Transgenic zebrafish that express tyrosine hydroxylase promoter in inner retinal cells
9
RESEARCH ARTICLE Transgenic Zebrafish That Express Tyrosine Hydroxylase Promoter in Inner Retinal Cells Yan Gao, Ping Li, and Lei Li * We have generated a transgenic zebrafish line [Tg(Th:GFP)] that expresses green fluorescence proteins (GFP) driven by rat tyrosine hydroxylase (TH) promoter. In zebrafish, the transgene was expressed as early as 16 hr postfertilization (hpf). The first transgene expression was detected in the midbrain. Within a few hours of development, the expression spread to the forebrain and hindbrain. In the retina, the first transgene expression was detected at approximately 40 hpf, at which time a single GFP-positive cell was seen in the ventral–nasal patch of the retina. In late development, GFP spread across the inner retina. GFP was found in retinal cells that expressed TH or phenylethanolamine N-methyl-transferase (PNMT), the first and last enzymes for synthesis of catecholamine, respectively. This suggests that the transgene is expressed in catecholaminergic neurons. Of interest, GFP was also detected in some retinal cells that release - aminobutyric acid. These latter data suggest that the transgene may also be expressed in noncatecholaminergic cells. Developmental Dynamics 233:921–929, 2005. © 2005 Wiley-Liss, Inc. Key words: catecholamine; transgenic; retina; development; zebrafish Received 21 January 2005; Revised 7 March 2005; Accepted 7 March 2005 INTRODUCTION Catecholamines (dopamine, and its derivatives norepinephrine and epi- nephrine) are abundant in the ner- vous systems, where they are involved in energy metabolism and the regula- tion of synaptic transmission, stress, addiction, motor function, and hor- monal homeostasis (Smeets and Reiner, 1994). Catecholamines are synthesized from the amino acid ty- rosine by a sequence of enzymatic steps (Wurtman and Axelrod, 1966; Axelrod, 1971; Armstrong et al., 1982). The first enzyme, known as the rate-limiting enzyme, is tyrosine hy- droxylase (TH). TH converts tyrosine to 3,4-dihydroxyphenylalanine (DOPA). Then, aromatic amino acid decarboxyl- ase (AACD) converts DOPA to dopa- mine, and dopamine -hyrodxylase (DBH) converts dopamine to norepi- nephrine. Lastly, phenylethanolamine N-methyl-transferase (PNMT) converts norepinephrine to epinephrine (Cooper et al., 1991; Smeets and Reiner, 1994). All the major enzymes are believed to be derived from a common gene family (Joh, 1985). The distribution of catecholamines in the central nervous system (CNS) has been carefully studied (Groves et al., 1995; Hynes et al., 1995; Puelles and Verney, 1998; Ma, 1994a,b, 1997, 2003). In mammals, catecholamines are primarily found in the forebrain and midbrain (Smeets and Reiner, 1994). In lower vertebrates, such as zebrafish, catecholamines are found in the forebrain, midbrain, and hind- brain (Guo et al., 1999a,b; Holzschuh et al., 2001; Ma and Lopez, 2003). Cat- echolamines, including dopamine and epinephrine, are also found in the neural retina. For example, dopamine is found in a subset of amacrine cells, whereas epinephrine is found in both amacrine cells and horizontal cells (Ehinger and Floren, 1976; Dowling and Ehinger, 1978; Adolph et al., 1980; Negishi et al., 1982; Witkovsky et al., 1984; Foster et al., 1985; Keyser et al., 1987; Baetge et al., 1988; Yazulla and Zucker, 1988; Baldridge and Ball, 1993). In the retina, catecholamines may function as either neurotransmit- ters or neuromodulators (for reviews, see Ehinger, 1982; Dowling, 1986; Wit- kovsky and Dearry, 1992). Recently, the course of development Department of Biological Sciences, University of Notre Dame, Notre Dame, Indiana Grant sponsor: NIH (NEI); Grant number: R01 EY13147; Grant number: R01 EY13680. *Correspondence to: Lei Li, Ph.D., Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556. E-mail: [email protected] DOI 10.1002/dvdy.20416 Published online 13 May 2005 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 233:921–929, 2005 © 2005 Wiley-Liss, Inc.
Transcript of Transgenic zebrafish that express tyrosine hydroxylase promoter in inner retinal cells

RESEARCH ARTICLE
Transgenic Zebrafish That Express TyrosineHydroxylase Promoter in Inner Retinal CellsYan Gao, Ping Li, and Lei Li*
We have generated a transgenic zebrafish line [Tg(Th:GFP)] that expresses green fluorescence proteins(GFP) driven by rat tyrosine hydroxylase (TH) promoter. In zebrafish, the transgene was expressed as earlyas 16 hr postfertilization (hpf). The first transgene expression was detected in the midbrain. Within a fewhours of development, the expression spread to the forebrain and hindbrain. In the retina, the firsttransgene expression was detected at approximately 40 hpf, at which time a single GFP-positive cell wasseen in the ventral–nasal patch of the retina. In late development, GFP spread across the inner retina. GFPwas found in retinal cells that expressed TH or phenylethanolamine N-methyl-transferase (PNMT), the firstand last enzymes for synthesis of catecholamine, respectively. This suggests that the transgene is expressedin catecholaminergic neurons. Of interest, GFP was also detected in some retinal cells that release �-aminobutyric acid. These latter data suggest that the transgene may also be expressed innoncatecholaminergic cells. Developmental Dynamics 233:921–929, 2005. © 2005 Wiley-Liss, Inc.
