
Transcriptome of Swarming Proteus mirabilis
12
INFECTION AND IMMUNITY, June 2010, p. 2834–2845 Vol. 78, No. 6 0019-9567/10/$12.00 doi:10.1128/IAI.01222-09 Copyright © 2010, American Society for Microbiology. All Rights Reserved. Transcriptome of Swarming Proteus mirabilis † Melanie M. Pearson, 1 David A. Rasko, 2 Sara N. Smith, 1 and Harry L. T. Mobley 1 * Department of Microbiology and Immunology, University of Michigan Medical School, Ann Arbor, Michigan, 1 and Institute for Genome Sciences, University of Maryland School of Medicine, Baltimore, Maryland 2 Received 28 October 2009/Returned for modification 14 December 2009/Accepted 28 March 2010 Swarming motility by the urinary tract pathogen Proteus mirabilis has been a long-studied but little under- stood phenomenon. On agar, a P. mirabilis colony grows outward in a bull’s-eye pattern formed by consecutive waves of rapid swarming followed by consolidation into shorter cells. To examine differential gene expression in these growth phases, a microarray was constructed based on the completed genome sequence and annota- tion. RNA was extracted from broth-cultured, swarming, and consolidation-phase cells to assess transcription during each of these growth states. A total of 587 genes were differentially expressed in broth-cultured cells versus swarming cells, and 527 genes were differentially expressed in broth-cultured cells versus consolidation- phase cells (consolidate). Flagellar genes were highly upregulated in both swarming cells and consolidation- phase cells. Fimbriae were downregulated in swarming cells, while genes involved in cell division and anaerobic growth were upregulated in broth-cultured cells. Direct comparison of swarming cells to consolidation-phase cells found that 541 genes were upregulated in consolidate, but only nine genes were upregulated in swarm cells. Genes involved in flagellar biosynthesis, oligopeptide transport, amino acid import and metabolism, cell division, and phage were upregulated in consolidate. Mutation of dppA, oppB, and cysJ, upregulated during consolidation compared to during swarming, revealed that although these genes play a minor role in swarming, dppA and cysJ are required during ascending urinary tract infection. Swarming on agar to which chloram- phenicol had been added suggested that protein synthesis is not required for swarming. These data suggest that the consolidation phase is a state in which P. mirabilis prepares for the next wave of swarming. Proteus mirabilis, a member of the Enterobacteriaceae and an opportunistic pathogen, is especially problematic as a urinary tract pathogen in catheterized patients, those with spinal cord injury, or those with anatomical abnormality of the urinary tract (reviewed in reference 16). This urease-positive bacte- rium causes an increase in urinary pH and the production of kidney and bladder stones (25, 39). In addition, urinary cath- eters become encrusted and even blocked during P. mirabilis infections (45). As our population continues to age, resulting in larger numbers of catheterized patients in hospitals and nurs- ing homes, this organism will likely be of special concern as an agent of nosocomial infections. Particularly worrisome is the more frequent appearance of multidrug-resistant strains of P. mirabilis (17, 47). P. mirabilis was first characterized in 1885 by G. Hauser for its ability to swarm over agar surfaces, resulting in a charac- teristic bull’s-eye pattern (reviewed in reference 65). During the swarm process, P. mirabilis differentiates into very long (50 m), multinucleate, highly motile hyperflagellated cells (65). At intervals, swarm cells slow down or cease movement and dedifferentiate into shorter rod-shaped cells in what is known as the consolidation phase. Repeated cycles of swarm- ing and consolidation lead to the bull’s-eye pattern. Reflecting this phenotype, P. mirabilis was named for the Greek god Proteus, who was able to change form at will to avoid ques- tioning (65). Flagellum-mediated swarming, accompanied by cell elongation, has been observed in other bacterial species, including Escherichia coli, Salmonella enterica serovar Typhi- murium, Serratia marcescens, Vibrio parahaemolyticus, and Pseudomonas aeruginosa (reviewed in references 22, 28, and 61). However, it should be noted that swarming by these spe- cies requires a much lower agar concentration and lacks the characteristic cyclic pattern of swarming and consolidation. Thus, swarming by these other species is analogous but not identical to the robust swarming activity of P. mirabilis. Numerous studies have been conducted in an effort to un- derstand how and why P. mirabilis swarms, yet despite some advances, much remains unknown. However, it has been de- finitively demonstrated that flagella (2, 8) and chemotaxis (12) are necessary for swarming. Polysaccharides (including lipo- polysaccharides [LPS]) (10, 26), extracellular matrix compo- nents (57), and fatty acids (40) may also play roles in helping P. mirabilis to move across solid surfaces. Putrescine (59) and glutamine (3) represent signals involved in the initiation of swarming. Cell density may also play a role in the transition between swarming and consolidation phases (53) and in the initiation of swarming (11); however, a luxS mutant of P. mira- bilis swarms normally (56). Mutagenesis and library screens have identified several regulators of swarming motility, includ- ing rsbA (11), ccmA (31), umoA, umoB, umoC, umoD (20), wosA (30), disA (58), and the Lon protease (14). Multiple virulence factors are also upregulated during swarming, includ- ing hemolysin, urease, and the Zap metalloprotease (4, 62). In the clinical setting, swarming may present an issue for patients with indwelling urinary catheters. P. mirabilis swarms across the surface of both latex and silicone urinary catheters (35, 54), thus gaining access to the bladder. Rapid colonization * Corresponding author. Mailing address: 5641 Medical Science Bldg. II, 1150 W. Medical Center Dr., Ann Arbor, MI 48109-0620. Phone: (734) 764-1466. Fax: (734) 763-7163. E-mail: hmobley@umich .edu. † Supplemental material for this article may be found at http://iai .asm.org/. Published ahead of print on 5 April 2010. 2834 Downloaded from https://journals.asm.org/journal/iai on 08 February 2022 by 208.107.36.47.
