
Probing the Role of Divalent Metal Ions in a Bacterial Psychrophilic Metalloprotease
9
JOURNAL OF BACTERIOLOGY, July 2003, p. 4195–4203 Vol. 185, No. 14 0021-9193/03/$08.000 DOI: 10.1128/JB.185.14.4195–4203.2003 Copyright © 2003, American Society for Microbiology. All Rights Reserved. Probing the Role of Divalent Metal Ions in a Bacterial Psychrophilic Metalloprotease: Binding Studies of an Enzyme in the Crystalline State by X-Ray Crystallography Stephanie Ravaud, Patrice Gouet, Richard Haser,* and Nushin Aghajari Laboratoire de BioCristallographie, Institut de Biologie et Chimie des Prote ´ines, UMR 5086, CNRS-Universite ´ Claude Bernard Lyon 1, 69367 Lyon Cedex 07, France Received 3 February 2003/Accepted 24 April 2003 The psychrophilic alkaline metalloprotease (PAP) produced by a Pseudomonas bacterium isolated in Ant- arctica belongs to the clan of metzincins, for which a zinc ion is essential for catalytic activity. Binding studies in the crystalline state have been performed by X-ray crystallography in order to improve the understanding of the role of the zinc and calcium ions bound to this protease. Cocrystallization and soaking experiments with EDTA in a concentration range from 1 to 85 mM have resulted in five three-dimensional structures with a distinct number of metal ions occupying the ion-binding sites. Evolution of the structural changes observed in the vicinity of each cation-binding site has been studied as a function of the concentration of EDTA, as well as of time, in the presence of the chelator. Among others, we have found that the catalytic zinc ion was the first ion to be chelated, ahead of a weakly bound calcium ion (Ca 700) exclusive to the psychrophilic enzyme. Upon removal of the catalytic zinc ion, the side chains of the active-site residues His-173, His-179 and Tyr-209 shifted 4, 1.0, and 1.6 A ˚ , respectively. Our studies confirm and also explain the sensitivity of PAP toward moderate EDTA concentrations and propose distinct roles for the calcium ions. A new crystal form of native PAP validates our previous predictions regarding the adaptation of this enzyme to cold environments as well as the proteolytic domain calcium ion being exclusive for PAP independent of crystallization conditions. The serralysin family constitutes a group of bacterial metal- loproteases with a molecular mass 50 kDa, including psy- chrophilic alkaline protease (PAP) from Pseudomonas sp. strain TAC II 18 (11). These metalloproteases are further classified into the clan of metzincins (http://merops.sanger .ac.uk/), encompassing astacins, matrix metalloproteinases (collagenases), and snake venom proteinases (5). The metz- incins are characterized by the zinc-binding consensus se- quence HEXXHXXGXXHZ (residues 169 to 180 in PAP), where the histidine residues are the zinc ligands and the Z residue is characteristic for the different subfamilies (in the case of the serralysins, Z Pro), and by a conserved methio- nine residue (Met-207 in PAP). Although the sequence iden- tity between the different subfamilies of the metzincins is very low, their catalytic domains do have similar three-dimensional (3D) structures (2–4, 6, 7, 19–21). The serralysins are secreted as inactive zymogens into the medium via the hemolysin pathway, involving an ABC trans- porter, which recognizes a C-terminal secretion signal. Activa- tion occurs in the medium autocatalytically in the presence of divalent ions, especially Zn 2 and Ca 2 (12–14). The implication of zinc metalloproteases in many diseases, e.g., cancer or arthrosis (15), makes structural studies of these enzymes highly interesting. Here we report the results from X-ray crystallographic stud- ies of PAP, an alkaline protease produced by the psychrophilic Pseudomonas sp. strain TAC II 18, which originated in Ant- arctica. This strain, identified as a gram-negative bacterium (18), is one of numerous bacteria living at temperatures close to 0°C. To survive and grow under such conditions, these organisms have evolved several adaptations of their cellular components, especially of their enzymes (16–18, 30). Thus, PAP is three times more active at 20°C than a mesophilic counterpart from Pseudomonas aeruginosa but rapidly inacti- vated at 45°C. The cold-adapted enzyme also displays a greater sensitivity to moderate EDTA concentrations, and fluores- cence studies have indicated enzyme unfolding after 2 h of incubation in 2 mM EDTA (11). The PAP molecule is made up of 463 amino acid residues distributed in two domains, and its tertiary structure is com- parable to mesophilic homologues from P. aeruginosa alkaline protease (AP) and Serratia marcescens protease (SMP) (1–3). Four -helices and a mixed five-stranded -sheet form the catalytic N-terminal domain to which the active-site Zn 2 ion and a Ca 2 ion (Ca 700, PAP numbering) are bound. This Ca 2 ion is not present in its mesophilic counterparts, AP and SMP, but it should be noticed that a calcium ion has been observed in the 3D structure of the metzincin adamalysin II in a position similar to that of Ca 700 (20). The C-terminal domain which may play a role in the folding of the molecule after transmembrane translocation forms an extended roll with 20 successive -strands wound in a right- handed spiral. The number of Ca 2 ions bound to this C- terminal domain varies from six to seven in the different or- ganisms and crystal forms. When we compared the 3D structure with those of the me- sophilic counterparts, AP and SMP, the most remarkable dif- ferences were found in the proteolytic domain. First, the active site is more accessible due, in part, to deletions in surrounding * Corresponding author. Mailing address: Laboratoire de BioCris- tallographie, Institut de Biologie et Chimie des Prote ´ines, UMR 5086, CNRS-UCBL1, 7 Passage du Vercors, 69367 Lyon, Cedex 07, France. Phone: 33(0)472722608. Fax: 33(0)472722616. E-mail: [email protected]. 4195 Downloaded from https://journals.asm.org/journal/jb on 22 January 2022 by 191.240.25.162.
Transcript of Probing the Role of Divalent Metal Ions in a Bacterial Psychrophilic Metalloprotease

JOURNAL OF BACTERIOLOGY, July 2003, p. 4195–4203 Vol. 185, No. 140021-9193/03/$08.00�0 DOI: 10.1128/JB.185.14.4195–4203.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Probing the Role of Divalent Metal Ions in a Bacterial PsychrophilicMetalloprotease: Binding Studies of an Enzyme in the
Crystalline State by X-Ray CrystallographyStephanie Ravaud, Patrice Gouet, Richard Haser,* and Nushin AghajariLaboratoire de BioCristallographie, Institut de Biologie et Chimie des Proteines, UMR 5086,
CNRS-Universite Claude Bernard Lyon 1, 69367 Lyon Cedex 07, France
Received 3 February 2003/Accepted 24 April 2003
The psychrophilic alkaline metalloprotease (PAP) produced by a Pseudomonas bacterium isolated in Ant-arctica belongs to the clan of metzincins, for which a zinc ion is essential for catalytic activity. Binding studiesin the crystalline state have been performed by X-ray crystallography in order to improve the understandingof the role of the zinc and calcium ions bound to this protease. Cocrystallization and soaking experiments withEDTA in a concentration range from 1 to 85 mM have resulted in five three-dimensional structures with adistinct number of metal ions occupying the ion-binding sites. Evolution of the structural changes observed inthe vicinity of each cation-binding site has been studied as a function of the concentration of EDTA, as well asof time, in the presence of the chelator. Among others, we have found that the catalytic zinc ion was the firstion to be chelated, ahead of a weakly bound calcium ion (Ca 700) exclusive to the psychrophilic enzyme. Uponremoval of the catalytic zinc ion, the side chains of the active-site residues His-173, His-179 and Tyr-209 shifted�4, 1.0, and 1.6 A, respectively. Our studies confirm and also explain the sensitivity of PAP toward moderateEDTA concentrations and propose distinct roles for the calcium ions. A new crystal form of native PAPvalidates our previous predictions regarding the adaptation of this enzyme to cold environments as well as theproteolytic domain calcium ion being exclusive for PAP independent of crystallization conditions.
