


Vibrio cholerae by PhoA/GFP fusion analysis 3 ACCEPTED
31
1 Membrane topology mapping of the Na + -pumping NADH:quinone oxidoreductase from 1 Vibrio cholerae by PhoA/GFP fusion analysis 2 3 Ellen B. Duffy and Blanca Barquera* 4 Department of Biology and Center for Biotechnology and Interdisciplinary Studies, 5 Rensselaer Polytechnic Institute. 6 7 8 Running Title: Membrane topology mapping of Na + -NQR from Vibrio cholerae 9 * Corresponding author: Mailing address: Rm. 2239, Biotechnology Center, 110 Eighth St, Troy, NY 12180. Tel.: (518) 276-3861. 10 E-mail:[email protected] . 11 ACCEPTED Copyright © 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. J. Bacteriol. doi:10.1128/JB.01383-06 JB Accepts, published online ahead of print on 13 October 2006 on April 3, 2019 by guest http://jb.asm.org/ Downloaded from
Transcript of Vibrio cholerae by PhoA/GFP fusion analysis 3 ACCEPTED

1
Membrane topology mapping of the Na+-pumping NADH:quinone oxidoreductase from 1
Vibrio cholerae by PhoA/GFP fusion analysis 2
3
Ellen B. Duffy and Blanca Barquera* 4
Department of Biology and Center for Biotechnology and Interdisciplinary Studies, 5
Rensselaer Polytechnic Institute. 6
7
8
Running Title: Membrane topology mapping of Na+-NQR from Vibrio cholerae 9
*Corresponding author: Mailing address: Rm. 2239, Biotechnology Center, 110 Eighth St, Troy, NY 12180. Tel.: (518) 276-3861. 10
E-mail:[email protected]. 11
ACCEPTED
Copyright © 2006, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01383-06 JB Accepts, published online ahead of print on 13 October 2006
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

2
ABSTRACT 1
The membrane topology of the six subunits of Na+-translocating NADH:quinone oxidoreductase (Na
+-NQR) from Vibrio 2
cholerae was determined by a combination of topology prediction algorithms and construction of C-terminal fusions. Fusion 3
expression vectors contained either bacterial alkaline phosphatase (phoA) or green-fluorescent protein (gfp) genes as reporters of 4
periplasmic and cytoplasmic localization, respectively. A majority of topology prediction algorithms did not predict any 5
transmembrane helices for NqrA. Lack of PhoA activity when fused to the C-terminus of NqrA and the observed fluorescence of the 6
GFP C-terminal fusion confirms that this subunit is localized to the cytoplasmic side of the membrane. Analysis of four PhoA fusions 7
for NqrB indicates that this subunit has nine transmembrane helices and that residue T236, the binding site for FMN, resides in the 8
cytoplasm. Three fusions confirm that the topology of NqrC consists of two transmembrane helices with the FMN binding site, 9
residue T225, on the cytoplasmic side. Fusion analysis of NqrD and NqrE showed almost mirror image topologies each consisting of 10
six transmembrane helices; the results for NqrD and NqrE are consistent with the topologies of Escherichia coli homologs YdgQ and 11
YdgL, respectively. The NADH, FAD, and Fe-S center binding sites of NqrF were localized to the cytoplasm. The determination of 12
the topology of the subunits of Na+-NQR provides valuable insights into the location of cofactors and identifies targets for 13
mutagenesis to characterize this enzyme in more detail. The finding that all the redox cofactors are localized to the cytoplasmic side 14
of the membrane is discussed. 15
16
17
18
19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

3
INTRODUCTION 1
The Na+-pumping NADH:quinone oxidoreductase (Na
+-NQR) is the main entry point for electrons into the aerobic respiratory 2
chain of many marine and pathogenic bacteria. The enzyme oxidizes NADH, reduces quinone, and uses the free energy released in 3
this redox reaction to generate a sodium motive force that can be used for motility and metabolic work (2, 5, 6, 8, 14, 19, 35, 43) 4
The Na+-NQR complex is made up of six subunits and accommodates a number of cofactors including several flavins (FAD, 5
FMN, and riboflavin), and a 2Fe-2S cluster (3, 4, 7). Two FMN’s are bound covalently to subunits B and C of the enzyme and a non-6
covalently bound FAD resides in subunit F (1, 15, 28). A molecule of ubiquinone-8 is believed to be bound near Gly-141 (Vibrio 7
cholerae numbering) of NqrB on the basis of inhibitor studies. (16). NqrF includes a motif typical of NADH binding sites. This 8
evidence together with mutant studies indicates that this subunit is the entry point of electrons into the enzyme (29, 31). 9
Na+-NQR is an integral membrane enzyme. Five of the six subunits that make up the complex (all but NqrA) apparently 10
include membrane spanning segments. In order to elucidate the mechanism that operates in this enzyme it is very important to know 11
where a given point in the amino acid sequence is situated with respect to the membrane; whether it is on the cytoplasmic side, within 12
the membrane itself, or on the periplasmic side. Topological information of this kind for subunits NqrB, NqrC and NqrF is important 13
to elucidate where, in relation to the membrane plane, the different cofactors are located, in particular the covalently bound FMN’s in 14
subunits B and C. This information is essential to studying the mechanism that couples the redox reaction of Na+-NQR with pumping 15
of sodium. To function correctly, an ion pump must move ions across the membrane in a specific direction. In Na+-NQR, sodium ions 16
are taken up from the cytoplasmic side of the membrane and released on the extracellular side, resulting in the outer side of the cell 17
membrane becoming positively charged with respect to the inner side. The directional (vectorial) nature of the ion pumping process 18
must be linked to an oriented placement of the protein with respect to the sides of the membrane. Localization of the redox cofactors 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

