Functional Characterization Extracellular Vesicles …iai.asm.org/content/55/1/111.full.pdf ·...
7
INFECTION AND IMMUNITY, Jan. 1987, p. 111-117 Vol. 55, No. 1 0019-9567/87/O10111-07$02.00/0 Copyright C) 1987, American Society for Microbiology Functional Characterization of Extracellular Vesicles Produced by Bacteroides gingivalis DANIEL GRENIERt AND DENIS MAYRAND* Groupe de Recherche en Ecologie Buccale and Departement de Biochimie, Faculte des Sciences et de Ge'nie, Universite Laval, Sainte-Foy, Quebec, Canada GIK 7P4 Received 3 June 1986/Accepted 6 October 1986 Extracellular vesicles of Bacteroides gingivalis (type strain 33277) were isolated, and some of their biological activities were characterized. The vesicles were obtained from a 2-day culture after ammonium sulfate precipitation, differential centrifugation, and dialysis. When viewed by electron microscopy, vesicles of approximately 50 nm predominated. The results indicated that the enriched vesicle fraction had a high proteolytic activity against collagen, Azocoll, and N-a-benzoyl-DL-arginine p-nitroanilide. The polypeptide pattern of the vesicles was similar but not identical to that of the outer membrane. The membrane vesicles could also promote bacterial adherence between homologous cells as well as mediate attachment between two noncoaggregating bacterial species. These vesicles could thus play an important role in periodontal diseases by serving as a vehicle for toxins and various proteolytic enzymes, as well as being involved in adherence. Recent electron-microscopic studies have shown that some gram-negative oral bacterial strains can release micro- vesicles into the culture supernatant (18, 19, 25, 36). These vesicles seem to consist of unmodified outer membrane and to have various bacterial activities associated with them. It was shown that vesicles of Actinobacillus actinomy- cetemcomitans exhibit a leukotoxic activity (13), as well as exhibiting endotoxic activity and containing a bone resorb- tion-inducing substance (25). Different types of phospha- tases were also found in membrane fragments of several Capnocytophaga species (28). More recently, Austin- Prather and Booth (1) showed that a bacteriocin is associated with vesicles of Bacteroides uniformis. B. gingivalis, a gram-negative anaerobe, has been found to predominate in some types of periodontal disease (reviewed in reference 31). A great variety of virulence factors have been attributed to this suspected periodontopathogen (32). The production of extracellular vesicles by B. gingivalis was recently reported by several groups (24, 27, 35). McKee et al. (24) showed that cells of B. gingivalis W50 grown under hemin limitation appeared as small rods and that a large number of extracellular vesicles (blebs) could be seen around the cells and free in the medium. Furthermore, studies in our laboratory on mixed anaerobic infections involving B. gingivalis revealed the presence of a large quantity of these structures in the exudate. The production of blebs by another oral asaccharolytic Bacteroides strain (presumably B. endodontalis) was reported by Haapasalo et al. (11). In this paper, we show that vesicles produced by B. gingivalis exhibit the same proteolytic activities as those shown by whole cells. We also found that vesicles could promote bacterial adherence between homologous cells and mediate adherence between two bacterial species which are otherwise noncoaggregating. * Corresponding author. t Present address: Department of Microbiology, University of British Columbia, Vancouver, British Columbia, Canada, V6T 1Z7. MATERIALS AND METHODS Bacterial strains. B. gingivalis ATCC 33277 was obtained from the American Type Culture Collection, Rockville, Md.; Capnocytophaga ochracea 1956c was kindly provided by M. Sebald, Institut Pasteur, Paris, France; and Eubacterium saburreum 162.4 was isolated and characterized in our laboratory. The growth of bacterial strains was carried out in a Trypticase (BBL Microbiology Systems, Cockeysville, Md.)-yeast extract medium containing glucose (2.5 g/liter), hemin (10 jig/ml), and vitamin K (1 p.g/ml) (30). All cultures were incubated in an anaerobic chamber (N2-CO2-H2, 80:10:10) at 37°C. Vesicles from B. gingivalis. Cells from a 1-liter culture (2-day culture) were first removed from the growth medium by centrifugation (10,000 x g for 15 min). Then, over a period of 2 h, 240 g of ammonium sulfate was added to the culture supernatant (40% saturation). After a second centrif- ugation (20,000 x g for 40 min), the pellet was suspended in 30 ml of Tris buffer (50 mM, pH 9.5) containing 0.5 mM dithiothreitol. This suspension was dialyzed for 16 h against 6 liters of the same buffer. The vesicles were then collected by centrifugation (27,000 x g for 40 min) and resuspended in 10 ml of Tris buffer (50 mM, pH 7.2) containing 0.5 mM dithiothreitol. A final centrifugation was done (27,000 x g for 40 min), and vesicles were resuspended in 1.5 ml of Tris buffer (pH 7.2) and kept at -20°C. Outer membranes from B. gingivalis. Outer membranes were prepared by the method of Boyd and McBride (3). Briefly, bacterial cells (2-day culture) were washed three times with 0.15 M phosphate buffer (pH 7.4) containing 0.15 M NaCl and 0.01 M EDTA. Outer membranes were removed by shearing whole cells through a 26-gauge hypodermic needle followed by mixing in a Waring blender for 10 s. The mixture was then centrifuged at 10,000 x g for 20 min to remove intact cells and debris. Outer membranes were isolated by centrifugation at 80,000 x g for 2 h. The pellet was finally lyophilized. Electron microscopy. Preparation of specimens for elec- tron microscopy of thin sections was carried out essentially 111 on May 7, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Functional Characterization Extracellular Vesicles …iai.asm.org/content/55/1/111.full.pdf ·...

