
Patterns of anatomical damage in a population of nine ...jloughry/Reprints/Mamm02.pdf · Patterns...
12
Patterns of anatomical damage in a population of nine-banded armadillos Dasypus novemcinctus (Xenarthra, Dasypodidae) W.J. LOUGHRY, CM. MCDONOUGH and E.G. ROBERTSON Department of Biology, Valdosta State University, Valdosta, GA 31698-0015 E-mail.. [email protected] Summary. - We described the types of anatomical damage exhibited by a population of nine-banded armadillos (Dasypus novemcinctus) in northern Florida USA, and quantify the extent and frequency of occurrence of damage among different subgroups of animals. Anatomi- cal damage included tail loss, missing portions of the carapace (notches), torn or missing ears, scarring of the carapace, and band abnormalities. Damage accumulated with age: adults exhibi- ted significantly more damage than either juveniles or yearlings, and, among adults, the amount of damage increased over time. There were few sex differences in the extent or incidence of any type of damage, nor was there any evidence that damage affected reproductive success, although lactating females did exhibit more damage than non-lactating females. Body weights of damaged and undamaged animals did not differ for juveniles or yearlings, but damaged adults were signi- ficantly heavier than non-damaged individuals. Possible causes of damage included predators, conspecific aggression, the physical environment, and developmental instability. Resume. - Description de la nature des dommages anatomiques observes dans une popula- tion de tatous it neuf bandes du nord de la Floride (Etats-Unis), et evaluation de l'etendue et de la frequence de tels dommages chez differents sous-groupes de cette population. Ces dommages concernent la peete de la queue, des pertes partielles de carapace (entailles), la dechirure ou la peete des oreilles, des cicatrices sur la carapace, ou des anomalies des bandes. Ces accidents se multiplient avec l'age: il y en a plus chez les adultes que chez les juveniles ou les jeunes de l'annee precedente ; l'importance des dommages augmente avec Ie temps chez les adultes. II y a peu de difference entre les sexes it ce point de vue. Ces blessures ne paraissent pas affecter la reproduction, bien que les femelles allaitantes soient plus atteintes que les femelles non allai- tantes. Le poids des animaux portant ou non des lesions ne differe pas chez les juveniles ou les jeunes de l'annee precedente. Cependant, les adultes portant des lesions sont significativement plus lourds que les autres. Les prectateurs, les agressions intraspecifiques, l'environnement phy- sique et l'instabilite du developpement figurent parmi les causes possibles de ces dommages. KEY WORDS: Dasypus novemcinctus, armadillos, morphology, phenotypic damages. INTRODUCTION It is a standard observation in evolutionary biology that individuals differ phenoty- pically from one another (Darwin 1859). While evolutionists focus on those variations Mammalia, t. 66. no 1, 2002.. 111-122.
Transcript of Patterns of anatomical damage in a population of nine ...jloughry/Reprints/Mamm02.pdf · Patterns...

Patterns of anatomical damage in a population of nine-bandedarmadillos Dasypus novemcinctus (Xenarthra, Dasypodidae)
W.J. LOUGHRY, CM. MCDONOUGH and E.G. ROBERTSON
Department of Biology, Valdosta State University, Valdosta, GA 31698-0015E-mail.. [email protected]
Summary. - We described the types of anatomical damage exhibited by a population ofnine-banded armadillos (Dasypus novemcinctus) in northern Florida USA, and quantify theextent and frequency of occurrence of damage among different subgroups of animals. Anatomi-cal damage included tail loss, missing portions of the carapace (notches), torn or missing ears,scarring of the carapace, and band abnormalities. Damage accumulated with age: adults exhibi-ted significantly more damage than either juveniles or yearlings, and, among adults, the amountof damage increased over time. There were few sex differences in the extent or incidence of anytype of damage, nor was there any evidence that damage affected reproductive success, althoughlactating females did exhibit more damage than non-lactating females. Body weights of damagedand undamaged animals did not differ for juveniles or yearlings, but damaged adults were signi-ficantly heavier than non-damaged individuals. Possible causes of damage included predators,conspecific aggression, the physical environment, and developmental instability.
Resume. - Description de la nature des dommages anatomiques observes dans une popula-tion de tatous it neuf bandes du nord de la Floride (Etats-Unis), et evaluation de l'etendue et dela frequence de tels dommages chez differents sous-groupes de cette population. Ces dommagesconcernent la peete de la queue, des pertes partielles de carapace (entailles), la dechirure ou lapeete des oreilles, des cicatrices sur la carapace, ou des anomalies des bandes. Ces accidents semultiplient avec l'age: il y en a plus chez les adultes que chez les juveniles ou les jeunes del'annee precedente ; l'importance des dommages augmente avec Ie temps chez les adultes. II y apeu de difference entre les sexes it ce point de vue. Ces blessures ne paraissent pas affecter lareproduction, bien que les femelles allaitantes soient plus atteintes que les femelles non allai-tantes. Le poids des animaux portant ou non des lesions ne differe pas chez les juveniles ou lesjeunes de l'annee precedente. Cependant, les adultes portant des lesions sont significativementplus lourds que les autres. Les prectateurs, les agressions intraspecifiques, l'environnement phy-sique et l'instabilite du developpement figurent parmi les causes possibles de ces dommages.
KEY WORDS: Dasypus novemcinctus, armadillos, morphology, phenotypic damages.
INTRODUCTION
It is a standard observation in evolutionary biology that individuals differ phenoty-pically from one another (Darwin 1859). While evolutionists focus on those variations
Mammalia, t. 66. no 1, 2002.. 111-122.