4
and other putative pump-related sites with respect to the sides of the membrane is important for understanding the pump mechanism 1
because it can reveal whether the work involved in moving Na+ up the membrane potential gradient is done during Na
+ uptake or 2
release. Furthermore, topological models of subunits NqrB, NqrD and NqrE can help to identify conserved amino acid residues 3
located within the membrane spanning regions which are likely to be involved in sodium pathways. 4
Computer prediction programs can be used to generate topological maps of membrane proteins. We employed web-based 5
topology prediction algorithms to create a set of seven models for each subunit. The predictions generated in this way disagree in 6
important respects. Importantly, the prediction of membrane spanning helices is more accurate than predictions of absolute sidedness, 7
i.e., whether the N-terminus is on the cytosolic or periplasmic side. These discrepancies can often be resolved by studying fusions of 8
reporters to the C-terminus of full-length or truncated membrane proteins. Typically, a reporter that is only active on one side of the 9
membrane is used in parallel with another reporter that is only active on the opposite side (38). For our work, we chose to use 10
bacterial alkaline phosphatase (PhoA) (22) as a reporter of periplasmic localization and green-fluorescent protein (GFP) (30), as a 11
reporter of cytoplasmic localization. All of the work with PhoA was done in E. coli since it requires an alkaline phosphatase deficient 12
strain, and no strain of this kind was available for V. cholerae. For the work with GFP, both E. coli and V. cholerae were used. The 13
use of GFP in E. coli is well established, but to our knowledge this is the first time that GFP fusions have been used in V. cholerae. In 14
several cases, this was successful, but some of the GFP-fusions products were apparently toxic to the cells. 15
By using reporter group gene-fusion methods to resolve the discrepancies among topology modeling predictions, we were able 16
to arrive at a single consensus model for each subunit of Na+-NQR (12, 18). This topological model will be useful for future 17
mechanistic studies because each of the redox cofactors for which the amino acid binding site is known, can now be localized with 18
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

5
respect to membrane sidedness. Interestingly, all of the localized cofactors are found on the cytoplasmic (negative) side of the 1
membrane, in contrast to what is seen in many other ion pumps. 2
3
MATERIALS AND METHODS 4
Bacterial strains, plasmids, and primers 5
Strains and plasmids used in this study are listed in Table 1. Primers were synthesized by either IDT, Inc. or Operon and were 6
HPLC purified. Primers included appropriate restriction cleavage sites to allow insertion of the PCR products into the pBAD vectors 7
(KpnI for pBAD-PhoA and NheI or SpeI (NqrD) for pBAD-GFP). 8
Membrane topology prediction methods 9
Seven current web-version topology prediction algorithms were utilized: TMHMMfix (27), MEMSAT3 (24), TOPPRED II 10
(10), TMpred (http://www.ch.embnet.org/software/TMPRED_form.html), HMMTOP 2.0 (37), DAS (11) and SOUSI (17). Primary 11
amino acid sequences of each subunit were individually submitted and default options were used. A summary of the predictions for 12
each of the six subunits of Na+-NQR is shown in Table 2. C-terminal fusions were then made to discriminate between the various 13
predictions, focusing on point of discrepancy. Using the results of the fusion studies together with the topological models, a 14
consensus model was constructed according to Ikeda et al. (18). 15
Molecular genetic techniques 16
V. cholerae O395N1 genomic DNA was used as source of nqr operon. Genomic DNA was extracted from an overnight 17
culture of V. cholerae using the Wizard purification kit (Promega) according to the manufacturer’s instructions. Full-length or partial 18
fragments of individual subunits of NQR were amplified by the polymerase chain reaction (PCR) using a high-fidelity polymerase 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

6
(Platinum SuperMix High Fidelity, Invitrogen), subjected to agarose gel electrophoresis, and single bands of the correct size were 1
purified from the agarose (MinElute or Qiaquick Gel Extraction Kit, Qiagen). Fragments were first cloned into pCR2.1-TOPO TA 2
vectors (Invitrogen) and transformed into E. coli Top10 cells (Invitrogen). Cloned PCR fragments were excised from the pCR2.1 3
TOPO TA vector with appropriate restriction enzymes (New England Biolabs, NEB). Resulting restriction enzyme digestion products 4
were purified from agarose gels. Fusion expression vectors were digested with corresponding restriction enzymes and 5
dephosphorylated with Antarctic Phosphatase (NEB), which was subsequently heat-inactivated (65 °C for 15 min). Vectors and 6
fragments were ligated using T4 ligase (NEB or Invitrogen) overnight at 16 °C. The resulting plasmids were transformed into E. coli 7
Top10 cells and the orientation of the insert checked by restriction enzyme digestion. Plasmids with correct orientation relative to the 8
reporter gene were transformed by electroporation (BioRad) into E. coli LMG194 for PhoA fusions and V. cholerae O395N1 or V. 9
cholerae ∆nqr for GFP fusions. 10
For GDNC and GDN125, an 18 amino acid linker was inserted in-frame between the nqr gene fragment and the GFP gene (40) 11
using a mutagenesis kit (Quikchange, Stratagene). Fusions for GDNC and GDN125 without the linker were also constructed. 12
Reporter gene assays 13
Overnight cultures of strains containing pBAD-PhoA/GFP fusions were re-inoculated into 50 mL cultures of Luria Broth 14
(Miller) with antibiotics (ampicillin, 100 µg/mL and, in addition in the case of V. cholerae, streptomycin, 50 µg/mL) and allowed to 15
grow to early log phase before addition of L-(+)-arabinose (Sigma) to a final concentration of 0.2%, to induce expression. Growth 16
was allowed to continue, and at late-log phase, cells were harvested for analysis of PhoA or GFP activity (see below). At the same 17
time, a 1-mL aliquot of culture was removed for SDS-PAGE and Western blot analysis. These cells were harvested by centrifugation 18
and the cell pellets stored at -20 °C until used. 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

7
A quantitative assay of alkaline phosphatase activity was conducted using the method of (21). After growth of cultures as 1
described above, a 5-mL aliquot of culture was removed, 50 µL of 100 mM Iodoacetamide (Sigma) was added, and the cells were 2
incubated for 10 min at 37 °C. Cells were then harvested by centrifugation, washed once with 5 mL cold PhoA buffer (10mM Tris, 3
pH 8.0, 10 mM MgSO4, 1mM Iodoacetamide) and resuspended in 1 mL 1M Tris, pH 8.0, 1mM Iodoacetamide. The cell density was 4
measured by absorbance at 600 nm in an Agilent spectrophotometer, Model 8453. PhoA activity was measured in cells permeabilized 5
with sodium dodecyl sulfate (SDS) and chloroform incubated for 45 min at 37 °C with p-nitrophenyl phosphate (pNPP) as substrate. 6
Absorbance at 420 nm and 550 nm was measured and used to calculate activity using the equation of Manoil (21). 7
To observe GFP fluorescence, 10 mL aliquots of late log-phase induced cells were harvested and resuspended in 0.5 mL of 10 8
mM Tris buffer, pH 8. Concentrated cells were mounted on 3% agarose pads, covered with a cover slip, and observed using a Zeiss 9
Axiovert 200M epifluorescent microscope equipped with a Plan-Neofluor 100X/1.3 N.A. oil immersion objective, 1.6X Optavar, and 10
Endow GFP bandpass filter set. Images were obtained using a Zeiss AxioCam MRm CCD digital camera and Axiovision software. 11
Different samples were photographed with the same exposure time. 12
Expression and analysis of fusions 13
For SDS-PAGE, thawed cell pellets were added to 100 µL of a 2X sample loading buffer and incubated in a boiling water bath 14
for 10 min. Denatured samples (10 µL) were loaded into a 4-12% Tris-Bis acrylamide gel (Invitrogen). Following electrophoresis, 15
the gel was first photographed under UV illumination to detect presence of fluorescent cofactors and then proteins were transferred to 16
a PVDF membrane. Western immunoblotting was performed on the PVDF membranes according to the protocol for the Western 17
Breeze kit (Invitrogen). The primary antibody was either mouse anti-alkaline phosphatase or mouse anti-GFP (Chemicon). 18
Membrane isolation 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

