were recorded in the bay. Inaddition to these species, Prorocentrumlima (Ehrenberg) Dodge was recordedas epipelic and epiphytic (on Bryopsishypnoides Lamour, Codium fragile(Sur) Hariot, Cystoseira barbata C.Agardh var. barbata, Cymadocea
nodosa (Uchria) Ascherson and Zosteranoltii (Horneman)) in the littoral zone.Prorocentrum lima is a benthicdinoflagellate usually found attached toor associated with macrophytes, float-ing detritus, debris or other substratesand less commonly in plankton. Thesespecies were common in Izmit Bay, buttoxic and noxious algal blooms were notrecorded during the sampling period.
References
1. Morkoc, E., et al., 2001. Environ. Int. 26: 157-161.
2. Shen, Z.L., 2001. Estuar. Coast. Shelf Sci. 52:211-224.
3. Smith, V.H., et al., 1999. Environ. Pollut. 100:179-196.
4. Koray, T., 2002. Workshop on LessepsianMigration, 20-21 July 2002, Gökçeada, 40-45.
5. Balkis, N. & Y. Aktan, 2004. Rapp. Comm. Int.Mer Médit. 37: 486 pp.
6. Aubert, M., et al., 1990. Etud. Hydrobiol.,Chim. Biolo., Tome 3, CERBOM (Nice), 47 pp.
7. Uysal, Z., 1996. Turk. J. Bot. 20: 321-327.8. Uysal, Z. & M. Ünsal, 1996. Turk. J. Bot. 20:
519-525.9. Tüfekçi, V. & E. Okus, 1998. Karadeniz-
Istanbul Bogazi kistagi, bogaz ve bogazinMarmara Denizi çikisindaki fitoplanktondagilimi. Büyüksehirlerde atiksu yönetimi vedeniz kirlenmesi kontrolu sempozyumu. 18-20Kasim 1998, ISKI, 157-172.
10. Balkis, N., 2000. Oebalia 26: 97-108.11. Balkis, N., 2003. J. Plankton Res. 25: 703-717.12. Okay, O.S., et al., 2001. Mar. Pollut. Bull. 42:
361-369.
Y. Aktan, Istanbul University,Fisheries Faculty, Ordu Cad., 200,Laleli, 34470 Istanbul, Turkey.Email: [email protected]
First record of Chattonella marina in Bahía de la Paz,Gulf of California
• Mexico
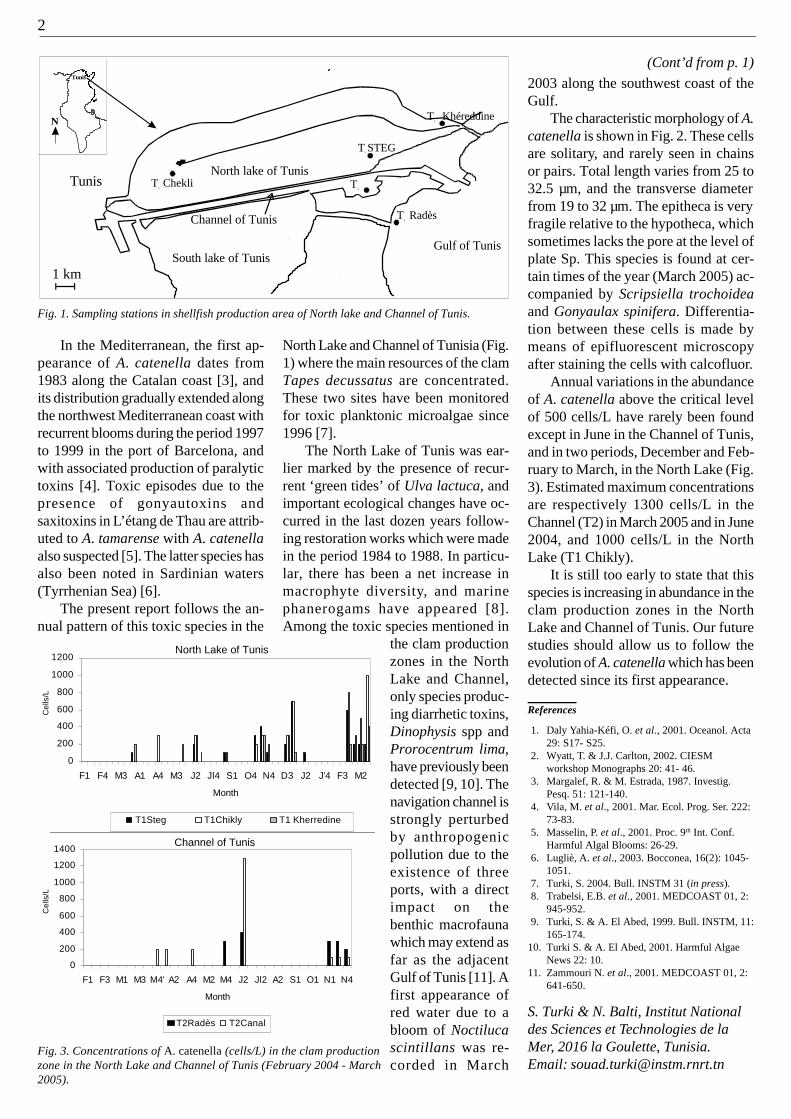
Harmful algal blooms are commonevents in Bahía de La Paz, a coastal la-goon located on the southwest side ofthe Gulf of Calfornia [1]. A massivebloom of Chattonella marina was ob-served from April 24 to May 9, 2005 inBahía de La Paz (Fig. 1). Live cells ofC. marina were identified in unfixed
samples. For cell enumeration watersamples were fixed using acid lugol(1%) and analyzed with the standardUtermöhl technique [2]. Fixed cellswere deformed and shrank making itdifficult to identify without observinglive cells (Fig. 2). Cell densities fluctu-ated between 1.9 and 3.5 x 106 cells L-1.
