
Zulato Et Al. - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
7
Review Metabolic effects of anti-angiogenic therapy in tumors Elisabetta Zulato 1 , Matteo Curtarello 1 , Giorgia Nardo, Stefano Indraccolo * Istituto Oncologico Veneto - IRCCS, via Gattamelata, 64 - 35128 Padova, Italy a r t i c l e i n f o Article history: Received 29 July 2011 Accepted 3 January 2012 Available online 11 January 2012 Keywords: Glycolysis Angiogenesis Anti-angiogenic therapy LKB1-AMPK pathway a b s t r a c t Anti-angiogenic therapy has recently been added to the panel of cancer therapeutics, but predictive biomarkers of response are still not available. In animal models, anti-angiogenic therapy causes tumor starvation by increasing hypoxia and impairing nutrients supply. It is thus conceivable that angiogenesis inhi biti on causes remark able met abol ic per tur bat ions in tumors , alt hou gh they remain lar gel y unchart ed. We review here recent acqu isitions about metabolic effects of angiog enesis blockade in tumors and discuss the possibility that some metabolic features of tumor cells - i.e. their dependency from glucose as primary energy substrate - might affect tumor responses to anti-VEGF treatment. 2012 Elsevier Masson SAS. All rights reserved. 1. Introduction The concept of targeting the vasculature of tumors has been vali dated clinic ally as ther apeu tic stra tegy with the appr ova l of several drug s that block the vasc ular endoth elial growth factor (VEGF)- VEGF receptor pathway [1]. In spit e of mu ch ef fort s, however , the ne mecha nisms of tumo r regr essi on caus ed by angiogenesis inhibition remain substantially unknown. In experi- mental tumors anti-VEGF drugs prune the newly formed vascula- ture, thus reducing microvessel density, blood ow and perfusion, and event uall y incr easi ng the hypo xic tumor fract ion [2e4]. In patients, mechanisms might be more complex, especially consid- ering that VEGF neutralization has therapeutic ef cacy mainly if comb ined with conv enti onal chemo ther apy [5]. It has been proposed that anti-VEGF therapy could transiently normalize the tumor vasculature thus improving delivery of drugs and oxygen to tumor cells [6]. Following this initial time window of vascular normalization , sustained VEGF blockade hypothetically leads to regressio n of the tumor vasculature followed by increased hypoxi a [6]. Althou gh it is assumed that in additi on to ox yg en anti- angiogenic drugs cut nutrients supply, the exact identity of the metab olites involved and thei r qua ntitative variatio ns have not been accurately reported so far. Here we present an overview of the predi ctive markers for anti- angiogenic therapy and of the key metabolic alterations occurring in cancer. Moreover, we review the emerging studies connecting anti-angiogenic therapy to metabolism. 2. Predictive mark ers for anti-angioge nic therapy Validated biomarkers to predict the optimal biological dose of anti-angiogenic agents and aiding selection of those patients who ar e mos t likely to benet from anti-angi ogen ic treat ment are currently not available, although a number of potential circulating, tissue and ima gin g biomarkers hav e eme rged fro m rec ent ly completed clinical trials [7]. Since most appr ove d anti- angio genic drugs targ et the VEGF pathw ay , VEGF itself has been the most extens ivel y exp lor ed biomarker. In a study with bevacizumab plus chemotherapy for metastatic breast cancer, lower circulating VEGF levels were asso- ciated with longer Time to Progression (TTP) [8]. Likewise, in lung cancer patients elevated baseline VEGF serum levels had a negative prognostic impact on survival [9] . In contrast, other studies have not found any correlation between VEGF blood concentration and the outcome of anti-angiogenic therapy. In a randomized phase II/ III trial Non-Small Cell Lung Cancer (NSCLC) patients with high levels of baseline plasma VEGF had an increased response to bev- acizumab, but VEGF levels were not predictive of survival bene t [10]. In three randomized phase III studies in metastatic colorectal cancer , lung cancer, and renal cell cancer investigator s assessed the valu e of circ ulati ng VEGF level as a prognost ic and pre dict ive biomarker for outcome of anti-angiogenic therapy. Baseline circu- lating VEGF levels were usefu l as a prognostic biomarke r, but not as a predictive biomarker for bevacizumab-based treatment benet [11]. * Corresponding author. Tel.: þ39 049 8215875; fax: þ39 049 8072854. E-mail address: stefano.indraccolo@un ipd.it (S. Indraccolo). 1 These Authors equally contributed to this work. Contents lists available at SciVerse ScienceDirect Biochimie journal homepage: www.elsevier.com/locate/biochi 0300-9084/$ e see front matter 2012 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.biochi.2012.01.001 Biochimie 94 (2012) 925e931
Transcript of Zulato Et Al. - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 17
Review
Metabolic effects of anti-angiogenic therapy in tumors
Elisabetta Zulato 1 Matteo Curtarello 1 Giorgia Nardo Stefano Indraccolo
Istituto Oncologico Veneto - IRCCS via Gattamelata 64 - 35128 Padova Italy
a r t i c l e i n f o
Article history
Received 29 July 2011
Accepted 3 January 2012
Available online 11 January 2012
Keywords
Glycolysis
Angiogenesis
Anti-angiogenic therapy
LKB1-AMPK pathway
a b s t r a c t
Anti-angiogenic therapy has recently been added to the panel of cancer therapeutics but predictive
biomarkers of response are still not available In animal models anti-angiogenic therapy causes tumor
starvation by increasing hypoxia and impairing nutrients supply It is thus conceivable that angiogenesisinhibition causes remarkable metabolic perturbations in tumors although they remain largely
uncharted We review here recent acquisitions about metabolic effects of angiogenesis blockade in
tumors and discuss the possibility that some metabolic features of tumor cells - ie their dependency
from glucose as primary energy substrate - might affect tumor responses to anti-VEGF treatment
2012 Elsevier Masson SAS All rights reserved
1 Introduction
The concept of targeting the vasculature of tumors has been
validated clinically as therapeutic strategy with the approval of
several drugs that block the vascular endothelial growth factor(VEGF)-VEGF receptor pathway [1] In spite of much efforts
however the 1047297ne mechanisms of tumor regression caused by
angiogenesis inhibition remain substantially unknown In experi-
mental tumors anti-VEGF drugs prune the newly formed vascula-
ture thus reducing microvessel density blood 1047298ow and perfusion
and eventually increasing the hypoxic tumor fraction [2e4] In
patients mechanisms might be more complex especially consid-
ering that VEGF neutralization has therapeutic ef 1047297cacy mainly if
combined with conventional chemotherapy [5] It has been
proposed that anti-VEGF therapy could transiently normalize the
tumor vasculature thus improving delivery of drugs and oxygen to
tumor cells [6] Following this initial time window of ldquovascular
normalizationrdquo sustained VEGF blockade hypothetically leads to
regression of the tumor vasculature followed by increased hypoxia[6] Although it is assumed that in addition to oxygen anti-
angiogenic drugs cut nutrients supply the exact identity of the
metabolites involved and their quantitative variations have not
been accurately reported so far
Here we present an overview of the predictive markers for anti-
angiogenic therapy and of the key metabolic alterations occurring
in cancer Moreover we review the emerging studies connecting
anti-angiogenic therapy to metabolism
2 Predictive markers for anti-angiogenic therapy
Validated biomarkers to predict the optimal biological dose of
anti-angiogenic agents and aiding selection of those patients who
are most likely to bene1047297t from anti-angiogenic treatment are
currently not available although a number of potential circulating
tissue and imaging biomarkers have emerged from recently
completed clinical trials [7]
Since most approved anti-angiogenic drugs target the VEGF
pathway VEGF itself has been the most extensively explored
biomarker In a study with bevacizumab plus chemotherapy for
metastatic breast cancer lower circulating VEGF levels were asso-
ciated with longer Time to Progression (TTP) [8] Likewise in lung
cancer patients elevated baseline VEGF serum levels had a negative
prognostic impact on survival [9] In contrast other studies have
not found any correlation between VEGF blood concentration andthe outcome of anti-angiogenic therapy In a randomized phase II
III trial Non-Small Cell Lung Cancer (NSCLC) patients with high
levels of baseline plasma VEGF had an increased response to bev-
acizumab but VEGF levels were not predictive of survival bene1047297t
[10] In three randomized phase III studies in metastatic colorectal
cancer lung cancer and renal cell cancer investigators assessed the
value of circulating VEGF level as a prognostic and predictive
biomarker for outcome of anti-angiogenic therapy Baseline circu-
lating VEGF levels were useful as a prognostic biomarker but
not as a predictive biomarker for bevacizumab-based treatment
bene1047297t [11]
Corresponding author Tel thorn39 049 8215875 fax thorn39 049 8072854
E-mail address stefanoindraccolounipdit (S Indraccolo)1 These Authors equally contributed to this work
Contents lists available at SciVerse ScienceDirect
Biochimie
j o u r n a l h o m e p a g e w w w e l s e v i e r c o m l o c a t e b i o c h i
0300-9084$ e see front matter 2012 Elsevier Masson SAS All rights reserved
doi101016jbiochi201201001
Biochimie 94 (2012) 925e931
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 27
Some studies tested the hypothesis that an association between
VEGF polymorphisms and response to anti-VEGF therapy could
exist In advanced breast cancer the VEGF-2578AA and VEGF-
1154AA genotypes predicted a favorable Overall Survival (OS) for
patients in the paclitaxel plus bevacizumab arm but did not predict
a better Progression Free Survival (PFS) interval [12] In ovarian
cancer VEGF SNPs did not correlate with PFS [13]
One of the common side-effects of anti-angiogenic drugs is
hypertension [14] Different retrospective studies involving NSCLC
colorectal and renal carcinoma patients reported a signi1047297cant
improve in OS or PFS among patients with bevacizumab-induced
hypertension [15e17] One limitation of these studies however is
the fact that consensus criteria to measure bevacizumab-induced
hypertension are not yet established [11]
Anti-angiogenic drugs are often cytostatic in action and tumor
shrinkage or regression may not be a realistic estimate of ef 1047297cacy
To overcome the lack of correspondence between the Response
Evaluation Criteria in Solid Tumors (RECIST) and survival in patients
treated with anti-VEGF therapy new radiological methods are
emerging as surrogate biomarkers One potential tool for biomarker
development is dynamic contrast-enhanced magnetic resonance
imaging (DCE-MRI) which could provide information about tumor
blood vessel structure and functions [18] The volume transferconstant of contrast agent (ktrans) is a measure of tumor perfusion
and permeability in DCE-MRI In one randomized trial of sorafenib
in renal cell carcinoma high baseline DCE-MRI parameters
including ktrans and V p (blood plasma volume fraction) correlated
with PFS whereas changes of DCE-MRI parameters after the start of
therapy did not predict PFS [19] In recurrent gliobastoma a marked
reduction in ktrans after one dose of cediranib was seen in patients
with increased PFS [20]
The [18F]1047298uorothymidine Positron Emission Tomography
(FLT-PET) is an imaging technique for measuring in vivo cellular
proliferation in malignant tumor and organ tissue and is used to
monitor tumor responses to cytostatic therapies [21] A prospective
study in patients with recurrent malignant gliomas suggests that
FLT-PET can predict responses to bevacizumab as early as 1e
2weeks after treatment [22]
Finally certain metabolism-associated biomarkers could be
helpful in selecting patients to bene1047297t from anti-angiogenic
therapy In the CONFIRM trials LDH-A GLUT-1 and VEGFR1
mRNA levels predicted responses of colorectal cancer patients to
chemotherapy plus vatalanib [23] In the same trials high tissue
LDH5 correlated with poor PFS in the placebo subgroup whereas
it correlated with improved PFS in the vatalanib subgroup [24]
Moreover although vatalanib did not improve either PFS or OS
compared with placebo when authors strati1047297ed patients by
serum LDH level before random assignment they observed that
patients with high serum LDH had longer median PFS when
treated with vatalanib than with placebo In conclusion the
authors proposed that high serum LDH levels may identify tumorsthat are more dependent on abnormal angiogenesis and may
be more susceptible to VEGF inhibition as also suggested by
others [25]
3 Warburg effect and other metabolic alterations in cancer
In the 1920s Otto Warburg af 1047297rmed the role of metabolism in
carcinogenesis by demonstrating that cancer cells - at variance
with normal cells - rely on glycolysis instead of mitochondrial
oxidative phosphorylation (OXPHOS) to produce ATP even under
aerobic conditions [26] Warburg originally hypothesized that the
glycolytic switch in cancer cells was a consequence of defects in
mitochondria which impair aerobic respiration Currently
