sintesis acidosgrasos
39
Síntesis de Novo
-
Upload
freddynetas -
Category
Education
-
view
1.132 -
download
0
Transcript of sintesis acidosgrasos

Síntesis de Novo

TEMARIO 1. DIGESTIÓN. 2. ABSORCIÓN 3. CATABOLISMO:
A. Metabolismo del glicerol. Β. β-Oxidación de Ácidos Grasos:
i. Saturados. ii. Insaturados cis y trans.
C. Cetogénesis. 4. ANABOLISMO:
A. Biosíntesis de Ácidos Grasos: I. Saturados (síntesis de Novo) II. Elongación de Ácidos Grasos. III. Biosíntesis de Ácidos Grasos insaturados.
5. Biosíntesis de Eicosanoides. 6. Biosíntesis de Colesterol
2

Una gran proporción de los ácidos grasos que requieren los organismos son adquiridos en la dieta.
Los glúcidos y las proteínas consumidos con exceso en la dieta pueden ser convertidos a ácidos grasos y almacenados como triacil-gliceroles.
En el adulto humano, la síntesis de ácidos grasos se realiza fundamentalmente en el hígado, en la glándula mamaria, y en menor cantidad en el tejido adiposo.
El proceso radica en la incorporación de grupos de dos carbonos a partir de la acetil-CoA a la cadena creciente de un ácido graso.
Este proceso requiere ATP como aporte energético y NADPH como agente reductor.

Síntesis de Ácidos Grasos La vía de síntesis de ácidos grasos es muy semejante a la
inversa de la β-oxidación, pero presenta algunas diferencias importantes: Se realiza en el citoplasma en vez de en la mitocondria.
○ La biosíntesis de ácidos grasos en plantas ocurre en los plastos Usa al NADPH como donador de electrones El acarreador de los grupos acilos es la Proteína Acarreadora de
Acilos (ACP), en vez de la Coenzima A.

Acetil-CoA carboxilasa La Acetil-CoA carboxilasa cataliza la primera etapa de la
biosíntesis de los ácidos grasos y es una etapa determinante que controla la velocidad.
En la ruta biosintética, la reacción de condensación se halla acoplada a la hidrólisis de ATP impulsando, por tanto, la reacción hasta completarse.
Este proceso implica dos etapas: a) La carboxilación de la acetil-CoA, dependiente del ATP,
por la Acetil-CoA carboxilasa para formar Malonil-CoA, b) La descarboxilación exergónica del grupo malonilo en la
reacción de condensación catalizada por la Ácido graso sintasa.

La síntesis de ácidos grasos utiliza a la acetil-CoA como su principal sustrato. Como el proceso es muy endergónico la acetil-CoA debe ser activada, lo cual
se realiza a través de su carboxilación, convirtiéndose en malonil-CoA. El malonil-CoA se forma a partir de acetil-CoA y de bicarbonato, reacción que
consume ATP y que está catalizada por la Acetil-CoA carboxilasa. Como otras carboxilasas (la Piruvato y la Propionil-CoA carboxilasas) la Acetil-
CoA carboxilasa, utiliza a la Biotina como grupo prostético.