The serralysin family constitutes a group of bacterial metal-loproteases with a molecular mass �50 kDa, including psy-chrophilic alkaline protease (PAP) from Pseudomonas sp.strain TAC II 18 (11). These metalloproteases are furtherclassified into the clan of metzincins (http://merops.sanger.ac.uk/), encompassing astacins, matrix metalloproteinases(collagenases), and snake venom proteinases (5). The metz-incins are characterized by the zinc-binding consensus se-quence HEXXHXXGXXHZ (residues 169 to 180 in PAP),where the histidine residues are the zinc ligands and the Zresidue is characteristic for the different subfamilies (in thecase of the serralysins, Z � Pro), and by a conserved methio-nine residue (Met-207 in PAP). Although the sequence iden-tity between the different subfamilies of the metzincins is verylow, their catalytic domains do have similar three-dimensional(3D) structures (2–4, 6, 7, 19–21).
The serralysins are secreted as inactive zymogens into themedium via the hemolysin pathway, involving an ABC trans-porter, which recognizes a C-terminal secretion signal. Activa-tion occurs in the medium autocatalytically in the presence ofdivalent ions, especially Zn2� and Ca2� (12–14).
The implication of zinc metalloproteases in many diseases,e.g., cancer or arthrosis (15), makes structural studies of theseenzymes highly interesting.
Here we report the results from X-ray crystallographic stud-ies of PAP, an alkaline protease produced by the psychrophilicPseudomonas sp. strain TAC II 18, which originated in Ant-
arctica. This strain, identified as a gram-negative bacterium(18), is one of numerous bacteria living at temperatures closeto 0°C. To survive and grow under such conditions, theseorganisms have evolved several adaptations of their cellularcomponents, especially of their enzymes (16–18, 30). Thus,PAP is three times more active at 20°C than a mesophiliccounterpart from Pseudomonas aeruginosa but rapidly inacti-vated at 45°C. The cold-adapted enzyme also displays a greatersensitivity to moderate EDTA concentrations, and fluores-cence studies have indicated enzyme unfolding after 2 h ofincubation in 2 mM EDTA (11).
The PAP molecule is made up of 463 amino acid residuesdistributed in two domains, and its tertiary structure is com-parable to mesophilic homologues from P. aeruginosa alkalineprotease (AP) and Serratia marcescens protease (SMP) (1–3).Four �-helices and a mixed five-stranded �-sheet form thecatalytic N-terminal domain to which the active-site Zn2� ionand a Ca2� ion (Ca 700, PAP numbering) are bound. ThisCa2� ion is not present in its mesophilic counterparts, AP andSMP, but it should be noticed that a calcium ion has beenobserved in the 3D structure of the metzincin adamalysin II ina position similar to that of Ca 700 (20).
The C-terminal domain which may play a role in the foldingof the molecule after transmembrane translocation forms anextended � roll with 20 successive �-strands wound in a right-handed spiral. The number of Ca2� ions bound to this C-terminal domain varies from six to seven in the different or-ganisms and crystal forms.
When we compared the 3D structure with those of the me-sophilic counterparts, AP and SMP, the most remarkable dif-ferences were found in the proteolytic domain. First, the activesite is more accessible due, in part, to deletions in surrounding
* Corresponding author. Mailing address: Laboratoire de BioCris-tallographie, Institut de Biologie et Chimie des Proteines, UMR 5086,CNRS-UCBL1, 7 Passage du Vercors, 69367 Lyon, Cedex 07, France.Phone: 33(0)472722608. Fax: 33(0)472722616. E-mail: [email protected].
4195
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

loops and to a region in the catalytic domain of PAP (residues107 to 116) that undergoes a conformational change with aloop movement as large as 13 A induced by the binding of theextra calcium ion, Ca 700. This change does not seem to berelated to crystal packing effects, since it was observed in crys-tal forms obtained under very different crystallization condi-tions. The role of this extra calcium ion in the activity orfolding of the PAP molecule still needs to be elucidated. Itsweak chelation in the proteolytic domain and its close proxim-ity to the solvent could partly explain the sensitivity of PAP toEDTA, which is also supported by a higher temperature factorfor this calcium ion compared to other calcium ions in the 3Dstructure. This sensitivity could also be related to the loweraffinity, compared to AP and SMP, for some calcium ionslocated in the C-terminal domain.
In order to address these questions, we solved the structuresof PAP (form 2) in the presence of various concentrations ofEDTA, as well as a new native form (form 3) by X-ray crys-tallography.
MATERIALS AND METHODS
Crystallization. PAP has been crystallized under three different conditions—form 1 (31), form 2 (1), and form 3 [0.2 M (NH4)2SO4, 22% PEG 4000, and 0.1M sodium acetate buffer at pH 4.6 at 16°C]—by the hanging-drop vapor diffusionmethod.
Cocrystallization experiments of PAP have been performed under conditionsderived from form 2 [1.6 M (NH4)2SO4 and 0.1 M HEPES (pH 7) at 16°C].EDTA was added to a final concentration of 1 mM (experiment A), 5 mM(experiment B), and 10 mM (experiment C) in the drops. Crystals grew within 1week to a final size of about 300 by 100 by 100 �m and were left another 2 monthsin the drops before data collection in experiments A and C. In experiment B thecrystals were left an additional 5 days (B1) or 2 months (B2) in the droplets,respectively, before data collection.
Soaking was achieved by directly adding a solution of EDTA to a final con-centration of 85 mM into a drop containing form 2 crystals and leaving it 4 h at16°C (experiment D). Higher concentrations of EDTA have been tested, but thecrystals cracked at concentrations of EDTA greater than 85 mM.
Cocrystallization or soaking with EDTA did not modify the space group, butsmall variations in the unit cell dimensions were observed (Table 2); these werecertainly due in part to flash freezing of the crystals. The large difference in unitcell dimensions between B1 and the remaining data is a consequence of the B1data being collected at 288 K and the rest being collected at 100 K.
