
Dieting for Muscle
119
Brad Pilon
-
Upload
francesco-de-giorgio -
Category
Documents
-
view
58 -
download
1
description
Dieting advice for muscle growth
Transcript of Dieting for Muscle

Brad Pilon

Copyright © 2013 by StrengthWorks International Publishing, Inc.
All rights Reserved
No portion of this book may be used, reproduced, or transmitted in any form or by any means, electronic or mechanical, including fax, photocopy, recording, or any information storage and retrieval system by anyone but the purchaser for their own personal use. This manual may not be reproduced in any form without the express written permission of Brad Pilon, except in the case of a reviewer who wishes to quote brief passages for the sake of a review written for inclusion in a magazine, newspaper, or journal, and all of these situations require the written approval of Brad Pilon prior to publication.
The information in this book is for educational purposes only. The information in this book is based on my own personal experiences and my own interpretation of available research. It is not medical advice and I am not a medical doctor.
The information within this book is meant for healthy adult individuals. You should consult with your physician to make sure it is appropriate for your individual circumstances. Keep in mind that nutritional needs vary from person to person, depending on age, sex, health status and total diet.
If you have any health issues or concerns please consult with your physician. Always consult your physician before beginning or making any changes in your diet or exercise program, for diagnosis and treatment of illness and injuries, and for advice regarding medications.



Your body has build-in regulators to prevent massive increases in muscle growth.
When we overeat our body turns on counter-regulatory factors to prevent the build-up of muscle mass.
Some Examples:
Overeating causes decreased testosterone levels in men
Overeating causes decreased insulin sensitivity in men and women.
Overeating also causes increased low grade systemic inflammation
While some inflammation is good for you, low grade systemic inflammation blocks muscle growth and is associated with a host of chronic diseases.



Latin: inflammare, to set on fire
There are two main types of inflammation.
Acute and Chronic
Acute Inflammation is a biological response to harmful stimuli
It is how your body removes the injurious stimuli and initiates the healing process
Without acute inflammation, wounds and infections would never heal
However, long term chronic inflammation is associated with many different disease states, including obesity, and the loss of muscle mass.
Especially when the chronic inflammation is systemic – throughout your whole body.


Other than the obvious health reasons (prevention of disease), you should also care about inflammation because it plays an important role in regulating muscle growth.
Based on the ‘Signal and Noise’ theory of inflammation’s role in regulating muscle growth, inflammation can both cause and prevent muscle growth.
It is also how many different lifestyle factors (like chronic endurance training) can actually prevent optimal muscle growth.
The role of inflammation in the muscle growth depends on whether or not the inflammation is systemic or localized.
Simply put - the Inflammation Theory of Muscle growth is as follows:

The Inflammation Theory of Muscle growth:

Above: The Acute Inflammation Response to periods of Resistance Training in a person with no chronic Inflammation. Each spike represents the inflammation caused by a workout. The spikes are large enough to initiate the signal for new muscle growth.
Below: The Same Acute Inflammation Response to periods of Resistance Training in a person WITH chronic inflammation. The Spikes are hidden by the ‘noise’ and thus muscle growth is not initiated.
Chronic inflammation is the body’s response to a chronic harmful stimuli, such as excess body fat or other chronic metabolic ‘insults’.
Low grade chronic inflammation is characterized by a 2 to 3 fold increase in the systemic concentrations of cytokines such as TNF-alpha, IL-6 and C-reactive Protein.
This type of inflammation is systemic, meaning it is not localized to any one part of your body.
In the case of Obesity the extra fat is the ‘harmful stimuli’ that is causing the inflammation; however our bodies cannot get rid of the fat with the inflammation response and thus will stay inflamed as long as the extra fat is still present.


Not only can obesity cause chronic inflammation, but the very act of overeating can also lead to systemic (whole body) inflammation.
A single large meal is enough to start to raise many markers of inflammation.
This inflammatory response is larger in an overweight person then it is in a lean person.
Having excess body fat can cause chronic inflammation AND increases the inflammatory response to a large meal.
In other words an overweight person will have a higher systemic inflammation response to a high calorie meal then a lean person.



Cytokines are small cell-signaling protein molecules
Typically cytokines are immunomodulating agents, such as interleukins and interferons.
Basically, a cytokine is either inflammatory or anti-inflammatory.
3 Common types of inflammatory cytokines: TNF-Alpha, IL-6 and CRP
TNF-Alpha is proposed as the main driver of chronic low grade inflammation
IL-6 increases several fold post-exercise after exhaustive exercise
C-Reactive Protein - acute-phase protein, elevated in inflammation and illnesses such as cancer

Muscle growth isn’t the only reason to keep low grade inflammation at bay.
Low grade systemic inflammation is also associated with many diseases, including:
Acne,
Rheumatoid arthritis,
Hypertension,
atherosclerosis,
Fatty liver,
Asthma,
Insulin resistance and diabetes,
Cardiovascular disease,
Alzheimer Disease,
And even the aging process itself.

Chronic Inflammation is a process that seems to be an underlying mechanism in many forms of cancer
It is also a possible link between obesity and cancer.
Surprisingly, this inflammation-cancer link was suggested as far back as the late 1800’s when German Pathologist Rudolf Virchow stated:
“Chronic irritation which is manifested by a chronic
inflammation is a key promoter of cancer.”


Chronic inflammation is widely observed in obesity.
People who are obese commonly have many elevated markers of systemic low-grade inflammation, including:
IL-6,
TNF-Alpha,
CRP,
Insulin,
Blood glucose,
Leptin
IL-18
This relationship also seems to be dose-dependent, meaning the more obese a person is the more inflammation they tend to produce.

One of the most important predictors of low-grade inflammation is your Waist circumference.
Waist circumferences over 50% of height are associated with low-grade inflammation and this relationship increases as waist circumference increases.
Adipose tissue has been shown to produce 10-35% of IL-6 in a resting individual, and this production increases with increasing adiposity.
Thus there is a link between increasing levels of obesity, and increasing levels of inflammation.
Within reason, the lower the body fat, the lower the overall levels of systemic inflammation.


To make matters even worse, obesity increases inflammation, which in turn promotes fat cells to secrete even more inflammatory cytokines.
Inactivity and chronic overeating only makes this problem worse.
Inflammation leads to lowered testosterone, insulin resistance, and possibly leptin resistance.
Excess fat leads to inflammation, which leads to more fat, and more fat leads to more inflammation.
This sets up the perfect downward spiral into disease (And definitely a complete lack of muscle growth potential).


Having high levels of blood sugar is called Hyperglycemia – and it occurs after a large sugar containing meal.
Hyperglycemia induces the release of the inflammatory cytokine IL-6 into your blood stream.
Thus high carbohydrate meals are associated with an increase in inflammation.
Meals containing high levels of saturated fat have also been associated with increased markers of inflammation.
High calorie meals containing carbohydrates, fats and proteins have been associated with increasing inflammation.
This response is elevated in people who are obese.




The acute inflammation response is the bodies response to insult and injury
(Think Bee sting)
It Involves the mobilization of Cytokines and other immune molecules to protect the body.
Role of acute inflammation is to remove the injurious stimuli and damaged tissue and to initiate the healing process
It is localized (happens in one spot), as opposed to systemic low grade inflammation which happens all throughout your body.

