
Purification and characterization of a soluble salicylic acid-binding ...


vi
Table of Contents
Abstract iii
Acknowledgements v
Table of contents vi
List of Figures xi
List of Tables xiii
List of Abbreviations xiv
Chapter 1: Introduction 1
1.1 MULTICELLULARITY 3
1.1.1 Origins of multicellular transition 3
1.1.2 Historical introduction of cell adhesion 5
1.1.3 Dictyostelium discoideum life cycle 8
1.1.4 Mechanisms involved in Dictyostelium pattern formation 19
1.1.5 Signal transduction and regulation of cell type differentiation in
Dictyostelium 22
1.1.6 Theories proposed for cell sorting in the multicellular development 27
1.1.6.1 Differential Adhesion Hypothesis (DAH) 28
1.1.6.2 Differential Surface Contraction (DSC) 29
1.1.6.3 Mechanistic explanations for cell sorting during morphogenesis (DAH
vs DSC) 31
1.1.6.4 Chemotaxis and differential adhesion combined as a model in
Dictyostelium 33
1.2 Regulation of Dictyostelium adhesion molecules during development 36
1.2.1 DdCAD-1 39
1.2.1.1 Gene structure and regulation of DdCAD-1 expression 39
1.2.1.2 Structural characteristics of DdCAD-1 41
1.2.1.3 Multiple roles of DdCAD-1 during Dictyostelium development 41
Calcium-dependent cell-cell adhesion 41 Anti-adhesion effect of secreted DdCAD-1 42 Cell type proportioning and Cell sorting 44
1.2.2 Adhesion complexes involved in the multicellular development 45
1.2.2.1 Adherens junction-like structure 46

vii
1.2.2.2 Other junctional complexes 47
1.3 SECRETORY PATHWAYS 50
1.3.1 Conventional secretory pathways 50
1.3.2 Unconventional secretory pathways 53
1.3.2.1 History of unconventional secretory processes 53
1.3.2.2 Unconventional secretion of signal-peptide-containing proteins 57
1.3.2.3 Unconventional secretion of soluble proteins from eukaryotic cells 58
Translocation across the plasma membrane 59
Lysosome-dependent pathway 61
Microvesicle-dependent secretion 62
1.3.3 The contractile vacuole system of Dictyostelium discoideum 63
1.3.3.1 Organization and function of the contractile vacuole system 63
1.3.3.2 Osmoregulation and Other Functions of the Contractile Vacuole 65
1.3.3.3 Is there a compartment corresponding to the CV system in higher eukaryotes? 69
1.3.4 Identifying unconventionally secreted proteins 71
1.4 SOCIAL INTERACTIONS OF MICROORGANISMS 72
1.4.1 Background information about social behaviours in microorganisms 72
1.4.1.1 Social interactions of Dictyostelium discoideum 73
1.4.1.2 Other microorganisms that are involved in the social interactions 76
1.4.2 The Problem of cooperation 77
1.4.3 Mechanisms that are involved in the altruistic cooperation 78
1.4.3.1 Limited dispersal 80
1.4.3.2 Kin discrimination 80
1.4.3.3 Green-beard genes 81
1.4.3.4 Other mechanisms of social evolution 84
Pleiotropy 84 Phoenix genes 85
1.5 HYPOTHESES AND RATIONALE OF THE THESIS 88
1.5.1 Characterization of the homophilic binding site of DdCAD-1 88
1.5.2 Elucidation of the DdCAD-1 transport mechanism during the early phase of
development 89

viii
1.5.3 Assessment of the role of DdCAD-1 in Dictyostelium pattern formation
89
Chapter 2: New insights into Ca2+-dependent cell-cell adhesion mediated by DdCAD-1 in Dictyostelium
91
2.1 Summary 92
2.2 Introduction 93
2.3 Experimental Procedures 95
Cell-to-substratum attachment assay 95 Fluorescent microspheres-to-cell binding assay 95 Antibody-induced cap formation 95 Construction and expression of His6-tagged fusion proteins 96 45Ca2+-overlay assay 96 Fluorescent microsphere-to-substratum attachment assay 96
2.4 Results 98
NMR structure of Ca2+-free and Ca2+-bound DdCAD-1 98 Ca 2+-binding sites 100 Structural comparison to other cell adhesion proteins 101 N-terminal domain mediates homophilic binding 103 C-terminal domain tethers DdCAD-1 to cell membrane 107 Reverse-charge mutations affect homophilic binding 107 Structural model of the Ca2+-bound DdCAD-1 dimer 109
2.5 Discussion 113
Ca2+-dependent adhesion mediated by DdCAD-1 114 Distinct roles for the two domains of DdCAD-1
114
Chapter 3: The Cell Adhesion Molecule DdCAD-1 is imported into contractile vacuoles by membrane invagination in a Ca2+- and conformation-dependent manner
118
3.1 Summary 119
3.2 Introduction 120
3.3 Experimental Procedures 123
Construction of DdCAD-1-GFP, N-GFP and C-GFP expression vectors and cell transfection
123
Immunofluorescence labeling of cells and laser scanning confocal microscopy 123 Isolation of contractile vacuoles and cytosol 125 Expression of His6-tagged mutant DdCAD-1 proteins 126

ix
In vitro reconstitution of DdCAD-1 import into contractile vacuoles 128 45Ca2+-overlay assay 128 Antibody-induced cap formation 128 Chemical cross-linking of DdCAD-1 129
3.4 Results 130
Import of DdCAD-1-GFP via invagination of vacuolar membrane 130 Import of DdCAD-1 into contractile vacuoles requires Both N- and C-terminal domains
133
Surface expression and secretion of DdCAD-1-GFP 135 In vitro import analysis of GFP-fusion protein 135 Requirement of Ca2+ in the import of DdCAD-1 into contractile vacuole 140 Effect of conformation on the import of DdCAD-1 into contractile vacuole 143
3.5 Discussion 149
Chapter 4: cadA Is a Single-Gene Green Beard that Regulates Morphogenesis through Differential Spatiotemporal Expression in Dictyostelium
154
4.1 Summary 155
4.2 Introduction 156
4.3 Experimental Procedures 159
Construction of DdCAD-1-GFP and mutant plasmids for cell transfection 159 Development on non-nutrient agar or soil plates 161 Cell cohesion assay 161 Temporal and spatial expression pattern of DdCAD-1 162 Analysis of cell sorting in slugs 162 In vitro reconstitution of DdCAD-1 using recombinant proteins 163 Antibody-induced cap formation 163 Flow cytometry analysis 164 Chemotactic cell migration assay 164
4.4 Results 165
The cadA gene displays both green beard and anti-green beard behaviour 165 Anti-green beard effects of the cadA gene led to cheating of cadA-null cells in chimeras
167
Preferential localization of cadA+ cells in prestalk region of chimeras during development
167
Rescue of cadA-null cells using wild-type and mutant constructs of DdCAD-1 171 Dynamic changes in the temporal and spatial distribution of DdCAD-1 during development
173
Enrichment of DdCAD-1 in the cell-cell contact regions of anterior cells 176 Rescue of cadA-null phenotype by in vitro reconstitution of DdCAD-1 178

x
Distinct chemotactic response of AX4 and cadA-null cells cAMP 184 4.5 Discussion 186
Chapter 5: Conclusions and Future Directions 192
5.1 Conclusions 193
5.1.1 Ca2+-dependent homophilic cell adhesion mediated by DdCAD-1 193 5.1.2 Mechanistic characterization of DdCAD-1 transport into contractile vacuole 195 5.1.3 cadA is a single-gene green beard that regulate morphogenesis through
differential cell adhesion in Dictyostelium
197
5.2 Future Directions 199
5.2.1 What is the mechanism by which DdCAD-1 is transported through invagination of the contractile vacuole?
199
5.2.2 Does calmodulin play a role in DdCAD-1 transport to the contractile vacuole?
201
5.2.3 What is the anchoring protein for DdCAD-1? 203 5.2.4 What are the mechanisms involved in the differential distribution of DdCAD-
1 in the prestalk and prespore cells? 205
5.2.5 Is DdCAD-1 present in the adherens junction-like structures in the constriction region of the culminant?
208
5.3 Concluding Remarks
210
References 211

1
Chapter 1
Introduction
Portions of this chapter have been published in the following book chapter:
Sriskanthadevan, S., Ivanov, I., Yang, C., and Siu, C. (2007). Novel Functions and Transport
mechanism associated with the Ca2+-dependent cell adhesion molecule DdCAD-1 in
Dictyostelium. Recent Research Developments in Cell Biology. 3: 9-21.

2
Adhesion molecules are of fundamental importance in the regulation of pattern
formation in multicellular organisms. An understanding of the structure-function
relationships, as well as the spatiotemporal expression patterns of adhesive molecules will
provide insights into the nature of their adhesive activity and role during multicellular
development. My thesis has focused on a unique soluble cell adhesion molecule DdCAD-1
expressed by Dictyostelium discoideum cells. My thesis research has three specific aims: (1)
to investigate the structure-function relationships of DdCAD-1, (2) to elucidate the transport
mechanism of DdCAD-1 mediated by contractile vacuole, and (3) to examine the role of
DdCAD-1 in pattern formation during development. Therefore, I have organized the thesis
introduction into five main sections. The Introduction begins with a historical background of
the origins of multicellularity with a focus on the involvement of adhesion molecules. A
detailed review of Dictyostelium discoideum as a multicellular model organism is included in
this section. The second section explores the role of adhesion molecules during development
of Dictyostelium. The third section contains a detailed review of the conventional and
unconventional secretory pathways, since DdCAD-1 is synthesized as a soluble cytoplasmic
protein and then transported through an unconventional pathway for secretion and surface
presentation. The fourth section is devoted to social interactions of microorganisms because
of the inherent function of multicellularity in the social behaviour of microbes. Finally, the
fifth section describes the thesis objectives and the rationale of the studies.

3
1.1 MULTICELLULARITY
1.1.1 Origins of Multicellular transitions
Organisms undergo transitions to more complex biological organization by means of
natural selection (Bonner, 1988; Bonner, 1998; McShea, 2002). There are several transitions
that occurred over the course of evolution that can be considered major evolutionary turning
points. Grosberg and Strathmann (2007) summarized the following as major evolutionary
transitions: (a) the compartmentalization of replicating molecules (first cells); (b) the
formation of chromosomes; (c) the use of DNA and proteins as the fundamental elements of
the genetic code and replication; (d) the generation of the first eukaryotic cell containing
choloroplasts and mitochondria; (e) evolution of sexual reproduction; (f) the evolution of
multicellular organisms from unicellular ancestors; and (g) the establishment of social groups.
Multicellularity originated at least 25 times independently from a variety of ancestral
unicellular lineages, once for the metazoan, and multiple times in plants, fungi and the
Eubacteria (Bonner, 1998; Grosberg and Strathmann, 2007). Multicellular forms exist in all
three of the life kingdoms. On the bacterial clade, cyanobacteria, myxobacteria and
actinobacteria are the three main multicellular lineages, while on the eukaryotic side these
include plants, animals and fungi, as well as several lineages of algae and slime molds
(Dictyostelid and Acraisid) (Fig. 1.1) (Rokas, 2008). Since the multicellularity originated
several times over history, there should be potential advantages of multicellular lineages over
unicellular ones. They include size related advantages, functional specialization and division
of labour. Further, metabolic cooperation also has contributed to the transition since key
metabolic processes such as photosynthesis and nitrogen fixation (Kaiser, 2001) cannot
concurrently take place within a cell. Motility-mitosis trade-offs: the loss of mitotic activity

EUKARYOTES
Fung
iNostoc
Lifestyle:Unicellular
Multicellular
BACTERIA
Figure 1.1 Multiple independent origins of multicellularity. A set of independently
evolved multicellular bacterial and eukaryotic lineages (blue) and their unicellular
Mechanism:Terrestrial & cell-aggregatory
Aquatic & non-divisional
relatives (green). On the bacterial clade, cyanobacteria, myxobacteria, and actinobacteria
are the three main multicellular lineages, whereas on the eukaryotic side these include
plants, animals, and fungi, as well as several lineages of algae and slime molds
(Dictyostelid and acrasid). Cell-aggregatory multicellular lineages whose origins are
terrestrial are shown in italics (Adapted from Rokas 2008)terrestrial are shown in italics. (Adapted from Rokas 2008)
4

5
in the somatic cells of large volvocaceans is associated with trade-off between cell division
and locomotion (Grosberg and Strathmann, 2007). In addition to these advantages, the
repeated inventions of multicellularity have given rise to a remarkable variety of
morphologies. The most significant difference between the various multicellular lineages is
that of complexity (Bonner, 1988; Carroll, 2001).
Multicellularity most likely evolved via one of the following mechanisms: clonal
development from unicellular spore or zygote or aggregative development (Bonner, 1998;
Bonner, 1999; Grosberg and Strathmann, 2007). Examples of ‘cell-aggregatory’ multicellular
lineages are the myxobacteria and the Dictyostelid slime molds. Based on the analysis of
number of independently evolved pairs of unicellular and multicellular relatedness, scientists
have come up with the idea that the proteins involved in cell adhesion, cell-cell signalling and
cell differentiation are genetic tool kits for multicellularity (King et al., 2007; Vogel and
Chothia, 2006).
1.1.2 Historical introduction of cell adhesion
Adhesion phenomena have been observed in a wide range of taxa, from prokaryotes to
eukaryotes and from unicellular organisms to metazoans. The study of the differential
binding and sorting out of cells in multicellular organisms began in 1907 when Wilson (1907)
showed that mechanically dissociated and re-mixed cells of two different species of marine
sponges sorted out to produce two aggregates, each consisting of the cells of only one species.
Later, a similar approach was applied by Holtfreter (1948a,b), who showed that embryonic
cells dissociated from different tissues could sort out in a mixture to form regions with
structures characteristic of parent tissues, the so-called histotypic aggregates. These
experiments demonstrate the presence of selective adherence among cells of various types. It

6
has been proposed that “ the phenomenon of cellular adhesion is the prerequisite, for the
evolution and ontogenesis of multicellular organisms” (Steinberg and Gilbert, 2004; Townes
and Holtfreter, 1955).
Since these early studies, many attempts have been made to develop more direct short
term assays in order to isolate molecular fractions that might be responsible for differential
selectivity or specificity of cell-cell adhesion (Balsamo and Lilien, 1974; Hausman and
Moscona, 1976; Merrell et al., 1975; Oppenheimer, 1975; Shur and Roth, 1975). Different
views about the nature of adhesion began to emerge. Some held that adhesion differences
were due to differences in interactions at the cell surface (such as electrostatic or van der
Waals interaction) (Curtis, 1967). Steinberg (1970) proposed that differential adhesion was
based on the principles of thermodynamic interactions, while others thought that cell
recognition was mediated by intermolecular specificity (Hausman and Moscona, 1976;
Moscona, 1962). These ideas formed the basis for a number of proposals (Turing, 1952;
Wolpert, 1971) concerning the nature of the specification of tissue pattern or positional
information in morphogenesis. The subsequent isolation and characterization of cell adhesion
molecules (CAMs) has helped clarify many of these ideas (Damsky et al., 1984; Edelman,
1984; Edelman, 1985; Edelman et al., 1983).
The original identification of most CAMs was based on the observation that
monovalent antibodies (Fabs) directed against CAMs interfere with cell-cell adhesion in in
vitro assays (Beug et al., 1970; Beug et al., 1973). Such an approach has led to the
identification of many candidates for CAMs. However, antibody inhibition of adhesion alone
is not a proof of adhesive function. The binding of antibodies to the cell surface may
sterically block access to molecules other than the specific antigen or may affect cell adhesion
indirectly by perturbing the function of a regulatory molecule that has global effects on cell

7
adhesion (Gerisch, 1986). An important conceptual development is that cell adhesion is
viewed as a cell-regulatory phenomenon with molecular specificity, that cell surface
modulation is a major mechanism in pattern formation (Edelman, 1976). This and other
conceptual developments have led to new criteria for CAM identification: (1) demonstration
of specific adhesive behaviour in an immunological assay, (2) sufficient characterization of
the molecular structure, binding mechanism, and specificity (Hoffman et al., 1982;
Rutishauser et al., 1982; Siu et al., 1986), (3) demonstration of CAMs on the membrane of
cells they interact, (4) the appearance of CAMs in definite sequences of expression during
embryogenesis consistent with the initial formation of cell collectives and boundaries with
morphological and functional significance (Gerisch, 1986), and (5) direct evidence for
morphogenetic function as shown by alteration of tissue structure after perturbation of CAM
binding function or CAM expression after mechanical or chemical disruption of morphology
(Edelman, 1986).
As discussed earlier, multicellularity arose more than once during eukaryotic
evolution. Did cell adhesion arise independently on each occasion, or are these ancient cell
adhesion systems conserved among different eukaryotic groups? Recent results show that cell
adhesion proteins related to cadherin, IgG-like CAM and C-type lectin are present in sponges
(the most distant animal branch), and in eukaryotic groups outside the metazoan lineage,
indicating that these forms of adhesion arose prior to animal evolution (Abedin and King,
2008). King et al. (2003) suggest that the discovery of diverse cadherins in choanoflagellates,
suggests that cadherins may have contributed to metazoan origins. Choanoflagellates are not
metazoans and did not evolve from sponges. Therefore, the ancestor of choanoflagellates and
metazoans was probably a unicellular organism that is capable of forming simple colonies
(Abedin and King, 2008).

8
Choanaflagellates and animals belong to a larger phylogenetic eukaryotic group, the
opisthokonts (Fig. 1.1), which also includes fungi and microsporidia. There is little evidence
for adhesion systems homologous to those of animals in either of these groups, although there
may be homologous proteins present. For example yeast contain a gene with similarities to α-
integrins (Sundstrom, 2002). Both α and β integrins have been cloned from sponges and an
aggregation factor (AF) protein contains an RGD integrin-binding motif (Wimmer et al.,
1999). Yeast contains a protein, Ax12p, with extracellular cadherin-like repeats (Dickens et
al., 2002) that is required for bud site selection during bipolar budding (Cullen and Sprague,
2002). Sponges also express both C-type lectins and calcium-independent lectins that help
provide a complex immune system in these organisms (Schroder et al., 2003). The
Dictyostelids are more distantly related to metazoan, belonging to the amoebozoa, which
diverged before the opisthokont lineage (Fig. 1.1). The genetic and cell-biological studies
indicate that cell adhesion play a major role in co-ordinating and patterning the fruiting body.
Detail of the CAMs present in Dictyostelium will be discussed in the following sections.
1.1.3 Dictyostelium discoideum life cycle
Several aspects of Dictyostelium life cycle (Fig. 1.2) make this cellular social amoeba
especially suitable for studies of cell-cell interactions, cell migration, signalling, development,
and social interactions. The Dictyostelium genome has been sequenced (Eichinger et al.,
2005). At a systems level, it provides a level of complexity that is greater than the yeast, but
much simpler than plants or animals. The gene-dense chromosomes encode approximately
12,500 predicted proteins, a high proportion of which have long, repetitive amino acid tracts
(Eichinger et al., 2005). A proteome-based phylogeny shows that the amoebozoa diverged

Fruiting body
0/24
618 Time (hrs)
12
Streaming
Slug
Streaming
Fi 1 2 Di li di id lif l M l i ll l d l iFigure 1.2 Dictyostelium discoideum life cyle. Multicellular development in
Dictyostelium is initiated by starvation and leads to the formation of a fruiting body,
which is composed of a ball of spores resting on top of a stalk containing vacuolated
cells. The transition between growth and aggregation is mediated by the chemotaxis of
cells towards cAMP to form a multicellular aggregate. During this process cells stream
towards a aggregation centre. Aggregation results in the formation of a multicellular
organism, mound. Within mound cells differentiate into prestalk and prespore cells and
sort out to form a tipped mound. Elongation of tip leads to the formation of a slug or
migrating pseudoplasmodium, which undergoes differentiation to form an early culminant
and finally the mature fruiting body Under some physiological conditions the slug stageand finally the mature fruiting body. Under some physiological conditions, the slug stage
is bypassed. (Redrawn from Chisholm & Firtel 2004)
9

10
from the animal–fungal lineage after the plant–animal split, but Dictyostelium seems to have
retained more of the diversity of the ancestral genome than have plants, animals or fungi
(Eichinger et al., 2005). Because of the small size and haploid state of the genome, it is
possible to study the cellular and molecular basis of these processes in great detail, by
mutational analysis of the genes involved. Thus, high-resolution molecular analysis in this
system may reveal control networks that are difficult to study in more complex systems and
foretell regulatory strategies used by higher organisms (Maeda et al., 2004; Soler-Lopez et
al., 2004; Thomason et al., 1998). In the following section, I will focus on the developmental
aspects of Dictyostelium and explain why Dictyostelium is a powerful model organism.
Vegetative Growth
Dictyostelium cells normally live as single cells in the soil leaf litter where they feed
on bacteria and divide by binary fission (Weijer, 2004). These cells can double their numbers
in four hours. The cell cycle of Dictyostelium cells is typical of lower eukaryotes. There is a
short mitosis (20 min), immediately followed by an S phase, which last up to 30 min. The S
phase is followed by a long and variable G2 phase, which in bacterially grown cells is
between 2 and 4 hr and in axenically grown cells is between 4 and 10 hr. There is no
detectable G1 phase (Araki et al., 1994; Gomer and Firtel, 1987). Cells can be synchronized
by arrest at a restriction point 1-2 hr before mitosis. During the initiation of starvation this
restriction point is activated. The cells continuously monitor both their own density and the
density of their food source by the secretion of a glycoprotein called prestarvation factor
(PSF). PSF binds to bacteria and thus allows the cells to titrate the amount of bacteria, so that
upon reaching high cell densities and a low food concentration the cells activate their
developmental program(Clarke et al., 1992; Clarke et al., 1988). Entry into development is
mediated by another factor called conditioned medium factor (CMF) which, together with

11
PSF, controls the activation of many genes required for aggregation (Clarke et al., 1992; Jain
et al., 1992). Additionally, Brook and Gomer (1996) have identified the counting factor (CF),
which allows cells to sense the number of cells during streaming and aggregate formation.
CF consists of multiple subunits: countin, CF45, and CF50 with different receptors and
different signal transduction pathways to help regulate group size (Brock et al., 2003).
Therefore, PSF, CMF, and CF act in diverse signalling pathways to establish the optimal
population size and timing of development.
Aggregation
A few hours after the initiation of starvation cells become sensitive to cAMP due to
the expression of the cell surface serpentine cAMP receptors. Four cAMP receptors (cAR1-
cAR4) are sequencially expressed thoroughout development (Parent and Devreotes, 1996).
cAR1 is expressed early in development. The mechanism of cAMP excitability involves two
different feedback loops: (1) a fast autocatalytic cAMP induced cAMP amplification, (2) A
slightly slower negative feedback loop (Devreotes, 1989; Martiel and Goldbeter, 1987). The
difference in excitation and adaptation is measured and leads to the activation of the
aggregation stage adenylyl cyclase; the enzyme that produces cAMP from ATP. Secreted
cAMP binds back to the receptor on the cell surface. This positive feedback loop leads to the
production of more cAMP. The cAMP amount is regulated by degrading (3’-5’ cAMP to the
inactive 5’AMP) continuously by an intracellular phosphodiesterase, regA, and a secreted
extracellular phosphodiesterase (ePDE) (Dormann et al., 2002a). Waves of cAMP move
outward through the lawn of cells with a periodicity of ~ 6 min (Parent and Devreotes, 1996),
and cAMP is only produced by more distal cells. As the wave is detected by cAR1, cells
become highly polarized in the direction of the chemoattractant gradient and move in that
direction for 1 min. The more inward cells have transiently adapted, and for a short period of

12
time (~ 5 min), cannot respond to the cAMP signal during which time they show random
motility and become unpolarized. This prevents these cells from responding to signals that
are distal to the centre of the aggregation territory. Cells therefore only move inward towards
the centre of the multicellular aggregate.
Similar chemotactic behaviour is observed in leukocytes. Dictyostelium cells are
responsive to cAMP, components found in bacterial extracts such as folic acid, platelet-
activating factor (PAF) and lysophosphatidic acid (LPA) (Jalink et al., 1993; Sordano C. et
al., 1993). Leukocytes respond to PAF and LPA, and in addition to, N-formylated peptides
(fMLPs), leukotrienes, complement factors and chemokines (van Es and Devreotes, 1999).
All of these chemoattractants in amoebae and leukocytes interact with specific seven
transmembrane domain surface receptors, which transduce signals by coupling to
heterotrimeric G-proteins (van Es and Devreotes, 1999). Therefore, despite their evolutionary
distance, both amoebae and immune cells sense chemoattactant by similar mechanisms.
Cell Streaming
Initially, the cells move towards the aggregation centre as individuals, but after 10-20
waves have passed they form bifurcating aggregation streams, in which the cells make head-
to-tail contacts via calcium-independent adhesion molecules, contact site A and side-to-side
contacts via a calcium-dependent contact molecule (DdCAD-1) (Gerisch, 1986). Stream
formation is dependent on the localization of aggregation-stage adenylyl cyclase (ACA) in the
rear of the aggregating cells, resulting in polarized cAMP secretion from the back of the cells
(Kriebel et al., 2003). cAMP wave propagation can be observed indirectly at the population
level, which can be seen as propagating optical density waves that are associated with the
periodic surges in cell movement of groups of cells in the direction of the cAMP signal.
Alternatively, the cAMP wave propagation can be observed at the individual cell level by

13
following the localized translocation of phosphatidylinositol 3,4,5-triphosphate (PIP3) at the
leading edge of the cell (Dormann et al., 2002b). As mentioned earlier, the number of cells in
aggregation streams appears to be controlled by the local concentration of a secreted
extracellular high molecular weight protein complex, counting factor, which control the
number of cells that stably migrate in an aggregation stream through modulation of movement
and adhesion (Tang et al., 2002).
Mound
Once the cells come into the aggregation centre they start to move on top of each other
and form a hemispherical structure known as the mound (Fig. 1.2). Mounds are characterized
by rotating waves of cAMP that direct the counter-rotational periodic movement of the cells
(Weijer, 2004). Cells start to differentiate into prespore and prestalk cells and these cells are
distributed randomly within the aggregate (Araki et al., 1997; Weening et al., 2003). In order
to go from a scattered pattern of the cell types to a highly organized anterior-posterior pattern,
not unlike the basic body plan of metazoans, the cells undergo a morphogenetic program that
involves cell sorting controlled by directed cell movements and differential cell adhesion
(Chisholm and Firtel, 2004; Weijer, 2004). The mechanistic details of Dictyostelium pattern
formation will be discussed in the following section.
Sorting is mediated by the differential abilities of the prestalk and prespore cells to
undergo chemotaxis towards cAMP (Weijer, 1999). Prestalk cells associate in a loose mass
near the base of the mound (Fig. 1.3A), which then moves upwards to form the apical region
of the mound (Clow et al., 2000). Further apical movement brings the cells to the top of the
mound where they form a tip. This tip functions as a signalling centre and produces cAMP
waves that pass through the mound (Dormann and Weijer, 2001; Siegert and Weijer, 1995).
In addition, the apical tip functions similarly to morphogenetic organizing centres in

Figure 1.3 Schematics of movement of prestalk cells during tipped mound formation,
slug migration and culmination. (A) Sorting of differentiating cells within the mound. g g ( ) g g
Prestalk cells (shown as blue dots) and prespore cells are initially distributed throughout
the mound and begin to sort. Prestalk cells move directionally towards the presumptive
apical tip of the mound, which functions as an embryonic organizer by serving as a centre
of cAMP secretion. Initially, prestalk cells coalesce and moves towards the apical tip.
Thi d ti d t d i t d i Fi 1 3A Th ti i di t d th hThis produces tipped aggregate as depicted in Fig. 1.3A. The sorting is mediated through
preferential chemotaxis of the prestalk cells, combined with differential cell adhesion
between the various cell populations. (B) Schematics showing the migration of a young
slug. It is characterized by lifting up and down of the slug tip. Cells in the tip rotate as
long as the tip is up in the air. (C) Schematics showing migration observed in older slugs.
These slugs reflects abortive attempts to culminate. Lifting off is initiated by the active
aggregation of anterior-like cells at the prestalk-prespore boundary which form a
stationary mass of cells. The pile forces the rest of the slug to lift off in the air. The pile
of cells is being left behind and gets translocated to the back of the slug. The slug falls
back onto the substratum the pile vanishes and the prestalk cells form a new pile at theback onto the substratum, the pile vanishes and the prestalk cells form a new pile at the
prestalk-prespore boundary. The cells in the prestalk O (pstO) region (refer 1.4A)
continue to rotate. (D) Schematics showing the formation of culmination. The first image
shows the formation of the pile, and the rotation of the cells in the posterior prestalk
(pstO) zone similar to that shown in B. The whole prespore mass shift on top of the pile.
The cells in the pile start to rotate. These cells will form the basal disc and the lower cup.
(Adapted from Dormann et al., 1996; Chisholm and Firtel 2004)
14

A
B
C
D
15

16
metazoans by organizing patterning and regulating cell-fate decisions. Extension of the tip
causes the organism to elongate which results in the formation of a standing finger. When
these finger-like structures collapse onto the substratum, they become the migrating slug.
Slug migration
Cell movement within the slug is also mediated by oscillatory waves of cAMP, which
are initiated from the anterior and propagate towards the posterior initially as radial waves
that pass through the prestalk domain (Dormann and Weijer, 2001). In the prespore domain,
they become planar waves through the activation of adenylyl cyclase by the same or a related
pathway that mediates cAMP relay throughout the lawn of cells during aggregation (Loomis,
1998; Mohanty et al., 2001). The anterior tip of the slug continues to function as a signalling
centre and organizer (Fig. 1.3B). Cells in the tip region often rotate perpendicularly to the
direction of slug migration, especially when it is lifted from the substrate. In the posterior
part of the slug, the cells move forward periodically and all cells move on average with the
speed of the whole slug. In the prestalk zone, cells rotate around the long axis slightly slanted
to the direction of slug migration (Fig. 1.3B,C). This rotational movement is especially
strong in the pstO zone (Fig. 1.4A) when the tip is lifted up from the substrate in the air. Due
to their twisted tracks, the speed of movement of the individual prestalk cells is greater than
the forward speed of the slug movement.
Modifications of slug tips such as removal of the tip or transplantation of a tip from
one slug onto the side of another lead to the formation new tips or the establishment of a
second organizing centre with appropriate prestalk and prespore proportion (Raper, 1940).
The role of extracellular signalling in these processes has been established through
experiments in which cAMP signalling has been repressed by drugs such as adenosine

A
Psp ALC PstB
PstO PstAB PstA
Prespore domain Prestalk domain
B
PstOUpper cup
Spore mass
B
Figure 1.4 Distribution of the prestalk-cell
types in slugs and fruiting body. The precursor
cell types include prespore cells (Psp) and
PstA
ALC
p
Lower cup
Stalk tube
cell types include prespore cells (Psp) and
prestalk cells (Pst), the latter ones being
subdivided into PstA, PstB, PstO, and PstAB
cells, plus anteriorlike cells (ALCs). These cell-
types differentiate at the mound-stage and sort
within the multicellular aggregate to generate a
defined spatial pattern. At the slug stage, the cell
types are organized along a well-defined
anteroposterior axis (A). The cells in the prestalk
region are represented as blocks of colour, as are
PstBPstO
Basal disc
g p ,
the prespore cells. The anterior-like cells
(ALCs) are shown individually, but the drawing
is not to scale. (B) Distribution of cell types in
fruiting body.
17

18
analogue IPA (2’,3’-o-isopropylidene adenosine), or by experiments in which a micropipette
is inserted into the slug and used as an oscillatory source of cAMP (Durston et al., 1979)
(Rietdorf et al., 1998). This triggers the recruitment of anterior-like cells (ALC) and prestalk
cells to the tip of the micropipette and the formation of a new centre (Dormann and Weijer,
2001). ALC cells are scattered within the prespore region (Fig. 1.4) and show many of the
properties of the prestalk cells (Devine and Loomis, 1985; Sternfeld and David, 1982). In
principle, cAMP wave propagation and chemotaxis in response to these waves is sufficient to
explain morphogenesis from single cells via aggregation, stream and mound formation to cell
sorting and slug formation. However, evidence suggests that morphogenesis is a complex
phenomenon. Since Umed and Inouye (2002) have reported that strains lacking the ACA can
still form slugs when they over-express the catalytic subunit of protein kinase A. This result
suggests that either there exists an ACA-independent mechanism to produce periodic cAMP
signals or that there exists different mechanisms that can control cell movement.
Culmination and fruiting body formation
Culmination is the final and most complex transformation that occurs during the
Dictyostelium developmental cycle (Fig. 1.2). On soil, environmental factors such as low
humidity, overhead light, and reduction of the local NH4+ concentration result in an arrest of
slug migration and in the initiation of culmination, the terminal differentiation of spores and
stalk cells, and formation of the mature fruiting body (Newell et al., 1969; Raper, 1940;
Schindler and Sussman, 1977). Dormann et al. (1996) have observed that culmination is
organized by two signalling centres: prestalk cells in the tip and the anterior-like cells in the
back of the slug (Fig. 1.3D). Culmination is initiated by a local aggregation of anterior-like
cells at the base of the slug at the prestalk-prespore boundary, where they form a stationary
mass of cells. During culmination, the majority of the cells follow the tip, moving over this

19
pile as a consequence are lifted up in the air. However, Dormann et al.(1996) reported that at
least some of these cells at the prespore-prestalk boundary are involved in the mechanics of
culmination. These cells are characterized by vigorous rotational cell movement. During this
process, the cells in the tip start to form a tube like structure from extracellular matrix
material including cellulose as a main component. This is the rudimentary structure of the
stalk.
It has been demonstrated that prestalk cells are motors for the culmination process.
Mutants in the cytoskeleton that disrupts the ability of prestalk cells to move properly result in
a failure of culmination (Chen et al., 1998; Noegel and Schleicher, 2000). During
culmination, some of the prestalk cells start to move into the stalk tube and move down until
they make contact with the substratum. They subsequently differentiate into mature highly
vacuolated stalk cells containing stiff cellulose walls. The stalk forms a mechanical structure
along which the other cells can move up. As cells moved up the stalk and into the stalk tube,
prestalk cells would undergo several distinct and dramatic cell shape changes during the
course of their travel (Grimson, 2000). As more cells enter the stalk tube, the stalk elongates
and eventually leads to the formation of fruiting body. The fruiting body consists of a sorus
surrounded by lower and upper cup cells, which are supported by a stalk tube with a basal
disc. The cells belonging to the basal centre will form the basal disc and the lower cup in the
fruiting body. The upper cup will be formed by the prestalk cells rotating most vigorously at
the prestalk-prespore boundary (Fig. 1.4B).
1.1.4 Mechanisms involved in the Dictyostelium pattern formation
Two major mechanisms, namely positional information based on morphogen gradient
and random differentiation followed by cell sorting out, have been proposed for Dictyostelium

20
pattern formation. The positional information model arose from the early embryological
studies that showed a patterning process depends on special “organizing” regions in the
embryo and led to the concept of morphogen gradients (Wolpert, 1996). Early evidence for
this model in Dictyostelium came from the transplantation experiments which showed the
presense of anterior-posterior morphogenetic gradients and the presence of an organizing
region, the tip, at the anterior (Raper 1940; Rubin and Robertson 1975). Pattern formation
based on the sorting out model is produced in two steps. First, different cell types are initially
specified from a precursor pool independent of their position to produce a salt and pepper
mixture. Second, the mixture of cell types is resolved into discrete tissues by the physical
movement and sorting out of the cells. Consequently, this mechanism does not involve
positional information. However, it can provide conditions such as sources and sinks for
signalling molecules for morphogen gradients to arise once the pattern has been formed (Kay
and Thompson, 2009).
The first challenge for the positional model arose when an experiment was performed
with cells which were grown in medium with or without glucose, and then mixed for
development. It was found that cells grown without glucose preferentially became stalk cells
and that these cells sorted out from their glucose-rich cells during the mound stage of
development (Leach et al., 1973; Tasaka and Takeuchi, 1981). These differences are biases,
not commitments, because when cells from a number of growth conditions are compared,
cells found to be “stalky” in one mixture, are “sporey” in another (Kay and Thompson, 2009).
In fact, cells start to differentiate into prespore and prestalk cells during aggregation, on the
basis of physiological biases like cell-cycle position at the time of starvation (Araki et al.,
1997; Weening et al., 2003). Cells in the periphery (starved at S phase or early G2 phase)
differentiate mostly into prestalk cells, while those in the inner core (starved at mid- or late

21
G2 phase) differentiate into prespore cells (Araki et al., 1994; Gomer and Firtel, 1987; Weijer
et al., 1984; Zimmermann and Weijer, 1993). These observations reveal that sorting must
have occurred at some stage in development, but can be explained in two different ways. On
the one hand, the cells sort out before being specified, such that one type is in the right place
to subsequently receive a positional signal directing it to cell type differentiation. On the
other hand, cells might differentiate first as intermingled prestalk and prespore cells, and later
sort according to their differentiated state. Tasaka and Tekeuchi (1981) favoured the second
possibility since they found that sorting occurred simultaneously with prestalk and prespore
cell differentiation, but not before it as a postional model predicted. Therefore, the positional
model proposes that prestalk and prespore cells should differentiate in distinct places, in
response to an underlying morphogen gradient, while a sorting model predicts that these cells
should be intermingled.
It is clear that prestalk and prespore cells first differentiate at the mound stage of
development before the slug formation, however the initial sites of prestalk and prespore cell
differentiation were unknown. Initial studies tended to support a positional model that was
based on antibodies against prespore and prestalk vesicles. In these studies, prespore cells
were first detectable in the upper part of the mound and prestalk cells were localized to the
basal cells (Krefft et al., 1984; Williams et al., 1989). In addition, pstA cells have been
described to differentiate at the mound periphery (Early et al., 1995). These observations
were affected by two problems. First, markers based on gene expression always take some
time to develop once the inductive event has occurred. Second, the cells in the mound are in
constant, rapid rotational movement (Kay and Thompson, 2009). Consequently, the apparent
separation of prestalk and prespore cells seen in the earlier experiments described above
probably reflect a sorting intermediate rather than positional differentiation. For instance,

22
when more sensitive markers, such as lacZ reporter genes, were developed, prestalk cells
were found scattered throughout the mound and both cell types are even detectable in the
streams of cells entering the mound (Early et al., 1995; Ozaki et al., 1993). Time lapse
studies have shown that prestalk and prespore cells arose in a spatially random manner
throughout the aggregates and clearly independent of any positional information (Nicol et al.,
1999). Recently, paralysis of cells using the actin-binding drug latrunculin (which still allows
efficient differentiation) shows that all prestalk and prespore cell types are scattered
throughout all parts of the aggregate (Thompson et al., 2004).
1.1.5 Signal transduction and regulation of cell type differentiation in Dictyostelium
Studies of the action of differentiation inducing factor (DIF-1) and cAMP provide
some molecular clues about how fates of cells are regulated. cAMP and DIF-1 are known to
differentially regulate cell type differentiation (Fig. 1.5). Prestalk differentiation is facilitated
by DIF-1, which is a small lipid-soluble chlorinated hexanone that also inhibits prespore
differentiation (Berks and Kay, 1990; Thompson and Kay, 2000; Williams et al., 1987).
Interestingly, prespore cells fail to respond to the DIF-1 they produce and at present it remains
unknown whether there is a cell type specific expression of components of the DIF-1 response
machinery. DIF-1 stimulates tyrosine phosphorylation of the signal transducer and activator
of transcription c (STATc) transcription factor, which is required for the differentiation of the
pstO cells. Within this pstO population, DIF-1, through STATc, inhibits the expression of the
pstA pathway, which provides a molecular mechanism that allows the spatially restricted
differentiation of pstA and pstO cells (Fukuzawa et al., 2003). Recently, a new DIF
responsive transcription factor, GataC was identified and shown to be only required for pstB
cell patterning (Keller and Thomson 2008).

OH OCl
A B
cAMPUncommitted cell Prespore cell
a b c
OH
Cl
H3COCl
C
DIF-1
Prestalk cell
DIFase
cAMP
cAR1
DIF-1
STATc
?
PstA
DIF-1
?
STATa
(CudA)tip
PstAPstO
Figure 1.5 Regulation of the prestalk pathway. (A) Structure of differentiation
inducing factor 1 (DIF-1, a chlorinated hexaphenone). (B) Cell-type-specific metabolism
of DIF-1. DIF-1 concentration is regulated by a negative-feedback loop, whereby it
rapidly induces the production of DIF-1 dechlorinase (DIFase), which catalyses DIF-1
inactivation. DIF-1 acts antagonistically to cAMP by repressing prespore differentiation
and directing a proportion of the cell population to differentiate as prestalk cells. (C)
Prestalk differentiation is controlled by two morphogens, DIF-1 and cyclic AMP, which
differentially regulate the prestalk domains. Among the main downstream regulators of
this pathway are the signal transducer and activator of transcription (STAT) factors,
STATa and STATc (a) DIF 1 induces pstO differentiation through an undefined pathwaySTATa and STATc. (a) DIF-1 induces pstO differentiation through an undefined pathway
that requires a bZIP/bRLZ (basic-leucine zipper/basic-region leucine zipper) transcription
factor (DimA). (b) DIF-1 also inhibits the differentiation of pstA cells by inducing the
STATc activity. The activator of pstA differentiation is unknown. (c) The anterior tip of
the slug is induced through STATa using the nuclear factor CudA. STATa is activated by
cAMP through the cAMP receptor cAR1 in a G-protein-independent manner. (Redrawn
from Chisholm & Firtel 2004; Williams 2006)
23

24
The extreme anterior tip of the prestalk domain is induced through cAMP-mediated
tyrosine phosphorylation of STATa and the activation of the nuclear factor CudA (Fukuzawa
and Williams, 2000). CudA, protein that regulates the slug/fruiting-body switch, is localized
in the prespore zone and in a cone of cells at the extreme anterior of the slug (Fukuzawa and
Williams, 2000). Two pieces of experimental evidence suggest that extreme anterior
population is made up of tip cells. First, the cudA-null mutant remains as a slug under
conditions in which wild-type slugs culminate; ‘slugger’ mutant. Second, the slugger
phenotype is reversed when CudA is expressed under the control of the ecmA promoter
(Fukuzawa and Williams, 2000). The cone of CudA expression overlaps with the anterior-
most pstA cells, suggesting sequential differentiation of a subset of the pstA cells into tip cells
(Williams, 2006).
The prespore pathway is activated by extracellular cAMP through cAR3 (encoded by
carC), a member of the seven-transmembrane-receptor family that controls chemotaxis (Plyte
et al., 1999). cAR3, through a Wnt-like pathway (Fig. 1.6A), leads to the activation of
glycogen-synthase kinase-3 (GSK3, encoded by gskA) which in turn is activated by the
tyrosine kinases zipper sterile motif kinase (ZAK ½, encoded by zakA). Subsequently,
nuclear localization of β-catenin homolog, aardvark (Aar) induces transcriptional changes
(Coates et al., 2002; Kim et al., 2002; Kim and Kimmel, 2000; Plyte et al., 1999). This
pathway shows strong similarities to the metazoan Wnt signalling pathway (Kim and
Kimmel, 2000; Moon et al., 2002) (Fig. 1.6). cAMP also functions through a prestalk-cell-
enriched cAMP receptor cAR4 (encoded by carD) to inhibit prestalk differentiation by
dephosphorylating ZAK1/2 using a yet-to-be identified protein tyrosine phosphatase (PTP)
(Coates et al., 2002; Moon et al., 2002).

Figure 1.6 A comparison of GSK3 signalling pathways and regulation of
developmental fate choice in D. discoideum, mammals and C. elegans. (A) In
Dictyostelium discoideum, the cyclic AMP receptors (cAR) promote (cAR3) or inhibit
(cAR4) the activation of glycogen-synthase kinase (GSK)3, which is controlled by
phosphorylation by the zipper sterile alpha motif kinases (ZAK)1 and ZAK2. cAR3
stimulates ZAK1/2, cAR4 activates a protein tyrosine phosphatase (PTPase) that
deactivates GSK3. The cAR3–ZAK–GSK3 pathway regulates differentiation of
prespore/spore fates and represses prestalk/stalk differentiation. The prespore pathway
requires GSK3 phosphorylation of the β-catenin homologue Aardvark (Aar). Inhibition
of GSK3 activity by cAR4 activates prestalk/stalk pathways and represses prespore/spore
differentiation. The cARs are closely related to mammalian Frizzled receptors (Fz),
which bind Wnt (B) Mammalian cells show GSK3 mediated activating and inhibitorywhich bind Wnt. (B) Mammalian cells show GSK3-mediated activating and inhibitory
pathways through Wnt–Fz signalling. In the absence of Wnt, β-catenin is de-stabilized by
phosphorylation by GSK3 in complex with Axin. Disheveled (Dvl) functionally inhibits
the activity of the Axin–GSK3 complex, stabilizing β-catenin, which accumulates and
facilitates interaction with transcription factors that activate genes required for
development and tumorigenesis. Adenomatous polyposis coli (APC), like Axin, functions
as a tumour suppressor, whereas the inhibitor of GSK3 FRAT and β-catenin are
oncogenic. A separate pathway that is activated by a different Wnt class antagonizes
‘canonical’ Wnt signalling. (C) The Caenorhabditis elegans Wnt pathway closely
resembles the ‘canonical’ pathway of other metazoa and regulates mesoderm/endodermresembles the canonical pathway of other metazoa and regulates mesoderm/endoderm
choice. As with D. discoideum cAR3, Wnt–Fz signalling activates GSK3. (Redrawn
from Chisholm & Firtel 2004)
25

D di idA
cAMP
cAR4 cAR3
D. discoideumA
GSK3
PTPase ZAK1/2
AAar
Prestalkfate
Presporefate
MammalsB C elegansCMammals
Wnt3 Wnt5a
Fz Fz
B
Mom-2 (Wnt)
MOM-5 (Fz)
C. elegansC
Axin
DvI
β t i
FRAT/GBPGSK3
APC
WRM 1 (β t i )
?
GSK3
β-catenin
TumoursuppressionTumorigenesis
Alternative fates
WRM-1 (β-catenin)
Mesoderm fate
Endoderm fate
26

27
cAR3 is preferentially expressed in prespore cells whereas cAR4 expression is more
specific to prestalk cells (Ginsburg and Kimmel, 1997). Developmental-fate changes in cells
that lack one or the other signalling pathways are reversible however different cell fate
changes are significantly reduced (Kim et al., 2002; Kim and Kimmel, 2000). Both cAR3
and cAR4 can interact with heterotrimeric G proteins and activate G-protein-dependent
pathways but the ZAK1/2 pathways are G-protein-independent. Thus, interactions of the
DIF-1 and cAMP signalling pathways control the final proportioning of the cell type during
development and this proportioning is very plastic. For instance, in classic experiments, after
removal of a portion of the slug, the proportion of prestalk and prespore cells remained
constant (Raper, 1940). Excision of the anterior of the slug led to the dedifferentiation of
some of the prespore cells into prestalk cells, which resulted in a properly proportioned
organism. Similar to metazoans, PKA has an essential role in controlling cell-fate decisions.
PKA is controlled by the intracellular levels of cAMP, which in turn, are controlled by the
rate of cAMP synthesis, the regulation of adenylyl cyclases, and the rate of cAMP
degradation by the cAMP-specific phosphodiesterase RegA (Harwood et al., 1992; Loomis,
1998).
1.1.6 Theories proposed for cell sorting in multicellular development
Ever since the early studies by Wilson (1907), Holtfreter (1943,1944), Moscona
(1952), and others who demonstrated tissue- and species-specific sorting out of embryonic
cells, it has been believed that specialized adhesive properties of cells play a key role in
morphogenesis. There is no doubt that cell adhesion is important in the dramatic cell
rearrangements that take place during gastrulation (Gerhart and Keller, 1986; Keller et al.,
1985), neurulation, neurogenesis (McClay and Ettensohn, 1987), and organ formation (Poole

28
and Steinberg, 1982; Zackson and Steinberg, 1986). However, little is known about the
extent to which cell adhesion proteins regulate or direct specific morphogenetic events during
development.
1.1.6.1 Differential Adhesion Hypothesis (DAH)
Steinberg proposed that the forces organizing the cells are closely analogous to those
organizing the molecules of immiscible fluids and put forward the differential adhesion
hypothesis (DAH) /thermodynamic hypothesis. The DAH proposes that the liquid-like tissue-
spreading and cell segregation phenomena of development arise from tissue surface tensions
that in turn arise from differences in intercellular adhesiveness (Fotya and Steinberg, 2005).
The physical explanation for this hypothesis is that a population of motile, mutually adhesive
cells will spontaneously tend to replace weaker intercellular adhesions with stronger ones
until it approaches the configuration in which adhesive bonding is maximized (Fotya and
Steinberg, 2005). The DAH makes no assumption about the specificity or selectivity of
intercellular adhesions, but it provides criteria by which the relative strengths of adhesion at
various kinds of cell-cell interfaces can be ranked within certain limits (Fotya and Steinberg,
2005). These criteria would determine whether two cell populations intermix or segregate; if
the later, which population would envelop the other and to what extent. It has been widely
believed that “cells expressing different cadherins sort out from each other by adhering only
to those expressing the same cadherin, the specificity of homophilic binding, a fundamental
mechanism by which cadherins influence the organization of various cell types into tissue”
(Fotya and Steinberg, 2005). However, Steinberg & others have demonstrated that sorting out
can result from mere quantitative differences in the expression level of a single cadherin type

29
(Fotya and Steinberg, 2005). Changing the adhesive relationship between cell types can
influence their relative positions in tissue.
Recent evaluation of the DAH has shown that tissue surface tension increases linearly
with the expression level of adhesion molecules such as cadherin (Lecuit and Lenne, 2007).
Surface tension can be defined as the free-energy change that occurs when the surface of a
medium is increased by a unit area. The tissue surface tension is the apparent surface tension
of a tissue, caused by adhesion between cells (Lecuit and Lenne, 2007). The binding
specificity between cadherin molecules is not sufficient to fully account for cell sorting.
Since the adhesion specificity and the strength of cell association are not fully dependent on
the extracellular interactions of cadherins, but also dependent on their dynamic interactions
with cortical actin and on actin organization (Lecuit and Lenne, 2007). For two contacting
cells, the increase of cortical tension due to the formation of a contractile acto-myosin
network at the zone of contact reduces the contact surface. Therefore, the dynamics of cell-
cell contacts requires an interplay between adhesion and cortical tension (Lecuit and Lenne,
2007).
1.1.6.2 Differential Surface Contraction (DSC)
Although, differential expression of the cell adhesion molecule cadherin is sufficient
to drive cell sorting in experimental systems involving isolated cells, to some the DAH was
only one of the possible mechanisms (Brodland, 2004). As a strong critique of DAH, Harris
proposed a differential surface contraction (DSC) model in which cortical tension (the force
generated within cells parallel to their surface) rather than adhesion between cells per se,
could drive cell sorting (Harris, 1976). In other words, adhesion is one of the forces
generated within cells. Cortical tension can be defined as the apparent cell surface tension

30
due to the contractile microfilament of the cell cortex and their interaction with the membrane
(Lecuit and Lenne, 2007). The cortical tension can be measured by observing cell
deformation within a conical micropipette or by pulling membrane nanotubes, also called
tethers(Perret et al., 2004; Sheetz, 2001).
Recently, Krieg et al. (2008) tested these two models by directly measuring the
adhesiveness and cortical tension of cells from the three germ layers of zebrafish embryos
using an atomic force microscope equipped with a tiny probe mounted and calibrated so that
its bending by an object at its tip can be measured and the corresponding bending force
determined. They measured cell adhesion by attaching one cell to the end of the microscope
probe and a second cell to a fixed substrate below, bringing the two cells together and
monitoring the force required to pull them apart. They also determined cortical tension by
measuring the force needed for a hard bead attached to the end of the probe to deform the
surface of a cell attached to the surface below. Measurements for cells from the ectoderm
(Ec), mesoderm (M) and endoderm (En) showed that homotypic adhesion was stronger in
mesoderm than in ectoderm, whereas endoderm values were in between (AdM > AdEn >
AdEc). Adhesion was calcium-dependent and correlated with cadherin expression at the
surface. The order of cortical tension values was different: CtEc > CtM > CtEn. In pairwise
sorting assays, ectoderm cells were always in the middle, opposite to what would be predicted
by DAH, under which their low homotypic adhesion would place them on the outside. The
central position of ectoderm cells within the aggregates correlated instead with higher cortical
tension. Mesoderm cells were surrounded by endoderm, extending the cortical tension
correlation: CtEc > CtM > CtEn. Authors (Green, 2008) have suggested that this observation
would seem to be a support DSC (Brodland, 2004), at least for these cell types, although it
fails to reproduce the in vivo configuration (ectoderm outside and endoderm innermost).

31
However, mechanisms involving differential adhesion and cortical tension are not mutually
exclusive. Differential adhesion forces may contribute to the cortical tension generated
between two cell surfaces.
1.1.6.3 Mechanistic explanations for cell sorting during morphogenesis (DAH vs DSC)
The work of Krieg et al. (2008) leads biologists to reconsider the role of differential
adhesion forces in cell sorting. For instance, the cortical actomyosin cytoskeleton becomes
more significant. Krieg et al. (2008) showed that disruption of this network, using
blebbistatin (an inhibitor of myosin II activity) and dominant-negative Rho kinase, blocks cell
sorting. This suggests that sorting is more similar to active migration, in which changes in
cell shape are crucial, whereas according to the DAH, cells are, effectively, structureless units
(Green, 2008). Additionally, integrins, adhesion molecules associated with migration, can be
crucial for cell sorting (Pearl et al., 2005). There are also instances when β-catenin regulation
of cadherin, the adhesion molecule traditionally thought to account for cell-sorting, might
play a secondary role (Reintsch et al., 2005). Second, the DSC model predicts that
contractility must be different and higher at cell-substratum interfaces than at internal
interfaces between cells (Harris, 1976). In other words, cortical tension should be localized
by cell polarization. For instance, in polarized cells actin-cytoskeleton is differentially
organizied in apical and basal surfaces therefore cortical tension might be different in these
surfaces. Krieg et al. (2008) addressed this prediction in two ways. First, using computer
models of cell sorting with and without localization of cortical tension (cortical tension can be
localized in polarized cells) they found that sorting operates only when the cortical tension is
localized. Second, by examining actin in live cell aggregates, they showed that it was
enriched at the cell-medium interface.