Data collection, structure solution, and refinement. (i) Native form. Diffrac-tion data on form 3 crystals (Table 1) were collected with a charge-coupleddevice camera by using synchrotron radiation at the beam-line D2AM at theEuropean Synchrotron Radiation Facility (ESRF; Grenoble, France) undercryogenic conditions (100 K). Initial data reduction and space group determina-tion were carried out with the program XDS (23), and further reduction of thedata was performed by using programs from the CCP4 package (10).
The known structure of the AP (3) served as model in a molecular replace-ment search performed with the program AMoRe (26). Refinement of thestructure was done by using the program CNS (9), applying an overall anisotropictemperature factor correction algorithm, followed by manual fitting. A summaryof the refinement statistics can be found in Table 1.
Due to poor or missing electron densities, as for form 2 (1), residues 1 to 2 andresidues 184 to 188 were not inserted into the electron density map.
(ii) Experiments A, B, C, and D. X-ray diffraction data of crystals fromexperiments A, B2, and C were collected at the BM30A beamline at the ESRFat cryogenic temperatures (100 K) at a wavelength of 0.98 A on a MarCCD.X-ray data on crystals from experiments B1 and D were collected in house at 288K and at cryogenic temperatures, respectively, on a MARresearch 345 imageplate system associated with a Nonius FR591 rotating anode (CuK� radiation)operating at 44 kV and 100 mA and coupled to Osmic confocal mirrors.
Diffracted intensities in experiments A, B2, C, and D were integrated with theprogram MOSFLM (25), as implemented in the CCP4 software package (10) andscaled with SCALA (10), whereas experiment B1 reflection data were processedand merged with DENZO/SCALEPACK (27).
Collection and processing statistics of experiments A, B, C, and D are sum-
marized in Table 2, and structures resulting from these experiments will hence-forward be referred to as structure A, B1, B2, C, and D, respectively. StructuresB1 and B2 were solved by the molecular replacement method by using theAMoRe software as implemented in the CCP4 suite (10), and the native form 2was resolved to a 1.96-A resolution (1) as a search model. Withdrawing diffrac-tion data in a resolution range of 15 to 4 A, a unique solution was obtained witha correlation coefficient of 58.8% and an R factor of 37.5% for B1 and acorrelation coefficient of 54.0% and an R factor of 37.5% for B2.
Since the crystals from experiments A, C, and D were isomorphous with B2crystals, their 3D structures were determined by direct phasing with the 2.0-Aresolution B2 structure, for which ions as well as water molecules in the activesite had been removed in order to avoid bias. The refinement of all models wasdone by using the simulated annealing protocol as implemented in the softwarepackage CNS (9), alternately with visual examination of electron density mapsand manual building by using the graphic software TURBO-FRODO (29).
Water molecules present at similar positions in the respective structures havethe same numbering.
In order to confirm the presence or absence of calcium or zinc ions, anomalousdifference Fourier maps were calculated by using the program CNS, in whichcalculated phases are derived from a refined model depleted of metal ions.
For all data set, free and conventional R factors were monitored (8) to avoidoverrefinement.
Compared to the native structure of form 2 (PDB entry 1G9K), residues notinserted in the electron density map due to poor or missing electron densitieswere residues 50 to 51 in structures A, B2, C, and D and Tyr-183 in all fivestructures. Moreover side chain residues for Asn-52, Asp-53, and Asn-184 werenot observed in electron density maps of structures A, B2, C, and D. Double
TABLE 1. Data collection and refinement statistics for crystal form3 of PAP
Statisticsa Valueb
Data statisticsWavelength (Å) ................................... 1.07Data collection temp (K) ................... 100Space group ........................................ R3Unit cell parameters ...........................a � b � 184.6 Å, c � 37.9 Å;
� � � � 90°, � � 120°Resolution range (Å).......................... 13.6–2.38Completeness of data (%) ................. 93.2 (59.0)Multiplicity of data.............................. 1.8 (1.4)Total no. of reflections ....................... 32,163No. of unique reflections.................... 17,980Rsym (%) ............................................... 4.0 (12.8)I/�(I) ..................................................... 15.5 (7.8)
Refinement statisticsR, factor (%)........................................ 19.5Rfree (%) ............................................... 24.6Resolution range (Å).......................... 50.0–2.38rmsd from ideal geometry..................
Bond length (Å) .............................. 0.0090Bond angle (°) ................................. 1.44
Ramachandran plot—residues in:Most favored regions (%).............. 90.1Additionally allowed regions (%) . 9.9
No. of protein atoms .......................... 3,371No. of water molecules....................... 359No. of zinc ions ................................... 1No. of calcium ions ............................. 8No. of sulfate ions............................... 1
a Rsym ��hkl�t
�Ihkli � �Ihkl��
�hkl�iIhkl
; R � *�h
�Fobsh� � k�Fcalch�
�h�Fobsh�
, where k is
a scaling factor; Rfree ��h
�Fobsh��k�Fcalch�
�h�Fobsh�
, where k is a scaling factor.
Values were calculated from a test set in which 10% of the total number ofreflections has been randomly selected.
b Values in parentheses are for highest-resolution shell.
4196 RAVAUD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

TA
BL
E2.
Data
collectionand
refinement
statisticsfor
experiments
A,B
1,B2,C
,andD
Condition
orstatistic
aE
xpt b
AB
1B
2C
D
Experim
entalconditionsC
ocrystallizationor
soakingc
CO
CO
CO
CO
SOE
DT
Aconc
(mM
)1
55
1085
Tim
eleft
indroplets
2m
o12
days2
mo
2m
o4
h
Data
statisticsW
avelength(Å
)0.9800
1.54180.9800
0.98001.5418
Data
collectiontem
p100
K288
K100
K100
K100
KSpace
groupR
3R
3R
3R
3R
3U
nitcellparam
etersa
�b
�179.9
Å,c
�37.3
Å;
��
��
90°,�
�120°
a�
b�
185.7Å
,c�
38.0Å
;�
��
�90°,
��
120°a
�b
�180.5
Å,c
�37.3
Å;
��
��
90°,�
�120°
a�
b�
181.7Å
,c�
37.5Å
;�
��
�90°,
��
120°a
�b
�182.9
Å,c
�37.6
Å;
��
��
90°,�
�120°
Resolution
range(Å
)25.9–2.2
32.3–2.534.1–2.0
27.1–2.034.5–2.8
Com
pleteness(%
)99.7
(99.9)99.1
(98.3)100.0
(100.0)99.8
(99.2)99.5
(99.3)M
ultiplicity3.6
(3.3)2.0
(2.0)4.0
(3.9)3.8
(3.6)2.0
(2.0)T
otalno.ofreflections
81,35533,198
123,884115,986
22,739N
o.ofunique
reflections22,772
16,74930,649
30,89711,326
Rsym
(%)
9.9(24.1)
11.6(37.4)
8.1(16.7)
9.1(19.5)
14.7(24.9)
I/�(I)
3.2(1.1)
5.6(1.8)
4.9(3.3)
3.6(1.8)
2.1(2.7)
Refinem
entstatistics
Rfacto
r(%
)18.9
16.718.5
17.723.6
Rfree
(%)
23.721.8
22.920.8
30.3R
esolutionrange
(Å)
50.0–2.250.0–2.5
50.0–2.050.0–2.0
50–2.8rm
sdfrom
idealgeometry
Bond
lengths(Å
)0.009
0.00960.0105
0.01040.0106
Bond
angles(°)
1.561.49
1.621.59
1.57R
amachandran—
residuesin:
Most
favoredregions
(%)
91.688.2
90.291.8
86.8A
dditionallyallow
edregions
(%)
8.411.8
9.88.2
13.2N
o.ofprotein
atoms
3,3323,357
3,3223,327
3,330N
o.ofw
aterm
olecules367
161379
369185
No.of
zincions
00
00
0N
o.ofcalcium
ions6
76
66
No.of
sulphateions
11
11
1
aSee
Table
1,footnotea.