Muscular contractions from a weight training workout are what sensitizes a muscle and allow it to increase in size.
After a person has grown to their full adult size, this is the only way to induce muscle growth, outside of using anabolic drugs.
This has been shown in research using:
Cross Sectional Area
Fractional Synthetic Rate
Amino Acid Markers
Increases in Muscular Strength
Increases in Muscular Weight
The right amount of work at the right effort can sensitize a muscle to the the anabolic signal that comes from eating a meal containing protein.
In other words it is weight training that makes eating protein effective at building muscle.

Workouts alone do not build muscle. They sensitize the muscle by initiating the anabolic process.
Eating protein alone will not build muscle. The muscle must be sensitized for eating protein to contribute to the muscle growth process.’
Working out sensitizes a muscle, allowing it to grow when given a high amount of amino acids.
This sensitization lasts for over 24 hours.
However, it is the ‘signal’ that sensitizes the muscle that we are interested in.


There is a known relationship between inflammation and muscle growth.
Acute localized inflammation is a response to muscular contractions and is necessary for muscle growth. It ‘starts’ the repair process.
Muscles can synthesize cytokines in response to contractions. So the inflammation that comes from a workout is localized – it happens inside the muscle that was exercised.
High doses of anti-inflammatory medicines are able to blunt muscle growth by blocking this inflammation response, so we know that it is a necessary component of muscle growth.

The inflammation response caused by a workout plays a role in the degeneration and regeneration process of muscle and surrounding connective tissue.
As a response to muscular contractions the acute inflammatory response initiates the breakdown and removal of damaged muscle tissue – the ‘good’ type of protein catabolism.
Elevations in localized inflammation can last up to 5 days after a workout, adding evidence to the fact that muscles continue to be sensitive to the anabolic affects of eating protein for an extended period after a workout.

The cytokine IL-6 is an important marker of acute inflammation.
It is released by muscle cells as a response to muscular contraction.
This increase can be up to 100 times above resting levels.
The amount of IL-6 released depends on total workload during the workout and intensities of the workout.
The IL-6 increase caused by a workout starts to increases about 4 hours after the workout and remains elevated well passed 24 hours after the workout.
How long IL-6 remains elevated also depends on the volume of work and the amount of effort or strain caused by the workout.


*The inflammation response from a workout causes increased glucose uptake, fat oxidation and satellite cell activation.

Satellite cells are like ‘dormant muscle seeds’ that sit on the outside of the muscle fiber (hence the name satellite cells).
They are present in all skeletal muscles and are associated with all muscle fiber types - albeit with unequal distribution (some muscles have more some have less)
You can think of satellite cells as the body’s reserve of muscle cell nuclei.
Satellite cells are crucial for skeletal muscle adaption to exercise.
Without satellite cell activation there would be very little possibility for muscle growth in the human body.


Satellite cells contribute to hypertrophy by providing the new myonuclei needed to repair damaged segments of mature muscle fibers for successful regeneration following injury or exercise induced muscle damage.
High level power-lifters can have up to 100% more satellite cells than untrained controls.
Supplemental testosterone also increases satellite cell number.
Without satellite cell activation muscle growth cannot occur.
Once a satellite cell has been incorporated into a muscle fiber it stays there for years, even without exercise, allowing the muscle to keep a memory of its previous size (this is often referred to as ‘muscle memory’)

Scientists have tested the importance of satellite cells by using low dose radiation in animal studies to destroy the satellite cells in one leg.
When this happens the muscles exposed to radiation can still adapt to exercise by becoming more efficient, but with the exception of a small amount of beginner growth, they simply cannot increase their size.
In other words, because they could not add more nuclei, they could not increase their size in response to resistance training.
Adding more exercise, or high doses of amino acids cannot induce muscle growth without the help of satellite cells acting as the ‘brain’ that puts it all together.

Satellite cells have receptors that detect localized acute inflammation, (IL-6 receptors) and these receptors increase following a workout.
The inflammation marker IL-6 has been shown to mediate muscle growth.
Unaccustomed exercise can increase IL-6 by up to 6 fold at 5 hours post exercise and 3 fold 8 days after exercise.
When Exercise induced inflammation is blocked by taking high doses of anti-inflammatory drugs satellite cells cannot response to exercise stimulus and post-exercise protein synthesis is decreased.




STEP 1: Weight-lifting workout
STEP 2: The Acute Inflammation Response
STEP 3: Activation of Satellite Cells
STEP 4: Multiple Protein feedings and Recovery Time
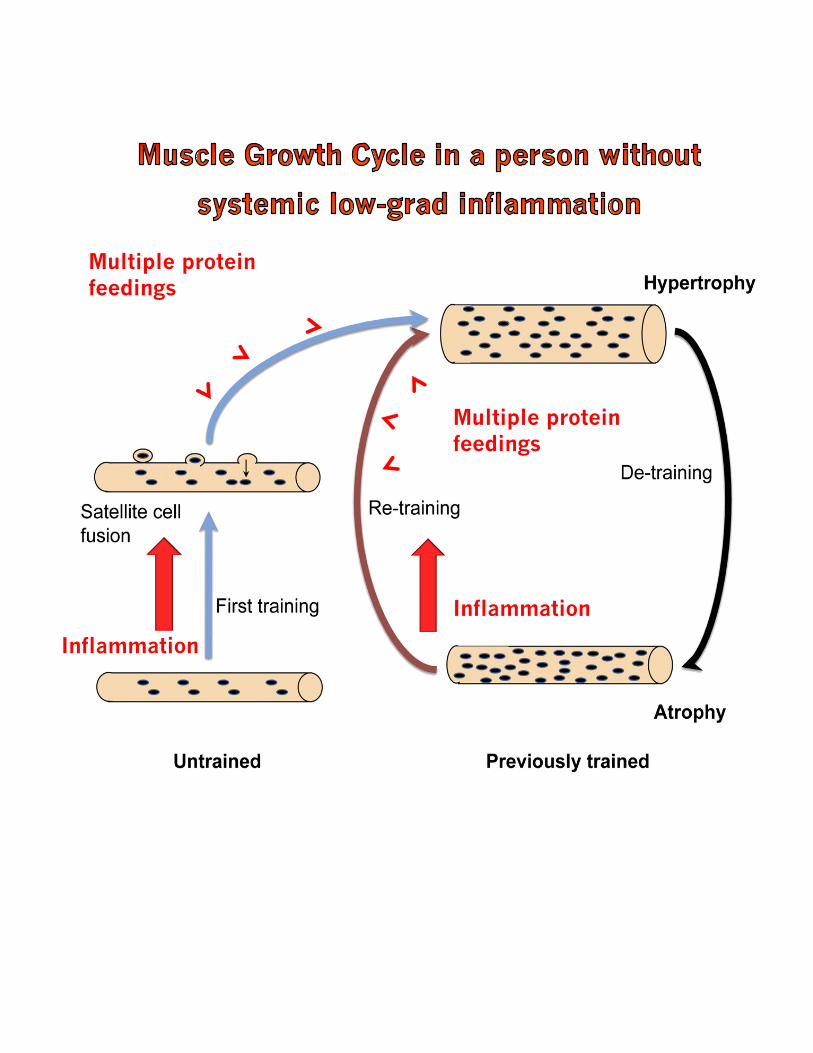
Inflammation
Multiple protein feedings
Inflammation
Multiple protein feedings


