
Leishmania infantum Visceral Infection with Mediates Heterologous ...
13
of April 16, 2018. This information is current as Leishmania infantum Visceral Infection with Mediates Heterologous Protection against Leishmania major Cutaneous Infection with L. Sacks and Nathan C. Peters Audrey Romano, Nicole A. Doria, Jonatan Mendez, David ol.1500752 http://www.jimmunol.org/content/early/2015/09/13/jimmun published online 14 September 2015 J Immunol Material Supplementary 2.DCSupplemental http://www.jimmunol.org/content/suppl/2015/09/13/jimmunol.150075 average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2015 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 16, 2018 http://www.jimmunol.org/ Downloaded from by guest on April 16, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Leishmania infantum Visceral Infection with Mediates Heterologous ...

of April 16, 2018.This information is current as
Leishmania infantumVisceral Infection with Mediates Heterologous Protection against
Leishmania majorCutaneous Infection with
L. Sacks and Nathan C. PetersAudrey Romano, Nicole A. Doria, Jonatan Mendez, David
ol.1500752http://www.jimmunol.org/content/early/2015/09/13/jimmun
published online 14 September 2015J Immunol
MaterialSupplementary
2.DCSupplementalhttp://www.jimmunol.org/content/suppl/2015/09/13/jimmunol.150075
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2015 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Cutaneous Infection with Leishmania major MediatesHeterologous Protection against Visceral Infection withLeishmania infantum
Audrey Romano,* Nicole A. Doria,* Jonatan Mendez,* David L. Sacks,* and
Nathan C. Peters†
Visceral leishmaniasis (VL) is a fatal disease of the internal organs caused by the eukaryotic parasite Leishmania. Control of VL
would best be achieved through vaccination. However, this has proven to be difficult partly because the correlates of protective
immunity are not fully understood. In contrast, protective immunity against nonfatal cutaneous leishmaniasis (CL) is well defined
and mediated by rapidly recruited, IFN-g–producing Ly6C+CD4+ T cells at the dermal challenge site. Protection against CL is
best achieved by prior infection or live vaccination with Leishmania major, termed leishmanization. A long-standing question is
whether prior CL or leishmanization can protect against VL. Employing an intradermal challenge model in mice, we report that
cutaneous infection with Leishmania major provides heterologous protection against visceral infection with Leishmania infantum.
Protection was associated with a robust CD4+ T cell response at the dermal challenge site and in the viscera. In vivo labeling of
circulating cells revealed that increased frequencies of IFN-g+CD4+ T cells at sites of infection are due to recruitment or retention
of cells in the tissue, rather than increased numbers of cells trapped in the vasculature. Shortly after challenge, IFN-g–producing
cells were highly enriched for Ly6C+T-bet+ cells in the viscera. Surprisingly, this heterologous immunity was superior to homol-
ogous immunity mediated by prior infection with L. infantum. Our observations demonstrate a common mechanism of protection
against different clinical forms of leishmaniasis. The efficacy of leishmanization against VL may warrant the introduction of the
practice in VL endemic areas or during outbreaks of disease. The Journal of Immunology, 2015, 195: 000–000.
The leishmaniases consist of a broad range of cutaneous,mucocutaneous, and visceral diseases caused by differentstrains of the eukarytotic parasite Leishmania. Leish-
mania, which is transmitted to the human host by the bite ofa sand fly vector, is an obligate intracellular pathogen that es-tablishes chronic infection within phagocytes. Visceral forms ofleishmaniasis are fatal when left untreated, and although drugs areavailable they are expensive, highly toxic, and drug resistance iscommon (1, 2). Visceral leishmaniasis caused by Leishmaniadonovani is endemic in northern India and East Africa, whereasLeishmania infantum, also known as Leishmania chagasi, is thecausative agent of visceral leishmaniasis (VL) in South Americaand the Mediterranean basin, which has experienced several recentoutbreaks (3–6). In some areas, L. infantum infection of dogs has
reached epidemic proportions and dogs are thought to be a majorreservoir of human disease (7).No vaccine is currently available for any form of leishmaniasis
in people. However, a deliberate, single needle inoculation ofinfectious Leishmania major into the skin without the disease-exacerbating factors coegested during natural sand fly transmis-sion, termed leishmanization, provides complete and long-lastinghomologous protection against sand fly–transmitted cutaneousdisease and has been used extensively as a live vaccine in humans(8–12). Despite its efficacy and the convenience of a single ad-ministration, leishmanization has largely been abandoned becauseof rare adverse reactions at the site of inoculation (13, 14); fur-thermore, the chronic nature of the infection raises concernsshould leishmanized individuals become immunocompromised,although there are no reports of reactivation or dissemination ofL. major in leishmanized individuals. A more justifiable use ofleishmanization would be to vaccinate against strains that causelethal visceral leishmaniasis (VL), for which the benefits ofleishmanization may outweigh any risks. Cross-protection con-ferred by leishmanization against VL would suggest a commonmechanism of resistance against Leishmania species that causedifferent clinical diseases, and it would suggest that differentLeishmania species share a sufficient number of protective Ags towarrant their use in pan-Leishmania vaccines (15, 16). However,evidence that L. major infection cross-protects against VL inpeople is rare or difficult to interpret (17–23).Experimentally, two prior studies have investigated this question
and found that leishmanization either provided no protection (24)or enhanced visceral infection (25) following L. infantum chal-lenge. However, these studies employed BALB/c mice that aresusceptible to L. major infection due to a defect in the generationof Th1 immunity, a condition not typically observed in people
*Laboratory of Parasitic Diseases, National Institute of Allergy and Infectious Dis-eases, National Institutes of Health, Bethesda, MD 20892; and †Snyder Institute forChronic Diseases, Department of Microbiology Immunology and Infectious Dis-eases, Cumming School of Medicine, University of Calgary, Calgary, Alberta T2N4Z6, Canada
Received for publication March 31, 2015. Accepted for publication August 17, 2015.
This work was supported by the Intramural Research Program of the National Insti-tute of Allergy and Infectious Diseases and by University of Calgary start-up funds(to N.C.P.).
Address correspondence and reprint requests to Prof. Nathan C. Peters, Snyder In-stitute for Chronic Diseases, Department of Microbiology Immunology and Infec-tious Diseases, Cumming School of Medicine, University of Calgary, 3280 HospitalDrive NW, Calgary, AB T2N 4Z6, Canada. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BFA, brefeldin A; CL, cutaneous leishmaniasis;dICS, direct intracellular staining; dLN, draining lymph node; GzB, granzyme B; i.d.,intradermal(ly); LDA, limiting dilution analysis; LST, leishmanin skin test; VL,visceral leishmaniasis.
Copyright� 2015 by The American Association of Immunologists, Inc. 0022-1767/15/$25.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1500752
Published September 14, 2015, doi:10.4049/jimmunol.1500752 by guest on A
pril 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

infected with L. major (26). In contrast, leishmanized C57BL/6mice more closely replicate the immune status of leishmanizedhumans (11). Therefore, we employed an intradermal (i.d.) chal-lenge model of visceral infection caused by L. infantum in C57BL/6mice leishmanized with L. major (27). We present evidence thatleishmanization provides robust protection and similar correlatesof protection against both cutaneous and visceral infection.Leishmanization may be a viable strategy for control of visceraldisease.
Materials and MethodsParasites
L. major Friedlin strain was isolated from a patient who acquired hisinfection in the Jordan Valley (MHOM/IL/80/Friedlin). L. infantum(MHOM/ES/92/LLM-320; isoenzyme typed MON-1) was isolated froma patient with VL in Spain and was provided by Diane MacMahon-Pratt.L. infantum–RFP was generated as previously described (28). Parasiteswere cultured in vitro at 26˚C in complete medium 199 supplementedwith 20% heat-inactivated FCS (Gemini Bio-Products), 100 U/ml peni-cillin, 100 mg/ml streptomycin, 2 mM L-glutamine, 40 mM HEPES,0.1 mM adenine (in 50 mM HEPES), 5 mg/ml hemin (in 50% trietha-nolamine), and 1 mg/ml 6-biotin. For L. infantum, the complete medium199 was further supplemented with 2 mg/ml 6-biopterin (Sigma-Aldrich,St Louis, MO). L. infantum and L. major infective-stage metacyclicpromastigotes were isolated from stationary cultures (4–6 d old) by cen-trifugation through a Ficoll-step gradient as described (29). For leishma-nization, L. major metacyclic promastigotes were isolated by negativeselection of noninfective forms using peanut agglutinin (Vector Labora-tories) (30).
Mice
Female C57BL/6 mice were obtained from Taconic. All mice weremaintained in the National Institute of Allergy and Infectious Diseasesanimal care facility under specific pathogen-free conditions.