however it is known that mitochondria are not damaged in most
cancer cells suggesting that aerobic glycolysis essentially repre-
sents an adaptive choice of tumors [27]
Glucose is a critical nutrient for proliferating cells and it is used
as primary substrate to generate ATP as well as to synthesize amino
acids nucleotides and fatty acid and to regulate the redox
potential so as to minimize the effects of reactive oxygen species
(ROS) that damage cellular membranes and proteins [2829]
Enhanced glucose uptake - visualized in the clinic by [18F]1047298uo-
rodeoxyglucose (FDG)-PET - correlates with poor prognosis in
certain tumor types suggesting that enhanced glycolysis confers
a substantial growth advantage [30]
In recent years there has been a number of studies indicating
that aerobic glycolysis is constitutively up-regulated in tumor cells
through genetic or epigenetic changes The 1047297rst documented
mechanistic link between an activated oncogene and altered
glucose metabolism was the transcriptional activation of lactate
dehydrogenase A (LDH-A) by the MYC oncogene [31] LDH-A
contributes a crucial component of the Warburg effect the
conversion of pyruvate e the end-point of glycolysis e to lactate
that is secreted by monocarboxylate carriers (MCTs) eliminating it
from the pool and keeping glycolysis active [32] The secreted
lactate lowers the extracellular pH which may in1047298uence remod-
eling of the matrix and facilitate invasion Furthermore acidosisallows for the selection of motile cells that can eventually break
through the basement membrane and metastasize [29] Indeed
high levels of lactate have been proposed as prognostic factor in
certain malignancies [33] MYC was also found to regulate other
glycolysis-associated genes such as hexokinase II (HK II ) as well as
glucose transporters [34]
The AKT signaling pathway links growth control to glucose
metabolism and several studies correlated its activity with high
glycolytic rates in cancer cells AKT regulates expression of glucose
transporters and HKII enhancing both glucose uptake and its
retention in the cell [35] Moreover AKT can also increase
activity of hypoxia-inducible factor (HIF) thus further enhancing
glycolysis [36]
HIF-1 - which can accumulate due to hypoxia as well as alter-ations of various signaling pathways in tumors - up-regulates
expression of genes involved in aerobic glycolysis including
glucose transporters glycolytic enzymes and LDH-A [37] In addi-
tion HIF-1 deviates pyruvate away from mitochondria by up-
regulating pyruvate dehydrogenase the rate-limiting enzyme for
pyruvate to acetyl-CoA conversion thus alleviating oxidative stress
derived from mitochondrial metabolism [36]
Activated RAS oncogene was initially linked to increase cellular
glucose uptake but recent studies suggest that metabolic effects of
RAS activation could be mediated by MYC and HIF although the
precise mechanism is not yet established [36]
Although the p53 tumor suppressor has been viewed as the
ldquoguardian of the genomerdquo recently it has been implicated in
metabolism control [36] The p53 protein represses transcription of GLUT-1 and GLUT-4 transporters and can in1047298uence the metabolic
balance between glycolysis and OXPHOS through the transcrip-
tional regulation of the 26 efructose bisphosphatase TP53-induce
glycolysis regulator (TIGAR) and synthesis of cytocrome c oxidase
(SCO2) subunit of complex IV of the electron transport chain
[3839]
Besides glucose glutamine could represent an important energy
substrate in cancer cells DeBerardinis et al recently proposed that
in transformed cells glucose accounts mainly for lipid and nucleo-
tide synthesis whereas glutamine is responsible for re-feeding of
the TCA cycle for amino acid synthesis and for nitrogen incorpo-
ration into purine and pyrimidine for nucleotide synthesis [40]
MYC-addicted tumor cells are particularly sensitive to glutamine
withdrawal [41] andgenes involvedin mitocondrial biogenesis and
E Zulato et al Biochimie 94 (2012) 925e931926
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 37
glutamine metabolism specially glutamine transporter and GLS
genes appear to be under both the direct and indirect transcrip-
tional control of MYC [42] The ability of MYC to induceboth aerobic
glycolysis and glutamine oxidation provides cancer cells with the
ability to accumulate biomass [36]
Many human tumor types show altered metabolism of certain
amino acids showing increased uptake and high levels of amino
acid transporter expression
Enhanced expression of L -type amino acid transporter (LAT-1)
which shows high af 1047297nity for several essential amino acids
including leucine tryptophan and methionine (MET) has been
reported in astrocytomas and correlated with poor prognosis [43]
Patients with glioblastoma or other extracranial malignant tumors
such as lung cancer head and neck cancers breast cancer sarcomas
and lymphomas showed increased uptake of MET [44] a feature
which is exploited to make diagnosis and assess therapeutic ef 1047297-
cacy by 18F-MET PET [45]
In addition to increased glycolytic and amino acid activities
recurrent alterations of lipid metabolism are found in cancer cells
conceivably due to their requirement of de novo synthesis of lipids
for membrane assembly Overexpression of fatty acid synthase
(FASN) that catalyzes the de novo synthesis of fatty acids has been
observed in many human cancers including breast prostate lungand colorectal cancers and high levels of FASN were associated
with poor prognosis [46]
Alterations in choline-metabolites (tCho) are also quite
common in cancer cells Tumor cell lines are characterized by an
increased content of phosphocholine (PCho) as compared with
normal epithelial cells [47] The alpha-isoform of Choline Kinase
(ChoK) is often over-expressed in cancer and it is required to
sustain the PCho pool in tumor cells [48] Choline phosphorilation
by ChoK represents the 1047297rst step of choline metabolism in which
choline is 1047297nally converted to phosphatidylcholine a major
constituent of the mammalian cell membrane Choline-
metabolites are of particular interest because they can be moni-
tored in patients by magnetic resonance spectral (MRS) which
detects endogenous PCho or PET which detects altered kinetics of labeled Cho
An interesting area for future studies is to investigate the
predictive and prognostic value of these metabolic features of
cancer cells and to clarify whether they are modulated by anti-
angiogenic therapy
4 Metabolic perturbations after anti-angiogenic therapy
Responses to anti-angiogenic drugs such as sunitinib or bev-
acizumab have been quite heterogeneous in cancer patients In
some cases tumors respond by decreasing tumor volume by more
than 33 qualifying it for a partial response according to RECIST
criteria In other patients however signi1047297cant changes in tumor
density with no decrease in tumor dimensions are observed [49]
This is often associated with central tumor cavitation and necrosis
an observation which suggests that VEGF blockade may perturb the
energy balance in cancer cells
In a recent study [50] we investigated how metabolic param-
eters contribute to determine the pathologic response to VEGF
blockade in tumor xenografts A landmark observation of our study
was that the level of ldquoglucose addictionrdquo of tumor cells dictates the
amount of necrosis caused by angiogenesis inhibition This was
explained by the fact that VEGF blockade acutely perturbs glucose
and ATP levels in tumor xenografts Measurements by biolumi-nescence metabolic imaging indicated that after anti-VEGF therapy
glucose and ATP concentrations in tumors were 130 mmolg and
110 mmolg respectively Values in control tumors were 330 mmol
g (glucose) and 150 mmolg (ATP) Notably glucose uptake
was maintained following anti-angiogenic therapy as shown by
FDG-PET imaging indicating that delivery of glucose through the
vasculature was not compromised despite a substantial decrease in
microvessel density [50] similarly to what has been observed in
patients after bevacizumab monotherapy [51] So it appears that
glucose steady-state levels are very low after anti-angiogenic
therapy whereas glucose uptake is high likely due to HIF-1a
accumulation in treated tumors Intriguingly a preliminary report
showed that a subset of breast cancer patients treated with short-
term 1047297rst-line bevacizumab strongly up-regulated the hypoxiametagene [52] lending support to our observations
Fig 1 Anti-VEGF therapy increases AMPK activation in tumors Representative pictures of pAMPK and pACC staining of ovarian cancer IGROV-1 xenografts following 1 week of
treatment with the anti-VEGF monoclonal antibody A461
E Zulato et al Biochimie 94 (2012) 925e931 927
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 47
A related study investigated metabolic changes in glioblastoma
following anti-VEGF treatment and observed a tendency toward
accumulation of lactate alanine choline myo-inositol creatine
taurine and mobile lipids together with induction of HIF-1a and
activation of the phosphatidyl-inositol-3-kinase pathway [53] This
combination of metabolic changes has previously been associated
with increased hypoxia in human brain tumor spectra [54] and
partially overlaps with our 1047297ndings in ovarian cancer xenografts
[50] In future studies global metabolic changes identi1047297ed by mass
spectrometry analysis (including LC-MSMS and GCeMS) will be
helpful to characterize more extensively metabolic changes
induced by anti-angiogenic therapy in tumors There is already
evidence that this technology enables to pick up speci1047297c oncome-
tabolites in prostate cancer [55] and in gliomas [56]
Fig 2 Parameters contributing to determine tumor responses to VEGF neutralization (A) Anti-angiogenic therapy has been demonstrated to perturb glucose levels in tumors
xenografts Levels of ldquoglucose addictionrdquo of tumor cells in1047298uence the amount of necrosis caused by VEGF blockade Highly glycolytic tumors show a signi1047297cant reduction in their sizeand develop large necrotic areas following short-term anti-angiogenic therapy In contrast poorly glycolytic tumors are only marginally affected in size and do not markedly
increase necrosis after anti-angiogenic therapy [based on [50]] (B) AMPK activation as consequence of perturbations of ATP levels in tumors and its effects on the outcome of VEGF
blockade AMPK activation following short-term anti-angiogenic therapy reduces anabolic processes and cell proliferation being associated with minimal necrosis areas In contrast
tumors that fail to activate this pathway (ie due to LKB1 loss or mutations) maintain a high metabolic demand and are committed to develop large necrotic areas (C) Possible
outlook following tumor necrosis Large necrotic areas may induce recruitment of pro-angiogenic bone marrow-derived cells followed by rapid tumor regrowth Alternatively
killing of the majority of tumor cells by necrosis may turn into a therapeutic effect if relapse mechanisms are not engaged
E Zulato et al Biochimie 94 (2012) 925e931928
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 57
Finally a novel role of the polyamine system in the hypoxic
response of cancer cells has recently been demonstrated The
polyamine system is up-regulated by hypoxia in a variety of cancer
cell lines and in hypoxic tumor regions and inhibition of polyamine
biosynthesis sensitizes cancer cells to hypoxia-induced apoptosis
in vitro [57] These 1047297ndings are relevant in this context if one
considers that the anti-tumor effect of bevacizumab was signi1047297-
cantly enhanced in mice receiving concomitant treatment with the
polyamine biosynthesis inhibitor DFMO which irreversibly inacti-
vates the key enzyme ornithine decarboxylase (ODC) [57] Thus
increased ODC expression and increased intracellular polyamine
levels may occur in tumors treated with anti-angiogenic drugs
likely contributing to protect tumor cells from hypoxia-induced
apoptosis Drugs which block polyamine synthesis could hypo-
thetically be used to increase the pro-apoptotic effects of anti-
vascular therapy
5 Anti-angiogenic therapy AMPK activation and Warburg
effect
AMP-activated protein kinase (AMPK) is a central metabolic
sensor found in all eukaryote systems that governs glucose and
lipid metabolism in response to alterations in nutrients supply andintracellular energy levels as well as cell polarity cell proliferation
and gene expression regulation [5859] In most species AMPK is
a heterotrimer that consists of a catalytic subunit (a) and two
regulatory subunits (b and g) In mammals there are two genes
encoding the AMPKa catalytic subunit two b genes and three g
subunit genes which differ in their tissue speci1047297city and subcel-
lular localization This serineethreonine kinase is mainly activated
in response to an increase in the AMPATP ratio within the cell and
it is phosphorylated at Thr-172 in the catalytic subunit by upstream
kinases including Liver Kinase B1 (LKB1) or calmodulin-dependent
protein kinase kinase beta (CAMKKb) [6061] In addition AMPK
can also be activated by a variety of pharmacological agents
including metformin which is used in the treatment of metabolic
disorders such as type 2 diabetes and obesity [62] AMPK activationreprograms cellular metabolism and enforces metabolic check-
points by acting on mTOR complex 1 (mTORC1) p53 and other
molecules [63] In particular AMPK acts to restore cellular energy