Key words: catecholamine; transgenic; retina; development; zebrafish
Received 21 January 2005; Revised 7 March 2005; Accepted 7 March 2005
INTRODUCTION
Catecholamines (dopamine, and itsderivatives norepinephrine and epi-nephrine) are abundant in the ner-vous systems, where they are involvedin energy metabolism and the regula-tion of synaptic transmission, stress,addiction, motor function, and hor-monal homeostasis (Smeets andReiner, 1994). Catecholamines aresynthesized from the amino acid ty-rosine by a sequence of enzymaticsteps (Wurtman and Axelrod, 1966;Axelrod, 1971; Armstrong et al.,1982). The first enzyme, known as therate-limiting enzyme, is tyrosine hy-droxylase (TH). TH converts tyrosineto 3,4-dihydroxyphenylalanine (DOPA).Then, aromatic amino acid decarboxyl-ase (AACD) converts DOPA to dopa-
mine, and dopamine �-hyrodxylase(DBH) converts dopamine to norepi-nephrine. Lastly, phenylethanolamineN-methyl-transferase (PNMT) convertsnorepinephrine to epinephrine (Cooperet al., 1991; Smeets and Reiner, 1994).All the major enzymes are believed tobe derived from a common gene family(Joh, 1985).
The distribution of catecholaminesin the central nervous system (CNS)has been carefully studied (Groves etal., 1995; Hynes et al., 1995; Puellesand Verney, 1998; Ma, 1994a,b, 1997,2003). In mammals, catecholaminesare primarily found in the forebrainand midbrain (Smeets and Reiner,1994). In lower vertebrates, such aszebrafish, catecholamines are found inthe forebrain, midbrain, and hind-
brain (Guo et al., 1999a,b; Holzschuhet al., 2001; Ma and Lopez, 2003). Cat-echolamines, including dopamine andepinephrine, are also found in theneural retina. For example, dopamineis found in a subset of amacrine cells,whereas epinephrine is found in bothamacrine cells and horizontal cells(Ehinger and Floren, 1976; Dowlingand Ehinger, 1978; Adolph et al.,1980; Negishi et al., 1982; Witkovskyet al., 1984; Foster et al., 1985; Keyseret al., 1987; Baetge et al., 1988; Yazullaand Zucker, 1988; Baldridge and Ball,1993). In the retina, catecholaminesmay function as either neurotransmit-ters or neuromodulators (for reviews,see Ehinger, 1982; Dowling, 1986; Wit-kovsky and Dearry, 1992).
Recently, the course of development
Department of Biological Sciences, University of Notre Dame, Notre Dame, IndianaGrant sponsor: NIH (NEI); Grant number: R01 EY13147; Grant number: R01 EY13680.*Correspondence to: Lei Li, Ph.D., Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556.E-mail: [email protected]
DOI 10.1002/dvdy.20416Published online 13 May 2005 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 233:921–929, 2005
© 2005 Wiley-Liss, Inc.

of catecholaminergic neurons hasbeen examined (Puelles and Verney,1998; Guo et al., 1999a,b; Holzschuhet al., 2001; Rink and Wullimann,2002). In zebrafish, the first THmRNA expression was detected in themidbrain at approximately 18 hourspostfertilization (hpf; Holzschuh etal., 2001). Within a few hours of devel-opment, TH mRNA expression spreadto the hindbrain and forebrain. THmRNA was also detected in the pe-ripheral nervous system (PNS), for ex-ample, in arch-associated neurons andsympathetic ganglia (Guo et al.,1999a,b; Holzschuh et al., 2001). Muta-tions in several genes that affect thedevelopment of catecholaminergic cells
in zebrafish have been identified (Guoet al., 1999a,b; Holzschuh et al., 2003).
In this study, we describe a trans-genic zebrafish model [Tg(Th:GFP)]that expresses green fluorescent pro-tein (GFP) under rat TH promoter.During development, TH-driven GFPexpression was first detected in themidbrain, then hindbrain and fore-brain. In the retina, GFP was detectedin the proximal inner nuclear layer(INL), ganglion cell layer (GCL), andinner plexiform layer (IPL). GFP wasfound in cells that expressed TH orPNMT, suggesting that the transgeneis expressed in catecholaminergicneurons. GFP was also found in someinner retinal cells that release GABA.
This is to our knowledge the firsttransgenic fish model that expressesGFP driven by the TH promoter.
RESULTS
Transgenic Fish
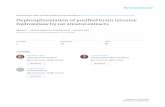
A 4.5-kb rat TH promoter (kindlyprovided by Dr. Dona Chikaraishi,Duke University) was cloned intoa pEGFP-1 vector (Fig. 1A, see alsoExperimental Procedures section).DNA was injected into one-cell stageembryos. After 24 hr, embryos wereexamined for transgene expressionunder a dissecting microscope thatwas connected to a fluorescent lightsource. Approximately 50% of injected
Fig. 1. Construction and expression of the trans-gene in developing zebrafish. A: A diagram show-ing the DNA fragment with restriction sites (HindIII/SphI) containing 4.5 kb of 5� DNA of rat tyrosinehydroxylase (TH) promoter, and 0.7 kb of cDNA ofenhanced green fluorescent protein (EGFP) with ashort poly-A (pA) tail. B,C: Transient TH-driven GFPexpression in the brain of founder fish at 26 hpf.Both cell bodies and axons can be identified. D:Polymerase chain reaction (PCR) showing germlinetransmission of the transgene. Lane 1, molecularmarkers; lane 2, positive control (plasmid DNA thatcontains 0.7 kb of cDNA of GFP); lane 3, wild-typecontrol. Lanes 4–15: PCR of genomic DNA from 12embryos selected from a cross between thefounder fish and a wild-type fish. Lanes 4–10: non-transgenic siblings; lanes 11–15, transgenic sib-lings. Each lane represents PCR from an individualembryo. Wnt expression (internal controls) was de-tected in every embryo. Scale bar � 20 �m in C(applies to B,C).
