Tyrosine 3-hydroxylase in rat brain and adrenal medulla ...
5
Proc. Natl. Acad. Sci. USA Vol. 83, pp. 6208-6212, August 1986 Neurobiology Tyrosine 3-hydroxylase in rat brain and adrenal medulla: Hybridization histochemistry and immunohistochemistry combined with retrograde tracing (in situ hybridization/immunofluorescence/dopamine neurons) M. SCHALLING*, T. HOKFELT*, B. WALLACEt, M. GOLDSTEINt, D. FILERS, C. YAMINt, AND D. H. SCHLESINGERt *Department of Histology, Karolinska Institutet, Stockholm, and tDepartment of Molecular Biology, UmeA University, Umef, Sweden; and tDepartment of Psychiatry, New York University, Medical Center, School of Medicine, New York, NY Contributed by T. Hokfelt, April 21, 1986 ABSTRACT Rat brain and adrenal gland were analyzed by hybridization histochemistry using an RNA probe comple- mentary to mRNA for tyrosine 3-hydroxylase (TyrOHase; tyrosine 3-monooxygenase, EC 1.14.16.2), by immunohisto- chemistry using TyrOHase antiserum, and by retrograde tracing using the fluorescent compound Fast blue. Cell bodies in the ventral mesencephalon contained mRNA for TyrOHase, and these cells were also TyrOHase immunoreactive. After injection of Fast blue into the striatum, such double-labeled cells in addition contained the retrograde tracer, showing that these cells send axonal projections to the injection site. These results show that hybridization histochemistry can be used to identify transmitter-specific neuron populations and that their projections can be established. Histochemical techniques have traditionally been important in analyzing catecholamine-containing cell systems. Thus, two populations of adrenal medullary gland cells containing, respectively, norepinephrine and epinephrine were early defined with histochemistry (1-3). Catecholamine neurons were visualized in the brain and periphery (4-6) with form- aldehyde-induced fluorescence (7) as well as with immuno- histochemical procedures (8) using the indirect im- munofluorescence technique of Coons and collaborators (9). More recently, in situ hybridization has been employed to visualize at the cellular level the localization of various types of mRNA (10-12), including mRNA for hormones in endo- crine cells (13) and for peptides in neurons (14-21). Further- more, two groups have isolated cDNA clones coding for tyrosine 3-hydroxylase (TyrOHase; tyrosine 3-monooxygen- ase, EC 1.14.16.2) (22, 23), opening up possibilities to localize catecholamine neurons with hybridization histochemistry. In the present study, we have analyzed mesencephalon and adrenal gland of rat, using a single-stranded RNA probe complementary to mRNA for TyrOHase, the rate limiting enzyme in catecholamine synthesis and a marker for all types of catecholamine neurons. The in situ hybridization has been combined with immunohistochemistry using antiserum to TyrOHase to correlate presence of mRNA and its product (cf. refs. 14-16, 18-21). Further, since mRNA, in contrast to its products, is not transported into the axonal processes, projections of cmRNA-labeled neurons have been studied with retrograde tracing, similar to studies of projections with retrograde tracing combined with immunohistochemistry (24). MATERIALS AND METHODS Tissue Treatment. Male Sprague-Dawley rats (150-200 g; ALAB, Solna, Sweden) were used. Some rats were injected unilaterally into the caudate nucleus with the retrograde tracer Fast blue (0.5 g4 of a 3% aqueous solution) (25) 48 hr before they were killed. The rats were anesthetized with Nembutal (50 mg/kg, i.p.) and perfused through the ascend- ing aorta with ice-cold 0.9% NaCl followed either (i) by ice-cold 10% (vol/vol) formalin in 0.1 M phosphate buffer (26) with or without 0.2% glutaraldehyde for 15 min or (it) by 10% formalin