En los mamíferos y en las aves estas dos actividades enzimáticas, así como el transportador de carboxil-biotina están contenidas en una sola cadena polipeptídica de 230 kD.
En E. coli, y en los procariotes en general, estas etapas están catalizadas por subunidades separadas, conocidas como Biotina carboxilasa y transcarboxilasa, respectivamente.
También en procariotes, la biotina se encuentra unida en forma de resto de biocitina a una tercera subunidad llamada proteína transportadora de la carboxil biotina.
Todas ellas tienen que formar un complejo activo para que la carboxilación de la acetil-CoA pueda realizarse en este tipo de organismos
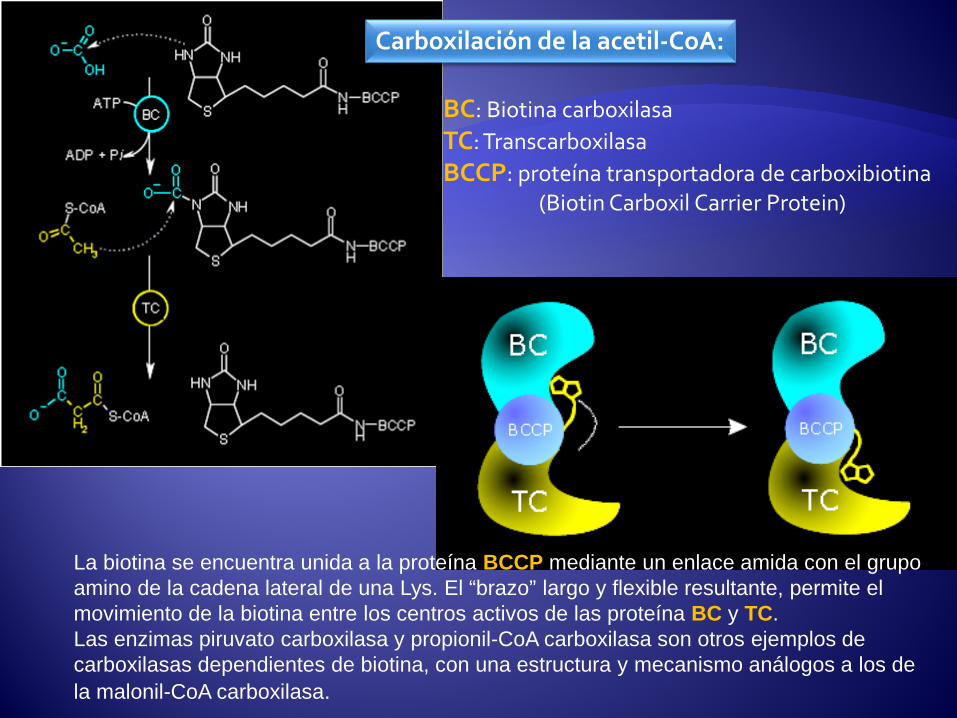
Carboxilación de la acetil-CoA:
BC: Biotina carboxilasa TC: Transcarboxilasa BCCP: proteína transportadora de carboxibiotina (Biotin Carboxil Carrier Protein)
La biotina se encuentra unida a la proteína BCCP mediante un enlace amida con el grupo amino de la cadena lateral de una Lys. El “brazo” largo y flexible resultante, permite el movimiento de la biotina entre los centros activos de las proteína BC y TC. Las enzimas piruvato carboxilasa y propionil-CoA carboxilasa son otros ejemplos de carboxilasas dependientes de biotina, con una estructura y mecanismo análogos a los de la malonil-CoA carboxilasa.

La síntesis de ácidos grasos se inicia con la carboxilación de la Acetil-CoA para formar Malonil-CoA. Éste, transferido a la ACP, será de aquí en adelante el donador activado de grupos de dos carbonos que se agregarán a la cadena creciente del ácido graso, de la misma manera que en este diagrama se muestra el primer paso de condensación entre el grupo acetilo, cedido temporalmente a la enzima Sintasa de ácidos grasos, con la Malonil-CoA

Ácido Graso Sintasa • Enlos animales cada paso de la síntesis de ácido palmítico (ácido graso
saturado de 16 carbonos) es catalizada por la Ácido graso Sintasa, un complejo enzimático de gran tamaño que posee todas las actividades enzimáticas necesarias.
• En las bacterias y otros procariotes, la enzimas involucradas se encuentran separadas y funcionan únicamente como un complejo enzimático en donde cada producto es cedido a la enzima siguiente para proseguir con su metabolismo.
• El Acetoacetil-ACP producido en la primera reacción se transforma en butirill-ACP.
• La secuencia de reacciones es la inversa de la β-oxidación: • reducción deshidratación hidrogenación.

Estequiometría • El Butirill-ACP puede después condensarse con otra
molécula de malonil-ACP. • Después de siete vueltas de este ciclo se produce palmitoil-
ACP, el cual es siempre el producto final en este mecanismo de síntesis. Ningún intermediario es liberado por el complejo enzimático antes de formar palmitoil-ACP.
• La hidrólisis del Palmitoil-ACP dará ácido palmítico que es enseguida convertido a Palmitoil-CoA.
• La estequiometría de la síntesis de ácido palmítico es: Acetil-CoA + 7 Malonil-CoA + 14 NADPH + 14 H+ ácido palmítico + 7 CO2 + 14 NADP+ + 8 CoA + 6 H2O • Ácidos grasos más largos, o insaturados, pueden ser
producidos a partir del ácido palmítico por elongasas y desaturasas.