INFECTION AND IMMUNITY, Jan. 1987, p. 111-117 Vol. 55, No. 10019-9567/87/O10111-07$02.00/0Copyright C) 1987, American Society for Microbiology
Functional Characterization of Extracellular Vesicles Produced byBacteroides gingivalis
DANIEL GRENIERt AND DENIS MAYRAND*Groupe de Recherche en Ecologie Buccale and Departement de Biochimie, Faculte des Sciences et de Ge'nie,
Universite Laval, Sainte-Foy, Quebec, Canada GIK 7P4
Received 3 June 1986/Accepted 6 October 1986
Extracellular vesicles of Bacteroides gingivalis (type strain 33277) were isolated, and some of their biologicalactivities were characterized. The vesicles were obtained from a 2-day culture after ammonium sulfateprecipitation, differential centrifugation, and dialysis. When viewed by electron microscopy, vesicles ofapproximately 50 nm predominated. The results indicated that the enriched vesicle fraction had a highproteolytic activity against collagen, Azocoll, and N-a-benzoyl-DL-arginine p-nitroanilide. The polypeptidepattern of the vesicles was similar but not identical to that of the outer membrane. The membrane vesicles couldalso promote bacterial adherence between homologous cells as well as mediate attachment between twononcoaggregating bacterial species. These vesicles could thus play an important role in periodontal diseases byserving as a vehicle for toxins and various proteolytic enzymes, as well as being involved in adherence.
Recent electron-microscopic studies have shown thatsome gram-negative oral bacterial strains can release micro-vesicles into the culture supernatant (18, 19, 25, 36). Thesevesicles seem to consist of unmodified outer membrane andto have various bacterial activities associated with them. Itwas shown that vesicles of Actinobacillus actinomy-cetemcomitans exhibit a leukotoxic activity (13), as well asexhibiting endotoxic activity and containing a bone resorb-tion-inducing substance (25). Different types of phospha-tases were also found in membrane fragments of severalCapnocytophaga species (28). More recently, Austin-Prather and Booth (1) showed that a bacteriocin is associatedwith vesicles of Bacteroides uniformis.
B. gingivalis, a gram-negative anaerobe, has been found topredominate in some types of periodontal disease (reviewedin reference 31). A great variety of virulence factors havebeen attributed to this suspected periodontopathogen (32).The production of extracellular vesicles by B. gingivalis wasrecently reported by several groups (24, 27, 35). McKee etal. (24) showed that cells of B. gingivalis W50 grown underhemin limitation appeared as small rods and that a largenumber of extracellular vesicles (blebs) could be seenaround the cells and free in the medium. Furthermore,studies in our laboratory on mixed anaerobic infectionsinvolving B. gingivalis revealed the presence of a largequantity of these structures in the exudate. The productionof blebs by another oral asaccharolytic Bacteroides strain(presumably B. endodontalis) was reported by Haapasalo etal. (11).
In this paper, we show that vesicles produced by B.gingivalis exhibit the same proteolytic activities as thoseshown by whole cells. We also found that vesicles couldpromote bacterial adherence between homologous cells andmediate adherence between two bacterial species which areotherwise noncoaggregating.
* Corresponding author.t Present address: Department of Microbiology, University of
British Columbia, Vancouver, British Columbia, Canada, V6T 1Z7.
MATERIALS AND METHODS
Bacterial strains. B. gingivalis ATCC 33277 was obtainedfrom the American Type Culture Collection, Rockville, Md.;Capnocytophaga ochracea 1956c was kindly provided by M.Sebald, Institut Pasteur, Paris, France; and Eubacteriumsaburreum 162.4 was isolated and characterized in ourlaboratory. The growth of bacterial strains was carried out ina Trypticase (BBL Microbiology Systems, Cockeysville,Md.)-yeast extract medium containing glucose (2.5 g/liter),hemin (10 jig/ml), and vitamin K (1 p.g/ml) (30). All cultureswere incubated in an anaerobic chamber (N2-CO2-H2,80:10:10) at 37°C.
Vesicles from B. gingivalis. Cells from a 1-liter culture(2-day culture) were first removed from the growth mediumby centrifugation (10,000 x g for 15 min). Then, over aperiod of 2 h, 240 g of ammonium sulfate was added to theculture supernatant (40% saturation). After a second centrif-ugation (20,000 x g for 40 min), the pellet was suspended in30 ml of Tris buffer (50 mM, pH 9.5) containing 0.5 mMdithiothreitol. This suspension was dialyzed for 16 h against6 liters of the same buffer. The vesicles were then collectedby centrifugation (27,000 x g for 40 min) and resuspended in10 ml of Tris buffer (50 mM, pH 7.2) containing 0.5 mMdithiothreitol. A final centrifugation was done (27,000 x g for40 min), and vesicles were resuspended in 1.5 ml of Trisbuffer (pH 7.2) and kept at -20°C.Outer membranes from B. gingivalis. Outer membranes
were prepared by the method of Boyd and McBride (3).Briefly, bacterial cells (2-day culture) were washed threetimes with 0.15 M phosphate buffer (pH 7.4) containing 0.15M NaCl and 0.01 M EDTA. Outer membranes were removedby shearing whole cells through a 26-gauge hypodermicneedle followed by mixing in a Waring blender for 10 s. Themixture was then centrifuged at 10,000 x g for 20 min toremove intact cells and debris. Outer membranes wereisolated by centrifugation at 80,000 x g for 2 h. The pelletwas finally lyophilized.