Leishmanization and challenge
Leishmanized mice were generated by injecting 104 L. major metacyclicpromastigotes s.c. in the hind footpad in a volume of 40 ml and used at 12–20 wk after primary infection when footpad lesions had completely re-solved. Mice with a primary L. infantum infection where generated in thesame manner. Naive mice, leishmanized mice, or L. infantum–infectedmice were challenged with 2 3 106 L. infantum metacyclic promastigotesi.d. in the ear in a volume of 10 ml. In some experiments, mice were in-jected i.v. in the tail vein with 23 106 L. infantum metacyclic promastigotesin a volume of 200 ml.
Processing of different sites of infection and parasitequantification
Mice were perfused via intracardiac injection of 20 ml cold PBS. Liverperfusion was performed by injection of 6 ml cold PBS into the liver portalvein. The spleen and ear draining lymph node (dLN) were removed, cut withtweezers, homogenized with a syringe plunger, and the cell suspension wasfiltered through a 70-mm strainer. Livers were incubated for 45 min with 2ml DMEM containing 250 mg/ml Liberase TL purified enzyme blend(Roche Diagnostics) and 10 mg/ml DNase. Liver cells were further purifiedusing a Percoll gradient (31). In experiments employing direct intracellularstaining (dICS), organs were processed in media prewarmed to 37˚Ccontaining 20 mg/ml brefeldin A (BFA) and incubated at 37˚C and 5% CO2
postprocessing for a total time in BFA of 4 h. Ear tissue was prepared aspreviously described (11). Briefly, ears were removed and placed in 70%ethanol for 2–5 min and then allowed to dry. Separated dorsal and ventralsheets of ears were then incubated at 37˚C for 90 min in 1 ml DMEMcontaining 160 mg/ml Liberase. Following Liberase treatment, tissue washomogenized for 3.5 min in a Medicon using a Medimachine (BectonDickinson). The tissue homogenate was then flushed from the Mediconwith 10 ml RPMI 1640 media containing 0.05% DNase and filtered usinga 50-mm pore size cell strainer. In experiments employing dICS, BFA at20 mg/ml was added to media prewarmed to 37˚C and the ear homogenatewas returned to 37˚C and 5% CO2 postprocessing for a total time in BFAof 4 h. In some experiments mice were not perfused and RBCs were re-moved from the spleen and liver using ACK lysing buffer for 5 min atroom temperature. Cells from each tissue were resuspended in completemedium 199 with 6-biopterin.
Parasite loads were determined by 2-fold serial dilutions in 96-wellflat-bottom microtiter plates by overlaying 100 ml of the diluted tissuesuspension onto 50 ml NNN medium containing 20% defibrinated rabbitblood. The dilutions were made in duplicate. The plates were scoredmicroscopically for growth and the number of parasites in each tissuewas determined from the highest dilution at which parasites could begrown out after 7–10 d incubation at 26˚C. In some experiments 25 mg/mlhygromycin B (Sigma-Aldrich) was added to the media to discriminatebetween parasites used for live vaccination versus parasites used forchallenge.
Restimulation of tissue-derived cells for cytokine analysis byflow cytometry
Tissue-derived cells were restimulated as described previously (11).Briefly, single-cell suspensions were incubated at 37˚C in 5% CO2 for12–14 h in flat-bottom 48-well plates with 0.5–1 3 106 T cell–depleted(Miltenyi Biotec) naive spleen cells (APCs), with or without 50 mg/mlfreeze-thaw Leishmania Ag in a total volume of 1 ml. During the final 4–6 h of culture, 1 mg/ml BFA (Sigma-Aldrich) was added. Cells were thenwashed and labeled with LIVE/DEAD Fixable Aqua Cell Stain Kit(ThermoFischer Scientific) at a 1:500 dilution of the manufacturersuggested stock solution (Invitrogen) to exclude dead cells and anti–FcIII/II (CD16/32) receptor Ab (2.4G2), followed by PE-Cy7 anti-mouseCD4 (RM4-5), and in some experiments allophycocyanin–eFluor 780CD8a (53-6.7) for 20 min. In some experiments cells were stained withPerCp-Cy5.5 anti-CD4 (RM4-5), FITC anti-CD44 (IM7), allophycocyanin–eFluor 780 anti-Ly6C (HK1.4) and PE-Cy7 anti-CD62L (MEL-14) for20 min. Cells were then fixed with BD Cytofix/Cytoperm (BD Bio-sciences) according to the manufacturer’s instructions and stained withFITC anti–IFN-g (XMG1.2), PerCP–eFluor 710 anti–TNF-a (MP6-XT22), and in some experiments allophycocyanin anti–TcR-b (145-2C11) and/or PE anti-human granzyme B (GzB; GRB04, Invitrogen)and/or V500 anti-CD3 (145-2C11). In some experiments, samples weretreated with Foxp3 fixation/permeabilization buffer (eBioscience) per themanufacturer’s instructions, and subsequently stained for 1 h at 4˚C withPE anti–IFN-g (XMG1.2) and eFluor 660 anti-T-bet (eBio4B10) Abs.Isotype controls employed were rat IgG1 (R3-34) and rat IgG2b (A95-1or eBR2a). All Abs were from eBioscience or BD Biosciences. Datawere collected using FACSDiva software on a FACSCanto flow cytom-eter (BD Biosciences) and analyzed using FlowJo software (Tree Star).Forward scatter and side scatter widths were employed to exclude celldoublets from analysis.
Perfusion and in vivo staining of circulating cells
In vivo labeling of circulating cells was done as described previously(32). Briefly, 0.6 mg anti–TCR-b BV421 (clone H57-597, BioLegend)was injected i.v. in 200 ml PBS via the tail vein. Three minutes after theinjection, mice were sacrificed by isoflurane asphyxiation and perfusedvia intracardiac injection of 20 ml cold PBS. Liver perfusion was per-formed by injection of 6 ml cold PBS into the liver portal vein. Organswere then harvested as described above. Following restimulation andintracellular staining the frequency of TcR-b+ cells labeled by i.v. ad-ministration of TcR-b Ab was not reduced on T cells from the liver(30.0 6 13 TcR-b+ before restimulation versus 29.6 6 12 TcR-b+ afterrestimulation and intracellular staining, n = 14) and only slightly re-duced on cells from the spleen (39.2 6 13 TcR-b+ before restimulationversus 34.0 6 12 TcR-b+ after restimulation and intracellular staining,n = 12).
Statistical analysis
Unless otherwise noted, data were compared using the Mann–Whitney Utest. Comparisons between multiple groups were done using ANOVAwitha Holm–Sidek posttest for comparisons between multiple groups. All pvalues are two-sided. Statistical calculations were done in GraphPad Prism5.0c (http://www.graphpad.com). For figures depicting pooled data, errorbars represent the SEM. For figures in which data points represent indi-vidual replicates, error bars represent the SD of the mean. A p value,0.05was considered significant.
Ethics statement
All animal experiments were performed under the LPD-68E Animal StudyProtocol approved by the National Institute of Allergy and InfectiousDiseases Animal Care and Use Committee using guidelines established bythe Animal Welfare Act and the Public Health Service Policy on HumaneCare and Use of Laboratory Animals.