Transcript of Transcriptome of Swarming Proteus mirabilis

INFECTION AND IMMUNITY, June 2010, p. 2834–2845 Vol. 78, No. 60019-9567/10/$12.00 doi:10.1128/IAI.01222-09Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Transcriptome of Swarming Proteus mirabilis�†Melanie M. Pearson,1 David A. Rasko,2 Sara N. Smith,1 and Harry L. T. Mobley1*
Department of Microbiology and Immunology, University of Michigan Medical School, Ann Arbor, Michigan,1 and Institute forGenome Sciences, University of Maryland School of Medicine, Baltimore, Maryland2
Received 28 October 2009/Returned for modification 14 December 2009/Accepted 28 March 2010
Swarming motility by the urinary tract pathogen Proteus mirabilis has been a long-studied but little under-stood phenomenon. On agar, a P. mirabilis colony grows outward in a bull’s-eye pattern formed by consecutivewaves of rapid swarming followed by consolidation into shorter cells. To examine differential gene expressionin these growth phases, a microarray was constructed based on the completed genome sequence and annota-tion. RNA was extracted from broth-cultured, swarming, and consolidation-phase cells to assess transcriptionduring each of these growth states. A total of 587 genes were differentially expressed in broth-cultured cellsversus swarming cells, and 527 genes were differentially expressed in broth-cultured cells versus consolidation-phase cells (consolidate). Flagellar genes were highly upregulated in both swarming cells and consolidation-phase cells. Fimbriae were downregulated in swarming cells, while genes involved in cell division and anaerobicgrowth were upregulated in broth-cultured cells. Direct comparison of swarming cells to consolidation-phasecells found that 541 genes were upregulated in consolidate, but only nine genes were upregulated in swarmcells. Genes involved in flagellar biosynthesis, oligopeptide transport, amino acid import and metabolism, celldivision, and phage were upregulated in consolidate. Mutation of dppA, oppB, and cysJ, upregulated duringconsolidation compared to during swarming, revealed that although these genes play a minor role in swarming,dppA and cysJ are required during ascending urinary tract infection. Swarming on agar to which chloram-phenicol had been added suggested that protein synthesis is not required for swarming. These data suggest thatthe consolidation phase is a state in which P. mirabilis prepares for the next wave of swarming.
Proteus mirabilis, a member of the Enterobacteriaceae and anopportunistic pathogen, is especially problematic as a urinarytract pathogen in catheterized patients, those with spinal cordinjury, or those with anatomical abnormality of the urinarytract (reviewed in reference 16). This urease-positive bacte-rium causes an increase in urinary pH and the production ofkidney and bladder stones (25, 39). In addition, urinary cath-eters become encrusted and even blocked during P. mirabilisinfections (45). As our population continues to age, resulting inlarger numbers of catheterized patients in hospitals and nurs-ing homes, this organism will likely be of special concern as anagent of nosocomial infections. Particularly worrisome is themore frequent appearance of multidrug-resistant strains of P.mirabilis (17, 47).
P. mirabilis was first characterized in 1885 by G. Hauser forits ability to swarm over agar surfaces, resulting in a charac-teristic bull’s-eye pattern (reviewed in reference 65). Duringthe swarm process, P. mirabilis differentiates into very long(�50 �m), multinucleate, highly motile hyperflagellated cells(65). At intervals, swarm cells slow down or cease movementand dedifferentiate into shorter rod-shaped cells in what isknown as the consolidation phase. Repeated cycles of swarm-ing and consolidation lead to the bull’s-eye pattern. Reflectingthis phenotype, P. mirabilis was named for the Greek godProteus, who was able to change form at will to avoid ques-
tioning (65). Flagellum-mediated swarming, accompanied bycell elongation, has been observed in other bacterial species,including Escherichia coli, Salmonella enterica serovar Typhi-murium, Serratia marcescens, Vibrio parahaemolyticus, andPseudomonas aeruginosa (reviewed in references 22, 28, and61). However, it should be noted that swarming by these spe-cies requires a much lower agar concentration and lacks thecharacteristic cyclic pattern of swarming and consolidation.Thus, swarming by these other species is analogous but notidentical to the robust swarming activity of P. mirabilis.
Numerous studies have been conducted in an effort to un-derstand how and why P. mirabilis swarms, yet despite someadvances, much remains unknown. However, it has been de-finitively demonstrated that flagella (2, 8) and chemotaxis (12)are necessary for swarming. Polysaccharides (including lipo-polysaccharides [LPS]) (10, 26), extracellular matrix compo-nents (57), and fatty acids (40) may also play roles in helping P.mirabilis to move across solid surfaces. Putrescine (59) andglutamine (3) represent signals involved in the initiation ofswarming. Cell density may also play a role in the transitionbetween swarming and consolidation phases (53) and in theinitiation of swarming (11); however, a luxS mutant of P. mira-bilis swarms normally (56). Mutagenesis and library screenshave identified several regulators of swarming motility, includ-ing rsbA (11), ccmA (31), umoA, umoB, umoC, umoD (20),wosA (30), disA (58), and the Lon protease (14). Multiplevirulence factors are also upregulated during swarming, includ-ing hemolysin, urease, and the Zap metalloprotease (4, 62).
In the clinical setting, swarming may present an issue forpatients with indwelling urinary catheters. P. mirabilis swarmsacross the surface of both latex and silicone urinary catheters(35, 54), thus gaining access to the bladder. Rapid colonization
* Corresponding author. Mailing address: 5641 Medical ScienceBldg. II, 1150 W. Medical Center Dr., Ann Arbor, MI 48109-0620.Phone: (734) 764-1466. Fax: (734) 763-7163. E-mail: [email protected].
† Supplemental material for this article may be found at http://iai.asm.org/.
� Published ahead of print on 5 April 2010.