Cysts of C. marina were not observedin these samples, which suggests thatenvironmental conditions were good forthis species. Sea surface temperatureduring the bloom was 22°C. Duringmonitoring on May 4th at a differentcollection site, C. marina cells were alsoobserved but in lower concentrations
7
Fig. 2. Light photomicrographs of Chattonella marina collected in Bahía de La Paz. Live cells(1-3), acid lugol fixed cells (4), and spherical cell showing flagella (3).
Fig. 1. Collection site showing location and size of Chattonella marina bloom (shaded area) inBahía de La Paz. * Indicates the site of additional sample.
which ranged from 11 to 23 x 103
cells L-1. As in other areas, different cellmorphologies were observed (Fig. 2).However, in the samples collected onMay 4th, round cells resembling tem-porary cysts were observed.
The presence of the ichthyotoxicphytoflagellates C. marina, C. cf ovata,
Fibrocapsa japonica, and Heterosigmaakashiwo (Raphidophyceae) have beenreported in various localities on theMexican Pacific coast as well as in theGulf of Mexico [3, 4, 5], including aprevious bloom report on the east coastof the Gulf of California [6]. Howeverthis is the first report of a bloom of C.
marina for Bahía de La Paz. The high-est abundances estimated for this eventwere an order of magnitude higher thanprevious reports [6].
Most marine Raphidophyceae areichthyotoxin producers, and havecaused severe fish mortalities with sig-nificant damage to the aquacultureeconomy in several countries. InMexico loss of marine resources wasassociated with the presence of C. ma-rina and C. cf ovata in Kun Kaak Bay,Sonora [6]. In Bahía de La Paz no fishor shellfish mortalities were observed,however the magnitude of this event isof special concern for fisheries andaquaculture activities in this area.
It is possible that these species havebeen undetected in most Mexican wa-ters due to traditional methods used forsampling and preserving phytoplankton.Raphidophyceans like other flagellates,are very delicate, and deform or disin-tegrate in formalin solution; samplespreserved with lugol have differentmorphology but can be recognized by awell-trained eye. We recommend thatduring the monitoring of harmful algae,non-preserved samples should be takento observe the presence of delicatephytoplankton forms. During thebloom, clonal cultures were isolated.These will be used in the near future toconfirm toxicity.
References
1. Gárate-Lizárraga, I., 2001. Oceánides 16(2):127-134.
2. Hasle, G.R., 1978. In: Sournia, A. (Ed.).Phytoplankton Manual, Monographs onOceanographic Methodology 6, UNESCO,Paris, pp. 191-196.
3. Cortés-Lara, C., et al., 2003. Harmful AlgaeNews 24: 1-4.
4. Alonso-Rodríguez, R., et al., 2004. In: Elfitoplancton en la camaronicultura ylarvicultura: importancia de un buen manejo.ICMyL & CESASIN, México. 147 pp.
5. Band-Schmidt, C.J., et al., 2004. Hydrobiología515: 79-84.
6. Barraza-Guardado, R., et al., 2004. HarmfulAlgae News 25: 7-9.
C. Band-Schmidt, A. Martínez-Lópezand I. Gárate-Lizárraga. Dpto.Plancton y Ecología Marina, CentroInterdisciplinario de CienciasMarinas (CICIMAR-IPN), Apdo. 592,La Paz, B.C.S. 23000, México.Email: [email protected],[email protected], [email protected].