balance by promoting ATP generating processes such as fatty acid
beta oxidation and simultaneously by inhibiting ATP consuming
processes such as fatty acid synthesis gluconeogenesis and protein
synthesis This is initially achieved by direct phosphorylation of
some key metabolic enzymes (such as Acetil-CoA carboxylase ACC)
and subsequently by modulation of gene expression [64] Decoding
substrates of AMPK that have roles in the various cellular processes
controlled by this kinase is a hot area of investigation in the 1047297eld
With respect to cancer several recent studies in cell culture
models and in vivo have shown that growth of tumor cell lines can
be inhibited by AMPK activation highlighting as this kinase mightbe a cancer relevant ldquodruggablerdquo target In particular combination
of chemotherapy with metformin is more effective than chemo-
therapy to suppress tumor growth and to inhibit metastasis in
xenografts of breast lung and ovarian cancer [6566]
Since AMPK is activated when intracellular levels of ATP decline
and intracellular levels of AMP increase as often happens during
nutrient starvation and hypoxia a certain level of AMPK activation
is commonly seen in solid tumors [67] and we also observed AMPK
activation in the peri-necrotic areas of control xenografts [50]
Moreover we found that anti-angiogenic therapy increased AMPK
activation levels in tumors probably as a consequence of the
dramatic glucose depletion and ATP level exhaustion as demon-
strated by immunohistochemistry analysis of pAMPK and pACC
levels in tumor xenografts treated with anti-VEGF (Fig 1 and [50])
Our results are in agreement with a clinical study that showed that
bevacizumab increased total AMPK and pAMPK levels in renal cell
carcinoma patients [68] Moreover in that study AMPK activation
correlated with longer OS and PFS of treated patients
Tumor cells bearing AMPK de1047297ciency are hypersensitive to
energy stress-inducing agents [6970] It is thus possible that
defects of AMPK activation may limit survival of tumor cells under
glucose starvation andor hypoxia in vitro or anti-angiogenic
therapy in vivo Indeed in preclinical models highly glycolytic
cells that failed to activate AMPK developed large necrotic areas
after short-term anti-VEGF therapy Moreover attenuation of
AMPKa2 in poorly glycolytic cells compromised their survival
under glucose deprivation in vitro and increased necrosis following
anti-angiogenic therapy of tumor xenografts [50]
With regard to cell metabolism AMPK activation may decrease
the glycolytic1047298ux for example by inducing the expression of TIGAR
through p53 phosphorylation and activation [38] In support of this
possibility we observed that AMPKa2 silencing increased glucose
consumption and lactate production in ovarian cancer cells [50] On
the other hand it was previously known that HIF-1a and its target
genes including genes encoding for several glycolytic enzymes are
up-regulated in LKB1- AMPK- and TSC-de1047297cient 1047297broblasts indi-
cating that loss of any of these genes is suf 1047297cient to alter cellmetabolism and to switch over a highly glycolytic phenotype
[7172] We indeed observed a slight increase (2-fold) in HIF-1a
activity following AMPKa2 silencing in tumor cells suggesting that
HIF-1a could in part account for modulation of cell metabolism in
this system
Finally given the mechanistic connections between cell prolif-
eration and glycolysis [27] it is possible that AMPK-mediated
inhibition of cellular growth under conditions where nutrients
are scarce could indirectly contribute to down-modulate glycolysis
in tumor cells
In conclusion the integrityof signaling pathwaysinvolved in the
control of cell metabolism and quiescence - such as AMPK - could
be important to sense changes in the tumor microenvironment
caused by angiogenesis inhibition and to instruct tumor cells toadapt
6 Conclusions
Based on the results reviewed here multiple factors appear to
orchestrate the pathologic responses of tumors to VEGF neutrali-
zation In tumor xenografts the level of ldquoglucose addictionrdquo is
certainly important to determine whether or not tumors will suffer
from shortage of this energy substrate caused by VEGF blockade
(Fig 2A) Whether or not this will be con1047297rmed in patients relies in
part on the feasibility to characterize the glycolytic phenotype of
tumors In patients some imaging techniques - including FDG-PET
and MRS - could be exploited to measure levels of glucose uptake
and lactate production respectively Moreover expression levels of glycolysis-associated transporters such as GLUT-1 or MCT-1 could
be used as surrogate IHC markers in retrospective studies
A second important parameter is the integrity of the LKB1-
AMPK pathway (Fig 2B) Genetic events leading to inactivation of
LKB1 are quite common in certain sporadic tumors such as lung
adenocarcinoma [73] Since these malignancies are also treated
with bevacizumab in combination with chemotherapy [74] there is
an opportunity to investigate whether loss of LKB1 is associated
with increased necrosis following anti-angiogenic therapy More-
over the observation that anti-angiogenic therapy activates AMPK
raises the question whether it might be appropriate to combine
anti-angiogenic therapy with other AMPK-activating drugs The
biguanide metformin which has shown ef 1047297cacy in preclinical
models of breast colon and prostate cancer [75] could reinforce
E Zulato et al Biochimie 94 (2012) 925e931 929
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 27
Some studies tested the hypothesis that an association between
VEGF polymorphisms and response to anti-VEGF therapy could
exist In advanced breast cancer the VEGF-2578AA and VEGF-
1154AA genotypes predicted a favorable Overall Survival (OS) for
patients in the paclitaxel plus bevacizumab arm but did not predict
a better Progression Free Survival (PFS) interval [12] In ovarian
cancer VEGF SNPs did not correlate with PFS [13]
One of the common side-effects of anti-angiogenic drugs is
hypertension [14] Different retrospective studies involving NSCLC
colorectal and renal carcinoma patients reported a signi1047297cant
improve in OS or PFS among patients with bevacizumab-induced
hypertension [15e17] One limitation of these studies however is
the fact that consensus criteria to measure bevacizumab-induced
hypertension are not yet established [11]
Anti-angiogenic drugs are often cytostatic in action and tumor
shrinkage or regression may not be a realistic estimate of ef 1047297cacy
To overcome the lack of correspondence between the Response
Evaluation Criteria in Solid Tumors (RECIST) and survival in patients
treated with anti-VEGF therapy new radiological methods are
emerging as surrogate biomarkers One potential tool for biomarker
development is dynamic contrast-enhanced magnetic resonance
imaging (DCE-MRI) which could provide information about tumor
blood vessel structure and functions [18] The volume transferconstant of contrast agent (ktrans) is a measure of tumor perfusion
and permeability in DCE-MRI In one randomized trial of sorafenib
in renal cell carcinoma high baseline DCE-MRI parameters
including ktrans and V p (blood plasma volume fraction) correlated
with PFS whereas changes of DCE-MRI parameters after the start of
therapy did not predict PFS [19] In recurrent gliobastoma a marked
reduction in ktrans after one dose of cediranib was seen in patients
with increased PFS [20]
The [18F]1047298uorothymidine Positron Emission Tomography
(FLT-PET) is an imaging technique for measuring in vivo cellular
proliferation in malignant tumor and organ tissue and is used to
monitor tumor responses to cytostatic therapies [21] A prospective
study in patients with recurrent malignant gliomas suggests that
FLT-PET can predict responses to bevacizumab as early as 1e
2weeks after treatment [22]
Finally certain metabolism-associated biomarkers could be
helpful in selecting patients to bene1047297t from anti-angiogenic
therapy In the CONFIRM trials LDH-A GLUT-1 and VEGFR1
mRNA levels predicted responses of colorectal cancer patients to
chemotherapy plus vatalanib [23] In the same trials high tissue
LDH5 correlated with poor PFS in the placebo subgroup whereas
it correlated with improved PFS in the vatalanib subgroup [24]
Moreover although vatalanib did not improve either PFS or OS
compared with placebo when authors strati1047297ed patients by
serum LDH level before random assignment they observed that
patients with high serum LDH had longer median PFS when
treated with vatalanib than with placebo In conclusion the
authors proposed that high serum LDH levels may identify tumorsthat are more dependent on abnormal angiogenesis and may
be more susceptible to VEGF inhibition as also suggested by
others [25]
3 Warburg effect and other metabolic alterations in cancer
In the 1920s Otto Warburg af 1047297rmed the role of metabolism in
carcinogenesis by demonstrating that cancer cells - at variance
with normal cells - rely on glycolysis instead of mitochondrial
oxidative phosphorylation (OXPHOS) to produce ATP even under
aerobic conditions [26] Warburg originally hypothesized that the
glycolytic switch in cancer cells was a consequence of defects in
mitochondria which impair aerobic respiration Currently
however it is known that mitochondria are not damaged in most
cancer cells suggesting that aerobic glycolysis essentially repre-
sents an adaptive choice of tumors [27]
Glucose is a critical nutrient for proliferating cells and it is used
as primary substrate to generate ATP as well as to synthesize amino
acids nucleotides and fatty acid and to regulate the redox
potential so as to minimize the effects of reactive oxygen species
(ROS) that damage cellular membranes and proteins [2829]
Enhanced glucose uptake - visualized in the clinic by [18F]1047298uo-
rodeoxyglucose (FDG)-PET - correlates with poor prognosis in
certain tumor types suggesting that enhanced glycolysis confers
a substantial growth advantage [30]
In recent years there has been a number of studies indicating
that aerobic glycolysis is constitutively up-regulated in tumor cells
through genetic or epigenetic changes The 1047297rst documented
mechanistic link between an activated oncogene and altered
glucose metabolism was the transcriptional activation of lactate
dehydrogenase A (LDH-A) by the MYC oncogene [31] LDH-A
contributes a crucial component of the Warburg effect the
conversion of pyruvate e the end-point of glycolysis e to lactate
that is secreted by monocarboxylate carriers (MCTs) eliminating it
from the pool and keeping glycolysis active [32] The secreted
lactate lowers the extracellular pH which may in1047298uence remod-
eling of the matrix and facilitate invasion Furthermore acidosisallows for the selection of motile cells that can eventually break
through the basement membrane and metastasize [29] Indeed
high levels of lactate have been proposed as prognostic factor in
certain malignancies [33] MYC was also found to regulate other
glycolysis-associated genes such as hexokinase II (HK II ) as well as
glucose transporters [34]
The AKT signaling pathway links growth control to glucose
metabolism and several studies correlated its activity with high
glycolytic rates in cancer cells AKT regulates expression of glucose
transporters and HKII enhancing both glucose uptake and its
retention in the cell [35] Moreover AKT can also increase
activity of hypoxia-inducible factor (HIF) thus further enhancing
glycolysis [36]
HIF-1 - which can accumulate due to hypoxia as well as alter-ations of various signaling pathways in tumors - up-regulates
expression of genes involved in aerobic glycolysis including
glucose transporters glycolytic enzymes and LDH-A [37] In addi-
tion HIF-1 deviates pyruvate away from mitochondria by up-
regulating pyruvate dehydrogenase the rate-limiting enzyme for
pyruvate to acetyl-CoA conversion thus alleviating oxidative stress
derived from mitochondrial metabolism [36]
Activated RAS oncogene was initially linked to increase cellular
glucose uptake but recent studies suggest that metabolic effects of
RAS activation could be mediated by MYC and HIF although the
precise mechanism is not yet established [36]
Although the p53 tumor suppressor has been viewed as the
ldquoguardian of the genomerdquo recently it has been implicated in
metabolism control [36] The p53 protein represses transcription of GLUT-1 and GLUT-4 transporters and can in1047298uence the metabolic
balance between glycolysis and OXPHOS through the transcrip-
tional regulation of the 26 efructose bisphosphatase TP53-induce
glycolysis regulator (TIGAR) and synthesis of cytocrome c oxidase
(SCO2) subunit of complex IV of the electron transport chain
[3839]
Besides glucose glutamine could represent an important energy
substrate in cancer cells DeBerardinis et al recently proposed that
in transformed cells glucose accounts mainly for lipid and nucleo-
tide synthesis whereas glutamine is responsible for re-feeding of
the TCA cycle for amino acid synthesis and for nitrogen incorpo-
ration into purine and pyrimidine for nucleotide synthesis [40]
MYC-addicted tumor cells are particularly sensitive to glutamine
withdrawal [41] andgenes involvedin mitocondrial biogenesis and
E Zulato et al Biochimie 94 (2012) 925e931926
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 37
glutamine metabolism specially glutamine transporter and GLS
genes appear to be under both the direct and indirect transcrip-