Electron microscopy. Preparation of specimens for elec-tron microscopy of thin sections was carried out essentially
111
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

112 GRENIER AND MAYRAND.~ -<,,#,..V,, ~;, ^
FIG. 1. Electron micrograph of ultrathin section of B. gingivalisATCC 33277. Bar, 100 nm.
by the method of Parent et al. (27). Cells or vesicles wereprefixed in a 0.15 M sodium cacodylate buffer (pH 7.4)containing 0.1% (vol/vol) glutaraldehyde and 2% (wt/vol)paraformaldehyde for 2 h at 37°C. Specimens were postfixedin buffered 1% (wt/vol) osmium tetroxide for 60 min at roomtemperature, dehydrated by passage through an ethanolseries and propylene oxide, and embedded in Epon 812(GBEM Services, Pointe-Claire, Quebec, Canada). Materialwas sectioned and double stained with uranyl acetate andlead citrate. Observation was done with a Philips 200 micro-scope (60 Kv).
Hemagglutination. The capacity to agglutinate sheeperythrocytes was evaluated by a method already described(23). Bacterial cells and vesicles were suspended in phos-phate-buffered saline PBS (pH 7.2), and the optical densityat 660 nm was adjusted to 0.5.
Proteolytic activity. Collagenase activity was measured bythe method of Gisslow and McBride (10). To limit the actionof nonspecific proteases on the radioactive substrate, theincubation period of the assay was kept to 2 h (21). Thecapacity of B. gingivalis vesicles to degrade Azocoll(Calbiochem-Behring, La Jolla, Calif.) was determined by a
previously described method (22). The presence of a trypsin-like activity was determined by using N-a-benzoyl-DL-arginine p-nitroanilide (BAPNA) as substrate (Sigma Chem-ical Co., St. Louis, Mo.). The incubation period of theassays with the two chromogenic substrates was also 2 h.The assays were done three times. Uninoculated mediumand buffer were used as controls.
Electrophoresis. Electrophoresis of outer membranes andvesicle polypeptides was carried out by using the buffersystem of Laemmli (17). Slab gels contained 12 or 7.5%(wt/vol) acrylamide in the resolving gel and 4.5% (wt/vol) inthe stacking gel. The lyophilized material or the vesiclepreparation was solubilized in 0.0625 M Tris hydrochloride(pH 6.8) containing 3% sodium dodecyl sulfate (SDS), 20%glycerol, 0.005 M EDTA, and 1% 2-mercaptoethanol. Thesamples were heated at 100°C for 10 min and applied (50 ,ug)to the gel. The electrophoresis was carried out at room
temperature and at a constant current of 35 mA until thebromophenol blue dye front reached the bottom of the gel.The polypeptides were stained with Coomassie blue.
Protease profile. The protease profile was determined by
using untreated samples and samples that had been treatedwith SDS. The proteins contained in the samples wereseparated on a 12% polyacrylamide gel (with SDS) or a 7.5%polyacrylamide gel (without SDS). The proteolytic activityof the bands was detected by laying the gel over an agarosegel containing skim milk (1%) by the method of Foltmann etal. (5). These procedures were done three times and gave thesame results.
Aggregation assay. After growth, the bacterial cells of E.saburreum 162.4 and C. ochracea 1956c were washed twicein PBS (pH 7.2) and finally resuspended to give an opticaldensity of 1.0 at 550 nm. Each bacterial suspension wasshaken vigorously before the assay. The mixtures consistedof 0.5 ml of each bacterial suspension, 0.5 ml of PBS, and 50,ul of the vesicle suspension; dilutions (1 in 2) were immedi-ately made. The diluted mixtures were incubated at 37°C for60 min in a slowly shaking bath. The highest aggregation titerwas noted, and the cells were observed with a phase-contrast microscope. The effect of various treatments ofboth cells and vesicles was verified to characterize thecomponents implicated in the aggregation. Cell suspensions(A550 of 2.0) in PBS and the vesicle preparation werepretreated for 30 min at 37°C in the presence of 1 mg ofprotease (no. 5380; Sigma) per ml, 1 mg of chymotrypsin (no.4124; Sigma) per ml, 0.1 N HCl, and 5 mM SDS. Aftertreatment but before use in the coaggregation assay, thebacterial cells and the vesicles were washed twice in PBS.The heat lability of the components was determined byexposure to temperatures of 50 and 80°C for 30 min. Finally,a series of potential inhibitors of coaggregation was added inthe coaggregation assay: 10 mM EDTA, various carbohy-drates and uronic acids at 100 mM (cellobiose, fructose,fucose, galactose, glucose, lactose, mannose, raffinose,rhamnose, mannitol, sorbitol, galacturonic acid, and gluc-uronic acid), and some amino acids at various concentra-tions (arginine, lysine, alanine, glycine, proline, valine,glutamine, and histidine, all in the L -form). The degree ofinhibition was expressed as the concentration at which thetested compound could inhibit the coaggregation assay.