2 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
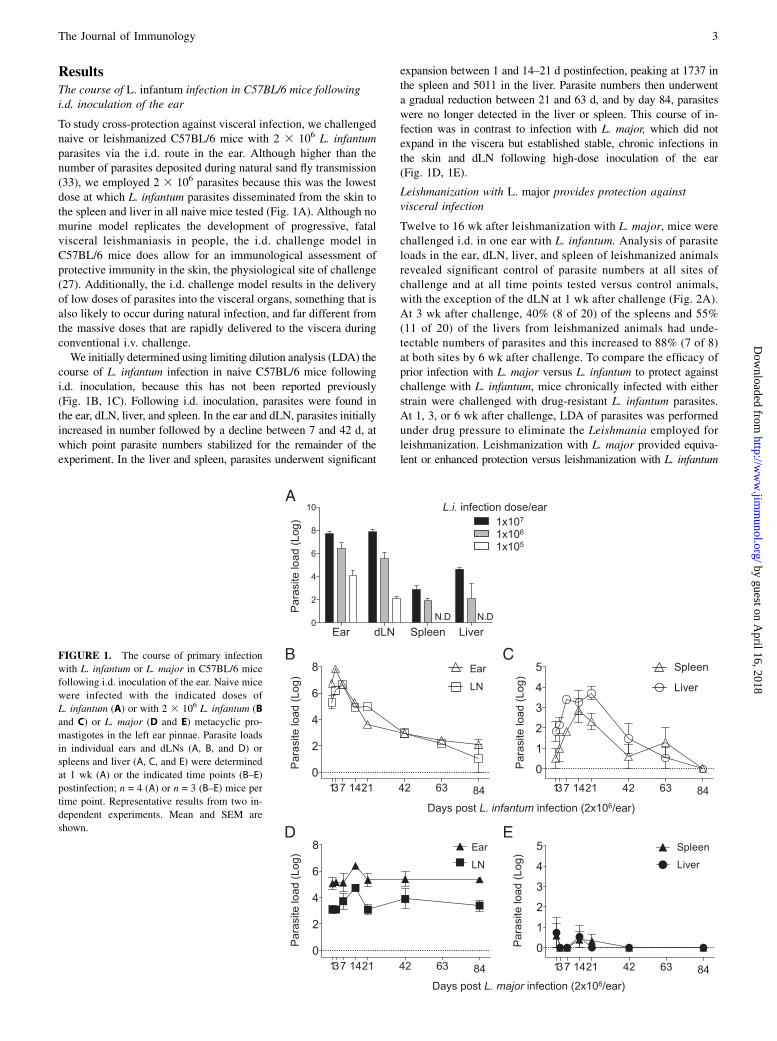
ResultsThe course of L. infantum infection in C57BL/6 mice followingi.d. inoculation of the ear
To study cross-protection against visceral infection, we challengednaive or leishmanized C57BL/6 mice with 2 3 106 L. infantumparasites via the i.d. route in the ear. Although higher than thenumber of parasites deposited during natural sand fly transmission(33), we employed 2 3 106 parasites because this was the lowestdose at which L. infantum parasites disseminated from the skin tothe spleen and liver in all naive mice tested (Fig. 1A). Although nomurine model replicates the development of progressive, fatalvisceral leishmaniasis in people, the i.d. challenge model inC57BL/6 mice does allow for an immunological assessment ofprotective immunity in the skin, the physiological site of challenge(27). Additionally, the i.d. challenge model results in the deliveryof low doses of parasites into the visceral organs, something that isalso likely to occur during natural infection, and far different fromthe massive doses that are rapidly delivered to the viscera duringconventional i.v. challenge.We initially determined using limiting dilution analysis (LDA) the
course of L. infantum infection in naive C57BL/6 mice followingi.d. inoculation, because this has not been reported previously(Fig. 1B, 1C). Following i.d. inoculation, parasites were found inthe ear, dLN, liver, and spleen. In the ear and dLN, parasites initiallyincreased in number followed by a decline between 7 and 42 d, atwhich point parasite numbers stabilized for the remainder of theexperiment. In the liver and spleen, parasites underwent significant
expansion between 1 and 14–21 d postinfection, peaking at 1737 inthe spleen and 5011 in the liver. Parasite numbers then underwenta gradual reduction between 21 and 63 d, and by day 84, parasiteswere no longer detected in the liver or spleen. This course of in-fection was in contrast to infection with L. major, which did notexpand in the viscera but established stable, chronic infections inthe skin and dLN following high-dose inoculation of the ear(Fig. 1D, 1E).
Leishmanization with L. major provides protection againstvisceral infection
Twelve to 16 wk after leishmanization with L. major, mice werechallenged i.d. in one ear with L. infantum. Analysis of parasiteloads in the ear, dLN, liver, and spleen of leishmanized animalsrevealed significant control of parasite numbers at all sites ofchallenge and at all time points tested versus control animals,with the exception of the dLN at 1 wk after challenge (Fig. 2A).At 3 wk after challenge, 40% (8 of 20) of the spleens and 55%(11 of 20) of the livers from leishmanized animals had unde-tectable numbers of parasites and this increased to 88% (7 of 8)at both sites by 6 wk after challenge. To compare the efficacy ofprior infection with L. major versus L. infantum to protect againstchallenge with L. infantum, mice chronically infected with eitherstrain were challenged with drug-resistant L. infantum parasites.At 1, 3, or 6 wk after challenge, LDA of parasites was performedunder drug pressure to eliminate the Leishmania employed forleishmanization. Leishmanization with L. major provided equiva-lent or enhanced protection versus leishmanization with L. infantum
FIGURE 1. The course of primary infection
with L. infantum or L. major in C57BL/6 mice
following i.d. inoculation of the ear. Naive mice
were infected with the indicated doses of
L. infantum (A) or with 2 3 106 L. infantum (B
and C) or L. major (D and E) metacyclic pro-
mastigotes in the left ear pinnae. Parasite loads
in individual ears and dLNs (A, B, and D) or
spleens and liver (A, C, and E) were determined
at 1 wk (A) or the indicated time points (B–E)
postinfection; n = 4 (A) or n = 3 (B–E) mice per
time point. Representative results from two in-
dependent experiments. Mean and SEM are
shown.
The Journal of Immunology 3
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(Fig. 2B). This was most significant in the ear where the het-erologous protection against L. infantum was similar to the levelof protection against homologous challenge with L. major (Fig.2C). At all time points tested, with the exception of the 6 wktime point in the liver when control mice are beginning to clearparasites from this organ, protection was associated with sig-nificantly greater numbers of leishmanized versus control miceexhibiting sterilizing immunity in the spleen and liver as de-termined by the inability to culture organisms from these tissues(p # 0.036, Fisher exact test). As far as we know, these obser-vations demonstrate the first experimental confirmation that
leishmanization with L. major provides protection against vis-ceral infection.
Leishmanization is associated with a rapid and robust CD4+
T cell response in the skin and visceral organs followingchallenge
IFN-g–producing CD4+ T cells mediate the protective responseconferred by leishmanization against homologous challenge withL. major (34, 35). Additionally, the rapidity of the CD4+ responseis the clearest correlate of protection against sand fly–transmitteddisease (11, 12). Therefore, we analyzed IFN-g production by
1w 3w 6w
0
1
2
3
###
###
1w 3w100
101
102
103
104
105
106
p
0
1
2
3
4
5
6
7 *** ****
**
1w 3w 6w0
1
2
3
4
5
6
7
Para
site
laod
(Log
)
1w 3w 6w
****
***
A
B
C
None L. maj L. inf
L. inf
Infection
L. majL. majL. inf L. inf
L. majL. inf
Infection
Weeks post challenge
None L. majL. inf L. inf
L. infL. inf
Infection
Weeks post challenge
Ear Ear dLN Spleen Liver
Ear
Fold
redu
ctio
n vs
. re
spec
tive
cont
rol
Par
asite
Loa
d (L
og)
Par
asite
Loa
d (L
og)
Ear Ear dLN
Spleen Liver
1w 3w 6w0
2
4
6 *** ***### ####
#
1w 3w 6w0
2
4
6####
###
1w 3w 6w
0
1
2
3
4
##
##
#
$
$
Weeks post challenge
0
1
2
3
4
1w 3w 6w
*******
**
0
1
2
3
4
5
6
Para
site
laod
(Log
)1w 3w 6w
**
***
FIGURE 2. Primary infection with L. major protects against visceral infection with L. infantum. Naive mice or mice with a healed L. major (A–C) or
L. infantum (B and C only) primary infection were challenged with 23 106 L. infantum (A–C) or L. major (C only) metacyclics in the ear. (A and B) Parasite
loads in individual ears, dLNs, spleens, or livers at the indicated time points postchallenge. Data are the pool of five independent experiments including at
least two time points per experiment. Mean and SD (A) or SEM (B) are shown. In (A), *0.01 , p , 0.05, **0.001 , p , 0.01, ***0.0001 , p , 0.001,
****p, 0.0001. In (B), ***0.0001, p, 0.001, refer to differences between L. major/L. infantum and L. infantum/L. infantum; #0.01, p, 0.05, ##0.001
, p , 0.01, ###0.0001 , p , 0.001, ####p , 0.0001 refer to differences between naive/L. infantum and L. major/L. infantum; $0.01 , p , 0.05 refers to
differences between naive/L. infantum and L. infantum/L. infantum. (C) Mean (6SEM) fold parasite load reduction at the site of secondary challenge (ear)
in mice with the indicated primary and secondary challenge infections versus the respective controls.
4 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CD4+ T cells at early time points following heterologous chal-lenge with L. infantum and found increased frequencies of CD4+
T cells with the capacity to make IFN-g in the skin, dLN, spleen,and liver of leishmanized versus control animals (Fig. 3). Aspreviously reported (11), a rapid CD4+ T cells response in the skinwas observed within 3 d of i.d. challenge (Fig. 3B). We also ob-served postchallenge responses in the viscera of leishmanizedmice that were significantly greater than prechallenge responsesstarting at day 3 (liver) or 1 wk (spleen) postchallenge withL. infantum. This early response was similar to the response tohomologous challenge with L. major (Supplemental Fig. 1), sug-gesting that the early protective responses to L. infantum orL. major are the same despite the different clinical outcomesof infection with these two parasites. Additionally, IFN-g produc-tion was elicited equally well employing freeze-thawed Ag fromL. infantum or L. major (ratio of response of L. major/L. infantum1:0.95, n = 4, p = 0.54, paired t test), suggesting these two para-sites also share immunogenic MHC class II–restricted epitopes, assuggested by work with the sterol 24-c-methyltransferase protein(36).