922 GAO ET AL.

embryos showed transient GFP ex-pression in the brain area (Fig. 1B,C).
Among 220 embryos that survived toadulthood (with or without transientGFP expression), we identified 44 fish(founders) that showed GFP expressionby performing polymerase chain reac-tion (PCR) with fin genomic DNA. Wecrossed each founder with wild-type ze-brafish, and screened their progeny byPCR for GFP expression. We identifiedone founder fish that showed germlinetransmission of the transgene; approxi-mately 50% of the progeny from crossesbetween this founder fish and wild-typefish showed TH-driven GFP expression(Fig. 1D). The progeny of this founderwere raised to adult and were used forbreeding colonies.
Transgene Expression in theBrain (16–18 hpf)
To determine the onset time and thelocation of TH-driven GFP transgeneexpression, we took live images oftransgenic embryos at different times ofdevelopment, beginning at 10 hpf, andrepeating at 2-hr intervals until 56 hpf.The embryos were treated with phenyl-2-thiourea (PTU, 0.003%) to preventthe development of melanin pigment.We found no obvious GFP expressionbefore 14 hpf (Fig. 2A). The first GFPexpression was seen at approximately16 hpf, at which time several clusters ofGFP-positive cells were seen scatteredin the brain areas, likely the midbrainand the boundary between the mid-brain and hindbrain (Fig. 2B). By 18hpf, more GFP expression was detected.Clusters of GFP-positive cells werefound in the forebrain, midbrain, andhindbrain (Fig. 2C).
Transgene Expression in theRetina (40–56 hpf)
In the retina, the transgene was firstexpressed at approximately 40 hpf, atwhich time a single GFP-positive cellwas detected in the ventral–nasal patchof the retina. The first GFP-positive cellwas always found in the inner retina, inclose proximity to the lens (Fig. 3A).Within 30 min of development, anotherGFP-positive cell was detected. It wasalways the case that the second cell ob-served was at a position anterior to thefirst cell (Fig. 3B).
Within a few hours of development,
GFP expression spread in the retina.At 46 hpf, GFP-positive cells were de-tected in both ventral–nasal and ante-rior areas (Fig. 4A). At 50 hpf, GFP-positive cells were seen in anterior–dorsal regions (Fig. 4B). By 56 hpf,nearly a full-circle of GFP expressionwas observed, except at the most ven-tral area where retinal ganglion cell(RGC) axons exit the retina (Fig. 4C).
In the retina, the first patch of GFPexpression was seen several hours afterthe onset time of differentiation ofRGCs. At 42 hpf, for example, only afew GFP-positive cells were detected inthe ventral–nasal patch of the retina(Fig. 5A). At this stage of development,differentiation of the ganglion cells wascompleted (Schmitt and Dowling, 1999).The 4�,6-diamidine-2-phenylidole-dihy-drochloride (DAPI) staining of the samesection revealed nuclei of both differen-tiated (in the inner retina) and undiffer-entiated cells (in the outer retina; Fig.5B).
GFP Was Expressed in theInner Retina
In the retina, GFP was confined to theinner retina (Fig. 6). In the proximalINL, GFP was seen in one to two lay-ers of cell bodies. Among them, someshowed strong GFP expression (ap-proximately 5% of the total GFP-pos-itive cells, arrows), whereas othersshowed weak expression. In the GCL,GFP was detected in cells, likely bothRGCs and displaced amacrine cells.Strong GFP expression was detectedin the IPL, where two narrow bandscould be readily identified. GFP wasalso detected in the optic nerve. Noobvious GFP expression was detectedin the distal INL, outer plexiformlayer, or outer nuclear layer.
GFP Was Found in Cells ThatExpressed TH or PNMT
To determine whether the transgene isexpressed in catecholaminergic cells,we labeled cryostat sections across theretina of transgenic fish with antibodiesagainst TH, DBH, and PNMT, threemajor enzymes found in the catechol-amine synthetic pathways. We identi-fied TH and PNMT immunoreactivity(IR) but no DBH IR. TH IR was de-tected in a subpopulation of amacrinecells in the proximal INL, known as do-
paminergic interplexiform cells (DA-IPC). While GFP was expressed in oneto two layers of cell bodies in the proxi-mal INL, TH IR was detected in only afew cells. No TH IR was detected in theGCL. Merged images showed that THIR and GFP were colocalized (Fig. 7, toppanel).
PNMT IR was detected in more celltypes. In the INL, PNMT antibodieslabeled one to two layers of cell bodies.Virtually all the cells that were la-beled with PNMT antibodies wereGFP-positive. Cells that were labeledwith PNMT antibodies were smallerin size than cells that were labeled byTH antibodies. In the GCL, all thecells were labeled with PNMT anti-bodies. Strong PNMT IR was also seenin the IPL. In the IPL, PNMT IRseemed to be distributed more broadlythan GFP. Merged images showedthat PNMT IR and GFP were colocal-ized (Fig. 7, bottom panel).
GFP Was Detected in InnerRetinal Cells That Released�-Aminobutyric Acid
Previous studies have shown that THand �-aminobutyric acid (GABA) can becoexpressed by the same neurons (Si-mon et al., 1989; Matsushita et al.,2002). We investigated whether this isthe case in zebrafish. We labeled retinalsections of transgenic fish with antibod-ies against GABA. GABA IR was de-tected in both inner and outer retinas.In the inner retina, GABA antibodieslabeled a subset of inner retinal cells.Merged images showed that GABA IRand GFP were colocalized. However,not all GFP-positive cells were labeledwith GABA antibodies (Fig. 8).
