
Research Article Evidence for Contribution of CD4+CD25...
7
Research Article Evidence for Contribution of CD4+CD25+ Regulatory T Cells in Maintaining Immune Tolerance to Human Factor IX following Perinatal Adenovirus Vector Delivery Megha S. Nivsarkar, 1 Suzanne M. K. Buckley, 1 Alan L. Parker, 2 Dany Perocheau, 1 Tristan R. McKay, 3 Ahad A. Rahim, 4 Steven J. Howe, 5 and Simon N. Waddington 1,6 1 Gene Transfer Technology Group, University College London, 86-96 Chenies Mews, London WC1E 6HXZ, UK 2 Institute of Cancer & Genetics, Cardiff University School of Medicine, Heath Park, Cardiff CF14 4XN, UK 3 Stem Cell Group, Cardiovascular & Cell Sciences Research Institute, St. George’s University of London, Cranmer Terrace, London SW17 0RE, UK 4 Department of Pharmacology, School of Pharmacy, University College London, 29-39 Brunswick Square, London WC1N 1AX, UK 5 Molecular and Cellular Immunology, UCL Institute of Child Health, 30 Guilford Street, London WC1N 1EH, UK 6 Antiviral Gene erapy Research Unit, Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa Correspondence should be addressed to Simon N. Waddington; [email protected] Received 26 November 2014; Accepted 12 January 2015 Academic Editor: Nejat K. Egilmez Copyright © 2015 Megha S. Nivsarkar et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Following fetal or neonatal gene transfer in mice and other species immune tolerance of the transgenic protein is frequently observed; however the underlying mechanisms remain largely undefined. In this study fetal and neonatal BALB/c mice received adenovirus vector to deliver human factor IX (hFIX) cDNA. e long-term tolerance of hFIX was robust in the face of immune challenge with hFIX protein and adjuvant but was eliminated by simultaneous administration of anti-CD25+ antibody. Naive irradiated BALB/c mice which had received lymphocytes from donors immunised with hFIX developed anti-hFIX antibodies upon immune challenge. Cotransplantation with CD4+CD25+ cells isolated from neonatally tolerized donors decreased the antibody response. In contrast, cotransplantation with CD4+CD25− cells isolated from the same donors increased the antibody response. ese data provide evidence that immune tolerance following perinatal gene transfer is maintained by a CD4+CD25+ regulatory population. 1. Introduction Fetal or neonatal gene transfer has been proposed as a potential therapeutic strategy with several advantages over adult gene transfer including increased likelihood of immune tolerization towards the transgenic protein [1, 2]. Absence of immune response aſter fetal or neonatal gene or stem cell delivery has been demonstrated in several studies [2–5], some of which have provided evidence for immune tolerance. We have previously observed prolonged expression of human factor IX (hFIX) aſter adenovirus-mediated gene transfer to fetal mice of the MF1 outbred strain. ere was transient and minimal production of anti-hFIX antibodies aſter immune challenge by repeated injection of purified hFIX protein and hFIX-adenovirus in adulthood [6]. To interrogate the immune mechanisms aſter fetal or neo- natal gene transfer more extensively we used inbred BALB/c mice. ese mice were chosen since they express relatively high levels of hFIX aſter neonatal retroviral gene transfer [4], exhibit a dominant 2 response [7], and, unlike outbred MF1 mice, can be used for adoptive transfer experiments. Adenovirus serotype 5 vector was used throughout. Hindawi Publishing Corporation Journal of Immunology Research Volume 2015, Article ID 397879, 6 pages http://dx.doi.org/10.1155/2015/397879
Transcript of Research Article Evidence for Contribution of CD4+CD25...

Research ArticleEvidence for Contribution of CD4+CD25+ Regulatory T Cellsin Maintaining Immune Tolerance to HumanFactor IX following Perinatal Adenovirus Vector Delivery
Megha S Nivsarkar1 Suzanne M K Buckley1 Alan L Parker2 Dany Perocheau1
Tristan R McKay3 Ahad A Rahim4 Steven J Howe5 and Simon N Waddington16
1Gene Transfer Technology Group University College London 86-96 Chenies Mews London WC1E 6HXZ UK2Institute of Cancer amp Genetics Cardiff University School of Medicine Heath Park Cardiff CF14 4XN UK3Stem Cell Group Cardiovascular amp Cell Sciences Research Institute St Georgersquos University of London Cranmer TerraceLondon SW17 0RE UK4Department of Pharmacology School of Pharmacy University College London 29-39 Brunswick Square London WC1N 1AX UK5Molecular and Cellular Immunology UCL Institute of Child Health 30 Guilford Street London WC1N 1EH UK6Antiviral Gene Therapy Research Unit Faculty of Health Sciences University of the Witwatersrand Johannesburg South Africa
Correspondence should be addressed to Simon N Waddington swaddingtonuclacuk
Received 26 November 2014 Accepted 12 January 2015
Academic Editor Nejat K Egilmez
Copyright copy 2015 Megha S Nivsarkar et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Following fetal or neonatal gene transfer in mice and other species immune tolerance of the transgenic protein is frequentlyobserved however the underlying mechanisms remain largely undefined In this study fetal and neonatal BALBc mice receivedadenovirus vector to deliver human factor IX (hFIX) cDNA The long-term tolerance of hFIX was robust in the face of immunechallenge with hFIX protein and adjuvant but was eliminated by simultaneous administration of anti-CD25+ antibody Naiveirradiated BALBcmice which had received lymphocytes from donors immunised with hFIX developed anti-hFIX antibodies uponimmune challenge Cotransplantation with CD4+CD25+ cells isolated from neonatally tolerized donors decreased the antibodyresponse In contrast cotransplantation with CD4+CD25minus cells isolated from the same donors increased the antibody responseThese data provide evidence that immune tolerance following perinatal gene transfer is maintained by a CD4+CD25+ regulatorypopulation
1 Introduction
Fetal or neonatal gene transfer has been proposed as apotential therapeutic strategy with several advantages overadult gene transfer including increased likelihood of immunetolerization towards the transgenic protein [1 2] Absence ofimmune response after fetal or neonatal gene or stem celldelivery has been demonstrated in several studies [2ndash5] someof which have provided evidence for immune tolerance Wehave previously observed prolonged expression of humanfactor IX (hFIX) after adenovirus-mediated gene transfer to
fetal mice of the MF1 outbred strain There was transient andminimal production of anti-hFIX antibodies after immunechallenge by repeated injection of purified hFIX protein andhFIX-adenovirus in adulthood [6]
To interrogate the immunemechanisms after fetal or neo-natal gene transfer more extensively we used inbred BALBcmice These mice were chosen since they express relativelyhigh levels of hFIX after neonatal retroviral gene transfer [4]exhibit a dominant Th2 response [7] and unlike outbredMF1 mice can be used for adoptive transfer experimentsAdenovirus serotype 5 vector was used throughout
Hindawi Publishing CorporationJournal of Immunology ResearchVolume 2015 Article ID 397879 6 pageshttpdxdoiorg1011552015397879
2 Journal of Immunology Research
2 Materials and Methods
Adenoviral vectors with CMV driving human factor FIX[6] (AdhFIX) and firefly luciferase [8] (AdLuc) genes wereprepared Vectors were titered by 293-cell plaque assays E16fetal mice received intravenous injection via the vitellinevessels [6] P0 neonates received injections under hypother-mic anaesthesia via the superficial temporal vein [9] Adultsreceived injections via the lateral tail vein Fetuses neonatesand adults received 1 times 109 1 times 109 and 3 times 1010 vectorgenomes per mouse Citrated blood was collected for plasmafor hFIX assay and for serum for antibody analysis Plasmaconcentrations of hFIX protein were determined using theAsserachrom FIX ELISA kit with pooled human plasmaas assay standard Antibodies against hFIX and adenoviruswere detected as previously described [6] Anti-CD25 IL2Rrat IgG1 antibody was produced by PC61 hybridoma cells(American Type Culture Collection Manassas USA) usinga capillary cell culture system as per manufacturerrsquos proto-col (CellMax Artificial Capillary Module Cellco Inc MDUSA) Antibody was purified and concentrated using affinitychromatography (Mab Trap Amersham Biosciences UKLimited) For vector genome quantitation 100 ng extractedDNAwas subjected to quantitative PCR (ABI Prism 7900HTApplied Biosystems Ltd Warrington UK) using hexon spe-cific primers (51015840CGCGGTGCGGCTGGTG31015840 and 51015840TGG-CGCATCCCATTCTCC31015840) normalized against a standardcurve ranging from 101 to 107 adenovirus particles Foradoptive transfer experiments recipients received sublethalirradiation (400 cGy) 24 h before cell transfer and hFIXimmunization Lymphocytes were harvested from inguinalcaudal and mesenteric lymph nodes and combined withsplenocytes CD4+CD25+ and CD4+CD25minus populationswere purified using magnetic beads (MACS Cell SeparationMiltenyi Biotec Surrey UK) Purity was confirmed by FACSanalysis From sensitized or naive pooled donors 3 times 107lymphocytes were injected intravenously into recipients 1 times104 or 1 times 105 CD4+CD25+ or CD4+CD25minus cells werecotransplanted both concentrations were equally effectivefor the two transplanted populations therefore antibodydata was pooled for subsequent analysis and presentationLuciferase expression in organ homogenates was assayedusing the Luciferase Assay System (Promega WI USA)and luminometer (Lucy1 Anthos Germany) as previouslydescribed [10]