Localization of IFN-g+CD4+ T cells associated with the liverand spleen employing in vivo i.v. staining
Leishmanized animals had low but detectable frequencies ofCD4+ T cells with the capacity to make IFN-g upon stimulationin the spleen and liver prior to challenge (Fig. 4A). However,similar cells were also found in the circulation, making it diffi-cult to conclude whether these cells were residing in the tissueprior to challenge or were simply trapped in the organ vascula-ture. Additionally, trapped circulating cells may contribute to theincreased frequencies of IFN-g+CD4+ T cells detected in thespleen and liver following challenge (Fig. 3B). Although weroutinely perfuse our mice, and perfusion does reduce the fre-quency of IFN-g+ cells in the liver and spleen (SupplementalFig. 2), recent observations have demonstrated that perfusioncan be incomplete (32, 37). Therefore, to discern cytokine pro-duction from cells present in visceral tissues versus those trappedin the vasculature, we in vivo labeled circulating cells prior toeuthanasia and perfusion (32). This technique labels cells on theluminal side of the liver and spleen vasculature as well as cells inthe splenic red pulp. This analysis is important in the context ofvaccine studies to determine whether Ag-specific cells have ac-tually infiltrated infected tissues to mediate effector functionversus those that are circulating. Analysis of live CD4+ T cellsfrom the blood following in vivo labeling revealed .99% werei.v.+ as expected (Fig. 4B). In the liver and spleen we found bothi.v.2 and i.v.+ cells, despite whole-body and liver-specific per-fusion (Fig. 4B). We then determined the frequency of cytokine-producing cells that were in the tissue versus those trapped in thevasculature of the ear, spleen, and liver prior to and at 1 wkfollowing i.d. challenge, the peak of the CD4+ IFN-g T cell re-sponse. We also employed i.v. challenge with 2 3 106 L. infantum,which delivers large numbers of parasites directly into the visceralorgans, particularly the liver (27), to act as a positive control forAg-driven recruitment and/or retention of IFN-g+CD4+ cells in thespleen and liver. Both prior to and following dermal challenge, thevast majority of IFN-g+ cells in the ear were i.v.2 (Fig. 4C, leftpanel), demonstrating that these cells were residing in the tissue.In contrast, most IFN-g+CD4+ cells in the spleen and liver werei.v.+, despite perfusion (i.v.2 versus i.v.+, p , 0.001), demon-strating that these cells were trapped, circulating cells. Thissuggests that most Leishmania-specific cytokine productionattributed to cells associated with the visceral organs in Fig. 3Bis due to cells trapped in the vasculature, not cells that are actually
mediating effector function in the organ. Whereas i.v. tail veinchallenge did not significantly change the data obtained fromthe spleen, there was a dramatic increase in the frequency ofIFN-g+CD4+ T cells from the liver that were i.v.2, suggestingthat Ag load can drive the recruitment and/or retention of IFN-g–producing cells residing in the liver parenchyma (Fig. 4C, rightpanel). Despite the fact that most IFN-g+CD4+ T cells associatedwith the viscera were trapped i.v.+ circulating cells, dermal chal-lenge significantly increased the frequency of IFN-g+CD4+ cellsthat were i.v.2 versus prechallenge mice in both the spleen andliver (compare open circles in Fig. 4C). This was most dramatic inthe liver following tail vein challenge (Fig. 4C, right panel). Tocorrelate responses in the actual tissue with the reductions inparasite numbers mediated by leishmanization observed in Fig. 1,we also determined the frequency of cytokine-producing cellswithin the total, i.v.2, or i.v.+ CD4+ T cell populations followingchallenge (Fig. 4D–F). In both the spleen and liver there wasa significant increase in the frequency of IFN-g+ cells withinthe i.v.2 population following i.d. challenge (Fig. 4E), dem-onstrating that leishmanization mediates a rapid response in thevisceral organs similar to that observed at the dermal site of chal-lenge. Analysis of i.v.+ cells revealed a trend toward increasedfrequencies of IFN-g+ cells following dermal challenge but thisdid not reach significance. These observations demonstrate thatleishmanization with L. major mediates an early CD4+ T cellsresponse in visceral sites of infection following i.d. challengewith L. infantum.
Intravenous2, IFN-g–producing cells in the viscera followingchallenge are enriched for Ly6C expression
We have previously reported that leishmanization maintains highfrequencies of short-lived CD44+CD62L2T-bet+Ly6C+CD4+
effector T cells that are required for protection against rein-fection with L. major in the ear dermis. These cells are notmemory cells reactivated by secondary challenge, and althoughthey produce cytokine in vivo in an Ag-dependent manner, theycan be rapidly recruited to dermal sites of challenge in an Ag-independent manner (35). Therefore, we wanted to determinewhether IFN-g–producing cells in the viscera at 3 d followingdermal challenge of leishmanized mice with L. infantum alsoexpressed the Ly6C+ effector T cell phenotype (Fig. 5). Weemployed dICS to identify IFN-g–producing cells (35) (Fig. 5A).Because dICS does not employ ex vivo Ag or pharmacologicalrestimulation, assessment of Ly6C and T-bet expression using thismethodology likely reflects patterns of in vivo expression. Non-circulating i.v.2CD44+CD62L2CD4+ T cells that expressed Ly6Cwere T-bethigh, as described previously (35) (Fig. 5B), and werehighly enriched for IFN-g–producing cells compared with Ly6C2
cells (Fig. 5C). Additionally, the frequency of T-bet+Ly6C+–expressing cells was significantly higher among IFN-g–producingi.v.2CD44+CD62L2CD4+ T cells versus IFN-g nonproducingi.v.2CD44+CD62L2CD4+ T cells (Fig. 5D). In contrast, IFN-gproduction in naive mice challenged with L. infantum was verylow, and no significant difference was observed between Ly6C+
and Ly6C2 cells. These data demonstrate that, similar to L. majorchallenge of the ear, IFN-g production is highly associated withLy6C-expressing CD4 T cells in the viscera at acute time pointsfollowing challenge with L. infantum.
Following challenge, responding CD8 T cells arepredominantly GzB+ but IFN-g2
Whereas Th1 CD4+ T cells are associated with mediating pro-tection against primary and secondary infection with L. major(35, 38), the role of CD8+ T cells in leishmaniasis remains
The Journal of Immunology 5
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 3. Kinetic of IFN-g–producing CD4+ T cells in the skin and viscera after challenge infection with L. infantum. At the indicated time post-
challenge, cells from mice as shown in Fig. 2Awere restimulated in vitro with APCs plus a mixture of L. major and L. infantum Ag and analyzed by flow
cytometry. (A) Representative intracellular staining contour plots for cytokine-producing CD4+TCRb+ T at 1 wk postchallenge. (B) Kinetic of IFN-g
production by CD4+TCRb+ T cells in ears, dLNs, spleens, and livers. *0.01 , p , 0.05, **0.001 , p , 0.01, ***0.0001 , p , 0.001 refer to differences
between leishmanized unchallenged mice and L. major/L. infantum; #0.01, p, 0.05, ##0.001, p, 0.01, ###0.0001, p, 0.001, ####p, 0.0001 refer to
differences between naive/L. infantum and naive control mice; $$0.001 , p , 0.01, $$$0.0001 , p , 0.001, $$$$p , 0.0001 refer to differences between
naive/L. infantum and L. major/L. infantum; n = 4–5 mice per group per time point. Data are the pool of five independent experiments including at least two
time points (day 3, 1 wk, and/or 3 wk) per experiment.
