
AberrantRetinoblastoma(RB)-E2FTranscriptionalRegulation ... ·...
15
Aberrant Retinoblastoma (RB)-E2F Transcriptional Regulation Defines Molecular Phenotypes of Osteosarcoma * Received for publication, July 24, 2015, and in revised form, September 11, 2015 Published, JBC Papers in Press, September 16, 2015, DOI 10.1074/jbc.M115.679696 Milcah C. Scott ‡§¶ , Aaron L. Sarver ‡§ , Hirotaka Tomiyasu ‡§¶ , Ingrid Cornax ‡¶ , Jamie Van Etten ¶ ** 1 , Jyotika Varshney ‡ ** ‡‡ , M. Gerard O’Sullivan ‡¶ , Subbaya Subramanian ‡¶ **, and Jaime F. Modiano ‡§¶§§¶¶2 From the ‡ Animal Cancer Care and Research Program, the Departments of § Veterinary Clinical Sciences and Veterinary Population Medicine and the ‡‡ Veterinary Medicine Graduate Program, College of Veterinary Medicine, the ¶ Masonic Cancer Center, the **Department of Surgery, School of Medicine, the §§ Stem Cell Institute, and the ¶¶ Center for Immunology, University of Minnesota, Minneapolis, Minnesota 55455 Background: Gene expression signatures define prognostically significant osteosarcoma phenotypes. Results: Deregulation of the RB-E2F pathway establishes more aggressive phenotype. Inhibitors of DNA and chromatin remod- eling promote comparable transcriptional changes as genetic restoration of RB. Conclusion: Aberrant RB-E2F pathway alters epigenetic landscape and biological behavior of osteosarcoma. Significance: Epigenetic remodeling regulated by RB-E2F gives rise to patterns of gene expression that are associated with different biological behavior and progression of osteosarcoma. We previously identified two distinct molecular subtypes of osteosarcoma through gene expression profiling. These subtypes are associated with distinct tumor behavior and clinical outcomes. Here, we describe mechanisms that give rise to these molecular subtypes. Using bioinformatic analyses, we identified a significant association between deregulation of the retinoblastoma (RB)-E2F pathway and the molecular subtype with worse clinical outcomes. Xenotransplantation models recapitulated the corresponding behavior for each osteosarcoma subtype; thus, we used cell lines to validate the role of the RB-E2F pathway in regulating the prognos- tic gene signature. Ectopic RB resets the patterns of E2F regulated gene expression in cells derived from tumors with worse clinical outcomes (molecular phenotype 2) to those comparable with those observed in cells derived from tumors with less aggressive out- comes (molecular phenotype 1), providing a functional association between RB-E2F dysfunction and altered gene expression in osteo- sarcoma. DNA methyltransferase and histone deacetylase inhibi- tors similarly reset the transcriptional state of the molecular phe- notype 2 cells from a state associated with RB deficiency to one seen with RB sufficiency. Our data indicate that deregulation of RB-E2F pathway alters the epigenetic landscape and biological behavior of osteosarcoma. Osteosarcoma is a genetically complex, heterogeneous dis- ease that occurs naturally in humans and dogs (1–3). In the past decade, the molecular basis of osteosarcoma has received sig- nificant attention, and a number of recurring chromosomal aberrations and changes in gene expression have been identi- fied (1–5). However, these findings have not yet translated into significant improvements in disease prognosis or outcome (2, 6 – 8), placing osteosarcoma among the “most wanted” for new and effective therapies (9). The ability to prospectively identify patients whose tumors have distinct gene expression profiles (molecular phenotypes) associated with clinical outcomes may offer insights to develop new therapeutic strategies adapted to tumor behavior. The conservation of disease mechanisms between canine and human osteosarcoma supports using the former as a compara- tive model to achieve this goal (2, 5, 10 –12). Previously, we identified a gene signature consisting of 250 genes that strat- ified canine osteosarcoma into subgroups predictive of patient outcome (5, 13). The gene signature and its prognostic value were conserved in tumors from human osteosarcoma patients (5). Tumors from patients with longer survival (henceforth called “molecular phenotype 1”) were characterized by decreased expression of genes associated with G 2 /M transition and DNA damage-induced cell cycle checkpoints. Conversely, decreased expression of genes associated with microenviron- ment interactions was observed in tumors from patients with shorter survival (henceforth called “molecular phenotype 2”). In the present study, we characterized mechanisms that are causally related to these distinct molecular phenotypes. Specif- ically, we show that deregulation of the RB-E2F 3 pathway is a major feature of molecular phenotype 2 tumors and that resto- ration of RB in cells from these tumors resets gene expression to * This work was supported by Morris Animal Foundation Grant D13CA-032, American Cancer Society Grant RSG-13-381-01, Karen Wyckoff Rein in Sar- coma Foundation Grant 2011-1, the Zach Sobiech Fund for Osteosarcoma Research of the Children’s Cancer Research Fund, and the Comparative Med- icine Signature Program of the College of Veterinary Medicine, University of Minnesota. National Institutes of Health Comprehensive Cancer Center Sup- port Grant P30 CA077598 to the Masonic Cancer Center provided support for bioinformatics, genomics, flow cytometry, bioimaging, cytogenetics, and comparative pathology services. This work was also supported by donors to the Animal Cancer Care and Research Program of the University of Minnesota, which helped support of the project. The authors declare that they have no conflicts of interest with the contents of this article. 1 Supported by Cancer Biology Training Grant T32 CA09138 from the National Institutes of Health. 2 Supported by the Alvin and June Perlman Chair in Animal Oncology. To whom correspondence should be addressed: Masonic Cancer Center, Univ. of Minnesota, 420 Delaware St., SE, MMC 806, Minneapolis, MN 55455. Tel.: 612-625-7436; Fax: 612-626-4915; E-mail: [email protected]. 3 The abbreviations used are: RB, retinoblastoma; IPA, ingenuity pathway analysis; DNMT, DNA methyltransferase; HDAC, histone deacetylase; Luc, luciferase; CDK, cyclin-dependent kinase; qRT-PCR, quantitative RT-PCR; SAHA, suberoylanilide hydroxamic acid; Zeb, zebularine; CDE, cell cycle-de- pendent element. crossmark THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 290, NO. 47, pp. 28070 –28083, November 20, 2015 © 2015 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A. 28070 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015 by guest on March 5, 2019 http://www.jbc.org/ Downloaded from
Transcript of AberrantRetinoblastoma(RB)-E2FTranscriptionalRegulation ... ·...

Aberrant Retinoblastoma (RB)-E2F Transcriptional RegulationDefines Molecular Phenotypes of Osteosarcoma*
Received for publication, July 24, 2015, and in revised form, September 11, 2015 Published, JBC Papers in Press, September 16, 2015, DOI 10.1074/jbc.M115.679696
Milcah C. Scott‡§¶, Aaron L. Sarver‡§, Hirotaka Tomiyasu‡§¶, Ingrid Cornax‡¶�, Jamie Van Etten¶**1,Jyotika Varshney‡**‡‡, M. Gerard O’Sullivan‡¶�, Subbaya Subramanian‡¶**, and Jaime F. Modiano‡§¶§§¶¶2
From the ‡Animal Cancer Care and Research Program, the Departments of §Veterinary Clinical Sciences and �VeterinaryPopulation Medicine and the ‡‡Veterinary Medicine Graduate Program, College of Veterinary Medicine, the ¶Masonic CancerCenter, the **Department of Surgery, School of Medicine, the §§Stem Cell Institute, and the ¶¶Center for Immunology, Universityof Minnesota, Minneapolis, Minnesota 55455
Background: Gene expression signatures define prognostically significant osteosarcoma phenotypes.Results: Deregulation of the RB-E2F pathway establishes more aggressive phenotype. Inhibitors of DNA and chromatin remod-eling promote comparable transcriptional changes as genetic restoration of RB.Conclusion: Aberrant RB-E2F pathway alters epigenetic landscape and biological behavior of osteosarcoma.Significance: Epigenetic remodeling regulated by RB-E2F gives rise to patterns of gene expression that are associated withdifferent biological behavior and progression of osteosarcoma.
We previously identified two distinct molecular subtypes ofosteosarcoma through gene expression profiling. These subtypesare associated with distinct tumor behavior and clinical outcomes.Here, we describe mechanisms that give rise to these molecularsubtypes. Using bioinformatic analyses, we identified a significantassociation between deregulation of the retinoblastoma (RB)-E2Fpathway and the molecular subtype with worse clinical outcomes.Xenotransplantation models recapitulated the correspondingbehavior for each osteosarcoma subtype; thus, we used cell lines tovalidate the role of the RB-E2F pathway in regulating the prognos-tic gene signature. Ectopic RB resets the patterns of E2F regulatedgene expression in cells derived from tumors with worse clinicaloutcomes (molecular phenotype 2) to those comparable with thoseobserved in cells derived from tumors with less aggressive out-comes (molecular phenotype 1), providing a functional associationbetween RB-E2F dysfunction and altered gene expression in osteo-sarcoma. DNA methyltransferase and histone deacetylase inhibi-tors similarly reset the transcriptional state of the molecular phe-notype 2 cells from a state associated with RB deficiency to one seenwith RB sufficiency. Our data indicate that deregulation of RB-E2Fpathway alters the epigenetic landscape and biological behavior ofosteosarcoma.
Osteosarcoma is a genetically complex, heterogeneous dis-ease that occurs naturally in humans and dogs (1–3). In the pastdecade, the molecular basis of osteosarcoma has received sig-nificant attention, and a number of recurring chromosomalaberrations and changes in gene expression have been identi-fied (1–5). However, these findings have not yet translated intosignificant improvements in disease prognosis or outcome (2,6 – 8), placing osteosarcoma among the “most wanted” for newand effective therapies (9).
