
Source: Proceedings of the Entomological Society of ... · Resumo.—Neste estudo...
20
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Variation of Male Secondary Sexual Structures and the Taxonomy of Theritas lisus and Relatives (Lepidoptera, Lycaenidae, Eumaeini) Author(s): Ananda Regina P. Martins, Christophe Faynel, and Robert K. Robbins Source: Proceedings of the Entomological Society of Washington, 118(4):555-573. Published By: Entomological Society of Washington DOI: http://dx.doi.org/10.4289/0013-8797.118.4.555 URL: http://www.bioone.org/doi/full/10.4289/0013-8797.118.4.555 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/ terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript of Source: Proceedings of the Entomological Society of ... · Resumo.—Neste estudo...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Variation of Male Secondary Sexual Structures and theTaxonomy of Theritas lisus and Relatives (Lepidoptera,Lycaenidae, Eumaeini)Author(s): Ananda Regina P. Martins, Christophe Faynel, and Robert K. RobbinsSource: Proceedings of the Entomological Society of Washington,118(4):555-573.Published By: Entomological Society of WashingtonDOI: http://dx.doi.org/10.4289/0013-8797.118.4.555URL: http://www.bioone.org/doi/full/10.4289/0013-8797.118.4.555
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

PROC. ENTOMOL. SOC. WASH.
118(4), 2016, pp. 555–573
VARIATION OF MALE SECONDARY SEXUAL STRUCTURES AND THETAXONOMY OF THERITAS LISUS AND RELATIVES (LEPIDOPTERA,
LYCAENIDAE, EUMAEINI)
ANANDA REGINA P. MARTINS, CHRISTOPHE FAYNEL, AND ROBERT K. ROBBINS
urn:lsid:zoobank.org:pub:496E1E98-EBFB-4554-831E-9BE8E4BAE13D
(ARPM) Museu de Zoologia, Universidade de Sao Paulo, Avenida Nazar�e 481,Ipiranga, 04263-000 Sao Paulo, SP, Brazil (e-mail: [email protected]);(ARPM) Redpath Museum, McGill University, 859 Sherbrooke Street West,Montreal, QC, H3A 2K6, Canada; (CF) 16 rue des Aspres, F-34160 Montaud,France (e-mail: [email protected]); (RKR) Department of Entomology,Smithsonian Institution, PO Box 37012, Washington, DC 20013-7012 USA(e-mail: [email protected])
ARPM: Isid:zoobank.org:author:F51CFF8B-0382-4D19-BA25-B3410A7F09E0CF: Isid:zoobank.org:author:38C2FC5F-7A0A-4A8B-A486-4E650745FD7DRKR: Isid:zoobank.org:author:44061674-2F12-49EE-BC5F-A264F51A3391
Abstract.—We assess variation of male secondary sexual structures, genitalia, andwing patterns in Theritas lisus (Stoll) and relatives to clarify the species level tax-onomy, which is needed for a phylogenetic analysis. Variation in the male ventralhindwing scent pouch was especially useful taxonomically. This pouch may beabsent, present without androconia, or present with androconia in T. lisus. Despitethis variation, pouch size (when present) distinguishes T. lisus from its closest rel-atives. As an overview, we propose distinguishing traits for a phenetic T. lisus speciesgroup, re-examine and confirm the proposed synonymies of T. hisbon and T. orsinawith T. lisus and of T. photeinoswith T. viresco, confirm the association of the sexes of T.lisus and T. viresco, and describe Theritas silma Martins, Faynel, & Robbins, newspecies, from French Guiana. CO1 mitochondrial sequences are consistent with thesetaxonomic results, but the recently introduced barcode index numbers (BINs) recognizedtoo many species. A nomenclatural list summarizes the taxonomic actions in the T. lisusspecies group.
Key Words: Atlides Section, DNA barcoding, scent pad, scent pouch, Theritasespiritosanto, Theritas silma, Theritas viresco
Resumo.—Neste estudo sao avaliadas as variacoes de estruturas sexuais secund�ariasmasculinas, genit�alias e padroes de asas em Theritas lisus (Stoll) e taxa pr�oximos, a fim deesclarecer a taxonomia do grupo ao n�ıvel de esp�ecie e, consequentemente fornecer sub-s�ıdios para futuras an�alises filogen�eticas. A variacao nas invaginacoes da membranaventral das asas posteriores de machos mostrou-se taxonomicamente �util. Essas in-vaginacoes podem estar ausentes, presentes e contendo androconias, ou ainda presentes

sem androconias em T. lisus. No entanto, apesar da variacao, o comprimento da in-vaginacao (quando presente) pode ser utilizado para distinguir T. lisus de taxa pr�oximos.Neste estudo sao propostos caracteres para o grupo fen�etico T. lisus; sao confirmadoscomo sinonimos T. hisbon, T. orsina e T. lisus, assim como T. photeinos e T. viresco; �econfirmada a associacao entre indiv�ıduos machos e femeas em T. lisus e T. viresco; e �edescrita uma nova esp�ecie daGuiana Francesa, Theritas silmaMartins, Faynel, &Robbins.Sequencias de CO1 mitocondrial mostraram-se congruentes com os resultados tax-onomicos, por�em os n�umeros de barcodes (BINs) recentemente introduzidos re-conhecem ummaior n�umero de esp�ecies. Uma lista nomenclatural sumariza as propostastaxonomicas do grupo T. lisus.
Palavras chave: Secao Atlides, barcoding DNA, �orgao odor�ıfero dorsal, �orgaoodor�ıfero ventral, Theritas espiritosanto, Theritas silma, Theritasviresco
DOI: 10.4289/0013-8797.118.4.555
The genus TheritasHubner (Lepidoptera,Lycaenidae, Eumaeini) contains 26 Neo-tropical hairstreak butterfly species (Robbins2004), many of which are spectacularlycolored with iridescent blue and green scales(Figs. 1-5). They are biologically interestingbecause species may possess a variety ofmale secondary sexual structures on thewings, as noted in the related genus ArcasSwainson (Robbins et al. 2012).
An analysis of nuclear and mitochondrialDNA sequences of exemplar Eumaeinispecies (including six species of Theritas)suggested that Theritas was paraphyletic interms of Arcas (Quental 2008). To test thisparaphyly hypothesis, we are completinga phylogenetic analysis of theAtlidesSection(sensuRobbins 2004), which is comprised ofgenera Atlides Hubner, Arcas Swainson,Pseudolycaena Wallengren, and TheritasHubner. As part of this project, we discov-ered new information that affects the currentspecies level classification of Theritas lisus(Stoll) and close relatives (Figs. 1-5).
The purpose of this paper is to presentnew information on variation of the ven-tral hindwing scent pouch and its andro-conia (Figs. 8-14), as well as other moretraditional wing pattern, genitalic, and
mitochondrial DNA sequence characters(Figs. 15-26).
The goal of this paper is to update thetaxonomy of T. lisus and close relatives sothat we have appropriate names to use in thephylogenetic analyses. Specifically, we phe-netically characterize the T. lisus speciesgroup and present new information for as-sociating the sexes of its species.We confirmthe synonymies of T. hisbon (Godman &Salvin) and T. orsina (Hewitson) with T. lisusand of T. photeinos (Druce) with T. viresco(Druce). Finally, we describe a new species,T. silma Martins, Faynel, & Robbins.
MATERIALS AND METHODS
Standard methods were used to dissectgenitalia and to make wing slides (Robbins1991). Genitalic terminology follows that inClench (1961) and Klots (1970), as modifiedfor the Eumaeini (Robbins 1991). Forewinglength was measured with a digital verniercaliper. Wing vein names follow Comstock(1918), and terminology for male secondarysexual characters follows Robbins et al.(2012). The holotype labels are given witha description of the labels in brackets. Oth-erwise, months are abbreviated by their firstthree letters in English. Genitalic dissections
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON556