32
Townes and Holfreter (1955) had already hinted at the importance of active cell-
surface contraction for self-sorting when they noticed that neurectodermal cells, whether as
single cells or organized as sheets, were surrounded by endodermal cells. Neurectodermal
sheets penetrated an endodermal mass by infolding or invagination, recapitulating the rolling-
up neurulation movements that make a tubular spinal cord. They proposed that the same
mechanism may drive cell sorting. Actomyosin-driven apical contraction is now recognized
as the main mechanism of epithelial folding in the neural plate, and support for actomyosin-
dependent DSC provides a mechanistic link between cell-sorting and epithelial folding.
Despite the support Krieg et al. (2008) provide for the DSC hypothesis, how can one
explain the observation that ectoderm or the mesoderm are inside the aggregates (CtEc > CtM
> CtEn)? To address this issue, the authors assayed cell sorting ‘in vivo’ using transplantation
experiments. They simulated progenitor cell sorting in the presense of the yolk cell and
enveloping cell layer cells. They found that strong interactions of germ-layer cells with the
yolk invert the inside-out cell sorting seen in vitro, thereby producing the endo-in/ecto-out
arrangement. Recently, Ninomiya and Winklbauer (2008) reported that tissue elongation in
mesodermal explants and cell aggregates is enhanced by a wrapping of epithelium. Epithelial
wrapping was performed by coating explants with ectodermal epithelial layer that was
manually peeled from Xenopus embryos. They showed that rod-shaped ectodermal
aggregates rapidly became spherical to reduce surface area. In contrast, isolated epithelial
layer folded irregularly, with the non-adhesive apical side facing outward. When aggregates
were wrapped in epithelium, an elongated shape was maintained by a reduction in surface
tension. These studies suggest that the epithelium facilitates tissue elongation by reducing the
tensions, intracellular or intercellular tensions that drive cell-sorting and aggregate rounding.
Interestingly, Green (2008) pointed out that other challenges should be addressed before

33
applying these findings in vivo: (1) the quality and strength of adhesions change with contact
time which is barely taken into account in most of the adhesion assays; (2) cortical tension is
only one of several factors determining cell deformability; (3) mesodermal cells in zebrafish
move as a loose population; (4) the authors examined the actin distribution at late-
differentiating stages rather than during sorting; (5) that adhesion of sheets of cells may be
different from that of individual cells.
In a further instance of how an epithelium helps cells sort, Ninomiya and Winklbauer
(2008) prepared aggregates, in which untreated ectodermal cells were mixed with similar
cells, and in which they expressed M-PAPC, a paraxial protocadherin derivative that reduces
cadherin-dependent adhesion since its cytoplasmic tail has been deleted (Chen and Gumbiner,
2006). As expected, M-PAPC-expressing cells, with their reduced cohesion, sorted to the
outside. Unexpectedly, wrapping with normal epithelial ectoderm sent M-PAPC-expressing
cells to the inside, whereas wrapping with M-PAPC-expressing ectoderm kept M-PAPC-
expressing cells on the outside. In short, cells with the same M-PAPC stick together,
suggesting that more complex cell interactions may influence the result of these experiments
(Davidson, 2008). These studies represent a new phase in the analysis of morphogenesis, in
which high-resolution force measurements and molecular analysis, in combination with more
physiological and multi-component models, will eventually leads to a better mechanistic
understanding of morphogenesis.
1.1.6.4 Chemotaxis and differential adhesion combined as a model in Dictyostelium
It has been proposed that differential cell motility based on chemotaxis and cell
adhesion play a role in cell sorting (Jiang et al., 1998). In chemotaxis, a diffusible chemical,
such as cAMP serves as a signal that instructs cells to move along the local chemical gradient

34
toward higher or lower chemical concentrations. During aggregation some cells
spontaneously emit cAMP, initiating an excitation wave that propagates outward as a
concentric ring or a spiral wave, as the signal is relayed by the surrounding cells (Caterina and
Devreotes, 1991). Individual cells respond to a temporal and spatial increase of cAMP and
start pulsatile chemotactic movement in the direction of higher cAMP concentration (Varnum
et al., 1986; Wessels et al., 1992). Unlike differential adhesion, chemotactic cell motion is
highly organized over a length scale significantly larger than the size of a single cell.
Intercellular adhesion only passively keeps cells together while diffusible signals regulate
morphogenesis. Alternatively, adhesive energy differences might drive cell motion, while
diffusible chemical gradients merely enhance the process or might even be absent. In fact,
live cell imaging of cells suggests that differential adhesion may be the dominant mode of
patterning (Nicol et al., 1999). Consistent with idea, dissociated prestalk and prespore cells
are differentially adhesive (Lam et al., 1981) and prestalk cells surround the mass of prespore
cells (Steinberg and Takeichi, 1994), an example of differential adhesion predicted by
Steinberg. In addition, a number of cell adhesion genes and regulators of cell adhesion have
been cloned and several mutants show sorting defects (Dynes et al., 1994; Parkinson et al.,
2009; Wong et al., 2002). Nevertheless, proper pattern formation probably requires the
collaboration of both mechanisms.
Based on Dictyostelium mathematical simulation studies, it has been shown that both
chemotaxis and differential cell adhesion play a role in pattern formation (Jiang et al., 1998).
In the mound stage, if differential adhesion alone regulated cell sorting, pre-stalk cells would
come to the surface of the mound but no tip would form. In other words, differential adhesion
alone cannot explain the formation of a sorted tip. Second, chemotaxis of cells to some
diffusible chemical radiated from the mound centre can result in tip formation. Since the tip

35
consists of both pre-stalk and pre-spore cells, sorting cannot be accomplished by chemotaxis
alone. Third, only under the regulation of both mechanisms can the cells move to form a tip
consisting of pre-stalk cells only. Therefore, there is very strong evidence for an essential
role of chemotaxis during all stages of development of the social amoebae. There is
increasing evidence in vertebrates supporting an important role for chemotactic movement in
response to growth factors of the FGF, PDGF and VEGF families during gastrulation
(Dormann and Weijer, 2006). A major challenge will be to investigate the mechanisms
underlying signal detection, cell polarization and movement during morphogenesis.

36
1.2 Regulation of Dictyostelium adhesion molecules during development
Adhesion is important in regulating morphogenesis and early studies in Dictyostelium
provided some of the first evidence for what is now recognized as an essential mechanism of
tissue morphogenesis in all organisms. Dictyostelium cells acquire the ability to sort out
according to cell type in mixed aggregates which suggests possible changes in cell adhesion
occurred with the onset of aggregation (Gerisch, 1986: Gerisch, 1961). There are at least two
separate mechanisms of cell-cell adhesion systems operating in Dictyostelium. One appears at
the aggregation stage and is responsible for the EDTA-resistant adhesion sites (contact site A)
and the other is responsible for the EDTA-sensitive adhesion sites in both growth-phase and
aggregation-stage cells (contact site B) (Beug et al., 1970; Beug et al., 1973). The EDTA-
sensitive adhesion sites are mediated by DdCAD-1 and they will be discussed in detail in the
next section. A list of Dictyostelium adhesion molecules is shown in Table 1.1. The EDTA-
resistant adhesion sites of aggregating cells are mediated by an integral membrane
glycoprotein of apparent molecular weight 80,000, known as gp80 or csA glycoprotein
(Bertholdt et al., 1985; Müller and Gerisch, 1978; Siu et al., 1985; Springer and Barondes,
1985). Siu and coworkers (Siu et al., 1986) have reported that polystyrene beads conjugated
with gp80 bind specifically to aggregation-stage cells. Their work also provided the first
evidence that the binding of gp80 is mediated by homophilic interaction. The site responsible
for gp80-gp80 binding has not yet been determined.
Another EDTA-resistant cell adhesion molecule gp150/LagC is expressed at the
aggregation stage of development. The lagC gene codes for a protein of 98 kDa, which
contains an N-terminal signal peptide, a single transmembrane domain and a short
cytoplasmic sequence (Dynes et al., 1994; Wang et al., 2000). gp150 does not show

37
Table 1.1 List of Dictyostelium adhesion molecules.
Gene name
Protein name and
size
mRNA Expression
Pattern (bacterially grown
NC4 cells)
Knockout phenotype
Characteristics & Functions References
cadA DdCAD-1 23.9 kDa
0 hr to 18 hr peak at 8 hr
decreased sporulation, delayed culmination, increased prestalk cell differentiation, multiple tips
calcium-dependent cell adhesion molecule-1 homophilic adhesion
Wong et al., 2002
csA gp80 53.7 kDa
4 hr to 20 hr
Culmination delayed on soil plates
contact site A protein cell adhesion molecule heterophilic adhesion green beard gene
Ponte et al., 1998; Queller et al., 2003
lagC gp150/lagC 97.4 kDa
8 hr to 24 hr
abolished sporulation development arrests at loose aggregate stage in the presence of EGTA
signal protein cell adhesion molecule IPT/TIG domain-containing protein Cell-cell recognition, heterophilic adhesion, cell migration, plays a role in kin discrimination and is highly polymorphic between different isolates of D. discoideum
Benabentos et al., 2009; Dynes et al., 1994; Kibler et al., 2003
lagB1 lagB 100.3 kDa
8 hr to 24 hr
Decreased sporulation, delayed sporulation and developmental arrest at loose aggregate stage
IPT/TIG domain found in cell surface receptors Cell-cell recognition, cell migration, plays a role in kin discrimination and is highly polymorphic between different isolates of D. discoideum
Benabentos et al., 2009

38
significant sequence similarities with known adhesion receptors but contains two IPT/TIG
domains. gp150 mediates cell–cell adhesion via heterophilic interactions (Gao et al., 1992;
Wang et al., 2000) and it has been implicated in the sorting out process between prespore
cells and prestalk cells (Siu et al., 1983). Disruption of the lagC gene results in the failure of
development beyond the loose mound stage (Dynes et al., 1994). Extracellular matrix
components are not synthesized and cell differentiation is arrested. gp150 is involved in
signaling events that regulate cell-type differentiation (Dynes et al., 1994; Sukumaran et al.,
1998). The lagC-null phenotype can be rescued by over-expressing the G-box binding factor
GBF, a transcription factor known to regulate the transcription of post-aggregation stage
genes (Sukumaran et al., 1998). Further analysis of the lagC and gbf knockout mutants
suggests a role for gp150 in the establishment of a signaling center in mound morphogenesis
(Sukumaran et al., 1998). Three-dimensional time-lapse microscopy reveals that mutant cells
exhibit random motions as opposed to the organized and rotational motion seen in mounds of
parental cells. Multiple wave centers are formed due to defects in cAMP signaling, resulting
in aberrant cell movements. Recent studies show the involvement of the comC and lagD
genes in the lagC signaling pathway (Kibler et al., 2003). ComC (COMmunication mutant) is
an EGF-like domain containing protein with 14 EGF domains. comC-null mutant fails to
develop beyond the loose aggregate stage with abberent streaming observed during early
stages of development. lagD (Loose AGgregate D) is also a IPT/TIG domain containing
protein with three IPT sequences similar to the mammalian plexin protein (Coates and
Harwood, 2001). lagD-null cells failed to develop beyond loose aggregation stage and shows
abberent cAMP signaling. The cells of comC, lagC and lagD-null mutants fail to sporulate in
pure populations or in chimeras with each other, but sporulate when codeveloped with wild-
type cells. Transcriptional and functional evidence indicate that comC inhibits lagC

39
expression, while lagC and lagD are mutually inductive, with lagC being the terminal node of
this signaling network (Kibler et al., 2003).
1.2.1 DdCAD-1
1.2.1.1 Gene structure and regulation of DdCAD-1 expression
DdCAD-1 is encoded by the cadA gene, which was isolated using a cDNA derived
from a λgt11 expression library (Wong et al., 1996). A genomic 3.8 kb EcoRI fragment
containing 2.5 kb of 5' flanking DNA and the entire coding region was isolated (Wong et al.,
2002). The cadA coding region contains two short introns (Fig. 1.7A), which share consensus
intron-exon boundary sequences with other Dictyostelium genes. DdCAD-1 belongs to a
group of early developmentally regulated proteins, which are synthesized soon after the
initiation of development (Knecht et al., 1987). DdCAD-1 expression is stimulated by the
prestarvation factor, PSF, which signals nutrient depletion during vegetative growth in axenic
cultures (Rathi et al., 1991; Yang et al., 1997). In bacterially grown cells, DdCAD-1 displays
a unique temporal pattern of expression, which combines the characteristics of early genes
and aggregation stage genes (Yang et al., 1997). Transcription of the cadA gene is initiated
soon after the onset of development and its expression is regulated by countin, a component
of the large protein complex known as “counting factor,” which is a part of a negative
feedback loop that regulates the expression of DdCAD-1 and gp80 (Tang et al., 2001). Later
at the cell streaming stage, DdCAD-1 expression is stimulated by exogenous cAMP pulses
(Desbarats et al., 1992; Yang et al., 1997). Deletion analysis has led to the identification of
an 80 bp sequence between -359 and -280, which contains the major PSF and cAMP response
activity (Fig. 1.7B) (Sriskanthadevan et al., 2007; Yang et al., 1997).

A
EcoRI XbaI HincII HaeIII MboI EcoRI
ATG TAAintron 1 intron 2
200 bp
B
GTAAGTGGGGTGTGAGATTTAAATCATTG(-359)
(-338)
b 2
B
GGCTATGTTGGGGTTGAAAAAAACGGTTT
TTTTGGATTATTTTCACACAAG
(-286)
(-280)
box 2
box 3
box 4
Figure 1.7 Structure of the cadA gene. (A) Schematic drawing of the 3.8 kb genomic
DNA fragment containing the cadA gene. The three exons are represented by black
rectangles. Several unique restriction sites are also shown. (B) Nucleotide sequence of
the cadA promoter region, showing G-rich elements (boxes 2, 3, 4) and the TTG motifs
(boldface type), which confer cAMP and PSF responsiveness, respectively.
40

41
DdCAD-1 is present primarily as a soluble protein in the cytosol and < 30% of the
molecules are associated with the cell surface at the aggregation stage (Sesaki and Siu, 1996).
Furthermore, DdCAD-1 is present in cell-cell contacts only during the early stages of cell
streaming, and it disappears from the contact regions at later stages. As the Ca2+/Mg2+-
independent contact site A protein gp80 moves into the cell-cell contact region, DdCAD-1
appear to redistributes and disappears from the contact zone (Sesaki and Siu, 1996). High
levels of DdCAD-1 remain in the cytoplasm throughout development. These changes in
DdCAD-1 distribution suggest that, in addition to cell-cell adhesion, DdCAD-1 may have
other roles during Dictyostelium development.
1.2.1.2 Structural characteristics of DdCAD-1
The primary structure of DdCAD-1 deduced from the cDNA contains 213 amino
acids, with a calculated molecular mass of 23,924 Da. The NMR-derived structure of
DdCAD-1 shows a β-strand rich protein with two distinct domains linked by a flexible hinge
region (Lin et al., 2004). N-terminal sequence shows moderate sequence similarity to
spherulin 3a (~23%) of Physarum polycephalum (Clout et al., 2001) and protein S, a Ca2+-
binding protein associated with the spore coat of Myxococcus xanthus (~17 %) (Teintze et al.,
1988). These proteins belong to the β/γ crystallin family of proteins which shows a typical
Greek key topology (Bagby et al., 1994). The overall topology of the C-terminal domain is
consistent with an immunoglobulin-like fold. The C-terminal domain of DdCAD-1 and the
extracellular domains of cadherins belong to the same fold family although their β-strand
connectivity is not completely identical.
1.2.1.3 Multiple roles of DdCAD-1 during Dictyostelium development
Ca2+-dependent cell-cell adhesion

42
Genetic knockout of the cadA gene results in loss of Ca2+-dependent cell-cell adhesion
(Wong et al., 2002). Ectopic expression of DdCAD-1 in the cadA-null strain restores the
Ca2+- dependent contact sites, confirming that DdCAD-1 is responsible for the Ca2+-
dependent adhesion sites (Wong et al., 2002). As the cells enter the aggregation stage, high
levels of DdCAD-1 partition to the cell periphery and become associated with F-actin-
containing membrane protrusions, such as filopodia and lamellipodia that emanate from the
leading edge of migrating cells (Sesaki and Siu, 1996). DdCAD-1 is enriched at initial
filopodia-mediated cell–cell contacts, while more stable contacts are formed with the
subsequent entry of gp80. DdCAD-1 seems to redistributes away from cell-cell contact
regions as gp80 moves into these regions (Sesaki and Siu, 1996). The redistribution of
DdCAD-1 may reflect the dynamic nature of Dictyostelium morphogenesis, which requires
weaker interactions that permit the constant “breaking” and “re-making” of contacts as the
cells migrate to form large aggregates. The contrasting distribution patterns of DdCAD-1 and
gp80 are especially apparent in multicellular streams and early aggregates, where DdCAD-1
is absent in the interior region but present at high levels at the periphery (Fig. 1.8Ab). High
concentrations of gp80 are present at cell-cell contacts throughout the cell stream. These
observations suggest that DdCAD-1 is involved primarily in the recruitment of cells into cell
streams, while gp80 maintains stable contacts among cells. How DdCAD-1 is excluded from
cell–cell contacts by gp80 is not known. Since gp80 prefers an ordered lipid environment
(Harris et al., 2001b; Harris et al., 2003; Harris et al., 2001c; Harris and Siu, 2002), such an
environment might not be compatible with DdCAD-1, which likely associates indirectly with
the actin cytoskeleton through a transmembrane protein.
Anti-adhesion effect of secreted DdCAD-1

A
B
4
3
4a
B
2
1
4b
DdCAD-1
Contractile vacuole
Figure 1.8 Membrane targeting of DdCAD-1 via contractile vacuoles. (A) Confocal
micrograph of a Dictyostelium cell at 3 h of development (a), showing DdCAD-1 in
membrane protrusions and the contractile vacuole (arrow). During cell aggregation (b),
DdCAD-1 is most prominent on cells lining the periphery of the cell stream. The bar
represents 5 µm in panels a & b (B) Schematic drawing depicting the major stepsrepresents 5 µm in panels a & b. (B) Schematic drawing depicting the major steps
involved in the transport of DdCAD-1: (1) docking of DdCAD-1 onto the cytoplasmic
surface of contractile vacuole, (2) import of DdCAD-1 into contractile vacuole, (3)
association of DdCAD-1 with the anchoring protein on the luminal surface of contractile
vacuole, and (4) fusion of contractile vacuole with the plasma membrane, leading to the
release of unbound DdCAD-1 (4a) or the lateral diffusion of the anchored DdCAD-1
molecules (Adapted from Sesaki et al., 1997)
43

44
DdCAD-1 is transported to the cell surface via contractile vacuoles. A detailed
discussion of contractile vacuoles and DdCAD-1 transport mechanisms will be provided in
the last section of the introduction. Inside contractile vacuoles, not all DdCAD-1 molecules
are anchored to the lumenal surface (Fig. 1.8B). Upon fusion of contractile vacuole with the
plasma membrane, molecules that are not associated with the membrane are released in the
medium as soluble protein. Since DdCAD-1 mediates cell-cell adhesion by homophilic
interaction, soluble extracellular DdCAD-1 has an anti-adhesive effect and inhibits cell
aggregation (Siu et al., 1997). Therefore, secreted DdCAD-1 may function to modulate and
fine tune the adhesive interactions among cells in the initial stages of development. On the
other hand, cell adhesion molecules secreted into the medium or fragments of their
extracellular domain released by endogenous proteases often become associated with the
extracellular matrix and promote cell-to-substratum adhesion. Whether the secreted form of
DdCAD-1 serves such a function remains to be determined.
Cell type proportioning and cell sorting
In addition to the loss of Ca2+-dependent cell-cell adhesion, cadA-null cells give rise to
slugs with abnormal morphology and the culmination stage was delayed. Although mutant
cells are able to complete development and form fruiting bodies, defects in cell-type
differentiation and proportioning are evident. The cadA-null strains give rise to fruiting
bodies with smaller sorus and a longer stalk. Spore yield is generally reduced by 50%, with a
corresponding increase in stalk cells (Wong et al., 2002). Since DdCAD-1 is internalized in
the post-aggregation stages (Sesaki and Siu, 1996), it is likely that DdCAD-1 has an
intracellular function that can influence cell type differentiation. The Ca2+-binding capability
of DdCAD-1 raises the possibility that the intracellular DdCAD-1 is involved in the

45
regulation of Ca2+ homeostasis. Additionally, when wild-type and cadA-null cells are
transfected with an expression vector containing GFP cDNA driven by the cotB promoter,
which is a strong promoter in prespore cells (Fosnaugh and Loomis, 1993), the distribution of
GFP-marked prespore cells can be traced in slugs. A variety of abnormal patterns are
observed in the cadA-null slugs (Wong et al., 2002). Most slugs show a partially sorted
pattern, with a higher level of non-GFP cells dispersed in the posterior zone. Quantitative
analysis by FACS shows a significant reduction in the proportion of prespore cells,
corresponding closely to the decrease in spore yield. The aberrant sorting pattern observed in
cadA-null slugs might be due to changes in the profile of cell adhesion molecules.
1.2.2 Adhesion complexes involved in the multicellular development
Adhesion complexes are critical for the physical coordination of cell-cell interactions
and the morphogenesis of tissues and organs (Harris and Siu, 2002). Cell adhesion complexes
are highly dynamic. Trans-interactions of adhesion molecules are relatively weak protein-
protein interactions that facilitate the rapid assembly, dissolution and re-assembly of these
adhesion complexes. However, stable cell-cell adhesion requires strong binding forces.
These strong binding forces can be formed by oligomerization of adhesion molecules such as
cadherin and axonin-1 into zippers and lattices within large cytoskeleton-linked adhesion
complexes (Harris and Siu, 2002). As a result, the receptor interactions are strengthened
through increased binding avidity.
Cell–cell interactions in Dictyostelium remain highly dynamic within cell streams and
in aggregates, as individual cells continually exchange partners while maintaining close but
transient cell–cell contacts (Shelden and Knecht, 1995; Siegert and Weijer, 1995; Weijer,
1999). Membrane proteins that are associated with the actin cytoskeleton are generally more
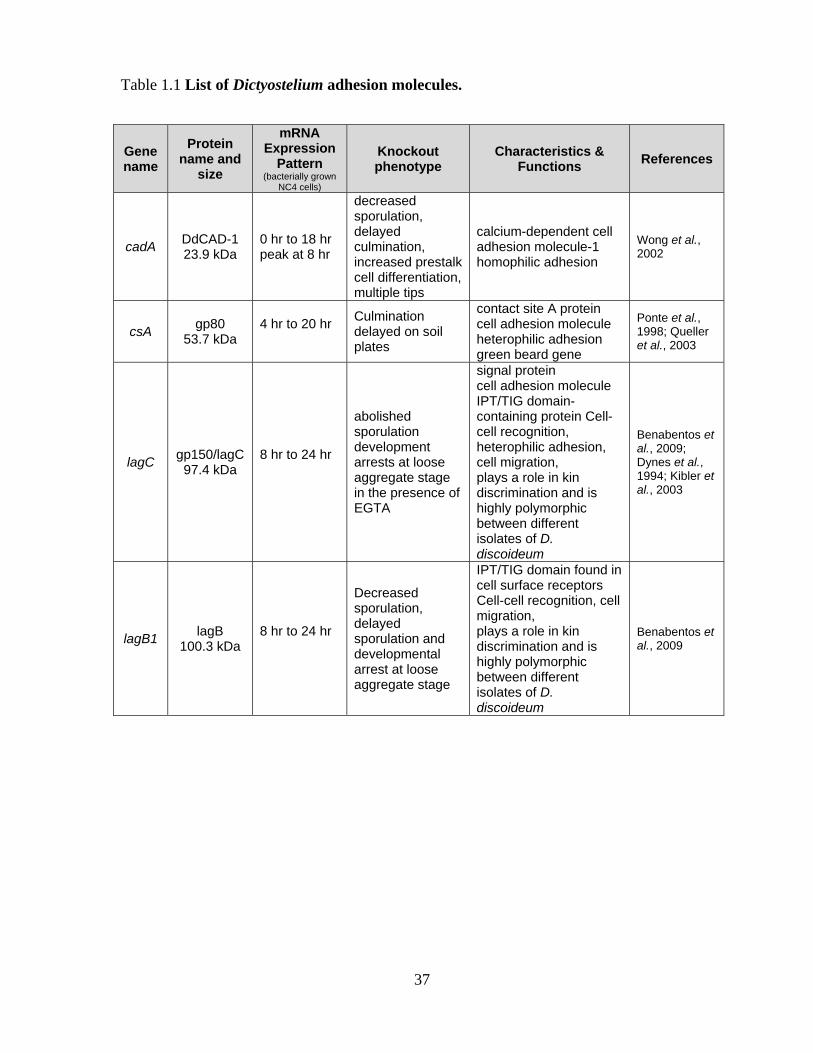
46
restricted in mobility. Many GPI-anchored membrane receptors are known to associate with
lipid rafts (Sharom and Lehto, 2002). gp80 has been found to be highly enriched in raft-like
membrane fractions (Harris et al., 2001b; Harris et al., 2001c). The oligomerization of gp80
has a stabilizing effect on rafts, which can facilitate the rapid movement of gp80 clusters in
the plasma membrane and the establishment of high avidity trans-interactions during the
initial stages of adhesion complex assembly (Harris et al., 2001c). While this model explains
how rapid assembly of gp80 adhesion complexes can be achieved, little is known about the
assembly of DdCAD-1 and gp150. Since both CAMs are not known to associate with rafts,
other mechanisms may exist.
The lack of functional epithelium with intercellular junctions is one important
difference that distinguishes Dictyostelium from the metazoa and, therefore, makes it a
potentially less powerful biological model for studying the basic cellular processes of higher
organisms (Grimson, 2000). Using a variety of conventional fixation procedures at different
developmental stages, Johnson et al. (1977) failed to find evidence of intercellular junctions
in Dictyostelium. Fuchs et al. (1993a,b) reported an epithelial-like electron dense layer of
cells on the periphery of the motile slug that appeared to have close cell-cell contacts similar
to tight junctions in higher organisms. They suggested that this distinctive cell layer might be
responsible for the secretion of the slime sheath, an ECM (extracellular matrix), and may also
act to maintain the shape and integrity of the slug. The peripheral layer of cells might also be
critical in providing a restrictive compartment in which processes, such as morphogenetic cell
movements and the establishment of chemical gradients, could occur (Fuchs et al., 1993b).
1.2.2.1 Adherens junction-like structure

47
Metazoan adherens junctions contain transmembrane cadherins, which engage in
homophilic interactions with neighbouring cells. Cadherins are connected to the actin
cytoskeleton by α- and β-catenin (Angst et al., 2001). With the advent of genome and cDNA
sequencing projects (Urushihara, 2002), database searches have led to the discovery of a β-
catenin homologue, aardvark (Aar), in Dictyostelium (Grimson et al., 2000). Aar contains ten
Armadillo (arm) repeats, which share 50% sequence similarity to those found in β-catenin.
Antibodies raised against Aar recognize specialized junctional structures that resemble
adherens junctions. These adherens junction-like structures (Fig. 1.9) are localized to a
population of cells that surround a constricted region at the top of the stalk tube (Grimson et
al., 2000; Williams and Harwood, 2003). The CAM that Aar binds to has not been identified.
It is unlikely that Aar binds directly to DdCAD-1, which does not contain a cytoplasmic
domain. The aar-null cells are able to complete development, but they form mechanically
weak fruiting bodies with a lower spore yield. Loss of Aar also leads to the formation of
branches arising from the main stalk tube (Coates et al., 2002). Similar to β-catenin, the Aar
protein has an intracellular signalling function. Aar has been found to regulate the spore cell
differentiation pathway, possibly through phosphorylation by GskA, the Dictyostelium
homologue of GSK-3β (Grimson et al., 2000). A better understanding of the function of Aar
in the architecture of the fruiting body will require a more detailed analysis of the association
of Aar with the plasma membrane and the identification of its binding partners.
1.2.2.2 Other junctional complexes
The focal adhesion protein Talin has been shown to play a role in the attachment of
cells to the substratum. However, its role in development was unraveled only recently. The
first evidence for the involvement of talin in development came from a study of a talin

A
Prestalkcells
Stalk F-actinfilaments
Adherensjunction
Stalk tubeStalk tube matrix
Transverse section
Stalk tube
Prestalkcells
Culminant
horizontal axisvertical axis
upper side basal side
B
lateral axis
junction
lower side
j
apical side
Figure 1.9 Schematics of culminant stage structures. (A) During culmination prestalk
cells move to the entrance to the stalk tube, where they differentiate into stalk cells. A
stalk of cells (marked in blue) forms constriction rings around the stalk tube. The cells in
each ring are held together by adherens junctions connected via F-actin filaments. The
formation of these junction generates cell polarity and facilitates localized deposition of
components of the stalk tube matrix (blue vesicles and arrow). (B) Schematic diagram of
an elongated prestalk cell at the constriction. The apex is adjacent to the stalk tube, the
base to the peripheral layer. (Adapted from Williams and Harwood 2003; Grimson 2000)
48
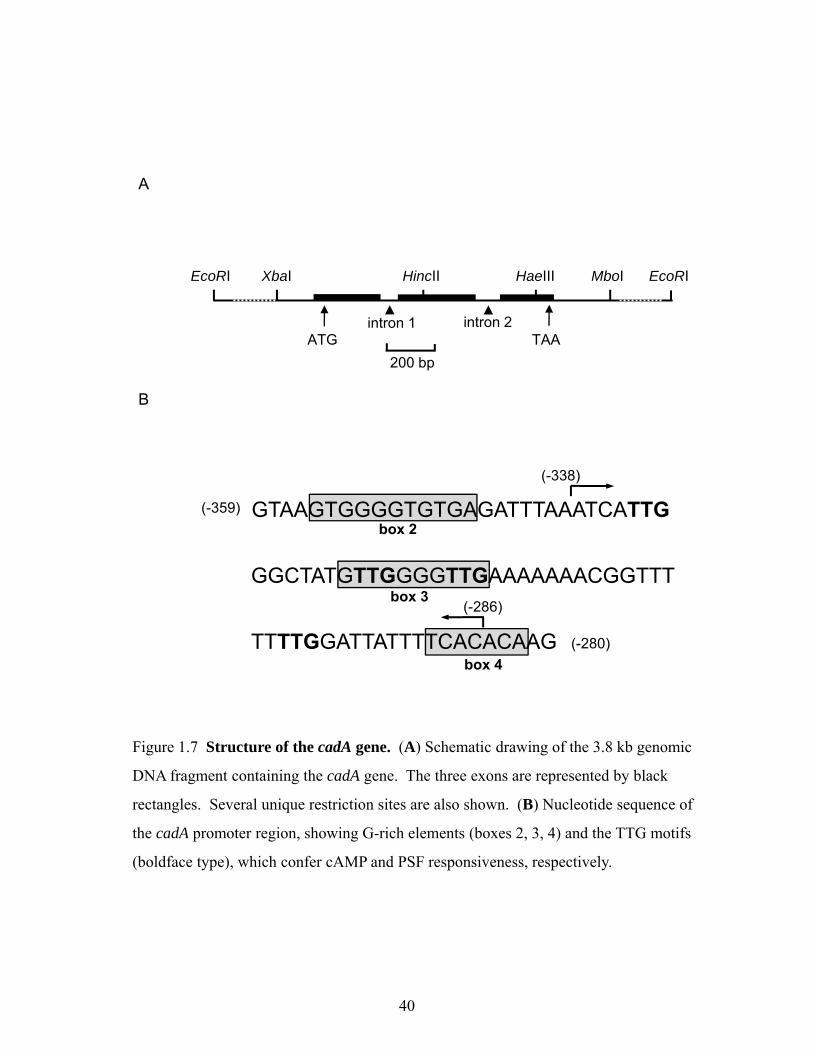
49
homologue in Dictyostelium (Tsujioka et al., 1999). Two genes (talA and talB) that encode
talin homologues have been identified in Dictyostelium (Kreitmeier et al., 1995; Tsujioka et
al., 1999). While talin A has a function during the growth phase of development (Merkel et
al., 2000; Niewohner et al., 1997; Simson et al., 1998), morphogenesis after the mound stage
is unaffected by the disruption of talA. By contrast, development of talB mutant cells is
completely blocked at the tight mound stage. These mutant cells differentiate normally within
the mounds and, when mixed with a sufficient number of wild-type cells, they participate in
the morphogenetic movement and differentiate into stalk cells and spores (Tsujioka et al.,
1999). Dictyostelium talin B, a direct homologue of the talin protein found in vertebrate focal
adhesions, is present at cell-substratum adhesion sites and is required for cell traction during
multicellular development (Tsujioka et al., 2004). A major function of talin B would be to
link mechanically the actin cytoskeleton to the substratum outside via an integrin-type
membrane protein suggesting that Dictyostelium may organize its actin cytoskeleton using a
mechanism similar to that seen in animal focal adhesions (Harwood and Coates, 2004).

50
1.3 SECRETORY PATHWAYS
Cellular proteins must be localized to the correct cellular membrane or aqueous
compartment to function properly. The process of directing each newly made polypeptide to
a particular destination – referred to as protein targeting, or protein sorting – is critical to the
organization and functioning of eukaryotic cells (Lodish et al., 2000; Pelham, 1995; Rothman
and Wieland, 1996). Proteins are sorted to the major organelles, compartments of the cell and
extracellular space. For the purpose of this thesis I will focus on the large group of proteins
that are synthesized and sorted in the secretory pathways. Secretory proteins are among the
most important messengers in the intercellular communications, which are fundamental for
many biological processes including signalling molecules for autocrine or paracrine
stimulation, cell adhesion, and cell locomotion (Alberts et al., 2007). There are two different
classes of secretory pathways that have been characterized; conventional/classical secretory
pathway and unconventional/non-classical secretory pathways.
1.3.1 Conventional secretory pathways
In conventional secretory pathways, proteins that are destined for secretion are
normally transported through ER-Golgi transport pathway. This pathway has been first
outlined by Palade and his colleagues almost forty years ago (Palade, 1975). Fig. 1.10 shows
the movement of proteins in the conventional secretory pathway. Proteins are transported to
the lumen of the ER, pass through the ER-Golgi transport vesicles to the Golgi complex for
post-translational modifications, and are stored in secretory or transport vesicles which fuse
with the plasma membrane for exocytosis (Palade, 1975; Rothman, 1994). Depending on the
cell type, exocytosis may be constitutive or regulated. Constitutively secreted vesicles will be
trafficked directly from the trans Golgi for fusion with the plasma membrane. In contrast,

Figure 1.10 Conventional secretory pathway. Ribosomes synthesizing proteins bearing
an ER signal sequence become bound to the rough ER. As translation is completed on the
ER, the polypeptide chains are inserted into the ER membrane or cross it into the lumen.
Some proteins remain resident in the ER. The remainder move into transport vesicles that
fuse together to form new cis-Golgi vesicles. Each cis-Golgi cisterna, with its protein
content, physically moves from the cis to the trans face of the Golgi stack. As this
cisternal progression occurs, many luminal and membrane proteins undergo
modifications, primarily to attach oligosaccharide chains. Some proteins remain in the
trans-Golgi cisternae, while others move via small vesicles to the cell surface or to
lysosomes. In certain cell types such as nerve cells and pancreatic acinar cells, some
soluble proteins are stored in secretory vesicles and are released only after the cell
receives an appropriate neural or hormonal signal (regulated secretion) In all cellsreceives an appropriate neural or hormonal signal (regulated secretion). In all cells,
certain proteins move to the cell surface in transport vesicles and are secreted
continuously (constitutive secretion). Integral membrane proteins move via transport
vesicles from the rough ER to the cis-Golgi and then on to their final destinations. The
orientation of a membrane protein, established when it is inserted into the ER membrane,
is retained during all the sorting steps; some segments always face the cytoplasm; others
always face the exoplasmic space (i.e. the lumen of the ER, Golgi cisternae, and vesicles
or the cell exterior). Retrograde movement via small transport vesicles retrieves ER
proteins that migrate to the cis-Golgi and returns them to the ER. Similarly, cis- or
medial-Golgi proteins that migrate to a later compartment are retrieved by smallmedial Golgi proteins that migrate to a later compartment are retrieved by small
retrograde transport vesicles. (Adapted from Glick and Malhotra, 1988; Lodish 2000;
Nickel and Rabouille 2009)
51
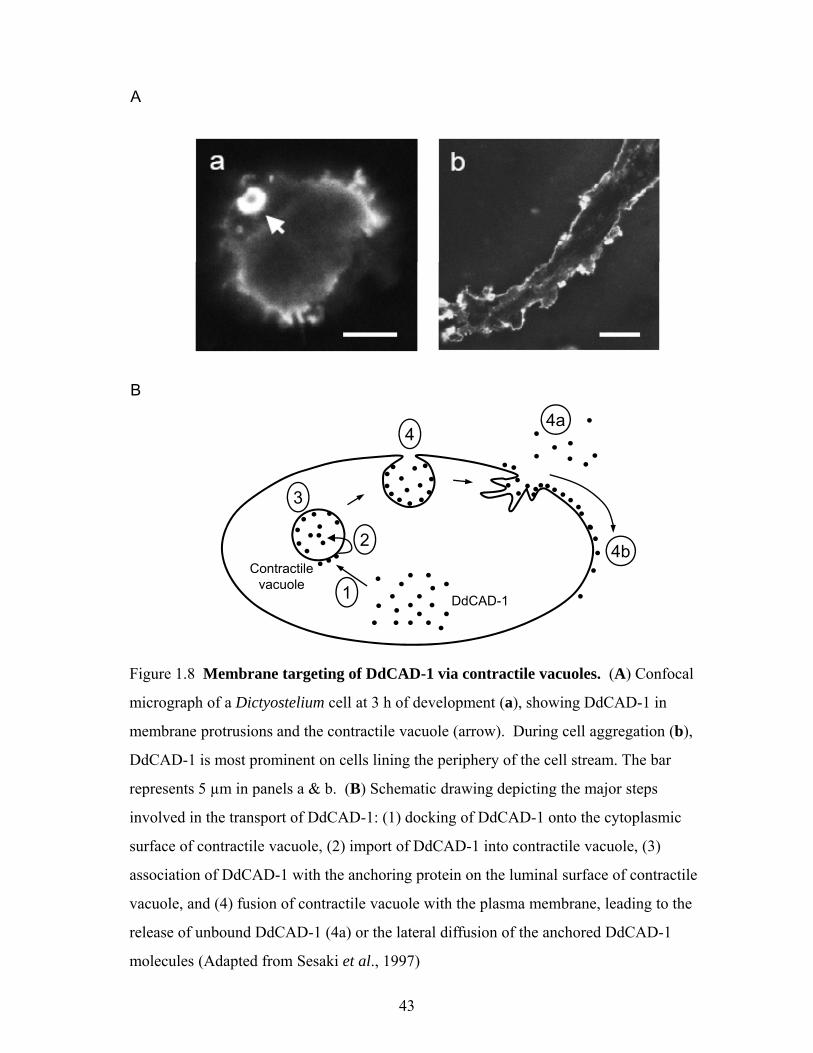
Signal peptide-
ER lumen
Signal peptide-recognition prticle
Ribosome
Endosome
g p pcontaining protein
COPI-coated vesicle
COPII-coated vesicle
Fusion events mediated bySyntaxin 5-G l i
Lysosome
mediated by SNARE proteins, NSF and SNAP
Syntaxin 5dependent
Constitutive secretion
Regulated secretion
Golgi
External stimulus
Plasma membrane
Extracellular space
52

53
regulated secretory vesicles are activated by extracellular signals such as neurotransmitters,
which signal synaptic vesicle release or a rise in intracellular calcium levels before fusion
with the plasma membrane (Coukell et al., 1992).
Nascent secretory and membrane proteins consist of N-terminal signal sequences that
mediate targeting to the ER in a signal recognition particle (SRP)-dependent manner. Signal
sequences do share common characteristics and have a tripartite structure, consisting of a
hydrophobic core region (H-region) flanked by N- and C-regions. The latter contains the
signal peptidase (SPase) consensus cleavage site (Hegde and Bernstein, 2006; Reithmeier,
1996; Reithrneier, 1996). This signal sequence directs the ribosomes that are synthesizing
these proteins to the rough ER. Once the ribosomes synthesizing these proteins become
bound to the rough ER, the proteins enter or cross the ER membrane cotranslationally during
their synthesis. This cotranslational import of proteins is unique to the ER. Other organelles
like the peroxisome and mitochondria, import proteins from the cytosol posttranslationally
(Hettema et al., 1999; Schatz and Dobberstein, 1996). Proteins meant for secretion are
completely translocated into the lumen of the ER, where they are exposed to signal peptidases
that selectively cleave off the signal peptide at a consensus site that requires small aliphatic
residues at positions -1 and -3 (von Heijne, 1990). Integral membrane proteins have one or
more additional stretches of hydrophobic amino acids that anchor the protein to the membrane
and act as transmembrane domains (Yost et al., 1983).
1.3.2 Unconventional secretory pathways
1.3.2.1 History of unconventional secretory processes
Interleukin 1β (Rubartelli et al., 1990) and galectin-1 (Cooper and Barondes, 1990)
were first demonstrated to be secreted by unconventional means almost 20 years ago. With

54
the ER-Golgi pathway processing and trafficking the majority of secreted proteins, the
identification of secreted proteins lacking a signal peptide was initially believed to occur as a
result of cell lysis, cell damage, or cell death (McNeil et al., 1989). However, further studies
clearly demonstrated that unconventional protein secretion is not paralleled by the release of
unrelated cytoplasmic proteins and, in many cases, represents a temperature-sensitive and
energy-consuming process (Cleves, 1997; Hughes, 1999; Nickel, 2003).
Originally these proteins were recognized as leaderless secretory proteins because they
lacked the classical signal peptide (Muesch et al., 1990; Rubartelli et al., 1990). In addition,
drugs which block the transport of proteins at different steps along the secretory route have
been instrumental in dissecting the process of secretion. For example, brefeldin A (BFA), a
drug that blocks the ER/Golgi-dependent secretory pathway, does not inhibit secretion of
unconventional secretory proteins (Cleves, 1997; Hughes, 1999; Misumi et al., 1986; Nickel,
2003; Prudovsky et al., 2003). Another drug, monensin blocks the transport of secretory
proteins within the Golgi complex but has no effect on unconventional secretory proteins
(Taktakoff, 1983). In short, unconventional secretory proteins usually shares several common
features, including the lack of conventional signal peptides, the absence of post-translational
modifications even if consensus NXS/T sites of glycosylation are present in their primary
amino acid sequences, and the presence of free cysteines not engaged in disulfide bridges
(Cleves, 1997; Hughes, 1999; Nickel, 2003). Many of these proteins play key roles in diverse
biological processes Table 1.2.
Recently, two types of unconventional protein transport to the cell surface of
eukaryotic cells have been characterized (Nickel and Rabouille, 2009). First, signal-peptide-
containing proteins are inserted into the ER but reach the cell surface in a coat protein
complex II (COPII) machinery-and/ or Golgi-independent manner. Second, cytoplasmic and

55
Table 1.2 List of unconventionally secreted proteins.
Protein kDa Function References
I Secretion of Signal-peptide containing proteins
BByyppaassssiinngg CCOOPPIIII vveessiicclleess Yeast heat shock protein 150 (Hsp150) 150 required for cell wall stability; induced by
heat shock, oxidative stress Fatal et al., 2002; Karhinen et al., 2005
ER degradation-enhancing a-manosidase-like protein 1 (EDEM1) (Homo sapiens)
74
It is directly involved in endoplasmic reticulum-associated degradation (ERAD) and targets misfolded glycoproteins for degradation in an N-glycan-independent manner
Cali et al., 2008
Interferon-induced GTP-binding protein Mx1 (MX1) (Ovis aries) 76 May regulate the calcium channel activity
of TRPCs Nickel and Rabouille, 2009
BByyppaassssiinngg tthhee GGoollggii
CD45 147 Protein tyrosine-protein phosphatase required for T-cell activation through the antigen receptor.
Baldwin and Ostergaard, 2001
cystic fibrosis transmembrane conductance regulator (CFTR) * transported from ER to cis-Golgi
then to endosomes, bypasses trans-Golgi
168 Involved in the transport of chloride ions. May regulate bicarbonate secretion Yoo et al., 2002
Increased sodium tolerance protein 2 (Ist2) - yeast multispan membrane protein
106 May involved in ion homeostasis Juschke et al., 2005
II Secretion of soluble proteins from eukaryotic cells
acyl-coenzyme A-binding protein (AcbA) - Dictyostelium discoideum 9
Binds to acyl-CoA. Processed into the SDF-2 (spore differentiation factor 2) a peptide which triggers sporulation.
Duran et al., 2010; Kinseth et al., 2007
Ciliary neurotrophic factor (CNTF) 24 Neurotrophic factor Reiness et al., 2001 Coagulation factor XIII, a chain 83 Coagulation Cordell et al., 2010 DdCAD-1 (Dictyostelium discoideum calcium dependent cell adhesion molecule 1)
24 Cell adhesion Sesaki et al., 1997; Sriskanthadevan et al., 2009
Endothelial/monocyte activating polypeptide II (EMAP II) 23 Activating factor for endothelium and
monocytes Tandlea et al., 2005
FGF-1 17 Differentiating and growth factor Involved in tumour induced angiogenesis
Carreira et al., 1998; LaVallee et al., 1998; Mignatti et al., 1992; Prudovsky et al., 2002
FGF-2 17 Differentiating and growth factor
Backhaus et al., 2004; Engling et al., 2002; Mignatti and Rifkin, 1991; Zehe et al., 2006
Galactin-1 (Homo sapiens) 15
May regulate apoptosis, cell proliferation and cell differentiation. Binds beta-galactoside and a wide array of complex carbohydrates.
Cho and Cummings, 1995; Cleves et al., 1996; Cooper and Barondes, 1990; Hughes, 1999; Seelenmeyer et al., 2005

56
Galactin-3 (Homo sapiens) 26
Galactose-specific lectin which binds IgE. May mediate with the alpha-3, beta-1 integrin the stimulation by CSPG4 of endothelial cells migration. Required for terminal differentiation of columnar epithelial cells during early embryogenesis
Mehul and Hughes, 1997; Sato et al., 1993; Sato and Hughes, 1994; Thery et al., 2001; Zhu and Ochieng, 2001
β-galactoside-binding protein 15 Antiproliferating factor Backhaus et al., 2004
protein high-mobility group box 1 (HMGB-1),
nuclear protein that binds to chromatin and regulates gene expression extracellular role of HMGB-1 as a proinflammatory cytokine (Erlandsson et al., 1998; Muller et al., 2001; Wang et al., 1999; Wang et al., 2004b)
Agresti and Bianchi, 2003; Bianchi and Agresti, 2005; Muller et al., 2001; Wang et al., 2004a
IL-1β 35-17 Proinflammatory and immunoregulatory cytokine Rubartelli et al., 1990
IL-18 24-18 g-interferon-inducing cytokine Gardella et al., 2000
Macrophage migration inhibitory factor (MIF) 12.5
Regulator of innate immunity, antimicrobial responses and tumor progression
Flieger et al., 2003; Merk et al., 2009
Mammary-derived growth inhibitor (MDGI) 15 Antiproliferative factor Specht et al., 1996
Platelet-derived endothelial cell growth factor (PD-ECGF) 45 Endothelial cell growth factor Schafer et al., 2004
Tat (HIV-1) 15 Viral transactivating factor: endothelial cell growth factor; immunomodulating factor
Nickel, 2003
Thioredoxin 10/12 Redox balance, catalyze thiol-disulfide exchange reactions
Arner and Holmgren, 2000; Holmgren, 1989; Rubartelli et al., 1992
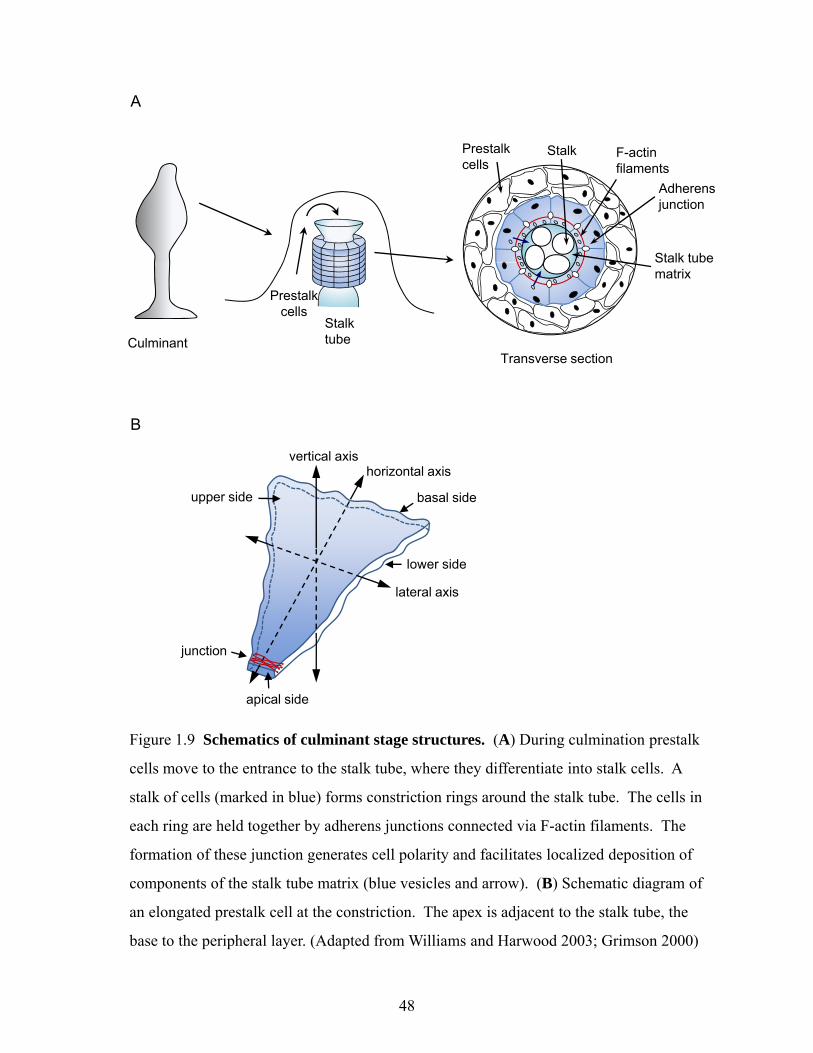
57
nuclear proteins that lack an ER-signal peptide have been shown to exit cells through ER-
Golgi-independent pathways.
1.3.2.2 Unconventional secretion of signal-peptide-containing proteins
It has been reported that some signal-peptide-containing proteins traffic
unconventionally either bypassing COPII vesicles when exiting ER or through Golgi-
independent processes (Nickel and Rabouille, 2009).
Bypassing COPII vesicles: Most of the signal-peptide-containing proteins use COPII-coated
vesicles to exit the ER. However, a number of signal-peptide-containing proteins have been
shown to bypass COPII vesicles. For example, the yeast protein Hsp150 (Fatal et al., 2002;
Karhinen et al., 2005) and ER degradation-enhancing α-mannosidase-like protein 1 (EDEM1)
are known to bypass COPII vesicles.
Bypassing Golgi: Transport of most plasma membrane proteins is sensitive to the drug BFA.
BFA is a potent inhibitor of membrane recruitment of the small GTPase ADP-ribosylation
factor 1 (ARF1). The recruitment of ARF1 is the first step in the formation of COPI-coated
vesicles from Golgi membrane (Helms and Rothman, 1992). COPI-coated vesicles mediate
retrograde transport of enzymes in the Golgi apparatus and back to the ER. Various studies
have reported evidence for Golgi-independent transport of membrane proteins to cell surface.
CD45 (Baldwin and Ostergaard, 2001), the simian rotavirus Rhesus monkey rhadino virus
(RRv) (Jourdan et al., 1997), and the transmembrane protein CFTR (Wang et al., 2004d) are
known to bypass Golgi during transport.
Both models involve the formation of ER-derived carriers with or without COPII-
coated vesicles (Nickel and Rabouille, 2009). These vesicles directly fuse with the plasma
membrane or with a late endosomal/lysosomal compartment for transport to the plasma

58
membrane. Direct fusion of ER-derived vesicles to either the endosomal/ lysosomal
compartment or the plasma membrane is dependent on the fusion machinery, which
comprises the formation of similar complexes of SNARE proteins, the ATPase NSF (N-
ethylmaleimide-Sensitive Fusion) and its cofactor synaptosomal-associated protein (SNAP)
(Jahn and Scheller, 2006; Sollner, 2004). Interestingly, a study on yeast SNARE pairings in
artificial membranes (liposomes) has revealed a functional SNARE pair that involves the ER
vesicle (v)-SNARE Sec22 and the plasma membrane t-SNARE Sec9 (Nickel and Rabouille,
2009). Moreover, yeast multispan membrane protein, Ist2 has also been shown to reach the
plasma membrane in a Golgi-independent manner. However, the transport route has been
suggested to be independent of SNAREs, NSF and SNAP (Juschke et al., 2004; Juschke et
al., 2005) and this route seems to be mediated by direct transfer of Ist2 from cortical ER
domains to the plasma membrane. It is unclear why signal-peptide containing proteins bypass
the Golgi apparatus during their transport to the cell surface. This prevents the processing of
their oligosaccharide chains as well as potential proteolytic cleavages. These two processes
can lead to a modulation of their biological activity. Therefore, it will be of interest to
analyse whether Golgi-independent transport routes of signal-peptide-containing proteins are
mechanisms that help to adapt secretory proteins to specific physiological needs.
1.3.2.3 Unconventional secretion of soluble proteins from eukaryotic cells
Many of the soluble unconventional secretory proteins are cytokines, growth factors,
or molecules with important signalling roles in physiological processes such as inflammation,
angiogenesis, cell differentiation, and proliferation (Nickel, 2003; Nickel and Seedorf,
2008a). Unconventional secretion of soluble proteins can be grouped into two subclasses
(Nickel and Seedorf, 2008a). The first class are factors that function primarily in the
extracellular space while the second class consists of proteins with both intracellular and

59
extracellular roles. The first class of proteins is represented by interleukin 1α (Siders et al.,
1993; Tarantini et al., 2001; Watanabe and Kobayashi, 1994) and interleukin 1β (Hamon et
al., 1997; Rubartelli et al., 1990; Zhou et al., 2002). The members of the second class of
unconventional secretory proteins normally localize in the cytoplasm or the nucleoplasm of
cells, where they mediate intracellular functions. However, in the presence of a specific
external stimulus, they are released from cells to mediate functions distinct from their
intracellular roles. Examples of cytoplasmic proteins that can become extracellular signaling
molecules are thioredoxin, a cytoplasmic factor involved in redox balance (Arner and
Holmgren, 2000), and the protein high-mobility group box 1 (HMGB-1), a nuclear protein
that binds to chromatin and regulates gene expression (Agresti & Bianchi 2003, Bianchi &
Agresti 2005). Although, intracellular and extracellular functions have been established for
many of these proteins, molecular details on their export mechanisms are poorly understood.
Six different mechanisms (Fig. 1.11) have been proposed to have potential roles in
unconventional secretion of soluble proteins (Nickel and Rabouille, 2009; Nickel and
Seedorf, 2008a).
Translocation across the plasma membrane
Translocation across the plasma membrane is best shown by the secretion of FGF2
(Fig. 1.11, mechanism 1). Pro-angiogenic growth factors (FGF1 and FGF2) promote
proliferation and differentiation of endothelial cells, smooth muscle cells, and fibroblasts.
During translocation across the plasma membrane, FGF2 is first recruited by the
phosphatidylinositol- 4,5-bisphosphate (PtdIns(4,5)P2) at the inner leaflet of the plasma
membrane (Temmerman et al., 2008). This interaction has been shown to be crucial for
FGF2 export because a reduction of cellular PtdIns(4,5)P2 levels results in a substantial
reduction in FGF2-export efficiency. Accordingly, mutations in FGF2 that prevent binding to

vacuoleCytoplasm
MicrovesicleDirect
Multivesicularbodies Secretory
lysosomes
1 2 3 4 5 6
Membrane flip flop
Microvesicleshedding
Direct translocation
Plasma membrane
Figure 1.11 Unconventional transport pathways of soluble proteins. Six different
mechanisms for unconventional secretion of soluble proteins have been proposed. The
Exosome releaseExtracellular space
first (mechanism 1) is non-vesicular, in which cytoplasmic proteins can be directly
translocated from the cytoplasm across the plasma membrane. For example, FGF2 is
transported via recruitment by phosphatidyl inositol-4,5-bisphosphate (Ptd Ins(4,5)P2)
and extracellular trapping by heparin sulphate proteoglycans (HPSGs), the sugar side of
which are bound by FGF2. Proteins can be captured from the cytoplasm during the y p y p g
formation of endosomal internal vesicles, which leads to the biogenesis of multivesicular
bodies (Mechanism 2). Mechanism 3 involves microvesicle shedding from the cell
surface. These internal vesicles are released into the extracellular space as exosomes. In
mechanism 4, proteins such as IL-1β is translocated into secretory lysosomes together
ith 1 Thi d t f f th t ki Th f i f thwith caspase 1. This produces a mature form of the cytokine. The fusion of these
lysosomes with the plasma membrane leads to the release of their content in the
extracellular space. Similar to mechanism 4, mechanism 5 also involves import of
soluble proteins into vacuoles and fusion of vacuoles with plasma membrane release the
proteins into extracellular space. Lastly, mechanism 6 proposes membrane flip flop of
proteins associated with the plasma membrane. (Nickel 2003; Nickel 2005)
60