8
The PhoA fusions ABNC and ACNC in V. cholerae ∆nqr were grown in LB medium (with the appropriate antibiotics) to the 1
beginning of early log-phase at which point arabinose was added to a final concentration of 0.2%. The cultures were allowed to grow 2
to the beginning of stationary phase and then harvested. Cells were washed with 10 mM Tris buffer, pH 8, 200 mM NaCl, 5 mM 3
MgSO4. Cells were broken using a Microfluidizer (Model 110S, Microfluidics) in the presence of DNAse I (Sigma) and the protease 4
inhibitor AEBSF (4-(2aminoethyl) benzenesulfonyl fluoride). Cell debris and unbroken cells were removed by centrifugation at 5
3500g for 30 min. The supernatant containing the membrane fraction was then centrifuged at 100000 g for 12 hours. The resulting 6
pellet was resuspended in the Tris buffer using a glass-Teflon homogenizer and membrane aliquots were frozen at -80oC until needed. 7
Protein concentration of the membrane fractions was determined by means of the BCA method (Pierce). 8
9
RESULTS 10
Cloning and expression 11
For all six subunits, full-length C-terminal fusions with PhoA were constructed. In addition, truncated C-terminal fusions with 12
PhoA were constructed for NqrB, NqrC, NqrD, and NqrF. No truncated C-terminal fusions were constructed for NqrA and NqrE 13
because topology prediction algorithms were in near agreement and could be discriminated based on the full-length fusion. A select 14
set of fusions with GFP were constructed to provide confirmation of PhoA results. C-terminal fusions of full-length NqrA, NqrC, 15
NqrD, and NqrE, and truncated C-terminal fusions of NqrC and NqrD to GFP were constructed. 16
All PhoA fusions were expressed upon addition of arabinose. Several GFP fusions, in particular the NqrD fusions, showed 17
inhibition of growth upon addition of arabinose, while still showing limited levels of expression (not shown). 18
PhoA fusion analysis 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

9
The activity of PhoA fusions are reported in Table 3. Based on PhoA activity of E. coli LMG194 cells (typically observed to 1
be between 0 to 20 units), a threshold of 40 units (a unit is the amount of substrate, pNPP, hydrolyzed per minute per OD600) was set 2
for positive activity; high PhoA activity was defined as >100 units. The full-length NqrA fusion showed no PhoA activity, indicating 3
a cytoplasmic location of the C terminus. Full-length fusions of NqrC and NqrE to PhoA yielded high PhoA activity. Only two of 4
nine truncated fusions showed positive PhoA activity: ABN377 and AFN80. 5
GFP fusion analysis 6
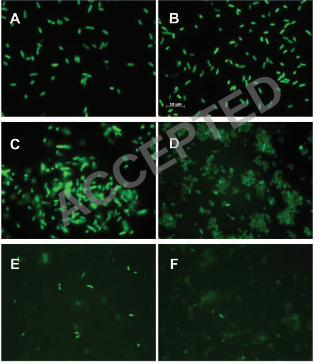
Observation of GFP activity in V. cholerae wild-type and ∆nqr confirmed the ability of GFP to serve as a reporter of 7
cytoplasmic localization (Fig. 1). Full-length C-terminal GFP fusions for NqrC and NqrE did not fluorescence under UV illumination 8
strengthening the conclusions from the PhoA analysis which show that the C-termini of NqrC and NqrE both lie on the periplasmic 9
side of the membrane (Fig. 3 and 5). The full-length C-terminal GFP fusion for NqrA fluoresced under UV illumination, again 10
confirming the results with the PhoA fusions and showing that C-terminus of this subunit lies on the cytoplasmic side of the 11
membrane (Fig. 1C and D). Two GFP fusions with NqrD (GDN125 and GDNC) where the reporter group is expected to be in the 12
cytoplasm, and therefore fluorescent, showed only low levels of fluorescence in the microscope (Fig. 1E and F). This is consistent 13
with the fact that the addition of inducer (arabinose) to these cells led to inhibition of growth and low levels of expression as judge by 14
Western blots (not shown). 15
Topology of Na+-NQR 16
The topology of the six subunits of Na+-NQR from V. cholerae was determined through a combination of topology prediction 17
algorithms and gene fusion analysis. Topology prediction algorithms and rules for generating a consensus model were useful in 18
reducing the total number of fusions to construct. C-terminal full-length and truncated fusions to PhoA were constructed and 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

10
analyzed, yielding data used to model the topologies of the six subunits. Fusions with GFP were used to confirm a subset of the PhoA 1
fusions. 2
The results of both the PhoA and GFP fusions with full-length NqrA are in agreement with a cytoplasmic location of this 3
subunit. No transmembrane helices were predicted by the majority of algorithms, which can be taken to indicate that NqrA is a 4
soluble protein. We can therefore conclude that the entire subunit A resides on the cytoplasmic side of the membrane. The fact that 5
NqrA consistently co-purifies with the other five subunits indicates that it must be tightly attached to the complex by noncovalent 6
interactions. 7
The PhoA activity results for NqrB are consistent with the topology model shown in Fig. 2. NqrB has nine transmembrane 8
helices containing seven negatively-charged residues that could be involved in sodium translocation. All of these residues are 9
conserved across the Na+-NQR sequences of several organisms (Fig. 2). Analysis of the NqrB subunit also showed that the putative 10
quinone binding site, G141 (G140 in Vibrio alginolyticus) (16) is localized to the cytoplasmic half of transmembrane helix III. The 11
FMN binding site, T236, was localized to the cytoplasmic loop between transmembrane helix V and VI. 12
The PhoA fusion constructs also provided evidence that the FMN cofactors of NqrB and C are incorporated even when the 13
remaining subunits of Na+-NQR are not present. When the membrane fraction from V. cholerae (∆nqr) containing the complete NqrB 14
subunit fused to alkaline phosphatase (ABNC fusion), was run on SDS-PAGE (Fig. 7), a fluorescent band was observed at the 15
predicted molecular weight for the fusion protein (~80 kDa) indicating the presence of the FMN. Similarly, the PhoA fusion for NqrC 16
(ACNC fusion) also gave a fluorescent band at predicted molecular weight for the fusion protein (~70 kDa) indicating that the FMN in 17
the subunit is also bound (Fig. 7). Since the SDS-PAGE gels denature the protein these results also indicate that the flavins in these 18
two subunits are already covalently bound. 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