bSee
Table
1,footnoteb.
cCO
,cocrystallization;SO,soaking.
VOL. 185, 2003 CRYSTAL STRUCTURES OF A PSYCHROPHILIC METALLOPROTEASE 4197
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

conformations for Ser-15, Ser-138, and Ser-332 were found in structure B2.Model qualities were examined with PROCHECK (24) and WHATCHECK(22). The details of the refinement statistics and model quality are given in Table2.
Relative B-factors were calculated by dividing the mean B-factor of everyresidue by the mean B-factor of the whole protein. Due to the rather poor qualityof structure D data, B-factors where initially set to 20 A2 and left out of therefinement.
Superimposition of structures was done by using the “rigid” option in TUR-BO-FRODO. The figures shown here were rendered by using TURBO-FRODOand VIEWERLITE 5.0 (freeware from Accelrys, Inc., San Diego, Calif.).
Coordinates. The coordinates and structure factors are deposited in the theprotein data bank as 1O0Q (A), 1O0T (B1), 1OM6 (B2), 1OM8 (C), 1OM7 (D),and 1OMJ (form 3).
RESULTS
Native form 3. In the crystal structure of form 3, 456 aminoacid residues, one Zn2� ion, and 8 Ca2� ions could be locatedin the electron density map. The superposition of 3D struc-tures of forms 2 and 3 using all C� atoms gives a root meansquare deviation (rmsd) of 0.19 A and thus two nearly identicalstructures, despite quite different crystallization conditions.The main difference between the two forms is an additionalcalcium-binding site in the C-terminal domain of form 3, whichwas observed in form 1 (1), as well as a loop region around Ca700 that seems more flexible in form 3, as judged from thediscontinuous electron density. Another difference is the pres-ence of a continuous electron density in the active site of form3 that can be interpreted as being either a peptide or a networkof mobile water molecules.
Effect of EDTA on ion binding. (i) Overall structures. Com-parative studies of native PAP (Fig. 1) with structures A, B1,B2, C, and D show that the overall 3D structures are verysimilar. The rmsd values between C�’s in these five structuresand the native form 2 of PAP were 0.42, 0.17. 0.43, 0.41, and0.37 Å, respectively. No density corresponding to a molecule ofEDTA can be seen in any of the calculated electron densitymaps. Significant differences, however, were observed in thenumber of metal ions ligated in the structures (Table 2). Infact, B1 reveals the structure of the apo-form of PAP, since theelectron density map calculated on the basis of the 2.5-A dataclearly showed that the active-site zinc ion had disappeared but
that all of the calcium ions were present relative to the nativestructure, as confirmed by an anomalous difference Fouriermap.
The electron density maps calculated for structures A, B2, C,and D clearly showed that the catalytic zinc ion and the calciumion located in the catalytic domain (Ca 700) were lacking.However, in the C-terminal domain all calcium ions could belocated and were inserted in the electron density of structuresA, B2 and C. In structure D, one calcium ion, Ca 702, isreplaced by a water molecule.
(ii) Changes in the Zn(II)-binding pocket. In the active siteof native PAP, the Zn2� ion is coordinated in a trigonal-bipyramidal geometry to four or five ligands depending on theposition of the Tyr-209. The Zn2�, when penta-coordinated, isbound to the side chains of three histidines (His-169, His-173,and His-179), Tyr-209, and a water molecule (1).
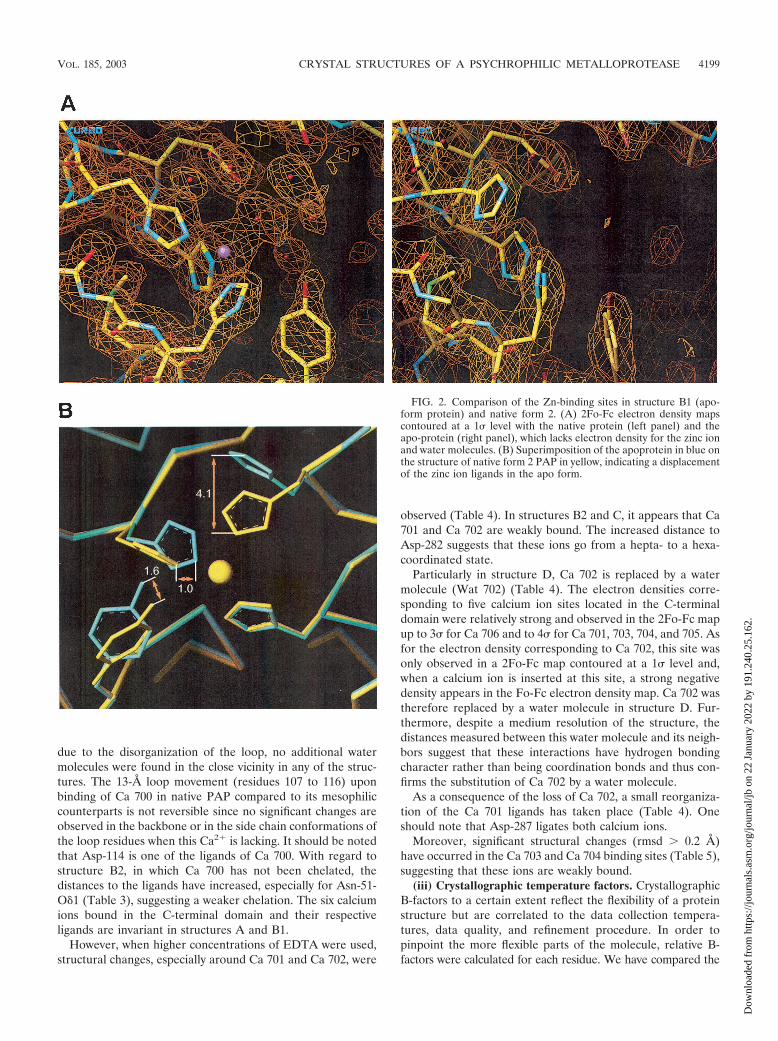
Electron density maps calculated for experiments A, B1, B2,C, and D, clearly show that no metal is present in the active-site region and that no additional water molecule replaces thezinc ion in the binding site (Fig. 2). As a consequence, dis-placements in the side chains of the zinc-ion ligands are ob-served, including His-173 and His-179. In the apo-form, theNε2 from His-173 undergoes a 4.1-A shift and the imidazolering of His-179 rotates 40° around its C�-C�, leading to a 1.0-Adisplacement of its Nε2. Similar changes for these histidinesare observed in the other structures. Upon removal of thecatalytic zinc ion, the phenol ring of Tyr-209 rotates ca. 60°around the C�-C� bond. The side chain of His-169 does notundergo significant conformational changes. Interestingly, amajor part of the water molecules constituting an organizednetwork in native PAP have disappeared, including Wat 1225,which ligates the zinc ion in the holo form (Fig. 2).