112 MAMMALIA
that might have a genetic foundation and thus be heritable, many deleterious alterationsof the phenotype may be acquired non-genetically during an individual's lifetimethrough interaction with the environment (biotic and abiotic). Such «damage» maystill be of interest by indicating which individuals are most often targets of conspecificaggression (Huntingford and Turner 1987) or have a higher risk of predation (e.g.,Juanes and Smith 1995; Johnson and Jakob 1999). In addition, anatomical damagemay be important if it negatively impacts an individual's fitness, either by limiting theability to compete for mates, or by generating a phenotype treated as less desirable bythe opposite sex (Andersson 1994). In this paper, we provide the first quantitative des-cription of anatomical damage in a population of nine-banded armadillos (Dasypusnovemcinctus).
Nine-banded armadillos are medium-sized, burrowing mammals (Mc Bee andBaker 1982). They possess relatively long, sharp claws used for digging in the soil ~after prey (primarily invertebrates, Kalmbach 1943; Clark 1951 ; Fitch et al. 1952; .
Breece and Dusi 1985 ; Redford 1985 ; Sikes et al. 1990; White 1992; Lippert 1994)and for digging burrows (Zimmerman 1990; McDonough et al. 2000). A summary of ...
the phenotype of D. novemcinctus can be found in McBee and Baker (1982; see alsoWetzel 1982, 1985). Briefly, the top and sides of the body are covered by a thick pro-tective carapace made of ossified dermal scutes and covered by a leathery skin. Thiscarapace is divided into 3 components: a front immovable scapular shield extendingfrom the head to the shoulders, followed by a series of 8-9 flexible bands, and finally aposterior immovable shield beginning at the back legs and ending at the tail base. Thetail is long and bony and normally comprised of about 14 segments and a tip ofvariable length. Nine-banded armadillos also have relatively large, thin ears protrudingfrom the top of the head. The carapace, ears, and tail are the parts of the armadillo phe-notype most exposed to the environment and, as described below, damage can occur toany of them.
METHODS
Data were collected on the nine-banded armadillos inhabiting Tall TimbersResearch Station, Leon County, Florida. With the exception of 1996, this populationhas been studied each summer between 1992 and 1999. Basic procedures are describedin detail elsewhere (McDonough and Loughry 1997 a; Loughry and McDonough1998; McDonough et al. 2000). Briefly, we censused the population each night by .walking or driving along roads and trails on the property. We attempted to capture allarmadillos observed during these censuses using large dip nets attached to 1.5-2mpoles. Once captured, animals were sexed, weighed, measured, and marked for perma- 'nent identification with ear tags and a passive induced transponder (PIT) tag injectedunder the skin and for long-range identification with reflective tape glued to the cara-pace. In addition, an ear notcher was used to obtain small tissue samples for geneticstudies. Once captured and marked, animals were not recaught again during the yearunless the reflective tape had worn off and they needed to be remarked. With theexception of ear tags, there was no evidence that our marking procedures generated anydamage to the animals. However, upon recapture, some individuals exhibited tears inthe ears from loss of an ear tag. In what follows, we only recorded instances of damagethat were not due to our handling procedures.

PHENOTYPIC DAMAGE IN ARMADILLOS 113
At first capture animals were assigned to age categories on the basis of body size(Loughry and McDonough 1996) as juveniles (young of the year), yearlings, andadults. The lactational status of yearling and adult females was recorded as lactating,possibly lactating, or not lactating using the criteria of Loughry et at. (1998 ; see alsoLoughry and McDonough 1996).
We identified five major categories of damage in the animals we captured: (1) Tailloss occured when some portion of the tail was missing. We subdivided tail loss intothose cases where just the 15th segment (i.e., the tail tip) was missing versus instanceswhere a larger section was gone. In the latter case we counted how many segments ofthe tail were still present. (2) Notches involved cases where a portion of the carapacewas gone (i.e., skin plus dermal scutes). Notches could occur anwhere on the surface ofthe carapace although most tended to be found on the movable bands. For each animal,we counted the number of notches present. (3) Damage to the ears entailed one of twopossibilities. In the first case, the ears were tom with slits that extended a variablelength down one or both ears. In this case, although tom, no ear tissue was missing. Thesecond case involved instances where some portion of one or both ears was gone. Foreach animal we recorded whether the ears were tom or not. If some ear tissue was mis-sing, we estimated how much was gone from both ears combined (e.g., if an animal wasmissing the top half of its right ear, this represented a loss of 0.25 of the total ear tissueof that individual). (4) Scars were similar to notches in that they involved damage to thecarapace, but no tissue was missing. Instead, the top skin layer was gone so that thewhite scutes underneath were visible. Scars varied in size from single, thin scratches orsmall circular patches to large, irregularly shaped patches that could cover large sectionsof the carapace (some scars nearly covered the entire back carapace). For each animal,we counted the number of scars and their size (small =scars less than 2 cm wide;large = scars> 2 cm). We combined these measures into an index of scarring by multi-plying the number of scars by their size (1 = small, 2 = large). (5) Band abnormalitieswere instances where the movable bands were not individually distinct (Newman andPatterson 1911). For example, two bands might fuse together to form a single band onone side of the body. It is arguable whether such abnormalities represent a type of ana-tomical damage, but we have included them here because they represent a form of phe-notypic asymmetry, and such asymmetries have been shown in many cases to have sub-stantial impacts on fitness (Moller and Swaddle 1997). For each animal, we recordedwhether or not any abnormalities were present (we did not record the number of abnor-malities because no animal exhibited more than one). Finally, we used these data togenerate an overall index of the damage exhibited by an individual. Total damage wascalculated as the sum of: tail loss (calculated as 15 - the number of segments
. remaining; individuals missing just the tail tip were assigned a value of 14) + number ofnotches + ears tom (yes = 1, no = 0) + amount of ears missing + scarring (number of
i scars x scar size as described above) + band abnormality (present =1, absent =0). Notethat more conspicuous types of damage (e.g., tail loss) were given heavier weighting incalculating this total index, rather than treating all types of damage as equivalent.