3 Results
31 Longevity of Expression and Immune Challenge Wemea-sured hFIX expression and anti-hFIX antibodies followingintravascular injection of AdhFIX into fetal (119899 = 20) neona-tal (119899 = 21) (Figure 1(a)) and adult (119899 = 4) (Figure 1(b))BALBc mice After perinatal delivery hFIX concentrationswere greater than 2 120583gmL at 48 hours but declined toa stable concentration of approximately 50 ngmL beyond150 days some mice (asymp15) lost expression None devel-oped significant anti-hFIX antibodies In contrast followingadult delivery hFIX concentrations were between 01 and
06 120583gmL at 48 hours and declined to zero within 7 daysHigh concentrations of anti-hFIX antibodies were detectedfrom day 14 onwards
As a stronger test of immune tolerance to hFIX adultmice which had received AdhFIX in utero received twosubcutaneous injections (119899 = 9) of purified hFIX plus MPL-TDM adjuvant (at 140 and 175 days of age) (four receivedan additional challenge of AdhFIX at 275 days (Figure 1(c))to test tolerization to vector described below) Eight miceshowedno significant decrease in hFIX expression or increasein anti-hFIX antibodies after each challenge however onemouse which had no hFIX expression at the time ofchallenge developed high titer anti-hFIX antibodies Thissupports the observation of others that continual antigenexpression is required for maintenance of tolerance followingperinatal gene transfer [4] All naive adult controls developedhigh anti-hFIX antibody titers following challenge (119899 = 4data not shown)
32 Depletion of CD4+CD25+ Cells The mechanism ofimmune tolerance was investigated Six months after fetalAdhFIX injection mice still expressing hFIX (range 24ndash148 ngmL) received three intravenous injections of anti-CD25+ antibody (119899 = 7) to deplete CD25+ regulatory Tcells controls received rat IgG antibody (119899 = 7) (1mgmouse)(0 4 and 29 days) Subsequent to each antibody injectionmice received a subcutaneous injection of a mix of hFIX(1 gmouse) ovalbumin (1 gmouse) and adjuvant (Freundrsquoscomplete adjuvant and then 2x Freundrsquos incomplete adju-vant) Blood was collected 9 days after the third immunechallenge All mice that received the anti-CD25+ antibodydeveloped high anti-hFIX titres unlike those that receivedcontrol rat IgG (Figure 1(e)) These results provide evidencethat CD4+CD25+ regulatory T cells contribute to maintain-ing tolerance after perinatal gene transfer To confirm that thedepleting antibody was antigen-specific in breaking immunetolerance to hFIX rather than acting as a global immunestimulus anti-ovalbumin antibody titres were measuredUnexpectedly mice receiving depleting antibody developed areduced immune response to ovalbumin (Figure 1(f)) SincePC61 also binds to recently activated cells which expressthe IL-2 receptor 120572 those reactive to ovalbumin may beinactivated or deleted
33 Adoptive Transfer Experiments The mechanism of im-mune tolerance was interrogated further A cohort of micestill expressing hFIX more than 250 days after neonatalAdhFIX injection received three subcutaneous immunisa-tions with hFIX and Freundrsquos adjuvant immune tolerancewas confirmed by hFIX expression and absence of antibodiesA second cohort of naive mice was sensitised by subcuta-neous immunisation three times with hFIX and Freundrsquosadjuvant Irradiated BALBc recipients received lymphocytesfrom sensitized mice (119899 = 4) Alternatively recipients re-ceived cotransplantation of sensitised lymphocytes pluseither CD4+CD25+ (119899 = 9) or CD4+CD25minus (119899 = 7) lym-phocytes from tolerized mice (119899 = 9) Two recipients re-ceived whole lymphocyte population from naive donors
Journal of Immunology Research 3
005
1
435
3
225
Days after injection
1
0 50 100 150 200 250 3000
2
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
Xan
tibod
ies (120583
gm
L)
(a)
0
0
02
04
06
10 20 30 40 50
500
150
250
350
450
Days after injection
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(b)
0
05
10
15
30
0
60
90
500
1000
Days after injection0 50 100 150
hFIX hFIX AdhFIX
200 250 300
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(c)
Days after injection
Ant
i-ade
novi
rus a
ntib
odie
s
0
04
08
12
16 S1-3
C1-3
hFIX hFIX AdhFIX
0 50 100 150 200 250 300
(rel
ativ
e abs
orba
nce)
(d)
Serum dilution110000110001100A
nti-h
FIX
antib
odie
s (re
lativ
e abs
orba
nce)
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(e)
Serum dilution110000110001100
Ant
i-ova
lbum
in an
tibod
ies
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(rel
ativ
e abs
orba
nce)
(f)
Figure 1 Measurement of concentrations of hFIX and anti-hFIX antibodies following perinatal and adult AdhFIX injection (a)Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) following fetal (119899 = 20 small squares) and neonatal(119899 = 21 large diamonds) injection of AdhFIX (b) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph)following adult injection of AdhFIX (c) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) in miceinjected in utero with AdhFIX Two immune challenges with hFIX plus adjuvant and one AdhFIX were administered at 119905 = 140 175 and 275(labelled arrows) (d) Concentration of anti-adenovirus antibody in naive controls (C1-3) positive control mice which received hFIX plusadjuvant (S1-3) andmice having received AdhFIX in utero (plotted lines) Mice having received AdhFIX in utero received immune challengeswith hFIX plus adjuvant and one AdhFIX was administered at 119905 = 140 175 and 275 (labelled arrows) (e) Anti-hFIX antibodies in seriallydiluted plasma of mice tolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closedcircles and unbroken line) or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plusadjuvant Nine days after the final challenge sera were collected for analysis (f) Anti-ovalbumin antibodies in serially diluted plasma of micetolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closed circles and unbroken line)or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plus adjuvant Nine days after thefinal challenge sera were collected for analysis
4 Journal of Immunology Research
00
10
20
30
40
Day 7
sensitised versus
sensitised versus
Day 17 Day 24
Ant
i-hFI
X an
tibod
ies
(rel
ativ
e abs
orba
nce a
t 11
00)
+Sensitized
+CD
25+
+CD
25minus
+N
aive
+N
aive
+Sensitized
+CD
25+
+CD
25minus
+Sensitized
+CD
25+
+CD
25minus
+N
aive
P = 0037
CD25+
P = 0040
CD25minus
CD25minus
P lt 0001CD25+ versus
(a)
Heart Liver Spleen Thymus
1 times 107
1 times 106
1 times 105
1 times 104
Vect
or g
enom
es100
ng D
NA
(b)
Luci
fera
se ac
tivity
(RLU
mg
prot
ein
s)
Heartheart
Thymusthymus
Liver Spleen
1 times 104
1 times 103
1 times 102
1 times 101
1 times 100
1 times 10minus1
1 times 10minus2
1 times 10minus3
P lt 001
P lt 001
minusve minusve
(c)
Figure 2 Adoptive transfer of regulatory cell populations tissue distribution of vector and transgene expression (a) Anti-hFIX antibodyconcentrations in serum diluted 1100 following adoptive transfer of lymphocytes from hFIX-sensitised mice (black bar) cotransplantationof sensitised cells plus CD4+CD25+ cells from neonatally tolerized mice (blue bar) or plus CD4+CD25minus cells from neonatally tolerized mice(red bar) or adoptive transfer of lymphocytes from naive donors (grey bars) Recipients underwent sublethal irradiation adoptive transferand hFIX-immunisation 24 hours later and blood collection on days 7 17 and 24 (b) Quantitative PCR analysis of vector genome distributionand (c) luciferase expression six weeks after neonatal injection of AdLuc
Following subcutaneous immunisation with hFIX and Fre-undrsquos adjuvant blood was collected on days 7 17 and 24 foranti-hFIX analysis (Figure 2(a)) At all times recipients ofsensitised cells produced substantially more anti-hFIX anti-body than those which had received naive cells Importantlycotransplantation of CD4+CD25+ cells from tolerized micesignificantly decreased antibody production (general linearmodel Tukey pairwise comparison 119875 = 0037) whereascotransplantation of CD4+CD25minus cells from tolerized micesignificantly increased antibody production (119875 = 0040)Thisprovides further evidence that after perinatal gene transfer
a CD4+CD25+ population downregulates immune reactivityto hFIX as measured by anti-hFIX antibody productionThis cell population appears to mediate tolerance in agreater number of gene therapy protocols than previouslyanticipated For example CD4+CD25+ regulatory T cellshave been implicated in immune tolerance following livergene transfer using adenoassociated virus vector [11] Sincecotransplantation of CD4+CD25minus cells did not abolish anti-body development in recipients the role of other regulatorycell populations or deletional tolerance cannot be ruledout Indeed others provide evidence for deletional tolerance
Journal of Immunology Research 5
following injection of hFIX retrovirus into neonatal mice[12] Nevertheless recently Gaensler and colleagues providedelegant demonstration for a role of CD4+CD25+ regulatoryT cells in maintaining immune tolerance following neonatalintraperitoneal injection of adenoassociated virus vector[13]
