
Use ofBacillus subtilis Minicells to Demonstrate …iai.asm.org/content/12/5/1189.full.pdf ·...
6
INFECTION AND IMMUNITY, Nov. 1975, p. 1189-1194 Copyright C) 1975 American Society for Microbiology Vol. 12, No. 5 Printed in USA. Use of Bacillus subtilis Minicells to Demonstrate an Antigenic Relationship Between the Poles and Lateral Cylindrical Regions of Rod-Shaped Cells SHEILA I. COYNE' AND NEIL H. MENDELSON* Department of Microbiology and Medical Technology and Graduate Committee on Genetics, University of Arizona, Tucson, Arizona 85721 Received for publication 19 June 1975 Purified minicells produced by Bacillus subtilis div IV-B1 mutants were used to immunize rabbits. Immune serum was obtained that agglutinated minicells and was able to form precipitin lines when reacted with minicell antigens in double-diffusion or immunoelectrophoretic procedures. Antiminicell serum ag- glutinated rod-shaped B. subtilis cells to long filaments produced by growth of a cell division-defective mutant at restrictive temperature. These findings indi- cate that minicells are immunogens capable of eliciting the production of anti- bodies that cross-react with the lateral, cylindrical regions of B. subtilis rods. It appears, therefore, that poles share a common antigen(s) with cylindrical re- gions of the cell. Rod-shaped bacteria consist of two geometri- cally distinct regions: cylindrical sides and hem- ispherical poles. During the growth cycle, cells elongate by the extension of the cylindrical re- gion and then compartmentalize into two daughter cells by the construction of a septum. Division is completed by the differentiation of the septum into two poles. Each daughter cell consists of two poles, one of which is a genera- tion older than the other, connected by a cylin- drical region. Septum formation, and conse- quently pole production, is known to be under specific genetic control distinct from that which regulates cell growth by cylindrical length ex- tension (7). It is of interest to determine whether the composition of the cell surface in the cylindrical portions of the cell is identical to that found in the poles. It has been demon- strated that wall regions isolated from the poles ofBacillus subtilis are more resistant to autoly- tic degradation than are wall regions from the cylindrical portion of the cell (3, 5). The reasons for this resistance are not clear since it has been reported that cell surface turnover occurs in cell pole regions, as well as in cylindrical portions of the cell (4). We have reported the isolation of a minicell- producing mutant of B. subtilis and have de- scribed the properties of purified minicells (8, 9). Minicells consist entirely of cell pole surface and, thus, offer a system for investigating differ- ences between poles and sides of rod-shaped bacteria (9). In this paper we report that mini- 1 Present address: Department of Microbiology, Univer- sity of Alabama in Birmingham, Birmingham, Ala. 35294. cells share a common antigen(s) found on cell cylindrical surfaces. MATERIALS AND METHODS Bacteria. Strain of B. subtilis and Escherichia coli used in these experiments are listed in Table 1. Media. Minimal medium consisted of: 0.0151 M (NH4)2S04, 0.0613 M K2HPO4-3H20, 0.0441 M KH2PO4, 0.0034 M sodium citrate 2H2O, 0.00081 M MgSO4-7H2O, and 0.0277 M glucose. Sterile supple- ments (20 ,ug/ml) were added as follows: thymine, isoleucine, and methionine for growth of CU403 cul- tures; thymine and methionine for growth of CU403 div IV-Bl and CU403 tms-12 cultures; and leucine, methionine, and adenine for growth of BC-101 cul- tures. Trypticase soy broth (BBL, Cockeysville, Md.; 30 g/liter) was supplemented with 20 ,ug of thymine per ml. Purification of minicells. Five 1-liter batches of B. subtilis CU403 divIV-Bl were grown overnight in minimal liquid medium at 30 C with shaking. Har- vesting and purification of minicells have been previ- ously described (8). Production of antiserum. Preimmunization se- rum was obtained by bleeding two rabbits from the marginal vein of the ear. Purified minicells sus- pended in 3 ml of saline were mixed with an equal volume of Freund incomplete adjuvant and injected subcutaneously into several sites on the back of each rabbit twice weekly for 3 weeks. Two weeks after the final injection, the rabbits were bled by cardiac punc- ture. The serum was collected after the blood clotted at room temperature for 1 h. The serum was de- canted and stored frozen until use. Before use, anti- serum was heated at 56 C for 30 min. All of the results reported in this paper were obtained by us- ing preimmune and antiminicell serum from one of the two rabbits. 1189 on September 17, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Use ofBacillus subtilis Minicells to Demonstrate …iai.asm.org/content/12/5/1189.full.pdf ·...