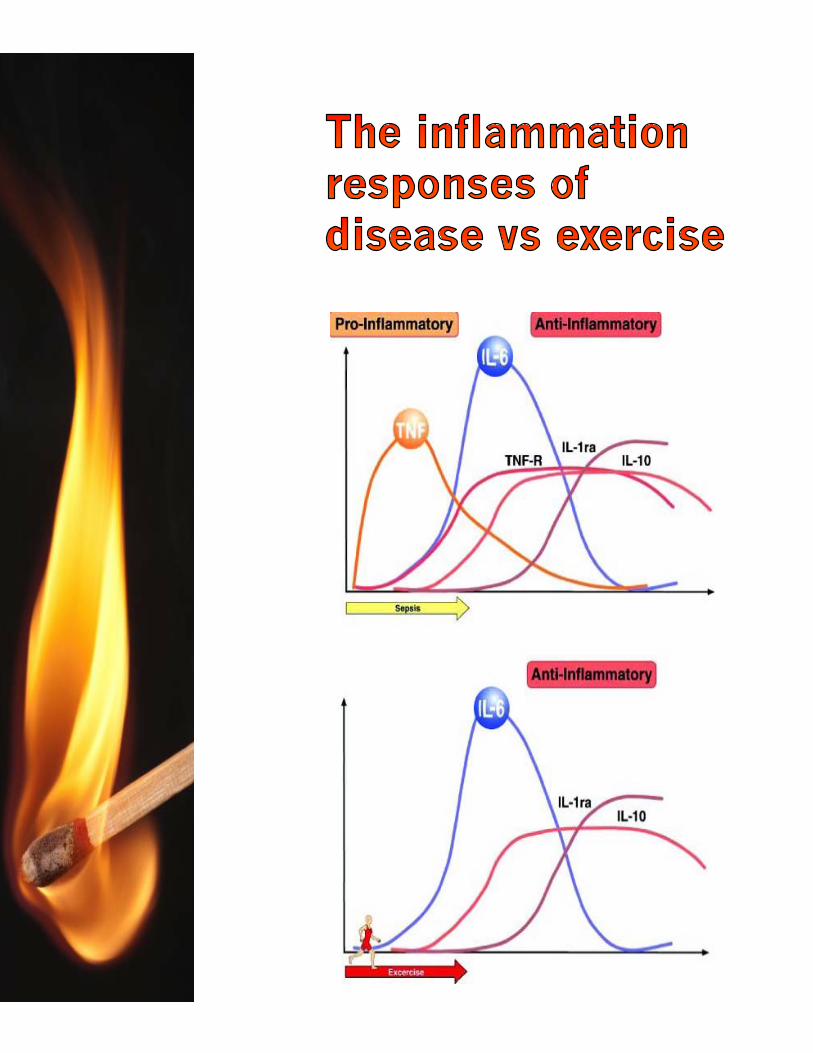
Both resistance training (weight lifting) and endurance training (long distance running) increase inflammation.
However, the response is uniquely different.
The cytokine response to endurance training such as running is slightly lower and more pronged, not reaching its full peak until the 12-24 hour mark.
While both types of exercise stimulate inflammation, the different ways in which inflammation is stimulated changes how each one is able to stimulate muscle growth.
Some form of resistance training must be completed on a consistent basis for muscle growth to occur.


Even a small increase in chronic inflammation can increase the risk of muscle strength loss and cause a decrease in your ability to build muscle.
Chronic inflammation has been implicated as part of the cause of the muscle loss that occurs with aging (sarcopenia).
Increased protein levels of myostatin have been described in patients with diseases characterized by chronic low-grade inflammation, limiting their ability to build muscle.
Increased levels ofinflammation can suppress the AKT/mTOR pathway - an important pathway in the muscle building process.
Inflammation may decrease the anabolic effects of IGF-1.

Sepsis is an extreme whole body inflammatory state that is able to inhibit the synthesis of both myofibrillar and sarcoplasmic proteins preferentially in muscles composed of fast twitch fibers.
(Sepsis prevents the creation of new muscle proteins)
Sepsis is able to prevent leucine from stimulating muscle protein synthesis.
In animal models when inflammation is created mTOR loses it’s ability to be stimulated by muscle growth



STEP 1: Weight-lifting workout
STEP 2: The Acute Inflammation Response
STEP 3: Activation of Satellite Cells
STEP 4: Multiple Protein feedings and Recovery Time
Blocked by Chronic Inflammation



Research has identified 6 major sources of inflammation that are considered ‘lifestyle factors’ since they do not come from a disease:
Chronic Exhaustive Exercise
Stress
Aging
Lack of Sleep
Overeating
Obesity
FIVE of these sources must be addressed for optimal muscle growth (Sadly, we can’t control aging)



Body fat:
Excess body fat is one of the strongest predictors of overall chronic inflammation, and the inflammation response to high calorie meals.
In order to keep low-grad inflammation at bay, keep your waist circumference below 50% of your height – especially when attempting to bulk.
Occasional periods of low calorie intake will serve to keep inflammation low, and keep your waist within acceptable limits.
TO OPTIMIZE MUSCLE GROWTH:
Keep your waist circumference UNDER 50% of your height at all times if you are trying to build muscle.


Weight Loss achieved through different diet programs with or without exercise resulted in decreases of markers of low grade inflammation by 7 to 48%.
Calorie restriction is anti-inflammatory, this includes brief periods of fasting.
Weight loss is effective at reducing inflammatory markers, specifically IL-6 and Leptin
Losing weight and maintaining a ‘healthy’ weight is your best solution for minimizing inflammation levels.
Maintain a ‘healthy’ weight is also your best solution for building muscle mass.
TO OPTIMIZE MUSCLE GROWTH:
Do not gain fat in an attempt to ‘force’ muscle growth. It does not work and you will regret it.


OVEREATING:
Extremely large meals cause an inflammation response, and this response is heightened in people who are already obese.
For the purpose of muscle gains, keep cheat days or cheat meals to a minimum until your waist circumference is under 50% of your height.
Even then, use this dieting technique sparingly if you are trying to gain muscle mass.
TO OPTIMIZE MUSCLE GROWTH:
No cheat days until your waist circumference is under 50% of your height.



LACK OF SLEEP:
Do not underestimate the power of sleep in the recovery process.
Sleep is far more important then massive amounts of calories or even overeating protein for recovery after a workout.
Lack of sleep (not enough in a night) or disrupted sleep (multiple periods of wakefulness during the night) area associated with: Increased inflammation
Decreased insulin sensitivity Overeating Weight gain
TO OPTIMIZE MUSCLE GROWTH:
If you are having trouble sleeping consider taking the supplement ZMA before bed.