Excepting the network of water molecules, similar significantstructural changes (rmsd � 0.2 A) have been noticed in struc-tures A, B2, C, and D. In structure A, Wat 1225, 1102, and1104 are present, whereas Wat 1102, 1104, and 1304 are foundin the catalytic site of structure B2, and Wat 1102 and 1134 arefound in structures C and D.
(iii) Calcium-binding sites. Of the seven calcium ions thatare bound in the native form 2, Ca 700 is found in the proteo-lytic domain where it is easily accessible to the solvent, and theothers are found in the C-terminal domain (Fig. 1).
Next to the proteolytic domain, Ca 701 and 702 are hepta-coordinated, and the four remaining calcium ions (Ca 703 toCa 706) are bound in the core of the parallel �-roll in ahexacoordinated manner. These are supposed to be essentialdeterminants of the parallel �-roll structure (1, 3).
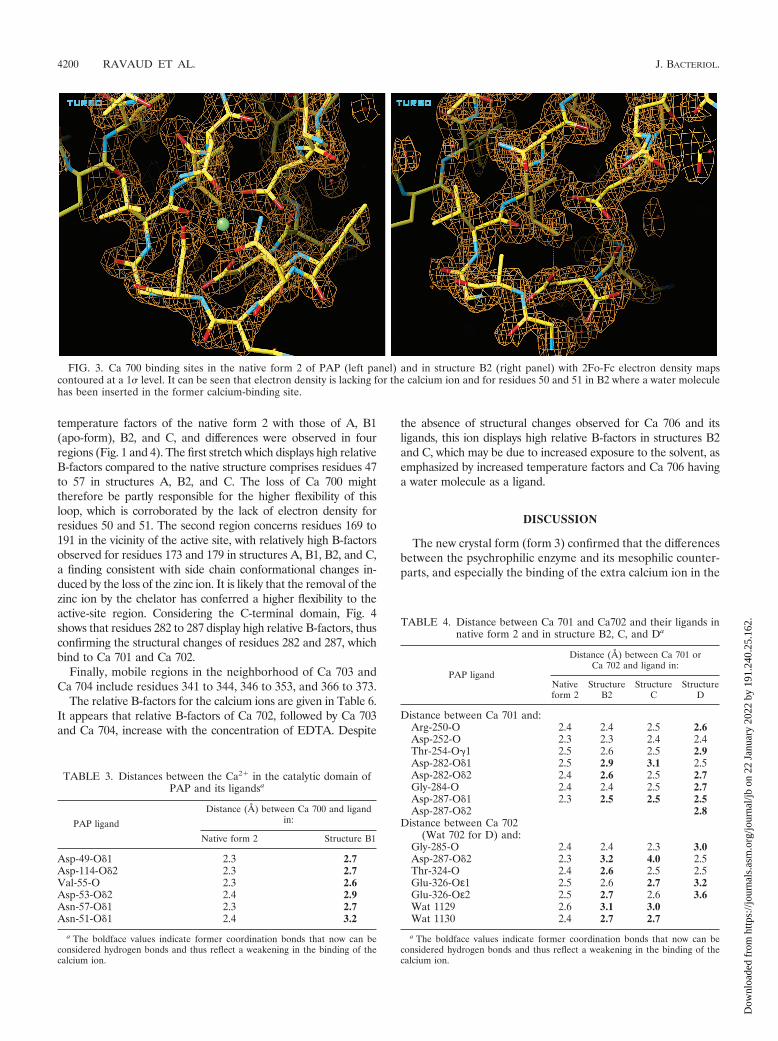
The loss of Ca 700 from the proteolytic domain in structuresA, B2, C, and D induces structural changes, including that ofAsn-57, which moves 1.5 A toward the former calcium posi-tion. Moreover, 2Fo-Fc electron density was lacking for resi-dues 50 and 51 and for the side chains of residues 52 and 53,all of which are located in the loop surrounding the Ca 700binding site (Fig. 3). The predominant change caused byEDTA chelating Ca 700 is a partial disorganization of thisloop.
From the highest-resolution structures (B2 and C at 2.0 A),it can be concluded that a water molecule replaces the calciumion; hence, Wat 1500 has been inserted (Fig. 3).
Although the Ca 700 binding site is accessible to the solvent
FIG. 1. Global view of native PAP form 2. Regions of interest forrelative B-factors are colored as a function of the degree of flexibility:yellow (residues 47 to 57) and cyan (residues 169 to 191) indicate areasthat undergo major changes upon EDTA ligation, dark blue represents“middle-flexibility” zones, and pink (region from residues 320 to 380)indicates the most rigid part of the molecule.
4198 RAVAUD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.
due to the disorganization of the loop, no additional watermolecules were found in the close vicinity in any of the struc-tures. The 13-A loop movement (residues 107 to 116) uponbinding of Ca 700 in native PAP compared to its mesophiliccounterparts is not reversible since no significant changes areobserved in the backbone or in the side chain conformations ofthe loop residues when this Ca2� is lacking. It should be notedthat Asp-114 is one of the ligands of Ca 700. With regard tostructure B2, in which Ca 700 has not been chelated, thedistances to the ligands have increased, especially for Asn-51-O 1 (Table 3), suggesting a weaker chelation. The six calciumions bound in the C-terminal domain and their respectiveligands are invariant in structures A and B1.
However, when higher concentrations of EDTA were used,structural changes, especially around Ca 701 and Ca 702, were
observed (Table 4). In structures B2 and C, it appears that Ca701 and Ca 702 are weakly bound. The increased distance toAsp-282 suggests that these ions go from a hepta- to a hexa-coordinated state.