RESULTS
Isolation and SDS-polyacrylamide gel electrophoresis(PAGE). When B. gingivalis was grown to stationary phasein a complex medium, extracellular vesicles were found tobe released into the medium. Preliminary observations indi-cated that B. gingivalis ATCC 33277 produced a much largerquantity of extracellular vesicles than did strain W83. Figure1 shows the presence of numerous vesicles originating fromB. gingivalis cells under normal growth conditions. Thesevesicles can be bound to the surface of the cells or be free inthe medium. The outer membrane resolution of both thecells and the vesicles allowed us to believe that the latteroriginate from an extrusion of the outer cell membrane. Asfor the whole cells, these membraneous vesicles appeared tobe covered by a moderately electron-dense material. Themethod described above permitted us to obtain large quan-tities of the membrane fragments. The use of Tris buffer atpH 9.5 helped to solubilize the pili and thus facilitated thespecific isolation of vesicles by centrifugation (4). It was
possible to obtain 35 mg (wet weight) of vesicles per liter ofculture. The vesicles had a strong tendency to form aggre-gates. We found that a light ultrasonic treatment was enoughto temporarily homogenize the suspension. Electron micros-copy revealed that the fraction was enriched in vesicles (Fig.2). The diameter of the vesicles varied between 20 and 150
INFECT. IMMUN.
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

B. GINGIVALIS EXTRACELLULAR VESICLES 113
ADB. ME .a
66-45-
FIG. 2. Electron micrograph of a vesicle preparation of B.gingivalis ATCC 33277. Bar, 100 nm.
nm; however, a predominance of vesicles of diameter 50 nmwas observed. The vesicles are not likely to be artifactsproduced during the isolation protocol, because it was notpossible to obtain these structures from a noninoculatedmedium. The polypeptide pattern of the vesicles was similarbut not identical to that of the outer membrane. Two majorsupplementary bands were present in the polypeptide pat-tern of the vesicles in comparison with the one of the outermembranes, while bands common to both varied slightly inintensity (Fig. 3).
Proteolytic activity. A strong proteolytic activity was as-sociated with vesicles. Azocoll was rapidly degraded. Simi-larly, collagenase and trypsinlike activity were also strong asmeasured by the degradation of the two substrates.
Protease profile. The protease profile of the membranevesicles was similar to the one found with outer membranes.For a denaturing system, only one band (molecular weight,
A B120-66-45-
29-
FIG. 3. SDS-PAGE polypeptide profiles of (A) outer membranesfrom B. gingivalis ATCC 33277 and (B) vesicles of B. gingivalisATCC 33277.
FIG. 4. Protease profile in a denaturing system. Proteins fromouter membranes and vesicles were separated on a 7.5% polyacryl-amide gel. When the tracking dye reached the bottom part of the gel,the current was stopped and the gel was laid over an agarose gelwhich contained skim milk (1%). The two gels were incubated at37°C for up to 6 h. (A) Outer membranes from B. gingivalis ATCC33277 and (B) vesicles of B. gingivalis ATCC 33277.
ca. 50,000) was found to be able to degrade casein (Fig. 4).However, in the absence of SDS, more proteolytic bandswere detected: three proteases (possibly four) of high mo-lecular weight and one of intermediate molecular weight(Fig. 5). The activity of the protease with the second-highestmolecular weight was the strongest. The incubation timeshould be no longer than 2.5 h, because at longer times thevisualization of the different proteases becomes difficult. Onthe other hand, the detection of the lowest band needed anincubation prolonged to 6 h. Concentrated culture superna-tant (2-day culture) showed the presence of two bands.