and wing slides are listed in the Appen-dix. Major biogeographical regions followBrown (1982). Brazilian states are noted bytheir standard two-letter abbreviations ex-cept in the type listings.
Legs from 18 adult Theritas were pre-pared for CO1 sequencing according to theprotocol outlined in Wilson (2012), which
also details each of the steps in sequencingthe CO1 gene. For species identificationand associating sexes, we visualized CO1DNA sequences phenetically using theneighbor joining methods in the BOLDdatabase (Ratnasingham & Hebert. 2007),which produces a phenogram of distancerelationships. We included the recently
Figs. 1–5. Adult male (left) and females, Theritas lisus species group. 1. T. lisus (Panama, Panama).
2. T. lisus (Peru, SC-Brazil). 3. T. silma (French Guiana holotype, Colombia paratype). 4. T. espiritosanto(SC-Brazil, ES-Brazil). 5. T. viresco (Peru, Ecuador). Scale 1 cm.
VOLUME 118, NUMBER 4 557

introduced barcode index number (BIN)(Ratnasingham&Hebert 2013) to see howwell it corresponded to taxonomy basedon traditional characters. Sequence sam-ples are listed with their BOLD processnumber (Ratnasingham & Hebert. 2007)in the Appendix.
Museum specimens cited in thisstudy are deposited in the followingcollections—museum acronyms fromEvenhuis (2015).
BMNH The Natural History Museum[formerly British Museum (Natural His-tory)], London, United Kingdom.CF Private collection of ChristopheFaynel, Montpellier, France.MNHN Mus�eum national d’Histoire na-turelle, Paris, France.MUSM Museo de Historia Natural, Uni-versidad Nacional Mayor de San Marcos,Lima, Peru.MZSP Museu de Zoologia da Uni-versidade de Sao Paulo, Sao Paulo, Brazil.USNM National Museum of Natural His-tory, Smithsonian Institution, Washington,DC, USA.
VARIATION AND TAXONOMY
Theritas lisus species group
We characterize the T. lisus speciesgroup as those species of Theritas thatlack a scent pad on the dorsal surface of
the male forewing (see Thomas 1893 forhistology, Robbins 1991 for morphologywith SEMs) and that lack a green discalend spot on the ventral surface of theforewing in both sexes (Figs. 1-5). Weemphasize that this is a phenetic group-ing of four species (listed below) that isconvenient for dealing with the specieslevel taxonomy of the included species.It should not be interpreted as a proposedmonophyletic grouping.
Historical Identification of T. lisus
Papilio lisus Stoll was named based onimages and description of an undisclosednumber of male specimens from Surinamthat could apply to many species (Fig. 6,compare with Figs. 1-5). Hewitson (1865:75) placed lisus in Thecla Fabricius andidentified it as a species without a malescent pad. He also illustrated a female thathe called T. lisus (Fig. 7), but it is actuallya female of T. viresco (compare with thefemale in Fig. 5). At that time, the male ofT. viresco was undescribed. In accordwith this misidentification, Hewitson(1877) later described the female of T. lisusas Thecla orsina. The evidence that He-witson mistakenly associated the sexes ofT. lisus and of T. viresco is detailed below.
Godman and Salvin (1887-1901) dif-ferentiated Central American specimens
Figs. 6–7. Historical illustrations. 6. Theritas lisus in Stoll’s original description. 7. Female ofT. viresco, misidentified in Hewitson as T. lisus.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON558

of Thecla lisus as Thecla hisbon. Draudt(1919-1920) synonymized the names,a treatment that was followed sub-sequently in faunal lists (e.g., Hoffmann1940, Robbins 2004), except by D’Abrera(1995), who remarked that T. hisbon isprobably a race of T. lisus. With theexception of the status of the nameT. hisbon, identification of T. lisus has
been consistent for almost a century.These names were treated in the genusTheritas in a checklist of the Neotropi-cal fauna (Robbins 2004).
Synonymy of T. lisus
To determine whether more than onegeographical species might be included
Figs. 8–14. Cleared male hindwings. 8. T. lisus (Costa Rica) without a pouch (arrow). 9. T. lisus(Panama) with a pouch, but without androconia (arrow). 10. T. lisus (SC, Brazil) with a pouch, but
without androconia (arrow). 11. T. lisus (eastern Ecuador) with a pouch containing a few dark androconia(arrow). 12. T. silma (PA, Brazil) with a pouch containing dark androconia at the distal end of the pouch
(arrow). 13. T. espiritosanto (SC, Brazil) with a pouch containing dark androconia at the distal end of thepouch (arrow). 14. T. viresco (Peru) with a long pouch-like fold in the wings without androconia (arrow).
VOLUME 118, NUMBER 4 559