Exportación de Acetil-CoA • La síntesis de ácidos grasos se realiza en el citoplasma, pero la
acetil-CoA se produce en la mitocondria. Por lo tanto la acetil-CoA debe cruzar la membrana mitocondrial interna y llegar al citosol antes de poder ser utilizado en la síntesis de ácidos grasos.
• Esto se realiza por medio del desvío, o lanzadera, del citrato: el citrato formado en la mitocondria difunde a través de la membrana mitocondrial al citoplasma, en donde es hidrolizado por la enzima Citrato-liasa dando acetil-CoA y oxaloacetato. Este último es reducido a malato y puede regresar a la matriz mitocondrial.
• El malato puede también ser procesado por la Enzima málica y de esta manera ser utilizado para producir parte del NADPH requerido para la síntesis de ácidos grasos
• El resto del NADPH requerido para la síntesis de ácidos grasos debe ser producido durante el ciclo de las pentosas fosfato.


Complejo Proteico de la Acido Graso Sintasa Proteína componente Actividad
Proteína portadora de acilo (ACP) Transporta grupos acilo en enlace tioéster
Acetil-CoA-ACP transacilasa (AT) Transfiere grupos acilo desde la CoA a la ACP en un residuo de Cys de KS
Malonil-CoA-ACP transacilasa (MT) Transfiere el grupo malonilo desde la CoA a la ACP
Beta-cetoacil-ACP sintasa (KS) Condensa grupos acilo y malonilo
Beta-cetoacil-ACP reductasa (KR) Reduce el grupo β-ceto a grupo β-hidroxilo
Beta-hidroxiacil-ACP deshidratasa (HD) Elimina H2O del β-hidroxiacil-ACP creando un doble enlace.
Enoil-ACP reductasa (ER) Reduce el doble enlace formando acil-ACP saturado
Palmitoil tioesterasa (Tease) Liberación del palmitato.
Las siglas entre paréntesis se refieren a los componentes del complejo ejemplificados en la siguiente figura que representa la estructura del dímero de la ácido graso sintasa

visualization by Kosi Gramatikoff
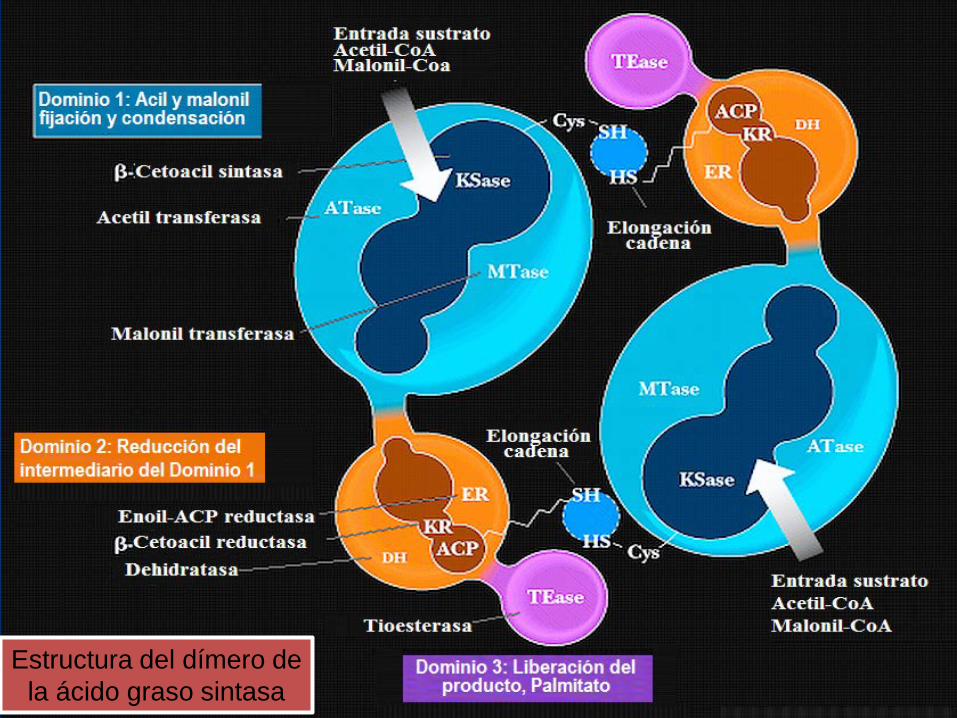
Estructura del dímero de la ácido graso sintasa