6 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 4. In vivo i.v. staining reveals localization of IFN-g+CD4+ T cells in the spleen and the liver. (A) Prechallenge production of IFN-g by CD4+
TCRb+ T cells from different organs of uninfected or leishmanized mice following ex vivo Ag restimulation as described in Fig. 3. (B) Representative
histograms of i.v staining with anti–TCRb-BV421 on CD4+ T cells in the blood, spleen and liver. (C) Proportion of i.v.2 or i.v.+ cells within the IFN-g+
CD4+ T cell population (following ex vivo Ag restimulation) from the ear, spleen, or liver of leishmanized mice following no challenge (Lm-⃠), i.d.
challenge in the ear (Lm-Li (Ear)), or i.v. challenge in the tail vein (Lm-Li (Tail Vein) with 2 3 106 L. infantum metacyclics. **0.001 , p , 0.01,
***0.0001 , p , 0.001, ****p , 0.0001 refer to differences between Lm-⃠ and Lm-Li (Ear); #0.01 , p , 0.05, ####p , 0.0001 refer to differences
between Lm-⃠ and Lm-Li (Tail Vein); $$$$p , 0.0001 refers to differences between Lm-Li (Ear) and Lm-Li (Tail Vein). (D–F) Percentage of IFN-g+ cells
in the total CD4+ T cell population (D), the i.v.2CD4+ T cell population (E), or the i.v.+CD4+ T cell population (F). One week after challenge data point is
shown. Data are the pool of three independent experiments employing four to five mice per experiment (two experiments employed tail vein challenge). Li,
L. infantum; Lm, L. major.
The Journal of Immunology 7
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

controversial and is dependent on the dose, strain, and infectionsetting (primary or secondary) (1, 38–41). In visceral infectionwith L. donovani, CD8+ T cells have been shown to be bothprotective and dispensable (40, 42). We determined IFN-g andGzB production by CD8+ T cells following challenge withL. infantum and, similar to observations in BALB/c mice (24),
found a very low frequency of IFN-g+ cells regardless of thesite of analysis (Fig. 6). In contrast, high frequencies of cells
were found to be GzB+. Similar observations were made fol-lowing challenge with L. major (Supplemental Fig. 3). The
postulated role of GzB in leishmaniasis is evolving. GranzymeA3B2/2 double knockout C57BL/6 mice are completely re-
sistant to L. major infection (43) whereas CD8+GzB+ T cellshave been associated with pathology, not parasite killing,
during cutaneous L. braziliensis infection (41, 44, 45). Despitethe high frequency of GzB+CD8+ cells we were unable to detectGzB by ELISA following Ag stimulation (data not shown).
Employing in vivo labeling of circulating cells, we detected an
increase in the frequency of GzB+CD8+ T cells in the viscera
following challenge, although this did not reach significance
(Supplemental Fig. 4).
DiscussionOur results demonstrate for the first time, as far as we know, thatleishmanization protects against heterologous visceral infection.Protection was associated with rapid activation and recruitment ofIFN-g–producing Th1 cells at both the cutaneous site of challengeand visceral sites of infection, demonstrating that protectionconferred by leishmanization is not site specific. Similar to whatwe have previously reported for L. major challenge in the skin,IFN-g production was highly associated with CD44+CD62L2
T-bet+Ly6C+CD4+ T cells within the i.v.2 population in the vis-cera. At early time points following challenge these cells arerecruited, infection-dependent, pre-existing effector T cells, not
FIGURE 5. i.v.2CD44+CD62L2Ly6C+CD4+ T cells in the viscera of leishmanized mice are enriched for IFN-g–producing cells following challenge.
Naive or leishmanized animals were challenged with L. infantum as described in Fig. 2. On day 3 postchallenge CD4+TcRb+ T cells were analyzed for
CD44, CD62L, Ly6C, T-bet, and IFN-g expression employing dICS. (A) Representative dot plots of IFN-g production by i.v.2CD4+ T cells. (B) Rep-
resentative histogram of T-bet expression by the indicated cell populations in the spleen. (C) Percentage of IFN-g+ cells within live i.v.2CD4+CD44+
CD62L2Ly6C+ or live i.v.2CD4+CD44+CD62L2Ly6C2 populations. (D) Percentage of Ly6C+T-bet+ cells within the live i.v.2CD4+CD44+CD62L2IFN-g+
or live i.v.2CD4+CD44+CD62L2IFN-g2 population. In (C) and (D), n = 4 mice/group. Data are representative of two independent experiments. *0.01 ,p , 0.05, **0.001 , p , 0.01, ***0.0001 , p , 0.001, ****p , 0.0001.
8 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
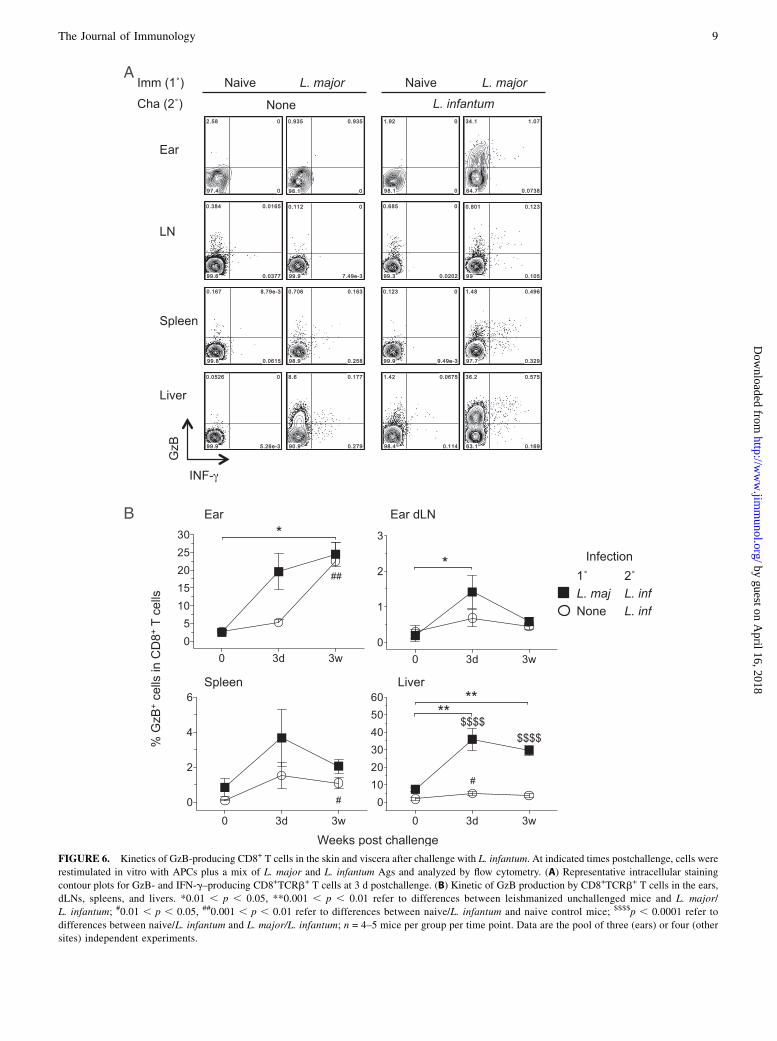
FIGURE 6. Kinetics of GzB-producing CD8+ T cells in the skin and viscera after challenge with L. infantum. At indicated times postchallenge, cells were
restimulated in vitro with APCs plus a mix of L. major and L. infantum Ags and analyzed by flow cytometry. (A) Representative intracellular staining
contour plots for GzB- and IFN-g–producing CD8+TCRb+ T cells at 3 d postchallenge. (B) Kinetic of GzB production by CD8+TCRb+ T cells in the ears,
dLNs, spleens, and livers. *0.01 , p , 0.05, **0.001 , p , 0.01 refer to differences between leishmanized unchallenged mice and L. major/
L. infantum; #0.01 , p , 0.05, ##0.001 , p , 0.01 refer to differences between naive/L. infantum and naive control mice; $$$$p , 0.0001 refer to
differences between naive/L. infantum and L. major/L. infantum; n = 4–5 mice per group per time point. Data are the pool of three (ears) or four (other
sites) independent experiments.