DISCUSSION
We report here a nonmosaic trans-genic zebrafish line that expressesGFP under rat TH promoter. Thesame TH promoter was characterizedpreviously in other vertebrate species.In mice, this promoter is expressed ina tissue-specific manner (Banerjee etal., 1992, 1994). In transgenic mice,however, this promoter drove gene ex-pressions in both type 1 (colocalizationwith TH antibodies) and type 2 (nocolocalization with TH antibodies) cat-echolaminergic cells (Gustincich et al.,1997; Zhang et al., 2004). We demon-
TH-DRIVEN GFP EXPRESSION IN ZEBRAFISH 923

strate that this same TH promoter canbe expressed in teleost. In zebrafish,the transgene is likely expressed inboth dopaminergic (colocalizationwith TH antibodies) and epinephri-nergic (colocalization with PNMTantibodies) cells. Some of the GFP-positive cells also express noncat-echolaminergic agents, e.g., GABA. Itis not clear whether this colocalizationis due to nonspecific transgene expres-sion in GABAergic cells or that GABA
is present in catecholaminergic cells.Coexistence of catecholamine andGABA was reported previously (Simonet al., 1989; Matsushita et al., 2002).
The distribution of catecholaminesthat was determined by transgene ex-pression in this study seemed to bebroader than the distribution of cat-echolamines determined using othermethods, e.g., immunocytochemistryand in situ hybridization (Ma,1994a,b, 1997, 2003; Guo et al., 1999a;
Holzschuh et al., 2001; Rink and Wul-limann, 2002; Ma and Lopez, 2003). Indeveloping zebrafish, only a fewgroups of cells can be identified in thebrain area by TH antibodies or by insitu hybridizations. Outside the CNS,catecholamines are found only in someenteric and sympathetic cells (Guo etal., 1999a; Holzschuh et al., 2001). Inour case, by contrast, TH-driven GFPexpression was detected in neuronsthat were scattered in much broader
Fig. 2. Live images showing green fluorescent protein (GFP) expres-sion in developing embryos. A: No obvious GFP was detected in theembryo at 14 hours postfertilization (hpf). B: At 16 phf, GFP-positive cellswere detected in the brain, likely the midbrain (arrow with solid line) andthe midbrain hindbrain boundary (arrow with dashed line). C: At 18 hpf,GFP-positive cells were seen in the forebrain, midbrain, and hindbrain.Dorsal is up and anterior is to the left. Tel, telencephalon; Dien, dien-cephalon; Hypo, hypothalamus; Mhb, midbrain hindbrain boundary.Scale bar in C� 200 �m in A, 100 �m in B,C.
924 GAO ET AL.

areas. This may be due to several rea-sons. First, under rat TH promoter thetransgene is expressed in all cat-echolaminergic cell types. Thus, notonly dopaminergic cells, but also othercatecholaminergic cells are labeled.Second, the transgene is expressed innoncatecholaminergic cells, such ascells that express GABA. Outside theCNS, GFP was found in cells along thelongitude of the body, dorsal to thenotochord and the spinal cord (datanot shown).
Despite that TH and PNMT func-tion in the same biosynthetic path-way, the two enzymes are not colocal-ized (Yazulla and Studholme, 2001). Alack of TH and PNMT colocalizationwas reported previously (Ross et al.,1984; Foster et al., 1985; Nguyen-Leg-ros et al., 1986; Keyser et al., 1987;Hammang et al., 1992). Only rarelywere TH and PNMT found to be colo-calized (Park et al., 1986). This findingraised the question of whether epi-nephrine is synthesized from tyrosine.
We offer three lines of suggestions.First, TH and PNMT may be colocal-ized, but because the concentration ofone of them is maintained at a lowlevel, colocalization cannot be readilydetected. The up- or down-regulationof TH activity was described in someearlier studies. In mice, for example,TH-driven GFP expression was de-tected at high levels during early de-velopment. However, GFP expressionwas markedly down-regulated in laterembryonic stages (Matsushita et al.,2002). In our case, TH antibodies la-beled a subpopulation of amacrinecells (DA-IPCs). These cells, however,showed only low levels of transgeneexpression, e.g., weak GFP (Fig. 7).Cells that showed bright GFP expres-sion, by contrast, were TH-negative.Second, TH and PNMT may be coex-pressed only transiently, and aftercertain periods of coexpression, eitherTH or PNMT activity is turned off dueto negative feedback mechanisms. Ithas been demonstrated that the enzy-
mic activity of TH is negatively con-trolled by catecholamine (Spector etal., 1967; Dairman et al., 1971; Na-gatsu et al., 1984; Phillips et al.,2001). Third, some epinephrine maybe synthesized from precursors otherthan tyrosine. It is suspected thatPNMT would not distribute morebroadly than TH if all epinephrine issynthesized from tyrosine. In our case,TH antibodies labeled only a smallportion of amacrine cells, whereasPNMT antibodies labeled one to twolayers of cell bodies in the proximalINL and GCL. It has been suggestedthat epinephrine may be synthesizedfrom octopamine and synephrine bymeans of different pathways (Fosteret al., 1985; Ruggiero et al., 1985).