34 Biodistribution of Vector Genomes and Gene Expres-sion Six to twelve weeks after neonatal AdLuc luciferaseexpression and vector genomes were quantitated in differentorgans Although fewer vector genomes were present inspleen (19880 plusmn 4600 vg100 ng DNA) and thymus (45072 plusmn14693 vg100 ng DNA) compared with those in other tissues(Figure 2(b)) significant luciferase expression was detected(Studentrsquos 119905-test 119875 lt 001) (Figure 2(c)) Therefore a role forcentral tolerance cannot be ruled out
35 Evidence for Tolerization to Vector Detection of anti-adenovirus antibodies was performed on four adult micefrom the experiment shown in Figure 1(d) (ie they receivedAdhFIX in utero two challenges of hFIX plus adjuvant on140 and 175 days and AdhFIX on 275 days) Only one ofthese mice developed anti-adenovirus antibodies after adultAdhFIX challenge as did all the naive adult mice receivingAdhFIX challenge (S1-3) whereas antibodies remained unde-tectable in the remaining three like in the nonchallengedcontrols (C1-3) (Figure 1(d)) This provides evidence forimmune tolerization to vector components This may beattributable to the use of first E1E3-deleted adenovirus whichmight lead to continuous expression of small amounts of viralproteins particularly since vector genomes can be detectedin thymuses from adult mice injected neonatally with AdLuc(Figure 2(b)) Alternatively it may arise from cotolerizationfrom simultaneous initial exposure to hFIX and adenovirusantigens
4 Discussion
Lifelong gene expression of various transgenes in mice hasbeen achieved following perinatal delivery of vectors derivedfrom adenovirus [6] gamma retrovirus [4] lentivirus [14]and adenoassociated virus [15 16] These studies have ledto improvements in the vector and expression cassette andprovidemotivation for childhood or even infant gene therapyfor early onset genetic diseases However understanding ofthe immune response to perinatal gene therapy remains poorHere we provide evidence for a contribution of regulatorytolerance to transgenic proteins following fetal and neonataldelivery in BALBc mice This supports recent observationsof regulatory tolerance following AAV vector delivery [13]One unexpected finding was evidence for immune toleranceto vector components Although this might permit multiplepostnatal administrations of the same vector it might alsorender the recipient vulnerable to infection by the archetypalvirus clearly an undesirable outcome One possible explana-tion for tolerization is residual and continuous expression ofviral proteinsThis provides strong justification for removal of
all viral sequences from vectors designed for long-term treat-ment of genetic diseases For example third generation HIV-based vectors contain approximately 18 kb of viral geneticmaterial This situation contrasts with recent observationsby Carlon and colleagues where perinatal delivery of AAVvector to murine airways benefitted from immune ignorance(rather than tolerance) to the capsid proteins permitting asecond administration at a later time point [17]
In these studies it is important to consider the develop-mental stage of the mouse immune system in late gestationrelative to the immune system of the fetal and newbornhuman Holladay and Smialowicz have provided an excellentcomparison of the developing mouse and human immunesystem By the end of the third trimester the human B cellsexpress surface IgG lymphocyte precursors are present inthe thymus thymocytes subsequently become responsive tomitogens and functional natural killer cells arise In contrastin the mouse lymphocyte precursors in the thymus aredetected at around 11 days of gestation whereas B cell sur-face IgG and thymocytes responsiveness to mitogen occursaround 17 days of gestation and NK cells arise after birth [18]In humans mature peripheral ab T cells first appear towardsthe end of the first trimester whereas inmice they only appearat the end of gestation [19] However the situation is morecomplex in that human serum IgM and IgG concentrationsdo not reach adult concentrations until 1-2 years of age and4ndash6 years of age respectively Moreover before 2 years ofage babies fail to mount humoral immune responses to someantigens [18] An additional complication is that recently ithas been shown that in humans as in mice and birds theimmune system is layered whereby hematopoiesis occurs inwaves specifically an early one which gives rise to fetal T cellsand a later one that gives rise to different adult T cells Thefetal and adult T cells possess different functional propertiesand gene expression patterns [20]
BALBc mice produce relatively small litters and fre-quently cannibalise their litters following maternal surgeryFor reasons of cost and to minimise animal wastage weswitched from fetal to neonatal gene transfer for adoptive Tcell transfer experiments as neonatal death following genetransfer is very rare Nevertheless it is important to recognisethat both qualitative and quantitative differences in immunefunction between E16 fetal and P0 newborn mouse exist
Over the course of the studywe switched fromMPL-TDMadjuvant to Freundrsquos adjuvant MPL-TDM is considered topose less of a risk of abscess formation and is therefore a betterchoice when complying with the 3Rs principle of refiningexperiments to minimise animal pain and distress Howeverwe chose to use Freundrsquos adjuvant for experiments involvingregulatory T cell depletion and adoptive T cell transfer asFreundrsquos adjuvant was considered likely to provide the mostpotent challenge [21]
Ex vivo luciferase expression correlated poorly with vec-tor copy number This may be explained by the observationthat hemoglobin interferes substantially with ex vivo lumi-nometry This is particularly apparent in the spleen whichretains a particularly high concentration of red blood cellsand hemoglobin even when perfused [22] An additionalexplanation may be that the CMV promoter which drives
6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Immunology Research
2 Materials and Methods
Adenoviral vectors with CMV driving human factor FIX[6] (AdhFIX) and firefly luciferase [8] (AdLuc) genes wereprepared Vectors were titered by 293-cell plaque assays E16fetal mice received intravenous injection via the vitellinevessels [6] P0 neonates received injections under hypother-mic anaesthesia via the superficial temporal vein [9] Adultsreceived injections via the lateral tail vein Fetuses neonatesand adults received 1 times 109 1 times 109 and 3 times 1010 vectorgenomes per mouse Citrated blood was collected for plasmafor hFIX assay and for serum for antibody analysis Plasmaconcentrations of hFIX protein were determined using theAsserachrom FIX ELISA kit with pooled human plasmaas assay standard Antibodies against hFIX and adenoviruswere detected as previously described [6] Anti-CD25 IL2Rrat IgG1 antibody was produced by PC61 hybridoma cells(American Type Culture Collection Manassas USA) usinga capillary cell culture system as per manufacturerrsquos proto-col (CellMax Artificial Capillary Module Cellco Inc MDUSA) Antibody was purified and concentrated using affinitychromatography (Mab Trap Amersham Biosciences UKLimited) For vector genome quantitation 100 ng extractedDNAwas subjected to quantitative PCR (ABI Prism 7900HTApplied Biosystems Ltd Warrington UK) using hexon spe-cific primers (51015840CGCGGTGCGGCTGGTG31015840 and 51015840TGG-CGCATCCCATTCTCC31015840) normalized against a standardcurve ranging from 101 to 107 adenovirus particles Foradoptive transfer experiments recipients received sublethalirradiation (400 cGy) 24 h before cell transfer and hFIXimmunization Lymphocytes were harvested from inguinalcaudal and mesenteric lymph nodes and combined withsplenocytes CD4+CD25+ and CD4+CD25minus populationswere purified using magnetic beads (MACS Cell SeparationMiltenyi Biotec Surrey UK) Purity was confirmed by FACSanalysis From sensitized or naive pooled donors 3 times 107lymphocytes were injected intravenously into recipients 1 times104 or 1 times 105 CD4+CD25+ or CD4+CD25minus cells werecotransplanted both concentrations were equally effectivefor the two transplanted populations therefore antibodydata was pooled for subsequent analysis and presentationLuciferase expression in organ homogenates was assayedusing the Luciferase Assay System (Promega WI USA)and luminometer (Lucy1 Anthos Germany) as previouslydescribed [10]
3 Results
31 Longevity of Expression and Immune Challenge Wemea-sured hFIX expression and anti-hFIX antibodies followingintravascular injection of AdhFIX into fetal (119899 = 20) neona-tal (119899 = 21) (Figure 1(a)) and adult (119899 = 4) (Figure 1(b))BALBc mice After perinatal delivery hFIX concentrationswere greater than 2 120583gmL at 48 hours but declined toa stable concentration of approximately 50 ngmL beyond150 days some mice (asymp15) lost expression None devel-oped significant anti-hFIX antibodies In contrast followingadult delivery hFIX concentrations were between 01 and
06 120583gmL at 48 hours and declined to zero within 7 daysHigh concentrations of anti-hFIX antibodies were detectedfrom day 14 onwards