8
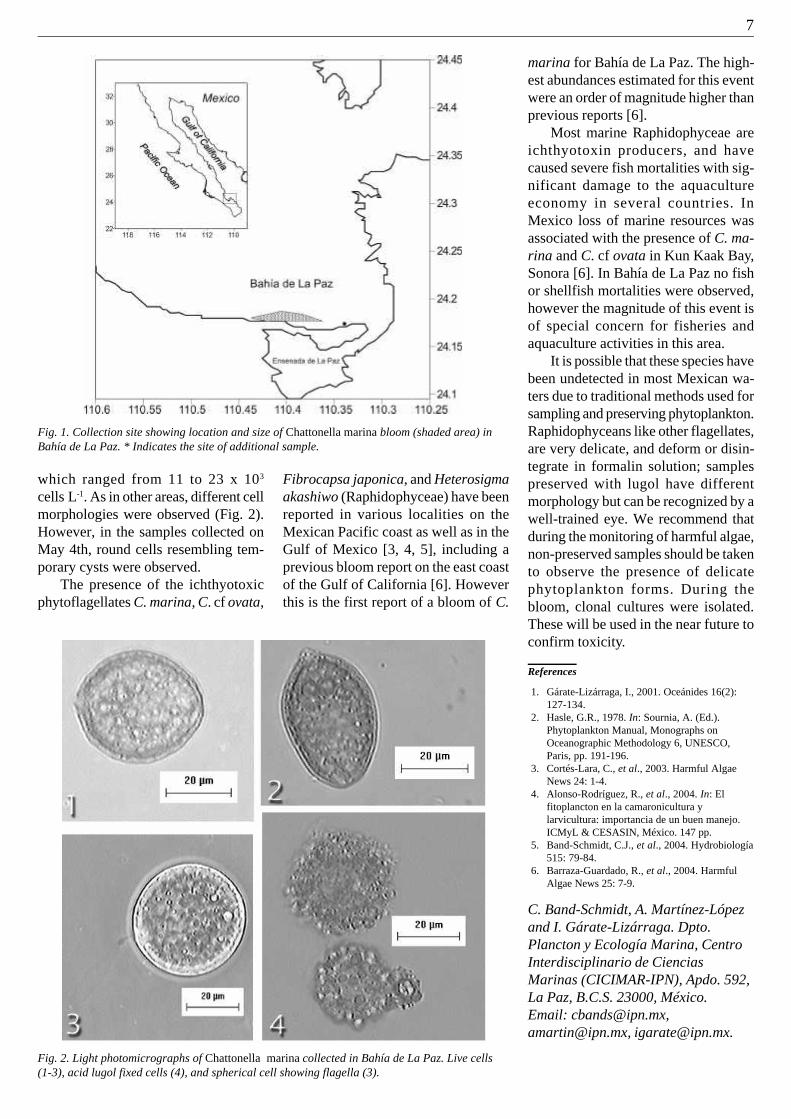
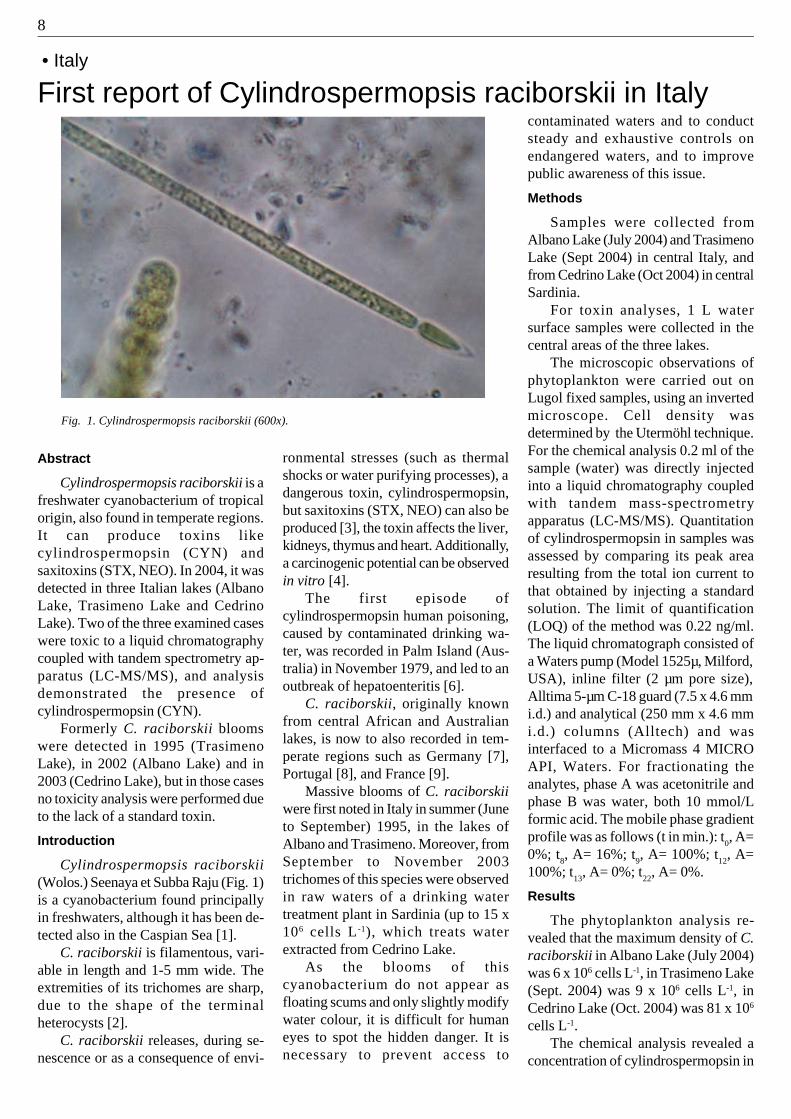
First report of Cylindrospermopsis raciborskii in Italy
Fig. 1. Cylindrospermopsis raciborskii (600x).