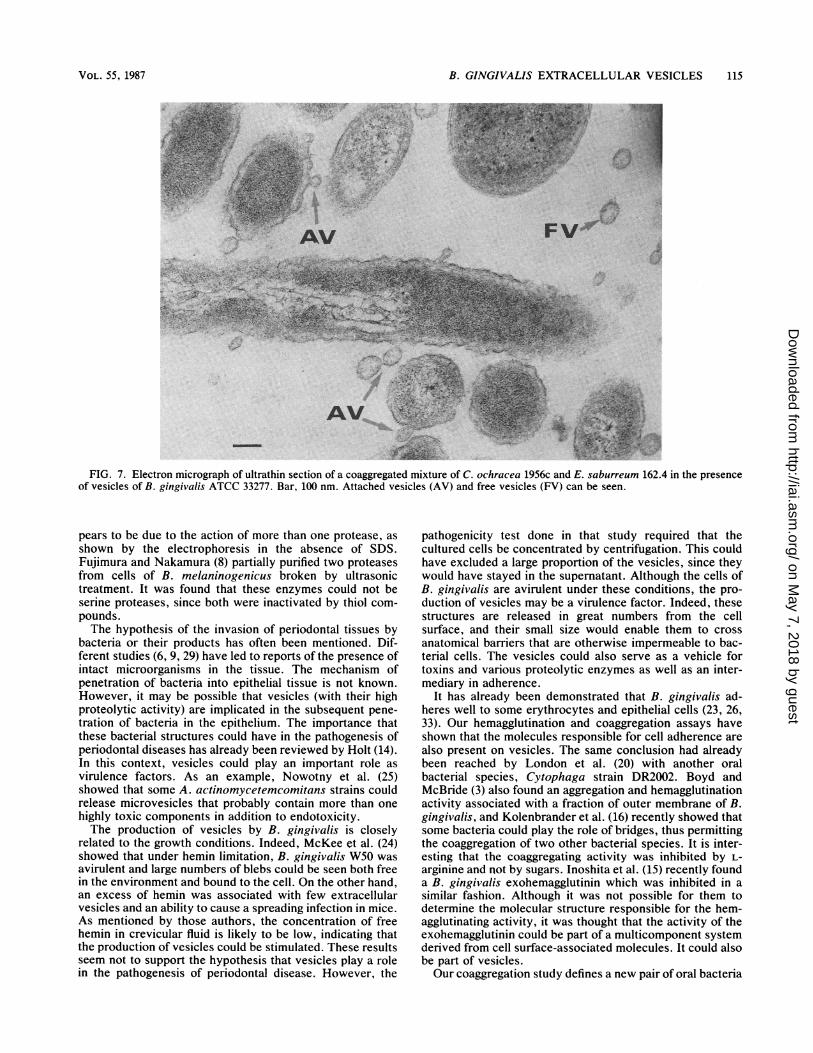
Attachment. The vesicles were shown to strongly aggluti-nate sheep erythrocytes (titer of 1 in 256). They also medi-ated coaggregation between bacterial cells of two species, E.saburreum, and C. ochracea (Fig. 6). Thus, the addition of asmall quantity of vesicles in the adherence assay permitted astrong attachment between these two noncoaggregating celltypes. An aggregation titer of 1:8 was noted; large coaggre-gates settled rapidly, leaving a clear supernatant fluid. Whenthe coaggregates were examined in the electron microscope,it could be seen that the vesicles served as a physical link orbridge between two bacterial cells (Fig. 7). The vesicles alsohad the capacity to aggregate pure cultures of these species.The two bacterial species could be stored at -20°C inPBS-glycerol (1:1) without any change in their ability tocoaggregate. This allowed a convenient cell supply. Varioustreatments of the vesicles completely inhibited coaggrega-tion. The component present on the vesicles was sensitive toheating (80°C), proteolytic treatment, low pH, and SDS,whereas such treatments of the coaggregating bacterial part-ners did not affect attachment. The effect of pH on coaggre-
VOL. 55, 1987
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

114 GRENIER AND MAYRAND
A B C
132-
66-
FIG. 5. Protease profile in a nondenaturing system. Preparationswere separated on a 12% polyacrylamide gel. The gel was then laidover an agarose gel which contained skim milk (1%). (A) Concen-trated culture supernatant (50 times by ultrafiltration), (B) outermembranes from B. gingivalis ATCC 33277, and (C) vesicles of B.gingivalis ATCC 33277. To obtain a better resolution, the uppersection of the gel was incubated (37°C) for 2 h, whereas the lowersection was incubated for 6 h. The arrows indicate the presence ofa proteolytic activity.
gation activity was examined in sodium acetate buffer (pH 3to 5), sodium phosphate buffer (pH 6 to 7), and Trishydrochloride buffer (pH 7 to 10). The results indicated thatthe attachment mediated by vesicles is stable in a pH rangeof 4.5 to 8.5. Coaggregation activity was unaffected by thepresence of sugars or uronic acids at a concentration of 100mM. EDTA (at 10 mM) also had no effect. Among the aminoacids, only L-arginine and, to a lesser degree, L-lysine had aninhibitory effect on the coaggregation. In this case, 2.5 mML-arginine completely inhibited the coaggregation and 50mM L-lysine was needed to significantly reduce but notcompletely inhibit the coaggregation activity of the vesicles.