11
The topologies of homologues of NqrD and NqrE (YdgQ and YdgL, respectively) in E. coli have been previously determined 1
and consist of six transmembrane helices each, but with opposite overall orientations (YdgQ has its N- and C- terminus in the 2
cytoplasm while YdgL has its N- and C-terminus in the periplasm) (33). Our results are in agreement with the topology of these 3
homologues (Figs. 4 and 5). While the consensus model for NqrD had five transmembrane helices, three of the seven predictions 4
included a sixth transmembrane helix at the N-terminus. Our method of constructing fusions (via PCR and subcloning into the fusion 5
vector) was unable to generate a fusion construct to test for the presence of this initial helix. However, analysis of the primary 6
sequence in conjunction with the “positive-inside” rule (39) suggest that the N-terminus is located in the cytoplasm, which would 7
indicate that the initial helix exists. Both subunits contain negatively-charged amino acids within their six transmembrane helices and 8
several of these residues are conserved. 9
The fusion activity results indicated that the topology of NqrC consists of two transmembrane helices with the FMN binding 10
site, T225, residing in the cytoplasm (Fig. 3). Fusion activity results for NqrF indicate a cytoplasmic localization of the cofactor 11
binding regions (Fig. 6). A single transmembrane helix is most likely for NqrF, despite the two helices predicted in the consensus. 12
All five topology algorithms that predicted a second transmembrane helix scored this helix as low probability of formation. Assuming 13
a single-helix topology is correct, an explanation of the positive PhoA activity for AFN80 is that the weakly hydrophobic region (the 14
second predicted helix, indicated by white squares in Fig. 6) and the lack of a strong positively-charged region preceding the fusion 15
site leads to translocation of the PhoA moiety to the periplasm resulting in the observed activity. However, the existence of this second 16
predicted helix is unlikely since this two-helix model would locate one of the four cysteines that ligate the 2Fe-2S in the middle of 17
membrane span. In addition, Turk et al. (36) was able to produce a soluble variant of NqrF by removing residues Threonine-3 to 18
Alanine-25, consistent with this stretch of the protein being the membrane anchor. 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

12
1
2
DISCUSSION 3
Protein reporter-group fusion is a well established and proven method of studying the topology of membrane proteins (12, 22). 4
In this method a marker is attached at a specific point in the amino acid sequence of a membrane protein so as to reveal the location of 5
that point with respect to the membrane. This is accomplished by making a fusion on the gene level. The gene for the membrane 6
protein is truncated at a point of interest, and the gene for a “reporter group” protein is spliced in. The objective is that when the 7
modified gene is expressed, the remaining part of the membrane protein will be inserted into the membrane in the normal fashion, 8
guiding the reporter group to one side of the membrane or the other, depending on where the membrane protein was truncated. A 9
reporter group must be a protein that exhibits different properties depending on whether it resides in the cytosol or the periplasmic 10
space. To be useful, the properties of the reporter group must manifest themselves in whole cells. We have used two reporter proteins 11
in this study: GFP as the cytoplasmic reporter, (12, 13) in both E. coli and V. cholerae (34) and PhoA (26) as the periplasmic reporter, 12
in a PhoA deficient strain of E . coli. 13
Using a combination of computer-based topology prediction and gene-fusion studies we have obtained a consensus model for 14
each of the six subunits of the Na+-NQR complex from V. cholerae. For each subunit at least one fusion was made providing an 15
experimental point of reference to establish sidedness for the computer predictions. In several cases additional fusions were made at 16
intermediate points in the nqr genes to resolve discrepancies among the predictions from different algorithms (Table 2). For example, 17
in the case of NqrC the seven topology algorithms gave four different predictions (Table 2) and the consensus algorithm produced a 18
prediction of one transmembrane helix with the C-terminus located in the periplasm. We constructed reporter fusions at three different 19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

13
points in the sequence of this subunit. The results indicate that NqrC has two transmembrane helices and that the C-terminus lies in 1
the periplasm, consistent with the predictions from the HMMTOP algorithm. This result allows us to definitively localize the FMN 2
cofactor to the cytoplasmic side of the membrane, result which would have been impossible to obtain from the theoretical models 3
alone. 4
The results presented in this paper indicate that all the redox-active cofactors of Na+-NQR are located on the cytoplasmic 5
(negative) side of the membrane, with the possible exception of riboflavin, whose location in the protein has not been established. 6
This topological placement is analogous to that of the H+-pumping NADH:quinone oxidoreductase (complex I) but contrasts with 7
some other redox-driven ion pumps . These differences almost certainly reflect important aspects of the ion pumping mechanism (9, 8
23, 32). In the case of cytochrome c oxidase, a proton pump, the redox cofactors all lie to the positive side of the membrane, that is, 9
toward the side where positively charged ions are ejected. The binuclear oxygen-reducing site, which is believed to play an important 10
role in proton pumping, is located near the electrical center of the membrane (Fig. 8). For protons to reach the binuclear center, they 11
must cross a fraction of the membrane, and therefore, a significant part of the work of ion translocation is done during proton uptake 12
(41, 42). In Na+-NQR, with the redox cofactors near the negative side of the membrane, if we assume that there is a discrete binding 13
site for sodium pumping associated with a redox cofactor; most of the electric work of ion translocation would need to be done during 14
sodium release. We hope to study this question further by biophysical methods. 15
The results in this paper will help to guide future structure-function studies on Na+-NQR. They will be of particular 16
importance for investigations of the chemiosmotic properties of this redox driven Na+ pump, where it is essential to know the location 17
of cofactors and amino acid residues with respect to the membrane dielectric. 18
19
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