• Italy
Abstract
Cylindrospermopsis raciborskii is afreshwater cyanobacterium of tropicalorigin, also found in temperate regions.It can produce toxins likecylindrospermopsin (CYN) andsaxitoxins (STX, NEO). In 2004, it wasdetected in three Italian lakes (AlbanoLake, Trasimeno Lake and CedrinoLake). Two of the three examined caseswere toxic to a liquid chromatographycoupled with tandem spectrometry ap-paratus (LC-MS/MS), and analysisdemonstrated the presence ofcylindrospermopsin (CYN).
Formerly C. raciborskii bloomswere detected in 1995 (TrasimenoLake), in 2002 (Albano Lake) and in2003 (Cedrino Lake), but in those casesno toxicity analysis were performed dueto the lack of a standard toxin.
Introduction
Cylindrospermopsis raciborskii(Wolos.) Seenaya et Subba Raju (Fig. 1)is a cyanobacterium found principallyin freshwaters, although it has been de-tected also in the Caspian Sea [1].
C. raciborskii is filamentous, vari-able in length and 1-5 mm wide. Theextremities of its trichomes are sharp,due to the shape of the terminalheterocysts [2].
C. raciborskii releases, during se-nescence or as a consequence of envi-
ronmental stresses (such as thermalshocks or water purifying processes), adangerous toxin, cylindrospermopsin,but saxitoxins (STX, NEO) can also beproduced [3], the toxin affects the liver,kidneys, thymus and heart. Additionally,a carcinogenic potential can be observedin vitro [4].
The first episode ofcylindrospermopsin human poisoning,caused by contaminated drinking wa-ter, was recorded in Palm Island (Aus-tralia) in November 1979, and led to anoutbreak of hepatoenteritis [6].
C. raciborskii, originally knownfrom central African and Australianlakes, is now to also recorded in tem-perate regions such as Germany [7],Portugal [8], and France [9].
Massive blooms of C. raciborskiiwere first noted in Italy in summer (Juneto September) 1995, in the lakes ofAlbano and Trasimeno. Moreover, fromSeptember to November 2003trichomes of this species were observedin raw waters of a drinking watertreatment plant in Sardinia (up to 15 x106 cells L -1), which treats waterextracted from Cedrino Lake.
As the blooms of thiscyanobacterium do not appear asfloating scums and only slightly modifywater colour, it is difficult for humaneyes to spot the hidden danger. It isnecessary to prevent access to
contaminated waters and to conductsteady and exhaustive controls onendangered waters, and to improvepublic awareness of this issue.
Methods
Samples were collected fromAlbano Lake (July 2004) and TrasimenoLake (Sept 2004) in central Italy, andfrom Cedrino Lake (Oct 2004) in centralSardinia.
For toxin analyses, 1 L watersurface samples were collected in thecentral areas of the three lakes.
The microscopic observations ofphytoplankton were carried out onLugol fixed samples, using an invertedmicroscope. Cell density wasdetermined by the Utermöhl technique.For the chemical analysis 0.2 ml of thesample (water) was directly injectedinto a liquid chromatography coupledwith tandem mass-spectrometryapparatus (LC-MS/MS). Quantitationof cylindrospermopsin in samples wasassessed by comparing its peak arearesulting from the total ion current tothat obtained by injecting a standardsolution. The limit of quantification(LOQ) of the method was 0.22 ng/ml.The liquid chromatograph consisted ofa Waters pump (Model 1525µ, Milford,USA), inline filter (2 µm pore size),Alltima 5-µm C-18 guard (7.5 x 4.6 mmi.d.) and analytical (250 mm x 4.6 mmi.d.) columns (Alltech) and wasinterfaced to a Micromass 4 MICROAPI, Waters. For fractionating theanalytes, phase A was acetonitrile andphase B was water, both 10 mmol/Lformic acid. The mobile phase gradientprofile was as follows (t in min.): t
0, A=
0%; t8, A= 16%; t
9, A= 100%; t
12, A=
100%; t13
, A= 0%; t22
, A= 0%.
