
UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS ...€¦ · meu projeto e, por me permitir...
137
UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA ALINE KELLY DE AQUINO LIMA CIPRIANO A INTERAÇÃO DO CAJUEIRO (Anacardium occidentale L.) COM O FUNGO Lasiodiplodia theobromae REPROGRAMA A EXPRESSÃO DE PROTEÍNAS NO CAULE, SÍTIO DE INFECÇÃO DO PATÓGENO FORTALEZA - CEARÁ 2014
Transcript of UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS ...€¦ · meu projeto e, por me permitir...

UNIVERSIDADE FEDERAL DO CEARÁ
CENTRO DE CIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
ALINE KELLY DE AQUINO LIMA CIPRIANO
A INTERAÇÃO DO CAJUEIRO (Anacardium occidentale L.) COM O FUNGO
Lasiodiplodia theobromae REPROGRAMA A EXPRESSÃO DE PROTEÍNAS
NO CAULE, SÍTIO DE INFECÇÃO DO PATÓGENO
FORTALEZA - CEARÁ
2014

ALINE KELLY DE AQUINO LIMA CIPRIANO
A interação do cajueiro (Anacardium occidentale L.) com o fungo Lasiodiplodia theobromae
reprograma a expressão de proteínas no caule, sítio de infecção do patógeno
Orientador: Prof. Dr. José Tadeu Abreu de Oliveira
Coorientadora: Dra. Darcy Mayra Furtado Gondim
FORTALEZA - CEARÁ
2014
Dissertação submetida à
Coordenação do Programa de Pós-
Graduação em Bioquímica da
Universidade Federal do Ceará,
como requisito parcial para
obtenção do grau de Mestre em
Bioquímica.


Esta dissertação de Mestrado foi apresentada ao Programa de Pós-Graduação em
Bioquímica da Universidade Federal do Ceará como requisito parcial para obtenção do grau
de Mestre em Bioquímica, outorgado pela Universidade Federal do Ceará, e encontrar-se-á à
disposição na Biblioteca Central da referida Universidade.
A transcrição ou utilização de qualquer trecho deste trabalho é permitida, desde que
seja feito de acordo com as normas de ética científica.
____________________________________
Aline Kelly de Aquino Lima Cipriano
Dissertação aprovada em: 21 de Março de 2014.
BANCA EXAMINADORA
...................................................
Dr. José Tadeu Abreu de Oliveira
Universidade Federal do Ceará
(Orientador)
.......................................................
Dr. José Emilson Cardoso
Embrapa Agroindústria Tropical
(Examinador)
.......................................................
Frederico Bruno Mendes Batista Moreno
Universidade de Fortaleza
(Examinador)

AGRADECIMENTOS
Ao meu bom Deus pelo zelo com que conduz a minha vida fornecendo os subsídios
que são necessários para alcançar meus propósitos: a força diante das fraquezas, a solução
diante das complicações e a saúde diante da possível fragilidade a que nós humanos somos
sujeitos. Enfim, por me fornecer na medida certa a “essência” que eu preciso para seguir em
frente.
Ao meu orientador, Prof. José Tadeu Abreu de Oliveira, que de forma muito especial
aceitou assumir e conduzir esta orientação. Pela confiança, cuidados e conselhos inerentes ao
meu projeto e, por me permitir compartilhar a certeza de que sua contribuição científica, na
minha formação, também foi graça de Deus na minha vida.
A minha coorientadora, Dra. Darcy Mayra, por todos os conselhos e ensinamentos
científicos. Por ter dividido as alegrias e aflições inerentes a este protejo, pelo incentivo na
condução dessa pesquisa.
Ao Dr. José Emilson pela pronta aceitação em participar da avaliação deste trabalho e
pelo entusiasmo sempre demonstrado nesse projeto, inicialmente conduzido pela Dra. Darcy
Mayra.
Ao Laboratório de Análises Proteômicas, da Universidade de Fortaleza, coordenado
pela professora Dra. Ana Cristina Moreira, pelas análises de espectrometria de massas. Em
especial ao Dr. Frederico Moreno pelo apoio durante os experimentos e pela aceitação em
participar da avaliação desse trabalho.
A todos que compõem o Laboratório de Fitopatologia da Embrapa Agroindústria
Tropical. Em especial aos estudantes Joilson e Glauber pela ajuda nos experimentos de
inoculação.
Ao Professor Francisco de Assis Paiva Campos e a todos do laboratório de Biologia
Molecular de Plantas, em especial a Manu, Fabiano e Camila.
Ao Laboratório de Fisiologia Animal- UFC, coordenado pelo Prof. Dr. Arlindo Moura
e, de forma muito especial, ao Jorge pelo apoio durante a análise proteômica.

Ao Laboratório de Toxinas Vegetais, coordenado pela Prof. Dra. Ilka Vasconcelos,
pelo apoio prestado e aos estudantes Helen, Paulo, Lucas, e, de forma mais que especial à
amiga Clídia.
À Iara Flávia, minha grande amiga e irmã de coração. Pela generosidade sempre
mostrada na nossa amizade e, por dividir comigo, a sobrecarga e aflições dos últimos seis
anos. Pela palavra amiga, confiança, respeito e, acima de tudo, por me aceitar com minhas
qualidades e limitações.
À Hudson Moura, meu grande parceiro de bancada e amigo. A quem sou
extremamente grata pelo abraço apertado de todos os dias, as discussões e entusiasmo
científico inerente a nossa linha de pesquisa. Por compartilhar comigo a palavra de força nos
momentos de maus resultados.
À grande Ana Luiza, pelo sorriso e energia positiva contagiante. Por ser essa pessoa
maravilhosa, indescritível e psicodélica.
À Anna Lídia, amiga linda. Pelas palavras consoladoras diante das angústias. Por
confiar e aceitar dividir comigo alguns momentos, que acredite, foram muito valiosos para
meu crescimento pessoal.
Ao querido Rodolpho Guedes (Rodolphinho). Pelo carinho e momentos
descontraídos que foram muito importantes para acalantar as preocupações científicas. Pelo
trabalho de designer na elaboração das figuras necessárias no meu artigo e, por ter sido, uma
pessoal muito especial durante esses dois anos.
Ao amigo Handerson (o grande), por ser essa pessoa tão gentil e atenciosa.
Aos queridos e demais colegas do Laboratório de Proteínas Vegetais de Defesa:
Thiago, Pedro, Raissa, Felipe, Emanoel e Larissa. Pelo acolhimento especial, excelente
convivência e carinho sempre demonstrado. Por terem se tornado a minha família de todos os
dias. E em particular, aos amigos Fredy Davi, por estar sempre solícito em compartilhar
opiniões e resultados científicos e, a Juliana Rodrigues (minha IC favorita), por todo o
carinho.
As minhas amigas, Joselena Mendonça e Janaína Daniela, por serem um grande
presente de Deus na minha vida.

Meus mais sinceros agradecimentos, ao meu melhor alicerce: minha família. De
maneira, extremamente especial, aos meus lindos e abençoados pais: Ivo Cipriano e Itacira
Nunes. A vocês eu não consigo expressar em palavras o sentimento de gratidão, por todo o
amor incondicional e dedicação que foram essenciais na minha formação pessoal e
profissional. Por encontrar, no apoio de vocês, a força para renovar minhas fraquezas.
A minha linda irmã Anielly Cristina e ao pequeno Yan Samuel, quem são bênçãos de
Deus na minha vida.
Aos meus tios e tias, que honrosamente são grandes exemplos de esforço e vitória.
A meus queridos avós de sangue e de coração. E, em particular, a minha querida
vozinha Noêmia Nunes.
Por fim, agradeço profundamente ao meu amado Ricardo Gonçalves, pelo exemplo de
dedicação e incentivo na iniciação científica. Por, todos os dias, me fazer experimentar da
alegria da cumplicidade e amor necessário para enfrentarmos as dificuldades.
A todos que direta e indiretamente contribuíram para a execução deste trabalho.

AGRADECIMENTOS INSTITUCIONAIS
Este trabalho foi realizado graças ao auxílio das seguintes Instituições:
Departamento de Bioquímica e Biologia Molecular do Centro de Ciências da Universidade
Federal do Ceará (DBBM-UFC).
Coordenação de Aperfeiçoamento de Pessoal e de Ensino Superior (CAPES), pela bolsa de
Pós-Graduação concedida ao autor, através de convênio com o Programa de Pós-Graduação
em Bioquímica do Departamento de Bioquímica e Biologia Molecular do Centro de Ciências
da Universidade Federal do Ceará.
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), através de
convênio com o Programa de Pós-Graduação em Bioquímica do Departamento de Bioquímica
e Biologia Molecular do Centro de Ciências da Universidade Federal do Ceará.
Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP),
através de convênio com o programa de Pós-Graduação em Bioquímica do Departamento de
Bioquímica e Biologia Molecular do Centro de Ciências da Universidade Federal do Ceará.

RESUMO
No Brasil, a indústria do caju é uma das principais fontes de renda e trabalho no campo e
representa a maior parcela da economia na região nordeste, que concentra 94% da produção,
destacando-se os estados do Ceará, Piauí e Rio Grande do Norte, respectivamente. Neste
contexto, para otimizar o setor produtivo, estratégias de melhoramento genético de cultivares
têm focado na seleção e desenvolvimento de clones anãos. Entretanto, essa prática tem
contribuído para diminuição da variabilidade genética e, consequentemente, para maior
vulnerabilidade ao ataque de patógenos. A resinose, causada pelo fungo Lasiodiplodia
theobromae (Pat.) Griff & Maubl. é considerada a principal doença do cajueiro nas condições
semiáridas. Métodos eficientes de controle da doença ainda não foram estabelecidos. Baseado
na ausência de dados publicados com relação às respostas bioquímicas e fisiológicas do
cajueiro infectado com L. theobromae, associado ao fato de que as plantas, para se
defenderem do ataque de patógenos, acionam/alteram vias metabólicas controladas por
diversas proteínas, o estudo da identidade dessas proteínas fornecem informações sobre os
mecanismos relacionados à interação de compatibilidade/incompatibilidade entre o cajueiro e
L. theobromae. Dessa forma, nesse estudo, foi realizada análise proteômica diferencial de
caules de cajueiro (Anacardium occidentale), clone BRS 226 (resistente), mantido em
condições controladas, em tempos iniciais pós-infecção com o L. theobromae, assim como, de
plantas de cajueiro resultantes da polinização aberta do BRS 226, classificadas como
resistentes e suscetíveis à resinose, em condições de campo, onde há alta pressão do patógeno.
Proteínas diferencialmente expressas foram identificadas por eletroforese bidimensional (2D-
PAGE) combinada com espectrometria de massas ESI-Q-TOF MS/MS. Plantas de cajueiro
infectadas com L. theobromae acionam respostas fisiológicas associadas à reprogramação da
expressão de um total de 73 proteínas, nos cenários investigados. Destas, 36 foram
identificadas no clone BRS 226, artificialmente inoculado, e 37 nas plantas de cajueiro,
cultivadas em condições de campo. Portanto, um número equivalente de proteínas é
responsivo dentro dos cenários analisados e compartilham funções celulares em rotas
metabólicas e de produção de energia, estresse/defesa, sinalização celular e
enovelamento/metabolismo de proteínas. Proteínas responsivas a hormônios e envolvidas com
estrutura celular foram diferencialmente expressas somente em plantas crescidas em condição
de campo, enquanto proteínas de transporte foram reprogramadas no clone BRS 226. Plantas
de cajueiro, cultivadas em campo, e inoculadas do clone BRS 226 compartilharam a
expressão reprogramada de 6 proteínas idênticas, dentre as quais, proteínas 14-3-3 e anexinas,
envolvidas com a sinalização celular, foram influenciadas ao mesmo tempo, aparentemente,
pelos estresses abiótico e biótico. A reprogramação de funções celulares comuns somado a
alteração na expressão de 6 proteínas idênticas, nas condições estudadas, mostrou
sobreposição de respostas, que parece ser indicativo da infecção pelo L. theobromae no tecido
caulinar. Essas observações revelam proteínas que são alvos dos mecanismos celulares
acionados em plantas de cajueiro desafiadas com L. theobromae e constituem uma base inicial
de resultados que podem ser, futuramente, integrados a programas de melhoramento genético
do cajueiro, visando resistência, particularmente, à resinose.
Palavras-chave: Anacardium occidentale, resinose, Lasiodiploidea theobromae, proteômica,
defesa de plantas.

ABSTRACT
The cashew industry in Brazil is one of the main sources of income and labor in the
countryside and contributes largely to Northeastern Brazil's economy. This region
concentrates 94% of cashew production and the biggest producers are the states of Ceará,
Piauí and Rio Grande do Norte. To optimize the productive sector, strategies for genetic
improvement of cashew cultivars have focused on the selection and development of dwarf
clones. However, this practice has contributed to the decrease in genetic variability and,
consequently, to increased vulnerability to pathogen attacks. Gummosis, caused by the fungus
Lasiodiplodia theobromae (Pat.) Griff & Maubl. is considered the major disease of cashew in
semiarid conditions. Efficient methods to control the disease have not yet been established.
Due to lack of published data on the biochemical and physiological responses of cashew
infected with L. theobromae together with the fact that to defend themselves from pathogen
attack plants trigger/alter pathways in decorrence of reprogrammed expression of certain
proteins, studies to identify these proteins can provides information on the mechanisms
related to both cashew compatible and incompatible interactions with L. theobromae. In this
study, the differential proteomic analysis of cashew (Anacardium occidentale) stems, clone
BRS 226 (resistant) cultivated under controlled conditions, was performed at early times
following L. theobromae infection, as well as, of stems from cashew plants, resulting from
open polinization of BRS 226, classified as resistant and susceptible to gummosis, and
planted under field conditions where there is high pressure of the studied pathogen.
Differentially expressed proteins were identified by two-dimensional electrophoresis (2D-
PAGE) combined with mass spectrometry ESI-Q-TOF MS/MS. Cashew plants infected with
L. theobromae trigger physiological responses associated with the reprogrammed expression
of 73 proteins in the investigated scenarios. Of these, 36 were identified in BRS 226
artificially inoculated and 37 in cashew plants grown under field conditions. Therefore, an
equivalent number of proteins is responsive within the analyzed scenarios and showed cellular
functions in pathways of: energy production and metabolism, stress/defense, cell signaling
and protein folding/metabolism. Proteins responsive to hormones and involved in cell
structure were differentially expressed only in plants grown under field conditions, while
transport proteins were reprogrammed in L. theobromae inoculated BRS 226. Cashew plants,
cultivated in the field, and inoculated of the clone BRS 226 have the reprogrammed
expression of 6 identical proteins, including annexins and 14-3-3 proteins, involved in cell
signaling, which appear to be influenced by abiotic/biotic stress, under field conditions. The
common reprogramming of cellular functions plus the change in expression of 6 identical
proteins, under conditions studied, shows an overlap of responses that seem to be indicative
of infection with L. theobromae in stem tissue. These observations reveal proteins that are
targets of the cellular mechanisms triggered in cashew plants challenged with L. theobromae
and constitute a baseline of results that may be, in future, integrated programs of genetic
improvement of cashew, seeking resistance, particularly to gummosis.
Key-words: Anacardium occidentale, gummosis, Lasiodiploidea theobromae, proteomics,
plant defense.

LISTA DE FIGURAS
PÁGINA
Figura 1 Interação planta-patógeno e desenvolvimento da resistência à
doença.
21
Figura 2 Plantas de cajueiro com sintomas típicos de resinose no caule. 30
Figura 3 Inoculação de caules dos clones de cajueiro com o fungo
Lasiodiplodia theobromae.
41
Figura 4 Coleta de caules de cajueiro do clone BRS 226 em diferentes
tempos após inoculação com L. theobromae.
42
Figura 5 Medida do comprimento da lesão interna provocada pelo
patógeno em caules de cajueiro dos clones BRS 226 e CCP
76, resistente e suscetível, respectivamente, à resinose.
44
Figura 6 Plantas de cajueiro, clone BRS 226, infectadas com L.
theobromae. A) Sintoma de exsudação de resina no ponto de
inoculação com o fitopatógeno. B) Caules de plantas
infectadas e controles com 72 horas após inoculação.
54
Figura 7 Sintomas de escurecimento interno nos caules de cajueiro,
clones BRS 226 e CCP 76, nas condições controle e infectado
com L. theobromae.
55
Figura 8 Teor de proteínas solúveis extraídas de caules de cajueiro,
clone BRS 266, controle e inoculado com L. theobromae, em
diferentes tempos de amostragem. Barras indicam a média
aritmética das três amostras experimentais. Os tratamentos
não apresentaram diferenças significativas (p≤0,05) entre as
médias.
56
Figura 9 Perfil bidimensional de proteínas extraídas de caules de
cajueiro, clone BRS 226, nas condições controle e infectado
com L. theobromae. Cada mapa bidimensional foi obtido por
separação de 350 μg de proteínas em strips de 13 cm, pH 3-
10, seguido por SDS-PAGE (13%). Os géis foram corados
com coomassie brilliant blue G-250. Spots diferenciais são
indicados no grupo onde foram superexpressos.
59

Figura 10 Classificação funcional (A) e localização celular (B) das
proteínas diferencialmente expressas de caules de cajueiro,
clone BRS 226, inoculados com L. theobromae. Barras cinza e
branca indicam o número de proteínas superexpressas e
subexpressas, respectivamente, após inoculação com L.
theobromae.
64
Figura 11 Rede de interação proteína-proteína analisada pelo software
STRING. Linhas de diferentes cores representam diferentes
evidências de associação: verde, evidência de vizinhança;
vermelha, evidência de fusão; azul, evidência de coocorrência;
preta, evidência de coexpressão; roxa, evidência experimental;
azul claro, evidência em banco de dados; amarela, evidência
em mineração de texto; roxa clara, evidência de homologia.
67
Figura 12 Teor de proteínas solúveis extraídas de caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à
resinose. As barras indicam desvio padrão. Letras iguais não
diferem significativamente pelo teste de Tukey a 5%.
70
Figura 13 Número total de spots proteicos, visualizados por 2D-PAGE,
de extratos de caules de plantas de cajueiro, crescidas em
campo, resistentes e suscetíveis à resinose. As barras indicam
desvio padrão. Letras iguais não diferem significativamente
pelo teste de Tukey a 5%.
70
Figura 14 Comparação dos mapas bidimensionais de proteínas extraídas
de caules de plantas de cajueiro, crescidas em campo,
resistentes e suscetíveis à resinose. 350 µg de proteínas foram
separadas em strips IPG, pH 3-10 (13 cm), seguido por SDS-
PAGE (13%). Para spots acompanhados da letra R, a proteína
foi superexpressa no perfil resistente comparado com o
suscetível, enquanto para spots S, o inverso é aplicado. A
identidade das proteínas indicadas nos mapas bidimensionais é
apresentada na tabela 5.
71
Figura 15 Categorias funcionais das proteínas diferencialmente
reprogramadas em caules de plantas de cajueiro, crescidas em
campo, associadas à resistência/suscetibilidade. A) O número
de proteínas identificadas em cada função biológica é
indicado, dando ênfase às proteínas pertencentes a categorias
também identificadas em plantas do clone BRS 226 infectadas
com L. theobromae. B) Proteínas superexpressas/subexpressas
são mostradas comparando mudanças na expressão observada
75

em plantas resistentes com relação às plantas suscetíveis.
Figura 16 Atividade da enzima superóxido dismutase (UA/g-1
MF) de
caules de plantas de cajueiro, crescidas em campo, resistentes
e suscetíveis à resinose. Barras indicam desvio padrão. Letras
diferentes indicam diferença estatística entre as médias pelo
teste de Tukey (p≤0,05).
77
Figura 17 Atividade da enzima fenilalanina amônia liase (nmol CA/g-1
MF) de caules de plantas de cajueiro, crescidas em campo,
resistentes e suscetíveis à resinose. Barras indicam desvio
padrão. Letras diferentes indicam diferença estatística entre as
médias pelo teste de Tukey (p≤0,05).
77
Figura 18 Atividade da enzima β-1,3-glucanase (nkat g-1
MF) de caules
de plantas de cajueiro, crescidas em campo, resistentes e
suscetíveis à resinose. Barras indicam desvio padrão. Letras
diferentes indicam diferença estatística entre as médias pelo
teste de Tukey (p≤0,05).
78
Figura 19 Expressão diferencial da ascorbato peroxidase (APX) em
caules de plantas de cajueiro, crescidas em campo, resistentes
e suscetíveis à resinose. (A) As proteínas foram separados por
SDS-PAGE e imonudetectados com anti-corpo anti-APX
citosólica. (B) Abundância relativa calculada a partir da
eletroforese bidimensional.
79

LISTA DE TABELAS
PÁGINA
Tabela 1 Comprimento médio da lesão interna (cm) em caules de cajueiro, clones
BRS 226 e CCP 76, inoculados com L.theobromae, avaliado no período
de 72 e 96 horas após inoculação (HAI).
55
Tabela 2 Número de spots identificados nos mapas proteicos de plantas de cajueiro,
clone BRS 226, em diferentes tempos após inoculação com o fungo L.
theobromae.
57
Tabela 3 Identificação por MS/MS de proteínas diferencialmente expressas em
caules de cajueiro, clone BRS 226, infectado com Lasiodiplodia
theobromae.
61
Tabela 4 Abreviações dos nomes específicos das proteínas da rede funcional de
interação (Fig. 11), proposta pelo programa STRING v. 9.05.
68
Tabela 5 Identidade das proteínas diferencialmente expressas em caules de plantas
de cajueiro, crescidas em campo, classificadas como resistentes e
suscetíveis à resinose.
72

LISTA DE ABREVIATURAS E SIGLAS
2D-PAGE Eletroforese bidimensional em gel de poliacrilamida
PEG Polietileno glicol
PVPP Polivinilpolipirrolidona
EDTA Ácido Etileno diaminotetracético
DTT Ditiotreitol
TFA Ácido Trifluoracético
CHAPS 3-[(3-Cholamidopropyl)dimethylammonio]-1-propane sulfonate
IEF Focalização isoelétrica
IPG Gradiente de pH imobilizados em géis de poliacrilamida
PMSF Fluoreto de Fenilmetilsulfonil
SDS Dodecil Sulfato de Sódio
SDS-PAGE Eletroforese em Gel de Poliacrilamida contendo SDS
TCA Ácido tricloroacético
HAI Horas Após a Inoculação
ACN Acetonitrila
MS Espectrometria de Massas
UPLC Sistema de Cromatografia Líquida de Ultraperformance
ESI Ionização por Eletrospray

SUMÁRIO
Página
RESUMO........................................................................................................................
VIII
ABSTRACT....................................................................................................................
IX
LISTA DE FIGURAS ....................................................................................................
X
LISTA DE TABELAS ...................................................................................................
XIII
LISTA DE ABREVIATURAS E SIGLAS ...................................................................
XIV
INTRODUÇÃO
1. INTRODUÇÃO .......................................................................................................
19
1.1 Mecanismos de defesa de plantas .........................................................................
19
1.2 Cajueiro ..................................................................................................................
25
1.3 Lasiodiplodia theobromae e resinose ....................................................................
28
1.4 Proteômica no estudo da interação planta-patógeno .........................................
32
1.5 Justificativa ............................................................................................................
35
HIPÓTESE E OBJETIVOS
2. HIPÓTESE E OBJETIVOS ...................................................................................
38
2.1 HIPÓTESE .............................................................................................................
38
2.2 OBJETIVOS ..........................................................................................................
38
2.2.1 Objetivo Geral ......................................................................................................
38
2.2.2 Objetivos Específicos ...........................................................................................
38
MATERIAIS E MÉTODOS
3. MATERIAIS E MÉTODOS ...................................................................................
39
3.1 Reagentes ................................................................................................................
39
3.2 Materiais Biológicos .............................................................................................. 39
3.2.1 Lasiodiplodia theobromae ...................................................................................
39
3.2.2 Clones de cajueiro BRS 226 e CCP 76 ...............................................................
39
3.2.3 Plantas de cajueiro resistentes e suscetíveis, em condições de campo ...............
40
3.3 Metodologia Experimental ....................................................................................
40
3.3.1 Inoculação de caules de cajueiro com L. theobromae .......................................
40
3.3.2 Coleta de caules de cajueiro do clone BRS 226 ..................................................
41
3.3.3 Coleta dos caules de cajueiro resistentes e suscetíveis .......................................
42
3.3.4 Área de lesão interna da doença em clones de cajueiro resistente e
suscetível ........................................................................................................................
43
3.3.5 Extração de proteínas do tecido caulinar para análise proteômica ...................
44
3.3.5.1 Precipitação com solução de acetona, TCA e 2-ME..........................................
44

3.3.5.2 Extração das proteínas totais, lavagem e secagem do precipitado proteico.....
45
3.3.5.3 Ressupensão do precipitado proteico ................................................................
45
3.3.6 Dosagem de Proteínas ..........................................................................................
46
3.3.7 Análise proteômica diferencial de caules de cajueiro infectados com L.
theobromae ....................................................................................................................
46
3.3.8 Análise estatística da expressão diferencial de proteínas ...................................
47
3.3.9 Processamento dos spots, espectrometria de massas e identificação de
proteínas diferencialmente expressas ...........................................................................
48
3.3.10 Rede de interação das proteínas diferencialmente expressas ..........................
49
3.3.11 Preparação dos extratos proteicos de caules de plantas de cajueiro,
crescidas em campo, resistentes e suscetíveis à resinose .................................
50
3.3.12 Determinação das atividades das enzimas superóxido dismutase, ascobarto
peroxidase, peroxidase do guaiacol, fenilalanina amônia liase e β-1,3-glucanase
de caules de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à
resinose .....................................................................................................
50
3.3.13 Western blot de expressão da ascobarto peroxidase de caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à
resinose...........................................................................................................................
53
RESULTADOS
4. RESULTADOS ........................................................................................................
54
4.1 Sintomas de lesão interna em caules de clones resistente e suscetível à
resinose ..........................................................................................................................
54
4.2 Proteômica diferencial da interação entre o clone BRS 226 e o fungo L.
theobromae ....................................................................................................................
56
4.2.1 Rede funcional de interação das proteínas diferencialmente expressas ...........
66
4.3 Alteração no proteoma de caules de plantas de cajueiro, crescidas em
campo, resistentes e suscetíveis à resinose .................................................................
69
4.4 Determinação das atividades das enzimas superóxido dismutase, ascobarto
peroxidase, peroxidase do guaiacol, fenilalanina amônia liase e β-1,3-glucanase
de caules de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à
resinose ............................................................................................................
76
4.5 Western blot de expressão da ascobarto peroxidase em caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à resinose ..........................
78
DISCUSSÃO
5. DISCUSSÃO .............................................................................................................
80
5.1 Interação incompatível entre o clone de cajueiro BRS 226 e o fungo L.
theobromae ....................................................................................................................
80
5.2 Reprogramação da expressão de proteínas em caules de plantas de cajueiro
infectadas com L. theobromae .....................................................................................
82
5.2.1 Mudanças no proteoma caulinar ........................................................................
83

5.2.1.1 Reprogramação da expressão de enzimas envolvidas com o metabolismo e
produção de energia ......................................................................................................
84
5.2.1.2 Proteínas induzidas por estresse e relacionadas à defesa ................................
89
5.2.1.3 Proteínas relacionadas à sinalização celular....................................................
95
5.2.1.4 Metabolismo de proteínas e enovelamento ........................................................
98
5.2.1.5 Proteínas relacionadas ao transporte ...............................................................
101
5.2.1.6 Mudanças na expressão de proteínas associadas a componentes estruturais e
responsivas a hormônios................................................................................................
102
5.3 Sobreposição da resposta proteômica do cajueiro à infecção pelo
Lasiodiplodia theobromae ............................................................................................
105
CONSIDERAÇÕES FINAIS
6. CONSIDERAÇÕES FINAIS .................................................................................. 106
REFERÊNCIAS
REFERÊNCIAS ........................................................................................................... 107

19
1. INTRODUÇÃO
1.1 Mecanismos de defesa de plantas
As plantas representam uma fonte rica de nutrientes para muitos organismos e, por este
motivo, são alvos de ampla gama de potenciais invasores, como vírus, bactérias, fungos,
insetos e nematóides (FREEMAN, 2008; WIRTHMUELLER et al., 2013). Apesar da
ausência de um sistema imunológico semelhante ao dos animais, as plantas desenvolveram
uma impressionante variedade de mecanismos de defesa constitutivos e induzidos para evitar
o acesso de patógenos, tornando a condição de doença uma exceção e não um resultado
comum da interação planta-patógeno (HOK et al., 2010; PALLAS, 2011).
A presença de barreiras físicas, como a cutícula/parede celular, somada a existência de
metabólitos secundários tóxicos, compõe a defesa constitutiva, geralmente, suficiente para
bloquear as tentativas de infecção por um potencial agressor (LAZNIEWSKA, 2012).
Contudo, a ruptura dessas barreiras pré-formadas por determinados fitopatógenos pode iniciar
outro conjunto de respostas de defesa baseadas na percepção, por parte da planta, da tentativa
de ataque. Assim, os patógenos são confrontados com mecanismos de reconhecimento e
consequente sinalização na planta que acionam vias de resposta de defesa induzida (HOK et
al., 2010).
A primeira destas vias, de reconhecimento do iminente ataque do patógeno, baseia-se na
percepção de moléculas características e conservadas entre muitas classes de microrganismos.
Tais elicitores reconhecidos pelas plantas são, em geral, formados por estruturas essenciais
para o ciclo de vida destes microrganismos e que não necessariamente desempenham um
papel na patogenicidade (BITTEL e ROBATZEK, 2007). Estas moléculas são conhecidas
como padrões moleculares associados ao patógeno ou microrganismo (do inglês
Pathogen/Microbe-Associated Molecular Patterns - PAMPs ou MAMPs). Alguns poucos
PAMPs, tais como, flagelina, lipopolissacarídeos (LPS), quitina e ergosterol têm sido
identificados entre várias espécies patogênicas (BOLLER e FELIX, 2009). Distintas famílias
ou espécies de plantas têm desenvolvido sistemas de reconhecimento para moléculas
microbianas adicionais. Exemplos representativos são: o fator de alongamento Tu (EF-Tu) e
proteínas bacterianas induzidas por choque de frio (CSP) (KUNZE et al., 2004; FELIX e
BOLLER, 2003). Muito embora, tanto EF-Tu como CSP sejam abundantes em bactérias, a
percepção de EF-Tu é restrita à família Brassicacae, enquanto que CSP é limitada para

20
membros da família Solanaceae, indicando sistemas de reconhecimento com sensibilidades
diferenciais para determinados padrões moleculares. O reconhecimento, pela planta, desses
padrões conservados dos patógenos (PAMPS/MAMPs) é referido como resistência basal ou
resistência primária, sendo considerada a primeira linha de defesa à infecção que, embora
considerada relativamente fraca, atua impedindo a colonização da planta por parte de larga
gama de patógenos (AHMAD et al., 2010, GURURANI et al., 2012).
Uma vez reconhecidos por receptores de reconhecimento de padrões (PRR), presentes na
membrana celular das plantas, a imunidade acionada por PAMPs, denominada PTI (do inglês,
Pattern Triggered Immunity) é ativada. O estabelecimento da interação PAMP- PRR aciona
eventos de sinalização que estimulam o influxo de cálcio a partir do apoplasto, que por sua
vez, atua como mensageiro secundário no citoplasma para abertura de outros canais de
membrana (BLUME et al., 2000; BRUNNER et al., 2002; RANF et al., 2008) e ativação de
proteínas quinases dependentes de cálcio (LUDWIG et al., 2005). Além disso, eventos como
cascatas de proteínas quinases ativadas por mitógeno (MAPKs), produção de espécies reativas
de oxigênio (ROS) e nitrogênio (NOS), alterações na parede celular, incluindo deposição de
calose e reforço desta, síntese de proteínas relacionadas à patogênese (PR-proteínas) e
indução de compostos antimicrobianos são observados associados à indução da
reprogramação transcricional de genes de defesa (ZIPFEL, 2009; NEWMAN et al., 2013).
Muitos patógenos têm conseguido promover a reprogramação celular do hospedeiro,
superando estas repostas de sinalização. Isto ocorre por meio da produção e liberação em
estágios iniciais de infecção de fatores de virulência ou efetores, codificados por genes
específicos, responsáveis pela supressão da PTI (DEMPSEY e KLESSIG, 2012). Dessa
forma, uma comparativa “corrida armamentista” entre plantas e patógenos tem sido observada
(BENT e MACKEY, 2007), onde o surgimento de novas estratégias de defesa na planta é
acompanhado pela evolução, no patógeno, de novos meios para superar tais estratégias. Tal
coevolução é referida como modelo “zig-zag” (JONES e DANGL, 2006; NISHIMURA e
DANGL, 2010).
Baseado neste modelo, moléculas efetoras alteram o estado fisiológico do hospedeiro a
fim de favorecer a colonização ou interromper as respostas de defesa acionadas pela PTI
(HAMMOND-KOSACK, 2007). Efetores são, portanto, proteínas codificadas pelos genes de
avirulência do patógeno, genes Avr, que podem permanecer no espaço extracelular ou entrar
no citoplasma do hospedeiro, atingindo diferentes compartimentos celulares, tal como o

21
núcleo (KOECK et al., 2011). Entretanto, as plantas desenvolveram uma forma de imunidade
baseada no reconhecimento direto ou indireto dos efetores por meio da interação com
proteínas de resistência, codificadas pelos genes de resistência (genes R) do hospedeiro
(RAFIQI et al., 2009). Este reconhecimento ativa uma segunda via de defesa, a imunidade
acionada pelo efetor, ETI (do inglês, Effector Triggered Immunity), conforme ilustrado na
Figura 1.
Nesse modelo de resistência, proposto inicialmente por Flor (1955) e denominado
resistência gene-a-gene, ocorre interação direta entre os produtos dos genes de resistência
(genes R) dominantes da planta com os produtos dos genes dominantes de avirulência (Avr)
do patógeno. Sabe-se, hoje, que os genes R da planta codificam receptores (proteínas R) que
compõem seu sistema imunológico e o genes Avr do patógeno codificam moléculas efetoras
reconhecidas pelos receptores dos genes R. A existência desse modelo explica a condição de
resistência ou incompatibilidade à doença em determinados patossistemas, de modo que,
plantas que possuem o gene R resistem a determinadas raças de fitopatógenos, que expressam
seus correspondentes efetores (KEEN, 1990; VAN e JONES, 1998).
As principais classes de genes R encontrados em plantas codificam proteínas
citoplasmáticas que apresentam um domínio contendo repetições ricas em leucina (LRR) na
porção C-terminal, um sítio de ligação a nucleotídeos (NBS) e um domínio CC (do inglês
coiled coil) na extremidade N-terminal (GURURANI et al., 2012). A principal função das
proteínas NB-LRRs é o reconhecimento de proteínas efetoras específicas do patógeno,
resultando, daí, a imunidade induzida por efetores (ETI). Nas plantas, NB-LRRs podem
reconhecer moléculas efetoras por interação física direta ou indiretamente via proteína
intermediária.
Figura 1. Interação planta-patógeno e desenvolvimento da resistência à doença.
Fonte: Gururani, et al. (2012).

22
Uma variação da hipótese gene-a-gene, a Hipótese Guarda (equivalente do Inglês, Guard
model), que tem suporte em alguns patossistemas, preconiza a existência de reconhecimento
indireto, que ocorre após interação do efetor do patógeno com determinada proteína alvo do
hospedeiro (proteína guardada, equivalente do Inglês guardee protein). Modificações
bioquímicas, nesta proteína alvo, permitem o reconhecimento delas por uma NB-LRR
(proteína guardiã) particular (DANGL e JONES, 2001; ELLIS et al., 2007). Resumidamente,
as proteínas R (proteínas guardiãs, equivalente do Inglês guard proteins) funcionariam para
guardar o alvo celular vegetal (proteína alvo guardada) da interação com o efetor do patógeno
(proteína Avr). Por sua vez, a proteína R aciona os mecanismos de resistência quando
reconhece modificações desse alvo pelo patógeno, ou seja, após percepção, pela proteína R,
de um ataque à proteína guardada. Portanto, está implícita nesse modelo (Guard model), a
noção de que a proteína alvo guardada é indispensável para a função de virulência da proteína
efetora do patógeno quando na ausência da proteína R cognata (VAN DEER HOORNA et al.,
2008; GURURANI et al., 2012).
Comparado com a PTI, a ETI é tipicamente uma resposta acelerada e amplificada, uma
vez que esta imunidade é específica para efetores que são altamente polimórficos entre as
diferentes linhagens de fitopatógenos e aciona uma forte resistência à doença, que aumenta as
reações de defesas basais (JONES e DANGL, 2006; SPOEL e DONG, 2012). A ETI é
tipicamente associada com a morte celular programada (PCD) das células infectadas e a
produção de moléculas antimicrobianas no tecido circundante, levando à resistência local ao
agente patogênico (SPOEL e DONG, 2012).
A PCD representa uma forma de resposta hipersensitiva (HR) rápida, localizada nos sítios
de infecção do patógeno, presumivelmente destinada a limitar a proliferação e propagação do
invasor durante a ETI. Nas plantas, o aumento da produção de espécies reativas de oxigênio
(ROS) é considerado uma das principais causas de necrose que leva à PCD (ZHANG et al.,
2009). ROS sintetizadas no apoplasto por NADPH oxidases, assim como, por cloroplastos,
mitocôndrias e peroxissomos são importantes para o desenvolvimento da HR. Essa
compartimentalização pode ser essencial para as funções de ROS na sinalização durante a
defesa (TORRES e DANGL, 2005; TORRES, 2010).
Na célula, a HR afeta a homeostase de cálcio (Ca2+
), alterando a permeabilidade e o
potencial de membrana. Além disso, durante a HR, várias proteases tipos caspases, como
enzimas de processamento vacuolar, são ativadas, atuando, principalmente, como efetores da
morte celular ou necrose (MUR et al., 2008). Embora a necrose seja considerada um evento

23
clássico da resistência mediada por HR, em determinados patossistemas sua ocorrência pode
ser desacoplada da condição de resistência (ISHIBASHI et al., 2007, 2009). Baseado nesta
situação, uma possível relação com as diferentes formas de obtenção dos nutrientes por
invasores também deve ser considerada.
Os fitopatógenos são categorizados de acordo com o modo de nutrição. Patógenos
necrotróficos promovem ativamente a morte do tecido hospedeiro, uma vez que prosperam
sobre o conteúdo das células mortas. Esse estilo de vida contrasta com o de agentes
patogênicos biotróficos, que drenam para si nutrientes de células vivas e, portanto, devem
manter a viabilidade do tecido vegetal para o sucesso de seu desenvolvimento. Por sua vez,
um terceiro grupo, os hemibiotróficos, exibem ambas as formas de aquisição de nutrientes
(LALUK e MENGISTE, 2010).
A HR, como estratégia de defesa pode ser observada durante interações com patógenos
biotróficos, hemibiotróficos e necrotróficos. No entanto, está principalmente relacionada com
a imunidade aos biotróficos, hemibiotróficos, uma vez que estes fitopatógenos adquirem
nutrientes do tecido vivo (KUZNIAK et al., 2013).
Além da HR, a ativação das vias de sinalização da ETI resulta na geração de sinais móveis
que são transportados a partir dos locais infectados para tecidos distais, induzindo a
resistência sistêmica adquirida (SAR), uma forma de imunidade de longa duração para uma
ampla variedade de patógenos (AN e MOU, 2011). O início da SAR é marcado pela
acumulação do hormônio ácido salicílico (SA), de modo que a remoção do SA por expressão
constitutiva de uma salicilato hidroxilase anula a SAR (YALPANI et al., 1991; GAFFNEY et
al., 1993). Essas evidências demonstram o mais bem estabelecido papel do SA como
molécula de sinalização na resposta imune da planta (VLOT et al., 2009; TSUDA et al.,
2009).
Alguns dados sugerem que o metilsalicilato (MeSA), forma conjugada do SA, é o sinal
móvel que ativa a SAR nos tecidos não infectados, após sua translocação pelo floema a partir
do local da infecção e síntese (AN e MOU, 2011). O metilsalicilato transportado pode ser
hidrolisado por esterases liberando ácido salicílico, necessário para a percepção do sinal nos
tecidos sistêmicos. O alto nível de SA, produzido em sítios de infecção, inibe a atividade da
MeSA esterase por meio da ligação do hormônio ao sítio ativo da enzima, facilitando assim o
acúmulo de MeSA transportado para tecidos distais (PARK et al., 2007). No entanto, é

24
possível que algumas outras moléculas também possam servir como sinais de longa distância
para a SAR (TRUMAN et al., 2007; JUNG et al. 2009; SHAH e ZEIER, 2013).
O ácido salicílico participa nestas respostas imunes, controlando a circulação de uma
proteína chamada NPR1 (do inglês, Non-expressor of pathogenesis-related genes 1). Na
ausência de SA ou desafio do patógeno, NPR1 permanece retida no citoplasma como um
oligômero, por meio de pontes dissulfeto intermoleculares sensíveis à condição redox celular.
Quando as plantas são desafiadas por determinado invasor, NPR1 é reduzido ao estado
monomérico, forma considerada ativa e translocada para o núcleo (MOU et al., 2003). Uma
vez dentro do núcleo, NPR1 interage fisicamente com fatores de transcrição TGA-bZIP
induzindo a expressão de genes de defesa (MUKHTAR et al., 2009). Portanto, a degradação
de NPR1 é vital para limitar a ativação transcricional da SAR, evitando assim as
consequências associadas a uma resposta de defesa constitutiva na ausência de infecção
(SPOEL et al., 2009).
A SAR oferece resistência de amplo espectro contra fungos patogênicos, oomicetos, vírus
e bactérias. A "memória" imunológica em plantas conferida pela SAR pode durar de semanas
a meses e, possivelmente, até mesmo toda a estação de crescimento (KUC, 1987). Esta
resposta sistêmica é conferida por um conjunto de proteínas relacionadas à patogênese (PR-
proteínas) induzidas coordenadamente, cuja secreção requer aprimoramento significativo da
função do retículo endoplasmático (WANG et al., 2005; WANG e DONG, 2011 ). Baseado
em dados de sequências de aminoácidos e funções bioquímicas, as PR-proteínas foram
classificados em 17 famílias até o presente momento (SELS et al., 2008) e podem apresentar
dois modos básicos de contribuição na defesa vegetal, o primeiro baseado no bloqueio direto
do desenvolvimento do patógeno e o segundo pela liberação de elicitores, ativando
rapidamente a resposta de defesa da planta.
Em Arabidopsis thaliana, a expressão das proteínas PR1 (função desconhecida), PR2 (β-
1,3-glucanase) e PR5 (proteína tipo taumatina) são induzidas por SA e usadas como alvo para
identificação da SAR (UKNES et al., 1992). A contribuição genética das PR-proteínas na
resistência ao patógeno é complexa, e testar a colaboração de cada uma delas é um desafio,
uma vez que essas proteínas trabalham em conjunto nestas vias de sinalização (VAN LOON
et al., 2006).
Além do ácido salicílico, a infecção de plantas com diversos patógenos resulta em
mudanças no nível de vários fitormônios, tais como, ácido jasmônico (AJ), etileno (ET) e