The Journal of Immunology 9
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

memory cells reactivated by challenge (35). Our observationssuggest that the correlates of protective immunity against cuta-neous leishmaniasis (CL) and VL may be sufficiently similar towarrant a pan-Leishmania vaccine provided the vaccine can rep-licate the correlates of immunity observed in leishmanized indi-viduals (11, 12, 35). Our data also provide the best evidence todate that heterologous immunity can be identical, or even superior,to homologous immunity.The controlled trials demonstrating the efficacy of leishmanization
in humans is restricted to protection against CL caused by L. major orLeishmania tropica. However, epidemiological studies suggest priorinfection of humans with L. major may provide cross-protectionagainst VL caused by L. donovani. Two longitudinal studies of VLin Sudan demonstrated that despite the presence of both leishmaninskin test (LST)–positive and –negative individuals at the beginning ofeach study, all cases of VL were reported among the LST-negativegroup (17, 18). Because LST positivity was highly associated withprior residence in an L. major endemic area, the authors hypothesizedthat prior infection with L. major provides protection against VLcaused by L. donovani. Related observations in Sri Lanka reportedthat districts with high levels of CL caused by a naturally attenuatedstrain of L. donovani are almost devoid of VL (19). Experimentally,susceptible BALB/c mice infected with the attenuated L. donovanistrain had significantly reduced parasite burden in the liver uponchallenge with the L. donovani strain responsible for Sri Lankan VL(46). In a vervet monkey model of disease, Gicheru et al. (47) re-ported the inverse result, that subclinical infection with L. donovaniprovided complete protection against cutaneous leishmaniasis fol-lowing experimental challenge with L. major.Experimental cross-protection mediated by infection (24, 25, 48–
54) as well as T cell cross-reactivity with Ags from differentLeishmania species (15, 16) have been studied extensively in animalmodels, primarily in an effort to define proteins that could be used ina pan-Leishmania vaccine. Genome sequencing has also revealedthat 90% of the genome is conserved between L. major andL. infantum, and 99% of the genes are syntenic (55, 56), suggestinga strong likelihood of shared Ags. Remarkably, only two experi-mental studies have investigated the cross-protection mediated byprimary infection with L. major against subsequent visceral infection(24, 25). These studies employed BALB/c mice chronically infectedwith L. major and showed no protection against either i.v. or i.d.challenge with L. infantum. In contrast, we were able to demonstratethe protective effect of leishmanization against visceral infectionemploying C57BL/6 mice with a healed L. major infection, whichmore closely replicates the immune status of leishmanized humans.Our observations are limited to an assessment of protection againstinfection, not disease, because mice infected i.d. with visceralizingstrains of Leishmania produce minimal pathology in the viscera.Additionally, the i.d. model of infection does not establish long-terminfection in naive mice as determined by the detection of parasites inthe liver and spleen by LDA at 12 wk postinfection (Fig. 1). Al-though this may be viewed as a limitation of the model, the use ofthe more conventional high-dose i.v. challenge model would bypassthe potentially important initial step of parasite deposition in theskin, the natural site of infection. Additionally, note that in humansthe immunobiology of asymptomatic infection with visceralizingparasites is understudied, and the scarcity of parasites in the visceramay in fact be a common finding in the field (57).Although the ideal challenge model would employ exposure to
the bites of L. infantum–infected sand flies, we have yet to es-tablish a reliable sand fly transmission model of L. infantum in ourlaboratory. Rather, we employed relatively early time points, thatis, 1 wk after needle challenge, to determine protective immunityin the viscera, as early control of parasite numbers in the skin
following needle challenge has proven to be an informative cor-relate of protection against cutaneous disease following challengewith L. major–infected sand flies (11, 12).Our observations suggest that most cytokine production typically
associated with adaptive T cell immunity in the spleen and liver isfrom circulating cells trapped in the vascular or the splenic red pulpat the time of euthanasia. Despite this observation, we were able toemploy a combination of in vivo labeling of circulating cells andprechallenge leishmanized mice to demonstrate that leishmani-zation does mediate a rapid Leishmania-specific immune responsein visceral tissue in response to dermal challenge with L. infantum.The use of the in vivo labeling technique would appear to becritical in future studies to determine the relative contribution ofcirculating cells versus tissue-resident cells to total cytokineproduction. Importantly, note that even before challenge, we foundlow frequencies of i.v.2 cells with the capacity to make IFN-gin the uninfected ear, non-dLN, blood, liver and spleen ofleishmanized mice, similar to what we have reported previously(35). Whether the i.v.2IFN-g+CD4+ T cells in the viscera aretissue-resident memory T cells, as has been reported for the lung(58), skin (59), and vaginal mucosa (60), or patrolling cells, as hasalso been reported for the skin (61), is unknown. Whereas pre-existing Ag-specific T cells are not required for the recruitment ofhighly protective Leishmania-specific T cells into a site of non-specific tissue damage (35), this does not preclude the possibilitythat they may enhance protection, regardless of their patrolling orresident nature as recently shown (59). Extensive studies haveidentified a role for pre-existing CD8+ tissue-resident memoryT cells in protective immunity (62). We found strong evidence forthe existence of i.v.2, resident, Leishmania-specific GzB+CD8+
T cells in the viscera of leishmanized mice prior to challenge. Therole of these cells in protection will be the topic of future research.Our data reinforces the rationale for reintroducing the practice of
leishmanization under conditions where the benefits, such as pro-tection against fatal visceral disease, are deemed to outweigh therisks associated with administering live L. major. Numerous ge-netically modified or avirulent Leishmania strains have been studiedin an effort to replicate the protective effect of leishmanizationwhile reducing or controlling for the virulence of the parasite (52,63–68). How effective these modified strains are compared withleishmanization with wild-type L. major has not been addressed,with a single exception (68). Past observations (34, 35, 69, 70)suggest that attenuation resulting in sterile clearance rather thanpersistence will compromise long-term protection, especiallyagainst the more stringent conditions of infected sand fly challenge.The ideal situation would be to leishmanize with a parasite thatminimizes pathology but also persists over the long term. An ad-ditional drawback of leishmanizing against VL is the wisdom ofintroducing L. major in areas where the parasite is not endemic,such as northeast India. However, L. major strains that establishpersistent infection in the mammalian host but cannot producetransmissible infections in the sand fly midgut, such as various lpgknockout parasites, have already been developed that would addressthese concerns (67, 71–73).
AcknowledgmentsWe thank Kim Beacht, Melanie Faivre-Charmoy, and Flavia L. Ribeiro-
Gomes for assistance with experiments. We especially thank Kristin
Anderson and David Masopust (University of Minnesota) and Dan Barber
(National Institute of Allergy and Infectious Diseases) for help with the i.v.
staining protocol.
DisclosuresThe authors have no financial conflicts of interest.
10 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

References1. Kaye, P., and P. Scott. 2011. Leishmaniasis: complexity at the host-pathogen
interface. Nat. Rev. Microbiol. 9: 604–615.2. Sundar, S., and J. Chakravarty. 2015. An update on pharmacotherapy for
leishmaniasis. Expert Opin. Pharmacother. 16: 237–252.3. Giorgobiani, E., N. Chitadze, G. Chanturya, M. Grdzelidze, R. C. Jochim,
A. Machablishvili, T. Tushishvili, Y. Zedginidze, M. K. Manjgaladze,N. Iashvili, et al. 2011. Epidemiologic aspects of an emerging focus of visceralleishmaniasis in Tbilisi, Georgia. PLoS Negl. Trop. Dis. 5: e1415.
4. Varani, S., R. Cagarelli, F. Melchionda, L. Attard, C. Salvadori, A. C. Finarelli,G. A. Gentilomi, R. Tigani, R. Rangoni, R. Todeschini, et al. 2013. Ongoingoutbreak of visceral leishmaniasis in Bologna Province, Italy, November 2012 toMay 2013. Euro Surveill. 18: 20530.
5. Gramiccia, M., A. Scalone, T. Di Muccio, S. Orsini, E. Fiorentino, andL. Gradoni. 2013. The burden of visceral leishmaniasis in Italy from 1982 to2012: a retrospective analysis of the multi-annual epidemic that occurred from1989 to 2009. Euro Surveill. 18: 20535.
6. Arce, A., A. Estirado, M. Ordobas, S. Sevilla, N. Garcia, L. Moratilla, S. de laFuente, A. M. Martinez, A. M. Perez, E. Aranguez, et al. 2013. Re-emergence ofleishmaniasis in Spain: community outbreak in Madrid, Spain, 2009 to 2012.Euro Surveill. 18: 20546.
7. Palatnik-de-Sousa, C. B., and M. J. Day. 2011. One Health: the global challengeof epidemic and endemic leishmaniasis. Parasit. Vectors 4: 197.
8. Melby, P. C. 1991. Experimental leishmaniasis in humans: review. Rev. Infect.Dis. 13: 1009–1017.
9. Schlein, Y., A. Warburg, L. F. Schnur, S. M. Le Blancq, and A. E. Gunders. 1984.Leishmaniasis in Israel: reservoir hosts, sandfly vectors and leishmanial strainsin the Negev, Central Arava and along the Dead Sea. Trans. R. Soc. Trop. Med.Hyg. 78: 480–484.
10. Nadim, A., E. Javadian, and M. Mohebali. 1997. The experience of leishmani-zation in the Islamic Republic of Iran. East. Mediterr. Health J. 3: 284–289.
11. Peters, N. C., N. Kimblin, N. Secundino, S. Kamhawi, P. Lawyer, andD. L. Sacks. 2009. Vector transmission of leishmania abrogates vaccine-inducedprotective immunity. PLoS Pathog. 5: e1000484.
12. Peters, N. C., S. Bertholet, P. G. Lawyer, M. Charmoy, A. Romano, F. L. Ribeiro-Gomes, L. W. Stamper, and D. L. Sacks. 2012. Evaluation of recombinantLeishmania polyprotein plus glucopyranosyl lipid A stable emulsion vaccinesagainst sand fly-transmitted Leishmania major in C57BL/6 mice. J. Immunol.189: 4832–4841.
13. Khamesipour, A., A. Abbasi, A. Firooz, A. M. Mohammadi, S. E. Eskandari, andM. R. Jaafari. 2012. Treatment of cutaneous lesion of 20 years’ duration causedby leishmanization. Indian J. Dermatol. 57: 123–125.
14. Nadim, A., E. Javadian, G. Tahvildar-Bidruni, and M. Ghorbani. 1983. Effec-tiveness of leishmanization in the control of cutaneous leishmaniasis. Bull. Soc.Pathol. Exot. 76: 377–383.