tional control of MYC [42] The ability of MYC to induceboth aerobic
glycolysis and glutamine oxidation provides cancer cells with the
ability to accumulate biomass [36]
Many human tumor types show altered metabolism of certain
amino acids showing increased uptake and high levels of amino
acid transporter expression
Enhanced expression of L -type amino acid transporter (LAT-1)
which shows high af 1047297nity for several essential amino acids
including leucine tryptophan and methionine (MET) has been
reported in astrocytomas and correlated with poor prognosis [43]
Patients with glioblastoma or other extracranial malignant tumors
such as lung cancer head and neck cancers breast cancer sarcomas
and lymphomas showed increased uptake of MET [44] a feature
which is exploited to make diagnosis and assess therapeutic ef 1047297-
cacy by 18F-MET PET [45]
In addition to increased glycolytic and amino acid activities
recurrent alterations of lipid metabolism are found in cancer cells
conceivably due to their requirement of de novo synthesis of lipids
for membrane assembly Overexpression of fatty acid synthase
(FASN) that catalyzes the de novo synthesis of fatty acids has been
observed in many human cancers including breast prostate lungand colorectal cancers and high levels of FASN were associated
with poor prognosis [46]
Alterations in choline-metabolites (tCho) are also quite
common in cancer cells Tumor cell lines are characterized by an
increased content of phosphocholine (PCho) as compared with
normal epithelial cells [47] The alpha-isoform of Choline Kinase
(ChoK) is often over-expressed in cancer and it is required to
sustain the PCho pool in tumor cells [48] Choline phosphorilation
by ChoK represents the 1047297rst step of choline metabolism in which
choline is 1047297nally converted to phosphatidylcholine a major
constituent of the mammalian cell membrane Choline-
metabolites are of particular interest because they can be moni-
tored in patients by magnetic resonance spectral (MRS) which
detects endogenous PCho or PET which detects altered kinetics of labeled Cho
An interesting area for future studies is to investigate the
predictive and prognostic value of these metabolic features of
cancer cells and to clarify whether they are modulated by anti-
angiogenic therapy
4 Metabolic perturbations after anti-angiogenic therapy
Responses to anti-angiogenic drugs such as sunitinib or bev-
acizumab have been quite heterogeneous in cancer patients In
some cases tumors respond by decreasing tumor volume by more
than 33 qualifying it for a partial response according to RECIST
criteria In other patients however signi1047297cant changes in tumor
density with no decrease in tumor dimensions are observed [49]
This is often associated with central tumor cavitation and necrosis
an observation which suggests that VEGF blockade may perturb the
energy balance in cancer cells
In a recent study [50] we investigated how metabolic param-
eters contribute to determine the pathologic response to VEGF
blockade in tumor xenografts A landmark observation of our study
was that the level of ldquoglucose addictionrdquo of tumor cells dictates the
amount of necrosis caused by angiogenesis inhibition This was
explained by the fact that VEGF blockade acutely perturbs glucose
and ATP levels in tumor xenografts Measurements by biolumi-nescence metabolic imaging indicated that after anti-VEGF therapy
glucose and ATP concentrations in tumors were 130 mmolg and
110 mmolg respectively Values in control tumors were 330 mmol
g (glucose) and 150 mmolg (ATP) Notably glucose uptake
was maintained following anti-angiogenic therapy as shown by
FDG-PET imaging indicating that delivery of glucose through the
vasculature was not compromised despite a substantial decrease in
microvessel density [50] similarly to what has been observed in
patients after bevacizumab monotherapy [51] So it appears that
glucose steady-state levels are very low after anti-angiogenic
therapy whereas glucose uptake is high likely due to HIF-1a
accumulation in treated tumors Intriguingly a preliminary report
showed that a subset of breast cancer patients treated with short-
term 1047297rst-line bevacizumab strongly up-regulated the hypoxiametagene [52] lending support to our observations
Fig 1 Anti-VEGF therapy increases AMPK activation in tumors Representative pictures of pAMPK and pACC staining of ovarian cancer IGROV-1 xenografts following 1 week of
treatment with the anti-VEGF monoclonal antibody A461
E Zulato et al Biochimie 94 (2012) 925e931 927
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 47
A related study investigated metabolic changes in glioblastoma
following anti-VEGF treatment and observed a tendency toward
accumulation of lactate alanine choline myo-inositol creatine
taurine and mobile lipids together with induction of HIF-1a and
activation of the phosphatidyl-inositol-3-kinase pathway [53] This
combination of metabolic changes has previously been associated
with increased hypoxia in human brain tumor spectra [54] and
partially overlaps with our 1047297ndings in ovarian cancer xenografts
[50] In future studies global metabolic changes identi1047297ed by mass
spectrometry analysis (including LC-MSMS and GCeMS) will be
helpful to characterize more extensively metabolic changes
induced by anti-angiogenic therapy in tumors There is already
evidence that this technology enables to pick up speci1047297c oncome-
tabolites in prostate cancer [55] and in gliomas [56]
Fig 2 Parameters contributing to determine tumor responses to VEGF neutralization (A) Anti-angiogenic therapy has been demonstrated to perturb glucose levels in tumors
xenografts Levels of ldquoglucose addictionrdquo of tumor cells in1047298uence the amount of necrosis caused by VEGF blockade Highly glycolytic tumors show a signi1047297cant reduction in their sizeand develop large necrotic areas following short-term anti-angiogenic therapy In contrast poorly glycolytic tumors are only marginally affected in size and do not markedly
increase necrosis after anti-angiogenic therapy [based on [50]] (B) AMPK activation as consequence of perturbations of ATP levels in tumors and its effects on the outcome of VEGF
blockade AMPK activation following short-term anti-angiogenic therapy reduces anabolic processes and cell proliferation being associated with minimal necrosis areas In contrast
tumors that fail to activate this pathway (ie due to LKB1 loss or mutations) maintain a high metabolic demand and are committed to develop large necrotic areas (C) Possible
outlook following tumor necrosis Large necrotic areas may induce recruitment of pro-angiogenic bone marrow-derived cells followed by rapid tumor regrowth Alternatively
killing of the majority of tumor cells by necrosis may turn into a therapeutic effect if relapse mechanisms are not engaged
E Zulato et al Biochimie 94 (2012) 925e931928
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 57
Finally a novel role of the polyamine system in the hypoxic
response of cancer cells has recently been demonstrated The
polyamine system is up-regulated by hypoxia in a variety of cancer
cell lines and in hypoxic tumor regions and inhibition of polyamine
biosynthesis sensitizes cancer cells to hypoxia-induced apoptosis
in vitro [57] These 1047297ndings are relevant in this context if one
considers that the anti-tumor effect of bevacizumab was signi1047297-
cantly enhanced in mice receiving concomitant treatment with the
polyamine biosynthesis inhibitor DFMO which irreversibly inacti-
vates the key enzyme ornithine decarboxylase (ODC) [57] Thus
increased ODC expression and increased intracellular polyamine
levels may occur in tumors treated with anti-angiogenic drugs
likely contributing to protect tumor cells from hypoxia-induced
apoptosis Drugs which block polyamine synthesis could hypo-
thetically be used to increase the pro-apoptotic effects of anti-
vascular therapy
5 Anti-angiogenic therapy AMPK activation and Warburg
effect
AMP-activated protein kinase (AMPK) is a central metabolic
sensor found in all eukaryote systems that governs glucose and
lipid metabolism in response to alterations in nutrients supply andintracellular energy levels as well as cell polarity cell proliferation
and gene expression regulation [5859] In most species AMPK is
a heterotrimer that consists of a catalytic subunit (a) and two
regulatory subunits (b and g) In mammals there are two genes
encoding the AMPKa catalytic subunit two b genes and three g
subunit genes which differ in their tissue speci1047297city and subcel-
lular localization This serineethreonine kinase is mainly activated
in response to an increase in the AMPATP ratio within the cell and
it is phosphorylated at Thr-172 in the catalytic subunit by upstream
kinases including Liver Kinase B1 (LKB1) or calmodulin-dependent
protein kinase kinase beta (CAMKKb) [6061] In addition AMPK
can also be activated by a variety of pharmacological agents
including metformin which is used in the treatment of metabolic
disorders such as type 2 diabetes and obesity [62] AMPK activationreprograms cellular metabolism and enforces metabolic check-
points by acting on mTOR complex 1 (mTORC1) p53 and other
molecules [63] In particular AMPK acts to restore cellular energy
balance by promoting ATP generating processes such as fatty acid
beta oxidation and simultaneously by inhibiting ATP consuming
processes such as fatty acid synthesis gluconeogenesis and protein
synthesis This is initially achieved by direct phosphorylation of
some key metabolic enzymes (such as Acetil-CoA carboxylase ACC)
and subsequently by modulation of gene expression [64] Decoding
substrates of AMPK that have roles in the various cellular processes
controlled by this kinase is a hot area of investigation in the 1047297eld
With respect to cancer several recent studies in cell culture
models and in vivo have shown that growth of tumor cell lines can
be inhibited by AMPK activation highlighting as this kinase mightbe a cancer relevant ldquodruggablerdquo target In particular combination
of chemotherapy with metformin is more effective than chemo-
therapy to suppress tumor growth and to inhibit metastasis in
xenografts of breast lung and ovarian cancer [6566]
Since AMPK is activated when intracellular levels of ATP decline
and intracellular levels of AMP increase as often happens during
nutrient starvation and hypoxia a certain level of AMPK activation
is commonly seen in solid tumors [67] and we also observed AMPK
activation in the peri-necrotic areas of control xenografts [50]
Moreover we found that anti-angiogenic therapy increased AMPK
activation levels in tumors probably as a consequence of the
dramatic glucose depletion and ATP level exhaustion as demon-
strated by immunohistochemistry analysis of pAMPK and pACC
levels in tumor xenografts treated with anti-VEGF (Fig 1 and [50])
Our results are in agreement with a clinical study that showed that
bevacizumab increased total AMPK and pAMPK levels in renal cell
carcinoma patients [68] Moreover in that study AMPK activation
correlated with longer OS and PFS of treated patients
Tumor cells bearing AMPK de1047297ciency are hypersensitive to
energy stress-inducing agents [6970] It is thus possible that
defects of AMPK activation may limit survival of tumor cells under
glucose starvation andor hypoxia in vitro or anti-angiogenic
therapy in vivo Indeed in preclinical models highly glycolytic
cells that failed to activate AMPK developed large necrotic areas
after short-term anti-VEGF therapy Moreover attenuation of
AMPKa2 in poorly glycolytic cells compromised their survival
under glucose deprivation in vitro and increased necrosis following
anti-angiogenic therapy of tumor xenografts [50]
With regard to cell metabolism AMPK activation may decrease
the glycolytic1047298ux for example by inducing the expression of TIGAR
through p53 phosphorylation and activation [38] In support of this
possibility we observed that AMPKa2 silencing increased glucose
consumption and lactate production in ovarian cancer cells [50] On
the other hand it was previously known that HIF-1a and its target
genes including genes encoding for several glycolytic enzymes are
up-regulated in LKB1- AMPK- and TSC-de1047297cient 1047297broblasts indi-
cating that loss of any of these genes is suf 1047297cient to alter cellmetabolism and to switch over a highly glycolytic phenotype
[7172] We indeed observed a slight increase (2-fold) in HIF-1a
activity following AMPKa2 silencing in tumor cells suggesting that
HIF-1a could in part account for modulation of cell metabolism in
this system
Finally given the mechanistic connections between cell prolif-
eration and glycolysis [27] it is possible that AMPK-mediated
inhibition of cellular growth under conditions where nutrients
are scarce could indirectly contribute to down-modulate glycolysis
in tumor cells
In conclusion the integrityof signaling pathwaysinvolved in the
control of cell metabolism and quiescence - such as AMPK - could
be important to sense changes in the tumor microenvironment
caused by angiogenesis inhibition and to instruct tumor cells toadapt
6 Conclusions
Based on the results reviewed here multiple factors appear to
orchestrate the pathologic responses of tumors to VEGF neutrali-
zation In tumor xenografts the level of ldquoglucose addictionrdquo is
certainly important to determine whether or not tumors will suffer
from shortage of this energy substrate caused by VEGF blockade