14
1
2
ACKNOWLEDGMENTS 3
This study was funded by NIH grant R01 GM069936 to B.B. The authors greatly appreciate the help, support and critical 4
reading of the manuscript of Dr. Joel Morgan. We thank Dr. Fern Finger and Ms. Christine Morton for use of and assistance with the 5
epifluorescence microscope. We are grateful to Chengyen Wu for his skillful assistance in the preparation of the nqr deletion strain. 6
We also thank Dr. Nadia Dolganov and Dr. Gary Schoolnik at Stanford University for their help in the preparation of strains. 7
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

15
REFERENCES 1
1. Barquera, B., Hase, C.C., Gennis, R.B. 2001. Expression and mutagenesis of the NqrC subunit of the NQR respiratory Na(+) 2
pump from Vibrio cholerae with covalently attached FMN. FEBS Lett. 492:45-49. 3
2. Barquera, B., Hellwig, P., Zhou, W., Morgan, J.E., Hase, C.C., Gosink, K.K., Nilges, M, Bruesehoff, P.J., Roth, A., 4
Lancaster, C.R., Gennis, R.B. 2002. Purification and characterization of the recombinant Na(+)-translocating NADH:quinone 5
oxidoreductase from Vibrio cholerae. Biochemistry. 2002 Mar 19; 41:3781-3789. 6
3. Barquera, B., Nilges, M.J., Morgan, J.E., Ramirez-Silva, L., Zhou, W., Gennis, R.B. 2004. Mutagenesis study of the 2Fe-7
2S center and the FAD binding site of the Na(+)-translocating NADH:ubiquinone oxidoreductase from Vibrio cholerae. 8
Biochemistry. 43:12322-12330. 9
4. Barquera, B., Zhou, W., Morgan JE, Gennis RB. 2002. Riboflavin is a component of the Na+-pumping NADH-quinone 10
oxidoreductase from Vibrio cholerae. Proc Natl Acad Sci U S A. 99:10322-10324. 11
5. Bertsova, Y., Bogachev, A.V. 2004. The origin of the sodium-dependent NADH oxidation by the respiratory chain of 12
Klebsiella pneumoniae. FEBS Lett. 563:207-212. 13
6. Bogachev, A., Bertsova YV, Barquera B, Verkhovsky, M.I. 2001. Sodium-dependent steps in the redox reactions of the 14
Na+-motive NADH:quinone oxidoreductase from Vibrio harveyi. Biochemistry. 40:7318-23. 15
7. Bogachev, A., Bertsova, Y.V., Ruuge, E.K., Wikstrom, M., Verkhovsky, M.I. 2002. Kinetics of the spectral changes during 16
reduction of the Na+-motive NADH:quinone oxidoreductase from Vibrio harveyi. Biochim Biophys Acta. 1556:113-20. 17
8. Bogachev, A., Verkhovsky, M.I. 2005. Na+-translocating NADH:quinone oxidoreductase:Progress achieved and prospects of 18
investigations. Biochemistry (Moscow) 70:143-149. 19
9. Brandt, U. 2006. Energy converting NADH:quinone oxidoreductase (Complex I). Annu Rev Biochem 75:69-92. 20
10. Claros, M. G., von Heijne, G. 1994. TopPred II: an improved software for membrane protein structure predictions. Comput 21
Appl Biosci 10:685-686. 22
11. Cserzo, M., Wallin, E., Simon, I., von Heijne, G., Elofsson, A. 1997. Prediction of transmembrane alpha-helices in 23
prokaryotic membrane proteins: the dense alignment surface method. Protein Eng 10:673-676. 24
12. Drew, D., Sjostrand, D., Nilsson, J., Urbig, T., Chin, C. N., de Gier, J. W., von Heijne, G. 2002. Rapid topology mapping 25
of Escherichia coli inner-membrane proteins by prediction and PhoA/GFP fusion analysis. Proc Natl Acad Sci USA 99:2690-26
2695. 27
13. Feilmeier, B. J., Iseminger, G., Schroeder, D., Webber, H., Phillips, G.J. 2000. Green fluorescent protein functions as a 28
reporter for protein localization in Escherichia coli. J Bacteriol 182:4068-4076. 29
14. Hayashi, M., Nakayama, Y., Unemoto, T. 2001. Recent progress in the Na(+)-translocating NADH-quinone reductase from 30
the marine Vibrio alginolyticus. Biochim Biophys Acta. 1505:37-44. 31
15. Hayashi, M., Nakayama, Y., Yasui, M., Maeda, M., Furuishi, K., Unemoto, T. 2001. FMN is covalently attached to a 32
threonine residue in the NqrB and NqrC subunits of Na(+)-translocating NADH-quinone reductase from Vibrio alginolyticus. 33
FEBS Lett. 488:5-8. 34
16. Hayashi, M., Shibata, N., Nakayama, Y., Yoshikawa, K, Unemoto, T. 2002. Korormicin insensitivity in Vibrio 35
alginolyticus is correlated with a single point mutation of Gly-140 in the NqrB subunit of the Na(+)-translocating NADH-36
quinone reductase. Arch Biochem Biophys. 401:173-177. 37
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