2834
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

of catheter surfaces, coupled with catheter encrustation result-ing from urease production, makes P. mirabilis a particularlytroublesome pathogen in the hospital setting.
The recent sequencing and annotation of the P. mirabilisHI4320 genome (52) allowed for the construction of a P. mira-bilis microarray. In this report, the first use of microarrays tostudy P. mirabilis gene expression, we investigated the tran-scriptome of this organism during its swarming and consolida-tion phases and contrasted gene expression during thesephases with expression during logarithmic-phase broth culture.Both swarming and consolidation phases were monitored inreal time. Only the edges of colonies in swarming or consoli-dation phases, confirmed by both Gram staining and micro-scopic observation of motility in intact colonies, were collectedso that only the youngest cells of the colony were analyzed.Selected genes identified by microarray analysis were mutatedand assessed for their effect on swarm behavior and virulencein a mouse model of ascending urinary tract infection. Thenecessity of active protein synthesis during swarming was alsoinvestigated.
MATERIALS AND METHODS
Bacterial strains and culture conditions. P. mirabilis HI4320 was isolated fromthe urine of an elderly, long-term catheterized woman (45). E. coli DH5� wasused as the host strain for cloning of mutagenesis constructs. All strains andmutants were cultured at 37°C with aeration in nonswarm LB broth (1% tryp-tone, 0.5% yeast extract, 0.05% NaCl) or on LB medium solidified with 1.5%agar. Media were supplemented with chloramphenicol (20 �g/ml), ampicillin (50�g/ml), or kanamycin (25 �g/ml) as necessary.
Swarm agar and collection of swarming P. mirabilis bacteria. Swarming mo-tility was assessed by spotting 5 �l late-logarithmic-phase bacterial culture (op-tical density at 600 nm [OD600] of 1.0) onto the center of an LB swarm plate (1%tryptone, 0.5% yeast extract, 1% NaCl, 1.5% agar) followed by overnight incu-bation at 30°C. P. mirabilis was assessed for swarm or consolidation behavior bymorphology of the colony edge, motility (using an inverted microscope), and byGram staining. In three representative experiments, the lengths of bacteria fromswarm and consolidation-phase samples were measured. In all three experi-ments, more than 99% of the consolidation-phase cells (consolidate) were lessthan 10 �m in length, and 88 to 97% of the swarming-phase cells were greaterthan 10 �m in length. Bacteria from the outermost edge of a swarming orconsolidating colony were scraped with a plastic loop into 1.5 ml RNA Protect(Qiagen); the colony perimeter from four 7.5-cm diameter plates was scraped foreach RNA sample. P. mirabilis was also cultured to mid-logarithmic phase(OD600 � 0.8) in LB broth at 30°C; 1 ml of culture was added to 2 ml RNAProtect. RNA was isolated using the RNeasy kit (Qiagen) according to themanufacturer’s instructions, except that 3 mg/ml lysozyme was added to cells for15 min. DNA was digested using Turbo DNAfree DNase (Ambion).
P. mirabilis microarray. A preliminary annotation of the incomplete HI4320genome located 3,722 potential open reading frames (ORFs) in the chromo-some. This annotation was used to design unique 70-mer oligonucleotide probes(Operon Technologies, Inc.) corresponding to each ORF. The completed ge-nome has 3,693 annotated ORFs; thus, the microarray contains probes for someregions that are now annotated as pseudogenes or were incorporated into neigh-boring ORFs. Our analysis accounts for these minor modifications. This microar-ray lacks probes corresponding to the genes in the 36-kb plasmid that is presentin P. mirabilis HI4320. Oligonucleotide probes were spotted in triplicate ontoUltraGaps II glass slides (Corning) by Microarrays, Inc. RNA (4 �g), obtainedfrom broth-cultured, swarm, or consolidation-phase P. mirabilis, was used as atemplate for cDNA synthesis and was labeled with either cyanine 3 or cyanine 5dye (GE Healthcare) according to standard operating procedure (SOP) no.M007 from The Institute for Genomic Research (TIGR) (ftp://ftp.jcvi.org/pub/data/PFGRC/pdf_files/protocols/M007.pdf). Labeled cDNA was mixed and hy-bridized to an array slide by using TIGR SOP no. M008 (ftp://ftp.jcvi.org/pub/data/PFGRC/pdf_files/protocols/M008.pdf). The microarray was scanned using aScanArray Express microarray scanner (Perkin Elmer) at 10-�m resolution andquantified using the ScanArray Express software. Five independent arrays wereeach analyzed for broth-cultured versus consolidate or swarm bacteria. Three
independent arrays were analyzed for the direct comparison of consolidate toswarm bacteria.
Microarray analysis. The total normalization algorithm (ScanArray Express;Perkin Elmer) was used to normalize microarray data. Median spot values foreach triplicate probe were extracted using MIDAS (TIGR). For the arrayscomparing broth-cultured P. mirabilis to swarm or consolidate, a gene was con-sidered differentially regulated if it was greater than 2-fold changed in four ormore of the arrays and with the median being greater than a 2-fold change. Forthe arrays directly comparing consolidate to swarm, genes were considereddifferentially regulated if they were up- or downregulated �2-fold in two of thethree datasets. The complete microarray data sets can be accessed at GEOdatabase number GSE17957.
qRT-PCR. RNA from broth-cultured, swarming, or consolidating P. mirabilisbacteria was converted into cDNA by using the Superscript first-strand synthesissystem (Invitrogen) according to the manufacturer’s protocol. PCR with primersspecific to the rpoA (RNA polymerase A) gene was performed on cDNA samplesprepared with and without reverse transcriptase to confirm lack of genomic DNAcontamination of the RNA preparations. Each quantitative reverse transcriptasePCR (qRT-PCR) was set up in duplicate and consisted of 30 ng cDNA template,150 nM each primer, and 12.5 �l 2� SYBR green PCR master mix (Stratagene).Target genes were amplified using an Mx3000P thermal cycler (Stratagene).Melting curve analysis was used to confirm a lack of primer dimers or genomicDNA contamination of reagents. Data were normalized to rpoA, and analyzed bythe threshold cycle (2���CT) method (42). Four independently isolated cDNAsamples were analyzed. Primer sequences are listed in Table S1 in the supple-mental material.