INFECTION AND IMMUNITY, Nov. 1975, p. 1189-1194Copyright C) 1975 American Society for Microbiology
Vol. 12, No. 5Printed in USA.
Use of Bacillus subtilis Minicells to Demonstrate an
Antigenic Relationship Between the Poles and LateralCylindrical Regions of Rod-Shaped Cells
SHEILA I. COYNE' AND NEIL H. MENDELSON*
Department of Microbiology and Medical Technology and Graduate Committee on Genetics, University of
Arizona, Tucson, Arizona 85721
Received for publication 19 June 1975
Purified minicells produced by Bacillus subtilis div IV-B1 mutants were usedto immunize rabbits. Immune serum was obtained that agglutinated minicellsand was able to form precipitin lines when reacted with minicell antigens indouble-diffusion or immunoelectrophoretic procedures. Antiminicell serum ag-
glutinated rod-shaped B. subtilis cells to long filaments produced by growth of acell division-defective mutant at restrictive temperature. These findings indi-cate that minicells are immunogens capable of eliciting the production of anti-bodies that cross-react with the lateral, cylindrical regions ofB. subtilis rods. Itappears, therefore, that poles share a common antigen(s) with cylindrical re-
gions of the cell.
Rod-shaped bacteria consist of two geometri-cally distinct regions: cylindrical sides and hem-ispherical poles. During the growth cycle, cellselongate by the extension of the cylindrical re-gion and then compartmentalize into twodaughter cells by the construction of a septum.Division is completed by the differentiation ofthe septum into two poles. Each daughter cellconsists of two poles, one of which is a genera-tion older than the other, connected by a cylin-drical region. Septum formation, and conse-quently pole production, is known to be underspecific genetic control distinct from that whichregulates cell growth by cylindrical length ex-tension (7). It is of interest to determinewhether the composition of the cell surface inthe cylindrical portions of the cell is identical tothat found in the poles. It has been demon-strated that wall regions isolated from the polesofBacillus subtilis are more resistant to autoly-tic degradation than are wall regions from thecylindrical portion of the cell (3, 5). The reasonsfor this resistance are not clear since it has beenreported that cell surface turnover occurs in cellpole regions, as well as in cylindrical portions ofthe cell (4).We have reported the isolation of a minicell-
producing mutant of B. subtilis and have de-scribed the properties of purified minicells (8,9). Minicells consist entirely of cell pole surfaceand, thus, offer a system for investigating differ-ences between poles and sides of rod-shapedbacteria (9). In this paper we report that mini-
1 Present address: Department of Microbiology, Univer-sity of Alabama in Birmingham, Birmingham, Ala. 35294.
cells share a common antigen(s) found on cellcylindrical surfaces.
MATERIALS AND METHODSBacteria. Strain of B. subtilis and Escherichia
coli used in these experiments are listed in Table 1.Media. Minimal medium consisted of: 0.0151 M
(NH4)2S04, 0.0613 M K2HPO4-3H20, 0.0441 MKH2PO4, 0.0034 M sodium citrate 2H2O, 0.00081 MMgSO4-7H2O, and 0.0277 M glucose. Sterile supple-ments (20 ,ug/ml) were added as follows: thymine,isoleucine, and methionine for growth of CU403 cul-tures; thymine and methionine for growth of CU403div IV-Bl and CU403 tms-12 cultures; and leucine,methionine, and adenine for growth of BC-101 cul-tures. Trypticase soy broth (BBL, Cockeysville,Md.; 30 g/liter) was supplemented with 20 ,ug ofthymine per ml.