61
PtdIns(4,5)P2 has been shown to block secretion. In the extracellular space, heparan sulphate
proteoglycans (HSPGs) have been shown to participate in FGF2 export to the extracellular
space (Zehe et al., 2006). It has been proposed that membrane-proximal HSPGs are
functioning as an extracellular trap that drives the net export of FGF2 (Nickel, 2007). The
membrane translocation of FGF2 does not depend on ATP hydrolysis or membrane potential
(Schafer et al., 2004). Therefore, whether FGF2 translocation across the plasma membrane is
mediated by a specific transporter or by an unrecognized ability of FGF2 to insert into plasma
membranes remains unknown (Nickel and Rabouille, 2009; Nickel and Seedorf, 2008a).
Lysosome-dependent pathway
Another pathway for unconventional protein secretion involves the sequestration of
soluble cytoplasmic factors by secretory lysosomes, a compartment that has features of both
lysosomes and secretory granules (Fig. 1.11, mechanism 4). Secretory lysosomes are
involved in the export of cytotoxic T lymphocytes or melanosomes of melanocytes when fuse
with plasma membrane (Stinchcombe et al., 2004). Lysosome-dependent pathway has been
documented for interleukin 1β (IL-1β). During an inflammatory response a precursor form of
IL-1β is first synthesized in response to bacterial lipopolysaccharides (Hamon et al., 1997;
Rubartelli et al., 1990; Zhou et al., 2002). Both IL-1β and caspase 1, the protease that is
known to convert the precursor into the mature form of IL-1β, are then translocated into
secretory lysosomes. A second trigger, extracellular ATP, is shown to promote the fusion of
secretory lysosomes to plasma membranes, which releases mature IL-1β and caspase 1 into
the extracellular space (Andrei et al., 1999; Andrei et al., 2004). IL-1β is an early
inflammatory factor while HMGB1 (protein high-mobility group box 1) secretion is triggered
by lysophosphatidylcholine and generated later in the inflammation site. Based on

62
immunofluorescence, subcellular fractionation and ultrastructural analysis, it was shown that
HMGB1 is secreted by lysosomes (Gardella et al., 2002).
Microvesicle-dependent secretion
Microvesicle-dependent secretion can occur through two different mechanisms: (1)
secretion by exosomes derived from multivesicular bodies (MVBs), and (2) secretion by
plasma membrane blebbing and vesicle shedding (Fig. 1.11, mechanism 2 and 3). The
formation of multivesicular bodies might explain the secretion of mature IL-1β, which
involves the shedding of microvesicles from the plasma membrane (MacKenzie et al., 2001;
Simon et al., 1992). In this case, caspase 1 activates IL-1β in the cytoplasm and is exported
along with the mature cytokine. The formation of endosomal intraluminal vesicles sequester
the complex made of cytoplasmic mature IL-1β–caspase, thereby leading to the formation of
IL-1β-enriched multi-vesicular bodies (MvBs). These internal vesicles are released into the
extracellular space following fusion with the plasma membrane. These vesicles, which are
known as exosomes, contain activated IL-1β as well as caspase 1 (Qu et al., 2007a). In
addition, Dictyostelium discoidin I has been shown to be transported through MvBs
(Barondes et al., 1985b).
Mechanism 5 also involves microvesicle-dependent secretion, however in this case
soluble proteins are first translocated into a vacuole similar to lysosome-dependent pathway.
Then, when vacuole fuses with plasma membrane, soluble proteins are released into the
extracellular space. Intriguigingly, Leishmania HASPB (hydrophobic acylated surface
protein B) has been shown to involve mechanism 6 for its cell surface expression. HASPB
has been found to associate with the outer leaflet of the plasma membrane only in the
infectious stages of the parasite life cycle (Nickel, 2003). It has been shown that HASPB is

63
transferred from the cytoplasm to the outer leaflet of the Golgi membrane, from where it is
transported to the inner leaflet of the plasma membrane. Based on various studies involving
fusion proteins, it has been revealed that a membrane flip flop mechanism might exit for the
expression of HASPB on the outer leaflet of the plasma membrane (Denny et al., 2000).
1.3.3 The contractile vacuole system of Dictyostelium discoideum
1.3.3.1 Organization and function of the contractile vacuole system
Contractile vacuole (CV) complex is an osmoregulatory organelle of free living
amoebae and protozoa, which controls the intracellular water balance by accumulating and
expelling excess water out of the cell, allowing cells to survive under hypotonic stress (Du et
al., 2008). In the absence of the CV complex, cells cannot expel water and they become
highly swollen and lyse. The CV system is a reticular network consisting of vacuolar
structures interconnected with tubular structures (Clarke and Heuser, 1997). Its morphology
is complex and extensive. A cell will typically contain one or two larger vacuoles with
several smaller vesicles. The number of vacuoles increases when grown in axenic (liquid)
medium and during mitosis. The CV system has been observed to split into many smaller
vesicles for stochastic partitioning during mitosis (Zhu et al., 1993b). Two different models
are proposed for these vacuolar and tubular structures; bipartite morphology or a
pleiomorphic structure. Pleiomorphic structure means that a structure is able to assume two
different forms.
Based on immunogold labelling of the CV, Nolta and Steck (1994) proposed that
Dictyostelium CV consists of two discrete structures; a large vacuole bladder and many
satellite vacuoles called spongiomes. In this bipartite model proton pumps are confined to
tubular elements (spongiome or acidosomes), while the vacuoles that fill with water
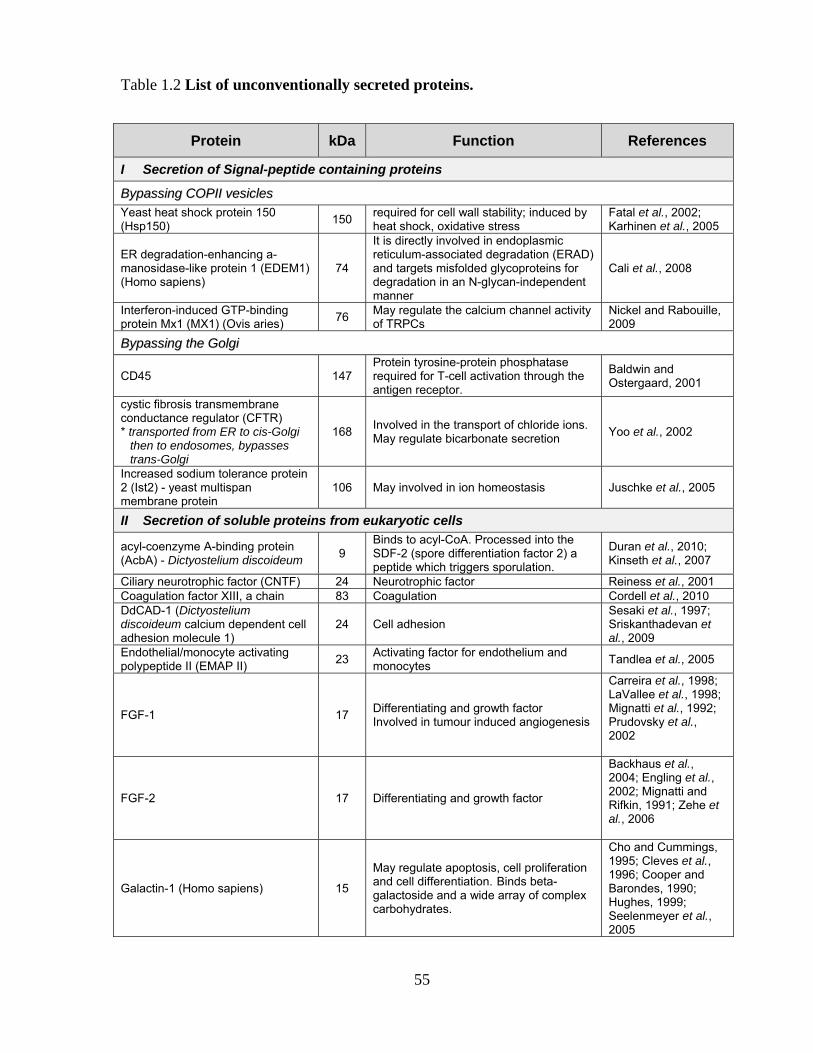
64
(bladders) are devoid of proton pumps (Bush et al., 1994; Nolta and Steck, 1994; Rhoads and
Friedberg, 1997). Spongiomes swell in response to changes in the osmotic environment and
then discharge contents by fusing with the bladder. The bladder then expels all accumulated
fluid into the extracellular space. In this model, spongiomes act as fluid uptake vesicles,
while the bladder acts as the reservoir and the final excretory organelle. On the other hand,
Clarke’s group has proposed that the contractile vacuole complex is a highly pleiomorphic
structure in which tubular and vacuolar membranes are continually undergoing
interconversion, with both forms being richly endowed with the vacuolar proton pumps
(vacuolar H+-ATPase, or V-ATPase) which allow the accumulation of water (Clarke and
Heuser, 1997; Gerisch et al., 2002; Heuser et al., 1993). The membrane composition of both
tubule and vacuole structures is of a single type. Proton pumps and alkaline phosphatase are
distributed across both structures (Heuser et al., 1993). A uniform composition of the CV
system allows fragmentation of the system into many smaller vesicles during mitosis for even
distribution into the daughter cells. After distribution, the CV system can reform by fusion of
the vesicles. Functionally, Clarke proposes that water is accumulated and retained by any part
of the interconnected network of tubular ducts and vacuolar reservoirs. In turn, any part of
the CV system can then fuse with the plasma membrane. During water accumulation, the
vacuoles can incorporate neighbouring tubules to form larger and more swollen vacuoles.
The tubules supply additional membrane material to accommodate for the growing size of the
vacuole. In this way, tubules and vacuoles can interconvert.
Clarke’s model is based on EM images of fixed and freeze-dried cells and on
observations of living cells. The EM images revealed a continuous system of interconnected
tubules and cisternae, the membranes of which are densely studded with vacuolar proton
pumps (Clarke and Heuser, 1997; Gerisch et al., 2002; Heuser et al., 1993). In addition, the

65
Gerisch group supported the pleiomorphic model using an artificial chimeric protein marker
of the contractile vacuole, dajumin-GFP, to visualize the in vivo characteristics of CV
structure (Gabriel et al., 1999). Labelling of the CV membrane with the styryl dye FM4-64
reveals that even after fusion with the plasma membrane, the dye is retained in the CV
membrane (Heuser et al., 1993), indicating very little lipid exchange between CV and plasma
membrane (Heuser, 2006). The basis of the bipartite model is the use of alkaline phosphatase
activity as the defining marker for contractile vacuole bladders, and V-ATPase activity as the
marker for spongiome membranes (Nolta and Steck, 1994). However, Clarke and Maddera
(2004) reported that alkaline phosphatase is not a suitable marker for contractile vacuoles in
Dictyostelium since antibody specific for alkaline phosphatase labels vesicles that lie at or
near the plasma membrane and only some times enriched near contractile vacuole
membranes. These observations removed the rationale for the bipartite model of contractile
vacuole structure and function. Therefore, it is more likely that the CV complex is a highly
pleiomorphic structure in which tubular and vacuolar membranes are continually undergoing
interconversion.
1.3.3.2 Osmoregulation and Other Functions of the Contractile Vacuole
Osmoregulation
Unlike the yeast vacuole, which acts as an osmoregulator and a pseudo-lysosome
containing hydrolytic enzymes, the Dictyostelium contractile vacuole is primarily an
osmoregulatory organelle (Moniakis et al., 1999). Vacuolar fusion with the plasma
membrane is believed to be a quick event. Soon after the expulsion of excess fluid, the
collapsed vacuole reforms beneath the plasmalemma and appears as a flattened tubular
structure that is ready for another cycle of water accumulation. An entire cycle of filling and
expulsion for one vacuole can take less than a minute (Clarke and Heuser, 1997). In cells

66
with multiple vacuoles, the vacuoles will alternatively fill and discharge, so that one vacuole
is always filling while another is discharging (Gabriel et al., 1999; Heuser et al., 1993). The
precise mechanism for fluid accumulation into the vacuole remains to be elucidated, but the
regulatory proteins involved in the CV fusion with plasma membrane are beginning to emerge
and will be discussed in the following section.
It has been proposed that V-H+-ATPase is involved in the retention of water in CV and
multiple ATP transporters have been localized to the CV membrane (Heuser et al., 1993;
Moniakis et al., 1999). The transmembrane electrochemical potential of H+ generated by V-
ATPase is used to transport Na+ and K+ ions or macromolecules (Beyenbach and Wieczorek,
2006), resulting in an inward flux of water (Heuser et al., 1993). Another model involves the
cotransport of H+ and HCO3- ions through the action of the V-H+-ATPase and a chloride-
bicarbonate anion exchanger, respectively (Giglione and Gross, 1995). In the CV lumen these
two components are osmotically active and can also form carbonic acid. Carbonic anhydrase
in the cytosol of the cells can readily replenish the supply of bicarbonate anions from CO2 and
H2O. Additionally, ammonia which is a by-product of protein degradation can diffuse as a
gas into the lumen of the CV, forming the ammonium cation and combine with the
bicarbonate anions (Heuser et al., 1993). The presence of these counter-ions raises the pH of
the CV. This is evidenced by the fact that CV lumens are much less acidic than endosomes.
Calcium homeostasis
In contrast to most mammalian cells, in Dictyostelium, there are two non-
mitochondrial calcium stores, one being sensitive to the second messenger inositol 1,4,5-
triphosphate (possibly ER) (Europe-Finner and Newell, 1986) and another acidic (Flaadt et
al., 1993; Malchow et al., 2006; Rooney and Gross, 1992). The acidic stores may consist of

67
the CV network, the endosomal compartment, and the acidocalcisomes (Malchow et al.,
2006). The Ca2+-transport activity of these vacuoles has been shown to be vanadate sensitive
(Milne and Coukell, 1988), thapsigargin-insensitive (Rooney et al., 1994) and facilitated by
the elevated intravesicular proton concentration (Rooney and Gross, 1992; Rooney et al.,
1994; Xie et al., 1996). Furthermore, a gene encoding the plasma membrane type Ca2+-
ATPase (PMCA), pat1 has been cloned, and its protein product, PAT1 was found to co-
localize with bound calmodulin to membranes of the CV (Moniakis et al., 1995; Moniakis et
al., 1999). The role of the CV system in Ca2+ homeostasis is supported by the upregulation of
the PAT1 and an increased localization to the CV membrane when cells are grown in a Ca2+-
rich medium. PAT1 is the Dictyostelium homologue of the yeast Ca2+ regulator Pmc lp,
which is upregulated through a calcineurin-dependent pathway. If PATl functions like Pmclp,
then it will be oriented on the CV membrane such that intracellular Ca2+ can be pumped into
the vacuole. PATI can also be found on the plasma membrane suggesting fusion of the CV
with the plasma membrane eliminates excess intracellular Ca2+ (Moniakis et al., 1999).
Furthermore Malchow et al. (2006) have found that CV is a highly efficient acidic Ca2+-store
that is required for cAMP-induced Ca2+-influx.
Regulation of CV system
Over the years, several putative regulatory proteins have been demonstrated to
associate with the Dictyostelium CV system, including vacuolar-H+-ATPase (V-ATPase),
LvsA, Drainin, Rab11A, and Rab14, but little is known about the mechanisms and signalling
pathways that control the regulation of the CV system (Becker et al., 1999; Bush et al., 1996;
Gerald et al., 2002; Harris et al., 2001a; Heuser et al., 1993; Wu et al., 2004). Fusion of
swollen vacuoles with the plasma membrane appears to be controlled by a volume sensing
protein located on the CV membrane (Becker et al., 1999). Becker et al. (1999) demonstrated

68
that Drainin, a TBC domain (RabGAP-domain)-containing protein, is required for proper CV
discharge. Drainin-null cells exhibit enlarged CV bladders and are partially sensitive to low
osmotic stress. The BEACH protein LvsA translocates to the CV membrane after the vacuole
reaches its maximum diameter and has been proposed to regulate CV biogenesis and
discharge (De Lozanne, 2003). In addition, LvsA is required for cytokinesis and phagocytosis
and for water homoeostasis (De Lozanne, 2003).
Rabs have been studied, and 16 of the 54 Dictyostelium total Rabs two, Rab11A and
Rab14, have been linked to the CV regulation (Parkinson et al., 2009). Rab11A localizes to
both CV bladder and tubular structures and cells expressing dominant-negative Rab11A
exhibit abnormal, enlarged CV bladders in hypotonic buffer and are sensitive to low
osmolarity stress (Harris et al., 2001a). Rab14 localizes to the endo-lysosomal and the CV
systems. Cells expressing dominant-negative Rab14 are defective in endocytosis, endosomal
membrane flow, homotypic lysosome fusion, and hypo-osmotic regulation (Parkinson et al.,
2009). Rab GTPases belong to the Ras superfamily of small GTPase proteins, that have
central functions in regulating membrane trafficking, including vesicle transport, formation,
movement, tethering, and fusion with target membranes (Grosshans et al., 2006; Jordens et
al., 2005). Similar to other small GTPases, Rabs cycle between GDP-bound (inactive) and
GTP-bound (active) states. The hydrolysis of GTP is stimulated by GTPase-activating
proteins (RabGAPs), most of which contain a conserved catalytic TBC (Tre/Bub2/Cdc16)
domain (Bernards, 2003). Through the examination of the cellular phenotypes of different
mutant strains and the genetic and biochemical interactions of the different CV components,
Du et al. (2008) provide new insights into the pathways regulating CV function and
formation. They reported that a novel Rab8A-GAP, Disgorgin, that along with Drainin and
its regulator Rab11A, control CV discharge. Drainin and Disgorgin/Rab8A sequentially
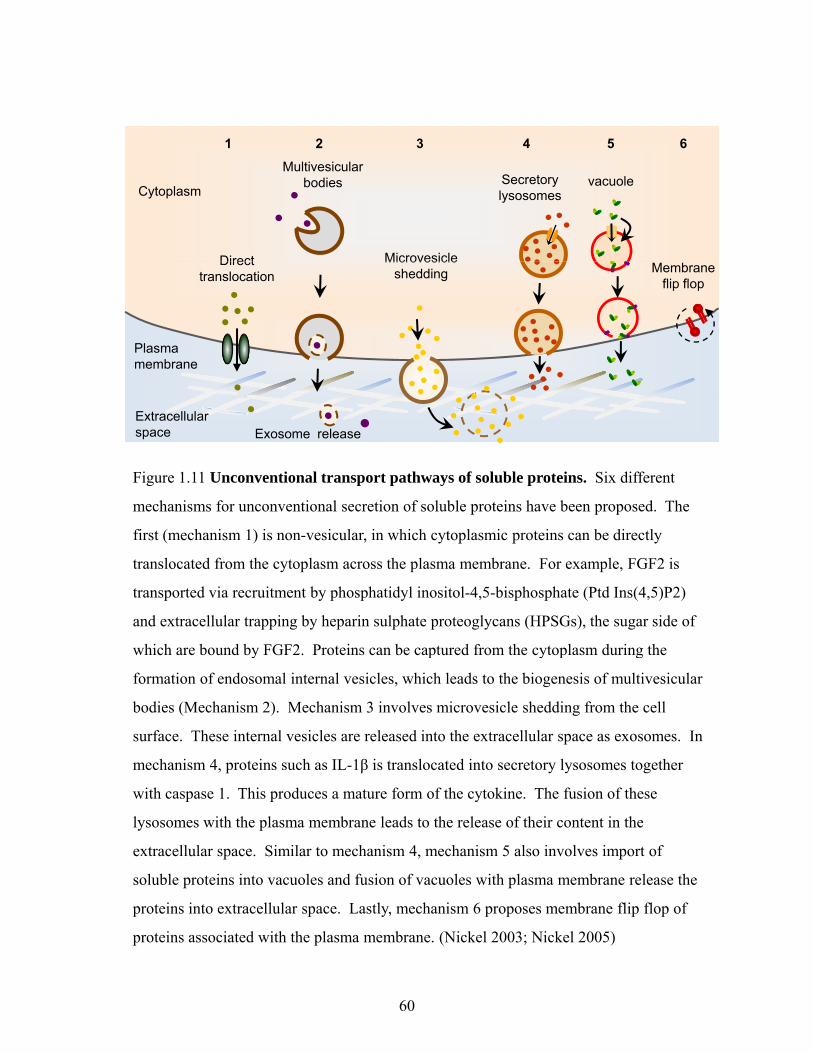
69
localize to the CV membrane at the late charging stage and control different stages of the
process. The model for the regulation of CV through the signalling pathway of Drainin and
Disgorgin is shown in Fig. 1.12. They also showed that two BEACH (Beige and Chediak-
Higashi) family proteins, LvsA and LvsD, exert an effect as suppressor and enhancer of
Disgorging-null phenotypes, respectively, and regulate CV formation.
1.3.3.3 Is there a compartment corresponding to the CV system in higher eukaryotes?
CVs are specialized organelles in protists living under hypo-osmotic conditions.
Drainin has been shown to specifically functions at the CV to expel water into the
environment. Drainin is the prototype of a protein family present in C.elegans and man
(Becker et al., 1999). Drainin family members are present in organisms whose
osmoregulation is based on mechanisms other than the periodic discharge of water from
specialized vacuoles. A compartment of apparent similarity is the sub-plasmalemmal
tubulocisternal system characterized in neuroendocrine cells (Schmidt et al., 2007). In
addition, it has been reported that giant vacuoles in the cytoplasm of frog urinary bladder
granular cells (Komissarchikf and Snigirevskaya, 2002). It has been proposed that these
vacuoles show similarity to contractile vacuoles of Protozoa and play an osmoregulatory role
in these cells. This membrane system is connected with the cell surface by narrow channels
and gives rise to synaptic vesicle-like microvesicles, the counterpart of neuronal synaptic
vesicles. It would be of interest to find out whether the CV complex, previously considered to
be a peculiarity of protozoa living under conditions of low osmolarity, is indeed a specialized
version of a compartment present in eukaryotic cells, which is connected to the plasma
membrane and functions in the conversion of ducts into vesicles.

DajuminRab11ARab11C
DajuminRab11ARab11C
Drainin Rab8A
DisgorginLvsA
Hypotonic stress
CV recharge
Figure 1.12 Contractile vacuole development in Dictyostelium cells. Based on the
kinetics of different proteins association with the CV system, different CV proteins were
assigned along the CV-charging and discharging cycles in order to define CV regulation.
Under isotonic conditions, the CV is not very active and Disgorgin, LvsA, LvsD, Rab8A, , y g g , , , ,
and Drainin are predominantly localized in the cytoplasm, whereas Dajumin, Rab11A,
and Rab11C are associated with the CV tubular and vacuolar structures. In response to
hypotonic stress, the CVs charge and enlarge by water uptake into the lumen. Late in this
charging process Drainin is recruited to the CV membrane by Rab11A-GTP. Drainin
recruitment is followed by Disgorgin and Rab8A recruitment. Once Disgorgin and
Rab8A are associated with the CV membrane, the vesicles stop charging and become
spherical as shown in the schematics. Shortly before CV fusion with plasma membrane,
LvsA is recruited to CV. Next, the CV bladder and plasma membranes fuse forming a
hole, which allows the CV contents to be released into the extracellular space. Rab8A,
Disgorgin, Drainin, and LvsA dissociate from the plasma membrane, whereas Dajumin,
Rab11A, and Rab11C remain on the CV patches. The CV patches eventually become re-
filled with water, and the next cycle begins. (Adapted from Du et al., 2008)
70

71
1.3.4 Identifying unconventionally secreted proteins
Despite the knowledge at alternative secretion pathways, no current approach directly
addresses the problem of predicting proteins entering the non-classical secretory pathway
(Bendtsen et al., 2004). Prediction methods based on amino acid composition are in principle
capable of predicting proteins entering the non-classical secretory pathway (Reinhardt and
Hubbard, 1998). However, a simple motif that defines the proteins that are secreted by
unconventional mechanisms has not been identified and may not be possible because of
diverse nature of the mechanisms identified so far. Several computational methods have
been developed to identify leaderless proteins, which might be secreted by unconventional
pathways in eukaryotic (Bendtsen et al., 2004) and prokaryotic (Bendtsen et al., 2005)
organisms. For example, Bendtsen et al. (2004) created a neural network that used sequence-
derived features to identify such proteins; features include the presence of potential sites of
post-translational modifications, predicted secondary structure, the abundance of charged
residues, the presence of predicted propeptides and other transmembrane helices, and regions
of low complexity. Using this network, they identified many proteins that are known to be
secreted by unconventional mechanisms, including fibroblast growth factor (FGF)-family
members, thioredoxin and galectin-1 (Bendtsen et al., 2004; Bendtsen et al., 2005). An
automated evaluation of known proteins or unknown protein sequences can be made at the
SecretomeP 2.0 Server website. A recent study has implicated caspase 1 as a mediator of
unconventional protein secretion, and a screen to identify proteins that transit the plasma
membrane in response to caspase 1activation has led to the identification of several proteins
that are known to exit the cell by unconventional means. These proteins include annexin A2,
macrophage migration inhibitory factor and high mobility group protein A2 (HMGA2)
(Keller et al., 2008).

72
1.4 SOCIAL INTERACTIONS OF MICROORGANISMS
1.4.1 Background information about social behaviours in microorganisms
Sociobiology, the study of social behaviour in an evolutionary framework (Wilson,
1975), provides a powerful means for explaining many kinds of social behaviour, including
mating systems, parental care, and group living. The majority of the evolutionary studies
have been focused on identifying and characterizing social behaviours in mammals, birds and
insects. It is often assumed that microorganisms lived relatively independent unicellular lives
without the cooperative behaviours. However, microbes are engaged in a variety of social
behaviours involving complex systems of cooperation, communication, and synchronization
(Crespi, 2001; West et al., 2007a). Most microbes have a multicellular stage at least at one
stage of their life cycle (West et al., 2007b). In addition, the emergence of the term
sociomicrobiology (Parsek and Greenberg, 2005) is evidence to the advances made through
the interface of sociobiology and microbiology over the past few years.
Microbiologists are making advances from a mechanistic perspective by examining
the molecular mechanisms involved in the social behaviours which have been made possible
by the availability of genomes of most microbes (Kolter and Greenberg, 2006; Lazdunski et
al., 2004; Parsek and Greenberg, 2005; Webb et al., 2003a; Williams et al., 2007).
Mechanistic studies stimulate evolutionary research because they demonstrate behaviours that
pose problems from an evolutionary perspective (Crespi, 2001; Keller and Surette, 2006).
Smith (2001) has suggested four ways to explain how microbes can be useful in the study of
social evolution. First, cooperation and communication is extremely important to microbes.
Second, the variety of social behaviours discovered in microbes offers a unique opportunity to
test how generally social evolution theory, which was largely developed to explain behaviours

73
in animals, insects and birds, can be applied to other taxa. Third, microbe systems are
uniquely amenable to experimental study since they have short generation time and genetic
mutants that do not cooperate can be created relatively easily. Fourth, the biology of
microbes means they offer some novel problems. For example, genes for social behaviours
can be transferred horizontally between different bacterial lineages, by mobile genetic
elements (Smith, 2001).
What is social behaviour? From an evolutionary point of view, behaviour is social if it
has consequences for both the actor and the recipient (West et al., 2006b). Social behaviours
can be categorized according to whether their consequences for the actor and recipient are
beneficial, increasing fitness, costly, or decreasing fitness (Table 1.3) (Hamilton, 1964b;
Hamilton, 1970). The term ‘cooperation’ refers to a behaviour that increases the fitness of the
recipient, so cooperation can be mutually beneficial or altruistically depending on the effect
on the actor (West et al., 2006b). Altruism is a social behaviour that is costly to the actor and
beneficial to the recipient (West et al., 2007c). A social behaviour is considered spite when
fitness of both the recipient and the actor decreases. Therefore, natural selection favours
mutually beneficial or selfish behaviours, whereas altruism and spite are more difficult to
explain and will be discussed later in this section.
1.4.1.1 Social interactions of Dictyostelium discoideum
Dictyostelium is a attractive model system for studying social interactions (Strassmann
et al., 2000). Social amoebae exhibit key social traits necessary for elucidating the
mechanisms of the evolution of sociality. It has a social stage where roughly 20% of the cells
die to form the stalk. The altruistic stalk cells give up reproduction in order to benefit the
spore cells, by lifting them above the hazards of the soil or increasing their chances of

74
Table 1.3 Classification of social behaviours. (Adapted from Velicer and Vos 2009; West et al., 2006)
Social behaviours
Fitness effect on
Actor Recipient
Mutually beneficial cooperation + +
Altruistic cooperation - +
Selfishness + -
Spite - -
Social behaviours
Social evolution
theory Examples
Mutually beneficial cooperation
Direct fitness benefits
Public goods: Siderophores – Iron-scavenging molecules (West and Buckling, 2003) Biosurfactants such as rhamnolipids – antiprotozoan defense mechanism (Cosson et al., 2002), mediate detachment from biofilms (Boles et al., 2005). Shiga toxins –break down host tissue (O'Loughlin and Robins-Browne, 2001) Microbial repellents – repels competitors (Burgess et al., 2003) Quorum-sensing molecules: cell-cell signals (Williams et al., 2007), iron chelation (Diggle et al., 2007; Kaufmann et al., 2005) Proteases – extracellular protein digestion (Hase and Finkelstein, 1993)
Altruistic cooperation
Indirect fitness benefits/kin selection
Fruiting bodies: Dictyostelium discoideum (Queller et al., 2003) and Myxococcus xanthus (Fiegna and Velicer, 2006; Velicer et al., 2000)
Spite (indirect altruism)
Indirect fitness benefits
Bacteriocins-proteins with antimicrobial killing activities (Gardner et al., 2004; Riley and Wertz, 2002) Red fire ant (Keller and Ross, 1998)
Whole-group traits
Both direct and indirect fitness benefits
Persisters- a small proportion of cells that are known to persist in the face of catastrophes such as antibiotic treatment (Cozens et al., 1986; Lewis, 2007) -provides a direct benefit by producing a phenotype that survives catastrophes. However, it reduces the short-term growth rate which reduces local competition for resources and hence provides an indirect benefit to relatives (West et al., 2007)
Cell death (altruistic characteristics)
Indirect fitness benefits
Saccharomyces cerevisiae – programmed cell death (PCD) provides resources that can be used by other cells for growth and survival (Fabrizio et al., 2006; Gourlay et al., 2006); Dictyostelium discoideum & Myxococcus xanthus – formation of fruiting bodies In biofilms, PCD may be useful for (a) creating channels within biofilms, which are responsible for transport of nutrients and waste and cells dispersal, and (b) releasing extracellular DNA that can be used for structuring biofilms ((Allesen-Holm et al., 2006; Webb et al., 2003; Whitchurch et al., 2002)

75
dispersal to a more favourable environment. This division between non-viable stalk cells and
viable spores leads to clear potential for conflict. Conflict is a concept that occurs when the
social disagreements arise between individuals or groups. Furthermore, aggregative
development exposes Dictyostelium to chimerism, including cheating, which raises the
questions about how the victims survive in nature and how social cooperation persists
(Benabentos et al., 2009). Cheating is a social behaviour that is carried out by individuals
who do not cooperate or who cooperate less than their fair share, but are potentially able to
gain the benefit of others cooperating (West et al., 2007c). If the slug is composed of a single
clone, then kin selection would lead to no conflict over cell fates since all the cells are initially
genetically identical. Kin selection is a process by which traits are favoured because of their
beneficial effects on the fitness of relatives (West et al., 2007c). However, when multiple
clones occur in a slug, one of clones is selected to make a relatively larger contribution to the
spore cells and a relatively smaller contribution to the stalk, at a cost to the other clones
(Strassmann et al., 2000). In addition, groups of non-clonemates can be found in nature and
cells from distinct clones or lineages will come together to form slugs and fruiting bodies,
suggesting that conflict exist in natural populations (Fortunato et al., 2003; Strassmann et al.,
2000).
Dictyostelium both cooperates and is involved in complex conflicts that are
comparable to those of social insects and vertebrates. Hence, it can be used as a model
system for conflict as well as cooperation. For example in vertebrates, meerkats generally
live in groups of up to 30 adults with their young where adults can be divided into the
dominant male and female (Griffin et al., 2003). While the dominant male and female do
most of the breeding, the subordinates help raise their offspring. When one of the
subordinates has found a scorpion, it hands it over to the offspring produced by the dominant

76
pair. How can this behaviour be explained as selfish interests? In this case, cooperation plays
a significant role to build societies (West et al., 2007c).
1.4.1.2 Other microorganisms that are involved in the social interactions
A system of differentiation analogous to the social amoebae occurs in the soil-
dwelling Gram-negative rod-shaped bacterium Myxococcus xanthus (Reichenbach, 1999;
Zusman et al., 2007). M. xanthus cells migrate on solid surfaces through gliding as a
combination of individuals and groups ranging from 2 to 3 cells up to 103 cells in a cohesive
pack. As in the Dictyostelium species, only a fraction of cells can become spores in these
fruiting bodies; many cells either undergo autolytic cell death (O'Connor and Zusman, 1988;
Wireman and Dworkin, 1977) or remain rod-shaped cells that circle the fruiting body
perimeter (O'Connor and Zusman, 1991). This population reduction within fruiting bodies
imposes strong selection on sporulation efficiency within chimeric groups, which will lead to
conflicts of interest between chimeric groups (Fiegna and Velicer, 2006a; Fiegna et al., 2006;
Kadam and Velicer, 2006). In addition, M. xanthus demonstrates a number of cooperative
traits between cells. For instance, cell density-dependent macromolecule degradation, cell–
cell exchange of motility components, and a higher efficiency of prey cell lysis when rippling
is observed, can only be observed in population when cells are cooperating for mutual
benefits (Berleman et al., 2006; Nudleman et al., 2005; Rosenberg et al., 1977; Stevens and
Søgaard-Andersen, 2005).
Pseudomonas aeruginosa, a gram-negative, aerobic, rod-shaped bacterium, capable of
causing disease in plants and animals, including humans (Griffin et al., 2004; Meyer et al.,
1997). The success and virulence of P. aeruginosa depends on quorum sensing (QS), a
process of communication by releasing and sensing small diffusible signal molecules. Diggle

77
et al. (2007) examined the social evolution of QS experimentally in Pseudomonas
aeruginosa, and showed that although QS can provide benefits at the group level, exploitative
individuals can avoid the cost of producing the QS signal or of performing the cooperative
behaviour that is coordinated by QS, and can therefore spread . Thus similar to Myxococcus
xanthus, Pseudomonas aeruginosa can also exhibit cooperative and competitive behaviours at
the population level.
1.4.2 The problem of cooperation
Cooperation is fundamental to the emergence of new levels of fitness in the biological
systems. Cooperation is an integral part of the evolution of multicellularity. While it may be
easy to agree on the basic role played by cooperation in the diversification of life, altruistic
cooperation remains a difficult interaction for evolutionary biologists to explain. Why should
an individual carry out a costly behaviour for the benefit of other individuals or the local
group? This seems to go completely against the Darwinian idea of ‘survival of the fittest.’ In
fact, Darwin identified the phenomena that were the greatest challenge to his ideas (Pennisi,
2005). Cooperation is one of these: “If it would be proved that any part of the structure of
any one species had been formed for the exclusive good of another species, it would
annihilate my theory, for such could not have been produced through natural selection”
(Darwin, 1859). Therefore, cooperation is a problem for evolution by natural selection
because according to this theory, individuals are predicted to act in a way that maximizes their
personal reproduction. However, there are several examples of cooperative behaviour in
nature (Table 1.3). Cooperative groups are vulnerable to exploitation by cheater individuals
that have access to group benefits without contributing their fair share. Cheaters are
individuals that reap the benefit of social traits while contributing less than average to the cost
(Foster et al., 2007). Therefore, cheaters would gain a competitive edge and be able to invade

78
and take over the population (Fig. 1.13). For example in the case of Dictyostelium
development acts of cooperation such as stalk formation are a challenge for evolutionary
biologists because of the potential for disruptive cheaters. Then, how do cooperative
behaviours evolve in nature?
1.4.3 Mechanisms that are involved in the altruistic cooperation
Kin selection theory provides an explanation for altruistic cooperation between
relatives (Hamilton, 1964b). By helping a close relative to reproduce, an individual is passing
on its own genes to the next generation, although indirectly. Hamilton called it ‘inclusive
fitness theory’, but it is more often referred to as ‘kin selection’, a term coined by Jon
Maynard Smith (West et al., 2006a). Altruism is a social behaviour that is costly to the actor
and beneficial to the recipient. Hamilton’s rule states that altruism is favoured when rb – c >
0; where c is the fitness cost to the altruist, b is the fitness benefit to the beneficiary and r is
their genetic relatedness (Hamilton, 1963; Hamilton, 1964b). Jerran Brown pointed out that
the inclusive fitness of an individual is divided into two components: ‘direct fitness’ and
‘indirect fitness’ (West et al., 2006a) (Fig. 1.14). Direct fitness is gained through the
production of offsprings, and indirect fitness through aiding the reproduction of nondecendent
relatives. A behaviour is only altruistic if it leads to a decrease in direct fitness, so altruism
can only be favoured when an indirect benefit outweighs this direct cost, as shown by
Hamilton’s rule (West et al., 2007d). According to his rule, greater levels of altruistic
cooperation are expected when r or b is high and c is low. Therefore, direct benefits explain
mutually beneficial cooperation whereas indirect benefits explain altruistic cooperation.
Hamilton suggested two possible mechanisms: (a) limited dispersal (population viscosity) and
(b) kin discrimination, through which a high relatedness could arise between social partners.

Mutation
C C
CC
C CC C
CCC D
D C
CDC D
D
DD
DD D
or migration Selection Selection
Declining average fitnessg g
Figure 1.13 The problem of cooperation. In the absence of cooperation, natural
selection favours selfish individuals who do not cooperate. Consider a population of
cooperators (‘C’) in which an uncooperative, selfish cheater or defector (‘D’) arises
through mutation or migration In a mixed population the selfish cheater benefits formthrough mutation or migration. In a mixed population, the selfish cheater benefits form
the cooperative behaviour of the cooperators, without paying the cost consequently the
selfish cheater has a higher fitness than the cooperators and spreads through the
population, despite the fact that this leads to a decline in mean fitness. (Redrawn from
Nowak 2006; West et al., 2007b)
COOPERATIONA behaviour that benefits another individual
(the recipient) and which is maintained because of its beneficial effect on the
recipient.
Direct fitness benefits(mutually beneficial cooperation)
Indirect fitness benefits/kin selection(altrustic cooperation)
Figure 1 14 A classification of the explanation for cooperation. (Adapted from West et
Non-enforced(shared interest in
cooperation)
Enforced/repression of competition
(when the selfish advantage of cheats is
removed)
Limited dispersal Kin Discrimination
Figure 1.14 A classification of the explanation for cooperation. (Adapted from West et
al., 2007b; West et al., 2006; Smith 1964)
79

80
1.4.3.1 Limited dispersal
Limited dispersal keeps relatives in spatial proximity to one another, allowing
cooperation to be directed indiscriminately toward all neighbours (who tend to be relatives)
(Hamilton, 1964b; Hamilton, 1971). There are two scenarios for this type of mechanism
based on how related are the neighbours. First, if only one clone colonizes in an area, then
relatedness will be high. Cooperation leads to greater growth rates and so cooperators will
out compete cheaters (Griffin et al., 2004). At high relatedness, cooperation is evolutionarily
stable. This predicted role of limited dispersal has been supported by an experimental study
on cooperation in bacteria. Many bacteria release siderophore molecules to scavenge for iron.
The release of siderophore for the common benefit is an example of a cooperative public
good; they are costly to the individual to produce, but iron bound to siderophores can be taken
up by any bacterium, providing a benefit to other individuals in the locality. Another example
of this scenario is demonstrated recently in social amobae. It has been shown that high
relatedness maintain multicellular cooperation by controlling cheater mutants in
Dictyostelium (Gilbert et al., 2007). They examined one cheater mutant (fbxA-) in the
laboratory and showed that it devastates cooperation at low relatedness but does not spread at
high relatedness. High genetic relatedness among cells can limit cheating, and promote the
cooperation that underlies multicellular development. These studies have initiated efforts to
explicitly estimate relatedness within natural populations of social microbes. Second, if
multiple colonies colonize each patch, then relatedness will be low, and each patch can
contain a mixture of cooperators and cheaters. In this case, cheaters can exploit and out-
compete cooperators and so cooperation will not be evolutionarily stable (Gilbert et al., 2007;
Griffin et al., 2004).
1.4.3.2 Kin discrimination

81
Kin discrimination happens when an individual can distinguish relatives from non-
relatives and preferentially direct aid toward them (nepotism) (Hamilton, 1964b). There are
numerous examples of kin discrimination in vertebrate species. For example, the feeding of
the young is preferentially directed towards close relatives. Long-tailed tits, where
individuals which fail to breed independently, help at the nest of closer relatives, based on
their vocal contact cues (Russell and Hatchwell, 2001). In contrast, meerkats (Griffin and
West, 2003) do not show kin discrimination when helping. However, there has been little
research on the importance of kin discrimination in microorganisms.
One of the few examples of kin discrimination is demonstrated experimentally in
Dictyostelium. Dictyostelids can minimize the cost of chimerism by preferential cooperation
with kin (Gilbert et al., 2007; Mehdiabadi et al., 2006), but the mechanisms of kin
discrimination are largely unknown. It has been shown that D. discoideum cells co-aggregate
more with genetically similar than dissimilar individuals, suggesting the existence of a
mechanism that discerns the degree of genetic similarity between individuals in this social
microorganism (Benabentos et al., 2009). It was demonstrated recently that polymorphic
members of lag gene family mediate kin discrimination in Dictyostelium (Benabentos et al.,
2009). Authors have shown that lagB1 and lagC1 are highly polymorphic in natural
populations and their sequence dissimilarity correlates well with wild-type strain segregation.
1.4.3.3 Green-beard genes
An extreme form of kin discrimination occurs in so called ‘green-beard’ gene, which
specifically direct cooperation towards other individuals that carry the same gene (West et al.,
2006b). A list of green-beard gene/gene complex identified to date is included in Table 1.4.
Hamilton initially theorized that true altruism might evolve if a supergene simultaneously

82
Table 1.4 List of green beard gene/gene complex that are involved in the self recognition.
Gene name organism Role of the gene(s) references
gp9 – gene complex
Solenopsis invicta (Red fire ants)
-a linked set of alleles, including one encoding an olfactory receptor, causes workers to kill queens that lack the green-beard allele
Keller and Ross, 1998
csA – single gene
Dictyostelium discoideum
-homophilic cell adhesion molecule -first single green beard gene Queller et al., 2003
OBY locus –gene complex
Uta stansburiana (side-blotched lizard)
-responsible for self-recognition and settlement behaviour Sinervo et al., 2006
ids (identification of self) – six-gene locus
Proteus mirabilis -involved in self, non-self recognition -localization not known Gibbs et al., 2008
FLO1 – single gene
Saccharomyces cerevisiae
-Responsible for the flocculation which protects the cells from multiple stresses, including antimicrobials and ethanol -undergoes hetereophilic adhesion
Smukalla et al., 2008
cadA – single gene
Dictyostelium discoideium
-homophilic cell-cell adhesion molecule
Sriskanthadevan et al… manuscript submitted for publication

83
affected a signal and recognition of the signal and that signal recognition elicited social acts
costly to donors but beneficial to recipients (Hamilton, 1964a). Dawkins coined Hamiton’s
social supergene a “green beard” gene in a hypothetical example of altruists that sport a green
beard distinct in color from other beards sported by non-altruists (Dawkins, 1976). Later,
Haig (1996) predicted that the homophilic functional class of protein will yield the first single
green beard gene. Classical green beard theory focuses on altruistic traits where the
behaviour imposes a net cost to the enactor and a net benefit to the recipient. In other words,
for altruism or spite to evolve, Hamilton’s rule rb > c must be satisfied. Green beard genes
remained a plausible thought experiment until an empirical example was reported by Keller
and Ross (1998). They discovered a gene cluster, gp9 linked to the display and
discrimination of identity in red fire ants, Solenopsis invicta. Gp-9 is linked to a green-beard
allele that preferentially induces workers bearing the allele to kill all queens that do not bear
it. However, this is an example of spite where red fire ants engaged in individually costly acts
of antagonism against individuals lacking the green beard trait. By using spiteful green beard
traits, microbes can actively target “unrelated” individuals that do not share the spiteful gene.
Many bacteria act rather like fire ants, using linked poison-antidote genes to kill members of
the same species that do not possess the antidote gene (Riley and Wertz, 2002).
According to Haig’s (1996) prediction, the first single gene green beard identified,
csaA, was found to code for a homophilic cell adhesion molecule in D. discoideum (Queller et
al., 2003). The csaA gene provided the first single-gene and first altruistic example of a
green-beard gene. The csaA gene encodes for a homophilic adhesion protein, gp80 that binds
to the same protein on other cells, thus excluding strains that do not express the protein from
the benefits of later altruism within the group. In this case, the green beard gene is fixed in
the population, and the effects of its absence are only seen in artificially constructed

84
knockouts. On the natural substrate of soil, the adhering cells pull each other into aggregates,
while knockout cells lacking gp80 are left behind (Ponte et al., 1998). However, on the
smooth surface of agar, the csaA knockout is not impeded from aggregating, and once in the
aggregate performs as a cheater, preferentially becoming a spore, not part of a stalk (Queller
et al., 2003). This behaviour is also partly due to its weakened intercellular binding, because
less-adhesive cells will slide to the back of the slug where they are more likely to become
spores. Thus, the greater adhesion of wild-type cells makes them more altruistic, but the
adhesion also insures that benefit does not go to less-adhesive cheaters, at least on the natural
substrate of soil.
In addition, Haig (1996) suggested that this single green beard gene could be a
heterophilic cell adhesion protein. Recently, Smukalla et al. (2008) have demonstrated this is
the case with the yeast flocculation gene, FLO1 which is a specific cell-surface protein that
interacts with mannan oligosaccharide chains to form lectin-like bonds resulting in cell-cell
adhesion which leads to the formation of flocs. The yeast FLO1 green-beard gene adds a new
level of interest to these studies because it is highly variable and the variation exerts a major
influence on protein binding strength. Conceivably, such variation could lead to multiple
recognition tags, although this possibility remains to be tested (Queller, 2008).
1.4.3.4 Other mechanisms of social evolution
Pleiotropy
The phenomenon that most genes affect many traits is known as pleiotropy. It is a
major evolutionary constraint because adaptive change in one trait may be prevented because
it would compromise other traits affected by the same genes (Barton, 1990; Foster et al.,
2004). The pleiotropic links between cheating and personal costs can stabilize cooperative

85
adaptation. For instance, the pleiotropy related form of cheater control occurs in bacteria by
tight linkage of genes in colicin production (Riley and Wertz, 2002). Colicins are produced
by bacteria through a plasmid that contains genes for both colicin production and resistance to
their toxic effects. This linkage of genes prevents loss of colicin production by plasmid loss
because this would also cause loss of resistance (Travisano and Velicer, 2004).
Another example of pleiotropy is shown in Dictyostelium where the pleiotropic
linkage of stalk and spore formation limits the potential for cheating because a defect on
prestalk cell production results in an even greater reduction in spores (Foster et al., 2004).
The complex interrelationships of developmental and biochemical processes mean that genes
have innumerable distinctive side effects that constrain adaptation (Barton, 1990;
Featherstone and Brodie, 2002; Otto, 2004). Foster et al. (2004) have found that these
constraints can benefit cooperative adaptations by limiting the potential for individual
selfishness. Social innovations will persist only when disruptive cheaters cannot invade the
population (Hurst et al., 1996). Pleiotropy provides one way to limit individual rebellion and
allow stable cooperation to evolve. Where pleiotropic costs are strong, other means of control
like high relatedness (Hamilton, 1964b) and policing (Frank, 2003) may be less necessary.
However, it is not expected that all cheating will be controlled by pleiotropy. Nevertheless,
pleiotropy will be a common form of cheater control because it is so ubiquitous (Foster et al.,
2004).
Phoenix genes
When cheater mutants were mixed with wild-type cells and cultured over multiple
cycles of fruiting-body formation, the populations became extinct in many cases. The cheater
mutants can dominate the population but could not sporulate when cultured alone. However,

86
in one example, a new strain called phoenix arose from the social collapse (Velicer et al.,
2006). Interestingly, this strain could form spores, resist the cheater mutant and even produce
more spores than the ancestral (wild type) strain (Velicer et al., 2006). Whole genome
sequencing has revealed that it took just single base change to go from the cheater strain to
the new super strain. The mutation increases expression of a gene that is predicted to encode
a member of the Gcn5-related N-acetyltransferase superfamily, some members of which
regulate both global and specific gene expression. Although the mechanism by which the
phoenix strain suppressed the cheater strain has not been determined, it is clear that a single
mutation could rescue the social system from the verge of collapse (Foster et al., 2007;
Velicer et al., 2006).
Although the same social evolution theory should apply to both macroscopic and
microscopic organisms, the details may be different (West et al., 2007b). For instance, based
on kin selection, limited dispersal may be the key mechanism in microorganisms (Griffin et
al., 2004; West et al., 2006b), whereas kin discrimination is the key mechanism in
macroorganisms such as cooperative breeding vertebrates (Griffin and West, 2003).
Greenbeard effects may be more important in microorganisms because the required signalling
and recognition can take place at the level of individual cells interacting with their neighbours
(Queller et al., 2003). Additionally, spite may be more common in microorganisms owing to
local competition for resources and extreme differences in relatedness (Gardner et al., 2004).
There have been several theories proposed to explain the cooperative behaviours in
organisms. These theories are not mutually exclusive and there are several examples of
explanations where behaviour is explained using two different theories. For example, whole-
group traits such as siderophore production provide a benefit to both the individual that
produces them and to nearby relatives. This cooperative behaviour can provide both a direct

87
and indirect fitness benefit (West et al., 2006b). Another example is that if by-product
benefits occur between relatives, this will lead to direct and indirect benefits of cooperation.
Similarly, although cooperation is favoured between relatives, if there is a mechanism for the
repression of competition between relatives, then there will be even higher levels of
cooperation (West et al., 2006b).

88
1.5 HYPOTHESES AND RATIONALE OF THE THESIS
My research project is focused on the Ca2+-dependent cell adhesion molecule
DdCAD-1, which is synthesized at the onset of Dictyoselium development. To investigate the
structure and functional relationships of DdCAD-1, my research project contains three main
objectives: (1) characterization of the homophilic binding site of DdCAD-1, (2) elucidation of
DdCAD-1 transport mechanism, and (3) assessment of the role of DdCAD-1 during pattern
formation.
1.5.1 Characterization of the homophilic binding site of DdCAD-1
Earlier work on DdCAD-1 (Wong et al., 1996) has suggested that DdCAD-1 might be
a distant homolog of cadherin. DdCAD-1 seems to be related to both the vertebrate and
invertebrate extracellular domain of cadherin and is involved in the morphogenesis of
Dictyostelium. Therefore, further investigation could provide insight into the evolutionary
biology of cadherins and the mechanism of binding of this unique protein. To determine the
NMR-structure of DdCAD-1 we collaborated with Dr. Yang from National University of
Singapore. The NMR-solution structure of DdCAD-1 has revealed that it consists of two β-
sandwich domains, with the N-terminal domain and the C-terminal domain resembling βγ-
crystallins and classical cadherins, respectively. Based on the structure of DdCAD-1, I first
tested which domain of DdCAD-1 is involved in the homophilic interaction. To investigate
the mechanism of dimer formation, we used site-directed mutational analysis in combination
with in vitro binding assays. Further, the dependence on Ca2+ was extensively examined
using EGTA to chelate Ca2+. The results are presented in Chapter 2.

89
1.5.2 Elucidation of the DdCAD-1 transport mechanism during the early phase of
development
In order to function as a cell adhesion molecule, DdCAD-1 has to be expressed on the
cell surface. DdCAD-1 is synthesized in the cytoplasm as a soluble protein and then
transported to the plasma membrane for surface presentation or secretion. Sesaki et al.
(Sesaki et al., 1997) have shown that DdCAD-1 is transported to the cell surface through a
non-classical transport pathway involving contractile vacuoles. However, the molecular
mechanism of DdCAD-1 expression on the cell surface through the non-classical pathway is
unknown. DdCAD-1-GFP fusion protein was expressed in cadA-null cells for further
investigation of this unconventional protein transport pathway. To assess the structural
requirements in this transport process, the N-terminal and C-terminal domains of DdCAD-1
were expressed separately in cadA-null cells as GFP-fusion proteins. The results are
presented in chapter 3.
1.5.3 Assessment of the role of DdCAD-1 in Dictyostelium pattern formation
A fundamental question in developmental biology is how a group of initially identical
cells forms a patterned adult organism with different cell types organized in correct positions.
During development of Dictyostelium, multiple cell types are formed and undergo a
coordinated series of morphogenetic cell movements to generate the fruiting body. DdCAD-1
is expressed on the cell surface in the initial stages of development and it is present in the
cytoplasm as a soluble protein throughout development (Sesaki and Siu, 1996; Wong et al.,
2002). The key question here is: What is/are the functions of DdCAD-1 during the later
stages of development. Studies on cadA-null cells have implicated DdCAD-1 in cell sorting
and cell-type proportioning (Wong et al., 2002). To investigate how DdCAD-1 is involved in

90
cell sorting, DdCAD-1-GFP was expressed ectopically using its endogenous promoter, -631.
The spatial and temporal changes in the subcellular distributions of DdCAD-1 were analyzed
using confocal microscopy. Ca2+-binding mutants are expressed in cadA-null strain and its
effects on cell sorting and cell-type proportioning are examined using both
immunofluorescence and biochemical techniques. Results are presented in chapter 4.