Mutant construction. A kanamycin resistance gene was inserted into dppA(PMI2847), oppB (PMI1474), cysJ (PMI2250), lrhA (PMI0629), and hexA(PMI1764) by using the TargeTron gene knockout system (Sigma) with a mod-ification previously described for P. mirabilis (51). Briefly, a group II intron wasreprogrammed according to the manufacturer’s protocol to specifically insertinto the target gene by mutagenic PCR. The retargeted intron was ligated intoplasmid pACD4K-C or pACD4K-CloxP to create plasmids pMP206, pMP207,pMP208, pMP196, and pMP197 for dppA, oppB, cysJ, lrhA, and hexA, respec-tively. To construct the lrhA hexA double mutant, the lrhA mutant, HI4320 lrhA�kan, was electroporated with pQL123 (41), which encodes an IPTG (isopropyl--D-thiogalactopyranoside)-inducible cre recombinase. IPTG induction led tothe removal of the kanamycin resistance cassette from the intron in lrhA, creatingthe mutant HI4320 lrhA�kan. This mutant was then electroporated with pMP197to the construct HI4320 lrhA�kan hexA�kan. For all mutants, growth in LB wasmeasured using a Bioscreen C growth curve analyzer (Growth Curves Ltd.).Mutants were screened for swarming ability on LB swarm agar or on minimal Aagar (9) containing glucose as a carbon source and supplemented with 1%tryptone (MinA-T). Swarm radii were measured 16.0 h after inoculation in fiveindependent experiments. Statistical significance was evaluated using the two-tailed paired t test.
Mouse model of ascending UTI. A modification (34) of the CBA/J mousemodel of ascending urinary tract infection (UTI) has been described previously(27, 38). Briefly, overnight cultures of wild-type P. mirabilis HI4230 and theisogenic mutant strain (dppA, oppB, cysJ, lrhA, or hexA) were individually ad-justed to an estimated density of 2 � 108 CFU/ml (OD600 � 0.2). Cultures ofmutants were each mixed in a 1:1 ratio with the wild-type parent strain. Six- to8-week-old female CBA/J mice were inoculated transurethrally with a 50-�lsuspension of the mixture (i.e., 1 � 107 CFU) via a sterile polyethylene catheter,using an infusion pump (Harvard Apparatus). At 7 days postinoculation, thebladders, kidneys, and spleens of these mice were homogenized in 3 ml phos-phate-buffered saline (PBS) by using an OMNI mechanical homogenizer (OMNIInternational) and plated onto LB agar or LB agar supplemented with kanamy-cin by using a spiral plater (Autoplate 4000; Spiral Biotech) to assess bacterialburden; urine was diluted in PBS and also plated. Colony counts were enumer-ated using a Qcount (Spiral Biotech). Wild-type HI4320 infection was deter-mined by subtracting the number of colonies on the kanamycin plate from thenumber of colonies on the plain LB plate. Statistical significance was assessedusing Wilcoxon’s matched-pairs test. The University of Michigan University Com-mittee on Use and Care of Animals approved all mouse protocols.
Swarming on agar containing antibiotic discs. Chloramphenicol or kanamycinwas added to 6-mm-diameter sterile blank paper discs (Becton, Dickinson andCompany) in specified amounts and allowed to dry. Swarm plates were inocu-lated with P. mirabilis HI4320 as described above, and the antibiotic discs wereplaced approximately 1 cm from the inoculation point. As a control, 50 �l of P.mirabilis HI4320 at an OD600 of 1.0 was spread plated onto nonswarm LB agar(0.5 g/liter NaCl) to create a lawn. Antibiotic discs were placed on the plate todetermine zones of inhibition. Plates were incubated at 30°C overnight.
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2835
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

RESULTS
P. mirabilis swarm and consolidate. When cultured onswarm agar, P. mirabilis alternates with regular periodicity be-tween an elongated, hyperflagellated “swarmer” form anda shorter “consolidation” form, resulting in a characteristicbull’s-eye pattern (Fig. 1A). The swarm and consolidationphases can be distinguished as they are forming at the colonyedge; consolidating colonies have a ragged edge (Fig. 1B) andhave little visible motion when examined microscopically(�50), while actively swarming colonies have a smooth edge(Fig. 1C) and are in vigorous motion (see Fig. S1 in the sup-plemental material). Gram stains of consolidate or swarm fromthe colony edge show the distinct cell morphologies corre-sponding to these well-defined phases (Fig. 1D and E). Usinga cell length of greater than 10 �m to define swarm cells (14,33, 44), more than 99% (298/300) of the bacteria shown in Fig.1D were in the consolidation phase, while 88% (120/137) of thecells shown in Fig. 1E had the swarmer morphology.