Purification of minicells. Five 1-liter batches ofB. subtilis CU403 divIV-Bl were grown overnight inminimal liquid medium at 30 C with shaking. Har-vesting and purification of minicells have been previ-ously described (8).
Production of antiserum. Preimmunization se-rum was obtained by bleeding two rabbits from themarginal vein of the ear. Purified minicells sus-pended in 3 ml of saline were mixed with an equalvolume of Freund incomplete adjuvant and injectedsubcutaneously into several sites on the back of eachrabbit twice weekly for 3 weeks. Two weeks after thefinal injection, the rabbits were bled by cardiac punc-ture. The serum was collected after the blood clottedat room temperature for 1 h. The serum was de-canted and stored frozen until use. Before use, anti-serum was heated at 56 C for 30 min. All of theresults reported in this paper were obtained by us-ing preimmune and antiminicell serum from one ofthe two rabbits.
1189
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1190 COYNE AND MENDELSON
TABLE 1. Strains ofB. subtilis and E. coli used in this study
GenotypeStrain Other Origin
Auxotrophic markers markers
B. subtilisCU403 thyA, thyB, ilvD, metB S. Zahler, Cornell Univ.CU403 div IV-B1 thyA, thyB, metB div IV-B1 Construction previously described
(9)BC-101 leu-8, metB, purA16 tms-12 (1)CU403 tms-12 thyA, thyB, metB tms-12 Congression. BC-101 used as donor
and CU403 as recipient; ilv+ trans-formants selected
E. coli S/6/5 Resistance to H. Bernstein, Univ. ofArizona Med-T6, T5 ical School
Adsorption of antiserum to lateral walls. Trypti-case soy broth (100 ml) was inoculated with a loopfulof B. subtilis CU403 tms-12 from a Trypticase soyagar plate grown overnight at 30 C. The liquid cul-ture was incubated at 30 C with shaking for severalhours. Upon reaching mid-logarithmic phase, theculture was transferred to 45 C and growth contin-ued for approximately 1 h, during which time thecells grew into long filaments, as determined byexamination under the phase-contrast microscope.The culture was pelleted by centrifugation at 5,000rpm for 10 min at 4 C and suspended in sodiumphosphate buffer, and flagella were then removedwith a Waring blender by the method of De-Pamphilis and Adler (2). The culture was again pel-leted by centrifugation and resuspended in l/lo vol-ume of buffer. A few drops of the cell suspensionwere mixed with an equal volume of undiluted anti-serum in two small test tubes and incubated for 20min at 4 C. At the same time, in other tubes a fewdrops of the cell suspension were mixed with serumcollected from the same rabbit before injection withminicells.
Meanwhile, cultures of CU403 and E. coli S/6/5were grown in Trypticase soy broth at 30 C withshaking until late logarithmic phase. Flagella wereremoved as above. A few drops of the CU403 wild-type cell suspension were added to one tube ofCU403 tms-12 filaments plus antiserum and to onetube of CU403 tms-12 filaments plus preimmunerabbit serum. Likewise, a few drops of the E. colicell suspension were added to other tubes containingCU403 tms-12 filaments plus antiserum or preim-mune rabbit serum. After incubation for 20 min, adrop of each of the cell-serum mixtures was placedonto slides, covered and observed under the phase-contrast microscope for agglutination.
RESULTSFlagella were sheared from the surface of
cells used in agglutination studies to eliminatethe possibility of false positive tests due to thepresence of antibody against flagellin in theantiminicell antisera. The presence ofsuch anti-bodies is unlikely, however, even though mini-
cells in producing cultures are flagellated. Thisis because of the purification procedure used toprepare minicells as antigens. Not only is motil-ity lost after the sonic treatment step of theminicell purification procedure, but also wehave never observed flagella in electron micro-graphs of purified minicells. Nevertheless, thisprecaution was taken, and for uniformityCU403 wild-type and E. coli S/6/5 cells, as wellas CU403 div IV-Bl cells, were also stripped offlagella in a Waring blender before use in agglu-tination tests.A CU403 tms-12 liquid culture (Trypticase
soy broth) was transferred from 30 to 45 C andallowed to grow into long filaments. After theconversion was complete, the cells were har-vested by centrifugation, the flagella were re-moved, and the cells were resuspended inbuffer. Antiminicell serum was added to theflagellaless filaments, and the mixture was in-cubated at 4 C for approximately 20 min. Incu-bation was carried out at this temperature tominimize autolysis of the filaments and tolessen the likelihood of flagella reorganizationduring the adsorption period. FlagellalessCU403 wild-type cells were then added to themixture. As shown in Fig. 1, the short wild-type cells become attached along the length ofthe long tms-12 filaments. Short rods can beseen lying lengthwise along both sides of andalso across the filaments. Attachment of shortrods to the filaments is not restricted to polarsites. Agglutination of the filaments, as well asthe CU403 wild-type cells with themselves, wasalso observed.The following controls were performed.