16
17. Hirokawa, T., Boon-Chieng, S., Mitaku, S. 1998. SOSUI: classification and secondary structure prediction system for 1
membrane proteins. Bioinformatics 14:378-379. 2
18. Ikeda, M., Arai, M., Lao, D. M., Shimizu, T. 2002. Transmembrane topology prediction methods: a re-assessment and 3
improvement by a consensus method using a dataset of experimentally-characterized transmembrane topologies. In Silico Biol 4
2:19-33. 5
19. Kojima, S., Yamamoto, K., Kawagishi, I., Homma, M. 1999. The polar flagellar motor of Vibrio cholerae is driven by an 6
Na+ motive force. J Bacteriol 181:1927-1930. 7
20. Lu, C., Bentley, W. E., Rao, G. 2004. A high-throughput approach to promoter study using green fluorescent protein. 8
Biotechnol Prog 20:1634-1640. 9
21. Manoil, C. 1991. Analysis of membrane protein topology using alkaline phosphatase and beta-galactosidase gene fusions. 10
Methods Cell Biol 34:61-75. 11
22. Manoil, C., Mekalanos, J. J., Beckwith, J. 1990. Alkaline phosphatase fusions: sensors of subcellular location. J Bacteriol 12
172:515-518. 13
23. Mathiesen, C., Hagerhall, C. 2002. Transmembrane topology of the NuoL, M and N subunits of NADH:quinone 14
oxidoreductase and their homologues among membrane-bound hydrogenases and bona fide antiporters. Biochem Biophys Acta 15
1556:121-132. 16
24. McGuffin, L. J., Bryson, K., Jones, D. T. 2000. The PSIPRED protein structure prediction server. Bioinformatics 16:404-17
405. 18
25. Mekalanos, J. J., Swartz, D.J., Pearson, G.D., Harford, N., Groyne, F., de Wilde, M. 1983. Cholera toxin genes: 19
nucleotide sequence, deletion analysis and vaccine development. Nature 306:551-557. 20
26. Melchers, K., Schuhmacher, A., Buhmann, A., Weitzenegger, T., Belin, D., Grau, S., Ehrmann, M. 1999. Membrane 21
topology of CadA homologous P-type ATPase of Helicobacter pylori as determined by expression of phoA fusions in 22
Escherichia coli and the positive inside rule. Res Microbiol 150:507-520. 23
27. Melen, K., Krogh, A., von Heijne, G. 2003. Reliability measures for membrane protein topology prediction algorithms. J 24
Mol Biol 327:735-744. 25
28. Nakayama, Y., Yasui M, Sugahara K, Hayashi M, Unemoto T. 2000. Covalently bound flavin in the NqrB and NqrC 26
subunits of Na(+)-translocating NADH-quinone reductase from Vibrio alginolyticus. FEBS Lett. 474:165-168. 27
29. Pfenninger-Li, X., Albracht, S.P., van Belzen, R., Dimroth, P. 1996. NADH:ubiquinone oxidoreductase of Vibrio alginolyticus: 28
purification, properties, and reconstitution of the Na+ pump. Biochemistry. 35:6233-42. 29
30. Phillips, G. J. 2001. Green fluorescent protein--a bright idea for the study of bacterial protein localization. FEMS Microbiol 30
Lett 204:9-18. 31
31. Rich, P. R., Meunier, B., Ward, F.B.1995. Predicted structure and possible ion motive mechanism of the sodium-linked NADH-32
ubiquinone oxidoreductase of Vibrio alginolyticus. FEBS Letters 375:5-10. 33 32. Roth, R., Hargerhall, C. 2001. Transmembrane orientation and topology of the NADH:quinone oxidoreductase putative 34
quinone binding subuunit NuoH. Biochem Biophys Acta 1504:352-362. 35
33. Saaf, A., Johansson, M., Wallin, E., von Heijne, G. 1999. Divergent evolution of membrane protein topology: the 36
Escherichia coli RnfA and RnfE homologues. Proc Natl Acad Sci USA 96:8540-8544. 37
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

17
34. Scott, M. E., Dossani, Z.Y., Sandkvist, M. 2001. Direct polar secretion of protease from single cells of Vibrio cholerae via 1
the type II secretion pathway. Proc Natl Acad Sci U S A 98:13978-13983. 2
35. Steuber, J. 2001. Na(+) translocation by bacterial NADH:quinone oxidoreductases: an extension to the complex-I family of 3
primary redox pumps. Biochim Biophys Acta. 1505:45-56. 4
36. Turk, K., Puhar, A., Neese, F., Bill, E., Fritz, G., Steuber, J. 2004. NADH oxidation by the Na+-translocating 5
NADH:quinone oxidoreductase from Vibrio cholerae: functional role of the NqrF subunit. J Biol Chem. 279:21349-21355. 6
37. Tusnady, G. E., Simon, I. 2001. The HMMTOP transmembrane topology prediction server. Bioinformatics 17:849-850. 7
38. van Geest, M., Lolkema, J. S. 2000. Membrane topology and insertion of membrane proteins: search for topogenic signals. 8
Microbiol Mol Biol Rev. 64:13-33. 9
39. von Heijne, G., Gavel, Y. 1988. Topogenic signals in integral membrane proteins. Eur J Biochem 174:671-678. 10
40. Waldo, G. S., Standish, B.M., Berendzen, J., Terwilliger, T.C. 1999. Rapid protein-folding assay using green fluorescent 11
protein. Nat Biotechnol. 17:691-695. 12
41. Wikstrom, M. 2004. Cytochrome c oxidase: 25 years of the elusive proton pump. Biochem Biophys Acta 1655:241-247. 13
42. Wikstrom, M. 1998. Proton translocation by bacteriorhodopsin and heme-copper oxidases. Curr Opin Struct Biol 8:480-484. 14
43. Zhou, W., Bertsova, Y.V., Feng, B., Tsatsos, P., Verkhovskaya, M.L., Gennis, R.B., Bogachev, A.V., Barquera, B. 1999. 15
Sequencing and preliminary characterization of the Na+-translocating NADH:ubiquinone oxidoreductase from Vibrio harveyi. 16
Biochemistry. 38:16246-52. 17
18
19
20
21
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

18
FIGURE LEGENDS 1
FIGURE 1. Epifluorescent micrograph of GFP fusions. (A) E. coli pBAD-GFP. (B) wild-type V. cholerae pBAD-GFP. (C) GFP 2
fusion to the complete nqrA gene (GANC) in E. coli TOP10. (D) GANC in wild-type V. cholerae. (E) GFP fusion to nqrD, from N 3
terminal to amino acid 125 (GDN125) in V. cholerae ∆nqr. (F) GFP fusion to the complete nqrD gene plus a linker (GDNC+L) in V. 4
cholerae ∆nqr. The scale bar in panel B applies to all panels. 5
6
FIGURE 2. Membrane topology model of NqrB. Conserved negatively-charged residues located in the membrane spans are 7
represented as black squares. The FMN binding site, T236 and a predicted quinone binding site, G141, are shown. C-terminal fusion 8
sites are indicated in the figure using the code shown in Table 1. 9
10
FIGURE 3. Membrane topology model of NqrC. The FMN binding site, T225, is shown. C-terminal fusion sites are indicated in the 11
figure using the code shown in Table 1. 12
13
FIGURE 4. Membrane topology model of NqrD. Conserved negatively-charged residues located in the membrane spans are 14
represented as black squares. C-terminal fusion sites are indicated in the figure using the code shown in Table 1. 15
16
FIGURE 5. Membrane topology model of NqrE. Conserved negatively-charged residues located in the membrane spans are 17
represented as squares. C-terminal fusion sites are indicated in the figure using the code shown in Table 1. 18
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