In summary, the Tg(Th:GFP) trans-genic fish provides a model for in vivoanalysis of the catecholaminergic sys-tem. It provides means for studyingnot only the development of cat-echolaminergic cells but also the dis-tribution of different classes of cat-
Fig. 3. Live images showing early green flu-orescent protein (GFP) expression in the ret-ina. Both images were taken from the sameretina. A: At 40 hours postfertilization (hpf), asingle GFP-positive cell was seen in the ven-tral–nasal patch of the retina (arrow). B: At40.5 hpf, another GFP-positive cell was iden-tified (arrow with dashed line). The second cellwas anterior to the first cell. Dashed circlesoutline the lens. Anterior is to the left anddorsal is up. L, lens. Scale bar � 20 �m in B(applies to A,B).
Fig. 4. Live images taken from a developing retina between 46 and 56 hours postfertilization (hpf). A: At 46 hpf, only a few green fluorescent protein (GFP)-positive cells were seen in the ventral–nasal patch of the retina. B: At 50 hpf, GFP was detected in the anterior and anterior–dorsal regions. C: By 56 hpf,nearly a full circle of GFP expression was detected, except at the most ventral area where the optic nerve exits. Dashed circles outline the lens, and dashedlines outline the optic nerve. Anterior is to the left and dorsal is up. L, lens; op, optic nerve. Scale bar � 50 �m in C (applies to A–C).
TH-DRIVEN GFP EXPRESSION IN ZEBRAFISH 925

echolamines. Furthermore, it providesmeans for studying the function of in-ner retinal cells that express thetransgene.
EXPERIMENTALPROCEDURES
Fish Care
Zebrafish (Danio rerio) were main-tained as described (Westerfield,1995). Breeding colonies were kept ina 14:10 light–dark cycle (room fluo-rescent light, 7:00 AM–9:00 PM). To setup crosses, paired male and female
fish were transferred to mating cagesthe day before they were allowed tomate. The mating cage was dividedinto two chambers by a plastic plateinserted in the middle of the cage. At7:00 AM on the following day, the plas-tic plate was removed, and the fishwere allowed to mate for 30 min. Eggswere collected immediately (within 30min) after mating and were lined upfor DNA injections. After DNA injec-tions, embryos were transferred to5-cm Petri dishes containing 100 ml ofE3 medium (Westerfield, 1995). PTU(0.003%; Sigma) was added to the me-
dium to prevent the development ofmelanin pigment (Westerfield, 1995).Embryos were staged according toKimmel et al. (1995).
Plasmid Construction andMicroinjection
A 4.5-kb rat TH promoter (Banerjee etal., 1992, 1994) was cloned into thepEGFP-1 vector (Clonetech). PlasmidDNA was prepared by using the Qia-gen plasmid kit. DNA fragments ofTH::GFP expression cassette (restric-tion sites, HindIII and SphI) were
Fig. 5. A,B: A sagittal section across the ret-ina of a transgenic embryo showing green flu-orescent protein (GFP, A) and 4�,6-diamidine-2-phenylidole-dihydrochloride (DAPI) staining(B) at 42 hours postfertilization (hpf). At thisstage of development, GFP was detected onlyin a few cells in the ventral–nasal areas of theretina (A, arrows). DAPI staining revealed nu-clei of both differentiated (inner retina) andundifferentiated (outer retina) cells. Anterior isto the left and dorsal is up. L, lens. Scale bar �50 �m in B (applies to A,B).
Fig. 6. A cryostat section across the retina of a trans-genic fish at 7 days postfertilization. Green fluorescentprotein (GFP) was seen in one to two layers of cellbodies in the proximal INL (pINL). GFP was also de-tected in the IPL, where two GFP-positive bands wereidentified. In the GCL, GFP was seen in both ganglioncells and displaced amacrine cells. The optic nerve wasalso GFP-positive. No GFP was detected in the distalINL (dINL), OPL, or ONL. The arrows point toward ret-inal cells that show strong GFP expression. The dashedcircle outlines the lens, and the dashed lines outline theOPL. GCL, ganglion cell layer; INL, inner nuclear layer;IPL, inner plexiform layer; ONL, outer nuclear layer;OPL, outer plexiform layer; op, optic nerve. Scale bar �50 �m.
926 GAO ET AL.

electrophoresed and recovered withthe Qiagen gel extraction kit. DNAwas dissolved in 1� Daniean buffer(58 mM NaCl, 0.7 mM KCl, 0.4 MMgSO4, 0.6 mM Ca(NO3)2, 5 mMHEPES, pH 7.6) and was injected (4.6nl, 50 ng/�l) into one-cell stage em-bryos (approximately 0.5 hpf) using a
glass pipette (tip diameter, approxi-mately 5 �m) held by a microinjector.
PCR
Genomic DNA was extracted by lysingthe embryos in 100 mM Tris, pH 8.3,200 mM NaCl, 0.4% SDS, 5 mM eth-
ylenediaminetetraacetic acid (EDTA),and 200 �g/ml proteinase K. We de-signed two pairs of primers, one forGFP (forward 5�-GGG CGA GGA GCTGTT CAC CGG, reverse 5�-CGG CGGCGG TCA CGA ACT CC, which am-plify a 674-bp band) and the other forWnt (forward 5�-CAG TTC TCA CGT
Fig. 7. Cryostat sections across the retina ofa transgenic fish (7 dpf) labeled with tyrosinehydroxylase (TH, top panel) or phenylethano-lamine N-methyl-transferase (PNMT, bottompanel) antibodies. TH antibodies labeled a do-paminergic interplexiform cells and its axons.The merged image shows that TH immunore-activity and GFP were colocalized (arrows).PNMT antibodies labeled one to two layers ofcell bodies in the proximal INL and GCL.PNMT also labeled the IPL. The merged imageshows that PNMT immunoreactivity and GFPwere colocalized (arrows). GCL, ganglion celllayer; INL, inner nuclear layer; IPL, inner plex-iform layer. Scale bar � 20 �m.