(Fig 2A) Whether or not this will be con1047297rmed in patients relies in
part on the feasibility to characterize the glycolytic phenotype of
tumors In patients some imaging techniques - including FDG-PET
and MRS - could be exploited to measure levels of glucose uptake
and lactate production respectively Moreover expression levels of glycolysis-associated transporters such as GLUT-1 or MCT-1 could
be used as surrogate IHC markers in retrospective studies
A second important parameter is the integrity of the LKB1-
AMPK pathway (Fig 2B) Genetic events leading to inactivation of
LKB1 are quite common in certain sporadic tumors such as lung
adenocarcinoma [73] Since these malignancies are also treated
with bevacizumab in combination with chemotherapy [74] there is
an opportunity to investigate whether loss of LKB1 is associated
with increased necrosis following anti-angiogenic therapy More-
over the observation that anti-angiogenic therapy activates AMPK
raises the question whether it might be appropriate to combine
anti-angiogenic therapy with other AMPK-activating drugs The
biguanide metformin which has shown ef 1047297cacy in preclinical
models of breast colon and prostate cancer [75] could reinforce
E Zulato et al Biochimie 94 (2012) 925e931 929
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931
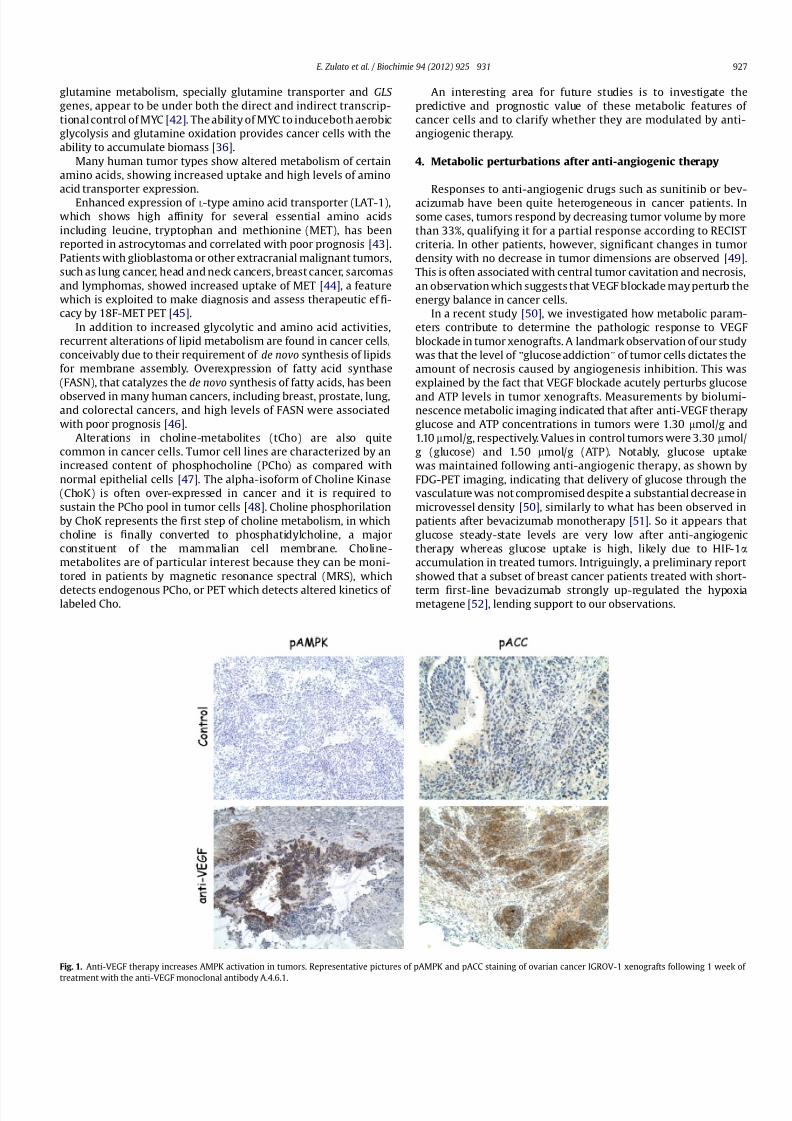
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 37
glutamine metabolism specially glutamine transporter and GLS
genes appear to be under both the direct and indirect transcrip-
tional control of MYC [42] The ability of MYC to induceboth aerobic
glycolysis and glutamine oxidation provides cancer cells with the
ability to accumulate biomass [36]
Many human tumor types show altered metabolism of certain
amino acids showing increased uptake and high levels of amino
acid transporter expression
Enhanced expression of L -type amino acid transporter (LAT-1)
which shows high af 1047297nity for several essential amino acids
including leucine tryptophan and methionine (MET) has been
reported in astrocytomas and correlated with poor prognosis [43]
Patients with glioblastoma or other extracranial malignant tumors
such as lung cancer head and neck cancers breast cancer sarcomas
and lymphomas showed increased uptake of MET [44] a feature
which is exploited to make diagnosis and assess therapeutic ef 1047297-
cacy by 18F-MET PET [45]
In addition to increased glycolytic and amino acid activities
recurrent alterations of lipid metabolism are found in cancer cells
conceivably due to their requirement of de novo synthesis of lipids
for membrane assembly Overexpression of fatty acid synthase
(FASN) that catalyzes the de novo synthesis of fatty acids has been
observed in many human cancers including breast prostate lungand colorectal cancers and high levels of FASN were associated
with poor prognosis [46]
Alterations in choline-metabolites (tCho) are also quite
common in cancer cells Tumor cell lines are characterized by an
increased content of phosphocholine (PCho) as compared with
normal epithelial cells [47] The alpha-isoform of Choline Kinase
(ChoK) is often over-expressed in cancer and it is required to
sustain the PCho pool in tumor cells [48] Choline phosphorilation
by ChoK represents the 1047297rst step of choline metabolism in which
choline is 1047297nally converted to phosphatidylcholine a major
constituent of the mammalian cell membrane Choline-
metabolites are of particular interest because they can be moni-
tored in patients by magnetic resonance spectral (MRS) which
detects endogenous PCho or PET which detects altered kinetics of labeled Cho
An interesting area for future studies is to investigate the
predictive and prognostic value of these metabolic features of
cancer cells and to clarify whether they are modulated by anti-
angiogenic therapy
4 Metabolic perturbations after anti-angiogenic therapy
Responses to anti-angiogenic drugs such as sunitinib or bev-
acizumab have been quite heterogeneous in cancer patients In
some cases tumors respond by decreasing tumor volume by more
than 33 qualifying it for a partial response according to RECIST
criteria In other patients however signi1047297cant changes in tumor
density with no decrease in tumor dimensions are observed [49]
This is often associated with central tumor cavitation and necrosis
an observation which suggests that VEGF blockade may perturb the
energy balance in cancer cells
In a recent study [50] we investigated how metabolic param-
eters contribute to determine the pathologic response to VEGF
blockade in tumor xenografts A landmark observation of our study
was that the level of ldquoglucose addictionrdquo of tumor cells dictates the
amount of necrosis caused by angiogenesis inhibition This was
explained by the fact that VEGF blockade acutely perturbs glucose
and ATP levels in tumor xenografts Measurements by biolumi-nescence metabolic imaging indicated that after anti-VEGF therapy
glucose and ATP concentrations in tumors were 130 mmolg and
110 mmolg respectively Values in control tumors were 330 mmol
g (glucose) and 150 mmolg (ATP) Notably glucose uptake
was maintained following anti-angiogenic therapy as shown by
FDG-PET imaging indicating that delivery of glucose through the
vasculature was not compromised despite a substantial decrease in
microvessel density [50] similarly to what has been observed in
patients after bevacizumab monotherapy [51] So it appears that
glucose steady-state levels are very low after anti-angiogenic
therapy whereas glucose uptake is high likely due to HIF-1a
accumulation in treated tumors Intriguingly a preliminary report
showed that a subset of breast cancer patients treated with short-
term 1047297rst-line bevacizumab strongly up-regulated the hypoxiametagene [52] lending support to our observations
Fig 1 Anti-VEGF therapy increases AMPK activation in tumors Representative pictures of pAMPK and pACC staining of ovarian cancer IGROV-1 xenografts following 1 week of
treatment with the anti-VEGF monoclonal antibody A461
E Zulato et al Biochimie 94 (2012) 925e931 927
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 47
A related study investigated metabolic changes in glioblastoma
following anti-VEGF treatment and observed a tendency toward
accumulation of lactate alanine choline myo-inositol creatine
taurine and mobile lipids together with induction of HIF-1a and
activation of the phosphatidyl-inositol-3-kinase pathway [53] This
combination of metabolic changes has previously been associated
with increased hypoxia in human brain tumor spectra [54] and
partially overlaps with our 1047297ndings in ovarian cancer xenografts
[50] In future studies global metabolic changes identi1047297ed by mass
spectrometry analysis (including LC-MSMS and GCeMS) will be
helpful to characterize more extensively metabolic changes
induced by anti-angiogenic therapy in tumors There is already
evidence that this technology enables to pick up speci1047297c oncome-
tabolites in prostate cancer [55] and in gliomas [56]
Fig 2 Parameters contributing to determine tumor responses to VEGF neutralization (A) Anti-angiogenic therapy has been demonstrated to perturb glucose levels in tumors
xenografts Levels of ldquoglucose addictionrdquo of tumor cells in1047298uence the amount of necrosis caused by VEGF blockade Highly glycolytic tumors show a signi1047297cant reduction in their sizeand develop large necrotic areas following short-term anti-angiogenic therapy In contrast poorly glycolytic tumors are only marginally affected in size and do not markedly
increase necrosis after anti-angiogenic therapy [based on [50]] (B) AMPK activation as consequence of perturbations of ATP levels in tumors and its effects on the outcome of VEGF
blockade AMPK activation following short-term anti-angiogenic therapy reduces anabolic processes and cell proliferation being associated with minimal necrosis areas In contrast
tumors that fail to activate this pathway (ie due to LKB1 loss or mutations) maintain a high metabolic demand and are committed to develop large necrotic areas (C) Possible
outlook following tumor necrosis Large necrotic areas may induce recruitment of pro-angiogenic bone marrow-derived cells followed by rapid tumor regrowth Alternatively
killing of the majority of tumor cells by necrosis may turn into a therapeutic effect if relapse mechanisms are not engaged
E Zulato et al Biochimie 94 (2012) 925e931928
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 57
Finally a novel role of the polyamine system in the hypoxic
response of cancer cells has recently been demonstrated The
polyamine system is up-regulated by hypoxia in a variety of cancer
cell lines and in hypoxic tumor regions and inhibition of polyamine
biosynthesis sensitizes cancer cells to hypoxia-induced apoptosis
in vitro [57] These 1047297ndings are relevant in this context if one
considers that the anti-tumor effect of bevacizumab was signi1047297-
cantly enhanced in mice receiving concomitant treatment with the
polyamine biosynthesis inhibitor DFMO which irreversibly inacti-
vates the key enzyme ornithine decarboxylase (ODC) [57] Thus
increased ODC expression and increased intracellular polyamine
levels may occur in tumors treated with anti-angiogenic drugs
likely contributing to protect tumor cells from hypoxia-induced
apoptosis Drugs which block polyamine synthesis could hypo-
thetically be used to increase the pro-apoptotic effects of anti-
vascular therapy
5 Anti-angiogenic therapy AMPK activation and Warburg
effect
AMP-activated protein kinase (AMPK) is a central metabolic
sensor found in all eukaryote systems that governs glucose and
lipid metabolism in response to alterations in nutrients supply andintracellular energy levels as well as cell polarity cell proliferation
and gene expression regulation [5859] In most species AMPK is
a heterotrimer that consists of a catalytic subunit (a) and two
regulatory subunits (b and g) In mammals there are two genes
encoding the AMPKa catalytic subunit two b genes and three g
subunit genes which differ in their tissue speci1047297city and subcel-
lular localization This serineethreonine kinase is mainly activated
in response to an increase in the AMPATP ratio within the cell and
it is phosphorylated at Thr-172 in the catalytic subunit by upstream
kinases including Liver Kinase B1 (LKB1) or calmodulin-dependent
protein kinase kinase beta (CAMKKb) [6061] In addition AMPK
can also be activated by a variety of pharmacological agents
including metformin which is used in the treatment of metabolic
disorders such as type 2 diabetes and obesity [62] AMPK activationreprograms cellular metabolism and enforces metabolic check-
points by acting on mTOR complex 1 (mTORC1) p53 and other
molecules [63] In particular AMPK acts to restore cellular energy
balance by promoting ATP generating processes such as fatty acid
beta oxidation and simultaneously by inhibiting ATP consuming
processes such as fatty acid synthesis gluconeogenesis and protein
synthesis This is initially achieved by direct phosphorylation of
some key metabolic enzymes (such as Acetil-CoA carboxylase ACC)
and subsequently by modulation of gene expression [64] Decoding
substrates of AMPK that have roles in the various cellular processes
controlled by this kinase is a hot area of investigation in the 1047297eld
With respect to cancer several recent studies in cell culture
models and in vivo have shown that growth of tumor cell lines can
be inhibited by AMPK activation highlighting as this kinase mightbe a cancer relevant ldquodruggablerdquo target In particular combination
of chemotherapy with metformin is more effective than chemo-
therapy to suppress tumor growth and to inhibit metastasis in
xenografts of breast lung and ovarian cancer [6566]
Since AMPK is activated when intracellular levels of ATP decline
and intracellular levels of AMP increase as often happens during
nutrient starvation and hypoxia a certain level of AMPK activation
is commonly seen in solid tumors [67] and we also observed AMPK
activation in the peri-necrotic areas of control xenografts [50]
Moreover we found that anti-angiogenic therapy increased AMPK
activation levels in tumors probably as a consequence of the
dramatic glucose depletion and ATP level exhaustion as demon-
strated by immunohistochemistry analysis of pAMPK and pACC
levels in tumor xenografts treated with anti-VEGF (Fig 1 and [50])
Our results are in agreement with a clinical study that showed that
bevacizumab increased total AMPK and pAMPK levels in renal cell
carcinoma patients [68] Moreover in that study AMPK activation
correlated with longer OS and PFS of treated patients
Tumor cells bearing AMPK de1047297ciency are hypersensitive to
energy stress-inducing agents [6970] It is thus possible that
defects of AMPK activation may limit survival of tumor cells under
glucose starvation andor hypoxia in vitro or anti-angiogenic
therapy in vivo Indeed in preclinical models highly glycolytic
cells that failed to activate AMPK developed large necrotic areas
after short-term anti-VEGF therapy Moreover attenuation of
AMPKa2 in poorly glycolytic cells compromised their survival
under glucose deprivation in vitro and increased necrosis following
anti-angiogenic therapy of tumor xenografts [50]
With regard to cell metabolism AMPK activation may decrease
the glycolytic1047298ux for example by inducing the expression of TIGAR
through p53 phosphorylation and activation [38] In support of this