The ability to prospectively identify patients whose tumorshave distinct gene expression profiles (molecular phenotypes)associated with clinical outcomes may offer insights to developnew therapeutic strategies adapted to tumor behavior. Theconservation of disease mechanisms between canine andhuman osteosarcoma supports using the former as a compara-tive model to achieve this goal (2, 5, 10 –12). Previously, weidentified a gene signature consisting of �250 genes that strat-ified canine osteosarcoma into subgroups predictive of patientoutcome (5, 13). The gene signature and its prognostic valuewere conserved in tumors from human osteosarcoma patients(5). Tumors from patients with longer survival (henceforthcalled “molecular phenotype 1”) were characterized bydecreased expression of genes associated with G2/M transitionand DNA damage-induced cell cycle checkpoints. Conversely,decreased expression of genes associated with microenviron-ment interactions was observed in tumors from patients withshorter survival (henceforth called “molecular phenotype 2”).In the present study, we characterized mechanisms that arecausally related to these distinct molecular phenotypes. Specif-ically, we show that deregulation of the RB-E2F3 pathway is amajor feature of molecular phenotype 2 tumors and that resto-ration of RB in cells from these tumors resets gene expression to
* This work was supported by Morris Animal Foundation Grant D13CA-032,American Cancer Society Grant RSG-13-381-01, Karen Wyckoff Rein in Sar-coma Foundation Grant 2011-1, the Zach Sobiech Fund for OsteosarcomaResearch of the Children’s Cancer Research Fund, and the Comparative Med-icine Signature Program of the College of Veterinary Medicine, University ofMinnesota. National Institutes of Health Comprehensive Cancer Center Sup-port Grant P30 CA077598 to the Masonic Cancer Center provided support forbioinformatics, genomics, flow cytometry, bioimaging, cytogenetics, andcomparative pathology services. This work was also supported by donors tothe Animal Cancer Care and Research Program of the University of Minnesota,which helped support of the project. The authors declare that they have noconflicts of interest with the contents of this article.
1 Supported by Cancer Biology Training Grant T32 CA09138 from the NationalInstitutes of Health.
2 Supported by the Alvin and June Perlman Chair in Animal Oncology. Towhom correspondence should be addressed: Masonic Cancer Center,Univ. of Minnesota, 420 Delaware St., SE, MMC 806, Minneapolis, MN55455. Tel.: 612-625-7436; Fax: 612-626-4915; E-mail: [email protected].
3 The abbreviations used are: RB, retinoblastoma; IPA, ingenuity pathwayanalysis; DNMT, DNA methyltransferase; HDAC, histone deacetylase; Luc,luciferase; CDK, cyclin-dependent kinase; qRT-PCR, quantitative RT-PCR;SAHA, suberoylanilide hydroxamic acid; Zeb, zebularine; CDE, cell cycle-de-pendent element.
crossmarkTHE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 290, NO. 47, pp. 28070 –28083, November 20, 2015
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
28070 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

a state comparable with that seen in tumors from patients withlonger survival.
One mechanism of RB-dependent gene regulation isthrough changing chromatin structure (14, 15). Thus, wehypothesized that the RB-E2F pathway might be functionallyrestored by pharmacologic alteration of DNA and chromatinstructure. We recently reported that DNA methyltransferase(DNMT) and histone deacetylase (HDAC) inhibitors hadsynergistic and selective cytotoxicity effects against humanand canine osteosarcoma cells (16). Here, we show thattreatment using DNMT inhibitor (zebularine) with theHDAC inhibitor (vorinostat) was sufficient to alter the tran-scriptional state of molecular phenotype 2 cells to oneresembling that seen with active RB.
Experimental Procedures
Cell Lines and Cell Culture—Canine and human osteosar-coma cell lines and the Jurkat T-cell leukemia line were estab-lished and maintained as previously described (5, 16, 17).OSCA-8, OSCA-32, and OSCA-40 cells were modified tostably express GFP and firefly luciferase (Luc) for in vivoexperiments (18). Fluorescence in situ hybridization wasused to determine the number of GFP/Luc copies in the celllines. Morphologic appearance, doubling time, and routineviability assays were used to confirm that growth propertiesof the derivative cell lines were comparable with those of theparental cell lines. Luciferase activity in the parental celllines and the GFP/Luc modified cells was measured in vitrowith the dual luciferase reporter assay system (Promega,Madison, WI) (19) using a Wallac 1420 microplate reader(PerkinElmer Life Sciences). Firefly luciferase was normal-ized to Renilla luciferase.
Expression Vectors and Transfections—A pGL3 luciferasereporter encoding Luc downstream from a 515-bp AURKBpromoter was a kind gift of Dr. Masashi Kimura (Gifu, Japan)(20). The 515-bp sequence contains full AURKB promoteractivity. Constructs encoding wild type, N-terminal truncatedRB (WT RB) or a cyclin-dependent kinase (CDK)-insensitive,N-terminal truncated mutant (PSM 7-LP) RB were provided byDr. Erik S. Knudsen (Dallas, TX and San Diego, CA) (21).Expression vectors encoding wild type p16 or p21 have beendescribed (19, 22). pGL4.73 hRenillaLuc/SV40 vector was pur-chased from Promega, and empty CMV-Neo-Bam vector waspurchased from Addgene (Cambridge, MA). Expression vec-tors were mixed with a 1/100 molar equivalent of hRenillaLuc/SV40 vector in 20 �l of supplemented SE solution (Lonza, Basel,Switzerland). These mixtures were added to 200,000 cells,which were then transfected using the Lonza 4D Nucleofector.The reactions were optimized to achieve �80% viability, andtransfection in all cell lines was monitored by GFP expression.Luciferase activity was measured using the dual luciferasereporter assay system.
Site-directed Mutagenesis—PCR site-directed mutagenesisof the [GGCGGG] E2F binding site in the AURKB promoterwas done using the mutagenic primers reported (20) and theGENEART site-directed mutagenesis system (Life Technolo-gies, Inc.).
Western Blotting—Immunoblotting was done as described(22), with detection and quantification using the Li-Cor Bio-sciences (Lincoln, NE) Odyssey system (Masonic CancerCenter Flow Cytometry Shared Resources). Anti-RB mono-clonal antibodies (clone G3-245 (catalog no. 554136); BDSciences, San Jose, CA, and clone IF8 (catalog no. sc-53566);Santa Cruz, Dallas, TX) were used to detect endogenouscanine RB (23). Human RB (PSM-7LP RB) was detected withan antibody that recognized the human but not the canineC-terminal domain of the protein (clone LM95.1 (catalog no.OP66 –100UG); EMD Millipore, Billerica, MA). �-Actin(clone AC-15 (catalog no. A5441); Sigma-Aldrich) was usedas a loading control.
RNA Preparation and Real Time Quantitative RT-PCR(qRT-PCR)—RNAs were prepared using the miRVANA kit(Life Technologies, Inc.), and cDNA was synthesized from totalRNA using a miScript reverse transcription kit. cDNAs werequantified using the miScript SYBR Green PCR kit (Qiagen)and the 7500 Real Time PCR system (Applied BioSystems, Fos-ter City, CA) protocol. Previously published primer sequenceswere used (24). GAPDH was used for normalization, and rela-tive levels of mRNA were established using the ��Ct method.
Inhibition of DNA Methylation and of Histone Dea-cetylation—Canine OSCA-40, OSCA-78, and OSCA-32 cellswere cultured in the presence of 1 �M suberoylanilidehydroxamic acid (SAHA/vorinostat; Cayman Chemical, AnnArbor, MI) and 10 �M zebularine (Zeb; Sigma-Aldrich) as pre-viously described (16)
Chromatin Immunoprecipitation—ChIP assays were per-formed using the ChIP-IT Express kit (Active Motif, Carlsbad,CA). Briefly, cells were cross-linked in culture medium contain-ing 1% formaldehyde, lysed, and then sheared to an average sizeof 250 –500 bp by sonication in shearing buffer using a Bransonsonicator (Thomas Scientific, Swedesboro, NJ). ChIP was per-formed by incubating 25 �g of chromatin/reaction with proteinG magnetic beads and 5 �g of anti-E2F1 antibody purchasedfrom Abcam (catalog no. ab112580; Cambridge, MA), anti-hu-man RB antibody (catalog no. OP66-100UG; EMD Millipore),or control IgG overnight at 4 °C. Immunoprecipitated chroma-tin was purified by magnetic separation, and proteins weredigested with proteinase K and enrichment of E2F1 sequences.To amplify the GGGCGG (CDE site) sequence of the humanAURKB (AC135178.13) promoter, the following primers wereused: 5�-GAGCCAATGGGAACTAGGCA (forward) and 5�-CCCTGGCCAAGGACTTTTCA (reverse). To amplify theTTTCCAGCCAAT E2F binding site in canine AURKB (NC_006587.3), the following primers were used: 5�-TTGGGTCC-CAAGGTCTACGT (forward) and 5�-AGGCCCTTTCAAAT-CTCCCG (reverse). To amplify the CGGCGCTAAA E2Fbinding site in canine CHEK1 (NC_006587.3), the followingprimers were used: 5�-TTGGGTCCCAAGGTCTACGT(forward) and 5�-AGGCCCTTTCAAATCTCCCG (reverse).For all primer pairs, PCR was performed at 60 °C, annealingtemperature for 40 cycles. For each sample, fold enrichment oftarget sequence in ChIP samples versus negative control wascalculated by the �Ct method. All ChIP reactions wereperformed in duplicate. The data represent the means � S.D. offold enrichment.
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28071
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Gene Expression Profiling—Hybridization to canine 4 � 44,000 microarray chips (Agilent Technologies, Santa Clara, CA)was done as described at the University of Minnesota GenomicsCenter (5, 18). Probe signal levels were quantile-normalizedand summarized as previously described (5) (data archive sub-mitted to the Gene Expression Omnibus). Two group t testswere done to determine differentially expressed genes.