Particularly in structure D, Ca 702 is replaced by a watermolecule (Wat 702) (Table 4). The electron densities corre-sponding to five calcium ion sites located in the C-terminaldomain were relatively strong and observed in the 2Fo-Fc mapup to 3� for Ca 706 and to 4� for Ca 701, 703, 704, and 705. Asfor the electron density corresponding to Ca 702, this site wasonly observed in a 2Fo-Fc map contoured at a 1� level and,when a calcium ion is inserted at this site, a strong negativedensity appears in the Fo-Fc electron density map. Ca 702 wastherefore replaced by a water molecule in structure D. Fur-thermore, despite a medium resolution of the structure, thedistances measured between this water molecule and its neigh-bors suggest that these interactions have hydrogen bondingcharacter rather than being coordination bonds and thus con-firms the substitution of Ca 702 by a water molecule.
As a consequence of the loss of Ca 702, a small reorganiza-tion of the Ca 701 ligands has taken place (Table 4). Oneshould note that Asp-287 ligates both calcium ions.
Moreover, significant structural changes (rmsd � 0.2 A)have occurred in the Ca 703 and Ca 704 binding sites (Table 5),suggesting that these ions are weakly bound.
(iii) Crystallographic temperature factors. CrystallographicB-factors to a certain extent reflect the flexibility of a proteinstructure but are correlated to the data collection tempera-tures, data quality, and refinement procedure. In order topinpoint the more flexible parts of the molecule, relative B-factors were calculated for each residue. We have compared the
FIG. 2. Comparison of the Zn-binding sites in structure B1 (apo-form protein) and native form 2. (A) 2Fo-Fc electron density mapscontoured at a 1� level with the native protein (left panel) and theapo-protein (right panel), which lacks electron density for the zinc ionand water molecules. (B) Superimposition of the apoprotein in blue onthe structure of native form 2 PAP in yellow, indicating a displacementof the zinc ion ligands in the apo form.
VOL. 185, 2003 CRYSTAL STRUCTURES OF A PSYCHROPHILIC METALLOPROTEASE 4199
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.
temperature factors of the native form 2 with those of A, B1(apo-form), B2, and C, and differences were observed in fourregions (Fig. 1 and 4). The first stretch which displays high relativeB-factors compared to the native structure comprises residues 47to 57 in structures A, B2, and C. The loss of Ca 700 mighttherefore be partly responsible for the higher flexibility of thisloop, which is corroborated by the lack of electron density forresidues 50 and 51. The second region concerns residues 169 to191 in the vicinity of the active site, with relatively high B-factorsobserved for residues 173 and 179 in structures A, B1, B2, and C,a finding consistent with side chain conformational changes in-duced by the loss of the zinc ion. It is likely that the removal of thezinc ion by the chelator has conferred a higher flexibility to theactive-site region. Considering the C-terminal domain, Fig. 4shows that residues 282 to 287 display high relative B-factors, thusconfirming the structural changes of residues 282 and 287, whichbind to Ca 701 and Ca 702.
Finally, mobile regions in the neighborhood of Ca 703 andCa 704 include residues 341 to 344, 346 to 353, and 366 to 373.
The relative B-factors for the calcium ions are given in Table 6.It appears that relative B-factors of Ca 702, followed by Ca 703and Ca 704, increase with the concentration of EDTA. Despite
the absence of structural changes observed for Ca 706 and itsligands, this ion displays high relative B-factors in structures B2and C, which may be due to increased exposure to the solvent, asemphasized by increased temperature factors and Ca 706 havinga water molecule as a ligand.
DISCUSSION
The new crystal form (form 3) confirmed that the differencesbetween the psychrophilic enzyme and its mesophilic counter-parts, and especially the binding of the extra calcium ion in the
FIG. 3. Ca 700 binding sites in the native form 2 of PAP (left panel) and in structure B2 (right panel) with 2Fo-Fc electron density mapscontoured at a 1� level. It can be seen that electron density is lacking for the calcium ion and for residues 50 and 51 in B2 where a water moleculehas been inserted in the former calcium-binding site.
TABLE 3. Distances between the Ca2� in the catalytic domain ofPAP and its ligandsa
PAP ligand
Distance (Å) between Ca 700 and ligandin:
Native form 2 Structure B1
Asp-49-O 1 2.3 2.7Asp-114-O 2 2.3 2.7Val-55-O 2.3 2.6Asp-53-O 2 2.4 2.9Asn-57-O 1 2.3 2.7Asn-51-O 1 2.4 3.2
a The boldface values indicate former coordination bonds that now can beconsidered hydrogen bonds and thus reflect a weakening in the binding of thecalcium ion.
TABLE 4. Distance between Ca 701 and Ca702 and their ligands innative form 2 and in structure B2, C, and Da
PAP ligand
Distance (Å) between Ca 701 orCa 702 and ligand in:
Nativeform 2
StructureB2
StructureC
StructureD
Distance between Ca 701 and:Arg-250-O 2.4 2.4 2.5 2.6Asp-252-O 2.3 2.3 2.4 2.4Thr-254-O�1 2.5 2.6 2.5 2.9Asp-282-O 1 2.5 2.9 3.1 2.5Asp-282-O 2 2.4 2.6 2.5 2.7Gly-284-O 2.4 2.4 2.5 2.7Asp-287-O 1 2.3 2.5 2.5 2.5Asp-287-O 2 2.8
Distance between Ca 702(Wat 702 for D) and:
Gly-285-O 2.4 2.4 2.3 3.0Asp-287-O 2 2.3 3.2 4.0 2.5Thr-324-O 2.4 2.6 2.5 2.5Glu-326-Oε1 2.5 2.6 2.7 3.2Glu-326-Oε2 2.5 2.7 2.6 3.6Wat 1129 2.6 3.1 3.0Wat 1130 2.4 2.7 2.7
a The boldface values indicate former coordination bonds that now can beconsidered hydrogen bonds and thus reflect a weakening in the binding of thecalcium ion.
4200 RAVAUD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

proteolytic domain, are independent of crystallization condi-tions or crystal packing.
Experiments A to D contribute to elucidate the role of di-valent metal ions in PAP. In light of the conditions of exper-iments B1 and B2 (contact times with EDTA of 12 days and 2months, respectively), it clearly appears that although the zincion was more strongly coordinated than Ca 700 (1), it was the
first to be chelated by EDTA. Moreover, the complexationconstant of Zn2� by EDTA is weaker than for Ca2� (1010.7 and1016.3, respectively, at 25°C).
Notwithstanding chelation of Ca 700 by EDTA appears fa-vorable from a thermodynamic point of view, our experimentssuggest that the action of EDTA on PAP predominantly iscontrolled by kinetics in agreement with studies on the �-rollperformed by Rose et al. (28). Thus, in order to explain theincreased sensitivity of PAP compared to mesophilic counter-parts, kinetic parameters such as steric hindrance should beconsidered with the increased accessibility to the active site inPAP as the key factor.
Our studies of all calcium-binding sites in structures A, B1,B2, C, and D and in the native form 2 of PAP have led us to
FIG. 4. Relative B-factors for native form 2 and structures A, B1, B2, and C, indicating the four regions displaying major changes: residues 47to 57, 169 to 191, 250 to 290, and 320 to 380. Gaps in the curves correspond to residues for which electron density was lacking.