25
ácido abscísico (ABA) (ROBERT-SEILANIANTZ et al., 2007; BARI e JONES, 2009;
PIETERSE et al., 2009). A resistência mediada pelo ácido salicílico é geralmente eficaz
contra patógenos biotróficos, enquanto que as respostas relacionadas à JA/ET são
predominantemente efetivas contra necrotróficos e insetos herbívoros (GLAZEBROOK,
2005). Vários estudos têm indicado interações mutuamente antagônicas entre a sinalização
dependente de SA e JA/ET, bem como, interações sinérgicas têm sido descritas (SCHENK et
al., 2000; KUNKEL e BROOKS 2002; BECKERS e SPOEL 2006; MUR et al., 2006). Isso
sugere, que a rede de sinalização de defesa ativada e utilizada pela planta é dependente da
natureza do patógeno e seu modo de patogenicidade. Além disso, o estilo de vida de
diferentes patógenos muitas vezes não é prontamente classificável como, exclusivamente,
biotrófico ou necrotrófico. Portanto, a interação positiva ou negativa entre vias do SA e
JA/ET, que determina o antagonismo ou sinergismo, pode ser regulada dependendo de
específicos patógenos (ADIE et al., 2007).
Por fim, baseado em todos os eventos citados, cuja interação molecular planta-patógeno
condiciona ou não a susceptibilidade a doença, recentes revisões têm sido propostas
enfatizando os mecanismos acionados para supressão das repostas de defesas de hospedeiros
por fungos, oomicetos e bactérias patogênicas (BIRCH, et al., 2009; ELLIS, 2009; HEIN et
al., 2009; SCHORNACK et al., 2009; MANSFIELD, 2009). A compreensão dos mecanismos
moleculares envolvidos nas vias de sinalização de defesa é um amplo campo de conhecimento
para pesquisas e aplicações, o que tem permitido o uso de abordagens inovadoras para o
desenvolvimento de espécies resistentes, por meio da engenharia genética de culturas (LI et
al., 2013).
1.2 Cajueiro
O cajueiro (Anacardium occidentale L.) pertence à família Anacardiaceae e caracteriza-se
por ser uma planta perene, de ramificação baixa e porte médio, cujo caule é comumente curto,
tortuoso e ramificado; as folhas são simples, inteiras, alternas, de aspecto subcoriáceo. É uma
planta andramonóica, com flores masculinas (estaminadas) e hermafroditas (perfeitas) numa
mesma panícula, e o fruto é um aquênio reniforme que se prende à panícula por um
pedúnculo hipertrofiado (LIMA, 1988; BARROS, 1993).
Segundo estudiosos, a origem brasileira do cajueiro é um fato, fundamentado em provas
circunstanciais, tais como, primeiras referências bibliográficas, distribuição geográfica,

26
comportamento ecológico, padrões de variação da espécie, utilização humana, dentre outros
(BARROS, 1993). Além disso, é provável que a origem do cultivo seja no Nordeste, onde os
primeiros colonizadores portugueses encontraram tradição de exploração da castanha e do
pedúnculo por parte das comunidades indígenas desta localidade (MAZZETTO et al.; 2009).
A região Amazônica é considerada, portanto, o centro da diversidade do gênero Anacardium,
enquanto que a maior diversidade de Anacardium occidentale L. está situada no Nordeste
brasileiro, com um centro secundário de diversidade nos cerrados (CRISÓSTOMO et al.,
2002; SOUSA et al., 2007).
Considerada uma das mais importantes espécies cultivadas das regiões tropicais, o
cajueiro ocupa no mundo, uma área estimada de 3,39 milhões de hectares, com uma produção
mundial de castanhas de aproximadamente 3,1 milhões de toneladas. Os principais produtos
de expressão econômica derivados do cajueiro são a amêndoa comestível e o líquido da
castanha. Além deste aspecto, os produtos derivados do cajueiro apresentam elevada
importância alimentar, observada pelo aumento de vendas e conquista de novos mercados
com a oferta de 30 subprodutos, entre os quais, se destacam o consumo do suco concentrado,
refrigerante gaseificado e cajuína (PAIVA e BARROS, 2004). Dentre os fatores associados a
essa tendência está à importância alimentar do consumo do pedúnculo in natura, que
apresenta por sua vez, elevados teores de vitamina C, fósforo, ferro e cálcio (SANTOS et al.,
2007; OLIVEIRA, 2008).
Os principais países responsáveis pelo suprimento mundial de amêndoas de castanha
do caju são a Índia, Vietnã e o Brasil, respectivamente. No Brasil a produção de castanha se
destina ao mercado externo, tendo como principais consumidores da amêndoa brasileira os
Estados Unidos e Canadá, com um mercado exportador gerando divisas em torno de 150
milhões de dólares anuais (FBB, 2010).
Enquanto espécie nativa do Brasil, 95% da área ocupada por esta cultura está situada
na região Nordeste, onde se destacam os estados do Ceará, Piauí e Rio Grande do Norte como
maiores produtores nacionais de castanha. Para a região semiárida, além de excelente
alternativa econômica, o cultivo do cajueiro apresenta uma significância social em virtude da
geração de empregos no campo, na entressafra de culturas tradicionais como o milho, feijão e
algodão, contribuindo com a redução do êxodo rural (CAVALCANTI et al., 2008;
GUANZIROLI et al., 2009). Diante destas condições, a cajucultura representa uma das
atividades agrícolas com maior potencial de crescimento sustentável (FERNANDES et al.,

27
2009). O desempenho produtivo em condições semiáridas é comprovadamente elevado, tanto
em quantidade como em qualidade do produto, o que permite inferir a ampla adaptabilidade
da espécie a estas condições (SOUZA et al., 2007; CARDOSO et al., 2009a).
Com relação à variabilidade genética existem na natureza dois tipos de cajueiro, o
comum e anão, classificados conforme o porte (CRISÓSTOMO, 2001). O tipo comum, mais
difundido, apresenta uma altura entre 8 e 15 metros com envergadura da copa atingindo 20
metros, caracterizando-se pela capacidade produtiva variável, com massa do pedúnculo entre
20 a 500 g. Por sua vez, o cajueiro anão, também conhecido como cajueiro de 6 meses, têm
porte baixo com altura inferior a 4 m, cuja massa do pedúnculo varia entre 20 a 160 g,
apresentando precocidade etária com início do florescimento entre 6 e 18 meses (BARROS,
2002; PAIVA, 2003; OLIVEIRA, 2008).
A cajucultura brasileira tem procurado alternativas para otimizar a produção e
competir com o mercador produtor de castanhas. Baseado nesta perspectiva, a exploração
inicialmente extrativista observada até a década de 1970, com técnicas rudimentares de
manejo, tem sido substituída nos últimos anos por investimentos em tecnologia agrícola, que
incluem preparo do solo, adubação, manejo de pragas, irrigação, colheita e pós-colheita, além
do desenvolvimento de pesquisas, por meio da criação em 1987 do Centro Nacional de
Pesquisa do Caju-CNPCa, atualmente chamado de Centro Nacional de Pesquisa de
Agroindústria Tropical, CNPAT (FBB, 2010). Uma das contribuições para o aumento na
produtividade da cajucultura está relacionada ao melhoramento genético de pomares. Em
linhas gerais, programas deste tipo têm como objetivo a seleção de cultivares com alto
potencial produtivo, características agronômicas superiores, adaptabilidade a diferentes
ambientes e estabilidade de produção (NETO et al., 2013).
Especial destaque tem sido dado ao melhoramento clonal, onde os clones de cajueiro
anão são importantes resultados desse método. A seleção destes clones se tornou uma
alternativa viável para a exploração econômica na região do semiárido nordestino, permitindo
a escolha de características favoráveis, tais como, uniformidade da castanha, pedúnculo,
produção, precocidade da colheita, facilidade no manuseio e condução dos pomares
(ROSSETTI e AQUINO, 2002; PAIVA et al., 2003).
A seleção dos primeiros clones de cajueiro ocorreu por meio de populações naturais
existentes na região litorânea do Nordeste, que foram introduzidas no Campo Experimental de
Pacajus - CE (OLIVEIRA, 2008; NETO et al., 2013). A seleção fenotípica individual e o

28
controle anual da produção nas plantas selecionadas permitiram a escolha dos clones de
cajueiro a serem difundidos no país (CCP 06, CCP 09, CCP 76 e CCP 1001), posteriormente,
métodos de melhoramento genético como policruzamento, seleção entre e dentro de progênies
e hibridização inter/intraespecífica, acabaram resultando na obtenção de outros clones
EMBRAPA 50, EMBRAPA 51, BRS 226 e BRS 265 (PAIVA et al., 2003; PAIVA e
BARROS, 2004; OLIVEIRA, 2008; FBB, 2010).
O uso de pomares de cajueiro tem gerado níveis produtivos no Brasil, que chegou a
contribuir com até 11% de todo o comércio mundial de castanhas (FAO, 2008). A alta
produção observada é devida, principalmente, ao uso de clones de cajueiro anão, que além de
atenderem aos parâmetros de qualidade comercial, citados anteriormente, ajudam a aumentar
a área de plantio e o tamanho da castanha. Contudo, levando em consideração o potencial
econômico desta cultura, a produtividade ainda permanece a níveis baixos, fato este associado
à deficiente infraestrutura na maior parte das áreas de cultivo e secas cíclicas nas regiões
produtoras (FREIRE e BARGUIL, 2001; MELO, 2002; OLIVEIRA, 2002). Somado a isso, o
uso de clones de cajueiro anão tem contribuído para redução da variabilidade genética e
consequentemente, a maior vulnerabilidade destas plantas ao ataque de patógenos (PAIVA et
al., 2002).
1.3 Lasiodiplodia theobromae e resinose
O fungo Lasiodiplodia theobromae (Pat.) Griffon & Maubl., pertence a família
Botryosphaeriaceae, que inclui espécies cosmopolitas e que habitam uma variedade de
hospedeiros de plantas (VON ARX, 1987). L. theobromae, formalmente referido como
Botryodiplodia theobromae Pat., é um fungo fitopatogênico de plantas tropicais e subtropicais
(TSUKADA et al., 2010), cuja infecção está frequentemente associada com plantas
submetidas a ferimentos naturais/provocados ou enfraquecidas por estresse (BRITTON e
HENDRIX, 1986; BAIRD e CARLING, 1998; PEREIRA et al., 2006).
Mais de 500 espécies de plantas podem ser infectadas por L. theobromae, entre as
quais se destacam as plantas lenhosas (MOHALI et al., 2005). A ampla variedade de espécies
vegetais que podem ser desafiadas por este fitopatógeno sugere, portanto, reduzida
especialização patogênica e grande variabilidade genética entre isolados (CARDOSO e
WILKINSON, 2008). Associado ao fato deste fungo ter se tornado um potencial invasor de
variadas culturas, uma hipótese tem sido proposta para ampla patogenicidade de L.

29
theobromae, segundo a qual é explicada como consequência da pressão ambiental,
especialmente nas regiões semiáridas, onde as condições climáticas lhes são muito favoráveis
(TAVARES, 2002; PEREIRA et al., 2006).
Em plantas de cajueiro, L. theobromae é responsável pela resinose, doença
relativamente nova, descrita inicialmente no município de Alto Santo - CE (FREIRE, 1991).
A doença foi primeiramente considerada de pouca importância devido a sua prevalência em
plantas de cajueiro comum com elevada idade e sob estresse. Contudo, os níveis epidêmicos
de pomares associados ao caráter destrutivo do patógeno tornam a resinose a principal doença
do cajueiro nas condições semiáridas do Nordeste brasileiro (CARDOSO et al. , 2010),
despertando a atenção de especialistas devido ao seu potencial de impacto adverso no
crescimento da indústria do cajueiro no Brasil (CYSNE et al., 2010).
Os primeiros sintomas de resinose ocorrem após a primeira safra comercial,
aproximadamente 24 a 34 meses após o plantio, com danos mais severos observados
preferencialmente no segundo ano (CARDOSO et al., 2006). Os sintomas caracterizam-se
pelo escurecimento, intumescimento e rachadura da casca, formando cancros pronunciados no
caule e ramos lenhosos, com posterior aparecimento de uma intensa exsudação de goma ou
resina, característica que confere o nome à doença (Figura 2). Sob a casca, observa-se um
escurecimento dos tecidos, o qual se estende, até atingir a região cortical e o câmbio vascular.
Com o progresso da doença, sintomas de deficiências nutricionais, murcha, queda de folhas e
morte dos ramos são observados, até o colapso total da planta (FREIRE et al., 2002;
CARDOSO et al., 2009a).
A importância da resinose para o cajueiro é acentuada devido à ausência de medidas
de controle. Muito embora, tenha sido recomendada a remoção cirúrgica do cancro quando a
infecção é localizada no tronco, seguido da aplicação de uma pasta fungicida a base de cobre
na superfície de corte (FREIRE, 1991), existe a prevalência de ramos infectados e a
observação de uma reinfecção no tronco no intervalo de 2 a 3 meses (CARDOSO et al.,
1995), o que contribui para baixa eficiência desse método. Além disso, o uso predominante do
clone CCP 76, reconhecidamente suscetível à resinose, associado às condições semiáridas,
com a prevalência do estresse hídrico e a ocorrência de coleobrocas do tronco e das raízes,
com as quais o fungo se associa sinergisticamente, tornam as plantas altamente vulneráveis
aos efeitos deletérios da doença (CARDOSO, 2009b).

30
Figura 2. Plantas de cajueiro com sintomas típicos de resinose no caule.
Fonte: Cardoso et al. (2009c) e Melo (2010).
A disseminação de L. theobromae em áreas isoladas tem sugerido o envolvimento do
besouro da raiz, Marshallus bondari, como vetor facilitador do processo de infecção
(FREIRE, 2002). Além disso, o vento, água, sementes, insetos e o homem, por meio de tratos
culturais, também estão associados com a propagação da doença. Fontes primárias do inóculo
têm sido obtidas a partir de sementes e mudas assintomáticas (FREIRE e CARDOSO, 1997;
CARDOSO et al., 2009c). Estudos de aspectos biológicos dos membros da família
Botryosphaeriaceae, baseados na distribuição taxonômica e na alta frequência de
sobrevivência endofítica, têm fornecido ferramentas para caracterizar várias espécies
examinadas, sugerindo que a grande maioria destas, se não todas, apresentam uma fase
endofítica de desenvolvimento (SLIPPERS e WINGFIELD, 2007), como evidenciado pelo
comportamento de L. theobromae.
Os danos causados pela resinose incluem redução no transporte de água e nutrientes,
com consequente destruição dos ramos e diminuição da fotossíntese, ocasionando perda da
produtividade do pomar, e ocorrendo, eventualmente, morte das plantas afetadas, reduzindo,
assim, a área de cultivo (BEZERRA et al., 2003; CARDOSO et al., 2004).
A resistência genética de plantas de cajueiro tem apresentado potencial perspectiva
para a redução epidêmica da resinose em pomares. Esta possibilidade está associada à
observação de que clones de cajueiro-anão se comportam diferentemente em relação à
incidência e à severidade da doença em condições de campo e com alta pressão de inóculo

31
(CARDOSO et al., 2006). Baseado nisto, o clone BRS 226, lançado por meio do programa de
melhoramento genético do cajueiro, mostrou ser resistente à resinose nas condições avaliadas
e com o rendimento da castanha semelhante ao clone comercial suscetível CCP 76 (PAIVA et
al., 2008). Além disso, o clone EMBRAPA 51 também tem se apresentado como uma
alternativa promissora de resistência nas mesmas condições de campo (CARDOSO et al.
2007), muito embora seu desempenho ainda seja considerado inferior ao clone BRS 226,
considerado o padrão comercial de resistência genética do cajueiro anão.
O possível surgimento de novas raças de patógenos associado à existência de um
único clone (BRS 226) de cajueiro com características produtivas e de resistência durável,
incentiva pesquisas para a seleção de novos genótipos que possibilitem aumentar a base
genética de resistência do cajueiro-anão para a região Nordeste do Brasil (MARTINS et al.,
2011; MOREIRA et al., 2013).
Nos últimos anos, pesquisas sobre o efeito de práticas agronômicas e a elucidação de
aspectos epidemiológicos associados ao progresso da resinose têm sido conduzidas pela
Empresa Brasileira de Pesquisa Agropecuária (Embrapa) (CARDOSO et al., 2009b). Além
disso, estudos sobre a especialização patogênica e genética têm revelado mais informações
sobre o fitopatógeno (CARDOSO e WILKINSON, 2008), assim como, a elucidação dos
meios de disseminação da doença foi alcançada por meio de estudos da interação enxerto e
porta-enxerto, bem como de sementes assintomáticas contaminadas pelo patógeno
(CARDOSO et al., 2009a; 2010). Efeitos da resinose na fotossíntese (BEZERRA et al.,
2003), seleção de genótipos resistentes (CARDOSO et al., 2006; 2007) e vias de colonização
do patógeno no tecido caulinar são outros alvos das pesquisas desenvolvidas (MUNIZ et al.,
2011). Contudo, é possível constatar a ausência de estudos publicados sobre os mecanismos
moleculares envolvidos na interação do cajueiro com L. theobromae e, mais especificamente,
sobre características moleculares associadas à incompatibilidade do clone BRS 226 ou de
populações caracterizadas como resistentes em condições de campo, com alta pressão do
inóculo. Essa observação destaca, portanto, a necessidade de estudos destinados à
compreensão das respostas bioquímicas de defesa de plantas de cajueiro desafiadas por L.
theobromae.

32
1.4 Proteômica no estudo da interação planta-patógeno
A interação planta-patógeno tem sido extensivamente estudada ao longo dos últimos anos,
envolvendo tanto o ponto de vista do hospedeiro como do parasita. Com o aumento dos
estudos genômicos e pós-genômicos, uma grande quantidade de informação está disponível
(RENSINK e BUELL, 2005; VARSHNEY et al., 2009; MOCHIDA e SHINOZAKI, 2010) e
avanços foram conseguidos no entendimento dos mecanismos de defesa das plantas, bem
como, nas estratégias de patogenicidade empregadas por microrganismos (MEHTA et al.,
2008).
Contudo, é possível constatar que os dados de expressão genética por si só não revelam a
completa complexidade de respostas moleculares a perturbações, tal como o ataque de
patógenos e, desse modo, o nível da expressão dos transcritos nem sempre é correlacionada
com o padrão celular das proteínas expressas (PIQUES et al., 2009; BAERENFALLER et al.,
2012). Tal perspectiva é justificada pelo fato de que alterações no comportamento celular
podem ser reguladas por proteínas pré-existentes, que são modificadas pós-traducionalmente
ou são degradadas, além disso, a possibilidade de variações no processamento do RNAm, por
splicing alternativo, permite a expressão de proteínas divergentes a partir de uma mesma
sequência gênica (QUIRINO et al., 2010). Levando em consideração que a interação planta-
patógeno envolve mecanismos moleculares associados a eventos de sinalização celular, que
culminam com cascatas de síntese, degradação e fosforilação de proteínas (DÓCZI et al.,
2007; JONES et al., 2006), a identificação de proteínas expressas pela planta após o desafio
de patógenos pode fornecer uma continuidade experimental com a informação contida no
genoma, permitindo a compreensão dos detalhes sobre as cascatas de sinalização envolvidas
na interação entre hospedeiros e fitopatógenos (MEHTA et al., 2008; QUIRINO et al., 2010).
O termo proteoma é definido como o conjunto de protéinas expressas pelo genoma de
uma célula, sob condições específicas. E a proteômica, conjunto de metodologias utilizadas
para caracterizar um proteoma, permite determinações qualitativas ou quantitativas de um
grande número de proteínas envolvidas no metabolismo celular (CAO et al., 2008; ELVIRA
et al., 2008). Nos últimos anos, a proteômica tem desempenhado um papel fundamental na
identificação de alterações nos níveis de proteínas em plantas hospedeiras sob infecção por
organismos patogênicos, e na caracterização de fatores celulares e extracelulares de virulência
produzidos por patógenos (LODHA et al., 2013).

33
Estudos proteômicos são iniciados com a preparação da amostra, por meio da extração de
proteínas do conteúdo celular. Em plantas, este processo é particularmente desafiador, uma
vez que células vegetais são ricas em constituintes, tais como polissacarídeos da parede
celular e polifenóis, além de proteases e enzimas oxidativas, cuja atividade pode interferir na
integridade do extrato protéico (ISAACSON et al., 2006; QUIRINO et al., 2010). A
dominância de certas proteínas, como a rubisco (ribulose-1,5-bifosfato carboxilase-oxigenase)
em folhas ou proteínas de armazenagem em sementes, pode comprometer a resolução de
proteínas de baixa abundância (JONES et al., 2004; CHEN e HARMON, 2006).
O aperfeiçoamento dos métodos de extração de proteínas do tecido vegetal tem sido
baseado no uso de diferentes passos, incluindo inicialmente, a escolha de diferentes tampões
de extração para amostras específicas, seguida de precipitação para concentrar proteínas e
eliminar compostos interferentes e, por fim, uma etapa de solubilização (ROCHA et al., 2005;
PAVOKOVIĆ et al., 2012). Muito dos protocolos, frequentemente empregados com sucesso,
utilizam para este fim a extração com ácido tricloroacético (TCA)/acetona e fenol-Tris,
seguido por precipitação com acetato de amônio/metanol (SARAVANAN e ROSE, 2004;
WANG et al., 2006; FAUROBERT et al, 2007; PAVOKOVIĆ et al., 2012). Como nenhum
único protocolo de extração de proteínas pode capturar o proteoma completo, protocolos têm
sido otimizados para determinados tecidos, e conforme objetivos da pesquisa
(MALDONADO et al., 2008).
A eletroforese bidimensional (2D-PAGE) é atualmente um dos métodos mais comumente
utilizados em estudos proteômicos (QUIRINO et al., 2010; GAUCI et al., 2013). Nesta
técnica, as proteínas da amostra de interesse são inicialmente separadas pelo seu ponto
isoelétrico (pI), na primeira dimensão, por meio da focalização isoelétrica, e posteriormente,
pela sua massa molecular na segunda dimensão (SDS-PAGE). A combinação dessas duas
etapas possibilita a separação de milhares de proteínas, culminando com a construção de ricos
mapas protéicos (ANDRADE, 2006).
Na 2D-PAGE, proteínas podem ser visualizadas em baixas concentrações por spot, e esta
visualização direta permite o controle da qualidade e reprodutibilidade do processo de
preparação da amostra (SMITH, 2009). Contudo, a principal razão da popularidade da
eletroforese bidimensional reside na facilidade da análise quantitativa de proteínas intactas.
Por quantificar a expressão das proteínas na imagem de gel 2D, é possível realizar
experimentos comparativos, por exemplo, detectando diferenças quantitativas e qualitativas

34
entre proteínas expressas em diferentes condições, amostras saudáveis versus doentes
(ROGOWSKA-WRZESINSKA et al., 2013) e, mais especificamente, no contexto da defesa,
plantas infectadas ou não com determinado invasor, a fim de entender a complexidade de
interações planta-patógeno (LODHA et al., 2013).
Para uma maior confiabilidade dos dados gerados em experimentos proteômicos, amostras
biológicas, bem como, repetições técnicas são comparadas usando vários programas
computacionais (por exemplo, PDQuest, BioNumerics, ImageMaster etc). Spots proteicos no
gel, considerados diferencialmente expressos com base na análise estatística dos géis, são
excisadas e processados para identificação por análise de espectrometria de massas
(QUIRINO et al., 2010).
A habilidade de uso de dados de espectrometria de massas (MS), inerentes a peptídeos,
para identificação de proteínas em bancos de dados é considerada uma das forças que
impulsiona a análise de proteomas (CANTÚ et al., 2008a). A MS é uma técnica sensível e
rápida, capaz de determinar a massa de moléculas, a partir da relação entre a massa e a carga
(m/z) de espécies ionizadas em fase gasosa (AEBERSOLD e MANN, 2003; CUNHA et al.,
2006). O espectrômetro de massas é um instrumento formado por uma fonte de íons, um
analisador de massas, um detector e um sistema de aquisição de dados. Atualmente,
analisadores de massas podem ser acoplados entre si, permitindo a ocorrência de
experimentos em sequência (MS/MS), sendo possível detectar determinado peptídeo e, em
seguida, submetê-lo a uma etapa de fragmentação, com consequente determinação da
sequência de aminoácidos (CANTÚ et al., 2008a).
Dados referentes à massa molecular dos peptídeos originados da digestão enzimática
(Peptide Mass Fingerprint - PMF), assim como, a informação referente à sequência de
aminoácidos dos peptídeos fragmentados (MS/MS), são comparados por bioinformática com
um banco de dados contendo sequências de proteínas conhecidas ou o genoma do organismo
(CHAMRAD et al., 2004; ELIAS et al., 2005). Isto é alcançado, por meio de programas, tais
como, MASCOT, Phenyx e OMSSA. Estes programas tem a capacidade de simular as
sequências primárias potenciais das proteínas, baseados na suposta predição da informação
contida em bancos de genes, e posteriormente, clivam teoricamente estas supostas proteínas
em peptídeos, calculando a massa absoluta dos peptídeos a partir de cada proteína. Portanto,
eles comparam massas de peptídeos da proteína alvo de estudo, desconhecida, à massa teórica

35
de peptídeos da proteína depositada em cada base de dados (CANTÚ et al., 2008a; QUIRINO
et al., 2010).
Recentes revisões têm demostrado à contribuição de estudos proteômicos na revelação de
detalhes sobre cascatas de sinalização e mecanismos moleculares envolvidos na interação de
plantas e patógenos (CURTIS, 2007; CHENG et al., 2010; BHADAURIA et al., 2010;
FERNÁNDEZ et al., 2010). Em algumas das pesquisas, a complexidade da amostra proteica
tem sido reduzida pelo uso da proteômica subcelular, baseada em estudos de proteínas
presentes em certos compartimentos celulares, tais como cloroplastos (Arai et al., 2008),
mitocôndrias (BRUGIERE et al., 2004), núcleo (PANDEY et al., 2008), vacúolo
(JAQUINOD et al., 2007) e parede celular (JAMET et al., 2008). A presença de certas
proteínas, como β-glicosidases e mirosinases, cuja localização nos peroxissomos de folhas era
anteriormente desconhecida, permitiu inferir o proposto papel destas organelas na defesa
contra patógenos e herbívoros (REUMANN et al., 2007).
Os dados de estudos proteômicos aliados às análises de validação funcional, podem
fornecer importantes contribuições na compreensão dos mecanismos complexos da interação
planta-patógeno. De acordo com esta perspectiva, o primeiro passo no entendimento da
incompatibilidade à doença está associado com identificação das proteínas expressas, durante
as interações planta-patógeno, seguindo, passos subsequentes de determinação de quais
proteínas conferem suscetibilidade ou resistência à doença, e os mecanismos atrelados a este
condicionamento (MEHTA et al., 2008). Todo este processo é bastante complexo e, em
virtude disto, alguns bons exemplos dessas informações podem ser integrados em estratégias
de melhoramento de culturas (VANDERSCHUREN et al., 2013).
1.5 Justificativa
A indústria do caju é uma das principais fontes de renda e trabalho no meio rural, e
representa a maior parcela da economia do nordeste brasileiro (MOREIRA et al., 2013). No
Brasil, a cajucultura mobiliza cerca de 280 mil pessoas e possui uma área cultivada de
740.000 ha (OLIVEIRA, 2008). Para permanecer no ranking mundial de produtores de
castanha, liderados pela Índia e Vietnã, a cajucultura brasileira tem buscado alternativas para
otimizar sua produção (FBB, 2010). Neste contexto, o uso de clones de cajueiro anão
desempenha interessantes contribuições, conferindo maior quantidade de castanhas por área

36
plantada, além de características nutricionais que favorecem o consumo dos seus pedúnculos
(CAJUNOR, 2013).
Plantas de cajueiro podem ser afetadas por várias doenças, que limitam o sistema de
produção agrícola (FREIRE et al., 2002; ADENIYI et al., 2011). Dentre elas, a resinose
causada pelo fungo Lasiodiplodia theobromae (Pat.) Griffon & Maubl. é considerada a
principal doença do cajueiro nas condições semiáridas do nordeste brasileiro. L. theobromae
infecta o caule levando à redução no transporte de água e nutrientes, com consequente
diminuição da fotossíntese e, eventualmente, a morte da planta (MUNIZ et al., 2011).
Nenhuma medida prática de controle da resinose, além da resistência genética, provou ser
eficaz em situações epidêmicas. Além disso, a expansão da área cultivada com o clone
suscetível CCP 76, associado com a descoberta de fontes primárias do inóculo, obtidas a
partir de sementes e mudas assintomáticas, tem contribuído para o surgimento de surtos
graves da doença (CARDOSO et al., 2009a; CARDOSO, 2010).
Estudos foram realizados para avaliar características ultraestruturais de infecção com L.
theobromae (MUNIZ et al., 2011), o efeito de diferentes combinações de enxerto e porta-
enxerto sobre a incidência de resinose (CARDOSO et al., 2010) e seleção de genótipos
resistentes (MOREIRA et al., 2013;. PAIVA et al., 2008). No entanto, não existem dados
publicados sobre as respostas fisiológicas e bioquímicas de plantas de cajueiro infectadas por
L. theobromae.
Nós últimos anos, a análise de proteomas tem sido empregada com sucesso para monitorar
respostas moleculares do hospedeiro ao desafio de patógeno (MANDELC et al., 2013). A
capacidade das plantas de se defenderem contra pragas e doenças está associada a uma série
de proteínas que podem diferencialmente reprogramadas (GERBER et al., 2008; YANG et
al., 2011; AFROZ et al., 2011). Uma vez que o proteoma representa a contribuição efetiva
das proteínas expressas para a função celular (MEHTA et al., 2008), a comparação com base
no perfil de expressão é necessária para obter uma compreensão mais completa dos
mecanismos envolvidos, e quais alterações são mais importantes na defesa das plantas
(ZIMARO et al., 2011).
Em nosso grupo de pesquisa, um estudo comparativo de mapas proteicos dos clones CCP
76 (sadio e doente) e BRS 226 (sadio e doente), coletados em campo, permitiu a identificação
de um conjunto de 80 proteínas que compõem o proteoma do cajueiro, assim como, de
proteínas diferencialmente expressas entre os grupos sadio e doente de cada clone. Em tal

37
estudo, um protocolo eficiente para elaboração de mapas bidimensionais de caules de cajueiro
foi desenvolvido, possibilitando a identificação de uma variedade de proteínas relacionadas à
defesa nos clones doentes (GONDIM, 2010).
Como os eventos moleculares de defesa associados à interação do cajueiro infectado com
L. theobromae permanecem desconhecidos, em especial, no que se refere a marcadores
proteicos de defesa, no presente estudo, a expressão de proteínas que possivelmente
justifiquem a condição de resistência/suscetibilidade foi explorada. Para tal proposito, foi
realizada análise proteômica diferencial de caules do clone BRS 226 (padrão comercial de
resistência), em tempos iniciais após inoculação com o fungo, e de plantas de cajueiro comuns
classificadas como resistentes e suscetíveis à resinose, em condições de campo com alta
pressão do patógeno.
Assim, esse estudo foi conduzido na perspectiva de identificar proteínas do caule de
cajueiro que são reprogramadas em resposta à infecção pelo L. theobromae e associá-las aos
mecanismos biológicos responsivos à presença do patógeno. Embora todo esse processo seja
bastante complexo, é esperado que esse estudo permita construir um conjunto de resultados
que possam ser, futuramente, integrados a programas de melhoramento genético do cajueiro
visando a geração, alicerçada em fundamentos bioquímicos, de novos genótipos resistentes à
resinose.

38
2. HIPÓTESE E OBJETIVOS
2.1 Hipótese
A resistência das plantas de cajueiro à infecção pelo fitopatógeno L. theobromae tem
como base a expressão diferencial de proteínas no caule envolvidas em alterações
bioquímicas e fisiológicas responsáveis pelo estabelecimento dessa relação de
incompatibilidade entre o fungo e a planta.
2.2 Objetivos
2.2.1 Objetivo Geral
Identificar proteínas diferencialmente expressas em caules de cajueiro do clone BRS
226, infectados pelo fungo Lasiodiplodia theobromae, bem como em caules de plantas
suscetíveis e resistentes à resinose, em condições de campo.
2.2.2 Objetivos Específicos
Extrair proteínas do tecido caulinar do clone BRS 226 (resistente), assim como de
caules de plantas resistentes e suscetíveis à resinose, em condições de campo;
Obter mapas bidimensionais com alta qualidade e reprodutibilidade das proteínas
extraídas dos caules de cajueiro;
Analisar os perfis bidimensionais obtidos de caules de cajueiro via programa PDQuest
8.0.1 (Bio-Rad Laboratories);
Identificar, por Espectrometria de Massas, as proteínas diferencialmente expressas nos
caules de cajueiro, após a infecção com L. theobromae;
Analisar a atividade de enzimas envolvidas com estresse oxidativo, metabolismo
secundário e PR-proteínas, em caules de plantas resistentes e suscetíveis à resinose.

39
3. MATERIAIS E MÉTODOS
3.1 Reagentes
Coomassie Brilhante Blue G-250, solução Fenol saturado com Tris, pH 8,0,
polivinilpolipirrolidona (PVPP), β-mercaptoetanol (2-ME), acrilamida e bis-acrilamida foram
obtidos da Sigma-Aldrich, Brasil. 3-[(3-Cholamidopropyl)dimethylammonio]-1-propane
sulfonate (CHAPS), Ditiotreitol (DTT), Fluoreto de Fenilmetil sulfonil (PMSF),
iodoacetamida, óleo mineral, tiouréia, uréia, tiras para focalização isoelétrica pH 3-10, , IPG
buffer 3 -10, dodecil sulfato de sódio (SDS) foram obtidos da GE Healthcare, Brasil. Ácido
Tricloroacético (TCA) e ácido trifluoracético (TFA), ambos possuindo grau analítico, foram
obtidos de diferentes fornecedores. A enzima tripsina foi adquirida da Promega, Madison, WI,
USA.
Os demais reagentes utilizados, de grau analítico, foram obtidos, comercialmente, de
diferentes fornecedores.
3.2 Materiais biológicos
3.2.1 Lasiodiplodia theobromae
O fungo L. theobromae utilizado nos experimentos foi previamente isolado de plantas
de cajueiro infectadas e situadas no município de Palhano-CE. Este fungo corresponde ao
isolado número 103 utilizado em testes de patogenicidade e pertence à Micoteca do
Laboratório de Fitopatologia da Embrapa Agroindústria Tropical, Fortaleza-CE. O fungo foi
mantido em meio de cultura batata-dextrose-ágar (BDA), que estimula a esporulação, e
incubado por 72 horas a 25 ± 1 ºC, com fotoperíodo de 12 horas claro/escuro.
3.2.2 Clones de cajueiro BRS 226 e CCP 76
Os clones de cajueiro utilizados neste estudo foram preparados sob condições de
viveiro na estação experimental da Embrapa Agroindústria Tropical, Pacajus-CE. Plantas dos
clones BRS 226 e CCP 76 foram enxertadas sobre o mesmo porta-enxerto, obtido de plântulas
do clone CCP 06 (SOUZA e ARAÚJO, 2001). Noventa dias pós-enxertia, as mudas foram

40
transportadas para a casa de vegetação do Laboratório de Fitopatologia da EMBRAPA
(Fortaleza - CE), onde permaneceram em condições de aclimatação durante 10 dias, com
radiação fotossinteticamente ativa (PAR) de 700 µm m−2
s−1
, temperatura média de 25 ºC
noite/ 35 ºC dia, umidade relativa de 30 - 70% e irrigação via aspersão com 8,60 mm de
água/planta.
3.2.3 Plantas de cajueiro resistentes e suscetíveis à resinose, em condições de campo
Para este estudo, foram utilizadas, também, plantas adultas classificadas como
resistentes e suscetíveis, em condições de campo, com aproximadamente 2 anos de idade.
Estas plantas fazem parte de uma população de cajueiro, estabelecida a partir da polinização
aberta do clone BRS 226, que foram cultivadas na Fazenda Planalto, pertencente à
Companhia Industrial de Óleos do Nordeste (CIONE), localizada na BR 020, km 4, município
de Pio IX-Piauí. As coordenadas geográficas da Fazenda são: latitude de 6° 34‟ e 24,5” S;
longitude de 40° 50‟ e 39” W; altitude de 730 m; temperatura média de 24 ºC (18 - 36 ºC) e a
pluviosidade média de 609,7 mm.
3.3 Metodologia Experimental
3.3.1 Inoculação de caules de cajueiro com L. theobromae
Caules dos clones de cajueiro, com aproximadamente 100 dias pós-enxertia, foram
inoculados com o micélio do fungo L. theobromae. A inoculação foi realizada perfurando o
caule com auxílio de uma broca de 1,6 mm de diâmetro, atingindo, aproximadamente, 0,5 mm
de profundidade em direção ao cerne da planta. Nesse orifício, um disco de 2 mm de diâmetro
do meio de cultura (BDA) contendo o micélio do fungo foi depositado, permitindo o contato
do fungo com o tecido vascular da planta. A deposição do inóculo no tecido caulinar foi
localizada 10 cm acima do ponto de enxertia. Após deposição, a região de inoculação foi
envolvida com vaselina e parafilm, para evitar a entrada de microrganismos oportunistas
(Figura 3). Plantas controles foram inoculadas apenas com o meio de cultura, sem conter a
estrutura do fungo (MARTINS et al., 2011). Os clones de cajueiro inoculados e controles
foram mantidos em casa de vegetação nas condições citadas acima (item 3.2.2).

41
Figura 3. Inoculação de caules dos clones de cajueiro com o fungo Lasiodiplodia
theobromae.
Legenda: A- Inoculação localizada 10 cm acima do ponto de enxertia; B- Perfuração do caule
com auxílio de uma broca de 1,6 mm de diâmetro, atingindo 0,5 mm de profundidade; C-
Disco de 2 mm de diâmetro contendo o micélio do fungo sendo depositado no ponto de
perfuração; D- A região de inoculação protegida com vaselina e parafilm; E- Lote de plantas
de cajueiro identificadas; F- Manutenção dos clones de cajueiro em casa de vegetação.
3.3.2 Coleta de caules de cajueiro do clone BRS 226
Para análise proteômica foram utilizados caules de cajueiro do clone BRS 226,
empregado comercialmente e resistente à resinose. A coleta dos caules desse clone foi
realizada em 12, 24, 48, 72 e 96 horas após inoculação (HAI), no Laboratório de
Fitopatologia (Embrapa-CE). As plantas controles e inoculadas foram submetidas à assepsia
da região de inoculação com etanol 70% embebido em algodão e, então, segmentos de caules
abrangendo 3 cm acima e abaixo do ponto de inoculação, perfazendo 6 cm de comprimento,
foram obtidos (Figura 4). Após coleta, os caules foram transportados para o Laboratório de
Proteínas Vegetais de Defesa, do Departamento de Bioquímica e Biologia Molecular
(DBBM)-UFC, onde foram armazenados a -20 °C para posterior processamento e extração de
proteínas.
O delineamento experimental adotado foi inteiramente casualizado (DIC), com 3
repetições, em esquema fatorial (2 x 5), sendo 2 tratamentos (plantas controles e inoculadas) e

42
5 tempos de análise (12, 24, 48, 72 e 96 HAI), com a parcela experimental composta por 4
plantas.
Figura 4. Coleta de caules de cajueiro do clone BRS 226 em diferentes tempos após
inoculação com L. theobromae.
Legenda: A- Remoção da fita de parafilm no ponto de inoculação; B- Assepsia da região de
inoculação com etanol 70%; C- Localização da região de corte 3 cm acima e abaixo do ponto
de inoculação; D- Corte do segmento caulinar com tesoura de poda; E- Segmentos de caules
com aproximadamente de 6 cm de comprimento; F- Amostras de caules identificadas para
posterior extração de proteínas.
3.3.3 Coleta dos caules de cajueiro resistentes e suscetíveis, em condições de campo
Plantas de cajueiro híbridas resultantes da polinização aberta do clone BRS 226
cultivadas de acordo com as condições citadas acima (item 3.2.3), foram mantidas em área
com alta pressão do patógeno (CYSNE et al., 2010). Uma característica da localização do
cultivo é a grande incidência de resinose, permitindo a origem de grupos característicos de
plantas com sintomatologia da doença. Em estudos de melhoramento genético com esta
população de plantas, realizados por pesquisadores da Embrapa (Fortaleza-CE), esta progênie
com 2 anos após semeadura foi avaliada e classificada (dados não publicados) de acordo com
escala numérica arbitrária de sintomas e severidade proposta por Cardoso et al. (2004).

43
Baseado nos resultados desta escala, 7 representantes de plantas classificadas como
resistentes, pertencentes ao grupo 0 ( ausência de sintomas), e 7 representantes de plantas
suscetíveis, avaliadas como grupo 2 (cancro rachados nos troncos e ramos, atingindo até 1/3
do diâmetro, com pouca ou ausência de exsudação de goma), foram escolhidos,
aleatoriamente, para deles serem seccionados segmentos de ramos caulinares medindo 6 cm
de comprimento. Após coleta, esses segmentos foram armazenados a -20 °C e transportados
para o Laboratório de Proteínas Vegetais de Defesa do DBBM-UFC.
3.3.4 Área de lesão interna da doença em clones de cajueiro resistente e suscetível
A análise comparativa da área de lesão interna da doença foi realizada com os clones
BRS 226 (resistente) e CCP 76 (suscetível), em 72 e 96 horas após inoculação com o L.
theobromae, intervalos de tempo quando os sintomas de exsudação de resina e lesão interna
do caule foram observados. Plantas controles e inoculadas desses clones foram obtidas como
citado acima (item 3.3.1).
O comprimento da lesão interna provocada pelo patógeno foi medido usando um
paquímetro, após corte longitudinal dos caules com auxílio de um estilete, na zona de
inoculação do patógeno, para expor a área lesionada (Figura 5).
O experimento foi montado em DIC, com 5 repetições, em esquema fatorial (4 x 2),
sendo 4 tratamentos (plantas controles e inoculadas do clone CCP 76, plantas controles e
inoculadas do clone BRS 226) e 2 tempos de análise (72 e 96 HAI), com cada parcela
experimental composta por 3 plantas.