possibility we observed that AMPKa2 silencing increased glucose
consumption and lactate production in ovarian cancer cells [50] On
the other hand it was previously known that HIF-1a and its target
genes including genes encoding for several glycolytic enzymes are
up-regulated in LKB1- AMPK- and TSC-de1047297cient 1047297broblasts indi-
cating that loss of any of these genes is suf 1047297cient to alter cellmetabolism and to switch over a highly glycolytic phenotype
[7172] We indeed observed a slight increase (2-fold) in HIF-1a
activity following AMPKa2 silencing in tumor cells suggesting that
HIF-1a could in part account for modulation of cell metabolism in
this system
Finally given the mechanistic connections between cell prolif-
eration and glycolysis [27] it is possible that AMPK-mediated
inhibition of cellular growth under conditions where nutrients
are scarce could indirectly contribute to down-modulate glycolysis
in tumor cells
In conclusion the integrityof signaling pathwaysinvolved in the
control of cell metabolism and quiescence - such as AMPK - could
be important to sense changes in the tumor microenvironment
caused by angiogenesis inhibition and to instruct tumor cells toadapt
6 Conclusions
Based on the results reviewed here multiple factors appear to
orchestrate the pathologic responses of tumors to VEGF neutrali-
zation In tumor xenografts the level of ldquoglucose addictionrdquo is
certainly important to determine whether or not tumors will suffer
from shortage of this energy substrate caused by VEGF blockade
(Fig 2A) Whether or not this will be con1047297rmed in patients relies in
part on the feasibility to characterize the glycolytic phenotype of
tumors In patients some imaging techniques - including FDG-PET
and MRS - could be exploited to measure levels of glucose uptake
and lactate production respectively Moreover expression levels of glycolysis-associated transporters such as GLUT-1 or MCT-1 could
be used as surrogate IHC markers in retrospective studies
A second important parameter is the integrity of the LKB1-
AMPK pathway (Fig 2B) Genetic events leading to inactivation of
LKB1 are quite common in certain sporadic tumors such as lung
adenocarcinoma [73] Since these malignancies are also treated
with bevacizumab in combination with chemotherapy [74] there is
an opportunity to investigate whether loss of LKB1 is associated
with increased necrosis following anti-angiogenic therapy More-
over the observation that anti-angiogenic therapy activates AMPK
raises the question whether it might be appropriate to combine
anti-angiogenic therapy with other AMPK-activating drugs The
biguanide metformin which has shown ef 1047297cacy in preclinical
models of breast colon and prostate cancer [75] could reinforce
E Zulato et al Biochimie 94 (2012) 925e931 929
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 47
A related study investigated metabolic changes in glioblastoma
following anti-VEGF treatment and observed a tendency toward
accumulation of lactate alanine choline myo-inositol creatine
taurine and mobile lipids together with induction of HIF-1a and
activation of the phosphatidyl-inositol-3-kinase pathway [53] This
combination of metabolic changes has previously been associated
with increased hypoxia in human brain tumor spectra [54] and
partially overlaps with our 1047297ndings in ovarian cancer xenografts
[50] In future studies global metabolic changes identi1047297ed by mass
spectrometry analysis (including LC-MSMS and GCeMS) will be
helpful to characterize more extensively metabolic changes
induced by anti-angiogenic therapy in tumors There is already
evidence that this technology enables to pick up speci1047297c oncome-
tabolites in prostate cancer [55] and in gliomas [56]
Fig 2 Parameters contributing to determine tumor responses to VEGF neutralization (A) Anti-angiogenic therapy has been demonstrated to perturb glucose levels in tumors
xenografts Levels of ldquoglucose addictionrdquo of tumor cells in1047298uence the amount of necrosis caused by VEGF blockade Highly glycolytic tumors show a signi1047297cant reduction in their sizeand develop large necrotic areas following short-term anti-angiogenic therapy In contrast poorly glycolytic tumors are only marginally affected in size and do not markedly
increase necrosis after anti-angiogenic therapy [based on [50]] (B) AMPK activation as consequence of perturbations of ATP levels in tumors and its effects on the outcome of VEGF
blockade AMPK activation following short-term anti-angiogenic therapy reduces anabolic processes and cell proliferation being associated with minimal necrosis areas In contrast
tumors that fail to activate this pathway (ie due to LKB1 loss or mutations) maintain a high metabolic demand and are committed to develop large necrotic areas (C) Possible
outlook following tumor necrosis Large necrotic areas may induce recruitment of pro-angiogenic bone marrow-derived cells followed by rapid tumor regrowth Alternatively
killing of the majority of tumor cells by necrosis may turn into a therapeutic effect if relapse mechanisms are not engaged
E Zulato et al Biochimie 94 (2012) 925e931928
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 57
Finally a novel role of the polyamine system in the hypoxic
response of cancer cells has recently been demonstrated The
polyamine system is up-regulated by hypoxia in a variety of cancer
cell lines and in hypoxic tumor regions and inhibition of polyamine
biosynthesis sensitizes cancer cells to hypoxia-induced apoptosis
in vitro [57] These 1047297ndings are relevant in this context if one
considers that the anti-tumor effect of bevacizumab was signi1047297-
cantly enhanced in mice receiving concomitant treatment with the
polyamine biosynthesis inhibitor DFMO which irreversibly inacti-
vates the key enzyme ornithine decarboxylase (ODC) [57] Thus
increased ODC expression and increased intracellular polyamine
levels may occur in tumors treated with anti-angiogenic drugs
likely contributing to protect tumor cells from hypoxia-induced
apoptosis Drugs which block polyamine synthesis could hypo-
thetically be used to increase the pro-apoptotic effects of anti-
vascular therapy
5 Anti-angiogenic therapy AMPK activation and Warburg
effect
AMP-activated protein kinase (AMPK) is a central metabolic
sensor found in all eukaryote systems that governs glucose and
lipid metabolism in response to alterations in nutrients supply andintracellular energy levels as well as cell polarity cell proliferation
and gene expression regulation [5859] In most species AMPK is
a heterotrimer that consists of a catalytic subunit (a) and two
regulatory subunits (b and g) In mammals there are two genes
encoding the AMPKa catalytic subunit two b genes and three g
subunit genes which differ in their tissue speci1047297city and subcel-
lular localization This serineethreonine kinase is mainly activated
in response to an increase in the AMPATP ratio within the cell and
it is phosphorylated at Thr-172 in the catalytic subunit by upstream
kinases including Liver Kinase B1 (LKB1) or calmodulin-dependent
protein kinase kinase beta (CAMKKb) [6061] In addition AMPK
can also be activated by a variety of pharmacological agents
including metformin which is used in the treatment of metabolic
disorders such as type 2 diabetes and obesity [62] AMPK activationreprograms cellular metabolism and enforces metabolic check-
points by acting on mTOR complex 1 (mTORC1) p53 and other
molecules [63] In particular AMPK acts to restore cellular energy
balance by promoting ATP generating processes such as fatty acid
beta oxidation and simultaneously by inhibiting ATP consuming
processes such as fatty acid synthesis gluconeogenesis and protein
synthesis This is initially achieved by direct phosphorylation of
some key metabolic enzymes (such as Acetil-CoA carboxylase ACC)
and subsequently by modulation of gene expression [64] Decoding
substrates of AMPK that have roles in the various cellular processes
controlled by this kinase is a hot area of investigation in the 1047297eld
With respect to cancer several recent studies in cell culture
models and in vivo have shown that growth of tumor cell lines can
be inhibited by AMPK activation highlighting as this kinase mightbe a cancer relevant ldquodruggablerdquo target In particular combination
of chemotherapy with metformin is more effective than chemo-
therapy to suppress tumor growth and to inhibit metastasis in
xenografts of breast lung and ovarian cancer [6566]
Since AMPK is activated when intracellular levels of ATP decline
and intracellular levels of AMP increase as often happens during
nutrient starvation and hypoxia a certain level of AMPK activation
is commonly seen in solid tumors [67] and we also observed AMPK
activation in the peri-necrotic areas of control xenografts [50]
Moreover we found that anti-angiogenic therapy increased AMPK
activation levels in tumors probably as a consequence of the
dramatic glucose depletion and ATP level exhaustion as demon-
strated by immunohistochemistry analysis of pAMPK and pACC
levels in tumor xenografts treated with anti-VEGF (Fig 1 and [50])
Our results are in agreement with a clinical study that showed that
bevacizumab increased total AMPK and pAMPK levels in renal cell
carcinoma patients [68] Moreover in that study AMPK activation
correlated with longer OS and PFS of treated patients
Tumor cells bearing AMPK de1047297ciency are hypersensitive to
energy stress-inducing agents [6970] It is thus possible that
defects of AMPK activation may limit survival of tumor cells under
glucose starvation andor hypoxia in vitro or anti-angiogenic
therapy in vivo Indeed in preclinical models highly glycolytic
cells that failed to activate AMPK developed large necrotic areas
after short-term anti-VEGF therapy Moreover attenuation of
AMPKa2 in poorly glycolytic cells compromised their survival
under glucose deprivation in vitro and increased necrosis following
anti-angiogenic therapy of tumor xenografts [50]
With regard to cell metabolism AMPK activation may decrease
the glycolytic1047298ux for example by inducing the expression of TIGAR
through p53 phosphorylation and activation [38] In support of this
possibility we observed that AMPKa2 silencing increased glucose
consumption and lactate production in ovarian cancer cells [50] On
the other hand it was previously known that HIF-1a and its target
genes including genes encoding for several glycolytic enzymes are
up-regulated in LKB1- AMPK- and TSC-de1047297cient 1047297broblasts indi-
cating that loss of any of these genes is suf 1047297cient to alter cellmetabolism and to switch over a highly glycolytic phenotype
[7172] We indeed observed a slight increase (2-fold) in HIF-1a
activity following AMPKa2 silencing in tumor cells suggesting that
HIF-1a could in part account for modulation of cell metabolism in
this system
Finally given the mechanistic connections between cell prolif-
eration and glycolysis [27] it is possible that AMPK-mediated
inhibition of cellular growth under conditions where nutrients
are scarce could indirectly contribute to down-modulate glycolysis
in tumor cells
In conclusion the integrityof signaling pathwaysinvolved in the
control of cell metabolism and quiescence - such as AMPK - could
be important to sense changes in the tumor microenvironment
caused by angiogenesis inhibition and to instruct tumor cells toadapt
6 Conclusions
Based on the results reviewed here multiple factors appear to
orchestrate the pathologic responses of tumors to VEGF neutrali-
zation In tumor xenografts the level of ldquoglucose addictionrdquo is
certainly important to determine whether or not tumors will suffer
from shortage of this energy substrate caused by VEGF blockade
(Fig 2A) Whether or not this will be con1047297rmed in patients relies in
part on the feasibility to characterize the glycolytic phenotype of
tumors In patients some imaging techniques - including FDG-PET
and MRS - could be exploited to measure levels of glucose uptake
and lactate production respectively Moreover expression levels of glycolysis-associated transporters such as GLUT-1 or MCT-1 could
be used as surrogate IHC markers in retrospective studies
A second important parameter is the integrity of the LKB1-
AMPK pathway (Fig 2B) Genetic events leading to inactivation of
LKB1 are quite common in certain sporadic tumors such as lung
adenocarcinoma [73] Since these malignancies are also treated
with bevacizumab in combination with chemotherapy [74] there is
an opportunity to investigate whether loss of LKB1 is associated
with increased necrosis following anti-angiogenic therapy More-
over the observation that anti-angiogenic therapy activates AMPK
raises the question whether it might be appropriate to combine
anti-angiogenic therapy with other AMPK-activating drugs The
biguanide metformin which has shown ef 1047297cacy in preclinical
models of breast colon and prostate cancer [75] could reinforce
E Zulato et al Biochimie 94 (2012) 925e931 929
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 57
Finally a novel role of the polyamine system in the hypoxic
response of cancer cells has recently been demonstrated The
polyamine system is up-regulated by hypoxia in a variety of cancer
cell lines and in hypoxic tumor regions and inhibition of polyamine
biosynthesis sensitizes cancer cells to hypoxia-induced apoptosis
in vitro [57] These 1047297ndings are relevant in this context if one