TABLE 5. Distances between the Ca 703 and Ca 704 and theirligands in native form 2 and in structure Da
PAP ligand
Distance (Å) between Ca 703 orCa 704 and ligand in:
Native form 2 Structure D
Distance between Ca 703 and:Gly-331-O 2.4 2.6Gly-333-O 2.3 2.5Asp-335-O 2 2.4 2.9Gly-348-O 2.3 2.5Ala-350-O 2.4 2.5Asp-353-O 1 2.4 2.6
Distance between Ca 704 and:Gly-349-O 2.3 4.3Gly-351-O 2.4 2.5Asp-353-O 2 2.6 2.9Gly-366-O 2.3 2.7Thr-368-O 2.4 2.8Asp-371-O 1 2.4 2.7
a See Table 4, footnote a.
TABLE 6. Relative B-factors for calcium ions
Calcium ion
% B-factor in:
Nativeform 2
StructureA
StructureB1
StructureB2
StructureC
Ca 700 159 162Ca 701 83 111.5 149 151 168Ca 702 97 112 116 367 359Ca 703 86 119 108 170 235Ca 704 103 142 123 161 203Ca 705 97.5 117 102 123 133.5Ca 706 101 164 146 216 215
VOL. 185, 2003 CRYSTAL STRUCTURES OF A PSYCHROPHILIC METALLOPROTEASE 4201
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

analyze their different roles in the global stability of the mol-ecule and their potential implication in the folding of PAP.
Structures B1 and B2 indicate that Ca 700 is present duringthe crystallization process and hereby when crystals are formedand is only chelated by EDTA after crystal formation. One canspeculate that if this site is not occupied, the molecule is tooflexible for crystal formation. It should also be stressed that themajor part of the intermolecular interactions to the proteolyticdomain is performed with residues from the region surround-ing this calcium ion. These findings, taken together with thefact that the 13-A loop movement upon binding of Ca 700 isnonreversible upon chelation in structures A, B2, C, and D, areindeed in agreement with an increased accessibility to theactive site, caused by this same loop movement, being a keyfactor of the adaptation of this enzyme to cold environments.Thus, Ca 700 does not appear to improve the global stability ofPAP, but rather the stability of the proteolytic domain asstrengthened by the partial disorganization of the loop sur-rounding the depleted calcium site.
On the basis of these findings, we have proposed a classifi-cation of calcium ions from the least to the most tightly bound.After 2 months, a 1 mM concentration of EDTA appears to besufficient for chelating the calcium ion located in the N-termi-nal proteolytic domain (Ca 700). When the concentration ofEDTA was increased to 5 mM, ligands of Ca 702 move awayfrom the calcium-binding site. This weakly bound calcium ionalso displays a high B-factor and is thought to be partiallyoccupied, especially in structure C. This effect increases withthe concentration of EDTA and, at 85 mM, Ca 702 seems to bereplaced by a water molecule. One ligand (Asp-287) is sharedby Ca 702 and Ca 701. When the Ca 702 binding site is mod-ified, structural changes in the Ca 701 binding site are concom-itant. After 2 months in 5 mM EDTA, the relative B-factor ofCa 706 increases by 60%, indicating a partial occupation of thissite.
Ca 703 and Ca 704 in turn seem to be affected when theconcentration of EDTA was increased to 10 mM (structure C),as indicated by their high relative B-factors. Structure D, forwhich conformational movements among the ligands of Ca 703and Ca 704 have been observed, confirms this observation.
As for the C-terminal domain, it can be concluded that Ca702 and hereafter Ca 701 are the less tightly bound, followedby Ca 706, Ca 703, and Ca 704. Finally, Ca 705 emerges as themost tightly bound ion since its binding site undergoes nosignificant changes in any of the experiments.
This classification is coherent with coordination states andspatial positions in the structures. Ca 701 and Ca 702, whichare located in one extremity of the C-terminal domain and nextto the catalytic domain, are relatively accessible to the solvent.Although Ca 706 is bound more internally in the central regionof the �-roll, this ion had one water molecule as ligand andthen was more exposed to the solvent. Finally, the last threecalcium ions (Ca 703 to 705) seem to hold the structure to-gether, thus confirming their putative implication in the foldingof the molecule after transmembrane translocation.
Our studies confirm that PAP is sensitive to low concentra-tions of EDTA, as described previously (11), since 1 mMEDTA in experiment A and 5 mM EDTA in experiment B1appear to be sufficient for chelating the zinc ion in the N-
terminal proteolytic domain and hence to inactivate the pro-tein.
However, our results do not explain previous fluorescencestudies that indicated enzyme unfolding after 2 h of incubationin 2 mM EDTA (11) but, because the experiments describedhere take place within a crystal, direct comparisons are notpossible.
From an overall structural point of view, relative exposure ofsome calcium ions located in the C-terminal domain to thesolvent seems to allow a higher conformational flexibility dueto the lack of protein-protein interactions by the bias of crystalcontacts.
The rigid nature of the C-terminal domain, which probablyplays an important role in the folding of the molecule, and thefact that Ca 703 to Ca 705 are essential determinants of theparallel �-roll structure (3), a finding supported by the presentstudy, promote restrictions in conformational changes withinthis domain. This can probably explain why the 3D structure ofPAP remains folded in high concentrations of EDTA (up to 85mM as found in the soaking experiments). Possibly, the che-lation of Ca 705 would result in the unfolding of the protein,which in turn may explain why form 2 crystals cracked in thesoaking experiments when the EDTA concentrations were be-yond 85 mM.
ACKNOWLEDGMENTS
This work was supported by the EU contracts BI04-CT96-0051 andBI04-CT97-0131, the Centre National d’Etudes Spatiales, and theCentre National de la Recherche Scientifique.
We thank C. Gerday for providing the enzyme for the crystallo-graphic studies and R. Kahn for assistance with data collection at theESRF.
REFERENCES
1. Aghajari, N., F. Van Petegem, V. Villeret, J. P. Chessa, C. Gerday, R. Haser,and J. Van Beeumen. 2003. Crystal structures of a psychrophilic metallopro-tease reveal new insights into catalysis by cold-adapted proteases. ProteinsStruct. Funct. Genet. 50:636–647.
2. Baumann, U. 1994. Crystal structure of the 50-kDa metalloprotease fromSerratia marcescens. J. Mol. Biol. 242:244–251.
3. Baumann, U., S. Wu, K. M. Flaherty, and D. B. McKay. 1993. Three-dimensional structure of the alkaline protease of Pseudomonas aeruginosa: atwo-domain protein with a calcium binding parallel beta roll motif. EMBOJ. 12:3357–3364.
4. Bode, W., F. X. Gomis-Ruth, R. Huber, R. Zwilling, and W. Stocker. 1992.Structure of astacin and implications for activation of astacins and zinc-ligation of collagenases. Nature 358:164–167.