containing 0.4% picric acid in 0.16 M phosphate buffer (27) for 6 min. The brains were dissected out and immersed in the respective fixative for 2 hr. After rinsing in 10% (wt/vol) sucrose in phosphate buffer for 18-24 hr, frontal sections of the midbrain were cut in a cryostat (Dittes, Heidelberg, F.R.G.) at 14 gum and thawed onto slides treated twice with chrom alum. Immediately after cutting, sections of Fast blue-injected brains were analyzed in a Zeiss fluores- cence microscope equipped with a dark-field oil condenser, a Schott UG1 excitation filter, and a Zeiss 41 stop filter. Fluorescent cells were photographed with Kodak Tri-X black-and-white film (Kodak). The sections were then processed essentially as described by Hafen and Levine (28): the sections were air-dried at room temperature for 1-2 hr, immersed in l1o formalin in phos- phate-buffered saline (PBS: 10 mM phosphate, pH 7.2-7.4/ 0.8% NaCl) at room temperature for 20 min, rinsed three times in PBS, dehydrated in ethanol, air-dried, and stored overnight at 40C. To increase permeability, the sections were incubated for 20 min at room temperature in 0.2 M HCl. The sections were then rinsed for 5 min in distilled water, incubated in 0.3 M sodium chloride/0.03 M sodium citrate at 70'C for 30 min, rinsed for 5 min in distilled water, and treated with Pronase (Calbiochem; 1.0 mg/ml in 50 mM Tris HCl buffer, pH 7.5, containing 5 mM EDTA). The Pronase was preincubated at 370C for 3-4 hr to eliminate nuclease activity. After a 30-sec rinse in PBS containing glycine (2 mg/ml), followed by two rinses in PBS, the sections were fixed in 10% formalin in PBS for 20 min at room temperature and finally rinsed in PBS and dehydrated in ethanol. Preparation of Probes. Two TyrOHase probes, pTH1 (792 base pairs) and pTH4 (400 base pairs) (22, 23) were used. The pTH1 probe was subcloned into the pGEM-1 vector (Pro- mega Biotec, Madison, WI) after digestion with Pst I endo- nuclease (New England Biolabs) and purification of the plasmid and the insert by gel electrophoresis (29). Run-off transcripts were prepared for T7 and SP6 transcription essentially according to the protocol from Promega Biotec, Abbreviation: TyrOHase, tyrosine 3-hydroxylase. 6208 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of Tyrosine 3-hydroxylase in rat brain and adrenal medulla ...

Proc. Natl. Acad. Sci. USAVol. 83, pp. 6208-6212, August 1986Neurobiology
Tyrosine 3-hydroxylase in rat brain and adrenal medulla:Hybridization histochemistry and immunohistochemistrycombined with retrograde tracing
(in situ hybridization/immunofluorescence/dopamine neurons)
M. SCHALLING*, T. HOKFELT*, B. WALLACEt, M. GOLDSTEINt, D. FILERS, C. YAMINt, AND D. H.SCHLESINGERt*Department of Histology, Karolinska Institutet, Stockholm, and tDepartment of Molecular Biology, UmeA University, Umef, Sweden; and tDepartment ofPsychiatry, New York University, Medical Center, School of Medicine, New York, NY
Contributed by T. Hokfelt, April 21, 1986
ABSTRACT Rat brain and adrenal gland were analyzedby hybridization histochemistry using an RNA probe comple-mentary to mRNA for tyrosine 3-hydroxylase (TyrOHase;tyrosine 3-monooxygenase, EC 1.14.16.2), by immunohisto-chemistry using TyrOHase antiserum, and by retrogradetracing using the fluorescent compound Fast blue. Cell bodiesin the ventral mesencephalon contained mRNA for TyrOHase,and these cells were also TyrOHase immunoreactive. Afterinjection of Fast blue into the striatum, such double-labeledcells in addition contained the retrograde tracer, showing thatthese cells send axonal projections to the injection site. Theseresults show that hybridization histochemistry can be used toidentify transmitter-specific neuron populations and that theirprojections can be established.