As a stronger test of immune tolerance to hFIX adultmice which had received AdhFIX in utero received twosubcutaneous injections (119899 = 9) of purified hFIX plus MPL-TDM adjuvant (at 140 and 175 days of age) (four receivedan additional challenge of AdhFIX at 275 days (Figure 1(c))to test tolerization to vector described below) Eight miceshowedno significant decrease in hFIX expression or increasein anti-hFIX antibodies after each challenge however onemouse which had no hFIX expression at the time ofchallenge developed high titer anti-hFIX antibodies Thissupports the observation of others that continual antigenexpression is required for maintenance of tolerance followingperinatal gene transfer [4] All naive adult controls developedhigh anti-hFIX antibody titers following challenge (119899 = 4data not shown)
32 Depletion of CD4+CD25+ Cells The mechanism ofimmune tolerance was investigated Six months after fetalAdhFIX injection mice still expressing hFIX (range 24ndash148 ngmL) received three intravenous injections of anti-CD25+ antibody (119899 = 7) to deplete CD25+ regulatory Tcells controls received rat IgG antibody (119899 = 7) (1mgmouse)(0 4 and 29 days) Subsequent to each antibody injectionmice received a subcutaneous injection of a mix of hFIX(1 gmouse) ovalbumin (1 gmouse) and adjuvant (Freundrsquoscomplete adjuvant and then 2x Freundrsquos incomplete adju-vant) Blood was collected 9 days after the third immunechallenge All mice that received the anti-CD25+ antibodydeveloped high anti-hFIX titres unlike those that receivedcontrol rat IgG (Figure 1(e)) These results provide evidencethat CD4+CD25+ regulatory T cells contribute to maintain-ing tolerance after perinatal gene transfer To confirm that thedepleting antibody was antigen-specific in breaking immunetolerance to hFIX rather than acting as a global immunestimulus anti-ovalbumin antibody titres were measuredUnexpectedly mice receiving depleting antibody developed areduced immune response to ovalbumin (Figure 1(f)) SincePC61 also binds to recently activated cells which expressthe IL-2 receptor 120572 those reactive to ovalbumin may beinactivated or deleted
33 Adoptive Transfer Experiments The mechanism of im-mune tolerance was interrogated further A cohort of micestill expressing hFIX more than 250 days after neonatalAdhFIX injection received three subcutaneous immunisa-tions with hFIX and Freundrsquos adjuvant immune tolerancewas confirmed by hFIX expression and absence of antibodiesA second cohort of naive mice was sensitised by subcuta-neous immunisation three times with hFIX and Freundrsquosadjuvant Irradiated BALBc recipients received lymphocytesfrom sensitized mice (119899 = 4) Alternatively recipients re-ceived cotransplantation of sensitised lymphocytes pluseither CD4+CD25+ (119899 = 9) or CD4+CD25minus (119899 = 7) lym-phocytes from tolerized mice (119899 = 9) Two recipients re-ceived whole lymphocyte population from naive donors
Journal of Immunology Research 3
005
1
435
3
225
Days after injection
1
0 50 100 150 200 250 3000
2
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
Xan
tibod
ies (120583
gm
L)
(a)
0
0
02
04
06
10 20 30 40 50
500
150
250
350
450
Days after injection
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(b)
0
05
10
15
30
0
60
90
500
1000
Days after injection0 50 100 150
hFIX hFIX AdhFIX
200 250 300
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(c)
Days after injection
Ant
i-ade
novi
rus a
ntib
odie
s
0
04
08
12
16 S1-3
C1-3
hFIX hFIX AdhFIX
0 50 100 150 200 250 300
(rel
ativ
e abs
orba
nce)
(d)
Serum dilution110000110001100A
nti-h
FIX
antib
odie
s (re
lativ
e abs
orba
nce)
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(e)
Serum dilution110000110001100
Ant
i-ova
lbum
in an
tibod
ies
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(rel
ativ
e abs
orba
nce)
(f)
Figure 1 Measurement of concentrations of hFIX and anti-hFIX antibodies following perinatal and adult AdhFIX injection (a)Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) following fetal (119899 = 20 small squares) and neonatal(119899 = 21 large diamonds) injection of AdhFIX (b) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph)following adult injection of AdhFIX (c) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) in miceinjected in utero with AdhFIX Two immune challenges with hFIX plus adjuvant and one AdhFIX were administered at 119905 = 140 175 and 275(labelled arrows) (d) Concentration of anti-adenovirus antibody in naive controls (C1-3) positive control mice which received hFIX plusadjuvant (S1-3) andmice having received AdhFIX in utero (plotted lines) Mice having received AdhFIX in utero received immune challengeswith hFIX plus adjuvant and one AdhFIX was administered at 119905 = 140 175 and 275 (labelled arrows) (e) Anti-hFIX antibodies in seriallydiluted plasma of mice tolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closedcircles and unbroken line) or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plusadjuvant Nine days after the final challenge sera were collected for analysis (f) Anti-ovalbumin antibodies in serially diluted plasma of micetolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closed circles and unbroken line)or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plus adjuvant Nine days after thefinal challenge sera were collected for analysis
4 Journal of Immunology Research
00
10
20
30
40
Day 7
sensitised versus
sensitised versus
Day 17 Day 24
Ant
i-hFI
X an
tibod
ies
(rel
ativ
e abs
orba
nce a
t 11
00)
+Sensitized
+CD
25+
+CD
25minus
+N
aive
+N
aive
+Sensitized
+CD
25+
+CD
25minus
+Sensitized
+CD
25+
+CD
25minus
+N
aive
P = 0037
CD25+
P = 0040
CD25minus
CD25minus
P lt 0001CD25+ versus
(a)
Heart Liver Spleen Thymus
1 times 107
1 times 106
1 times 105
1 times 104
Vect
or g
enom
es100
ng D
NA
(b)
Luci
fera
se ac
tivity
(RLU
mg
prot
ein
s)
Heartheart
Thymusthymus
Liver Spleen
1 times 104
1 times 103
1 times 102
1 times 101
1 times 100
1 times 10minus1
1 times 10minus2
1 times 10minus3
P lt 001
P lt 001
minusve minusve
(c)
Figure 2 Adoptive transfer of regulatory cell populations tissue distribution of vector and transgene expression (a) Anti-hFIX antibodyconcentrations in serum diluted 1100 following adoptive transfer of lymphocytes from hFIX-sensitised mice (black bar) cotransplantationof sensitised cells plus CD4+CD25+ cells from neonatally tolerized mice (blue bar) or plus CD4+CD25minus cells from neonatally tolerized mice(red bar) or adoptive transfer of lymphocytes from naive donors (grey bars) Recipients underwent sublethal irradiation adoptive transferand hFIX-immunisation 24 hours later and blood collection on days 7 17 and 24 (b) Quantitative PCR analysis of vector genome distributionand (c) luciferase expression six weeks after neonatal injection of AdLuc
Following subcutaneous immunisation with hFIX and Fre-undrsquos adjuvant blood was collected on days 7 17 and 24 foranti-hFIX analysis (Figure 2(a)) At all times recipients ofsensitised cells produced substantially more anti-hFIX anti-body than those which had received naive cells Importantlycotransplantation of CD4+CD25+ cells from tolerized micesignificantly decreased antibody production (general linearmodel Tukey pairwise comparison 119875 = 0037) whereascotransplantation of CD4+CD25minus cells from tolerized micesignificantly increased antibody production (119875 = 0040)Thisprovides further evidence that after perinatal gene transfer
a CD4+CD25+ population downregulates immune reactivityto hFIX as measured by anti-hFIX antibody productionThis cell population appears to mediate tolerance in agreater number of gene therapy protocols than previouslyanticipated For example CD4+CD25+ regulatory T cellshave been implicated in immune tolerance following livergene transfer using adenoassociated virus vector [11] Sincecotransplantation of CD4+CD25minus cells did not abolish anti-body development in recipients the role of other regulatorycell populations or deletional tolerance cannot be ruledout Indeed others provide evidence for deletional tolerance
Journal of Immunology Research 5
following injection of hFIX retrovirus into neonatal mice[12] Nevertheless recently Gaensler and colleagues providedelegant demonstration for a role of CD4+CD25+ regulatoryT cells in maintaining immune tolerance following neonatalintraperitoneal injection of adenoassociated virus vector[13]
34 Biodistribution of Vector Genomes and Gene Expres-sion Six to twelve weeks after neonatal AdLuc luciferaseexpression and vector genomes were quantitated in differentorgans Although fewer vector genomes were present inspleen (19880 plusmn 4600 vg100 ng DNA) and thymus (45072 plusmn14693 vg100 ng DNA) compared with those in other tissues(Figure 2(b)) significant luciferase expression was detected(Studentrsquos 119905-test 119875 lt 001) (Figure 2(c)) Therefore a role forcentral tolerance cannot be ruled out
35 Evidence for Tolerization to Vector Detection of anti-adenovirus antibodies was performed on four adult micefrom the experiment shown in Figure 1(d) (ie they receivedAdhFIX in utero two challenges of hFIX plus adjuvant on140 and 175 days and AdhFIX on 275 days) Only one ofthese mice developed anti-adenovirus antibodies after adultAdhFIX challenge as did all the naive adult mice receivingAdhFIX challenge (S1-3) whereas antibodies remained unde-tectable in the remaining three like in the nonchallengedcontrols (C1-3) (Figure 1(d)) This provides evidence forimmune tolerization to vector components This may beattributable to the use of first E1E3-deleted adenovirus whichmight lead to continuous expression of small amounts of viralproteins particularly since vector genomes can be detectedin thymuses from adult mice injected neonatally with AdLuc(Figure 2(b)) Alternatively it may arise from cotolerizationfrom simultaneous initial exposure to hFIX and adenovirusantigens