44
Figura 5. Medida do comprimento da lesão interna provocada pelo patógeno em caules de
cajueiro dos clones BRS 226 e CCP 76, resistente e suscetível, respectivamente, à resinose.
3.3.5 Extração de proteínas do tecido caulinar para análise proteômica
Segmentos de caules de cajueiro foram liofilizados (FreeZone 6 benchtop freeze dry
system, Labconco, Kansas City, USA) durante 5 dias e, em seguida, pulverizados em moinho
até a obtenção de um pó com textura fina. Os extratos proteicos foram produzidos de acordo
com protocolos previamente descritos (WANG et al., 2003; YAO et al., 2006) e adaptados
por Gondim (2010). Na extração, foi utilizado Fenol-Tris, pH 8,0 (Acros Organic), que
permite a formação de uma fase fenólica superior, originada por meio do gradiente de
densidade estabelecido pela sacarose (30%) contida nesse tampão de extração, onde se
encontram as proteínas solubilizadas. Abaixo, estão descritas, resumidamente, as etapas do
protocolo de extração usado:
3.3.5.1 Precipitação com solução de acetona, TCA e 2-ME
Inicialmente, 1,5 g do pó do caule foi homogeneizado em 20 mL de uma solução
gelada de acetona contendo 10% de TCA e 2% de β-Mercaptoetanol (2-ME) durante 1 h a 4
°C. Em seguida, a precipitação foi realizada por meio da armazenagem do homogeneizado a -
80 ºC durante 2 h. Após centrifugação (15,000 x g, 20 min, 4 ºC) o precipitado resultante foi
lavado duas vezes com 15 mL de acetona contendo 2 % de 2-ME, para remoção do TCA, e

45
centrifugado como anteriormente. O precipitado foi, por fim, submetido à secagem em
dessecador na presença de sílica gel, a 4 ºC, durante à noite.
3.3.5.2 Extração das proteínas totais, lavagem e secagem do precipitado proteico
Ao precipitado obtido na etapa anterior, foram adicionados 10 mL do Tampão Tris-
HCl 0,1 M, pH 8,65, contendo 30% de sacarose, 2% de SDS, 0,001 M de PMSF, 2% de 2-ME
e 1% de PVPP. A amostra foi homogeneizada por 1 h e 30 min e, posteriormente,
centrifugada a 10.000 x g, 10 min, 4 ºC. O sobrenadante foi armazenado temporariamente a 4
ºC, e o precipitado submetido à re-extração com 5 mL do tampão de extração sob agitação
durante 30 min. Após centrifugação (10.000 x g, 10 min, 4 ºC), o sobrenadante obtido foi
adicionado ao sobrenadante coletado anteriormente, e o conjunto considerado o Extrato
proteico. Posteriormente, a esse extrato foi adicionado o mesmo volume do reagente Fenol-
Tris, pH 8,0. A mistura obtida foi agitada durante 15 min a 4 ºC. As proteínas presentes na
fase fenólica superior, gerada após centrifugação (10.000 x g, 10 min, 4 ºC), foram
precipitadas pela adição de quatro volumes de 0,1 M de acetato de amônio em metanol, a -80
ºC, durante 2 h. Após centrifugação (15.000 x g por 30 min, 4 ºC), o precipitado foi lavado
com 0,1 M de acetato de amônio em metanol gelado e, em seguida, três vezes com acetona
80% gelada. Entre cada lavagem com acetona, o precipitado foi incubado a 4 °C por 20 min e
a suspensão centrifugada a 10.000 x g, 10 min, 4 ºC. O precipitado proteico resultante foi seco
em dessecador na presença de sílica gel, a 4 ºC, durante à noite.
3.3.5.3 Ressupensão do precipitado proteico
O precipitado foi ressuspendido em 200 µL de Úreia: Tiouréia: Chaps [7 M: 2 M: 2%
(m/v)] por meio de agitação a 120 rpm por 20 min, seguido de sonicação durante 10 min e
repouso à temperatura ambiente, no intervalo de 20 min. Após centrifugação (10.000 x g, 10
min, 4 ºC), o sobrenadante obtido foi armazenado a 4 °C e o precipitado resultante submetido
a uma nova ressuspensão pela adição de 70 µL da mesma solução de Úreia: Tiouréia: Chaps,
citada acima. Ao término da centrifugação (10.000 x g, 10 min, 4 ºC), o sobrenadante foi
adicionado ao obtido na ressuspensão inicial, sendo o teor de proteína do extrato final dosado.

46
3.3.6 Dosagem de Proteínas
O teor de proteínas totais nos extratos obtidos foi determinado conforme metodologia
descrita por Bradford (1976). 2,5 mL do reagente Bradford foram adicionados a alíquotas de
0,1 mL dos extratos totais. Após 10 min de repouso, a absorbância foi determinada a 595 nm
em espectofotômetro (Novaspec III da Pharmacia). Albumina sérica bovina foi utilizada para
obtenção de uma curva padrão e definição do fator de correção, necessário para determinar o
teor de proteínas solúveis nos extratos.
3.3.7 Análise proteômica diferencial de caules de cajueiro infectados com L. theobromae
A análise proteômica da expressão diferencial de proteínas em caules de cajueiro
infectados com L. theobromae foi realizada por meio da eletroforese bidimensional
(O'FARREL; KLOSE, 1975). Para tanto, foram utilizadas plantas do clone BRS 226
(resistente) controles e inoculadas, assim como plantas resistentes e suscetíveis à resinose, em
condições de campo com alta pressão do patógeno. A eletroforese bidimensional consistiu,
inicialmente, da separação das proteínas de acordo com seu ponto isoelétrico (pI), por meio de
focalização isoelétrica. Nesta etapa, foram utilizadas fitas de gradientes de pH imobilizados
em géis de poliacrilamida (IPG) de 13 cm, pH 3-10 (GE Healthcare). As amostras proteicas
foram solubilizadas em solução de rehidratação [uréia 7M, tiouréia 2M, DTT 0,065 M,
CHAPS 2% (m/v), tampão IPG 2% (v/v) e azul de bromofenol 0,02% (m/v)] durante 16 h em
cubas de rehidratação (Reswelling Tray II, Pharmacia Biotech). Em seguida, as fitas
rehidratadas foram submetidas à separação isoelétrica utilizando a seguinte programação: 200
V por 45 min; 500 V por 45 min; 1000 V por 1 h; 5000 V por 1 h; 8000 V até atingir 18000
Volts/hora totais.
Após focalização isoelétrica, as fitas foram equilibradas sob agitação lenta, por 20 min
em solução de equilíbrio e redução [Tris-HCl 50 mM, pH 8.8, contendo glicerol 30% (v/v),
ureia 6 M, SDS 2% (m/v), DTT 2% (m/v) e azul de bromofenol] e, posteriormente, lavadas
com solução de alquilação [Tris-HCl 50 mM pH 8.8, glicerol 30% (v/v), ureia 6 M, SDS 2%
(m/v), iodoacetamida 2,5% (m/v) e azul de bromofenol] por 20 min, sob leve agitação.
A 2º dimensão (SDS-PAGE) foi realizada em gel vertical homogêneo (14 x 14 cm)
por meio da fixação das fitas focalizadas sobre o gel de poliacrilamida, utilizando solução de
agarose 0,5% (m/v). A separação das proteínas, de acordo com suas massas moleculares, foi

47
realizada a 5 °C, usando a fonte Power-Pac 3000 (Bio-Rad). A corrida foi composta de duas
etapas: 15 mA/gel durante 1 h, e 25 mA/gel até que o indicador (azul de bromofenol) saísse
do gel (aproximadamente, 5:30 h). As proteínas foram visualizadas com Comassie Brilliant
Blue (CBB) coloidal (CANDIANO, 2004). As imagens dos géis com proteínas coradas foram
obtidas por meio do ImageScanner (Amersham Biosciences) e analisadas pelo programa
LabScan v. 5.0 (GE Healthcare). As imagens dos géis foram processados e analisados
utilizando o programa PDQuest, versão 7.3.1 (Bio-Rad, EUA). Para tanto, as etapas de
processamento incluíram filtração da imagem, detecção dos spots e quantificação da
intensidade, além da eliminação de back-ground. Os mapas bidimensionais foram avaliados
como um conjunto único, representado por um gel de referência (máster), gerado a partir dos
demais mapas experimentais. Dessa forma, os spots consistentemente presentes nos géis
remanescentes foram adicionados ao máster, de modo que pudessem ser combinados para
todas as amostras. Além disso, a massa molecular (kDa) de cada proteína foi estimada
utilizando um conjunto padrão de marcadores e os pontos isoelétricos foram determinados
pelas posições dos spots ao longo das tiras com gradiente de pH imobilizado. Então, os spots
marcados nos géis, com intensidade registrada pelo PDQuest, foram submetidos à análise
estatística.
3.3.8 Análise estatística da expressão diferencial de proteínas
As intensidades dos spots proteicos (medidas em pixels) registrados nos perfis
bidimensionais foram avaliadas quanto à normalidade da sua distribuição utilizando o teste de
Shapiro-Wilk, enquanto que para avaliação da assimetria e curtose, o procedimento
UNIVARIATE foi empregado com a opção NORMAL do aplicativo estatístico SAS (v 9.0,
2002). Transformações logarítmicas [log (x+1)] foram realizadas quando necessário. As
variáveis que apresentaram distribuição normal e as que foram ajustadas à normalidade, após
transformação, foram consideradas paramétricas, enquanto aquelas que não apresentaram
distribuição normal foram consideradas não paramétricas.
As variáveis paramétricas foram submetidas à análise de variância por meio do
procedimento GLM (do inglês, General Linear Model) do SAS. Médias das intensidades dos
spots dos grupos resistente e suscetível, em condição de campo, foram comparadas usando o
teste t de Student. Para plantas do clone BRS 226, a análise de variância realizada pelo GLM
utilizou como fontes de variação a interação entre o tratamento avaliado (controle e

48
inoculado) e o tempo de análise (12, 24, 48, 72 e 96 HAI), e as médias das intensidades dos
spots foram comparadas por meio do Método dos Quadrados Mínimos, entre os diferentes
tratamentos dentro de cada tempo avaliado.
As variáveis não paramétricas, tanto para perfis bidimensionais da condição de campo
como para perfis do clone BRS 226, foram avaliadas por meio do teste de Mann-Whitney
utilizando o procedimento NPAR1WAY. Os resultados foram apresentados na forma de
médias e desvios padrão. Diferenças foram consideradas significativas quando p < 0,05.
3.3.9 Processamento dos spots, espectrometria de massas e identificação de proteínas
diferencialmente expressas
Os spots correspondentes às proteínas diferencialmente expressas foram excisados do
gel e digeridos com tripsina (Promega, Madison, WI, USA). Para tanto, os spots de interesse
excisados foram descorados com 400 l de 0,025 M de NH4HCO3 (m/v) e acetonitrila (ACN)
50% (v/v) até a remoção completa do corante, CBB coloidal. Posteriormente, as partículas do
gel foram desidratadas com 200 l de ACN 100% (v/v) por 5 min, até tornarem-se opacas e,
então, secas a vácuo em Speed Vac (Savant) por 15 min. Os pedaços de gel secos foram
incubados com tripsina grau sequenciamento (20 ng/μL em NH4HCO3 50 mM) a 37 °C em
banho-maria, por 17 h. Os peptídeos tripsinizados foram extraídos de cada spot-gel com 30 l
da solução de NH4HCO3 0,025 M, ACN 50% (v/v) e ácido trifluoracético 5% (v/v), com
auxílio de vórtex a 90 rpm, por 30 min (2 vezes). O sobrenadante contendo os peptídeos
trípticos extraídos foi, então, concentrado em Speed Vac (Savant).
Os peptídeos secos foram dissolvidos em solução de ácido fórmico 0,1% (v/v) e,
posteriormente, submetidos à análise por espectrometria de massas. Análises de MS/MS
foram realizadas em um espectrômetro de massas caracterizado por uma fonte de ionização
por eletrospray (ESI), dois analisadores de massas - um quadrupolo (Q) associado a um tubo
no qual se mede o tempo de vôo dos íons (TOF) e um detector de íons. Um sistema de
cromatografia líquida de ultra performace - UPLC (Waters, Milford, US) foi acoplado on-line
ao ESI-Q-TOF. Os peptídeos foram separados em uma coluna capilar nano -C18 (75 μm x 10
cm) com uma razão de fluxo de 0.35 µL/min. Os espectros de massa foram adquiridos pelo
instrumento Synapt G1 HDMS Acquity UPLC (Waters Co., Milford, MA, EUA), utilizando
Aquisição de Dados Dependente (DDA), onde os três principais picos foram selecionados e
sujeitos a MS/MS.

49
Os peptídeos foram eluidos por meio de um gradiente de água-ACN, segundo o qual
as fases móveis A e B foram constituídas de 0,1% de ácido fórmico em água e 0,1% de ácido
fórmico em acetonitrila, respectivamente. As condições do gradiente foram: 1 min, com 5%
de B, aumentando linearmente para 40% de B em 30 min, e posteriormente, para 80% de B
em 10 min, onde permaneceu até 35 minutos e nos 5 minutos seguintes houve uma redução
para 3% de B. Os espectros de íons resultantes foram coletadas e processadas utilizando o
programa Protein Lynx Global Server 2, sendo posteriormente convertidos para arquivos de
micromassa (PKL) usados para a pesquisa de sequências contra o banco de dados do NCBI
(National Center for Biotechnology Information) por meio do programa MASCOT v.2.2
(Matrix Science – www.matrixscience.com).
Para a correta indentificação das proteínas, as buscas de similaridade com proteínas
depositadas no NCBI foram realizadas com limitação taxonômica para “Viridiplantae taxa”,
além disso, os peptídeos foram considerados identificados, quando o valor do score excedeu
valor limiar de extensa similaridade ou identidade calculado pelo Mascot (p<0,05). Cada
proteína identificada foi, então, categorizada de acordo com o processo biológico com o qual
está envolvida e compartimentalização celular usando dados do UniProt
(http://www.ebi.uniprot.org). Quando as informações não estavam disponíveis, as proteínas
foram anotadas manualmente com base em pesquisas bibliográficas.
3.3.10 Rede de interação das proteínas diferencialmente expressas
A rede de interação funcional das proteínas identificadas neste estudo foi realizada
usando o programa STRING versão 9.05 (Search Tool for the Retrieval of Interacting
Genes/Proteins), disponível online (http://string-db.org/). A lista de proteínas
diferencialmente expressas foi explorada contra o banco de dados de Arabidopsis thaliana
depositados no STRING. As relações físicas e funcionais das proteínas responsáveis pela rede
de interação proteína-proteína foram estabelecidas com o apoio de associações decorrentes de
quatro fontes possíveis: contexto genômico, dados experimentais/bioquímicos, co-expressão e
informações previamente estabelecidas. Portanto, estes dados de interação são
quantitativamente integrados pelo STRING formando uma rede funcional de proteínas.

50
3.3.11 Preparação dos extratos enzimáticos de caules de plantas de cajueiro, crescidas
em campo, resistentes e suscetíveis à resinose
Os extratos enzimáticos foram obtidos por meio do contato do pó do caule,
previamente liofilizado e pulverizado em moinho, com o tampão de extração [acetato de sódio
0,05 M, pH 5,2, contendo 0,15 M de NaCl e 3% (v/v) de PEG] sob leve agitação a 4 °C, por 1
h. A relação massa (tecido): volume (tampão) utilizada foi 1:15 (m/v). Posteriormente, a
suspensão foi centrifugada (15.000 x g, 4 ºC, 20 min) e o sobrenadante coletado dialisado
contra o tampão de extração sem PEG, em membrana de diálise (Sigma) com poros de
exclusão de 14,3 KDa. Os extratos enzimáticos foram novamente centrifugados (10.000 x g,
4 °C por 20 min) e o sobrenadante usado para determinação das atividades enzimáticas.
3.3.12 Determinação das atividades das enzimas superóxido dismutase, ascobarto
peroxidase, peroxidase do guaiacol, fenilalanina amônia liase e β-1,3-glucanase de caules
de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à resinose
A atividade da superóxido dismutase (SOD, EC 1.15.1.1) foi avaliada segundo
metodologia descrita por Van Rossum et al. (1997). O princípio do ensaio consiste na ação do
radical superóxido (O2•-), formado no meio reacional, em converter o nitro azul de tetrazólio
(NBT) para formazana azul. A presença da SOD na amostra inibe essa reação final, portanto,
a medida de inibição da fotorredução do NBT é usada como parâmetro para determinação da
presença e atividade da enzima. No ensaio, alíquotas de 50 µL de extrato caulinar dialisado
foram adicionadas em uma placa de micropoços contendo uma mistura composta por 0,02 mL
de tampão fosfato de sódio 0,5 M, pH 7,8, 0,01 mL de Triton X-100 0,50% (v/v), 0,02 mL de
L-metionina 0,13 M, 0,01 mL de EDTA 0,002 M, 0,02 mL de NBT 0,001 M, 0,02 mL de
riboflavina 0,00075 M e 300 µL água grau Milli-Q. A adição do NBT e da riboflavina foi
feita na ausência de luz. A mistura reacional foi exposta à luz (lâmpada fluorescente de 32
W), dando início à reação, por 5 min. A produção de formazana azul foi medida pela leitura
de absorbância a 630 nm, em intervalos de 1 min, por uma leitora de ELISA (“Automated
Microplate Reader”, modelo ELX800-Bio-Tek Instruments®, Inc.). No branco da reação,
todos os reagentes, com exceção do extrato caulinar, foram colocados, permitindo redução
máxima do NBT. Uma unidade de atividade (UA) da SOD foi definida como a quantidade da
enzima necessária para inibir 50% da fotorredução do NBT à formazana azul, em relação ao

51
controle positivo, e foi expressa como unidade de atividade por g de massa fresca do caule
(UA/g MF).
A atividade da ascobarto peroxidase (APX, EC. 1.11.1.11) presente no extrato caulinar
foi realizado segundo Koshiba (1993) e Peixoto et al. (1999), com algumas modificações. 0,1
mL do extrato enzimático foi adicionado a 0,8 mL de tampão fosfato de potássio 0,05 M, pH
6,0, contendo 0,0005 M de ácido ascórbico e previamente incubado a 30 °C. A reação foi
iniciada pela adição de 0,1 mL de uma solução de peróxido de hidrogênio 0,002 M. O
decréscimo na leitura de absorbância a 290 nm, no intervalo de 10-120 s, foi medido como
índice de oxidação do ascorbato. A atividade da APX foi determinada utilizando uma curva
padrão construída a partir de concentrações conhecidas de ascorbato (0,1 - 1,0 µmoL
ascorbato/mL) e expressa em ηmol de ascorbate oxidado/min por grama de massa fresca
(ηmol Asc g-1
FW).
Para determinação da atividade da peroxidase do guaiacol (POX, EC 1.11.1.7) foi
utilizada a metodologia escrita por Urbanek et al. (1991). Como substratos para a reação
foram utilizados guaicol como doador de prótons e o peróxido de hidrogênio como receptor.
A mistura reacional consistiu de alíquotas de 0,1 mL do extrato total, 0,9 mL de tampão
acetato de sódio 0,05 M, pH 5,2, 0,5 mL de guaiacol 0,02 M e 0,5 mL de peróxido de
hidrogênio 0,06 M, perfazendo um volume total de 2,0 mL. A mistura foi incubada em banho-
maria a 30 °C e a leitura da absorbância do composto colorido 3,3‟-dimetoxi-4-4‟-
bifenolquinona, formado na reação, foi medida a 480 nm (espectrofotômetro Genesys 10S
UV-VIS; Thermo Scientific), durante 2 min em intervalos de 20 segundos. A variação de 1,0
unidade de absorbância por min foi assumida como sendo 1,0 unidade (UA) de atividade
peroxidásica, sendo expressa em unidades de atividade por g de massa fresca do caule (UA/g
MF).
A atividade da fenilalanina amônia liase (PAL, EC 4.3.1.5) foi determinada de acordo
com a metodologia descrita por El-Shora (2002) e Mori et al. (2001), com algumas
modificações. A atividade foi medida pela quantidade de ácido trans-cinâmico produzido, a
partir da desaminação da L-fenilalanina. A mistura reacional consistiu de 0,1 mL do extrato
enzimático, 0,2 mL de L-fenilalanina 0,04 M, 0,02 mL de β-mercaptoetanol 0,05 M, 0,58 mL
de tampão Tris-HCl 0,1 M, pH 8,8. Essa mistura foi incubada por 1 h, a 30 °C. A reação foi
parada pela adição de 0,1 mL de HCl 6 M e a mistura centrifugada a 10.000 x g por 10 min, a
25 °C. O branco do ensaio consistiu na adição da L-fenilalanina após parada da reação com

52
HCL. Após leituras de absorbância a 290 nm, a atividade da PAL foi determinada utilizando
uma curva padrão construída a partir de concentrações conhecidas de ácido trans-cinâmico
(0,01 – 0,1 µg ácido trâns-cinâmico/mL). A atividade da PAL foi expressa como a quantidade
(pmol) de ácido trans-cinâmico produzido por grama de massa fresca do caule por segundo
(pmol/g MF/s).
A atividade da enzima β-1,3-glucanase (GLU, EC 3.2.1.39) foi determinada segundo o
método descrito por Boller (1992) e medida em função da velocidade de formação de glucose
a partir da degradação da laminarina, polissacarídeo usado como substrato. Uma vez
liberados, os monômeros de glucose reduzem sais de cobre, em solução alcalina, que irão
produzir um composto de coloração azul, passível de ser quantificado por colorimetria. A
solução de laminarina (2,0 mg/L) foi dissolvida em água grau Milli-Q, aquecida a 60 ºC, por
10 minutos e, em seguida, dialisada exaustivamente contra água grau Milli-Q para remoção
da glucose livre. No ensaio, 0,1 mL de extrato enzimático foi incubado com 0,9 mL da
solução de laminarina, a 50 °C por 30 min. A seguir, 1,0 mL da solução “D” [1,0 mL da
solução “B” (15,0 g de sulfato de cobre pentahidratado, 0,02 mL de ácido sulfúrico e água
grau Milli-Q q.s.p. 100 mL) mais 25 mL da solução “A” (25,0 g de carbonato de sódio anidro,
25,0 g de tartarato de sódio e potássio, 20,0 g de bicarbonato de sódio, 200,0 g de sulfato de
sódio anidro e água grau Milli-Q q.s.p. 1000 mL)], preparada no momento do ensaio, foram
adicionadas e a mistura aquecida a 98 °C, em banho-maria, por 20 min. Após resfriamento em
água corrente, por 5 min, 1,0 mL da solução “C” [3,0 g de arseniato de sódio e água grau
Milli-Q, q.s.p. 25,0 mL] foi acrescido e, logo em seguida, os tubos foram agitados
vigorosamente, em vórtex, até a completa remoção dos gases formados na reação. Leituras de
absorbância a 520 nm foram feitas e a quantidade de monômeros de glucose liberados
determinada utilizando uma curva padrão construída a partir de concentrações conhecidas de
glucose, variando de 7,5 a 240 µg/mL, em tampão acetato de sódio 0,05 M, pH 5,2. A
atividade β-1,3-glucanásica foi expressa em nanokatal por grama de massa caulinar (ηkat/g
MF), sendo 1,0 ηKat equivalente a 1,0 nmol de glucose liberado por segundo, nas condições
do ensaio.

53
3.3.13 Western blot de expressão da ascobarto peroxidase de caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à resinose
20 µg do extrato total de proteínas a partir de plantas resistentes e suscetíveis foram
aplicados em gel SDS-PAGE (12,5%) e em seguida transferidos para membrana de
nitrocelulose (GE Healthcare, USA) usando um sistema de transferência (TE 22 tank transfer
unit, GE Healthcare, USA) a 100 V, 200 mA e 40 W, durante 2 horas. Posteriormente, as
membranas foram bloqueadas, durante à noite, com tampão PBS (50 mL de fosfato de sódio
0,1 M, pH 7,4, contendo 0,13 M de NaCl) sob agitação suave, a 4 °C e, então, incubadas por 2
horas com o anticorpo primário produzido em coelhos contra ascorbato peroxidase citosólica
(anti-APX, Agrisera) e utilizado para a imunodetecção na diluição de 1: 10.000.
As membranas foram novamente lavadas três vezes com PBS e incubadas com um
anticorpo secundário específico para imunoglobulinas de coelho (Agrisera), conjugado com
fosfatase alcalina, usado na diluição de 1:2.000 por mais 2 h. Seguindo-se as três lavagens
com PBS, a imunorreação foi visualizada por exposição das membranas ao tampão Tris-HCl
(0,1 M), pH 9,0, contendo BCIP® (fosfato de tetrazólio-5-bromo-4-cloro-3-indolilo, 0,15
mg/mL), NBT (azul de nitro-tetrazólio, 0,30 mg / mL ), NaCl (0,15 M) e MgCl2 (0,001 M). A
reação foi parada por lavagem exaustiva das membranas com água grau Milli-Q.

54
4. RESULTADOS
4.1 Sintomas de lesão interna em caules de clones resistente e suscetível à resinose
Plantas do clone BRS 226, usadas para avaliação da expressão diferencial de proteínas
após infecção com L. theobromae, exibiram sintomas de exsudação de resina e escurecimento
interno do lenho, conforme demonstrado na figura 6. Tais sintomas são característicos da
presença do fitopatógeno e foram observados em 72 e 96 HAI, de modo que plantas controles
não apresentaram qualquer tipo de exsudação de resina, indicando ausência de infecção.
Figura 6. Plantas de cajueiro, clone BRS 226, infectadas com L. theobromae. A) Sintoma de
exsudação de resina no ponto de inoculação com o fitopatógeno. B) Caules de plantas
infectadas e controles com 72 horas após inoculação.
Para avaliar a progressão comparativa destes sintomas entre os clones BRS 226
(resistente) e CCP 76 (suscetível) foi realizada, em 72 e 96 HAI, medida do comprimento da
lesão interna da doença no caule, tomando como critério de medição a região escurecida do
lenho ao redor do ponto de inoculação, uma vez que a presença de cor amarronzada pode ser
observada atingindo o câmbio vascular, quando cortes transversais ou longitudinais são feitos
no tronco e ramos lenhosos (FREIRE et al. 2002, 2004; FREIRE e CARDOSO, 2003).
A análise de variância mostrou que houve interação significativa entre os fatores
tratamento e tempo, como observado na Tabela 1. Baseado nos dados obtidos foi possível
constatar que com 72 HAI, plantas controles e inoculadas dos clones BRS 226 e CCP 76
apresentaram valores médios de lesão interna que não diferem estatisticamente entre si.

55
Contudo, com 96 HAI, a progressão da lesão nas plantas inoculadas do clone CCP 76 mostrou
maior extensão de necrose no caule (Figura 7), o que é compatível com a característica de
suscetibilidade, registrada em outros trabalhos (CARDOSO et al., 2010; MUNIZ et al., 2011).
Tabela 1. Comprimento médio da lesão interna (cm) em caules de cajueiro, clones BRS 226 e
CCP 76, inoculados com L.theobromae, avaliado no período de 72 e 96 horas após inoculação
(HAI).
Clone Tratamento Comprimento da lesão (cm)
72 HAI 96 HAI
CCP 76 (S) Controle 0,50 Aa 0,76 Ba
Inoculado 0,81 Ab 1,47 Aa
BRS 226 (R) Controle 0,43 Aa 0,53 Ba
Inoculado 0,71 Aa 0,72 Ba
Desv. Padrão Geral 0,23
Médias seguidas da mesma letra maiúscula na coluna e minúscula na linha não diferem entre si
pelo teste de Tukey (p≤ 0,05). S, suscetível; R, resistente ao L. theobromae.
Figura 7. Sintomas de escurecimento interno nos caules de cajueiro, clones BRS 226 e CCP
76, nas condições controle e infectado com L. theobromae.

56
4.2 Proteômica diferencial da interação entre o clone BRS 226 e o fungo L. theobromae
Amostras de caules do clone BRS 226, na condição controle e inoculado, foram
coletados nos tempos de 12, 24, 48, 72 e 96 HAI, conforme descrito anteriormente e então,
submetidos à extração de proteínas para análise proteômica. Os extratos foram dosados e o
conteúdo médio de proteínas solúveis obtido. Resultados da quantificação de proteínas
extraídas mostraram valores médios uniformes ao longo do experimento, não havendo,
portanto, diferença significativa (p≤0,05) entre os tratamentos (Figura 8).
Para cada amostragem experimental representada pelo grupo, controle e inoculado no
referido tempo, foram obtidos mapas bidimensionais representativos destas condições. No
conjunto do experimento, 6 géis bidimensionais representam cada tempo experimental (12,
24, 48, 72 e 96 horas): 3 géis pertencentes ao grupo controle e 3 géis para o grupo inoculado,
totalizando 30 géis, que foram escaneados e analisados com relação à expressão diferencial de
proteínas em cajueiros inoculados com L. theobromae.
Figura 8. Teor de proteínas solúveis extraídas de caules de cajueiro, clone BRS 266, controle
e inoculado com L. theobromae, em diferentes tempos de amostragem. Barras indicam a
média aritmética das três amostras experimentais. Os tratamentos não apresentaram
diferenças significativas (p≤0,05) entre as médias.
Com relação ao número total de spots, foram encontrados no experimento 729 ± 24 e
721 ± 25 para os tratamentos controle e inoculado, respectivamente. Conforme demonstrado
na tabela 2, não houve diferença significativa (p ≤ 0,01) entre os grupos controle e inoculado
nos respectivos tempos de estudo. Contudo, a análise estatística da interação do número total

57
de spots, ao longo do tempo, demonstrou que a quantidade de spots observada foi maior nos
grupos experimentais com 72 HAI, sendo esse resultado diferente dos demais tempos, mas
semelhante ao observado para 24 HAI. Esses resultados apontam que, nesse intervalo de
tempo após inoculação, uma variedade maior de proteínas está sendo mobilizada pela planta.
Tabela 2. Número de spots identificados nos mapas proteicos de plantas de cajueiro, clone
BRS 226, em diferentes tempos após inoculação com o fungo L. theobromae.
Tratamento
Horas após inoculação (HAI)
MÉDIA 12 HAI 24 HAI 48 HAI 72 HAI 96 HAI
Controle
Inoculado
726,33 A
711,33 A
727,33 A
742,33 A
707,66 A
693,33 A
770,00 A
772,33 A
717,33 A
689,00 A
729,73 A
721,66 A
Média 718,83 b 734,83 ab 700,50 b 771,16 a 703,16 b
C.V. 3,67
Médias seguidas da mesma letra maiúscula na coluna e minúscula na linha não diferem entre si pelo teste de Tukey (p ≤
0,01).
A abundância relativa dos spots nos respectivos tratamentos, ao longo do tempo, foi
submetida à análise estatística e baseado no uso das fontes de interação tratamento e tempo,
64 spots proteicos apresentaram expressão alterada. Para análise proteômica uma seleção da
condição de alteração foi realizada e de acordo com isso, foram considerados para
excisão/identificação os spots que se enquadravam nos seguintes parâmetros:
1. Alteração da expressão decorrente do fator tratamento (controle e inoculado) dentro do
tempo, sendo desconsiderados spots em que a expressão foi alterada em virtude somente do
tempo.
2. Devida localização de registro no software PDQuest, por meio da conferência dos spots nos
mapas proteicos. Aqueles decorrentes de marcação manual erroneamente registrada não
foram selecionados.
3. Consenso na expressão ao longo dos tempos de estudo, observada por gráficos que
representam a expressão relativa.
Baseado nesta seleção, 24 spots foram eliminados, a maior parte deles por não
atenderem ao parâmetro 1. O grande número de proteínas, com expressão alterada em virtude
do tempo, revela um panorama de alterações decorrentes da condição fisiológica da planta no
ambiente e constituem dados que fogem do escopo inicial da pesquisa, ou seja, a identificação

58
de proteínas cuja expressão é reprogramada devido à infecção pelo fungo. Dessa forma, um
total de 40 spots foram excisados dos géis bidimensionais, digeridos com tripsina e
submetidos à identificação por Espectrometria de Massas (ESI-Q-TOF MS/MS).
A análise por Espectrometria de Massas (MS) permitiu a identificação de 36 proteínas,
perfazendo, aproximadamente, 90% da quantidade de spots diferencialmente expressos
considerados para análise. O estabelecimento da interação do clone BRS 226 com o fungo
levou a um aumento significativo da expressão de 19 proteínas no grupo inoculado. Por outro
lado, 17 proteínas apresentaram expressão diminuída após o contato com o L. theobromae
sendo, portanto, mais expressas nos perfis de plantas controles. No experimento, não foi
possível observar a presença de proteínas que sejam exclusivas de somente um grupo de
estudo. Na figura 9, são representados os mapas proteicos de cada condição (controle e
inoculado) e os spots cuja abundância relativa foi aumentada são indicados nos perfis
bidimensionais em que houve reprogramação da expressão.

59
Figura 9. Perfil bidimensional de proteínas extraídas de caules de cajueiro, clone BRS 226, nas
condições controle e infectado com L. theobromae. Cada mapa bidimensional foi obtido por
separação de 350 μg de proteínas em strips de 13 cm, pH 3-10, seguido por SDS-PAGE (13%).
Os géis foram corados com coomassie brilliant blue G-250. Spots diferenciais são indicados no
grupo onde foram superexpressos.

60
Do total de spots expressos diferencialmente e numerados de 1 a 40 (Fig. 9), somente
4 (spots 1, 2, 4 e 12) não foram identificados por MS. A maior parte das proteínas
identificadas apresentou expressão alterada em 72 e 96 horas após a inoculação do patógeno,
período em que sintomas de exsudação de resina foram observados.
De acordo com a análise dos dados relativos à intensidade dos spots em cada gel,
registrados pelo programa PDQuest, foi calculado o índice de variação de cada proteína, com
expressão diferencial, em comparação aos respectivos valores de intensidade do spot nos géis
referentes ao grupo de plantas controle.
A identidade das proteínas está representada na tabela 3, conforme busca de
similaridade com outras sequencias de proteínas já depositadas no NCBI. Na tabela consta
também, o número do spot, organismo vegetal de referência, número de acesso no banco de
dados, os valores de ponto isoelétrico e massa molecular (teóricos e experimentais), score,
cobertura da sequência e o índice de variação.
As 36 proteínas identificadas na análise diferencial foram agrupadas de acordo com o
processo biológico que desempenham no organismo vegetal. Baseado nesta classificação, a
condição de infecção provoca reprogramação da expressão de proteínas envolvidas em vários
processos celulares e, portanto, categorizadas nas seguintes funções: metabolismo e energia
(13), estresse e defesa (8), sinalização celular (3), metabolismo e enovelamento de proteínas
(5), transporte (2) e proteínas cuja função permanece desconhecida (5). Estes resultados
mostram que um grande número de proteínas envolvidas com o metabolismo e produção de
energia alteram sua expressão em resposta a L. theobromae, seguido por proteínas
relacionadas a estresse e defesa.

61
Tabela 3. Identificação por MS/MS de proteínas diferencialmente expressas em caules de cajueiro, clone BRS 226, infectado com Lasiodiplodia
theobromae.
N° do spot
Proteína
Acesso
(NCBI)
Organismo de
Referência
Mr/pI
Teórico
Mr/pI
Experimental
Cobertura
(%)
Score
(a)
Índice de variação
Infectado vs. Controle (b)
12 hai 24 hai 48 hai 72 hai 96 hai
Energia e metabolism
11 NADH dehydrogenase, putative gi|255545146 Ricinus
communis 18.97/4.76 21.6/4.7 24 107 1.58
15 PREDICTED: 1,2-dihydroxy-3-keto-5-
methylthiopentene dioxygenase 2-like gi|470121006
Fragaria
vesca 23.67/5.14 28.3/4.8 40 435 1.36
16
PREDICTED: 3-isopropylmalate
dehydratase small subunit-like isoform
1
gi|460392785 Solanum
lycopersicum 27.07/6.52 26.2/4.8 16 156 1.55 1.44
17 PREDICTED: chalcone--flavonone
isomerase-like isoform 1 gi|225448675 Vitis vinifera 23.31/4.81 29.0/4.9 19 166 1.56
18 Fructokinase gi|45550051 Citrus unshiu 37.48/5.11 45.6/4.9 14 282 1.54
20 PREDICTED: pyruvate decarboxylase
isozyme 2-like isoform X1 gi|502104230
Cicer
arietinum 65.06/5.62 75.5/6.0 6 196 1.99
23 S-adenosylmethionine synthetase gi|17529621 Oryza sativa 43.19/5.93 54.4/6.0 18 206 0.93
25 Pyruvate decarboxylase, putative gi|255579310 Ricinus
communis 64.71/5.71 76.9/5.9 9 237
0.40
32 Alpha-glucan phosphorylase isozyme
H gi|15232704
Arabidopsis
thaliana 95.10/5.79 97.5/7.1 8 187 0.64
35 Fructose-bisphosphate Aldolase
cytoplasmic isozyme gi|474256276
Triticum
urartu 38.88/6.85 44.3/8.0 21 498 3.86
37 Alcohol dehydrogenase class III gi|400177690 Lactuca sativa 40.43/6.51 53.3/7.6 8 118 0.85
38 Serine hydroxymethyltransferase,
putative gi|255563608
Ricinus
communis 51.84/7.59 65.4/7.5 25 561 0.64
39
Small subunit of ribulose-1,5-
bisphosphate carboxylase
gi|218210
Oryza sativa
Japonica
Group
19.67/9.11 16.4/9.6 9 128 2.66
Estresse e defesa
07 Protein disulfide isomerase gi|171854980 Glycine max 58.55/5.13 70.2/4.5 7 193 1.43 1.83 1.54
09 Heat shock protein 70 gi|1143427 Cucumis
sativus 75.36/5.15 89.4/4.5 16 530 1.33 1.65
10 2-cys peroxiredoxin-like protein gi|9758409 Arabidopsis
thaliana 29.54/5.55 24.5/4.4 26 159 1.29 1.76
21 HSP20-like chaperones superfamily
protein gi|508786804
Theobroma
cacao 17.96/5.57 19.9/5.7 25 161 0.46
27 Glutathione-s-transferase theta, gst,
putative gi|255578691
Ricinus
communis 24.85/6.24 27.0/6.2 15 66 0.56 0.59
28 PREDICTED: CBS domain-containing gi|449444316 Cucumis 22.68/7.03 18.7/6.6 20 240 0.51 0.27
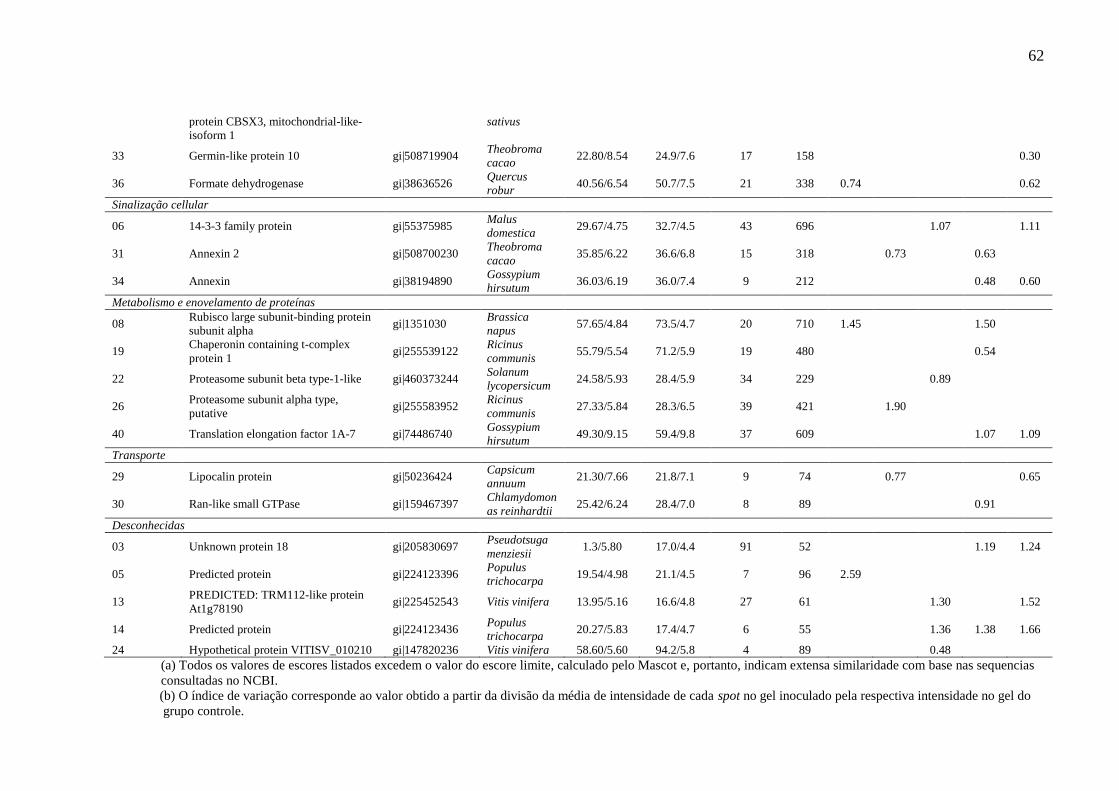
62
protein CBSX3, mitochondrial-like-
isoform 1
sativus
33 Germin-like protein 10 gi|508719904 Theobroma
cacao 22.80/8.54 24.9/7.6 17 158 0.30
36 Formate dehydrogenase gi|38636526 Quercus
robur 40.56/6.54 50.7/7.5 21 338 0.74 0.62
Sinalização cellular
06 14-3-3 family protein gi|55375985 Malus
domestica 29.67/4.75 32.7/4.5 43 696 1.07 1.11
31 Annexin 2 gi|508700230 Theobroma
cacao 35.85/6.22 36.6/6.8 15 318 0.73 0.63
34 Annexin gi|38194890 Gossypium
hirsutum 36.03/6.19 36.0/7.4 9 212 0.48 0.60
Metabolismo e enovelamento de proteínas
08 Rubisco large subunit-binding protein
subunit alpha gi|1351030
Brassica
napus 57.65/4.84 73.5/4.7 20 710 1.45 1.50
19 Chaperonin containing t-complex
protein 1 gi|255539122
Ricinus
communis 55.79/5.54 71.2/5.9 19 480 0.54
22 Proteasome subunit beta type-1-like gi|460373244 Solanum
lycopersicum 24.58/5.93 28.4/5.9 34 229 0.89
26 Proteasome subunit alpha type,
putative gi|255583952
Ricinus
communis 27.33/5.84 28.3/6.5 39 421 1.90
40 Translation elongation factor 1A-7 gi|74486740 Gossypium
hirsutum 49.30/9.15 59.4/9.8 37 609 1.07 1.09
Transporte
29 Lipocalin protein gi|50236424 Capsicum
annuum 21.30/7.66 21.8/7.1 9 74 0.77 0.65
30 Ran-like small GTPase gi|159467397 Chlamydomon
as reinhardtii 25.42/6.24 28.4/7.0 8 89 0.91
Desconhecidas
03 Unknown protein 18 gi|205830697 Pseudotsuga
menziesii 1.3/5.80 17.0/4.4 91 52 1.19 1.24
05 Predicted protein gi|224123396 Populus
trichocarpa 19.54/4.98 21.1/4.5 7 96 2.59
13 PREDICTED: TRM112-like protein
At1g78190 gi|225452543 Vitis vinifera 13.95/5.16 16.6/4.8 27 61 1.30 1.52
14 Predicted protein gi|224123436 Populus
trichocarpa 20.27/5.83 17.4/4.7 6 55 1.36 1.38 1.66
24 Hypothetical protein VITISV_010210 gi|147820236 Vitis vinifera 58.60/5.60 94.2/5.8 4 89 0.48
(a) Todos os valores de escores listados excedem o valor do escore limite, calculado pelo Mascot e, portanto, indicam extensa similaridade com base nas sequencias
consultadas no NCBI.
(b) O índice de variação corresponde ao valor obtido a partir da divisão da média de intensidade de cada spot no gel inoculado pela respectiva intensidade no gel do
grupo controle.