91
Chapter 2
Solution structures of the adhesion molecule DdCAD-1 reveal new
insights into Ca2+-dependent cell-cell adhesion.
The work presented in this chapter has been published in the following paper:
Lin, Z., Sriskanthadevan, S., Huang, H., Siu, C.H., and Yang, D. (2006). Solution structures
of the adhesion molecule DdCAD-1 reveal new insights into Ca2+-dependent cell-cell
adhesion. Nat. Struct. Mol. Biol. 13: 1016-1022.
(Z.L. contributed to NMR sample preparations, assignments, structure calculations,
mutagenesis studies, in vitro homoassociation studies (Figures 2.1, 2.2 and 2.6); H.H.
contributed to molecular cloning and protein expression; C.-H.S contributed to project
guidance and manuscript preparation; and D.Y. contributed to project guidance, NMR data
collection and manuscript preparation.)

92
2.1 Summary
DdCAD-1 is a novel Ca2+-dependent cell adhesion molecule that lacks a hydrophobic
signal peptide and a transmembrane domain. DdCAD-1 is expressed by the social amoeba
Dictyostelium discoideum at the onset of development. It is synthesized as a soluble protein
and then transported to the plasma membrane by contractile vacuoles. Here we describe the
novel features of the solution structures of Ca2+-free and Ca2+-bound monomeric DdCAD-1.
DdCAD-1 contains two β-sandwich domains, belonging to the βγ-crystallin and
immunoglobulin fold classes, respectively. Whereas the N-terminal domain has a major role
in homophilic binding, the C-terminal domain tethers the protein to the cell membrane. From
structural and mutational analyses, we propose a model for the Ca2+-bound DdCAD-1 dimer
as a basis for understanding DdCAD-1–mediated cell-cell adhesion at the molecular level.
Our results provide new insights into Ca2+-dependent mechanisms for cell-cell adhesion.

93
2.2 Introduction
Development of eukaryotes is characterized by the coordinated expression of a variety
of cell adhesion systems, which provide the mechanical forces that regulate cell shape, cell
motility and the formation of three-dimensional tissue structures. Cell adhesion molecules
not only contribute to the formation of specialized junctional complexes that maintain tissue
integrity, but also can serve as signaling centers that regulate cell proliferation, differentiation,
apoptosis and many important cellular processes. Recent studies of metazoan and
nonmetazoan adhesion systems have revealed the diversity of roles played by adhesion
molecules as well as common principles that underlie the mechanism and function of this
important class of molecules (Bowers-Morrow et al., 2004).
Our study seeks to investigate the evolutionary origins of adhesion molecules and the
characteristics of their structure-function relationships shared by organisms across the phyla.
The social amoeba Dictyostelium discoideum is a nonmetazoan related to both animals and
fungi, situated favorably at an evolutionary position close to Acanthamoebae and the acellular
slime mold Physarum polycephalum (Baldauf and Doolittle, 1997; Baldauf et al., 2000). For
several decades, Dictyostelium has been a good model organism for multifaceted study of
cell-cell adhesion, as it is amenable to both genetic and biochemical manipulations (Coates
and Harwood, 2001; Siu et al., 2004a).
The onset of Dictyostelium development is characterized by chemotactic migration to
aggregate in response to cAMP. Multicellularity is maintained by the expression of several
adhesion systems. Early studies distinguished two major classes of cell adhesion sites (Beug
et al., 1973). One class is sensitive to low concentrations of EDTA, whereas the other is
stable in the presence of 10–15 mM EDTA. The EDTA-sensitive sites are mediated by the

94
cell adhesion molecule DdCAD-1 (Brar and Siu, 1993), whereas csA (also called gp80)
(Kamboj et al., 1989) and LagC (also called gp150) (Wang et al., 2000) are respectively
responsible for the EDTA-resistant sites at the aggregation stage and the mound stage of
development. In this paper, we focus on DdCAD-1, which is expressed at the onset of
development (Yang et al., 1997). DdCAD-1 is a unique cell-cell adhesion molecule because
it lacks a hydrophobic signal peptide and a transmembrane domain (Wong et al., 1996). It is
synthesized as a soluble protein of 213 amino acid residues and then transported to the plasma
membrane by contractile vacuoles (Sesaki et al., 1997). DdCAD-1 molecules on the cell
surface can be induced to form ‘caps’ by antibody cross-linking, suggesting that they are
linked to the cytoskeleton by a transmembrane component (Wong et al., 1996). DdCAD-1
shows limited sequence similarity with the spore coat protein of Myxococcus xanthus, protein
S, and with classical cadherins (Wong et al., 1996). Like cadherins, DdCAD-1 is a Ca2+-
binding protein and its adhesive activity is dependent on Ca2+. Knockout of the cadA gene,
which encodes DdCAD-1, not only results in the loss of the Ca2+-dependent cell-cell adhesion
but also gives rise to aberrant cell sorting and a reduction in spore yield during Dictyostelium
development (Wong et al., 2002). These results indicate that, in addition to cell-cell
adhesion, DdCAD-1 has a role in cell-type proportioning and pattern formation during
development.
We now report the previously uncharacterized NMR solution structures of Ca2+-free
and Ca2+-bound DdCAD-1 in their monomeric forms. We ascribe distinct binding
interactions to the two domains of DdCAD-1. In addition, we describe a model structure for
the Ca2+-bound DdCAD-1 dimer as a basis for understanding DdCAD-1–mediated cell-cell
adhesion at the molecular level.

95
2.3 Experimental Procedures
Cell-to-substratum attachment assay
Aliquots of 50 μl each were taken from 4 μM protein samples and placed on 1-cm-
diameter spots on nitrocellulose coated Petri dishes for 1.5 h, then blocked with 1% (w/v)
BSA for 1 h. The Dictyostelium strain KAx-3 was cultured in liquid medium and collected at
3 h of development for the assay as described (Siu et al., 2004a). All values were normalized
to those of cells bound on the intact protein (WT) after background subtraction.
Fluorescent microspheres-to-cell binding assay
The cadA-null cells were developed for 3 h and resuspended at 2 x 106 cells ml–1.
After blocking with 1% (w/v) BSA for 15 min, red microspheres conjugated with different
proteins (0.5 ml) were added to 0.1 ml cell samples and rotated at room temperature for 10
min. Cells were allowed to attach onto 0.1% (w/v) poly(l-lysine)–coated slides for 10 min
before fixation with 3.7% (v/v) formaldehyde and staining with DiO (green) for microscopic
observation.
Antibody-induced cap formation
The cadA-null cells were developed for 3 h and then incubated with 40 μg of His-
tagged protein for 30 min. Antibody to His tag (TeckniScience) was added to the cells for 30
min. Next, Alexa-568 goat secondary antibody to mouse (Molecular Probes) was added at
1:100 dilution and the sample was rotated for 30 min at 22 oC to allow antibody-induced
clustering of the antigen. Cells were fixed and stained with DiO. Routinely, 100–200 cells
were scored for cap formation.

96
Construction and expression of His6-tagged fusion proteins
His6-WT (intact DdCAD-1) was constructed by ligating a 0.7-kb cDNA fragment
encoding the DdCAD-1 sequence between Ser2 and Lys213 into the expression vector pQE-9
(Qiagen). Both deletion mutations and point mutations of DdCAD-1 were created by PCR
amplification. The His6-N (N-terminal domain between Val3 and Gly94) and His6-C (C-
terminal domain between Ala95 and Lys213) were also cloned into pQE-9. Site-directed
point mutations were created by overlapping extension PCR method. All constructs were
sequenced to ascertain sequence fidelity. Protein expression was carried out in the E. coli
strain BL21 (DE3).
45Ca2+-overlay assay
Recombinant wildtype and mutant DdCAD-1 proteins were blotted onto nitrocellulose
membrane using a slot blot apparatus (BioRad). 45Ca2+-binding assays were carried out as
described previously (Maruyama et al., 1984; Sesaki et al., 1997). The membrane was first
washed with an overlay buffer containing 60 mM KCl, 5 mM MgCl2, and 10 mM imidazole,
pH 6.8, for 1 h in four changes. The blot was then incubated with 10 μCi/ml 45Ca2+ in the
overlay buffer for 30 min at room temperature on a platform shaker, followed by three 10-min
washes with deionized water. Autoradiography was carried out by exposure of the air-dried
45Ca2+-labeled membrane to Bioflex-MRI Films (Clonex Corp, AR) for 24 h at 70 °C.
Fluorescent microsphere-to-substratum attachment assay
Red fluorescent 0.5 μm diameter microspheres (250 μl) were covalently conjugated
with 250 μg of His6-tagged WT, N or C protein using the water soluble carbodiimide reagent,

97
EDAC (Molecular Probes). Recombinant proteins (5 μl of a 5 μM solution) were used to coat
spots of 6 mm in diameter on poly-L-lysine-coated glass slides for binding with protein-
conjugated microspheres as previously described (Zhao and Siu, 1995). Digital images of
attached microspheres were recorded and analyzed using the NIH Image software.

98
2.4 Results
NMR structure of Ca2+-free and Ca2+-bound DdCAD-1
The structures of DdCAD-1 in the absence and presence of Ca2+ were determined
(Fig. 2.1). DdCAD-1 consists of an N-terminal domain (residues 2–93), a C-terminal domain
(98–213) and a short linker of 4 residues connecting the two domains. The N-terminal
domain comprises two Greek-key motifs, each with four β-strands, and two short helical
elements between the two motifs, forming a two-layer sandwich architecture. The C-terminal
domain is also a two-layer sandwich composed mainly of β-strands, with nine strands
organized in two sheets. The relative orientation of the two domains is primarily determined
by hydrophobic interactions, salt bridges and hydrogen bonds. The hydrophobic interdomain
contacts involve residues Leu92 and Pro93 from the N-terminal domain, Ala95 and Phe96
from the interdomain linker and Ala99 and Pro139 from the C-terminal domain. In addition
to hydrophobic interactions, positively charged residues Lys88 and Arg30 from the N-
terminal domain and negatively charged residues Asp101 and Asp201 from the C-terminal
domain appear to form interdomain salt bridges. Hydrogen bonding characterized by
hydrogen-deuterium exchange studies also contributes to interdomain interactions, such as the
hydrogen bond between the ε proton of Trp55 in the N-terminal domain and the carbonyl
oxygen of Thr179 in the C-terminal domain. These interactions result in an exclusive
arrangement of the two domains.
Comparison of the Ca2+-bound structure with the Ca2+-free one reveals remarkable
similarities. Ca2+-bound DdCAD-1 has the same β-sheet structure as the Ca2+-free form, and
its domain orientation shows nearly no change. The average backbone r.m.s. deviation with
respect to the Ca2+-free structure is 1.55 A° for the N-terminal domain (residues 2–55 and 68–

a
b
Figure 2.1 Ribbon diagrams of Ca2+-free (a) and Ca2+ -bound (b) DdCAD-1. Blue, N-
terminal domain; green, C-terminal domain; red, helical structures in the N-terminal
domain. In each domain, β-strands are sequentially labeled with letters and numbers.
Strands A1, B1, D1 and G1 form motif 1; C1, E1, F1 and H1 form motif 2; A2, D2, I2 and
H2 form sheet 1; B2, C2, E2, F2 and G2 form sheet 2. (Additional figures of DdCAD-1
NMR structure can be found in Lin et al., 2006)
99

100
93) and 1.10 A° for the C-terminal domain (residues 98–213). Major differences between the
NMR data obtained for Ca2+-free and Ca2+-bound DdCAD-1 exist only in the Ca2+-binding
regions. Binding of Ca2+ results in more ordered side chain packing in the Ca2+-binding
pockets and stabilizes the whole structure of DdCAD-1. This was also evidenced by the full
backbone assignment of DdCAD-1 in the presence of Ca2+, in contrast to the incomplete
backbone assignment in the absence of Ca2+.
Ca 2+-binding sites
Ca2+ titration monitored with 1H-15N HSQC spectra reveals three potential Ca2+-
binding pockets. Structure calculation reveals that the first Ca2+-binding site, site I, involves
side chain oxygen atoms of Asp39, Thr81 and Asn84 and the backbone carbonyl oxygen of
Phe41 in the N-terminal domain. The second site, site II, which consists of the side chain
oxygen atoms of Asp35, Asn38, Glu56 and Ser87, is also able to coordinate one Ca2+ in the
N-terminal domain of DdCAD-1. The last binding site, site III, is unique, as it is situated at
the interface of the two domains. It comprises the E1-F1 loop from the N-terminal domain
and the F2-G2 and H2-I2 turns from the C-terminal domain. Side chain oxygen atoms of
Glu59 and Asp61 from the N-terminal domain and Thr179, Gln181 and Asn202 from the C-
terminal domain are involved in chelating Ca2+ in site III. These Ca2+-binding residues are
further confirmed by Mn2+-induced relaxation experiments. The dependence of chemical
shifts of amides in the binding sites on Ca2+ concentration fits well to a moderately fast
exchange model with average dissociation constants (Kd) of 14.9 ± 3.4 μM, 14.4 ± 2.9 μM
and 12.1 ± 2.1 μM for site I, site II and site III, respectively. These Kd values are statistically
identical, implying nearly identical binding affinities for the three Ca2+-binding pockets and a
noncooperative Ca2+-binding mode unlike the cooperative binding observed in cadherins

101
(Alattia et al., 1997). These results are consistent with those obtained from equilibrium
binding studies, which also suggest noncooperative binding, with an apparent Kd of 24 μM.
Structural comparison to other cell adhesion proteins
Although the two domains of DdCAD-1 adopt similar structural architectures, the
structural topologies of the N-terminal and C-terminal domains are totally different. Closest-
fold searching using both the GRATH (Pearl et al., 2005) and DALI (Holm and Sander, 1998)
servers shows that the N-terminal domain of DdCAD-1 has a βγ-crystallin fold, whereas the
topology of the C-terminal domain is consistent with an immunoglobulin-like fold (Fig. 2.2),
revealing a novel global fold of the two-domain DdCAD-1. More notably, structural
comparison with other Ca2+-dependent cell adhesion proteins shows that the N-terminal
domain of DdCAD-1 shares a very similar topology with the N-terminal domain of protein S
(Bagby et al., 1994), but it lacks one α-helix and has a relatively long loop between β-strands
E1 and F1. This loop contains 12 residues (Glu56–Lys67) and forms a unique domain
interface with the F2-G2 and H2-I2 turns of the C-terminal domain. The C-terminal domain
of DdCAD-1 and the extracellular domains of cadherins (Boggon et al., 2002) belong to the
same fold family, although their β-strand connectivities are not completely identical (Fig.
2.2).
Structure-based sequence alignment demonstrates that the N-terminal domain of
DdCAD-1 and the N-terminal domain of protein S share 88% of structural overlay with an
SSAP score (http://www.cathdb.info/cgi-bin/cath/SsapServer.pl) of 86. The superimposed
structures are shown in Figure 2.2. These results indicate that the N-terminal domain of
DdCAD-1 and protein S can be clustered into the same homologous superfamily and further
suggest their evolutionary relatedness. The evolutionary relationship is also evidenced by the

Figure 2.2 Structural comparisons of DdCAD-1 with protein S and cadherin. (a) β-
sheet topology of Ca2+-bound DdCAD-1. The first and last residues in each β-strand are
indicated. (b) Stereoview of superimposed structure of the N-terminal domain of
d ( ) i h h i l d i f i ( ) (DdCAD-1(cyan) with the N-terminal domain of protein S (cyan, PDB: 1NPS). (c)
Superimposition of the C-terminal domain of DdCAD-1 (blue) with the first N-terminal
domain of C-cadherin (cyan, PDB: 1L3W).
102

103
fact that these proteins have adhesive properties and are capable of forming dimers or
oligomers in a Ca2+-dependent manner (Bagby et al., 1994; Inouye et al., 1979).
Additionally, the two Ca2+-binding sites in the N-terminal domain of DdCAD-1 show
similarity to those in the N-terminal domain of protein S (Wenk et al., 1999). According to
structure-based sequence alignment, Asn84 and Ser87 in DdCAD-1 are evolutionarily
retained, and the equivalent residues in protein S are similarly located in two Ca2+-binding
sites in the N-terminal domain (Wenk et al., 1999); DdCAD-1 Asp39 and Glu56 are partially
conserved.
Structure-based multiple sequence alignment among the C-terminal domain of
DdCAD-1 and the first N-terminal domains of cadherins reveals limited structural similarity.
Thr179 and Gln181, which are involved in forming the interdomain Ca2+-binding site (site III)
in DdCAD-1, are conserved and in the same alignment positions as their counterparts
involved in forming one Ca2+-binding site in cadherins. However, the C-terminal domain of
DdCAD-1 and the first N-terminal domain of cadherins may not belong to the same
homologous superfamily, given their low sequence identity and differences in strand
connectivity.
N-terminal domain mediates homophilic binding
Although gel-filtration results suggest that both Ca2+-free and Ca2+-bound DdCAD-1
exist in a monomeric form in solution, dynamic light-scattering (DLS) data demonstrate that
monomers and dimers coexist in a concentration-dependent equilibrium in the presence of
Ca2+. An obvious increase in the apparent size of DdCAD-1 upon binding Ca2+ was observed
when the protein concentration was > 0.2 mM. On the basis of hydrodynamic radius (Rh)
measured by DLS, the apparent molecular weight of DdCAD-1 at 0.5 mM was estimated to

104
be 27.5 ± 0.4 kDa in the absence of Ca2+, a measurement that increased to 37.3 ± 4.3 kDa in
the presence of 10 mM Ca2+. DLS size distribution shows that the increase in apparent
molecular weight results from the formation of dimers instead of larger oligomers, which
agrees well with our earlier observation that DdCAD-1 mediates cell-cell adhesion through
homophilic interactions in vivo (Brar and Siu, 1993). DLS results also show that DdCAD-1
does not self-associate in the presence of 10 mM Mg2+, regardless of protein concentration,
which is consistent with the observation that the adhesion function of DdCAD-1 is dependent
on Ca2+.
Direct binding assays show that DdCAD-1 present on cells or conjugated to
fluorescent microspheres can bind DdCAD-1 coated on the substratum (Fig. 2.3). To
determine which of the two domains is involved in homophilic binding, recombinant proteins
containing either the N-terminal or C-terminal domain were prepared from bacteria (Fig.
2.3a,b). Consistent with the structural data, the N-terminal domain retained its Ca2+-binding
capability, whereas the C-terminal domain had none (Fig. 2.3c). Results from both cell to
substratum and microsphere-to-substratum binding assays demonstrate that the N-terminal
domain alone is sufficient for homophilic binding (Fig. 2.3d–f). In both assays, the C-
terminal domain retained ~20% of the binding capability of the intact protein. Consistent with
our previous in vivo observation (Brar and Siu, 1993), the homophilic interaction of DdCAD-
1 is dependent on Ca2+ (Fig. 2.3d).
To assess the contribution of ionic interactions to homophilic binding, the
microsphere-to-substratum binding assay was performed in the presence of various
concentrations of NaCl. Binding was inhibited by 50% at 0.2 M NaCl, whereas 0.5 M NaCl
reduced it to the background level (Fig. 2.3g). Once the complex was formed, the homophilic
interaction of DdCAD-1 became resistant to NaCl. The complex was relatively stable even

Figure 2.3 The N-terminal domain of DdCAD-1 mediates cell-cell adhesion via
homophilic binding. (a) Schematic drawing depicts the His6-DdCAD-1 recombinant
proteins (WT, wild-type; N, N-terminal domain; C, C-terminal domain). (b) Coomassie
blue–stained gel profiles of the purified recombinant proteins. (c) Autoradiograms of the 45Ca2+-overlay assay done with different amounts of protein. BSA was included as a
negative control. (d) The N-terminal domain of DdCAD-1 promotes cell attachment.
Cells were seeded onto spots on a Petri dish coated with His tagged WT N or CCells were seeded onto spots on a Petri dish coated with His-tagged WT, N or C
recombinant protein (white bars). In competition experiments, cells were first incubated
with either 50 μg ml–1 (gray) or 100 μg ml–1 (black) of recombinant protein for 30 min at
4 oC before deposition on the protein coated spot. The requirement for Ca2+ was examined
by including either 2 mM CaCl2 or 2 mM EGTA in the assay. Data represent means ± s.d.
(n = 9). (e) Micrographs showing binding of fluorescent microspheres conjugated with
different recombinant proteins to slides coated with different proteins. (f) Quantification
of microspheres attached to recombinant WT, N or C, or BSA, coated on the Petri dish.
Results were normalized to amount of beads bound to spots coated with WT protein. (g)
Salt effects on DdCAD-1 homophilic binding in the bead-binding assay The fluorescentSalt effects on DdCAD 1 homophilic binding in the bead binding assay. The fluorescent
microsphere-to-substratum binding assay was carried out in the presence of various
concentrations of NaCl. (h) Effects of salt washes on WT-beads bound on spots coated
with WT protein. After each assay, the bound beads were incubated for 1 min with
various concentrations of NaCl. Alternatively, the bound beads were incubated with 0.1%
or 0.5% (v/v) NP-40 or Triton X-100 for 1 min. Beads remaining on the protein-coated
spots were fixed for quantification. Data in f–h represent means ± s.d. (n = 3 to 6).
105

106

107
when 0.5 M NaCl was included in one of the washing steps (Fig. 2.3h). Notably, the
inclusion of low concentrations of a nonionic detergent disrupted the adhesion complex
effectively (Fig. 2.3h). The data suggest that the initial binding reaction involves mainly
ionic interactions, and subsequent hydrophobic interactions contribute to the stability of the
complex.
C-terminal domain tethers DdCAD-1 to cell membrane
As DdCAD-1 does not contain a transmembrane domain, a segment of the protein
must be involved in tethering it to the cell membrane. To determine whether the C-terminal
domain has such a role, fluorescent microspheres conjugated with various DdCAD-1
recombinant proteins were incubated with cadA-null cells, which do not express DdCAD-1,
with the assumption that the putative anchoring protein for DdCAD-1 was fully available for
binding with exogenous DdCAD-1. The results show that beads conjugated to both the C-
terminal domain and the intact DdCAD-1 attached to cadA-null cells, whereas N-terminal
domain–conjugated beads did not bind (Fig. 2.4a,b). Alternatively, cells were incubated with
soluble recombinant proteins, and the binding of intact proteins and C-terminal domains was
assessed by antibody-induced clustering of the membrane component involved in tethering
DdCAD-1 to the cell surface. Neither BSA nor the N-terminal domain was efficient in
inducing cap formation, whereas 60%–70% of cells incubated with either intact protein or C-
terminal domain formed caps (Fig. 2.4c,d). In contrast to N-terminal domain–N-terminal
domain interactions, the binding of the C-terminal domain to the cell membrane was
insensitive to EGTA (Fig. 2.4e).
Reverse-charge mutations affect homophilic binding

Figure 2.4 The C-terminal domain tethers DdCAD-1 to the cell membrane. (a)
Epifluorescence micrographs showing the binding of fluorescent microspheres (red)
conjugated with WT, N or C fusion protein (as in Fig. 2.2a) to cadA-null cells (green).
Scale bar, 5 μm. (b) Quantification of data from cells with at least five beads bound on
the cell surface. Routinely, 200–300 cells were scored per coverslip. (c) Antibody-induced
cap formation (red) after the binding of recombinant proteins to the cadA-null cells
(green). Scale bar, 5 μm. (d) Quantification of data from cells with cap structures on the
cell surface (e) Effects of Ca2+ and EGTA on the binding of DdCAD-1 to cadA-null cellscell surface. (e) Effects of Ca and EGTA on the binding of DdCAD 1 to cadA null cells.
Data in b, d and e represent means ± s.d. (n = 6).
108

109
The availability of three-dimensional structures of Ca2+-free and Ca2+-bound DdCAD-
1 allows rationalization of mutagenesis experiments to assess the importance of ionic
interactions in the homophilic binding of DdCAD-1. Site-specific mutations were introduced
in both hydrophobic and charged residues (Fig. 2.5). The reverse-charge mutations were
designed on the basis of the surface charge of Ca2+-free DdCAD-1. Conversion of two crucial
positively charged residues to negative ones (K40E and K36E) around the Ca2+-binding sites
in the N-terminal domain substantially reduced its relative binding activity in the cell-to-
substratum attachment assay (Fig. 2.5a) without disturbing the secondary structure or
abolishing Ca2+ binding (Figs. 2.6 & 2.7). Reverse-charge mutations (D133K, D134K and
D125K) in the other negatively charged region in the C-terminal domain also had a negative
effect, albeit to a lesser extent, on the homophilic binding activity of DdCAD-1. Similar
results were obtained with the microsphere-to substratum binding assay. The data suggest the
involvement of both N-terminal and C-terminal domains during dimerization and further
highlight the importance of charge interactions between two DdCAD-1 molecules.
Structural model of the Ca2+-bound DdCAD-1 dimer
The Ca2+-bound DdCAD-1 structure was employed to assess the sites of homophilic
interaction. The experimental data obtained from mutagenesis studies were used as docking
restraints and were sufficient to characterize a unique relative orientation for the two
molecules by HADDOCK (Dominguez et al., 2003) calculations. Multiple refinements
converged to a mean backbone r.m.s. deviation of 0.78 ± 0.19A°. Ramachandran analysis of
the resulting model shows that the dimer structure, including the intermolecular interface, still
occupies the energetically preferred conformation. The pairwise backbone r.m.s. deviation
from the starting structures is less than 0.9 A°, indicating that the structures of the two Ca2+-

a
b
Figure 2.5 Structural model of Ca2+-bound DdCAD-1 dimer. (a) Effects of mutations
on the cell-binding activity of DdCAD-1 in the cell-to-substratum binding assay.
Asterisks indicate significant differences: *P < 0.001; **P > 0.1 (Student’s t-test for
unpaired samples). (b) Ribbon diagram of a representative structure of DdCAD-1 dimer.
The partner molecules are colored in blue and green, respectively.
110

Figure 2.6 CD spectra of DdCAD-1 mutants. Wildtype (WT): blue; mutants:
pink.
111

a
b
Figure 2.7 Binding of 45Ca2+ to wildtype and mutant His6-DdCAD-1 proteins. (a)
Autoradiograms of the 45Ca2+ overlay assay. Different amounts of recombinant protein
were blotted onto nitrocellulose membrane using the BioRad slot blot apparatus. The blot
was incubated with 45Ca2+ (10 μCi/ml) for 30 min at room temperature. BSA was
included as the negative control and purified Dictyostelium calmodulin (CaM) was
included as the positive control. Filters were washed with several changes of distilled
water and the then exposed to X-ray film at –70 oC for 24 h. (b) Quantification of the
autoradiograms using the NIH Image software. Pixel values were normalized to that of
the WT protein and plotted against the amount of protein loaded in the slot Datathe WT protein and plotted against the amount of protein loaded in the slot. Data
represent the average of three experiments.
112

113
bound DdCAD-1 molecules do not undergo any considerable change in the dimer form. The
backbone ribbon drawing of the lowest-energy conformer and an overview of the
intermolecular interface are presented in Figure 2.5b. The model structure of Ca2+-bound
DdCAD-1 dimer adopts C2 symmetry. Analysis of intermolecular contacts reveals that the
hydrophobic contact between Trp37 and the C-terminal domain of the partner is most
prominent, appearing in ~90% of the model structures. Indeed, the single W37A mutation,
which did not abolish Ca2+ binding, inhibited cell attachment by ~80% in the cell-to-
substratum binding assay, whereas the C16A mutation had little effect (Fig. 2.5a).

114
2.5 Discussion
Ca2+-dependent adhesion mediated by DdCAD-1
Earlier work on E-cadherin points out that binding of Ca2+ to cadherin results in a rod-
like structure, which is crucial for cis-dimerization, and this structure would collapse in the
absence of calcium ions (Pertz et al., 1999). We did not observe such conformational changes
in DdCAD-1, although the relative rearrangement of the two domains is slightly better
defined by the binding of Ca2+, especially to site III. As Ca2+ does not induce major
conformational changes in DdCAD-1, what is its role in the adhesion function of DdCAD-1?
Binding of Ca2+ markedly changes the electrostatic potential only on the Ca2+ binding sites in
the N-terminal domain and has no effect on the negative charge surface of the C-terminal
domain. Taking all of the available data together, we propose that the homophilic interaction
of DdCAD-1 could be driven by a change in electrostatic potential caused by the binding of
Ca2+ to the three binding sites. In the absence of Ca2+, electrostatic repulsion between the N-
terminal domain of one molecule and the C-terminal domain of the other prevents two
DdCAD-1 molecules from docking together. Ca2+ would greatly reduce repulsion forces, and
the basic side chains surrounding the Ca2+-binding sites in the N-terminal domain would be
involved in electrostatic interactions with the negative ones in the C-terminal domain of the
partner molecule. These interactions probably constitute the initial binding reaction, followed
by further stabilization of the dimer structure by hydrophobic interactions between the N-
terminal domain and the partner N- and C-terminal domains.
Distinct roles for the two domains of DdCAD-1

115
High concentrations of DdCAD-1 lead to homoassociation in solution. However,
attempts to identify the intermolecular interface by transferred cross-saturation measurements
or transferred NOEs have been unsuccessful, possibly owing to the relatively short lifetime of
the dimer and the native weakness of DdCAD-1 homophilic interaction in vitro. Similar
observations have been made for protein S and cadherins, whose solution structures are both
monomers (Bagby et al., 1994; Overduin et al., 1995).
Subdomain studies demonstrate that the N-terminal domain is crucial for homophilic
binding and the C-terminal domain also contributes, albeit to a lesser extent, to dimer
formation. Microsphere-to-cell binding assays using the cadA-null cells and capping of the
putative anchor protein via binding of the C-terminal domain to the null cells show that the C-
terminal domain of DdCAD-1 is involved in tethering DdCAD-1 to a transmembrane protein
on the cell surface.
Although protein S is capable of self-assembly, presumably through cis-interaction, on
the spore coat of myxobacteria (Bagby et al., 1994; Inouye et al., 1979), it is not clear
whether it can undergo trans-interactions as seen for DdCAD-1. Classical cadherins are best
characterized in their trans-homophilic interactions. Although trans-homophilic interactions
are centered in the first extracellular domain (Boggon et al., 2002), cadherin adhesion
involves the interdigitation of multiple extracellular domains, and it has been suggested that
the first two N-terminal domains are a minimal unit for adhesion (Koch et al., 2004; Perret et
al., 2004; Shan et al., 2004). The C-terminal domain of DdCAD-1 contributes to homophilic
binding, although it is responsible primarily for tethering DdCAD-1 to the cell membrane. It
is noteworthy that both E-cadherin and NCAM can interact heterophilically with other
membrane receptors (Karecla et al., 1996; Kleene and Schachner, 2004), even though it is
well established that they mediate cell-cell adhesion by homophilic binding. It remains to be

116
determined whether the heterophilic and homophilic binding capabilities associated with
adhesion molecules have evolved independently or one from the other.
Several crystal structures of cadherins have provided models for cadherin-mediated
adhesion, whereas the exact molecular basis for protein S multimerization is still open to
speculation. Although questions concerning cadherin-mediated homophilic specificity still
remain, it has been reported that tryptophan-mediated interactions are crucial for cadherin
dimerization, and Trp2 is engaged in both cis and trans-interactions with the partner N-
terminal domain (He et al., 2003). The model structure of DdCAD-1 dimer indicates only
one type of homophilic interaction (trans-interaction). Notably, Trp37, near the Ca2+-binding
region in the N-terminal domain, is also important in DdCAD-1–mediated cell-cell adhesion.
Although minor structural perturbations caused by the W37A mutation cannot be ruled out,
our data show that the mutant protein still retains nearly full Ca2+-binding capacity (Fig. 2.7),
suggesting that the configuration of the Ca2+-binding sites remains intact. Therefore, it is
likely that the much weaker binding of W37A to the cell in the cell-to-substratum binding
assay is primarily due to the loss of hydrophobic interactions between Trp37 and the C-
terminal domain. In contrast to classical cadherins, the tryptophan side chain interacts mainly
with the partner C-terminal domain rather than the partner N-terminal domain in the model
structure. As the C-terminal domain does not undergo homophilic binding efficiently, the
function of Trp37 may depend on the initial electrostatic interactions between the two partner
molecules. Therefore, the mechanism of DdCAD-1 interaction seems to differ from the
tryptophan-docking model proposed for classical cadherins (Boggon et al., 2002). It is
possible that the Trp37 interaction contributes to the specificity of DdCAD-1 homophilic
interactions.

117
The NMR structures and dimeric model provide new insights into the unique
structure-function relationships of DdCAD-1, which are distinct from other Ca2+-dependent
adhesion proteins. The structural characterization of the Ca2+-binding sites of DdCAD-1
provides the basis for future studies on its potential role in the regulation of Ca2+ homeostasis,
which is important for cell-type differentiation and proportioning during development.

118
Chapter 3
The Cell Adhesion Molecule DdCAD-1 Is Imported into Contractile
Vacuoles by Membrane Invagination in a Ca2+- and Conformation-
dependent Manner
The work presented in this chapter has been published in the following paper:
Sriskanthadevan, S., Lee, T., Lin, Z., Yang, D., and Siu, C.H. (2009). The Cell Adhesion
Molecule DdCAD-1 Is Imported into Contractile Vacuoles by Membrane Invagination in a
Ca2+- and Conformation-dependent Manner. J. Biol. Chem. 284: 36377-36386.
(T.L. contributed to some of the in vitro import assays, Z.L. and D.Y. contributed to circular
dichroism measurements (Figure 3.11); C.-H.S contributed to project guidance and
manuscript preparation.)

119
3.1 Summary
The cadA gene in Dictyostelium encodes a Ca2+-dependent cell adhesion molecule
DdCAD-1 which contains two β-sandwich domains. DdCAD-1 is synthesized in the
cytoplasm as a soluble protein and then transported by contractile vacuoles to the plasma
membrane for surface presentation or secretion. DdCAD-1-GFP fusion protein was expressed
in cadA-null cells for further investigation of this unconventional protein transport pathway.
Both morphological and biochemical characterizations showed that DdCAD-1-GFP was
imported into contractile vacuoles. Time-lapse microscopy of transfectants revealed the
transient appearance of DdCAD-1-GFP-filled vesicular structures in the lumen of contractile
vacuoles, suggesting that DdCAD-1 could be imported by invagination of contractile vacuole
membrane. To assess the structural requirements in this transport process, the N-terminal and
C-terminal domains of DdCAD-1 were expressed separately in cells as GFP-fusion proteins.
Both fusion proteins failed to enter the contractile vacuole, suggesting that the integrity of
DdCAD-1 is required for import. Such a requirement was also observed in in vitro
reconstitution assays using His6-tagged fusion proteins and purified contractile vacuoles.
Import of DdCAD-1 was compromised when two of its three Ca2+-binding sites were
mutated, indicating a role for Ca2+ in the import process. Spectral analysis showed that
mutations in the Ca2+-binding sites resulted in subtle conformational changes. Indeed,
proteins with altered conformation failed to enter the contractile vacuole, suggesting that the
import signal is somehow integrated in the three-dimensional structure of DdCAD-1.

120
3.2 Introduction
In eukaryotes, soluble secretory proteins are typically transported through the classical
ER-Golgi transport pathway (Palade, 1975; Rothman, 1994). However, it has become evident
in recent years that a growing number of soluble proteins synthesized in the cytoplasm are
targeted for secretion (Nickel and Rabouille, 2009; Nickel and Seedorf, 2008b). This group
of secretory proteins usually share several common features, including the lack of
conventional signal peptides, the absence of post-translational modifications, and the presence
of free cysteines (Cleves, 1997; Nickel, 2003). Many of these proteins play key roles in
diverse biological processes. Among them are galectins (Cho and Cummings, 1995; Cleves
et al., 1996), interleukin-1β (Orci et al., 1991), thioredoxin (Rubartelli et al., 1992),
macrophage migration inhibitory factor (Flieger et al., 2003), and fibroblast growth factor 1
and 2 (FGF-1 and FGF-2) (Engling et al., 2002; Mignatti P, 1992). Several viral proteins,
such as HIV-Tat and Herpes simplex VP22, are also known to be secreted via ER/Golgi-
independent routes (Denny et al., 2000; Elliott and O’Hare, 1997; Mann and Frankel, 1991).
Multiple unconventional transport mechanisms have been discovered and cells can
utilize one or more pathways to target soluble proteins for secretion. The IL-1β export
mechanism involves intracellular vesicles in its secretory pathway (Andrei et al., 1999;
Rubartelli et al., 1990), while the muscle lectin galectin-1 is externalized by the shedding of
membrane vesicles (Cooper and Barondes, 1990) or through a protein transporter
(Seelenmeyer et al., 2005). Secretion of the growth factor FGF-2 is also likely mediated by a
plasma membrane-resident transporter (Schafer et al., 2004). Additionally, cell surface
counter receptors are essential components in the export machinery of galectin-1 since they
can provide an extracellular trap mechanism (Seelenmeyer et al., 2005).

121
Several soluble proteins expressed by the social amoeba Dictyostelium discoideum have been
found to be targeted for secretion by unconventional pathways (Siu et al., 2004b;
Sriskanthadevan et al., 2007). At the onset of development, amoeboid cells undergo
chemotactic migration to form large aggregates of ~105 cells, which eventually culminate in
the formation of fruiting bodies (Aubry and Firtel, 1999). The endogenous soluble lectin
discoidin-I, which is externalized to facilitate cell-substratum adhesion (Crowley et al., 1985;
Springer et al., 1984), is targeted for secretion through vesicular structures (Barondes et al.,
1985a). Multicellularity during Dictyostelium development is maintained by the expression
of several cell adhesion molecules (Siu et al., 2004b). One of them is the Ca2+-dependent cell
adhesion molecule DdCAD-1 which is encoded by the cadA gene. DdCAD-1 lacks the
classical signal peptide and is synthesized as a soluble protein in the cytoplasm (Brar and Siu,
1993; Wong et al., 1996; Yang et al., 1997). It is imported into contractile vacuoles for
transport to the plasma membrane (Sesaki et al., 1997). DdCAD-1 contains two distinct
domains with β-sandwich architecture. While the N-terminal domain is involved in
homophilic binding, the C-terminal domain tethers the protein to a membrane anchor, thus
allowing it to function as a cell adhesion molecule on the cell surface (Lin et al., 2006). Both
DdCAD-1 and discoidin-I have been found to be enriched in contractile vacuoles (Sesaki et
al., 1997), the osmoregulatory organelles that regulate water balance during the growth and
the initial stages of development (Gerisch et al., 2002; Heuser et al., 1993). As they fuse with
the plasma membrane, the contents of the contractile vacuoles are released into the medium.
In this report, we investigated the structural requirements for the import of DdCAD-1
into contractile vacuoles. Using transfectants that express DdCAD-1-GFP fusion proteins, we
observe that DdCAD-1 is imported through the invagination of contractile vacuole
membranes. However, the whole protein is required since neither the N-terminal domain nor

122
the C-terminal domain alone is sufficient for transport into the contractile vacuole.
Additionally, the import mechanism is dependent on Ca2+ and proper protein conformation,
suggesting that the secretory targeting motif of DdCAD-1 lies in its three-dimensional
structure rather than in a linear stretch of amino acids.

123
3.3 Experimental Procedures
Construction of DdCAD-1-GFP, N-GFP and C-GFP expression vectors and cell
transfection
For DdCAD-1-GFP expression, a 645 bp fragment of DdCAD-1 cDNA was cloned
into the HindIII and EcoRI sites of pA15/NIGFP expression vector (kindly provided by Dr.
David Knecht, University of Connecticut, CN). The N-terminal domain (N, 288 bp) and the
C-terminal domain (C, 366 bp) of DdCAD-1 were amplified by PCR methods before cloning
into pA15/NIGFP. Primers used in this study are listed in Fig. 3.1. The expression constructs
were transfected into cadA-null cells by electroporation as described by Pang et al. (Pang et
al., 1999). Cells were harvested at log phase, washed two times in cold H50 solution (20 mM
Hepes, 50 mM KCl, 10 mM NaCl, 1 mM MgSO4, 5 mM NaHCO3, 1 mM NaH2PO4, pH 7.0)
and then resuspended in H50 at a 2 x 107 cells/ml. Cell samples (100 µl each) were mixed
with ~5 µg of plasmid DNA and incubated for 5 min on ice in 1-mm cuvettes.
Electroporation was carried out by two consecutive pulses of 0.85 kV with a capacitance of
25 mF applied to the cuvette with a 5-sec recovery between pulses. After 5 min of incubation
on ice, the cells from each cuvette were deposited on to a six well tissue culture plate
containing 3 ml of HL5 (0.5% Difco proteose peptone No. 2, 0.5% BBL Thiotone E peptone,
1% glucose, 0.5% yeast extract, 2.5 mM KH2PO4, 2.5 mM Na2HPO4, pH 6.7) in each well
(Pang et al., 1999). The cells became adherent and transfectants were then selected by
sequential incubation in 2, 5, 10 and 20 μg/ml G418 (Sigma Chemical Co., St. Louis, MO).
Immunofluorescence labeling of cells and laser scanning confocal microscopy

A
…ATG GAT GGT GAA GAT GTT Caa gct tCT GTT…………………C TTT AAC TCT
GAA AAA gaa ttc ATG
Act15 Start codon cadA coding regionHindIII
A
GAA AAA gaa ttc ATG ….
EcoRI GFP coding region
B
Name Sequence
DdCAD-1-GFP Forward 5’ -> 3’ (HindIII)GGC Caa gct tCT GTT GAT GCA AAT AAA G
Reverse 5’ -> 3’ (EcoRI)CG tt TTT TTC AGA GTT TAA GTT AAA GCG gaa ttc TTT TTC AGA GTT TAA GTT AAA G
N-GFP Forward 5’ -> 3’ (HindIII)- same as DdCAD-1-GFPGGC Caa gct tCT GTT GAT GCA AAT AAA G
Reverse 5’ -> 3’ (EcoRI)CG gaa ttc TCC TGG TAA GAC TTG GAA CTT TGA AAG ACC
C-GFP Forward 5’ -> 3’ (HindIII)GGT Caa gct tTT CAA TGG GCA GTT GAT G
Reverse 5’ -> 3’ (EcoRI)- same as DdCAD-1-GFPCG gaa ttc TTT TTC AGA GTT TAA GTT AAA G
Figure 3.1 Construction of DdCAD-1-GFP, N-GFP and C-GFP Plasmids for Cell
Transfection. (A) Schematic diagram for the multiple cloning site of the GFP constructs.
(B) Table listing primers used for constructing the GFP fusion proteins.
124

125
Transfected cells expressing DdCAD-1-GFP, N-GFP, C-GFP and GFP were
developed in 17 mM phosphate buffer, pH 6.4, for 1-3 h at 2 x 107 cells/ml. Then, cell
samples (1 x 106 cells) were allowed to attach on positively charged coverslips (Fisher
Scientific, Pittsburgh, PA, cat. no. 12-545-84) for 30-60 min. Cells were then fixed in 4%
paraformaldehyde (PFA), 50% HL-5 and 0.1% DMSO for 30 min at room temperature. The
cells were permeabilized with 0.5% Triton X-100 and 1% PFA for 30 min (Heuser et al.,
1993; Zhu and Clarke, 1992). Nonspecific binding was blocked by incubation with 1% BSA
in MCG buffer (50 mM MES, pH 6.4, 0.2 mM CaCl2, 2 mM MgCl2) for 30 min. Cells were
labeled with the mouse anti-calmodulin (CaM) mAb 6D4 (Sigma Chemical Co., St. Louis,
MO) (1:200 dilution in MCG containing 0.1% BSA) for 1 h, washed three times with TBS
containing 0.1% Tween-20, and then stained with Alexa-568-conjugated goat anti–mouse
antibodies (1:400 dilution) (Molecular Probes, Eugene, OR) for 1 h. Coverslips were
mounted in DAKO fluorescent mounting medium (DakoCytomation, Glostrup, Denmark).
Confocal images were acquired using the Zeiss LSM 510 microscope.
For live cell imaging, axenically grown transfected cells were suspended in 17 mM
Na/Na2 phosphate buffer at ~2 x 105 cells/ml and 100 µl was deposited on a concavity glass
slide for 30 to 60 min. The medium was then replaced with 8.5 mM Na/Na2 phosphate buffer
containing the styryl dye FM4-64 (Molecular Probes) at 1 mg/ml. Images were recorded
between 5 and 20 min after staining to visualize both the CV and the plasma membrane.
Time-lapse sections were collected on the Zeiss LSM510 microscope equipped with a 100X
water immersion objective. Images were processed using the Zeiss LSM Image Browser
Software (Version 4.0.0.157).
Isolation of contractile vacuoles and cytosol

126
Contractile vacuoles were isolated according to Sesaki et al. (Sesaki et al., 1997) with
minor modifications. Cells (2 x 109) were developed for 6 h in suspension and then
homogenized in 12 ml of TM buffer (2 mM MgCl2, 10 mM Tris-HCl, pH 7.5) at room
temperature. Samples (5 ml each) were layered on top of discontinuous (28% and 48%
(w/w), 4 ml each) sucrose density gradients and then centrifuged for 1 h at 40,000 rpm at 4ºC
in an SW40 rotor. Contractile vacuoles enriched at the interface were collected and aliquots
were stored at -70ºC. Also, cytosol derived from cadA-null cells was obtained after
centrifugation of the cell lysate at 40,000 rpm and aliquots were stored at -70ºC. Protein
concentration was determined using the bicinchoninic acid assay kit (Pierce Chemical Co.,
Rockford, IL). Proteolytic digestion of contractile vacuoles was carried out by incubating
samples (containing ~100 µg protein) with 0.01 mg/ml of proteinase K in the presence or
absence of 0.05% SDS at 37ºC for 1 h. After the addition of 2 mM phenylmethylsulfonyl
fluoride, samples were boiled for 10 min and subjected to SDS-PAGE, followed by western
blot analysis using either rabbit antibodies against DdCAD-1 (32) or rabbit antibodies against
GFP (Molecular Probe).
Expression of His6-tagged mutant DdCAD-1 proteins
His-tagged wild-type DdCAD-1 (His6-WT), N and C domains (His6-N, His6-C) were
expressed and purified as soluble native proteins as described previously (Lin et al., 2006).
The Ca2+-binding site mutants were created using site-directed mutagenesis by overlapping
extension PCR method to substitute two to three residues with alanine in each Ca2+-binding
site (SI, SII, and SIII): His6-SI(D39A, T81A), His6-SII(D35A, E56A), His6-SIII(E59A,
D61A) and His6-S(I+II)(D35A, N38A, D39A). The primers used to generate these constructs
are shown in Fig. 3.2. All constructs were sequenced to ascertain sequence fidelity. Protein

NAME SEQUENCE
SI(D39A,T81A)
D39A Forward:5’ ----GGT GAT AAA TGG AAT GCT AAA TTC ATG TCA TGT TTG GTT GG--- 3’ D39A Reverse:5’ ----CC AAC CAA ACA TGA CAT GAA TTT AGC ATT CCA TTT ATC ACC--- 3’ T81A Forward:5’---- GGC AGT ACA AAC AAT GAT TTA GCC TCA ATA AAT GGT CTT TC --- 3T81A Reverse: 5’---- GA AAG ACC ATT TAT TGA GGC TAA ATC ATT GTT TGT ACT GCC ---3’
SII (D35A, E56A)
D35A Forward: 5’ ---GGT GAA ACT GTA AGA TTC AAC AAT GGT GCT AAA TGG AAT GAT AAA TTC --- 3’D35A Reverse:5’--- GAA TTT ATC ATT CCA TTT AGC ACC ATT GTT GAA TCT TAC AGT TTC ACC --- 3’E56A Forward:E56A Forward:5’------ GTT AGA TGT AAC ATT TGG GCG CAT AAT GAA ATT GAT ACT CC ----3’ E56A Reverse:5’------ GG AGT ATC AAT TTC ATT ATG CGC CCA AAT GTT ACA TCT AAC ---3’
SIII (E59A,D61A)
E59A, D61A Forward: 5’----- C ATT TGG GAG CAT AAT GCA ATT GCT ACT CCA ACT CCA GG ---- 3’ E59A D61A RD61A) E59A, D61A Reverse:5’----- CC TGG AGT TGG AGT AGC AAT TGC ATT ATG CTC CCA AAT G ---- 3’
S(I+II) (D35A,N38A, D39A)
D35A, N38A, D39A Forward: 5’ ---GA TTC AAC AAT GGT GCT AAA TGG GCT GCT AAA TTC ATG TCA TG ---3’D35A, N38A, D39A Reverse:5’--- CA TGA CAT GAA TTT AGC AGC CCA TTT AGC ACC ATT GTT GAA TC ---3’3
W37A W37A Forward:5’- GGT GAA ACT GTA AGA TTC AAC AAT GGT GAT AAA GCG AAT GAT AAA TTC ATG TCA TG -3’W37A Reverse:5’-- CA TGA CAT GAA TTT ATC ATT CGC TTT ATC ACC ATT GTT GAA TCT TAC AGT TTC ACC - 3’
W55A W55A Forward:5’-- GGT TCA AAT GTT AGA TGT AAC ATT GCG GAG CAT AAT GAA ATT GAT ACT CC – 3’W55A Reverse:5’-- GG AGT ATC AAT TTC ATT ATG CTC CGC AAT GTT ACA TCT AAC ATT TGA ACC – 3’
Fi 3 2 Li f i d h C 2+ bi di i i dFigure 3.2 List of primers used to create the Ca2+-binding site mutations and
Tryptophan mutations. Codons with base substitutions are shown in bold fonts.
127

128
expression was carried out in the E. coli strain BL21(DE3) and His-tagged proteins were
purified using Ni-NTA resin (Qiagen Inc., Valencia, CA).
In vitro reconstitution of DdCAD-1 import into contractile vacuoles
Contractile vacuoles and cytosol fractions derived from cadA-null cells were mixed at
1:1 ratio in terms of protein amount in a 600-µl reaction sample to give a final concentration
of 2 mg/ml. His6-DdCAD-1 and the different mutant forms of DdCAD-1 were added to a final
concentration of 2 µM and the mixture was incubated for 1 h at room temperature. The
contractile vacuoles were pelleted at 12000 rpm for 10 min at 4ºC, washed twice with TM
buffer, and subjected to proteinase K (10 µg/ml) digestion for 1 h at 37ºC in the presence or
absence of 0.1% SDS. Samples were subjected to SDS-PAGE and western blot analysis.
45Ca2+-overlay Assay
45Ca2+-overlay assays were carried out as described previously (Lin et al., 2006).
Recombinant DdCAD-1 proteins were blotted onto nitrocellulose membrane using a slot blot
apparatus (Bio-Rad, Hercules, CA). The membrane was washed for 1 h in four changes with
the overlay buffer (60 mM KCl, 5 mM MgCl2, and 10 mM imidazole, pH 6.8). The blot was
then incubated with 10 ml of 45Ca2+ (10 µCi/ml) in the overlay buffer for 30 min at room
temperature on a platform shaker, followed by three 10-min washes with deionized water.
Autoradiography was carried out by exposure of the air-dried membrane to Bioflex-MRI
Films (Clonex Corp, AR) for 24 h at -70ºC.
Antibody-induced Cap Formation

129
Cells (2x107 cells/ml) were developed in liquid medium and collected at 3 h. Anti-
GFP antiserum (1:100 dilution) was added to 1.2 x 106 cells suspended in 300 µl of 50 mM
MES buffer, pH 6.3, and incubated for 30 min at room temperature. Alexa-568-conjugated
goat anti-mouse antibody was added at 1:400 dilution and rotated at room temperature for
another 30 min. Next, 300 µl of cells were deposited on a positively charged cover slip
(Fisher Scientific, Pittsburgh, PA) and allowed to attach for 15 min. The coverslips were
washed gently with MCG buffer, fixed with 3.7% formaldehyde and mounted for
fluorescence microscopy.
Analysis of DdCAD-1 Secretion - Axenically grown cadA-null transfectants (2 x 107 cells/ml)
were developed in 17 mM phosphate buffer for 6 h at room temperature. During
development, 1-ml cell samples were collected every hour and the cells were pelleted at
15,000g for 1 min to obtain a clear supernatant. Aliquots of the supernatant were mixed with
sample buffer (1:1) and prepared for SDS-PAGE, followed by western blot analysis.
Chemical cross-linking of DdCAD-1
His6-DdCAD-1 (30 µg) was cross-linked in the presence of 1 mM disuccinimidyl
suberate (DSS) for 30 min at room temperature. The excess DSS in the reaction sample was
quenched with 50 mM Tris-HCl, pH 7.6. The cross-linked His6-DdCAD-1 was subjected to
the contractile vacuole import assay.