Mecanismo 1) Se transfiere una molécula de acetato de la Acetil-CoA al grupo –
SH de la ACP. Dominio 1: Acetil-CoA-ACP acetiltransacetilasa. 2) En seguida este fragmento de dos carbones es transferido a un
sitio de fijación temporal, el grupo tiol del residuo de cisteína en la estructura enzimática.
3) El sitio, ahora vacante, en la ACP acepta un residuo malonato de tres carbonos de la malonil-CoA. Dominio 1: Malonil-CoA-ACP transacilasa.
4) El grupo malonil pierde el HCO3- originalmente agregado por la
CoA carboxilasa, facilitando su ataque nucleofílico sobre el enlace tioéster que une el grupo acetilo al residuo de cisteína. El resultado será la formación de una unidad de cuatro carbonos unida al dominio de la ACP. La pérdida de energía libre de la descarboxilación dirige la reacción. Dominio 1: β-Cetoacil-ACP-Sintasa.

Mecanismo • Las siguientes tres reacciones convierten el grupo 3-cetoacil al
correspondiente grupo acilo saturado, por medio de un par de reacciones de reducción separadas por un paso de deshidratación:
1) El grupo ceto es reducido a un grupo alcohol. Dominio 2: β-cetoacil-ACP-Reductasa.
2) Se suprime una molécula de agua para introducir un doble enlace entre los carbonos 2 y 3 (carbonos α- y β-). Dominio 2: β-Hidroxiacil-ACP dehidratasa.
3) Se reduce el doble enlace. Dominio 2: Enoil-ACP reductasa • Finalmente cuando el ciclo se ha repetido un número adecuado de
veces para terminar con el Palmitil-CoA este es liberado por la hidrólisis del enlace tioester. Dominio 3: Tioesterasa

El primer paso es la condensación de la acetil-ACP y La malonil-ACP catalizada por la enzima β-Ceto-acil-ACP Sintasa.
Previamente la acetil-CoA ha cedido el radical acetilo para formar un enlace con la enzima condensante (KS)
El resultado es la formación de acetoacetil-ACP con liberación de CO2.
Aunque esta reacción es termodinámicamente desfavorable, la evolución de CO2 conduce la reacción hacia delante

Redución del Acetoacetil-ACP
• En este paso, la acetoacetil-ACP es reducida por la β-Cetoacil-ACP reductasa a D-3-hidroxibutiril-ACP, utilizando NADPH.
• El doble enlace se reduce a un grupo hidroxilo.
• Solo se forma el isómero D. Recordemos que durante la β-oxidación el isomero que se forma es el L

Deshidratación
• En esta reacción, el D-3-hidroxibutiril-ACP es deshidratado a crotonil-ACP, por la 3-Hidroxiacil-ACP deshidratasa.