63
Uma vez que a localização celular fornece importantes informações sobre a função
fisiológica das proteínas, a predição desta localização também foi realizada. Muitas das
proteínas identificadas pertencem a diferentes compartimentos celulares, entre os quais,
citoplasma (17), cloroplasto (7), mitocôndria (3), membrana celular (4), retículo
endoplasmático (1) e região extracelular (1). No conjunto de proteínas diferencialmente
expressas, somente 3 delas apresentam localização celular desconhecida.
Na figura 10, a classificação das proteínas identificadas é indicada com relação ao
processo biológico que desempenham, assim como o componente celular a que pertencem.
Nesta figura, é colocado em destaque o número de proteínas cuja alteração está relacionada ao
aumento (“up-regulated”) ou diminuição da expressão (“down-regulated”) em virtude da
condição de infecção com o patógeno.
Nesse estudo, várias classes funcionais de proteínas parecem ser responsivas à
infecção por L. theobromae indicando reprogramação de eventos celulares no clone BRS 226.
A diversidade de proteínas envolvidas com metabolismo e geração de energia, responsivas ao
estresse e associadas à defesa, assim como relacionadas ao enovelamento de proteínas e
sinalização celular sugere certa importância às funções desses processos durante a resposta ao
ataque do patógeno.
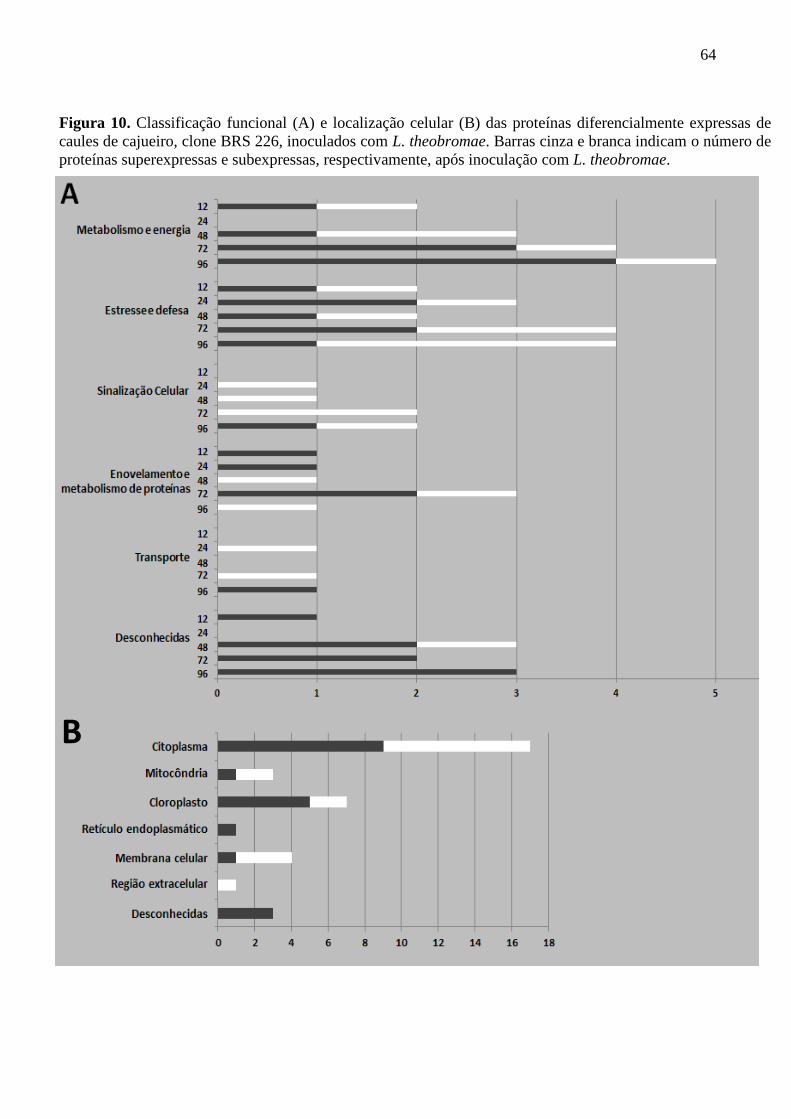
64
Figura 10. Classificação funcional (A) e localização celular (B) das proteínas diferencialmente expressas de
caules de cajueiro, clone BRS 226, inoculados com L. theobromae. Barras cinza e branca indicam o número de
proteínas superexpressas e subexpressas, respectivamente, após inoculação com L. theobromae.
B

65
De um modo geral, mudanças na expressão de proteínas metabólicas e relacionadas à
geração de energia correspondem à categoria com maior número de proteínas identificadas
neste estudo. Essa aparente reprogramação parece evidenciar os custos da condição de
infecção, além de mostrar a importância destes processos para o crescimento e
desenvolvimento da planta. Nesses processos biológicos, estão incluídas proteínas do
complexo NADH desidrogenase (spot 11), superexpressas sob condição de infecção, além de
enzimas envolvidas com o metabolismo de carboidratos, como frutoquinase (spot 18), alfa
glucano fosforilase (spot 32), frutose bifosfato aldolase (spot 35), ou com vias fermentativas
representadas pela atuação da piruvato descarbolixase (spots 20, 25) e álcool desidrogenase
(spot 37). Enzimas associadas ao metabolismo de aminioácidos também foram identificadas,
assim como, uma chalcona isomerase (spot 17), responsável pela biossítense de flavonóides.
Um segundo grande grupo de proteínas com expressão alterada é responsivo a
condição de estresse. Nesta categoria, estão 2 proteínas de choque térmico [do Inglês, Heat
shock proteins (HSPs)] com massas moleculares de 70 (spot 9) e 20 kDa (spot 21), além de
proteínas de detoxificação, como a glutationa-s-transferase (spot 27). A ocorrência de
explosão oxidativa como resposta a inoculação do cajueiro com L. theobromae é sugerida
pela observação de que houve superexpresão de enzimas associadas à homeostase redox,
dentre elas, a dissulfeto isomerase (spot 7) e 2-cis peroxirredoxina (spot 10).
Interessantemente, proteínas clássicas de defesa parecem não sofrer reprogramação durante a
infecção. Somente um membro está incluído nesta categoria e foi identificado como uma
proteína tipo germina (spot 33), tendo sua expressão reduzida em virtude da presença de L.
theobromae.
Com relação às vias de sinalização celular, foram encontradas 3 proteínas com
expressão alterada, dentre as quais, uma proteína 14-3-3 (spot 6) superexpressa em 48 e 96
horas após inoculação, e duas anexinas(spots 31 e 34) que são proteínas ligantes a Ca2+
e cuja
expressão foi reduzida sob infecção em diferentes tempos (24, 72 e 96 HAI). O metabolismo
de proteínas foi alterado por componentes de rotas de degradação, identificados pelas
subunidades alfa (spot 26) e beta (spot 22) do proteassoma 26S, além do fator de elongação
da tradução (spot 40). Em contrapartida, a atuação funcional no transporte de componentes no
ambiente celular parece ser reprimida com o desafio do patógeno, observado pela
subexpressão de uma small GTPase tipo Ran (spot 30) e lipocalina (spot 29).

66
4.2.1 Rede funcional de interação das proteínas diferencialmente expressas
As proteínas não atuam nos processos celulares como entidades únicas, mas formando
uma rede funcional baseada na interação proteína-proteína. Para determinar como a infecção
de plantas do clone BRS 226 com L. theobromae afeta a reprogramação de proteínas e,
consequentemente, as funções celulares, as proteínas diferencialmente identificadas nesse
estudo foram analisadas por meio do banco de dados disponível no programa STRING.
Parceiros funcionais destas proteínas foram preditos e, com base nisso, a ligação dos
processos celulares foi evidenciada (Figura 11). As abreviações referentes aos nomes
específicos das proteínas mostradas na rede são indicadas na tabela 4, com destaque para 5
parceiros funcionais propostos com base na informação disponível no programa.
Resumidamente, algumas das interações propostas incluem a relação entre a GTPase
tipo Ran (spot 30, RAN-1) com parceiros representados por subunidades do proteassoma
(PBG1, PAG1, RPT2a, RPN10) incluindo a subunidade beta (spot 22, PBD2) e alfa (spot 26,
At5g35590) identificadas neste trabalho. Uma ligação entre estas proteínas com a chaperona
Hsp70 (spot 9, HSP70) foi proposto, que por sua vez, interage com a dissulfeto isomerase
(spot 7, ATPDIL2-2). Interação entre proteínas envolvidas com o metabolismo fermentativo
foram observadas para as enzimas álcool desidrigenase (spot 37, HOT 5) e piruvato
descarboxilase (spots 20 e 25, PCD1, AT5GO1320). As duas anexinas (spots 31 e 34,
ANNATA1, ANNATA2) diferencialmente expressas apresentaram várias evidências de
interação, decorrentes, por exemplo, de dados experimentais e disponíveis em banco de
dados, vizinhança e co-expressão gênica.
Por fim, um último conjunto de interação foi observado entre a NADH desidrogenase
(spot 11, AT5G11770, TYKY), formato desidrogenase (spot 36, FDH), 1,2-dihidroxi-3-ceto-
5-metiltiopentano dioxigenase 2 (spot 15, ATARD2), serina hidroximetiltransferase (spot 38,
SHM3) e o fator de elongação da tradução (spot 40, AT1G45332).

67
Figura 11. Rede de interação proteína-proteína analisada pelo software STRING. Linhas de diferentes
cores representam diferentes evidências de associação: verde, evidência de vizinhança; vermelha,
evidência de fusão; azul, evidência de coocorrência; preta, evidência de coexpressão; roxa, evidência
experimental; azul claro, evidência em banco de dados; amarela, evidência em mineração de texto;
roxa clara, evidência de homologia.
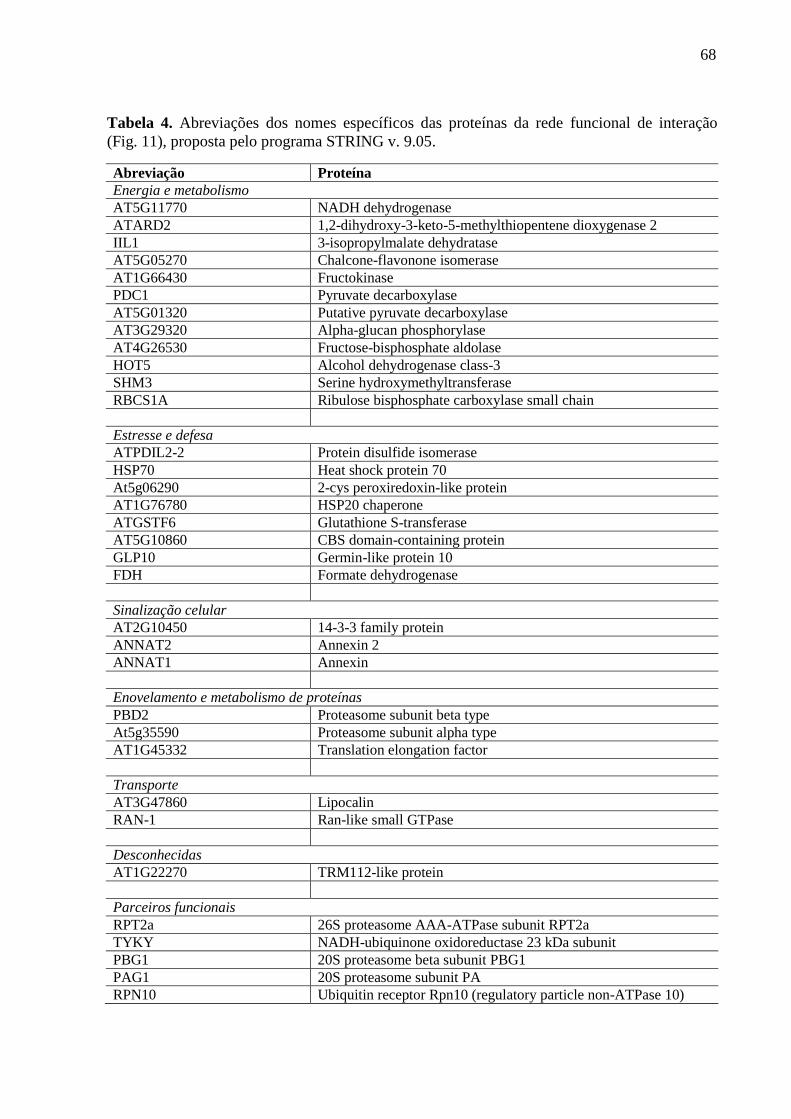
68
Tabela 4. Abreviações dos nomes específicos das proteínas da rede funcional de interação
(Fig. 11), proposta pelo programa STRING v. 9.05.
Abreviação Proteína
Energia e metabolismo
AT5G11770 NADH dehydrogenase
ATARD2 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase 2
IIL1 3-isopropylmalate dehydratase
AT5G05270 Chalcone-flavonone isomerase
AT1G66430 Fructokinase
PDC1 Pyruvate decarboxylase
AT5G01320 Putative pyruvate decarboxylase
AT3G29320 Alpha-glucan phosphorylase
AT4G26530 Fructose-bisphosphate aldolase
HOT5 Alcohol dehydrogenase class-3
SHM3 Serine hydroxymethyltransferase
RBCS1A Ribulose bisphosphate carboxylase small chain
Estresse e defesa
ATPDIL2-2 Protein disulfide isomerase
HSP70 Heat shock protein 70
At5g06290 2-cys peroxiredoxin-like protein
AT1G76780 HSP20 chaperone
ATGSTF6 Glutathione S-transferase
AT5G10860 CBS domain-containing protein
GLP10 Germin-like protein 10
FDH Formate dehydrogenase
Sinalização celular
AT2G10450 14-3-3 family protein
ANNAT2 Annexin 2
ANNAT1 Annexin
Enovelamento e metabolismo de proteínas
PBD2 Proteasome subunit beta type
At5g35590 Proteasome subunit alpha type
AT1G45332 Translation elongation factor
Transporte
AT3G47860 Lipocalin
RAN-1 Ran-like small GTPase
Desconhecidas
AT1G22270 TRM112-like protein
Parceiros funcionais
RPT2a 26S proteasome AAA-ATPase subunit RPT2a
TYKY NADH-ubiquinone oxidoreductase 23 kDa subunit
PBG1 20S proteasome beta subunit PBG1
PAG1 20S proteasome subunit PA
RPN10 Ubiquitin receptor Rpn10 (regulatory particle non-ATPase 10)

69
4.3 Alteração no proteoma de caules de plantas de cajueiro, crescidas em campo,
resistentes e suscetíveis à resinose
Os mecanismos moleculares responsáveis pela suscetibilidade ou resistência à doença,
também foram examinados estudando plantas de cajueiro cultivadas em um pomar comercial
localizado no município de Pio IX (PI), que tem como característica a alta pressão do
patógeno, L. theobromae, evidenciada pela grande incidência de resinose na região. Plantas de
cajueiro dessa localidade foram avaliadas com relação ao desenvolvimento de sintomas e,
então, classificadas de acordo com uma escala de severidade em resistentes (escala 0) e
suscetíveis (escala 2) à resinose.
Extratos proteicos do tecido caulinar de plantas pertencentes a estes grupos foram
obtidos e usados para gerar os mapas proteicos representativos de cada condição. Na figura 12
o conteúdo de proteínas solúveis extraídas destas amostras é apresentado e, conforme
observado, não foi possível detectar diferença estatística (p ≤ 0,05) na quantidade de proteínas
solúveis entre os respectivos grupos de plantas.
Nesse estudo, para cada condição experimental (resistente e suscetível), foram obtidos
7 mapas bidimensionais, totalizando 14 géis escaneados e analisados quanto à ocorrência de
alteração significativa na intensidade dos spots. A média de spots totais quantificados em cada
condição foi de 522 ± 9 e 516 ± 8 para o proteoma de plantas resistentes e suscetíveis à
resinose, respectivamente. Os perfis bidimensionais destas plantas não apresentaram interação
significativa entre o número total de spots e a condição de resistência ou suscetibilidade à
doença (Figura 13).
A comparação do proteoma do caule destas plantas revelou que 54 proteínas tiveram a
expressão reprogramada associada à condição de resistência ou suscetibilidade. Dos 27 spots
que foram mais abundantes no grupo resistente, 19 foram identificados com sucesso, ao passo
que foram identificados 18 dos 27 spots cuja maior abundância reside em plantas suscetíveis.
Como observado, plantas classificadas como resistentes e suscetíveis exibiram número igual
de proteínas com expressão alterada no extrato caulinar, contudo, um total de 17 proteínas não
foram identificadas por MS/MS, com nível significativo de confiança. Perfis representativos
de cada grupo de estudo são indicados na figura 14, juntamente com os spots cuja abundância
relativa foi aumentada em virtude da condição de resistência ou suscetibilidade à doença.

70
Figura 12. Teor de proteínas solúveis extraídas de caules de plantas de cajueiro, crescidas em
campo, resistentes e suscetíveis à resinose. As barras indicam desvio padrão. Letras iguais não
diferem significativamente pelo teste de Tukey a 5%.
Figura 13. Número total de spots proteicos, visualizados por 2D-PAGE, de extratos de caules
de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à resinose. As barras
indicam desvio padrão. Letras iguais não diferem significativamente pelo teste de Tukey a
5%.
A tabela 5 lista as proteínas identificadas por MS/MS agrupadas de acordo o processo
biológico que desempenham. Nessa tabela, os spots proteicos cujos números correspondentes
são precedidos da letra “R” representam proteínas mais abundantes no proteoma caulinar da
amostra resistente em comparação com a suscetível. Por sua vez, para os spots precedidos da
letra “S”, o inverso é aplicado, ou seja, eles são mais abundantes no grupo suscetível. Esses
dados podem ser observados pelo índice de variação da expressão relativa da proteína,
também indicado na tabela, e calculado a partir da comparação do valor médio da intensidade
de cada spot na amostra resistente em comparação ao valor obtido na amostra suscetível.
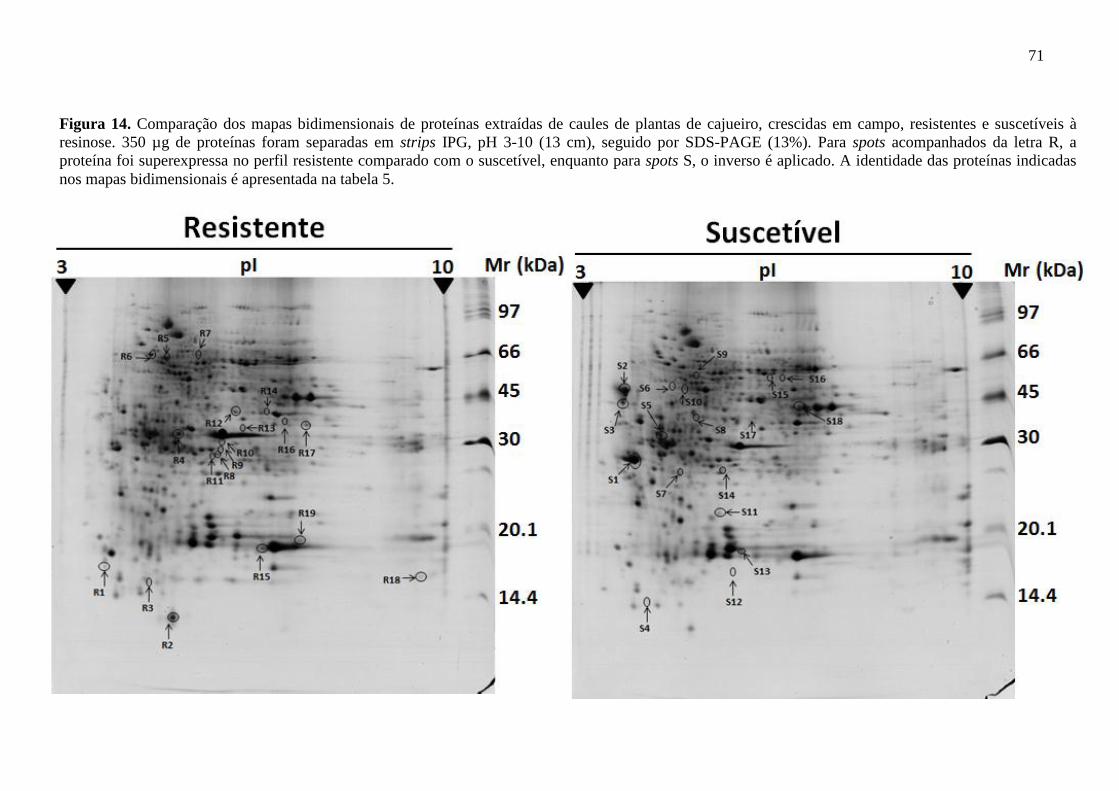
71
Figura 14. Comparação dos mapas bidimensionais de proteínas extraídas de caules de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à
resinose. 350 µg de proteínas foram separadas em strips IPG, pH 3-10 (13 cm), seguido por SDS-PAGE (13%). Para spots acompanhados da letra R, a
proteína foi superexpressa no perfil resistente comparado com o suscetível, enquanto para spots S, o inverso é aplicado. A identidade das proteínas indicadas
nos mapas bidimensionais é apresentada na tabela 5.

72
Tabela 5. Identidade das proteínas diferencialmente expressas em caules de plantas de cajueiro, crescidas em campo, classificadas como
resistentes e suscetíveis à resinose.
N° do spot
(a) Proteína
Acesso
(NCBI)
Organismo de
Referência
Mr/pI
Teórico
Mr/pI
Experimental
Cobertura
(%)
Score
Índice de
variação (b)
Energia e metabolism
S6 Transaldolase gi|217795376 Dimocarpus longan 47.88/6.34 53.05/4.29 7 184 0.66
S7 Adenine phosphoribosyl Transferase gi|1402894 Arabidopsis thaliana 21.01/5.69 21.16/4.46 7 58 0.34
S10 Transaldolase gi|217795376 Dimocarpus longan 47.88/6.34 52.94/4.52 7 164 0.47
S11 Asparaginase gi|735918 Arabidopsis thaliana 33.05/5.41 18.24/5.26 5 71 0.48
S17 PREDICTED: 6-Phosphogluconate dehydrogenase,
decarboxylating 3-like gi|502102108 Cicer arietinum 54.27/5.88 58.32/6.22 6 77 0.42
S18 Glyceraldehyde 3-phosphate dehydrogenase gi|357463093 Medicago truncatula 37.03/6.97 44.39/6.73 30 463 0.20
R7 D-3-phosphoglycerate dehydrogenase isoform 1 gi|508706453 Theobroma cacao 63.52/8.12 69.00/5.15 8 223 2.17
R4 Chloroplast photosynthetic water oxidation complex 33kDa
subunit precursor gi|152143640 Morus nigra 28.24/5.48 32.76/4.77 16 154 3.08
R11 PREDICTED: triosephosphate isomerase, chloroplastic-like gi|502111535 Cicer arietinum 33.57/6.36 23.25/5.47 23 284 2.38
R12 Fructose-bisphosphate aldolase 3 gi|15226185 Arabidopsis thaliana 42.30/8.19 38.19/5.89 7 87 1.79
R18 Chloroplast ribulose-1,5-bisphosphate
carboxylase/oxygenase small gi|76574181 Musa balbisiana 13.58/8.36 15.18/9.64 19 78 2.70
Estresse e defesa
S2 PREDICTED: peroxidase 2-like gi|449467745 Cucumis sativus 37.22/5.64 51.93/3.37 6 53 0.22
S4 PREDICTED: thioredoxin-like protein CXXS1 gi|225461146 Vitis vinifera 14.17/5.15 14.78/3.99 7 51 0.34
S13 HSP20-like chaperones superfamily protein gi|508786804 Theobroma cacao 17.96/5.57 16.61/5.69 25 162 0.66
S16 Cytosolic glutathione reductase gi|94494355 Phaseolus vulgaris 54.72/6.32 61.15/6.66 7 144 0.56
R9 Ascorbate peroxidase gi|39939493 Pinus pinaster 27.25/5.40 26.92/5.59 12 149 1.84
R14 PREDICTED: serpin-ZX gi|359492076 Vitis vinifera 48.17/8.67 39.08/6.50 4 87 1.68
R13 PREDICTED: S-formylglutathione hydrolase-like gi|356500537 Glycine max 32.10/6.55 33.68/6.05 8 86 1.55
R15 Class I small heat shock protein 20.1 gi|349591294 Solanum lycopersicum 17.62/5.82 16.25/6.51 14 155 3.60
R19 Heat-shock protein, putative gi|255558876 Ricinus communis 17.50/5.34 16.48/7.22 9 90 2.29
Sinalização cellular
S5 PREDICTED: 14-3-3-like protein D-like gi|449438105 Cucumis sativus 29.37/4.76 34.63/4.07 17 173 0.22
S12 Protein kinase C inhibitor-like protein gi|7594527 Arabidopsis thaliana 14.09/6.18 15.7/5.57 13 148 0.53
R3 PREDICTED: probable calcium-binding protein CML13-
like gi|449460417 Cucumis sativus 16.72/4.82 14.89/4.23 36 243 3.26
R16 Annexin, partial gi|16973318 Gossypium hirsutum 14.33/8.70 35.57/6.84 17 108 1.98
R17 Annexin gi|429326384 Populus tomentosa 36.11/6.18 34.08/7.25 8 177 1.36
Metabolismo de proteínas
S14 PREDICTED: proteasome subunit beta type-1-like gi|460373244 Solanum lycopersicum 24.58/5.93 22.24/5.3 8 113 0.27
R1 60S acidic ribosomal protein P gi|730583 Parthenium argentatum 11.49/4.13 15.44/3.46 10 64 2.28
R8 PREDICTED: proteasome subunit beta type-1 gi|225453909 Vitis vinifera 24.61/6.88 22.08/5.68 17 135 1.31
Estrutura cellular
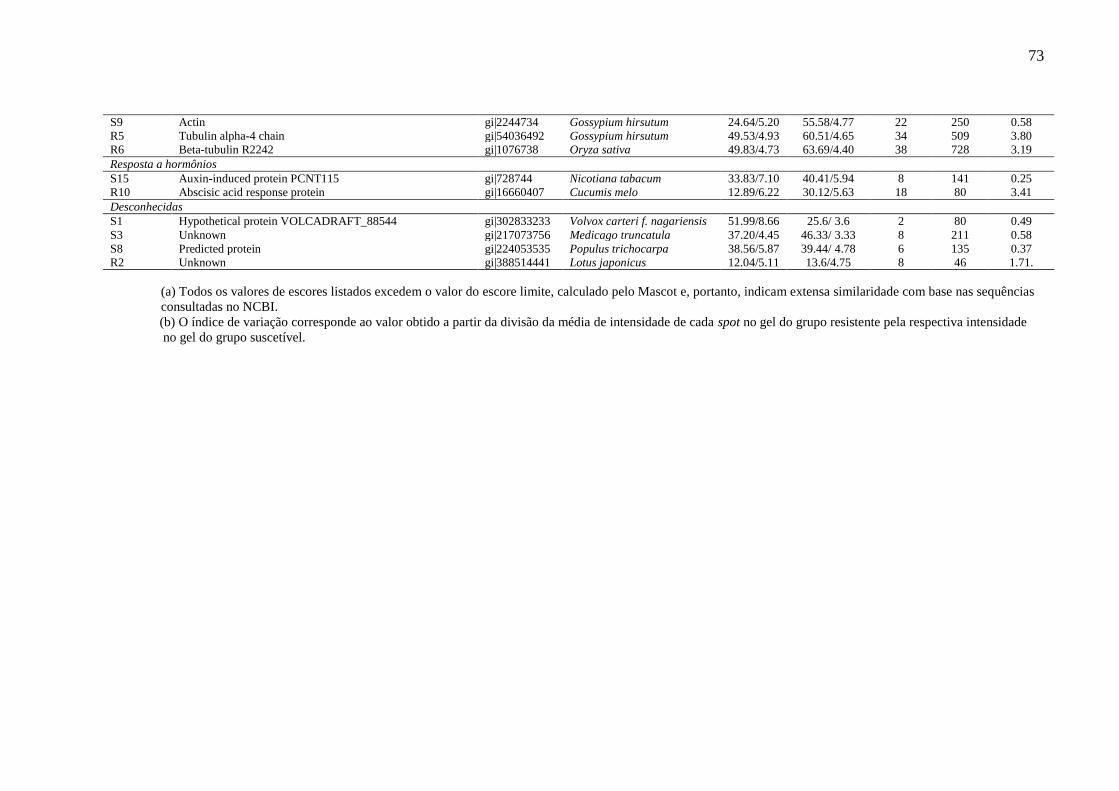
73
S9 Actin gi|2244734 Gossypium hirsutum 24.64/5.20 55.58/4.77 22 250 0.58
R5 Tubulin alpha-4 chain gi|54036492 Gossypium hirsutum 49.53/4.93 60.51/4.65 34 509 3.80
R6 Beta-tubulin R2242 gi|1076738 Oryza sativa 49.83/4.73 63.69/4.40 38 728 3.19
Resposta a hormônios
S15 Auxin-induced protein PCNT115 gi|728744 Nicotiana tabacum 33.83/7.10 40.41/5.94 8 141 0.25
R10 Abscisic acid response protein gi|16660407 Cucumis melo 12.89/6.22 30.12/5.63 18 80 3.41
Desconhecidas
S1 Hypothetical protein VOLCADRAFT_88544 gi|302833233 Volvox carteri f. nagariensis 51.99/8.66 25.6/ 3.6 2 80 0.49
S3 Unknown gi|217073756 Medicago truncatula 37.20/4.45 46.33/ 3.33 8 211 0.58
S8 Predicted protein gi|224053535 Populus trichocarpa 38.56/5.87 39.44/ 4.78 6 135 0.37
R2 Unknown gi|388514441 Lotus japonicus 12.04/5.11 13.6/4.75 8 46 1.71.
(a) Todos os valores de escores listados excedem o valor do escore limite, calculado pelo Mascot e, portanto, indicam extensa similaridade com base nas sequências
consultadas no NCBI.
(b) O índice de variação corresponde ao valor obtido a partir da divisão da média de intensidade de cada spot no gel do grupo resistente pela respectiva intensidade
no gel do grupo suscetível.

74
No conjunto das 37 proteínas com expressão diferencial em caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis, a maioria delas pertence à categoria
funcional de metabolismo e energia (11), enquanto o restante foi classificado como de
estresse e defesa (9), sinalização celular (5), metabolismo de proteínas (3), estrutura celular
(3) e proteínas responsivas a hormônios (2). Para 4 proteínas, o processo celular que
desempenham não está claro, sendo, portanto, classificadas na categoria de desconhecidas.
Na figura 15A, as proteínas diferencialmente reprogramadas em caules de plantas de cajueiro,
em virtude da condição de resistência/suscetibilidade, foram organizadas colocando em
destaque o número de proteínas categorizadas em processos biológicos que, também, tiveram
suas expressões alteradas em plantas do clone BRS 226. O número de spots
superexpressos/subexpressos é evidenciado comparando plantas resistentes em relação ao
grupo suscetível, em condições de campo (Figura 15B).
As proteínas identificadas neste estudo participam de vários processos metabólicos,
dentre os quais, vias anabólicas de aminoácidos (spots S11 e R7) ou catabolismo de
carboidratos (spots S18, R11, R12), além da geração de poder redutor (spots S6, S10, S17)
para rotas de biossíntese. Uma forte tendência à reprogramação da expressão de proteínas
responsivas ao estresse foi observada entre os grupos de plantas analisados, de modo que,
houve uma considerável expressão de enzimas removedoras de peróxido de hidrogênio ou
associadas à regeneração de substratos usados na explosão oxidativa (spots S2, S4, S16 e R9).
Somente uma proteína relacionada à defesa, a serpina (spot R14), foi superexpressa em
plantas. A atividade no controle da proteólise sugere papel de defesa contra insetos e outros
patógenos para essa proteína identificada.
Proteínas envolvidas com rotas de sinalização celular englobam proteínas ligantes a
Ca2+
(spots R3, R16 e R 17), induzidas no grupo resistente, além de outras duas proteínas que
foram mais expressas nas plantas suscetíveis (spots S5 e S12). Das proteínas associadas com a
manutenção da estrutura celular por envolvimento com componentes do citoesqueleto, 2 das 3
identificadas tiveram expressão aumentada em plantas resistentes (spots R5 e R6). Eventos de
síntese e degradação envolvidos no metabolismo de proteínas também foram influenciados
pela condição de infecção nas plantas de cajueiro. Nessa categoria, proteínas constituintes do
ribossomo (spot R1) e subunidades do proteassoma (spots S14 e R8) parecem ser
diferencialmente reguladas. O envolvimento de hormônios neste contexto é indicado por
mudanças no perfil de proteínas responsivas a auxina (spot S15) e ácido abscísico (spot R10).

75
Figura 15. Categorias funcionais das proteínas diferencialmente reprogramadas em caules de plantas de
cajueiro, crescidas em campo, associadas à resistência/suscetibilidade. A) O número de proteínas
identificadas em cada função biológica é indicado, dando ênfase às proteínas pertencentes a categorias
também identificadas em plantas do clone BRS 226 infectadas com L. theobromae. B) Proteínas
superexpressas/subexpressas são mostradas comparando mudanças na expressão observada em plantas
resistentes com relação às plantas suscetíveis.
Proteínas de caules de plantas de cajueiro, crescidas em campo, categorizadas em processos biológicos também identificados no clone BRS 226
4
22
33
28
1111 99
33 55
A
Energia e metabolismo
Estresse e defesa
Sinalização celular
Metabolismo de proteínas
Estrutura celular
Resposta a hormônios
Desconhecidas
B

76
4.4 Determinação das atividades das enzimas superóxido dismutase, ascobarto
peroxidase, peroxidase do guaiacol, fenilalanina amônia liase e β-1,3-glucanase de caules
de plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à resinose
O ensaio da atividade de enzimas envolvidas com a patogênese, associadas ao estresse
oxidativo e metabolismo secundário foi realizado com o objetivo de comparar diferenças nas
respostas bioquímicas entre os grupos de plantas categorizadas como resistentes e suscetíveis,
em condições de campo.
Na categoria de proteínas responsivas ao estresse oxidativo foram avaliadas as
enzimas superóxido dismutase (SOD), peroxidase do guaiacol (POX) e ascobarto peroxidase
(APX), enquanto, que em relação às proteínas relacionadas à patogênese (PR-proteínas)
apenas a atividade da enzima β-1,3-glucanase (PR-2) foi avaliada. A fenilalanina amônia liase
(PAL), enzima chave no metabolismo dos fenilpropanoídes, foi analisada entre os grupos de
estudo como representante de enzimas envolvidas com o metabolismo secundário e cujo
produto da atividade é, em geral, induzido sob infecção por patógenos.
Os dados resultantes do ensaio da atividade das enzimas citadas são mostrados nas
Figuras 16, 17 e 18. A atividade da enzima APX, bem como da POX, não foi detectada sob as
condições empregadas, indicando possível desnaturação. Muito embora as demais proteínas
apresentem papel direto ou indireto na defesa vegetal, não foi possível constatar diferenças
significativas (p≤ 0,05) nas atividades destas enzimas entre plantas resistentes e suscetíveis,
cultivadas em condições de campo. Estes resultados são coerentes com aqueles obtidos a
partir da abordagem proteômica, uma vez que proteínas como SOD, PAL e β-1,3-glucanase
não foram identificadas dentre aquelas diferencialmente expressas em caules de plantas
resistentes e suscetíveis à resinose, confirmando a ausência de reprogramação da expressão,
tendo em vista que todas as proteínas usadas para determinação da atividade enzimática foram
previamente identificadas nos mapas bidimensionais de proteínas extraídas de caules de
cajueiro por análise proteômica descritiva realizada pelo nosso grupo de pesquisa (GONDIM,
2010). A ausência de diferença significativa na atividade dessas proteínas, tanto em plantas
resistentes como suscetíveis, somada a não identificação delas no conjunto de proteínas
analisadas por proteômica diferencial, fornece evidências de que estas enzimas parecem não
participar de respostas moleculares em plantas de cajueiro infectadas com L. theobromae,
uma vez que elas também não fazem parte das proteínas responsivas no clone BRS 226
inoculado com o fitopatógeno.

77
Figura 16. Atividade da enzima superóxido dismutase (UA/g-1
MF) de caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à resinose. Barras indicam desvio
padrão. Letras diferentes indicam diferença estatística entre as médias pelo teste de Tukey
(p≤0,05).
Figura 17. Atividade da enzima fenilalanina amônia liase (nmol CA/g-1
MF) de caules de
plantas de cajueiro, crescidas em campo, resistentes e suscetíveis à resinose. Barras indicam
desvio padrão. Letras diferentes indicam diferença estatística entre as médias pelo teste de
Tukey (p≤0,05).
a a
a
a
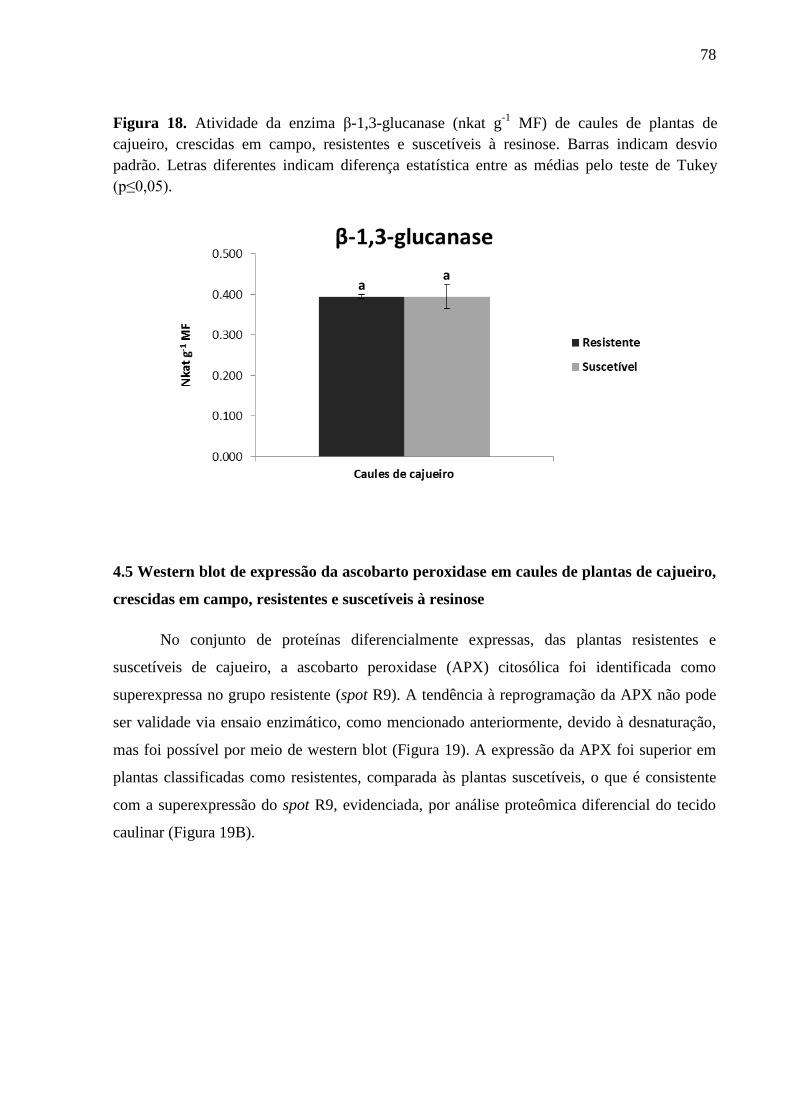
78
Figura 18. Atividade da enzima β-1,3-glucanase (nkat g-1
MF) de caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à resinose. Barras indicam desvio
padrão. Letras diferentes indicam diferença estatística entre as médias pelo teste de Tukey
(p≤0,05).
4.5 Western blot de expressão da ascobarto peroxidase em caules de plantas de cajueiro,
crescidas em campo, resistentes e suscetíveis à resinose
No conjunto de proteínas diferencialmente expressas, das plantas resistentes e
suscetíveis de cajueiro, a ascobarto peroxidase (APX) citosólica foi identificada como
superexpressa no grupo resistente (spot R9). A tendência à reprogramação da APX não pode
ser validade via ensaio enzimático, como mencionado anteriormente, devido à desnaturação,
mas foi possível por meio de western blot (Figura 19). A expressão da APX foi superior em
plantas classificadas como resistentes, comparada às plantas suscetíveis, o que é consistente
com a superexpressão do spot R9, evidenciada, por análise proteômica diferencial do tecido
caulinar (Figura 19B).
a a

79
Figura 19. Expressão diferencial da ascorbato peroxidase (APX) em caules de plantas de
cajueiro, crescidas em campo, resistentes e suscetíveis à resinose. (A) As proteínas foram
separados por SDS-PAGE e imonudetectados com anti-corpo anti-APX citosólica. (B)
Abundância relativa calculada a partir da eletroforese bidimensional.

80
5. DISCUSSÃO
5.1 Interação incompatível entre o clone de cajueiro BRS 226 e o fungo L. theobromae
A inoculação artificial dos caules de cajueiro anão, clones BRS 226 (resistente) e CCP
76 (suscetível), com L. theobromae, resultou no aparecimento de lesão interna, no lenho, ao
redor do ponto de aplicação do inóculo. A análise comparativa da área de escurecimento
interno entre os clones, realizada em 72 e 96 HAI, mostrou que, embora ambos os clones
tenham desenvolvido os sintomas de lesão necrótica no tecido caulinar, o clone BRS 226,
com 96 HAI, manteve o comprimento da necrose similar aquele quantificado nas plantas
controles com 72 HAI (Figura 7), não tendo, portanto, progredido, como observado para o
clone CCP 76, que com 96 HAI apresentou comprimento significativamente (p ≤ 0,05) maior
em comparação com o clone resistente (Tabela 1).
Uma vez que L. theobromae é um patógeno com hábitos necrotróficos (THAKKAR,
2004), a ausência de expansão da necrose define a interação desse patossistema como
incompatível (MUKHERJEE et al., 2010). Esta característica é coerente com outros estudos
relacionados ao monitoramento da condição de resistência desse clone em pomares
comerciais com alta pressão do patógeno (PAIVA et al., 2008; CYSNE et al., 2010).
Portanto, o clone BRS 226 parece não sofrer alterações fisiológicas drásticas, durante a
infecção, que prejudiquem a produtividade do pomar nas condições avaliadas, apresentando
um modelo de resistência durável no decorrer do tempo, com a manutenção do rendimento da
castanha semelhante ao clone comercial CCP 76, que é considerado suscetível ao
desenvolvimento da doença (CARDOSO et al., 2006).
A manutenção da homeostase vegetal diante da presença do patógeno é uma
característica relevante quando se trata da resinose, uma vez que existe uma aparente
tendência em plantas de cajueiro à ocorrência endofítica de L. theobromae, recentemente
registrada pela presença de hifas em vasos do xilema de plantas sem sintomas aparentes da
doença (MUNIZ et al., 2011), condição biológica que outros membros da família
Botryosphaeriaceae também apresentam (SLIPPERS e WINGFIELD, 2007). Assim, plantas
contendo o patógeno nos tecidos internos desenvolvem sintomas da doença somente após um
sinal externo, caracterizado em geral por estresse biótico ou abiótico.