Fig. 8. A cryostat section across the inner retina of a transgenic fish (7days postfertilization) showing green fluorescent protein (GFP) expression(top panel), �-aminobutyric acid immunoreactivity (GABA IR, middle panel),and the colocalization of GFP and GABA IR (bottom panel). In the innerretina, GABA IR was detected in a subpopulation of amacrine cells. GFPwas seen in all GABA IR-positive cells. Note that some of the GFP-positivecells (arrows) were not labeled by GABA antibodies. IPL, inner plexiformlayer. Scale bar � 20 �m.
TH-DRIVEN GFP EXPRESSION IN ZEBRAFISH 927

CTG CTA CTT GCA, reverse 5�-ACTTCC GGC GTG TTG GAG AAT TC,which amplify a 387-bp band). Wntwas used for an internal control. ThePCR was run in 1� PCR buffer, with0.25 units Taq polymerase (Invitro-gen), 1.5 mM of MgCl2, 0.2 mM ofdNTP, and 0.1 �M of each primer. Thereaction was performed with an initial2-min denaturation step at 94°C fol-lowed by 30 cycles of 94°C for 45 sec,65°C for 30 sec, and 72°C for 1 min,and a final extension at 72°C for 10min.
Live Imaging
Live images were taken from PTU-treated transgenic embryos that wereembedded in E3 culture medium con-taining 0.5% low-melting agarose and0.01% 3-amino benzoic acid methyl-ester. Embryos were first dechorion-ated and then transferred to the aga-rose solution. The agarose wasallowed to cool to room temperature,then trimmed to 2 � 1 � 1 mm blockswith the embryo in the center of eachblock. The agarose block was placed tohanging-drop glass slides and coveredwith glass coverslips. The coverslipwas not sealed at the edges, thus al-lowing oxygen to pass onto the aga-rose block. Live images were takenusing a Nikon microscope that wasconnected to a fluorescent light sourceand a CCD camera with autoexpo-sure. Images were taken at differenttimes of development, beginning at 10hpf, and repeating at 2-hr intervalsuntil 56 hpf.
DAPI Staining
DAPI staining was performed usingcryostat sections. Embryos were em-bedded in OCT compound (Sigma).Cryostat sections of 16 �m weremounted on gelatin-treated slides andwere air-dried for 2 hr. Specimenswere rinsed in phosphate buffered sa-line (PBS) for three times, 5 min each,and were incubated with DAPI (0.01%in PBS; Sigma) for 10 min. Specimenswere washed with PBS for threetimes, 5 min each, and covered withglass coverslips. Specimens wereviewed under a Nikon microscope thatwas connected to a fluorescent lightsource.
Immunocytochemistry
Protocols for immunocytochemistrywere similar as described (Schmittand Dowling, 1996; Li and Dowling,1997, 2000a,b). All the antibodieswere purchased from Chemicon. Em-bryos were fixed in 4% paraformalde-hyde in PBS for 2 hr at room temper-ature. After three washes in PBS, 5min each, the embryos were trans-ferred to 15% and 30% sucrose in PBS,30 min each, and were embedded inOCT compound. Cryostat sections of16 �m were mounted on gelatin-treated glass slides and were air-driedfor 2 hr. After three washes with PBS,5 min each, specimens were incubatedwith blocking solutions containing 5%normal goat serum and 0.1%Tween-20 in PBS. Specimens were in-cubated with primary antibodies at4°C overnight (anti-GABA, 1:200; an-ti-TH, 1:200; anti-DBH, 1:200; anti-PNMT, 1:500; the anti-PNMT anti-bodies we used in this study have notbeen tested for their specificity in ze-brafish, but their staining patternswere similar to those described in anearlier study that used PNMT anti-bodies from a different source; see Ya-zulla and Studholme, 2001). Afterthree washes in PBS, 10 min each,specimens were incubated with rho-damine-conjugated secondary anti-bodies for 30 min at room tempera-ture. After five washes in PBS, 5 mineach, specimens were covered withglass coverslips and viewed under aconfocal microscope.
ACKNOWLEGMENTSThe authors thank Dr. Dona Chikara-ishi of Duke University for providingus with the rat TH promoter. The au-thors thank Drs. Steve Massey (UTHouston), Steve Yazulla (SUNY StonyBrook), and Tom Vihtelic (NotreDame) for suggestions on the immu-nocytochemical experiments. The au-thors also thank Drs. John Dowling(Harvard) and Berndt Ehinger (Uni-versity of Lund) for providing com-ments on the manuscript, and AprellCarr and Sitar Shah for proofreading.The authors thank Deb Bang at theCenter for Zebrafish Research at theUniversity of Notre Dame for main-taining the zebrafish colonies. L.L.was funded by the NIH (NEI).
REFERENCES
Adolph A, Dowling JE, Ehinger B. 1980.Monoaminergic neurons of the mudp-uppy retina. Cell Tissue Res 210:269–282.
Armstrong DM, Ross CA, Pickel VM, JohTH, Reis DJ. 1982. Distribution of dopa-mine-, noradrenaline-, and adrenaline-containing cell bodies in the rat medullaoblongata: demonstrated by the immu-nocytochemical localization of catechol-amine biosynthetic enzymes. J CompNeurol 212:173–187.
Axelrod J. 1971. Noradrenaline: fate andcontrol of its biosynthesis. Science 173:598–606.
Baetge EE, Behringer RR, Messing A,Brinster RL, Palmiter RD. 1988. Trans-genic mice express the human phe-nylethanolamine N-methyltransferasegene in adrenal medulla and retina. ProcNatl Acad Sci U S A 85:3648–3652.