Preimmune rabbit serum was applied to flag-ellaless tms-12 filaments to which flagellalesswild-type cells were added in a manner analo-gous to that described above with antiminicellserum. As shown in Fig. 2, some small wild-type cells have collided with the filaments, and
INFECT. IMMUN.
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CELL POLE AND LATERAL CYLINDRICAL ANTIGENS 1191
i
FIG. 1. Adsorption of antiminicell serum to lateral walls of CU403 tms-12 filaments. CU403 tms-12 cellswere incubated at 45 C to promote growth into filaments. Flagella were removed in a Waring blender.Antiminicell serum was allowed to adsorb to filaments, and then CU403 tms-12+ rods (after removal offlagella) were added to the filaments plus antiserum. Bar represents 10 ,um.
some have been trapped by the filaments.These cells are not bound together as those heldby antiminicell serum. In the normal serum
controls, short rods are not found attached toboth sides of the same filaments, nor do they lie
across the filaments. A few free short rods arealso evident in Fig. 2. A second control con-sisted of adding E. coli S/6/5 cells to incubationmixtures of flagella-free filaments plus anti-minicell serum. Figure 3 shows that the short
VOL. 12, 1975
.If
6a,.- .1
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
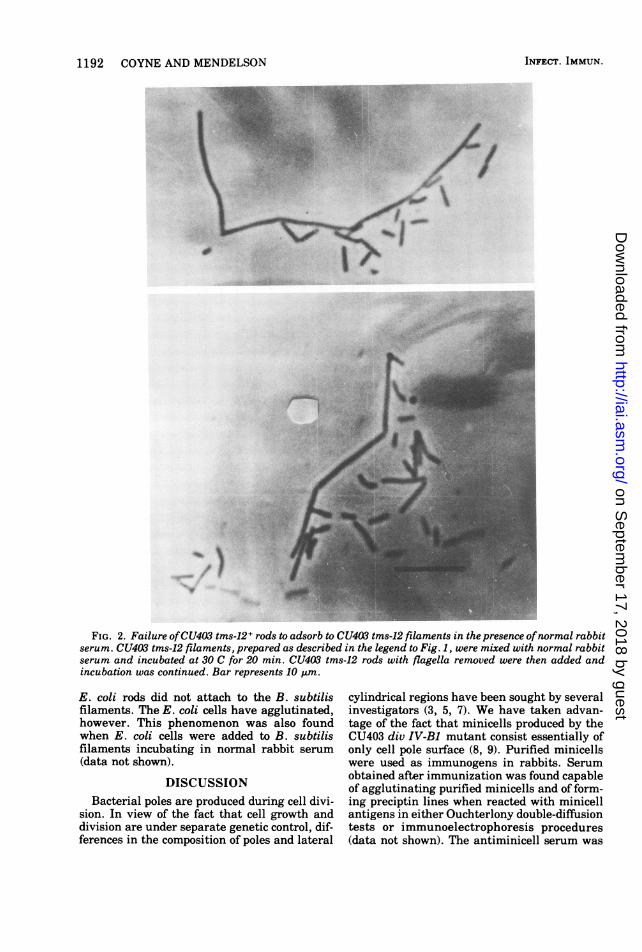
1192 COYNE AND MENDELSON
/k
l
)
FIG. 2. Failure ofCU403 tms-12+ rods to adsorb to CU403 tms-12 filaments in the presence ofnormal rabbitserum. CU403 tms-12 filaments, prepared as described in the legend to Fig. 1, were mixed with normal rabbitserum and incubated at 30 C for 20 min. CU403 tms-12 rods with flagella removed were then added andincubation was continued. Bar represents 10 Am.