19
FIGURE 6. Membrane topology model of NqrF. The binding sites for NADH, the 2Fe-2S center and FAD are indicated. The 1
residues of the weak hydrophobic region predicted as a possible second transmembrane helix are shown as white squares. C-terminal 2
fusion sites are indicated in the figure using the code shown in Table 1. 3
4
FIGURE 7. Coomassie blue-stained and UV-illuminated SDS-PAGE gel of membrane fractions of the alkaline phosphatase fusion to 5
the complete nqrB gene (ABNC) and the alkaline phosphatase fusion to the complete nqrC gene (ACNC) in V. cholerae ∆nqr. The 6
fluorescent bands indicate presence of the covalently bound FMN’s. 30 µg of membrane protein were used in each lane. 7
8
FIGURE 8. Scheme to illustrate relationship between redox cofactor sidedness and the directionality of ion translocation in primary 9
pumps. A. H+-pumping NADH:quinone oxidoreductase (Complex I). B. Na
+-pumping NADH:quinone oxidoreductase (Na
+-NQR). 10
C. cytochrome c oxidase. 11
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

20
TABLE 1. Strains and plasmids used in this study. 1
Strain or plasmid Genotype or description
Source or
reference
STRAIN
E. coli Top10 F- mcrA ∆(mrr-hsdRMS-mcrBC) Φ80lacZ∆M15 ∆lacX74 recA1 araD139∆(ara-
leu)7697 galU galK rpsL (StrR) endA1 nupG
Invitrogen
E. coli LMG194 F- ∆lacX74 galE thi rpsL ∆phoA (Pvu II) ∆ara714 leu::Tn10 Invitrogen
V. cholerae O395N1 Wild-type, Smr (25)
V. cholerae O395N1 ∆nqr Smr, ∆nqr Unpublished results
PLASMID
pBAD-PhoA pBAD22 derivative containing signal sequenceless phoA with a 5’ KpnI site (26)
pBAD-GFP pBAD plasmid with cycle-3 gfp (20)
AANC pBAD-PhoA, nqrA, full-length C-terminal fusion This study
ABN240 pBAD-PhoA, nqrB, C-terminal fusion at residue 240 This study
ABN255 pBAD-PhoA, nqrB, C-terminal fusion at residue 255 This study
ABN377 pBAD-PhoA, nqrB, C-terminal fusion at residue 377 This study
ABNC pBAD-PhoA, nqrB, full-length C-terminal fusion This study
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

21
ACN90 pBAD-PhoA, nqrC, C-terminal fusion at residue 90 This study
ACN180 pBAD-PhoA, nqrC, C-terminal fusion at residue 180 This study
ACNC pBAD-PhoA, nqrC, full-length C-terminal fusion This study
ADN110 pBAD-PhoA, nqrD, C-terminal fusion at residue 110 This study
ADN125 pBAD-PhoA, nqrD, C-terminal fusion at residue 125 This study
ADNC pBAD-PhoA, nqrD, full-length C-terminal fusion This study
AENC pBAD-PhoA, nqrE, full-length C-terminal fusion This study
AFN80 pBAD-PhoA, nqrF, C-terminal fusion at residue 80 This study
AFN160 pBAD-PhoA, nqrF, C-terminal fusion at residue 160 This study
AFNC pBAD-PhoA, nqrF, full-length C-terminal fusion This study
GANC pBAD-GFP, nqrA, full-length C-terminal fusion This study
GCN90 pBAD-GFP, nqrC, C-terminal fusion at residue 90 This study
GCN180 pBAD-GFP, nqrC, C-terminal fusion at residue 180 This study
GCNC pBAD-GFP, nqrC, full-length C-terminal fusion This study
GDN125 pBAD-GFP, nqrD, C-terminal fusion at residue 125 This study
GDN125+L pBAD-GFP, nqrD, C-terminal fusion at residue 125, with 18 amino acid linker This study
GDNC pBAD-GFP, nqrD, full-length C-terminal fusion This study
GDNC+L pBAD-GFP, nqrD, full-length C-terminal fusion, with 18 amino acid linker This study
GENC pBAD-GFP, nqrE, full-length C-terminal fusion This study
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

22
TABLE 2. Summary of membrane topology predictions among seven web-based algorithms for each of the six subunits of 1
Na+-NQR. 2
Subunit TMHMMfix MEMSAT3 TopPred TMPred HMMTOP DASa SOUSI
a Consensus
b
NqrA 0P 1C 0P 0P 0P 0 0 0
NqrB 9C 10C 10C 8P 11C 9 9 9C
NqrC 1P 1P 2C 3C 2P 1 1 1P
NqrD 5C 6C 5P 5P 6C 6 5 5C
NqrE 6C 5C 6P 6P 6P 6 6 6P
NrqF 1P 1C 3C 2P 2P 2 2 2P
Values indicate predicted number of transmembrane helices and the P or C denotes predicted periplasmic or cytoplasmic C-terminus 3
location, respectively. 4
a DAS and SOUSI do not provide prediction of N-terminus location therefore C-terminal location can not be predicted. 5
b Consensus model based on rules from Ikeda et al (18)6
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

23
TABLE 3. Alkaline phosphatase (PhoA) activity of constructed fusions. 1
Fusiona Alkaline
phosphatase
activityb
Locationc
AANC -0.14 C
ABN240 33 C
ABN255 24 C
ABN377 390 P
ABNC -7.4 C
ACN90 20 C
ACN180 0.2 C
ACNC 630 P
ADN110 20 C
ADN125 1.4 C
ADNC 9.5 C
AENC 400 P
AFN80 230 P
AFN160 6.2 C
AFNC 6.0 C
a Descriptions given in Table 1 2
b Units = pNPP hydrolyzed per minute per OD600 of cells 3
c C= cytoplasm, P = periplasm 4
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