We analyzed the frequency of occurrence of damage in different age and sexgroups of armadillos using Fisher's exact (for 2 x 2 comparisons) and chi-square tests(for larger analyses). These analyses were performed for (a) the numbers of individualsexhibiting any type of damage versus no damage and (b) for the numbers of indivi-duals exhibiting tailloss, notches, tom ears, missing ears, scars, and band abnormalitiesversus the number not exhibiting each type of damage. We also compared the distribu-tion of types of tail loss (tip only versus more) and scars (large versus small) betweenage/sex groups.

114 MAMMALIA
The extent of damage (number of tail segments remaining, number of notches,amount of ears missing, overall scarring, and total damage) was compared between agegroups with ANOVAs and between males and females within age groups with t-tests.Initially, these comparisons included data from all individuals (damaged and undama-ged). However, this introduced a potential bias into the analyses because the frequencyof occurrence of damage sometimes differed between groups. To evaluate this, weredid the analyses using data from just individuals exhibiting each particular type ofdamage. With just a few exceptions (reported below), we found no significant diffe-rences between groups in these analyses, suggesting that, when damage occurs, it isequally severe for all individuals. Because so few significant differences in the extentof damage were obtained in these latter analyses, we only report data from compari-sons using data from all individuals.
Next, we tried to identify potential influences on damage. Weights of damaged ver- J'sus undamaged individuals were compared with t-tests. Because we found few sex diffe-
rences in damage within age groups (see below), data for these analyses were pooled"across males and females. We also performed linear regression analyses of the extent ofdamage (number of notches, scarring, and total damage) with body weight. Multipleweight measurements for the same animal in the same age category Guvenile, yearling,adult) were averaged to produce a single value for the analyses. Finally, for adults andyearlings, we compared the extent and frequency of occurrence of damage between ani-mals that reproduced at least once during the study versus those that had not, and bet-ween females that were either lactating, possibly lactating, or not lactating. Comparisonsof reproductive versus non-reproductive individuals used data from 1992-1995 wheremicrosatellite DNA data were available to identify parents of particular litters (Loughryet at. 1998; Prodohl et at. 1998). Linear regression was used to determine whether fit-ness covaried with the total damage exhibited by an individual. Fitness was measuredfirst as the number of juveniles caught that were attributed to a particular individual(= realized fitness). However, because female D. novemcinctus invariably give birth tolitters of genetically identical quadruplets (Newman and Patterson 1910 ; Patterson 1913; Hamlett 1932; Storrs and Williams 1968; Prodohl et at. 1996), we estimated maxi-mum fitness as the total number of offspring that could possibly have been produced byan individual in cases where some, but not all, members of a litter were captured (e.g., iftwo juveniles from two separate litters were assigned to a particular adult, that animal'srealized fitness was 2, but its maximum fitness was 8 ; see Loughly et at. 1998). Forcomparisons of lactational status, females were classified as (1) lactating if they wereidentified as lactating during at least one year of the study, (2) possibly lactating if theywere identified as possibly lactating in at least one year, but never identified as defini-tely lactating, and (3) not lactating if they were never identified as lactating in any year .of the study. We used data from both yearlings and adults in analyzing the relationshipbetween reproduction, lactation and anatomical damage. This was done primarily,because there were insufficient samples to analyse yearlings separately (for reproductivesuccess n = 8 yearlings; for lactational status, n = 15 females, all of which were non-lactating). Yearlings are physiologically capable of reproduction (McCusker 1977 ; Pep-pler 1985), so it may be reasonable to lump them with adults. Nonetheless, we also ana-lyzed adult data separately to determine if effects were due simply to the inclusion ofdata from yearlings. Because we obtained exactly the same pattern of results when year-lings were included as when they were excluded, in this paper we only present resultsfrom analyses using pooled data from adults and yearlings.
We attempted to determine whether adults continue to accumulate more damageas they age by performing a linear regression between the total damage exhibited by an

PHENOTYPIC DAMAGE IN ARMADILLOS 115
individual and its current age. Age was determined as follows: all adults were arbitra-rily assigned an age of 2 years old when first captured, with age increasing yearly withcaptures in subsequent years. Thus, an individual first caught as an adult in 1992 andrecaptured in 1999 was considered to be 9 years old. Because we did not know theactual ages of these animals, our estimates represent the minimum age of an individual.Not knowing the «true» age of adults is a problem for our analyses, but we do notconsider the problem fatal because, regardless of true age, if damage accumulates withage, animals first caught in 1992 should exhibit more damage than those caught subse-quently. Nonetheless, as a further analysis, we performed a regression between age andtotal damage using just data from juveniles recruited into the population (n =39 ani-mals captured 81 times), where the true age of each individual was known.
RESULTS
Data were available for 631 animals (444 adults, 31 yearlings, and 156 juveniles).Of these, 443 (70.21 %) exhibited some form of anatomical damage (table 1). The fre-quency of occurrence of damaged individuals differed significantly among age groups(X2 = 84.29, P < 0.0001 ; table 1) as did the occurrence of tail loss (loss of tip plusmore combined, X2 = 71.34, P < 0.0001), notches (X2 = 42,50 P < 0.0001), and tomand missing ears (X2 = 33.61 and 15.61, p < 0.0001 and 0.0004 respectively; df = 2 inall analyses). However, the occurrence of band abnormalities, scars (large and smallscars combined), and the distribution of scars (large versus small) and tail loss (tip ver-sus more; juveniles were excluded from this comparison) did not (Chi-square tests,p > 0.20, df = 2 in all comparisons; table 1).