Figs. 15–19. Male genitalia in ventral (top) and lateral (middle) aspect, with penis (bottom) in lateral
aspect. 15. T. lisus (Panama). 16. T. lisus (Brazil). 17. T. silma (Peru), arrow points to “finger-like” ventral processof the fused tegumen and vinculum. 18. T. espiritosanto (Brazil). 19. T. viresco (Brazil). Scale 1 mm.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON560
in the concept of T. lisus in Robbins(2004), we assessed geographic varia-tion in the (1) scent pouch on the ventralsurface of the hindwing in cell Cu2-2A,(2) wing pattern and venation, and(3) genitalic structures. We also looked atCO1 sequences from Central and SouthAmerica.
In Central America, a hindwing scentpouch in cell Cu2-2A (discovered byGodman & Salvin 1887-1901, char-acterized morphologically in Robbins
et al. 2012) may or may not be present(Figs. 8-9), but when present, it does notcontain androconia (Fig. 9). In SouthAmerica east of the Andes, a scentpouch occurs, but androconia may ormay not be present (Figs. 10-11). Whenthey are present, there are very few an-droconia (Fig. 11). In other words, malesfrom both Central and South Americamay possess a pouch without androconia,so this structure cannot be used to dis-tinguish them.
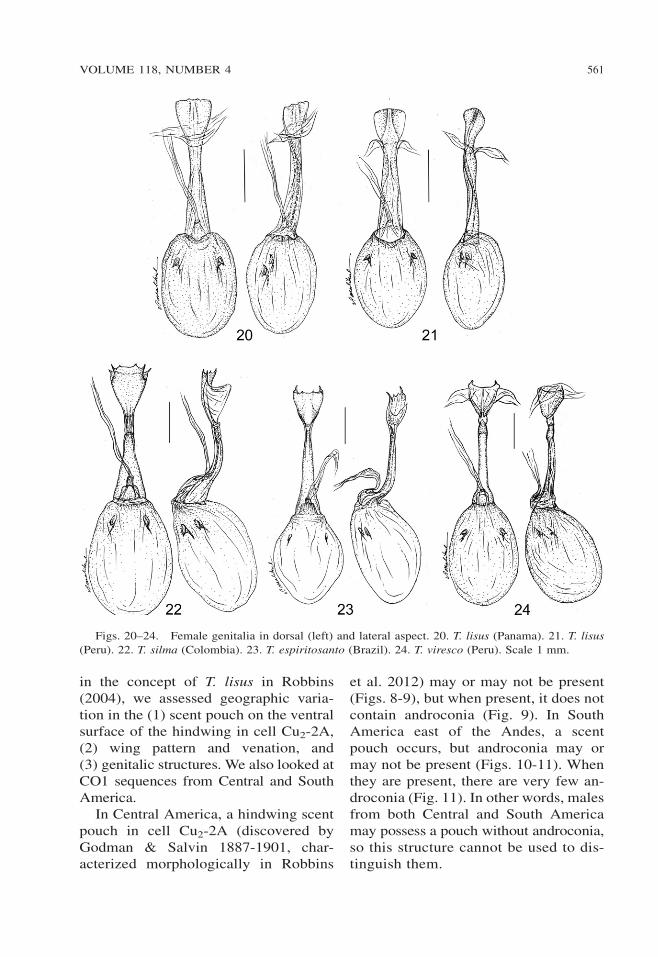
Figs. 20–24. Female genitalia in dorsal (left) and lateral aspect. 20. T. lisus (Panama). 21. T. lisus
(Peru). 22. T. silma (Colombia). 23. T. espiritosanto (Brazil). 24. T. viresco (Peru). Scale 1 mm.
VOLUME 118, NUMBER 4 561
The size and wing pattern markingsof T. lisus are highly variable at any onelocality, as is the width of the dorsalforewing apical border in males. The
amount of greenish sheen overlayingthe brown ground color on male ventralwings is highly variable, often beingmost pronounced in larger individuals,
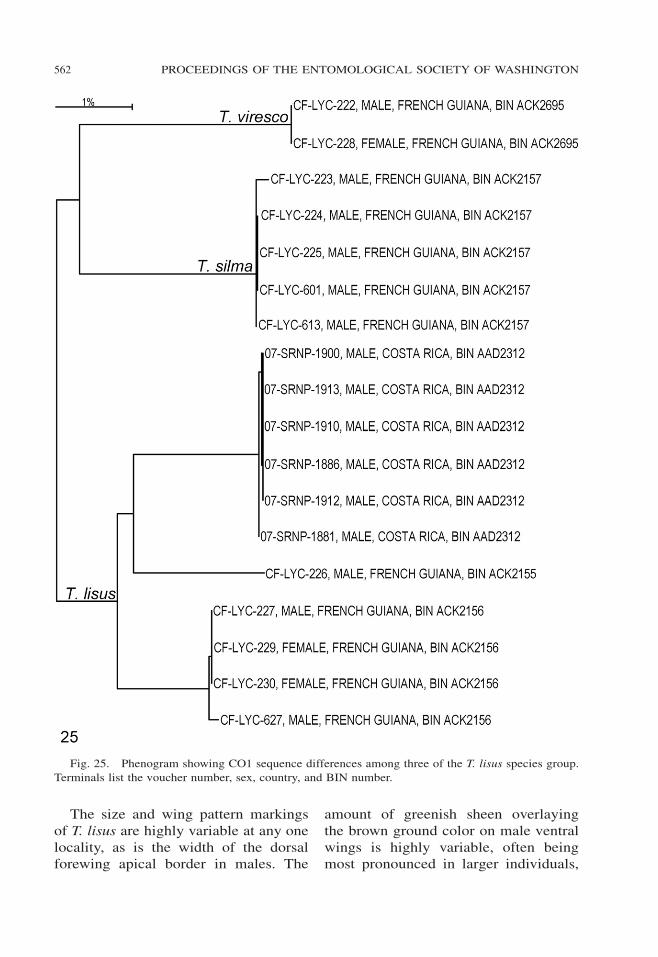
Fig. 25. Phenogram showing CO1 sequence differences among three of the T. lisus species group.
Terminals list the voucher number, sex, country, and BIN number.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON562

especially from lower montane forest onthe eastern slopes of the Andes. Theorigin of forewing vein R3 in malesvaries continuously from the discal cellto being stalked with vein M1. Some ofthis variation occurs among males rearedfrom a gregarious lot of caterpillars (seebelow).
There is minor geographic varia-tion, especially among females. CentralAmerican females have, on average, awider distal border on the dorsal surfaceof the forewing than do South Americanfemales from east of the Andes (Figs.1-2). Females from southern Brazil have,on average, a shorter tail at the end ofvein Cu1 (Figs. 1-2). However, thesegeographical patterns are not matched byvariation of male wing patterns.
Male and female genitalia of in-dividuals from throughout the range ofT. lisus are indistinguishable, based on16 male and 13 female dissections (Ap-pendix). The small differences illus-trated (Figs. 15-16, 20-21) representindividual variation. The rounded distaltips of the valvae distinguish the malegenitalia from those of the other mem-bers of the T. lisus complex (Figs. 17-19).Similarly, the rounded lamella post-vaginalis lacking spines distinguishesthe female genitalia from those of theother members of the T. lisus complex(Figs. 22-24).
CO1 sequences from individuals ofT. lisus from Central and South Americacluster (Fig. 25), albeit they have threedifferent BIN numbers.
Fig. 26. Wing venation of T. silma. Male on left.
VOLUME 118, NUMBER 4 563