5. Bode, W., F. X. Gomis-Ruth, and W. Stockler. 1993. Astacins, serralysins,snake venom and matrix metalloproteinases exhibit identical zinc-bindingenvironments (HEXXHXXGXXH and Met-turn) and topologies andshould be grouped into a common family, the “metzincins.” FEBS Lett.331:134–140.
6. Bode, W., P. Reinemer, R. Huber, T. Kleine, S. Schnierer, and H. Tschesche.1994. The X-ray crystal structure of the catalytic domain of human neutro-phil collagenase inhibited by a substrate analogue reveals the essentials forcatalysis and specificity. EMBO J. 13:1263–1269.
7. Borkakoti, N., F. K. Winkler, D. H. Williams, A. D’Arcy, M. J. Broadhurst,P. A. Brown, W. H. Johnson, and E. J. Murray. 1994. Structure of thecatalytic domain of human fibroblast collagenase complexed with an inhib-itor. Nat. Struct. Biol. 1:106–110.
8. Brunger, A. T. 1992. Free R value: a novel statistical quantity for assessingthe accuracy of crystal structures. Nature 355:472–475.
9. Brunger, A. T., P. D. Adams, G. M. Clore, W. L. DeLano, P. Gros, R. W.Grosse-Kunstleve, J. S. Jiang, J. Kuszewski, M. Nilges, N. S. Pannu, R. J.Read, L. M. Rice, T. Simonson, and G. L. Warren. 1998. Crystallography andNMR system: a new software suite for macromolecular structure determi-nation. Acta Crystallogr. D54:905–921.
10. CCP4 Collaborative Computational Project. 1994. The CCP4 suite: pro-grams for protein crystallography. Acta Crystallogr. D50:760–763.
11. Chessa, J. P., I. Petrescu, M. Bentahir, J. Van Beeumen, and C. Gerday.2000. Purification, physico-chemical characterization and sequence of a heat
4202 RAVAUD ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.

labile alkaline metalloprotease isolated from a psychrophilic Pseudomonasspecies. Biochim. Biophys. Acta 1479:265–274.
12. Delepelaire, P. 1994. PrtD, the integral membrane ATP-binding cassettecomponent of the Erwinia chrysanthemi metalloprotease secretion system,exhibits a secretion signal-regulated ATPase activity. J. Biol. Chem. 269:27952–27957.
13. Delepelaire, P., and C. Wandersman. 1989. Protease secretion by Erwiniachrysanthemi: proteases B and C are synthesized and secreted as zymogenswithout a signal peptide. J. Biol. Chem. 264:9083–9089.
14. Delepelaire, P., and C. Wandersman. 1990. Protein secretion in gram-neg-ative bacteria. The extracellular metalloprotease B from Erwinia chrysan-themi contains a C-terminal secretion signal analogous to that of Escherichiacoli alpha-hemolysin. J. Biol. Chem. 265:17118–17125.
15. Dive, V., M. Kaczorek, C. Roussel, and F. Roux. 1997. Les nouveaux inhibi-teurs des metalloproteinases a zinc. Biofutur 167:29–33.
16. Feller, G., and C. Gerday. 1997. Psychrophilic enzymes: molecular basis ofcold adaptation. Cell. Mol. Life Sci. 53:830–841.
17. Feller, G., E. Narinx, J. L. Arpigny, M. Aittaleb, E. Baise, S. Genicots, andC. Gerday. 1996. Enzymes from psychrophilic organisms. FEMS Microbiol.Rev. 18:189–202.
18. Gerday, C., M. Aittaleb, J. L. Arpigny, E. Baise, J. P. Chessa, G. Garsoux, I.Petrescu, and G. Feller. 1997. Psychrophilic enzymes: a thermodynamicchallenge. Biochim. Biophys. Acta 1342:119–131.
19. Gomis-Ruth, F. X., L. F. Kress, and W. Bode. 1993. First structure of a snakevenom metalloproteinase: a prototype for matrix metalloproteinases/collag-enases. EMBO J. 12:4151–4157.
20. Gomis-Ruth, F. X., L. F. Kress, J. Kellermann, I. Mayr, X. Lee, R. Huber,and W. Bode. 1994. Refined 2.0 A X-ray crystal structure of the snake venomzinc-endopeptidase adamalysin II: primary and tertiary structure determina-tion, refinement, molecular structure, and comparison with astacin, collage-nase and thermolysin. J. Mol. Biol. 239:513–544.
21. Gomis-Ruth, F. X., W. Stocker, R. Huber, R. Zwilling, and W. Bode. 1993.Refined 1.8 A X-ray crystal structure of astacin, a zinc-endopeptidase fromthe crayfish Astacus astacus L.: structure determination, refinement, molec-ular structure, and comparison with thermolysin. J. Mol. Biol. 229:945–968.
22. Hooft, R. W., G. Vriend, C. Sander, and E. E. Abola. 1996. Errors in proteinstructures. Nature 381:272.
23. Kabsch, W. 1993. Automatic processing of rotation diffraction data fromcrystals of initially unknown symmetry and cell constants. J. Appl. Crystal-logr. 26:795–800.
24. Laskowski, R. A., M. W. MacArthur, and D. S. Moss. 1993. PROCHECK: aprogram to check the stereochemical quality of protein structures. J. Appl.Crystallogr. 26:283–291.
25. Leslie, A. G. W. 1991. Molecular data processing, p. 50–61. In D. Moras,A. D. Podjarny, and J. C. Thierry (ed.), Crystallography computing. OxfordUniversity Press, Oxford, United Kingdom.
26. Navaza, J. 2001. Implementation of molecular replacement in AMoRe. ActaCrystallogr. D57:1367–1372.
27. Otwinoswki, Z., and W. Minor. 1997. Processing of X-ray diffraction datacollected in oscillation mode. Methods Enzymol. 276:307–326
28. Rose, T., P. Sebo, J. Bellalou, and D. Ladant. 1995. Interaction of calciumwith Bordetella pertussis adenylate cyclase toxin: characterization of multiplecalcium-binding sites and calcium-induced conformational changes. J. Biol.Chem. 270:26370–26376.
29. Roussel, A., and C. Cambillau. 1992. TURBO-FRODO. BiographicsAFMB, Marseille, France.
30. Russell, N. J. 1998. Molecular adaptations in psychrophilic bacteria: poten-tial for biotechnological applications. Adv. Biochem. Eng. Biotechnol. 61:1–21.
31. Villeret, V., J. P. Chessa, C. Gerday, and J. Van Beeumen. 1997. Preliminarycrystal structure determination of the alkaline protease from the Antarcticpsychrophile Pseudomonas aeruginosa. Protein Sci. 6:2462–2464.
VOL. 185, 2003 CRYSTAL STRUCTURES OF A PSYCHROPHILIC METALLOPROTEASE 4203
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Janu
ary
2022
by
191.
240.
25.1
62.


