Histochemical techniques have traditionally been importantin analyzing catecholamine-containing cell systems. Thus,two populations of adrenal medullary gland cells containing,respectively, norepinephrine and epinephrine were earlydefined with histochemistry (1-3). Catecholamine neuronswere visualized in the brain and periphery (4-6) with form-aldehyde-induced fluorescence (7) as well as with immuno-histochemical procedures (8) using the indirect im-munofluorescence technique of Coons and collaborators (9).More recently, in situ hybridization has been employed to
visualize at the cellular level the localization of various typesof mRNA (10-12), including mRNA for hormones in endo-crine cells (13) and for peptides in neurons (14-21). Further-more, two groups have isolated cDNA clones coding fortyrosine 3-hydroxylase (TyrOHase; tyrosine 3-monooxygen-ase, EC 1.14.16.2) (22, 23), opening up possibilities to localizecatecholamine neurons with hybridization histochemistry. Inthe present study, we have analyzed mesencephalon andadrenal gland of rat, using a single-stranded RNA probecomplementary to mRNA for TyrOHase, the rate limitingenzyme in catecholamine synthesis and a marker for all typesof catecholamine neurons. The in situ hybridization has beencombined with immunohistochemistry using antiserum toTyrOHase to correlate presence of mRNA and its product(cf. refs. 14-16, 18-21). Further, since mRNA, in contrast toits products, is not transported into the axonal processes,projections of cmRNA-labeled neurons have been studiedwith retrograde tracing, similar to studies of projections withretrograde tracing combined with immunohistochemistry(24).
MATERIALS AND METHODS
Tissue Treatment. Male Sprague-Dawley rats (150-200 g;ALAB, Solna, Sweden) were used. Some rats were injectedunilaterally into the caudate nucleus with the retrogradetracer Fast blue (0.5 g4 of a 3% aqueous solution) (25) 48 hrbefore they were killed. The rats were anesthetized withNembutal (50 mg/kg, i.p.) and perfused through the ascend-ing aorta with ice-cold 0.9% NaCl followed either (i) byice-cold 10% (vol/vol) formalin in 0.1 M phosphate buffer(26) with or without 0.2% glutaraldehyde for 15 min or (it) by10% formalin containing 0.4% picric acid in 0.16 M phosphatebuffer (27) for 6 min. The brains were dissected out andimmersed in the respective fixative for 2 hr. After rinsing in10% (wt/vol) sucrose in phosphate buffer for 18-24 hr,frontal sections ofthe midbrain were cut in a cryostat (Dittes,Heidelberg, F.R.G.) at 14 gum and thawed onto slides treatedtwice with chrom alum. Immediately after cutting, sections ofFast blue-injected brains were analyzed in a Zeiss fluores-cence microscope equipped with a dark-field oil condenser,a Schott UG1 excitation filter, and a Zeiss 41 stop filter.Fluorescent cells were photographed with Kodak Tri-Xblack-and-white film (Kodak).The sections were then processed essentially as described
by Hafen and Levine (28): the sections were air-dried at roomtemperature for 1-2 hr, immersed in l1o formalin in phos-phate-buffered saline (PBS: 10 mM phosphate, pH 7.2-7.4/0.8% NaCl) at room temperature for 20 min, rinsed threetimes in PBS, dehydrated in ethanol, air-dried, and storedovernight at 40C. To increase permeability, the sections wereincubated for 20 min at room temperature in 0.2 M HCl. Thesections were then rinsed for 5 min in distilled water,incubated in 0.3 M sodium chloride/0.03 M sodium citrate at70'C for 30 min, rinsed for 5 min in distilled water, and treatedwith Pronase (Calbiochem; 1.0 mg/ml in 50 mM Tris HClbuffer, pH 7.5, containing 5 mM EDTA). The Pronase waspreincubated at 370C for 3-4 hr to eliminate nuclease activity.After a 30-sec rinse in PBS containing glycine (2 mg/ml),followed by two rinses in PBS, the sections were fixed in 10%formalin in PBS for 20 min at room temperature and finallyrinsed in PBS and dehydrated in ethanol.