On the basis of these results, we con-tinue to treat T. lisus as one geographi-cally variable species with T. hisbon andT. orsina as synonyms of T. lisus.
Theritas viresco and Theritasphoteinos
Druce (1907: 573) differentiated T.photeinos (from Brazil) from T. viresco(from Guyana) by “upper side brilliantshining indigo blue and forewing belowopalescent blue in place of green.”Draudt (1919-1920: 749) treated thesenames as forms, questioning whetherthey were distinct species, and Robbins(2004) synonymized them. In the studyseries of 18 males, wing color variescontinuously from blue to green, al-though most are green as in Fig. 5. Thereis no evidence from four male genitalicdissections and two male wing slides(Appendix) for more than one taxon.In lieu of evidence to the contrary,we continue to treat these names assynonyms.
Both sexes of T. viresco are easilydistinguished from other members of theT. lisus species group. As noted by Druce(1907), the postmedian line on the ven-tral surface of the male forewing variesin T. viresco from a series of dashes toremnant vestigial dashes, which differsconspicuously from other males of theT. lisus species group in which thepostmedian line is a band basally edgedwith brown (Figs. 1-5). The ventralhindwing has a long pouch (it looksmore like an incomplete fold in the wingin some individuals) that lacks androconia(Fig. 14), which distinguishes malesfrom others in the T. lisus species group(Figs 8-13). Finally, the female ventralhindwing basal black spot in cellSc+R1-Rs is consistently more con-spicuous than in other females of theT. lisus species group (Figs. 1-5).
Associating the sexes of T. lisus andT. viresco
The first line of evidence for associ-ating the sexes of T. lisus is from rearedimmatures. Three male and six femaleadults of T. lisus were reared by G.Bondar on 22 March 1937 from larvaeeating leaves of cacao (Theobroma ca-cao, Sterculiaceae) in �Agua Preta, Bahia,Brazil (vouchers in USNM). Similarly,three adults of T. lisus were reared byJ. A. J. Jarama on 27 August 1952 fromlarvae eating leaves of cacao in TingoMar�ıa, Hu�anuco, Peru (vouchers inUSNM). Similarly again, D. Janzenreared 14 males and 17 females in Cor-covado National Park, Osa Peninsula,Costa Rica on 16-26 November 1978(vouchers in USNM, additional foodplant rearing data lost). Finally, two maleand three female adults of T. lisus werereared by P. J. DeVries from gregariouslarvae eating flowers of Gustavia su-perba (Lecythidaceae) on Pipeline Road,northwest of Gamboa, Canal Area, Pan-ama (vouchers in USNM & DeVries’scollection, Lot 88-14). The immatureswere found as egg clusters, and theadults eclosed 16-18 March 1988. Asnoted by DeVries (1990) under thename Thecla hisbon, the caterpillars arenot myrmecophilous. Other recordedcaterpillar food plants for T. lisus canbe found in Zik�an (1956), Muyshondt(1973), and Janzen and Hallwachs (2016),including images of the gregariouslarvae.
The second line of evidence for as-sociating the sexes of T. lisus is distri-bution. All females in the T. lisus speciesgroup from Central America are T. lisusbecause males of the other species do notoccur there. As noted, females fromCentral America have the same distinctgenitalia as those from other parts of theNeotropics.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON564

The third line of evidence for associ-ating the sexes of T. lisus is a copulatingpair collected on the Tena-Pano Road,Napo, Ecuador at 600 m on 23 May 1988(vouchers in USNM, leg. D. Ahrenholz).
The fourth line of evidence for asso-ciating the sexes of T. lisus is the clus-tering of CO1 sequences (Fig. 25).
The evidence for associating the sexesof T. viresco is more limited than that forT. lisus. Both males and females ofT. viresco have the same AmazonianRegion distribution, and their CO1 se-quences cluster (Fig. 25).
Neotype of T. lisus
There is a male specimen of T. lisus inthe BMNH that has had a neotype labelfor a decade, but the designation has notbeen published.
It is premature and invalid to desig-nate a neotype of Papilio lisus Stoll,1790. Identification of the name hasbeen consistent for more than a century(Godman and Salvin 1887-1901, Druce1907, Draudt 1919-1920, D’Abrera1995, Robbins 2004). Papers on the bi-ology of the immatures of this species(Zik�an 1956, Muyshondt 1973, DeVries1990, Janzen & Hallwachs 2016) usethis identification or that of its geo-graphic synonym T. hisbon. Now thatgeographical variation of taxonomiccharacters has been assessed, the con-sistent use of this name is likely tocontinue. Further, at such point thatanalysis of nuclear DNA sequences canconfirm or falsify these results, a choiceof neotype, if one should be deemednecessary, can then be made based onmore information. Finally, according toArticle 75 of the Code (ICZN 1999),a neotype “is validly designated whenthere is an exceptional need”. This papermakes clear that no such exceptionalneed exists.
Theritas silma Martins, Faynel, &Robbins, new species
urn:lsid:zoobank.org:act:EBC455E7-381E-4F98-AD40-460083FF00B2
(Figs. 3, 12, 17, 22, 25-27)
Diagnosis.—We describe this speciesin Theritas, pending completion of aphylogeny of the Atlides Section (inprep.), because it possesses the charac-teristics of the T. lisus species group, asnoted above.
Theritas silma differs from the othermembers of the T. lisus species group.The male scent pouch on the ventralsurface of the hindwing in T. silma ex-tends from the base of the hindwing toa point more distal than the origin ofvein Cu2, which distinguishes it fromT. lisus (Figs. 8-12). The pouch containsconspicuous androconia, which differ-entiates it from T. viresco (Figs. 12, 14).More than half of the androconia aremore distal than the origin of vein Cu2in T. silma while more than half aremore basal in T. espiritosanto (B�alint &Moser) (Figs 12-13), but this differenti-ation is based on only six wing slides(Appendix).
Theritas silma is distinguished fromT. espiritosanto by the absence of whitescales dispersed on the ventral forewing(present in T. espiritosanto), and theyellow male ventral abdomen (brown inT. espiritosanto).
Theritas silma is distinguished fromthe other members of the T. lisus speciesgroup by its male genitalia. Most con-spicuously, the male genitalia of T. silmapossesses a conspicuous process on theventral edge of the fused vinculum/tegumen (Fig. 17), originally referred to asa “finger-like” projection (McDunnough1942). This process is lacking in themale genitalia of T. viresco andT. espiritosanto. There is an indicationof a process in T. lisus. Also, the male
VOLUME 118, NUMBER 4 565
genitalia valvae of T. silma are slenderand tapering to a fine point in lateralaspect while they are broad and blunt-tipped in the other members of thisgroup (Figs. 15-19). In ventral aspect,the tapered posterior tip of the valvaeof T. silma contrast markedly with thebroad valvae of T. lisus and T. espir-itosanto (Figs. 15-18). The distalcornutus of T. silma is slightly
bifurcate posteriorly. It is not bifurcate inT. espiritosanto, but is conspicuouslybifurcate in T. lisus.
Theritas silma is distinguished fromT. lisus by its female genitalia (Figs. 20-22). The posterior edge of the lamellapostvaginalis of T. silma has four “spinelike” short projections while that of T.lisus is smooth. Further, T. silma pos-sesses a constricted transparent, lightly
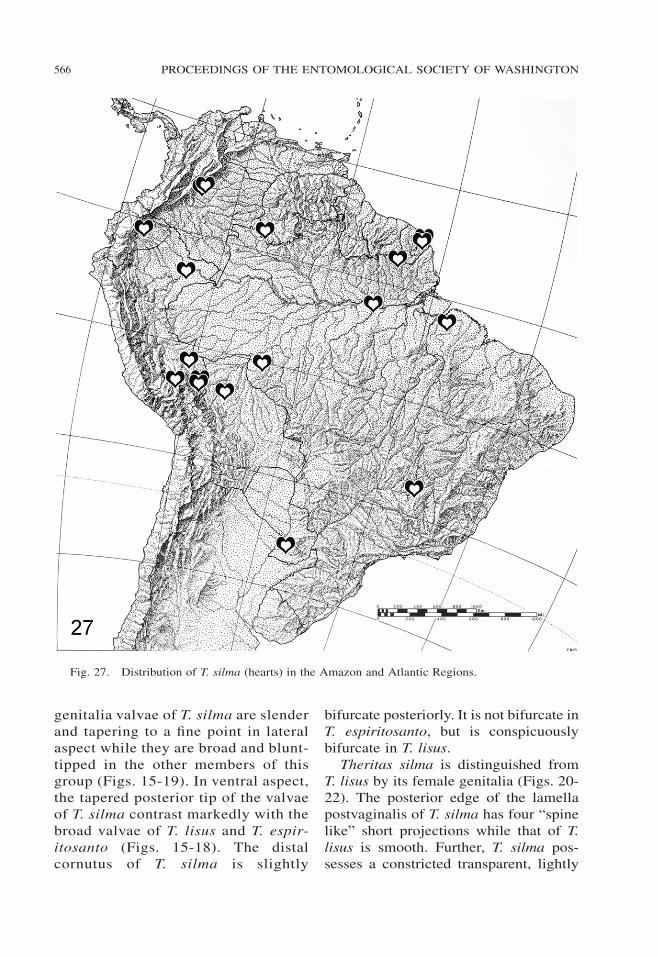
Fig. 27. Distribution of T. silma (hearts) in the Amazon and Atlantic Regions.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON566