15. Goto, Y., A. Bhatia, V. S. Raman, H. Liang, R. Mohamath, A. F. Picone,S. E. Vidal, T. S. Vedvick, R. F. Howard, and S. G. Reed. 2011. KSAC, the firstdefined polyprotein vaccine candidate for visceral leishmaniasis. Clin. VaccineImmunol. 18: 1118–1124.
16. Duthie, M. S., V. S. Raman, F. M. Piazza, and S. G. Reed. 2012. The develop-ment and clinical evaluation of second-generation leishmaniasis vaccines. Vac-cine 30: 134–141.
17. Zijlstra, E. E., A. M. el-Hassan, A. Ismael, and H. W. Ghalib. 1994. Endemickala-azar in eastern Sudan: a longitudinal study on the incidence of clinical andsubclinical infection and post-kala-azar dermal leishmaniasis. Am. J. Trop. Med.Hyg. 51: 826–836.
18. Khalil, E. A., E. E. Zijlstra, P. A. Kager, and A. M. El Hassan. 2002. Epide-miology and clinical manifestations of Leishmania donovani infection in twovillages in an endemic area in eastern Sudan. Trop. Med. Int. Health 7: 35–44.
19. Ranasinghe, S., W. W. Zhang, R. Wickremasinghe, P. Abeygunasekera,V. Chandrasekharan, S. Athauda, S. Mendis, S. Hulangamuwa, G. Matlashewski,and F. Pratlong. 2012. Leishmania donovani zymodeme MON-37 isolated froman autochthonous visceral leishmaniasis patient in Sri Lanka. Pathog. Glob.Health 106: 421–424.
20. Manson-Bahr, P. E., R. B. Heisch, and P. C. Garnham. 1959. Studies in leish-manifasis in East Africa. IV. The Montenegro test in kala-azar in Kenya. Trans.R. Soc. Trop. Med. Hyg. 53: 380–383.
21. Senekji, H. A. 1943. Hematologic and immunologic studies on natural and in-duced leishmaniasis in paretics. Am. J. Trop. Med. Hyg. 23: 53–58.
22. Adler, S., and A. E. Gunders. 1964. Immunity to Leishmania mexicana followingspontaneous recovery from oriental sore. Trans. R. Soc. Trop. Med. Hyg. 58: 274–277.
23. Manson-Bahr, P. E. 1961. Immunity in kala-azar. Trans. R. Soc. Trop. Med. Hyg.55: 550–555.
24. Streit, J. A., T. J. Recker, F. G. Filho, S. M. Beverley, and M. E. Wilson. 2001.Protective immunity against the protozoan Leishmania chagasi is induced bysubclinical cutaneous infection with virulent but not avirulent organisms. J.Immunol. 166: 1921–1929.
25. Nation, C. S., B. Dondji, and G. A. Stryker. 2012. Previous exposure to a lowinfectious dose of Leishmania major exacerbates infection with Leishmaniainfantum in the susceptible BALB/c mouse. Parasitol. Res. 111: 1407–1415.
26. Sacks, D., and N. Noben-Trauth. 2002. The immunology of susceptibility andresistance to Leishmania major in mice. Nat. Rev. Immunol. 2: 845–858.
27. Ahmed, S., M. Colmenares, L. Soong, K. Goldsmith-Pestana, L. Munstermann,R. Molina, and D. McMahon-Pratt. 2003. Intradermal infection model forpathogenesis and vaccine studies of murine visceral leishmaniasis. Infect.Immun. 71: 401–410.
28. Romano, A., E. Inbar, A. Debrabant, M. Charmoy, P. Lawyer, F. Ribeiro-Gomes,M. Barhoumi, M. Grigg, J. Shaik, D. Dobson, et al. 2014. Cross-species geneticexchange between visceral and cutaneous strains of Leishmania in the sand flyvector. Proc. Natl. Acad. Sci. USA 111: 16808–16813.
29. Spath, G. F., and S. M. Beverley. 2001. A lipophosphoglycan-independentmethod for isolation of infective Leishmania metacyclic promastigotes by den-sity gradient centrifugation. Exp. Parasitol. 99: 97–103.
30. Sacks, D. L., S. Hieny, and A. Sher. 1985. Identification of cell surface carbo-hydrate and antigenic changes between noninfective and infective developmentalstages of Leishmania major promastigotes. J. Immunol. 135: 564–569.
31. Ramalingam, T. R., J. T. Pesce, F. Sheikh, A. W. Cheever, M. M. Mentink-Kane,M. S. Wilson, S. Stevens, D. M. Valenzuela, A. J. Murphy, G. D. Yancopoulos,et al. 2008. Unique functions of the type II interleukin 4 receptor identified inmice lacking the interleukin 13 receptor a1 chain. Nat. Immunol. 9: 25–33.
32. Anderson, K. G., K. Mayer-Barber, H. Sung, L. Beura, B. R. James, J. J. Taylor,L. Qunaj, T. S. Griffith, V. Vezys, D. L. Barber, and D. Masopust. 2014. Intra-vascular staining for discrimination of vascular and tissue leukocytes. Nat.Protoc. 9: 209–222.
33. Kimblin, N., N. Peters, A. Debrabant, N. Secundino, J. Egen, P. Lawyer,M. P. Fay, S. Kamhawi, and D. Sacks. 2008. Quantification of the infectious doseof Leishmania major transmitted to the skin by single sand flies. Proc. Natl.Acad. Sci. USA 105: 10125–10130.
34. Zaph, C., J. Uzonna, S. M. Beverley, and P. Scott. 2004. Central memory T cellsmediate long-term immunity to Leishmania major in the absence of persistentparasites. Nat. Med. 10: 1104–1110.
35. Peters, N. C., A. J. Pagan, P. G. Lawyer, T. W. Hand, E. Henrique Roma,L. W. Stamper, A. Romano, and D. L. Sacks. 2014. Chronic parasitic infectionmaintains high frequencies of short-lived Ly6C+CD4+ effector T cells that arerequired for protection against re-infection. PLoS Pathog. 10: e1004538.
36. Goto, Y., A. Bhatia, V. S. Raman, S. E. Vidal, S. Bertholet, R. N. Coler,R. F. Howard, and S. G. Reed. 2009. Leishmania infantum sterol 24-c-methyltransferase formulated with MPL-SE induces cross-protection againstL. major infection. Vaccine 27: 2884–2890.
37. Sakai, S., K. D. Kauffman, J. M. Schenkel, C. C. McBerry, K. D. Mayer-Barber,D. Masopust, and D. L. Barber. 2014. Cutting edge: control of Mycobacteriumtuberculosis infection by a subset of lung parenchyma-homing CD4 T cells. J.Immunol. 192: 2965–2969.
38. Okwor, I. B., P. Jia, Z. Mou, C. Onyilagha, and J. E. Uzonna. 2014. CD8+ T cellsare preferentially activated during primary low dose leishmania major infectionbut are completely dispensable during secondary anti-Leishmania immunity.PLoS Negl. Trop. Dis. 8: e3300.
39. Uzonna, J. E., K. L. Joyce, and P. Scott. 2004. Low dose Leishmania majorpromotes a transient T helper cell type 2 response that is down-regulated byinterferon g-producing CD8+ T cells. J. Exp. Med. 199: 1559–1566.
40. Stager, S., and S. Rafati. 2012. CD8+ T cells in leishmania infections: friends orfoes? Front. Immunol. 3: 5.
41. Novais, F. O., L. P. Carvalho, J. W. Graff, D. P. Beiting, G. Ruthel, D. S. Roos,M. R. Betts, M. H. Goldschmidt, M. E. Wilson, C. I. de Oliveira, and P. Scott.2013. Cytotoxic T cells mediate pathology and metastasis in cutaneous leish-maniasis. PLoS Pathog. 9: e1003504.
42. Bunn, P. T., A. C. Stanley, F. de Labastida Rivera, A. Mulherin, M. Sheel,C. E. Alexander, R. J. Faleiro, F. H. Amante, M. Montes De Oca, S. E. Best, et al.2014. Tissue requirements for establishing long-term CD4+ T cell-mediatedimmunity following Leishmania donovani infection. J. Immunol. 192: 3709–3718.
43. Eisert, V., U. M€unster, M. M. Simon, and H. Moll. 2002. The course of Leish-mania major infection in mice lacking granzyme-mediated mechanisms.Immunobiology 205: 314–320.
44. Faria, D. R., P. E. Souza, F. V. Duraes, E. M. Carvalho, K. J. Gollob,P. R. Machado, and W. O. Dutra. 2009. Recruitment of CD8+ T cells expressinggranzyme A is associated with lesion progression in human cutaneous leish-maniasis. Parasite Immunol. 31: 432–439.