Preparation of Probes. Two TyrOHase probes, pTH1 (792base pairs) and pTH4 (400 base pairs) (22, 23) were used. ThepTH1 probe was subcloned into the pGEM-1 vector (Pro-mega Biotec, Madison, WI) after digestion with Pst I endo-nuclease (New England Biolabs) and purification of theplasmid and the insert by gel electrophoresis (29). Run-offtranscripts were prepared for T7 and SP6 transcriptionessentially according to the protocol from Promega Biotec,
Abbreviation: TyrOHase, tyrosine 3-hydroxylase.
6208
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 83 (1986) 6209
using the restriction enzymes HindIII and BamHI (PromegaBiotec). Transcripts were labeled with uridine [a-[35S]thio]-triphosphate (1000 Ci/mmol, New England Nuclear; 1 Ci =37 GBq) to reach a final activity of =4 x 107 cpm per ml ofhybridization fluid. These transcripts were then digested tofragments ranging from 50 to 150 bases and used for hybrid-ization histochemistry after removal of unincorporated ra-dioactivity centrifugation thru a Sephadex G-50 spin column(29).
Hybridization Procedure. The hybridization procedure wasessentially based on protocols by Hafen and Levine (28) andMoench et al. (30). In brief, hybridization was performed at30'C for 40-50 hr, using a hybridization buffer containing50% (vol/vol) deionized formamide (J. T. Baker Chemicals,Deventer, Netherlands), 0.6 M NaCl, 10 mM Tris HCl (pH7.5), 1 mM EDTA and lx Denhardt's solution (29). Eachsection was covered with 80 Al of hybridization fluid (3.2 x106 cpm) and a cover glass to diminish evaporation. The slideswere put in boxes humidified with 50% formamide and 0.6 MNaCl and were incubated as described above. After removalof the coverslips, the sections were washed for 30-40 hr at30'C in the above hybridization buffer, with five or sixchanges. The sections were then either rinsed in PBS orsubjected to RNase A digestion (20 Ag/ml in 500 mMNaCl/10 mM Tris HC1, pH 7.5) for 30 min at 370C followedby 30 min at 37°C in buffer without RNase (31). The sectionswere finally dehydrated in ethanol, air-dried, and coveredwith LKB Ultrofilm (LKB) or Fuji RX-G x-ray film (FujiPhotofilm, Tokyo, Japan) for autoradiography. After expo-sure for 1-2 days, the films were developed with LX24(Kodak) for 4 min and fixed with AL4 (Kodak). Thereafter,NTB2 nuclear track emulsion (Kodak; diluted 1:1 withdistilled water) was applied by the loop technique (32, 33).After exposure for 7-10 days at -20°C, the sections weredeveloped with D19 (Kodak) for 2 min and fixed with AL4 for
W-ia J--- 7_'Frn,>_
4 min. The sections were analyzed in a Leitz light micro-scope, using a dark-field condenser.Immunohistochemistry. After development, the slides were
processed for indirect immunohistochemistry, essentiallyaccording to Coons and collaborators (see ref. 9). In brief, thesections were incubated at 4°C in a humid atmosphere for 30min with a 1:400 dilution of antiserum to TyrOHase (34),rinsed in PBS, incubated with tetramethylrhodamine isothio-cyanate-conjugated donkey anti-rabbit antibodies (Amer-sham; dilution 1:20), rinsed in PBS, mounted in glycerol/PBS(3:1) containing 0.1% p-phenylenediamine (35, 36), andexamined in a Zeiss fluorescence microscope equipped withproper filter combinations. The cells were photographed withKodak Tri-X film.