considers that the anti-tumor effect of bevacizumab was signi1047297-
cantly enhanced in mice receiving concomitant treatment with the
polyamine biosynthesis inhibitor DFMO which irreversibly inacti-
vates the key enzyme ornithine decarboxylase (ODC) [57] Thus
increased ODC expression and increased intracellular polyamine
levels may occur in tumors treated with anti-angiogenic drugs
likely contributing to protect tumor cells from hypoxia-induced
apoptosis Drugs which block polyamine synthesis could hypo-
thetically be used to increase the pro-apoptotic effects of anti-
vascular therapy
5 Anti-angiogenic therapy AMPK activation and Warburg
effect
AMP-activated protein kinase (AMPK) is a central metabolic
sensor found in all eukaryote systems that governs glucose and
lipid metabolism in response to alterations in nutrients supply andintracellular energy levels as well as cell polarity cell proliferation
and gene expression regulation [5859] In most species AMPK is
a heterotrimer that consists of a catalytic subunit (a) and two
regulatory subunits (b and g) In mammals there are two genes
encoding the AMPKa catalytic subunit two b genes and three g
subunit genes which differ in their tissue speci1047297city and subcel-
lular localization This serineethreonine kinase is mainly activated
in response to an increase in the AMPATP ratio within the cell and
it is phosphorylated at Thr-172 in the catalytic subunit by upstream
kinases including Liver Kinase B1 (LKB1) or calmodulin-dependent
protein kinase kinase beta (CAMKKb) [6061] In addition AMPK
can also be activated by a variety of pharmacological agents
including metformin which is used in the treatment of metabolic
disorders such as type 2 diabetes and obesity [62] AMPK activationreprograms cellular metabolism and enforces metabolic check-
points by acting on mTOR complex 1 (mTORC1) p53 and other
molecules [63] In particular AMPK acts to restore cellular energy
balance by promoting ATP generating processes such as fatty acid
beta oxidation and simultaneously by inhibiting ATP consuming
processes such as fatty acid synthesis gluconeogenesis and protein
synthesis This is initially achieved by direct phosphorylation of
some key metabolic enzymes (such as Acetil-CoA carboxylase ACC)
and subsequently by modulation of gene expression [64] Decoding
substrates of AMPK that have roles in the various cellular processes
controlled by this kinase is a hot area of investigation in the 1047297eld
With respect to cancer several recent studies in cell culture
models and in vivo have shown that growth of tumor cell lines can
be inhibited by AMPK activation highlighting as this kinase mightbe a cancer relevant ldquodruggablerdquo target In particular combination
of chemotherapy with metformin is more effective than chemo-
therapy to suppress tumor growth and to inhibit metastasis in
xenografts of breast lung and ovarian cancer [6566]
Since AMPK is activated when intracellular levels of ATP decline
and intracellular levels of AMP increase as often happens during
nutrient starvation and hypoxia a certain level of AMPK activation
is commonly seen in solid tumors [67] and we also observed AMPK
activation in the peri-necrotic areas of control xenografts [50]
Moreover we found that anti-angiogenic therapy increased AMPK
activation levels in tumors probably as a consequence of the
dramatic glucose depletion and ATP level exhaustion as demon-
strated by immunohistochemistry analysis of pAMPK and pACC
levels in tumor xenografts treated with anti-VEGF (Fig 1 and [50])
Our results are in agreement with a clinical study that showed that
bevacizumab increased total AMPK and pAMPK levels in renal cell
carcinoma patients [68] Moreover in that study AMPK activation
correlated with longer OS and PFS of treated patients
Tumor cells bearing AMPK de1047297ciency are hypersensitive to
energy stress-inducing agents [6970] It is thus possible that
defects of AMPK activation may limit survival of tumor cells under
glucose starvation andor hypoxia in vitro or anti-angiogenic
therapy in vivo Indeed in preclinical models highly glycolytic
cells that failed to activate AMPK developed large necrotic areas
after short-term anti-VEGF therapy Moreover attenuation of
AMPKa2 in poorly glycolytic cells compromised their survival
under glucose deprivation in vitro and increased necrosis following
anti-angiogenic therapy of tumor xenografts [50]
With regard to cell metabolism AMPK activation may decrease
the glycolytic1047298ux for example by inducing the expression of TIGAR
through p53 phosphorylation and activation [38] In support of this
possibility we observed that AMPKa2 silencing increased glucose
consumption and lactate production in ovarian cancer cells [50] On
the other hand it was previously known that HIF-1a and its target
genes including genes encoding for several glycolytic enzymes are
up-regulated in LKB1- AMPK- and TSC-de1047297cient 1047297broblasts indi-
cating that loss of any of these genes is suf 1047297cient to alter cellmetabolism and to switch over a highly glycolytic phenotype
[7172] We indeed observed a slight increase (2-fold) in HIF-1a
activity following AMPKa2 silencing in tumor cells suggesting that
HIF-1a could in part account for modulation of cell metabolism in
this system
Finally given the mechanistic connections between cell prolif-
eration and glycolysis [27] it is possible that AMPK-mediated
inhibition of cellular growth under conditions where nutrients
are scarce could indirectly contribute to down-modulate glycolysis
in tumor cells
In conclusion the integrityof signaling pathwaysinvolved in the
control of cell metabolism and quiescence - such as AMPK - could
be important to sense changes in the tumor microenvironment
caused by angiogenesis inhibition and to instruct tumor cells toadapt
6 Conclusions
Based on the results reviewed here multiple factors appear to
orchestrate the pathologic responses of tumors to VEGF neutrali-
zation In tumor xenografts the level of ldquoglucose addictionrdquo is
certainly important to determine whether or not tumors will suffer
from shortage of this energy substrate caused by VEGF blockade
(Fig 2A) Whether or not this will be con1047297rmed in patients relies in
part on the feasibility to characterize the glycolytic phenotype of
tumors In patients some imaging techniques - including FDG-PET
and MRS - could be exploited to measure levels of glucose uptake
and lactate production respectively Moreover expression levels of glycolysis-associated transporters such as GLUT-1 or MCT-1 could
be used as surrogate IHC markers in retrospective studies
A second important parameter is the integrity of the LKB1-
AMPK pathway (Fig 2B) Genetic events leading to inactivation of
LKB1 are quite common in certain sporadic tumors such as lung
adenocarcinoma [73] Since these malignancies are also treated
with bevacizumab in combination with chemotherapy [74] there is
an opportunity to investigate whether loss of LKB1 is associated
with increased necrosis following anti-angiogenic therapy More-
over the observation that anti-angiogenic therapy activates AMPK
raises the question whether it might be appropriate to combine
anti-angiogenic therapy with other AMPK-activating drugs The
biguanide metformin which has shown ef 1047297cacy in preclinical
models of breast colon and prostate cancer [75] could reinforce
E Zulato et al Biochimie 94 (2012) 925e931 929
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 67
AMPK activation caused by bevacizumab perhaps leading to more
ef 1047297cient control of tumor growth compared with either drug alone
Alternatively since metformin acts as mild inhibitor of complex I of
the respiratory chain [64] forcing the cells to exploit glycolysis to
produce ATP it could transiently make tumors ldquoglucose addictedrdquo
thus increasing treatment-induced tumor necrosis following anti-
angiogenic therapy Future experimental work is needed to clarify
these issues and design appropriate drug combinations
Finally a third critical parameter intimately linked to the energy
balance in tumor is necrosis It is well established treatment with
vascular disrupting agents (VDA) causes massive tumor necrosis
and this has been associated with rapid regrowth promoted by
recruitment of myeloid cells and increased expression of pro-
angiogenic factors (Fig 2C) [7677] On the other hand ablation of
the large majority of tumor cells by necrosis could also be consid-
ered an evidence of therapeutic response and it may not neces-
sarily be followed by tumor relapse (Fig 2C) Necrosis - sometimes
with central cavitation - is also observed in a subset of patients
treated with sunitinib [49] or bevacizumab [52] Therefore an
important goal of future studies is to clarify the prognostic value of
treatment-induced tumor necrosis by utilizing new protocols
which have been optimized to estimate necrosis in CT scans [78]
In conclusion identi1047297cation of clinically feasible methods todetermine the glycolytic phenotype of tumors interrogate the
status of the LKB1AMPK pathway and accurately measure the
extent of necrosis remain a priority in order to test the predictive
value of these upcoming markers in patients treated with anti-
angiogenic drugs Moreover it is certainly possible that a broader
view of the metabolic changes induced by anti-angiogenic drugs in
tumors by using mass spectrometry or other emerging technolo-
gies will uncover additional molecular sensors engaged by meta-
bolic stress and offer an opportunity to understand how
metabolism-based approaches could improve cancer therapy
Acknowledgment
This work was supported in part by grants from Progetto
Oncologico di Medicina Molecolare i tumori femminili Universitagrave
di Padova - Progetto drsquoAteneo 2010 EZ and GN are recipient of
AIRC fellowships
References
[1] VL Heath R Bicknell Anticancer strategies involving the vasculature NatRev Clin Oncol 6 (2009) 395e404
[2] BM Fenton SF Paoni I Ding Effect of VEGF receptor-2 antibody on vascularfunction and oxygenation in spontaneous and transplanted tumors RadiotherOncol 72 (2004) 221e230
[3] M Franco S Man L Chen U Emmenegger Y Shaked AM CheungAS Brown DJ Hicklin FS Foster RS Kerbel Targeted anti-vascular endo-thelial growth factor receptor-2 therapy leads to short-term and long-term
impairment of vascular function and increase in tumor hypoxia Cancer Res66 (2006) 3639e3648
[4] F Winkler SV Kozin RT Tong SS Chae MF Booth I Garkavtsev L XuDJ Hicklin D Fukumura E di Tomaso LL Munn RK Jain Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response toradiation role of oxygenation angiopoietin-1 and matrix metalloproteinasesCancer Cell 6 (2004) 553e563
[5] A Grothey E Galanis Targeting angiogenesis progress with anti-VEGFtreatment with large molecules Nat Rev Clin Oncol 6 (2009) 507e518
[6] RK Jain DG Duda JW Clark JS Loef 1047298er Lessons from phase III clinical trialson anti-VEGF therapy for cancer Nat Clin Pract Oncol 3 (2006) 24e40
[7] RK Jain DG Duda CG Willett DV Sahani AX Zhu JS Loef 1047298erTT Batchelor AG Sorensen Biomarkers of response and resistance to anti-angiogenic therapy Nat Rev Clin Oncol 6 (2009) 327e338
[8] HJ Burstein YH Chen LM Parker J Savoie J Younger I Kuter PD Ryan JE Garber H Chen SM Campos LN Shulman LN Harris R GelmanEP Winer VEGF as a marker for outcome among advanced breast cancerpatients receiving anti-VEGF therapy with bevacizumab and vinorelbine
chemotherapy Clin Cancer Res 14 (2008) 7871e
7877
[9] E Carrillo de Santa Pau FC Arias E Caso Pelaez GM Munoz MolinaI Sanchez Hernandez I Muguruza Trueba R Moreno Balsalobre S SacristanLopez A Gomez Pinillos M del Val Toledo Lobo Prognostic signi1047297cance of theexpression of vascular endothelial growth factors A B C and D and theirreceptors R1 R2 and R3 in patients with nonsmall cell lung cancer Cancer115 (2009) 1701e1712
[10] A Dowlati R Gray AB Sandler JH Schiller DH Johnson Cell adhesionmolecules vascular endothelial growth factor and basic 1047297broblast growthfactor in patients with non-small cell lung cancer treated with chemotherapywith or without bevacizumabean Eastern Cooperative Oncology Group Study
Clin Cancer Res 14 (2008) 1407e
1412[11] AM Jubb AL Harris Biomarkers to predict the clinical ef 1047297cacy of bev-
acizumab in cancer Lancet Oncol 11 (2010) 1172e1183[12] BP Schneider M Wang M Radovich GW Sledge S Badve A Thor
DA Flockhart B Hancock N Davidson J Gralow M Dickler EA PerezM Cobleigh T Shenkier S Edgerton KD Miller Association of vascularendothelial growth factor and vascular endothelial growth factor receptor-2genetic polymorphisms with outcome in a trial of paclitaxel compared withpaclitaxel plus bevacizumab in advanced breast cancer ECOG 2100 J ClinOncol 26 (2008) 4672e4678
[13] KD Steffensen M Waldstrom I Brandslund A Jakobsen The relationship of VEGF polymorphisms with serum VEGF levels and progression-free survival inpatients with epithelial ovarian cancer Gynecol Oncol 117 (2010) 109e116
[14] CS Facemire AB Nixon R Grif 1047297ths H Hurwitz TM Coffman Vascularendothelial growth factor receptor 2 controls blood pressure by regulatingnitric oxide synthase expression Hypertension 54 (2009) 652e658
[15] P Bono H Elfving T Utriainen P Osterlund T Saarto T Alanko H JoensuuHypertension and clinical bene1047297t of bevacizumab in the treatment of advanced renal cell carcinoma Ann Oncol 20 (2009) 393e394