Identification of Transcriptional Regulators—The ingenuitypathway analysis (IPA) suite (Ingenuity Systems, RedwoodCity, CA) was used to identify potential driver upstream tran-scriptional regulators responsible for gene signatures or differ-entially expressed genes. IPA upstream regulator analysis isbased on prior knowledge of predictable effects between tran-scriptional regulators and their target genes stored in the Inge-nuity Knowledge Base. IPA provides two statistical measures:the p value and regulation Z score to detect potential upstreamtranscriptional regulators. First, the p value was calculatedbased on how many known targets of each transcriptional reg-ulator were present in the gene signature. Second, the knowneffect (repression or activation) of a transcriptional regulatoron each target gene was compared with the observed changes ingene expression in the signature. A Z score was calculated fromthe concordance of the known effects of transcriptional regula-tors and the observed changes in gene expression. A Z score of�2 indicated activation of the transcriptional regulator,whereas a Z score of �2 indicated repression of the transcrip-tional regulator. The predicted upstream regulators were lim-ited to those known to be a “transcriptional regulator” or a“group.”
DNA Motif Identification—The hg19_genes_2012-03-09GTF file (University of California, Santa Cruz GenomeBrowser) was used for retrieval of 1000 to 1 nucleotideregions relative to the predicted ATG translation start site foreach ORF (25, 26). The 5� promoter sequences of 143 genes ofthe G2/M cell cycle transition and DNA damage cluster and 108genes of the microenvironment interactions cluster (5) wereavailable for motif discovery using the Multiple ExpectationMaximization for Motif Elicitation Suite (version 4.9.0) in theGalaxy platform (27). Motifs with zero or one occurrence ineach promoter and a length of between 5 and 10 nucleotideswere identified.
Orthotopic Model of Canine Osteosarcoma Cell Lines—Pro-cedures using laboratory animals were done according to theguidelines and under the supervision, of the University of Min-nesota Institutional Animal Care and Use Committee (protocol1207A17293). Six-week-old (�20 grams) female athymic nudemice (NCr-nu/nu; NCI, National Institutes of Health, Fredrick,MD) anesthetized with xylazine (10 mg/kg) and ketamine (100mg/kg) were injected intratibially with OSCA-8, OSCA-32, orOSCA-40 cells (105 per mouse). Buprenorphine (0.075 mg/kgq.8 h) was used for pain control over the first 24 h, and Tylenoladministered in the water was used as needed for pain controlthereafter. Routine tumor end points (ill thrift, or a tumorreaching 1 cm in the largest diameter for any animal in a group)or the inability to control pain or discomfort (visible lamenessor difficulty moving in the cage) triggered termination of theexperiment and humane euthanasia of the mice for thatgroup. Tumor growth was monitored using caliper measure-
ments and in vivo imaging as described (18). For histologicalconfirmation, tumors were collected immediately upon sac-rifice, fixed in 10% neutral buffered formalin, and evaluatedgrossly and histologically by board-certified veterinarypathologists (Masonic Cancer Center Comparative Pathol-ogy Shared Resource Core).
Statistical Analysis—Graphs were created using Prism (ver-sion 5.0; GraphPad Software, Inc., La Jolla, CA). The results arepresented as the means � S.D. Student’s two-tailed t test wasused to assess significance. p values � 0.05 were consideredsignificant.
Results
Deregulation of the RB-E2F Pathway Is Associated withMolecular Phenotype That Predicts Worse Clinical Outcomes—We anticipated that one or few upstream transcriptional regu-lators were likely responsible for the previously published,observed expression changes that segregate osteosarcomasamples into two distinct molecular phenotypes predictiveof tumor behavior and outcome (Fig. 1A) (5). To identifypotential candidates, we used upstream regulator analysiswithin the IPA suite. The direction of gene expressionchanges in the molecular phenotype 2 samples (shorter
FIGURE 1. Osteosarcoma in vivo models recapitulate the biologicalbehavior of two distinct molecular phenotypes. A, sample dendrogramfrom previously published gene expression profiling of canine osteosarcomashowing two molecularly distinct sample clusters, denoted as molecular phe-notype 1 and molecular phenotype 2, which have significantly different sur-vival times of 14 and 2.83 months, respectively (5). Representative cell linesfrom each phenotype (OSCA-32, OSCA-40, OSCA-78, and OSCA-8) were usedto identify mechanisms driving sample stratification. B, serial in vivo imagingof athymic nude mice harboring intratibial xenografts of OSCA-32 cells(molecular phenotype 1) or OSCA-40 or OSCA-8 cells (molecular phenotype2).
Aberrant RB-E2F Defines Molecular Phenotypes
28072 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

median survival times) was consistent with inactive RB andp53 tumor suppressor genes (Table 1, activation Z scores of3.801 and 3.791, respectively). Other significant, pre-dicted altered regulators included E2F transcription factorsand chromatin remodelers: E2F-1, E2F-2, E2F-3, E2F4, E2F6,SMARCB1, KDM5B, and HDAC1. E2F4 was the most signif-icantly altered transcriptional regulator of the gene signa-ture (Table 1). However, because E2F4 up-regulates some ofthese genes and down-regulates others, a direction of activ-ity (Z score) could not be determined.
An important role for E2F regulation of the target genes inthe gene signature became more evident when we searched forconserved DNA response elements in 5� upstream promotersequences. More than 70% of the 5� upstream promotersequences of the genes comprising the prognostic signaturecontained the E2F consensus binding motif sequence, CCAG-GCTGG (data not shown). The CCAGGCTGG sequence waspresent in 106 of 143 promoters of genes in the G2/M cluster (Evalue 8.1E-105) and in 80 of 108 promoters of the genes associ-ated with microenvironment interactions (E value 5.0E-32) (5).Importantly, the 9 base pair dyad sequence also is one of themost common sequence motifs in promoters of E2F4 targetgenes (28).
Orthotopic Xenografts Using Cell Lines Derived from Molec-ular Phenotype 1 and Molecular Phenotype 2 OsteosarcomasRecapitulate Their Clinical Behavior—To test the tumorigenicpotential and outcome of cells from tumors from each molec-ular phenotype and to establish suitable cell lines for down-stream functional studies, we evaluated tumor growth in ortho-topic xenografts. The pattern of outcomes and tumor behaviorwas maintained in vivo: the OSCA-32 cell line, which was
derived from a molecular phenotype 1 tumor, progressed moreslowly and generated less local bone destruction than theOSCA-40 and OSCA-8 cell lines, which were derived from dogswith molecular phenotype 2 (shorter median survival times)tumors (Fig. 1B). Microscopic findings for these tumors wereconsistent with clinical outcome. The OSCA-32 tumor cellsshowed relatively well differentiated tumor cells laying downosteoid seams in an orderly fashion (Fig. 2, A and C); in contrast,OSCA-40 had highly anaplastic cells embedded in a poorlyorganized osteoid matrix and extensive areas of necrosis (Fig. 2,B and D).
Characterization of Representative Cell Lines of Each Osteo-sarcoma Phenotype—The results from the orthotopic xeno-grafts supported the use of these cell lines to elucidate patho-genetic mechanisms responsible for the biological behavior ofthe two molecular phenotypes of osteosarcoma. Deregulationof E2F transcriptional activity could result from direct or indi-rect mechanisms upon loss of or reduced RB function. This lossof or reduced RB function, in turn, might be due to mutationsthat decrease or eliminate RB-1 expression or that render theRB protein inactive; however, RB is a component of a complexpathway, and its activity can be influenced by several regulatoryfactors (29). The steady state levels of RB protein were repro-ducibly lower in cell lines derived from molecular phenotype 2tumors (OSCA-40, OSCA-78, and OSCA-8) as compared withthose seen in the cell line derived from a molecular phenotype 1tumor (OSCA-32) (Fig. 3A).
To test the hypothesis that functional deregulation ofRB-E2F transcriptional regulation was causally related to theosteosarcoma molecular phenotypes, we measured the effect ofRB protein abundance on expression levels of AURKB (aurora
TABLE 1Regulators of the gene signature that identifies two osteosarcoma phenotypesIPA was applied to identify upstream transcriptional regulators of the gene signature consisting of approximately 250 genes (5). A Z score of �2 indicates activation, whereasa Z score of �2 indicates inactivation.
Upstream regulator Molecule typePredicted activation state in samples
with worse patient outcomes Activation Z score p value of overlap
E2F4 Transcription regulator NA NA 7.86E-39Vegf Group Activated 5.751 2.79E-30E2F1 Transcription regulator Activated 5.032 1.18E-31TBX2 Transcription regulator Activated 4.583 9.26E-27FOXM1 Transcription regulator Activated 4.127 3.70E-20CCND1 Transcription regulator Activated 3.755 6.74E-28MED1 Transcription regulator Activated 3.550 2.88E-09FOXO1 Transcription regulator Activated 3.503 4.12E-09E2f Group Activated 3.382 3.61E-19MYC Transcription regulator Activated 3.309 3.18E-10STAT3 Transcription regulator Activated 3.239 2.24E-04E2F3 Transcription regulator Activated 3.124 1.28E-23NFKBIA Transcription regulator Activated 3.011 2.16E-09E2F2 Transcription regulator Activated 3.000 1.20E-18NRIP1 Transcription regulator Inhibited 2.219 2.32E-06SMARCB1 Transcription regulator Inhibited 2.393 2.73E-16TOB1 Transcription regulator Inhibited 2.449 5.86E-06ATF3 Transcription regulator Inhibited 2.577 3.17E-07HOXA10 Transcription regulator Inhibited 2.588 2.20E-03HDAC1 Transcription regulator Inhibited 2.611 2.24E-11E2F6 Transcription regulator Inhibited 2.828 1.27E-11TCF3 Transcription regulator Inhibited 3.317 3.95E-08Rb Group Inhibited 3.348 2.94E-19RBL1 Transcription regulator Inhibited 3.379 8.36E-15KDM5B Transcription regulator Inhibited 3.734 9.06E-17TP53 Transcription regulator Inhibited 3.791 2.03E-38RB1 Transcription regulator Inhibited 3.801 2.51E-23CDKN2A Transcription regulator Inhibited 4.358 2.27E-16NUPR1 Transcription regulator Inhibited 4.849 6.56E-17
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28073
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

kinase B), a target gene that was among those most highlyexpressed in molecular phenotype 2 samples (5). As shown inFig. 3B, differential expression of AURKB between molecularphenotypes 1 and 2 was maintained; molecular phenotype 1cells had lower expression levels of AURKB, whereas molecularphenotype 2 cells had higher expression levels, providing therationale to next assess the activity of a AURKB-Luc reporterconstruct (20). The AURKB 515 base pair minimal promoteralone showed basal activity in all four osteosarcoma cell linesthat was consistent with observed endogenous AURKB tran-script abundance (Fig. 3C). Reporter activity was not furtherenhanced when we used an AURKB promoter that included1000 base pairs upstream from the transcriptional start site(20), indicating that the full complement of activity for thispromoter in osteosarcoma cells was contained within the min-imal promoter sequence.