RESULTS
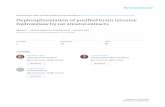
After hybridization of frontal sections of the mesencephalonand adjacent cortical areas with single-stranded RNA (fromprobe pTH1) complementary to TyrOHase mRNA, autora-diographs with varying activity were obtained. Thus, highactivity was generally seen in areas such as the zonacompacta of the substantia nigra and in the cellular layers ofthe hippocampal formation, and often there was high activityin many gray-matter areas of the mesencephalon as well as incerebral cortex (Fig. la). Treatment with RNase A reducedactivity, with the exception of the zona compacta of thesubstantia nigra (Fig. le). However, a weak but distinctactivity was always seen over the cell layers in the hip-pocampal formation (Fig. le). Hybridization of sections ofthe adrenal gland with the same probe gave results of similartype. Thus, before RNase A treatment it was often difficultto distinguish the difference between adrenal cortex andmedulla (Fig. lb), whereas enzyme treatment reduced activ-
FIG. 1. Micrographs of frontal section of rat brain at the mesencephalic (m) level (a, c, e, and g) and of rat adrenal gland (b, d, f, and h)after hybridization with a single-stranded RNA probe complementary to TyrOHase mRNA (a, b, e, andf) and a single-stranded RNA probeidentical to TyrOHase mRNA (c, d, g, and h). All autoradiographs in Figs. 1 and 2 are from sections hybridized with RNA obtained from thepTH1 probe. a and e, b andf, and c and g show, respectively, the same section. Micrographs a-d show x-ray film autoradiographs before RNaseA treatment, and e-h, after enzyme treatment. Note strong, widely distributed high activity before enzyme treatment, with particularly highintensity over hippocampal (HI) cell layers (arrowheads) with both types of probes (compare a and c), as well as a diffuse activity over the entireadrenal gland (b and d). After RNase treatment (e-h), activity is mainly seen over the zona compacta in the substantia nigra (open arrow ine) and over adrenal medulla (between arrowheads inf). Note, however, a distinct band of activity in hippocampus (arrowhead in e). No specificactivity is seen in the zona compacta or in the adrenal medulla after incubation with a sense probe (g and h). (Bar = 500 ,um; all micrographshave same magnification.)
Neurobiology: Schalling et al.

6210 Neurobiology: Schalling et al.
FIG. 2. X-ray-film autoradiographs (a and b), nuclear emulsion track autoradiographs (c andf), and fluorescence (e) and immunofluorescence(d, g, and h) micrographs showing frontal sections of ventral mesencephalon and adjacent cortical structures (a and b) and the ventralmesencephalon (c-h). a, f, and g are taken from the same section. b-e and h are taken from the same section of formalin/picric acid-fixed brain(shown in b), which received an intrastriatal Fast blue injection. (a and b) Distinct activity is seen over the zona compacta of the substantianigra (arrowheads in a and box in b). HI, hippocampus; m, mamillary body. (f and g) With nuclear track emulsion, an intense labeling can beseen over single cell bodies (f) that in almost all cases also contain TyrOHase-like immunoreactivity (g). Arrowheads point to TyrOHase-negative cells covered by grains, and double arrowheads point to TyrOHase-positive cell lacking grains. (c-e and h) Area with high activityindicated by rectangle in b is shown to contain numerous TyrOHase-positive cell bodies with a similar distribution (h). Arrowheads in b andh point to densely packed group of TyrOHase-positive cells covered by large number of grains. Rectangle in h approximately indicates the areashown in d. These TyrOHase-positive cells are covered by grains (c) and several ofthem also contain retrogradely transported Fast blue (arrowsin c-e). Asterisks indicate the same blood vessels. (Bar in a = 500 ,um, bars in e andf= 50 pum. a and b, c-e and h, andfand g have, respectively,the same magnification.)
ity over the cortex, leaving a strong activity only over themedulla (Fig. 1f).
Hybridization with a probe of the sense mRNA revealedsimilar results as the ones described for the complementary
RNA probe, with high activity over the entire gray matter,including the zona compacta of the substantia nigra, thecellular layers in the hippocampal region, and the cortex (Fig.1c), as well as diffuse activity over the entire adrenal gland
Proc. Natl. Acad. Sci. USA 83 (1986)

Proc. Natl. Acad. Sci. USA 83 (1986) 6211
(Fig. 1d). After treatment with RNase A, a loss of activitywas seen over the entire section of the brain, including thezona compacta (Fig. ig). However, a band of reducedactivity remained over the hippocampal cell layers andcortical areas (Fig. ig), and in the adrenal gland no distinctionbetween medulla and cortex could be made (Fig. lh).Of the fixatives used, formalin (Fig. 2a) and formalin
combined with picric acid (Fig. 2 b andf), the former solutiongave the most consistent result and the most distinct labeling(compare Fig. 2 a and b). Addition of glutaraldehyde resultedin somewhat higher background activity.