DISCUSSION
Some bacteria from the oral cavity are capable of produc-ing extracellular vesicles. The release of these vesicles fromwhole cells depends on the bacterial strain as well asnutritional conditions (37). These vesicles could be morpho-logically similar to those (30 to 200 nm) previously reportedto be present in dental plaque samples (7, 12). This phenom-enon seems to be the result of a blebbing or extrusion fromthe outer cell membrane. Production of vesicles by B.gingivalis was observed by Listgarten and Lai (19) andParent et al. (27). Williams and Holt (35) recently isolatedblebs from various oral Bacteroides species including B.gingivalis; the isolation method that they used allowed themto obtain vesicles or membrane fragments of various sizes,but vesicles of approximately 50 nm in diameter predomi-nated. The SDS-PAGE profiles revealed that the polypeptidepatterns of these vesicles were similar but not identical tothose of outer membrane preparations isolated from thecorresponding cells. Generally speaking, fewer polypeptideswere seen on profiles from outer membranes. Two additionalbands were found in the PAGE profile of the vesicles, and atlea,st two peptides were found in lower amounts as deter-mined by the intensity of protein staining. The presence of a
few additional bands in the profile of the vesicles which arenot present in outer membranes can be explained the follow-ing ways: either these proteins were part of the cells andwere concentrated in the process of bleb formation or thesebands are the result of action of protease(s) on surfacemolecules of the vesicles. Boyd and McBride (3) found thatparticulate material containing hemagglutinating and bacte-rial aggregating activity could be isolated from culture su-pernatants of B. gingivalis by centrifugation. SDS-PAGE ofthis material revealed the presence of several bands not seenin isolated membrane preparations. These bands werethought to be contaminants due to cell lysis. A possibleconsequence of this characteristic is that vesicles Couldcompete for antibodies and thus impede the specific antibac-terial immune defense.Our results indicate that whole cells as well as vesicles or
blebs degraded the proteinaceous substrates tested. It is nowevident that the proteolytic system of B. gingivalis is presenton the surface of cells as well as vesicles. A similar distri-bution of the cellulolytic system of Clostridium thermocel-lum was recently observed (2). Similarly Thompson et al.(34) showed that blebs associated with the growth of Pseu-domonas fragi contain high concentrations of neutralproteases. Vesicles could thus constitute a good startingmaterial for the subsequent purification and characterizationof B. gingivalis proteases or molecules that take part inaggregation mechanisms. The protease profile of the vesicleswas found to be identical to the one found for whole cells.Furthermore, the degradation of the various substrates ap-
I
V4,
A
i^
IS
.: ..
~4
.i
C
'A. ..
.k.
4%N.4,N
B./
{-C DFIG. 6. Phase-contrast microscopy of (A) C. ochracea 1956c, (B)
E. saburreum 162.4, (C) mixture of C. ochracea 1956c and E.saburreum 162.4, and (D) mixture of C. ochracea 1956c and E.saburreum 162.4 in the presence of vesicles of B. gingivalis ATCC33277.
INFECT. IMMUN.
-0
.,le.I 'a 'e
a I
a.o' 'i
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
B. GINGIVALIS EXTRACELLULAR VESICLES 115
AV FV~
._
'A^Ijkl ._'.iE,za
;-:
AV*V:4>p."
,.1.
r,,,,I,,.. .,, ,..afFIG. 7. Electron micrograph of ultrathin section of a coaggregated mixture of C. ochracea 1956c and E. saburreum 162.4 in the presence
of vesicles of B. gingivalis ATCC 33277. Bar, 100 nm. Attached vesicles (AV) and free vesicles (FV) can be seen.
pears to be due to the action of more than one protease, asshown by the electrophoresis in the absence of SDS.Fujimura and Nakamura (8) partially purified two proteasesfrom cells of B. melaninogenicus broken by ultrasonictreatment. It was found that these enzymes could not beserine proteases, since both were inactivated by thiol com-pounds.The hypothesis of the invasion of periodontal tissues by
bacteria or their products has often been mentioned. Dif-ferent studies (6, 9, 29) have led to reports of the presence ofintact microorganisms in the tissue. The mechanism ofpenetration of bacteria into epithelial tissue is not known.However, it may be possible that vesicles (with their highproteolytic activity) are implicated in the subsequent pene-tration of bacteria in the epithelium. The importance thatthese bacterial structures could have in the pathogenesis ofperiodontal diseases has already been reviewed by Holt (14).In this context, vesicles could play an important role asvirulence factors. As an example, Nowotny et al. (25)showed that some A. actinomycetemcomitans strains couldrelease microvesicles that probably contain more than onehighly toxic components in addition to endotoxicity.The production of vesicles by B. gingivalis is closely
related to the growth conditions. Indeed, McKee et al. (24)showed that under hemin limitation, B. gingivalis W50 wasavirulent and large numbers of blebs could be seen both freein the environment and bound to the cell. On the other hand,an excess of hemin was associated with few extracellularvesicles and an ability to cause a spreading infection in mice.As mentioned by those authors, the concentration of freehemin in crevicular fluid is likely to be low, indicating thatthe production of vesicles could be stimulated. These resultsseem not to support the hypothesis that vesicles play a rolein the pathogenesis of periodontal disease. However, the
pathogenicity test done in that study required that thecultured cells be concentrated by centrifugation. This couldhave excluded a large proportion of the vesicles, since theywould have stayed in the supernatant. Although the cells ofB. gingivalis are avirulent under these conditions, the pro-duction of vesicles may be a virulence factor. Indeed, thesestructures are released in great numbers from the cellsurface, and their small size would enable them to crossanatomical barriers that are otherwise impermeable to bac-terial cells. The vesicles could also serve as a vehicle fortoxins and various proteolytic enzymes as well as an inter-mediary in adherence.