Baldridge WH, Ball AK. 1993. A new typeof interplexiform cell in the goldfish ret-ina is PNMT-immunoreactive. Neurore-port 4:1015–1018.
Banerjee SA, Hoppe P, Brilliant M,Chikaraishi DM. 1992. 5� flanking se-quences of the rat tyrosine hydroxylasegene target accurate tissue-specific, de-velopmental, and transsynaptic expres-sion in transgenic mice. J Neurosci 12:4460–4467.
Banerjee SA, Roffler-Tarlov S, Szabo M,Frohman L, Chikaraishi DM. 1994. DNAregulatory sequences of the rat tyrosinehydroxylase gene direct correct cat-echolaminergic cell-type specificity of ahuman growth hormone reporter in theCNS of transgenic mice causing a dwarfphenotype. Brain Res Mol Brain Res 24:89–106.
Cooper JR, Bloom FE, Roth RH. 1991. Thebiochemical basis of neuropharmacology.New York: Oxford University Press.
Dairman W, Christenson JG, UdenfriendS. 1971. Decrease in liver aromatic L-amino-acid decarboxylase produced bychronic administration of L-dopa. ProcNatl Acad Sci U S A 68:2117–2120.
Dowling JD. 1986. Dopamine: a retinalneuromodulator. Trends Neurosci 9:236–240.
Dowling JE, Ehinger B. 1978. Synaptic or-ganization of the dopaminergic neuronsin the rabbit retina. J Comp Neurol 180:203–220.
Ehinger B. 1982. Neurotransmitter sys-tems in the retina. Retina 2:305–321.
Ehinger B, Floren I. 1976. Indoleamine-accumulating neurons in the retina ofrabbit, cat and goldfish. Cell Tissue Res175:37–48.
Foster GA, Hokfelt T, Coyle JT, GoldsteinM. 1985. Immunohistochemical evidencefor phenylethanolamine-N-methyltrans-ferase-positive/tyrosine hydroxylase-negative neurons in the retina and theposterior hypothalamus of the rat. BrainRes 330:183–188.
Groves AK, George KM, Tissier-Seta JP,Engel JD, Brunet JF, Anderson DJ.1995. Differential regulation of tran-
928 GAO ET AL.

scription factor gene expression and phe-notypic markers in developing sympa-thetic neurons. Development 121:887–901.
Guo S, Wilson SW, Cooke S, Chitnis AB,Driever W, Rosenthal A. 1999a. Muta-tions in the zebrafish unmask sharedregulatory pathways controlling the de-velopment of catecholaminergic neurons.Dev Biol 208:473–487.
Guo S, Brush J, Teraoka H, Goddard A,Wilson SW, Mullins MC, Rosenthal A.1999b. Development of noradrenergicneurons in the zebrafish hindbrain re-quires BMP, FGF8, and the homeodo-main protein soulless/Phox2a. Neuron 24:555–566.
Gustincich S, Feigenspan A, Wu DK, Koop-man LJ, Raviola E. 1997. Control of do-pamine release in the retina: a trans-genic approach to neural networks.Neuron 18:723–736.
Holzschuh J, Ryu S, Aberger F, Driever W.2001. Dopamine transporter expressiondistinguishes dopaminergic neuronsfrom other catecholaminergic neurons inthe developing zebrafish embryo. MechDev 101:237–243.
Holzschuh J, Hauptmann G, Driever W.2003. Genetic analysis of the roles of Hh,FGF8, and nodal signaling during cat-echolaminergic system development inthe zebrafish brain. J Neurosci 23:5507–5519.
Hammang JP, Bohn MC, Messing A. 1992.Phenylethanolamine N-methyltransferase(PNMT)-expressing horizontal cells in therat retina: a study employing double-labelimmunohistochemistry.JCompNeurol316:383–389.
Hynes M, Porter JA, Chiang C, Chang D,Tessier-Lavigne M, Beachy PA,Rosenthal A. 1995. Induction of mid-brain dopaminergic neurons by Sonichedgehog. Neuron 15:35–44.
Joh TH. 1985. Genes for neurotransmittersynthesis, storage and release. Prog ClinBiol Res 192:423–429.
Keyser KT, Karten HJ, Katz B, Bohn MC.1987. Catecholaminergic horizontal andamacrine cells in the ferret retina.J Neurosci 7:3996-4004.
Kimmel CB, Ballard WW, Kimmel SR, Ul-lmann B, Schilling TF. 1995. Stages ofembryonic development of the zebrafish.Dev Dyn 203:253–310.
Li L, Dowling JE. 1997. A dominant form ofinherited retinal degeneration caused bya non- photoreceptor cell-specific muta-tion. Proc Natl Acad Sci U S A 94:11645–11650.
Li L, Dowling JE. 2000a. Disruption of theolfactoretinal centrifugal pathway mayrelate to the visual system defect innight blindness b mutant zebrafish.J Neurosci 20:1883–1892.
Li L, Dowling JE. 2000b. Effects of dopa-mine depletion on visual sensitivity ofzebrafish. J Neurosci 20:1893–1903.
Ma PM. 1994a. Catecholaminergic systemsin the zebrafish. I. Number, morphology,and histochemical characteristics of neu-rons in the locus coeruleus. J Comp Neu-rol 344:242–255.
Ma PM. 1994b. Catecholaminergic systemsin the zebrafish. II. Projection pathwaysand pattern of termination of the locuscoeruleus. J Comp Neurol 344:256–269.
Ma PM. 1997. Catecholaminergic systemsin the zebrafish. III. Organization andprojection pattern of medullary dopami-nergic and noradrenergic neurons.J Comp Neurol 381:411–427.