Microarray analysis of broth-cultured P. mirabilis andswarm or consolidate. The transcriptome of broth-cultured P.mirabilis HI4320 was compared to consolidation- or swarming-phase P. mirabilis HI4320. For all agar-cultured samples, cellmorphology was confirmed as swarm or consolidate by Gramstaining. Bacteria for both swarm and consolidation sampleswere collected from the leading edge of the colony. Analysis of
data from five arrays indicated that 587 genes (16% of thegenome) were differentially expressed in broth-grown cells ver-sus swarm cells, and 527 genes (14%) were differentially ex-pressed in broth-grown cells versus consolidate. Most genes(470 of 644 [73%]) were found to be regulated similarly inswarm or consolidate cells compared to broth-grown cells (Fig.2); that is, genes that were upregulated in consolidate com-pared to broth-cultured cells (Table 1; see also Table S2 in thesupplemental material) also were likely to be upregulated inswarmer cells compared to broth-cultured cells (Table 2; seealso Table S3 in the supplemental material). Genes that wereupregulated on swarm agar compared to broth included nu-merous genes previously correlated with swarming behavior,such as flagella, the Zap metalloprotease, and hemolysin (4,62). In fact, flaA (PMI1620), encoding flagellin, the majorsubunit of flagella, was among the most highly expressed genesin both consolidate (sixth) and swarm (third) cells. Fimbrialgenes were upregulated in broth compared to either swarm orconsolidate. See Tables S2, S3, S4, and S5 in the supplementalmaterial for the complete list of genes differentially regulatedat least 2-fold in broth compared to consolidate or swarm. SeeTables S6, S7, and S8 in the supplemental material for lists ofthe most highly expressed genes in consolidate, swarm, andbroth-cultured cells, respectively.
Microarray analysis of swarm compared to consolidate.When the consolidate morphotype was compared directly tothat of swarm, 541 genes were found to have transcriptionincreased �2-fold in consolidate (see Table S9 in the supple-mental material; the 50 most upregulated genes are listed inTable 3). Only nine genes had increased transcription in swarmcells compared to consolidate cells (Table 4).
Genes encoding proteins involved in peptide uptake andamino acid uptake and biosynthesis were highly upregulated inconsolidate compared to swarm. Genes encoding stress pro-teins, as well as the general stress response sigma factor RpoS,were also upregulated during consolidation, while three genesencoding cold shock proteins were upregulated during swarm-ing. Numerous proteases, including Lon, which has previously
FIG. 1. (A) P. mirabilis swarm plate, with sample collection timesindicated for microarray analysis. The edge of a newly forming con-solidation ring forms finger-like projections (B) while the edge of anactive swarm front is smooth (C). When bacteria are visualized byGram staining from the edge of a consolidating colony (D), vegetativecells are found, while bacteria isolated from an active swarm edgedisplay the elongated swarm morphology (E). In panel D, more than99% of the bacteria are less than 10 �m in length, while in panel E,88% of the bacteria are greater than 10 �m in length. The photographsin panels B and C were taken at �50 magnification, and the photo-graphs in panels D and E were taken at �400 magnification. The sizebar in panels D and E indicates 50 �m.
FIG. 2. Venn diagrams showing the numbers of differentially reg-ulated genes during broth culture compared to consolidation orswarming phases. (A) Genes with increased transcription on swarmagar (consolidation or swarming) compared to broth. (B) Genes withdecreased transcription on swarm agar (consolidation or swarming)compared to broth.
2836 PEARSON ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

been identified as active in swarm behavior (13, 14, 43), wereupregulated during consolidation. In addition, genes involvedin central metabolic pathways, including gluconeogenesis andthe tricarboxylic acid (TCA) cycle, were upregulated duringconsolidation (see Table S6 in the supplemental material).Surprisingly, many flagellar genes were also upregulated dur-ing consolidation. Classes of genes found to be differentiallyregulated during swarming are summarized in Fig. 3.
Validation of microarray data by qRT-PCR. To indepen-dently confirm data obtained by microarray analysis, qRT-PCRwas conducted using cDNA templates obtained from consoli-dation-phase or swarming-phase P. mirabilis with primers cor-
responding to genes predicted to be up- or downregulatedunder each condition examined. Table 5 shows the results forgenes predicted to be differentially regulated in consolidatecompared to swarm. Nine of the 10 genes analyzed respondedas predicted by the microarray.
Swarming-regulated genes are not necessarily required forswarming. Because many genes involved in amino acid orpeptide uptake were upregulated during consolidation, we hy-pothesized that these processes might play a role in swarmsignaling. In particular, the entire opp oligopeptide permeaseoperon, oppA2ABCDF, was upregulated greater than 5-fold inconsolidate compared to swarm by microarray analysis (Table
TABLE 1. Fifty genes most upregulated in consolidate compared to broth-cultured cells
Gene Annotation Swarma Fold change