The extent of anatomical damage also differed between age groups. Adults hadsignificantly shorter tails and more total damage than either juveniles or yearlings, andhad significantly more notches and more ear tissue missing than did juveniles (table 2).Although the extent of damage did not differ between age groups when data from just
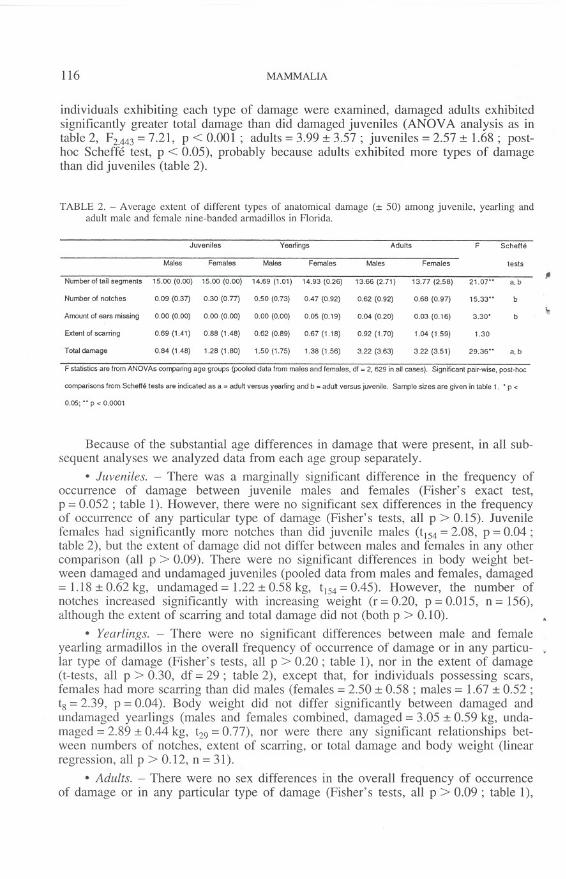
TABLE 1. - Frequency of occurence of different types of anatomical damage among juvenile, yearling andadult male and female nine-banded armadillos in Florida.
Damage Tail loss Notches Ears Ears Scars Band
Yes No Tip More torn missing Small Large abnormalities
Juvenile males 25 50 0 0 5 2 0 16 5 2
Juvenile females 40 41 0 0 14 1 0 24 3 8
Yearling males 11 5 1 1 6 1 0 5 1 0
Yearling females 9 6 1 0 4 0 1 3 1 2
Adult males 202 42 30 51 98 56 18 66 22 16
Adult females 156 44 21 42 83 33 17 60 23 15
116 MAMMALIA
individuals exhibiting each type of damage were examined, damaged adults exhibitedsignificantly greater total damage than did damaged juveniles (ANOYA analysis as intable2, F2.443= 7.21, P < 0.001; adults= 3.99:t 3.57; juveniles=2.57:t 1.68; post-hoc Scheffe test, p < 0.05), probably because adults' exhibited more types of damagethan did juveniles (table 2).
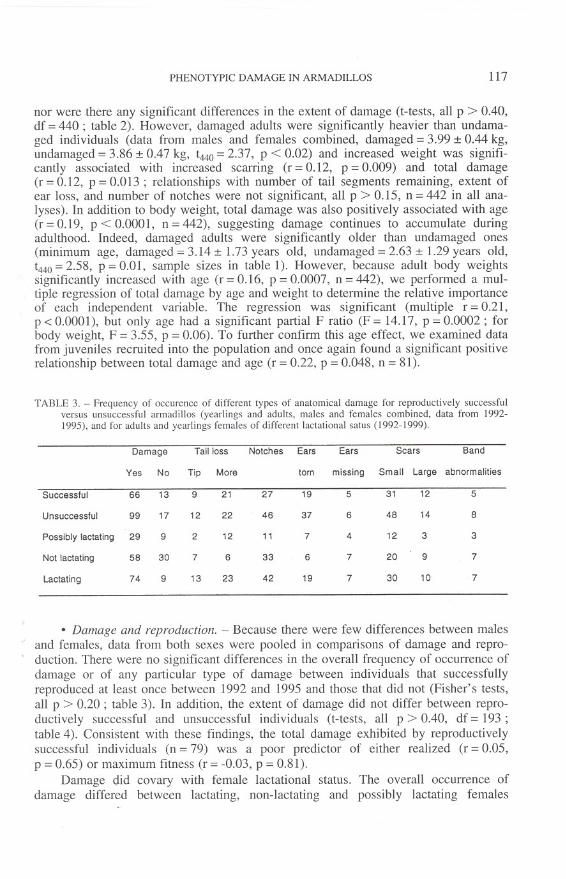
TABLE 2. - Average extent of different types of anatomical damage (ot 50) among juvenile, yearling andadult male and female nine-banded armadillos in Florida.
Because of the substantial age differences in damage that were present, in all sub-sequent analyses we analyzed data from each age group separately.