Ma PM. 2003. Catecholaminergic systemsin the zebrafish. IV. Organization andprojection pattern of dopaminergic neu-rons in the diencephalon. J Comp Neurol460:13–37.
Ma PM, Lopez M. 2003. Consistency in thenumber of dopaminergic paraventricularorgan- accompanying neurons in the pos-terior tuberculum of the zebrafish brain.Brain Res 967:267–272.
Matsushita N, Okada H, Yasoshima Y, Ta-kahashi K, Kiuchi K, Kobayashi K. 2002.Dynamics of tyrosine hydroxylase pro-moter activity during midbrain dopami-nergic neuron development. J Neuro-chem 82:295–304.
Nagatsu T, Yamaguchi T, Rahman MK,Trocewicz J, Oka K, Hirata Y, Nagatsu I,Narabayashi H, Kondo T, Iizuka R.1984. Catecholamine-related enzymesand the biopterin cofactor in Parkinson’sdisease and related extrapyramidal dis-eases. Adv Neurol 40:467–473.
Negishi K, Teranishi T, Kato S. 1982. Newdopaminergic and indoleamine-accumu-lating cells in the growth zone of goldfishretinas after neurotoxic destruction. Sci-ence 216:747–749.
Nguyen-Legros J, Martin-Martinelli E, Si-mon A, Denoroy L, Vigny A. 1986. Co-localization of tyrosine hydroxylase andphenylethanolamine-N-methyltrans-ferase immunoreactivity in the rat ret-ina: a re-examination using double label-ing on semi-thin sections. Exp Eye Res43:575–584.
Park DH, Teitelman G, Evinger MJ, WooJI, Ruggiero DA, Albert VR, Baetge EE,Pickel VM, Reis DJ, Joh TH. 1986. Phe-nylethanolamine N-methyltransferase-containing neurons in rat retina: immu-nohistochemistry, immunochemistry, andmolecular biology. J Neurosci 6:1108–1113.
Phillips JK, Goodchild AK, Dubey R, Se-siashvili E, Takeda M, Chalmers J,Pilowsky PM, Lipski J. 2001. Differen-tial expression of catecholamine biosyn-thetic enzymes in the rat ventrolateralmedulla. J Comp Neurol 432:20–34.
Puelles L, Verney C. 1998. Early neuro-meric distribution of tyrosine-hydroxy-lase- immunoreactive neurons in humanembryos. J Comp Neurol 394:283–308.
Rink E, Wullimann MF. 2002. Develop-ment of the catecholaminergic system inthe early zebrafish brain: an immunohis-tochemical study. Brain Res Dev BrainRes 137:89–100.
Ross CA, Ruggiero DA, Meeley MP, ParkDH, Joh TH, Reis DJ. 1984. A new groupof neurons in hypothalamus containingphenylethanolamine N-methyltrans-ferase (PNMT) but not tyrosine hydrox-ylase. Brain Res 306:349–353.
Ruggiero DA, Ross CA, Anwar M, ParkDH, Joh TH, Reis DJ. 1985. Distributionof neurons containing phenylethano-lamine N-methyltransferase in medullaand hypothalamus of rat. J Comp Neurol239:127–154.
Schmitt EA, Dowling JE. 1996. Compari-son of topographical patterns of ganglionand photoreceptor cell differentiation inthe retina of the zebrafish, Danio rerio.J Comp Neurol 371:222–234.
Schmitt EA, Dowling JE. 1999. Early reti-nal development in the zebrafish, Daniorerio: light and electron microscopicanalyses. J Comp Neurol 404:515–536.
Simon A, Versaux-Botteri C, Denoroy L,Vigny A, Nguyen-Legros J. 1989. Doubleantigen localization of two catechol-amine enzymes and GABA in amacrinecells of the rat retina in semi-thin sec-tions. J Neurosci Methods 27:181–189.
Smeets WJAJ, Reiner A. 1994. Phylogenyand development of catecholamine sys-tem in the CNS of vertebrates. Cam-bridge: Cambridge University Press.
Spector S, Gordon R, Sjoerdsma A, Uden-friend S. 1967. End-product inhibition oftyrosine hydroxylase as a possible mech-anism for regulation of norepinephrinesynthesis. Mol Pharmacol 3:549–555.
Westerfield M. 1995. The zebrafish book: aguide for the laboratory use of zebrafish(Brachydanio rerio). Eugene: Universityof Oregon Press.
Witkovsky P, Dearry A. 1992. Functionalroles of dopamine in the vertebrate ret-ina. Prog Retinal Res 11:247–292.
Witkovsky P, Eldred W, Karten HJ. 1984.Catecholamine- and indoleamine-con-taining neurons in the turtle retina.J Comp Neurol 228:217–225.
Wurtman RJ, Axelrod J. 1966. Control ofenzymatic synthesis of adrenaline in theadrenal medulla by adrenal cortical ste-roids. J Biol Chem 241:2301–2305.
Yazulla S, Studholme KM. 2001. Neuro-chemical anatomy of the zebrafish retinaas determined by immunocytochemistry.J Neurocytol 30:551–592.
Yazulla S, Zucker CL. 1988. Synaptic orga-nization of dopaminergic interplexiformcells in the goldfish retina. Vis Neurosci1:13–29.
Zhang DQ, Stone J, Zhou TR, Ohta H, Mc-Mahon DG. 2004. Characterization of ge-netically labeled catecholaminergic neu-rons in the mouse retina. Neuroreport15:1761–1765.
TH-DRIVEN GFP EXPRESSION IN ZEBRAFISH 929