Reducción del Crotonil-ACP Este es el paso final del ciclo de síntesis de ácidos grasos,
en este caso, el crotonil-ACP es reducido por el NADPH a butiril-ACP. La enzima que participa es la Enoil-ACP reductasa. A partir de este intermediario el ciclo se repite.
El producto final del proceso es siempre ácido palmítico, un ácido graso saturado de 16 carbonos, que es inmediatamente esterificado con la coenzima A, para formar palmitoil-CoA. Este mismo proceso se hace con cualquier ácido graso proveniente de la dieta. Para ser convertido, si es necesario, en otros ácidos grasos, el palmitoil-CoA sintetizado en el citoplasma, debe ser transportado al retículo endoplásmico.
Crotonil-ACP Butiril-ACP
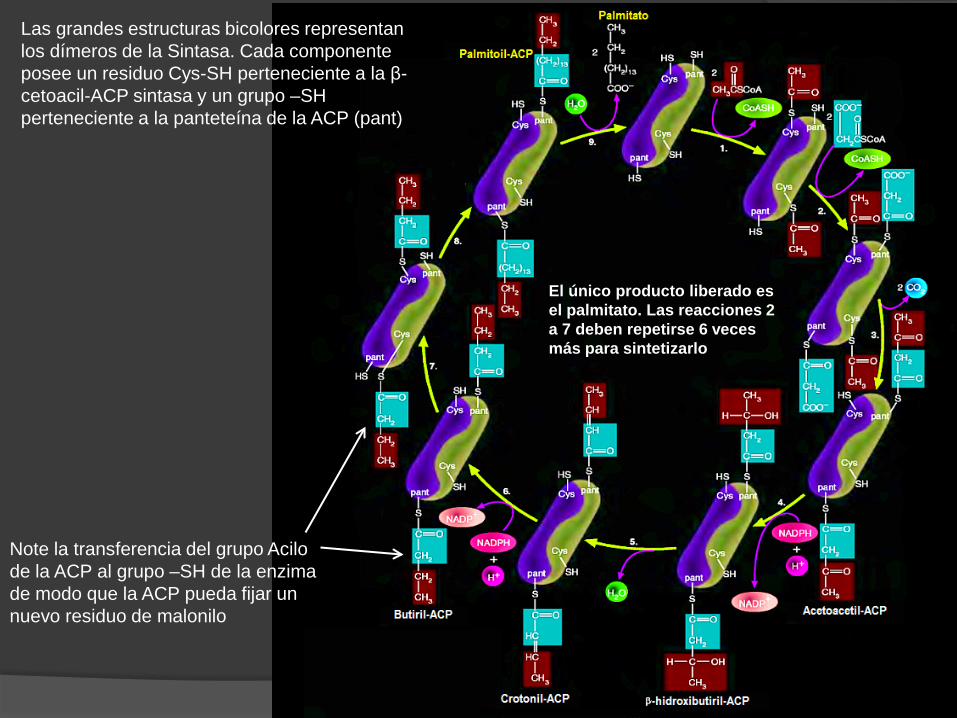
Note la transferencia del grupo Acilo de la ACP al grupo –SH de la enzima de modo que la ACP pueda fijar un nuevo residuo de malonilo
Las grandes estructuras bicolores representan los dímeros de la Sintasa. Cada componente posee un residuo Cys-SH perteneciente a la β-cetoacil-ACP sintasa y un grupo –SH perteneciente a la panteteína de la ACP (pant)
El único producto liberado es el palmitato. Las reacciones 2 a 7 deben repetirse 6 veces más para sintetizarlo


Diferencias entre Biosíntesis y Biodegradación de ácidos grasos

Hay tres niveles de control: 1) Dependiente de las concentraciones de metabolitos clave,
mediante inhibiciones o activaciones de tipo alostérico 2) Dependiente del control hormonal, mediante fosforilación o
defosforilación 3) A nivel de expresión génica, dependiente de la dieta.

Puntos de Control Alostérico • El punto principal de control es la acetil-CoA carboxilasa . • La malonil-CoA solo se emplea en la biosíntesis de ácidos grasos, por lo
que es lógico que el punto principal de control sea el de su síntesis. • Por otra parte, la inhibición alostérica que ejerce la malonil-CoA sobre la
acil-CoA: carnitina aciltransferasa I impide el paso de los acil-CoA a la mitocondria y su degradación mediante la β-oxidación, con lo que se evita que los acil-CoA se sinteticen y degraden simultáneamente

La insulina y el glucagón actúan induciendo la desfosforilación o la fosforilación, respectivamente, de la acetil-CoA carboxilasa mediante sus correspondientes cascadas de señalización dependientes de proteín cinasas.
En resumen, la insulina aumenta la síntesis de ácidos grasos, mientras que el glucagón o la adrenalina inhiben dicha síntesis.
La forma fosforilada de la enzima es un protómero inactivo formado por la asociación de 2 cadenea polipeptídicas, mientras que la forma fosforilada es una cadena de 20 a 40 protómeros (ver más adelante)

Los ácidos grasos polienoicos inhiben la expresión de los genes de las enzimas lipogénicas hepáticas.
Por otra parte, las dietas ricas en hidratos de carbono y bajas en lípidos inducen la expresión de estas enzimas, con lo que se favorece la síntesis de lípidos saturados a partir del exceso de hidratos de carbono.
No se conocen todavía los mecanismos responsables de estos efectos.