81
No processo de infecção com L. theobromae, durante o estágio inicial, há assimilação
pelo patógeno de componentes proteicos, pectinas, amido e açúcares livres (ENCINAS e
DANIELL, 1976). Em seguida, subsequente degradação dos raios parenquimáticos e parede
celular são observadas, o que tem sugerido a ação sinérgica de um conjunto de enzimas
hidrolíticas, dentre as quais, glucanases, celobiohidrolases, celulases, xilanases e xilosidases
(MUNIZ et al., 2011). O uso de estratégias baseadas na atuação de enzimas líticas,
combinadas com a produção de altos níveis de espécies reativas de oxigênio, tem sido
relatado em fungos necrótroficos, após a entrada nos tecidos internos da planta, conduzindo a
um estado subsequente de necrose (LALUK e MENGISTE, 2010). No caso de L.
theobromae, a intensidade da capacidade de degradação é diferenciada conforme a
agressividade do isolado (LIMA et al., 2012).
A diversidade de estratégias de virulência empregadas por espécies de patógenos
necrotróficos, como L. theobromae, correspondem por sua vez, a uma variedade de
mecanismos de reposta imune acionados no hospedeiro. Quando eficaz, o sistema
imunológico da planta é capaz de “desarmar” o arsenal patogênico do invasor ou atenuar seus
efeitos, restringindo a entrada e o desenvolvimento de sintomas da doença (MENGISTE et
al., 2012).
Muito embora, outros clones de cajueiro tenham apresentado potencial de resistência à
resinose (CARDOSO et al., 2007), o desempenho produtivo do clone BRS 226 o torna padrão
comercial de resistência, além de importante alternativa de exploração em programas de
melhoramento genético (MARTINS et al., 2011) e para estudos baseados nos mecanismos
moleculares associados a esta condição. Contudo, a existência de apenas um clone (BRS 226)
com características agronômicas e de resistência durável à resinose, mostra a necessidade de
aumento da base genética do cajueiro (MARTINS et al., 2011), uma vez que o surgimento de
novas raças do fitopatógeno representam uma ameaça a resistência adquirida por novas
variedades ou híbridos (ALZATE-MARIN et al., 2005), o que tem contribuído para a seleção
de novos genótipos (MOREIRA et al., 2013; PAIVA et al., 2008).
Baseado na ausência de informações publicadas com relação aos mecanismos
bioquímicos e fisiológicos do cajueiro infectado com L. theobromae, associado ao fato de que
a habilidade das plantas de se defenderem do ataque de patógenos tem como base alterações
em vias metabólicas chaves, em decorrência da reprogramação da expressão de certas
proteínas (AFROZ et al., 2011), o estudo da identidade dessas moléculas, ou seja, do

82
proteoma do caule de cajueiro, sítio de infecção do patógeno, fornece um conjunto de
informações que ajudam a entender como essas proteínas contribuem, efetivamente, com os
mecanismos relacionados à interação cajueiro-L. theobromae.
5.2 Reprogramação da expressão de proteínas em caules de plantas de cajueiro
infectadas com L. theobromae
Os mecanismos moleculares associados à resistência e suscetibilidade foram estudados
por meio da identificação de proteínas cuja expressão foi alterada em caules de cajueiro sob
infecção. A análise da abundância relativa de proteínas presentes no proteoma caulinar foi
realizada utilizando eletroforese bidimensional como abordagem, uma vez que o uso desta
técnica tem sido empregado com sucesso no monitoramento das respostas moleculares do
hospedeiro ao ataque de patógenos (MANDELC et al., 2013). Para alcançar tal propósito, no
presente estudo, dois cenários de investigação foram abordados. No primeiro deles, foram
avaliadas possíveis alterações no proteoma, e suas consequências para a resistência, dos
caules de plantas de cajueiro anão, clone BRS 226, altamente resistente à resinose, em tempos
iniciais após a infecção artificial com L. theobromae (12, 24, 48, 72 e 96 HAI). No segundo
cenário, o proteoma do caule foi estudado em plantas de cajueiro híbridas, crescidas a partir
de sementes resultantes de polinização aberta do BRS 226, com dois anos de idade após
plantio em condições de campo, onde há alta pressão do patógeno, e fazendo parte de grupos
classificados como resistentes e suscetíveis à resinose, com base na observação dos sintomas
característicos da doença.
Para ambos os cenários de estudo, o conteúdo de proteínas solúveis extraídas do tecido
caulinar não apresentou diferenças quantitativamente significantes, entre o grupo controle e
inoculado (Figura 8), no caso do primeiro cenário, e os outros dois grupos comparados entre
si, do segundo cenário, um com sintomas e outro livre da doença, na condição de crescimento
em campo (Figura 12). Além disso, o número de spots proteicos totais visualizados nos mapas
bidimensionais, desses grupos, não mostrou diferenças relacionadas à condição de infecção
com o fungo, no clone BRS 226 (Tabela 2), ou a resistência/suscetibilidade observada no
campo (Figura 13). Este resultado é coerente com a ausência de proteínas exclusivas,
mostrando uma tendência a sútil reprogramação da expressão de proteínas que refletem a
composição do proteoma caulinar compartilhado entre as amostras diferenciais. A eficiência
das repostas de expressão diferencial dessas proteínas parece, portanto, estar relacionada a

83
alterações fisiológicas que condicionam mudanças no conteúdo de proteínas pré-existentes,
que por sua vez, apresentam interações funcionais entre si (Figura 11).
Por outro lado, a limitação dos métodos de extração no que se refere à solubilização de
todas as proteínas presentes nas amostras estudadas (MALDONADO et al., 2008), não
permite descartar a hipótese da existência de proteínas exclusivas de baixa abundância nas
condições estudadas que, no entanto, podem não ser suficientemente extraídas em
quantidades que permitem a sua visualização no mapa bidimensional. Entretanto, vale
salientar que do conjunto de protocolos previamente estudados, em nosso laboratório, para a
obtenção da melhor resolução bidimensional de proteínas do caule de cajueiro, o método
baseado no uso da solução fenol equilibrada com Tris-HCl, pH 8,0, foi o mais eficiente na
obtenção de conteúdo compatíveis de proteínas, assim como na melhor resolução dos perfis
bidimensionais (GONDIM, 2010). De fato, usando essa metodologia de extração, a
visualização direta das proteínas no gel permitiu a constatação de um padrão de mapa
bidimensional, característico de plantas de cajueiro e, portanto, bastante semelhante entre as
condições de campo e do clone BRS 226. A resolução desses géis evidenciou, também, o alto
controle de qualidade obtido na preparação das amostras. A degradação não controlada de
proteínas, modificação química, ou problemas de solubilização podem ser facilmente
detectados por mudanças na distribuição dos spots no perfil bidimensional (ROGOWSKA-
WRZESINSKA et al., 2013), o que não foi observado ao longo dos mapas de proteínas
obtidos nos experimentos (Figuras 9 e 14). Assim, a qualidade dos resultados obtidos permitiu
avaliar alterações quantitativas da expressão relativa das proteínas.
5.2.1 Mudanças no proteoma caulinar
Plantas de cajueiro infectadas com L. theobromae acionam respostas moleculares
associadas à reprogramação da expressão de proteínas no tecido caulinar, sítio de infecção do
patógeno. Isso sugere que a percepção do patógeno pela planta elicita alterações fisiológicas
que induzem a expressão diferencial de proteínas do caule com funções moleculares
relacionadas a variados processos biológicos no organismo vegetal.
Um número praticamente equivalente de spots proteicos se apresentou responsivo para
os dois cenários analisados. Na condição de incompatibilidade com o clone BRS 226, 40
proteínas foram diferencialmente expressas no grupo artificialmente infectado com L.

84
theobromae comparado com plantas controles não inoculadas. Em campo, a condição de
resistência/suscetibilidade induziu alteração no padrão de expressão de 54 proteínas presentes
no proteoma caulinar. Contudo, foram identificadas por MS/MS, 37 proteínas nas plantas com
alta pressão do patógeno, em condições de campo, e 36 proteínas associada à infecção no
clone de cajueiro anão, BRS 226, infectado artificialmente.
As proteínas sub/superexpressas estão indicadas nos mapas bidimensionais
representativos da condição em que houve reprogramação da expressão (Figuras 9 e 14).
Interessantemente, em plantas do clone BRS 226 inoculado artificialmente, o maior número
de proteínas com expressão reprogramada foi verificado nos intervalos de tempo nos quais
sintomas de exsudação de resina e necrose interna do caule foram observados (72 e 96 HAI).
Essa tendência à alteração da expressão foi evidenciada para proteínas que participam em
todos os processos biológicos identificados, com exceção de proteínas relacionadas à
categoria de transporte (Figura 10A). As respostas associadas à alteração da expressão de
proteínas, nesses intervalos de tempo estudados, podem estar relacionadas à tentativa da
planta em restringir o patógeno ao local de inoculação, evitando a progressão dos sintomas da
doença no clone BRS 226 (Figura 7). Assim, a rápida restrição do fungo em torno dos locais
iniciais de infecção pode ser extremamente importante para respostas de resistência
(HASEGAWA et al., 2010).
Para melhor compreensão dos papéis das proteínas com expressão reprogramada nas
condições estudadas, a discussão de suas funções nos processos biológicos do vegetal são, em
seguida, apresentados de acordo com as categorias identificadas neste estudo e indicadas nas
tabelas 3 e 4. Conforme observado, a alteração na expressão de proteínas relacionadas a
diversos processos fisiológicos e bioquímicos é coerente com a complexidade do proteoma
caulinar e revelam possível ativação/supressão das vias biológicos em plantas de cajueiro
infectadas com L. theobromae.
5.2.1.1 Reprogramação da expressão de enzimas envolvidas com o metabolismo e produção
de energia
Neste trabalho, um grande número de proteínas metabólicas e relacionadas à geração
de energia foi reprogramado em plantas de cajueiro nos dois cenários de investigação, o que
sugere a importância desses processos na reposta à infecção. Em geral, a indução das

85
respostas de defesa pela planta tem mostrado custo elevado na mobilização de alterações
metabólicas para superar ou suprimir o ataque de patógenos (SWARBRICK et al., 2006).
A identificação das proteínas que tiveram expressão alterada, por MS/MS, revelou
mudanças na abundância relativa de várias delas envolvidas com o metabolismo de
aminoácidos (spots 15, 16, 23, 38, S11 e R7). Interessantemente, em plantas do clone BRS
226 inoculadas artificialmente, dois desses spots (15 e 16) correspondem a proteínas
envolvidas com processos anabólicos e que são superexpressas em 72 e 96 HAI, período
coincidente com a superexpressão do fator de elongação da tradução (spot 40). Por outro lado,
nos tempos iniciais após a infecção, as enzimas s-adenosilmetionina sintetase (spot 23) e
serina hidrometiltransferase (spot 38), que participam das vias do metabolismo de
aminoácidos foram reprimidas em plantas do clone BRS 226 inoculadas com L. theobromae.
Essas enzimas se mostraram, também, com expressão reprogramada em plantas de grão de
bico após infecção com o nematoíde Meloidogyne artiellia (PALOMARES-RIUS, 2011).
As observações citadas sugerem que plantas do clone BRS 226 infectadas com L.
theobromae, regulam o metabolismo de aminoácidos de forma diferenciada conforme o
intervalo de tempo pós-infecção. Essa dinâmica observada para proteínas envolvidas com o
metabolismo de aminoácidos sugere haver regulação sútil da disponibilidade dos monômeros
de aminoácidos em períodos necessários para reprogramação da expressão de proteínas
associadas à resistência. Esse controle espacial-temporal também foi observado na biossíntese
do aminoácido glutamina em plantas desafiadas com patógenos (Pageau et al., 2006; PEREZ-
GARCIA et al., 1995, 1998). Evidências de que a homeostase nos níveis de aminoácidos
desempenham papel relevante na defesa de plantas foram reveladas em estudos com
aminotransferases e homosserina quinases (SONG et al., 2004; TALER et al., 2004; VAN
DAMME et al., 2009) ou na investigação do metabolismo primário de aminoácidos
específicos (LIU et al., 2010).
Enzimas associadas ao metabolismo de carboidratos foram diferencialmente reguladas
na interação cajueiro-L. theobromae. No clone BRS 226, essa reprogramação foi
significativamente relevante em 72 HAI (spots 18, 32 e 35) e entre estas proteínas somente
uma alfa-glucano fosforilase foi subexpressa (spot 32). Em contrapartida, nas condições de
campo, o estímulo da via glicolítica foi evidenciado pela superexpressão, em plantas
resistentes, de enzimas responsáveis pela catálise de passos sequenciais na oxidação de
carboidratos, como a aldolase (spot R12) e a triose fostato isomerase (spots R11). Das

86
proteínas diferencialmente analisadas por MS/MS e categorizadas como enzimas do
metabolismo de carboidratos, a frutose-bifosfato aldolase foi igualmente superexpressa no
clone BRS 226 (spot 35) e em plantas de cajueiro resistentes a resinose (spot R12), em
condições de campo. A indução do estado de “priming”, em plantas de tomate tratadas com
vários elicitores de defesa, mostrou a superexpressão de várias proteínas do metabolismo
primário, incluindo a frutose-bifosfato aldolase (ARASIMOWICZ-JELONEK et al., 2013).
Curiosamente, a subexpressão desta enzima foi relatada no estudo proteômico de plantas de
ervilha suscetíveis desafiadas com o parasita Orobanche crenata (CASTILLEJO et al., 2004).
As alterações no metabolismo de carboidratos, decorrentes da interação planta-
patógeno, são entendidas como parte do recurso usado na geração de energia pela planta
(LÓPEZ-GRESA et al., 2009). Além disso, o envolvimento de açúcares que são precursores
para a biossíntese de metabólitos secundários tem sido proposta com base, por exemplo, no
acúmulo dos ácidos p- cumárico, cafeico e ferúlico formando ésteres com a glicose nas
plantas de Cucumis sativus e Cucumis melo infectadas com os vírus PNRSV (Prunus necrotic
ringspot vírus) e MNSV (Melon necrotic spot vírus), respectivamente (BELLÉS et al., 2008).
Somado a isso, mudanças nos níveis de açúcares são uma típica indicação de alteração no
metabolismo de carboidratos após a infecção (ABDEL-FARID et al., 2009). Dessa forma, o
conteúdo de carboidratos pode afetar os mecanismos de resistência à doença por meio de
mudanças no metabolismo geral da planta (BERGER et al., 2007).
A atuação funcional de enzimas envolvidas com via glicolítica, mencionadas
anteriormente, representam o primeiro estágio da respiração celular, seguido do ciclo do ácido
tricarboxílico (TCA) e cadeia transportadora de elétrons. Estas vias interligadas funcionam
para gerar equivalentes de energia e esqueletos de carbono que podem ser utilizados na
biossíntese de vários metabólitos (BOLTON, 2009).
A oxidação de piruvato proveniente da glicólise é realizada pelo complexo da piruvato
desidrogenase (PDH), formando o acetil-CoA que entra no ciclo de Krebs. A PDH é o ponto
regulatório chave no fluxo de entrada do piruvato para a completa oxidação (FERNIE et al.,
2004). NADH e FADH2, gerados nesta etapa são oxidados na cadeia transportadora de
elétrons, formada por quatro complexos, entre os quais, o complexo da NADH desidrogenase
(spot 11), referido como complexo I, sendo responsável pela transferência de elétrons do
NADH para coenzima Q. O fluxo concomitante de elétrons pelos demais complexos fornece
um gradiente de pH necessário para a síntese ATP, pela ATPsintase. A mudança na

87
abundância relativa de uma putativa NADH desidrogenase, com 12 HAI, revela uma demanda
energética no metabolismo das plantas de cajueiro infectadas.
O requerimento de poder redutor para vias anabólicas parece ser estimulado pela
superexpressão em plantas de cajueiro suscetíveis, cultivadas em campo, de duas
transaldolases (spots S6 e S10), envolvidas na via das pentoses fosfato, que gera NADPH por
oxidação da glicose-6-fosfato. O estímulo à oxidação de hexoses fosfato, por esta via, é
tipicamente uma fonte de NADPH, que pode ser utilizado pela NADPH oxidase (PUGIN et
al., 1997). Sabendo do papel das NADPH oxidases na produção de espécies reativas de
oxigênio (ROS) (TORRES e DANGL, 2005; SUZUKI et al., 2012), o aumento dos níveis de
ROS pode, potencialmente, favorecer a infecção por L.theobromae, dado o caráter
necrotrófico deste patógeno (THAKKAR et al., 2003). Além da geração de poder redutor na
forma de NADPH, a via das pentoses fosfato pode contribuir com a disponibilidade de ribose
para componentes como ATP, DNA e RNA. Coerente com este papel no metabolismo de
nucleotídeos, em plantas suscetíveis houve superexpressão de uma adenine fosforil
transferase (spot S7), que catálise a incorporação de purinas a nucleotídeos.
Como mencionado acima, o metabolismo oxidativo do piruvato pela PDH forma o
acetil-CoA que ingressa no ciclo de Krebs durante a respiração aeróbica. Em condições de
hipóxia, o piruvato pode ser oxidado pela lactato desidrogenase (LDH) para formar lactato e
NAD+. Contudo, o aumento na concentração de lactato provoca diminuição no pH, levando à
inibição da LDH e ativação da piruvato descarboxilase (PDC) (DAVIES et al., 1974).
A PDC catalisa a conversão irreversível do piruvato a acetaldeído e CO2, enquanto
uma subsequente álcool desidrogenase converte o acetaldeído em etanol e NAD+
durante a
fermentação etanólica, que é essencial na manutenção da glicólise e produção de ATP
(BOLTON, 2009). Na verdade, a fermentação etanólica foi sugerida como uma via de
produção de energia, mesmo na presença de oxigênio sob condições de estresse abiótico ou
no desenvolvimento do pólen (KURSTEINER et al., 2003).
Spots 20 e 25, referentes à PDC, foram diferencialmente expressos em plantas
inoculadas do clone BRS 226, ao mesmo tempo em que uma álcool desidrogenase classe III
foi subexpressa com 96 HAI. Em Arabidopsis thaliana, a glicólise, respiração e fermentação
foram alteradas no sítio de infecção do oídio, sendo especulado que a fermentação é
favorecida sob condições celulares associadas com a aquisição de nutrientes pelo parasita
(CHANDRAN et al., 2010; WILDERMUTH, 2010). Nas plantas de cevada, a superexpressão

88
do gene que codifica uma ADH (HvADH1) modula a suscetibilidade ao fungo Blumeria
graminis f.sp. hordei, enquanto a diminuição da sua expressão mediada por RNAi resulta na
redução do sucesso de penetração pelo patógeno. Baseado nessas observações, a ADH pode
cumprir função na manutenção da via glicolítica/fermentativa em plantas estressadas por
patógenos, o que pode ser crucial para o abastecimento de hexoses necessário para
desenvolvimento do fungo (PATHURI et al., 2011). Neste sentido, a redução da expressão
desta proteína pode ser favorável ao estabelecimento da relação incompatível nas plantas de
cajueiro anão, clone BRS 226, inoculadas com L. theobromae.
Além do metabolismo primário, plantas têm uma grande variedade de metabólitos
secundários, também conhecidos como metabólitos especializados, que facilitam as interações
com o meio biótico e abiótico, incluindo papel essencial na defesa química contra herbívoros
e patógenos (NEILSON et al., 2013). No clone BRS 226, houve superexpressão da enzima
chalcona isomerase (spot 17) envolvida na biossíntese de flavonóides.
Os flavonóides são sintetizados a partir de derivados da fenilalanina gerados,
inicialmente, por meio da rota do ácido chiquímico, com subsequente entrada na via dos
fenilpropanóides (TOHGE et al., 2013). Nas reações iniciais desta via, a chalcona sintase
(CHS) condensa 4-cumaroil-CoA e malonil-CoA formando o flavonóide naringenina
chalcona de cadeia aberta, que é convertido para a naringenina pela chalcone isomerase
(KANG et al., 2014). A identificação de proteínas envolvidas com a biossíntese dos
flavonóides tem sido relatada em análises proteômicas de plantas sob estresse biótico
(PALOMARES-RIUS et al., 2011; MANDELC et al., 2013). Nas plantas, os flavonóides
podem atuar como fitoalexinas, compostos de baixa massa molecular com atividade
antimicrobiana e induzidos em reposta à presença de patógenos (IBRAHEEM et al., 2010;
GROßKINSKY et al., 2012). Além disso, a rota dos fenilpropanóides pode gerar metabólitos
que atuam como componentes estruturais da parede celular, incluindo lignina (LI et al.,
2010). A lignificação torna a parede celular mais resistente à pressão mecânica aplicada por
apressórios fúngicos durante a tentativa de penetração, bem como, mais resistente à água e,
portanto, menos acessível à ação de enzimas degradantes da parede celular (BHUIYAN et al.,
2009).

89
5.2.1.2 Proteínas induzidas por estresse e relacionadas à defesa
Em plantas de cajueiro, proteínas categorizadas como relacionadas ao estresse
correspondem ao segundo maior grupo com expressão alterada para ambos os cenários
investigados. Quando se trata de resinose, a manutenção da homeostase celular é uma
característica potencialmente importante, uma vez que o processo infeccioso iniciado por L.
theobromae é induzido por estresses, principalmente de ordem fisiológica (CARDOSO et al.,
2009a). As pressões ambientais são fatores predisponentes ao desenvolvimento de resinose,
aliado ao caráter oportunista do patógeno (ALVES et al., 2013). Na região semiárida, o
estresse hídrico é um fenômeno recorrente, afetando os processos bioquímicos e fisiológicos
do cajueiro (BEZERRA et al., 2007) e tornando estas plantas mais propensas a infecção.
O estresse térmico, também prevalente em condições semiáridas, tem sido associado a
mudanças no padrão de transcritos que resultam no aumento da síntese de um grupo de
proteínas conhecidas como heat schock (Hsp) (AL-WHAIBI, 2011). O acúmulo destas
proteínas, no ambiente celular, ocorre quando os mecanismos moleculares de termolerância
são acionados para prevenir ou reparar os danos ocasionados em proteínas e membranas,
lábeis ao calor (SCHOFFL et al., 1998; LARKINDALE e VIERLING, 2008). As Hsps, mais
abundantemente expressas, correspondem às famílias altamente conservadas de chaperones
moleculares Hsp100, Hsp90, Hsp70, Hsp60 e as pequenas Hsps, com massa molecular entre
17 e 30 kDa (EFEOGLU, 2009). No presente estudo, 5 Hsps tiveram sua expressão alterada
em cajueiros cultivados em condições de campo (spots R15, R19 e S13) e no clone BRS 226
(spots 9 e 21). Entre elas, uma chaperona Hsp20 foi igualmente subexpressa em plantas
inoculados do clone BRS 226 (spot 21), assim como, no grupo de plantas resistentes (spot
S13), em condições de campo, indicando alteração da expressão associada à infecção com L.
theobromae, uma vez que estas plantas compartilham este estresse biótico comum. Algumas
dessas chaperonas, por sua vez, reconhecem proteínas deformadas atuando no controle da
qualidade e evitando a formação de agregados potencialmente tóxicos durante o estresse
(TIMPERIO et al., 2008; SAIDI et al., 2009). A subexpressão da Hsp20 pode estar
relacionada à tentativa do patógeno em interferir nos mecanismos moleculares associados à
prevenção de danos celulares decorrentes da condição de estresse, tornando as plantas de
cajueiro mais suscetíveis ao desenvolvimento da infecção.
Poucos relatos tem associado o papel das sHsps (do inglês, small Heat Schock
Proteins) na interação planta-patógeno. Maimbo et al. (2007) demonstraram que a expressão

90
de uma sHsp de 17 kDa foi aumentada em folhas de tabaco na interação compatível e
incompatível com Ralstonia solanacearum e Pseudomonas cichorii, respectivamente. O
crescimento da bactéria R. solanacearum foi acelerado nas plantas de Nicotiana benthamiana
com expressão silenciada da sHsp (Nbshsp17), sugerindo importante papel na resistência à
doença. Essa observação foi consistente com a redução na expressão de genes relacionados à
defesa, incluindo PR1 (do Inglês, Pathogenesis-related 1), PR4 (do Inglês, Pathogenesis-
related 4), e EREBP (do Inglês, Et-Responsive element-binding protein) nas plantas
Nbshsp17-silenciadas (MAIMBO et al., 2007).
Garofalo et al. (2009) especularam que sHsps induzidas em plantas de laranja e
pimenta desafiadas, respectivamente, com Xanthomonas axonopodis e Xanthomonas
campestris, podem participar na resposta imune basal, uma vez que são expressas em fases
iniciais da infecção. Plantas expostas a um agente patogênico mostram rápida expressão de
proteínas envolvidas na resposta imune, o que pode sobrecarregar, no ambiente celular, as
chaperonas Hsp70 e Hsp60, também identificadas previamente nos patossistemas
mencionados (GARAVAGLIA et al., 2009). Dessa forma, foi sugerido que sHsps podem
ajudar a planta a se ajustar aos organismos invasores, contribuindo com a manutenção das
proteínas sintetizadas numa conformação aberta, até que elas possam ser corretamente
dobradas pelas chaperonas Hsp70 e Hsp60 (GAROFALO et al., 2009).
Uma Hsp70 foi superexpressa em plantas de cajueiro do clone BRS 226, 48 e 72 HAI.
Moléculas efetoras HopI1 de Pseudomonas syringae apresentam um domínio J que interage
com Hsps 70. Uma possível hipótese para interação específica deste efetor com Hsp70 foi
levantada, segundo a qual, proteínas Hsp70 afetam o enovelamento/montagem de um
complexo de defesa localizado no cloroplasto, possivelmente representado por componentes
da biossíntese do ácido salicílico ou relacionados ao seu transporte. Dessa forma, a estratégia
de P. syringae estaria centrada no fato de que a presença de HopI1 pode interferir com a
defesa porque pode sobrecarregar Hsp70, facilitando a degradação ou desmontagem do
complexo de promoção da defesa (JELENSKA et al., 2007). Contudo, o papel das Hsps70 na
resistência basal foi evidenciado pelo aumento da suscetibilidade a bactéria não patogênica P.
syringae pv. maculicola em plantas mutantes ou down-reguladas para expressão de Hsp70
(JELENSKA et al., 2010).
Em outros modelos, proteínas Hsp70, também, identificadas na interação do clone
BRS 226 com L. theobromae, são, na verdade, alvos comuns visados pelos patógenos de

91
plantas e animais, que suprimem ou exploram sua atividade como estratégias de infecção
(MULTHOFF, 2006; NOËL et al., 2007; CHEN et al., 2008; AXSEN et al., 2009). Os
mecanismos empregados por Hsps70 em resposta à interação de plantas com diferentes
patógenos, incluindo proteínas alvos e os processos celulares envolvidos, ainda não são
compreendidos, porém seus resultados influenciam, fortemente, no sucesso ou não da
infecção (JELENSKA et al., 2010).
Em plantas de tomate, óxido nítrico (NO) e peróxido de hidrogênio (H2O2) participam
na regulação da produção e acúmulo de Hsps70 sob condições de estresses abióticos e
bióticos, mas, o tipo de resposta depende, fortemente, da capacidade da planta em acionar
diferentes mecanismos de defesa (PITERKOVÁ et al., 2013).
Coerente com a contribuição das espécies reativas de oxigênio (ROS) na transdução de
sinal para expressão de Hsps em plantas (VOLKOV et al., 2006; KÖNIGSHOFER et al.,
2008; BANTI et al., 2010), no presente estudo, proteínas diretamente relacionadas ao
estresse oxidativo foram reprogramadas nas condições de campo (spots S2, S4, S16, R9) e no
clone BRS 226 (spots 07, 10, 27 e 28), contudo estas respostas parecem ser diferencialmente
reguladas.
Sabe-se que após o desafio da planta por patógenos, várias enzimas são acionadas
como NADPH oxidase, peroxidases, superóxido dismutase (SOD), oxalato oxidases,
lipoxigenases e amino oxidases, todas envolvidas na geração de ROS (SHETTY et al., 2008).
Eventos precoces de sinalização, incluindo o fluxo de íons através da membrana plasmática,
aumento dos níveis de Ca2 +
no citoplasma, ativação de MAPKs (do Inglês, mitogen-activated
protein kinase) e consequente produção de ROS podem ser ativados dentro de poucos
minutos após a exposição ao estímulo (BENSCHOP et al., 2007; MILLER et al., 2009;
FINKA et al., 2012; BAXTER et al., 2013). Essa sobrecarga de ROS na planta proporciona
uma condição de estresse oxidativo, conhecido como explosão oxidativa, podendo levar a
danos celulares e em componentes como ácidos nucléicos, proteínas e lipídeos (GILL e
TUTEJA, 2010; KOVALCHUK, 2010). Portanto, o acúmulo de ROS deve ser
contrabalanceado por sistemas antioxidantes, dentre os quais, a ação de enzimas como
catalase, ascorbate peroxidase e peroxiredoxinas (NAVROT et al., 2011).
Algumas moléculas geradas durante a explosão oxidativa são consideradas moléculas
de sinalização, sendo percebidas por diferentes receptores, proteínas ou enzimas da planta
(MITTLER et al., 2011). A geração destas espécies tem papel significativo para a resistência

92
a patógenos biotróficos e hemibiotróficos, no entanto, tem sido sugerido que as ROS
funcionam como fator de virulência para fungos necrotróficos (WEN, 2013), uma vez que seu
acúmulo induz à morte celular, favorecendo o crescimento do patógeno e desenvolvimento da
doença (GOVRIN e LEVINE, 2000).
A expressão alterada de enzimas envolvidas no processo de remoção dessas espécies,
observada neste estudo, indica que houve estresse oxidativo em plantas de cajueiro quando
desafiadas com L. theobromae. De fato, no clone BRS 226, uma proteína tipo 2-cys
peroxirredoxina (spot 10) foi superexpressa em 12 e 24 HAI, evidenciando possível explosão
oxidativa em tempos iniciais após infecção com L. theobromae. Proteínas deste tipo
pertencem à família das peroxirredoxinas, que usam o conservado resíduo de cisteína para
reduzir o peróxido de hidrogênio, com consequente formação de pontes de dissulfeto (KIM et
al., 2009). Um novo estado reduzido, destas pontes, garante o mecanismo catalítico das 2-cis
peroxirredoxinas e é alcançado com o auxílio de sistemas de redução baseados na atuação de
tiorredoxinas e glutarredoxinas (ROUHIER et al., 2008; MEYER et al., 2009). Em condições
de campo, proteínas envolvidas com estes sistemas foram mais expressas em plantas
suscetíveis e correspondem aos spots S4 e S16.
Coerente com a expressão da 2-cys peroxirredoxina, outra proteína envolvida com a
homeostase redox foi superexpressa em plantas inoculadas do clone BRS 266,
correspondendo a uma proteína dissulfeto isomerase (spot 07). Esta enzima teve sua
expressão reprogramada no período de 24 HAI, em paralelo com a superexpressão da 2-cys
peroxirredoxina. Contudo, esta alteração na expressão foi também evidenciada no intervalo de
tempo em que sintomas característicos de infecção com L. theobromae foram observados, isto
é, com 72 e 96 HAI. A proteína dissulfeto isomerase (PDI) catalisa a formação, redução e
isomerização de pontes dissulfeto em proteínas secretórias nascentes no lúmem do retículo
endoplasmático, exibindo atividade de chaperona (CHRISTIS et al., 2008; HATAHET e
RUDDOCK, 2009).
A superexpressão da PDI em plantas desafiadas com fitopatógenos tem sido observada
em estudos proteômicos diferenciais (AFROZ et al., 2009; FANG et al., 2012;
PALOMARES-RIUS et al., 2011) e sua participação na interação-planta patógeno pode estar
relacionado ao correto enovelamento de proteínas relacionadas à defesa (GRUBER et al.,
2007). De acordo com esse papel, o significado biológico da expressão das dissulfeto
isomerases (NbERp57 e NbP5) na resitência do tabaco ao TMV (do Inglês, Tobacco mosaic

93
vírus) foi estudado por meio do silenciamento destas proteínas, seguido de infecção, o que
resultou em baixo fenótipo de resistência, indicando a necessidade destas isomerases na
completa ativação da resposta imune (CAPLAN et al., 2009).
Um especial interesse tem sido dado à expressão de tiorredoxinas na resposta de
plantas a fungos necrotróficos. No nosso estudo, em condições de campo, uma proteína tipo
tiorredoxina (spot S4) foi superexpressa no grupo suscetível de cajueiros, em relação ao grupo
resistente. Estas proteínas, juntamente com glutarredoxinas, têm sido sugeridas como alvos
diretos ou indiretos de virulência para patógenos necrotróficos em Arabidopsis thaliana (LAI
e MENGISTE, 2013). De acordo com esta premissa, TRX-h5 e TRX-h3 são requeridas para a
transição do estado oligomérico de NPR1 (do Inglês, Non expressor of pathogenesis-related
genes 1) para o monomérico (TADA et al., 2008), necessário na interação física com os
fatores transcricionais TGA, que induzem a expressão dos genes de defesa (LINDERMAYR
et al., 2010). A ativação de NPR1, por sua vez, aumentou a suscetibilidade de A. thaliana a B.
cinerea (EL OIRDI et al., 2011).
A aparente contradição desse cenário é explicada pela suscetibilidade acionada por
toxina, segunda a qual, muitos patógenos necrotróficos secretam toxinas que servem como
fatores de virulência sensíveis à percepção do hospedeiro e que por sua vez, aumentam a
suscetibilidade (MENGISTE, 2012). De fato, as respostas observadas nesta interação
apresentam um significado inversamente correlacionado a ETI (OLIVER e SOLOMON,
2010). Nesse contexto, a resistência contra patógenos necrotróficos é sugerida pela existência
de genes dominantes que codificam proteínas que desintoxicam toxinas secretadas pelo
invasor, que garantem à insensibilidade a presença delas ou que aumentam a tolerância à
morte celular induzida (LAI e MENGISTE, 2013).
Baseados nesse modelo, a superexpressão de uma proteína tipo tiorredoxina em
plantas de cajueiro suscetíveis à resinose, em condições de campo, pode atuar regulando o
estado redox de proteínas alvos em decorrência da perturbação por fatores de virulência de
L.theobromae, influenciando na suscetibilidade. Uma evidência direta dessa condição de
suscetibilidade para patógenos necrotróficos é exemplificada pela toxina vitorina de
Cochliobolus victoriae que interage com a tiorredoxina TRX-h5 da planta, permitindo o
reconhecimento desta toxina pela proteína R LOV1, o que condiciona à sensibilidade a essa
toxina, com consequente suscetibilidade ao agente patogênico (LORANG et al., 2012).

94
Alteração no estado redox em plantas de cajueiro suscetíveis foi evidenciada pela
observação de que as enzimas peroxidase (spot S2) e glutationa redutase (spot S16) foram
superexpressas. Em contrapartida, nas plantas de cajueiro resistentes somente uma enzima
removedora de peróxido, identificada como uma ascobarto peroxidase citosólica (spot R9),
foi superexpressa em relação ao grupo suscetível. A tendência na superexpressão desta
enzima no grupo de plantas resistentes foi confirmada por análise de western blot (Figura 19).
A ascobarto peroxidase atua com alta afinidade na eliminação de H2O2, usando como
doador de elétrons o ascobarto, que é restaurado ao seu estado reduzido pela MDHAR
(monodeidroascorbato redutase) de forma dependente de NADPH (NAVROT et al., 2011). O
genótipo resistente de Theobroma cacao (TSH1188) mostrou aumento na expressão de genes
que codificam ascobarto peroxidases citosólicas, em comparação com o genótipo suscetível
(Catongo), sugerindo que estas enzimas atuam na defesa dessas plantas quando são infetadas
pelo fungo Moniliophthora perniciosa (CEITA et al., 2007; DIAS et al., 2011).
Curiosamente, somente duas proteínas diretamente relacionadas à defesa vegetal
foram identificadas nas plantas de cajueiro do clone BRS 226 e em plantas de cajueiro
cultivadas em condição de campo. No clone BRS 226, houve subexpressão de uma proteína
tipo germina (spot 33) com 96 HAI. Proteínas desse tipo foram originalmente identificadas
em trigo, como marcadores específicos do estado de germinação (LANE et al., 1993).
Registros do aumento da expressão de genes relacionados a essas proteínas foram mostrados
no ataque de plantas a fungos (MANOSALVA et al., 2008), nematóides (KNECHT et al.,
2010), insetos (RAMPUTH et al., 2002; Lou e Baldwin, 2006), bactérias e vírus (Park et al.,
2003; BARRERA-PACHECO et al., 2008).
O estudo do papel fisiológico das GLPs (do Inglês, Germin-like proteins) na defesa
celular tem focado na expressão e mecanismo de defesa dessas proteínas no apoplasto
(FELLE et al., 2005), onde sua interação com a parede celular é capaz de responder, em
estágios iniciais, ao desafio de patógenos e em regiões próximas ao sítio de infecção
(DAVIDSON et al., 2010). GLPs podem atuar no reforço da parede celular e exibir atividade
única ou combinada das enzimas oxalato oxidase e superóxido dismutase, que resulta na
produção de H2O2 (DUNWELL et al., 2008; BANERJEE e MAITI, 2010). O papel
pressuposto dessas atividades pode explicar a possível subexpressão dessa proteína em
plantas inoculadas do clone BRS 226, uma vez que o H2O2, juntamente com outras ROS, atua
em vias de sinalização para percepção do patógeno acionando, por exemplo, a morte celular

95
favorável para o desenvolvimento de invasores necrotróficos. A manutenção de baixos níveis
de ROS pode ser pensada como um mecanismo importante nas plantas de cajueiro, em
períodos de tempo onde o patógeno parece desenvolver sintomas severos de infecção, como
observado com 96 HAI.
Outra proteína relacionada à resposta basal foi superexpressa em plantas resistentes de
cajueiro, infectadas em condições de campo. As serpinas (spot R14) têm um papel no
controle da proteólise por inibição irreversível de proteínas alvos. Dessa forma, elas são
pensadas como inibidores, elicitados na defesa, contra proteases presentes no intestino de
insetos ou secretadas por microrganismos, o que determina a redução na disponibilidade de
aminoácidos necessários para o crescimento desses organismos (ROBERTS e HEJGAARD,
2008). Na interação planta-patógeno proteínas dessa classe foram identificadas sendo
superexpressas em raízes de Actinidia chinensis durante a infecção sistêmica com
Pseudomonas syringae pv. actinidiae (PETRICCIONE et al., 2013). Outros inibidores de
protease responderam com superexpressão em vários patossistemas (CANTÚ et al., 2008b;
FAN et al., 2011), sugerindo que a intervenção da proteólise faz parte dos mecanismos de
resistência à doença. Por exemplo, nosso grupo de pesquisa observou, recentemente, haver
aumento da expressão de um inibidor de proteinase cisteínica em plantas de feijão-de-corda
resistente, quando desfiado pelo nematóide das galhas, Meloidogyne incognita (OLIVEIRA et
al., 2012).
5.2.1.3 Proteínas relacionadas à sinalização celular
Todos os processos biológicos das plantas são controlados por vias de transdução de
sinal e, em geral, a regulação das rotas metabólicas parece ocorrer através da transição do
estado de proteínas alvos, mediado por fosforilação (CHEN et al., 2006). Em muitos casos,
para completar o mecanismo regulatório, proteínas fosforiladas devem se associar fisicamente
com outras proteínas, conhecidas como 14-3-3 (FULGOSI et al., 2002; SEHNKE et al.,
2002). Nas condições avaliadas neste trabalho, proteínas 14-3-3 foram reprogramadas nas
plantas de cajueiro infectadas, em ambas as condições de estudo, campo e clone BRS 226
inoculado artificialmente.
O mecanismo de ação destas proteínas envolve, inicialmente, um estímulo que aciona
rotas de sinalização celular, com consequente fosforilação de proteínas que são, por sua vez,

96
reconhecidas pelas 14-3-3 (DENISON et al., 2011). O número de proteínas identificadas em
plantas e que são candidatas à regulação mediada por membros desta família é superior a 300
(CHANG et al., 2009; OECKING e JASPERT, 2009; PAUL et al., 2009; SCHOONHEIM et
al., 2009). A crescente expansão de dados relacionados a estas proteínas sugerem seu
envolvimento em vários processos fisiológicos e vias de sinalização nas plantas.
No clone BRS 226, a expressão de uma proteína 14-3-3 foi aumentada em 48 e 72
HAI. A ligação de proteínas 14-3-3 a alvos, de maneira dependente de fosforilação, sugere o
envolvimento nas respostas de defesa do organismo vegetal. Esta hipótese é reforçada, por
exemplo, na importância das proteínas quinases e fosfatases na transdução de sinas para as
respostas de defesa da PTI e ETI em plantas, cujas atividades dessas enzimas fosforiladas
podem ser reguladas por meio da ligação com as 14-3-3 (OH, 2010). Recentemente foram
relatados, em plantas, dois casos em que proteínas 14-3-3 desempenham um papel crucial na
resistência à doença ou imunidade associada à morte celular programada (YANG et al, 2009;
OH et al., 2010).
Nas condições de campo, plantas resistentes de cajueiro apresentaram menor
abundância relativa dessa proteína em relação às plantas suscetíveis. Esta repressão de 14-3-3
foi observada em A. thaliana de maneira dependente da presença de etileno e ácido abscísico
(LANCIEN e ROBERTS, 2006). Coerente com essa observação, uma proteína responsiva ao
ABA também foi superexpressa no grupo resistente de plantas de cajueiro, demonstrando
possível contribuição da sinalização com este fitohormônio à regulação da expressão protéica.
Respostas ao estresse abiótico são em grande parte controlados pelo hormônio ABA,
enquanto que a defesa contra estresse biótico é especificada pelo antagonismo entre as vias do
ácido salicílico (SA) e ácido jasmónico (JA)/etileno. No entanto, descobertas recentes
sugerem que o ABA atua tanto de forma sinérgica como antagônica na sinalização induzida
por estresse biótico, criando uma complexa rede de respostas em diferentes níveis (FUJITA et
al., 2006; ASSELBERGH et al., 2008; YASUDA et al., 2008) o que explica, portanto,
repostas divergentes nas plantas de cajueiro mantidas em condições controladas (clone BRS
226) ou sob campo (plantas resistentes).
Concernente com esta linha de observação, proteínas chamadas de anexinas foram
igualmente responsivas nos cenários analisados, porém, com expressão divergente nos grupos
experimentais, sendo subexpressas em plantas inoculadas do BRS 226 (spots 31 e 32),
enquanto superexpressas em plantas resistentes cultivadas em campo (spots R16 e R17). A