[16] SE Dahlberg AB Sandler JR Brahmer JH Schiller DH Johnson Clinicalcourse of advanced non-small-cell lung cancer patients experiencing hyper-tension during treatment with bevacizumab in combination with carboplatinand paclitaxel on ECOG 4599 J Clin Oncol 28 (2010) 949e954
[17] M Scartozzi E Galizia S Chiorrini R Giampieri R Berardi C PierantoniS Cascinu Arterial hypertension correlates with clinical outcome in colorectalcancer patients treated with 1047297rst-line bevacizumab Ann Oncol 20 (2009)227e230
[18] AR Padhani Dynamic contrast-enhanced MRI in clinical oncology currentstatus and future directions J Magn Reson Imaging 16 (2002) 407e422
[19] OMHahnC YangM Medved G KarczmarE KistnerT KarrisonE ManchenM Mitchell MJ Ratain WM Stadler Dynamic contrast-enhanced magneticresonance imaging pharmacodynamic biomarker study of sorafenib in meta-static renal carcinoma J Clin Oncol 26 (2008) 4572e4578
[20] AG Sorensen TT Batchelor WT Zhang PJ Chen P Yeo M WangD Jennings PY Wen J Lahdenranta M Ancukiewicz E di TomasoDG Duda RK Jain A vascular normalization index as potential mechanisticbiomarker to predict survival after a single dose of cediranib in recurrent
glioblastoma patients Cancer Res 69 (2009) 5296e
5300[21] A Salskov VS Tammisetti J Grierson H Vesselle FLT measuring tumor cellproliferation in vivo with positron emission tomography and 3rsquo-deoxy-3rsquo-[18F]1047298uorothymidine Semin Nucl Med 37 (2007) 429e439
[22] W Chen S Delaloye DH Silverman C Geist J Czernin J SayreN Satyamurthy W Pope A Lai ME Phelps T Cloughesy Predicting treat-ment response of malignant gliomas to bevacizumab and irinotecan byimaging proliferation with [18F] 1047298uorothymidine positron emission tomog-raphy a pilot study J Clin Oncol 25 (2007) 4714e4721
[23] PM Wilson D Yang MM Shi W Zhang C Jacques JC Barret K Danene-berg T Trarbach G Folprecht G Meinhardt HJ Lenz Use of intratumoralmRNA expression of genes involved in angiogenesis and HIF1 pathway topredict outcome to VEGFR tyrosine Kinase inhibitor (TKI) in patients enrolledin CONFIRM1 and CONFIRM2 ASCO Annu Meet (2008)
[24] MI Koukourakis A Giatromanolaki E Sivridis KC Gatter T TrarbachG Folprecht MM Shi D Lebwohl T Jalava D Laurent G MeinhardtAL Harris Prognostic and predictive role of lactate dehydrogenase 5expression in colorectal cancer patients treated with PTK787ZK 222584(Vatalanib) antiangiogenic therapy Clin Cancer Res 17 (2011) 4892e4900
[25] JR Hecht T Trarbach JD Hainsworth P Major E Jager RA Wolff K Lloyd-Salvant G Bodoky K Pendergrass W Berg BL Chen T Jalava G MeinhardtD Laurent D Lebwohl D Kerr Randomized placebo-controlled phase IIIstudy of 1047297rst-line oxaliplatin-based chemotherapy plus PTK787ZK 222584an oral vascular endothelial growth factor receptor inhibitor in patients withmetastatic colorectal adenocarcinoma J Clin Oncol 29 (2011) 1997e2003
[26] O Warburg On the origin of cancer cells Science 123 (1956) 309e314[27] MG Vander Heiden LC Cantley CB Thompson Understanding the warburg
effect the metabolic requirements of cell proliferation Science 324 (2009)1029e1033
[28] V Fritz L Fajas Metabolism and proliferation share common regulatorypathways in cancer cells Oncogene 29 (2010) 4369e4377
[29] AJ Levine AM Puzio-Kuter The control of the metabolic switch in cancers byoncogenes and tumor suppressor genes Science 330 (2010) 1340e1344
[30] SS Gambhir Molecular imaging of cancer with positron emission tomog-raphy Nat Rev Cancer 2 (2002) 683e693
[31] H Shim C Dolde BC Lewis CS Wu G Dang RA Jungmann R Dalla-FaveraCV Dang c-Myc transactivation of LDH-A implications for tumor metabolismand growth Proc Natl Acad Sci U S A 94 (1997) 6658 e6663
E Zulato et al Biochimie 94 (2012) 925e931930
8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931

8162019 Zulato Et Al - 2012 - Metabolic Effects of Anti-Angiogenic Therapy in Tumors
httpslidepdfcomreaderfullzulato-et-al-2012-metabolic-effects-of-anti-angiogenic-therapy-in-tumors 77
[32] DA Tennant RV Duran E Gottlieb Targeting metabolic transformation forcancer therapy Nat Rev Cancer 10 (2010) 267e277
[33] S Walenta M Wetterling M Lehrke G Schwickert K Sundfor EK RofstadW Mueller-Klieser High lactate levels predict likelihood of metastases tumorrecurrence and restricted patient survival in human cervical cancers CancerRes 60 (2000) 916e921
[34] RC Osthus H Shim S Kim Q Li R Reddy M Mukherjee Y Xu D WonseyLA Lee CV Dang Deregulation of glucose transporter 1 and glycolytic geneexpression by c-Myc J Biol Chem 275 (2000) 21797e21800
[35] RJ DeBerardinis JJ Lum G Hatzivassiliou CB Thompson The biology of
cancer metabolic reprogramming fuels cell growth and proliferation CellMetab 7 (2008) 11e20
[36] WH Koppenol PL Bounds CV Dang Otto warburgrsquos contributions tocurrent concepts of cancer metabolism Nat Rev Cancer 11 (2011) 325e337
[37] GL Semenza Targeting HIF-1 for cancer therapy Nat Rev Cancer 3 (2003)721e732
[38] K Bensaad A Tsuruta MA Selak MN Vidal K Nakano R BartronsE Gottlieb KH Vousden TIGAR a p53-inducible regulator of glycolysis andapoptosis Cell 126 (2006) 107e120
[39] S Matoba JG Kang WD Patino A Wragg M Boehm O GavrilovaPJ Hurley F Bunz PM Hwang p53 regulates mitochondrial respirationScience 312 (2006) 1650e1653
[40] RJ DeBerardinis A Mancuso E Daikhin I Nissim M Yudkoff S WehrliCB Thompson Beyond aerobic glycolysis transformed cells can engage inglutamine metabolism that exceeds the requirement for protein and nucle-otide synthesis Proc Natl Acad Sci U S A 104 (2007) 19345e19350
[41] M Yuneva N Zamboni P Oefner R Sachidanandam Y Lazebnik De1047297ciencyin glutamine but not glucose induces MYC-dependent apoptosis in humancells J Cell Biol 178 (2007) 93e105
[42] P Gao I Tchernyshyov TC Chang YS Lee K Kita T Ochi KI Zeller AM DeMarzo JE Van Eyk JT Mendell CV Dang c-Myc suppression of miR-23ab enhances mitochondrial glutaminase expression and glutamine metabo-lism Nature 458 (2009) 762e765
[43] HNawashiroN OtaniN Shinomiya SFukui HOoigawaK ShimaH MatsuoY Kanai H Endou L-type amino acid transporter 1 as a potential moleculartarget in human astrocytic tumors Int J Cancer 119 (2006) 484e492
[44] C Plathow WA Weber Tumor cell metabolism imaging J Nucl Med 49(Suppl 2) (2008) 43Se63S
[45] T Singhal TK Narayanan V Jain J Mukherjee J Mantil 11C-L-methioninepositron emission tomography in the clinical management of cerebralgliomas Mol Imaging Biol 10 (2008) 1e18
[46] JA Menendez R Lupu Fatty acid synthase and the lipogenic phenotype incancer pathogenesis Nat Rev Cancer 7 (2007) 763e777
[47] E Iorio D Mezzanzanica P Alberti F Spadaro C Ramoni S D rsquoAscenzoD Millimaggi A Pavan V Dolo S Canevari F Podo Alterations of cholinephospholipid metabolism in ovarian tumor progression Cancer Res 65 (2005)9369e9376
[48] E Iorio A Ricci M Bagnoli ME Pisanu G Castellano M Di Vito E VenturiniK Glunde ZM Bhujwalla D Mezzanzanica S Canevari F Podo Activation of phosphatidylcholine cycle enzymes in human epithelial ovarian cancer cellsCancer Res 70 (2010) 2126e2135
[49] S Faivre G Demetri W Sargent E Raymond Molecular basis for sunitinibef 1047297cacy and future clinical development Nat Rev Drug Discov 6 (2007)734e745
[50] G Nardo E Favaro M Curtarello L Moserle E Zulato L Persano E RossiG Esposito M Crescenzi O Casanovas UG Sattler W Mueller-KlieserB Biesalski O Thews R Canese E Iorio P Zanovello A AmadoriS Indraccolo Glycolytic phenotype and amp kinase modify the pathologicresponse of tumor xenografts to vegf neutralization Cancer Res (2011)
[51] CG Willett Y Boucher E di Tomaso DG Duda LL Munn RT TongDC Chung DV Sahani SP Kalva SV Kozin M Mino KS CohenDT Scadden AC Hartford AJ Fischman JW Clark DP Ryan AX ZhuLS Blaszkowsky HX Chen PC Shellito GY Lauwers RK Jain Directevidence that the VEGF-speci1047297c antibody bevacizumab has antivasculareffects in human rectal cancer Nat Med 10 (2004) 145e147
[52] S Mehta NP Hughes FM Buffa SP Li RF Adams A Adwani NJ Taylor
NC Levitt AR Padhani A Makris AL Harris Assessing early therapeuticresponse to bevacizumab in primary breast cancer using magnetic resonanceimaging and gene expression pro1047297les J Natl Cancer Inst Monogr 2011(2011) 71e74
[53] O Keunen M Johansson A Oudin M Sanzey SA Rahim F Fack F ThorsenT Taxt M Bartos R Jirik H Miletic J Wang D Stieber L Stuhr I MoenCB Rygh R Bjerkvig SP Niclou Anti-VEGF treatment reduces blood supplyand increases tumor cell invasion in glioblastoma Proc Natl Acad Sci U S A108 (2011) 3749e3754
[54] FA Howe SJ Barton SA Cudlip M Stubbs DE Saunders M MurphyP Wilkins KS Opstad VL Doyle MA McLean BA Bell JR Grif 1047297thsMetabolic pro1047297les of human brain tumors using quantitative in vivo 1Hmagnetic resonance spectroscopy Magn Reson Med 49 (2003) 223e232
[55] A Sreekumar LM Poisson TM Rajendiran AP Khan Q Cao J YuB Laxman R Mehra RJ Lonigro Y Li MK Nyati A Ahsan S Kalyana-
Sundaram B Han X Cao J Byun GS Omenn D Ghosh S PennathurDC Alexander A Berger JR Shuster JT Wei S Varambally C BeecherAM Chinnaiyan Metabolomic pro1047297les delineate potential role for sarcosinein prostate cancer progression Nature 457 (2009) 910e914
[56] ZJ Reitman G Jin ED Karoly I Spasojevic J Yang KW Kinzler Y HeDD Bigner B Vogelstein H Yan Pro1047297ling the effects of isocitrate dehydro-genase 1 and 2 mutations on the cellular metabolome Proc Natl Acad Sci US A 108 (2011) 3270e3275
[57] KJ Svensson JE Welch P Kucharzewska P Bengtson M BjurbergS Pahlman GB Ten Dam L Persson M Belting Hypoxia-mediated induction
of the polyamine system provides opportunities for tumor growth inhibitionby combined targeting of vascular endothelial growth factor and ornithinedecarboxylase Cancer Res 68 (2008) 9291e9301
[58] A Alexander CL Walker The role of LKB1 and AMPK in cellular responses tostress and damage FEBS Lett 585 (2011) 952e957
[59] DB Shackelford RJ Shaw The LKB1-AMPK pathway metabolism andgrowth control in tumour suppression Nat Rev Cancer 9 (2009) 563e575
[60] SA Hawley DA Pan KJ Mustard L Ross J Bain AM EdelmanBG Frenguelli DG Hardie Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinaseCell Metab 2 (2005) 9e19
[61] A Woods SR Johnstone K Dickerson FC Leiper LG Fryer D NeumannU Schlattner T Wallimann M Carlson D Carling LKB1 is the upstreamkinase in the AMP-activated protein kinase cascade Curr Biol 13 (2003)2004e2008
[62] BB Zhang G Zhou C Li AMPK an emerging drug target for diabetes and themetabolic syndrome Cell Metab 9 (2009) 407e416
[63] Z Luo M Zang W Guo AMPK as a metabolic tumor suppressor control of metabolism and cell growth Future Oncol 6 (2010) 457e470
[64] DG Hardie AMP-activated protein kinase as a drug target Annu Rev Phar-macol Toxicol 47 (2007) 185e210
[65] R Rattan RP Graham JL Maguire S Giri V Shridhar Metformin suppressesovarian cancer growth and metastasis with enhancement of cisplatin cyto-toxicity in vivo Neoplasia 13 (2011) 483e491
[66] GZ Rocha MM Dias ER Ropelle F Osorio-Costa FA Rossato AE VercesiMJ Saad JB Carvalheira Metformin Ampli1047297es chemotherapy-induced AMPKactivation and Antitumoral growth Clin Cancer Res 17 (2011) 3993e4005
[67] KR Laderoute K Amin JM Calaoagan M Knapp T Le J Orduna M ForetzB Viollet 5rsquo-AMP-activated protein kinase (AMPK) is induced by low-oxygenand glucose deprivation conditions found in solid-tumor microenvironmentsMol Cell Biol 26 (2006) 5336e5347
[68] D Tsavachidou-Fenner N Tannir P Tamboli W Liu D Petillo B TehGB Mills E Jonasch Gene and protein expression markers of response tocombined antiangiogenic and epidermal growth factor targeted therapy inrenal cell carcinoma Ann Oncol 21 (2010) 1599e1606
[69] J Carretero PP Medina R Blanco L Smit M Tang G Roncador L MaestreE Conde F Lopez-Rios HC Clevers M Sanchez-Cespedes Dysfunctional
AMPK activity signalling through mTOR and survival in response to energeticstress in LKB1-de1047297cient lung cancer Oncogene 26 (2007) 1616e1625[70] RJ Shaw M Kosmatka N Bardeesy RL Hurley LA Witters RA DePinho
LC Cantley The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress ProcNatl Acad Sci U S A 101 (2004) 3329e3335
[71] JB Brugarolas F Vazquez A Reddy WR Sellers WG Kaelin Jr TSC2regulates VEGF through mTOR-dependent and -independent pathwaysCancer Cell 4 (2003) 147e158
[72] DB Shackelford DS Vasquez J Corbeil S Wu M Leblanc CL Wu DR VeraRJ Shaw mTOR and HIF-1alpha-mediated tumor metabolism in an LKB1mouse model of Peutz-Jeghers syndrome Proc Natl Acad Sci U S A 106(2009) 11137e11142
[73] M Sanchez-Cespedes P Parrella M Esteller S Nomoto B Trink JM EnglesWH Westra JG Herman D Sidransky Inactivation of LKB1STK11 isa common event in adenocarcinomas of the lung Cancer Res 62 (2002)3659e3662
[74] AB Lima LT Macedo AD Sasse Addition of bevacizumab to chemotherapyin advanced non-small cell lung cancer a systematic review and meta-anal-
ysis PLoS One 6 (2011) e22681[75] AM Gonzalez-Angulo F Meric-Bernstam Metformin a therapeutic oppor-
tunity in breast cancer Clin Cancer Res 16 (2010) 1695e1700[76] M Crescenzi L Persano G Esposito E Zulato L Borsi E Balza A Ruol
E Ancona S Indraccolo A Amadori Vandetanib improves anti-tumor effectsof L19mTNFalpha in xenograft models of esophageal cancer Clin Cancer Res17 (2011) 447e458
[77] Y Shaked A Ciarrocchi M Franco CR Lee S Man AM Cheung DJ HicklinD Chaplin FS Foster R Benezra RS Kerbel Therapy-induced acuterecruitment of circulating endothelial progenitor cells to tumors Science 313(2006) 1785e1787
[78] AA van der Veldt MR Meijerink AJ van den Eertwegh JB HaanenE Boven Choi response criteria for early prediction of clinical outcome inpatients with metastatic renal cell cancer treated with sunitinib Br J Cancer102 (2010) 803e809
E Zulato et al Biochimie 94 (2012) 925e931 931


