4 Discussion
Lifelong gene expression of various transgenes in mice hasbeen achieved following perinatal delivery of vectors derivedfrom adenovirus [6] gamma retrovirus [4] lentivirus [14]and adenoassociated virus [15 16] These studies have ledto improvements in the vector and expression cassette andprovidemotivation for childhood or even infant gene therapyfor early onset genetic diseases However understanding ofthe immune response to perinatal gene therapy remains poorHere we provide evidence for a contribution of regulatorytolerance to transgenic proteins following fetal and neonataldelivery in BALBc mice This supports recent observationsof regulatory tolerance following AAV vector delivery [13]One unexpected finding was evidence for immune toleranceto vector components Although this might permit multiplepostnatal administrations of the same vector it might alsorender the recipient vulnerable to infection by the archetypalvirus clearly an undesirable outcome One possible explana-tion for tolerization is residual and continuous expression ofviral proteinsThis provides strong justification for removal of
all viral sequences from vectors designed for long-term treat-ment of genetic diseases For example third generation HIV-based vectors contain approximately 18 kb of viral geneticmaterial This situation contrasts with recent observationsby Carlon and colleagues where perinatal delivery of AAVvector to murine airways benefitted from immune ignorance(rather than tolerance) to the capsid proteins permitting asecond administration at a later time point [17]
In these studies it is important to consider the develop-mental stage of the mouse immune system in late gestationrelative to the immune system of the fetal and newbornhuman Holladay and Smialowicz have provided an excellentcomparison of the developing mouse and human immunesystem By the end of the third trimester the human B cellsexpress surface IgG lymphocyte precursors are present inthe thymus thymocytes subsequently become responsive tomitogens and functional natural killer cells arise In contrastin the mouse lymphocyte precursors in the thymus aredetected at around 11 days of gestation whereas B cell sur-face IgG and thymocytes responsiveness to mitogen occursaround 17 days of gestation and NK cells arise after birth [18]In humans mature peripheral ab T cells first appear towardsthe end of the first trimester whereas inmice they only appearat the end of gestation [19] However the situation is morecomplex in that human serum IgM and IgG concentrationsdo not reach adult concentrations until 1-2 years of age and4ndash6 years of age respectively Moreover before 2 years ofage babies fail to mount humoral immune responses to someantigens [18] An additional complication is that recently ithas been shown that in humans as in mice and birds theimmune system is layered whereby hematopoiesis occurs inwaves specifically an early one which gives rise to fetal T cellsand a later one that gives rise to different adult T cells Thefetal and adult T cells possess different functional propertiesand gene expression patterns [20]
BALBc mice produce relatively small litters and fre-quently cannibalise their litters following maternal surgeryFor reasons of cost and to minimise animal wastage weswitched from fetal to neonatal gene transfer for adoptive Tcell transfer experiments as neonatal death following genetransfer is very rare Nevertheless it is important to recognisethat both qualitative and quantitative differences in immunefunction between E16 fetal and P0 newborn mouse exist
Over the course of the studywe switched fromMPL-TDMadjuvant to Freundrsquos adjuvant MPL-TDM is considered topose less of a risk of abscess formation and is therefore a betterchoice when complying with the 3Rs principle of refiningexperiments to minimise animal pain and distress Howeverwe chose to use Freundrsquos adjuvant for experiments involvingregulatory T cell depletion and adoptive T cell transfer asFreundrsquos adjuvant was considered likely to provide the mostpotent challenge [21]
Ex vivo luciferase expression correlated poorly with vec-tor copy number This may be explained by the observationthat hemoglobin interferes substantially with ex vivo lumi-nometry This is particularly apparent in the spleen whichretains a particularly high concentration of red blood cellsand hemoglobin even when perfused [22] An additionalexplanation may be that the CMV promoter which drives
6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 3
005
1
435
3
225
Days after injection
1
0 50 100 150 200 250 3000
2
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
Xan
tibod
ies (120583
gm
L)
(a)
0
0
02
04
06
10 20 30 40 50
500
150
250
350
450
Days after injection
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(b)
0
05
10
15
30
0
60
90
500
1000
Days after injection0 50 100 150
hFIX hFIX AdhFIX
200 250 300
hFIX
conc
entr
atio
n (120583
gm
L)
Ant
i-hFI
X an
tibod
ies (120583
gm
L)
(c)
Days after injection
Ant
i-ade
novi
rus a
ntib
odie
s
0
04
08
12
16 S1-3
C1-3
hFIX hFIX AdhFIX
0 50 100 150 200 250 300
(rel
ativ
e abs
orba
nce)
(d)
Serum dilution110000110001100A
nti-h
FIX
antib
odie
s (re
lativ
e abs
orba
nce)
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(e)
Serum dilution110000110001100
Ant
i-ova
lbum
in an
tibod
ies
0
1
2
3
4 P lt 001 anti-CD25 versus rat IgG
(rel
ativ
e abs
orba
nce)
(f)
Figure 1 Measurement of concentrations of hFIX and anti-hFIX antibodies following perinatal and adult AdhFIX injection (a)Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) following fetal (119899 = 20 small squares) and neonatal(119899 = 21 large diamonds) injection of AdhFIX (b) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph)following adult injection of AdhFIX (c) Concentrations of hFIX (red upper graph) and anti-hFIX antibodies (black lower graph) in miceinjected in utero with AdhFIX Two immune challenges with hFIX plus adjuvant and one AdhFIX were administered at 119905 = 140 175 and 275(labelled arrows) (d) Concentration of anti-adenovirus antibody in naive controls (C1-3) positive control mice which received hFIX plusadjuvant (S1-3) andmice having received AdhFIX in utero (plotted lines) Mice having received AdhFIX in utero received immune challengeswith hFIX plus adjuvant and one AdhFIX was administered at 119905 = 140 175 and 275 (labelled arrows) (e) Anti-hFIX antibodies in seriallydiluted plasma of mice tolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closedcircles and unbroken line) or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plusadjuvant Nine days after the final challenge sera were collected for analysis (f) Anti-ovalbumin antibodies in serially diluted plasma of micetolerized to hFIX by fetal AdhFIX Mice received three injections of anti-CD25+ depleting antibody (black closed circles and unbroken line)or rat IgG negative control (blue open circles and dotted line) simultaneously with hFIX plus ovalbumin plus adjuvant Nine days after thefinal challenge sera were collected for analysis
4 Journal of Immunology Research
00
10
20
30
40
Day 7
sensitised versus
sensitised versus
Day 17 Day 24
Ant
i-hFI
X an
tibod
ies
(rel
ativ
e abs
orba
nce a
t 11
00)
+Sensitized
+CD
25+
+CD
25minus
+N
aive
+N
aive
+Sensitized
+CD
25+
+CD
25minus
+Sensitized
+CD
25+
+CD
25minus
+N
aive
P = 0037
CD25+
P = 0040
CD25minus
CD25minus
P lt 0001CD25+ versus
(a)
Heart Liver Spleen Thymus
1 times 107
1 times 106
1 times 105
1 times 104
Vect
or g
enom
es100
ng D
NA
(b)
Luci
fera
se ac
tivity
(RLU
mg
prot
ein
s)
Heartheart
Thymusthymus
Liver Spleen
1 times 104
1 times 103
1 times 102
1 times 101
1 times 100
1 times 10minus1
1 times 10minus2
1 times 10minus3
P lt 001
P lt 001
minusve minusve
(c)
Figure 2 Adoptive transfer of regulatory cell populations tissue distribution of vector and transgene expression (a) Anti-hFIX antibodyconcentrations in serum diluted 1100 following adoptive transfer of lymphocytes from hFIX-sensitised mice (black bar) cotransplantationof sensitised cells plus CD4+CD25+ cells from neonatally tolerized mice (blue bar) or plus CD4+CD25minus cells from neonatally tolerized mice(red bar) or adoptive transfer of lymphocytes from naive donors (grey bars) Recipients underwent sublethal irradiation adoptive transferand hFIX-immunisation 24 hours later and blood collection on days 7 17 and 24 (b) Quantitative PCR analysis of vector genome distributionand (c) luciferase expression six weeks after neonatal injection of AdLuc
Following subcutaneous immunisation with hFIX and Fre-undrsquos adjuvant blood was collected on days 7 17 and 24 foranti-hFIX analysis (Figure 2(a)) At all times recipients ofsensitised cells produced substantially more anti-hFIX anti-body than those which had received naive cells Importantlycotransplantation of CD4+CD25+ cells from tolerized micesignificantly decreased antibody production (general linearmodel Tukey pairwise comparison 119875 = 0037) whereascotransplantation of CD4+CD25minus cells from tolerized micesignificantly increased antibody production (119875 = 0040)Thisprovides further evidence that after perinatal gene transfer