. Juveniles. - There was a marginally significant difference in the frequency ofoccurrence of damage between juvenile males and females (Fisher's exact test,p = 0.052 ; table 1). However, there were no significant sex differences in the frequencyof occurrence of any particular type of damage (Fisher's tests, all p> 0.15). Juvenilefemales had significantly more notches than did juvenile males (t154= 2.08, P= 0.04;table 2), but the extent of damage did not differ between males and females in any othercomparison (all p > 0.09). There were no significant differences in body weight bet-ween damaged and undamaged juveniles (pooled data from males and females, damaged= 1.18 :t 0.62 kg, undamaged = 1.22 :t 0.58 kg, t154= 0.45). However, the number ofnotches increased significantly with increasing weight (r = 0.20, P = 0.015, n = 156),although the extent of scarring and total damage did not (both p > 0.10).
. Yearlings. - There were no significant differences between male and femaleyearling armadillos in the overall frequency of occurrence of damage or in any particu- .lar type of damage (Fisher's tests, all p> 0.20 ; table 1), nor in the extent of damage(t-tests, all p > 0.30, df = 29; table 2), except that, for individuals possessing scars,females had more scarring than did males (females =2.50 :t 0.58 ; males = 1.67 :t 0.52 ;ts =2.39,P=0.04). Body weight did not differ significantlybetween damaged andundamaged yearlings (males and females combined, damaged =3.05 :t 0.59 kg, unda-maged=2.89:t 0.44kg, t29 =0.77), nor were there any significant relationships bet-ween numbers of notches, extent of scarring, or total damage and body weight (linearregression, all p > 0.12, n = 31).
. Adults. - There were no sex differences in the overall frequency of occurrenceof damage or in any particular type of damage (Fisher's tests, all p > 0.09 ; table 1),
Juveniles Yearlings Adults F Scheffe
Males Females Males Females Males Females tests
-Number of tail segments 15.00 (0.00) 15.00 (0.00) 14.69 (1.01) 14.93 (0.26) 13.66 (2.71) 13.77 (2.58) 21.07" a.b
Number of notches 0.09 (0.37) 0.30 (0.77) 0.50 (0.73) 0.47 (0.92) 0.62 (0.92) 0.68 (O.g7) 15.33" b
Amount of ears missing 0.00 (0.00) 0.00 (0.00) 0.00 (0.00) 0.05 (0.19) 0.04 (0.20) 0.03 (0.16) 3.30' b..
Extent of scarring 0.69 (1.41) 0.88 (1.48) 0.62 (0.89) 0.67 (1.18) 0.92 (1.70) 1.04 (1.59) 1.30
Total damage 0.84 (1.48) 1.28 (1.80) 1.50 (1.75) 1.38 (1.56) 3.22 (3.63) 3.22 (3.51) 29.36" a,b
F statistics are from ANOVAs comparing age groups (pooled data from males and temales, df = 2, 629 in all cases), Significant pair.wise, post-hoc
comparisons from Scheffe tests are indicated as a = adun versus yearling and b = adun versus iuvenile. Sample sizes are given in table 1. .P <
0.05; ..P < 0.0001
PHENOTYPIC DAMAGE IN ARMADILLOS 117
nor were there any significant differences in the extent of damage (Hests, all p > OAO,df = 440 ; table 2). However, damaged adults were significantly heavier than undama-ged individuals(data from males and females combined,damaged=3.99:t OA4kg,undamaged=3.86:t OA7kg, t440= 2.37, P < 0.02) and increasedweight was signifi-cantly associated with increased scarring (r = 0.12, P = 0.009) and total damage(r = 0.12, P = 0.013 ; relationships with number of tail segments remaining, extent ofear loss, and number of notches were not significant, all p > 0.15, n =442 in all ana-lyses). In addition to body weight, total damage was also positively associated with age(r = 0.19, P < 0.0001, n = 442), suggesting damage continues to accumulate duringadulthood. Indeed, damaged adults were significantly older than undamaged ones(minimum age, damaged = 3.14:t 1.73 years old, undamaged =2.63 :t 1.29 years old,t440= 2.58, P = 0.01, sample sizes in table 1). However, because adult body weightssignificantly increased with age (r = 0.16, P = 0.0007, n = 442), we performed a mul-tiple regression of total damage by age and weight to determine the relative importanceof each independent variable. The regression was significant (multiple r =0.21,P < 0.0001), but only age had a significant partial F ratio (F = 14.17,P = 0.0002; forbody weight, F = 3.55, P = 0.06). To further confirm this age effect, we examined datafrom juveniles recruited into the population and once again found a significant positiverelationship between total damage and age (r = 0.22,P = 0.048,n = 81).
TABLE 3. - Frequency of occurence of different types of anatomical damage for reproductively successfulversus unsuccessful armadillos (yearlings and adults, males and females combined, data from 1992-1995), and for adults and yearlings females of different lactational satus (1992-1999).
. Damage and reproduction. - Because there were few differences between males< and females, data from both sexes were pooled in comparisons of damage and repro-, duction. There were no significant differences in the overall frequency of occurrence of
damage or of any particular type of damage between individuals that successfullyreproduced at least once between 1992 and 1995 and those that did not (Fisher's tests,all p > 0.20 ; table 3). In addition, the extent of damage did not differ between repro-ductively successful and unsuccessful individuals (t-tests, all p > OAO, df = 193 ;table 4). Consistent with these findings, the total damage exhibited by reproductivelysuccessful individuals (n =79) was a poor predictor of either realized (r = 0.05,p = 0.65) or maximum fitness (r = -0.03,p = 0.81).