After application of the nuclear track emulsion, a stronglabeling was seen over cell bodies in the zona compacta andventral tegmental area (Fig. 2 c andf). Processing for indirectimmunofluorescence with TyrOHase antiserum revealed adistinct labeling of cell bodies in these areas (Fig. 2 d, g, andh), and there was an almost complete correlation betweengrain-labeled cells and TyrOHase-positive cells (compareFig. 2 c with d, and f with g). However, single TyrOHasemRNA-positive cells lacked the enzyme, and there weresome TyrOHase-positive cells that were not covered bygrains (Fig. 2 f and g). After injection of Fast blue into thecorpus of the caudate nucleus, numerous blue fluorescentcells were observed in the ventral mesencephalon (Fig. 2e).Many of these cells contained TyrOHase-like immunoreac-tivity (Fig. 2d) and were covered by grains indicating thepresence of TyrOHase mRNA (Fig. 2c).None of the TyrOHase-positive structures described were
observed after incubation with control serum.
DISCUSSIONThe present results show that hybridization histochemistryusing an RNA probe complementary to TyrOHase mRNArepresents a potentially valuable tool in the further analysisof central and peripheral catecholamine cell systems, com-plementary to the formaldehyde-induced-fluorescence tech-nique (37) and immunohistochemistry (38). Thus, the presentresults demonstrate localization of TyrOHase mRNA in cellbodies in the zona compacta of the substantia nigra, as wellas in the ventral tegmental area (cell groups A9 and A10, ref.6). After RNase A treatment, this activity was not seen whenthe sense mRNA probe was used for hybridization. This wasshown both with x-ray film and with liquid nuclear trackemulsion, the latter providing resolution at the cellular level.The identity of these cells was established by combining insitu hybridization with immunohistochemistry using antise-rum to TyrOHase on the same sections. The detailed analysisrevealed some cells containing TyrOHase immunoreactivitybut apparently lacking TyrOHase mRNA, as well as someTyrOHase mRNA-positive cells apparently devoid ofTyrOHase immunoreactivity. The number of such single-labeled cells was very small and may represent artifacts dueto, for example, lack of penetration of the probe or antibody,respectively. It could also reflect low levels of mRNA orenzyme, respectively, perhaps related to differences in turn-over. Furthermore, the processing during the hybridizationprocedure may destroy TyrOHase antigenicity.The present findings support earlier demonstrations that
treatment with RNase A represents a valuable tool in reduc-ing nonspecific background (17, 31), but some activity re-mained, mainly over the hippocampal cell layers and to acertain extent in cortical areas. This was seen also afterhybridization with the sense mRNA probe. This suggests thatthe activity in the hippocampal layers represents binding ofthe complementary mRNA probe with partially homologoussequences not related to catecholamine cell systems (39, 40)or nonspecific labeling in cell layers with unusually highcellular density.
Of the three fixatives tested, formalin (26) gave the bestresults with regard to hybridization histochemistry. Thepicric acid-containing fixative (27) often provided betterresults in immunohistochemistry, whereas with the glutaral-dehyde-containing fixative, there was a tendency for higherbackground.The potential of hybridization histochemistry as a tool for
mapping neuronal systems is clearly reduced by the fact thatthe marker (mRNA) is confined to cell bodies and cannot befound in axons or terminal ramifications. The present studyshows that this can be overcome in part by combininghybridization histochemistry with retrograde tracing usingfluorescent compounds (see also ref. 41). Thus, after injec-tion of Fast blue into the caudate nucleus, many nigralTyrOHase mRNA-positive cells also contained this retro-grade tracer, providing evidence that these neurons project tothe caudate nucleus. With the additional detection ofTyrOHase-like immunoreactivity in these cells, it is thuspossible to demonstrate the occurrence of three markers-the retrograde tracer, the dopamine-synthesizing enzymeTyrOHase, and the mRNA for this enzyme-in the samesection.