sclerotized area in the ductus bursae(absent in T. lisus). The female genitaliaof T. silma differ slightly from thoseof T. espiritosanto and T. viresco (Figs.22-25), but our sample size for T. silmais not sufficiently large to conclude thatthese differences are distinguishing.
Description.—Head: ♂ muted moss-green forehead with blue para-oculararea; yellowish compound eyes; mutedmoss-green labial palp with scatteredwhite scales; short third labial palp seg-ment. ♀ muted brown forehead withwhitish para-ocular area; brown com-pound eyes; muted brown labial palpwith scattered white scales; long thirdlabial palp segment.
Thorax: ♂ thorax dorsally dark-blueiridescent with long blue bristles andventrally moss-green. ♀ thorax dorsallyblue-lilac, ventrally brown.
Wings: ♂ dark-blue iridescent dorsalforewing and hindwing with black fore-wing apex border; moss-green groundcolor of forewing and hindwing ventralsurface; ventral forewing with darkbrown marginal, submarginal and post-median bands, each one reaching Cu2vein; postmedian band with bluish scalesalong the band; ventral hindwing withdark brown marginal, submarginal andpostmedian band; submarginal bandwith iridescent bluish scales along theband; postmedian band formed by sep-arated dark brown dots with iridescentbluish scales; hindwing with two tails(Cu1 and Cu2 veins) (Fig. 3). ♀ irides-cent blue-lilac dorsal forewing andhindwing with black apex and marginalborder; brown ground color of forewingand hindwing ventral surface; ventralforewing with dark brown marginal, sub-marginal and postmedian bands; forewingpostmedian band with bluish scales alongthe band; ventral hindwing with darkbrown marginal, submarginal and post-median band; hindwing postmedian band
formed by separated dark brown dotswith iridescent bluish scales; hindwingwith two tails (Cu1 and Cu2 veins) thatare longer than those in males (Fig. 3).Venation and wing shape of each sexillustrated (Fig. 26).
Abdomen: ♂ abdomen dark-blue dor-sally, brown and yellow ventrally. ♀abdomen drab-blue dorsally, brown lat-erally, and yellow ventrally.
Genitalia: ♂ Eumaeini typical geni-talia with a narrow brush organ abovevinculum (secondary sexual organ);narrow valvae with angular posteriormargin (ventral view); medium saccus;tegumen with a ventral projection (Fig.17). ♀ genitalia with a straight ductusbursae with a membranous area forminga constriction (“neck”); presence ofa sclerotized process on lateral side ofthe ductus bursae; lamella postvaginaliswith spines (Fig. 22).
Size: ♂ mean forewing length (mea-sured from base to apex): 17.9 mm (sd =1.4; n = 3). ♂ mean hindwing length(measured from base to anal angle): 7.8mm (sd = 0.8; n = 3). ♀ mean forewinglength: 16.7 mm (sd = 0.7; n= 2).♀meanhindwing length: 8.2 mm (sd = 0; n = 2).
Type material.—Holotype. ♂ (Fig. 3,left). [white printed label] FRENCHGUIANA: Matoury/0 -100m. 4° 50’N,52°21’W/30 October 1988/leg. D. J.Harvey. [printed label on red paper]Holotype/Theritas silma/Martins, Faynel, &Robbins. Deposited USNM.
Paratypes.—(25♂, 4♀) French Guiana:1♂ Cayenne (USNM). 2♂ Mont GrandMatoury, 8 Sep 2012, 16 Sep 2012(USNM). 1♂ Route de Kaw, PK 15,24 Aug 2005 (MZSP). 1♀ Route de Kaw,PK 37, 27 Aug 1999 (MZSP). 2♂ Routede Kaw, PK 29, 6 Oct 1999, 23 Oct 2006(USNM, CF). 1♀ Route de Kaw, PK 7,27 Aug 2015 (CF). 1♂ Route de Kaw,5 Jan 2003 (CF). 2♂ Roura 4 Jan 2003(CF). 1♂ Crique Toussaint, 12 Feb
VOLUME 118, NUMBER 4 567