Damage did covary with female lactational status. The overall occurrence ofdamage differed between lactating, non-lactating and possibly lactating females
Damage Tail loss Notches Ears Ears Scars Band
Yes No Tip More torn missing Small Large abnormalities
Success1Ur----- 66 13 9 21 27 19 5 31 12 5
Unsuccessful 99 17 12 22 46 37 6 48 14 8
Possibly lactating 29 9 2 12 11 7 4 12 3 3
Not lactating 58 30 7 6 33 6 7 20 9 7
Lactating 74 9 13 23 42 19 7 30 10 7

118 MAMMALIA
(X2 = 13.6, P = 0.002, df = 2) as did the frequency of tom ears (X2=8.86, p = 0.01)and the distribution of tail loss (tip versus more, X2= 17.58, p. =0.0002; table 3).There was a marginally significant difference in the frequency of occurrence of notches(X2 =5.88, P =0.053). The extent of damage also varied among females of differinglactational status for tail loss (ANOY A, F2.2O6=6.54, P =0.002) and total damage(F2.2O6=5.48, P =0.005 ; table 4). Post-hoc pair-wise comparisons with Scheff6 testsshowed that non-lactating females had significantly longer tails than either possibly ordefinitely lactating females (p < 0.05 in both cases) and had less total damage thandefinitely lactating females (p < 0.05).
For reproductive success, data are from all adult and yearling males and females present from 1992-1995, while dataon lactational status are from all adult and yearling females sampled between 1992-1999. Sample size are in Table 3.
Differences due to lactation may have been tied to age because lactating femaleswere significantly older than other females (ANOVA, F2.206=17.35,P < 0.0001,meanminimum age of lactating females =3.73:t 2.23 years, possibly lactating females =3.18:t 1.69years, not lactating females =2.20 :t 1.03 years; significant post-hoc Scheff6comparions were obtained between lactating and non-lactating females and betweenpossibly lactating and non-lactating females). This difference was not solely due to theinclusion of yearling females in the analysis. Lactating females were still significantly'older when only data from adult females were used (mean ages for lactating and possi- ,bly lactatingfemaleswere the sameas above,for not lactating,age=2.45:t 0.96years, .ANOV A F2.19l=10.57, P < 0.0001).
DISCUSSION
Anatomical damage was widespread in our population, with at least 70 % of theanimals exhibiting some type of damage (table 1). The most common types of damagewere, in descending order: scarring, notches, and tail loss. The main difference indamage between individuals seemed to be in the occurrence of damage rather than its
TABLE 4. - Average extent of different types of anatomical damage (:t 50) among reproductively successfulnine-banded armadillos and among females of differing lactational status. I
Reproductive success Lactational status\
Successful Unsuccessful Possibly Not Defintely
lactating lactating lactating
Number of tail segments 13.43 (2.81) 13.74 (2.75) 13.05 (3.58) 14.55 (1.60) 13.47 (2.58)
Number of notches 0.52 (0.83) 0.58 (0.86) 0.39 (0.72) 0.69 (1.08) 0.78 (0.94)
Amount of ears missing 0.03 (0.13) 0.03 (0.19) 0.03 (0.08) 0.04 (0.19) 0.04 (0.16)
Extent of scarring 1.50 (2.02) 1.34 (1.81) 1.05 (1.68) 0.68 (1.59) 1.16 (1.53)
Total damage 3.90 (3.85) 3.60 (3.83) 3.68 (4.60) 2.21 (2.85) 3.82 (3.27)

PHENOTYPIC DAMAGE IN ARMADILLOS 119
extent. For example, fewer juveniles exhibited notches than did adults, but, if a juve-nile had notches, they were likely to be just as numerous as those exhibited by anadult. Such findings suggest that, whatever the cause of a particular type of damage, itimpacts all individuals with roughly equal severity.
We can not definitely identify the causes of anatomical damage described in thispaper, but certain possibilities seem likely. For example, tail loss, especially of portionsgreater than just the tail tip, most probably results from encounters with predators. Alli-gators (Alligator mississippiensis) are the only confirmed predators of armadillos thatoccur at Tall Timbers (McNease and Joanen 1977; Shoop and Ruckdeschel 1990).However, other potential predators include coyotes (Canis latrans), bobcats (Lynxrufus), domestic dogs (Canis familiaris), and various raptors (McDonough and Loughry1997b). It is possible that tail breakage might occur during the winter as a result ofexposure to freezing temperatures, but this seems unlikely at our study site where days
0- with consistently freezing temperatures are relatively rare (36.1 days per year withdaily minima below freezing, but only 0.2 days per year with daily maxima below free-zing, based on data from the last 30 years, US National Climatic Data Center). If tailloss does result from encounters with predators, our data suggest that such encountersoccur at similar frequencies for males and females. However, adults apparently hadmore encounters than juveniles or yearlings. This may seem contradictory given dataon carapaces of dead animals collected in the field that indicated juveniles sufferedmore from predation than did adults (McDonough and Loughry 1997 b). The two datasets can be reconciled by noting that tail loss represents only a partially successful pre-dation attempt, while the carapaces that were collected presumably represented indivi-duals killed by a predator. If so, then the obvious conclusion is that juveniles are lesslikely to survive an encounter with a predator than are adults.
Other forms of damage may also be due to predators. For example, ear loss, largescars on, or large notches out of, the carapace might result from encounters with preda-tors. Indeed, we have captured animals with fresh scars or with fresh chunks taken outof the carapace that seem to have been generated by a predator. However, predation isnot the only possible cause of these types of damage. Armadillos have sharp clawswhich they use in fights with conspecifics (McDonough 1994) and some scarring,notches, and ear damage may result from conspecific aggression. In addition, the habi-tat at our study site contains many thorny shrubs and bushes that could also generatethese types of damage. Predation events and con specific aggression are both relativelyrare events, so it will be difficult to obtain direct evidence about the sources of damageexhibited by a particular armadillo.