97
expressão controversa das anexinas por estresses bióticos combinados foi registrada pela
redução da expressão dessa proteína em plantas resistentes de Cicer arietinum L. (Grão-de-
bico) infectadas com Fusarium oxysporum f. sp.ciceris. Contudo, a coinfecção dessa planta
com Meloidogyne artiellia resultou na supereexpressão dessa proteína (PALOMARES-RIUS
et al., 2011). Essa observação fornece indícios de que uma suposta combinação do ataque de
patógenos em plantas resistentes, cultivadas em campo, pode alterar o padrão de expressão
observado no clone BRS 226.
A expressão das anexinas também tem sido implicada em condições de estresse salino,
oxidativo ou modulado positivamente por ABA (WATKINSON et al., 2003; HOSHINO et
al., 2004). Uma especial característica dessas proteínas é sua potencial ligação a fosfolipídios
de maneira dependente ou não de cálcio (Ca 2+
) (MADUREIRA e WAISMAN, 2013), o que
as torna potencialmente aptas à atuação em vias de sinalização celular envolvendo cálcio
citosólico livre e espécies reativas de oxigênio (MORTIMER et al., 2008).
Diversas condições de estresse biótico e abiótico resultam em um rápido aumento de
ROS que podem atuar como mensageiros secundários juntamente com Ca 2+
citosólico
(FRAIRE-VELÁZQUEZ et al., 2011). O pressuposto papel das anexinas na resposta ao
estresse oxidativo tem sido investigado em plantas (MADUREIRA e WAISMAN, 2013;
MORTIMER et al., 2008). Em campo, o aumento da expressão desta proteína foi observado
em plantas de cajueiro resistentes, como mencionado acima. Coerente com a resistência desse
grupo, a expressão ectópica de uma anexina de Brassica juncea, em tabaco transgênico,
conferiu tolerância aos estresses abióticos e bióticos (JAMI et al., 2008). Resultados
relacionados à ocorrência de estresse abiótico na indução da translocação de anexinas do
citosol para a membrana, associado ao turnover de expressão desta proteína, foram
relacionados à sinalização por ABA dependente de cálcio (LEE et al., 2004; CLARK et al.,
2010), também pressuposta para plantas resistentes de cajueiro pela superexpressão de uma
proteína ligante a cálcio (spot R3).
Cascatas de sinalização para rotas de defesa vegetal tem envolvido a participação de
proteínas quinases, responsáveis pela percepção direta de elicitores e produtos dos genes Avr,
evento que condiciona a ativação de fatores de transcrição e respostas sistêmicas no vegetal
(ROMEIS, 2001). Participando do conjunto de proteínas quinases que são induzidas por
PAMPs ou efetores derivados de patógenos, as quinases ativadas por mitógeno (MAPKs)
estão potencialmente envolvidas na sinalização para a resistência à doença, tanto na PTI como

98
ETI (RODRIGUEZ et al., 2010; MENG e ZHANG et al., 2013). Dessa forma, a regulação
positiva ou negativa das respostas de defesa tem sido atribuída à expressão dessas proteínas
(NÜRNBERGER e SCHEEL, 2001).
Em plantas de cajueiro suscetíveis, uma proteína inibidora de quinase (spot S12) teve
sua expressão aumentada em relação ao grupo resistente. A superexpressão desta proteína
inibidora pode influenciar nas rotas de sinalização acionadas por proteínas quinases e na
consequente condição de suscetibilidade. Em plantas de tomate a redução da expressão de
uma proteína quinase TPK1b por RNA de interferência resultou no aumento da
suscetibilidade ao fungo necrotrófico B. cinerea e herbivoria por larvas de Manduca sexta
(ABUQAMAR et al., 2008). Curiosamente, em A. thaliana o comprometimento da resistência
a Peronospora parasítica e Pseudomonas syringae, evidenciado por silenciamento da
proteína quinase MPK6, demostrou que a função molecular dessa proteína não pode ser
complementada por outras MAPKs endógenas (MENKE et al., 2004), o que é indicativo de
uma sofisticada regulação da resistência vegetal a patógenos mediada por proteínas quinases
(MELVIN et al., 2014).
5.2.1.4 Metabolismo de proteínas e enovelamento
As plantas constantemente monitoram o desafio de patógenos e utilizam diversificados
mecanismos adaptativos de defesa, incluindo a regulação diferencial de proteínas (GERBER
et al., 2008). Essas alterações podem estar relacionadas ao turnover de proteínas pelos
processos de biossíntese e degradação, bem como, por mudanças pós-traducionais que
influenciam na atividade de proteínas pré-existentes, somado a alterações no enovelamento
que favorecem seus mecanismos funcionais. De acordo com esse potencial evento da
interação planta-patógeno, vários trabalhos têm discutido a regulação de proteínas
categorizadas nesses processos biológicos (VALCU et al., 2009; FANG et al., 2012; ZHAO
et al., 2012).
Proteínas que são componentes do sistema proteolítico foram expressas
diferencialmente tanto nas plantas de cajueiro resistentes e suscetíveis em campo (spots R8 e
S14), como no grupo controle e inoculado do clone BRS 226 (spots 22 e 26). A sobreposição
de respostas nesses grupos, com relação à expressão das subunidades alfa e beta do

99
proteassoma 26S parece ser responsiva à infecção pelo L. theobromae, muito embora não
exista consenso na indução ou repressão da expressão dessas proteínas.
Crescentes evidências indicam que as plantas utilizam a via do sistema de degradação
proteassoma 26S/ubiquitina (USP) na resposta imune à invasão de patógenos, enfatizando a
importância dessa via durante as interações planta-patógeno (DEVOTO et al., 2003;
DELAURÉ et al., 2008; CITOVSKY et al. , 2009; TRUJILLO E SHIRASU, 2010). Contudo,
respostas de degradação proteolítica têm mostrado diferenciadas contribuições conforme o
patossistema estudado. Por exemplo, o efetor AvrPto induz a atividade proteolítica endógena
responsável pela degradação de RIN4, um regulador negativo da resposta de defesa basal, em
Solanum lycopersicum e Nicotiana benthamiana. Essa interação é, por sua vez, dependente de
genes de resistência nestas plantas, sendo induzida por múltiplos efetores bacterianos (LUO et
al., 2009). Na análise proteômica da interação entre Mentha arvensis-Alternaria alternata, a
subexpressão das subunidades do proteassoma 26S sugere possível destruição desse
complexo, associada à infecção, com o colapso das proteínas oxidadas mediado por outros
sistemas proteolíticos (SINHA e CHATTOPADHYAY, 2011).
Em outros casos o USP do hospedeiro é utilizado por patógenos para seu próprio
benefício (ANGOT et al. 2007). O mais bem estudado caso em que efetores do patógeno
usam o proteassoma da planta para degradação de proteínas alvos do hospedeiro é
representado pelo efetor AvrPtoB secretado por P. syringae (ABRAMOVITCH et al., 2006,
TORRES et al., 2006, JANJUSEVIC et al., 2006). O AvrPtoB tem a atividade ligase das
proteínas E3, uma das últimas enzimas envolvidas na cascata de ubiquitinação, fato que
potencialmente favorece supressão da PTI e ETI (DUDLER, 2013). Esse efetor AvrPtoB
ubiquitina também, FLS2 e CERK1, dois receptores tipo PRRs de A. thaliana responsáveis
pelo reconhecimento dos PAMPs flagelina e quitina, respectivamente, orientando-os para
destruição proteossomal (GOHRE et al., 2008; GIMENEZ-IBANEZ et al., 2009). Em tomate,
AvrPtoB também marca a proteína de resistência Fen para proteólise, interferindo, assim com
ETI (ROSEBROCK et al., 2007).
Outras proteínas, envolvidas com a síntese proteica foram superexpressas em plantas
de cajueiro resistentes (spot R1), em campo, assim como no clone de cajueiro anão, BRS 226,
infectado artificialmente (spot 40). No clone BRS 226, a superexpressão do fator de
elongação da tradução 1A no grupo inoculado foi observada em 72 e 96 HAI, período de

100
tempo onde houve maior concentração de proteínas identificadas com expressão
reprogramada associada à infecção.
A família de fatores de elongação dos eucariotos compreendem as proteínas eEF1A e
EF1B em plantas (RODNINA e WINTERMEYER, 2009). O fator de elongação da tradução
eEF1A é uma das mais abundantes proteínas nas células eucarióticas, e tem papel no
transporte do complexo aminoacil- tRNA no sítio ribossômico ao longo da biossíntese de
proteínas (LE SOURD et al., 2006). Além disso, outras funções adicionais têm sido atribuídas
a esse fator, incluindo o controle de qualidade das proteínas recém-produzidas, degradação de
proteínas dependente de ubiquitina, organização do citoesqueleto de actina, elicitação da
imunidade inata e resistência a bactérias patogênicas em plantas (KUNZE et al., 2004,
CHUANG et al., 2005; GROSS e KINZY, 2005; NEWTON et al., 2010; FU et al., 2012). A
subexpressão destes fatores foi observada na interação compatível entre o lúpulo (Humulus
lupulus L.) e o fungo Verticillium albo-atrum (MANDELC et al., 2013).
A proteína ribossomal 60S acídica (spot R1) foi superexpressa no grupo de cajueiro
resistente, cultivado em campo, e tem também função relacionada ao metabolismo proteico.
Essa proteína é constituinte do ribossomo e existe em múltiplas cópias, formando um
complexo hetero-oligomérico (P1/P2)2 reconhecido como uma protuberância lateral sobre a
subunidade ribossomal 60S e que está diretamente envolvida nas interações com fatores de
alongamento durante a biossíntese de proteínas (TCHÓRZEWSKI, 2002). Recentemente, a
atuação funcional como alérgeno foi atribuída a uma proteína ribossomal 60S acídica de
tomate (LÓPEZ-MATAS et al., 2011).
Análise proteômica do genótipo resistente de feijão de corda (genótipo BR-3)
infectado com o fungo hemiotrófico Colletotrichum gloeosporioides evidenciou alterações
associadas à superexpressão de proteínas envolvidas em vias anabólicas de aminoácidos e
proteínas, como observado no presente estudo. A ocorrência desses eventos, em plantas de
feijão-de-corda inoculadas, foi relacionada à disponibilização de aminoácidos para a síntese
de novo de proteínas acionadas nos eventos de resistência à doença (MOURA et al., 2014).
Além dos processos de síntese e degradação de proteínas, mencionados anteriormente,
é fundamental para a viabilidade celular o estabelecimento de um correto equilíbrio entre os
eventos de enovelamento e degradação de proteínas deformadas (MCCLELLAN et al., 2005).
De forma especial, com o desafio de patógenos a garantia da atuação funcional das proteínas

101
mobilizadas decorrentes da interação pode ser alcançada, por exemplo, pela correta
conformação associada à estrutura ativa.
Respostas associadas ao processo de enovelamento proteico incluem a atuação de uma
elaborada maquinaria enzimática baseada no mecanismo de proteínas conhecidas como
chaperonas (FRYDMAN, 2001; HARTL, 2002). As chaperonas moleculares interagem
seletivamente com certas sequências e elementos estruturais, podendo contribuir tanto com a
limitação da conformação espacial das proteínas alvos como com o favorecimento de
determinadas formas de enovelamento (GIDALEVITZ et al., 2013). Alguns fatores são
relacionados ao incorreto dobramento proteico, entre os quais, uma resposta adaptativa celular
acionada por estresse biótico, já relatada em plantas (FANATA et al., 2013). Nesse contexto,
em plantas de cajueiro do clone BRS 226 a infecção com L. theobromae alterou a expressão
de proteínas envolvidas com o enovelamento (spots 08 e 19), sugerindo uma resposta
bioquímica ao sinal da presença do patógeno.
5.2.1.5 Proteínas relacionadas ao transporte
Proteínas envolvidas com esse processo biológico foram diferencialmente responsivas
somente em plantas do clone BRS 226, sendo subexpressas no grupo inoculado com o L.
theobromae. Nesta categoria, foram incluídas uma lipocalina (spot 29) reprogramada 24 HAI
e, posteriormente 96 HAI, além de uma Ran-like small GTPase (spot 30), subexpressa 72
HAI.
Lipocalinas de plantas são classificadas em dois grupos, as lipocalinas induzidas por
temperatura (TILs), localizados na membrana plasmática (CHARRON et al., 2005; GRZYB
et al., 2006) e as cloroplásticas (CHL), encontradas no lúmen dos tilacóides (LEVESQUE-
TREMBLAY et al., 2009). Essas proteínas possuem estrutura terciária simples que lhes
confere a capacidade de se ligar a moléculas pequenas, geralmente hidrofóbicas, auxiliando
no transporte (GANFORNINA et al., 2000; CHARRON et al., 2008). Nas plantas, os alvos
moleculares de lipocalinas são desconhecidos (ABO-OGIALA, 2014). Contudo, esta família
de proteínas está emergindo como um alvo interessante para o melhoramento de plantas, uma
vez que, estão envolvidas na proteção a uma vasta gama de sinais abióticos (CHARRON et
al., 2008; CHI et al., 2009; KJELLSEN et al., 2010). Em plantas de A. thaliana tem sido
sugerido que a TLI1 é capaz de mediar a resistência a estresse, sugerindo sua participação na

102
defesa constitutiva inata ao estresse abiótico (BRINKER et al., 2010), somado a isto, o
nocaute de CHL foi associado a uma forte tendência à peroxidação de lipídeos (LEVESQUE-
TREMBLAY et al., 2009).
Recentemente, foi relatado que o silenciamento de plantas resistentes de tomate no
gene SlVRSLip, que codifica uma proteína tipo lipocalina, levou ao colapso da resistência
após inoculação com o vírus TYLCV (Tomato yellow leaf curl virus), associado à ocorrência
de necrose ao longo das hastes e pecíolos, em pararelo com a geração de ROS (SADE et al.,
2012).
Uma segunda proteína de transporte subexpressa em plantas de cajueiro inoculadas,
clone BRS 226, corresponde ao spot 30. Proteínas tipo Ran são uma das quatro subfamílias
das pequenas GTPases (small GTPases) (VERNOUD et al., 2003). Uma diversidade de
funções foram atribuídas a essas proteínas, dentre as quais, as Ran GTPases estão envolvidos
na mediação do transporte de proteínas e RNA através do envelope nuclear (DASSO, 2002).
Coerente com este papel, essas proteínas são requeridas para importação ou exportação entre
o núcleo e citoplasma de proteínas contendo o sinal de localização nuclear, além disso, a
atividade de hidrólise de GTP no citoplasma é importante para a dissociação de complexos de
exportação (GASIOROSWSKI e DEAN, 2003).
Levando em consideração, que alguns dados sugerem que a dinâmica de importação
de proteínas R para o núcleo é importante para iniciar a resposta de sinalização a doença em
plantas (BURCH-SMITH et al., 2007; SHEN et al., 2007; WIRTHMUELLER et al., 2007),
componentes envolvidos nesse tráfego, como as Ran GTPases, são sugeridos estarem
envolvidos nesse processo (LIU e COAKER, 2008). Baseado nisso, usando abordagem
proteômica, uma proteína R tipo NB-LRR, em batata, foi implicada no tráfico núcleo-
citoplasmático através de sua associação com uma Ran GTPase (SACCO et al., 2007;
TAMELING e BAULCOMBE, 2007).
5.2.1.6 Mudanças na expressão de proteínas associadas a componentes estruturais e
responsivas a hormônios
Os hormônios vegetais têm um papel essencial na integração de estímulos ambientais
e de desenvolvimento em uma rede de sinalização elaborada, que não só molda a arquitetura
da planta, mas, também a prepara para responder, apropriadamente, aos potenciais estresses

103
(BARI e JONES, 2009; KAZAN e MANNERS, 2009). Em plantas de cajueiro, cultivadas em
campo, duas proteínas responsivas a hormônios foram identificadas, demonstrando uma
possível condição de sinalização em reposta à presença de auxina (spot S15) e ácido abscísico
(spot R10).
A proteína responsiva ao ABA (spot R10) foi mais expressa em plantas de cajueiro
classificadas como resistentes. Dessa forma, respostas relacionadas a esse hormônio são
sugeridas neste grupo de plantas. Além de modular a possível divergência na expressão de
proteínas relacionadas à sinalização celular, como proposto anteriormente (spots S5, R16 e
R17), o ABA atua como um mensageiro endógeno comum em vias acionadas por estresse
biótico e abiótico (ADIE et al., 2007; HIRAYAMA e SHINOZAKI, 2007; FUJII e ZHU,
2009), ambos presentes em condição de campo, com alta pressão do patógeno.
Interessantemente, o papel conhecido do ABA no fechamento estomático durante
condições de seca (SCHROEDER et al., 2001; OSAKABE et al., 2013) parece contribuir
com uma espécie de imunidade estomática acionada conjuntamente com a via do ácido
salicílico (CAO et al., 2011), justificada em virtude do fato dos estômatos representarem uma
importante rota de entrada para fitopatógenos (ZENG et al., 2010).
Coerente com essa sobreposição de funções em condições de estresse biótico e
abiótico, os níveis de ABA aumentam rapidamente durante a infecção com B. cinerea (PAN
et al., 2008) e a colonização com P. syringae (TORRES-ZABALA et al., 2007). Plantas
mutantes deficientes na biossíntese de ABA aumentam a resistência aos patógenos biotróficos
Hyaloperonospora parasítica e Blumeria graminis (MOHR e CAHILL, 2003; JENSEN et al.,
2008), enquanto, esse evento é comprometido para os necrotróficos A. brassicicola e Pythium
irregulare (TON e MAUCH-MANI, 2004; ADIE, et al., 2007). Baseado nisso, manifestações
específicas do papel emergente do ABA na modulação da resistência a doença pode depender
não só do estilo de vida do patógeno e biologia geral da infecção, mas, também, dos recursos
especializados de cada interação, indicando complexos mecanismos subjacentes à sinalização
por ABA na resposta das plantas a estresses bióticos (FAN et al., 2009; CAO et al., 2011).
Em contrapartida, uma proteína induzida por auxina (spot 15) foi superexpressa em
plantas suscetíveis de cajueiro, cultivadas em campo. Os efeitos da infecção de patógenos têm
sido mostrados, pelo menos em parte, como mediado por hormônios da planta, ou pela
própria capacidade do patógeno em produzir hormônios vegetais, durante a infecção, que
interferem no desenvolvimento e respostas de defesa (ROBERT-SEILANIANTZ et al., 2007;

104
NAVARRO et al., 2008; TSUKADA et al., 2010), sugerindo interação íntima entre a
regulação do crescimento e desenvolvimento, mediado por hormônios como a auxina, e a
reposta de resistência da planta contra patógenos (AGNEW et al., 2000; KORVES e
BERGELSON, 2003; POTTERS et al., 2007).
O efeito da auxina nas vias de sinalização para resistência à doença pode ser direto ou
indireto. Efeitos diretos podem envolver a interferência com vias de defesa da planta, por
exemplo, na sinalização por ácido salicílico, enquanto efeitos indiretos estão relacionados às
mudanças na progressão da interação hospedeiro-patógeno, causada por interferência da
auxina no desenvolvimento da planta (KAZAN e MANNERS, 2009).
Existem relatos de que a sinalização por auxina afeta, diferencialmente, a resistência
de plantas a grupos separados de patógenos, tornando-as mais resistentes a patógenos
necrotróficos (LLORENTE et al., 2008; MAH et al., 2012) e mais propensas à suscetibilidade
por patógenos biotróficos (NAVARRO et al., 2006; WANG et al., 2007). A superexpressão
de uma proteína induzida por auxina e, portanto, a possível presença de maiores
concentrações deste fitohormônio em plantas de cajueiro suscetível, podem tornar estas
plantas mais propensas ao ataque de patógenos secundários, induzindo condição de estresse
biótico, também favorável ao processo infeccioso iniciado por L. theobromae.
As proteínas responsivas a hormônios, citadas anteriormente, juntamente com
componentes relacionados à estrutura celular (spots S9, R5 e R6) foram diferencialmente
expressas somente em plantas de cajueiro cultivadas em condições de campo. Filamentos de
actina e microtúbulos formam uma estrutura tridimensional complexa e dinâmica nas células
vegetais (SCHMIDT e PANSTRUGA, 2004).
Componentes do citoesqueleto têm sido previamente identificados na análise
proteômica de plantas desafiadas com fitopatógenos (SINHA e CHATTOPADHYAY, 2011;
CASTILLEJO et al., 2012, MANDELC et al., 2013; SUN et al., 2014; PETRICCIONE et al.,
2013). Estímulos mecânicos que mimetizam o ataque de microrganismos são suficientes para
induzir a concentração dos microfilamentos de actina em células da epiderme foliar de A.
thalina, o que sugere a importância de sinais mecânicos de infecção na reorganização desses
filamentos (HARDHAM et al., 2008). Assim, tem sido sugerido que os microfilamentos de
actina se concentram no sítio de infecção e funcionam orientando o transporte polar de
compostos antimicrobianos (HIGAKI et al., 2011).

105
A presença de proteínas relacionadas a componentes estruturais identificadas em
plantas de cajueiro, cultivadas em campo, sugere haver eventos de reestruturação celular que
podem ser reprogramados nestas plantas por interação com o fitopatógeno L. theobromae.
5.3 Sobreposição da resposta proteômica do cajueiro à infecção pelo Lasiodiplodia
theobromae
Nosso estudo mostrou uma aparente sobreposição de respostas em plantas de cajueiro
infectadas com o L. theobromae. Essa observação se apoia no fato de que tanto plantas
inoculadas do clone BRS 226 (resistente), quanto à progênie desse clone, cultivada em
condições de campo, induzem mudanças fisiológicas no perfil de expressão das proteínas
associadas ao metabolismo e produção de energia, estresse e defesa, sinalização celular e
metabolismo/enovelamento de proteínas. Entre as proteínas diferencialmente expressas em
plantas de cajueiro cultivadas em condições de campo, 76% delas pertencem às categorias
citadas e que, por sua vez, foram diferencialmente reprogramadas nas plantas do clone BRS
226 infectadas (Figura 14A).
Interessantemente, além da alteração fisiológica de processos biológicos comuns, a
identidade compartilhada entre 6 proteínas diferencialmente reguladas, reforça essas
observações, sugerindo sobreposição e fornecendo consequente validação da reprogramação
em reposta a presença de L. theobromae, uma vez que, as plantas de cajueiro analisadas
nesses dois cenários compartilham este estresse biótico comum. Curiosamente, como
discutido acima, outros fatores abióticos e bióticos, propensos em condições de campo,
parecem alterar o padrão de expressão evidenciado em plantas classificadas como resistentes,
quando comparado ao clone BRS 226 inoculado.
A sobreposição, por análise proteômica comparativa de ecótipos resistentes e
suscetíveis de trigo, revelou a reprogramação de proteínas envolvidas nas mesmas funções
celulares, quando plantas pertencentes a estes grupos foram infectadas com o vírus SCMV
(Sugarcane mosaic virus). Os dados mostrados discutem uma possível expressão de proteínas
marcadoras de infecção, em plantas de trigo, com o vírus SCMV (WU et al., 2013).
Assim, a identidade comum de 6 proteínas (frutose-bifosfato aldolase, Hsp20-
chaperona, proteínas 14-3-3, subunidade beta do proteassoma e duas anexinas) apresentadas
nessa presente investigação, em especial daquelas envolvidas com sinalização celular
(anexinas e 14-3-3), parecem revelar possíveis alvos das respostas celulares envolvidas na
interação cajueiro-L. theobromae.

106
6. CONSIDERAÇÕES FINAIS
Mudanças no proteoma caulinar de plantas de cajueiro infectadas com L. theobromae
sugerem possível percepção do patógeno, seguido por alterações fisiológicas que induzem à
expressão diferencial de proteínas envolvidas em diversos processos celulares.
A possível ativação/supressão em componentes de vias biológicos do metabolismo,
produção de energia, sinalização celular ou induzidas sob estresse, em especial o oxidativo,
foram igualmente observadas para plantas de cajueiro cultivadas em campo, com alta pressão
do patógeno, bem como, para plantas inoculadas do clone BRS 226. A sobreposição de
respostas moleculares, em especial no que concerne a proteínas envolvidas com sinalização
celular, fornecem possíveis alvos resultantes da interação cajueiro-L. theobromae e reforçam
os eventos moleculares observados.
Dessa forma, os dados proteômicos apresentados neste trabalho fornecem informações
valiosas sobre os mecanismos biológicos subjacentes à infecção com L. theobromae e se
constituem numa base inicial de resultados que podem ser, futuramente, integrados a
programas de melhoramento genético do cajueiro, visando resistência, particularmente, à
resinose.

107
REFERÊNCIAS
ABDEL-FARID I. B.; JAHANGIR, M.; VAN DEN HONDEL, C. A. M. J.; KIM, H. K.;
CHOI, Y. H.; VERPOORTE, R. Fungal infection-induced metabolites in Brassica rapa.
Plant Science, v. 176, p. 608-615, 2009.
ABO-OGIALA, A.; CARSJENS, C.; DIEKMANN, H.; FAYYAZ, P.; HERRFURTH, C.;
FEUSSNER, I.; POLLE, A. Temperature-induced lipocalin (TIL) is translocated under
salt stress and protects chloroplasts from ion toxicity. Journal of Plant Physiology, v.
171, p. 250-259, 2014.
ABRAMOVITCH, R. B.; JANJUSEVIC, R.; STEBBINS, C. E.; MARTIN, G. B. Type III
effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and
immunity. Proceedings of the National Academy of Sciences, v. 103, p. 2851-2856, 2006.
ABUQAMAR, S.; CHAI, M.-F.; LUO, H.; SONG, F.; MENGISTE, T. Tomato protein kinase
1b mediates signaling of plant responses to necrotrophic fungi and insect herbivory. The Plant
Cell, v. 20, p. 1964-1983, 2008.
ADENIYI, D. O.; ORISAJO, S. B.; FADEMI, O. A.; ADENUGA, O. O.; DONGO, L. N.
Physiological studies of fungi complexes associated with cashew diseases. Journal of
Agricultural and Biological Science, v. 6, p. 34-38, 2011.
ADIE, B. A.; PÉREZ-PÉREZ, J.; PÉREZ-PÉREZ, M. M.; GODOY, M.; SÁNCHES-
SERRANO, J. J.; SCHMELS, E. A.; SOLANO, R. ABA is an essential signal for plant
resistance to pathogens affecting JA biosynthesis and the activation of defences in
Arabidopsis. Plant Cell, v. 19, p. 1665-1681, 2007.
AEBERSOLD, R.; MANN, M. Mass spectrometry-based proteomics. Nature, v. 422, p. 198-
207, 2003.
AFROZ, A.; ALI, G. M.; MIR, A.; KOMATSU, S. Application of proteomics to investigate
stress induced proteins for improvement in crop protection. Plant Cell Reports., v. 30, p. 745-
63, 2011.
AGNEW, P.; KOELLA, J. C.; MICHALAKIS, Y. Host life history responses to parasitism.
Microbes and Infection, v. 2, p. 891-896, 2000.
AHMAD S.; GORDON-WEEKS R.; PICKETT J.; TON J. Natural variation in priming of
basal resistance: from evolutionary origin to agricultural exploitation. Molecular Plant
Pathology, v. 11, p. 817-827, 2010.
ALVES, E. S.; CARDOSO, J. E.; SILVA, L. G. C.; LIMA, J. S. Interação das condições
edafoclimáticas com a resinose do cajueiro. Enciclopédia biosfera, v. 9, p. 1320-1345, 2013.
AL-WHAIBI, M. H. Plant heat-shock proteins: A mini review. Journal of King Saud
University-Science, v. 23, p. 139-150, 2011.

108
ALZATE-MARIN, A. L.; CERVIGNI, G. D. L.; MOREIRA, M. A.; BARROS, E. G. Seleção
assistida por marcadores moleculares visando ao desenvolvimento de plantas resistentes a
doenças, com ênfase em feijoeiro e soja. Fitopatologia Brasileira, v. 30, p. 333-342, 2005.
AN, C.; MOU, Z. Salicylic acid and its function in plant immunity. Journal of Integrative
Plant Biology, v. 53, p. 412-428, 2011.
ANDRADE, A. Sequenciamento, identificação e análise de proteínas do caule de mudas de
Eucalyptus grandis. 120p. Tese (Doutorado em Agronomia)- Escola Superior de Agricultura
“Luiz de Queiroz”, Universidade de São Paulo, Piracicaba – SP. 2006.
ANGOT, A.; VERGUNST, A.; GENIN, S.; PEETERS, N. Exploitation of eukaryotic
ubiquitin signaling pathways by effectors translocated by bacterial type III and type IV
secretion systems. PLoS Pathogens, v. 3, p. e3, 2007.
ARAI, Y.; HAYASHI, M.; NISHIMURA, M. Proteomic analysis of highly purified
peroxisomes from etiolated soybean cotyledons. Plant and Cell Physiology, v. 49, p. 526-539,
2008.
ARASIMOWICZ-JELONEK, M.; KOSMALA, A.; JANUS, Ł.; ABRAMOWSKI, D.;
FLORYSZAK-WIECZOREK, J. The proteome response of potato leaves to priming
agents and S-nitrosoglutathione. Plant Science, v. 198, p. 83-90, 2013.
ASAI, S.; YOSHIOKA , H. Nitric oxide as a partner of reactive oxygen species participates in
disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana.
Molecular Plant-Microbe Interactions, v. 22, p. 619-629, 2009.
ASSELBERGH, B.; DE VLEESSCHAUWER, D.; HOFTE, M. Global switches and fine-
tuning-ABA modulates plant pathogen defence. Molecular Plant-Microbe Interaction, v. 6,
p.709-719, 2008.
AXSEN, W. S.; STYER, C. M.; SOLNICK, J. V. Inhibition of heat shock protein expression
by Helicobacter pylori. Microbial Pathogenesis, v. 47, p. 231-236, 2009.
BAERENFALLER, K.; MASSONNET, C.; WALSH, S.; BAGINSKY, S.;
BUHLMANN, P,; HENNIG, L. et al. Systems-based analysis of Arabidopsis leaf growth
reveals adaptation to water deficit. Molecular Systems Biology, v. 8, p. 606, 2012.
BAIRD, R.; CARLING, D. Survival of parasitic anal saprophytic fungi or intact
senescent cotton roots. Journal of Cotton Science, v. 2, p. 27-34, 1998.
BANERJEE, J.; MAITI, M. K. Functional role of rice germin-like protein1 in regulation of
plant height and disease resistance. Biochemical and Biophysical Research Communications,
v. 394, p. 178-183, 2010.
BANTI, V.; MAFESSONI, F.; LORETI, E.; ALPI, A.; PERATA, P. The heat-inducible
transcription factor HsfA2 enhances anoxia tolerance in Arabidopsis. Plant Physiology,
v. 152, p. 1471-1483, 2010.

109
BARI, R.; JONES, J. D. G. Role of plant hormones in plant defense response. Plant
Molecular Biology, v. 69, p. 473-488, 2009.
BARROS, L. M. et al. Recomendações técnicas para a cultura do cajueiro anão precoce.
Fortaleza: EMBRAPA/CNPAT, 1993. 65p. (EMBRAPA-CNPAT. Circular
Técnica, 1).
BARROS, L.M. Caju. Produção: aspectos técnicos/editor técnico: Embrapa
Agroindústria Tropical (Fortaleza -CE). Brasília: Embrapa Informação Tecnológica,
2002. 148 p.; (Frutas do Brasil; 30).
BAXTER, A.; MITTLER, R.; SUZUKI, N. ROS as key players in plant stress signalling.
Journal of Experimental Botany, v.19, p. 1-12, 2013.
BECKERS, G. J.; SPOEL, S. H. Fine-tuning plant defence signaling: salicylate versus
jasmonate. Plant Biology Stuttg, v. 8, p.1-10, 2006.
BELLÉS, J. M.; LOPEZ-GRESA, M. P.; FAYOS, J.; PALLAS, V.; RODRIGO, I.;
CONEJERO, V. Induction of cinnamate 4-hydroxylase and phenylpropanoids in virus-
infected cucumber and melon plants. Plant Science, v. 174, p. 524-533, 2008.
BENSCHOP, J. J.; MOHAMMED, S.; O‟FLAHERTY, M.; HECK, A. J.; SLIJPER, M.;
MENKE, F. L. Quantitative phosphoproteomics of early elicitor signaling in Arabidopsis.
Molecular and Cellular Proteomics, v. 6, p. 1198-1214, 2007.
BENT, A. F.; MACKEY; D. Elicitors, effectors, and R genes: the new paradigm and a
lifetime supply of questions. Annual Review Phytopathology, v. 45, p. 399–436, 2007.
BERGER, S.; SINHA, A. K.; ROITSCH, T. Plant physiology meets phytopathology: plant
primary metabolism and plant-pathogen interactions. Journal of Experimental Botany, v. 58,
p. 4019-4026, 2007.
BEZERRA, M. A.; CARDOSO, J. E.; SANTOS, A. A.; VIDAL, J. C.; ALENCAR, E. S.
Efeito da resinose na fotossíntese do cajueiro anão precoce. Fortaleza: Embrapa
Agroindústria Tropical (Boletim de Pesquisa e Desenvolvimento, 8), 2003, 11p.
BEZERRA, M. A.; LACERDA, C. F.; GOMES FILHO, E. ABREU, C. E. B.; PRISCO, J. T.
Physiology of cashew plants grown under adverse conditions. Brazilian Journal of Plant
Physiology, v. 19, , p. 449-461, 2007.
BHADAURIA, V.; BANNIZA, S.; WANG, L. X.; WEI, Y. D.; PENG, Y. L. Proteomic
studies of phytopathogenic fungi, oomycetes and their interactions with hosts. European
Journal of Plant Pathology, v. 126, p. 81-89, 2010.
BHUIYAN, N. H.; SELVARAJ, G.; WEI, Y.; KING, J. Role of lignification in plant defense.
Plant Signaling & Behavior, v. 4, p. 158-159, 2009.
BIRCH, P. R.; ARMSTRONG, M.; BOS, J.; BOEVINK, P.; GILROY, E. M.; TAYLOR, R.
M.; WAWRA, S.; PRITCHARD, L.; CONTI, L.; EWAN, R.; WHISSON, S. C.; VAN
WEST, P.; SADANANDOM, A.; KAMOUN, S. Towards understand-ing the virulence

110
functions of RXLR effectors of the oomycete plant pathogen Phytophthora infestans.
Journal of Experimental Botany, v. 60, p.1133-1140, 2009.
BITTEL, P.; ROBATZEK, S. Microbe-associated molecular patterns (MAMPs) probe plant
immunity. Current Opinion in Plant Biology, v. 10, p.335–341, 2007.
BLUME, B.; NURNBERGER, T.; NASS, N.; SCHEEL, D. Receptor-mediated increase in
cytoplasmic free calcium required for activation of pathogen defense in parsley. Plant Cell, v.
12, p.1425-1440, 2000.
BOLLER, T. Biochemical analysis of chitinases and β-1,3-glucanases. In: Molecular Plant
Pathology. Edited by S.J. GURR, M. J. McPHERSON, and D. J. BOWLES, IRL Press, New
York, v. 2, p. 23-30, 1992.
BOLLER, T.; FELIX, G. A renaissance of elicitors: perception of microbe-associated
molecular patterns and danger signals by pattern-recognition receptors. Annual Review in
Plant Biology, v. 60, p. 379-406, 2009.
BOLTON, M. D. Primary Metabolism and Plant Defense-Fuel for the Fire. Molecular Plant-
Microbe Interactions, v. 22, p. 487-497, 2009.
BRADFORD, M. M. A rapid and sensitive method for quantification of micrograms
quantities of protein utilizing the principle of protein-dye binding. Analytical
Biochemistry, v. 72, p. 248-254, 1976.
BRINKER, M.; BROSCHE, M.; VINOCUR, B.; ABO-OGIALA, A.; FAYYAZ, P.; JANZ,
D.; OTTOW, E. A.; CULLMANN, A. D.; SABOROWSKI, J.; KANGASJARVI, J.;
ALTMAN, A.; POLLE, A. Linking the salt transcriptome with physiological responses of
a salt-resistant Populus species as a strategy to identify genes important for stress
acclimation. Plant Physiology, v. 154, p. 1697-709, 2010.
BRITTON, K. O.; HENDRIX, F. F. Population dynamics of Botryosphaeria spp. in peach
gummosis canker. Plant Disease, v. 70, p. 134-136, 1986.
BRUGIERE, S.; KOWALSKI, S.; FERRO, M.; SEIGNEURIN-BERNY, D.; MIRAS, S.;
SALVI, D.; RAVANEL, S.; D‟HERIN, P.; GARIN, J.; BOURGUIGNON, J.; JOYARD, J.;
ROLLAND, N. The hydrophobic proteome of mitochondrial membranes from Arabidopsis
cell suspensions. Phytochemistry, v. 65, p. 1693-1707, 2004.
BRUNNER, F.; ROSAHL, S.; LEE, J.; RUDD, J. J.; GEILER, C. et al. Pep-13, a plant
defense-inducing pathogen-associated pattern from Phytophthora transglutaminases. EMBO
Journal, v. 21, p.6681–6688, 2002.
BURCH-SMITH, T. M.; SCHIFF, M.; CAPLAN, J. L.; TSAO, J.; CZYMMEK, K.;
DINESH-KUMAR, S. P. A novel role for the TIR domain in association with pathogen-
derived elicitors. PLoS Biology, v. 5, p. e68, 2007.
CAJUNOR. Especificações técnicas: cajueiro anão precoce. Disponível em: http://www.cajunor.com.br/noticia_detalhe.php?id=19. Acesso em: 20/12/2013.
CANDIANO, G.; BRUSCHI, M.; MUSANTE, L. et al., Blue silver: a very sensitive

111
coloidal coomassie G-250 staining for proteome analysis. Electrophoresis, v. 25, p.
1327-1333, 2004.
CANTÚ, M. D.; CARRILHO, E.; WULFF, N. A.; PALMA, M. S. Seqüenciamento de
peptídeos usando espectrometria de massas: um guia prático. Química nova, v. 31, p. 669-
675, 2008a.
CANTÚ, M. D.; MARIANO, A. G.; PALMA, M. S.; CARRILHO, E.; WULFF, N. A.
Proteomic analysis reveals suppression of bark chitinases and proteinase inhibitors in citrus
plants affected by the citrus sudden death disease. Phytopathology, v. 98, p.1084-1092,
2008b.
CAO, F. Y.; YOSHIOKA, K.; DESVEAUX, D. The roles of ABA in plant-pathogen
interactions. Journal of Plant Research, v. 124, p. 489-499, 2011.
CAO, T.; SRIVASTAVA, S.; RAHMAN, M. H.; KAV, N. N. V. M.; HOTTE, N.;
DEYHOLOS, M. K.; STRELKOV, S. E. Proteome-level changes in roots of Brassica
napus as a result of Plasmodiophora brassica infection. Plant Science, v. 174, p. 97-115,
2008.
CAPLAN, J. L.; ZHU, X.; MAMILLAPALLI, P.; MARATHE, R.; ANANDALAKSHMI, R.;
DINESH-KUMAR, S. P. Induced ER Chaperones Regulate a Receptor-like Kinase to
Mediate Antiviral Innate Immune Response in Plants. Cell Host & Microbe, v. 6, p. 457-469,
2009.
CARDOSO, J. E.; ARAGÃO, M. L.; BLEICHER, E.; CAVALCANTE, M. J. B. Efeito de
práticas agronômicas na ocorrência da resinose do cajueiro. Fitopatologia Brasileira, v. 20, p.
242, 1995.
CARDOSO, J. E.; FREIRE, F.C. O. Doenças. In: BARROS, L. M. (Coord.).
Caju produção: aspectos técnicos. Fortaleza: Embrapa Agroindústria Tropical,
2002. p. 132-136. (Frutas do Brasil, 30).
CARDOSO, J. E.; SANTOS, A. A.; ROSSETTI, A. G.; VIDAL, J. C. Relationship
between incidence and severity of gummosis in semiarid northeastern Brazil. Plant
Pathology, v. 53, p. 363-367, 2004.
CARDOSO, J. E.; PAIVA, J. R.; CAVALCANTI, J. J. V.; SANTOS, A. A.; VIDAL, J. C.
Evaluation of resistance in dwarf cashew to gummosis in northeastern Brasil. Crop
Protection, v. 25, p. 855-859, 2006.
CARDOSO, J. E.; VIANNA, F. M. P.; CYSNE, A. Q.; FARIAS, F. C.; SOUZA, R. N. M.
Clone Embrapa 51: uma alternative para resistência à resinose-do-cajueiro.
Fortaleza: Embrapa Agroindústria Tropical (Comunicado Técnico, 130), 2007, 3p.
CARDOSO, J. E.; WILKINSON, M. J. Development and characterisation of microsatellite
markers for the fungus Lasiodiplodia theobromae. Summa
Phytopathologica, v.34, p. 55-57, 2008.