PMI1629 fliE flagellar hook-basal body complex protein FliE Yes 10.20PMI1654 flgB flagellar basal body rod protein FlgB Yes 10.17PMI1618 fliA RNA polymerase sigma factor for flagellar operon Yes 9.86PMI1961 ccm putative membrane protein (Ccm1 protein) Yes 9.49PMI1649 flgG flagellar basal body rod protein FlgG (distal rod protein) Yes 9.24PMI1653 flgC flagellar basal body rod protein FlgC Yes 8.96PMI2898 Putative amino acid ABC transporter, substrate-binding protein Yes 8.86PMI1650 flgF flagellar basal body rod protein Yes 8.85PMI1631 fliG flagellar motor switch protein FliG Yes 8.50PMI1637 fliM flagellar motor switch protein FliM Yes 8.48PMI1636 fliL flagellar protein FliL Yes 8.37PMI1621 flaD flagellar hook-associated protein 2 Yes 8.19PMI1617 fliZ putative alternative sigma factor regulatory protein Yes 8.19PMI1359 Putative attachment invasion locus protein Yes 8.13PMI0182 Putative transcriptional regulator (MrpJ homolog) Yes 7.90PMI1630 fliF flagellar M-ring protein No 7.82PMI1651 flgE flagellar hook protein FlgE Yes 7.76PMI1318 nlpA lipoprotein 28 No 7.71PMI1638 fliN flagellar motor switch protein FliN Yes 7.39PMI1442 Hypothetical protein Yes 7.28PMI1647 flgI flagellar P-ring protein precursor Yes 7.21PMI0876 umoD upregulator of flagellar master operon Yes 7.18PMI1660 flhB flagellar biosynthetic protein FlhB No 7.18PMI1671 flhC flagellum biosynthesis transcription activator No 6.85PMI1624 Conserved hypothetical protein No 6.82PMI3460 Putative lipoprotein Yes 6.63PMI0808 ddg cold-induced palmitoleoyl transferase No 6.62PMI2496 rsmC rRNA small subunit methyltransferase C Yes 6.49PMI1908 Putative colicin No 6.38PMI3036 sodA superoxide dismutase (Mn) Yes 6.31PMI1248 Putative cystathionine gamma-synthase Yes 6.09PMI1669 motB chemotaxis motility protein B Yes 5.96PMI1657 flgN flagellar synthesis protein FlgN Yes 5.92PMI1666 cheD methyl-accepting chemotaxis protein No 5.76PMI0994 Putative lipoprotein Yes 5.72PMI1645 flgK flagellar hook associated protein 1 Yes 5.72PMI1661 cheZ chemotaxis phosphatase Yes 5.65PMI1644 flgL flagellar hook-associated protein 3 Yes 5.51PMI0272 Probable transport protein Yes 5.41PMI1622 fliS flagellar protein FliS Yes 5.40PMI1635 fliK flagellar hook length control protein Yes 5.39PMI0044 Putative outer membrane protein No 5.37PMI0842 TonB-dependent receptor Yes 5.33PMI1670 motA chemotaxis motility protein A Yes 5.29PMI1633 fliI flagellum-specific ATP synthase No 5.18PMI0029 exbB biopolymer transport protein Yes 5.03PMI1662 cheY chemotaxis response regulator Yes 5.01PMI3384 fklB FkbP-type peptidyl-prolyl cis-trans isomerase No 4.99PMI2808 Methyl-accepting chemotaxis protein Yes 4.92PMI1835 cysP sulfate/thiosulfate ABC transporter, thiosulfate-binding protein Yes 4.91
a Genes that are also listed on Table 2.
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2837
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

3; see also Table S9 in the supplemental material), and thistrend was confirmed for oppA by qRT-PCR (Table S5C).Therefore, three genes, dppA (dipeptide permease, PMI2847,upregulated 4.08-fold in consolidate compared to swarm),oppB (oligopeptide permease, PMI1474, upregulated 6.70-foldin consolidate compared to swarm), and cysJ (sulfite reductaseflavoprotein subunit, PMI2250, upregulated 4.26-fold in con-solidate compared to swarm) were inactivated by insertion of akanamycin resistance cassette. Because each of the targetedgenes is predicted to be involved with peptide uptake or aminoacid biosynthesis, the mutant strains were tested for swarmingon minimal A medium supplemented with 1% tryptone as a
protein source (MinA-T). P. mirabilis swarms on MinA-T agarwith a bull’s-eye pattern of swarming and consolidation similarto that seen on LB swarm agar (Fig. 4, top panel), although theswarm edge is thinner on MinA-T. As on LB agar, P. mirabilisalternates between short consolidate and long swarmer formson MinA-T agar (Fig. 4, center panel). Swarming initially pro-ceeds more slowly on MinA-T but then advances at a ratecomparable to that seen on LB agar (Fig. 4, bottom panel).Gene expression on MinA-T agar, analyzed by qRT-PCR, issimilar to expression on LB agar; amino acid biosyntheticgenes gltI and cysJ and the outer membrane protein OmpA areall upregulated during consolidate on MinA-T (fold changes of
TABLE 2. Fifty genes most upregulated in swarm compared to broth-cultured cells
Gene Annotation Consolidatea Fold change
PMI1618 fliA RNA polymerase sigma factor for flagellar operon Yes 10.11PMI1359 Putative attachment invasion locus protein Yes 9.95PMI1629 fliE flagellar hook-basal body complex protein FliE Yes 9.17PMI2898 Putative amino acid ABC transporter, substrate-binding protein Yes 9.17PMI1654 flgB flagellar basal body rod protein FlgB Yes 8.94PMI1617 fliZ putative alternative sigma factor regulatory protein Yes 8.91PMI1653 flgC flagellar basal body rod protein FlgC Yes 8.40PMI3460 Putative lipoprotein Yes 8.21PMI2057 hpmA hemolysin No 8.12PMI1438 Putative membrane protein No 7.96PMI1650 flgF flagellar basal body rod protein Yes 7.76PMI1637 fliM flagellar motor switch protein FliM Yes 7.48PMI1442 Hypothetical protein Yes 7.32PMI0993 Putative lipoprotein No 7.21PMI1649 flgG flagellar basal body rod protein FlgG (distal rod protein) Yes 7.19PMI1647 flgI flagellar P-ring protein precursor Yes 6.96PMI1644 flgL flagellar hook-associated protein 3 Yes 6.90PMI1961 ccm putative membrane protein (Ccm1 protein) Yes 6.90PMI0182 Putative transcriptional regulator (MrpJ homolog) Yes 6.78PMI1621 flaD flagellar hook-associated protein 2 Yes 6.73PMI3036 sodA superoxide dismutase (Mn) Yes 6.71PMI2496 rsmC rRNA small subunit methyltransferase C Yes 6.65PMI1651 flgE flagellar hook protein FlgE Yes 6.63PMI1636 fliL flagellar protein FliL Yes 6.39PMI1638 fliN flagellar motor switch protein FliN Yes 6.36PMI2808 Methyl-accepting chemotaxis protein Yes 6.31PMI1631 fliG flagellar motor switch protein FliG Yes 6.20PMI1669 motB chemotaxis motility protein B Yes 6.16PMI0842 TonB-dependent receptor Yes 6.16PMI0994 Putative lipoprotein Yes 6.10PMI1622 fliS flagellar protein FliS Yes 6.02PMI0187 dsdA D-serine dehydratase No 6.00PMI1634 fliJ flagellar protein FliJ No 5.89PMI1835 cysP sulfate/thiosulfate ABC transporter, thiosulfate-binding protein Yes 5.74PMI1661 cheZ chemotaxis phosphatase Yes 5.45PMI2385 terC tellurite resistance protein No 5.42PMI1665 tap methyl-accepting chemotaxis protein No 5.36PMI1645 flgK flagellar hook-associated protein 1 Yes 5.34PMI2196 Putative membrane-associated CAAX amino-terminal protease No 5.19PMI0876 umoD upregulator of flagellar master operon Yes 5.17PMI1005 Putative membrane protein No 5.11PMI1657 flgN flagellar synthesis protein FlgN Yes 5.10PMI0272 Probable transport protein Yes 5.07PMI1248 Putative cystathionine gamma-synthase Yes 5.05PMI1635 fliK flagellar hook length control protein Yes 5.00PMI1670 motA chemotaxis motility protein A Yes 4.95PMI0276 zapD type I secretion outer membrane protein No 4.94PMI1662 cheY chemotaxis response regulator Yes 4.92PMI2383 terE tellurite resistance protein No 4.88PMI0029 exbB biopolymer transport protein Yes 4.83
a Genes that are also listed on Table 1.