STRESS:
The effect of stress on chronic inflammation is VERY real.
Plasma levels of inflammation markers are higher in workers suffering from ‘burnout’ then in those not suffering ‘burnout’
Elevated markers of inflammation are associated with excessive daytime sleepiness, disturbed night time sleep and daily lethargy.
It is speculated that many of the people who claim to be suffering ‘adrenal fatigue’ are actually suffering from chronic-low grade inflammation.
TO OPTIMIZE MUSCLE GROWTH:
Inflammation, adrenal fatigue, and overtraining are all similar terms to describe and overall feeling of chronic exhaustion



Chronic Exhaustive Exercise
There more than100 original scientific reports concerning exercise and inflammation Showing a strong relationship between exhaustive exercise and chronic low grade inflammation
Marathon running may enhance IL-6 levels as much as 100 times over normal and increases total leuckocyte count and neutrophil mobilization.
Low grade inflammation caused by chronic exhaustive exercise can blunt your ability to gain muscle by blunting the inflammation response and your anabolic response to eating protein.
TO OPTIMIZE MUSCLE GROWTH:
If you are trying to gain muscle, then you should avoid exhaustive exercise, especially if you are currently overweight (waist circumference above 50% of your height)



NUTRITION:
Other than eating less, the are very little types of dietary changes that need to be made in order to lower grade systemic Inflammation.
Adjusting your protein, fat or carbohydrate intake causes very little change in markers of inflammation.
When it comes to reducing inflammation it truly is how much you eat more than it is what you eat.
Herbs and spices may have anti-inflammatory effects or at least lessen some of the negative effects of inflammation, so if you were going to make one change on top of eating less, I would recommend eating more herbs and spices on a day-to-day basis.



SUPPLEMENTS:
You must be careful with your use of supplements as some may actually decrease the acute inflammation response to exercise (possibly lowering muscle building potential).
Don’t drink carbohydrates during your workouts - Post or during workout Carbohydrate supplementation has been shown to be able to decrease the IL-6 Response to high intensity running.
Avoid taking high dose vitamin C - Daily high dose Vitamin C supplementation may also blunt the contraction induced acute inflammation.
Whey protein is a ‘safe’ supplement - Neither acute nor long-term whey protein supplementation have any effect on the major markers of inflammation.
Avoid CLA while attempting to build muscle. -Conjugated Linoleic Acid (CLA) seems to be able to increase markers of inflammation
Fish Oils are good, but do little to reduce inflammation - Studies of dose-response between fish oils and inflammation have found that even high dose (3.4 g/d of EPA and DHA) did not change inflammatory status over 8 weeks.
Irvingia gabonesis has a lot of potential - it seems to have the ability to decrease low-grade inflammation


Regular physical activity is reported to decrease markers of inflammation.
Baseline measurements of circulating inflammatory markers do not seem to differ greatly between healthy trained and untrained adults
However long-term chronic training may help reduce chronic low grade inflammation
Levels of inflammatory markers (IL-6) remained elevated longer into the recovery period following and acute bout of exercise in patients with inflammatory diseases as opposed to healthy controls.


Research has shown that there is a strong trend towards reduced post absorptive muscle protein synthesis associated with aging.
This means as we get older we get less of a muscle growth response from eating protein.
Evidence suggests that this is related to increased circulating levels of inflammatory cytokines TNF-Alpha, IL-6 and CRP
High levels of circulating IL-6 can predict muscle atrophy in the elderly
The increased inflammation associated with aging blunts our ability to gain muscle.





The affect of exercise on inflammation seems to scale the level of exhaustion associated with the exercise.
Low intensity training such as walking can reduce resting pro-inflammatory markers.
Moderate exercise can have some anti-inflammatory benefits, protecting against the development of low grade inflammation.
Strenuous or exhaustive exercise can increase inflammation.





There is a strong relationship between Testosterone and inflammation in men.
Low levels of testosterone are associated with increased levels of inflammation.
Testosterone injections result in profound declines in markers of inflammation
Testosterone is able to suppress the expression of many different inflammatory cytokines (IL-6, IL-1B and TNF-Alpha)
Testosterone can also stimulate the production of anti-inflammatory IL-10
Testosterone is able to decrease inflammation and increase anti-inflammatory markers.




Overeating has been purported to add in the muscle building process in young, non-steroid using athletes.
This does work for a short period, however this effect seems to decrease with time, leading to speculation that the slow build up of inflammation eventually reaches a point where muscle growth is blunted.
This time course would depend on the degree of overeating and speed of fat gain in the individual.
The faster the fat gain, the quicker the muscle gains stop.
However, the use of testosterone could allow for a prolonged anabolic signal from overeating, as it would prevent both the decrease in testosterone usually associated with overeating, as well as the increase in inflammation induced by overeating.
Bottom line: Don’t take bulking advice from people using steroids.


Gaining fat and grossly overeating is NOT the answer!
Controlling Inflammation is the answer.
Low grade chronic inflammation brought about by any combination of overeating, obesity, stress, lack of sleep, aging, and exhaustive exercise will blunt muscle growth signals form both exercise and diet
Decreasing chronic inflammation may allow for a return to proper anabolic signaling.
However, it seems wiping out inflammation completely also prevents the acute local inflammation needed for muscle growth (as evidenced by high does NSAID Studies)

Utilizing the signal/noise theory of the role of inflammation in muscle growth it seems very plausible that the best course of action for long term muscle growth is the opposite of what we have been lead to believe.
Keep body fat low, get lots of sleep, and avoid excessive use of exhaustive exercise for optimal muscle growth and long term health.
If your waist circumference is above 50% start with a goal of decreasing your body fat before you start to attempt to ‘eat up’ to build muscle.
Once your waist is below 50% of your height, eat as much as you can WITHOUT gaining fat (Eat but do NOT over eat).
Any time your waist reaches 50% of your height, lower calories again, in order to keep from building up chronic low grade inflammation levels.
Train consistently, and match strenuous exercise with periods of light exercise (active recovery).
Consume 1-4 protein meals in-between each workout to take optimal advantage of the sensitization that occurs when you workout while in a low-inflammation state.

The occasional brief fast may help keep inflammation low, while still allowing for a calorie intake the allows for muscle growth on the days you are not fasting. For this reason fasting once or twice a week during a muscle building phase is ideal
Use anti-inflammatory drugs sparingly after a workout. Use them when they are needed, but keep in mind they may blunt the inflammation response to exercise.
Carbohydrate drinks during a workout may also blunt the exercise induced inflammation response.
Creatine monohydrate may increase the amount or sensitivity of your satellite cells.
Keep your waist circumference below 50% of your height at all times.
Avoid periods of excessive overeating.


Adams GR. Satellite cell proliferation and skeletal muscle hypertrophy. Appl Physiol Nutr Metab. 2006 Dec;31(6):782-90. Review.
Afroundeh R, Siahkouhian M, Khalili A. The effect of post-exercise carbohydrate ingestion on inflammatory responses to short time, high-force eccentric exercise. J Sports Med Phys Fitness. 2010 Jun;50(2):182-8.
Andersson A, Tengblad S, Karlström B, Kamal-Eldin A, Landberg R, Basu S, Aman P, Vessby B. Whole-grain foods do not affect insulin sensitivity or markers of lipid peroxidation and inflammation in healthy, moderately overweight subjects. J Nutr. 2007 Jun;137(6):1401-7.
Barbieri M, Ferrucci L, Ragno E, Corsi A, Bandinelli S, Bonafè M, Olivieri F, Giovagnetti S, Franceschi C, Guralnik JM, Paolisso G. Chronic inflammation and the effect of IGF-I on muscle strength and power in older persons. Am J Physiol Endocrinol Metab. 2003 Mar;284(3):E481-7. Epub 2002 Nov 5.
Bastard JP, Maachi M, Lagathu C, Kim MJ, Caron M, Vidal H, Capeau J, Feve B. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur Cytokine Netw. 2006 Mar;17(1):4-12. Review.
Basu A, Devaraj S, Jialal I. Dietary factors that promote or retard inflammation. Arterioscler Thromb Vasc Biol. 2006 May;26(5):995-1001. Epub 2006 Feb 16. Review.
Bharti AC, Donato N, Aggarwal BB. Curcumin (diferuloylmethane) inhibits constitutive and IL-6-inducible STAT3 phosphorylation in human multiple myeloma cells. J Immunol. 2003 Oct 1;171(7):3863-71.