Results
The phytoplankton analysis re-vealed that the maximum density of C.raciborskii in Albano Lake (July 2004)was 6 x 106 cells L-1, in Trasimeno Lake(Sept. 2004) was 9 x 106 cells L-1, inCedrino Lake (Oct. 2004) was 81 x 106
cells L-1.The chemical analysis revealed a
concentration of cylindrospermopsin in

9
Albano Lake of 15 ng ml-1, toxin tracein Trasimeno Lake (0.46 ng ml-1) andno toxicity was found in Cedrino Lake.
Discussion
Cylindrospermopsin is an unusualalkaloid (m.w. 415 Da) and consists ofa trycilic guanidine moiety combinedwith hydroxymethyluracil [10]. Apartfrom liver toxicity, mutagenicity [11]was shown in vitro and evidence alsoexists for its carcinogenicity in vivo[12]. The LD
50 (i.p.) in mice is cells/kg
2,1 mg/kg over 24 h [13]. The occur-rence of C. raciborskii on the three Ital-ian lakes is correlated with a toxic pres-ence of cylindrospermopsin only in twocases. The maximum density of C.raciborskii was found in the CedrinoLake but the maximum toxin concen-tration was registered in Albano Lake:this demonstrates that wild populationsof cyanobacteria species, catalogued astoxic, not necessarily or always, areimplicated in toxin production.
Few years ago, the presence of C.raciborskii was revealed in Europe,only as far as Germany [6] Portugal [7],France [8] and now the presence in Italyis confirmed by observations over a ten-year period. The extending geographi-cal distribution of this species could belinked to the migration of water birds
like gulls, as they are important agentsof dispersal for many aquatic organisms,by carrying resistant forms within theirintestinal tract. The dispersal of fresh-water species not capable of active over-land transport and lacking resistantforms, while not so well documentedhas largely been attributed to transportvia external surfaces of birds [14]. Com-parative studies on ecological condi-tions and on phylogenetic relations ofEuropean wild populations of C.raciborskii could help to explain theirdifference in toxin production, too.
Anyway, these data show for thefirst time that C. raciborskii can alsobe found in Italian water bodies, alsofor drinking use, and that monitoringthis cyanobacterium and associatedcylindrospermopsin is a real emergentnecessity.
References
1. Neilan, B.A., et al., 2003. Mol. Ecol. 12: 133-140.
2. Shafik, H.M., 2001. Acta Biol. Hung. 54: 121-136.
3. Pomati, F., et al., 2004. Microbiology 150: 455-461.
4. Rao, P.V., et al., 2002. J. Environ. Biol. 23:215-224.
5. Komarkova, J., et al., 1999. Algol. Stud. 94/Arch. Hydrobiol. Suppl. 129: 207-222.
6. Dilwyn J.G. & M.L. Saker, 2003. Environ.Toxicol. 18: 78-93.
7. Fastner, J., et al., 2003. Toxicon 42: 313-321.8. Saker, M.L., et al., 2003. Ecotoxicol. Environ.
Saf. 55(2): 243-250.9. Briand, J.F., et al., 2002. Water Res. 36: 3183-
3192.10. Shaw, G.R., et al., 2000. Ther. Drug Monit.
22(1): 89-92.11. Shen, X., et al., 2002. Toxicon 40: 1499-1501.12. Falconer, I.R. & A.R. Humpage, 2001. Environ.
Toxicol. 16: 192-195.13. Ohtani, I., et al. 1992. J. Am. Chem. Soc. 114:
7941-7942.14. Proctor, V.W. & C. Malone, 1965. Ecology 46:
728-729.
G. Manti, D. Mattei, V. Messineo andS. Melchiorre, Dept. of Environ. &Primary Prevention, Nat. Institute ofHealth, via Regina Elena, 299 - 00161Rome, Italy.