Ectopic RB Partially Rescues the Effects of RB-E2F Deregula-tion in Vitro—The minimal AURKB promoter reporter doesnot contain the CCAGGCTGG sequence or the canonical E2Fbinding DNA motif (TTTCCCGC). However, it contains thecell cycle-dependent element (CDE; GGGCGG) that is respon-sive to E2F-mediated transcriptional activation (20, 30). Toevaluate the role of RB-E2F1, we used both the AURKB-Lucreporter and a CDE mutant AURKB-Luc reporter.
Canine osteosarcoma cell lines were co-transfected withAURKB reporter and an N-terminally truncated, RB pocketprotein domain (WT RB) or the same construct containingseven mutations that render the protein insensitive to CDKinactivation (PSM-7LP RB). Introduction of a CMV empty vec-tor control (21) did not alter the activity of the AURKB reporter,whereas ectopic expression of RB inhibited AURKB promoteractivity in the four cell lines (Fig. 4, A and B). However, only theCDK-insensitive RB construct decreased the activity of the
AURKB reporter in OSCA-32 cells, and the effect was modestwhen compared with that observed in the OSCA-40, OSCA-78,and OSCA-8 cell lines, where both RB plasmids showedapproximately equal repression (Fig. 4B).
Repression by ectopic PSM-7LP RB was significantly attenu-ated in each of the four cell lines when we used a CDE mutantAURKB promoter, although partial repression was stillobserved in the molecular phenotype 2 cell lines (Fig. 4C). ChIPanalysis confirmed that endogenous canine E2F1 binds to theCDE in the AURKB reporter and that E2F1 had lower affinityfor the mutant CDE site (Fig. 4C).
The effect of ectopic PSM-7LP RB to suppress the AURKBreporter also was rapidly saturable in OSCA-32 cells, consistentwith the presence of active endogenous RB (Fig. 4D). In con-trast, ectopic PSM-7LP RB showed dose-dependent suppres-sion in OSCA-40, OSCA-78, and OSCA-8 cells (Fig. 4D), aswould be predicted by absence of endogenous functional RB.
Ectopic RB Displaces E2F1 from Endogenous Promoters—Given the repressive effect of ectopic PSM-7LP RB on theectopic AURKB vector in cells representing the two molecularphenotypes of osteosarcoma, we investigated whether theseeffects were relevant and reproducible in the endogenous con-text. Ectopic PSM-7LP RB protein was detectable in all of thecell lines (Fig. 5A), and the presence of ectopic RB consistentlyreduced transcript abundance of AURKB and three other genes(AURKA, BUB1B, and TOP2A) that were part of the signaturethat identified osteosarcoma molecular phenotypes (5)GAPDH transcript abundance was not affected by the presenceof ectopic PSM-7LP RB (Fig. 5B).
To further validate the importance of the E2F-dependentinteractions in the study context, we next confirmed that E2F1was bound to the endogenous TTTCCAGCCAAT motif in thecanine AURKB promoter and that its presence on the promotercontributed to transcription of AURKB. ChIP assays (Fig. 6A)showed greater enrichment for E2F1 bound to endogenousAURKB promoter in OSCA-8 cells (molecular phenotype 2)than in OSCA-32 cells (molecular phenotype 1). Furthermore,E2F1 binding to the promoter in both molecular phenotypeswas reduced in the presence of ectopic PSM-7LP RB. Quanti-tative assessment of AURKB transcript abundance by qRT-PCRwas consistent with what was observed in our ChIP data, show-ing that ectopic PSM-7LP RB caused a greater magnitude ofreduction of AURKB transcript in OSCA-8 cells versusOSCA-32 cells (data not shown). We observed a similar effectupon analysis of E2F1 binding to the CGGCGCTAAA motif ofthe endogenous CHEK1 promoter (Fig. 6B), illustrating thatectopic PSM-7LP RB was repressing the binding of E2F1 pro-tein to genes associated with G2/M progression and the DNAdamage checkpoint.
Importantly, we did not see any difference in E2F1 proteinabundance in the cells transfected with either the ectopic PSM-7LP RB or CMV plasmids, suggesting that ectopic RB was notsimply reducing E2F1 protein levels. PSM-7LP RB also was notfound in complexes bound to the E2F1 response elements in theendogenous AURKB promoter as determined by ChIP inOSCA-8 cells using an antibody that specifically recognized theectopic human RB protein (data not shown). We similarly didnot find evidence of complex formation between ectopic PSM-
FIGURE 2. Histopathology findings. A and B show low magnificationphotomicrographs of OSCA-32 and OS-40 tumors, respectively. Noteorderly deposition of osteoid seams (indicated by asterisk) in an OSCA-32tumor (A) in contrast to the poorly organized osteoid matrix in an OSCA-40tumor (B); also note area of necrosis (indicated by asterisk). Bars, 200 �m. Cand D show high magnification photomicrographs of OSCA-32 and OS-40tumors, respectively. Relatively well differentiated osteoblastic cells laydown osteoid (indicated by asterisk) in an OSCA-32 tumor (C). In contrast,note proliferating spindle cells with numerous mitotic figures (arrow-heads) within poorly organized matrix (osteoid) of an OSCA-40 tumor (D).Bars, 50 �m.
Aberrant RB-E2F Defines Molecular Phenotypes
28074 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7LP RB and endogenous E2F1 in co-immunoprecipitationassays; yet we also did not see a quantitative difference in theamount of immunoprecipitated E2F1 protein in cells fromeither molecular phenotype transfected with PSM-7LP RB orwith the CMV empty vector, suggesting that ectopic PSM-7LPRB did not compete with or sterically hinder binding of theanti-E2F1 antibody (data not shown). Thus, we favor the inter-pretation that displacement of E2F1 from the AURKB pro-moter was an indirect effect of RB.
Ectopic RB Alters the Transcriptional Landscape of Molecu-lar Phenotype 2 Osteosarcoma Cells toward One ResemblingMolecular Phenotype 1—The selective effects of RB on molec-ular phenotype 2 osteosarcoma cells led us to hypothesize thatrestoration of RB in these cells would shift the genome-widetranscriptional state in these cells to one resembling that ofmolecular phenotype 1 cells. Genome-wide expression pro-
filing highlighted 78 genes that were differentially expressedin OSCA-78 cells transfected with PSM-7LP RB versus theCMV control; these genes were significantly associated withfunctions of DNA replication, metabolism of DNA, and bindingof chromatin (Table 2). The most significant canonical pathwayidentified by IPA was cell cycle control (data not shown).Importantly, active RB, and inactive E2F1 and E2F1 wereamong the most significant predicted transcriptional regulatorsassociated with the differential expression of these 78 genes(Table 3). In addition, IPA yielded a number of predicted inac-tivated oncogenes that have been shown to be aberrantlyexpressed in osteosarcoma, including MYC (31), as well as tran-scriptional regulators associated with DNA damage repair andthe mitotic checkpoint, including TBX2 (32) (Table 3). Intrigu-ingly, the gene expression profiles resulting from restored RBactivity in these cells reflected an apparent recovery of activityfor the TP53 tumor suppressor gene (Table 3).
Regulation of AURKB by RB Does Not Require p16 —Whereashuman U2-OS cells retain wild type functional RB, humanSAOS-2 cells have a mutated RB (33, 34). As shown in Fig. 5A,consistent with observations from canine samples, RB-repleteU2-OS cells had lower basal AURKB luciferase activity in com-parison to RB-deficient SAOS-2 cells (average AURKB-Lucif-erase/Renilla RLU: 0.43 and 41.01, respectively) (Fig. 7A).Ectopic PSM-7LP RB did not significantly repress expression ofthe AURKB reporter in U2-OS cells after 6 h (p � 0.150),whereas AURKB reporter activity was significantly (p � 0.002)reduced (�30%) in SAOS-2 cells after 6 h (Fig. 7B).
In U2-OS cells, p16 protein is silenced by methylation of thepromoter (33). Loss of p16 can lead to unrestrained activationof CDK4/6 and may impair function of RB at the G0/G1 transi-tion checkpoint, during G1 progression and at cell cycle exit(re-entry to G0). We hypothesized that p16 deficiency wouldnot be equivalent to complete loss of RB, as other mechanismsof control are operative during progression through the S phaseand the G2/M phase (35, 36). Therefore, we tested whetherCDK inhibition occurred in p16-deficient U2-OS cells upon RBactivation. First, we confirmed that U2-OS cells do not expressp16 protein by Western blot (Fig. 7C). We then determinedwhether RB was completely inactivated in U2-OS cells by cul-turing them under conditions of serum deprivation, whichleads to growth arrest. Within 72 h of serum withdrawal, RBwas predominantly present in the active, faster migrating(hypophosphorylated) form, indicating that, despite silencingof p16, CDKs can still be inhibited in U2-OS cells (Fig. 7D).
As we observed in canine OSCA-32 cells, transfection ofU2-OS cells with the CDK-insensitive PSM-7LP RB led to mod-est repression (30 – 40%; p � 0.047) of the AURKB reporterafter 24 h (Fig. 7E). Ectopic expression of the CMV empty vec-tor or p21 (CDKN1A) pan-CDK inhibitor did not significantlyreduce AURKB Luc activity (p � 0.261, and p � 0.128, respec-tively). Similarly, ectopic expression of the p16 (CDKN2A)CDK4/6 inhibitor had no effect (p � 0.484) (Fig. 7E).