Bhutani M, Pathak AK, Nair AS, Kunnumakkara AB, Guha S, Sethi G, Aggarwal BB. Capsaicin is a novel blocker of constitutive and interleukin-6-inducible STAT3 activation. Clin Cancer Res. 2007 May 15;13(10):3024-32.
Blackburn P, Després JP, Lamarche B, Tremblay A, Bergeron J, Lemieux I, Couillard C. Impact of postprandial variation in triglyceridemia on low-density lipoprotein particle size. Metabolism. 2003 Nov;52(11):1379-86.
Bladbjerg EM, Larsen TM, Due A, Stender S, Astrup A, Jespersen J. Effects on markers of inflammation and endothelial cell function of three ad libitum diets differing in type and amount of fat and carbohydrate: a 6-month randomised study in obese individuals. Br J Nutr. 2011 Jan 28:1-8. [Epub ahead of print]
Brandt C, Pedersen BK. The role of exercise-induced myokines in muscle homeostasis and the defense against chronic diseases. J Biomed Biotechnol. 2010;2010:520258. Epub 2010 Mar 9. Review.
Bruunsgaard H, Galbo H, Halkjaer-Kristensen J, Johansen TL, MacLean DA, Pedersen BK. Exercise-induced increase in serum interleukin-6 in humans is related to muscle damage. J Physiol. 1997 Mar 15;499 ( Pt 3):833-41.
Bruunsgaard H, Pedersen AN, Schroll M, Skinhøj P, Pedersen BK. TNF-alpha, leptin, and lymphocyte function in human aging. Life Sci. 2000 Oct 20;67(22):2721-31.
Cannon JG, St Pierre BA. Cytokines in exertion-induced skeletal muscle injury. Mol Cell Biochem. 1998 Feb;179(1-2):159-67. Review.

Cao H, Urban JF Jr, Anderson RA. Cinnamon polyphenol extract affects immune responses by regulating anti- and proinflammatory and glucose transporter gene expression in mouse macrophages. J Nutr. 2008 May;138(5):833-40.
Carpenter LL, Gawuga CE, Tyrka AR, Lee JK, Anderson GM, Price LH. Association between plasma IL-6 response to acute stress and early-life adversity in healthy adults. Neuropsychopharmacology. 2010 Dec;35(13):2617-23. Epub 2010 Sep 29.
Chan MH, McGee SL, Watt MJ, Hargreaves M, Febbraio MA. Altering dietary nutrient intake that reduces glycogen content leads to phosphorylation of nuclear p38 MAP kinase in human skeletal muscle: association with IL-6 gene transcription during contraction. FASEB J. 2004 Nov;18(14):1785-7. Epub 2004 Sep 2.
Corcoran MP, Meydani M, Lichtenstein AH, Schaefer EJ, Dillard A, Lamon-Fava S. Sex hormone modulation of proinflammatory cytokine and C-reactive protein expression in macrophages from older men and postmenopausal women. J Endocrinol. 2010 Aug;206(2):217-24. Epub 2010 May 18.
D'Agostino P, Milano S, Barbera C, Di Bella G, La Rosa M, Ferlazzo V, Farruggio R, Miceli DM, Miele M, Castagnetta L, Cillari E. Sex hormones modulate inflammatory mediators produced by macrophages. Ann N Y Acad Sci. 1999 Jun 22;876:426-9. No abstract available.
de Salles BF, Simão R, Fleck SJ, Dias I, Kraemer-Aguiar LG, Bouskela E. Effects of resistance training on cytokines. Int J Sports Med. 2010 Jul;31(7):441-50. Epub 2010 Apr 29. Review.
Eriksson A, Kadi F, Malm C, Thornell LE. Skeletal muscle morphology in power-lifters with and without anabolic steroids. Histochem Cell Biol. 2005 Aug;124(2):167-75. Epub 2005 Sep 29.

Ershler WB, Keller ET. Age-associated increased interleukin-6 gene expression, late-life diseases, and frailty. Annu Rev Med. 2000;51:245-70. Review.
Ershler WB, Sun WH, Binkley N. The role of interleukin-6 in certain age-related diseases. Drugs Aging. 1994 Nov;5(5):358-65. Review.
Esposito K, Nappo F, Marfella R, Giugliano G, Giugliano F, Ciotola M, Quagliaro L, Ceriello A, Giugliano D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: role of oxidative stress. Circulation. 2002 Oct 15;106(16):2067-72.
Esposito K, Pontillo A, Ciotola M, Di Palo C, Grella E, Nicoletti G, Giugliano D. Weight loss reduces interleukin-18 levels in obese women. J Clin Endocrinol Metab. 2002 Aug;87(8):3864-6.
Esposito K, Pontillo A, Di Palo C, Giugliano G, Masella M, Marfella R, Giugliano D. Effect of weight loss and lifestyle changes on vascular inflammatory markers in obese women: a randomized trial. JAMA. 2003 Apr 9;289(14):1799-804.
Fantuzzi G, Faggioni R. Leptin in the regulation of immunity, inflammation, and hematopoiesis. J Leukoc Biol. 2000 Oct;68(4):437-46. Review.
Febbraio MA, Hiscock N, Sacchetti M, Fischer CP, Pedersen BK. Interleukin-6 is a novel factor mediating glucose homeostasis during skeletal muscle contraction. Diabetes. 2004 Jul;53(7):1643-8.
Ferrucci L, Cavazzini C, Corsi A, Bartali B, Russo CR, Lauretani F, Ferrucci L, Cavazzini C, Corsi AM, Bartali B, Russo CR, Lauretani F, Bandinelli S, Bandinelli S, Guralnik JM. Biomarkers of frailty in older persons. J Endocrinol Invest. 2002;25(10 Suppl):10-5. Review.