Diferencias Estructurales Aunque las reacciones de biosíntesis de ácidos
grasos son comunes en eubacterias y en los diferentes tipos de eucariotas, las enzimas muestran una interesante variación estructural.
Acetil-CoA Carboxilasa

Diferencias Estructurales Ácido Graso Sintasa
Bacterias y plastos: 6 enzimas aisladas + ACP en un complejo multienzimático
Hongos: 2 cadenas polipeptídicas multifuncionales asociadas A: ACP, sintasa y β-ceto reductasa B: dehidrasa, β -hidroxireductasa, acil-ACP transacilasa y
malonil-ACP transacilasa ○ Liberan palmitil-CoA (no tienen tioesterasa)
Animales: 2 cadenas polipeptídicas (240 kD c/u), cada una con TODAS las actividades enzimáticas, incluyendo la tioesterasa, y la ACP Cada subunidad es una cadena polipeptídica con 8
actividades

Elongación Las elongasas se hallan presentes
tanto en la mitocondria como en el retículo endoplásmico, pero los mecanismos de elongación en ambos sitios son diferentes.
La elongación mitocondrial (un proceso que es independiente de la ruta de la ácido graso sintasa) se produce por adición sucesiva y reducción de unidades de acetilo en una inversión de la oxidación del ácido graso; la única diferencia entre estas dos rutas tiene lugar en la etapa de reducción final en la que interviene el NADPH en lugar de FADH2 como coenzima redox terminal.

Elongación La elongación en el retículo endoplásmico
comprende la condensación sucesiva del malonil-CoA con acil-CoA. Estas reacciones van seguidas cada una por reducciones asociadas al NADPH, semejantes a las catalizadas por la ácido graso sintasa, la única diferencia consiste en que el ácido graso resulta alargado en forma de su derivado CoA en lugar del derivado ACP.

Ácidos Grasos no Saturados La síntesis de ácidos grasos no saturados implica una
reacción de desaturación, durante la cual se introduce una doble ligadura dentro de la cadena del ácido graso.
Poe ejemplo, en los humanos, la desaturación del ácido esteárico por la estearoil-CoA desaturasa-1, produce ácido oleico.
El ácido linoleico, doble insaturado, así como el triple insaturado ácido α-linolénico, no pueden ser sintetizados por los tejidos de los mamíferos, por lo que se consideran como ácidos grasos esenciales y deben ser suministrados en la dieta.
El ácido linoleico es el precursor del ácido araquidónico, el sustrato indispensable para la síntesis de prostaglandinas y otros compuestos llamados eicosanoides. Por supuesto, si la dieta es insuficiente en la cantidad de linoleico, el ácido araquidónico debe ser también aportado en la dieta.

Probablemente el papel más importante de los ácidos grasos poli-insaturados (AGPI) omega-3, EPA y DHA, es que sirven como precursores para la síntesis de un potente anti-inflamatorio llamado resolvins (Rvs) lípidos y protectins (PD). El Rvs ejercer sus acciones anti-inflamatorias mediante la promoción de la resolución de la inflamación del ciclo, de ahí la derivación de sus nombres como resolvins. El resolvins se sintetizan a partir de EPA o DHA. La serie D resolvins se derivan de DHA y la serie E de la EPA. Un adicionales antiinflamatorios lípidos derivados de DHA es protectin D1 (PD1).

Ácidos Grasos no Saturados Las enzimas responsables de la insaturación de los ácidos
grasos (es decir agregar cis-dobles enlaces) se encuentran en el retículo endoplásmico.
Reciben el nombre de desaturasas y pertenecen a la clase de oxidasas de función múltiple.
Las desaturasas requieren NADH y O2. Su actividad, combinada con la actividad de las elongasas son capaces de sintetizar una gran variedad de ácidos grasos.
Los humanos poseen desaturasas para los carbonos 9, 6, 5 y 4, pero carecen de la habilidad de introducir dobles enlaces desde el carbono 10 hasta el extremo ω de la cadena.
Esta es la razón de que algunos ácidos grasos poli-insaturados, como el linoleico y el linolénico, deban ser consumidos en la dieta y sean considerados esenciales.