While there may be a variety of causes for most types of anatomical damage, deve-lopmental instability seems the only possible explanation for band abnormalities. At pre-
.' sent, we can not determine whether these abnormalities are due to a genetic mutation or, some epigenetic influence during development. However, it seems unlikely that a gene-
tic mutation can be the sole cause. Nine-banded armadillos are polyembryonic, givingbirth to litters of genetically-identical quadruplets (Newman and Patterson 1910; Patter-son 1913; Hamlett 1932; Storrs and Williams 1968; Prodohl et al. 1996). If bandabnormalities have a genetic basis, one would expect all members of a litter to exhibitthem. In fact, we identified five instances where band abnormalities occurred within acloneship containing multiple members (3 groups of 4 sibs, 2 groups of 2 sibs), but in nocase did all members of the cloneship exhibit the abnormality (in 4 of the 5 litters only 1sib exhibited the abnormality, in the other litter, 3 of 4 did). Such findings are consistentwith other reports documenting substantial phenotypic variability among clonemates(Newman and Patterson 1911 ; Burchfield 1967; Bagatto et al. 2000).

120 MAMMALIA
The primary influence on the damage exhibited by an individual was age. This isnot surprising because increased longevity should increase the probability of encounterswith the agents that can cause anatomical damage. However, there were a few instancesof sex differences. Juvenile females exhibited more notches than did juvenile males, andyearling females exhibited more scarring than yearling males (for those yearlings pos-sessing scars at all). It is difficult to interpret these results. The lack of differences indamage between adult males and females suggest that differences occuring earlier in lifemay be transitory and thus, of questionable significance. Alternatively, our results mayindicate that females are exposed to more risks early in life, although it is not obviouswhy this would occur. Additional data are required to clarify this issue.
At first glance, anatomical damage might seem an obvious cue armadillos coulduse in identifying potential mates, either positively because damage is a handicap indi-cating an individual's ability to survive (Zahavi and Zahavi 1997), or negatively Ibecause damage indicates an inferior phenotype (Andersson 1994). However, compari- -sons of anatomical damage with reproductive success suggest that armadillos do not 'tuse damage as a cue for discriminating among potential mates. Coupled with earlieranalyses that found few influences on armadillo fitness in this population (Loughry etal. 1998), it appears that, at least over the years studied, reproduction is relatively ran-dom. Other studies have also suggested that anatomical damage need not negativelyimpact fitness (e.g., Johnson and Jakob 1999).
To conclude, our data indicate that, while armadillos in our population may exhi-bit extensive and diverse types of phenotypic damage, there appear to be few functio-nal consequences to the animals of being damaged. To our knowledge, this study is thefirst to fully describe and quantify the types of anatomical damage that can occur inarmadillos. It will be interesting to see if the patterns we report are confirmed in otherpopulations.
ACKNOWLEDGEMENTS
We thank T. Engstrom and L. Brennan of Tall Timbers Research Station for their continuedsupport of our work. This study was supported by a Tall Timbers Research Station internshipawarded to E.G.R. and Valdosta State University faculty research awards to C.M.M. andW.J.L. We thank C. Denys, J.B. Pascarella, and an anonymous reviewer for their comments onthe manuscript and F. Delsuc for his invaluable help in translating the abstract into French.
BIBLIOGRAPHY ..
ANDERSSON,M., 1994. - Sexual selection. Princeton University Press, Princeton.
BAGATTO,B., CROSSLEY,D.A. II and W.W. BURGGREN,2000. - Physiological variability in neo-natal armadillo quadruplets: within- and between-litter differences. J. Exp. BioI., 203 :1733-1740.
BREECE,G.A. and J.L. Dus!, 1985. - Food habits and home ranges of the common long-nosedarmadillo Dasypus novemcinctus in Alabama. Pp.419-427 in: The evolution and eco-logy of armadillos, sloths, and vermilinguas. Ed. Montgomery. Smithsonian InstitutionPress, Washington, D.C.
BURCHFIELD,E.E.S., 1967. - Individuality in monozygotic quadruplets of the armadillo, Dasypusnovemcinctus, Linn. Ph. D. : dissertation. Univ. Texas, Austin, USA.

PHENOTYPIC DAMAGE IN ARMADILLOS 121
CLARK, W.K, 1951. - Ecological life history of the armadillo in the eastern Edwards Plateauregion. Am. MidI. Nat., 46 : 337-358.
DARWIN,e., 1859. - On the origin of species. John Murray, London; facsimile edition, HarvardUniversity Press, Cambridge, 1964.
FITCH, H.S., P. GOODRUMand C. NEWMAN, 1952. - The armadillo in the southeastern U.S.J. Mammal., 33 : 21-37.
HAMLETI, G.W.D., 1932. - The reproductive cycle of the armadillo. Z. Wissenschaft., Zool.,141 : 143-157.
HUNTINGFORD,EA. and A.K TURNER, 1987. - Animal conflict. Chapman and Hall, London.
JOHNSON,S.A. and EM. JAKOB, 1999. - Leg autonomy in a spider has minimal costs in competi-tive ability and development. Anim. Behav., 57 : 957-965.
JuANES, E and LD. SMITH, 1995. - The ecological consequences of limb damage and loss indecapod crustaceans: a review and prospectus. 1. Exper. Mar. BioI. Ecol., 193: 197-223.
.- KALMBACH,ER, 1943. - The armadillo: its relation to agriculture and game. Texas Game,Fish and Oyster Commission, Austin, Texas.
LIPPERT,KJ., 1994. - Food habits, distribution and impact of the nine-banded armadillo in Mis-souri. M.S. thesis. Southwest Missouri State University, Springfield, USA.