a CD4+CD25+ population downregulates immune reactivityto hFIX as measured by anti-hFIX antibody productionThis cell population appears to mediate tolerance in agreater number of gene therapy protocols than previouslyanticipated For example CD4+CD25+ regulatory T cellshave been implicated in immune tolerance following livergene transfer using adenoassociated virus vector [11] Sincecotransplantation of CD4+CD25minus cells did not abolish anti-body development in recipients the role of other regulatorycell populations or deletional tolerance cannot be ruledout Indeed others provide evidence for deletional tolerance
Journal of Immunology Research 5
following injection of hFIX retrovirus into neonatal mice[12] Nevertheless recently Gaensler and colleagues providedelegant demonstration for a role of CD4+CD25+ regulatoryT cells in maintaining immune tolerance following neonatalintraperitoneal injection of adenoassociated virus vector[13]
34 Biodistribution of Vector Genomes and Gene Expres-sion Six to twelve weeks after neonatal AdLuc luciferaseexpression and vector genomes were quantitated in differentorgans Although fewer vector genomes were present inspleen (19880 plusmn 4600 vg100 ng DNA) and thymus (45072 plusmn14693 vg100 ng DNA) compared with those in other tissues(Figure 2(b)) significant luciferase expression was detected(Studentrsquos 119905-test 119875 lt 001) (Figure 2(c)) Therefore a role forcentral tolerance cannot be ruled out
35 Evidence for Tolerization to Vector Detection of anti-adenovirus antibodies was performed on four adult micefrom the experiment shown in Figure 1(d) (ie they receivedAdhFIX in utero two challenges of hFIX plus adjuvant on140 and 175 days and AdhFIX on 275 days) Only one ofthese mice developed anti-adenovirus antibodies after adultAdhFIX challenge as did all the naive adult mice receivingAdhFIX challenge (S1-3) whereas antibodies remained unde-tectable in the remaining three like in the nonchallengedcontrols (C1-3) (Figure 1(d)) This provides evidence forimmune tolerization to vector components This may beattributable to the use of first E1E3-deleted adenovirus whichmight lead to continuous expression of small amounts of viralproteins particularly since vector genomes can be detectedin thymuses from adult mice injected neonatally with AdLuc(Figure 2(b)) Alternatively it may arise from cotolerizationfrom simultaneous initial exposure to hFIX and adenovirusantigens
4 Discussion
Lifelong gene expression of various transgenes in mice hasbeen achieved following perinatal delivery of vectors derivedfrom adenovirus [6] gamma retrovirus [4] lentivirus [14]and adenoassociated virus [15 16] These studies have ledto improvements in the vector and expression cassette andprovidemotivation for childhood or even infant gene therapyfor early onset genetic diseases However understanding ofthe immune response to perinatal gene therapy remains poorHere we provide evidence for a contribution of regulatorytolerance to transgenic proteins following fetal and neonataldelivery in BALBc mice This supports recent observationsof regulatory tolerance following AAV vector delivery [13]One unexpected finding was evidence for immune toleranceto vector components Although this might permit multiplepostnatal administrations of the same vector it might alsorender the recipient vulnerable to infection by the archetypalvirus clearly an undesirable outcome One possible explana-tion for tolerization is residual and continuous expression ofviral proteinsThis provides strong justification for removal of
all viral sequences from vectors designed for long-term treat-ment of genetic diseases For example third generation HIV-based vectors contain approximately 18 kb of viral geneticmaterial This situation contrasts with recent observationsby Carlon and colleagues where perinatal delivery of AAVvector to murine airways benefitted from immune ignorance(rather than tolerance) to the capsid proteins permitting asecond administration at a later time point [17]
In these studies it is important to consider the develop-mental stage of the mouse immune system in late gestationrelative to the immune system of the fetal and newbornhuman Holladay and Smialowicz have provided an excellentcomparison of the developing mouse and human immunesystem By the end of the third trimester the human B cellsexpress surface IgG lymphocyte precursors are present inthe thymus thymocytes subsequently become responsive tomitogens and functional natural killer cells arise In contrastin the mouse lymphocyte precursors in the thymus aredetected at around 11 days of gestation whereas B cell sur-face IgG and thymocytes responsiveness to mitogen occursaround 17 days of gestation and NK cells arise after birth [18]In humans mature peripheral ab T cells first appear towardsthe end of the first trimester whereas inmice they only appearat the end of gestation [19] However the situation is morecomplex in that human serum IgM and IgG concentrationsdo not reach adult concentrations until 1-2 years of age and4ndash6 years of age respectively Moreover before 2 years ofage babies fail to mount humoral immune responses to someantigens [18] An additional complication is that recently ithas been shown that in humans as in mice and birds theimmune system is layered whereby hematopoiesis occurs inwaves specifically an early one which gives rise to fetal T cellsand a later one that gives rise to different adult T cells Thefetal and adult T cells possess different functional propertiesand gene expression patterns [20]
BALBc mice produce relatively small litters and fre-quently cannibalise their litters following maternal surgeryFor reasons of cost and to minimise animal wastage weswitched from fetal to neonatal gene transfer for adoptive Tcell transfer experiments as neonatal death following genetransfer is very rare Nevertheless it is important to recognisethat both qualitative and quantitative differences in immunefunction between E16 fetal and P0 newborn mouse exist
Over the course of the studywe switched fromMPL-TDMadjuvant to Freundrsquos adjuvant MPL-TDM is considered topose less of a risk of abscess formation and is therefore a betterchoice when complying with the 3Rs principle of refiningexperiments to minimise animal pain and distress Howeverwe chose to use Freundrsquos adjuvant for experiments involvingregulatory T cell depletion and adoptive T cell transfer asFreundrsquos adjuvant was considered likely to provide the mostpotent challenge [21]
Ex vivo luciferase expression correlated poorly with vec-tor copy number This may be explained by the observationthat hemoglobin interferes substantially with ex vivo lumi-nometry This is particularly apparent in the spleen whichretains a particularly high concentration of red blood cellsand hemoglobin even when perfused [22] An additionalexplanation may be that the CMV promoter which drives
6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
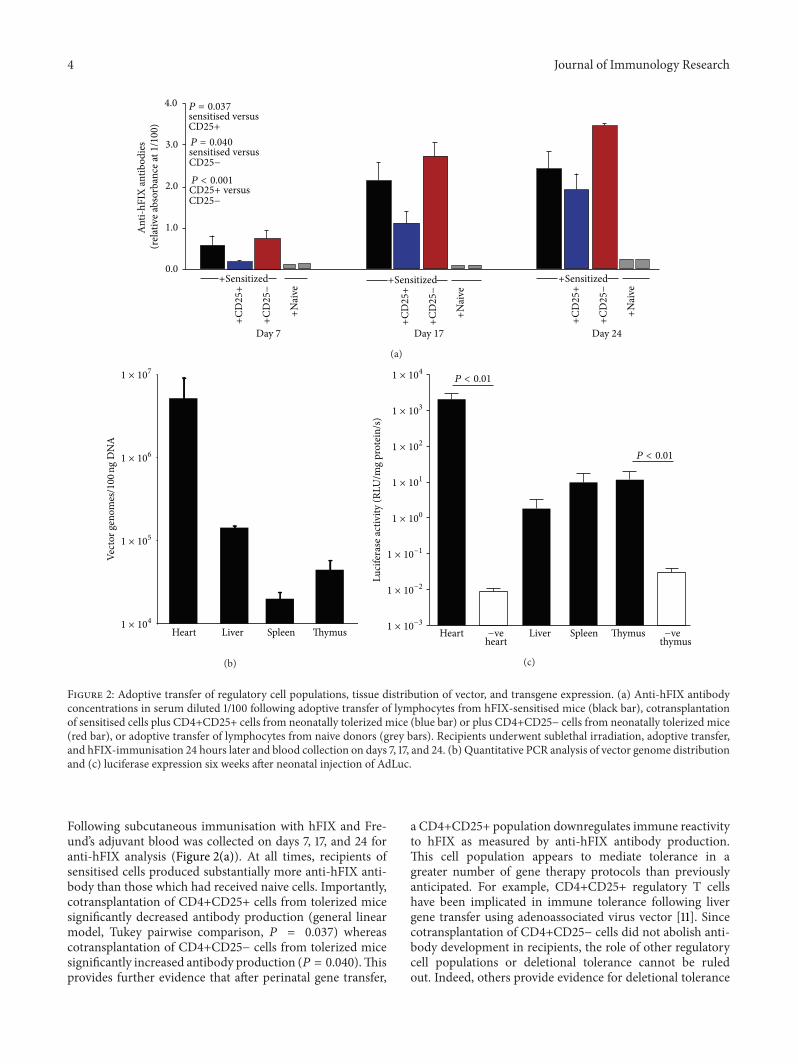
4 Journal of Immunology Research
00
10
20
30
40
Day 7
sensitised versus
sensitised versus
Day 17 Day 24
Ant
i-hFI
X an
tibod
ies
(rel
ativ
e abs
orba
nce a
t 11
00)
+Sensitized
+CD
25+
+CD
25minus
+N
aive
+N
aive
+Sensitized
+CD
25+
+CD
25minus
+Sensitized
+CD
25+
+CD
25minus
+N
aive
P = 0037
CD25+
P = 0040
CD25minus
CD25minus
P lt 0001CD25+ versus
(a)
Heart Liver Spleen Thymus
1 times 107
1 times 106
1 times 105
1 times 104
Vect
or g
enom
es100
ng D
NA
(b)
Luci
fera
se ac
tivity
(RLU
mg
prot
ein
s)
Heartheart
Thymusthymus
Liver Spleen
1 times 104
1 times 103
1 times 102
1 times 101
1 times 100
1 times 10minus1
1 times 10minus2
1 times 10minus3
P lt 001
P lt 001
minusve minusve
(c)
Figure 2 Adoptive transfer of regulatory cell populations tissue distribution of vector and transgene expression (a) Anti-hFIX antibodyconcentrations in serum diluted 1100 following adoptive transfer of lymphocytes from hFIX-sensitised mice (black bar) cotransplantationof sensitised cells plus CD4+CD25+ cells from neonatally tolerized mice (blue bar) or plus CD4+CD25minus cells from neonatally tolerized mice(red bar) or adoptive transfer of lymphocytes from naive donors (grey bars) Recipients underwent sublethal irradiation adoptive transferand hFIX-immunisation 24 hours later and blood collection on days 7 17 and 24 (b) Quantitative PCR analysis of vector genome distributionand (c) luciferase expression six weeks after neonatal injection of AdLuc
Following subcutaneous immunisation with hFIX and Fre-undrsquos adjuvant blood was collected on days 7 17 and 24 foranti-hFIX analysis (Figure 2(a)) At all times recipients ofsensitised cells produced substantially more anti-hFIX anti-body than those which had received naive cells Importantlycotransplantation of CD4+CD25+ cells from tolerized micesignificantly decreased antibody production (general linearmodel Tukey pairwise comparison 119875 = 0037) whereascotransplantation of CD4+CD25minus cells from tolerized micesignificantly increased antibody production (119875 = 0040)Thisprovides further evidence that after perinatal gene transfer