2000 (CF). 2♂ Maripasoula, Massifdu Mitaraka, N 2°13’590 O 54°27’460,11- 21 Aug 2015, La Planete Revisit�ee –MNHN / PNI Guyane 2015, APA 973-1(MNHN, CF). Venezuela: 1♂ Ama-zonas, San Carlos, 11 Mar 1984 (USNM).Colombia: 1♂ Cundinamarca, 5 km E.Medina, 4°30’N, 73°81W, 800 m(USNM). 1♀ Meta, Rio Negro, 750m,17 Aug 1980 (USNM). Ecuador: 1♂Napo, Cotundo, 800 m, 3 Sep 1977(USNM). Peru: 1♂ Loreto, 40 kmNE Iquitos, 14-20 Jan 1991 (USNM).1♂ Madre de Dios, Erika, 12°53’S,71°14’W, 550-650 m, 5 Sep 1989(MUSM). 2♂ Madre de Dios, PuertoMaldonado, 290 m, 15 Oct 1983(USNM). Brazil: 1♂ Par�a, Obidos, Sep1966 (USNM), 1♂&1♀ Par�a, SantoAntonio do Taua, Reserva “SonhoAzul”, PA 140, km 15, 4 Feb 2005 (CF).4♂ Rondonia, vicinity Caucalandia (20km S Ariquemes), 10°32’S, 62°48’W,160-350 m, 13 Oct 1991, 25 Oct 1991,30 Oct 1991, 19-29 Sep 1996 (USNM).
Other material examined.—Peru: 1♂Madre de Dios, Tambopata Reserve,12°50’S, 69°70’W, 300 m, 25 Oct 1990[excluded from the type series becauseit is a right hindwing that was found onthe ground]. Peru: 1♂ Madre de Dios,Pakitza (cf. Robbins et al. 1996 for de-tails about the locality). 9 Nov 1990[excluded from the type series becausethe data label, other than the date, waslost]. Bolivia: 1♂ “Peru” [excluded fromthe type series because this specimen isfrom the Schaus Collection, where thePeru labels appear to refer to a Bolivianlocality (Lamas & Robbins, in prep.)].Paraguay: 13 Sapucay, 18 Sep 1946[excluded from the type series becausethis specimen is not from the AmazonianRegion]. 1♂ Minas Gerais, Tres MariasDam, 1 May 1973 [excluded from thetype series because this specimen is notfrom the Amazonian Region]. French
Guiana: 10♂&6♀ in CF whose data islargely redundant with the paratypes al-ready listed. There are likely other fe-males of this species in museums curatedas T. lisus – they can easily be distin-guished by their genitalia (Figs. 15-24).
Etymology.—O nome da esp�ecie �euma homenagem a geneticista brasileiraDra. Silma Regina Ferreira Pereira, quecom amor e entusiasmo tem apoiadoa carreira dos seus filhos bi�ologos que sededicam ao estudo dos insetos. [Thisspecies is named for Brazilian geneticistDr. Silma Regina Ferreira Pereira. Withloving forbearance and enthusiasm shehas supported the entomological career ofher daughter and son.] The name is a non-latinized feminine noun in apposition.
Habitat, elevation, and distribution(Fig. 27) .—Theritas silma occurs in wetlowland forest under 800 m elevationthroughout the Amazon Region and inparts of the Atlantic Region, as charac-terized in Brown (1982).
Biology and behavior.—Both sexesof T. silma feed on the flowers of Cordiaschomburgkii A.DC. (Boraginaceae)during the morning in French Guiana(Faynel 2010, where this species wascalled “Theritas espiritosanto ?”). Maleterritorial behavior was observed at16:00 hours on hilltops or on the edgesof forest gaps (Faynel 2010). Thisspecies also is occasionally attracted bylight traps in French Guiana (vouchersin CF).
Remarks.—The relationship betweenT. silma and T. espiritosanto needs fur-ther investigation. Morphology of themale ventral hindwing scent pouch isvery similar (Figs. 12-13). They alsohave similar wing patterns (except forthe scattered white scale at the apex onthe ventral forewing of T. espiritosanto)and female genitalia that are probablyindistinguishable (Figs. 22-23). How-ever, the conspicuously different shape
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON568

of the male genitalia valvae and thepresence/absence of ventral finger pro-cess (as described above) immediatelydistinguish them. The two species arenot known to be sympatric, but it ispossible that the type series of T. espir-itosanto includes specimens of T. silma.
Nomenclatural list for the pheneticT. lisus species group
Theritas lisus (Stoll, 1790) (Papilio),type locality: Surinam
orsina (Hewitson, 1877) (Thecla),type locality: Bolivia
hisbon (Godman & Salvin, 1887)(Thecla), type localities: Guatemala;Costa Rica; Panama
Theritas silma Martins, Faynel, &Robbins, new species, type locality:French Guiana
Theritas espiritosanto (B�alint &Moser, 2007) (Denivia), type locality:Brazil (RJ)
Theritas viresco (H.H. Druce, 1907)(Thecla), type locality: Brazil (MA)
photeinos (H.H. Druce, 1907)(Thecla), type locality: Guyana
DISCUSSION
Aventral hindwing scent pouch in cellCu2-2A was discovered by Godman andSalvin (1887-1901) in Theritas lisus(treated as Thecla hisbon) and relatives,but its morphology was not investigatedfor more than a century (Robbins et al.2012). In genus Arcas the pouch may bepresent or absent, and when present, itmay or may not contain androconia(Robbins et al. 2012). This range ofvariation also occurs in the Theritas lisusspecies group, and in fact, all threeconditions occur intraspecifically withinT. lisus. Despite this intraspecific varia-tion, the structure was taxonomicallyuseful in the T. lisus species group.
Each of the species that we recognizeas distinct on the basis of morphologywas a distinct cluster in the CO1 phe-netic tree. However, the barcode indexnumbers (BIN, Ratnasingham and He-bert 2013) were discordant with thistaxonomy in T. lisus, and it is possiblethat the recognition algorithms need tobe modified to deal with widespreadspecies that have geographically vari-able CO1 sequences.
Robbins (2004) placed 23 describedand three undescribed species in Ther-itas. Two of the undescribed Theritasspecies (#112 & 127 in Robbins 2004)have been given the names T. boliboyerus(B�alint &Wojtusiak) and T. espiritosanto,respectively (B�alint & Wojtusiak 2006,B�alint & Moser 2007). The name T.anna Druce does not appear to belong toTheritas (Quental 2008). The discoveryof a distinct male of T. dabrerus B�alintelevates it from synonymy (B�alint &Wojtusiak 2006). Finally, with the de-scription of T. silma (#128 in Robbins2004) and confirmation of the classifi-cation of T. lisus and T. viresco in thispaper, there are now 26 described spe-cies in Theritas. Formally partitioningthese species, plus those of Arcas, intomonophyletic lineages awaits comple-tion of the phylogenetic analyses.
ACKNOWLEDGMENTS
The senior author did much of theresearch for this project while pursuinga graduate degree under the direction ofMarcelo Duarte. In addition, Marceloprovided us with original images ofPapilio lisus. The authors acknowledgeour gratitude to him.
The senior author thanks the SaoPaulo Research Foundation – FAPESP,Brazil (grants: 2012/03854-8 e 2013/00952-1; 2002/13898-0; 2010/14682-8;2011/50225-3) and the National Council
VOLUME 118, NUMBER 4 569