HDAC and DNMT Inhibitors Alter Genome Wide GeneExpression in Molecular Phenotype 2 Cells to a Status Associ-ated with a Functional RB-E2F Regulatory Network (MolecularPhenotype 1)—RB protein is known to associate with HDACsand DNMTs (14, 15). We reasoned that the absence of func-
FIGURE 3. Expression of AURKB in osteosarcoma phenotypes is inverselyrelated to endogenous RB protein and cells from tumors derived from dogswith shorter median survival times (molecular phenotype 2) behaveaggressively in vivo. A, steady state levels of RB protein as determined by West-ern blotting (�-actin, loading control). B, average value (�S.D.) for AURKB expres-sion from two Affymetrix probes (Gene Expression Omnibus accession numberGSE27217) (5). C, AURKB minimal promoter as determined by dual luciferaseassays in the four cell lines (means � S.D. of duplicate experiments).
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28075
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

tional RB would hinder the function of these chromatin-re-modeling enzymes and that RB activity might be restoredthrough pharmacological modulation. To assess the effects ontranscription caused by treatment with DNMT and HDAC
inhibitors, we did genome-wide expression profiling of molec-ular phenotype 2 cells treated with Zeb and SAHA. Fig. 8 (A andB) shows that in molecular phenotype 2 cells (OSCA-78) cells,the transcriptional state of the defining prognostic signature
Aberrant RB-E2F Defines Molecular Phenotypes
28076 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from
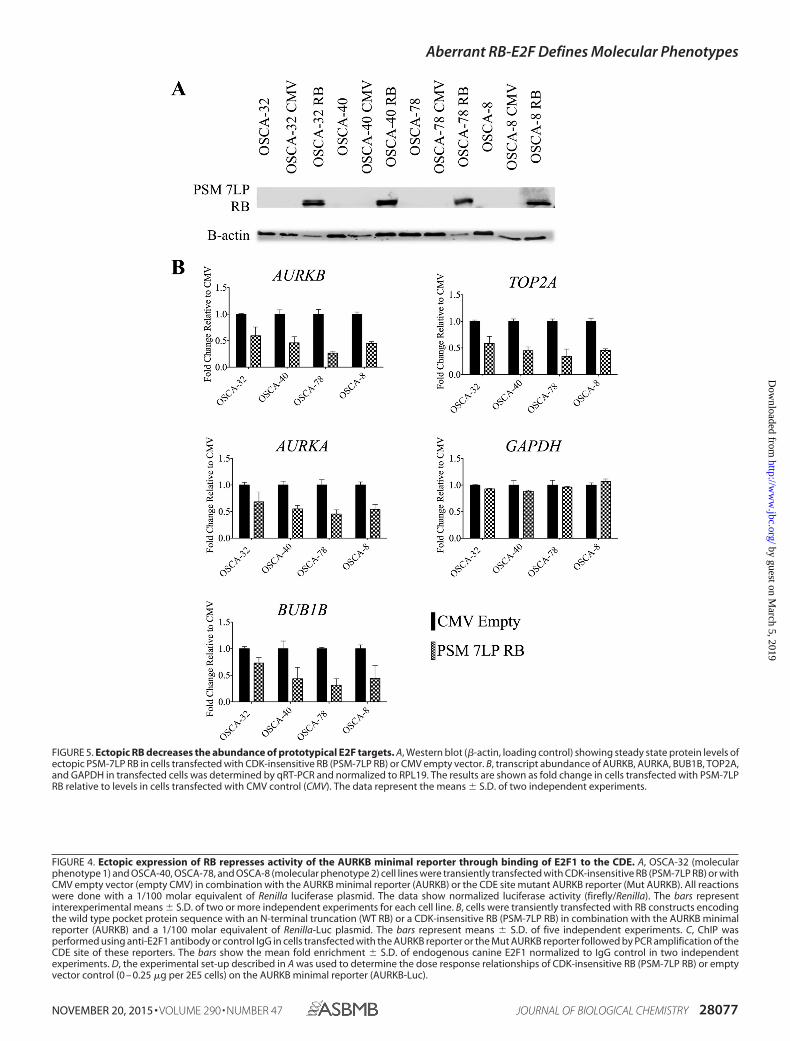
FIGURE 4. Ectopic expression of RB represses activity of the AURKB minimal reporter through binding of E2F1 to the CDE. A, OSCA-32 (molecularphenotype 1) and OSCA-40, OSCA-78, and OSCA-8 (molecular phenotype 2) cell lines were transiently transfected with CDK-insensitive RB (PSM-7LP RB) or withCMV empty vector (empty CMV) in combination with the AURKB minimal reporter (AURKB) or the CDE site mutant AURKB reporter (Mut AURKB). All reactionswere done with a 1/100 molar equivalent of Renilla luciferase plasmid. The data show normalized luciferase activity (firefly/Renilla). The bars representinterexperimental means � S.D. of two or more independent experiments for each cell line. B, cells were transiently transfected with RB constructs encodingthe wild type pocket protein sequence with an N-terminal truncation (WT RB) or a CDK-insensitive RB (PSM-7LP RB) in combination with the AURKB minimalreporter (AURKB) and a 1/100 molar equivalent of Renilla-Luc plasmid. The bars represent means � S.D. of five independent experiments. C, ChIP wasperformed using anti-E2F1 antibody or control IgG in cells transfected with the AURKB reporter or the Mut AURKB reporter followed by PCR amplification of theCDE site of these reporters. The bars show the mean fold enrichment � S.D. of endogenous canine E2F1 normalized to IgG control in two independentexperiments. D, the experimental set-up described in A was used to determine the dose response relationships of CDK-insensitive RB (PSM-7LP RB) or emptyvector control (0 – 0.25 �g per 2E5 cells) on the AURKB minimal reporter (AURKB-Luc).
FIGURE 5. Ectopic RB decreases the abundance of prototypical E2F targets. A, Western blot (�-actin, loading control) showing steady state protein levels ofectopic PSM-7LP RB in cells transfected with CDK-insensitive RB (PSM-7LP RB) or CMV empty vector. B, transcript abundance of AURKB, AURKA, BUB1B, TOP2A,and GAPDH in transfected cells was determined by qRT-PCR and normalized to RPL19. The results are shown as fold change in cells transfected with PSM-7LPRB relative to levels in cells transfected with CMV control (CMV). The data represent the means � S.D. of two independent experiments.
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28077
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

shifted to a state that resembled molecular phenotype 1(OSCA-32) after treatment with Zeb and SAHA (5). Next, weexamined whether the observed effect was associated with dis-placement of E2F1 from relevant promoters. ChIP showed thatthe amount of E2F1 bound to the endogenous AURKB (Fig. 8C)promoter was decreased by �90% in molecular phenotype 1cells and by 95% in molecular phenotype 2 cells, after treatmentwith these drugs. Similarly, the amount of E2F1 bound to theendogenous CHEK1 promoter decreased by �80% in bothmolecular phenotype 1 and molecular phenotype 2 cells aftertreatment (Fig. 8C).
In addition, we identified 1047 statistically significant differ-entially expressed genes (p � 0.05 and average fold change of2.0) in untreated and treated OSCA-78 cells. These genes clus-tered into two groups (data not shown). The first group ofgenes, which was significantly associated with functions of cellcycle progression and proliferation, was down-regulated intreated cells. The second group, which was significantly associ-ated with functions of cellular organization, maintenance, andcell-cell interactions, was up-regulated in treated cells. As asingle group, these genes were significantly associated withfunctions related to cell cycle, proliferation, and cancer func-tions (data not shown).
The IPA transcription factor module was used to predictupstream regulators of the 1047 genes that were differentiallyexpressed between cells treated with DNMT and HDAC inhib-itors and those that were not. RB was a predicted upstreamtranscriptional regulator (Table 4) and was predicted as beingactive in treated cells. The predicted activity of other upstreamtranscriptional regulators for these 1047 genes paralleled thatobserved in molecular phenotype 1 cells (Table 1), as well asthat seen upon ectopic reintroduction of RB into molecular
phenotype 2 cells (Table 2). SMARCB1 was predicted as beingactivated in treated cells, whereas E2F1, MYC, and FOXM1were predicted as being inactivated (Table 4).
Discussion
Here we provide insight into mechanisms that account for aprognostic gene signature that reduces the heterogeneity asso-ciated with osteosarcoma. As shown in our model Fig. 9, thesignature allowed us to group the disease into two subgroups(molecular phenotype 1 and molecular phenotype 2) that differin their biological behavior; i.e. time to progression and clinicaloutcomes (5).
We observed that molecular phenotype 1- and molecularphenotype 2-derived xenograft tumors recapitulated the grossand histologic features of spontaneous canine osteosarcoma.
FIGURE 6. RB-mediated repression of endogenous AURKB associated with displacement of E2F1. A, ChIP was performed using anti-E2F1 antibody orcontrol IgG and enrichment of the E2F1 DNA binding motif TTTCCAGCCAAT in the endogenous canine AURKB promoter in molecular phenotype 1 (OSCA-32,left panel) and molecular phenotype 2 (OSCA-8, right panel) cells transfected with the CDK-insensitive RB (PSM-7LP RB) or empty vector control was determinedby qRT-PCR. The bars represent the means � S.D. of duplicate reactions from one of six experiments done with similar results. B, ChIP to determine enrichmentof E2F1 DNA binding to the endogenous CHEK1 promoter in OSCA-32 cells (left panel) and OSCA-8 cells (right panel) transfected with CMV empty vector or withthe CDK-insensitive RB (PSM-7LP RB).
TABLE 2Gene function enrichment analysis after RB is ectopically restored inmolecular phenotype cellsGenome-wide expression profiling was used to compare OSCA-78 cells transfectedwith CDK-insensitive RB (PSM-7LP RB) to OSCA-78 cells transfected with CMVempty vector. Genes with a p value of �0.05 and an average fold change of 1.53 wereidentified for further analysis. IPA was applied to identify biological functions asso-ciated with differentially expressed genes in molecular phenotype 2 osteosarcomaRB restored cells.