LOUGHRY,W.J. and e.M. McDoNOUGH, 1996. - Are road-kills valid indicators of armadillopopulation structure? Am. MidI. Nat., 135: 53-59.
LOUGHRY,W.J. and e.M. McDoNOUGH, 1998. - Spatial patterns in a population of nine-bandedarmadillos. Am. Midi. Nat., 140: 161-169.
LOUGHRY,W.J., P.A. PRODOHL,e.M.McDONOUGH, W.S. NELSONand J.e. AVISE, 1998. - Cor-relates of reproductive success in a population of nine-banded armadillos. Can. J. Zool.,76: 1815-1821.
McBEE, K and R.J. BAKER, 1982. - Dasypus novemcinctus. Mamm. Spec., 162: 1-9.
MCCUSKER, J.S., 1977. - The nine-banded armadillo, Dasypus novemcinctus, in north centralTexas with emphasis on the reproductive biology of the male. M.S. thesis, Texas Chris-tian University, Fort Worth, USA.
McDoNOUGH, C.M., 1994. - Determinants of aggression in nine-banded armadillos. J. Mammal.,75: 189-198.
McDONOUGH, e.M. and W.J. LOUGHRY,1997 a. - Influences on activity patterns in a populationof nine-banded armadillos. J. Mammal., 78 : 932-941.
McDoNOUGH, e.M. and W.J. LOUGHRY,1997 b. - Patterns of mortality in a population of nine-banded armadillos, Dasypus novemcinctus. Am. MidI. Nat., 138 : 299-305.
McDoNOUGH, C.M., M.A. DELANEY, P.Q. LE, M.S. BLACKMOREand W.J. LOUGHRY, 2000. -Burrow characteristics and habitat associations of armadillos in Brazil and the United
States. Rev. BioI. Tropic., 48 : 109-102.
. McNEASE,L and T.JOANEN,1977.- Alligator diets in relation to marsh salinity. Proc. Annu.Y Coni Southeast. Assoc. Fish Wild. Agen., 31: 36-40.
"MOLLER, A.P. and J.P. SWADDLE, 1997. - Asymmetry, developmental stability, and evolution.Oxford University Press, New York.
NEWMAN, H.H. and J.T. PATIERSON, 1910. - The development of the nine-banded armadillofrom primitive streak to birth, with special reference to the question of polyembryony. J.Morphol., 21 : 359-423.
NEWMAN,H.H. and J.T. PATTERSON,1911. - The limits of hereditary control in armadillo qua-druplets : a study of blastogenic variation. 1. Morphol., 22 : 855-926.
PATIERSON,J.T., 1913. - Polyembryonic development in Tatusia novemcinctus. J. Morphol., 24 :559-684.
PEPPLER,RD., 1985. - Reproductive parameters in the nine-banded armadillo: correlation of folli-cular development and plasma progesterone concentration with age. Anat. Rec., 211 : 351.

122 MAMMALIA
PRODOHL, P.A., W.J. LOUGHRY, e.M. MCDONOUGH, W.S. NELSON and J.e. AVISE, 1996. - Mole-
cular documentation of polyembryony and the micro-spatial dispersion of clonal sibshipsin the nine-banded armadillo, Dasypus novemcinctus. Proc. R. Soc. London, Ser. B,263: 1643-1649.
PRODOHL, P.A., W.J. LOUGHRY, e.M. MCDONOUGH, W.S. NELSON, E.A. THOMPSON andJ.e. AVISE, 1998. - Genetic maternity and paternity in a local population of armadillosassessed by microsatellite DNA markers and field data. Am. Nat., 151 : 7-19.
REDFORD,K.H., 1985. - Food habits of armadillos (Xenarthra: Dasypodidae). Pp.429-437 in:The evolution and ecology of armadillos, slots, and vermilinguas. Ed. Montgomery.Smithsonian Institution Press, Washington, D.e.
SHOOP, C.R and e.A. RUCKDESCHEL,1990. - Alligators as predators on terrestrial mammals.Am. Mid!. Nat., 124: 407-412.
SIKES, R.S., G.A. HEIDT and D.A. ELROD, 1990. - Seasonal diets of the nine-banded armadillo(Dasypus novemcinctus) in a northern part of its range. Am. Mid!. Nat., 123 : 383-389. j
STORRS,E.E. and RJ. WILLIAMS, 1968. - A study of monozygotic quadruplet armadillos in reIa- '>-tion to mammalian inheritance. Proc. Natl. Acad. Sci., 60 : 910-914.
WETZEL, R.M., 1982. - Systematics, distribution, ecology, and conservation of South Americanedentates. Pp. 345-375, in: Mammalian biology in South America. Eds. Mares andGenoways, Special Publication Series, Pymatuning Laboratory of Ecology, University ofPittsburgh.
WETZEL, RM., 1985. - Taxonomy and distribution of armadillos, Dasypodide. Pp.23-46, in:The evolution and ecology of armadillos, sloths, and vermilinguas. Ed. Montgomery.Smithsonian Institution Press, Washington, D.e.
WHITE, LL, 1992. - Seasonal dietary habits of the nine-banded armadillo in southern Alabamaand the surrounding area. M.S. thesis. University of South Alabama, Mobile, USA.
ZAHAVI, A. and A. ZAHAVI, 1997. - The handicap principle. Oxford University Press,New York.
ZIMMERMAN,J.W., 1990. - Burrow characteristics of the nine-banded armadillo, Dasypus novem-cinctus. Southwest. Nat., 35 : 226-227.
't
..
~"


