for Scientific and Technological Dev-elopment - CNPq/ SISBIOTA, Brazil(grant: 563332/2010-7) for financialsupport. Paratypes were collected duringthe “Our Planet Reviewed” Guyane-2015 expedition in the Mitaraka range,in the core area of the French GuianaAmazonian Park, organized by theMNHN and Pro-Natura International.The expedition was funded by the Eu-ropean Regional Development Fund(ERDF), the Conseil r�egional deGuyane, the Conseil g�en�eral de Guyane,the Direction de l’Environnement, del’Am�enagement et du Logement and bythe Ministere de l’�Education nationale,de l’Enseignement sup�erieur et de laRecherche, France. It was realized incollaboration with the Parc amazoniende Guyane and the Soci�et�e ento-mologique Antilles-Guyane (SEAG),France. For expertise with illustrationand other technical aspects of thisproject, we are grateful to KarieDarrow, Brian Harris, and VichaiMalikul at the USNM. For sharingspecimens and rearing data, we areindebted to D. Ahrenholz, R. Busby,P. DeVries, W. Hallwachs, and D. Janzen.We acknowledge the Canadian Cen-tre for DNA Barcoding (CCDB),Canada, especially the gracious helpof Jeremy DeWaard, Vlad Dinca, andE. V. Zakharov.
Literature Cited
B�alint, Z. and A. Moser. 2007. Description of
a Denivia species from south and southeast
Brazil with notes on the genus (Lepidoptera,
Lycaenidae: Eumaeini). Folia Entomologica
Hungarica 68: 147–156.B�alint, Z. and J. Wojtusiak 2006. Contribution to
the knowledge of Neotropical Lycaenidae -
reinstating names of three recently described
species and description of a new one (Lepi-
doptera: Eumaeini). Genus 17(2): 273–282.Brown, K. S. Jr. 1982. Historical and ecological
factors in the biogeography of aposematic
neotropical butterflies. American Zoologist
22(2): 453–471.Clench, H. K. 1961. Tribe Theclini. pp. 177-220.
In P. R. Ehrlich & A. H. Ehrlich, How to
know the butterflies, Brown Co., Dubuque,
Iowa, 262 pp., 525 figs.
Comstock, J. H. 1918. The wings of insects.
The Comstock Publishing Company, Ithaca.
430 pp.DeVries, P. J. 1990. Enhancement of symbioses
between butterfly caterpillars and ants by
vibrational communication. Science 248:
1104–1106.D’Abrera, B. 1995. Butterflies of the Neotropical
Region. Part VII. Lycaenidae. Hill House,
Victoria, pp. 1098–1270.
Draudt, M. 1919-1920. “Theclini F”. In: Die
Gross-Schmetterlinge der Erde, ed. A. Seitz,
vol. 5, Die amerikanischen Tagfalter,
pp.744–812. Stuttgart: Alfred Kernen.
Druce, H.H. 1907. On Neotropical Lycaenidae,
with descriptions of new species. Pro-
ceedings of the Zoological Society of London
1907: 566–632.
Evenhuis E. 2015. The insect and spider col-
lections of the world website. http://hbs.
bishopmuseum.org/codens/, (accessed 19
January 2016).Faynel, C. 2010. 3eme note sur les Theclinae de
Guyane (Lepidoptera: Lycaenidae). Lambil-
lionea 110 (1)(1): 9-16.
Godman, F.D. and O. Salvin. 1887-1901.
Biologia Centrali-Americana. Insecta. Lepidoptera-
Rhopalocera. Fam. Lycaenidae. 2: 1–112.Hewitson, W. C. 1865. Illustrations of Diurnal
Lepidoptera. Part I. Lycaenidae. London,
John Van Voorst, (2): 37–76, pl. 17-30.
Hewitson, W.C. 1877. Illustrations of Diurnal
Lepidoptera. Part I. Lycaenidae. London,
John Van Voorst, (7): 185–209, pl. 74-83.Hoffmann, C. C. 1940. Cat�alogo sistem�atico y
zoogeogr�afico de los lepid�opteros mexicanos.
Primera parte. Papilionoidea. Anales del In-
stituto de Biolog�ıa. Universidad nacional de
M�exico 11(2): 639–739.
ICZN. 1999. International Code of Zoological
Nomenclature, Fourth Edition. The Interna-
tional Trust for Zoological Nomenclature.Janzen, D. H. and W. Hallwachs. 2016. Dynamic
database for an inventory of the macrocater-
pillar fauna, and its food plants and parasit-
oids, of Area de Conservaci�on Guanacaste
(ACG), northwestern Costa Rica. http://janzen.
sas.upenn.edu. (accessed March 2016.
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON570

Klots, A. B. 1970. “Lepidoptera.” In Taxonomist’s
glossary of genitalia in insects, ed. S. L. Tuxen,pp. 115–130. Munksgaard, Copenhagen.
McDunnough, J. 1942. A New Canadian Strymon(Lycaenidae, Lepidoptera). The Canadian
Entomologist 74: 1.Muyshondt, A. 1973. Notes on the life cycle and
natural history of butterflies of El Salvador.I. Prepona omphale octavia (Nymphalidae).
Journal of the Lepidopterists’ Society27:210–219.
Quental, T.B. 2008. Systematics, sexual selectionand evolution of secondary sexual characters
in Neotropical Eumaeini butterflies (Lepi-doptera: Lycaenidae). Dissertation, Harvard
University, 226 pages.Ratnasingham, S., and P. D. N. Hebert. 2007. BOLD:
The Barcode of Life Data System (www.barcodinglife.org). Molecular Ecology Notes
10 pages. doi: 10.1111/j.1471-8286.2006.01678.x.Ratnasingham, S., and P. D. N. Hebert. 2013. A
DNA-Based Registry for All Animal Species:The Barcode Index Number (BIN) System.
PLoS ONE 8(8): e66213. doi:10.1371/journal.pone.066213. Ratnasingham & Hebert 2013.
Robbins, R. K. 1991. Evolution, comparativemorphology, and identification of the eu-
maeine butterfly genus Rekoa Kaye (Lycae-nidae: Theclinae). Smithsonian Contributions
to Zoology #498, 64 pp.
Robbins, R. K., G. Lamas, O. H. H. Mielke, D. J.Harvey, and M. Casagrande. 1996. Taxonomic
composition and ecological structure of the
species-rich butterfly community at Pakitza,
Parque Nacional del Manu, Peru. Pp. 217–
252, in Manu: The biodiversity of South-
eastern Peru (D. E. Wilson and A. Sandoval,
eds.). Smithsonian Institution Press, Wash-
ington DC. 679 pp. Robbins 2004.
Robbins, R. K. 2004. Lycaenidae. Theclinae.
Tribe Eumaeini, pp. 118–137. In: Lamas, G.
(Ed.), Checklist: Part 4A. Hesperioidea -
Papilionoidea. In: Heppner, J. B. (Ed.), Atlas
of Neotropical Lepidoptera. Volume 5A.
Gainesville, Association for Tropical Lepi-
doptera; Scientific Publishers.
Robbins, R. K., A. R. Martins, R. C. Busby, and
M. Duarte. 2012. Loss of male secondary
sexual structures in allopatry in the Neotropical
butterfly genus Arcas (Lycaenidae: Theclinae:
Eumaeini). Insect Systematics & Evolution 43:
35–65. DOI 10.1163/187631212X626195.
Thomas, M.B. 1893. The androchonia of Lepi-
doptera. The American Naturalist 27: 1018–
1021.Wilson, J. J. 2012. “DNA Barcodes for Insects.” In
DNABarcodes:Methods and Protocols,Methods
in Molecular Biology, eds. W. John Kress, and
David L. Erickson, pp. 17–46 (Chapter 3),
vol. 858. doi 10.1007/978-1-61779-591-6_3.Zik�an, J.F. 1956. Beitrage zur Biologie von 12
Theclinen-Arten. Dusenia (Curitiba) 7:
139–148.
VOLUME 118, NUMBER 4 571