130
3.4 Results
Import of DdCAD-1-GFP via invagination of vacuolar membrane
To facilitate the in vivo analysis of DdCAD-1 transport, cadA-null cells were
transfected with a DdCAD-1-GFP construct (Fig. 3.3A). Transfectants were selected using
G418 and by direct visualization under a fluorescence microscope. The expression of
DdCAD-GFP was confirmed by probing protein blots with antibodies against DdCAD-1 and
GFP (Fig. 3.3B). Both antibodies detected a protein band corresponding to the expected size
of the fusion protein. Transfected cells showed uniform green fluorescence in the cytoplasm.
In order to determine whether the fusion protein was imported by contractile vacuoles, cell
samples were fixed and permeabilized to induce partial loss of the cytoplasmic GFP fusion
protein. Some samples were also stained with anti-calmodulin antibodies. Calmodulin serves
as a marker of contractile vacuoles because it is associated with their cytoplasmic surface
(Zhu et al., 1993a). Confocal microscopy showed that an abundance of DdCAD-1-GFP
associated with the contractile vacuole network and the cortical region (Fig. 3.3C). Notably,
DdCAD-1-GFP was present in the lumen of contractile vacuoles, where calmodulin staining
was absent.
Confocal images of fixed specimens of DdCAD-1-GFP transfectants occasionally
displayed a punctate staining pattern of DdCAD-1 inside contractile vacuoles. To examine
the nature of these stained structures, live cells were incubated with the vital dye FM4-64 to
label cellular membranes (Heuser et al., 1993), followed by time lapse microscopy. The
confocal series of a number of cells revealed novel vesicular structures inside their contractile
vacuoles (Fig. 3.4, A&B). These vesicles adopted either a tubular or rounded appearance and
were often filled with DdCAD-1-GFP. The membrane of these vesicles was marked by FM4-

BA
270 bp 369 bp 720 bpAct15 promoter
DdCAD-1-GFP
GFPCN
kDa
51GFPCN
C α DdCAD-1
4138
26
α GFP
C MDdCAD 1 GFP M
Figure 3.3 Construction and expression of DdCAD-1-GFP in cadA-null cells. (A)
Schematic drawing of the DdCAD-1-GFP construct. (B) Western blots of cadA-null cells
transfected with plasmid DNA. Cell lysates of stable G418-resistant clones and parental
CaMDdCAD-1-GFP Merge
transfected with plasmid DNA. Cell lysates of stable G418 resistant clones and parental
cells (cadA-) were prepared from cells at 3 h of development for SDS-PAGE and protein
blots, which were probed with rabbit antisera against either DdCAD-1 or GFP. The
expected size of the fusion protein is indicated by an arrowhead. The lower molecular
weight band probably represents a degraded product. Molecular weight markers are
shown on the left. (C) Confocal micrographs showing the association of DdCAD-1-GFP
with the contractile vacuole network. Cells were collected at 3 h of development, fixed in
4% PFA, and permeabilized with 0.5% Triton X-100 for 5 min. Cells were labeled with
mouse anti-calmodulin (CaM) mAb (red) and subjected to laser scanning confocal
microscopy. Arrows indicate contractile vacuoles. Bar, 10 µm.
131

A
0 s 54 s 108 s 162 s
5 µm5 µm5 μm
BB
0 s 38 s 76 s 114 s
5 µm
5 µm
Figure 3.4 Budding of vesicles into the lumen of the contractile vacuoles. (A and B)
Transfectants expressing DdCAD-1-GFP were collected from HL-5 medium, washed and
suspended in 17 mM phosphate buffer at ~2 x 105 cells/ml. Live cells (100 µl) were
deposited on slides for attachment. The styryl dye FM4-64 (red) was added at 1 mg/ml todeposited on slides for attachment. The styryl dye FM4 64 (red) was added at 1 mg/ml to
visualize both the contractile vacuoles and the plasma membrane. Time-lapse sequences
of confocal images were recorded between 5 to 20 min after dye addition. Confocal
images of the boxed area in the light micrograph are shown. Arrows point to membrane
protrusions in the contractile vacuole lumen, where DdCAD-1-GFP and FM4-64
colocalize. Schematic drawings of the contractile vacuole are shown below the confocal
images.
132

133
64 and showed continuity with the contractile vacuole membrane, indicating that they had
originated from the contractile vacuole. These structures were relatively transient as they
remained for only 2-3 minutes inside the lumen of contractile vacuoles and then disappeared.
After that, the GFP fluorescence in the lumen became diffuse and it was accompanied by the
loss of FM4-64 staining, suggesting that the vesicles might have pinched off from the
vacuolar membrane and ruptured to release the cargo in the lumen. Thus, membrane
invagination might serve as a pathway by which DdCAD-1 is imported into contractile
vacuoles.
Import of DdCAD-1 into contractile vacuoles requires both N- and C-terminal domains
DdCAD-1 contains two distinct domains linked by a short stretch of amino acids (Lin
et al., 2004; Lin et al., 2006). To investigate the structural requirements for its import into
contractile vacuoles, constructs containing GFP fused to either the N-terminal domain (N) or
the C-terminal domain (C) were transfected into cadA-null cells (Fig. 3.5A). As a control,
cadA-null cells were also transfected with a GFP construct. The expression of the GFP fusion
proteins was confirmed by protein blot analysis (Fig. 3.5B). Antibodies against DdCAD-1
and GFP detected protein bands corresponding to the expected molecular size of the
respective fusion protein. Transfectants expressing comparable levels of fusion protein were
selected for further analysis. The two GFP fusion proteins appeared to colocalize with
calmodulin to the periphery of contractile vacuoles, while GFP showed a more diffuse
staining pattern in the cytoplasm (Fig. 3.5C). In all three types of transfectants, green
fluorescence was not detected inside contractile vacuoles, suggesting that neither the N-
terminal domain nor the C-terminal domain alone was sufficient for import into contractile
vacuoles.

A B
N-GFP
C-GFP
GFP GFP
GFPN
GFPC51
kDa
4138
26
C
α DdCAD-1 α GFP
CaMN-GFP Merge
C-GFP CaM Merge
GFP CaM Merge
Figure 3.5 Expression of N-GFP and C-GFP fusion proteins in transfected cells. (A)
Schematic drawings of GFP-fusion protein constructs. (B) Western blots of transfectants
probed with antibodies against DdCAD-1 and GFP. (C) Cells were collected at 3 h of
d l d h fi d d l b l d f f l i A i di
GFP CaM Merge
development and then fixed and labeled for confocal microscopy. Arrows indicate
contractile vacuoles. Bars, 10 µm.
134

135
Surface expression and secretion of DdCAD-1-GFP
Our previous work show that DdCAD-1 binds to an anchoring protein on the luminal
surface of contractile vacuoles for presentation on the cell surface, while the unbound protein
is targeted for secretion (Sesaki et al., 1997; Sriskanthadevan et al., 2007). To determine
whether N-GFP and C-GFP were capable of reaching the cell membrane independent of the
contractile vacuole route, live cells were treated with antibodies against GFP to induce
antigen clustering, which would eventually lead to the formation of “caps.” Caps were only
observed in transfectants expressing DdCAD-1-GFP, indicating that it was associated with the
cell surface (Fig. 3.6A). In contrast, neither N-GFP nor C-GFP was detectable on the surface
of their respective transfectants.
To determine whether fusion proteins were secreted into the medium, cells were
incubated in phosphate buffer and samples were collected at hourly intervals. After the
removal of cells, the supernatants were subjected to SDS-PAGE and western blot analysis.
Among the three fusion proteins, only DdCAD-1-GFP was found in the conditioned medium
(Fig. 3.6B). Neither N-GFP nor C-GFP was detected in the medium. As a positive control,
protein blots were probed with antibodies against the endogenous lectin discoidin-I, which is
also targeted for secretion through the contractile vacuole. Discoidin-I was found in the
conditioned media of all three transfectants (Fig. 3.6B), indicating that this unconventional
secretory pathway functioned normally in these transfectants.
In vitro import analysis of GFP-fusion protein
To further investigate the import of DdCAD-1-GFP, contractile vacuoles were isolated
from transfectants and then subjected to western blot analysis. Consistent with our
morphological observation, DdCAD-1-GFP, N-GFP and C-GFP co-purified with contractile

A
N-GFP C-GFP GFPDdCAD-1-GFP
B
5 μm
α DdCAD 1 α Discoidin I
26
26
64 49
49 37
α DdCAD-1 α Discoidin-I
DdCAD-1-GFP transfectant
N-GFP transfectant
kDa kDa
2649 37
C-GFP transfectant
0 1 2 30 1 2 3Hours of development
Figure 3.6 Differential cell surface expression and secretion of the DdCAD-1 fusion
proteins in transfectants. (A) fluorescence micrographs showing antibody induced cap
formation of DdCAD-1-GFP on live cells. cadA-null cells expressing the different GFP
fusion proteins were incubated with anti-GFP antibodies for 30 min. After washing, cells
were incubated at room temperature for another 30 min with a secondary antibody to
induce ‘cap’ formation (red). The corresponding light micrographs are shown in the
lower panels. Bars, 5 µm. (B) DdCAD-1 secretion during development. Transfectants
were developed in 17 mM phosphate buffer at 2 x 107 cells/ml and the conditioned media
were collected at 1 h intervals for western blot analysis using rabbit antibodies against
DdCAD 1 or discoidin I The arrowheads indicate the position of N GFP (38 kDa) andDdCAD-1 or discoidin-I. The arrowheads indicate the position of N-GFP (38 kDa) and
C-GFP (41 kDa), respectively.
136

137
vacuoles (Fig. 3.7A). If fusion proteins were present in the lumen, they should be protected
from protease digestion unless the vacuolar membrane was disrupted by detergent. To test
this, contractile vacuoles isolated from different transfectants were subjected to proteinase K
digestion in the presence or absence of SDS, followed by protein blot analysis (Fig. 3.7A).
As a positive control, blots were probed with antibodies against discoidin-I. Whereas N-GFP
and C-GFP were completely degraded in the absence of SDS, DdCAD-1-GFP and discoidin-I
were reduced by only 40-50%, indicating that a substantial amount of these two proteins were
protected from proteolysis. In the presence of detergent, both DdCAD-1 and discoidin-I were
reduced by >90% (Fig. 3.7A). The data provided biochemical evidence demonstrating that
only DdCAD-1-GFP and discoidin-I were able to enter the contractile vacuole.
A cell-free reconstitution assay was employed to examine the selective import of
DdCAD-1 into contractile vacuoles. His6-tagged fusion proteins were used in these assays to
avoid potential steric hindrance due to the bulky GFP. Contractile vacuoles and cytosol were
isolated from cadA-null cells and then incubated with one of the three fusion proteins, His6-
DdCAD-1, His6-N and His6-C. At the end of the incubation period, the contractile vacuoles
were washed and then subjected to proteinase K digestion in the presence or absence of
detergent. Western blot analysis showed that although all three proteins bound to contractile
vacuoles, only His6-DdCAD-1 was protected from proteolysis in the absence of detergent,
indicating the selective import of His6-DdCAD-1 (Fig. 3.7B). The blots were also probed
with antibodies against calmodulin. Calmodulin was digested completely in the absence of
detergent, demonstrating the effectiveness of proteinase K in the removal of proteins
associated with the surface of these vacuoles (Fig. 3.7B).
Since both His6-N and His6-C were able to bind contractile vacuoles, an important
question was whether they would compete for the DdCAD-1 binding sites on contractile

Figure 3.7 Import of DdCAD-1 fusion proteins into contractile vacuoles. (A)
Association of GFP fusion proteins with purified contractile vacuoles. Transfectants were
developed for 6 h in liquid culture and then homogenized. The post-nuclear supernatant
was fractionated on a 28% and 48% step sucrose density gradient by centrifugation at
40,000 rpm for 1 h at 4ºC. Contractile vacuoles banded in the interface were collected
and subjected to digestion with proteinase K in the presence or absence of 0.05% SDS for
1 h at 37ºC. Panels a, b and c were probed with rabbit antibody against GFP (Molecular
Probe). Panel d was probed with rabbit antibody against discoidin-I, which served as aProbe). Panel d was probed with rabbit antibody against discoidin I, which served as a
positive control. (B) In vitro import of His-tagged DdCAD-1 fusion proteins into
contractile vacuoles. Contractile vacuoles (2 mg/ml) and cytosol (2 mg/ml) fractions
derived from cadA-null cells were mixed together in 600 µl reaction volume. His6-
DdCAD-1, His6-N or His6-C was added to a final concentration of 2 μM and the assay
was carried out as described in Experimental Procedures. Samples were subjected to
SDS-PAGE and the protein blots were probed with rabbit antibodies against either
DdCAD-1 (29) (upper panels) or anti-calmodulin mAb (Sigma) (lower panels). The
arrows indicate the position of the His-tagged proteins. A higher molecular weight band
representing the dimer form of His6-DdCAD-1 and His6-N samples was frequently
observed. (C) In vitro import assays of the His6-DdCAD-1 in the presence of the
individual domains. His6-N or His6-C (4 µM) was added to the import assay samples the
mixture was incubated at room temperature for 30 min. Then, His6-DdCAD-1 was added
to a final concentration of 2 µM and the mixture was incubated for 1 h at room
temperat re The contractile ac oles ere pelleted and s bjected to proteinase Ktemperature. The contractile vacuoles were pelleted and subjected to proteinase K
digestion in the presence or absence of detergent and the protein blots were probed with
rabbit anti-DdCAD-1 antibody.
138

A
B
C
139

140
vacuoles. Import assays were carried out for His6-DdCAD-1 in the presence of a 2-fold
higher concentration of either His6-N or His6-C (Fig. 3.7C). In all cases, comparable levels of
His6-DdCAD-1 were found inside the contractile vacuoles, although both His6-N and His6-C
bound efficiently to the contractile vacuole. The fact that they did not interfere with the
import of His6-DdCAD-1 suggests that both His6-N and His6-C bound non-specifically or to
components that are not involved in DdCAD-1 transport.
Requirement of Ca2+ in the import of DdCAD-1 into contractile vacuole
DdCAD-1 is a Ca2+-binding protein which mediates cell-cell adhesion via homophilic
binding in a Ca2+-dependent manner (Lin et al., 2006; Wong et al., 1996). To assess the role
of Ca2+ in DdCAD-1 transport, the import assay was performed in the presence of EGTA.
DdCAD-1 import was reduced by ~80% in the presence of 5 mM EGTA (Fig. 3.8A). The
inhibitory effects of EGTA could be reversed by the addition of Ca2+. Furthermore, the
efficiency of DdCAD-1 import was enhanced as the Ca2+ level increased (Fig. 3.8A). In
contrast, Ca2+ did not have any effect on His6-N in the import assay.
DdCAD-1 consists of three Ca2+-binding pockets (Lin et al., 2006). Site I (SI)
involves the residues Asp39, Thr81, Asn84 and Phe41 in the N-terminal domain, while Site II
(SII) consists of Asp35, Asn38, Glu56 and Ser87. Site III (SIII) is unique as it is situated at
the interface of the two domains, comprising Glu59 and Asp61 from the N-terminal domain
and Thr179, Gln181 and Asn202 from the C-terminal domain. To further assess the
requirement of Ca2+ in DdCAD-1 transport, mutations were introduced in the Ca2+-binding
sites of DdCAD-1, with SI mutant containing D39A, T81A, SII mutant D35A, E56A, SIII
mutant E59A, D61A, and S(I+II) mutant D35A, D39A, N38A. His-tagged mutant proteins
were expressed and purified from bacteria (Fig. 3.8B). Their ability to bind Ca2+ was

Figure 3.8 Role of Ca2+ on DdCAD-1 import into contractile vacuole. (A) Effects of g p ( )
EGTA on DdCAD-1 import into CV. DdCAD-1 import assays were performed in the
presence of 5 mM EGTA plus different concentrations of CaCl2. Samples were subjected
to SDS-PAGE and protein blots were probed with anti-DdCAD-1 antibody. In a separate
experiment, import of His6-N was tested in the presence of 5 mM EGTA and 10 mM
C Cl Th i di h i i f Hi DdCAD 1 hil h h d i diCaCl2. The arrow indicates the position of His6-DdCAD-1, while the arrowhead indicates
the position His6-N. (B) Coomassie stained gel of purified His-tagged wild-type (WT) and
mutant DdCAD-1 containing mutations in one of the Ca2+-binding sites (SI, SII, SIII,
S(I+II)). (C) Binding of 45Ca2+ to His6-DdCAD-1 and Ca2+-binding site mutant DdCAD-
1. Protein samples were blotted onto nitrocellulose membrane. The blot was incubated
with 10 ml of 45Ca2+ (10 μCi/ml) for 30 min at room temperature. After washing
extensively with water, the blot was exposed to X-ray film at –70ºC for 24 h. (D) In vitro
import of His-tagged wildtype (WT) and mutant (SI, SII, SIII and S(I+II)) DdCAD-1 into
contractile vacuoles. Protein blots of import assay samples were probed with anti-
DdCAD 1 antibody (E) Secretion of mutant DdCAD 1 during developmentDdCAD-1 antibody. (E) Secretion of mutant DdCAD-1 during development.
Transfectants expressing SI- or S(1+II) mutant DdCAD-1 were developed in 17 mM
phosphate buffer at 2 x 107 cells/ml and the conditioned media were collected at 2 h
intervals for western blot analysis using rabbit antibodies against DdCAD-1. The same
samples were also probed with mAb against actin. The very low level of protein detected
in the medium was probably due to cell lysis, as indicated by the actin blots.
141

A
- 5 5 5 5 5EGTA (mM)- - 1 3 5 10CaCl2 (mM)
510
His6-DdCAD-1 His6-N
B WT SI SII SIII S(I II)
- - - - - - - - - - - - + +- -- +Proteinase K
SDS- + - + - + - + - + + + +-
WT: wildtype His6-DdCAD-1SI mutations: D39A, T81A SII mutations: D35A, E56ASIII mutations: E59A, D61A S(I+II) mutations: D35A, N38A, D39A
B
C WT SI SII SIII S(I+II)0.3
nmol
WT SI SII SIII S(I+II)
D
Proteinase K - + + - + + - + +- + +- + +
WT SI SII S(I+II)SIII
0.30.10.05
SDS - - + - - + - - +- - +- - +
E pAb α GFPGFP
pAb α DdCAD-1S(I+II)-GFPSI-GFP
cell lysate
supernatant
0 2 4 60 2 4 60 2 4 6
mAb α Actin
supernatant
supernatant
Hours of development
142
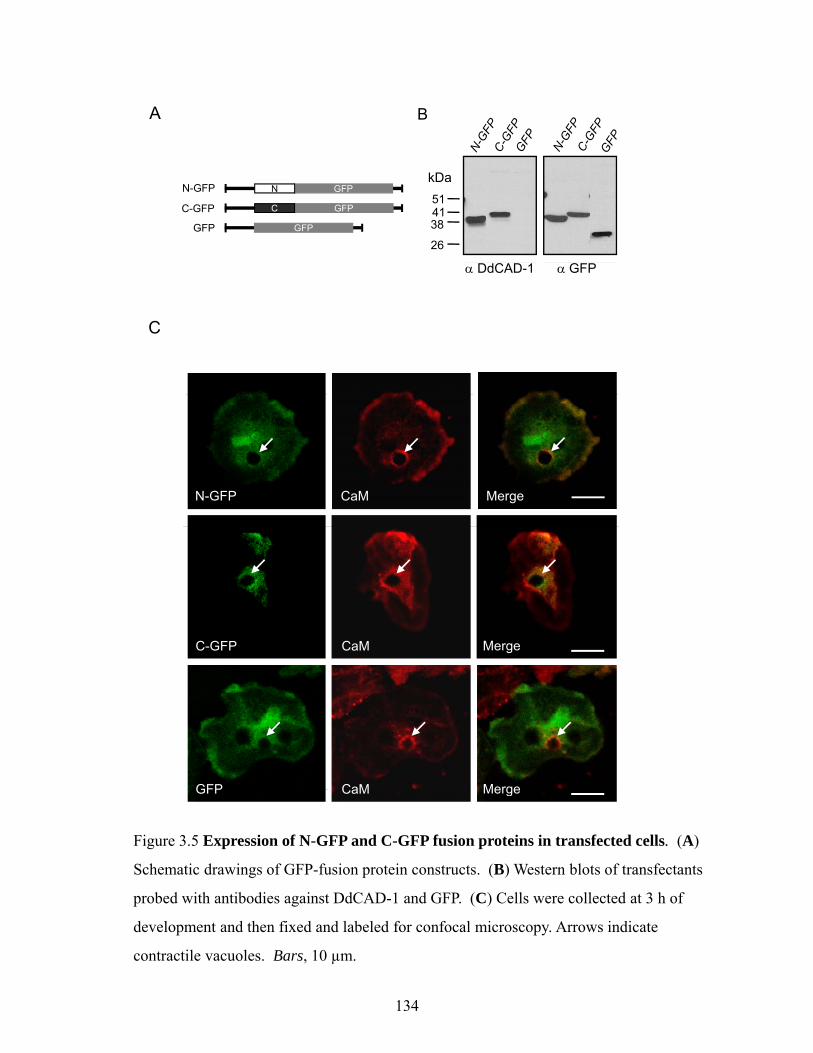
143
assessed using the 45Ca2+-overlay assay (Fig. 3.8C). His6-SI and His6-SII showed substantial
reduction in 45Ca2+-binding, while His6-SIII exhibited a similar level of bound 45Ca2+ as the
wild-type protein, indicating that the side-chains of residues Glu59 and Asp61 do not
contribute significantly to the Ca2+ binding affinity. Binding of 45Ca2+ was not observed in
the His6-S(I+II) mutant.
Import assays showed that the Ca2+-binding site mutants that displayed reduced
binding of Ca2+ failed to enter the contractile vacuole, while His6-SIII was imported into the
contractile vacuole at a level similar to that of the wild-type protein (Fig. 3.8D). Therefore,
the import of DdCAD-1 into contractile vacuoles likely depends on the function of Ca2+-
binding sites SI and SII. Consistent with this observation, cadA-null transfectants expressing
the SI and S(I+II) mutant DdCAD-1 (see Fig. 3.9) did not secrete significant amounts of these
mutant proteins (Fig. 3.8E).
Effect of conformation on the import of DdCAD-1 into contractile vacuole
CD analysis was carried out to determine whether mutations in the Ca2+-binding sites
had any effect on the conformation of DdCAD-1, since loss of Ca2+ binding could lead to
subtle conformational changes which might in turn influence the import process. A
comparison between the CD spectra of the Ca2+-binding impaired mutant His6-SII and Ca2+-
binding competent mutant His6-SIII showed a loss of the positive peak between 220 nm and
230 nm in the His6-SII spectra (Fig. 3.10). In contrast, the His6-SIII mutant, which showed
no impairment in its import into contractile vacuoles, exhibited a CD spectrum similar to that
of the wild-type protein (Fig. 3.10). Since aromatic residues are known to contribute to the
positive signal in the far UV region of CD spectra (Andersson et al., 2001; Clark et al., 1996),
the data suggest that the mutations in SI, SII, and S(I+II) might have caused conformation

SI-GFP CaM Merge
S(I+II)-GFP CaM Merge
Figure 3.9 Confocal images of transfectants expressing DdCAD-1 with different Ca2+-
binding site mutations. Cells were collected at 3 h of development and then fixed and
labeled for confocal microscopy. Calmodulin (CaM) (red) demarcate the contour of
contractile vacuoles (arrows). Bars, 10 µm.
144

10
20
30
y(m
deg)
-40
-30
-20
-10
0
SIII: E59A, D61ASII: D35A, E56AMol
ecul
ar E
llipic
ity
-50190 200 210 220 230 240 250 260 270
Wavelength (nm)
Figure 3.10 Conformational changes in Ca2+-binding mutants of DdCAD-1. The CD
spectra of 40 µM His6-SII(D35A, E56A) (purple) and His6-SIII(E59A, D61A) (blue)
were plotted together for comparison. The open arrow indicates loss of the positive peak
between 220 and 230 nm.
145

146
perturbations around Trp37, which is situated close to the Ca2+-binding sites SI and SII (Lin et
al., 2006).
To investigate whether conformation plays a role in DdCAD-1 import, we first
examined the import of denatured His6-DdCAD-1 and a His6-tagged mutant DdCAD-1
containing five mutations (N58A, E59A, D61A, D201A, and N202A), which displayed severe
distortions in secondary structure (see Fig. 3.11F). The results showed that neither of them
was able to enter the contractile vacuole (Fig. 3.12A). Next, we used a His6-DdCAD-1
mutant protein with Trp-37 substituted with Ala (W37A) since it is situated close to SI and
SII. The CD spectrum of His6-DdCAD-1 (W37A) showed a reduced positive peak between
220 nm and 230 nm similar to those of the SI and SII mutant proteins (see Fig. 3.11, panels
A,B and E). Import assays showed that the W37A mutant protein failed to enter the
contractile vacuole (Fig. 3.12B). As a control, Trp-55 which is situated further from the SI
and SII was substituted with Ala. The W55A mutation did not cause any adverse effect in
import assays, though it showed similar changes in the CD spectra (Fig. 3.11G). Therefore,
the integrity of the SI and SII Ca2+-binding sites are crucial to DdCAD-1 import.
Our previous studies suggest that W37 is involved in the homophilic interactions of
DdCAD-1 (Lin et al., 2006). This observation suggested the possibility that DdCAD-1 might
be imported into contractile vacuoles as a dimer. To test this hypothesis, we cross-linked
His6-DdCAD-1 with disuccinimidyl suberate before the import assay. Import studies showed
that a substantial level of dimer and cross-linked oligomers were able to enter the vacuole
(Fig. 3.12C). However, the monomeric form of DdCAD-1 appeared to be imported more
efficiently.

A
eg)
B
Mol
ecul
ar E
llipic
ity(m
deM
ar E
llipi
city
(mde
g)
C D
E
ty(m
deg)
Mol
ecul
a
F
Mol
ecul
ar E
llipi
cit
G H
)
WTW55A
Ca2+ free WTCa2+ bound WT
lecu
lar E
llipi
city
(mde
g)
Wavelength (nm) Wavelength (nm)
Mol
Figure 3.11 Conformational changes in DdCAD-1 mutants. The CD spectra of Ca2+-
binding site mutants of DdCAD-1 (A to D) and three other mutant forms of DdCAD-1 (E
to G) were plotted with the spectra of wildtype protein for comparison. CD spectra of the to G) we e p otted w t t e spect a o w dtype p ote o co pa so . C spect a o t e
Ca2+-bound and Ca2+-free forms of DdCAD-1 are shown in panel H for comparison.
147
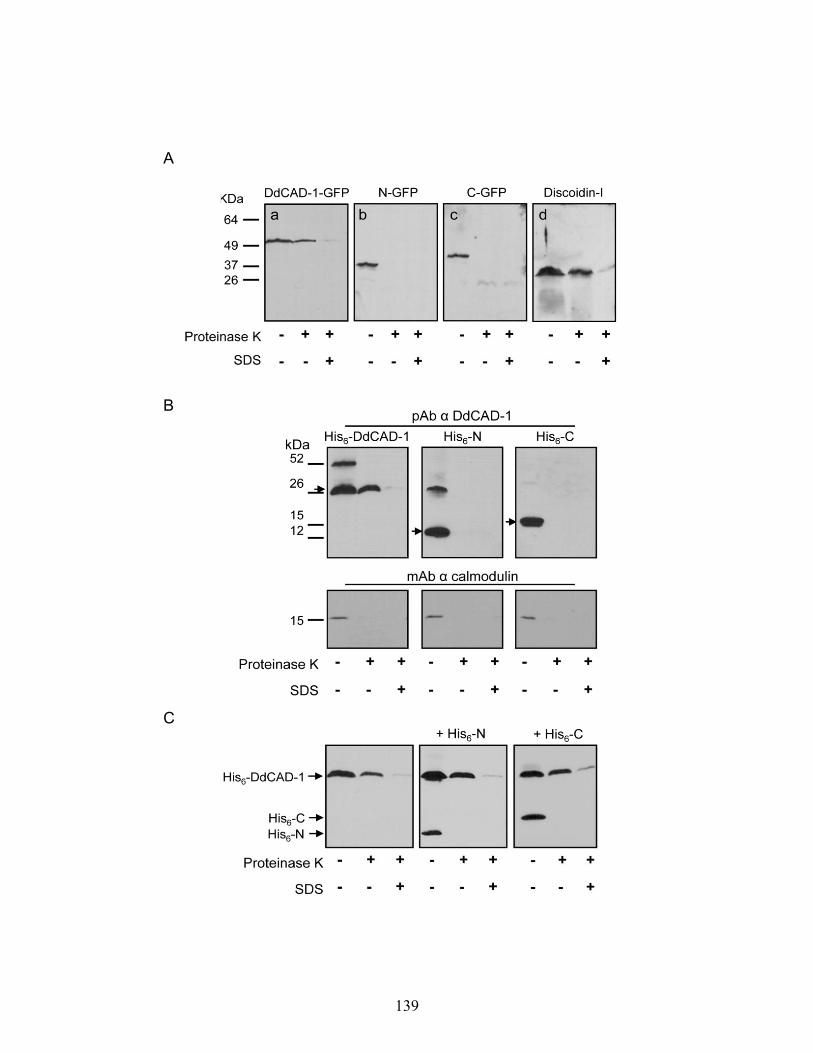
ADenatured DdCAD-1 Penta-mutation of
- + +Proteinase K
DdCAD-1 DdCAD-1
- + + - + +
B
- - +SDS - - + - - +
DdCAD-1 W37A W55A
C DSS
kDa
- + - +
- + +- - +
Proteinase K
SDS
- + +- - +
- + +- - +
644937
2619
kDa11282
dimer
monomer
Figure 3.12 Effects of DdCAD-1 conformational changes on the import process. (A)
His6-DdCAD-1 was denatured by boiling for 10 min followed by quick cooling on ice.
The denatured form of His6-DdCAD-1 and the penta-mutant (N58A, E59A, D61A,
Proteinase KSDS -
+--
-+
--
6
D201A, and N202A) protein were subjected to the in vitro import assay. (B) Effects of
the tryptophan mutations, W37A and W55A, on DdCAD-1 import into contractile
vacuoles were examined. Protein blots were probed with anti-DdCAD-1 antibody. (C)
Import of DdCAD-1 dimers into contractile vacuoles. His6-DdCAD-1 was cross-linked
i di i i id l b t (DSS) d th bj t d t th i t P t i bl tusing disuccinimidyl suberate (DSS) and then subjected to the import assay. Protein blots
were probed with anti-DdCAD-1 antibody.
148

149
3.5 Discussion
In Dictyostelium, the contractile vacuoles serve as the major vehicle for the transport
of DdCAD-1 to the plasma membrane for either secretion or cell surface presentation. Our in
vivo analysis using time-lapse microscopy has revealed transient membrane budding into the
lumen of contractile vacuoles. DdCAD-1 docked on the contractile vacuoles is mobilized to
fill these invaginations, which are then pinched off to become vesicles inside the lumen.
DdCAD-1 is released upon the burst of these vesicles. In many ways, the export of DdCAD-1
via contractile vacuole is similar to the budding of vesicles in yeast vacuoles (Muller et al.,
2000) or the formation multivesicular bodies in murine macrophages (Qu et al., 2007a). In
yeast, the tubular structures facilitate that lateral sorting of proteins and lipids into vesicles
inside the vacuole (Muller et al., 2000). In macrophages, the multivesicular bodies are the
primary vehicle for the transport of IL-1β trapped inside the exosomes (Qu et al., 2007a).
However, both vesicles and tubules are only transient structures in the lumen of contractile
vacuoles in Dictyostelium cells and multivesicular structures are rarely observed. It is
possible that the water collected in the contractile vacuole creates a hypo-osmotic
environment that can cause the rapid rupture of vesicles soon after they bud off into the
lumen.
In order to further investigate the structural mechanisms involved in DdCAD-1 import
into contractile vacuoles, individual domains were transfected into cadA-null cells. Both in
vivo and in vitro analyses show that neither the N-terminal domain nor the C-terminal domain
of DdCAD-1 alone is sufficient for import into the contractile vacuole. Since the docking of
His6-DdCAD-1 to contractile vacuoles is not affected by the presence by either His6-N or
His6-C, the import signal is likely integrated in the 3-dimensional structure of DdCAD-1.

150
Further analysis has revealed a role for Ca2+ since the import of DdCAD-1 is
abrogated by EGTA. Mutations that disrupt the Ca2+-binding capability of DdCAD-1 also
inhibit its import. Also, Ca2+ enhances the binding of DdCAD-1 to contractile vacuoles as
well as its entry into the lumen. DdCAD-1 is a Ca2+-binding protein. However, the Ca2+
binding affinity of DdCAD-1 is relatively low (Kd 12-15 µM) (Lin et al., 2006). Since the
intracellular free Ca2+ concentration is estimated to be ~56 nM in aggregation-competent cells
(Yumura et al., 1996), most of the intracellular DdCAD-1 probably exists in the Ca2+-free
form. On the other hand, contractile vacuoles are enriched in Ca2+-pumps (Moniakis et al.,
1999) and they constitute a highly efficient acidic Ca2+ store (Malchow et al., 2006). It is
conceivable that the release of Ca2+ from contractile vacuoles may serve as an attractant to
DdCAD-1. Moreover, the negatively charged surface of membranes is known to determine
the targeting of proteins with polybasic clusters (McLaughlin and Murray, 2005; Yeung et al.,
2008). The bound Ca2+ would increase the overall positive charge on the surface of DdCAD-
1. The positively charged regions as well as the His6-tag would promote electrostatic
interactions with acidic lipids and facilitate the binding of DdCAD-1 to the contractile
vacuole membrane. Structural studies have shown that Ca2+-binding interfacial membrane
proteins, such as annexin V, complex with phospholipid head groups via Ca2+-bridges
(Swairjo et al., 1995). Such a phenomenon may explain the non-specific binding of the C-
and N-terminal fusion proteins.
Since neither the C-terminal domain nor the N-terminal domain competes with
DdCAD-1 for binding to the contractile vacuole, it is likely that DdCAD-1 binds to a specific
docking element in the midst of negatively charged lipids on the vacuolar membrane. In
addition to Ca2+-pumps, an abundance of the ubiquitous Ca2+ regulator, calmodulin, is
associated with the surface of contractile vacuoles (Zhu and Clarke, 1992; Zhu et al., 1993a).

151
Calmodulin is also found on endosomal and lysosomal membranes (Enrich et al., 1996;
Nielsen et al., 1987) and is known to play a role in the endosome-mediated transport system
(Pryor et al., 2000). Interestingly, bioinformatic analysis shows that DdCAD-1 contains a
putative binding site for calmodulin in the C-terminal domain, suggesting that the Ca2+-bound
form of calmodulin may serve as a potential docking partner for DdCAD-1. Indeed, direct
interaction between DdCAD-1 and calmodulin has been detected in far western blots, while
pharmacological inhibition of calmodulin function by W7 leads to a reduction in DdCAD-1
binding to contractile vacuoles.
DdCAD-1 contains three Ca2+-binding pockets and the NMR solution structures of
DdCAD-1 have shown that binding of Ca2+ leads to more ordered side chain packing in the
Ca2+-binding sites and stabilizes the whole structure of DdCAD-1 (Lin et al., 2006). An
analysis of the CD spectra of the Ca2+-binding site mutants suggests conformational
perturbations in regions that contain tryptophan. A tryptophan (W37) is situated in the loop
structure connecting SI and SII. Substitution of W37 with alanine results in alterations in CD
spectra similar to those observed for the SI and SII mutants. The W37A mutation as well as
mutations in either the SI or SII Ca2+-binding site results in the loss of DdCAD-1 import into
contractile vacuoles, implicating a role for the 3-dimensional structure in DdCAD-1 transport.
It is possible that the Ca2+-binding sites may serve a dual function. While contributions by
Ca2+ to the surface charge can influence the docking of DdCAD-1 to contractile vacuoles,
conformational changes induced by Ca2+-binding may regulate its import into the lumen.
In both animal and plant cells, several unconventional pathways are known to utilize
transporters to import or export soluble proteins across cellular membranes (Teter and
Klionsky, 1999). Matrix components of peroxisomes are transported in folded form from the
cytoplasm into the peroxisomal matrix (Brocard et al., 2003; Nickel, 2005). Other pathways

152
that can accommodate folded substrates include the bacterial twin-arginine translocation (Tat)
pathway (Cline and McCaffery, 2007; Pugsley, 1992), the ∆pH-dependent pathway of plant
plastids (Clark and Theg, 1997), the cytoplasm-to-vacuole targeting pathway of
Saccharomyces cerevisiae (Kim et al., 1997) and the direct translocation of proteins, such as
FGF-2 and galectin-1, across the plasma membrane in mammalian cells (Schafer et al., 2004;
Seelenmeyer et al., 2005). Among them, only the peroxisomal and Tat pathways involve
targeting signals, while the others do not seem to require a generalized signal for protein
transport (Schatz and Dobberstein, 1996; Wickner and Schekman, 2005). Similarly, there has
been no targeting motif detected in the primary sequence of DdCAD-1, the import signal is
likely integrated in its 3-dimensional structure.
The export of soluble proteins may involve more than one unconventional pathway
(Nickel and Rabouille, 2009; Nickel and Seedorf, 2008b). Our previous in vitro studies
suggest that the translocation of DdCAD-1 across the contractile vacuole membrane may
involve a membrane transporter (31). Exogenously added ATP and an ATP regeneration
system enhance the import of DdCAD-1, suggesting the involvement of specific ATP-
dependent transporter(s) in the contractile vacuole membrane. Also, the import of DdCAD-1
is inhibited by 7-chloro-4-nitrobenzo-2-oxa-1,3-diazole, a specific inhibitor of vacuolar-type
H+-ATPase (Mellman et al., 1986; Padh et al., 1989), implicating a role for the V-ATPase in
the import mechanism. Among other secreted proteins in Dictyostelium, the acyl-CoA
binding protein (AcbA) which is required for the terminal differentiation of prespore cells is
also externalized through an unconventional protein secretion pathway. Interestingly, this
pathway involves the Golgi reassembly stacking protein (GRASP) and a membrane
transporter (Kinseth et al., 2007). However, the identity of the transporter for AcbA remains
to be elucidated. It is therefore evident that multiple unconventional pathways are employed
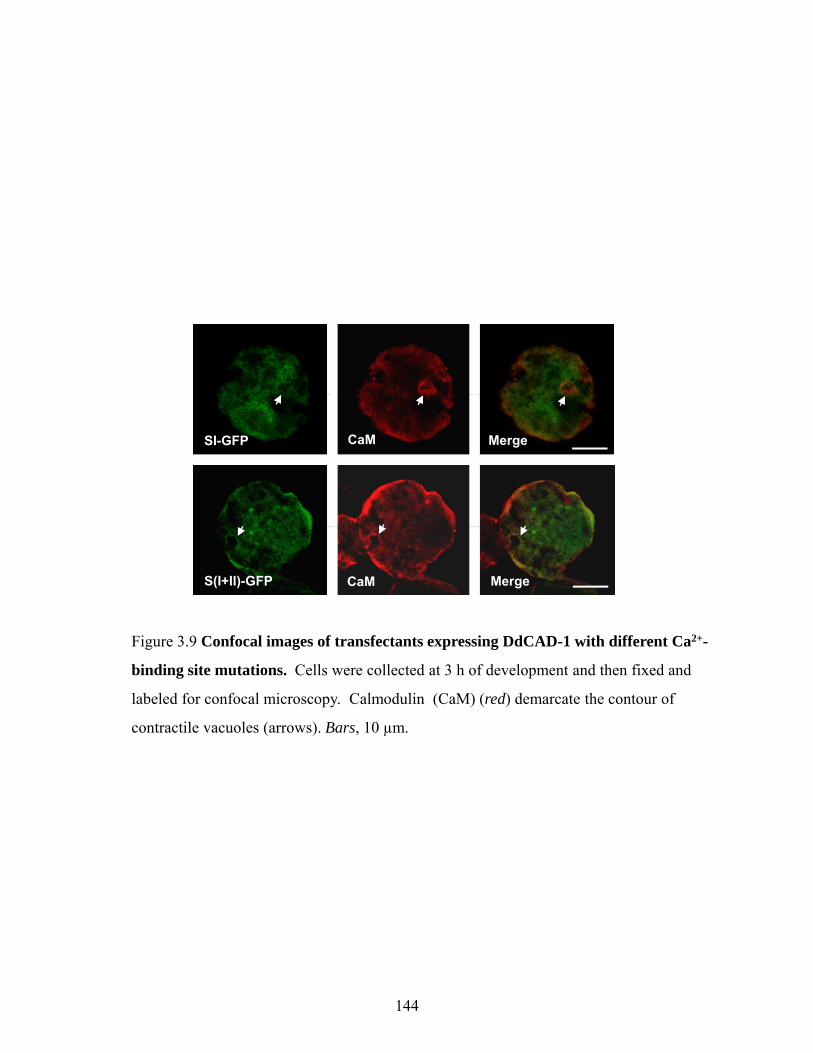
153
by eukaryotic cells to target soluble proteins for surface expression and secretion. The
combination of genetic and biochemical analyses in Dictyostelium should provide a useful
model for the future dissection of these pathways.

154
Chapter 4
cadA Is a Single-Gene Green Beard that Regulates Morphogenesis
through Differential Spatiotemporal Expression in Dictyostelium
The work presented in this chapter has been submitted for publication:
Sriskanthadevan, S., Zhu, Y., Manoharan, K., Yang, C., and Siu, C.H. (2010). cadA Is a
Single-Gene Green Beard that Regulates Morphogenesis through Differential Spatiotemporal
Expression in Dictyostelium.
(Y.Z. contributed to some of the molecular cloning and mutagenesis studies; K.M. and C.Y.
contributed to some of the morphological studies; C.-H.S contributed to project guidance and
manuscript preparation.)

155
4.1 Summary
During development of Dictyostelium, multiple cell types are formed and undergo a
coordinated series of morphogenetic movements guided by their adhesive properties.
DdCAD-1 is a unique homophilic cell adhesion molecule encoded by the cadA gene and
synthesized in the cytoplasm. In chimeras developed on soil plates, cells expressing DdCAD-
1 were more likely recruited into aggregates and developed into fruiting bodies than cadA-
null cells. Thus, cadA behaved as a single-gene green beard, directing benefit towards a copy
of itself in another cell. However, cadA exhibited anti-green beard behaviour when
development was carried out on non-nutrient agar. Wild-type cells differentiated mostly into
stalk cells and eventually died, whereas cadA-null cells survived as spores. Analysis of cadA-
null cells showed that their defect in cell sorting was rescued by the ectopic expression of
DdCAD-1. Morphological studies of cells which expressed DdCAD-1-GFP, revealed major
spatial and temporal changes in the subcellular distribution of DdCAD-1 during development.
Whereas DdCAD-1 became internalized in most cells in the post-aggregation stages, it was
prominent in the contact regions of anterior cells. Additionally, cell sorting was restored in
cadA-null slugs by externally applied recombinant DdCAD-1. Interestingly, DdCAD-1
remained on the cell surface of anterior cells, while it was internalized in the posterior cells.
Further analysis showed that DdCAD-1-expressing cells migrated slower than cadA-null cells
and sorted to the anterior region of chimeric slugs. Taken together, these results suggest that
DdCAD-1 regulates the social behaviour of Dictyostelium through its differential subcellular
distribution in prespore and prestalk cells.

156
4.2 Introduction
Cooperation among cells is a challenge for evolution by natural selection. According
to Darwin (1859), individuals act in a way to maximize their own survival. Hamilton (1964)
proposed that cooperative behaviours evolve because they help individuals who share more
alleles with one another than predicted by chance (Hamilton, 1964a; Queller, 1984). From an
evolutionary point of view, behaviour is altruistic if it increases the fitness of the recipient
while the donor’s fitness decreases (Velicer and Vos, 2009; West et al., 2006b). Hamilton
described three conditions that would allow a single gene to direct altruistic benefits toward a
copy of itself in another individual. The gene should bear a phenotype that advertises the
presence of the allele, and it must facilitate the recognition of that phenotype in others, and act
altruistically towards those that have been recognized (Hamilton, 1964a). Dawkins coined the
metaphor “green beard” to denote the phenotype expressed by an altruistic gene (Dawkins,
1976).
Green beard genes remained a hypothetical phenomenon until a recent study showed
that all components of a green beard effect are present in the red fire ant, Solenopsis invicta
(Keller and Ross, 1998). It has been predicted that the homophilic functional class of proteins
would constitute the single-gene green beard (Haig, 1996). Indeed, the first single gene green
beard identified was found to code for the homophilic cell adhesion molecule contact site
A/gp80 in Dictyostelium (Queller et al., 2003). Interestingly, Haig (1996) has also suggested
that a heterophilic cell adhesion protein could also exhibit the single-gene green beard effect,
which was demonstrated recently in the yeast flocculation gene, FLO1 (Smukalla et al.,
2008). FLO1 is a cell-surface protein that interacts with mannan oligosaccharide chains to
form lectin-like bonds, resulting in cell-cell adhesion that leads to flocculation.

157
Dictyostelium has been recognized as an attractive model system for the study of
social behaviour and evolution (Strassmann et al., 2000). During development, the stalk cells
display altruism and give up reproduction in order to benefit the spore cells by lifting them
above the hazards of the soil, thus increasing their chances of dispersal to a more favourable
environment. In addition, aggregative development exposes Dictyostelium to chimerism,
which includes cheating, whereby individuals have access to group benefits without
contributing their fair share. This phenomenon raises the issues of how social cooperation
persists in nature and what factors would permit individuals with a compromised phenotype
to survive.
Several adhesion systems that can influence the social behaviour of Dictyostelium
have been identified. Mutations in these genes have major effects on aggregate formation and
morphogenesis (Siu et al., 2004c). The Ca2+-dependent cell-cell contact sites are mediated by
the adhesion molecule DdCAD-1, which is encoded by the cadA gene and expressed soon
after the initiation of development (Brar and Siu, 1993; Yang et al., 1997). The Ca2+/Mg2+-
independent sites are mediated by the contact site A protein gp80 (Kamboj et al., 1989; Siu et
al., 1985; Stein and Gerisch, 1996) and TgrC1/lagC/gp150 (Dynes et al., 1994; Gao et al.,
1992; Wang et al., 2000). The csA gene which codes for gp80 has been identified as the first
single gene green beard (Queller et al., 2003) and the tgr gene family which includes tgrC1
and tgrB1, has been implicated in kin discrimination and provides an early evolutionary origin
of kin discrimination (Benabentos et al., 2009).
DdCAD-1 is unique among cell adhesion molecules because it is synthesized as a
soluble protein and then transported to the plasma membrane by contractile vacuoles (Sesaki
et al., 1997; Sriskanthadevan et al., 2009). DdCAD-1 contains two distinct domains with β-
sandwich architecture. While the N-terminal domain is involved in homophilic binding, the

158
C-terminal domain tethers the protein to a membrane anchor (Lin et al., 2006). Studies on the
cadA-null cells have implicated DdCAD-1 in cell sorting and cell-type proportioning (Wong
et al., 2002). Since DdCAD-1 is expressed transiently on the plasma membrane during cell
aggregation (Sesaki and Siu, 1996), it is not known how DdCAD-1 regulates cell
differentiation and cell sorting.
In this report, we examined chimeras consisting of cadA-null cells and the parental
AX4 cells (cadA+). Results show that cadA behaved as a single-gene green beard when
development was carried out on soil plates, which mimicked the natural environment.
However, cadA exhibited anti-green beard effects when development was carried out on non-
nutrient agar plates. In the latter case, the cadA+ cells differentiated into stalk cells, while
most of cadA- cells differentiated into spores and survived. To investigate mechanisms that
underlie the anti-green beard behaviour of cadA, DdCAD-1-GFP was expressed ectopically in
cadA-null cells. Morphological studies have revealed remarkable changes in the temporal and
spatial expression of DdCAD-1 during development. Whereas DdCAD-1 is enriched in the
cell-cell contact regions in the anterior zone, it is present primarily in the cytoplasm in the
posterior zone. The aberrant cell sorting behaviour of cadA-null cells could be rescued by the
addition of recombinant DdCAD-1. Cells that moved into the anterior zone during tip
formation showed that recombinant DdCAD-1 was enriched in their contact regions. The
data suggest that cell sorting results from the differential distribution of membrane-associated
DdCAD-1 and provide new insights into the cellular basis of the anti-green bread
phenomenon.

159
4.3 Experimental Procedures
Construction of DdCAD-1-GFP and Mutant Plasmids for Cell Transfection
PCR products were obtained from the cadA cDNA and the cadA genomic DNA cloned
in pBluescript SK+ vectors. To construct the -631::cadA-GFP expression vector, the cadA
promoter starting at -631 (GenBank accession number: AAFI02000079.1) was ligated to the
cadA cDNA and then inserted before GFP in the pIGFP vector which contains the neomycin
marker (kindly provided by Dr. David Knecht, University of Connecticut, CN) (Fig. 4.1).
Mutations in cadA were constructed as described previously (Sriskanthadevan et al., 2009)
and all constructs were sequenced from both ends. Plasmid DNAs were transfected into
cadA-null cells by electroporation (Pang et al., 1999). Cells were collected at mid-log phase,
washed twice in cold H50 buffer (20 mM Hepes, 50 mM KCl, 10 mM NaCl, 1 mM MgSO4, 5
mM NaHCO3, 1 mM NaH2PO4, pH 7.0) and then suspended in H50 at 2 x 107 cells/ml. Cell
samples (100 µl each) were mixed with ~5 µg of plasmid DNA and incubated for 5 minutes
on ice in 1-mm cuvettes. Electroporation was carried out by two consecutive pulses of 0.85
kV with a capacitance of 25 mF applied to the cuvette and with a 5-second recovery between
pulses. After 5 minutes of incubation on ice, the cells from each cuvette were transferred to 3
ml of HL5 (0.5% Difco proteose peptone No. 2, 0.5% BBL Thiotone E peptone, 1% glucose,
0.5% yeast extract, 2.5 mM KH2PO4, 2.5 mM Na2HPO4, pH 6.7) in wells of a six-well tissue
culture plate (Pang et al., 1999). Transfectants were selected by sequential incubation in 2, 5,
10 and 20 μg/ml G418 (Sigma Chemical Co., St. Louis, MO) over a period of 3 weeks. The
expression of wild-type or mutant DdCAD-1 was monitored by western blot analysis.
Transfectants were maintained in 20 µg/ml of G418.

A
… TGA AGA TGT TCa agc ttT GCA ACT TGT TTC…….ATG TCT
GTT………….C TTT AAC TCT GAA AAA gaa ttc ATG ….
-631 Start codonHindIII
B
cadA coding regionEcoRI GFP coding region
Name Sequence
-631-cadA-pBSK-631-SI-pBSK-631-S(I+II)-pBSK
Forward 5’ -> 3’ (BamHI and AccI)TCC gga tcc ATG TCT gtc gac GCA AAT AAA GTA AAA TTC
Reverse 5’ -> 3’ (XhoI and BglII)Reverse 5 -> 3 (XhoI and BglII)GG ctc gag aga tct TTA TTT TTC AGA GTT TAA GTT AAA GAT GAA AG
-631-cadA-GFP-631-SI-GFP-631-S(I+II)-GFP
Forward 5’ -> 3’ (HindIII)GCa agc ttT GCA ACT TGT TTC AAC TTA
Reverse 5’ -> 3’ (EcoRI)( )CG gaa ttc TTT TTC AGA GTT TAA GTT AAA G
-631-GFP Forward 5’ -> 3’ (HindIII)GCa agc ttT GCA ACT TGT TTC AAC TTA
Reverse 5’ -> 3’ (EcoRI)GCG CGT gaa ttc AGA CAT TTT TTA ATT TTT AAT ACT ATA CC
Figure 4.1 Construction of DdCAD-1-GFP and Mutant Plasmids for Cell
Transfection. (A) Schematic diagram showing the multiple cloning site region of the
DdCAD-1-GFP construct (B) Table listing primers used in the construction of GFP
ATA CC
DdCAD 1 GFP construct. (B) Table listing primers used in the construction of GFP
fusion proteins.
160

161
Development on Non-nutrient Agar or Soil Plates
Dictyostelium strains, AX4 (parental strain of the cadA-null strain), cadA-null and
various transfectants, were cultured either in association with Klebsiella aerogenes or in HL5
axenic medium (Sussman, 1987). Cells were grown and developed at 22°C. To determine
the percentage of cadA-null spores in chimeric fruiting bodies, axenically cultured AX4 and
act15::GFP;cadA- cells were mixed to give a final concentration of 1 x 108 cells/ml in the
developmental buffer DB (25 mM sodium phosphate buffer, pH 6.5, 1 mM CaCl2, 2 mM
MgCl2). Cell mixtures at 20:80, 50:50 and 80:20 ratios were developed on either 2% non-
nutrient agar buffered with KK-2 (2.2g/L KH2PO4 and 0.7g/L K2HPO4, pH 6.4) or soil plates.
Soil plates were prepared using commercially available garden soil (neutral pH) which was
sieved to obtain particles of homogenous size (0.4 cm in diameter) and then autoclaved (Ponte
et al., 1998). Aliquots of 20 g were distributed homogeneously on 90-mm Petri dishes which
were then moistened with 4 ml of sterile water. Aliquots of 0.25 ml (2.5 x 107 cells) of
mixed cells were pipetted onto soil plates to cover an area of ~ 2 cm2. Four samples were
placed on the each dish and then incubated at room temperature in a moisture box covered
with aluminum foil. The cells were developed for 48 hour. Pictures were taken from two
random regions for each example using a dissecting microscope for the quantification of
fruiting bodies. Sori were picked randomly, treated with 0.05% SDS in DB, and the
percentage of fluorescent cadA-null spores was estimated.
Cell Cohesion Assay
Cells were collected for development in 17 mM phosphate buffer (pH 6.4) at 2x107
cells/ml. After 4 hr, cells were resuspended at ~2.5x106 cells/ml. Cells in 200-µl samples
were dispersed by vortexing for 15 seconds. Cells were allowed to re-form aggregates on a

162
platform shaker rotating at 180 rpm at 22°C. At regular time intervals, the numbers of non-
aggregated cells (singles and doublets) were scored using a hemocytometer and the
percentage of cell aggregation was calculated.
Temporal and Spatial Expression Pattern of DdCAD-1
Axenically grown -631::cadA-GFP transfectants were washed free of medium and
resuspended in DB at 4 x 106 cells/ml. Aliquots of 0.5 ml were deposited on positively
charged coverslips (Fisher Scientific, Pittsburgh, PA, cat. no. 12-545-84) for 30-60 minutes
and excess buffer was removed. Aggregates were fixed in 3.7% formaldehyde at different
time points and permeabilized with 0.5% Triton X-100 for 15 minutes. Samples were
incubated in 1% BSA in MCG buffer (50 mM MES, pH 6.4, 0.2 mM CaCl2, 2 mM MgCl2)
for 30 minutes. Then, samples were stained with phalloidin to mark the cell periphery (1:400
dilution) (Molecular Probes, Eugene, OR) for 30 minutes. Coverslips were mounted in
DAKO fluorescent mounting medium (DakoCytomation, Glostrup, Denmark). Confocal
images were acquired using the Zeiss LSM 510 microscope.
Analysis of Cell Sorting in Slugs
Cells were cultured either in association with K. aerogenes or axenically in HL5
medium. Cells were collected, washed, and resuspended in 17 mM phosphate buffer at107
cells/ml. The cells were stained in 0.05% Neutral Red for 5 minutes at room temperature
(Weijer et al., 1987). Prestalk cells stains more strongly with Neutral Red that stains the large
intracellular acidic vesicles (Gross, 1994). After washing, cells (2 x 108cells/ml) were deposited
on 1.5% non-nutrient agar (Eichinger et al., 2005). Development was carried out at 22°C in
the dark. Slugs were photographed under light microscope at different time intervals. The

163
distribution pattern of Neutral Red-stained cells within the slug was examined.
Approximately 200 slugs were analyzed in each experiment. The lengths of the whole slug
(y) and the intensely stained anterior zone (x) were measured using Image J. The x/y ratio (R)
was calculated and the frequencies of occurrence of the R values were determined.
In vitro Reconstitution of DdCAD-1 using Recombinant Proteins
Cells were developed for 4-6 hours before staining with Neutral Red. After staining,
cells were incubated at 2 x 108 cells/ml with wild-type or mutant recombinant DdCAD-1
protein at 0.5 mg/ml or 3 mg/ml for 30 minutes. Then 100 µl of cells were placed in a straight
line ~5 cm long on 1.5% non-nutrient agar plates for development until the slug stage. For
microscopic analysis, His-DdCAD-1 was labelled with sulfodicholorophenol esters (SDP)
conjugated with Alexa fluor 488 (Molecular Probes). Conjugated proteins were separated
from un-reacted labelling reagent using ZebaTM desalt spin columns (Pierce, Rockford, IL cat.
no. 89890). The protein was resuspended at 6 mg/ml in 20 mM PIPES, pH 6.4. Then, cadA-
null cells (2 x 106 cells) were developed for 4-6 hours and incubated with Alexa-488-
conjugated His6-DdCAD-1 proteins at 3 mg/ml for 12 hours on coverslips and then excess
buffer was removed. Cells were developed until the culmination stage before fixation with
3.7% formaldehyde for confocal microscopy.
Antibody-induced Cap Formation
Slug or culminant stage cells were dissociated and antibody-induced cap formation
was performed. Anti-GFP mAb (1:100 dilution) (Sigma Chemical Co., St. Louis, MO) was
added to 1.2 x 106 cells suspended in 300 μl of 50 mM MES buffer, pH 6.3, and incubated for
30 minutes at room temperature. Alexa-568-conjugated goat anti-mouse antibody was added

164
at 1:400 dilution and rotated at room temperature for another 30 minutes. Aliquots (300 μl
each) of cells were deposited on coverslips and allowed to attach for 15 minutes. The
coverslips were washed gently with MCG buffer, fixed in 3.7% formaldehyde, washed and
mounted for fluorescence microscopy.
Flow Cytometry Analysis
Axenically grown -631:: cadA-GFP transfectants were washed free of the medium and
was resuspended in DB at 2 x 108 cells/ml. Cells were deposited on 1.5% non-nutrient agar
and development was carried out at 22°C in the dark. Cells were dissociated from mounds,
slugs and culminants for further analysis. Dissociated cells were analyzed using a LSRII
(Becton Dickson) flow cytometer using excitation wavelengths of 488 nm. Results were
analyzed using the WinMDI software.
Chemotactic cell migration assay
The chemotactic migration assay was performed according to Wallace and Frazier
(1979). Cells were collected were collected at 4 hours of development and resuspended at 2.5
x 108 cells/ml in 17 mM sodium phosphate buffer. The cAMP stock solution (10 mM) was
diluted to 100 μM or 250 μM. Non-nutrient 1.5% agar plates were prepared fresh and fully
hydrated with buffer before the experiment. Four wells were created in the centre of the
quadrants of an agar plate. Then, the wells were filled with 30 μl of cAMP and 1 μl aliquots
the cell suspension were placed 5 mm away from the well. The plates were incubated at room
temperature in a moisture box covered with aluminum foil. Cell migration was monitored
using a dissecting microscope and the distance of migration was measured after 16 hours.