45. Santos, Cda. S., V. Boaventura, C. Ribeiro Cardoso, N. Tavares, M. J. Lordelo,A. Noronha, J. Costa, V. M. Borges, C. I. de Oliveira, J. Van Weyenbergh, et al.2013. CD8+ granzyme B+-mediated tissue injury vs. CD4+IFNg+-mediatedparasite killing in human cutaneous leishmaniasis. J. Invest. Dermatol. 133:1533–1540.
46. McCall, L. I., W. W. Zhang, S. Ranasinghe, and G. Matlashewski. 2013.Leishmanization revisited: immunization with a naturally attenuated cutaneousLeishmania donovani isolate from Sri Lanka protects against visceral leish-maniasis. Vaccine 31: 1420–1425.
47. Gicheru, M. M., J. O. Olobo, and C. O. Anjili. 1997. Heterologous protection byLeishmania donovani for Leishmania major infections in the vervet monkeymodel of the disease. Exp. Parasitol. 85: 109–116.
48. Alexander, J., and R. S. Phillips. 1978. Leishmania tropica and Leishmaniamexicana: cross-immunity in mice. Exp. Parasitol. 45: 93–100.
49. Perez, H., B. Arredondo, and R. Machado. 1979. Leishmania mexicana andLeishmania tropica: cross immunity in C57BL/6 mice. Exp. Parasitol. 48: 9–14.
50. Lainson, R., and J. J. Shaw. 1977. Leishmaniasis in Brazil: XII. Observations oncross-immunity in monkeys and man infected with Leishmania mexicana mex-icana, L. m. amazonensis, L. braziliensis braziliensis, L. b. guyanensis and L. b.panamensis. J. Trop. Med. Hyg. 80: 29–35.
51. Porrozzi, R., A. Teva, V. F. Amaral, M. V. Santos da Costa, and G. Grimaldi, Jr.2004. Cross-immunity experiments between different species or strains ofLeishmania in rhesus macaques (Macaca mulatta). Am. J. Trop. Med. Hyg. 71:297–305.
The Journal of Immunology 11
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

52. Dey, R., P. K. Dagur, A. Selvapandiyan, J. P. McCoy, P. Salotra, R. Duncan, andH. L. Nakhasi. 2013. Live attenuated Leishmania donovani p27 gene knockoutparasites are nonpathogenic and elicit long-term protective immunity in BALB/cmice. J. Immunol. 190: 2138–2149.
53. Dey, R., G. Natarajan, P. Bhattacharya, H. Cummings, P. K. Dagur, C. Terrazas,A. Selvapandiyan, J. P. McCoy, Jr., R. Duncan, A. R. Satoskar, andH. L. Nakhasi. 2014. Characterization of cross-protection by geneticallymodified live-attenuated Leishmania donovani parasites against Leishmaniamexicana. J. Immunol. 193: 3513–3527.
54. Mizbani, A., T. Taheri, F. Zahedifard, Y. Taslimi, H. Azizi, K. Azadmanesh,B. Papadopoulou, and S. Rafati. 2009. Recombinant Leishmania tarentolaeexpressing the A2 virulence gene as a novel candidate vaccine against visceralleishmaniasis. Vaccine 28: 53–62.
55. Rochette, A., F. Raymond, J. M. Ubeda, M. Smith, N. Messier, S. Boisvert,P. Rigault, J. Corbeil, M. Ouellette, and B. Papadopoulou. 2008. Genome-widegene expression profiling analysis of Leishmania major and Leishmania infan-tum developmental stages reveals substantial differences between the two spe-cies. BMC Genomics 9: 255.
56. Peacock, C. S., K. Seeger, D. Harris, L. Murphy, J. C. Ruiz, M. A. Quail,N. Peters, E. Adlem, A. Tivey, M. Aslett, et al. 2007. Comparative genomicanalysis of three Leishmania species that cause diverse human disease. Nat.Genet. 39: 839–847.
57. Singh, O. P., E. Hasker, D. Sacks, M. Boelaert, and S. Sundar. 2014. Asymp-tomatic Leishmania infection: a new challenge for Leishmania control. Clin.Infect. Dis. 58: 1424–1429.
58. Teijaro, J. R., D. Turner, Q. Pham, E. J. Wherry, L. Lefrancois, and D. L. Farber.2011. Cutting edge: tissue-retentive lung memory CD4 T cells mediate optimalprotection to respiratory virus infection. J. Immunol. 187: 5510–5514.
59. Glennie, N. D., V. A. Yeramilli, D. P. Beiting, S. W. Volk, C. T. Weaver, andP. Scott. 2015. Skin-resident memory CD4+ T cells enhance protection againstLeishmania major infection. J. Exp. Med. 212: 1405–1414.
60. Iijima, N., and A. Iwasaki. 2014. T cell memory. A local macrophage chemokinenetwork sustains protective tissue-resident memory CD4 T cells. Science 346:93–98.
61. Gebhardt, T., P. G. Whitney, A. Zaid, L. K. Mackay, A. G. Brooks, W. R. Heath,F. R. Carbone, and S. N. Mueller. 2011. Different patterns of peripheral mi-gration by memory CD4+ and CD8+ T cells. Nature 477: 216–219.
62. Mueller, S. N., T. Gebhardt, F. R. Carbone, and W. R. Heath. 2013. MemoryT cell subsets, migration patterns, and tissue residence. Annu. Rev. Immunol. 31:137–161.
63. Selvapandiyan, A., R. Dey, S. Nylen, R. Duncan, D. Sacks, and H. L. Nakhasi.2009. Intracellular replication-deficient Leishmania donovani induces longlasting protective immunity against visceral leishmaniasis. J. Immunol. 183:1813–1820.
64. Zhang, W. W., and G. Matlashewski. 2001. Characterization of the A2–A2relgene cluster in Leishmania donovani: involvement of A2 in visceralizationduring infection. Mol. Microbiol. 39: 935–948.
65. Mendez, S., K. Tabbara, Y. Belkaid, S. Bertholet, D. Verthelyi, D. Klinman,R. A. Seder, and D. L. Sacks. 2003. Coinjection with CpG-containing immu-nostimulatory oligodeoxynucleotides reduces the pathogenicity of a live vaccineagainst cutaneous leishmaniasis but maintains its potency and durability. Infect.Immun. 71: 5121–5129.
66. Silvestre, R., A. Cordeiro-Da-Silva, N. Santarem, B. Vergnes, D. Sereno,and A. Ouaissi. 2007. SIR2-deficient Leishmania infantum induces a definedIFN-g/IL-10 pattern that correlates with protection. J. Immunol. 179: 3161–3170.
67. Liu, D., I. Okwor, Z. Mou, S. M. Beverley, and J. E. Uzonna. 2013. Deficiency ofLeishmania phosphoglycans influences the magnitude but does not affect thequality of secondary (memory) anti-Leishmania immunity. PLoS One 8: e66058.
68. Davoudi, N., A. Khamesipour, F. Mahboudi, and W. R. McMaster. 2014. A dualdrug sensitive L. major induces protection without lesion in C57BL/6 mice.PLoS Negl. Trop. Dis. 8: e2785.
69. Uzonna, J. E., G. Wei, D. Yurkowski, and P. Bretscher. 2001. Immune elimi-nation of Leishmania major in mice: implications for immune memory, vacci-nation, and reactivation disease. J. Immunol. 167: 6967–6974.
70. Belkaid, Y., K. F. Hoffmann, S. Mendez, S. Kamhawi, M. C. Udey, T. A. Wynn,and D. L. Sacks. 2001. The role of interleukin (IL)-10 in the persistence ofLeishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 194: 1497–1506.
71. Kamhawi, S., M. Ramalho-Ortigao, V. M. Pham, S. Kumar, P. G. Lawyer,S. J. Turco, C. Barillas-Mury, D. L. Sacks, and J. G. Valenzuela. 2004. A role forinsect galectins in parasite survival. Cell 119: 329–341.
72. Secundino, N., N. Kimblin, N. C. Peters, P. Lawyer, A. A. Capul, S. M. Beverley,S. J. Turco, and D. Sacks. 2010. Proteophosphoglycan confers resistance ofLeishmania major to midgut digestive enzymes induced by blood feeding invector sand flies. Cell. Microbiol. 12: 906–918.
73. Dobson, D. E., S. Kamhawi, P. Lawyer, S. J. Turco, S. M. Beverley, andD. L. Sacks. 2010. Leishmania major survival in selective Phlebotomus papatasisand fly vector requires a specific SCG-encoded lipophosphoglycan gal-actosylation pattern. PLoS Pathog. 6: e1001185.
12 CUTANEOUS LEISHMANIASIS PREVENTS VISCERAL LEISHMANIASIS
by guest on April 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