We thank Professor J. Mallet (Laboratory of Cellular Neurobiol-ogy, Gif-sur-Yvette, France) for supplying pTH1, which was used forall illustrations in this paper. We thank Dr. D. M. Chikaraishi(Department of Neurology, Neuroscience Laboratories, Tufts Uni-versity, Boston, MA) for supplying pTH4, for reading the manu-script, and for valuable advice. We thank Dr. R. Olsson (Departmentof Chemistry, Oslo University, Oslo, Norway) and Dr. E. Egyhazi(Department of Histology, Karolinska Institutet, Stockholm, Swe-den) for support. We thank Dr. J. Dixon (Purdue University, WestLafayette, IN) for valuable advice. We thank Ms. W. Hiort, Ms. S.Nilsson, Ms. A. Peters, and Ms. S. Soltesz-Mattisson for valuabletechnical assistance and Ms. E. Bjorklund for expert secretarial help.This study was supported by the Swedish Medical Research Council(04X-2887), Magnus Bergvalls Stiftelse, Petrus och AugustaHedlunds Stiftelse, and Fredrik och Ingrid Thurings Stiftelse, as wellas by Grant 06801-19 from the National Institute of Neurological andCommunicative Disorders and Stroke, by Grant 02717-28 from theNational Institute of Mental Health, and by a grant from the MentalHealth Clinical Research Center.
1. Eranko, 0. (1952) Acta Anat. 16, Suppl. 17, 1-60.2. Eranko, 0. (1953) Endocrinology 57, 363-368.3. Hillarp, N.-A. & Hckfelt, B. (1953) Acta Physiol. Scand. 30,
55-68.4. Carlsson, A., Falck, B. & Hillarp, N.-A. (1962) Acta Physiol.
Scand. 56, Suppl. 196, 1-27.5. Falck, B. (1962) Acta Physiol. Scand. 56, Suppl. 197, 1-25.6. Dahlstrom, A. & Fuxe, K. (1964) Acta Physiol. Scand. 62,
Suppl. 232, 1-55.7. Falck, B., Hillarp, N.-A., Thieme, G. & Torp, A. (1962) J.
Histochem. Cytochem. 10, 348-354.8. Geffen, L. B., Livett, D. G. & Rush, R. A. (1969) J. Physiol.
(London) 204, 593-605.9. Coons, A. H. (1958) in General Cytochemical Methods, ed.
Danielli, J. F. (Academic, New York), pp. 394-422.10. Gall, J. G. & Pardue, M. L. (1971) Methods Enzymol. 38,
470-480.11. Brahic, M. & Haase, A. T. (1978) Proc. Natl. Acad. Sci. USA
75, 6125-6129.12. Gee, C. E. & Roberts, J. L. (1983) DNA 2, 157-163.13. Hudson, P., Penshow, J., Shine, J., Ryan, G., Niall, H. &
Coghlan, J. P. (1981) Endocrinology 108, 353-356.14. Gee, C. E., Chen, C.-L. C. & Roberts, J. L. (1983) Nature
(London) 306, 374-376.15. Bloch, B., Le Guellec, D. & De Keyzer, Y. (1985) Neurosci.
Lett. 53, 141-148.16. Nojiri, H., Sato, M. & Urano, A. (1985) Neurosci. Lett. 58,
101-105.17. Siegel, R. E. & Young, W. S., III (1985) Neuropeptides 6,
573-580.18. Wolfson, B., Manning, R. W., David, L. G., Arentzen, R. &
Baldino, F., Jr. (1985) Nature (London) 315, 59-61.
Neurobiology: Schaffing et al.