112
CARDOSO, J. E.; BEZERRA, M. A.; VIANA, F. M. P.; SOUSA, T. R. M.; CYSNE, A. Q.; F
ARIAS, F. C. Ocorrência endofítica de Lasiodiplodia theobromae em tecidos de cajueiro e
sua transmissão por propágulos. Summa phytopathologica, v. 354, p. 262-266, 2009a.
CARDOSO, J. E.; VIANA, F. M. P.; BEZERRA, M. A.; SOUSA, T. R. M.; CYSNE, A. Q.;
FARIAS, F. C. Transmissão de Lasiodiplodia theobromae, agente da resinose, em
propágulos de cajueiro. Fortaleza: Embrapa Agroindústria Tropical (Boletim de Pesquisa e
Desenvolvimento, 34), 2009 b, 21p.
CARDOSO, J. E.; VIANA, F. M. P.; FREIRE, F. C. O.; CYSNE, A. Q.; FARIAS, F. C.;
CAVALCANTI, J. J. V. Manejo da resinose do cajueiro. Fortaleza: Embrapa Agroindústria
Tropical (Comunicado Técnico, 154), 2009c, 3p.
CARDOSO, J. E.; CAVALCANTI, J. J. V.; CYSNE, A. Q.; SOUSA, T. R. M.; CORRÊA, M.
C. M. Interação enxerto e porta-enxerto na incidência da resinose do cajueiro. Revista
Brasileira de Fruticultura, v. 32, p. 847-854, 2010.
CASTILLEJO, M. A.; AMIOUR, N.; DUMAS-GAUDOT, E.; RUBIALES, D.; JORRÍN, J.
V. A proteomic approach to studying plant response to crenate broomrape (Orobanche
crenata) in pea (Pisum sativum). Phytochemistry, v. 65, p. 1817-1828, 2004.
CASTILLEJO, M. A.; FERNÁNDEZ-APARICIO, M.; RUBIALES, D. Proteomic analysis
by two-dimensional differential in gel electrophoresis (2D DIGE) of the early response of
Pisum sativum to Orobanche crenata. Journal of Experimental Botany, v. 63, p. 107-119,
2012.
CAVALCANTI, M. L. F.; FERNANDES, P. D.; GHEYI, H. R.; JÚNIOR, G. B. Fisiologia do
Cajueiro Anão precoce submetido à estresse hídrico em fases fenológicas . Revista de
Biotecnologia e Ciências da Terra, v. 8, p.42-53, 2008.
CEITA, G. O.; MACÊDO, J. N. A.; SANTOS, T. B.; ALEMANNO, L.; GESTEIRA, A. S.;
MICHELI, F.; MARIANO, A. C.; GRAMACHO, A. P.; SILVA, D. C.; MEINHARDT, L.;
MAZZAFERA, P.; PEREIRA, G. A. G. Involvement of calcium oxalate degradation during
programmed cell death in Theobroma cacao tissues triggered by the hemibiotrophic fungus
Moniliophthora perniciosa. Plant Science, v. 173, p. 106-117, 2007.
CITOVSKY, V.; ZALTSMAN, A.; KOZLOVSKY, S. V.; GAFNI, Y.; KRICHEVSKY, A.
Proteasomal degradation in plant-pathogen interactions. Seminars in Cell & Developmental
Biology, v. 20, p. 1048-1054, 2009.
CHAMRAD, D. C.; KORTING, G.; STUHLER, K.; MEYER, H. E.; KLOSE, J.; BLUGGEL,
M. Evaluation of algorithms for protein identification from sequence databases using mass
spectrometry data. Proteomics, v. 4, p. 619- 628, 2004.
CHANDRAN, D; INADA, N.; HATHER, G.; KLEINDT, C. K.; WILDERMUTH, M. C.
Laser microdissection of Arabidopsis cells at the powdery mildew infection site reveals site-
specific processes and regulators. Proceedings of the National Academy of Sciences, v. 107, p.
460-465, 2010.

113
CHANG, I. F.; CURRAN, A.; WOOLSEY, R.; QUILICI, D.; CUSHMAN, J. C.; MITTLER,
R.; HARMON, A.; HARPER, J. F. Proteomic profiling of tandem affinity purified 14-3-3
protein complexes in Arabidopsis thaliana. Proteomics, v. 9, p. 2967-2985, 2009.
CHARRON, J.-B. F.; OUELLET, F.; PELLETIER, M.; DANYLUK, J.; CHAUVE, C.;
SARHAN, F. Identification, expression, and evolutionary analyses of plant lipocalins.
Plant Physiology, v. 139, p. 2017-2028, 2005.
CHARRON, J.-B. F.; OUELLET, F.; HOUDE, M.; SARHAN, F. The plant Apolipoprotein D
ortholog protects Arabidopsis against oxidative stress. BMC Plant Biology, v. 8, p. 1-12,
2008.
CHEN, F.; LI, Q.; SUN, L.; HE, Z. The rice 14-3-3 gene family and its involvement in
responses to biotic and abiotic stress. DNA Research, v.13, p. 53-63, 2006.
CHEN, S.; HARMON, A. L. Advances in plant proteomics. Proteomics, v. 6, p. 5504-5516,
2006.
CHEN, Z. R., ZHOU, T.; WU, X.; HONG, Y.; FAN, Z.; LI, H. Influence of cytoplasmic heat
shock protein 70 on viral infection of Nicotiana benthamiana. Molecular Plant Pathology, v.
9, p. 809-817, 2008.
CHENG, Z.; MCCONKEY, B. J.; GLICK, B. R. Proteomic studies of plant-bacterial
interactions. Soil Biology & Biochemistry, v. 42, p. 1673-1684, 2010.
CHI, W.-T.; FUNG, R. W. M.; LIU, H.-C.; HSU, C.-C.; CHARNG, Y.-Y. Temperature-
induced lipocalin is required for basal and acquired thermotolerance in Arabidopsis.
Plant Cell and Environment, v. 32, p. 917-927, 2009.
CHRISTIS, C.; LUBSEN, N. H.; BRAAKMAN, I. Protein folding includes oligomerization-
examples from the endoplasmic reticulum and cytosol. FEBS Journal, v.275, p. 4700-4727,
2008.
CHUANG, S.M.; CHEN, L.; LAMBERTSON, D.; ANAND, M.; KINZY, T. G.; MADURA,
K. Proteasome-mediated degradation of cotranslationally damaged proteins involves
translation elongation factor 1A. Molecular and Cell Biology, v. 25, p. 403-413, 2005.
CLARK, G.; KONOPKA-POSTUPOLSKA, D.; HENNIG, J.; ROUX, S. Is annexin 1 a
multifunctional protein during stress responses? Plant Signaling & Behavior, v. 5, p. 303-307,
2010.
COLLINS, R. M.; AFZAL, M.; WARD, D. A.; PRESCOTT, M. C.; SAIT, S. M.; REES, H.
H.; TOMSETT, A. B. Differential proteomic analysis of Arabidopsis thaliana genotypes
exhibiting resistance or susceptibility to the insect herbivore, Plutella xylostella. Plos one, v.
5, p.1-14, 2010.
CRISÓSTOMO, L. A.; SANTOS, F. J. S.; OLIVEIRA, V. H.; RAIJ, V. B.; BERNARDI, A.
C. C.; SILVA, C. A.; SOARES, I. Cultivo do cajueiro anão precoce: aspectos fitotécnicos
com ênfase na adubação e na irrigação. Fortaleza: Embrapa Agroindústria Tropical
(Comunicado Técnico, 10), 2001, 10p.

114
CRISÓSTOMO, J. R.; CAVALCANTI, J. J. V.; BARROS, L. M., ALVES, R. E.; FREITAS,
J. G.; OLIVEIRA, J. N. Melhoramento do cajueiro-anão-precoce: avaliação da qualidade do
pedúnculo e a heterose dos seus híbridos. Revista Brasileira de Fruticultura, v. 24, p. 477-
480, 2002.
CUNHA, R.B., CASTRO, M.S., FONTES, W. Espectrometria de massa de proteínas.
Biotecnologia, Ciência e Desenvolvimento, ano IX, n. 36, 2006.
CURTIS, R. H. C. Plant parasitic nematode proteins and the host-parasite interaction.
Briefings in Functional Genomics and Proteomics, v. 6, p. 50 -58, 2007.
CYSNE, A. Q.; CARDOSO, J. E.; MAIA, A. H. N.; FARIAS, F. C. Spatial-temporal analysis
of gummosis in three cashew clones at Northeastern Brazil. Journal of Phytopathology, v.158,
p. 676-682, 2010.
DANGL, J.L.; JONES, J.D. Plant pathogens and integrated defence responses to infection.
Nature, v. 411, p. 826–833, 2001.
DASSO, M. The Ran GTPase: theme and variations. Current Biology, v. 12, p. R502-508,
2002.
DAVIDSON, R. M.; MANOSALVA, P. M.; SNELLING, J.; BRUCE, M.; LEUNG, H.;
LEACH, J. E. Rice germin-like proteins: allelic diversity and relationships to early stress
responses. Rice, v. 3, p. 43-55, 2010.
DAVIES, D. D.; GREGO, S.; KENWORTHY, P. The control of the production of lactate and
ethanol by higher plants. Planta, v. 118, p. 297-310, 1974.
DELAURÉ, S. L.; HEMELRIJCK, W. V.; DE BOLLE, M. F. C.; CAMMUE, B. P. A.; DE
CONINCK, B. M. A. Building up plant defenses by breaking down proteins. Plant Science, v.
174, p. 375-385, 2008.
DEMPSEY, D. A.; KLESSIG, D. F. SOS – too many signals for systemic acquired
resistance? Trends in Plant Science, v. 17(9), p. 538-545, 2012.
DENISON, F. C.; PAUL, A.-L.; ZUPANSKA, A. K.; FERL, R. J. 14-3-3 proteins in plant
physiology. Seminars in Cell & Developmental Biology, v. 22, p. 720-727, 2011.
DEVOTO, A.; MUSKETT, P. R.; SHIRASU, K. Role of ubiquitination in the regulation of
plant defence against pathogens. Current Opinion in Plant Biology, v. 6, p. 307-311, 2003.
DIAS, C.V.; MENDES, J. S.; DOS SANTOS, A. C.; PIROVANI, C. P.; SILVA, A.;
GESTEIRA, F. MICHELI, GRAMACHO, K. P.; HAMMERSTONE, J.; MAZZAFERAF, P.; CASCARDO, J. C. M. Hydrogen peroxide formation in cacao tissues infected by the
hemibiotrophic fungus Moniliophthora perniciosa. Plant Physiology and Biochemistry, v. 49,
p. 917-922, 2009.
DÓCZI, R.; BRADER, G.; PETTKÓ-SZANDTNER, A.; RAJH, I.; DJAMEI, A.;
PITZSCHKE, A.; TEIGE, M.; HIRT, H. The Arabidopsis mitogen-activated protein kinase

115
kinase MKK3 is upstream of group C mitogen-activated protein kinases and participates in
pathogen signaling. Plant Cell, v.19, p. 3266–3279, 2007.
DUDLER, R. Manipulation of host proteasomes as a virulence mechanism of plant
pathogens. Annual Review of Phytopathology, v. 51, p. 521-542, 2013.
DUNWELL, J.; GIBBINGS, J.; MAHMOOD, T.; NAQVI, S. S. Germin and germin-like
proteins: evolution, structure, and function. Critical Reviews in Plant Sciences, v. 27, p. 342-
375, 2008.
EFEOĞLU, B. Heat Shock Proteins and Heat Shock Response in Plants. Journal of Science,
v. 22, p. 67-75, 2009.
ELIAS, J. E.; HAAS, W.; FAHERTY, B. K.; GYGI, S. P. Comparative evaluation of mass
spectrometry platforms used in large-scale proteomics investigations. Nature Methods, v. 2, p.
667-675, 2005.
ELLIS, J.G.; DODDS, P.N.; LAWRENCE, G.J. Flax rust resistance gene specificity is based
on direct resistance-avirulence protein interactions. Annual Review of Phytopathology, v. 45,
p. 289–306, 2007.
ELLIS, J. G.; RAFIQI, M.; GAN, P.; CHAKRABARTI, A.; DODDS, P. N. Recent progress
in discovery and functional analysis of effector proteins of fungal and oomycete plant
pathogens. Current Opinion in Plant Biology, v. 12, p. 399-405, 2009.
EL OIRDI, M.; EL RAHMAN, T. A.; RIGANO, L.; EL HADRAMI, A.;
RODRIGUEZ, M. C.; DAAYF, F.; VOJNOV, A.; BOUARAB, K. Botrytis cinerea
manipulates the antagonistic effects between immune pathways to promote disease
development in tomato. Plant Cell, v. 23, p. 2405-2421, 2011.
EL-SHORA , H. M. Properties of phenylalanine ammonia-lyase from Marrow cotyledons.
Plant Science, v. 162, p. 1-7, 2002.
ELVIRA, M. I.; GALDEANO, M. M.; GILARDI, P.; GARCIA-LUQUE, I.; SERRA,
M.T. Proteomic analysis of pathogenesis-related proteins (PRs) induced by compatible
and incompatible interaction of pepper mild mottle virus (PMMoV) in Capsicum
chinense L plants. Journal of Experimental Botany, v. 59, p. 1253-1265, 2008.
ENCINAS, O., 1996. Development and Significance of Attack by Lasiodiplodia theobromae
(Pat.)Griff.&Maubl. In Caribbean Pine Wood and Some Other Wood Species.Uppsala,
Sweden: Swedish University of Agricultural Sciences, PhD Tese.
FAN, J.; HILL, L.; CROOKS, C.; DOERNER, P.; LAMB, C. Abscisic acid has a key role in
modulating diverse plant-pathogen interactions. Plant Physiology, v. 150, p. 1750-1761,
2009.
FAN, J.; CHEN, C.; YU, Q.; BRLANSKY, R. H.; LI, Z.-G.; GMITTER JR, F. G.
Comparative iTRAQ proteome and transcriptome analyses of sweet orange infected by
„„Candidatus Liberibacter asiaticus‟‟. Physiologia Plantarum, v. 143, p. 235-245, 2011.

116
FANATA, W. I. D.; LEE, S. Y.; LEE, K. O. The unfolded protein response in plants: a
fundamental adaptive cellular response to internal and external stresses. Journal of
Proteomics, v. 93, p. 356-368, 2013.
FANG, X.; Chen, W.; XINA, Y.; ZHANG, H.; YAN, C.; YU, H.; LIU, H.; XIAO, W.;
WANGA, S.; ZHENG, A.;, LIU, H.; JIN, L.; MA, H.; RUAN, S. Proteomic analysis of
strawberry leaves infected with Colletotrichum fragariae. Journal of Proteomics, v. 75,
p. 4074-4090, 2012.
FAO (Food and Agriculture Organization of the United Nations). FAOSTAT, 2008.
Disponível em: http://faostat.fao.org/. Acesso em 24/10/2008.
FAUROBERT, M.; PELPOIR, E.; CHAÏB, J. Phenol extraction of proteins for proteomic
studies of recalcitrant plant tissues. In: THIELLEMENT, H.; ZIVY, M.; DAMERVAL, C.;
MÉCHIN, V. In: Plant Proteomics: methods and protocols. Totowa : Humana Press, 2007,
416p.
FELIX, G.; BOLLER, T. Molecular sensing of bacteria in plants. The highly conserved RNA-
binding motif RNP-1 of bacterial cold shock proteins is recognized as an elicitor signal in
tobacco. Journal of Biological Chemistry, v. 278, p. 6201-6208, 2003.
FELLE, H. H.; HERRMANN, A.; HÜCKELHOVEN, R.; KOGEL, K.-H. Root-to-shoot
signalling: apoplastic alkalinization, a general stress response and defence factor in barley
(Hordeum vulgare). Protoplasma, v. 227, p. 17-24, 2005.
FERNANDES, J. B.; HOLANDA, J. S. de.; CHAGAS, M. C. M. das.; LIMA, J. M. P.;
OLIVEIRA, J. S. F.; Recomendações Técnicas para o Cultivo do Cajueiro. Natal: EMPARN,
2009. 21p.
FERNÁNDEZ, R. G.; JESÚS, E. P.; JORRÍN-NOVO, V. Proteomics of Plant Pathogenic
Fungi. Journal of Biomedicine and Biotechnology, v. 2010, p. 1-36, 2010.
FERNIE, A. R.; CARRARI, F.; SWEETLOVE, L. J. Respiratory metabolism: Glycolysis, the
TCA cycle and mitochondrial electron transport. Current Opinion in Plant Biology, v. 7, p.
254-261, 2004.
FINKA, A.; CUENDET, A. F.; MAATHUIS, F. J.; SAIDI, Y.; GOLOUBINOFF, P. Plasma
membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and
acquired thermotolerance.The Plant Cell, v. 24, p. 3333-3348, 2012.
FLOR, H. H. Host-parasite interaction in flax rust: its genetic and other implications.
Phytopathology, v. 45, p. 680-685, 1955.
FRAIRE-VELÁZQUEZ, S.; RODRÍGUEZ-GUERRA, R.; SÁNCHEZ-CALDERÓN, L.
2011. Abiotic and Biotic Stress Response Crosstalk in Plants, Abiotic Stress Response in
Plants - Physiological, Biochemical and Genetic Perspectives, Prof. Arun Shanker (Ed.),
ISBN: 978-953-307-672-0. In Tech, Disponível em:
http://www.intechopen.com/books/abiotic-stress-response-in-plants-physiological-
biochemical-and-genetic-perspectives/abiotic-and-biotic-stress-response-crosstalk-in-plants.

117
FREEMAN, B.C.; BEATTIE, G.A. 2008. An Overview of Plant Defenses against Pathogens
and Herbivores. The Plant Health Instructor. DOI: 10.1094/PHI-I-2008-0226-01.
FREIRE, F. C. O. A resinose do cajueiro. Caju Informativo, Fortaleza: 4 (1-2), 1991.
FREIRE, F. C. O.; CARDOSO, J. E. Doenças das Spondias-cajarana (S. cytherera
Sonn.), cajazeira (S. mombin L.), ciriguela (S. purpurea L.), umbu (S. tuberosa A.
Camp.) e umbuguela (Spondia sp.) no Brasil. Agrotrópica, v. 9, p.75-82, 1997.
FREIRE, F. C. O.; CARDOSO, J. E. Doenças do coqueiro. In: FREIRE, F. C. O.;
CARDOSO, J. E.; VIANA, F. M. P. (Ed.) Doenças de fruteiras tropicais de interesse
agroindustrial. Brasília: EMBRAPA, p.191-226, 2003.
FREIRE, F. C. O.; VIANA, F. M. P.; CARDOSO, J. E.; SANTOS, A. A. Novos hospedeiros
dos fungo Lasiodiplodia theobromae no estado do Ceará. Fortaleza: Embrapa Agroindústria
Tropical, (Comunicado Técnico, 91), 2004. 6 p.
FRYDMAN, J. Folding of newly translated proteins in vivo: the role of molecular
chaperones. Annual Review of Biochemistry, v. 70, p. 603-647, 2001.
FU, J.; MOMCILOVIC, I.; PRASAD, P. V. V. Roles of protein synthesis elongation factor
ef-tu in heat tolerancein plants. Journal of Botany, v. 2012, p. 1-8, 2012.
FUJII, H.; ZHU, J.-K. Arabidopsis mutant deficient in three abscisic acid-activated protein
kinases reveals critical roles in growth, reproduction and stress. Proceedings of the National
Academy of Sciences, v. 106, p. 8380-8385, 2009.
FUJITA, M.; FUJITA, Y.; NOUTOSHI, Y.; TAKAHASHI, F.; NARUSAKA, Y.;
YAMAGUCHI-SHINOGAKI K.; SHINOZAKI, K. Crosstalk between abiotic and biotic
stress responses: a current view from the points of convergence in the stress signaling
networks. Current Opinion in Plant Biology, v. 9, p. 436-442, 2006.
FULGOSI, H.; SOLL, J.; DE FARIA MARASCHIN, S.; KORTHOUT, H. A.; WANG, M.;
TESTERINK, C. 14-3-3 proteins and plant development. Plant Molecular Biology, v. 50, p.
1019-1029, 2002.
FUNDAÇÃO BANCO DO BRASIL. Os frutos sociais do caju. N T Mendes Editora.
203p., 2010.
GAFFNEY, T.; FRIEDRICH L.; VERNOOIJ, B.; NEGROTTO, D.; NYE, G.; UKNES
S.;WARD, E.; KESSMANN, H.; RYALS, J. Requirement of salicylic acid for the induction
of systemic acquired resistance. Science, v. 261, p. 754-756, 1993.
GANFORNINA, M. D.; GUTIÉRREZ, G.; BASTIANI, M.; SÁNCHEZ, D. A phylogenetic
analysis of the lipocalin protein family. Molecular Biology and Evolution, v. 17, p. 114-126,
2000.
GARAVAGLIA, B. S.; GAROFALO, C. G.; ORELLANO, E. G.; OTTADO, J. Hsp70 and
Hsp90 expression in citrus and pepper plants in response to Xanthomonas

118
axonopodis pv. citri, European Journal Plant Pathology, v. 123, p. 91-97, 2009.
GAROFALO, C. G.; GARAVAGLIA, B. S.; DUNGER, G.; GOTTIG, N.; ORELLANO, E.
G.; OTTADO, J. Expression analysis of small heat shock proteins during compatible and
incompatible plant-pathogen interactions. Advanced Studies in Biology, v. 1, p. 197-205,
2009.
GASIOROWSKI, J. Z.; DEAN, D. A. Mechanisms of nuclear transport and interventions.
Advances Drug Delivery Review, v. 55, p. 703-716, 2003.
GAUCI, V. J.; PADULA, M. P.; COORSSEN, J. R. Coomassie blue staining for high
sensitivity gel-based proteomics. Journal of proteomics, v. 9 0, p. 96-106, 2013.
GERBER, I. B.; LAUKENS, K.; DE VIJLDER, T.; WITTERS, E.; DUBERY, I. A.
Proteomic profiling of cellular targets of lipopolysaccharide-induced signalling in Nicotiana
tabacum BY-2 cells. Biochimica et Biophysica Acta, v. 1784, p. 1750-1762, 2008.
GIDALEVITZ, T.; STEVENS, F.; ARGON, Y. Orchestration of secretory protein folding by
ER chaperones. Biochimica et Biophysica Acta, v. 1833, p. 2410-2424, 2013.
GILL, S. S.; TUTEJA, N. Reactive oxygen species and antiox-idant machinery in abiotic
stress tolerance in crop plants. Plant Physiology and Biochemistry, v. 48, p. 909-930, 2010.
GIMENEZ-IBANEZ, S.; HANN, D. R., NTOUKAKLS, V.; PETUTSCHNIG, E.; LIPKA,
V.; RATHJEN, J. P. AvrPtoB targets the LysM receptor kinase CERK1 to promote bacterial
virulence on plants. Current Biology, v. 19, p.423-429, 2009.
GLAZEBROOK, J. Contrasting mechanisms of defense against biotrophic and
necrotrophic pathogens. Annual Review of Phytopathology, v. 43, p. 205-227, 2005.
GONDIM, D. M. F. Bases Bioquímicas da Interação do Cajueiro Anão Precoce com o
Fungo Lasiodiplodia theobromae causador da Resinose. 143p. Tese (Doutorado em
Bioquímica). Departamento de Bioquímica e Biologia Molecular, Universidade
Federal do Ceará, Fortaleza - CE. 2010.
GOHRE, V.; SPALLEK, T.; HAEWEKER, H.; MERSMANN, S.; MENTZEL, T., TOREES,
M., MANSFIELD, J. W.; ROBATZEK, S. Plant pattern-recognition receptor FLS2 is directed
for degradation by the bacterial ubiquitin ligase AvrPtoB. Current Biology, v.18, p. 1824-
1832, 2008.
GOVRIN, E. M.; LEVINE, A. The hypersensitive response facilitates plant infection by the
necrotrophic pathogen Botrytis cinerea. Current Opinion in Plant Biology, v. 10, p. 751-757,
2000.
GROSS, S. R.; KINZY, T. G. Translation elongation factor 1A is essential for regulation of
the actin cytoskeleton and cell morphology. Nature Structural & Molecular Biology, v. 12, p.
772-778, 2005.
GROßKINSKY, D. K.; GRAAFF, E. V. D.; ROITSCH, T. Phytoalexin transgenics in crop
protection-Fairy tale with a happy end? Plant Science, v. 195, p. 54-70, 2012.

119
GRUBER, C. W.; CEMAZAR, M.; CLARK, R. J.; HORIBE, T.; RENDA, R. F.;
ANDERSON, M. A.; CRAIK, D. J. A novel plant protein-disulfide isomerase involved in the
oxidative folding of cystine knot defense proteins. Journal of Biological Chemistry, v. 282, p.
20435-2046, 2007.
GRZYB, J.; LATOWSKI, D.; STRZAŁKA, K. Lipocalins - a family portrait. Journal of
Plant Physiology, v. 163, p. 895-915, 2006.
GUANZIROLI, C. E.; SOUZA, H. M.; JÚNIOR, A. V.; BASCO, C. A. Entraves ao
desenvolvimento da cajucultura no nordeste: margens de comercialização ou aumentos de
produtividade e de escala? Revista Extensão Rural, v. 16, p. 96-122, 2009.
GURURANI, M. A.; VENKATESH, J.; UPADHYAYA, C. P., et al. Plant disease resistance
genes: Current status and future directions. Physiological and Molecular Plant Pathology, v.
78, p. 51-65, 2012.
HAMMOND-KOSACK, K.E.; KANYUKA, K. Resistance genes (R genes) in plants. eLS,
2007.
HARDHAM, A. R.; TAKEMOTO, D.; WHITE, R. G. Rapid and dynamic subcellular
reorganization following mechanical stimulation of Arabidopsis epidermal cells mimics
responses to fungal and oomycete attack. BMC Plant Biology, v. 8, p. 63, 2008.
HARTL, F. U.; HAYER-HARTL, M. Molecular chaperones in the cytosol: from nascent
chain to folded protein. Science, v. 295, p. 1852-1858, 2002.
HASEGAWA, M.; MITSUHARA, I.; SEO, S.; IMAI, T.; KOGA, J.; OKADA, K.;
YAMANE, H.; OHASHI, Y. Phytoalexin accumulation in the interaction between rice and
the blast fungus. Molecular Plant-Microbe Interactions, v. 23, p. 1000-1011, 2010.
HATAHET, F.; RUDDOCK, L. W. Protein disulfide isomerase: a critical evaluation of its
function in disulfide bond formation. Antioxidant Redox Signaling, v. 11, p. 2807-2850, 2009.
HEIN, I.; GILROY, E. M.; ARMSTRONG, M. R.; BIRCH, P. R. The zig-zag-zig in
oomycete-plant interactions. Molecular Plant Pathology, v. 10, p. 547-562, 2009.
HIGAKI, T.; KURUSU, T.; HASEZAWA, S.; KUCHITSU, K. Dynamic intracellular
reorganization of cytoskeletons and the vacuole in defense responses and hypersensitive cell
death in plants. Journal of Plant Research, v. 124, p. 315-24, 2011.
HIRAYAMA, T.; SHINOZAKI, K. Perception and transduction of abscisic acid signals: keys
to the function of the versatile plant hormone ABA. Trends in Plant Science, v. 12, p. 343-
351, 2007.
HOK, S.; ATTARD, A.; KELLER, H. Getting the most from the host: How pathogens force
plants to cooperate in disease. Molecular Plant-Microbe Interactions, v. 23, p. 1253-1259,
2010.
HOSHINO, D.; HAYASHI, A.; TEMMEI, Y.; KANZAWA, N.; TSUCHIYA, T.
Biochemical and immunohistochemical characterization of Mimosa annexin. Planta, v. 219,
p. 867-875, 2004.

120
IBRAHEEM, F.; GAFFOOR, I.; CHOPRA, S. Flavonoid phytoalexin-dependent resistance to
anthracnose leaf blight requires a functional yellow seed1in Sorghum bicolor. Genetics, v.
184, p. 915-926, 2010.
ISAACSON, T.; DAMASCENO, C. M. B.; SARAVANAN, R. S.; HE, Y.; CATALÁ, C.;
SALADIÉ, M.; ROSE, J. K. Sample extraction techniques for enhanced proteomic analysis of
plant tissues. Nature Protocols, v. 1, p. 769-774, 2006.
ISHIBASHI, K.; MASUDA, K.; NAITO, S.; MESHI, T.; ISHIKAWA, M. An inhibitor of
viral RNA replication is encoded by a plant resistance gene. Proceedings of the National
Academy of Sciences, v. 104, p.13833-13838, 2007.
ISHIBASHI, K.; NAITO, S.; MESHI, T.; ISHIKAWA, M. An inhibitory interaction between
viral and cellular proteins underlies the resistance of tomato to non adapted tobamoviruses.
Proceedings of the National Academy of Sciences, v. 106, p. 8778-8783, 2009.
JAMET, E.; ALBENNE, C.; BOUDART, G.; IRSHAD, M.; CANUT, H.; PONT-LEZICA,
R. Recent advances in plant cell wall protomics. Proteomics, v. 8, p. 893-908, 2008.
JAMI, S. K.; CLARK, G. B.; TURLAPATI, S. A.; HANDLEY, C.; ROUX, S. J.; KIRTI, P.
B. Ectopic expression of an annexin from Brassica juncea confers tolerance to abiotic and
biotic stress treatments in transgenic tobacco. Plant Physiology and Biochemistry, v. 46, p.
1019-1030, 2008.
JANJUSEVIC, R.; ABRAMOVITCH, R. B.; MARTIN, G. B.; STEBBINS, C. E. A bacterial
inhibitor of host programmed cell death defenses is an E3 ubiquitin ligase. Science, v. 311, p.
222–2226, 2006.
JAQUINOD, M.; VILLIERS, F.; KIEFFER-JAQUINOD, S.; HUGOUVIEUX, V.; BRULEY,
C.; GARIN, J.; BOURGUIGNON, J. A proteomics dissection of Arabidopsis thaliana
vacuoles isolated from cell culture. Molecular & Cellular Proteomics, v. 6, p. 394-412, 2007.
JELENSKA, J.; YAO, N.; VINATZER, B. A.; WRIGHT, C. M.; BRODSKY, J.
L.; GREENBERG, J. T. A J domain virulence effector of Pseudomonas syringae
remodels host chloroplasts and suppresses defenses. Current Biology, v.17, p.499-508, 2007.
JELENSKA, J.; VAN, J. A. H.; GREENBERG, J. T. Pseudomonas syringae hijacks plant
stress chaperone machinery for virulence. Proceedings of the National Academy of Sciences,
v. 107 , p. 13177-1382, 2010.
JENSEN, M. K.; HAGEDORN, P. H.; TORRES-ZABALA, M.; GRANT, M. R.; RUNG, J.
H.; COLLINGE, D. B.; LYNGKJAER, M. F. Transcriptional regulation by an NAC (NAM–
ATAF1,2–CUC2) transcription factor attenuates ABA signalling for efficient basal defence
towards Blumeria graminis f. sp. hordei in Arabidopsis. The Plant Journal, v. 56, p. 867-880,
2008.
JONES, A. M.; THOMAS, V.; TRUMAN, B.; LILLEY, K.; MANSFIELD, J.; GRANT, M.
Specific changes in the Arabidopsis proteome in response to bacterial challenge:
differentiating basal and R-gene mediated resistance. Phytochemistry, v. 65, p. 1805–1816,
2004.

121
JONES, J. D.; DANGL, J. L. The plant immune system. Nature, v. 444, p. 323-329, 2006.
JUNG, H. W.; TSCHAPLINSKI, T. J.; WANG, L.; GLAZEBROOK, J.; GREENBERG, J. T.
Priming in systemic plant immunity. Science, v. 324, p. 89-91, 2009.
KANG, J.-H.; MCROBERTS, J.; SHI, F.; MORENO, J. E.; JONES, A. D.; HOWE, G. A.
The flavonoid biosynthetic enzyme chalcone isomerase modulates terpenoid production in
glandular trichomes of tomato. Plant Physiology, v. 164, p.1161-1174, 2014.
KAZAN, K.; MANNERS, J. M. Linking development to defense: auxin in plant-pathogen
interactions. Trends in Plant Science, v.14, p. 373-382, 2009.
KEEN, N.T. Gene-for-gene complementarity in plant-pathogen interactions. Annual Review
of Genetics,v. 24, p. 447-463, 1990.
KIM, S. Y.; JANG, H. H.; LEE, J. R.; SUNG, N. R.; LEE, H. B.; LEE, D. H.; PARK, D.-J.;
KANG, C. H.; CHUNG, W. S.; LIM, C. O.; YUN, D.-J.; KIM, W. Y.; LEE, K. O.; LEE, S.Y.
Oligomerization and chaperone activity of a plant 2-Cys peroxiredoxin in
response to oxidative stress. Plant Science, v. 177, p. 227-232, 2009.
KJELLSEN, T. D.; SHIRYAEVA, L.; SCHRÖDER, W. P.; STRIMBECK, G. R. Proteomics
of extreme freezing tolerance in Siberian spruce (Picea obovata). Journal of Proteomics, v.
73, p. 965-975, 2010.
KLOSE, J. Protein mapping by combined isoelectric focusing and electrophoresis in mouse
tissues. A novel approach to testing for induced point mutations in mammals. Humangenetik,
v. 26, p. 231-243, 1975.
KNECHT, K.; SEYFFARTH, M.; DESEL, C.; THURAU, T.; SHERAMETI, I.; LOU, B.;
OELMÜLLER, R.; CAI, D. Expression of bvglp-1 encoding a germin-like protein from sugar
beet in Arabidopsis thaliana leads to resistance against phytopathogenic fungi. Molecular
Plant-Microbe Interactions, v. 23, p. 446-457, 2010.
KOECK, M.; HARDHAM, A.R.; DODDS, P.N. The role of effectors of biotrophic and
hemibiotrophic fungi in infection. Cell Microbiology, v. 13, p.1849-1857, 2011.
KÖNIGSHOFER, H.; TROMBALLA, H. W.; LÖPPERT, H. G. Early events in
signaling high-temperature stress in tobacco BY2 cells involve alterations in membrane
fluidity and enhanced hydrogen peroxide production. Plant Cell Environment, v. 31, p.
1771-1780, 2008.
KORVES, T. M.; BERGELSON, J. A developmental response to pathogen infection in
Arabidopsis. Plant Physiology, v. 133, p. 339-347, 2003.
KOSHIBA, T. Cytosolic ascorbate peroxidase in seedlings and leaves of maize (Zea
mays). Plant and Cell Physiology, v. 34(5), p. 713-721, 1993.
KOVALCHUK, I. Multiple roles of radicles in plants in Reactive Oxygen Species and
Antioxidants in Higher Plants,S. Dutta Gupta, Ed., p. 31-44, CRC Press, New York, NY,
USA, 2010.

122
KUC, J. Translocated signals for plant immunization. Annals of the New York
Academy of Sciences, v. 494, p. 221–223, 1987.
KUNKEL, B.N.; BROOKS, D. M. Cross talk between signaling pathways in pathogen
defence. Current Opinion in Plant Biology, v. 5, p. 325-331, 2002.
KUNZE, G.; ZIPFEL, C.; ROBATZEK S.; NIEHAUS. K; BOLLER, T.; FELIX, G. The N
terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant
Cell, v.16, p.3496-3507, 2004.
KURSTEINER, O.; DUPUIS, I.; KUHLEMEIER, C. The pyruvate decarboxylase 1 gene of
Arabidopsisis required during anoxia but not other environmental stresses. Plant Physiology,
v. 132, p. 968-978, 2003.
KUZNIAK, E.; KAZMIERCZAK, A.; WIELANEK, M.; GLOWACKI R.; KORNAS, A.
Involvement of salicylic acid, glutathione and protein S -thiolation in plant cell death-
mediated defence response of Mesembryanthemum crystallinum against Botrytis cinerea.
Plant Physiology and Biochemistry, v. 63, p. 30-38, 2013.
LAI, Z; MENGISTE, T. Genetic and cellular mechanisms regulating plant responses to
necrotrophic pathogens. Current Opinion in Plant Biology, v. 16, p. 1-8, 2013.
LALUK, K.; MENGISTE, T. Necrotroph Attacks on Plants: Wanton Destruction or
Covert Extortion? The Arabidopsis Book, v. 10, p. 1-34, 2010.
LANCIEN, M.; ROBERTS, M. R. Regulation of Arabidopsis thaliana 14-3-3 gene expression
by r-aminobutyric acid. Plant Cell and Environment, v. 29, p.1430-1436, 2006.
LANE, B. G.; DUNWELL, J. M.; RAY, J. A.; SCHMITT, M. R.; CUMING, A. C. Germin, a
protein marker of early plant development, is an oxalate oxidase. Journal of Biological
Chemistry, v. 268, p. 12239-12242, 1993.
LARCHER, W. Physiological plant ecology. 4° edition. Berlin, Heidelberg: Springer Verlag;
2003.
LARKINDALE, J.; VIERLING, E. Core genome responses involved in acclimation to high
temperature. Plant Physiology, v. 146, p. 748-761, 2008.
LAZNIEWSKA, J.; MACIOSZEK, V. K.; KONONOWICZ, A. K. Plant-fungus interface:
The role of surface structures in plant resistance and susceptibility to pathogenic fungi.
Physiological and Molecular Plant Pathology, v. 78, p. 24-30, 2012.
LE SOURD, F.; BOULBEN, S.; LE BOUFFANT, R.; CORMIER, P.; MORALES, J.;
BELLE, R.; MULNER-LORILLON, O. eEF1B: at the dawn of the 21st century. Biochimica
et Biophysica Acta- Gene Structure and Expression, v. 1759, p.13-31, 2006.
LEE, S.; LEE, E. J.; YANG, E. J.; LEE, J. E.; PARK, A. R.; SONG, W. H.; PARK, O. K.
Proteomic identification of annexins, calcium-dependent membrane binding proteins that
mediate osmotic stress and abscisic acid signal transduction in Arabidopsis. The Plant Cell, v.
16, p. 1378-1391, 2004.

123
LEVESQUE-TREMBLAY, G.; HAVAUX, M.; OUELLET, F. The chloroplastic lipocalin
AtCHL prevents lipid peroxidation and protects Arabidopsis against oxidative stress.
The Plant Journal, v. 60, p. 691-702, 2009.
LIMA, V. P. M. S. A cultura do cajueiro no nordeste do Brasil. Fortaleza: Banco do
Nordeste do Brasil, 301-319, 1988.
LIMA, J. S.; CARDOSO, J. E.; MOREIRA, R. C.; ALVES, R. C.; SOUZA, E.; MELO, J. G.
M. Caracterização cultural de isolados de Lasiodiplodia theobromae e patogenicidade em
plantas de aceroleira. Revista Trópica -Ciências Agrárias e Biológicas, v. 6, p. 10-16, 2012.
LINDERMAYR, C.; SELL, S.; MULLER, B.; LEISTER, D.; DURNER, J. Redox
regulation of the NPR1-TGA1 system of Arabidopsis thaliana by nitric oxide. Plant
Cell, v. 22, p. 2894-2907, 2010.
LIU, G.; JI, Y.; BHUIYAN, N. H.; PILOT, G.; SELVARAJ, G.; ZOU, J.; WEI, Y. Amino
Acid Homeostasis Modulates Salicylic Acid-Associated Redox Status and Defense Responses
in Arabidopsis. The Plant Cell, v. 22, p. 3845–3863, 2010.
LIU, J.; COAKER, G. Nuclear Trafficking During Plant Innate Immunity. Molecular Plant,
v. 1, p. 411-422, 2008.
LLORENTE, F.; MUSKETT, P.; SÁNCHEZ-VALLET, A.; LÓPEZ, G.; RAMOS, B.;
SÁNCHEZ-RODRÍGUEZ, C.; JORDÁ, L.; PARKER, J.; MOLINA, A. Repression of the
auxin response pathway increases Arabidopsis susceptibility to necrotrophic fungi. Molecular
Plant, v.1, p. 496-509, 2008.
LODHA, T. D.; HEMBRAM, P.; TEP, N.; BASAK, J. Proteomics: a successful approach to
understand the molecular mechanism of plant-pathogen interaction. American Journal of
Plant Sciences, v. 4, p. 1212-1226, 2013.
LÓPEZ-GRESA, M. P.; MALTESE, F.; BELLÉS, J. M.; CONEJERO, V.; KIM, H. K.;
CHOI, Y. H.; VERPOORTE, R. Metabolic response of tomato leaves upon different plant-
pathogen interactions. Phytochemical Analysis, v. 21, p. 89-94, 2010.
LÓPEZ-MATAS, M. A.; FERRER, A.; LARRAMENDI, C. H.; HUERTAS, A. J.; PAGÁN,
J. A.; GARCÍA-ABUJETA, J. L.; BARTRA, J.; ANDREU, C.; LAVÍN, J. R.; CARNÉS, J.
Acidic ribosomal protein 60S: a new tomato allergen. Food Chemistry, v. 127, p. 638-640,
2011.
LORANG, J.; KIDARSA, T.; BRADFORD, C. S.; GILBERT, B.; CURTIS, M.; TZENG, S.
C.; MAIER, C. S.; WOLPERT, T. J. Tricking the guard: exploiting plant defense for
disease susceptibility. Science, v. 338, p. 659-662, 2012.
LOU, Y.; BALDWIN, I. T. Silencing of a germin-like gene in nicotiana attenuata improves
performance of native herbivores. Plant Physiology, v. 140, p. 1126-1136, 2006.
LUDWIG, A. A.; SAITOH, H.; FELIX, G.; FREYMARK, G.; MIERSCH, O. et al. Ethylene-
mediated cross-talk between calcium-dependent protein kinase and MAPK signaling controls

124
stress responses in plants. Proceedings of the National Academy of Sciences, v. 102, p.
10736–10741, 2005.
LUO, Y.; CALDWELL, K. S.; WROBLEWSKI, T.; WRIGHT, M. E.; MICHELMORE, R.
W. Proteolysis of a negative regulator of innate immunity is dependent on resistance genes in
tomato and Nicotiana benthamiana and induced by multiple bacterial effectors. The Plant
Cell, v. 21, p. 2458-2472, 2009.
MADUREIRA, P. A.; WAISMAN, D. M. Annexin A2: The Importance of Being Redox
Sensitive. International. Journal of Molecular Sciences, v. 14, p. 3568-3594, 2013.
MAH, K. M.; UPPALAPATI, S. R.; TANG, Y.; ALLEN, S.; SHUAI, B. Gene expression
profiling of Macrophomina phaseolina infected Medicago truncatula roots reveals a role for
auxin in plant tolerance against the charcoal rot pathogen. Physiological and Molecular Plant
Pathology, v. 79, p. 21-30, 2012.
MAIMBO, M.; OHNISHI, K.; HIKICHI, Y.; YOSHIOKA, H., KIBA, A. Induction of a
small heat shock protein and its functional roles in Nicotiana plants in the defense
response against Ralstonia solanacearum. Plant Physiology, v. 145, p. 1588-1599, 2007.
MALDONADO, A. M.; ECHEVARRÍA-ZOMEÑO, S.; JEAN-BAPTISTE, S.;
HERNÁNDEZ, M.; JORRÍN-NOVO, J. V. Evaluation of three different protocols of protein
extraction for Arabidopsis thaliana leaf proteome analysis by two-dimensional
electrophoresis. Journal of Proteomics, v. 71, p. 461-472, 2008.
MANDELC, S.; TIMPERMAN, I.; RADISEK, S.; DEVREESE, B.; SAMYN, B.;
JAVORNIK, B. Comparative proteomic profiling in compatible and incompatible interactions
between hop roots and Verticillium albo-atrum. Plant Physiology and Biochemistry, v. 68, p.
23-31, 2013.
MANOSALVA, P.; DAVIDSON, R.; LIU, B.; ZHU, X.; HULBERT, S.; LEUNG, H.;
LEACH, J. A germin-like protein gene family functions as a complex qtl conferring broad-
spectrum disease resistance in rice. Plant Physiology, v. 149, p. 286-296, 2008.
MANSFIELD, J.W. From bacterial avirulence genes to effector functions via the hrp
delivery system: An overview of 25 years of progress in our understanding of plant
innate immunity. Molecular Plant Pathology, v.10, p. 721-734, 2009.
MARTINS, M. V. V.; CARDOSO, J. E.; CAVALCANTI, J. J. V.; NETO, F. C. V.; MELO,
J. G. M.; LIMA, J. S. Caracterização de progênie híbrida de cajueiro-anão precoce quanto à
infecção por Lasiodiplodia theobromae. Fortaleza: Embrapa Agroindústria Tropical (Boletim
de Pesquisa e Desenvolvimento, 41), 2011, 19p.
MAZZETTO, S. E.; LOMONACO, D.; MELE, G. Óleo da castanha de caju:
oportunidades e desafios no contexto do desenvolvimento e sustentabilidade industrial.
Química Nova, v. 32,, p. 1-10, 2009.
MCCLELLAN, A. J.; TAM, S.; KAGANOVICH, D.; FRYDMAN, J. Protein quality control:
chaperones culling corrupt conformations. Nature Cell Biology, v. 7, p. 736-741, 2005.