165
4.4 Results
The cadA gene displays both green beard and anti-green beard behaviour
Cell adhesion molecules that undergo homophilic binding are known to exhibit green
beard effects (Haig, 1996; Queller et al., 2003). Although DdCAD-1 mediates cell-cell
adhesion via homophilic interaction, it appears to function transiently at the aggregation stage
of development. DdCAD-1 redistributes from the cell-cell contact regions as the Ca2+/Mg2+-
independent contact site A protein gp80 moves into these regions (Sesaki and Siu, 1996).
DdCAD-1 is therefore not expected to behave as a green beard gene like the csA gene.
Contrary to this prediction, when the parental AX4 cells and cadA-null cells were mixed for
development on soil plates, disproportionate spore yields were observed in fruiting bodies
collected at 48 hours of development (Fig. 4.2A). The yield of cadA-null spores was about
40% lower than the expected value if cells from both strains were equally represented in the
chimeras. These results showed that the cadA+ cells promoted the survival of other cadA+
cells, but not cadA-null cells. Therefore, the cadA gene displays green beard behaviour when
chimeras were developed under conditions that mimicked their natural habitat (Ponte et al.,
1998). It is possible that the presence of DdCAD-1 on the cell surface allows AX4 cells to
adhere preferentially to each other and facilitates aggregate formation. Indeed, when AX4
cells and cadA-null cells were developed separately on soil plates, cadA-null cells developed
much slower than AX4 cells. Whereas most of the AX4 cells completed development by 24
hours, only a few fruiting bodies were observed for cadA-null cells (Fig. 4.2B). At 48 hours,
the number of cadA-null fruiting bodies reached only 40% of that of the AX4 cells, indicating
that the loss of DdCAD-1 expression hampered the aggregation and differentiation processes.

A20 : 80 80 : 20 50 : 50
% AX4 : % GFP;cadA-
24 h
r
ntsp
ores
ras
100
80
60248
hr
% AX4 : % GFP;cadA-
% o
f flu
ores
cen
in c
him
er
40
20
050:5080:20 20:80
B AX4 cadA-
ve n
o. o
f fru
iting
bod
ies
(%)
24 h
rhr
14012010080604020
Rel
ativ
48 0
024 hr 48 hr
Figure 4.2 Green beard effects of the cadA gene. (A) Spore formation by
act15::GFP;cadA- cells in the fruiting body of chimeras developed on soil plates. AX4
cells were mixed with act15::GFP;cadA- cells at different ratios and allowed to develop
for 48 hours. Bars, 1 mm. The percentages of fluorescent spores in chimeric fruiting
bodies were scored (n=3) (gray bars). The data were compared to the theoretical values if
both types of cells were distributed evenly in the sorus (black bars). (B) Development of
cadA- cells and the parental AX4 cells (cadA+) on soil plates. Images were taken at 24
hours and 48 hours of development. Bars, 1 mm. The number of fruiting bodies formed
by AX4 (gray bars) and cadA- (black bars) cells were scored and normalized to that of
AX4 ll t 24 h f d l t ( 8)AX4 cells at 24 hours of development (n=8).
166

167
Anti-green beard effects of the cadA gene led to cheating of cadA-null cells in chimeras
Interestingly, cadA-null cells displayed anti-green beard behaviour when cell mixtures
were deposited on non-nutrient agar plates for development. Contrary to the data obtained
with soil plates, the cadA-null cells differentiated primarily into spores (Fig. 4.3Aa,b). Only
a few of the fluorescent cadA-null cells were found occasionally in the stalk. The quantitative
data showed that almost all of the cadA-null cells in the chimeras formed spores when
chimeras were made up of 20% cadA-null cells (Fig. 4.3Ac). When chimeras were made up
of 80% cadA-null cells, almost all spores came from the cadA-null strain. The anti-green
beard behaviour of the cadA gene allowed the cadA-null to cheat and survive as they
preferentially differentiated into spores while the cadA+ cells differentiated into stalk cells and
eventually died.
Preferential localization of cadA+ cells in prestalk region of chimeras during
development
In order to investigate how DdCAD-1 exert anti-green beard effects during
development, cadA-null cells were transfected with plasmid DNA containing the act15::cadA-
GFP construct (Fig. 4.4A,B) to ensure even expression of the fusion protein in cells (Fig.
4.4C,D). Normal sorting pattern was restored in slugs by the ectopic expression of DdCAD-1
(Fig. 4.4E,F). Transfected cells were mixed with cadA-null cells at different ratios and the
location of the DdCAD-1-GFP-expressing cells in the chimeras was monitored at different
stages. The green cells appeared mostly in the periphery at the mound stage. Most of them
were present in the anterior prestalk region by the slug stage. To quantify the distribution of
green cells, images of slugs were collected and divided into three segments for the
measurement of fluorescence intensity (Fig. 4.3B). The data confirmed the preferential

Figure 4.3 Anti-green beard behaviour of the cadA gene. (A) Spore formation by
act15::GFP;cadA- cells in the fruiting body of chimeras developed on non-nutrient agar.
(a) Confocal image showing the predominant presence of fluorescent spores inside the
sorus of a chimeric fruiting body. Bar, 50 µm. (b) A pair of fluorescence and phase
images of spores collected from a chimeric fruiting body made up of 20% AX4 and 80%
act15::GFP;cadA- cells. (c) Quantification of fluorescent spores in different chimeras
(n=3) (gray bars), which are compared to the theoretical values if both types of cells were
distributed evenly throughout the sorus (black bars) or if the cadA-null cells cheat and
constitute to 100% of the spores (white bars). (B) The act15::cadA-GFP;cadA-
transfectant cells were mixed at different ratios with cadA- cells for development on
coverslips. Developing structures were fixed for confocal microscopy. The micrographs
show the spatial distribution of act15::cadA GFP;cadA- cells (green) in the slug structuresshow the spatial distribution of act15::cadA-GFP;cadA cells (green) in the slug structures
of chimeras. Bars, 50 µm. Fluorescence intensity per unit area was quantified using the
Image J software (NIH) for the anterior, middle and posterior regions of the slug as shown
in the schematic diagram. The relative fluorescence intensities were calculated for each
region of the different chimeras and compared in the bar graph (n = 20 to 25). (C)
Confocal images of chimeric slugs containing act15::GFP;cadA- cells and AX4 cells
mixed at different ratios. Bars, 50 µm. The bar graph shows the relative fluorescence
intensities of the three slug regions (n = 20). (D) Confocal images of chimeric slugs
containing act15::GFP;cadA- and cadA- cells mixed at different ratios. Bars, 50 µm.
Relative fluorescence intensities were measured for each slug regions (n = 20). (E)Relative fluorescence intensities were measured for each slug regions (n 20). (E)
Confocal images of the different chimeras at the culmination stage. Bars, 50 µm.
168

A
GFP
;cad
A-
GFP
;cad
A-
ntsp
ores
in
as
a b c1201008060
ant. post.mid.dA GFP dA
B20
% A
X4
: 80%
80%
AX
4: 2
0% G
% o
f flu
ores
cen
chim
era
% AX4 : % GFP;cadA-
604020
050:5080:20 20:80
C
e
cadA-GFP : cadA-
20 : 80 80 : 20 R
elat
ive
fluor
esce
nce
in
tens
ity (
%)
anterior middle posterior
10080604020
050:5020:80 80:20
100
D
Rel
ativ
e flu
ores
cenc
ein
tens
ity (%
)en
ce
GFP;cadA- : AX4
20 : 80 80 : 20
GFP;cadA- : cadA-
20 : 80 80 : 20
20 : 80 80 : 20
100806040
200
ant. mid. post.
20 : 80100
Rel
ativ
e flu
ores
cein
tens
ity (%
)
E
20 : 80 80 : 20
cadA-GFP : cadA- GFP;cadA- : AX4 GFP;cadA- : cadA-
20: 80 20: 8020: 80
20 : 80 80 : 20
ant. mid. post.
80604020
0
169

A B
cadA GFP639 bp 734 bp
Act15 promoter
A B
C D
52 kDa
DdCAD 1 pAbC D
20µm
DdCAD-1 pAb
Eve
nts
E F
sortedpartially sortednon sorted
100
80
GFP
KAX-3 DdCAD-1-GFP cadA-
% to
tal s
lugs
non-sorted
KAX-3 DdCAD-1-GFP
cadA-
60
40
20
0
Figure 4.4 Expression of cadA-GFP rescues the cell-sorting defects of cadA- cells. (A)
Schematic drawing of DdCAD-1-GFP fusion construct. (B) Western blot profiles
showing the expression of DdCAD-1-GFP in transfectants. (C) A pair of fluorescence
and phase images of cells dissociated from the mound stage of development. (D) FACS
analysis showing normal distribution of DdCAD-1-GFP expression at 0 hour of
development. (E) The pattern of of prestalk and prespore cell distribution in slugs of
Neutral Red-stained AX4, act15::cadA-GFP;cadA- cells and cadA- cells. (F)
Quantification cell sorting patterns in slugs based on the Neutral Red-stained wild-type
and mutant cellsand mutant cells.
170

171
localization of green cells in the anterior zone of slugs. Spore yield was also identical to that
of AX4 cells in similar chimeras.
As a control, cadA-null cells were transfected with an act15::GFP construct. The
GFP;cadA- cells were mixed with the AX4 (cadA+) cells for development. Most of the
GFP;cadA- cells were found in the middle and posterior region of the slug, whereas the
anterior region was occupied primarily by the AX4 cells (Fig. 4.3C). Even within the 80:20
chimeras, a distinctly “dark” area was evident at the tip region. In a second control, GFP-
cadA-null cells were mixed with cadA-null cells. As expected, the green cells distributed
more or less evenly along the whole length of the slug (Fig. 4.3D). At the early culmination
stage, all three types of chimeras showed patterns of green cell distribution similar to those
observed at their respective slug stage (Fig. 4.3E). These results suggest that cadA+ cells
preferentially sort to the periphery of chimeric aggregates and then move to the anterior
region and differentiate into prestalk cells, whereas cadA- cells occupy primarily the posterior
two-third of the slug and eventually differentiate into spores.
Rescue of cadA-null cells using wild-type and mutant constructs of DdCAD-1
The notion that DdCAD-1 plays an important role in the sorting out of prestalk cells
from prespore cells would predict the presence of DdCAD-1 on the cell surface during
morphogenesis. To investigate how DdCAD-1 might be involved in cell sorting during
development, cadA cDNA was fused to GFP and the fusion construct was ligated to the
endogenous cadA promoter, -631 (Yang et al., 1997) (Fig. 4.5A) for transfection into cadA-
null cells. Using the endogenous cadA promoter ensured that expression occurs according to
the normal temporal program of development. G418-resistant clones were screened by direct
visualization under a fluorescence microscope. The proper temporal expression of DdCAD-

A
cadA GFP639 bp 734 bp
-631 promoter
B
52 kDa
DdCAD-1-GFP
hr 0 2 4 6
C
greg
atio
n
60
80
100DdCAD-1-GFPDdCAD-1-GFP + EGTAcadA-nullDdCAD-1-GFP + EDTA
pAb α GFP
Time (min)
% a
gg
0
20
40
0 20 40 60 80
Figure 4.5 Expression of DdCAD-1-GFP restores the Ca2+-dependent adhesion sites
in transfectants. (A) Schematic drawings of DdCAD-1-GFP fusion construct driven by
the endogenous cadA promoter. (B) Western blots probed with rabbit polyclonal
antibodies (pAb) against GFP showed the expression of the fusion protein in transfectantsantibodies (pAb) against GFP showed the expression of the fusion protein in transfectants
during development. (C) Cell cohesion assays performed in the presence and absence of
EGTA, showing that the Ca2+-dependent adhesion sites were restored in the transfectants.
The residual cohesiveness in both transfectants and the parental cadA-null cells were
sensitive to EDTA inhibition. Addition of EGTA to the cadA-null cells in this assay has
172
no effect (Wong et al., 2002).

173
1-GFP in transfectants was confirmed by probing protein blots with antibodies against
DdCAD-1and GFP (Fig. 4.5B). While both transfectants and the parental cadA-null cells
expressed the EDTA-sensitive/EGTA-resistant adhesion sites, cell cohesion assays showed
that the EDTA/EGTA-sensitive or Ca2+-dependent cell-cell adhesion sites were observed only
in the transfectants (Fig. 4.5C).
Since DdCAD-1-mediated cell-cell adhesion requires Ca2+, we tested whether the
Ca2+-binding capability of DdCAD-1 is required for proper cell adhesion and cell sorting
during development. cadA-null cells were transfected with two constructs of DdCAD-1
which contained mutations in the Ca2+-binding site I and S(I+II). Expression of the mutant
proteins were confirmed with western blots (Fig. 4.6Bb). When cell sorting experiments
were performed using Neutral Red stained cells, cadA-nulls expressing DdCAD-1-GFP
ectopically showed the normal sorting pattern similar to AX4 cells (Fig. 4.6Ab-d). However,
ectopic expression of DdCAD-1 containing mutated Ca2+-binding sites failed to rescue the
cell sorting defects of the cadA-null cells. Most of the mutant slugs showed non-sorted
patterns (Fig. 4.6Bc,d). These results indicate that Ca2+-binding capability of DdCAD-1 is
required for the proper sorting out of prestalk and prespore cells at the slug stage.
Dynamic changes in the temporal and spatial distribution of DdCAD-1 during
development
Transfectants that expressed DdCAD-1-GFP at levels comparable to the DdCAD-1
level in AX4 cells were selected for development and the temporal and spatial expression of
DdCAD-1-GFP was monitored by confocal microscopy. Developmental structures were
fixed and F-actin was stained with phalloidin to demarcate the cell boundaries (Fig. 4.7).
Confocal microscopy revealed that DdCAD-1-GFP was synthesized inside the cytoplasm

a cb d
A
x
y
R = x/y
% o
f tot
al s
lugs 100
80604020
0
RR R.1 .3 .5 .7 .9 .1 .3 .5 .7 .9 .1 .3 .5 .7 .9
KAx-3 DdCAD-1-GFP cadA-
SI GFP-631 D39A, T81A
S(I+II) GFP-631 D35A, N38A, D39A
B
ac
b
d
otal
slu
gs 100806040
SI-GFP S(I+II)-GFP
52 kDa
SI-GFP S(I+II)-GFP
pAb α DdCAD-1R
% o
f to
R
4020
0.1 .3 .5 .7 .9 .1 .3 .5 .7 .9
4hr 6hr 4hr 6hr
Figure 4.6 Rescue of cell sorting in cadA- slugs by DdCAD-1 depends on its Ca2+-
binding capability (A) Expression of DdCAD 1 GFP driven by its endogenousbinding capability. (A) Expression of DdCAD-1-GFP driven by its endogenous
promoter restored normal cell sorting in cadA- slugs. (a) A light micrograph showing
Neutral Red-stained slugs formed by -631::cadA-GFP;cadA- transfected cells. The
relative length of the stained anterior zone was measured and the R values were plotted
against the frequency of occurrence of the R values: (b) AX4 slugs; (c) -631::cadA-
GFP;cadA- slugs; (d) cadA- slugs. Slugs without a clear sorting pattern were given an R
value of 0.9. (n = 200 to 300 slugs for each strain) (B) Aberrant cell sorting patterns in
slugs expressing DdCAD-1 containing mutations in its Ca2+-binding sites. (a) Schematic
drawings depicting the mutant Ca2+-binding form of cadA (SI, S(I+II), with amino acid
substitutions indicated on top of the schematic drawing) fused to GFP. (b) Expression ofsubstitutions indicated on top of the schematic drawing) fused to GFP. (b) Expression of
mutant DdCAD-1-GFP in transfectants. Transfectants derived from cadA-null cells were
cultured on bacteria lawn and collected for development. Cell lysates were prepared from
4- and 6-hour cells for SDS-PAGE and the protein blots were probed with rabbit anti-
DdCAD-1 antiserum. (c, d) Sorting patterns of Neutral Red-stained transfectants. The
relative lengths of the anterior zones were measured and the R values were plotted against
the frequency of occurrence.
174

20 µm
cell streamDdCAD-1-GFP F-actin Merge
mound
tip mound
20 µm
50 µm
slug
50 µm
culminant
50 µm
Figure 4 7 Changes in the pattern of DdCAD 1 expression during developmentFigure 4.7 Changes in the pattern of DdCAD-1 expression during development.
The -631::cadA-GFP;cadA- transfectant was developed on coverslips for 10 to 22 hours.
The developing structures were fixed and permeabilized with 0.5% Triton X-100 before
staining with phalloidin (red). Confocal images show changes in the temporal and spatial
expression of DdCAD-1-GFP at different developmental stages.
175

176
shortly after the initiation of development and became associated with the plasma membrane
and filopodial structures as previously reported (Sesaki and Siu, 1996). At the mound stage,
DdCAD-1-GFP was present mostly in the cytoplasm while enrichment in the cell-cell contact
regions was not observed. A similar pattern of DdCAD-1 expression was evident at the tip
mound stage. Additionally, DdCAD-1 appeared to be absent from a subset of cells scattered
throughout the aggregate. At the migrating slug stage, cells with higher levels of DdCAD-1
appeared in the anterior region. This difference in DdCAD-1 expression between the anterior
cells and the posterior cells became more obvious in culminants.
When cells were subjected to FACS analysis, a single group of fluorescent cells was
observed at the mound stage (Fig. 4.8). However, two groups of fluorescent cells became
evident at the culmination stage. About 30% of the cells had a much reduced level of
DdCAD-1, while the other group showed a broad range of higher fluorescence intensity.
Taken together, the data indicate heterogeneity in DdCAD-1 expression as development
progressed and that cells with a higher level of DdCAD-1 expression had the propensity of
differentiating into prestalk cells.
Enrichment of DdCAD-1 in the cell-cell contact regions of anterior cells
A role for DdCAD-1 in cell sorting would imply the presence of DdCAD-1 on the cell
surface. However, the high level of cytoplasmic DdCAD-1-GFP prevented the morphological
analysis of membrane-associated DdCAD-1. Therefore, transfectants at the slug stage were
treated with detergent for a longer period of time to allow intracellular DdCAD-1-GFP to leak
out from cells. Confocal images of these cells revealed differential association of DdCAD-1-
GFP with the plasma membrane along the long axis of the slug. An abundance of DdCAD-1-
GFP was observed in cell-cell contact regions in the anterior zone, with close coincidence

A
Eve
nts
mound
BGFP
culminant
GFP
Eve
nts
GFP
Figure 4.8 FACS analysis showing DdCAD-GFP-expressing populations at mound
(A) and culminant (B) stages of the development.
177

178
with the phalloidin-staining pattern, while the intracellular green fluorescence was relatively
low (Fig. 4.9A). However, DdCAD-1-GFP was present mostly in the cytoplasm in the
posterior zone where membrane association of DdCAD-1 became less evident (Fig. 4.9B).
During culmination, DdCAD-1-GFP was present predominantly in cell-cell contacts of the
upper cup region surrounding the descending stalk tube (Fig. 4.9C). In fruiting bodies, cells
expressing high levels of DdCAD-1-GFP were found concentrated in the upper cup and lower
cup regions of the sorus (Fig. 4.9D).
To determine whether DdCAD-1-GFP was associated with the surface of anterior
cells, the anterior and posterior zones of slugs were cut and the cells were dissociated for
further analysis. DdCAD-1-GFP was found enriched in cell-cell contacts of anterior cells but
not posterior cells (Fig. 4.9E). When antibody-induced clustering of DdCAD-1 was
performed on living cells, DdCAD-1 “caps” were observed on the elongated anterior cells,
indicating that DdCAD-1 was present on the cell surface (Fig. 4.9Fa). In contrast, antibody
treatment of cells isolated from the posterior region failed to induce cap formation (Fig.
4.9Fb), confirming that DdCAD-1-GFP was present at a very low level or absent from the
cell surface.
Rescue of cadA-null phenotype by in vitro reconstitution of DdCAD-1
DdCAD-1 is synthesized as a soluble protein and exported by contractile vacuoles to
the plasma membrane (Sesaki et al., 1997). DdCAD-1 adheres to the cell surface via
interaction with an as yet unidentified membrane anchoring protein (Lin et al., 2006). Since
this membrane anchoring protein is expected to be present in cadA-null cells, exogenously
added DdCAD-1 should bind to cadA-null cells. We therefore examined whether
exogenously added recombinant DdCAD-1 could rescue the mutant phenotype of cadA-null

Figure 4.9 Enrichment of DdCAD-1-GFP at cell-cell contacts in the anterior zone of
slugs and culminants. (A-B) Differences in subcellular localization of DdCAD-1-GFP
in slugs. The act15::cadA-GFP;cadA- transfectants were developed on coverslips for 18
to 20 hours. DdCAD-1-GFP was enriched in cell-cell contacts in the anterior zone of the
l (A) hil DdCAD 1 GFP t tl i th t l f ll i thslug (A), while DdCAD-1-GFP was present mostly in the cytoplasm of cells in the
posterior zone (B). The areas boxed by dashed lines are shown at higher magnification in
the two adjacent panels. Bars, 20 µm. (C) Confocal image showing enrichment of
DdCAD-1-GFP in cell-cell contact regions in the anterior zone of -631::cadA-GFP;cadA-
culminant. ST, stalk tube. Bar, 20 µm. (D) Confocal mage of the sorus of a fixed fruiting
body, showing an abundance of DdCAD-1-expressing cells ion the upper cup (thin arrow)
and lower cup (thick arrow) regions. (E) Surface localization of DdCAD-1-GFP in the
anterior cells of act15::cadA-GFP;cadA- slugs: (a) cells dissociated from the anterior
zone, showing DdCAD-1-GFP enrichment in the cell-cell contact region (arrowhead); (b)
cells dissociated from the posterior region of slugs Bars 5 µm (F) Capping of DdCAD-cells dissociated from the posterior region of slugs. Bars, 5 µm. (F) Capping of DdCAD-
1-GFP in act15::cadA-GFP;cadA- cells. Capping was induced by incubating cells with
mouse anti-GFP mAb followed by Alexa-568-conjugated secondary antibody (red)
(arrowhead): (a) cells derived from the anterior zone; (b) cells derived from the posterior
zone. Bars, 5 µm.
179

ADdCAD-1-GFP phalloidinmerge pg
BDdCAD-1-GFP phalloidinmerge
C D
ST
E
a a
F
b b
180

181
cells. Recombinant His6-DdCAD-1 (wild-type) and His6-S(I+II) DdCAD-1 (containing
mutations in the Ca2+-binding pockets SI and SII) were purified from E. coli (Fig. 4.10A).
Protein blots showed that both proteins bound to cadA-null cells with similar efficiency (Fig.
4.10B). To monitor the cell sorting phenomenon, cells were incubated with Neutral Red for 5
minutes before the addition of recombinant protein. The wild-type His6-DdCAD-1 protein
rescued the cell sorting defect in cadA-null cells, whereas the mutant protein failed to do so
(Fig. 4.10C). It was evident that exogenously applied DdCAD-1 was sufficient in restoring
the wild-type cell sorting pattern in null cells and its rescue function was dependent of Ca2+.
To monitor the fate of the bound recombinant DdCAD-1 during development,
DdCAD-1 was labelled with the fluorescent probe Alexa-488 before adding to cells and the
distribution patterns of the labelled protein was followed by confocal microscopy. At the
aggregation stage, DdCAD-1 was associated primarily with the cell membrane (Fig.
4.10Da,b). The honeycomb patterns of early mound and tight mound structures showed that
the recombinant DdCAD-1 was present primarily in the cell-cell contact regions (Fig.
4.10Dc,d). However, in the center of the tight mound (Fig. 4.10Dd), diffuse fluorescence in
the cytoplasm became evident, providing the first sign of DdCAD-1 internalization. Also,
cells in the outermost layer of the mound structure showed a higher level of fluorescence
intensity associated with the cell membrane. At the slug stage, a lot of cells began to show
fluorescence in their cytoplasm, indicative protein internalization. Also, cells in the anterior
zone appeared to be more elongated, suggesting that they might be moving toward the tip
region. At the early culmination stage, DdCAD-1 was highly enriched in the contact regions
among cells in the anterior upper cup region and the periphery region, while DdCAD-1 was
mostly in the cytoplasm in the posterior cells. These results confirmed the extracellular role

A BA B
a b
C
% o
f slu
gs
1008060
4020
00 1 0 50 3 0 90 7 0 1 0 50 3 0 90 7
ba
R R0.1 0.50.3 0.90.7 0.1 0.50.3 0.90.7
Figure 4.10 Rescue of the cell sorting defects in cadA- cells by DdCAD-1 recombinant
protein. (A) Coomassie-stained gel profiles (a) and western blots (b) of purified His6-
DdCAD 1 and His S(I+II) recombinant proteins (1 25 g) (B) Cell l sates (25 g)DdCAD-1 and His6-S(I+II) recombinant proteins (1.25 µg). (B) Cell lysates (25 µg)
derived from slugs previously incubated with DdCAD-1 recombinant proteins at 3 µg/µl
were prepared for SDS-PAGE and protein blots were probed with rabbit antiserum against
DdCAD-1. (C) Cell sorting in the cadA- slugs was examined in the presence of
exogenous recombinant proteins. Cells were stained with Neutral Red and the length of
the anterior region was measured and the R values were determined. (a) cadA- slugs
developed in presence of 0.5 µg/µl (gray bars) and 3 µg/µl (black bars) of His6-DdCAD-1
protein in 100 µl of cell suspension. (b) cadA- slugs developed in the presence of 0.5
µg/µl (gray bars) and 3µg/µl (black bars) of His6-S(I+II) protein. (n = 200 to 300 slugs)
182
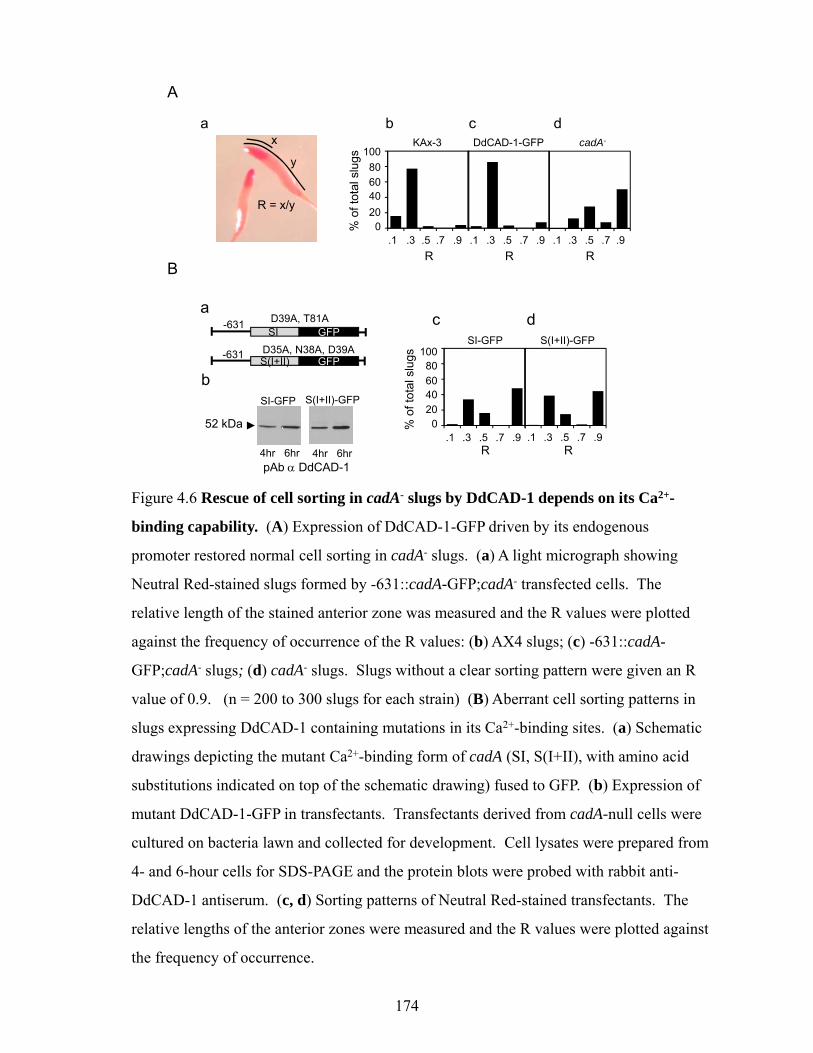
D
a b
*c d
e f
Figure 4.10 Rescue of the cell sorting defects in cadA- cells by DdCAD-1 recombinant
protein. (D) cadA- cells were mixed with 3 µg/µl of Alexa-488-conjugated His6-
DdCAD-1 (green) in 100 µl of cell suspension for development on coverslips. At 12
hours, excess buffer was removed and cells were developed for another 8 hours. Slugs at
different stages of development were prepared for confocal microscopy: (a) early
aggregation stage, (b) late aggregation stage; (c) early mound stage, (d) late mound stage,
(e) early slug stage, (f) slug stage. An enrichment of recombinant DdCAD-1 was
observed in the cell-cell contacts of anterior cells (arrow). Bars, 50 µm.
183

184
of DdCAD-1 in cell sorting. Additionally, posterior cells were able to internalize membrane-
bound DdCAD-1 at the slug stage.
Distinct chemotactic response of AX4 and cadA-null cells to cAMP
Differential chemotactic response to cAMP has been implicated in the sorting out of
prespore and prestalk cells (Jiang et al., 1998). When AX4 cells and cadA-null cells were
subjected to the chemotaxis assay, AX4 cells migrated at a rate about 20% slower than cadA-
null cells (Fig. 4.11A). These results would predict that AX4 cells took more time to reach the
aggregation center than cadA-null cells. Indeed, when cadA+ and cadA- cells were mixed at
equal numbers for development on agar, the cadA+ cells were found primarily in the
periphery of the mound structure (Fig. 4.11B).

50% cadA-GFP;cadA- : 50% cadA-
A B
* *on (%
) 120
100
*
Rel
ativ
e m
igra
tio
cadA- AX4
80
60
40
20
0
Fig. 4.11 AX4 and cadA- cells show distinct chemotactic response to cAMP. (A)
Relative chemotactic migration of cadA- and AX4 cells towards 100 µM (gray bars) or
250 M (bl k b ) f AMP D t t th ± S D ( 14 * < 0 05) (B)250 µM (black bars) of cAMP. Data represent the mean ± S.D. (n = 14; * p < 0.05). (B)
The act15::cadA-GFP;cadA- transfected cells were mixed at 1:1 ratio with cadA- cells for
development on coverslips. Developmental structures were fixed for confocal
microscopy. The pair of micrographs show the spatial distribution of act15::cadA-
GFP;cadA- cells (green) in a chimeric mound structure. Bars, 50 µm.
185

186
4.5 Discussion
Despite previous finding that DdCAD-1 appears only transiently during early cell
aggregation and becomes hardly detectable on the cell surface in the post aggregation stages
of development (Sesaki and Siu, 1996), studies presented in this paper have provided
evidence that DdCAD-1 can exert opposite influences on the social behaviour of
Dictyostelium depending on the environment encountered by the organism. When
development is carried out under conditions that mimicked its natural habitat, cadA behaves
as a single-gene green beard in chimeras and promote the survival of cells that express the
same gene product on the cell surface. However, on non-nutrient agar plates, cadA confers
anti-green beard effects on cells, allowing the cadA-null cells to behave as cheaters and
survive while the cadA+ cells undergo terminal differentiation to form stalk cells and
eventually die.
DdCAD-1 is expressed at high levels at the onset of development. During cell
streaming, DdCAD-1 is enriched especially on membrane protrusions such as lamellipodia
and filopodia, which are dynamic structures that are known to mediate initial contacts among
Dictyostelium cells (Choi and Siu, 1987; Sesaki and Siu, 1996) as well as mammalian cells
(Sandig et al., 1997; Vasioukhin et al., 2000). After entry into the cell stream, DdCAD-1
redistributes from the contact regions but remains associated with membranes on the outer
edge of the stream to make contacts with incoming cells (Sesaki and Siu, 1996). The loss of
DdCAD-1 would hinder the recruitment of cells into streams due to compromised cell-cell
adhesion. This defect is exacerbated by the presence of the rough and uneven surfaces of soil
particles, resulting in the delay of cell aggregation and subsequent developmental processes
(Fig. 4.2). In contrast, wild-types cells complete development in 24 hours on soil plates, just

187
as efficiently as development on agar surface. As the ability of the knockout cells to enter cell
streams is compromised on soil plates, chimeric cell aggregates would contain mostly wild-
type cells, resulting in a significant reduction of cadA- spores. In this way, the cadA gene
confers green beard effects in the natural habitat and favours the survival of individuals that
express DdCAD-1.
However, cadA switches to anti-green beard behaviour in chimeras developed on agar
surfaces and promote the survival of the knockout cells. An explanation of the anti-green
effect would involve two intertwining developmental processes, cell sorting and cell
differentiation, both of which are defective in the cadA-null cells (Wong et al., 2002). In
order that the cadA-null cells may form spores, the wild-types cells have to sort out from the
knockout cells to occupy the anterior zone and differentiate into stalk cells. Both differential
cell cohesiveness and chemotactic migration have been implicated in the cell sorting process
(Jiang et al., 1998). Our results show that the cadA-null cells undergo faster chemotactic
migration than wild-type cells. Although the difference is small, it is significant and should
allow the knockout cells to reach the aggregation center earlier than the wild-type cells (Fig.
4.11). This should account for the abundance of wild-type cells in the periphery of chimeric
aggregates as illustrated in the schematic drawings in Figure 4.12.
Normally, cells that show greater cohesiveness for one another would stay in the core
while the less cohesive ones would sort to the periphery in chimeric aggregates (Steinberg,
1975; Steinberg and Gilbert, 2004; Steinberg and Takeichi, 1994). But, how could the
knockout cells remain in the core of the chimeric mounds? Interestingly, a similar sorting
pattern has been observed when wild-type cells and cadA-null cells are mixed and gyrated in
suspension cultures (Wong et al., 2002). Further analysis has revealed that a higher level of
csA/gp80 is expressed at the later stages of cell aggregation by the knockout cells. The

aggregation
mound
tipped moundlate culminant
Figure 4.12 Schematic drawings depicting the distribution of DdCAD-1 expressing
cells (green) in different developmental structures. During aggregation, DdCAD-1-
slug
expressing cells migrate slower than the knockout cells on the agar surface. Consequently,
the cadA+ cells (green dots) reach the aggregation center later than cadA- cells and
become localized primarily in the periphery of the mound. In tipped mounds, most
DdCAD-1-expressing cells sort out from the cadA- cells and become concentrated in the
tip region. Migrating slugs show similar pattern as the tipped mound with enrichment oftip region. Migrating slugs show similar pattern as the tipped mound with enrichment of
DdCAD-1-expressing cells in the anterior region. However, a small number of cells are
observed in the posterior region of the slug, and they may eventually differentiate into the
basal disc cells. Finally, DdCAD-1-expressing cells are enriched in the upper cup, lower
cup and basal disc regions at the late culminant stage. These observations reveal that
188
DdCAD-1-expressing cells preferentially differentiate into stalk cells while cadA-null
cells differentiate into spore cells resulting in the anti-green beard effect of the cadA gene.

189
increased level of csA/gp80 compensates for the loss of DdCAD-1 expression, allowing the
knockout cells to stay in the aggregation core due to higher mutual adhesion. On the other
hand, wild-type cells inside the core of the mound structure eventually sort out to the
periphery due to their lower level of csA/gp80 expression. Cells in the periphery of the
mound have the propensity of differentiating into prestalk cells and migrate to the anterior
zone of the slug (Siu et al., 1983). Eventaully, the cadA+ cells undergo terminal
differentiation and die in order to confer altruistic benefits to the knockout cells. Therefore,
the anti-green beard effects of the cadA gene in chimeras appear to rely on an intricate
balance in the expression of the various cell adhesion molecules. Since the loss of csA/gp80
expression elicits the precocious expression of TgrC1/LagC in csaA-knockout cells (Wang et
al., 2000), it is conceivable that this phenomenon may also account for the anti-green beard
behaviour of the csaA gene in chimeras of csaA-null cells and wild-type cells (Queller et al.,
2003).
The anti-green beard behaviour of cadA is also corroborated by rescue studies, which
involves the ectopic expression of DdCAD-1 in cadA-null cells. Ectopically expressed
DdCAD-1 also rescued the cell sorting defect observed in the knockout cells. In addition, the
sorting defect can be rescued by exogenous recombinant DdCAD-1. Rescued cells display
the normal ratio in the anterior-posterior pattern of slugs. In both types of experiments, the
cell sorting function of DdCAD-1 is dependent on the integrity of its Ca2+-binding sites.
Several mechanisms have been proposed for the formation of the anterior-posterior pattern in
Dictyostelium. This pattern may arise by the sorting out of prestalk cells primarily to the
anterior zone and the prespore cells to the posterior zone of the slug as a consequence of
differential cell adhesiveness (Araki et al., 1994; Leach et al., 1973; Siu et al., 1983;
Steinberg, 1975; Steinberg and Gilbert, 2004). Studies from several laboratories have

190
suggested that prestalk and prespore cells arise in a spatially random manner throughout the
aggregate and sorting results from cell-type specific alterations in adhesion (Abe et al., 1994;
Nicol et al., 1999). However, it has also been proposed that pattern can be generated by
position-dependent differentiation of the different cell types (Krefft et al., 1984; Wolpert,
1981). The positional information model states that the value related to a position in a
coordinate system determines cell differentiation (Wolpert, 1989; Wolpert and Szathmary,
2002). During Dictyostelium development, cells that are starved in different cell cycle phases
tend to reach the aggregate at different times and therefore occupy different positions within
the aggregate. Cells in the periphery, which are usually starved at S phase or early G2 phase,
differentiate mostly into prestalk cells, suggesting that a combination of cell cycle stage and
positional information dictates cell fate (Gomer and Firtel, 1987; Maeda et al., 2003; Weijer,
2009; Weijer et al., 1984; Zimmermann and Weijer, 1993). Although both wild-type and
cadA-null cells are capable of differentiating into the different cell types required for fruiting
body formation, the cadA-null cells achieve 100 % cheating and form spores exclusively in
chimeras. These results are consistent with the notion of position-dependent differentiation
during development.
Contrary to the previous report of the transient expression of DdCAD-1 on the cell
surface (Sesaki and Siu, 1996), results shown in this paper have revealed heterogeneity in the
temporal and spatial expression of DdCAD-1. Whereas most of the cells have internalized or
shed their membrane DdCAD-1, a distinct subset of cells retain the expression of DdCAD-1
on the surface whether DdCAD-1 is expressed ectopically or added exogenously. These cells
are localized in the periphery of the aggregate and eventually move to the anterior zone
occupying the tip region of the slug. Peripheral cells usually follow a circular path to move
around the mound to the tip region (Nicol et al., 1999; Siegert and Weijer, 1995). In

191
chimeras, the presence of DdCAD-1 on the surface of peripherally located wild-type cells
would help them pull other DdCAD-1-expressing cells out toward them during the rotational
movement, whereas prespore cells in the core would re-establish contact and continue to
rotate uninterrupted.
Our results provide evidence for a direct extracellular role of DdCAD-1 in cell sorting,
which in turn explains the anti-green beard behaviour of the cadA gene. Cell sorting during
development is achieved by differential subcellular distribution of DdCAD-1, although the
mechanism involved and its relationship with the differentiation signal remain unknown.
Cells in the anterior zone with high levels of DdCAD-1 on the cell surface display altruism
and differentiate into stalk cells and die, while the posterior cells with DdCAD-1 localized
mainly in the cytoplasm survive as spores. Remarkably similar observation has been made in
chimeras using different ratios of cadA+ and cadA- cells, where cadA+ cells becomes stalk
cells and display altruism while cadA- cells survive as spores. Additionally, in vitro rescue
experiments have revealed that cadA+ cells that retain DdCAD-1 in the contact region move
into the anterior zone and become prestalk cells. Therefore, the anti-green beard effects of
cadA are evident in chimeras as well as in normal development of wild-type cells.
Dictyostelium express several adhesion systems to govern the social behavior of cells
during morphogenesis. Future studies on the mechanisms that regulate the spatiotemporal
expression of these proteins should lead to a better understanding of how cells communicate
and influence behavior and yield insights into the complexity of development.

192
Chapter 5
Conclusions and Future Directions

193
5.1 Conclusions
My doctoral research has investigated many aspects of the structural and functional
relationships of DdCAD-1 and has led to the mechanistic understanding of homophilic
adhesion, unconventional transport pathway, and cell sorting during Dictyostelium
morphogenesis.
5.1.1 Ca2+-dependent homophilic cell adhesion mediated by DdCAD-1
Although, earlier work (Wong et al., 1996) has suggested that DdCAD-1 could be a
distant homolog of cadherin, our current structural results indicate that DdCAD-1 may not be
evolutionarily related to the cadherin extracellular domain. First, DdCAD-1 consists of an N-
terminal domain of residues 2-93, a C-terminal domain of residues 98-213, and a short linker
of 4 residues connecting the two domains. The N-terminal domain comprises two Greek key
motifs, each with 4 β-strands and two short helical elements between the two motifs to yield a
two-layer sandwich architecture. The C-terminal domain is also a two-layer sandwich with 9
β-strands organized in two sheets. Initial comparisons based on the immunoglobulin-like
topology of the C-terminal domain of DdCAD-1 and the extracellular domain of the cadherin
showed some similarities. However, their β-strand connectivities are different (chapter 2).
Second, it was shown previously that the binding of Ca2+ to E-cadherin results in a rod-like
structure, which is crucial for cis-dimerization (Pertz et al., 1999). However, a similar
conformational change has not been observed in DdCAD-1 in the presence of Ca2+, although
the relative rearrangement of the two domains is slightly better defined by the binding of Ca2+
(chapter 2). Recent work by Harrison and coworkers (2005) suggests that structural changes
due to Ca2+ binding at the interface between two extracellular domains alter the exchange of

194
tryptophans between cadherins. A similar exchange of tryptophans does not occur in the case
for DdCAD-1. The cooperativity between Ca2+ binding sites reported in cadherin (Alattia et
al., 1997) is not present in DdCAD-1 since all three Kd values for the Ca2+-binding sites are
very similar. Furthermore, RMS (root-mean-square) coordinate difference of 3.8 Å found
between a cadherin domain and the C-terminal domain of DdCAD-1 is outside the range
found for related β-sandwich domains. Only one of the three Ca2+-binding sites is similar to
those in cadherin.
Based on our results (chapter 2), we propose that both hydrophobic and electrostatic
interactions mediate DdCAD-1 homophilic adhesion. Since the N-terminal domain alone
shows sufficient binding while the C-terminal domain retained only ~20% of the intact wild-
type binding capability, an examination of the binding interface suggests that hydrophobic
interactions could constitute the major force for adhesion. However, electrostatic interactions
may act as a ‘switch’ to regulate the homophilic binding by a change in electrostatic potential
caused by the binding of Ca2+ to the three binding sites. In the absence of Ca2+, electrostatic
repulsion between the N-terminal domain of one molecule and the C-terminal domain of the
other prevents two DdCAD-1 molecules from docking together. Ca2+ would greatly reduce
repulsion forces and the basic side chains surrounding the Ca2+ -binding sites in the N-
terminal domain would be involved in electrostatic interactions with the negative ones in the
C-terminal domain of the partner molecule. These interactions probably constitute the initial
binding reaction, followed by further stabilization of the dimer structure by hydrophobic
interactions between the N-terminal domain and its partner N- terminal domain. Dependence
on electrostatic interactions has been confirmed by the in vitro binding studies in the presence
of high salt which disrupted homophilic binding. These results support the proposal that
electrostatic interactions regulate binding, although the exact binding interface has not been

195
defined. In short, the NMR structure and dimeric model provide new insights into the unique
structure-function relationships of DdCAD-1, which are distinct from other Ca2+-dependent
adhesion proteins. This structural characterization of the Ca2+-binding sites and homophilic
interaction of DdCAD-1 provides the basis for future studies on its potential role in cell type
differentiation and cell sorting during development.
5.1.2 Mechanistic characterization of DdCAD-1 transport into contractile vacuole
Although DdCAD-1 is a soluble cytoplasmic protein, it has to be expressed on the cell
surface in order to function as a cell adhesion molecule. The second part of my thesis project
addresses the question of how DdCAD-1 is transported to the cell surface. Previously, Sesaki
et al. (2007) have demonstrated that DdCAD-1 is secreted by an unconventional transport
pathway involving contractile vacuoles. Unconventional protein secretion is an important but
poorly understood process in eukaryotes. These pathways might facilitate integration of
intracellular and extracellular functions by allowing flexible and dynamic regulation of
protein localization in response to various internal and external stimuli (Radisky et al., 2009).
Results derived from my thesis work show that the import of DdCAD-1 into contractile
vacuoles involves invagination of contractile vacuole membrane into the lumen (Fig. 5.1) and
the subsequent release by hypotonic lysis of the invaginated vesicles. However, neither the
N-terminal domain nor the C-terminal domain alone is sufficient for entry into the contractile
vacuole, suggesting that the integrity of DdCAD-1 is required for import (chapter 3). Binding
of Ca2+ leads to more ordered side chain packing in the Ca2+-binding sites and stabilizes the
whole structure of DdCAD-1. Import of DdCAD-1 was compromised when two of its three
Ca2+-binding sites were mutated, indicating a role for Ca2+ in the import process. The Ca2+-
binding sites might have a dual function. While contributions by Ca2+ to the surface charge

Extracellular region
C
N
Cytosol
?
Putative anchoringprotein X
X
XCVCV
Figure 5.1 Schematic diagram showing DdCAD-1 transport to the cell membrane.
The N-terminal domain is shown in light green color while the C-terminal domain is
shown in dark green color. The homophilic binding site that mediates cell-cell adhesion
is mainly located in the N-terminal domain of DdCAD-1 (N). Homophilic interaction of
DdCAD-1 is driven by changes in electrostatic potential caused by the binding of Ca2+ to
the 3 binding sites. These interactions are further stabilized by hydrophobic interactions.
The anchoring protein binding site is located in the C-terminal domain (C) and it is
Ca2+-independent. Both N- and C-terminal domains are required for DdCAD-1 transport
into contractile vacuoles. The import mechanism is dependent on Ca2+ and proper
protein conformation, suggesting that the secretory targeting motif of DdCAD-1 is in its
three dimensional structure. Time-lapse microscopic studies revealed that DdCAD-1 is
transported via invagination of contractile vacuole membrane.
196

197
can influence the docking of DdCAD-1 on to contractile vacuoles, conformational changes or
at least more ordered side chain packing induced by Ca2+-binding may regulate its import
into the lumen. If this is the case, then the import signal is likely integrated in the three-
dimensional structure of DdCAD-1. Other pathways that can accommodate folded substrates
without requiring a generalized signal for protein transport include ΔpH-dependent pathway
of plant plastids (Clark and Theg, 1997), the cytoplasm-to-vacuole targeting pathway of
Saccharomyces cerevisiae (Kim et al., 1997), and the direct translocation of proteins, such as
FGF-2 and galectin-1, across the plasma membrane in mammalian cells (Schatz and
Dobberstein, 1996; Wickner and Schekman, 2005). In many ways, the export of DdCAD-1
via contractile vacuoles is similar to the budding of vesicles in yeast vacuoles (Muller et al.,
2000) or the formation of multivesicular bodies in murine macrophages (Qu et al., 2007b).
However, whether ceramide/lipids plays a role in this invagination process as suggested in
multivesicular body formation (Trajkovic et al., 2008) remains to be elucidated.
5.1.3 cadA is a single-gene green beard that regulate morphogenesis through differential
cell adhesion in Dictyostelium
Specialized adhesive properties of cells have been shown to play a key role in
morphogenesis (Edelman, 1986). However, limited advance has been made in the
understanding of how adhesive interactions regulate or direct specific morphogenetic events
in vivo. Studies on cadA-null cells have implicated DdCAD-1 in cell sorting and cell-type
proportioning (Wong et al., 2002). The final part of my thesis research has yielded important
insights for the role of DdCAD-1 in Dictyostelium pattern formation (Fig. 4.12). We show
how the cadA gene which codes for a homophilic cell adhesion protein, acts as a single gene
green beard (Chapter 4). In chimera experiments, the cells expressing DdCAD-1 on the cell

198
surface were more likely recruited into aggregates than cadA-null cells, when development
was carried out on soil plates. Here, cadA behaved as a single gene green beard. However,
cadA exhibited anti-green beard behaviour when development was carried out on non-nutrient
agar. Wild-type cells differentiated mostly into prestalk cells and eventually died for the sake
of the organism whereas the cadA-null cells survived as spores. Morphological studies have
revealed remarkable differences in the temporal and spatial expression of DdCAD-1.
DdCAD-1 is enriched in the cell-cell contact regions in the anterior zone, but it is present
primarily in the cytoplasm of cells in the posterior zone. The aberrant cell sorting behaviour
of cadA-null cells can be rescued by the addition of recombinant DdCAD-1. Cells that move
into the anterior zone during tip formation retain DdCAD-1 in their contact regions, while
cells remaining in the posterior zone either shed or internalize their membrane-associated
DdCAD-1. These results convincingly demonstrate that DdCAD-1 plays a direct role in cell
sorting through differential cell-cell adhesion based on differential distribution of the protein.
Therefore, the presence of DdCAD-1 on the cell surface of prestalk cells is crucial for its role
in cell sorting, which in turn explains the anti-green beard effect observed in chimeras
containing cadA+ and cadA- cells.

199
5.2 Future Directions
My thesis project has revealed new insights into the structural and functional
relationships of DdCAD-1. However, in order to understand the precise mechanism of
DdCAD-1-mediated adhesion in Dictyostelium morphogenesis, several questions remain to be
addressed. I will focus on the following five major questions and propose some future
experiments. First, what is the mechanism by which DdCAD-1 is transported through
invagination of the contractile vacuole? Second, does calmodulin play a role in DdCAD-1
transport into the lumen of contractile vacuoles? Third, what is the anchoring protein of
DdCAD-1? Fourth, what is the mechanism involved in the differential distribution of
DdCAD-1 in the prestalk and prespore cells? Fifth, is DdCAD-1 present in the adherens
junction-like structures in the constriction region of the culminant?
5.2.1 What is the mechanism by which DdCAD-1 is transported through invagination of
the contractile vacuole?
DdCAD-1 is transported to the cell surface through invagination of contractile vacuole
membranes that form intraluminal vesicles which eventually lyse to release DdCAD-1 into
the lumen. This process might be similar to sorting of proteins such as EGFR into
multivesicular endosomes (MVEs) (Qu et al., 2007). Sorting of proteins into MVEs has been
shown to depend on ESCRT (endosomal sorting complex required for transport) (Williams
and Urbe, 2007). However, Trajkovir et al. (2008) reported recently that cargo is segregated
into distinct subdomains on the endosomal membrane and that the transfer of exosome-
associated domains into the lumen of the endosome did not depend on the function of the
ESCRT machinery, but required the sphingolipid ceremide. They show that the amount of

200
purified exosomes is reduced after the inhibition of neutral sphingomyelinases. These results
suggest an alternative pathway in intraendosomal membrane transport and exosome
formation. Proteins and lipids that are destined for lysosomal degradation are first
incorporated into intraluminal vesicles of MVEs and then delivered to lysosomes for
digestion. Alternatively, MVEs can directly fuse with the plasma membrane, which leads to
release of the intraluminal vesicles to the extracellular environment as exosomes. These data
support an ESCRT-independent pathway for sorting cargo into MVEs, which may depend on
raft-based microdomains for the lateral segregation of cargo within the endosomal membrane.
These microdomains may contain high concentrations of sphingolipids from which ceramides
are formed. Then, ceramide induces the coalescence of small microdomains into larger
domains, which in turn promote domain-induced budding. The cone-shaped structure of
ceramide might induce spontaneous negative curvature by creating an area difference between
the two membrane leaflets.
It is possible that DdCAD-1 mediated invagination of the contractile vacuole
membrane may use similar mechanisms through its interaction with lipids. Based on previous
observations, DdCAD-1 seems to be associated with lipids on the contractile vacuole of
membrane. In addition, Alexander et al. (2006) have shown that Dictyostelium has homologs
to most of the genes in sphingolipid synthesis and degradation pathway and consists of acidic
spingomyelinase (aSMase) even though neutral sphingomyelinase (nSMase) has not been
identified in Dictyostelium. To investigate the invagination mechanisms, one can test whether
sphingomyelinase participates in DdCAD-1 import using the pharmacological inhibitor
SR33557 to block the activity of acidic sphingomyelinase (Higuchi et al., 1996). If a
significant reduction is observed in the import of DdCAD-1 into contractile vacuoles in in
vitro assays, genetic approaches will be used to either knock out or knock down specific

201
enzymes such as acidic sphingomyelinase, ceramidase and dihydroceramide desaturase in the
sphingolipid synthesis pathway (Alexander et al., 2006). This will lead to the identification
of the signalling pathway involved in the invagination process. To further explore the role of
lipids in the formation of invagination, one can perform experiments using giant unilamelar
versicles (GUVs) that consist of known lipid composition as described in Trajkovi et al.
(2008). GUVs are particularly adapted to investigate membrane deformations, such as
membrane invagination, induced by mechanical constraints. Their size, ranging from 10 –
100 μm in diameter, with spherical smooth membrane surface is well suited for optical
microscopy studies (Solon et al., 2005). GUVs can be prepared from pure DOPC (1,2-
dioleoyl-sn-glycero-3-phosphocholine) or from a mixture of DOPC, DOPS (1,2-dioleoyl-sn-
glycero-3-[phosphor-1-serine]), SM (sphingomyelin) and sterol with electro-formation
technique according to Mathivet et al. (1996). DdCAD-1 can be incubated with negatively
charged GUVs to investigate the budding curvature formation inside the vesicle. The
experiment can be performed in an observation chamber, which consists of two glass
coverslips held together by a spacer of melted Parafilm. In order to limit nonspecific
adhesion of the vesicles on the glass, the bottom slide will be coated with a layer of Mal-PEG
covalently bound to the glass (Solon et al., 2005). Purified DdCAD-1 solution will be
transferred to the chamber and then GUV solution will be injected. The formation of the
curvature of the GUVs will be followed by a phase-contrast microscopy. These data should
provide further insights into the role of lipids in DdCAD-1 transport into contractile vacuole.
5.2.2 Does calmodulin play a role in DdCAD-1 transport to the contractile vacuole?
Calmodulin is bound to yeast vacuoles in a Ca2+-dependent manner and Ca2+ enhances
calmodulin binding to the vacuole membrane (Peters et al., 2001; Peters and Mayer, 1998). A

202
large proportion of the bound calmodulin was released from the vacuoles at Ca2+
concentrations below 500 nM, which includes the concentration range present in the cytosol
of the yeast cells (100 – 150 nM) (Peters and Mayer, 1998). Interestingly, calmodulin is also
localized to the contractile vacuole network and its association with the cytoplasmic surface
of the vacuolar membrane is sensitive to EGTA. Studies involving calmodulin antagonists
have suggested that this protein is important in endocytosis in mammalian cells, fusion of
early endosomes, recycling to the cell surface, delivery to the Golgi complex, and lysosomal
degradation of endocytosed immune complexes (Pryor et al., 2000). Localization of
intracellular calmodulin on both endosomal (Enrich et al., 1996) and lysosomal (Nielsen et
al., 1987) membranes has been reported. Interestingly, the observation that the calmodulin
antagonist W-7 inhibits late endosome–lysosome fusion is consistent with experiments on
yeast vacuole fusion where calmodulin plays a role on the postdocking events (Peters and
Mayer, 1998). Subsequent work has shown that calmodulin is a component of a high
molecular weight complex on the yeast vacuole that also contains protein phosphatase 1, an
essential protein for vacuolar bilayer mixing (Peters et al., 1999).
We have demonstrated (chapter 3) that Ca2+ is required for DdCAD-1 import into
contractile vacuoles. Bioinformatic analysis has suggested that DdCAD-1 consists of sixteen
1-10, including five 1-5-10 and fourteen 1-14, including five 1-8-14 and two 1-5-8-14, and
fourteen 1-16 Ca2+-dependent putative calmodulin binding motifs, suggesting a possible
direct interaction of DdCAD-1 with calmodulin (Rhoads and Friedberg, 1997). Therefore, we
propose that calmodulin plays a role in DdCAD-1 import into contractile vacuole by enabling
DdCAD-1 to dock and be imported more efficiently. In order to test this hypothesis, in vitro
import assays (chapter 3) will be performed in the presence of recombinant calmodulin to
examine its effect on import efficiency of DdCAD-1. In addition, pharmacological inhibitors

203
of calmodulin, such as compound 48/80 and W-7 will be used in import assays to investigate
the role of calmodulin in DdCAD-1 import. DdCAD-1 has been shown to interact with
calmodulin in in vivo binding assays (unpublished data). Whether DdCAD-1 directly
interacts with calmodulin will be tested using far western blot analysis. Once direct
interaction between DdCAD-1 and calmodulin is confirmed, the binding region for
calmodulin and the requirement of Ca2+ will be investigated using recombinant proteins, such
as His6-N, His6-C, and Ca2+-binding site mutants using in vitro binding assays.
The calmodulin binding site can be mapped using chemical cross-linking and mass
spectrometry analysis. Recombinant calmodulin will be covalently coupled with sulfo-SBED
cross-linker (Pierce). Sulfo-SBED is a trifunctional reagent that contains an amine- reactive
sulfo-NHS-ester on one arm, photo-reactive phenyl azide on the other side and a biotin
handle. The arm containing the sulfo-NHS-ester has a cleavable disulfide bond, which
permits transfer of the biotin component to any interacting proteins. This modified
calmodulin will be mixed with the recombinant DdCAD-1 and the biotin label will be
transferred to DdCAD-1 by photoactivation. Then the cross-linked material will be analyzed
by SDS-PAGE under reducing condition. The DdCAD-1 band will be excised, digested with
trypsin and then subjected to MALDI-TOF mass spectrometry. The identity of the modified
peptide based on the biotin mass (244.3 Da) difference will be determined by comparing the
peptide masses to the control DdCAD-1 peptides without modification (Howarth et al., 2006).
These results should demonstrate whether calmodulin enhances the efficiency of DdCAD-1
import into the contractile vacuole and provide better mechanistic understanding of this
unconventional transport pathway.
5.2.3 What is the anchoring protein for DdCAD-1?