It has already been demonstrated that B. gingivalis ad-heres well to some erythrocytes and epithelial cells (23, 26,33). Our hemagglutination and coaggregation assays haveshown that the molecules responsible for cell adherence arealso present on vesicles. The same conclusion had alreadybeen reached by London et al. (20) with another oralbacterial species, Cytophaga strain DR2002. Boyd andMcBride (3) also found an aggregation and hemagglutinationactivity associated with a fraction of outer membrane of B.gingivalis, and Kolenbrander et al. (16) recently showed thatsome bacteria could play the role of bridges, thus permittingthe coaggregation of two other bacterial species. It is inter-esting that the coaggregating activity was inhibited by L-arginine and not by sugars. Inoshita et al. (15) recently founda B. gingivalis exohemagglutinin which was inhibited in asimilar fashion. Although it was not possible for them todetermine the molecular structure responsible for the hem-agglutinating activity, it was thought that the activity of theexohemagglutinin could be part of a multicomponent systemderived from cell surface-associated molecules. It could alsobe part of vesicles.Our coaggregation study defines a new pair of oral bacteria
VOL. 55, 1987
A. N,
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

116 GRENIER AND MAYRAND
and, more importantly, the potential role of vesicles to act as
coaggregation bridges which could be implicated in bacterialcolonization of the oral cavity. The coaggregation seems toresult from a specific interaction between a protein orglycoprotein molecule on vesicles and a nonproteinaceouscomponent on bacterial cells.
ACKNOWLEDGMENTS
We thank Louise Lamonde for her help with the electron micro-scope and Gene Bourgeau for his help in the preparation of themanuscript.D.G. was supported by a studentship from the Fonds de la
Recherche en Sante du Qudbec. The work was supported by theMedical Research Council of Canada.
LITERATURE CITED
1. Austin-Prather, S. L., and S. J. Booth. 1984. Evidence for amembrane-bound form of a bacteriocin of Bacteroides uniformisT1-1. Can. J. Microbiol. 30:268-272.
2. Bayer, E. A., E. Setter, and R. Lamed. 1985. Organization anddistribution of the cellulosome in Clostridium thermocellum. J.Bacteriol. 163:552-559.
3. Boyd, J., and B. C. McBride. 1984. Fractionation of hemagglu-tinating and bacterial binding adhesins of Bacteroidesgingivalis. Infect. Immun. 45:403-409.
4. Brinton, C. C., J. Bryan, J.-A. Dillon, N. Guerina, L. J.Jacobson, A. Labik, S. Lee, A. Levine, S. Lim, J. McMichael, S.Polen, K. Rogers, A. C.-C. To, and S. C.-M. To. 1978. Uses ofpili in gonorrhea control: role of bacterial pili in disease,purification and properties of gonococcal pili, and progress inthe development of a gonococcal pilus vaccine for gonorrhea, p.
155-178. In G. F. Brooks, E. C. Gotschlich, K. K. Holmes,W. D. Sawyer, and F. E. Young (ed.), Immunobiology ofNeisseria gonorrhoeae. American Society for Microbiology,Washington, D.C.
5. Foltmann, B., P. B. Szecsi, and N. I. Tarasova. 1985. Detectionof proteases by clotting of casein after gel electrophoresis. Anal.Biochem. 146:353-360.
6. Frank, R. M. 1980. Bacterial penetration in the apical pocketwall of advanced periodontitis. J. Periodontal Res. 15:563-573.
7. Frank, R. M., and G. Hoover. 1970. An ultrastructural study ofhuman supragingival dental plaque formation, p. 85-108. In:W. D. McHugh. (ed.), Dental plaque. S. Livingstone Ltd.,London.
8. Fujimura, S., and T. Nakamura. 1981. Isolation and character-ization of proteases from Bacteroides melaninogenicus. Infect.Immun. 33:738-742.
9. Gillett, R., and N. W. Johnson. 1982. Bacterial invasion of theperiodontium in a case of juvenile periodontitis. J. Clin.Periodontol. 9:93-100.
10. Gisslow, M. T., and B. C. McBride. 1975. A rapid sensitivecollagenase assay. Anal. Biochem. 68:70-78.
11. Haapasalo, M., H. Ranta, H. Shah, K. Ranta, and K. Lounat-maa. 1984. Isolation and characterization of a new variant ofblack-pigmented asaccharolytic Bacteroides. FEMS Microbiol.Lett. 23:269-274.
12. Halhoul, N., and J. R. Colvin. 1975. The ultrastructure ofbacterial plaque attached to the gingiva of man. Arch. Oral Biol.20:115-118.
13. Hammond, B. C., and R. H. Stevens. 1982. Capnocytophaga andActinobacillus actinomycetemcomitans: occurrence and patho-genic potential in juvenile periodontitis, p. 46-61. In: R. J.Genco and S. E. Mergenhagen (ed.), Host-parasite interactionsin periodontal diseases. American Society for Microbiology,Washington, D.C.
14. Holt, S. C. 1982. Bacterial surface structures and their role inperiodontal disease, p. 139-150. In R. J. Genco and S. E.Mergenhagen (ed.), Host-parasite interactions in periodontaldisease. American Society for Microbiology, Washington, D.C.
15. Inoshita, E., A. Amano, T. Hanioka, H. Tamagawa, S. Shizuku-
ishi, and A. Tsunemitsu. 1986. Isolation and some properties ofexohemagglutinin from the culture medium of Bacteroidesgingivalis 381. Infect. Immun. 52:421-427.
16. Kolenbrander, P. E., R. N. Andersen, and L. V. Holdeman.1985. Coaggregation of oral Bacteroides species with otherbacteria: central role in coaggregation bridges and competitions.Infect. Immun. 48:741-746.
17. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
18. Lai, G. H., M. A. Listgarten, and B. F. Hammond. 1981.Comparative ultrastructure of leukotoxic and non-leukotoxicstrains of Actinobacillus actinomycetemcomitans. J. Periodon-tal Res. 16:379-389.
19. Listgarten, M. A., and C. H. Lai. 1979. Comparative ultrastruc-ture of Bacteroides melaninogenicus subspecies. J. PeriodontalRes. 14:332-340.
20. London, J., R. Celesk, and P. Kolenbrander. 1982. Physiologicaland ecological properties of the oral gram-negative glidingbacteria capable of attaching to hydroxyapatite, p. 76-85. InR. J. Genco and S. E. Mergenhagen (ed.), Host-parasite inter-actions in periodontal diseases. American Society for Microbi-ology, Washington, D.C.
21. Mayrand, D., and D. Grenier. 1985. Detection of collagenaseactivity in oral bacteria. Can. J. Microbiol. 31:134-138.
22. Mayrand, D., and B. C. McBride. 1980. Ecological relationshipsof bacteria involved in a simple mixed anaerobic infection.Infect, Immun. 27:44-50.
23. Mayrand, D., B. C. McBride, T. Edwards, and S. Jensen. 1980.Characterization of Bacteroides asaccharolyticus and B.melaninogenicus oral isolates. Can. J. Microbiol. 26:1178-1183.
24. McKee, A. S., A. S. McDermid, A. Baskerville, A. B. Dowsett,D. C. Ellwood, and P. D. Marsh. 1986. Effect of hemin on thephysiology and virulence of Bacteroides gingivalis W50. Infect.Immun. 52:349-355.
25. Nowotny, A., U. H. Behling, B. Hammond, C.-H Lai, M.Listgarten, P. H. Pham, and F. Sanavi. 1982. Release of toxicmicrovesicles by Actinobacillus actinomycetemcomitans. In-fect. Immun. 37:151-154.
26. Okuda, K., J. Slots, and R. J. Genco. 1981. Bacteroidesgingivalis, Bacteroides asaccharolyticus, and Bacteroides mel-aninogenicus subspecies: cell surface morphology and adher-ence to erythrocytes and human buccal epithelial cells. Cur.Microbiol. 6:7-12.
27. Parent, R., C. Mouton, L. Lamonde, and D. Bouchard. 1986.Human and animal serotypes of Bacteroides gingivalis definedby crossed immunoelectrophoresis. Infect. Immun. 51:909-918.
28. Poirier, T. P., and S. C. Holt. 1983, Acid and alkaline phospha-tases of Capnocytophaga species. I. Production and cytologicallocalization of the enzymes. Can. J. Microbiol. 29:1369-1381.
29. Saglie, R., M. G. Newman, F. A. Carranza Jr., and G. L.Pattison. 1982. Bacterial invasion of gingiva in advanced peri-odontitis in humans. J. Periodontol. 53:217-222.
30. Sawyer, S. J., J. B. MacDonald, and R. J. Gibbons. 1962.Biochemical characteristics of Bacteroides melaninogenicus. Astudy of thirty-one strains. Arch. Oral Biol. 7:685-691.
31. Slots, J. 1982. Importance of black-pigmented Bacteroides inhuman periodontal disease, p. 27-45. In R. J. Genco and S. E.Mergenhagen (ed.), Host-parasite interactions in periodontaldiseases. American Society for Microbiology, Washington,D.C.
32. Slots, J., and R. J. Genco. 1984. Black-pigmented Bacteroidesspecies, Capnocytophaga species,-and Actinobacillus actinomy-cetemcomitans in human periodontal disease: virulence factorsin colonization, survival, and tissue destruction. J. Dent. Res.63:412-421.
33. Slots, J., and R. J. Gibbons. 1978. Attachment of Bacteroidesmelaninogenicus subsp. asaccharolyticus to oral surfaces andits possible role in colonization of the mouth and of periodontalpockets. Infect. Immun. 19:254-264.
34. Thompson, S. S., Y. M. Naidu, and J. J. Petska. 1985. Ultra-structural localization of an extracellular protease in Pseudomo-nasfragi by using the peroxidase-antiperoxidase reaction. Appl.
INFECT. IMMUN.
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

B. GINGIVALIS EXTRACELLULAR VESICLES 117
Environ. Microbiol. 50:1038-1042.35. Williams, G. D., and S. C. Holt. 1985. Characteristics of the
outer membrane of selected oral Bacteroides species. Can. J.Microbiol. 31:238-250.
36. Woo, D. P., S. C. Holt, and E. R. Leadbetter. 1979. Ultrastruc-ture of Bacteroides species: Bacteroides asaccharolyticuis, Bac-
teroides fragilis, Bacteroides inelaninogenicius, subspeciesmelaninogenicus and B. inelaninogenicus, subspeciesintermedius. J. Infect. Dis. 139:534-546.
37. Work, E., K. W. Knox, and M. Vesk. 1966. The chemistry andelectron microscopy of an extracellular lipopolysaccharide fromEscherichia coli. Ann. N.Y. Acad. Sci. 133:438-449.
VOL. 55, 1987
on May 7, 2018 by guest
http://iai.asm.org/
Dow
nloaded from