125
MEHTA, A.; BRASILEIRO, A.C.M.; SOUZA, D.S.L.; ROMANO, E.; CAMPOS, M.A.;
GROSSI-DE-SÁ, M.F.; SILVA, M.S.; FRANCO, O.L.; FRAGOSO, R.R.; BEVITORI, R.;
ROCHA, T. L. Plant–pathogen interactions: what is proteomics telling us? FEBS Journal, v.
275, p. 3731-3746, 2008.
MELO, Q. M. S. (Ed.) Caju: Fitossanidade. Brasília: Embrapa Informação Tecnológica, 62p.
(Frutas do Brasil, 26), 2002.
MELO, J. G. M. Diversidade genética e patogênica de Lasiodiplodia theobromae associado
ao cajueiro. 2010. 60p. Dissertação (Mestrado em Agronomia). Departamento de Ciências
Agrárias, Universidade Federal do Ceará, Fortaleza – CE.
MELVIN, P.; PRABHU, S. A.; ANUP, C. P.; SHAILASREE, S.; SHETTY, H. S.; KINI, K.
R. Involvement of mitogen-activated protein kinase signalling in pearl millet-downy
mildew interaction. Plant Science, v. 214, p. 29-37, 2014.
MENGISTE, T. Plant Immunity to Necrotrophs. Annual Review of Phytopathology, v. 50, p.
267-94, 2012.
MENG, X.; ZHANG, S. MAPK cascades in plant disease resistance signaling. Annual Review
of Phytopathology, v. 51, p. 245-266, 2013.
MENKE, F. L. H.; VAN PELT, J. A.; PIETERSE, C. M. J.; KLESSIG, D. F. Silencing of the
mitogen-activated protein kinase mpk6 compromises disease resistance in Arabidopsis. The
Plant Cell, v. 16, p. 897-907, 2004.
MEYER, Y.; BUCHANAN, B. B.; VIGNOLS, F.; REICHHELD, J.-P. Thioredoxins and
Glutaredoxins: unifying elements in redox biology. Annual Review of Genetics, v. 43, p. 335-
367, 2009.
MILLER, G.; SCHLAUCH, K.; TAM, R.; CORTES, D.; TORRES, M. A.; SHULAEV, V.;
DANGL, J. L.; MITTLER, R. The plant NADPH oxidase RBOHD mediates rapid systemic
signaling in response to diverse stimuli. Science Signaling, v. 2, p. ra45, 2009.
MILLI, A.; CECCONI, D.; BORTESI, L.; PERSI, A.; RINALDUCCI, S.; ZAMBONI, A.;
ZOCCATELLI, G.; LOVATO, A.; ZOLLA, L.; POLVERARI, A. Proteomic analysis of the
compatible interaction betweenVitis vinifera and Plasmopara viticola. Journal of Proteomics,
v. 75, p. 1284-1302, 2012.
MITTLER, R.; VANDERAUWERA, S.; SUZUKI, N.; MILLER, G.; TOGNETTI, V.;
VANDEPOELE, K.; GOLLERY, M.; SHULAEV, V.; BREUSEGEM, F. V. ROS signaling:
the new wawe? Trends in Plant Science, v.16, p. 300-309, 2011.
MOCHIDA, K.; SHINOZAKI, K. Genomics and bioinformatics resources for crop
improvement. Plant Cell Physiology, v. 51, p. 497-523, 2010.
MOHALI, S.; BURGESS, T. I.; WINGFIELD, M. J. Diversity and host association of the
tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence
repeat markers. Forest Pathology, v. 35, p. 385-396, 2005.

126
MOHR, P. G.; CAHILL, D. M. Abscisic acid influences the susceptibility of Arabidopsis
thaliana to Pseudomonas syringae pv. tomato and Peronospora parasitica. Functional Plant
Biology, v. 30, p. 461-469, 2003.
MOREIRA, R. C.; LIMA, J. S.; SILVA, L. G. C.; CARDOSO, J. E. Resistance to gummosis
in wild cashew genotypes in northern Brazil. Crop Protection, v. 52, p. 10-13, 2013.
MORI, T.; SAKURAI, M.; SAKUTA, M. Effects of conditioned medium on activities of
PAL, CHS, DAHP syntase (DS-Co and Ds-Mn) and anthocyanin production in
suspension cultures of Fragaria ananassa. Plant Science, v. 160, p. 355-360, 2001.
MORTIMER, J. C.; LAOHAVISIT, A.; MACPHERSON, N.; WEBB, A.; BROWNLEE, C.;
BATTEY, N. H.; DAVIES, J. M. Annexins: multifunctional components of growth and
adaptation. Journal of Experimental Botany, v. 59, p. 533-544, 2008.
MOU, Z.; FAN, W.; DONG, X. Inducers of plant systemic acquired resistance regulate
NPR1 function through redox changes. Cell, v. 113, p. 935–944, 2003.
MUKHERJEE, A. K.; CARP, M.-J.; ZUCHMAN, R.; ZIV, T.; HORWITZ, B. A.;
GEPSTEIN, S. Proteomics of the response of Arabidopsis thaliana to infection with
Alternaria brassicicola. Journal of proteomics, v. 73, p. 709-720, 2010.
MOURA, H. F. N.; VASCONCELOS, I. M.; SOUZA, C. E. A.; SILVA, F. D. A.; MORENO,
F. B. M. B.; LOBO, M. D. P.; MONTEIRO-MOREIRA, A. C. O.; MOURA, A. A.; COSTA,
J. H. Proteomics changes during the incompatible interaction between cowpea and
Colletotrichum gloeosporioides (Penz.) Penz and Sacc. Plant Science, v. 217-218, p. 158-
175, 2014.
MUKHTAR, M. S.; NISHIMURA, M. T.; DANGL, J. NPR1 in Plant Defense: It‟s not over
‟til It‟s Turned over. Cell, v. 137, p. 804-806, 2009.
MULTHOFF , G. Heat shock proteins in immunity. Hand b Exper Pharmacology, v. 172, p.
279-304, 2006.
MUNIZ, C. R.; FREIRE, F. C. O.; VIANA, F. M. P.; CARDOSO, J. E.; COOKE , P.;
WOOD, D.; GUEDES, M. I. F. Colonization of cashew plants by Lasiodiplodia theobromae:
Microscopical features. Micron, v. 42, p. 419-428, 2011.
MUR, L. A.; KENTON, P.; LLOYD, A. J; OUGHAM, H.; PRATS, E. The hypersensitive
response: the centenary is upon us but how much do we know? Journal of Experimental
Botany, v. 59, p. 501–520, 2008.
NAVARRO, L.; DUNOYER, P.; JAY, F.; ARNOLD, B.; DHARMASIRI, N.; ESTELLE,
M.; VOINNET, O.; JONES, J. D. A plant miRNA contributes to antibacterial resistance by
repressing auxin signaling. Science, v. 312, p. 436-439, 2006.
NAVROT, N.; FINNIE, C.; SVENSSON, B.; HÄGGLUND, P. Plant redox proteomics.
Journal of Proteomics, v. 74, p. 1450-1462, 2011.
NEILSON E. H., GOODGER J. Q. D., WOODROW I. E., MOLLER, B. L. Plant

127
Chemical defense: at what cost? Trends in Plant Science, v. 18, p. 250-258, 2013.
NETO, V. F. C.; BARROS, L. M.; CAVALCANTI, J. J. V.; MELO, D. S.Melhoramento
genético e cultivares de cajueiro. In: ARAÚJO, J. P. P. de (Ed.). Agronegócio caju: práticas
e inovações. Brasília, DF: Embrapa, 2013. parte 7, cap. 2, p. 481-508.
NEWMAN, M. A.; SUNDELIN, T.; NIELSEN, J. T.; ERBS, G. MAMP (microbe-associated
molecular pattern) triggered immunity in plants. Frontiers in Plant Science, v. 4, p. 1-14,
2013.
NEWTON, A. C.; FITT, B. D. L.; ATKINS, S. D.; WALTERS, D. R.; DANIELL, T. J.
Pathogenesis, parasitism and mutualism in the trophic space of microbe-plant interactions.
Trends in Microbiology, v. 18, p. 365-373, 2010.
NISHIMURA, M. T.; DANGL, J. L. Arabidopsis and the plant immune system. Plant
Journal, v. 61, p. 053-1066, 2010.
NOËL, L. D. et al. Interaction between SGT1 and cytosolic/nuclear HSC70 chaperones
regulates Arabidopsis immune responses. Plant Cell, v. 19, p. 4061-4076, 2007.
NÜRNBERGER, T.; SCHEEL, D. Signal transmission in the plant immune response. Trends
in Plant Science, v. 6, p. 372-379, 2001.
OECKING, C.; JASPERT, N. Plant 14-3-3 proteins catch up with their mammalian
orthologs. Current Opinion in Plant Biology, v. 12, 760-765, 2009.
O'FARRELL, P. H. High resolution two-dimensional electrophoresis of proteins. Journal of
Biological Chemistry, v. 250, p. 4007-4021, 1975.
OH, C. S. Characteristics of 14-3-3 proteins and their role in plant immunity. The Plant
Pathology Journal, v. 26, p. 1-7, 2010.
OH, C. S.; PEDLEY, K. F.; MARTIN, G. B. Tomato 14-3-3 protein 7 positively regulates
immunity-associated programmed cell death by enhancing protein abundance and signaling
ability of MAPKKKα. Plant Cell, v. 22, p. 260-272, 2010.
OLIVEIRA, V. H. (Ed.) Cultivo do cajueiro anão precoce. Fortaleza: Embrapa Agroindústria
Tropical (Sistemas de Produção, 1), 2002.
OLIVEIRA, V. H. Cajucultura. Revista Brasileira de Fruticultura, v. 30, p. 1-3, 2008.
OLIVEIRA J. T. A.; ANDRADE N. C.; MARTINS-MIRANDA A. S.; SOARES, A. A.;
GONDIM D. M. F.; ARAUJO-FILHO J. H.; FREIRE-FILHO F. R.; VASCONCELOS I. M.
Differential expression of antioxidant enzymes and PR-proteins in compatible and
incompatible interactions of cowpea (Vigna unguiculata) and the root-knot nematode
Meloidogyne incognita. Plant Physiology and Biochemistry, v. 51, p. 145-152, 2012.
OLIVER, R. P.; SOLOMON, P. S. New developments in pathogenicity and virulence of
necrotrophs. Current Opinion in Plant Biology, v. 13, p. 415-419, 2010.

128
OSAKABE, Y.; YAMAGUCHI-SHINOZAKI, K.; SHINOZAKI, K.; TRAN, L.-S. P. ABA
control of plant macroelement membrane transport systems in response to water deficit and
high salinity. New Phytologist, v. 202, p. 35-49, 2013.
PAIVA, J. R.; CARDOSO, J. E.; BARROS, L. M.; CRSÓSTOMO, J. R.; CAVALCANTI, J.
J. V.; ALENCAR, E. S. Clone de cajueiro-anão-precoce BRS 226 ou Planalto: nova
alternativa para o plantio na região semi-árida do nordeste. Fortaleza: Embrapa
Agroindústria Tropical, 4p. (Comunicado Técnico, 78), 2002.
PAIVA, J.R., CRISÓSTOMO, J.R., BARROS, L.M. Recursos genéticos do cajueiro:
coleta, conservação, caracterização e utilização. Fortaleza: Embrapa Agroindústria
Tropical, 43p. (Documentos, 65), 2003.
PAIVA, J. R.; BARROS, L. M. Clones de Cajueiro: Obtenção, Características e
Perspectivas. Embrapa Agroindústria Tropical (Documentos, 82), 2004.
PAIVA, J. R.; CARDOSO, J. E.; MESQUITA, A. L. M.; CAVALCANTI, J. J. V.; SANTOS,
A. A. Desempenho de clones de cajueiro-anão precoce no semiárido do Estado do Piauí.
Revista Ciência Agronômica, v. 39, p. 295-300, 2008.
PALLAS, V.; GARCÍA, J. A. How do plant viruses induce disease? Interactions and
interference with host components. Journal of General Virology, v. 92, p. 2691–2705, 2011.
PALOMARES-RIUS, J. E.; CASTILLO, P.; NAVAS-CORTÉS, J. A.; JIMÉNEZ-DÍAZ, R.
M.; TENA, M. A proteomic study of in-root interactions between chickpea pathogens: The
root-knot nematode Meloidogyne artiellia and the soil-borne fungus Fusarium oxysporum f.
sp.ciceris race 5. Journal of Proteomics, v. 74, p. 2034-2051, 2011.
PAN, X.; WELTI, R.; WANG, X. Simultaneous quantification of major phytohormones and
related compounds in crude plant extracts by liquid chromatography-electrospray
tandem mass spectrometry. Phytochemistry, v. 69, p. 1773-1781, 2008.
PANDEY, A.; CHAKRABORTY, S.; DATTA, A.; CHAKRABORTY, N. Proteomics
approach toidentify dehydration responsive nuclear proteins from chickpea (Cicer arietinum
L.). Molecular & Cellular Proteomics, v. 7, p. 88-107, 2008.
PAGEAU, K.; REISDORF-CREN, M.; MOROT-GAUDRY, J. F.; MASCLAUX-
DAUBRESSE, C.The two senescence-related markers, GS1and GDH, involved in nitrogen
mobilization, are differentially regulated during pathogen attack and by stress hormones and
reactive oxygen species in Nicotiana tabacum L. leaves. Journal of Experimental Botany, v.
57, p. 547-557, 2006.
PARK, C.-J.; KIM, K.-J.; SHIN, R.; PARK, J. M.; SHIN, Y.-C.; PAEK, K.-H. Pathogenesis-
related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral
pathway. The Plant Journal, v. 37, p. 186-198, 2003.
PARK, S.W.; KAIMOYO, E.; KUMAR, D.; MOSHER, S.; KLESSIG, D. F. Methyl
salicylate is a critical mobile signal for plant systemic acquired resistance. Science, v. 318,
p. 113–116, 2007.

129
PATHURI, I. P.; REITBERGER, I. E.; HUCKELHOVEN, R.; PROELS, R. K. Alcohol
dehydrogenase 1 of barley modulates susceptibility to the parasitic fungus Blumeria graminis
f.sp.hordei. Journal of Experimental Botany, v. 62, p. 3449-3457, 2011.
PAUL, A. L.; LIU, L.; LAUGHNER, B.; MCCLUNG, S.; CHEN, S.; FERL, R. Comparative
interactomics: analysis of Arabidopsis 14-3-3 complexes reveals highly conserved 14-3-
3 interactions between humans and plants. Journal of Proteome Research, v. 8, p. 1913-
1924, 2009.
PAVOKOVIĆ, D.; KRIŽNIK, B.; KRSNIK-RASOL, M. Evaluation of protein extraction
methods for proteomic analysis of non-model recalcitrant plant tissues. Croatica Chemica
Acta, v. 85, p. 177–183, 2012.
PEREIRA, A. L.; SILVA, G. S.; RIBEIRO, V. Q. Caracterização fisiológica, cultural e
patogênica de diferentes isolados de Lasiodiplodia theobromae. Fitopatologia Brasileira, v.
31, p. 572-578, 2006.
PEIXOTO, P. H. P.; CAMBRAIA, J.; SANT‟ANNA, R.; MOSQUIM, P. R.; MOREIRA, M.
A. Aluminum effects on lipid peroxidation and on the activities of enzymes of oxidative
metabolism in sorghum. Revista Brasileira de Fisiologia Vegetal, v. 11, p. 137-143, 1999.
PEREZ-GARCIA, A.; CANOVAS, F. M.; GALLARDO, F.; HIREL, B.; DEVICENTE, A.
Differential expression of glutamine-synthetase isoforms in tomato detached leaflets infected
with Pseudomonas syringae pv tomato. Molecular Plant Microbe Interact, v.8, p.96-103,
1995.
PEREZ-GARCIA, A.; PEREIRA, S.; PISSARRA, J.; GUTIERREZ, A. G.; CAZORLA, F.
M.; SALEMA, R.; DE VICENTE, A.; CANOVAS, F. M. Cytosolic localization in tomato
mesophyll cells of a novel glutamine synthetase induced in response to bacterial infection or
phosphinothricin treatment. Planta, v. 206, p. 426-434, 1998.
PETRICCIONE, M.; CECCO, I. D.; ARENA, S.; SCALONI, A.; SCORTICHINI, M.
Proteomic changes in Actinidia chinensis shoot during systemic infection with a pandemic
Pseudomonas syringae pv. actinidiae strain. Journal of Proteomics, v. 78, p. 461-476, 2013.
PIETERSE, C.M.; LEON-REYES, A.; VAN, D. E. S.; VAN, W. S. C. M. Networking by
small-molecule hormones in plant immunity. Nature Chemical Biology, v.5, p. 308-316,
2009.
PIQUES, M.; SCHULZE, W. X.; HOHNE, M.; USADEL, B.; GIBON, Y.; ROHWER, J.;
STITT, M. Ribosome and transcript copy numbers, polysome occupancy and enzyme
dynamics in Arabidopsis. Molecular Systems Biology, v. 5, p. 314, 2009.
PITERKOVÁ, J.; LUHOVÁ, L.; MIESLEROVÁB, B.; LEBEDA, B.; PETRIVALSK, M.
Nitric oxide and reactive oxygen species regulate the accumulation of heat shock
proteins in tomato leaves in response to heat shock and pathogen infection. Plant
Science, v. 207, p. 57-65, 2013.

130
POTTERS, G.; PASTERNAK, T. P.; GUISEZ, Y.; PALME, K. J.; JANSEN, M. A. K. Stress-
induced morphogenic responses: growing out of trouble? Trends in Plant Science, v. 12, p.
98-105, 2007.
PUGIN, A.; FRACHISSE, J. M.; TAVERNIER, E.; BLIGNY, R.; GOUT, E.; DOUCE, R.;
GUERN, J. Early events induced by the elicitor cryptogein in tobacco cells: Involvement of a
plasma membrane NADPH oxidase and activation of glycolysis and the pentose phosphate
pathway. Plant Cell, v. 9, p. 2077-2091, 1997.
QUIRINO, B. F.; CANDIDO, E. S.; CAMPOS, P. F.; FRANCO, O. L.; KRÜGER, R. H.
Proteomic approaches to study plant pathogen interactions. Phytochemstry, v. 71, p. 351-362,
2010.
RAFIQI, M.; BERNOUX, M.; ELLIS, J.G.; DODDS, P. N. In the trenches of plant pathogen
recognition: Role of NB-LRR proteins. Seminars in Cell & Developmental Biology, v. 20, p.
1017–1024, 2009.
RAMPUTH, A.; ARNASON, J.; CASS, L.; SIMMONDS, J. Reduced herbivory of the
european corn borer (Ostrinia nubilalis) on corn transformed with germin, a wheat oxalate
oxidase gene. Plant Science, v. 162, p. 431-440, 2002.
RANF, S.; WUNNENBERG, P.; LEE, J.; BECKER, D.; DUNKEL, M. et al. Loss of the
vacuolar cation channel, AtTPC1, does not impair Ca2+
signals induced by abiotic and biotic
stresses. Plant Journal, v. 53, p.287–299, 2008.
RENSINK, W. A.; BUELL, C. R. Microarray expression profiling resources for plant
genomics. Trends in Plant Science, v. 10, p. 603-609, 2005.
REUMANN, S.; BABUJEE, L.; MA, C.; WIENKOOP, S.; SIEMSEN, T.; ANTONICELLI,
G. E.; RASCHE, N.; LUDER, F.; WECKWERTH, W.; JAHN, O. Proteome analysis of
Arabidopsis leaf peroxisomes reveals novel targeting peptides, metabolic pathways, and
defense mechanisms. Plant Cell, v. 19, p. 3170-3193, 2007.
ROBERT-SEILANIANTZ, A.; NAVARRO, L.; BARI, R.; JONES, J. D. Pathological
hormone imbalances. Current Opinion in Plant Biology, v. 10, p. 372–379, 2007.
ROBERTS, T. H.; HEJGAARD, J. Serpins in plants and green algae. Functional and
Integrative Genomics, v. 8, p. 1-27, 2008.
ROCHA, T. L.; COSTA, P. H. A.; MAGALHÃES, J. C. C.; EVARISTO, R. G. S.;
VASCONCELOS, E. A. R.; COUTINHO, M. V.; PAES, N. S.; SILVA, M. C. M.; GROSSI-
DE-SÁ, M. F. Eletroforese bidimensional e análise de proteomas. Brasília: Embrapa
Recursos Genéticos e Biotecnologia, 12p. (Comunicado Técnico, 136), 2005.
RODNINA, M. V.; WINTERMEYER, W. Recent mechanistic insights into eukaryotic
ribosomes. Current Opinion in Cell Biology, v. 21, p. 435-443, 2009.
RODRIGUEZ, M. R. S.; PETERSEN, M.; MUNDY, J. Mitogen-activated protein
kinase signaling in plants. Annual Review Plant Biology, v. 61, p. 621-649, 2010.

131
ROGOWSKA-WRZESINSKA, A.; LE BIHAN, M. C.; THAYSEN-ANDERSEN, M.;
ROEPSTORFF, P. 2D gels still have a niche in proteomics. Journal of Proteomics, v.
88, p. 4-13, 2013.
ROMEIS, T. Protein kinases in the plant defence response. Current Opinion in Plant Biology,
v. 4, p. 407-414, 2001.
ROSEBROCK, T. R.; ZENG, L. R.; BRADY, J. J.; ABRAMOVITCH, R. B.; XIAO, F. M.;
MARTIN, G. B. A bacterial E3 ubiquitin ligase targets a host protein kinase to disrupt plant
immunity. Nature, v. 448, p. 370-374, 2007.
ROSSETTI, A. G.; AQUINO, A. R. L. Influência do tipo de ramo sobre o crescimento e
produção do cajueiro anão precoce de copa substituída. Revista Brasileira de Fruticultura, v.
24, 2002.
ROUHIER, N.; KOH, C. S.; GELHAYE, E.; CORBIER, C.; FAVIER, F.; DIDIERJEAN, C.;
JACQUOT, J.-P. Redox based anti-oxidant systems in plants: biochemical and structural
analyses. Biochimica et Biophysica Acta, v.1780, p. 1249-1260, 2008.
SACCO, M.; MANSOOR, S.; MOFFETT, P. A RanGAP protein physically interacts with the
NB-LRR protein Rx, and is required for Rx-mediated viral resistance. The Plant Journal, v.
52, p. 82-93, 2007.
SADE, D.; EYBISHTZ, A.; GOROVITS, R.; SOBOL, I.; CZOSNEK, H. A developmentally
regulated lipocalin-like gene is overexpressed in Tomato yellow leaf curl virus-resistant
tomato plants upon virus inoculation, and its silencing abolishes resistance. Plant Molecular
Biology, v. 80, p. 273-287, 2012.
SAIDI, Y.; FINKA, A.; MURISET, M.; BROMBERG, Z.; WEISS, Y. G.; MAATHUIS, F. J.
M.; GOLOUBINOFF, P. The heat shock response in moss plants is regulated by specific
calcium-permeable channels in the plasma membrane. The Plant Cell, v. 21, p. 2829-2843,
2009.
SANTOS, R. P.; SANTIAGO, A. A. X.; GADELHA, C. A. A.; CAJAZEIRAS, J. B.;
CAVADA, B. S.; MARTINS, J. L.; OLIVEIRA, T. M.; BEZERRA, G. A.; SANTOS, R. P.;
FREIRE, V. N. Production and characterization of the cashew (Anacardium occidentale L.)
peduncle bagasse ashes. Journal of Food Engineering, v. 79, p. 1432-1437, 2007.
SARAVANAN, R. S.; ROSE, J. K. C. A critical evaluation of sample extraction techniques
for enhanced proteomic analysis of recalcitrant plant tissues. Proteomics, v. 4, p. 2522-2532,
2004.
SCHENK, P.M.; KAZAN, K. et al. Coordinated plant defence responses in Arabidopsis
revealed by microarray analysis. Proceedings of the National Academy of Sciences, v. 97, p.
11655-11660, 2000.
SCHMIDT, S. M.; PANSTRUGA, R. Cytoskeletal functions in plant-microbe interactions.
Physiological and Molecular Plant Pathology, v. 71, p. 135-148, 2007.
SCHOFFL, F.; PRANDL, R.; REINDL, A. Regulation of the heat-shock response. Plant
Physiology, v.117, p. 1135-1141, 1998.

132
SCHOONHEIM, P. J.; VEIGA, H.; PEREIRA, D. C; FRISO, G.; VAN WIJK, K. J.; DE
BOER, A. H. A comprehensive analysis of the 14-3-3 interactome in barley leaves using
a complementary proteomics and two-hybrid approach. Plant Physiology, v.143, p. 670-
683, 2007.
SCHORNACK, S.; HUITEMA, E.; CANO, L. M.; BOZKURT, T. O.; OLIVA, R.; VAN
DAMME, M.; SCHWIZER, S.; RAFFAELE, S.; CHAPARRO-GARCIA, A.; FARRER, R.;
SEGRETIN, M. E.; BOS, J.; HAAS, B. J.; ZODY, M. C.; NUSBAUM, C.; WIN, J.;
THINES, M.; KAMOUN, S. Ten things to know about oomycete effectors. Molecular Plant
Pathology, v. 10, p. 795-803, 2009.
SCHROEDER, J. I.; KWAK, J. M.; ALLEN, G. J. Guard cell abscisic acid signaling and
engineering drought hardiness in plants. Nature, v. 410, p. 317-330, 2001.
SEHNKE, P. C.; DELILLE, J. M.; FERL, R. J. Consummating signal transduction: the role of
14-3-3 proteins in the completion of signal-induced transitions in protein activity. Plant Cell,
v. 14, p. S339–S354, 2002.
SELS, J.; MATHYS, J.; CONINCK, B. M.A.; CAMMUE, B. P.A.; BOLLE, M. F.C. Plant
pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiology and
Biochemistry, v. 46, p. 941–950, 2008.
SHAH, J.; ZEIER, J. Long-distance communication and signal amplification in
systemic acquired resistance. Frontiers in Plant Science, v. 4, p. 1-16, 2013.
SHEN, Q. H.; SAIJO, Y.; MAUCH, S.; BISKUP, C.; BIERI, S.; KELLER, B.; SEKI, H.;
ULKER, B.; SOMSSICH, I. E.; SCHULZE-LEFERT, P. Nuclear activity of MLA immune
receptors links isolate-specific and basal disease-resistance responses. Science, v. 315, p.
1098-1103, 2007.
SHETTY, N. P.; JØRGENSEN, H. J. L.; JENSEN, J. D., COLLINGE, D. B.; SHETTY, H. S.
Roles of reactive oxygen species in interactions between plants and pathogens, European
Journal Plant Pathology, v. 121, p. 267-280, 2008.
SINHA, R.; CHATTOPADHYAY, S. Changes in the leaf proteome profile of Mentha
arvensis in response to Alternaria alternate infection. Journal of Proteomics, v. 74, p. 327-
336, 2011.
SLIPPERS, B.; WINGFIELD, M. J. Botryosphaeriaceae as endophytes and latent
pathogens of woody plants: diversity, ecology and impact. Fungal Biology Reviews, v. 21, p.
90-106, 2007.
SMITH R. Two-Dimensional Electrophoresis: An Overview. In: TYTHER, R.; SHEEHAN,
D. Two-Dimensional Electrophoresis Protocols. Humana Press; 2009. p. 2-17.
SONG, J. T.; LU, H.; MCDOWELL, J. M.; GREENBERG, J. T. A key role for ALD1 in
activation of local and systemic defenses in Arabidopsis. The Plant Journal, v. 40, p. 200-
212, 2004.

133
SOUSA. L.B., FEITOZA, L.L., GOMES, R.L.F., LOPES, A.C.A., SOARES, E.B., SILVA,
E.M.P. Aspectos de biologia floral de cajueiros anão precoce e comum. Ciência Rural, v. 37,
p. 882-885, 2007.
SPOEL, S. H.; MOU, Z.; TADA, Y. ; SPIVEY, N. W.; GENSCHIK, P.; DONG, X.
Proteasome-Mediated Turnover of the Transcription Co-Activator NPR1 Plays Dual Roles in
Regulating Plant Immunity. Cell, v. 137, p. 860–872, 2009.
SPOEL, S. H.; DONG, X. How do plants achieve immunity? Defence without specialized
immune cells. Nature Review Immunology, v.12, p. 89-100, 2012.
SUN, C.; WANG, L.; HU, D.; RIQUICHO, A. R. M.; LIU, T.; HOU, X.; LI, Y. Proteomic
analysis of non-heading Chinese cabbage infected with Hyaloper-onospora parasitica.
Journal of Proteomics, v. 98, p. 15-30, 2014.
SUZUKI, N.; KOUSSEVITZKY, S.; MITTLER, R.; MILLER, G. ROS and redox signalling
in the response of plants to abiotic stress. Plant, Cell and Environment, v. 35, p. 259-270,
2012.
SWARBRICK, P.L.; SCHUZE-LEFERT, P.; SCHOLES, J. D. Metabolic consequences of
susceptibility and resistance (race specific and broad-spectrum) in barley leaves challenged
with powdery mildew. Plant Cell Environment, v. 29, p. 1061-1076, 2006.
TADA, Y.; SPOEL, S. H.; PAJEROWSKA-MUKHTAR, K.; MOU, Z.; SONG, J.;
WANG, C.; ZUO, J.; DONG, X. Plant immunity requires conformational changes
[corrected] of NPR1 via S-nitrosylation and thioredoxins. Science, v. 321, p. 952-956,
2008.
TALER, D.; GALPERIN, M.; BENJAMIN, I.; COHEN, Y.; KENIGSBUCH, D. Plant eR
genes that encode photorespiratory enzymes confer resistance against disease. Plant Cell, v.
16, p.172-184, 2004.
TAMELING, W. I.; BAULCOMBE, D. C. Physical association of the NB-LRR resistance
protein Rx with a Ran GTPase-activating protein is required for extreme resistance to Potato
virus X. The Plant Cell, v. 19, p. 1682-1694, 2007.
TAVARES, S.C.C.H. Epidemiologia e manejo integrado de Botryodiplodia theobromae-
situação atual no Brasil e no mundo. Fitopatologia Brasileira, v. 27, p. 46-52. 2002.
TCHÓRZEWSKI, M. The acidic ribosomal P proteins. The International Journal of
Biochemistry & Cell Biology , v. 34, p. 911-915, 2002.
THAKKAR, V. R.; SUBRAMANIAN, R. B.; KOTHARI, I. L. Culture filtrate
of Lasiodiplodia theobromae restricts the development of natural resistance in Brassica nigra
plants. Indian Journal of Experimental Biology, v. 42, p. 111-114, 2004.
TIMPERIO, A. M.; EGIDI, M. G.; ZOLLA, L. Proteomics applied on plant abiotic stresses:
Role of heat shock proteins (HSP). Journal of Proteomics, v. 71, p. 391-411, 2008.

134
TOHGE, T.; WATANABE, M.; HOEFGEN, R.; FERNIE, A. R. The evolution of
phenylpropanoid metabolism in the green lineage. Critical Review in Biochemistry and
Molecular Biology, v. 48, p. 123-152, 2013.
TON, J.; MAUCH-MANI, B. b-Amino-butyric acid-induced resistance against necrotrophic
pathogens is based on ABA-dependent priming for callose. The Plant Journal, v. 38, p. 119-
130, 2004.
TORRES, M. A.; DANGL, J. L. Functions of the respiratory burst oxidase in biotic
interactions, abiotic stress and development. Current Opinion in Plant Biology, v.8, p. 397-
403, 2005.
TORRES, M.; MANSFIELD, J. W.; GRABOV, N.; BROWN, I. R.; AMMOUNEH, H.;
TSIAMIS, G.; FORSYTH, A.; ROBATZEK, S.; GRANT, M.; BOCH, J. Pseudomonas
syringae effector AvrPtoB suppresses basal defence in Arabidopsis. The Plant Journal, v. 47,
p. 368-382, 2006.
TORRES, M.A. ROS in biotic interactions. Plant Physiology, v.138, p. 414–429, 2010.
TORRES-ZABALA, M.; TRUMAN , W.; BENNETT, M. H.; LAFFORGUE, G.;
MANSfiELD, J. W.; EGEA, P. R.; BOGRE, L.; GRANT, M. Pseudomonas syringae pv.
tomato hijacks the Arabidopsis abscisic acid signalling pathway to cause disease. The EMBO
Journal, v. 26, p.1434-1443, 2007.
TRUJILLO, M.; SHIRASU, K. Ubiquitination in plant immunity. Current Opinion in Cell
Biology, v. 13, p. 402-408, 2010.
TRUMAN, W.; BENNETT, M. H.; KUBIGSTELTIG, I.; TURNBULL, C.; GRANT, M.
Arabidopsis systemic immunity uses conserved defense signaling pathways and is mediated
by jasmonates. Proceedings of the National Academy of Sciences, v. 104, p. 1075–1080,
2007.
TSUDA, K.; SATO, M.; STODDARD, T.; GLAZEBROOK, J.; KATAGIRI, F. Network
properties of robust immunity in plants. Plos Genetics, v.5, p. 1000772-1000788, 2009.
TSUKADA, K.; TAKAHASHI, K.; NABETA, K. Biosynthesis of jasmonic acid in a plant
pathogenic fungus, Lasiodiplodia theobromae. Phytochemistry, v. 71, p. 2019–2023, 2010.
UKNES, S.; MAUCH-MANI, B.; MOYER, M.; POTTER, S.; WILLIAMS, S. et al. Acquired
resistance in Arabidopsis. Plant Cell, v. 4, p. 645–56, 1992.
URBANEK H.; KUZNIAK-GEBAROWSKA, E.; HERKA K. Elicitation of defense
responses in bean leaves by Botrytis cinerea polygalacturonase. Acta Physiologiae
Plantarum, v. 13, p. 43-50, 1991.
VALCU, C.-M.; JUNQUEIRA, M.; SHEVCHENKO, A.; SCHLINK, K. Comparative
proteomic analysis of responses to pathogen infection and wounding in Fagus sylvatica.
Journal of Proteomics research, v.8, p. 4077-4091, 2009.

135
VAN, D. B. E. A; JONES, J.D.G. Plant disease-resistance proteins and the gene- for-gene
concept. Trends in Biochemical Sciences, v. 12, p 454- 456, 1998.
VAN DAMME, M.; ZEILMAKER, T.; ELBERSE, J.; ANDEL, A.; DE SAIN-VAN DER
VELDEN, M.; VAN DEN ACKERVEKEN, G. Downy mildew resistance in Arabidopsis by
mutation of HOMOSERINE KINASE. Plant Cell, v. 21, p. 2179-2189, 2009.
VAN DER HOORNA, R. A. L.; KAMOUN, S. From Guard to Decoy: A New Model for
Perception of Plant Pathogen Effectors. The Plant Cell, v. 20, p. 2009-2017, 2008.
VAN LOON, L. C.; REP, M.; PIETERSE, C. M. J. Significance of inducible defense-related
proteins in infected plants. Annual Review of Phytopathology, v. 44, p. 135-162, 2006.
VAN ROSSUM, G.; JANSEN J.; MULLENDER K.S.; BULTERMAN, D. CMIFed: A
Presentation Environment for Portable Hypermedia Documents. ACM
Multimedia’93, Anaheim, EUA, Agosto de 1993.
VANDERSCHUREN, H.; LENTZ, E.; ZAINUDDIN, I.; GRUISSEM, W. Proteomics of
model and crop plant species: status, current limitations and strategic advances for crop
improvement. Journal of proteomics, v. 93, p. 5-19, 2013.
VARSHNEY, R. K.; NAYAK, S. N.; MAY, G. D.; JACKSON, S. A. Next-generation
sequencing technologies and their implications for crop genetics and breeding. Trends in
Biotechnology, v. 27, p. 522-530, 2009.
VERNOUD, V.; HORTON, A. C.; YANG, Z.; NIELSEN, E. Analysis of the small GTPase
gene superfamily of Arabidopsis. Plant Physiology, v. 131, p. 1191-1208, 2003.
VLOT, A. C.; DEMPSEY, D. A.; KLESSIG, D. F. Salicylic acid, a multifaceted hormone to
combat disease. Annual Review of Phytopathology, v. 47, p. 177-206, 2009.
VOLKOV, R. A.; PANCHUK, I. I.; MULLINEAUX, P. M.; SCHÖFFL, F. Heat stress-
induced H2O2 is required for effective expression of heat shock genes in Arabidopsis.
Plant Molecular Biology, v. 61, 733-746, 2006.
VON ARX, J. A. Plant Pathogenic Fungi. Beheifte zur Nova Hedwigia. 87, J.
Cramer, Berlin, 1987.
WANG, W.; SCALI, M.; VIGNANI, R.; SPADAFORA, A.; et al. Protein extraction
for two-dimensional electrophoresis from olive leaf, a plant tissue containing high
levels of interfering compounds. Electrophoresis, v. 24, p. 2369-2375, 2003.
WANG, D.; WEAVER, N. D.; KESARWANI, M.; DONG, X. Induction of protein secretory
pathway is required for systemic acquired resistance. Science, v. 308, p. 1036-1040, 2005.
WANG, W.; VIGNANI, R.; SCALI, M.; CRESTI, M. A universal and rapid protocol for
protein extraction from recalcitrant plant tissues for proteomic analysis. Electrophoresis, v.
27, p. 2782–2786, 2006.

136
WANG, D.; PAJEROWSKA-MUKHTAR, K.; CULLER, A. H.; DONG, X. Salicylic acid
inhibits pathogen growth in plants through repression of the auxin signaling pathway. Current
Biology, v. 17, p.1784-1790, 2007.
WANG, D.; DONG, X. A highway for war and peace: the secretory pathway in plant-microbe
interactions. Molecular Plant, v. 4, p. 581-87, 2011.
WATKINSON, J. I.; SIOSON, A. A.; VASQUEZ-ROBINET, C.; SHUKLA, M.; KUMAR,
D.; ELLIS, M.; HEATH, L. S.; RAMAKRISHNAN, N.; CHEVONE, B.; WATSON, L. T.;
VAN ZYL, L.; EGERTSDOTTER, U.; SEDEROFF, R. R.; GRENE, R. Photosynthetic
acclimation is reflected in specific patterns of gene expression in drought-stressed loblolly
pine. Plant Physiology, v. 133, p. 1702-1716, 2003.
WEN, L. Cell Death in Plant Immune Response to Necrotrophs. Journal Plant Biochemistry
and Physiology, v. 1, p. 1-3, 2013.
WILDERMUTH, M. C. Modulation of host nuclear ploidy: a common plant biotroph
mechanism. Current Opinion in Plant Biology, v.13, p. 1-10, 2010.
WIRTHMUELLER, L.; MAQBOOL, A.; BANFIELD, M. J. On the front line: structural
insights into plant–pathogen interactions. Nature, v. 11, p. 761-776, 2013.
WIRTHMUELLER, L.; ZHANG, Y.; JONES, J. D.; PARKER, J. E. Nuclear accumulation of
the Arabidopsis immune receptor RPS4 is necessary for triggering EDS1-dependent defense.
Current Biology, v. 17, p. 2023-029, 2007.
WU, L.; HAN, Z.; WANG, S.; WANG, X.; SUN, A.; ZU, X.; CHEN, Y. Comparative
proteomic analysis of the plant–virus interaction in resistant and susceptible ecotypes of
maize infected with sugarcane mosaic virus. Journal of Proteomics, v. 89, p. 124-140, 2013.
YALPANI, N.; SILVERMAN, P.; WILSON, T.M.A.; KLEIER, D.A.; RASKIN, I. Salicylic
acid is a systemic signal and an inducer of pathogenesis-related proteins in virus-infected
tobacco. Plant Cell, v. 3, p. 809-818, 1991.
YANG, L.; LIN, H.; TAKAHASHI, Y.; CHEN, F.; WALKER, M. A.; CIVEROLO, E. L.
Proteomic analysis of grapevine stem in response to Xylella fastidiosa inoculation.
Physiological and Molecular Plant Pathology, v. 75, p. 90-99, 2011.
YANG, X.; WANG, W.; COLEMAN, M.; ORGIL, U.; FENG, J.; MA, X.; FERL, R. J.;
TURNER, J. G.; XIAO, S. Arabidopsis 14-3-3 lambda is a positive regulator of RPW8-
mediated disease resistance. Plant Journal, v. 60, p. 539-550, 2009.
YAO, Y.; YANG, Y. W.; LIU, J. Y. An efficient protein preparation for proteomic analysis of
developing cotton fibers by 2-DE. Electrophoresis, v. 27, p. 4559-4569, 2006.
YASUDA, M.; ISHIKAWA, A.; JIKUMARU, Y.; UMEZAWA, T.; ASAMI, T.;
MARUYAMA-NAKASHITA, A.; KUDO, T.; SHINOZAKI, K.; YOSHIDA, S.;
NAKASHITA, H. Antagonistic interaction between systemic acquired resistance and the
abscisic acid-mediated abiotic stress responses in Arabidopsis. Plant Cell, v. 20, p.1678-1692,
2008.

137
ZENG, W.; MELOTTO, M.; HE, S. Y. Plant stomata: a checkpoint of host immunity and
pathogen virulence. Current Opinion in Biotechnology, v. 21, p. 599-603, 2010.
ZHAO, F.; FANG, W.; XIE, D.; ZHAO, Y.; TANG, Z.; LI, W.; NIE, L.; LV, S. Proteomic
identification of differentially expressed proteins in Gossypium thurberi inoculated with
cotton Verticillium dahlia. Plant Science, v.185-186, p. 176-184, 2012.
ZHANG, D.W.; SHAO, J.; LIN, J.; ZHANG, N.; LU, B.J.; LIN, S.C.; DONG, M.Q.; HAN, J.
RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to
necrosis. Science, v. 325, p. 332–336, 2009.
ZIMARO, T.; GOTTIG, N.; GARAVAGLIA, B. S.; GEHRING, C.; OTTADO, J. Unraveling
plant responses to bacterial pathogens through proteomics. Journal of Biomedicine and
Biotechnology, v. 2011, p. 1-12, 2011.
ZIPFEL, C. Early molecular events in PAMP-triggered immunity. Current Opinion in Plant
Biology, v. 12, p.414-420, 2009.


