S. Bogialli, Dept. of Chemistry,University of Rome «La Sapienza»,P.le Aldo Moro, 5 - 00185 Rome, Italy.
N. Sechi, P. Casiddu and A. Luglié,Dept. of Botany and Plant Ecology,via Muroni, 25 - 07100 Sassari, Italy.
M. Di Brizio, ARPA umbria, ViaPievaiola – Loc. S.Sisto - 06132Perugia, Italy.
M. Bruno, Dept. of Environment andPrimary Prevention, Nat. Institute ofHealth, via Regina Elena, 299 - 00161Rome, Italy, Email: [email protected].
IOC Training Course on Qualitative and QuantitativeDetermination of Algal Toxins, Germany, 22 February to 2March 2005 The IOC-BMU-AWI-DZMB-FSU
JENA Training Course on Qualitativeand Quantitative Determination of Al-gal Toxins was held at theWattenmeerstation Sylt, AWI in List/Sylt, Germany from 22nd February to 2nd
March 2005. The course was the firstin a series of five cooperative trainingcourses implemented by IOC andForschungsinstitut Senckenberg(DZMB), Friedrich-Schiller-UniversitätJena (FSU) und Stiftung Alfred-Wegener-Institut für Polar- undMeeresforschung in der Helmholtz-Gesellschaft (AWI). It was financed byIOC and Germany: Bundesministeriumfür Umwelt, Naturschutz undReaktorsicherheit (BMU); AppliedBiosystems Darmstadt and Mr J. Gosch,Gosch-Seafood Tourist Service GMBH,List/Sylt, supported the course.
The course was hosted at theWattenmeerstation Sylt of AWI and or-

10
Marine and Freshwater Toxins Analysis First JointSymposium and AOAC, Baiona, Spain, 11 - 14 April 05
Task Force Meeting was held April11-14, 2005, in Baiona, Spain. The con-ference was convened under blue skiesat the Hotel Talaso Atlantico just abovethe rugged coastline of the prolific mus-sel-producing Galicia region. The meet-ing was well attended by a global mixof academics, government regulators,seafood and analytical industries andindustry associations. Over 120 partici-pants pre-registered and over 150 at-tended in total.
The Minister of Fisheries of theGalician Government as well as theformer Minister of Health of Galicia,who was instrumental to success in hissupportive role working closely with the
University of Vigo since the first effortsto manage the threat of marine toxinsin Galicia, opened the conference. Alsopresent was the Vice Rector of Institu-tional Affairs. Symposium Chair AnaGago Martinez, and Task Force ChairJames Hungerford spoke briefly andnext warm recollections were given byJose Antonio Rodriguez Vasquez, chairof the Analytical and Food ChemistryDept., University of Vigo, of excitingand productive collaborations of theUniversity with visiting researchersfrom Japan and Canada. Other keynotepresentations followed on the currentstate of marine biotoxin monitoring inGalicia and an overview and progress
report of the Marine and FreshwaterToxins Task Force.
Many excellent keynote and con-tributed presentations were giventhroughout the meeting. A unique aspectof the conference in Baiona was theintegration of the Symposium and TaskForce meetings, with the symposiumpresentations (72 total, 35 oral)preceding subgroup discussions in rel-evant toxin areas. This allowed for dis-cussion of method applicability, currentvalidation efforts, future directions, anda variety of other topics relating to toxinanalysis methods and their validation.Discussion threads from the new AOACon-line forums of the Task Force sup-
ganized by Dr. Malte Elbrächter (FIS)Germany and in cooperation with Dr.Henrik Enevoldsen, IOC, Paris, Prof.Dr. Allen Cembella, AWI-Bremerhaven, Germany and Prof.Bernd Luckas, FSU-Jena, Germanywith his team. Applications arrivedfrom 96 persons from 46 nations. Dueto financial and space limitations only12 participants originating from 11countries could attend the trainingcourse.
The goal of the course was to pro-vide participants with an overview ofthe state of the art of different analyti-cal methods for qualitative and quanti-tative determination of phycotoxins.Lectures and demonstrations on all rel-evant toxins were performed, and theparticipants were encouraged to estab-lish scientific collaboration.
Qualitative and quantitative deter-mination of algal toxins using differentanalytical methods (HPLC with selec-
tive detection, HPLC/MS coupling andLC-MS) were made on samples of al-gae and mussels provided mainly by theparticipants. The laboratory experi-ments focused on the analysis of Para-lytic Shellfish Poisoning (PSP),Diarrhetic Shellfish Poisoning (DSP),Amnesic Shellfish Poisoning (ASP),and cyanobacterial toxins by applicationof chromatographic methods.
The next training course is plannedfor October 2006.