2838 PEARSON ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

2.02, 2.86, and 3.14, respectively, compared to swarm). How-ever, the oligopeptide transporter gene oppA was not signifi-cantly upregulated during consolidation phase compared toswarm (fold change of 1.10). We hypothesized that if peptide
and amino acid uptake contributed to swarm behavior,MinA-T agar might reveal subtle defects in swarming by pep-tide or amino acid uptake mutants. On this medium, all mu-tants tested had a defect in swarming (Fig. 5), with the mostpronounced phenotype in the cysJ mutant. It should be notedthat P. mirabilis HI4320 does not swarm on unaugmentedminimal A agar. However, none of these mutants had a statis-tically significant defect in swarming behavior when tested onLB swarm agar (data not shown).
Sequencing and annotation of the P. mirabilis HI4230 ge-nome (52) led to the observation that this species encodes twohomologs of lrhA, a LysR family transcriptional regulator.These two genes, PMI0629 (lrhA) and PMI1764 (hexA), en-code proteins with 54% amino acid sequence identity and 74%similarity to each other. Only one of the two homologs, hexA,was found to be upregulated in consolidate compared to swarm(3.60-fold); lrhA was not found to be differentially regulatedgreater than 2-fold under any of the conditions tested by mi-croarray. To learn what roles lrhA and hexA might play inswarming, lrhA and hexA mutants and an lrhA hexA doublemutant were tested for swarming on MinA-T agar (Fig. 5).While the lrhA mutant had only a modest decrease in swarm-ing, the hexA and lrhA hexA mutants both swarmed signifi-cantly less than did the wild-type parent (P 0.01). Interest-
TABLE 3. Fifty genes most upregulated in consolidatecompared to swarm
Gene Annotation Foldchange
PMI0785 ompA outer membrane protein A 9.09PMI1908 Putative colicin 8.30PMI1475 oppA periplasmic oligopeptide-binding protein
precursor8.10
PMI1592 fbaB fructose-bisphosphate aldolase class I 7.86PMI0250 Putative lipoprotein 7.73PMI2019 gcvP glycine dehydrogenase 7.58PMI3037 acs acetyl-coenzyme A synthetase 7.52PMI2236 rpoS RNA polymerase sigma factor RpoS 7.48PMI2662 chb secreted chitin-binding protein 6.96PMI0437 gltI glutamate/aspartate ABC transporter,
substrate-binding protein6.91
PMI3579 Conserved hypothetical protein 6.89PMI1056 Putative TRAP-type transport system substrate-
binding protein6.86
PMI2114 Putative sodium:alanine symporter 6.82PMI1474 oppB oligopeptide transport system permease
protein6.70
PMI1421 ppsA phosphoenolpyruvate synthase 6.69PMI1722 Phage protein 6.68PMI1615 putP proline permease 6.49PMI2246 cysD sulfate adenylyltransferase subunit 2 6.37PMI2052 coaE dephospho-CoA kinase 6.36PMI2255 Conserved hypothetical protein 6.34PMI2254 mdeA methionine gamma-lyase 6.33PMI3177 kbl 2-amino-3-ketobutyrate coenzyme A ligase 6.28PMI1075 Hypothetical protein 6.27PMI0968 Isocitrate dehydrogenase (NADP) (partial) 6.20PMI3559 accA3 putative bifunctional acetyl-/propionyl-
coenzyme A carboxylase alpha chain6.17
PMI3556 Putative transmembrane transport protein 6.16PMI0564 gltA citrate synthase GltA 6.11PMI0971 Putative lipase 6.03PMI1451 uspG2 universal stress protein G 6.02PMI3178 tdh threonine 3-dehydrogenase 6.01PMI2256 Putative oxidoreductase 5.99PMI1471 oppF oligopeptide transport ATP-binding
protein5.95
PMI1473 oppC oligopeptide transport system permeaseprotein
5.90
PMI1171 Conserved hypothetical protein 5.77PMI1472 oppD oligopeptide transport ATP-binding
protein5.76
PMI1616 putA proline dehydrogenase/delta-1-pyrroline-5-carboxylate
5.76
PMI0360 Putative general stress response protein 5.69PMI1730 rcsB two-component system response regulator,
capsular synthesis5.62
PMI2268 Short-chain dehydrogenase 5.60PMI0047 Putative secreted 5’-nucleotidase 5.54PMI2036 Putative exported protein 5.49PMI1737 Putative lipoprotein 5.49PMI3401 sfsB sugar fermentation stimulation protein 5.43PMI2890 greB transcription elongation factor 5.42PMI1509 dadA D-amino acid dehydrogenase small subunit 5.38PMI3701 Putative sodium:dicarboxylate symporter 5.37PMI0952 Putative isocitrate dehydrogenase (NADP)
(partial)5.34
PMI3204 sbp exported sulfate-binding protein 5.33PMI3115 umoA putative upregulator of flagellar operon 5.32PMI3449 Conserved hypothetical protein 5.30
TABLE 4. Genes upregulated �2-fold in swarm comparedto consolidate
Gene Annotation Foldchange
PMI0875 ndh NADH dehydrogenase 4.67PMI0913 cspA cold shock protein 2.60PMI3584 sugE quaternary ammonium compound
resistance protein2.39
PMI2309 recB exodeoxyribonuclease V -chain 2.36PMI0807 cspB major cold shock protein 2.24PMI1140 Rhs family protein 2.24PMI0972 cspG cold shock protein 2.18PMI2667 Putative mannosyl-glycoprotein endo-
-N-acetylglucosaminidase2.07
PMI2732 atfE fimbrial adhesin 2.06
FIG. 3. Functional categories of genes differentially expressed dur-ing swarming or consolidation. The number of genes differentiallyregulated for each class is shown on the y axis.