204
The main puzzle in the cell adhesion function of DdCAD-1 is how it is tethered to the
plasma membrane? Based on the evidence that labelled recombinant protein is enriched in
the cell-cell contact regions of the prestalk cells (Fig. 4.10) it should be feasible to identify
and isolate the anchoring protein from these prestalk cells. Identification, isolation and
characterization of the membrane-anchoring complex will be crucial to achieve a better
understanding of how DdCAD-1 functions on the cell surface. One of the methods that we
can use to identify the putative anchoring protein is chemical cross-linking. First,
recombinant His6-DdCAD-1 protein will be covalently cross-linked with sulfo-SBED. Then
the labelled protein can be used to bind to the putative anchoring protein on differentiated
cadA-null cells. The interacting proteins will be cross-linked on the cell surface by
photoactivation. After chemical cross-linking, the DdCAD-1 interacting protein(s) will be co-
immunoprecipitated with DdCAD-1 or purified using streptavidin beads, and then analyzed
by SDS-PAGE and MALDI-TOF mass spectrometry. Alternatively, two-dimensional gels
can be used to identify cross-linked proteins in a complex. First, the proteins can be
fractionated on an SDS-gel in the absence of reducing agents. Then, a second dimension of
the SDS-gel is run after treatment of the gel with a reducing agent to cleave the central
disulfide bond. Proteins that are not cross-linked have the same mobility in both dimensions
of the SDS gel and will form a diagonal. However, proteins that are cross-linked migrate
slowly in the first dimension; after cleavage of the cross-link with mercaptoethanol, these
proteins migrate at their native positions in the second dimension and are off the diagonal
(Phizicky and Fields, 1995). Based on mass spectrometric analysis, the identity of each
protein will be determined by submitting peptide masses to database search for matches. This
approach should lead to the identification of the putative anchoring protein and other proteins
involved in linking of DdCAD-1 to the actin cytoskeleton. It is possible that the putative

205
anchoring protein might be interacting with adapter proteins in the cytoplasmic side of the
plasma membrane in order to interact with actin cytoskeleton. The candidate proteins will be
subject to the in vitro binding assays such as pull down experiments using His6 or GST tagged
DdCAD-1 protein or far western blot analysis to examine their binding specificity with
DdCAD-1.
A major disadvantage of using the chemical cross-linking method to investigate
protein-protein interactions is that it detects all adjacent (~22 Å) neighbours of DdCAD-1.
Any interaction detected by cross-linking should be independently assessed using direct
protein-protein binding assays. Also, cross-linking is a completely empirical process. One
cannot predict which proteins will be cross-linked by which reagents under certain conditions.
Several experimental conditions and reagents will be screened to achieve optimal cross-
linking. Nevertheless, cross-linking has three advantages over other methods. First, it can
identify weak interactions that would otherwise escape detections by other methods. Second,
it can be used to detect transient contacts with different proteins by freezing the cellular
process at different stages. Third, cross-linking can be done in vivo with membrane
permeable cross-linking reagents such as disuccinimidyl glutarate (DSG) and disuccinimidyl
suberate (DSS) (Phizicky and Fields, 1995).
5.2.4 What are the mechanisms involved in the differential distribution of DdCAD-1 in
the prestalk and prespore cells?
An interesting observation in the later stages of Dictyostelium development is that
DdCAD-1 is localized in the cytoplasm of the prespore cells and the externally added
recombinant DdCAD-1 is internalized in these cells. How is DdCAD-1 internalized in the
prespore cell? Also, it has been demonstrated that DdCAD-1 transiently appears on the cell

206
surface during the early stages of development and later replaced by gp80. However, the fate
of cell surface DdCAD-1 is unknown and it has been speculated that DdCAD-1 might be
internalized through endocytosis. Therefore, it will be of interest to investigate the
mechanisms involved in the process of DdCAD-1 internalization.
There are two possible routes whereby DdCAD-1 can be internalized in Dictyostelium
cells, via either the contractile vacuole network or the endocytic pathway. Although
contractile vacuole activity is significantly decreased in the later stages of development, it is
possible that DdCAD-1 can still be internalized through the contractile vacuole network
during the first 9 hours of development. To investigate the role of contractile vacuole in these
processes, time-lapse confocal imaging will be used. For microscopic analysis, His6-DdCAD-
1 will be labelled with sulfodicholorophenol esters (SDP) conjugated with Alexa fluor 488
(green) (Molecular Probes). First, cadA-null cells at the early aggregate stage will be
incubated with labelled DdCAD-1 and then fixed with paraformaldehyde and permeabilized
with Triton-X 100. Samples will be stained with calmodulin antibody as described in chapter
3 to monitor the contractile vacuoles. Co-localization of labelled DdCAD-1 with calmodulin
will suggest that DdCAD-1 might be internalized through contractile vacuoles. Then, cadA-
null cells, developed for at least 6 hours or with the slug stage prespore cells, will be
incubated with Alexa fluor-conjugated DdCAD-1 (green) and FM4-64 dye (red) to observe
the DdCAD-1 internalization into contractile vacuoles. FM4-64 dye labels the contractile
vacuole in vivo (Heuser et al., 1993). Finally, time-lapse confocal imaging will be performed
to examine the co-localization of FM4-64 (red) and the labelled DdCAD-1 (green) protein as
described in chapter 3. Since it is possible that FM4-64 can also label the endosomes if
incubated for more than 15 min, phase contrast images will be taken simultaneously to
confirm the presence of phase lucent images of the contractile vacuoles. If these experiments

207
show that Alexa fluor conjugated DdCAD-1 is associated with contractile vacuole, His6-
DdCAD-1 can be added externally and the contractile vacuoles can be isolated from these
cells. Whether His6-DdCAD-1 is present in the lumen will be determined after proteinase K
digestion.
Endocytosis pathways could be subdivided into three categories: clathrin-mediated
endocytosis, caveolar-mediated endocytosis, and macropinocytosis (Mukherjee et al., 1997).
Clathrin-mediated endocytosis is mediated by small vesicles that are mainly associated with
the cytosolic protein clathrin. Clathrin-coated vesicles are found in virtually all cells and
from domains of the plasma membrane termed clathrin-coated pits. Caveolar-mediated
endocytosis engrosses invagination of cholesterol-enriched microdomains within the plasma
membrane that may contain a coat protein, caveolin. These structures are referred to as lipid
rafts and/or caveolae (Bonifacino and Glick, 2004; Mukherjee et al., 1997). Interestingly,
caveolae-like structures with cholesterol-like sterol enrichment are found in Dictyostelium
(Xiao and Devreotes, 1997). Macropinocytosis, which usually occurs from highly ruffled
regions of the plasma membrane, is the invagination of the cell membrane to form a pocket,
which then pinches off into the cell to form a vesicle filled with large volume of extracellular
fluid and molecules within it (Bonifacino and Glick, 2004).
Many components of this clathrin-mediated endocytosis machinery exist in
Dictyostelium (Stavrou and O’Halloran, 2006). Endocytosis of fluid in Dictyostelium
depends largely on a pathway in which clathrin and actin are involved. In cell lines, where
clathrin heavy-chain expression is impaired by antisense techniques (Ruscetti et al., 1994) or
by gene disruption (Ruscetti et al., 1994), fluid-phase endocytosis is reduced by about 80% as
compared to the parental strain. Also, 3 mM cytochalasin A, which can dissociate the actin
cytoskeleton, is sufficient to inhibit fluid-phase uptake completely (Hacker et al., 1997).

208
To investigate whether the components of endosomal vesicles are involved in
DdCAD-1 internalization, cells will be incubated with Alexa fluor-conjugated DdCAD-1 and
at the same time with two ligands known to be internalised by clathrin-mediated endocytosis
(BODIPYFL-labelled transferrin and fluorescein-labelled Semliki Forest Virus (FLXSFV)) or
a fluorescent fluid phase marker (FITC-dextran). After internalisation, the overlap between
DdCAD-1 and the markers will be observed using fluorescent microscopy. Whether the
DdCAD-1-containing vesicles are distinct from classical endosomes will be tested with
antibodies against endosomal marker p80 in Dictyostelium (Ravanel et al., 2001). If co-
localization is observed with endosomal markers and DdCAD-1, then endosomes will be
purified using gradient centrifugation for further biochemical analysis. Once the particular
pathway involved in DdCAD-1 internalization is identified, further mechanistic details can be
derived by genetic and morphological approaches.
5.2.5 Is DdCAD-1 present in the adherens junction-like structures in the constriction
region of the culminant?
Grimson et al. (2000) have reported that adherens junction like structures are present
among a population of cells that surround the constriction region at the top of the stalk tube
(Fig. 1.9). In addition, a β-catenin homologue, Aar, has been localized to these structures.
The cell adhesion molecule that Aar binds to has not been identified. Although, it is unlikely
that Aar bind to DdCAD-1 directly, it is conceivable that it might bind to the putative
anchoring protein for DdCAD-1. Interestingly, DdCAD-1 is enriched in the constriction
region of the culminant. Therefore, it will be of interest to test whether DdCAD-1 is also
enriched in these junctions. In order to examine this, co-localization experiment can be
performed with antibodies raised against Aar protein. Alternatively, aar cDNA fused with

209
RFP in an expression vector can be transfected into the DdCAD-1-GFP transfected cadA-null
cells to investigate the co-localization and interactions of these proteins in vivo. Further,
electron microscopic studies after colloidal gold labelling can be performed to prove
unequivocally that DdCAD-1 is indeed present in these adherens junction-like structures.
After the identification of the anchoring protein for DdCAD-1, the interaction of
DdCAD-1 anchoring protein and Aar can be investigated using biochemical, immunological
and genetic approaches. For instance, in order to investigate the potential biological role of
the putative anchoring protein, its gene will be knocked out by homologous recombination
using the REMI (restriction enzyme mediated integration) approach (Kuspa and Loomis,
1992; Wong et al., 2002). Furthermore, cDNA for the anchoring protein will be fused with
GFP in an expression vector for expression in the knockout cells and the spatial and temporal
expression pattern will be analysed as described in chapter 4. Also, potential interaction
between the putative anchoring protein, DdCAD-1 and Aar will be investigated by co-capping
experiments. Upper cup cells will be isolated by manually cutting the tip of the culminants or
using specific membrane markers for these cells. Antibodies against DdCAD-1 and GFP will
be used to cap DdCAD-1 and putative anchoring protein, respectively. The cells will be
permeablized and stained with antibody against Aar to test whether DdCAD-1 and Aar or
anchoring protein and Aar are in the same complex. Alternatively, caps will be isolated from
these cells after mild detergent treatment and the presence of anchoring protein, DdCAD-1
and Aar in this complex can be confirmed by western blot analysis. Direct interaction
between putative anchoring protein and Aar will be tested in vitro by pull down assays using
recombinant proteins. If all these experiments are successful, the data will provide evidence
that DdCAD-1 together with its anchoring protein is present in these adherens junction-like

210
structures and show that DdCAD-1 indeed behaves as a remote distant homolog of cadherin
in this simple model organism.
5.3 Concluding Remarks
The highlights of my thesis are three-fold with DdCAD-1 in the centre of all these
findings. First, the dimer model proposed based on hydrophobic and electrostatic interaction
has contributed to the mechanistic understanding of homophilic adhesion. Second, on the
basis of quantitative in vitro and in vivo assays, advanced imaging techniques and genetic
studies, the molecular mechanisms of unconventional transport of DdCAD-1 is revealed. Our
findings imply that protein conformation plays a significant role in this unusual transport of
DdCAD-1 through the invagination of contractile vacuole membrane. Third, the long
standing question in developmental biology is how multiple cell types are formed and
undergo a coordinated series of morphogenetic cell movements to generate a patterned
organism. We have provided evidence for DdCAD-1 mediated cell sorting in Dictyostelium.
DdCAD-1 regulates social behaviour through its differential distribution in prestalk and
prespore cells, which eventually leads to cell sorting.

211
References

212
Abe, T., Early, A., Siegert, F., Weijer, C., and Williams, J. (1994). Patterns of cell movement within the Dictyostelium slug revealed by cell type-specific, surface labeling of living cells. Cell. 77:687-699.
Abedin, M., and King, N. (2008). The Premetazoan Ancestry of Cadherins. Science. 319:946-948. Agresti, A., and Bianchi, M.E. (2003). HMGB proteins and gene expression. Curr. Opin. Genet. Dev.
13:170. Alattia, J.R., Ames, J.B., Porumb, T., Tong, K.I., Heng, Y.M., Ottensmeyer, P., Kay, C.M., and Ikura,
M. (1997). Lateral self-assembly of E-cadherin directed by cooperative calcium binding. FEBS Lett. 417:405-408.
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., and Watson, J.D. (2007). Molecular biology of the cell. Garland Science, New York. 1728 pp.
Alexander, S., Min, J., and Alexander, H. (2006). Dictyostelium discoideum to human cells: Pharmacogenetic studies demonstrate a role for sphingolipids in chemoresistance. Biochim. Biophys. Acta. 1760:301-309.
Allesen-Holm, M., Barken, K.B., Yang, L., Klausen, M., and Webb, J.S. (2006). A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol. 59:1114-1128.
Andersson, D., Carlsson, U., and Freskgard, P. (2001). Contribution of tryptophan residues to the CD spectrum of the extracellular domain of human tissue factor. Eur. J. Biochem. 268:1118-1128.
Andrei, C., Dazzi, C., Lotti, L., Torrisi, M.R., Chimini, G., and Rubartelli, A. (1999). The secretory route of the leaderless protein interleukin 1b involves exocytosis of endolysosome-related vesicles. Mol. Biol. Cell. 10:1463-1475.
Andrei, C., Margiocco, P., Poggi, A., Lotti, L.V., Torrisi, M.R., and Rubartelli, A. (2004). Phospholipases C and A2 control lysosome-mediated IL-1beta secretion: implications for inflammatory processes. Proc. Natl. Acad. Sci. USA. 101:9745.
Angst, B.D., Marcozzi, C., and Magee, A.I. (2001). The cadherin superfamily. J. Cell Sci. 114:625-626.
Araki, T., Abe, T., Williams, J.G., and Maeda, Y. (1997). Symmetry breaking in Dictyostelium morphogenesis: evidence that a combination of cell cycle stage and positional information dictates cell fate. Dev. Biol. 192:645-648.
Araki, T., Nakao, H., Takeuchi, I., and Maeda, Y. (1994). Cell-Cycle-Dependent Sorting in the Development of Dictyostelium Cells. Dev. Biol. 162:221-228.
Arner, E.S., and Holmgren, A. (2000). Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 267:6102.
Aubry, L., and Firtel, R.A. (1999). Integration of signaling networks that regulate Dictyostelium differentiation. Ann. Rev. Cell Devel. Biol. 15:469-517.
Bagby, S., Harvey, T.S., Eagle, S.G., Inouye, S., and Ikura, M. (1994). Structural similarity of a developmentally regulated bacterial spore coat protein to beta gamma-crystallins of the vertebrate eye lens. Proc. Natl. Acad. Sci. USA. 91:4308-4312.
Baldauf, S.L., and Doolittle, W.F. (1997). Origin and evolution of the slime molds (Mycetozoa). Proc. Natl. Acad. Sci. USA. 94:12007-12012.
Baldauf, S.L., Roger, A.J., Wenk-Siefert, I., and Doolittle, W.F. (2000). A kingdom-level phylogeny of eukaryotes based on combined protein data. Science. 290:972-977.
Baldwin, T.A., and Ostergaard, H.L. (2001). Developmentally regulated changes in glucosidase II association with, and carbohydrate content of the protein tyrosine phosphatase CD45. J. Immunol. 167:3829-3835.

213
Balsamo, J., and Lilien, J. (1974). Functional identification of three components which mediate tissue-type specific embryonic cell adhesion. Nature. 251:522-524.
Barondes, S., Haywood-Reid, P., and Cooper, D. (1985a). Discoidin I, an endogenous lectin, is externalized from Dictyostelium discoideum in multilamellar bodies. J Cell Biol. 100:1825-1833.
Barondes, S.H., Haywood-Reid, P.L., and Cooper, D.N.W. (1985b). Discoidin I, an endogenous lectin, is externalized from Dictyostelium discoideum in multilamellar bodies. J. Cell Biol. 100:1825-1833.
Barton, N.H. (1990). Pleiotropic models of quantitative variation. Genetics. 124:773-782. Becker, M., Matzner, M., and Gerisch, G. (1999). Drainin required for membrane fusion of the
contractile vacuole in Dictyostelium is the prototype of a protein family also represented in man. EMBO J. 18:3305-3316.
Benabentos, R., Hirose, S., Sucgang, R., Curk, T., Katoh, M., Ostrowski, E.A., Strassman, J.E., Queller, D.C., Zupan, B., Shaulsky, G., et al. (2009). Polymorphic Members of the lag Gene Family Mediate Kin Discrimination in Dictyostelium. Curr. Biol. 19:567-572.
Bendtsen, J.D., Jensen, L.J., Blom, N., Von Heijne, G., and Brunak, S. (2004). Feature based prediction of non-classical and learderless protein secretion. Protein Eng. Des. Sel. 17:349-356.
Bendtsen, J.D., Kiemer, L., Fausboll, A., and Brunak, S. (2005). Non-classical protein secretion in bacteria. BMC Microbiol. 5:58.
Berks, M., and Kay, R.R. (1990). Combinatorial control of cell differentiation by cAMP and DIF-1 during development of Dictyostelium discoideum. Development. 110:977-984.
Berleman, J.E., Chumley, T., Cheung, P., and Kirby, J.R. (2006). Rippling Is a Predatory Behavior in Myxococcus xanthus. J. Bacteriol. 188:5888-5895.
Bernards, A. (2003). GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila Biochim. Biophys. Acta. 1603:47-82
Bertholdt, G., Stadler, J., Bozzaro, S., Fichtner, B., and Gerisch, G. (1985). Carbohydrate and other epitopes of the contact site A glycoprotein of Dictyostelium discoideum as characterized by monoclonal antibodies. Cell Diff. 16:187-202.
Beug, H., Gerisch, G., Kempff, S., Riedel, V., and Cremer, G. (1970). Specific inhibition of cell contact formation in Dictyostelium by univalent antibodies. Exp. Cell Res. 63:147-158.
Beug, H., Katz, F.E., and Gerisch, G. (1973). Dynamics of antigenic membrane sites relating to cell aggregation in Dictyostelium discoideum. J. Cell Biol. 56:647-658.
Beyenbach, K.W., and Wieczorek, H. (2006). The V-type H+ ATPase: molecular structure and function, physiological roles and regulation. J. Exp. Biol. 209:577-589.
Bianchi, M.E., and Agresti, A. (2005). HMG proteins: dynamic players in gene regulation and differentiation. Curr. Opin. Genet. Dev. 15:496.
Boggon, T.J., Murray, J., Chappuis-Flament, S., Wong, E., Gumbiner, B.M., and Shapiro, L. (2002). C-cadherin ectodomain structure and implications for cell adhesion mechanisms. Science. 296:1308-1313.
Boles, B.R., Thoendel, M., and Singh, P.K. (2005). Rhamnolipids mediate detachment of Pseudomonas aeruginosa from biofilms. Mol. Microbiol. 57:1210.
Bonifacino, J.S., and Glick, B.S. (2004). The mechanisms of vesicle budding and fusion. Cell. 116:153-166.
Bonner, J.T. (1988). The Evolution of Complexity by Means of Natural Selection. 272 pp. Bonner, J.T. (1998). The origins of multicellularity. Integr. Biol. 1:27-36.

214
Bowers-Morrow, V.M., Ali, S.O., and Williams, K.L. (2004). Comparison of molecular mechanisms mediating cell contact phenomena in model developmental systems: an exploration of universality. Biol. Rev. Camb. Philos. Soc. 79:611-642.
Brar, S.K., and Siu, C.-H. (1993). Characterization of the cell adhesion molecule gp24 in Dictyostelium discoideum. Mediation of cell-cell adhesion via a Ca2+-dependent mechanism. J. Biol. Chem. 268:24902-24909.
Brocard, C.B., Jedeszko, C., Song, H.C., Terlecky, S.R., and Walton, P.A. (2003). Protein structure and import into the peroxisomal matrix. Traffic. 4:74-82.
Brock, D.A., Ehrenman, K., Ammann, R., Tang, Y., and Gomer, R.H. (2003). Two Components of a Secreted Cell Number-counting Factor Bind to Cells and Have Opposing Effects on cAMP Signal Transduction in Dictyostelium. J. Biol. Chem. 278:52262-52272.
Brodland, G.W. (2004). Computational modeling of cell sorting, tissue engulfment, and related phenomena: A review. Appl. Mech. Rev. 57:47-77.
Burgess, J.G., Boyd, K.G., Armstrong, E., Jiang, Z., and Yan, L. (2003). The development of a marine natural product-based antifouling paint. Biofouling. 19 (Suppl.):197.
Bush, J., Nolta, K., Rodriguez-Paris, J., Kaufmann, N., O'Halloran, T., Ruscetti, T., Temesvari, L., Steck, T., and Cardelli, J. (1994). A Rab4-like GTPase in Dictyostelium discoideum colocalizes with V-H+-ATPases in reticular membranes of the contractile vacuole complex and in lysosomes. J. Cell Sci. 107:2801-2812.
Bush, J., Temesvari, L., Rodriguez-Paris, J., Buczynski, G., and Cardelli, J. (1996). A role for a Rab4-like GTPase in endocytosis and in regulation of contractile vacuole structure and function in Dictyostelium discoideum. . Mol. Biol. Cell. 7:1623-1638.
Cali, T., Galli, C., Olivari, S., and Molinari, M. (2008). Segregation and rapid turnover of EDEM1 by and autophagy-like mechanism modulates standard ERAD and folding activities. Biochem. Biophys. Res. Commun. 371:405-410.
Carreira, C.M., LaVallee, T.M., Tarantini, F., Jackson, A., and Lathrop, J.T. (1998). S100A13 is involved in the regulation of fibroblast growth factor-1 and p40 synaptotagmin-1 release in vitro. J. Biol. Chem. 273:22224.
Carroll, S.B. (2001). Chance and necessity: the evolution of morphological complexity and diversity. Nature. 409:1102-1109.
Caterina, M.J., and Devreotes, P.N. (1991). Molecular insights into eukaryotic chemotaxis. FASEB J. 5:3078-3085.
Chen, T.L.L., Wolf, W.A., and Chisholm, R.L. (1998). Cell-Type-Specific Rescue of Myosin Function During Dictyostelium Development Defines Two Distinct Cell Movements Required for Culmination. Development. 125:3895-3903.
Chen, X.J., and Gumbiner, B.M. (2006). Paraxial protocadherin mediates cell sorting and tissue morphogenesis by regulating C-cadherin adhesion activity. J. Cell Biol. 174:301-313.
Chisholm, R.L., and Firtel, R.A. (2004). Insights into morphogenesis from a simple developmental system. Nat. Rev. Mol. Cell Biol. 5:531-541.
Cho, M., and Cummings, R.D. (1995). Galectin-1, a beta-galactoside-binding lectin in Chinese hamster ovary cells. II. Localization and biosynthesis. J. Biol. Chem. 270:5207.
Choi, A.H.C., and Siu, C.-H. (1987). Filopodia are enriched in a cell cohesion molecule of Mr 80,000 and participate in cell-cell contact formation in Dictyostelium discoideum. J. Cell Biol. 104:1375-1387.
Clark, P., Liu, Z., Zhang, J., and Gierasch, L. (1996). Intrinsic tryptophans of CRABPI as probes of structure and folding. Prot. Sci. 5:1108-1117.

215
Clark, S., and Theg, S. (1997). A Folded Protein Can Be Transported Across the Chloroplast Envelope and Thylakoid Membranes. Mol Biol Cell. 8 923-934.
Clarke, M., Dominguez, N., Yuenb, I.S., and Gomer, R.H. (1992). Growing and starving Dictyostelium cells produce distinct density-sensing factors. Dev. Biol. 152:403-406.
Clarke, M., and Heuser, J. (1997). Water and ion transport. In Maeda,Y., Inouye,K. and Takeuchi,I. (eds), Dictyostelium. A Model System for Cell and Developmental Biology. University Academic Press, Inc., Tokyo. 75-91 pp.
Clarke, M., and Maddera, L. (2004). Distribution of alkaline phosphatase in vegetative Dictyostelium cells in relation to the contractile vacuole complex. Eur. J. Cell Biol. 83:289-296.
Clarke, M., Yang, J., and Kayman, S.C. (1988). Analysis of the prestarvation response in growing cells of Dictyostelium discoideum. Dev. Gen. 9:315 -326.
Cleves, A., Cooper, D., Barondes, S., and Kelly, R. (1996). A new pathway for protein export in Saccharomyces cerevisiae. J. Cell Biol. 133:1017-1026.
Cleves, A.E. (1997). Protein transports: the nonclassical ins and outs. Curr. Biol. 7:R318-320. Cline, K., and McCaffery, M. (2007). Evidence for a dynamic and transient pathway through the Tat
protein transport machinery. EMBO J. 26:3039-3049. Clout, N.J., Kretschmar, M., Jaenicke, R., and Slingsby, C. (2001). Crystal structure of spherulin S3a
homodimer in the calcium-loaded form sheds light on the evolution of the eye lens βγ-crystallin Greek-key domain fold. Structure. 9:115-124.
Clow, P.A., Chen, T., Chisholm, R.L., and McNally, J.G. (2000). Three-dimensional in vivo analysis of Dictyostelium mounds reveals directional sorting of prestalk cells and defines a role for the myosin II regulatory light chain in prestalk cell sorting and tip protrusion. Development. 127:2715 - 2728.
Coates, J.C., Grimson, M.J., Williams, R.S., Bergman, W., Blanton, R.L., and Harwood, A.J. (2002). Loss of the beta-catenin homologue aardvark causes ectopic stalk formation in Dictyostelium. Mech. Dev. 116:117-127.
Coates, J.C., and Harwood, A.J. (2001). Cell-cell adhesion and signal transduction during Dictyostelium development. J. Cell Sci. 114:4349-4358.
Cooper, D., and Barondes, S. (1990). Evidence for export of a muscle lectin from cytosol to extracellular matrix and for a novel secretory mechanism. J. Cell Biol. 110:1681-1691.
Cordell, P.A., Kile, B.T., Standeven, K.F., Josefsson, E.C., Pease, R.J., and Grant, P.J. (2010). Association of coagulation factor XIII-A with Golgi proteins within monocyte-macrophages: implications for subcellular trafficking and secretion. Blood. 115:2674-2681.
Cosson, P., Zulianello, L., Join-Lambert, O., Faurisson, F., and Gebbie, L. (2002). Pseudomonas aeruginosa virulence analyzed in a Dictyostelium discoideum host system. J. Bacteriol. 184:3027.
Coukell, M.B., Cameron, A.M., and Adames, N.R. (1992). Involvement of intracellular calcium in protein secretion in Dictyostelium discoideum. J. Cell Sci. 103:371-380.
Cozens, R.M., Tuomanen, E., Tosch, W., Zak, O., Suter, J., and Tomasz, A. (1986). Evaluation of the bactericidal activity of ss-lactam antibiotics on slowly growing bacteria cultured in the chemostat. Animicrob. Agents Chemother. 29:797.
Crespi, B.J. (2001). The evolution of social behavior in microorganisms. Trends Ecol. Evol. 16:178-183.
Crowley, T., Nellen, R., and Comer, R. (1985). Phenocopy of discoidin I-minus mutants by antisense transformation in Dictyostelium. Cell. 43:633-641.
Cullen, P.J., and Sprague, G.F.J. (2002). The roles of bud-site-selection proteins during haploid invasive growth in yeast. Mol. Biol. Cell. 13:2990-3004.

216
Curtis, A.S.G. (1967). The Cell Surface : Its Molecular Role in Morphogenesis. Academic, New York. Damsky, C.H., Knudsen, K.A., and Buck, C.A. (1984). Integral membrane proteins in cell-cell and
cell-substratum adhesion. Plenum, New York/London. pp. 1-64 pp. Darwin, C. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of
Favoured Races in the Struggle for Life. John Murray, London. 502 pp. Davidson, L.A. (2008). Taming the tiger of tissue aggregation: how epithelia control structural
assembly of underlying cells. Dev. Cell. 14:152-154. Dawkins, R. (1976). The Selfish Gene. 224 pp. De Lozanne, A. (2003). The role of BEACH proteins in Dictyostelium. Traffic. 4:6-12. Denny, P.W., Gokool, S., Russell, D.G., Field, M.C., and Smith, D.F. (2000). Acylation-dependent
protein export in Leishmania. J. Biol. Chem. 275:11017-11025. Desbarats, L., Lam, T.Y., Wong, L.M., and Siu, C.-H. (1992). Identification of a unique cAMP-
response element in the gene encoding the cell adhesion molecule gp80 in Dictyostelium discoideum. J. Biol. Chem. 267:19655-19664.
Devine, K.M., and Loomis, W.F. (1985). Molecular characterization of anterior-like cells in Dictyostelium discoideum. Dev. Biol. 107:364-372.
Devreotes, P. (1989). Cell-cell interactions in Dictyostelium development. Trends Genet. 5:242-245. Dickens, N.J., Beatson, S., and Ponting, C.P. (2002). Cadherin-like domains in a-dystroglycan, a/e-
sarcoglycan and yeast and bacterial proteins. Curr. Biol. 12:R197-R199. Diggle, S.P., Griffin, A.S., Campbell, G.S., and West, S.A. (2007). Cooperation and conflict in
quorum-sensing bacterial populations. Nature. 450:411-414. Dominguez, C., Boelens, R., and Bonvin, A.M. (2003). HADDOCK: a protein-protein docking
approach based on biochemical or biophysical information. J. Am. Chem. Soc. 125:1731-1737. Dormann, D., Siegert, F., and Weijer, C.J. (1996). Analysis of cell movement during the culmination
phase of Dictyostelium development. Development. 122:761-769. Dormann, D., Vasiev, B., and Weijer, C.J. (2002a). Becoming Multicellular by Aggregation; The
Morphogenesis of the Social Amoebae Dictyostelium discoideum. J. Biol. Phy. 28:765-780. Dormann, D., and Weijer, C.J. (2001). Propagating chemoattractant waves coordinate periodic cell
movement in Dictyostelium slugs. Development. 128:4535-4543. Dormann, D., and Weijer, C.J. (2006). Chemotactic cell movement during Dictyostelium development
and gastrulation. Curr. Opin. Gen. Dev. 16:367-373. Dormann, D., Weijer, G., Parent, C.A., Devreotes, P.N., and Weijer, C.J. (2002b). Visualizing PI3
kinase-mediated cell-cell signaling during Dictyostelium development. Curr. Biol. . 12:1178-1188.
Du, F., Edwards, K., Shen, Z., Sun, B., De Lozanne, A., Briggs, S., and Firtel, R.A. (2008). Regulation of contractile vacuole formation and activity in Dictyostelium. EMBO J. 27:2064 - 2076.
Duran, J.M., Anjard, C., Stefan, C., Loomis, W.F., and Malhotra, V. (2010). Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell Biol. . 188:4527-536.
Durston, A.J., Vork, F., and Weinberger, C. (1979). The control of later morphogenesis by chemotactic signals in Dictyostelim discoideum. In Biophysical and Biochemical Information Transfer in Recognition. J.G. Vassileva-Popova and E.V. Jensen, editors. Plenum, New York. 693-708.
Dynes, J.L., Clark, A.M., Caulks, G., Cusp, A., Loomis, W.F., and Firtel, R.A. (1994). LagC is required for cell-cell interactions that are essential for cell type differentiation in Dictyostelium. Genes Devel. 8:948-958.

217
Early, A., Abe, T., and Williams, J. (1995). Evidence for positional differentiation of prestalk cells and for a morphogenetic gradient in Dictyostelium. Cell. 83:91-99.
Edelman, G.M. (1976). Surface modulation in cell recognition and cell growth. Science. 192:218-226. Edelman, G.M. (1984). Modulation of cell adhesion during induction, histogenesis, and perinatal
development of the nervous system. Ann. Rev. Neurosci. 7:339-377. Edelman, G.M. (1985). Cell adhesion and the molecular processes of morphogenesis. Ann. Rev.
Biochem. 54:135-169. Edelman, G.M. (1986). Cell adhesion molecules in the regulation of animal form and tissue pattern
Ann. Rev. Cell BioI. 2:81- 116. Edelman, G.M., Gallin, W.J., Delouvee, A., Cunningham, B.A., and Thiery, J.-P. (1983). Early
epochal maps of two different cell adhesion molecules. Proc. Natl. Acad. Sci. USA. 80:4384-4388.
Eichinger, L., Pachebat, J.A., Glockner, G., Rajandream, M.A., Sucgang, R., Berriman, M., Song, J., Olsen, R., Szafranski, K., Xu, Q., et al. (2005). The genome of the social amoeba Dictyostelium discoideum. Nature. 435:43-57.
Elliott, G., and O’Hare, P. (1997). Intercellular trafficking and protein delivery by a herpesvirus structural protein. Cell. 88:223-233.
Engling, A., Backhaus, R., Stegmayer, C., Zehe, C., Seelenmeyer, C., Kehlenbach, A., Schwappach, B., Wegehingel, S., and Nickel, W. (2002). Biosynthetic FGF-2 is targeted to non-lipid raft microdomains following translocation to the extracellular surface of CHO cells. J Cell Sci. 115:3619-3631.
Enrich, C., Jackle, S., and Havel, R.J. (1996). The polymeric immunoglobulin receptor is the major calmodulin-binding protein in an endosome fraction from rat liver enriched in recycling recetor. Hepatology. 24:226-232.
Erlandsson, L., Andersson, K., Sigvardsson, M., Lycke, N., and Leanderson, T. (1998). Mice with an inactivated joining chain locus have perturbed IgM secretion. Eur. J. Immunol. 28:2355.
Europe-Finner, G.N., and Newell, P.C. (1986). Inositol 1,4,5-triphosphate induces calcium release from a non-mitochondrial pool in amoebae in Dictyostelium. Biochim. Biophys. Acta. 887:335-340.
Fabrizio, P., Battistella, L., Vardavas, R., Gattazzo, C., and Liou, L. (2006). Superoxide is a mediator of an altruistic aging program in Saccharomyces cerevisiae. J. Cell Biol. 166:1055-1067.
Fatal, N., Suntio, T., and Makarow, M. (2002). Selective protein exit from yeast endoplasmic reticulum in absence of functional COPII coat component Sec13p. Mol. Biol. Cell. 13:4130.
Featherstone, D.E., and Brodie, K. (2002). Wrestling with pleiotropy: genomic and topological analysis of the yeast expression network. BioEssays. 24:267-274.
Fiegna, F., and Velicer, G.J. (2006a). Exploitative and hierarchial antagonism in a cooperative bacteria. PLoS Biol. 3:e370.
Fiegna, F., and Velicer, G.J. (2006b). Exploitative and hierarchical antagonism in a cooperative bacteria. PLoS Biol. 3:e370.
Fiegna, F., Yu, Y.T.N., Kadam, S.V., and Velicer, G.J. (2006). Evolution of an obligate social cheater to a superior cooperator. Nature. 441:310-314.
Flaadt, H., Jaworski, E., and Malchow, D. (1993). Evidence for two intracellular calcium pools in Dictyostelium: the cAMPinduced calcium influx is directed into a NBD-Cl- and 2,5-di-(tert-butyl)-1,4-hydroquinone-sensitive pool. J. Cell Sci. 105:1131-1135.
Flieger, O., Engling, A., Bucala, R., Lue, H., Nickel, W., and Bernhagen, J. (2003). Regulated secretion of macrophage migration inhibitory factor is mediated by a non-classical pathway involving an ABC transporter. FEBS Letters. 551:78-86.

218
Fortunato, A., Strassmann, J.E., Santorelli, L., and Queller, D.C. (2003). Co-occurrence in nature of different clones of the social amoeba, Dictyostelium discoideum. Mol. Ecol. 12:1031-1038.
Fosnaugh, K.L., and Loomis, W.F. (1993). Enhancer regions for temporal and cell-type-specific expression of a spore coat gene in Dictyostelium. Dev. Biol. 157:38-48.
Foster, K.R., Parkinson, K., and Thompson, C.R.L. (2007). What can microbial genetics teach sociobiology? Trends in Genetics. 23:74-80.
Foster, K.R., Shaulsky, G., Strassmann, J.E., Queller, D.C., and Thompson, C.R.L. (2004). Pleiotropy as a mechanism to stabilize cooperation. Nature. 431:693-696.
Fotya, R.A., and Steinberg, M.S. (2005). The differential adhesion hypothesis: a direct evaluation. Dev. Biol. 278:255- 263.
Frank, S.A. (2003). Repression of competition and the evolution of cooperation. Evolution. 57:693-705.
Fuchs, M., Jones, M.K., and Williams, K.L. (1993a). Characterisation of an epithelium like layer of cells in the multicellular Dictyostelium discoideum slug. J. Cell Sci. 105:243-253.
Fuchs, M., Jones, M.K., and Williams, K.L. (1993b). Freeze-substitution for the preservation of cell-to-cell contact regions in die multicellular Dictyostelium discoideum slug. J. Comp. Asst. Microsc. 5:135-137.
Fukuzawa, M., Abe, T., and Williams, J.G. (2003). The Dictyostelium prestalk cell inducer DIF regulates nuclear accumulation of a STAT protein by controlling its rate of export from the nucleus. Development. 130:797-804.
Fukuzawa, M., and Williams, J.G. (2000). Analysis of the promoter of the cudA gene reveals novel mechanisms of Dictyostelium cell type differentiation. Development. 127:2705-2713.
Gabriel, D., Hacker, U., Kohler, J., Muller-Taubenberger, A., Schwartz, J.M., Westphal, M., and Gerisch, G. (1999). The contractile vacuole network of Dictyostelium as a distinct organelle: its dynamics visualized by a GFP marker protein. J. Cell Sci. 112:3995-4005
Gao, E.N., Shier, P., and Siu, C.-H. (1992). Purification and partial characterization of a cell adhesion molecule (gp150) involved in postaggregation stage cell-cell binding in Dictyostelium discoideum. J. Biol. Chem. 267:9409-9415.
Gardella, S., Andrei, C., Ferrera, D., Lotti, L.V., and Torrisi, M.R. (2002). The nuclear protein HMGB1 is secreted by monocytes via a nonclassical, vesicle-mediated secretory pathway. EMBO Rep. 3:995.
Gardner, A., West, S.A., and Buckling, A. (2004). Bacteriocins, spite and virulence. Proc. R. Soc. London Ser. B. 271:1529.
Gerald, N.J., Siano, M., and De Lozanne, A. (2002). The Dictyostelium LvsA protein is localized on the contractile vacuole and is required for osmoregulation. Traffic. 3:50-60.
Gerhart, J., and Keller, R. (1986). Region-specific cell activities in amphibian gastrulation. Ann. Rev. Cell BioI. 2:201-229.
Gerisch, G. (1961). Cell functions and change in cell function in the development of Dictyostelium discoideum. V. Stagespecific cell contact formation and its quantitative evaluation. Exp. Cell Res. 25:535-554.
Gerisch, G. (1986). Interrelation of cell adhesion and differentiation in Dictyostelium discoideum. J. Cell Sci. Suppl. 4:201-209.
Gerisch, G., Heuser, J., and Clarke, M. (2002). Tubular–vesicular transformation in the contractile vacuole system of Dictyostelium. Cell Biol. Int. 26:845-852.
Giglione, C., and Gross, J.D. (1995). Anion effects on vesicle acidification in Dictyostelium. Biochem. Mol. Biol. Int. 36:1057-1065.

219
Gilbert, O.M., Foster, K.R., Mehdiabadi, N.J., Strassmann, J.E., and Queller, D.C. (2007). High relatedness maintains multicellular cooperation in a social amoeba by controlling cheater mutants. Proc. Natl Acad. Sci. USA. 104:8913-8917.
Ginsburg, G.T., and Kimmel, A.R. (1997). Autonomous and nonautonomous regulation of axis formation by antagonistic signaling via 7-span cAMP receptors and GSK3 in Dictyostelium. Genes Dev. 11:2112-2123.
Gomer, R.H., and Firtel, R.A. (1987). Cell-autonomous determination of cell-type choice in Dictyostelium development by cell-cycle phase. Science. 237:758-762.
Gourlay, C.W., Du, W., and Ayscough, K.R. (2006). Apoptosis in yeast--mechanisms and benefits to a unicellular organism. Mol. Microbiol. 62:1515-1521.
Green, J.B.A. (2008). Sophistications of cell sorting. Nat. Cell Biol. 10:375-377. Griffin, A.S., Pemberton, J.M., Brotherton, P.N.M., McIlrath, G., Gaynor, D., Kansky, R., O'Riain, J.,
and Clutton-Brock, T.H. (2003). A genetic analysis of breeding success in the cooperative meerkat (Suricata suricatta). Behav. Ecol. 14:472-480.
Griffin, A.S., and West, S.A. (2003). Kin discrimination and the benefit of helping in cooperatively breeding vertebrates. Science. 302:634-636.
Griffin, A.S., West, S.A., and Buckling, A. (2004). Cooperation and competition in pathogenic bacteria. Nature. 430:1024-1027.
Grimson, M.J. 2000. A new view of culmination in Dictyostelium: Morphogenetic roles of cell shapes, intercellular junctions, and extracellular matrices In Biology. Vol. Doctor of Philosophy. Texas Tech University, Lubbock. 234.
Grimson, M.J., Coates, J.C., Reynolds, J.P., Shipman, M., Blanton, R.L., and Harwood, A.J. (2000). Adherens junctions and beta-catenin-mediated cell signaling in a non-metazoan organism. Nature. 408:727-731.
Grosberg, R.K., and Strathmann, R.R. (2007). The Evolution of Multicellularity: A Minor Major Transition? Ann. Rev. Ecol. Evol. Systematics. 38:621-654.
Gross, J.D. (1994). Developmental Decisions in Dictyostelium discoideum. Microbiol. Rev. . 58:330-351.
Grosshans, B.L., Ortiz, D., and Novick, P. (2006). Rabs and their effectors: achieving specificity in membrane traffic. Proc. Natl. Acad. Sci. USA. 103:11821-11827.
Hacker, U., Albrecht, R., and Maniak, M. (1997). Fluid-phase uptake by macropinocytosis in Dictyostelium. J. Cell Sci. 110:105-112.
Haig, D. (1996). "Gestational drive and the green-bearded placenta." Proc. Natl. Acad. Sci. USA. 93:6547-6551.
Hamilton, W.D. (1963). The evolution of altruistic behaviour. Am. Nat. 97:354-356. Hamilton, W.D. (1964). The genetical evolution of social behavior I and II. J. Theor. Biol. 7:1-52. Hamilton, W.D. (1970). Selfish and spiteful behavior in an evolutionary model. Nature. 228:1218. Hamilton, W.D. (1971). "Geometry for the Selfish Herd,". J. Theor. Biol. 31:295-311. Hamon, Y., Luciani, M.F., Becq, F., Verrier, B., Rubartelli, A., and Chimini, G. (1997). Interleukin-
1beta secretion is impaired by inhibitors of the Atp binding cassette transporter, ABC1. Blood. 90:2911.
Harris, A.K. (1976). Is cell sorting caused by differences in the work of intercellular adhesion? A critique of the steinberg hypothesis. J. Theor. Biol. 61:267-285.
Harris, E., Yoshida, K., Cardelli, J., and Bush, J. (2001a). Rab11-like GTPase associates with and regulates the structure and function of the contractile vacuole system in Dictyostelium. J. Cell Sci. 114:3035-3045.

220
Harris, T.J., Awrey, D.E., Cox, B.J., Ravandi, A., Tsang, A., and Siu, C.H. (2001b). Involvement of a triton-insoluble floating fraction in Dictyostelium cell-cell adhesion. J. Biol. Chem. 276:18640-18648.
Harris, T.J., Ravandi, A., Awrey, D.E., and Siu, C.H. (2003). Cytoskeleton interactions involved in the assembly and function of glycoprotein-80 adhesion complexes in Dictyostelium. J. Biol. Chem. 278:2614-2623.
Harris, T.J., Ravandi, A., and Siu, C.H. (2001c). Assembly of glycoprotein-80 adhesion complexes in Dictyostelium. Receptor compartmentalization and oligomerization in membrane rafts. J. Biol. Chem. 276:48764-48774.
Harris, T.J., and Siu, C.H. (2002). Reciprocal raft-receptor interactions and the assembly of adhesion complexes. Bioessays. 24:996-1003.
Harrison, O.J., Corps, E.M., and Kilshaw, P.J. (2005). Cadherin adhesion depends on a salt bridge at the N-terminus. J. Cell Sci. 118:4123-4130
Harwood, A., and Coates, J.C. (2004). A prehistory of cell adhesion. Curr. Opin. Cell Biol. 16:470-476.
Harwood, A.J., Hopper, N.A., Simon, M.N., Bouzid, S., Veron, M., and Williams, J.G. (1992). Multiple roles for cAMP-dependent protein kinase during Dictyostelium development. Dev. Biol. 149:90-99.
Hase, C., and Finkelstein, R.A. (1993). Bacterial extracellular zinc-containing metalloproteases. Microbiol. Rev. 57:823.
Hausman, R.E., and Moscona, A.A. (1976). Isolation of a retina-specific cell-aggregating factor from membranes of embryonic neural retina tissue. Proc. Natl. Acad. Sci. USA. 73:3594-3598.
He, W., Cowin, P., and Stokes, D.L. (2003). Untangling desmosomal knots with electron tomography. . Science. 302:109-113.
Hegde, R.S., and Bernstein, H.D. (2006). The surprising complexity of signal sequences. Trends Biochem. Sci. 31:563-571.
Helms, J.B., and Rothman, J.E. (1992). Inhibition by brefeldin A of a Golgi membrane enzyme that catalyses exchange of guanine nucleotide bound to ARF. Nature. 360:352-354.
Hettema, E.H., Distel, B., and F., T.H. (1999). Import of proteins into peroxisomes. Biochem. Biophys. Acta. 1451:17-34.
Heuser, J. (2006). Evidence for recycling of contractile vacuole membrane during osmoregulation in Dictyostelium amoebae-a tribute to Gunther Gerisch. Eur. J. Cell Biol. 85:859-871.
Heuser, J., Zhu, Q., and Clarke, M. (1993). Proton Pumps Populate the Contractile Vacuoles of Dictyostelium Amoebae. J. Cell Biol. 121:1311-1327.
Higuchi, M., Singh, S., Jaffrezou, J.-P., and Aggarwal, B.B. (1996). Acidic Sphingomyelinase-Generated Ceramide Not Sufficient for TNF-induced Apoptosis and is Needed But Nuclear Factor-rcB Activation'. J. Immun. 156:297-304.
Hoffman, S., Sorkin, R.C., White, P.C., Brackenbury, R., Mailhammer, R., and et aI. (1982). Chemical characterization of a neural cell adhesion molecule purified from embryonic brain membrane. J. Biol. Chem. 257:7720-7729.
Holm, L., and Sander, C. (1998). Touring protein fold space with Dali/FSSP. Nucleic Acids Res. 26:316-319.
Holtfreter, J. (1943). A study of the mechanics of gastrulation. Part I. J. Exp. Zool. 94:261-318. Holtfreter, J. (1944). A study o f the mechanics of gastrulation. Part II. J. Exp. Zool. 95:171-212. Holtfreter, J. (1948a). The mechanism of embryonic induction and its relation to parthenogenesis and
malignancy. . Cambridge Univ., Cambridge. 17 pp.

221
Holtfreter, J. (1948b). Significance of the cell membrane in embryonic processes. Ann. NY Acad. Sci. 49:709--760.
Howarth, M., Chinnapen, D.J.-F., Gerrow, K., Dorrestein, P.C., Grandy, M.R., Kelleher, N.L., El-Husseini, A., and Ting, A.Y. (2006). A monovalent streptavidin with a single femtomolar biotin binding site. Nature Methods. 3:267 - 273
Hughes, R.C. (1999). Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim. Biophys. Acta. 1473:172.
Hurst, L.D., Atlan, A., and Bengtsson, B.O. (1996). Genetic conflicts. Q. Rev. Biol. 71:317-364. Inouye, M., Inouye, S., and Zusman, D.R. (1979). Biosynthesis and self-assembly of protein S, a
development-specific protein of Myxococcus xanthus. Proc. Natl. Acad. Sci. USA. 76:209-213.
Jahn, R., and Scheller, R.H. (2006). SNAREs-engines for membrane fusion. Nature Rev. Mol. Cell Biol. 7:631-643.
Jain, R., Yuen, I.S., Taphouse, C.R., and Gomer, R.H. (1992). A density-sensing factor controls development in Dictyostelium. Genes Dev. 6:390-400.
Jalink, K., Moolenaar, W.H., and van Duijn, B. (1993). Lysophosphatidic acid is a chemoattractant for Dictyostelium discoideum amoebae. Proc. Natl. Acad. Sci. USA. 90:1857-1861.
Jiang, Y., Levine, H., and Glazier, J. (1998). Possible Cooperation of Differential Adhesion and Chemotaxis in Mound Formation of Dictyostelium. Biophysical J. 75:2615-2625.
Johnson, G., Johnson, R., Miller, M., Borysenko, J., and Revel, J. (1977). Do cellular slime molds form intercellular junctions? Science. 197:1300.
Jordens, I., Marsman, M., Kuijl, C., and Neefjes, J. (2005). Rab proteins, connecting transport and vesicle fusion. Traffic. 6:1070-1077
Jourdan, N., Maurice, M., Delautier, D., Quero, A.M., Servin, A.L., and Trugnan, G. (1997). Rotavirus is released from the apical surface of cultured human intestinal cells through nonconventional vesicular transport that bypasses the Golgi apparatus. J. Virol. 71:8268-8278.
Juschke, C., Ferring, D., Jansen, R.P., and Seedorf, M. (2004). A novel transport pathway for a yeast plasma membrane protein encoded by a localized mRNA. Curr. Biol. 14:406-411.
Juschke, C., Wachter, A., Schwappach, B., and Seedorf, M. (2005). SEC18/NSF-independent, protein-sorting pathway from the yeast cortical ER to the plasma membrane. J. Cell Biol. 169:613-622.
Kadam, S.V., and Velicer, G.J. (2006). Variable patterns of density-dependent survival in social bacteria. Behav. Ecol. 17:833-838.
Kaiser, D. (2001). Building a multicellular organism. Annu. Rev. Genet. 35:103-123. Kamboj, R.K., Gariepy, J., and Siu, C.-H. (1989). Identification of an octapeptide involved in
homophilic interaction of the cell adhesion molecule gp80 of Dictyostelium discoideum. Cell. 59:615-625.
Karecla, P.I., Green, S.J., Bowden, S.J., Coadwell, J., and Kilshaw, P.J. (1996). Identification of a binding site for integrin aEb7 in the N-terminal domain of E-cadherin. J. Biol. Chem. 271:30909-30915.
Karhinen, L., Bastos, R.N., Jokitalo, E., and Makarow, M. (2005). Endoplasmic reticulum exit of a secretory glycoprotein in the absence of Sec24p family proteins in yeast. Traffic. 6:562.
Kaufmann, G.F., Sartorio, R., Lee, S.H., Rogers, C.J., and Meijler, M.M. (2005). Revisiting quorum sensing: discovery of additional chemical and biological functions for 3-oxo-N-acylhomoserine lactones. Proc. Natl. Acad. Sci. USA. 102:309.