E. coli rods did not attach to the B. subtilisfilaments. The E. coli cells have agglutinated,however. This phenomenon was also foundwhen E. coli cells were added to B. subtilisfilaments incubating in normal rabbit serum(data not shown).
DISCUSSIONBacterial poles are produced during cell divi-
sion. In view of the fact that cell growth anddivision are under separate genetic control, dif-ferences in the composition of poles and lateral
cylindrical regions have been sought by severalinvestigators (3, 5, 7). We have taken advan-tage of the fact that minicells produced by theCU403 div IV-B1 mutant consist essentially ofonly cell pole surface (8, 9). Purified minicellswere used as immunogens in rabbits. Serumobtained after immunization was found capableof agglutinating purified minicells and of form-ing preciptin lines when reacted with minicellantigens in either Ouchterlony double-diffusiontests or immunoelectrophoresis procedures(data not shown). The antiminicell serum was
INFECT. IMMUN.
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CELL POLE AND LATERAL CYLINDRICAL ANTIGENS 1193
7q/ ~ /'odI*k
-_A_0Ioi
-
q 'I
FIG. 3. Failure ofE. coli Sf615 to adsorb to CU403 tms-12 filaments in the presence ofantiminicell serum.E. coli cells grown at30 C were added to filaments plus antiserum in the usual manner. Bar represents 10 ,um.
then used to determine whether the lateral cy-lindrical surfaces of the cell and the cell polesshare common antigens.To carry out such determinations we have
made use of a temperature-sensitive cell divi-sion initiation mutant (tms-12) that grows intolong filaments when incubated at 45 C (1). Thismutant was chosen because the ratio of cylindri-cal region to poles is much increased in the longfilaments over that found in typical B. subtilisrods and it is, therefore, easy to determinewhether the cylindrical regions cross-react withantiminicell serum. Simple agglutination tests,for example, clearly demonstrate that wild-type
B. subtilis cells adhere along their cylindricalregions to the long cylindrical regions of themutant filaments when antiminicell antibodyis present. Preimmune rabbit serum does notfacilitate such adhesions. Furthermore, an un-related antigen (E. coli) is not bound to thefilaments in the presence of antiminicell se-rum. Thus, the reaction of wild-type B. subtilisrods, with the cylindrical regions of B. subtilisfilaments facilitated by antiminicell serum, isspecific and indicative of a shared antigen(s) onpole and cylindrical cell surface regions. Simi-lar findings have been reported in the Salmo-nella minicell system (10).
VOL. 12, 1975
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

1194 COYNE AND MENDELSON
One additional feature of the minicell immu-nogen system is worthy of mention. Minicellsare unique not only because they consist of aspecific cell compartment of pole material, butalso because they lack the cell genome. As aresult, they are unable to grow. In this regardthey resemble other anucleate bacterial cellsproduced by certain temperature-sensitiveDNA- mutants (6). Minicells are, however, eas-ier to separate from the nucleated cells in theculture than are anucleate cells produced by ts-DNA- mutants since the latter produce anu-cleate cells of normal size. If, indeed, minicellsshare an adequate antigenic similarity to thenucleated cells that produce them, then puri-fied minicells might prove to be of use in immu-nization since the inability to grow precludesinfection of the host by the immunogen. Itmight be of value, therefore, to search for mini-cell-producing mutants in pathogenic species inwhich adequate immunogens are unavailable.There is no reason to believe that such mutantscannot be found in all bacteria.
ACKNOWLEDGMENTSS.I.C. was the recipient of HEA Title IX-B Fellowship
(formerly NDEA Title IV). This work was supported byPublic Health Service Career Development award K04 GM70555 and research grant 1 R01 GM 18735, both from theNational Institute of General Medical Sciences, to N.H.M.We wish to acknowledge the advice and technical assist-
ance of A. Paquet, Jr., in matters concerning immunologyprocedures.