11
plemented these in-person subgroupmeetings and discussion periods. Sub-group areas included in the Baiona pro-gram included assays for saxitoxins, LCmethodology for saxitoxins and domoicacids, okadaiates and azaspiracids,yessotoxins, and ciguatoxins. BenjaminSuarez-Isla, James Lawrence, Ana GagoMartinez, Aurelia Tubaro, and co-chairRobert Dickey (for ciguatoxin chairRichard Lewis) chaired these sessions.Two other areas discussed werecyanobacterial toxins and LC-MS meth-ods for multiple toxin groups. Thesevery active areas are new to the TaskForce and will soon become formal sub-groups when chairs are appointed thisyear.
Benjamin Suarez-Isla, the subgroupchair, led discussion of saxitoxin assayswith the primary focus on an IAEA-funded initiative to validate a receptor-binding assay (RBA) for the saxitoxinsusing radiolabeled saxitoxin. He pre-sented results of a Single LaboratoryValidation (SLV) study submitted byFran Van Dolah, the RBA assay projectleader. Participants also suggested pur-suit of receptor binding assays usingalternative (non radioactive) labels.James Lawrence led subgroup discus-sion on LC methods for saxitoxins anddomoic acids. A positive vote by theTask Force last year has moved a pre-column oxidation LC method for thesaxitoxins into review by AOAC forofficial method status This method isnow in the final stages of First Actionapproval following minor revisions.Still, post-column methods forsaxitoxins also remain an area of inter-est, with recent advances made by Japa-nese and Canadian research groups.
Several presentations on thecyanobacterial toxins were also givenand the availability of toxin test kits wasdiscussed, building on responses ob-tained following a request for this in-formation in the on-line forum. Partici-pants shared some of their experienceswith the kits and additional kits werebrought into the discussion, as was theneed for cyanobacterial reference stand-ards. The ciguatoxin subgroup sessionbegan with a proposal put forward inthe ciguatoxin on-line forum by RobertDickey to establish a reference collec-tion of ciguatoxic fish tissues. The fish
tissues would cover a range ofciguatoxicity and tissue origin andwould be characterized by a combina-tion of cytotoxicity assay and LC-MSmethods. Practical issues including sta-bility, storage requirements, costs, andissues regarding tissue sample transportwere discussed.
Okadaic acids, their congeners andalso azaspiracids were discussed in asubgroup chaired by Ana GagoMartinez and this addressed a range oftopics. These included both PP2A as-say marketed in Japan based on enzymeinhibition and a Biosense method basedon competitive binding assay, also us-ing PP2A. Also the relative merits ofthese two functional assays versus LC-MS-MS became the subject of a livelydiscussion. At the conclusion of thissession was a statement that validationof both classes of methods, those basedon assays and also LC-MS, should bepursued without competition betweenthe two. At this stage each group, con-sisting of the LC-MS enthusiasts andthose working on functional assays likePP2A, should reach a consensus on ex-traction conditions, which congeners toinclude, etc. Then validation plansshould continue to move ahead as theoverall process can be time consuming.The toxin sessions concluded with pres-entations on yessotoxin toxicology andyessotoxin detection methods followedby a yessotoxin subgroup meetingchaired by Aurelia Tubaro of Italy. Ex-isting methodology and validation planswere discussed from the standpoint ofsingle laboratory validation, and the is-sue of which YTX forms to include inmethods was also discussed. The sym-posium was concluded with presenta-tions by European government, indus-try, and industry associationstakeholders. In these talks the need forvalidated methods was stressed, as wasthe need to make them stakeholder-friendly.
Comments from those who attendedthis four-day combined symposium andTask Force meeting were very positiveand discussions and plans made at theconference rekindled hopes in the vali-dated of additional methods.
The on-line forums will be used toreview and carry forward what was dis-cussed in Spain and to engage in fur-
ther discussions both the Baiona partici-pants and all interested in development,validation and application of methodsfor the toxins. As another step in ourcontinuous path towards validatedmethods, a meeting will be held June22-24, 2005 in Seattle, Washington,USA, in connection with the PacificNorthwest Regional AOAC Meeting. InOrlando, Florida, USA, a full TaskForce meeting consisting of severaltoxin subgroups, will meet as part of oursecond symposium-Task Forceconference at the AOAC Annual Meet-ing September 11-15, 2005. Two sym-posia, including a full day devoted toMarine and Freshwater Toxins Analy-sis: Quality Methods for Public Healthand International Trade, and also a half-day session to honor Wiley Award win-ner Mike Quilliam, will precede theseTask Force meetings. As with all ourmeetings, we encourage both research-ers and stakeholders (and potentialstakeholders) to attend.
SEPTEMBER 2006
12th INTERNATIONAL CON-FERENCE ON HARMFUL ALGAE
September 4-8, 2006. Copenhagen,Denmark.
Call for submission of abstracts andregistration form: February 1, 2006.