Finch CE, Morgan TE. Systemic inflammation, infection, ApoE alleles, and Alzheimer disease: a position paper. Curr Alzheimer Res. 2007 Apr;4(2):185-9. Review.
Fischer CP, Hiscock NJ, Penkowa M, Basu S, Vessby B, Kallner A, Sjöberg LB, Pedersen BK. Supplementation with vitamins C and E inhibits the release of interleukin-6 from contracting human skeletal muscle. J Physiol. 2004 Jul 15;558(Pt 2):633-45. Epub 2004 May 28.
Fischer CP, Plomgaard P, Hansen AK, Pilegaard H, Saltin B, Pedersen BK. Endurance training reduces the contraction-induced interleukin-6 mRNA expression in human skeletal muscle. Am J Physiol Endocrinol Metab. 2004 Dec;287(6):E1189-94. Epub 2004 Aug 10.
Fontana L. The scientific basis of caloric restriction leading to longer life. Curr Opin Gastroenterol. 2009 Mar;25(2):144-50. Review.
Gabay C, Marinova-Mutafchieva L, Williams RO, Gigley JP, Butler DM, Feldmann M, Arend WP. Increased production of intracellular interleukin-1 receptor antagonist type I in the synovium of mice with collagen-induced arthritis: a possible role in the resolution of arthritis. Arthritis Rheum. 2001 Feb;44(2):451-62.
Garcia-Martinez C, Lopez-Soriano FJ, and Argiles JM. Acute treatment with tumor-necrosis-factor-alpha induces changes in protein metabolism in rat skeletal-muscle. Molecular and Cellular Biochemistry 125: 11-18, 1993.
Giltay EJ, Haider A, Saad F, Gooren LJ. C-reactive protein levels and ageing male symptoms in hypogonadal men treated with testosterone supplementation. Andrologia. 2008 Dec;40(6):398-400.
Gleeson M. Immune system adaptation in elite athletes. Curr Opin Clin Nutr Metab Care. 2006 Nov;9(6):659-65. Review.
Grounds MD, White JD, Rosenthal N, Bogoyevitch MA. The role of stem cells in skeletal and cardiac muscle repair. J Histochem Cytochem. 2002 May;50(5):589-610. Review.

Hatakeyama H, Nishizawa M, Nakagawa A, Nakano S, Kigoshi T, Uchida K. Testosterone inhibits tumor necrosis factor-alpha-induced vascular cell adhesion molecule-1 expression in human aortic endothelial cells. FEBS Lett. 2002 Oct 23;530(1-3):129-32.
Hawke TJ, Garry DJ. Myogenic satellite cells: physiology to molecular biology. J Appl Physiol. 2001 Aug;91(2):534-51. Review. Erratum in: J Appl Physiol 2001 Dec;91(6):2414.
Hawke TJ. Muscle stem cells and exercise training. Exerc Sport Sci Rev. 2005 Apr;33(2):63-8. Review.
Hiscock N, Chan MH, Bisucci T, Darby IA, Febbraio MA. Skeletal myocytes are a source of interleukin-6 mRNA expression and protein release during contraction: evidence of fiber type specificity. FASEB J. 2004 Jun;18(9):992-4. Epub 2004 Apr 1.
Hiscock N, Petersen EW, Krzywkowski K, Boza J, Halkjaer-Kristensen J, Pedersen BK. Glutamine supplementation further enhances exercise-induced plasma IL-6. J Appl Physiol. 2003 Jul;95(1):145-8. Epub 2003 Feb 28
Hood DA, Hawke TJ. Skeletal muscle stem cells: a symposium. Appl Physiol Nutr Metab. 2006 Dec;31(6):771-2.
Janssen SPM, Gayan-Ramirez G, Van Den Bergh A, Herijgers P, Maes K, Verbeken E, and Decramer M. Interleukin-6 causes myocardial failure and skeletal muscle atrophy in rats. Circulation 111: 996-1005, 2005.
Jones TH, Saad F. The effects of testosterone on risk factors for, and the mediators of, the atherosclerotic process. Atherosclerosis. 2009 Dec;207(2):318-27. Epub 2009 Apr 24. Review.
Jurasinski CV, Vary TC. Insulin-like growth factor I accelerates protein synthesis in skeletal muscle during sepsis. Am J Physiol. 1995 Nov;269(5 Pt 1):E977-81.
Kadi F, Eriksson A, Holmner S, Butler-Browne GS, Thornell LE. Cellular adaptation of the trapezius muscle in strength-trained athletes. Histochem Cell Biol. 1999 Mar;111(3):189-95.
Keller P, Penkowa M, Keller C, Steensberg A, Fischer CP, Giralt M, Hidalgo J, Pedersen BK. Interleukin-6 receptor expression in contracting human skeletal muscle: regulating role of IL-6. FASEB J. 2005 Jul;19(9):1181-3. Epub 2005 Apr 18

Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab. 2004 Jun;89(6):2548-56. Review.
Kondo T, Kobayashi I, Murakami M. Effect of exercise on circulating adipokine levels in obese young women. Endocr J. 2006 Apr;53(2):189-95.
Kroczynska B, Kaur S, Platanias LC. Growth suppressive cytokines and the AKT/mTOR pathway. Cytokine. 2009 Oct-Nov;48(1-2):138-43.
Lang CH, Frost RA, Bronson SK, Lynch CJ, Vary TC. Skeletal muscle protein balance in mTOR heterozygous mice in response to inflammation and leucine. Am J Physiol Endocrinol Metab. 2010 Jun;298(6):E1283-94. Epub 2010 Apr 13.
Lang CH, Frost RA, Vary TC. Regulation of muscle protein synthesis during sepsis and inflammation. Am J Physiol Endocrinol Metab. 2007 Aug;293(2):E453-9. Epub 2007 May 15. Review.
Lang CH, Frost RA. Glucocorticoids and TNFalpha interact cooperatively to mediate sepsis-induced leucine resistance in skeletal muscle. Mol Med. 2006 Nov-Dec;12(11-12):291-9.
Lang CH, Frost RA. Sepsis-induced suppression of skeletal muscle translation initiation mediated by tumor necrosis factor alpha. Metabolism. 2007 Jan;56(1):49-57.
Lang CH, Hong-Brown L, Frost RA. Cytokine inhibition of JAK-STAT signaling: a new mechanism of growth hormone resistance. Pediatr Nephrol. 2005 Mar;20(3):306-12. Epub 2004 Nov 10. Review.
Levinger I, Goodman C, Peake J, Garnham A, Hare DL, Jerums G, Selig S. Inflammation, hepatic enzymes and resistance training in individuals with metabolic risk factors. Diabet Med. 2009 Mar;26(3):220-7.
Li ZG, Danis VA, Brooks PM. Effect of gonadal steroids on the production of IL-1 and IL-6 by blood mononuclear cells in vitro. Clin Exp Rheumatol. 1993 Mar-Apr;11(2):157-62.

Liva SM, Voskuhl RR. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. J Immunol. 2001 Aug 15;167(4):2060-7.
Loffreda S, Yang SQ, Lin HZ, Karp CL, Brengman ML, Wang DJ, Klein AS, Bulkley GB, Bao C, Noble PW, Lane MD, Diehl AM. Leptin regulates proinflammatory immune responses. FASEB J. 1998 Jan;12(1):57-65.
Louis E, Raue U, Yang Y, Jemiolo B, Trappe S. Time course of proteolytic, cytokine, and myostatin gene expression after acute exercise in human skeletal muscle. J Appl Physiol. 2007 Nov;103(5):1744-51. Epub 2007 Sep 6.
MacIntyre DL, Sorichter S, Mair J, Berg A, McKenzie DC. Markers of inflammation and myofibrillar proteins following eccentric exercise in humans. Eur J Appl Physiol. 2001 Mar;84(3):180-6.
Mackinnon LT. Chronic exercise training effects on immune function. Med Sci Sports Exerc. 2000 Jul;32(7 Suppl):S369-76. Review.
Maki T, Pham NM, Yoshida D, Yin G, Ohnaka K, Takayanagi R, Kono S. The relationship of coffee and green tea consumption with high-sensitivity C-reactive protein in Japanese men and women. Clin Chem Lab Med. 2010 Jun;48(6):849-54.
Manna SK, Mukhopadhyay A, Aggarwal BB. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappa B, activator protein-1, and apoptosis: potential role of reactive oxygen intermediates and lipid peroxidation. J Immunol. 2000 Jun 15;164(12):6509-19.
Marimuthu K, Murton AJ, Greenhaff PL. Mechanisms regulating muscle mass during disuse atrophy and rehabilitation in humans. J Appl Physiol. 2011 Feb;110(2):555-60.