E
H
K
G
G
P
E
F
H
H
E
I
DE
L
F
K
K
L
G
M
K W F A
L Y E
A A A T
L F Y
T P G L
V T K
R
S
S H V R D S V D L
K
R
I M I M
V W L
A V F P
A M F
W G M Y
N A G
G
Q
A
I
A
A
L
N
H L
Y
S
G
D
Q
L
A A
I
V
A
G
N
W
H Y
W
L
T
E
M
L
G
G
T
M
S
S
D
A
G
W
G
S
K
M
L
L
G A T Y
F L P
I Y A T
V F I
V G G F
W E V
L
F
C M V R K H E V N
E
G
F F V T
S I L
F A L I
V P P
T L P L
W Q A
A
L
G
I
T F
G
V
V V
A
K
E V
F
G
G T
G
R
N
F
L
N
P
A L A G
R A F
L F F A
Y P A
Q I S G
D L V
W
T
A
A
D
G
Y
S
G A
T
A
L
S
Q
W
A
Q
G
G
A
G
A
L
I
N
N
A
T
G
Q
T
I
T
W
M
D
A
F
I
G
N
I
P
G
S
I G E V
S T L
A L M I
G A A
F I V Y
M G I
A S
W R I I
G G V
M I G M
I L L
S T L F
N V I
G
S D T N A M F N M
P
W H W H
L V L
G G F A
F G M
F F M A
T D P
V
S
A
S
F
T N
S
G
K
W A
Y
G
I
L I
G
V
M
C V
L
I
R
V V
N
P
A
Y
P
E G M M
L A I
L F A N
L F A
P L F D
H V V
V
E
R
N
I
K
R
R
L
A
R
Y
G K
Q
out
in
FMN ligand
ABN377
ABNC
quinone binding site
(putative) ABN240 ABN255
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

T
K
K
I
S
D
N
N
S
A
M
L F V V
I A L S
L V C S
I I V
S A A A
V G L
R
D
K
Q
K
E
N
A
A
L
D
K
Q
S
KILQVAGIEAKG
S
K
Q
I
V
E
L
F
N K
S
I
E
P
R
L
V D
F
N
T
G
D
F
V E
G
D
A
A
N
Y
D Q
R
K
A
A
K
E
A S
E
S
I
K
L
T
A E
Q
D
K
A
K
I
Q
R
R
A
N
V
G
V
V
Y
L
V
K
D
G
D
K
T
S
K
V
I
L
P
V
H
G
N
G
L
W
S
M
M
Y
A
F
V
A
V
E
T
D
G
N
T
V
S
G
L
T
Y
Y
E
Q
G
E
T
P
G
L
G
G
E
V
E
N
P
A
W
R
A
Q
W
V
G
K
K
L
F
D
E
N
H
K
P
A
I
K
I
V
K
G
GAPQGSEHGVDG
L
S
G
A
T
L
T
S
N
G
V
Q
N
T
F D F W
L G D M
G F G P
F L T
K V R D
G G L
N
FMN ligand
A/GCNC
A/GCN90A/GCN180
out
in
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

P
A
L
V
S
K
K
L
E
K
A
S
SM
V L D N
N P I
A L Q V
L G V
C S A
L A V
T T K L E
T A F V
M T L A
V M F V
T A L S
N F F V
S L I
R N H I P N S
V R I
I V Q
M A I
I A S
L V I
VV
D
Q
I
L K A Y L Y D I
S
K
Q
L S V F
V G L
I I T N
C I V
M G R A
E A F
A
M
K S E P I P S F
I
D
G I G N
G L G Y
G F V L
M T V G
F F R E
L L G
S
G
K
L
F
G L E V L P L I S
N
G
G
W
Y
Q P N G
L M L L
A P S A
F F L I
G F M I
W A I
R
T
F
K
P
E
Q
V
E
A
K
E
A/GDN125
ADN110
out
in
A/GDNC
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

K
V
L
L
S
I
Y
H
E
M
S I F I
E N M
A L S F
F L G
M C T F
L A V
S K K V K T S
F G L G
I A V
I V V L
T I S
V P V N
N L V
Y
N
L
V L K P D A L V E
G
V
D
L S F L
N F I T
F I G V
I A A L
V Q I L
E M I L
D R F F P P L
Y N A L G
I F L P
L I T V
N C A I
F G G V
S F M V
Q R D Y S F A E S
V V Y G
F G S
G V G W
M L A
I V A L
A G I
R
E
K
M K Y S D V P P
G
L
R
G L G I
T F I
T A G L
M A L
G F M
S F S
G
V
Q
L
out
in
A/GENC
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

F
I
I
T
S
M
G V V M
F T L
I I L
A L V
L V I
L F A
K
S
K
L
V
P
T
G
D
I
T
I
S
I
N
G
D
P
E
K
A
I
V
T
Q
P
G
G
K
L
L
T
A L
A
G
A
G
V
F
V
S
S
A
C
G
G
G
G
S
C
G
Q
C
R
V
K
I
K
S
G
G
G
D
I L
P
T
E
L
D
H
I
S
K
G
E
A
R
E
G
E
R
L
A
C
Q
V
A
V
K
A
D
M
D
L
E L
P
E
E
I
F
G
V
K
K
W
E
C
T
V
I
S
N
D
N
K
A
T
F
I
K
E
L
K
L
A
I P
D
G
E
S
V
P
F
R
A
G
G
Y
I
Q
I
E
A
P
A
H
H
V
K
Y
A
D
F
D
V
P
E K
Y
R
G
D
W
D
K
F
N
L
F
R
Y
E
S
K
V
D
E
P
I
I
R
A
Y
S
M
A
N
Y
P E
E
F
G
I
I
M
L
N
V
R
I
A
T
P
P
P
N
N
P
N
V
P
P
G
Q
M
S
S
Y
I
W S
L
K
A
G
D
K
C
T
I
S
G
P
F
G
E
F
F
A
K
D
T
D
A
E
M
V
F
I
G
G
G A
G
M
A
P
M
R
S
H
I
F
D
Q
L
K
R
L
K
S
K
R
K
M
S
Y
W
Y
G
A
R
S
K R
E
M
F
Y
V
E
D
F
D
G
L
A
A
E
N
D
N
F
V
W
H
C
A
L
S
D
P
Q
P
E
D N
W
T
G
Y
T
G
F
I
H
N
V
L
Y
E
N
Y
L
K
D
H
E
A
P
E
D
C
E
Y
Y
M
C G
P
P
M
M
N
A
A
V
I
N
M
L
K
N
L
G
V
E
E
E
N
I
L
L
D
D
F
G
G
low probability
helix
2Fe2S ligands
NADH binding site
FAD ligands
AFNC
AFN160
AFN80
out
in
ACCEPTED
on April 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from


![Vibrio cholerae use pili and flagella synergistically to ...wonglab.seas.ucla.edu/pdf/2014 Nat Commun [Utada, Wong] Vibrio... · Vibrio cholerae use pili and flagella synergistically](https://static.fdocuments.in/doc/165x107/5afa9d177f8b9a32348e07cc/vibrio-cholerae-use-pili-and-flagella-synergistically-to-nat-commun-utada.jpg)