Functions annotation p value
Metabolism of DNA 6.95E-14DNA replication 7.76E-14Synthesis of DNA 3.22E-12Initiation of replication of DNA 1.13E-09Interphase 1.52E-08Binding of chromatin 2.33E-08Checkpoint control 3.98E-08S phase 5.79E-08Repair of DNA 1.11E-07Proliferation of tumor cell lines 1.73E-07
Aberrant RB-E2F Defines Molecular Phenotypes
28078 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from
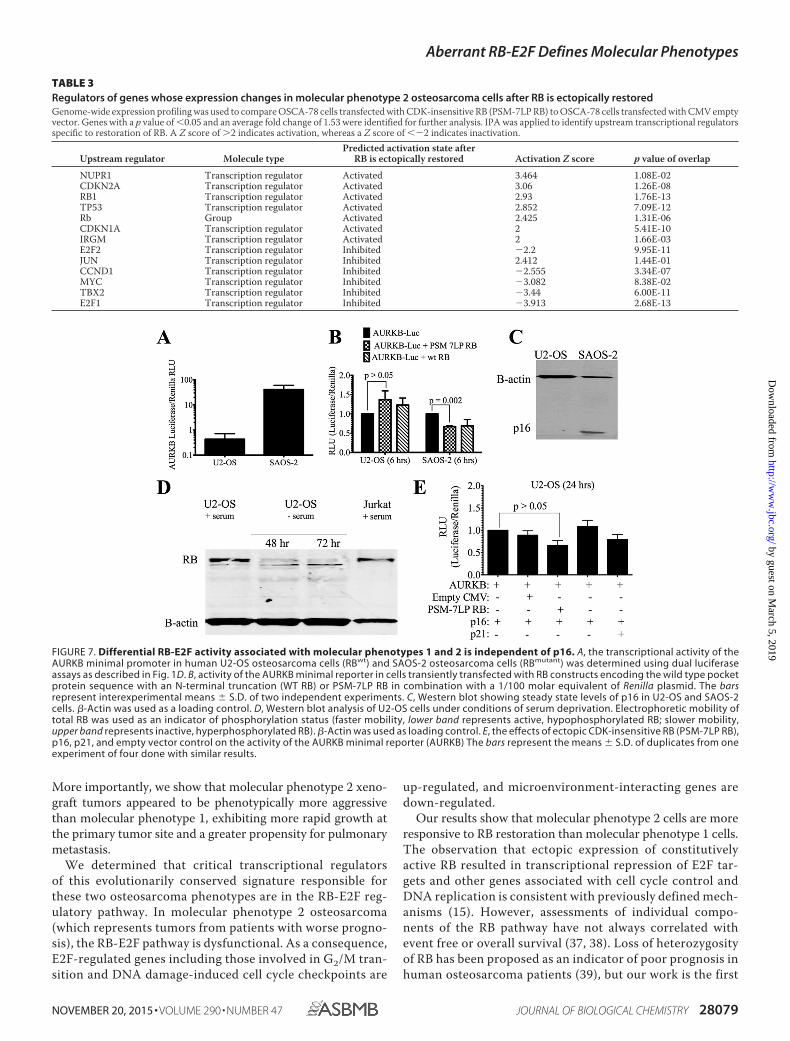
More importantly, we show that molecular phenotype 2 xeno-graft tumors appeared to be phenotypically more aggressivethan molecular phenotype 1, exhibiting more rapid growth atthe primary tumor site and a greater propensity for pulmonarymetastasis.
We determined that critical transcriptional regulatorsof this evolutionarily conserved signature responsible forthese two osteosarcoma phenotypes are in the RB-E2F reg-ulatory pathway. In molecular phenotype 2 osteosarcoma(which represents tumors from patients with worse progno-sis), the RB-E2F pathway is dysfunctional. As a consequence,E2F-regulated genes including those involved in G2/M tran-sition and DNA damage-induced cell cycle checkpoints are
up-regulated, and microenvironment-interacting genes aredown-regulated.
Our results show that molecular phenotype 2 cells are moreresponsive to RB restoration than molecular phenotype 1 cells.The observation that ectopic expression of constitutivelyactive RB resulted in transcriptional repression of E2F tar-gets and other genes associated with cell cycle control andDNA replication is consistent with previously defined mech-anisms (15). However, assessments of individual compo-nents of the RB pathway have not always correlated withevent free or overall survival (37, 38). Loss of heterozygosityof RB has been proposed as an indicator of poor prognosis inhuman osteosarcoma patients (39), but our work is the first
TABLE 3Regulators of genes whose expression changes in molecular phenotype 2 osteosarcoma cells after RB is ectopically restoredGenome-wide expression profiling was used to compare OSCA-78 cells transfected with CDK-insensitive RB (PSM-7LP RB) to OSCA-78 cells transfected with CMV emptyvector. Genes with a p value of �0.05 and an average fold change of 1.53 were identified for further analysis. IPA was applied to identify upstream transcriptional regulatorsspecific to restoration of RB. A Z score of �2 indicates activation, whereas a Z score of �2 indicates inactivation.
Upstream regulator Molecule typePredicted activation state after
RB is ectopically restored Activation Z score p value of overlap
NUPR1 Transcription regulator Activated 3.464 1.08E-02CDKN2A Transcription regulator Activated 3.06 1.26E-08RB1 Transcription regulator Activated 2.93 1.76E-13TP53 Transcription regulator Activated 2.852 7.09E-12Rb Group Activated 2.425 1.31E-06CDKN1A Transcription regulator Activated 2 5.41E-10IRGM Transcription regulator Activated 2 1.66E-03E2F2 Transcription regulator Inhibited 2.2 9.95E-11JUN Transcription regulator Inhibited 2.412 1.44E-01CCND1 Transcription regulator Inhibited 2.555 3.34E-07MYC Transcription regulator Inhibited 3.082 8.38E-02TBX2 Transcription regulator Inhibited 3.44 6.00E-11E2F1 Transcription regulator Inhibited 3.913 2.68E-13
FIGURE 7. Differential RB-E2F activity associated with molecular phenotypes 1 and 2 is independent of p16. A, the transcriptional activity of theAURKB minimal promoter in human U2-OS osteosarcoma cells (RBwt) and SAOS-2 osteosarcoma cells (RBmutant) was determined using dual luciferaseassays as described in Fig. 1D. B, activity of the AURKB minimal reporter in cells transiently transfected with RB constructs encoding the wild type pocketprotein sequence with an N-terminal truncation (WT RB) or PSM-7LP RB in combination with a 1/100 molar equivalent of Renilla plasmid. The barsrepresent interexperimental means � S.D. of two independent experiments. C, Western blot showing steady state levels of p16 in U2-OS and SAOS-2cells. �-Actin was used as a loading control. D, Western blot analysis of U2-OS cells under conditions of serum deprivation. Electrophoretic mobility oftotal RB was used as an indicator of phosphorylation status (faster mobility, lower band represents active, hypophosphorylated RB; slower mobility,upper band represents inactive, hyperphosphorylated RB). �-Actin was used as loading control. E, the effects of ectopic CDK-insensitive RB (PSM-7LP RB),p16, p21, and empty vector control on the activity of the AURKB minimal reporter (AURKB) The bars represent the means � S.D. of duplicates from oneexperiment of four done with similar results.
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28079
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

to establish a direct link between a conserved, prognosticgenome-wide gene expression signature and deregulation ofthe RB-E2F pathway in osteosarcoma (5). More specifically,current models of the RB-E2F pathway do not consistentlyaccount for expression levels of genes during the G2/M phaseof cell cycle (40, 41); yet many of the genes that are overex-pressed in molecular phenotype 2 osteosarcoma, includingAURKB, are associated with the G2/M transition. Recentdata suggest that, unlike canonical E2F response elementsthat operate primarily in G1/S, the binding of E2F to the CDEis stronger in G2/M (30).
One interesting finding from our study was that restorationof RB compensated for other common genetic alterations (e.g.TP53 and MYC) associated with osteosarcoma. The findingsuggests that RB loss or deregulation can modify tumor behav-ior and disease progression downstream of oncogenes that areoften altered in osteosarcoma. Although some of these effectscould be due to direct modulation of E2F activity, it also is likelythat indirect changes in the epigenetic landscape that are estab-lished by functional RB contribute to these effects. The inter-pretation that functional RB utilizes mechanisms that are inde-pendent of E2F binding is consistent with the observation thatmutating the CDE sequence did not completely abrogatereduced AURKB activity upon reintroduction of a functionalRB gene to molecular phenotype 2 cells. Additional support forour interpretation was the observation that mutating the CDEsequence also did not completely abolish E2F1 binding and thatwe did not see evidence of RB binding to the E2F-responsiveelements in the AURKB promoter.
An important finding from our study was that treatmentwith DNMT and HDAC inhibitors was sufficient to supplant
RB-E2F pathway function in molecular phenotype 2 cells.Treatment with these inhibitors altered genome wide geneexpression in molecular phenotype 2 cells to a status associ-ated with a functional RB-E2F regulatory network (as seen inmolecular phenotype 1). Moreover, we show that treatmentwith DNMT and HDAC inhibitors achieved comparabletranscriptional changes as genetic restoration of RB, albeitthrough regulation of somewhat different gene sets. Still,these gene sets were concentrated in or near control nodesfor overlapping biochemical pathways, which is not entirelyunexpected, given the expectation that chromatin-remodel-ing enzymes would have a broader effect to modulate geneexpression.
The observation that p16-deficient U2-OS cells maintainedat least a partially functional RB, possibly through compensa-tion of other CDK inhibitors like p21, indicates that by itself,the status of p16 cannot explain the RB-dependent hetero-geneity of osteosarcoma. Specifically, our findings show thatRB-E2F pathway are still able to regulate genes associatedwith the G2/M transition in U2-OS cells, probably throughinhibition of the S phase and G2/M CDKs. Nevertheless,CDKN2A deletion or silencing could contribute to deregula-tion of the E2F pathway in osteosarcoma (42). Not surpris-ingly, CDKN2A was a predicted transcriptional regulator ofthe prognostic gene signature. The CDKN2A locus wasrecently linked to osteosarcoma risk in dogs, and the riskallele is “fixed” in certain breeds like Rottweilers and IrishWolfhounds (43). This finding illuminates the need to inves-tigate an explanation for why canine osteosarcoma is sooften a highly aggressive disease (5).