McFarlin BK, Flynn MG, Campbell WW, Stewart LK, Timmerman KL. TLR4 is lower in resistance-trained older women and related to inflammatory cytokines. Med Sci Sports Exerc. 2004 Nov;36(11):1876-83. Erratum in: Med Sci Sports Exerc. 2005 Feb;37(2):345.
McFarlin BK, Flynn MG, Phillips MD, Stewart LK, Timmerman KL. Chronic resistance exercise training improves natural killer cell activity in older women. J Gerontol A Biol Sci Med Sci. 2005 Oct;60(10):1315-8.
McKay BR, De Lisio M, Johnston AP, O'Reilly CE, Phillips SM, Tarnopolsky MA, Parise G. Association of interleukin-6 signalling with the muscle stem cell response following muscle-lengthening contractions in humans. PLoS One. 2009 Jun 24;4(6):e6027.
Mikkelsen UR, Langberg H, Helmark IC, Skovgaard D, Andersen LL, Kjaer M, Mackey AL. Local NSAID infusion inhibits satellite cell proliferation in human skeletal muscle after eccentric exercise. J Appl Physiol. 2009 Nov;107(5):1600-11. Epub 2009 Aug 27.
Mikkelsen UR, Schjerling P, Helmark IC, Reitelseder S, Holm L, Skovgaard D, Langberg H, Kjær M, Heinemeier KM. Local NSAID infusion does not affect protein synthesis and gene expression in human muscle after eccentric exercise. Scand J Med Sci Sports. 2010 Aug 24. [Epub ahead of print]
Mohamed-Ali V, Goodrick S, Rawesh A, Katz DR, Miles JM, Yudkin JS, Klein S, Coppack SW. Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J Clin Endocrinol Metab. 1997 Dec;82(12):4196-200.
Monteiro R, Azevedo I. Chronic inflammation in obesity and the metabolic syndrome. Mediators Inflamm. 2010;2010. pii: 289645. Epub 2010 Jul 14. Review.
Morgan TE, Wong AM, Finch CE. Anti-inflammatory mechanisms of dietary restriction in slowing aging processes. Interdiscip Top Gerontol. 2007;35:83-97. Review.

Moschen AR, Molnar C, Geiger S, Graziadei I, Ebenbichler CF, Weiss H, Kaser S, Kaser A, Tilg H. Anti-inflammatory effects of excessive weight loss: potent suppression of adipose interleukin 6 and tumour necrosis factor alpha expression. Gut. 2010 Sep;59(9):1259-64. Epub 2010 Jul 21.
Mullington JM, Simpson NS, Meier-Ewert HK, Haack M. Sleep loss and inflammation. Best Pract Res Clin Endocrinol Metab. 2010 Oct;24(5):775-84.
Murton AJ, Greenhaff PL. Physiological control of muscle mass in humans during resistance exercise, disuse and rehabilitation. Curr Opin Clin Nutr Metab Care. 2010 May;13(3):249-54. Review.
Naghii MR, Mofid M, Asgari AR, Hedayati M, Daneshpour MS. Comparative effects of daily and weekly boron supplementation on plasma steroid hormones and proinflammatory cytokines. J Trace Elem Med Biol. 2010 Dec 1. [Epub ahead of print]
Nakhai-Pour HR, Grobbee DE, Emmelot-Vonk MH, Bots ML, Verhaar HJ, van der Schouw YT. Oral testosterone supplementation and chronic low-grade inflammation in elderly men: a 26-week randomized, placebo-controlled trial. Am Heart J. 2007 Dec;154(6):1228.e1-7
Nehlsen-Cannarella SL, Fagoaga OR, Nieman DC, Henson DA, Butterworth DE, Schmitt RL, Bailey EM, Warren BJ, Utter A, Davis JM. Carbohydrate and the cytokine response to 2.5 h of running. J Appl Physiol. 1997 May;82(5):1662-7.
Nicklas BJ, Hsu FC, Brinkley TJ, Church T, Goodpaster BH, Kritchevsky SB, Pahor M. . Exercise training and plasma C-reactive protein and interleukin-6 in elderly people. J Am Geriatr Soc. 2008 Nov;56(11):2045-52.
Ostrowski K, Rohde T, Zacho M, Asp S, Pedersen BK. Evidence that interleukin-6 is produced in human skeletal muscle during prolonged running. J Physiol. 1998 May 1;508 ( Pt 3):949-53.

Pace TW, Mletzko TC, Alagbe O, Musselman DL, Nemeroff CB, Miller AH, Heim CM. Increased stress-induced inflammatory responses in male patients with major depression and increased early life stress. Am J Psychiatry. 2006 Sep;163(9):1630-3
Pal S, Ellis V. Acute effects of whey protein isolate on blood pressure, vascular function and inflammatory markers in overweight postmenopausal women. Br J Nutr. 2011 Jan 28:1-8. [Epub ahead of print]
Pal S, Ellis V. The chronic effects of whey proteins on blood pressure, vascular function, and inflammatory markers in overweight individuals. Obesity (Silver Spring). 2010 Jul;18(7):1354-9. Epub 2009 Nov 5.
Payette H, Roubenoff R, Jacques PF, Dinarello CA, Wilson PW, Abad LW, Harris T. Insulin-like growth factor-1 and interleukin 6 predict sarcopenia in very old community-living men and women: the Framingham Heart Study. J Am Geriatr Soc. 2003 Sep;51(9):1237-43.
Pedersen BK, Hoffman-Goetz L. Exercise and the immune system: regulation, integration, and adaptation. Physiol Rev. 2000 Jul;80(3):1055-81. Review.
Pedersen BK, Toft AD. Effects of exercise on lymphocytes and cytokines. Br J Sports Med. 2000 Aug;34(4):246-51. Review.
Petersen AM, Pedersen BK. The anti-inflammatory effect of exercise. J Appl Physiol. 2005 Apr;98(4):1154-62. Review.
Ploeger HE, Takken T, de Greef MH, Timmons BW. The effects of acute and chronic exercise on inflammatory markers in children and adults with a chronic inflammatory disease: a systematic review. Exerc Immunol Rev. 2009;15:6-41. Review.
Postprandial variations of plasma inflammatory markers in abdominally obese men. Obesity (Silver Spring). 2006 Oct;14(10):1747-54.
Reardon KA, Davis J, Kapsa RM, Choong P, Byrne E. Myostatin, insulin-like growth factor-1, and leukemia inhibitory factor mRNAs are upregulated in chronic human disuse muscle atrophy. Muscle Nerve. 2001 Jul;24(7):893-9.