APPENDIX
Theritas lisus species group genitalia dissection listings
Male GenitaliaWDF2182 ♂ T. viresco Peru
L378 ♂ T. viresco Eastern Colombia2013:24 ♂ (Martins) T. viresco Brazil (RO) (drawn)
1992:142 ♂ T. viresco Peru2013:25 ♂ (Martins) T. silma Peru: Loreto, 40Km NE Iquitos
WDF2183 ♂ T. silma Peru1992:140 ♂ T. silma Peru (drawn)
2013:75♂ T. silma USNM Brazil, MG2013:33 ♂ (Martins) T. espiritosanto Brazil (RJ) (drawn)
2013:26 ♂ (Martins) T. espiritosanto Brazil (SC)WDF2269♂ T. lisus (hisbon) Mexico
WDF2270♂ T. lisus (hisbon) GuatemalaWDF2268♂ T. lisus (hisbon) Guatemala
WDF2188♂ T. lisus (hisbon) GuatemalaWDF2185♂ T. lisus (hisbon) Costa Rica
WDF2187♂ T. lisus (hisbon) Costa RicaRKR3♂ T. lisus (hisbon) Panama (drawn)
2013:22 ♂ (Martins) T. lisus (hisbon) PanamaWDF2267♂ T. lisus (hisbon) Western Colombia
1982:26 ♂ T. lisus Eastern Colombia1992:145 ♂ T. lisus Peru
WDF5225 ♂ T. lisus Peru2013:43 ♂ (Martins) T. lisus Brazil (PA) (drawn)
WDF2324 ♂ T. lisus Brazil (BA)WDF2271 ♂ T. lisus Brazil (RJ)
WDF2189 ♂ T. lisus Brazil (SC)
Female Genitalia
1992: 143 ♀ T. viresco Peru1992: 144 ♀ T. viresco Peru (drawn)
WDF2181 ♀ T. viresco Eastern Ecuador1992: 151 ♀ T. silma Colombia (drawn)
WDF2184 ♀ T. silma Paraguay2013:47 ♀ (Martins) T. espiritosanto Brazil (ES) (drawn)
WDF2271♀ T. lisus (hisbon) MexicoWDF2273♀ T. lisus (hisbon) Guatemala
WDF2186♀ T. lisus (hisbon) Costa RicaWDF2274♀ T. lisus (hisbon) Costa Rica
2013:22 ♀ (Martins) T. lisus (hisbon) Panama (drawn)1982:27 ♀ T. lisus Eastern Colombia
1992:152 ♀ T. lisus Eastern Colombia1992:141 ♀ T. lisus Peru
1992:146 ♀ T. lisus Peru (drawn)1992:147 ♀ T. lisus Peru
WDF2276 ♀ T. lisus Brazil (BA)WDF2277 ♀ T. lisus Brazil (BA)
L379♀ T. lisus Brazil (MT)
PROCEEDINGS OF THE ENTOMOLOGICAL SOCIETY OF WASHINGTON572

Theritas lisus species group wing slide listings
Male Wing slides
WDF2182 ♂ T. viresco Peru (digitized)Martins 10 ♂ T. viresco Brazil (RO)
Martins 11 ♂ T. silma Peru: Loreto, 40Km NE IquitosMartins 21 ♂ T. silma Brazil, RO.
Martins 26 ♂ T. silma Brasil: Para (digitized)WDF2183 ♂ T. silma Peru
Martins 12 ♂ T. espiritosanto Brazil (SC)Martins 29 ♂ T. espiritosanto Brazil (SC) (digitized)
WDF2185♂ T. lisus (hisbon) Costa Rica (digitized)Martins 12 ♂ T. lisus (hisbon) Panama
Martins 30 ♂ T. lisus (hisbon) Panama (digitized)Martins 20 ♂ T. lisus Eastern Ecuador (digitized)
WDF2196 ♂ T. lisus Brazil (BA)Martins 31 ♂ T. lisus Brazil (SC) (digitized)
Female Wing slides
WDF2181 ♀ T. viresco Eastern EcuadorWDF2184 ♀ T. silma Paraguay
WDF2273 ♀ T. lisus (hisbon) GuatemalaWDF2277 ♀ T. lisus Brazil (BA)
Theritas lisus species group CO1 sequence information
CO1 sequence samples by species, sex, country, voucher number, and BOLDsequence/process ID. Vouchers in USNM and CF.
Theritas lisus, m, Costa Rica 07-SRNP-1913 MHMXK445-07Theritas lisus, m, Costa Rica 07-SRNP-1912 MHMXK449-07
Theritas lisus, m, Costa Rica 07-SRNP-1910 MHMXK447-07Theritas lisus, m, Costa Rica 07-SRNP-1900 MHMXK444-07
Theritas lisus, m, Costa Rica 07-SRNP-1886 MHMXK448-07Theritas lisus, m, Costa Rica 07-SRNP-1881 MHMXK446-07
Theritas lisus, m, French Guiana CF-LYC-226 NLYCA226-13Theritas lisus, m, French Guiana CF-LYC-227 NLYCA227-13
Theritas lisus, f, French Guiana CF-LYC-229 NLYCA229-13Theritas lisus, f, French Guiana CF-LYC-230 NLYCA230-13
Theritas lisus, m, French Guiana CF-LYC-627 NLYCA627-15Theritas silma, m, French Guiana CF-LYC-223 NLYCA223-13
Theritas silma, m, French Guiana CF-LYC-224 NLYCA224-13Theritas silma, m, French Guiana CF-LYC-225 NLYCA225-13
Theritas silma, m, French Guiana CF-LYC-601 NLYCA601-15Theritas silma, m, French Guiana CF-LYC-613 NLYCA613-15
Theritas viresco, m, French Guiana CF-LYC-222 NLYCA222-13
Theritas viresco, f, French Guiana CF-LYC-228 NLYCA228-13
VOLUME 118, NUMBER 4 573











![[11] Estados Intersexuais e Síndromes Sexuais Especiais - Dra. Sônia](https://static.fdocuments.in/doc/165x107/5695d2d31a28ab9b029bdbb0/11-estados-intersexuais-e-sindromes-sexuais-especiais-dra-sonia.jpg)