a CD4+CD25+ population downregulates immune reactivityto hFIX as measured by anti-hFIX antibody productionThis cell population appears to mediate tolerance in agreater number of gene therapy protocols than previouslyanticipated For example CD4+CD25+ regulatory T cellshave been implicated in immune tolerance following livergene transfer using adenoassociated virus vector [11] Sincecotransplantation of CD4+CD25minus cells did not abolish anti-body development in recipients the role of other regulatorycell populations or deletional tolerance cannot be ruledout Indeed others provide evidence for deletional tolerance
Journal of Immunology Research 5
following injection of hFIX retrovirus into neonatal mice[12] Nevertheless recently Gaensler and colleagues providedelegant demonstration for a role of CD4+CD25+ regulatoryT cells in maintaining immune tolerance following neonatalintraperitoneal injection of adenoassociated virus vector[13]
34 Biodistribution of Vector Genomes and Gene Expres-sion Six to twelve weeks after neonatal AdLuc luciferaseexpression and vector genomes were quantitated in differentorgans Although fewer vector genomes were present inspleen (19880 plusmn 4600 vg100 ng DNA) and thymus (45072 plusmn14693 vg100 ng DNA) compared with those in other tissues(Figure 2(b)) significant luciferase expression was detected(Studentrsquos 119905-test 119875 lt 001) (Figure 2(c)) Therefore a role forcentral tolerance cannot be ruled out
35 Evidence for Tolerization to Vector Detection of anti-adenovirus antibodies was performed on four adult micefrom the experiment shown in Figure 1(d) (ie they receivedAdhFIX in utero two challenges of hFIX plus adjuvant on140 and 175 days and AdhFIX on 275 days) Only one ofthese mice developed anti-adenovirus antibodies after adultAdhFIX challenge as did all the naive adult mice receivingAdhFIX challenge (S1-3) whereas antibodies remained unde-tectable in the remaining three like in the nonchallengedcontrols (C1-3) (Figure 1(d)) This provides evidence forimmune tolerization to vector components This may beattributable to the use of first E1E3-deleted adenovirus whichmight lead to continuous expression of small amounts of viralproteins particularly since vector genomes can be detectedin thymuses from adult mice injected neonatally with AdLuc(Figure 2(b)) Alternatively it may arise from cotolerizationfrom simultaneous initial exposure to hFIX and adenovirusantigens
4 Discussion
Lifelong gene expression of various transgenes in mice hasbeen achieved following perinatal delivery of vectors derivedfrom adenovirus [6] gamma retrovirus [4] lentivirus [14]and adenoassociated virus [15 16] These studies have ledto improvements in the vector and expression cassette andprovidemotivation for childhood or even infant gene therapyfor early onset genetic diseases However understanding ofthe immune response to perinatal gene therapy remains poorHere we provide evidence for a contribution of regulatorytolerance to transgenic proteins following fetal and neonataldelivery in BALBc mice This supports recent observationsof regulatory tolerance following AAV vector delivery [13]One unexpected finding was evidence for immune toleranceto vector components Although this might permit multiplepostnatal administrations of the same vector it might alsorender the recipient vulnerable to infection by the archetypalvirus clearly an undesirable outcome One possible explana-tion for tolerization is residual and continuous expression ofviral proteinsThis provides strong justification for removal of
all viral sequences from vectors designed for long-term treat-ment of genetic diseases For example third generation HIV-based vectors contain approximately 18 kb of viral geneticmaterial This situation contrasts with recent observationsby Carlon and colleagues where perinatal delivery of AAVvector to murine airways benefitted from immune ignorance(rather than tolerance) to the capsid proteins permitting asecond administration at a later time point [17]
In these studies it is important to consider the develop-mental stage of the mouse immune system in late gestationrelative to the immune system of the fetal and newbornhuman Holladay and Smialowicz have provided an excellentcomparison of the developing mouse and human immunesystem By the end of the third trimester the human B cellsexpress surface IgG lymphocyte precursors are present inthe thymus thymocytes subsequently become responsive tomitogens and functional natural killer cells arise In contrastin the mouse lymphocyte precursors in the thymus aredetected at around 11 days of gestation whereas B cell sur-face IgG and thymocytes responsiveness to mitogen occursaround 17 days of gestation and NK cells arise after birth [18]In humans mature peripheral ab T cells first appear towardsthe end of the first trimester whereas inmice they only appearat the end of gestation [19] However the situation is morecomplex in that human serum IgM and IgG concentrationsdo not reach adult concentrations until 1-2 years of age and4ndash6 years of age respectively Moreover before 2 years ofage babies fail to mount humoral immune responses to someantigens [18] An additional complication is that recently ithas been shown that in humans as in mice and birds theimmune system is layered whereby hematopoiesis occurs inwaves specifically an early one which gives rise to fetal T cellsand a later one that gives rise to different adult T cells Thefetal and adult T cells possess different functional propertiesand gene expression patterns [20]
BALBc mice produce relatively small litters and fre-quently cannibalise their litters following maternal surgeryFor reasons of cost and to minimise animal wastage weswitched from fetal to neonatal gene transfer for adoptive Tcell transfer experiments as neonatal death following genetransfer is very rare Nevertheless it is important to recognisethat both qualitative and quantitative differences in immunefunction between E16 fetal and P0 newborn mouse exist
Over the course of the studywe switched fromMPL-TDMadjuvant to Freundrsquos adjuvant MPL-TDM is considered topose less of a risk of abscess formation and is therefore a betterchoice when complying with the 3Rs principle of refiningexperiments to minimise animal pain and distress Howeverwe chose to use Freundrsquos adjuvant for experiments involvingregulatory T cell depletion and adoptive T cell transfer asFreundrsquos adjuvant was considered likely to provide the mostpotent challenge [21]
Ex vivo luciferase expression correlated poorly with vec-tor copy number This may be explained by the observationthat hemoglobin interferes substantially with ex vivo lumi-nometry This is particularly apparent in the spleen whichretains a particularly high concentration of red blood cellsand hemoglobin even when perfused [22] An additionalexplanation may be that the CMV promoter which drives
6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 5
following injection of hFIX retrovirus into neonatal mice[12] Nevertheless recently Gaensler and colleagues providedelegant demonstration for a role of CD4+CD25+ regulatoryT cells in maintaining immune tolerance following neonatalintraperitoneal injection of adenoassociated virus vector[13]
34 Biodistribution of Vector Genomes and Gene Expres-sion Six to twelve weeks after neonatal AdLuc luciferaseexpression and vector genomes were quantitated in differentorgans Although fewer vector genomes were present inspleen (19880 plusmn 4600 vg100 ng DNA) and thymus (45072 plusmn14693 vg100 ng DNA) compared with those in other tissues(Figure 2(b)) significant luciferase expression was detected(Studentrsquos 119905-test 119875 lt 001) (Figure 2(c)) Therefore a role forcentral tolerance cannot be ruled out
35 Evidence for Tolerization to Vector Detection of anti-adenovirus antibodies was performed on four adult micefrom the experiment shown in Figure 1(d) (ie they receivedAdhFIX in utero two challenges of hFIX plus adjuvant on140 and 175 days and AdhFIX on 275 days) Only one ofthese mice developed anti-adenovirus antibodies after adultAdhFIX challenge as did all the naive adult mice receivingAdhFIX challenge (S1-3) whereas antibodies remained unde-tectable in the remaining three like in the nonchallengedcontrols (C1-3) (Figure 1(d)) This provides evidence forimmune tolerization to vector components This may beattributable to the use of first E1E3-deleted adenovirus whichmight lead to continuous expression of small amounts of viralproteins particularly since vector genomes can be detectedin thymuses from adult mice injected neonatally with AdLuc(Figure 2(b)) Alternatively it may arise from cotolerizationfrom simultaneous initial exposure to hFIX and adenovirusantigens
4 Discussion
Lifelong gene expression of various transgenes in mice hasbeen achieved following perinatal delivery of vectors derivedfrom adenovirus [6] gamma retrovirus [4] lentivirus [14]and adenoassociated virus [15 16] These studies have ledto improvements in the vector and expression cassette andprovidemotivation for childhood or even infant gene therapyfor early onset genetic diseases However understanding ofthe immune response to perinatal gene therapy remains poorHere we provide evidence for a contribution of regulatorytolerance to transgenic proteins following fetal and neonataldelivery in BALBc mice This supports recent observationsof regulatory tolerance following AAV vector delivery [13]One unexpected finding was evidence for immune toleranceto vector components Although this might permit multiplepostnatal administrations of the same vector it might alsorender the recipient vulnerable to infection by the archetypalvirus clearly an undesirable outcome One possible explana-tion for tolerization is residual and continuous expression ofviral proteinsThis provides strong justification for removal of