FIGURE 8. Treatment of molecular phenotype 2 osteosarcoma cells with HDAC and DNMT inhibitors shifts their transcriptional state to resemble thatof molecular phenotype 1 osteosarcoma cells. A, genome-wide microarray transcriptional profiling was used to define the effects of treatment with DNMTand HDAC inhibitors (Zeb and SAHA) in molecular phenotype 2 cells (OSCA-78) and to compare these transcriptional changes to the basal state of molecularphenotype 1 cells (OSCA-32). Left panel, expression levels of evolutionary conserved prognostic gene signature genes (n � 255) in molecular phenotype 1 cells(OSCA-32) and molecular phenotype 2 cells (OSCA-78) (Gene Expression Omnibus accession number GSE27217). Right panel, expression levels in replicates ofZeb SAHA-treated and untreated OSCA-78 cells (5). Up-regulated genes are in red, and down-regulated genes are in green. B, subset of G2/M transition andDNA damage genes, including AURKB, from previously reported prognostic signature. C, ChIP was done as in Fig. 3A to determine enrichment of the E2F1 DNAbinding motif (TTTCCAGCCAAT) in the endogenous canine AURKB promoter in untreated and Zeb SAHA-treated molecular phenotype 1 cells (OSCA-32, leftpanel) and molecular phenotype 2 cells (OSCA-8, right panel).
Aberrant RB-E2F Defines Molecular Phenotypes
28080 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

It is widely accepted that RB inactivation is not necessaryfor the development of osteosarcoma but rather acceleratesits development and progression (44, 45). Here, we show thatthe integrity of the RB-E2F pathway is mechanistically asso-ciated with the biological behavior of tumor cells derivedfrom spontaneous canine osteosarcoma in vitro and in vivoand that the RB-E2F molecular regulatory network extendsto human osteosarcoma cells. Specifically, our data suggestthat alternative treatment options that create a state analo-gous to that seen with functional RB could improve out-comes in osteosarcoma patients, especially those with theworst prognoses. Our data provide support for further eval-uation of the mechanistic role of RB-E2F pathway in chro-matin remodeling and the contribution of the epigeneticlandscape in osteosarcoma pathogenesis.
Author Contributions—M. C. S. generated data, led the analysis andinterpretation of data, prepared manuscript figures, and wrote thepaper. A. L. S. generated data, contributed to bioinformatics dataanalysis and interpretation, and performed a critical analysis of themanuscript. H. T. conducted xenograft experiments, provided tech-nical assistance, and contributed to the preparation of the xenograftfigures and text. J. V. E. and J. V. conducted qRT-PCR analysis andrevised the manuscript. I. C. and M. G. O. did histopathology ofxenograft tumors and prepared the histopathology figure and text.S. S. offered advice on interpretation of data and gave a critical anal-ysis of the manuscript during its preparation. J. F. M. conceived anddesigned the study, supervised laboratory experiments, and assistedin writing the manuscript. All authors reviewed the results andapproved the final version of the manuscript.
Acknowledgments—We thank the Minnesota Supercomputing Insti-tute for computational resources and especially Dr. Ying Zhang forproviding support and advice. We thank Dr. Aric Frantz for genera-tion of OSCA-40-G/L cells, Dr. Ramesh Kovi for assistance with path-ological analyses, Mitzi Lewellen for assistance with in vivo experi-ments, and LeAnn Oseth and the MCC cytogenetics core for assistancewith FISH. We also thank Rachit Gupta and Frances Phan for tech-nical assistance and Drs. Dai Ito, Ali Khammanivong, and Siu ChiuChan for technical advice. We also thank Drs. Eric Knudsen andMasashi Kimura for providing constructs and Drs. David Largaes-pada, Logan Spector, Scott Dehm, Tim Hallstrom, and Richard Gor-lick for helpful discussions and for review of the manuscript.
References1. Martin, J. W., Squire, J. A., and Zielenska, M. (2012) The genetics of os-
teosarcoma. Sarcoma 2012, 6272542. Dobson, J. M. (2013) Breed-predispositions to cancer in pedigree dogs.
ISRN Vet. Sci. 2013, 9412753. Tan, M. L., Choong, P. F., and Dass, C. R. (2009) Osteosarcoma: Conven-
tional treatment vs. gene therapy. Cancer Biol. Ther. 8, 106 –1174. Thomas, R., Wang, H. J., Tsai, P. C., Langford, C. F., Fosmire, S. P., Jubala,
C. M., Getzy, D. M., Cutter, G. R., Modiano, J. F., and Breen, M. (2009)Influence of genetic background on tumor karyotypes: evidence for breed-associated cytogenetic aberrations in canine appendicular osteosarcoma.
FIGURE 9. Working model of RB-E2F regulated gene expression in thetwo molecular phenotypes of osteosarcoma. In molecular phenotype 1osteosarcoma (left side), functional RB down-regulates E2F activity andrestricts expression of genes associated with cell cycle progressionthrough its interaction with the E2F DNA binding sequences. In molecularphenotype 2 osteosarcoma (right side), RB is absent or nonfunctional andcannot form stable RB-E2F complexes, leading to deregulation of E2F-responsive targets (indicated by black shapes in figure). Restoration of RBor treatment with DNMT and HDAC inhibitors shifts the transcriptionalstate of genes associated with molecular phenotype 2 osteosarcoma(rapid progression and worse prognosis) to a transcriptional state associ-ated with functional RB and molecular phenotype 1 osteosarcoma (lessrapid progression and better prognosis).
TABLE 4Regulators of genes whose expression significantly changes in molecular phenotype 2 osteosarcoma cells after treatment with DNMT and HDACinhibitorsGenome-wide expression profiling was used to compare OSCA-78 cells treated with DNMT and HDAC inhibitors to untreated OSCA-78 cells. Genes with a p value of�0.05 and an average fold change of 1.53 were identified for further analysis. IPA was applied to identify upstream transcriptional regulators specific to treatment withDNMT and HDAC inhibitors. A Z score of �2 indicates activation, whereas a Z score of �2 indicates inactivation.
Upstream regulator Molecule typePredicted activation status after treatment
with DNMT and HDAC inhibitors Activation Z score p value of overlap
FOXO3 Transcription regulator Activated 2.224 6.58E-06RB1 Transcription regulator Activated 2.200 1.19E-05HNF4A Transcription regulator Activated 2.114 1.17E-04EPAS1 Transcription regulator Activated 2.362 6.75E-04Rb Group Activated 2.528 4.46E-03SMARCB1 Transcription regulator Activated 2.877 1.89E-02HIF1A Transcription regulator Activated 2.063 2.19E-02TBX2 Transcription regulator Inhibited 3.053 4.34E-06Vegf Group Inhibited 2.882 1.23E-05MYC Transcription regulator Inhibited 3.386 3.16E-04FOXM1 Transcription regulator Inhibited 3.082 4.13E-04E2F1 Transcription regulator Inhibited 2.417 3.52E-03EIF4E translation regulator Inhibited 2.517 5.61E-03ZNF217 Transcription regulator Inhibited 2.000 5.78E-03Ras Group Inhibited 2.150 1.59E-02KLF5 Transcription regulator Inhibited 2.197 3.19E-02JUN Transcription regulator Inhibited 2.013 5.03E-02Hdac Group Inhibited 2.345 7.14E-02ETS1 Transcription regulator Inhibited 2.076 8.35E-02
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28081
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Chromosome Res. 17, 365–3775. Scott, M. C., Sarver, A. L., Gavin, K. J., Thayanithy, V., Getzy, D. M.,
Newman, R. A., Cutter, G. R., Lindblad-Toh, K., Kisseberth, W. C.,Hunter, L. E., Subramanian, S., Breen, M., and Modiano, J. F. (2011)Molecular subtypes of osteosarcoma identified by reducing tumor het-erogeneity through an interspecies comparative approach. Bone 49,356 –367
6. Gorlick, R., Janeway, K., Lessnick, S., Randall, R. L., Marina, N., andCOG Bone Tumor Committee (2013) Children’s Oncology Group’s2013 blueprint for research: bone tumors. Pediatr. Blood Cancer 60,1009 –1015
7. Janeway, K. A., Barkauskas, D. A., Krailo, M. D., Meyers, P. A., Schwartz,C. L., Ebb, D. H., Seibel, N. L., Grier, H. E., Gorlick, R., and Marina, N.(2012) Outcome for adolescent and young adult patients with osteosar-coma: a report from the Children’s Oncology Group. Cancer 118,4597– 4605
8. Rainusso, N., Wang, L. L., and Yustein, J. T. (2013) The adolescent andyoung adult with cancer: state of the art: bone tumors. Curr. Oncol. Rep.15, 296 –307
9. Janeway, K. A., and Maki, R. G. (2012) New strategies in sarcoma therapy:linking biology and novel agents. Clin. Cancer Res. 18, 5837–5844
10. Ranieri, G., Gadaleta, C. D., Patruno, R., Zizzo, N., Daidone, M. G., Hans-son, M. G., Paradiso, A., and Ribatti, D. (2013) A model of study for humancancer: Spontaneous occurring tumors in dogs. Biological features andtranslation for new anticancer therapies. Crit. Rev. Oncol. Hematol. 88,187–197