222
Kay, R.R., and Thompson, C.R.L. (2009). Forming Patterns in Development without Morphogen Gradients: Scattered Differentiation and Sorting Out. Cold Spring Harb. Perspect. Biol. 1:a001503.
Keller, L., and Ross, K.G. (1998). Selfish genes: a green beard in the red fire ant. Nature. 394:573-575.
Keller, L., and Surette, M.G. (2006). Communication in bacteria: an ecological and evolutionary perspective. Nat. Rev. Microbiol. 4:249.
Keller, M., Ruegg, A., Werner, S., and Beer, H.D. (2008). Active caspase-1 is a regulator of unconventional protein secretion. Cell. 132:818-831.
Keller, R.E., Danilchik, M., Gimlich, R., and Shih, J. (1985). The function and mechanism of convergent extension during gastrulation of Xenopus laevis. J. Embryol. Exp. Morphol. 89:185-209.
Kibler, K., Nguyen, T.L., Svetz, J., Van Driessche, N., Ibarra, M., Thompson, C.R., Shaw, C., and Shaulsky, G. (2003). A novel developmental mechanism in Dictyostelium revealed in a screen for communication mutants. Dev. Biol. 259:193-208.
Kim, J., Scott, S.V., Oda, M.N., and Klionsky, D.J. (1997). Transport of a large oligomeric protein by the cytoplasm to vacuole protein targeting pathway. J Cell Biol. 137:609-618.
Kim, L., Harwood, A., and Kimmel, A.R. (2002). Receptor-dependent and tyrosine phosphatase-mediated inhibition of GSK3 regulates cell fate choice. Dev. Cell. 3:523-532.
Kim, L., and Kimmel, A.R. (2000). GSK3, a master switch regulating cell-fate specification and tumorigenesis. Curr. Opin. Genet. Devel. 10:508-514.
King, N., Hittinger, C.T., and Carroll, S.B. (2003). Evolution of key cell signaling and adhesion protein families predates animal origins. Science. 301:361-363.
King, N., Westbrook, M.J., Young, S.L., Kuo, A., Abedin, M., Chapman, J., Fairclough, S., Hellsten, U., Isogai, Y., Letunic, v., et al. (2007). The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature. 451:783-788.
Kinseth, M.A., Anjard, C., Fuller, D., Guizzunti, G., Loomis, W.F., and Malhotra, V. (2007). The Golgi-associated protein GRASP is required for unconventional protein secretion during development. Cell. 130:524-534.
Kleene, R., and Schachner, M. (2004). Glycans and neural cell interactions. Nat. Rev. Neurosci. 5:195-208.
Knecht, D.A., Fuller, D.L., and Loomis, W.F. (1987). Surface glycoprotein, gp24, involved in early adhesion of Dictyostelium discoideum. Dev. Biol. 121:277-283.
Koch, A.W., Manzur, K.L., and Shan, W. (2004). Structure-based models of cadherin-mediated cell adhesion: the evolution continues. Cell. Mol. Life Sci. 61:1884-1895.
Kolter, R., and Greenberg, E.P. (2006). The superficial life of microbes. Nature. 441:300. Komissarchikf, Y.Y., and Snigirevskaya, E.S. (2002). Giant vacuoles arising during ADH-induced
transcellular bulk water flow across the epithelium of the frog urinary bladder. Cell Biology International. 26:873-883.
Krefft, M., Voet, L., Gregg, J.H., Mairhofer, H., and Williams, K.L. (1984). Evidence that positional information is used to establish the prestalk-prespore pattern in Dictyostelium discoideum aggregates. EMBO J. 3:201-206.
Kreitmeier, M., Gerisch, G., Heizer, C., and Muller-Taubenberger, A. (1995). A talin homologue of Dictyostelium rapidly assembles at the leading edge of cells in response to chemoattractant. J. Cell Biol. 129:179-188.
Kriebel, P.W., Barr, V.A., and Parent, C.A. (2003). Adenylyl cyclase localization regulates streaming during chemotaxis. Cell. 112:549-560.

223
Krieg, M., Arboleda-Estudillo, Y., Puech, P.H., Kafer, J., Graner, F., Muller, D.J., and Heisenberg, C.P. (2008). Tensile forces govern germ-layer organization in zebrafish. Nat. Cell Biol. 10:429-436.
Kuspa, A., and Loomis, W.F. (1992). Tagging developmental genes in Dictyostelium by restriction enzyme-mediated integration of plasmid DNA. Proc. Natl Acad. Sci. USA. 89:8803-8807.
Lam, T.Y., Pickering, G., Geltosky, J., and Siu, C.-H. (1981). Differential cell cohesiveness expressed by prespore and prestalk cells of Dictyostelium discoideum. Differentiation. 20:22-28.
LaVallee, T.M., Tarantini, F., Gamble, S., Carreira, C.M., Jackson, A., and Maciag, T. (1998). Synaptotagmin-1 is required for fibroblast growth factor-1 release. J. Biol. Chem. 273:22217.
Lazdunski, A.M., Ventre, I., and Sturgis, J.N. (2004). Regulatory circuits and communication in gram-negative bacteria. Nat. Rev. Microbiol. 2:581.
Leach, C.K., Ashworth, J.M., and Garrod, D.R. (1973). Cell sorting out during the differentiation of mixtures of metabolically distinct populations of Dictyostelium discoideum. J. Embryol. Exp. Morphol. 29:647-661.
Lecuit, T., and Lenne, P.-F. (2007). Cell surface mechanics and the control of cell shape, tissue patterns and morphogenesis. Nat. Rev. Mol. Cell Biol. 8:633-644.
Lewis, K. (2007). Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 5:48. Lin, Z., Huang, H., Siu, C.-H., and Yang, D. (2004). (1)H, (13)C and (15)N resonance assignments of
Ca(2+)-free DdCAD-1: a Ca(2+)-dependent cell-cell adhesion molecule. J Biomol NMR. 3:375-376.
Lin, Z., Sriskanthadevan, S., Huang, H., Siu, C.-H., and Yang, D. (2006). Solution structures of the adhesion molecule DdCAD-1 reveal new insights into Ca2+-dependent cell-cell adhesion. Nat Struct Mol Biol. 13:1016-1022.
Lodish, H., S, A.B., Zipursky, L., Matsudaira, P., Baltimore, D., and Darnell, J. (2000). Molecular Cell Biology. W.H. Freeman, New York. 1084 pp.
Loomis, W.F. (1998). Role of PKA in the Timing of Developmental Events in Dictyostelium Cells. Micro. Mol. Biol. Rev. 62:684-694.
MacKenzie, A., Wilson, H.L., Kiss-Toth, E., Dower, S.K., North, R.A., and Surprenant, A. (2001). Rapid secretion of interleukin-1beta by microvesicle shedding. Immunity. 15:825.
Maeda, M., Lu, S., Shaulsky, G., Miyazaki, Y., Kuwayama, H., Tanaka, Y., Kuspa, A., and Loomis, W.F. (2004). Periodic signaling controlled by an oscillatory circuit that includes protein kinases ERK2 and PKA. Science. 304:875-878.
Maeda, M., Sakamoto, H., Iranfar, N., Fuller, D., Maruo, T., Ogihara, S., Morio, T., Urushihara, H., Tanaka, Y., and Loomis, W.F. (2003). Changing Patterns of Gene Expression in Dictyostelium Prestalk Cell Subtypes Recognized by In Situ Hybridization with Genes from Microarray Analyses. Eukaryot. Cell. 2:627-637.
Malchow, D., Lusche, D.F., Schlatterer, C., Lozanne, A.D., and Muller-Taubenberger, A. (2006). The contractile vacuole in Ca2+-regulation in Dictyostelium:its essential function for cAMP-induced Ca2+-influx. BMC Dev. Biol. 6:31-38.
Mann, D., and Frankel, A. (1991). Endocytosis and targeting of exogenous HIV-1 Tat protein. EMBO J. 10:1733-1739.
Martiel, J.L., and Goldbeter, A. (1987). A model based on receptor desensitization for cyclic AMP signaling in Dictyostelium cells. Biophys. J. 52:807-828.
Maruyama, K., Mikawa, T., and Ebashi, S. (1984). Detection of calcium binding proteins by 45Ca autoradiography on nitrocellulose membrane after sodium dodecyl sulfate gel electrophoresis. J. Biochem. 95:511-519.

224
Mathivet, L., Cribier, S., and Devaux, P.F. (1996). Shape change and physical properties of giant phospholipid vesicles prepared in the presence of an AC electric field. Biophys. J. 70:1112-1121.
McClay, D.R., and Ettensohn, C.A. (1987). Cell Adhesion in Morphogenesis. Ann. Rev. Cell Biol. 3:319-45.
McLaughlin, S., and Murray, D. (2005). Plasma membrane phosphoinositide organization by protein electrostatics. Nature. 438:605-611.
McNeil, P.L., Muthukrishnan, L., Warder, E., and D'Amore, P.A. (1989). Growth factors are released by mechanically wounded endothelial cells. J. Cell Biol. 109:811.
McShea, D.W. (2002). A complexity drain on cells in the evolution of multicellularity. Evolution. 56:441-452.
Mehdiabadi, N.J., Jack, C.N., Farnham, T.T., Platt, T.G., Kalla, S.E., Shaulsky, G., Queller, D.C., and Strassmann, J.E. (2006). Social evolution: Kin preference in a social microbe. Nature. 442:881-882.
Mehul, B., and Hughes, R.C. (1997). Plasma membrane targetting, vesicular budding and release of galectin 3 from the cytoplasm of mammalian cells during secretion. J. Cell Sci. 110:1169.
Mellman, I., Fuchs, R., and Helenius, A. (1986). Acidification of the endocytic pathways. Annu. Rev. Biochem. 55:663-700.
Merkel, R., Simson, R., Simson, D.A., Hohenadl, M., Boulbitch, A., Wallraff, E., and Sackmann, E. (2000). A micromechanic study of cell polarity and plasma membrane cell body coupling in Dictyostelium. Biophys. J. 79:707-719.
Merrell, R., Gottlieb, D.I., and Glaser, L. (1975). Embryonal cell surface recognition. Extraction of an active plasma membrane component. J. Biol. Chern. 250:5655-5659.
Meyer, J.M., Stintzi, A., Vos, D.D., Cornellis, P., and Tappe, R. (1997). Use of siderophores to type pseudomonads: the three Pseudomonas aeruginosa pyoverdine systems. Microbiology. 143:35-43.
Mignatti P, M.T., Rifkin DB. . (1992). Basic fibroblast growth factor, a protein devoid of secretory signal sequence, is released by cells via a pathway independent of the endoplasmic reticulum-Golgi complex. J. Cell Physiol. 151:81-93.
Milne, J.L., and Coukell, M.B. (1988). Isolation and characterization of a plasma membrane calcium pump from Dictyostelium discoideum. Biochem. J. 249:223-230.
Misumi, Y., Misumi, Y., Miki, K., Takatsuki, A., Tamura, G., and Ikehara, Y. (1986). Novel blockade by brefeldin A of intracellular transport of secretory proteins in cultured rat hepatocytes. J. Biol. Chem. 261:11398.
Mohanty, S., Lee, S., Yadava, N., Dealy, M.J., Johnson, R.S., and Firtel, R.A. (2001). Regulated protein degradation controls PKA function and cell-type differentiation in Dictyostelium. Genes Dev. 15:1435-1448.
Moniakis, J., Coukell, M.B., and Forer, A. (1995). Molecular cloning of an intracellular P-type ATPase from Dictyostelium that is up-regulated in calcium-adapted cells. J. Biol. Chem. 270:28276-28281.
Moniakis, J., Coukell, M.B., and Janiec, A. (1999). Involvement of the Ca2+-ATPase PAT1 and the contractile vacuole in calcium regulation in Dictyostelium discoideum. J Cell Sci. 112:405-414.
Moon, R.T., Bowerman, B., Boutros, M., and Perrimon, N. (2002). The promise and perils of Wnt signaling through β-catenin. Science. 296:1644-1646.
Moscona, A.A. (1952). Cell suspensions from organ rudiments of chick embryos. Exp. Cell Res. 3:536-539.

225
Moscona, A.A. (1962). Analysis of cell recombinations in experimental synthesis of tissues in vitro. J. Cell Compo. Physiol. 60:65-80.
Müller, K., and Gerisch, G. (1978). A specific glycoprotein as the target site of adhesion blocking Fab in aggregating Dictyostelium cells. Nature. 274:445-449.
Muesch, A., Hartmann, E., Rohde, K., Rubartelli, A., Sitia, R., and Rapoport, T.A. (1990). A novel pathway for secretory proteins? Trends Biochem. Sci. 15:86.
Mukherjee, S., Ghosh, R.N., and Maxfield, F.R. (1997). “Endocytosis”. Physiol. Rev. 77:759-803. Muller, O., Sattler, T., Flotenmeyer, M., Schwarz, H., Plattner, H., and Mayer, A. (2000). Autophagic
tubes: vacuolar invaginations involved in lateral membrane sorting and inverse vesicle budding. J. Cell Biol. 151:519-528.
Muller, S., Scaffidi, P., Degryse, B., Bonaldi, T., and Ronfani, L. (2001). The double life of HMGB1 chromatin protein architectural factor and extracellular signal. EMBO J. 20:4337.
Newell, P.C., Telser, A., and Sussman, M. (1969). Alternative developmental pathways determined by environmental conditions in the cellular slime mold Dictyostelium discoideum. J. Bacteriol. 100:763-768.
Nickel, W. (2003). The mystery of nonclassical protein secretion. Eur J Biochem. 270:2109-2119. Nickel, W. (2005). Unconventional secretory routes: Direct protein export across the plasma
membrane of mammalian cells. Traffic. 6:607-614. Nickel, W. (2007). Unconventional secretion: an extracellular trap for export of fibroblast growth
factor 2. J. Cell Sci. 120:2295. Nickel, W., and Rabouille, C. (2009). Mechanisms of regulated unconventional protein secretion. Nat.
Rev. Mol. Cell Biol. 10:234-234. Nickel, W., and Seedorf, M. (2008). Unconventional mechanisms of protein transport to the cell
surface of eukaryotic cells. Annu. Rev. Cell Dev. Biol. 24:287-308. Nicol, A., Rappel, W.-J., Levine, H., and Loomis, W.F. (1999). Cell-sorting in aggregates of
Dictyostelium discoideum. J. Cell Sci. 112:3923-3929. Nielsen, T.B., Field, J.B., and Dedman, J.R. (1987). Association of calmodulin with lysosomes. J. Cell
Sci. 87:327-336. Niewohner, J., Weber, I., Maniak, M., Muller-Taubenberger, A., and Gerisch, G. (1997). Talin-null
cells of Dictyostelium are strongly defective in adhesion to particle and substrate surfaces and slightly impaired in cytokinesis. J. Cell Biol. 138:349-361.
Ninomiya, H., Elinson, R.P., and Winklbauer, R. (2004). Antero-posterior tissue polarity links mesoderm convergent extension to axial patterning. Nature. 430:364-367.
Noegel, A.A., and Schleicher, M. (2000). The Actin Cytoskeleton of Dictyostelium: A Story Told by Mutants. J. Cell Sci. 113:759-766.
Nolta, K.V., and Steck, T.L. (1994). Isolation and initial characterization of the bipartite contractile vacuole complex from Dictyostelium discoideum. J. Biol. Chem. 269:2225-2233.
Nudleman, E., Wall, D., and Kaiser, D. (2005). Cell-to-Cell Transfer of Bacterial Outer Membrane Lipoproteins. Science. 309:125-127.
O'Connor, K.A., and Zusman, D.R. (1988). Reexamination of the role of autolysis in the development of Myxococcus xanthus. J. Bacteriol. 170:4103-4112.
O'Connor, K.A., and Zusman, D.R. (1991). Behavior of peripheral rods and their role in the life cycle of Myxococcus xanthus. J. Bacteriol. 173:3342-3355.
O'Loughlin, E.V., and Robins-Browne, R.M. (2001). Effect of Shiga toxin and Shiga-like toxins on eukaryotic cells. Microbes Infect. 3:493.

226
Oppenheimer, S.B. (1975). Functional involvement of specific carbohydrate in teratoma cell adhesion factor. Exp. Cell Res. 92:122-126.
Orci, L., Tagaya, M., Amherdt, M., Perrelet, A., Donaldson, J., Klausner, R., and Rothman, J. (1991). Brefeldin A, a drug that blocks secretion, prevents the assembly of non-clathrin-coated buds on Golgi cisternae. Cell. 64:1183-1195.
Otto, S.P. (2004). Two steps forward, one step back: the pleiotropic effects of favoured alleles. Proc. R. Soc. Lond. B. 271:705-714.
Overduin, M., Harvey, T.S., Bagby, S., Tong, K.I., Yau, P., Takeichi, M., and Ikura, M. (1995). Solution structure of the epithelial cadherin domain responsible for selective cell adhesion. . Science. 267:386-389.
Ozaki, T., Nakao, H., Orii, H., Morio, T., Takeuchi, I., and Tasaka, M. (1993). Developmental regulation of transcription of a novel prespore-specific gene (Dp87) in Dictyostelium discoideum. Development. 117:1299 - 1308.
Padh, H., Lavasa, M., and Steck, T.L. (1989). Characterization of vacuolar proton ATPase in Dictyostelium discoideum. Biochim. Biophys. Acta. 982:271-278.
Palade, G.E. (1975). Intracellular aspects of the process of protein synthesis. Science. 189:347-358. Pang, K.M., Lynes, M.A., and Knecht, D.A. (1999). Variables controlling the expression level of
exogenous genes in Dictyostelium. Plasmid. 41:187-197. Parent, C.A., and Devreotes, P.N. (1996). Molecular genetics of signal transduction in Dictyostelium.
Annu. Rev. Biochem. 65:411-440. Parkinson, K., Bolourani, P., Traynor, D., Aldren, N.L., Kay, R.R., Weeks, G., and Thompson, C.R.
(2009). Regulation of Rap1 activity is required for differential adhesion, cell-type patterning and morphogenesis in Dictyostelium. J. Cell Sci. 122:335-344.
Parsek, M.R., and Greenberg, E.P. (2005). Sociomicrobiology: the connections between quorum sensing and biofilms. Trends Microbiol. 13:27-33.
Pearl, F., Todd, A., Sillitoe, I., Dibley, M., Redfern, O., Lewis, T., Bennett, C., Marsden, R., Grant, A., Lee, D., et al. (2005). The CATH Domain Structure Database and related resources Gene3D and DHS provide comprehensive domain family information for genome analysis. Nucleic Acids Res. 33:D247-D251.
Pelham, H.R. (1995). Sorting and retrieval between the endoplasmic reticulum and Golgi apparatus. Curr. Opin. Cell Biol. 7:530-535.
Pennisi, E. (2005). How Did Cooperative Behavior Evolve? Science. 309:93. Perret, E., Leung, A., Feracci, H., and Evans, E. (2004). Trans-bonded pairs of E-cadherin exhibit a
remarkable hierarchy of mechanical strengths. Proc. Natl. Acad. Sci. USA. 101:16472-16477. Pertz, O., Bozic, D., Koch, A.W., Fauser, C., Brancaccio, A., and Engel, J. (1999). A new crystal
structure, Ca2+ dependence and mutational analysis reveal molecular details of E-cadherin homoassociation. EMBO J. 18:1738-1747.
Peters, C., Andrews, P.D., Stark, M.J.R., Cesaro- Tadic, S., Glatz, A., Podtelejnikov, A., Mann, M., and Mayer, A. (1999). Control of the Terminal Step of Intracellular Membrane Fusion by Protein Phosphatase 1. Science. 285:1084 - 1087.
Peters, C., Bayer, M.J., Buhler, S., Andersen, J.S., Mann, M., and Mayer, A. (2001). Trans-complex formation by proteolipid channels in the terminal phase of membrane fusion. Nature. 409:581-588.
Peters, C., and Mayer, A. (1998). Ca2+/calmodulin signals the completion of docking and triggers a late step of vacuole fusion. Nature. 396:575-580.
Phizicky, E.M., and Fields, S. (1995). Protein-protein interactions: Methods for detection and analysis. Microbiol. Rev. 59:94-123.

227
Plyte, S.E., O’Donovan, E., Woodgett, J.R., and Harwood, A.J. (1999). Glycogen synthase kinase-3 (GSK-3) is regulated during Dictyostelium development via the serpentine receptor cAR3. Development. 126:325-333.
Ponte, E., Bracco, E., Faix, J., and Bozzaro, S. (1998). Detection of subtle phenotypes: the case of the cell adhesion molecule csA in Dictyostelium. Proc. Natl Acad. Sci. USA. 95:9360-9365.
Poole, T.J., and Steinberg, M.S. (1982). Evidence for the guidance of pronephric duct migration by a craniocaudally travelling adhesion gradient. Dev. BioI. 92:144-158.
Prudovsky, I., Mandinova, A., Soldi, R., Bagala, C., and Graziani, I. (2003). The nonclassical export routes: FGF1 and IL-1alpha point the way. J. Cell Sci. 116:4871.
Pryor, P.R., Mullock, B.M., Bright, N.A., Gray, S.R., and Luzio, J.P. (2000). The Role of Intraorganellar Ca2+ in Late Endosome-Lysosome Heterotypic Fusion and in teh Reformation of Lysosomes from Hybrid Organelles. J. Cell Biol. 149:1053-1062.
Pugsley, A.P. (1992). Translocation of a folded protein across the outer membrane in Escherichia coli. Proc. Natl. Acad. Sci. USA. 89:12058-12062.
Qu, Y., Franchi, L., Nunez, G., and Dubyak, G.R. (2007). Nonclassical IL-1 beta secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. J. Immunol. 179:1913-1925.
Queller, D.C. (1984). Kin selection and frequency dependence: a game theoretic approach. Biological Journal of Linnean Society. 23:133-143.
Queller, D.C. (2008). Behavioural ecology: The social side of wild yeast. Nature. 456:589-590. Queller, D.C., Ponte, E., Bozzaro, S., and Strassmann, J.E. (2003). Single-gene Greenbeard effects in
the social amoeba Dictyostelium discoideum. Science. 299:105-106. Radisky, D.C., Stallings-Mann, M., Hirai, Y., and Bissell, M.J. (2009). Single proteins might have
dual but related functions in intracellular and extracellular microenvironments. Nat. Rev. Mol. Cell Biol. 10:228-234.
Raper, K.B. (1940). Pseudoplasmodium formation and organization in Dictyostelium discoideum. Journal of the Elisha Mitchell Scientific Society. 56:241-282.
Rathi, A., Kayman, S.C., and Clarke, M. (1991). Induction of gene expression in Dictyostelium by prestarvation factor, a factor secreted by growing cells. Dev. Genet. 12:82-87.
Ravanel, K., Chassey, B.d., Cornillon, S., Benghezal, M., Zulianello, L., Gebbie, L., Letourneur, F., and Cosson, P. (2001). Membrane sorting in the endocytic and phagocytic pathway of Dictyostelium discoideum. Eur. J. Cell Biol. 80:754-764.
Reichenbach, H. (1999). The ecology of the myxobacteria. Environ. Microbiol. 1:15-21. Reiness, C.G., Seppa, M.J., Dion, D.M., Sweeney, S., Foster, D.N., and Nishi, R. (2001). Chick ciliary
neurotrophic factor is secreted via a nonclassical pathway. Mol. Cell Neurosci. 17:931-944. Reinhardt, A., and Hubbard, T. (1998). Using neural networks for prediction of the subcellular
location of proteins. Nucleic Acids Res. 26:2230-2236. Reintsch, W.E., Habring-Mueller, A., Wang, R.W., Schohl, A., and Fagotto, F. (2005). beta-Catenin
controls cell sorting at the notochord-somite boundary independently of cadherin-mediated adhesion. J. Cell Biol. 170:675-686.
Reithmeier, R.A.F. (1996). Assembly of proteins into membrane. In Biochemistry of Lipids, Lipoproteins and Membranes. Elsevier Science B. V., Amsterdam. 425-471 pp.
Reithrneier, R.A.F. (1996). Assembly of proteins into membrane. In Biochemistry of Lipids, Lipoproteins and Membranes. Elsevier Science B. V., Amsterdam. 425-471 pp.
Rhoads, A.R., and Friedberg, F. (1997). Sequence motifs for calmodulin recognition. FASEB J. 11:331-340.

228
Rietdorf, J., Siegert, F., and Weijer, C.J. (1998). Induction of optical density waves and chemotactic cell movement in Dictyostelium discoideum by microinjection of cAMP pulses. Dev. Biol. 204:525-536.
Riley, M.A., and Wertz, J.E. (2002). Bacteriocins: evolution, ecology, and application. Annu. Rev. Microbiol. 56:117-137.
Rokas, A. (2008). The molecular origins of multicellular transitions. Curr. Opin. Gen. & Dev. 18:472-478.
Rooney, E.K., and Gross, J.D. (1992). ATP-driven Ca2+/H+ antiport in acid vesicles from Dictyostelium. PNAS. 89:8025-8029.
Rooney, E.K., J.D., G., and Satre, M. (1994). Characterisation of an intracellular Ca2+ pump in Dictyostelium. Cell Calcium. 16:509-522.
Rosenberg, E., Keller, K.H., and Dworkin, M. (1977). Cell density-dependent growth of Myxococcus xanthus on casein. J. Bacteriol. 129:770-777.
Rothman, J.E. (1994). Mechanisms of intracellular protein transport. Nature. 372:55-63. Rothman, J.E., and Wieland, F.T. (1996). Protein sorting by transport vesicles. Science. 272:227-234. Rubartelli, A., Bajetto, A., Allavena, G., Wollman, E., and Sitia, R. (1992). Secretion of thioredoxin
by normal and neoplastic cells through a leaderless secretory pathway. J. Biol. Chem. 267:24161-24164.
Rubartelli, A., Cozzolino, F., Talio, M., and Sitia, R. (1990). A novel secretory pathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J. 9:1503-1510.
Ruscetti, T., Cardelli, J.A., Niswonger, M.L., and O’Halloran, T.J. (1994). Clathrin heavy-chain functions in sorting and secretion of lysosomalenzymes in Dictyostelium discoideum. J. Cell Biol. 126:343-352.
Russell, A.F., and Hatchwell, B.J. (2001). Experimental evidence for kin-biased helping in a cooperatively breeding vertebrate. Proc. R. Soc. Lond. B. 268:2169-2174.
Rutishauser, U., Hoffman, S., and Edelman, G.M. (1982). Binding properties of a cell adhesion molecule from neural tissue. Proc. Natl. Acad. Sci. USA. 79:685-689.
Sandig, M., Kalnins, V.I., and Siu, C.-H. (1997). Role of NCAM, cadherins, and microfilaments in cell-cell contact formation in TM4 immature mouse Sertoli cells. Cell Motil. Cytoskel. 36:149-163.
Schafer, T., Zentgraf, H., Zehe, C., Brugger, B., Bernhagen, J., and Nickel, W. (2004). Unconventional secretion of fibroblast growth factor 2 is mediated by direct translocation across the plasma membrane of mammalian cells. J. Biol. Chem. 279:6244-6251.
Schatz, G., and Dobberstein, B. (1996). Common principles of protein translocation across membranes. Science. 271:1519-1526.
Schindler, J., and Sussman, M. (1977). Ammonia determines the choice of morphogenetic pathways in Dictyostelium discoideum. J. Molec. Biol. 116:161-169.
Schmidt, M., Paes, K., Mazière, A.D., Smyczek, T., Yang, S., Gray, A., French, D., Kasman, I., Klumperman, J., Rice, D.S., et al. (2007). EGFL7 regulates the collective migration of endothelial cells by restricting their spatial distribution. Development. 134:2913-2923.
Schroder, H.C., Ushijima, H., Krasko, A., Gamulin, V., Thakur, N.L., Diehl-Seifert, B., Muller, I.M., and Muller, W.E. (2003). Emergence and disappearance of an immune molecule, an antimicrobial lectin, in basal metazoa. A tachylectin-related protein in the sponge Suberites domuncula. J. Biol. Chem. 278:32810-32817.
Seelenmeyer, C., Wegehingel, S., Tews, I., Kunzler, M., Aebi, M., and Nickel, W. (2005). Cell surface counter receptors are essential components of the unconventional export machinery of galectin-1. J. Cell Biol. 171:373-381.

229
Sesaki, H., and Siu, C.-H. (1996). Novel redistribution of the Ca2+-dependent cell adhesion molecule DdCAD-1 during development of Dictyostelium discoideum. Dev. Biol. 177:504-516.
Sesaki, H., Wong, E., and Siu, C.-H. (1997). The cell adhesion molecule DdCAD-1 in Dictyostelium is targeted to the cell surface by a nonclassical transport pathway involving contractile vacuoles. J Cell Biol. 138:939-951.
Shan, W., Yagita, Y., Wang, Z., Koch, A., Svenningsen, A.F., Gruzglin, E., Pedraza, L., and Colman, D.R. (2004). The minimal essential unit for cadherin-mediated intercellular adhesion comprises extracellular domains 1 and 2. J. Biol. Chem. 279:55914-55923.
Sharom, F.J., and Lehto, M.T. (2002). Glycosylphosphatidylinositol-anchored proteins: structure, function, and cleavage by phosphatidylinositolspecific phospholipase C. Biochem. Cell Biol. 80:535-549.
Sheetz, M.P. (2001). Cell control by membrane-cytoskeleton adhesion. Nature Rev. Mol. Cell Biol. 2:392-396.
Shelden, E., and Knecht, D.A. (1995). Mutants lacking myosin II cannot resist forces generated during multicellular morphogenesis. J. Cell Sci. 108:1105-1115.
Shur, B.D., and Roth, S. (1975). Cell surface glycosyltransferases. Biochim. Biophys. Acta. 415:473-512.
Siders, W.M., Klimovitz, J.C., and Mizel, S.B. (1993). Characterization of the structural requirements and cell type specificity of IL-1alpha and IL-1beta secretion. J. Biol. Chem. 268:22170.
Siegert, F., and Weijer, C.J. (1995). Spiral and concentric waves organize multicellular Dictyostelium mounds. Curr. Biol. 5:937-943.
Simon, M.N., Pelegrini, O., Veron, M., and Kay, R.R. (1992). Mutation of protein kinase-A causes heterochronic development of Dictyostelium. Nature. 356:171-172.
Simson, R., Wallraff, E., Faix, J., Niewohner, J., Gerisch, G., and Sackmann, E. (1998). Membrane bending modulus and adhesion energy of wildtype and mutant cells of Dictyostelium lacking talin or cortexillins. Biophys. J. 74:514-522.
Sinervo, B., Chaine, A., Clobert, J., Calsbeek, R., Hazard, L., Lancaster, L., McAdam, A.G., Alonzo, S., Corrigan, G., and Hochberg, M.E. (2006). Self-recognition, color signals, and cycles of greenbeard mutualism and altruism. Proc Natl Acad Sci USA. 103:7372-7377.
Siu, C.-H., Des Roches, B., and Lam, T.Y. (1983). Involvement of a cell surface glycoprotein in the cell sorting process of Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA. 80:6596-6600.
Siu, C.-H., Harris, T., Wang, J., and Wong, E. (2004a). Regulation of cell-cell adhesion during Dictyostelium development. Semin. Cell Dev. Biol. 15:633-641.
Siu, C.-H., Harris, T., Wang, J., and Wong, E. (2004b). Regulation of cell-cell adhesion during Dictyostelium development. Semin Cell Dev Biol. 15:633-641.
Siu, C.-H., Harris, T.J.C., Wang, J., and Wong, E. (2004c). Regulation of cell-cell adhesion during Dictyostelium development. Seminars in Cell & Developmental Biology. 15:633-641.
Siu, C.-H., Harris, T.J.C., Wong, E.F.S., Yang, C., Sesaki, H., and Wang, J. (1997). Cell adhesion molecules in Dictyostelium. In Dictyostelium - A Model System for Cell and Development Biology. Universal Academy Press, Tokyo.
Siu, C., Lam, T., and Choi, A. (1985). Inhibition of cell-cell binding at the aggregation stage of Dictyostelium discoideum development by monoclonal antibodies directed against an 80,000-dalton surface glycoprotein. J. Biol. Chem. 260:16030-16036.
Siu, C.H., Cho, A.S., and Choi, A. (1986). Mechanism of action of the contact site A glycoprotein during development of Dictyostelium discoideum J. Cell BioI. 103:3a.
Smith, J. (2001). The social evolution of bacterial pathogenesis. Proc. R. Soc. London Ser. B. 268:61.

230
Smukalla, S., Caldara, M., Pochet, N., Beauvais, A., Guadagnini, S., Yan, C., Vinces, M.D., Jansen, A., Prevost, M.C., Latgé, J.-P., et al. (2008). FLO1 Is a Variable Green Beard Gene that Drives Biofilm-like Cooperation in Budding Yeast. Cell. 135:726-737.
Soler-Lopez, M., Petosa, C., Fukuzawa, M., Ravelli, R., Williams, J.G., and Müller, C.W. (2004). Structure of an activated Dictyostelium STAT in its DNA-unbound form. Mol. Cell 13:791-804.
Sollner, T.H. (2004). Intracellular and viral membrane fusion: a uniting mechanism. Curr. Opin. Cell Biol. 16:429-435.
Solon, J., Gareil, O., Bassereau, P., and Gaudin, Y. (2005). Membrane deformations induced by the matrix protein of vesicular stomatitis virus in a minimalsystem. J. General Virol. 86:3357-3363.
Sordano C., Cristino, E., Bussolino, F., Wurster, B., and Bozzaro, S. (1993). Platelet activating factor modulates signal transduction in Dictyostelium. J. Cell Sci. 104:197-202.
Specht, B., Bartetzko, N., Hohoff, C., Kuhl, H., Franke, R., Börchers, T., and Spener, F. (1996). Mammary Derived Growth Inhibitor Is Not a Distinct Protein but a Mix of Heart-type and Adipocyte-type Fatty Acid-binding Protein. J. Biol. Chem. . 271:19943-19949.
Springer, W.R., and Barondes, S.H. (1985). Protein-linked oligosaccharide implicated in cell-cell adhesion in two Dictyostelium species. Dev. Biol. 109:102-110.
Springer, W.R., Cooper, D.N., and Barondes, S.H. (1984). Discoidin I is implicated in cell-substratum attachment and oriented cell migration of Dictyostelium discoideum and resembles fibronectin. Cell. 39:557-564.
Sriskanthadevan, S., Ivanov, I., Yang, C., and Siu, C.-H. (2007). Novel Functions and Transport mechanism associated with the Ca2+- dependent cell adhesion molecule DdCAD-1 in Dictyostelium. In Recent Research Developments in Cell Biology. Vol. 3. S.G. Pandalai, editor. Research Signpost, Trivandrum. 9-21.
Sriskanthadevan, S., Lee, T., Lin, Z., Yang, D., and Siu, C.-H. (2009). The Cell Adhesion Molecule DdCAD-1 Is Imported into Contractile Vacuoles by Membrane Invagination in a Ca2+- and Conformation-dependent Manner. J. Biol. Chem. 284:36377-36386.
Stavrou, I., and O’Halloran, T.J. (2006). The Monomeric Clathrin Assembly Protein, AP180, Regulates Contractile Vacuole Size in Dictyostelium discoideum. Mol. Biol. Cell. 17:5381-5389.
Stein, T., and Gerisch, G. (1996). Oriented Binding of a Lipid-Anchored Cell Adhesion Protein onto a Biosensor Surface Using Hydrophobic Immobilization and Photoactive Crosslinking. Analytical Biochemistry. 237:252-259.
Steinberg, M.S. (1970). Does differential adhesion govern self-assembly processes in histogenesis? Equilibrium configurations and the emergence of a hierarchy among populations of embryonic cells. J. Exp. Zool. 173:395-434.
Steinberg, M.S. (1975). Adhesion-guided multicellular assembly: A commentary upon the postulates, real and imagined, of the differential adhesion hypothesis, with special attention to computer simulations of cell sorting. J. Theor. Biol. 55:431-443.
Steinberg, M.S., and Gilbert, S.F. (2004). Townes and Holtfreter (1955): directed movements and selective adhesion of embryonic amphibian cells. J. Exp. Zool. A Comp. Exp. Biol. 301:701-706.
Steinberg, M.S., and Takeichi, M. (1994). Experimental specification of cell sorting, tissue spreading, and specific spatial patterning by quantitative differences in cadherin expression. Proc. Natl Acad. Sci. USA. 91:206-209.
Sternfeld, J., and David, C.N. (1982). Fate and regulation of anterior-like cells in Dictyostelium slugs. Dev. Biol. 93:111-118.

231
Stevens, A., and Søgaard-Andersen, L. (2005). Making waves: Pattern formation by a cell surface-associated signal. Trends Microbiol. 13:249-252.
Stinchcombe, J., Bossi, G., and Griffiths, G.M. (2004). Linking Albinism and Immunity: The Secrets of Secretory Lysosomes. Science. 305:55 - 59.
Strassmann, J.E., Zhu, Y., and Queller, D.C. (2000). Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature. 408:965-967.
Sukumaran, S., Brown, J.M., Firtel, R.A., and McNally, J.G. (1998). lagC-null and gbf-null cells define key steps in the morphogenesis of Dictyostelium mounds. Dev. Biol. 200:16-26.
Sundstrom, P. (2002). Adhesion in Candida spp. Cell Microbiol. 4:461-469. Swairjo, M.A., Concha, N.O., Kaetzel, M.A., Dedman, J.R., and Seaton, B.A. (1995). Ca2+-bridging
mechanism and phospholipid head group recognition in the membrane-binding protein annexin V. Nat. Struct. Biol. 2 968-974.
Taktakoff, A.M. (1983). Perturbation of vesicular traffic with the carboxylic ionophore monensin. Cell. 32:1026-1028.
Tandlea, A.T., Mazzantia, C., Alexandera, H.R., Robertsb, D.D., and Libutti, S.K. (2005). Endothelial monocyte activating polypeptide-II induced gene expression changes in endothelial cells. Cytokine. 30:347-358.
Tang, L., Ammann, R., Gao, T., and Gomer, R.H. (2001). A Cell Number-counting Factor Regulates Group Size in Dictyostelium by Differentially Modulating cAMP-induced cAMP and cGMP Pulse Sizes. J. Biol. Chem. 276:27663-27669.
Tang, L., Gao, T., McCollum, C., Jang, W., Vicker, M.G., Ammann, R.R., and Gomer, R.H. (2002). A cell number-counting factor regulates the cytoskeleton and cell motility in Dictyostelium. Proc. Natl. Acad. Sci. USA. 99:1371-1376.
Tarantini, F., Micucci, I., Bellum, S., Landriscina, M., and Garfinkel, S. (2001). The precursor but not the mature form of IL1alpha blocks the release of FGF1 in response to heat shock. J. Biol. Chem. 276:5147.
Tasaka, M., and Takeuchi, I. (1981). Role of cell sorting in pattern formation in Dictyostelium discoideum. Differentiation. 18:191-196.
Teintze, M., Inouye, M., and Inouye, S. (1988). Characterization of calcium-binding sites in development-specific protein S of Myxococcus xanthus using site-specific mutagenesis. J. Biol. Chem. 263:1199-1203.
Temmerman, K., Ebert, A.D., Mller, H.M., Sinning, I., Tews, I., and Nickel, W. (2008). A direct role for phosphatidylinositol 4,5 bisphosphate in unconventional secretion of fibroblast growth factor 2. Traffic. 9:1204.
Teter, S.A., and Klionsky, D.J. (1999). How to get a folded protein across a membrane. Trends Cell Biol. 9:428-431.
Thery, C., Boussac, M., Veron, P., Ricciardi-Castagnoli, P., and Raposo, G. (2001). Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J. Immunol. 166:7309.
Thomason, P.A., Traynor, D., Cavet, G., Chang, W.T., Harwood, A.J., and Kay, R.R. (1998). An intersection of the cAMP/PKA and two-component signal transduction systems in Dictyostelium. EMBO J. 17:2838-2845.
Thompson, C.R.L., Fu, Q., Buhay, C., Kay, R.R., and Shaulsky, G. (2004). A bZIP/bRLZ transcription factor required for DIF signaling in Dictyostelium. Development. 131:513-523.
Thompson, C.R.L., and Kay, R.R. (2000). Cell-fate choice in Dictyostelium: intrinsic biases modulate sensitivity to DIF signaling. Dev. Biol. 227:56-64.

232
Townes, P.L., and Holtfreter, J. (1955). Directed movements and selective adhesion of embryonic amphibian cells. J. Exp. Zool. 128:53-120.
Trajkovic, K., Hsu, C., Chiantia, S., Rajendran, L., Wenzel, D., Wieland, F., Schwille, P., Brügger, B., and Simons, M. (2008). Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science. 319:1244-1247.
Travisano, M., and Velicer, G.J. (2004). Strategies of microbial cheater control. Trends Microbiol. 12:72-78.
Tsujioka, M., Machesky, L.M., Cole, S.L., Yahata, K., and Inouye, K. (1999). A unique talin homologue with a villin headpiece-like domain is required for multicellular morphogenesis in Dictyostelium. Curr. Biol. 9:389-392.
Tsujioka, M., Yoshida, K., and Inouye, K. (2004). Talin B is required for force transmission in morphogenesis of Dictyostelium. EMBO J. 23:2216-2225.
Turing, A.M. (1952). The chemical basis of morphogenesis. Phi/os. Trans. R. Soc. London Ser. B237:37-72.
Umeda, T., and Inouye, K. (2002). Possible Role of Contact Following in the Generation of Coherent Motion of Dictyostelium Cells. J. Theor. Biol. 219:301-308.
Urushihara, H. (2002). Functional genomics of the social amoebae Dictyostelium discoideum. Mol. Cells. 13:1-4.
van Es, S., and Devreotes, P.N. (1999). Molecular basis of localized responses during chemotaxis in amoebae and leukocytes. Cell. Mol. Life Sci. 55:1341-1351.
Varnum, B., Edwards, K.B., and Soll, D.R. (1986). The developmental regulation of single-cell motility in Dictyostelium discoideum. Dev. Biol. 113:218-227.
Vasioukhin, V., Bauer, C., Yin, M., and Fuchs, E. (2000). Directed actin polymerization is the driving force for epithelial cell-cell adhesion. Cell. 100:209-219.
Velicer, G.J., Kroos, L., and Lenski, R.E. (2000). Developmental cheating in the social bacterium Myxococcus xanthus. Nature. 404:598-601.
Velicer, G.J., Raddatz, G., Keller, H., Deiss, S., Lanz, C., Dinkelacker, I., and Schuster, S.C. (2006). Comprehensive mutation identification in an evolved bacterial cooperator and its cheating ancestor. Proc. Natl Acad. Sci. USA. 103:8107-8112.
Velicer, G.J., and Vos, M. (2009). Sociobiology of the Myxobacteria. Annu. Rev. Microbiol. 63:599-623.
Vogel, C., and Chothia, C. (2006). Protein family expansions and biological complexity. PLoS Comput. Biol. 2:e48.
von Heijne, G. (1990). The signal peptide. J. Membr. Biol. 115:195-201. Wallace, L.J., and Frazier, W.A. (1979). Photoaffinity labeling of cyclic-AMP- and AMP-binding
proteins differentiating Dictyostelium discoideum cells. Proc. Natl. Acad. Sci. USA. 76:4250-4254.
Wang, H., Bloom, O., Zhang, M., Vishnubhakat, J.M., and Ombrellino, M. (1999). HMG-1 as a late mediator of endotoxin lethality in mice. Science. 285:248.
Wang, H., Liao, H., Ochani, M., Justiniani, M., and Lin, X. (2004a). Cholinergic agonists inhibit HMGB1 release and improve survival in experimental sepsis. Nat. Med. 10:1216.
Wang, H., Yang, H., and Tracey, K.J. (2004b). Extracellular role of HMGB1 in inflammation and sepsis. J. Intern. Med. 255:320.
Wang, J., Hou, L., Awrey, D., Loomis, W.F., Firtel, R.A., and Siu, C.-H. (2000). The membrane glycoprotein gp150 is encoded by the lagC gene and mediates cell-cell adhesion by heterophilic binding during Dictyostelium development. Dev. Biol. 227:734-745.

233
Wang, X., Matteson, J., An, Y., Moyer, B., Yoo, J., Bannykh, S., Wilson, I.A., Riordan, J.R., and Balch, W.E. (2004c). COPII-dependent export of cystic fibrosis transmembrane conductance regulator from the ER uses a di-acidic exit code. J. Cell BioI. 167:165-174.
Wang, X., Matteson, J., An, Y., Moyer, B., Yoo, J., Bannykh, S., Wilson, I.A., Riordan, J.R., and Balch, W.E. (2004d). COPII-dependent export of cystic fibrosis transmembrane conductance regulator from the ER uses a di-acidic exit code. JCB. 167:165-174.
Watanabe, N., and Kobayashi, Y. (1994). Selective release of a processed form of interleukin 1α. Cytokine. 6:597-601.
Webb, J.S., Givskov, M., and Kjelleberg, S. (2003a). Bacterial biofilms: prokaryotic adventures in multicellularity. Curr. Opin. Microbiol. 6:578.
Webb, J.S., Thompson, L.S., James, S., Charlton, T., and Tolker-Nielsen, T. (2003b). Cell death in Pseudomonas aeruginosa biofilm development. J. Bacteriol. 185:4582-4592.
Weening, K.E., Wijk, I.V., Thompson, C.R., Kessin, R.H., Podgorski, G.J., and Schaap, P. (2003). Contrasting activities of the aggregative and late PDSA promoters in Dictyostelium development. Dev. Biol. 255:373-382.
Weijer, C.J. (1999). Morphogenetic cell movement in Dictyostelium. Semin. Cell Dev. Biol. 10:609-619.
Weijer, C.J. (2004). Dictyostellium morphogenesis. Curr. Opin. Gen. Dev. 14:392-398. Weijer, C.J. (2009). Collective cell migration in development. J. Cell Sci. 122:3215-3223. Weijer, C.J., Duschl, G., and David, C.N. (1984). Dependence of cell-type proportioning and sorting
on cell cycle phase in Dictyostelium discoideum. J. Cell Sci. 70:133-145. Wenk, M., Baumgartner, R., Holak, T.A., Huber, R., Jaenicke, R., and Mayr, E.M. (1999). The
domains of protein S from Myxococcus xanthus: structure, stability and interactions. J. Mol. Biol. 286:1533-1545.
Wessels, D., Murray, J., and Soll, D.R. (1992). Behavior of Dictyostelium amoebae is regulated primarily by the temporal dynamic of the natural cAMP wave. Cell Motil. Cytoskel. 23:145-156.
West, S.A., and Buckling, A. (2003). Cooperation, virulence and siderophore production in bacterial parasites. Proc. R. Sci. London Ser. B. 270:37-44.
West, S.A., Diggle, S.P., Buckling, A., Gardner, A., and Griffin, A.S. (2007a). The Social Lives of Microbes. Ann. Rev. Ecol. Evol. Systematics. 38:53 -77
West, S.A., Diggle, S.P., Buckling, A., Gardner, A., and Griffin, A.S. (2007b). The Social Lives of Microbes. Annual Review of Ecology, Evolution, and Systematics. 38:53-77.
West, S.A., Gardner, A., and Griffin, A.S. (2006a). Altruism. Curr. Biol. 16:R482-R483. West, S.A., Griffin, A.S., and Gardner, A. (2007c). Evolutionary Explanations for Cooperation.
Current Biology. 17:R661-R672. West, S.A., Griffin, A.S., and Gardner, A. (2007d). Social semantics: altruism, cooperation,
mutualism, strong reciprocity and group selection. J. Evol. Biol. 20:415. West, S.A., Griffin, A.S., Gardner, A., and Diggle, S.P. (2006b). Social evolution theory for
microorganisms. Nat Rev Micro. 4:597-607. Whitchurch, C.B., Tolker-Nielsen, T., Ragas, P.C., and Mattick, J.S. (2002). Extracellular DNA
required for bacterial biofilm formation. Science. 295:1487. Wickner, W., and Schekman, R. (2005). Protein translocation across biological membranes. Science.
310:1452-1456. Williams, H.P., and Harwood, A.J. (2003). Cell polarity and Dictyostelium development. Curr. Opin.
Microbiol. 6:621-627.

234
Williams, J.G. (2006). Transcriptional regulation of Dictyostelium pattern formation. EMBO Rep. 7:694-698.
Williams, J.G., Ceccarelli, A., McRobbie, S., Mahbubani, H., Kay, R.R., Farly, A., Berks, M., and Jermyn, K.A. (1987). Direct induction of Dictyostelium prestalk gene expression by D1F provides evidence that D1F is a morphogen. Cell. 49:185-192.
Williams, J.G., Duffy, K.T., Lane, D.P., McRobbie, S.J., Harwood, A.J., Traynor, D., Kay, R.R., and Jermyn, K.A. (1989). Origins of the prestalk-prespore pattern in Dictyostelium development. . Cell. 59:1157-1163.
Williams, P., Winzer, K., Chan, W., and Cmara, M. (2007). Look who's talking: communication and quorum sensing in the bacterial world. Philos. Trans. R. Soc. London Ser. B. 362(1483):1119.
Williams, R.L., and Urbe, S. (2007). The emerging shape of the ESCRT machinery. Nature Rev. Mol. Cell Biol. 8:355-368.
Wilson, E.O. (1975). Sociobiology: The New Synthesis Harvard University Press, Cambridge, MA. Wilson, H.V. (1907). On some phenomena of coalescence and regeneration in sponges. J. Exp. Zool.
5:245-258. Wimmer, W., Blumbach, B., Diehl-Seifert, B., Koziol, C., Batel, R., Steffen, R., Muller, I.M., and
Muller, W.E. (1999). Increased expression of integrin and receptor tyrosine kinase genes during autograft fusion in the sponge Geodia cydonium. Cell Adhes. Commun. 7:111-124.
Wireman, J.W., and Dworkin, M. (1977). Developmentally induced autolysis during fruiting body formation by Myxococcus xanthus. J. Bacteriol. 129:798-802.
Wolpert, L. (1971). Positional information and pattern formation Curr. Top. Dev. Biol. . 6 183-224. Wolpert, L. (1981). Positional information and pattern formation. Phil. Trans. Roy. Soc. B. 295:441-
450. Wolpert, L. (1989). Positional information revisited. Development. 107:3-12. Wolpert, L. (1996). One hundred years of positional information. Trends Genet. 12:359-364. Wolpert, L., and Szathmary, E. (2002). Multicellularity: Evolution and the egg. Nature. 420:745. Wong, E., Yang, C., Wang, J., Fuller, D., Loomis, W.F., and Siu, C.-H. (2002). Disruption of the gene
encoding the cell adhesion molecule DdCAD-1 leads to aberrant cell sorting and cell-type proportioning during Dictyostelium development. Development. 129:3839-3850.
Wong, E.F., Brar, S.K., Sesaki, H., Yang, C., and Siu, C.-H. (1996). Molecular cloning and characterization of DdCAD-1, a Ca2+-dependent cell-cell adhesion molecule, in Dictyostelium discoideum. J. Biol. Chem. 271:16399-16408.
Wu, W.I., Yajnik, J., Siano, M., and De Lozanne, A. (2004). Structure-function analysis of the BEACH protein LvsA. Traffic. 5:346-355.
Xiao, Z., and Devreotes, P.N. (1997). Identification of Detergent-resistant Plasma Membrane Microdomains in Dictyostelium: Enrichment of Signal Transduction Proteins. Cell. 8:855-869.
Xie, Y., Coukell, M.B., and Gombos, Z. (1996). Antisense RNA inhibition of the putative vacuolar H+-ATPase proteolipid of Dictyostelium reduces intracellular Ca2+ transport and cell viability. J. Cell Sci. 109:489-497.
Yang, C., Brar, S.K., Desbarats, L., and Siu, C.-H. (1997). Synthesis of the Ca(2+)-dependent cell adhesion molecule DdCAD-1 is regulated by multiple factors during Dictyostelium development. Differentiation. 61:275-284.
Yeung, T., Gilvert, G.E., Shi, J., Silvius, J., Kapus, A., and Grinstein, S. (2008). Membrane Phosphatidylserine Regulates Surface Charge and Protein Localization. Science. 319:210-213.

235
Yoo, J.S., Moyer, B.D., Banngkh, S., Yoo, H.M., Riordan, J.R. and Balch, W.E. (2002) Non-conventional trafficking of the cystic fibrosis transmembrane conductance regulator through the early secretory pathway. J. Biol. Chem. 277: 11401-11409.
Yost, C.S., Hedgpeth, J., and Lingappa, V.R. (1983). A stop transfer sequence confers predictable transmembrane orientation to a previously secreted protein in cell-free system. Cell. 34:759-766.
Yumura, S., Furuya, K., and Takeuchi, I. (1996). Intracellular free calcium responses during chemotaxis of Dictyostelium cells. J. Cell Sci. 109:2673-2678.
Zackson, S.L., and Steinberg, M.S. (1986). Cranial neural crest cells exhibit directed migration on the pronephric duct pathway: Further evidence for an in vivo adhesion gradient Dev. BioI. 117:342-353.
Zehe, C., Engling, A., Wegehingel, S., Schafer, T., and Nickel, W. (2006). Cell-surface heparan sulfate proteoglycans are essential components of the unconventional export machinery of FGF-2. Proc. Natl. Acad. Sci. USA. 103:15479.
Zhao, X., and Siu, C.-H. (1995). Colocalization of the homophilic binding site and the neuritogenic activity of the cell adhesion molecule L1 to its second Ig-like domain. J. Biol. Chem. 270:29413-29421.
Zhou, X., Engel, T., Goepfert, C., Erren, M., Assmann, G., and von Eckardstein, A. (2002). The ATP binding cassette transporter A1 contributes to the secretion of interleukin 1beta from macrophages but not from monocytes. Biochem. Biophys. Res. Commun. 291:598.
Zhu, Q., and Clarke, M. (1992). Association of calmodulin and an unconventional myosin with the contractile vacuole complex of Dictyostelium discoideum. J. Cell Sci. 118:347-358.
Zhu, Q., Liu, T., and Clarke, M. (1993). Calmodulin and the contractile vacuole complex in mitotic cells of Dictyostelium discoideum. J. Cell Sci. 104:1119-1127.
Zhu, W.Q., and Ochieng, J. (2001). Rapid release of intracellular galectin-3 from breast carcinoma cells by fetuin. Cancer Res. 61:1869.
Zimmermann, W., and Weijer, C.J. (1993). Analysis of cell cycle progression during the development of Dictyostelium and its relationship to differentiation. Dev. Biol. 160:178-185.
Zusman, D.R., Scott, A.E., Yang, Z., and Kirby, J.R. (2007). Chemosensory pathways, motility and development in Myxococcus xanthus. Nat. Rev. Microbiol. 5:862-872.


