6212 Neurobiology: Schalling et al.
19. Uhl, G. R., Zingg, H. H. & Habener, J. F. (1985) Proc. Natl.Acad. Sci. USA 82, 5555-5559.
20. Shivers, B. D., Harlan, R. E., Pfaff, D. W. & Schachter, B. S.(1986) J. Histochem. Cytochem. 34, 39-43.
21. McCabe, J. T., Morrell, J. I., Ivell, R., Schmale, H., Richter,D. & Pfaff, D. W. (1986) J. Histochem. Cytochem. 34, 45-50.
22. Lamouroux, A., Faucon Biguet, N., Samolyk, D., Privat, A.,Salomon, J. C., Pujol, J. F. & Mallet, J. (1982) Proc. Natl.Acad. Sci. USA4 79, 3881-3885.
23. Lewis, E. J., Tank, A. W., Weiner, N. & Chikaraishi, D. M.(1983) J. Biol. Chem. 258, 14632-14637.
24. Skirboll, L., Hdkfelt, T., Norell, G., Phillipson, O., Kuypers,H. G. J. M., Bentivoglio, M., Catsman-Berrevoets, C. E.,Visser, T. J., Steinbusch, H., Verhofstad, A., Cuello, A. C.,Goldstein, M. & Brownstein, M. (1984) Brain Res. Rev. 8,99-127.
25. Bentivoglio, M., Kuypers, H. G. J. M., Catsman-Berrevoets,C. E., Loewe, H. & Dann, 0. (1980) Neurosci. Lett. 18, 25-30.
26. Pease, P. C. (1962) Anat. Rec. 142, 342 (abstr.).27. Zamboni, L. & de Martino, C. (1967) J. Cell Biol. 148A, 35
(abstr.).28. Hafen, E. & Levine, M. (1983) EMBO J. 2, 617-623.29. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular
Cloning: A Laboratory Manual (Cold Spring Harbor Labora-tory, Cold Spring Harbor, NY).
30. Moench, T. R., Gendelman, H. E., Clements, J. E., Narayan,
Proc. Natl. Acad. Sci. USA 83 (1986)
0. & Griffin, D. E. (1985) J. Virol. Methods 11, 119-130.31. McGowan, R. A., Kane-Haas, C. & Gross, K. W. (1985) N.
Engl. Nuclear Prod. News 4, 1-8.32. Miller, 0. L., Jr., Stone, G. E. & Prescott, D. M. (1964) J.
Cell Biol. 23, 654-658.33. Nagata, T. & Nawa, T. (1966) Histochemie 7, 370-371.34. Markey, K. A., Kondo, S., Shenkman, L. & Goldstein, M.
(1980) Mol. Pharmacol. 17, 79-85.35. Johnson, D. G. & Nogueira Araujo, G. M. de C. (1981) J.
Immunol. Methods 43, 349.36. Platt, J. L. & Michael, A. F. (1983) J. Histochem. Cytochem.
31, 840-842.37. Bjorklund, A. (1983) in Handbook of Chemical Neuroanat-
omy: Methods in Chemical Neuroanatomy, eds. Bjorklund, A.& Hokfelt, T. (Elsevier, Amsterdam), Vol. 1, pp. 50-121.
38. Hokfelt, T., Johansson, 0. & Goldstein, M. (1984) in Hand-book of Chemical Neuroanatomy: Classical Transmitters inthe CNS, eds. Bjorklund, A. & Hbkfelt, T. (Elsevier, Amster-dam), Vol. 2, Part 1, pp. 157-276.
39. Lewis, E. J., Delano, M. L. & Aldrich, W. R. (1985) Neuro-science 11, abstr. 316.3.
40. Mahalic, T. J., Chaudhari, N., Hahn, W. E. & Finger, T. E.(1985) Neuroscience 11, abstr. 316.1.
41. Wilcox, J. N., Chronwall, B. M., O'Donohue, T. L. &Roberts, J. L. (1985) Neuroscience 11, abstr. 46.9.