11. Withrow, S. J., and Wilkins, R. M. (2010) Cross talk from pets to people:translational osteosarcoma treatments. ILAR J. 51, 208 –213
12. Modiano, J. F., Breen, M., Lana, S. E., Ehrhart, E. J., Schaack, J., Duke, R. C.,Cutter, G. C., and Bellgrau, D. (2006) Naturally occurring translationalmodels for development of cancer gene therapy. Gene Ther. Mol. Biol. 10,31– 40
13. Sarver, A. L., Thayanithy, V., Scott, M. C., Cleton-Jansen, A. M., Hogen-doorn, P. C., Modiano, J. F., and Subramanian, S. (2013) MicroRNAs at thehuman 14q32 locus have prognostic significance in osteosarcoma. Orpha-net J. Rare Dis. 8, 7
14. Fiorentino, F. P., Marchesi, I., and Giordano, A. (2013) On the role ofretinoblastoma family proteins in the establishment and maintenance ofthe epigenetic landscape. J. Cell. Physiol. 228, 276 –284
15. Harbour, J. W., and Dean, D. C. (2000) Chromatin remodeling and Rbactivity. Curr. Opin. Cell Biol. 12, 685– 689
16. Thayanithy, V., Park, C., Sarver, A. L., Kartha, R. V., Korpela, D. M., Graef,A. J., Steer, C. J., Modiano, J. F., and Subramanian, S. (2012) Combinatorialtreatment of DNA and chromatin-modifying drugs cause cell death inhuman and canine osteosarcoma cell lines. PLoS One 7, e43720
17. Jubala, C. M., Wojcieszyn, J. W., Valli, V. E., Getzy, D. M., Fosmire, S. P.,Coffey, D., Bellgrau, D., and Modiano, J. F. (2005) CD20 expression innormal canine B cells and in canine non-Hodgkin lymphoma. Vet. Pathol.42, 468 – 476
18. Kim, J. H., Frantz, A. M., Anderson, K. L., Graef, A. J., Scott, M. C., Rob-inson, S., Sharkey, L. C., O’Brien, T. D., Dickerson, E. B., and Modiano, J. F.(2014) Interleukin-8 promotes canine hemangiosarcoma growth by regu-lating the tumor microenvironment. Exp. Cell Res. 323, 155–164
19. Ritt, M. G., Mayor, J., Wojcieszyn, J., Smith, R., Barton, C. L., and Modiano,J. F. (2000) Sustained nuclear localization of p21/WAF-1 upon growtharrest induced by contact inhibition. Cancer Lett. 158, 73– 84
20. Kimura, M., Uchida, C., Takano, Y., Kitagawa, M., and Okano, Y. (2004)Cell cycle-dependent regulation of the human aurora B promoter.Biochem. Biophys. Res. Commun. 316, 930 –936
21. Knudsen, E. S., and Wang, J. Y. (1997) Dual mechanisms for the inhibitionof E2F binding to RB by cyclin-dependent kinase-mediated RB phosphor-ylation. Mol. Cell. Biol. 17, 5771–5783
22. Modiano, J. F., Mayor, J., Ball, C., Fuentes, M. K., and Linthicum, D. S.(2000) CDK4 expression and activity are required for cytokine responsive-ness in T cells. J. Immunol. 165, 6693– 6702
23. Ritt, M. G., Wojcieszyn, J., and Modiano, J. F. (1998) Functional loss ofp21/Waf-1 in a case of benign canine multicentric melanoma. Vet. Pathol.35, 94 –101
24. Modiano, J. F., Bellgrau, D., Cutter, G. R., Lana, S. E., Ehrhart, N. P.,Ehrhart, E., Wilke, V. L., Charles, J. B., Munson, S., Scott, M. C.,Pozniak, J., Carlson, C. S., Schaack, J., and Duke, R. C. (2012) Inflam-mation, apoptosis, and necrosis induced by neoadjuvant fas ligandgene therapy improves survival of dogs with spontaneous bone cancer.Mol. Ther. 20, 2234 –2243
25. Kent, W. J., Sugnet, C. W., Furey, T. S., Roskin, K. M., Pringle, T. H.,Zahler, A. M., and Haussler, D. (2002) The human genome browser atUCSC. Genome Res. 12, 996 –1006
26. Kent, W. J., Sugnet, C. W., Furey, T. S., Roskin, K. M., Pringle, T. H.,Zahler, A. M., Haussler, D. (2002) The human genome browser at UCSC.Genome Res. 12, 996 –1006
27. Goecks, J., Nekrutenko, A., Taylor, J., and Galaxy, T. (2010) Galaxy: acomprehensive approach for supporting accessible, reproducible, andtransparent computational research in the life sciences. Genome Biol. 11,R86
28. Weinmann, A. S., Yan, P. S., Oberley, M. J., Huang, T. H., and Farnham,P. J. (2002) Isolating human transcription factor targets by coupling chro-matin immunoprecipitation and CpG island microarray analysis. GenesDev. 16, 235–244
29. Chitko-McKown, C. G., and Modiano, J. F. (1997) Clues to immune func-tion and oncogenesis provided by events that activate the cell cycle ma-chinery in normal human T cells. J. Leukocyte Biol. 62, 430 – 437
30. Müller, G. A., and Engeland, K. (2010) The central role of CDE/CHRpromoter elements in the regulation of cell cycle-dependent gene tran-scription. FEBS J. 277, 877– 893
31. Thayanithy, V., Sarver, A. L., Kartha, R. V., Li, L., Angstadt, A. Y.,Breen, M., Steer, C. J., Modiano, J. F., and Subramanian, S. (2012)Perturbation of 14q32 miRNAs-cMYC gene network in osteosarcoma.Bone 50, 171–181
32. Wansleben, S., Davis, E., Peres, J., and Prince, S. (2013) A novel role for theanti-senescence factor TBX2 in DNA repair and cisplatin resistance. CellDeath Dis. 4, e846
33. Park, Y. B., Park, M. J., Kimura, K., Shimizu, K., Lee, S. H., and Yokota,J. (2002) Alterations in the INK4a/ARF locus and their effects on thegrowth of human osteosarcoma cell lines. Cancer Genet. Cytogenet.133, 105–111
34. Rogatsky, I., Trowbridge, J. M., and Garabedian, M. J. (1997) Glucocorti-coid receptor-mediated cell cycle arrest is achieved through distinct cell-specific transcriptional regulatory mechanisms. Mol. Cell. Biol. 17,3181–3193
35. Lukas, J., Herzinger, T., Hansen, K., Moroni, M. C., Resnitzky, D., Helin,K., Reed, S. I., and Bartek, J. (1997) Cyclin E-induced S phase withoutactivation of the pRb/E2F pathway. Genes Dev. 11, 1479 –1492
36. Mann, D. J., and Jones, N. C. (1996) E2F-1 but not E2F-4 can overcomep16-induced G1 cell-cycle arrest. Curr. Biol. 6, 474 – 483
37. Heinsohn, S., Evermann, U., Zur Stadt, U., Bielack, S., and Kabisch, H.(2007) Determination of the prognostic value of loss of heterozygosity atthe retinoblastoma gene in osteosarcoma. Int. J. Oncol. 30, 1205–1214
38. Maitra, A., Roberts, H., Weinberg, A. G., and Geradts, J. (2001) Loss ofp16INK4a expression correlates with decreased survival in pediatric os-teosarcomas. Int. J. Cancer 95, 34 –38
39. Feugeas, O., Guriec, N., Babin-Boilletot, A., Marcellin, L., Simon, P., Babin,S., Thyss, A., Hofman, P., Terrier, P., Kalifa, C., Brunat-Mentigny, M.,Patricot, L. M., and Oberling, F. (1996) Loss of heterozygosity of the RBgene is a poor prognostic factor in patients with osteosarcoma. J. Clin.Oncol. 14, 467– 472
40. Dyson, N. (1998) The regulation of E2F by pRB-family proteins. GenesDev. 12, 2245–2262
41. Sadasivam, S., and DeCaprio, J. A. (2013) The DREAM complex: mastercoordinator of cell cycle-dependent gene expression. Nat. Rev. Cancer 13,585–595
42. Ashizawa, S., Nishizawa, H., Yamada, M., Higashi, H., Kondo, T., Ozawa,H., Kakita, A., and Hatakeyama, M. (2001) Collective inhibition of pRBfamily proteins by phosphorylation in cells with p16INK4a loss or cyclin Eoverexpression. J. Biol. Chem. 276, 11362–11370
43. Karlsson, E. K., Sigurdsson, S., Ivansson, E., Thomas, R., Elvers, I., Wright,J., Howald, C., Tonomura, N., Perloski, M., Swofford, R., Biagi, T., Fryc, S.,
Aberrant RB-E2F Defines Molecular Phenotypes
28082 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 290 • NUMBER 47 • NOVEMBER 20, 2015
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Anderson, N., Courtay-Cahen, C., Youell, L., Ricketts, S. L., Mandlebaum,S., Rivera, P., von Euler, H., Kisseberth, W. C., London, C. A., Lander, E. S.,Couto, G., Comstock, K., Starkey, M. P., Modiano, J. F., Breen, M., andLindblad-Toh, K. (2013) Genome-wide analyses implicate 33 loci in her-itable dog osteosarcoma, including regulatory variants near CDKN2A/B.Genome Biol. 14, R132
44. Berman, S. D., Calo, E., Landman, A. S., Danielian, P. S., Miller, E. S., West,
J. C., Fonhoue, B. D., Caron, A., Bronson, R., Bouxsein, M. L., Mukherjee,S., and Lees, J. A. (2008) Metastatic osteosarcoma induced by inactivationof Rb and p53 in the osteoblast lineage. Proc. Natl. Acad. Sci. U.S.A. 105,11851–11856
45. Calo, E., Quintero-Estades, J. A., Danielian, P. S., Nedelcu, S., Berman,S. D., and Lees, J. A. (2010) Rb regulates fate choice and lineage commit-ment in vivo. Nature 466, 1110 –1114
Aberrant RB-E2F Defines Molecular Phenotypes
NOVEMBER 20, 2015 • VOLUME 290 • NUMBER 47 JOURNAL OF BIOLOGICAL CHEMISTRY 28083
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Jyotika Varshney, M. Gerard O'Sullivan, Subbaya Subramanian and Jaime F. ModianoMilcah C. Scott, Aaron L. Sarver, Hirotaka Tomiyasu, Ingrid Cornax, Jamie Van Etten,
Phenotypes of OsteosarcomaAberrant Retinoblastoma (RB)-E2F Transcriptional Regulation Defines Molecular
doi: 10.1074/jbc.M115.679696 originally published online September 16, 20152015, 290:28070-28083.J. Biol. Chem.
10.1074/jbc.M115.679696Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/290/47/28070.full.html#ref-list-1
This article cites 45 references, 12 of which can be accessed free at
by guest on March 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from


