all viral sequences from vectors designed for long-term treat-ment of genetic diseases For example third generation HIV-based vectors contain approximately 18 kb of viral geneticmaterial This situation contrasts with recent observationsby Carlon and colleagues where perinatal delivery of AAVvector to murine airways benefitted from immune ignorance(rather than tolerance) to the capsid proteins permitting asecond administration at a later time point [17]
In these studies it is important to consider the develop-mental stage of the mouse immune system in late gestationrelative to the immune system of the fetal and newbornhuman Holladay and Smialowicz have provided an excellentcomparison of the developing mouse and human immunesystem By the end of the third trimester the human B cellsexpress surface IgG lymphocyte precursors are present inthe thymus thymocytes subsequently become responsive tomitogens and functional natural killer cells arise In contrastin the mouse lymphocyte precursors in the thymus aredetected at around 11 days of gestation whereas B cell sur-face IgG and thymocytes responsiveness to mitogen occursaround 17 days of gestation and NK cells arise after birth [18]In humans mature peripheral ab T cells first appear towardsthe end of the first trimester whereas inmice they only appearat the end of gestation [19] However the situation is morecomplex in that human serum IgM and IgG concentrationsdo not reach adult concentrations until 1-2 years of age and4ndash6 years of age respectively Moreover before 2 years ofage babies fail to mount humoral immune responses to someantigens [18] An additional complication is that recently ithas been shown that in humans as in mice and birds theimmune system is layered whereby hematopoiesis occurs inwaves specifically an early one which gives rise to fetal T cellsand a later one that gives rise to different adult T cells Thefetal and adult T cells possess different functional propertiesand gene expression patterns [20]
BALBc mice produce relatively small litters and fre-quently cannibalise their litters following maternal surgeryFor reasons of cost and to minimise animal wastage weswitched from fetal to neonatal gene transfer for adoptive Tcell transfer experiments as neonatal death following genetransfer is very rare Nevertheless it is important to recognisethat both qualitative and quantitative differences in immunefunction between E16 fetal and P0 newborn mouse exist
Over the course of the studywe switched fromMPL-TDMadjuvant to Freundrsquos adjuvant MPL-TDM is considered topose less of a risk of abscess formation and is therefore a betterchoice when complying with the 3Rs principle of refiningexperiments to minimise animal pain and distress Howeverwe chose to use Freundrsquos adjuvant for experiments involvingregulatory T cell depletion and adoptive T cell transfer asFreundrsquos adjuvant was considered likely to provide the mostpotent challenge [21]
Ex vivo luciferase expression correlated poorly with vec-tor copy number This may be explained by the observationthat hemoglobin interferes substantially with ex vivo lumi-nometry This is particularly apparent in the spleen whichretains a particularly high concentration of red blood cellsand hemoglobin even when perfused [22] An additionalexplanation may be that the CMV promoter which drives
6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Journal of Immunology Research
luciferase expression in this vector may vary in activity indifferent tissue types
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Megha S Nivsarkar and Suzanne M K Buckley contributedequally to this work Megha S Nivsarkar prepared vectorMegha S Nivsarkar and Suzanne M K Buckley performedELISA and adoptive transferMegha SNivsarkar SuzanneMK Buckley and Simon NWaddington performed preclinicaldelivery Alan L Parker and Steven J Howe performed quan-titative PCR Ahad A Rahim Tristan R McKay and SimonN Waddington planned experiments and interpreted dataMegha S Nivsarkar and Simon N Waddington prepared thepaper
Acknowledgments
The authors thank Professor Ted Tuddenham for his helpfuldiscussion and review of the data Simon N WaddingtonSuzanne M K Buckley and Tristan R McKay are fundedby the European Research Council grant ldquoSomabiordquo SimonN Waddington Ahad A Rahim and Tristan R McKay arefunded by NC3Rs Grant NCL0017801 Ahad A Rahim wasfunded by MRC Grant G1000709
References
[1] S N Waddington M G Kramer R Hernandez-Alcoceba etal ldquoIn utero gene therapy current challenges and perspectivesrdquoMolecular Therapy vol 11 no 5 pp 661ndash676 2005
[2] N D Tran C D Porada G Almeida-Porada H A GlimpW French Anderson and E D Zanjani ldquoInduction of stableprenatal tolerance to 120573-galactosidase by in utero gene transferinto preimmune sheep fetusesrdquo Blood vol 97 no 11 pp 3417ndash3423 2001
[3] S Hayashi W H Peranteau A F Shaaban and A W FlakeldquoComplete allogeneic hematopoietic chimerism achieved by acombined strategy of in utero hematopoietic stem cell trans-plantation and postnatal donor lymphocyte infusionrdquo Bloodvol 100 no 3 pp 804ndash812 2002
[4] J Zhang L XuM E Haskins and K P Ponder ldquoNeonatal genetransfer with a retroviral vector results in tolerance to humanfactor IX in mice and dogsrdquo Blood vol 103 no 1 pp 143ndash1512004
[5] S N Waddington M S Nivsarkar A R Mistry et al ldquoPerma-nent phenotypic correction of Hemophilia B in immunocom-petent mice by prenatal gene therapyrdquo Blood vol 104 no 9 pp2714ndash2721 2004
[6] S N Waddington S M K Buckley M Nivsarkar et al ldquoInutero gene transfer of human factor IX to fetal mice can inducepostnatal tolerance of the exogenous clotting factorrdquo Blood vol101 no 4 pp 1359ndash1366 2003
[7] F Zhang Z Liang N Matsuki et al ldquoA murine locus onchromosome 18 controls NKT cell homeostasis and Th cell
differentiationrdquo The Journal of Immunology vol 171 no 9 pp4613ndash4620 2003
[8] S N Waddington J H McVey D Bhella et al ldquoAdenovirusserotype 5 hexon mediates liver gene transferrdquo Cell vol 132 no3 pp 397ndash409 2008
[9] A A Rahim A M S Wong K Hoefer et al ldquoIntravenousadministration of AAV29 to the fetal and neonatal mouseleads to differential targeting of CNS cell types and extensivetransduction of the nervous systemrdquo FASEB Journal vol 25 no10 pp 3505ndash3518 2011
[10] SM K Buckley S J Howe A A Rahim et al ldquoLuciferin detec-tion after intranasal vector delivery is improved by intranasalrather than intraperitoneal luciferin administrationrdquo HumanGene Therapy vol 19 no 10 pp 1050ndash1056 2008
[11] O Cao E Dobrzynski L Wang et al ldquoInduction and role ofregulatory CD4+CD25+ T cells in tolerance to the transgeneproduct following hepatic in vivo gene transferrdquo Blood vol 110no 4 pp 1132ndash1140 2007
[12] L Xu M Mei M E Haskins et al ldquoImmune response afterneonatal transfer of a human factor IX-expressing retroviralvector in dogs cats and micerdquo Thrombosis Research vol 120no 2 pp 269ndash280 2007
[13] Y Shi R Falahati J Zhang L Flebbe-Rehwaldt and K M LGaensler ldquoRole of antigen-specific regulatory CD4+ CD25+ Tcells in tolerance induction after neonatal IP administration ofAAV-hFIXrdquo Gene Therapy vol 20 no 10 pp 987ndash996 2013
[14] N J Ward S M K Buckley S N Waddington et al ldquoCodonoptimization of human factor VIII cDNAs leads to high-levelexpressionrdquo Blood vol 117 no 3 pp 798ndash807 2011
[15] R J Chandler and C P Venditti ldquoAdenovirus-mediated genedelivery rescues a neonatal lethal murine model of mut0methylmalonic acidemiardquo Human Gene Therapy vol 19 no 1pp 53ndash60 2008
[16] K D Foust X Wang V L McGovern et al ldquoRescue of thespinal muscular atrophy phenotype in a mouse model by earlypostnatal delivery of SMNrdquoNature Biotechnology vol 28 no 3pp 271ndash274 2010
[17] M S Carlon D Vidovic J Dooley et al ldquoImmunologicalignorance allows long-term gene expression following perinatalrAAV-mediated gene transfer to murine airwaysrdquoHuman GeneTherapy vol 25 no 6 pp 517ndash528 2014
[18] S D Holladay and R J Smialowicz ldquoDevelopment of themurine and human immune system differential effects ofimmunotoxicants depend on time of exposurerdquo EnvironmentalHealth Perspectives vol 108 no 3 pp 463ndash473 2000
[19] J E Mold and J M McCune ldquoImmunological toleranceduring fetal development from mouse to manrdquo Advances inImmunology vol 115 pp 73ndash111 2012
[20] J E Mold S Venkatasubrahmanyam T D Burt et al ldquoFetaland adult hematopoietic stem cells give rise to distinct T celllineages in humansrdquo Science vol 330 no 6011 pp 1695ndash16992010
[21] H F Stills Jr ldquoAdjuvants and antibody production dispellingthe myths associated with Freundrsquos complete and other adju-vantsrdquo ILAR Journal vol 46 no 3 pp 280ndash293 2005
[22] M Colin S Moritz H Schneider J Capeau C Coutelle andM C Brahimi-Horn ldquoHaemoglobin interferes with the ex vivoluciferase luminescence assay consequence for detection ofluciferase reporter gene expression in vivordquo Gene Therapy vol7 no 15 pp 1333ndash1336 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