LITERATURE CITED
1. Copeland, J. C., and J. Marmur. 1968. Identification ofconserved genetic functions in Bacillus by use of tem-perature-sensitive mutants. Bacteriol. Rev. 32:302-312.
2. DePamphilis, M. L., and J. Adler. 1971. Purification ofintact flagella from Escherichia coli and Bacillus sub-tilis. J. Bacteriol. 105:376-383.
3. Fan, D. P., and B. E. Beckman. 1973. Structural differ-ence between walls from hemispherical caps and par-tial septa of Bacillus subtilis. J. Bacteriol. 114:790-797.
4. Fan, D. P., B. E. Beckman, and M. M. Beckman. 1974.Cell wall turnover at the hemispherical caps ofBacil-lus subtilis. J. Bacteriol. 117:1330-1334.
5. Fan, D. P., M. C. Pelvit, and W. P. Cunningham. 1972.Structural difference between walls from ends andsides of the rod-shaped bacterium Bacillus subtilis. J.Bacteriol. 109:1266-1272.
6. Hirota, Y., F. Jacob, A. Ryter, G. Buttin, and T. Nakai.1968. On the process of cellular division in Esche-richia coli. I. Asymmetrical cell division and produc-tion of deoxyribonucleic acid-less bacteria. J. Mol.Biol. 35:175-192.
7. Mendelson, N. H., and R. M. Cole. 1972. Genetic regula-tion of cell division initiation in Bacillus subtilis. J.Bacteriol. 112:994-1003.
8. Mendelson, N. H., J. N. Reeve, and R. M. Cole. 1974.Physiological studies ofBacillus subtilis minicells. J.Bacteriol. 117:1312-1319.
9. Reeve, J. N., N. H. Mendelson, S. I. Coyne, L. L.Hallock, and R. M. Cole. 1973. Minicells of Bacillussubtilis. J. Bacteriol. 114:860-873.
10. Tankersley, W. G., and J. M. Woodward. 1974. Induc-tion and isolation of a minicell-producing strain ofSalmonella typhimurium. Proc. Soc. Exp. Biol. Med.145:802-805.
INFECT. IMMUN.
on Septem
ber 17, 2018 by guesthttp://iai.asm
.org/D
ownloaded from




![Escherichia - PNAS · ,-O-Ic FIG. 3. Autoradiograms of proteins synthesized in E. coli mini-cells. (A) [85S]Methionine-labeled proteinsfromtrpR' minicells con-taining pAS621 (lane](https://static.fdocuments.in/doc/165x107/5f1351b6bb0c4115e926b9cf/escherichia-pnas-o-ic-fig-3-autoradiograms-of-proteins-synthesized-in-e-coli.jpg)







![Estimation of Resected Liver Regions Using a Tumor ... · (MeVis Medical Solutions AG, Bremen, Germany)[3] , OVA ... B. Preim, A. Schenk, et al., “Analysis of vasculature for liver](https://static.fdocuments.in/doc/165x107/5b9f447d09d3f2fc778d2e02/estimation-of-resected-liver-regions-using-a-tumor-mevis-medical-solutions.jpg)


![6 DAYS ARIRANG KOREA (S.I.C INCHEON IN/OUT) · 6d5n arirang korea [6cju] audlt & child with room child no bed s. supplement tips per head tour fare usd625 per person usd520 per child](https://static.fdocuments.in/doc/165x107/5b66ca3b7f8b9a87148d7699/6-days-arirang-korea-sic-incheon-inout-6d5n-arirang-korea-6cju-audlt.jpg)

![, Pierre Ermesƒ Rob van der Rijt , Harrie van den … · bulletin board pin-like shape with a small ag on top of it. ... 1 INTRODUCTION AND MEDICAL BACKGROUND ... Preim et al. [15],](https://static.fdocuments.in/doc/165x107/5b9f447d09d3f2fc778d2e26/-pierre-ermesf-rob-van-der-rijt-harrie-van-den-bulletin-board-pin-like-shape.jpg)