Deadlines:• Abstracts reception: May 1, 2006• Early registration: June 1, 2006• Late registration: July 15, 2006
Further information can be foundat: www.issha.org.

12
Compiled and edited by Tim Wyatt, Instituto de Investigacio-nes Marinas, CSIC, Eduardo Cabello 6, 36208 Vigo, Spain;Tel.: +34 986 23 19 30/23 19 73; Fax: +34 986 29 27 62;E-mail: [email protected] and Mónica Lion, CentroCientífico y de Comunicación sobre Algas Nocivas COI-IEO, Apdo. 1552, 36200 Vigo, Spain; Tel.: +34 986 49 2111; Fax: +34 986 49 20 03; E-mail: [email protected]
The opinions expressed herein are those of the authorsindicated and do not necessarily reflect the views ofUNESCO or its IOC. Texts may be freely reproduced andtranslated (except when reproduction or translation rightsare indicated as reserved), provided that mention is made ofthe author and source and a copy sent to the Editors.
Project Coordinator: Henrik Enevoldsen,IOC Science and Communication Centre on Harmful AlgaeUniversity of Copenhagen, Botanical Institute ØsterFarimagsgade 2D, DK-1353 Copenhagen K, DenmarkTel.: +45 33 13 44 46, Fax.: +45 33 13 44 47,E-mail: [email protected] Editor: Institute of Biology, Compenhagen
HARMFUL ALGAE NEWS
News from the 11th HAB meeting inCapetown, South Africa
Announcement of election of offic-ers for the next 2 years include PatriciaTester as President, Karen Steidingeras acting past President, GustaffHallegraeff and Beatriz Reguera asVice President, Tracy Villareal as Sec-retary and Nina Lundholm as Treasurer.
ISSHA’s emphasis during the nexttwo years will be to update the web site.Tracy Villareal is working with StephenBates, former ISSHA Secretary, to ac-complish this. Take a look atwww.issha.org and see how that is com-ing. ISSHA is committed to increas-ing its visibility and will be featured ina poster during the Third Symposiumon Harmful Algae in the U.S., 2-7 Oc-tober 2005 (www.visitasilomar.com).Also, ISSHA is a co-host for the sum-mer ASLO (American Society ofLimnology and Oceanography) meet-ing in Victoria 6-9 June 2006. Increas-ing our membership base is key to oursuccess as an organization and we wel-come new members, especially, studentmembers and say thank you to our life-
time members for their strong show ofsupport.
Maureen Keller Awards
Five Maureen Keller StudentAwards were presented by ISSHA at the11th Conference on Harmful Algae heldin Cape Town, South Africa. Pleasejoin me in congratulating these youngresearchers for their excellent oral pres-entations and posters. Also, thanks goto the other students who presented 67posters and 19 oral presentations andwhose work helped spark many discus-sions during the meeting. The 32 judgeswho generously agreed to help evalu-ate the efforts of this diverse body ofstudent work were recognized individu-ally at the banquet. However, I wouldlike to say again how much I appreciatetheir assistance to the ISSHA awardscommittee.
Though no biography is present,mention should also be given to the fifthawardee, M-L Chateau-Degat for herpresentation on Neurological Signs ofCiguatera Disease.
Pat Tester, ISSHA President.
ISSHA President’s Corner
Future events
Members of the ISSHA Council during the IV General Assembly of the Society (Cape Town, SouthAfrica, 18 Nov. 2004). From left to right: Beatriz Reguera, Gustaaf Hallegraeff, Don Anderson,Adriana Zingone, Ojvind Moestrup, Edna Granéli, Allan Cembella, Max Taylor and Pat Tester.
DECEMBER 2005
GEOHAB: OPEN SCIENCEMEETING. HABS AND STRATIFI-CATION
December 5-8, 2005. UNESCOHeadquarters, Paris, France.
The conference topics include:• Physical processes relevant to strati-
fication• Maintenance of HAB populations
in thin layers• Selection of assemblages by differ-
ent turbulent regimes• Changes in small scale physical
properties caused by the phyto-plankton community
• Implications for sampling, monitor-ing and operational oceanography
• Detection systems
Important dates:• Abstract deadline: October 1, 2005.• Early registration deadline: Novem-
ber 1, 2005.
Further information can be found at:www.geohab.info.
Harmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae NewsHarmful Algae News
Previous issues of HAN andnewsletters of the IOC HABProgramme can be downloadedat http://ioc.unesco.org/hab/news.htm
Requests for subscription
Subscription to HAN is made bysending a request with a com-plete address to Ms V. Bonnet
([email protected]).([email protected]).([email protected]).([email protected]).
©UNESCO 2005. Printed in France ISSN 0020-7918


