Reed JL, De Souza MJ, Williams NI. Effects of exercise combined with caloric restriction on inflammatory cytokines. Appl Physiol Nutr Metab. 2010 Oct;35(5):573-82.
Robson-Ansley PJ, Blannin A, Gleeson M. Elevated plasma interleukin-6 levels in trained male triathletes following an acute period of intense interval training. Eur J Appl Physiol. 2007 Mar;99(4):353-60. Epub 2006 Dec 13.
Rogowski O, Shapira I, Bassat OK, Chundadze T, Finn T, Berliner S, Steinvil A. Waist circumference as the predominant contributor to the micro-inflammatory response in the metabolic syndrome: a cross sectional study. J Inflamm (Lond). 2010 Jul 26;7:35.
Roubenoff R, Parise H, Payette HA, Abad LW, D'Agostino R, Jacques PF, Wilson PW, Dinarello CA, Harris TB. Cytokines, insulin-like growth factor 1, sarcopenia, and mortality in very old community-dwelling men and women: the Framingham Heart Study. Am J Med. 2003 Oct 15;115(6):429-35.
Roubenoff R. Catabolism of aging: is it an inflammatory process? Curr Opin Clin Nutr Metab Care. 2003 May;6(3):295-9. Review.
Roubenoff R. Exercise and inflammatory disease. Arthritis Rheum. 2003 Apr 15;49(2):263-6. Review. No abstract available.
Schlitt A, Heine GH, Blankenberg S, Espinola-Klein C, Dopheide JF, Bickel C, Lackner KJ, Iz M, Meyer J, Darius H, Rupprecht HJ. CD14+CD16+ monocytes in coronary artery disease and their relationship to serum TNF-alpha levels. Thromb Haemost. 2004 Aug;92(2):419-24.
Serrano AL, Baeza-Raja B, Perdiguero E, Jardí M, Muñoz-Cánoves P. Interleukin-6 is an essential regulator of satellite cell-mediated skeletal muscle hypertrophy. Cell Metab. 2008 Jan;7(1):33-44.
Smedman A, Basu S, Jovinge S, Fredrikson GN, Vessby B. Conjugated linoleic acid increased C-reactive protein in human subjects. Br J Nutr. 2005 Nov;94(5):791-5.
Smith JK, Dykes R, Douglas JE, Krishnaswamy G, Berk S. Long-term exercise and atherogenic activity of blood mononuclear cells in persons at risk of developing ischemic heart disease. JAMA. 1999 May 12;281(18):1722-7.
Starkie RL, Arkinstall MJ, Koukoulas I, Hawley JA, Febbraio MA. Carbohydrate ingestion attenuates the increase in plasma interleukin-6, but not skeletal muscle interleukin-6 mRNA, during exercise in humans. J Physiol. 2001 Jun 1;533(Pt 2):585-91.
Steensberg A, Febbraio MA, Osada T, Schjerling P, van Hall G, Saltin B, Pedersen BK. Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content. J Physiol. 2001 Dec 1;537(Pt 2):633-9.

Steensberg A, Keller C, Starkie RL, Osada T, Febbraio MA, Pedersen BK. IL-6 and TNF-alpha expression in, and release from, contracting human skeletal muscle. Am J Physiol Endocrinol Metab. 2002 Dec;283(6):E1272-8. Epub 2002 Aug 20.
Stewart LK, Flynn MG, Campbell WW, Craig BA, Robinson JP, McFarlin BK, Timmerman KL, Coen PM, Felker J, Talbert E. Influence of exercise training and age on CD14+ cell-surface expression of toll-like receptor 2 and 4. Brain Behav Immun. 2005 Sep;19(5):389-97.
Stewart LK, Flynn MG, Campbell WW, Craig BA, Robinson JP, Timmerman KL, McFarlin BK, Coen PM, Talbert E. The influence of exercise training on inflammatory cytokines and C-reactive protein. Med Sci Sports Exerc. 2007 Oct;39(10):1714-9.
Suzuki K, Ito Y, Ochiai J, Kusuhara Y, Hashimoto S, Tokudome S, Kojima M, Wakai K, Toyoshima H, Tamakoshi K, Watanabe Y, Hayakawa N, Maruta M, Watanabe M, Kato K, Ohta Y, Tamakoshi A; JACC Study Group. Relationship between obesity and serum markers of oxidative stress and inflammation in Japanese. Asian Pac J Cancer Prev. 2003 Jul-Sep;4(3):259-66.
Suzuki K, Nakaji S, Yamada M, Liu Q, Kurakake S, Okamura N, Kumae T, Umeda T, Sugawara K. Impact of a competitive marathon race on systemic cytokine and neutrophil responses. Med Sci Sports Exerc. 2003 Feb;35(2):348-55.
Suzuki K, Nakaji S, Yamada M, Totsuka M, Sato K, Sugawara K. Systemic inflammatory response to exhaustive exercise. Cytokine kinetics. Exerc Immunol Rev. 2002;8:6-48. Review.
Timmerman KL, Flynn MG, Coen PM, Markofski MM, Pence BD. Exercise training-induced lowering of inflammatory (CD14+CD16+) monocytes: a role in the anti-inflammatory influence of exercise? J Leukoc Biol. 2008 Nov;84(5):1271-8. Epub 2008 Aug 12.
Toth MJ, Ades PA, Tischler MD, Tracy RP, LeWinter MM. Immune activation is associated with reduced skeletal muscle mass and physical function in chronic heart failure. Int J Cardiol. 2006 May 10;109(2):179-87. Epub 2005 Jul 15.
Toth MJ, Matthews DE, Tracy RP, Previs MJ. Age-related differences in skeletal muscle protein synthesis: relation to markers of immune activation. Am J Physiol Endocrinol Metab. 2005 May;288(5):E883-91. Epub 2004 Dec 21.
Trappe TA, White F, Lambert CP, Cesar D, Hellerstein M, Evans WJ. Effect of ibuprofen and acetaminophen on postexercise muscle protein synthesis. Am J Physiol Endocrinol Metab. 2002 Mar;282(3):E551-6.

van Dijk SJ, Feskens EJ, Bos MB, Hoelen DW, Heijligenberg R, Bromhaar MG, de Groot LC, de Vries JH, Müller M, Afman LA. A saturated fatty acid-rich diet induces an obesity-linked proinflammatory gene expression profile in adipose tissue of subjects at risk of metabolic syndrome. Am J Clin Nutr. 2009 Dec;90(6):1656-64. Epub 2009 Oct 14.
Varady KA, Bhutani S, Church EC, Klempel MC. Short-term modified alternate-day fasting: a novel dietary strategy for weight loss and cardioprotection in obese adults. Am J Clin Nutr. 2009 Nov;90(5):1138-43. Epub 2009 Sep 30.
Vary TC, Kimball SR. Regulation of hepatic protein synthesis in chronic inflammation and sepsis. Am J Physiol. 1992 Feb;262(2 Pt 1):C445-52.
Visser M, Pahor M, Taaffe DR, Goodpaster BH, Simonsick EM, Newman AB, Nevitt M, Harris TB. Relationship of interleukin-6 and tumor necrosis factor-alpha with muscle mass and muscle strength in elderly men and women: the Health ABC Study. J Gerontol A Biol Sci Med Sci. 2002 May;57(5):M326-32.
Vozarova B, Weyer C, Hanson K, Tataranni PA, Bogardus C, Pratley RE. Circulating interleukin-6 in relation to adiposity, insulin action, and insulin secretion. Obes Res. 2001 Jul;9(7):414-7.
You T, Nicklas BJ. Chronic inflammation: role of adipose tissue and modulation by weight loss.


