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2839
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

ingly, the double mutant had a slightly greater defect than didthe hexA single mutant (P � 0.0318). On LB swarm agar, boththe hexA and lrhA hexA mutants consistently had a decreasedswarm radius compared to that of the wild-type parent, al-though these differences were not significant (data not shown).
The growth rates for all six mutants constructed for thisstudy were also determined in LB broth. Only the oppB mutanthad a modest growth deficiency in broth culture, both growingmore slowly and not reaching the same density as that reachedby the wild-type parent (data not shown).
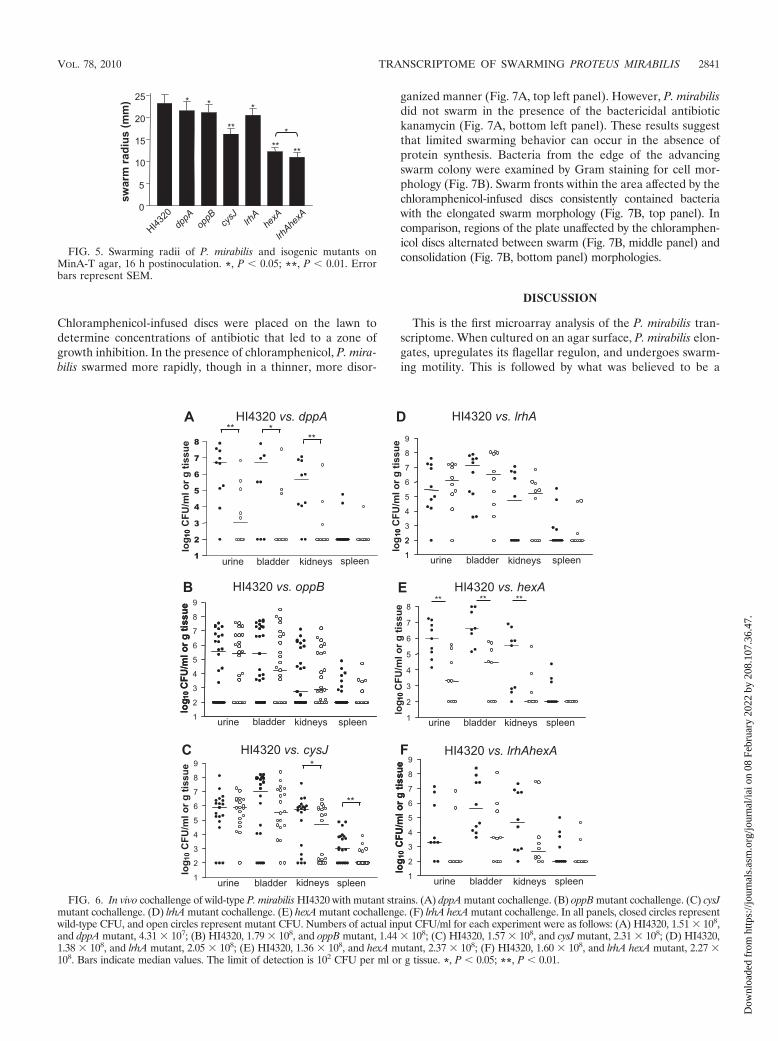
Swarming-regulated genes contribute to in vivo fitness. Vir-ulence factors, including hemolysin and the Zap metallopro-tease, are upregulated during culture on swarm agar (4, 62). Totest whether the six mutants described above (dppA, oppB, cysJ,lrhA, hexA, and lrhA hexA) were attenuated in the ability tocause infection, we transurethrally inoculated mice with a 1:1mixture of each mutant with the wild-type parent strain (totalinoculum, 1 � 107 CFU/mouse). After 7 days, bacteria wereenumerated in the urine, bladders, kidneys, and spleens (Fig.6). The dppA, cysJ, and hexA mutants had significant coloniza-tion defects relative to the wild-type parent strain (Fig. 6A, C,and E), indicating that these genes contribute to fitness duringinfection of the urinary tract. Specifically, the dppA mutant wasrecovered in reduced numbers from the urine, bladder, andkidneys (P values of 0.0039, 0.0156, and 0.0078, respectively),the cysJ mutant was defective in colonization of the kidneysand spleen (P values of 0.0119 and 0.0052, respectively), andthe hexA mutant was found in lower quantities in the urine,bladder, and kidneys (P values of 0.0039, 0.0039, and 0.0078,respectively). Likewise, the lrhA hexA mutant tended to berecovered in lower numbers from these same sites, althoughthese differences were not significant compared to the wildtype (P � 0.05). This suggests that cysteine biosynthesis orsulfate assimilation may be more important during infection ofthe upper urinary tract (i.e., pyelonephritis), while dipeptideuptake and regulation by HexA may be necessary throughoutthe urinary tract. In contrast, the oppB mutant survived at allsites in numbers similar to those of the wild-type parent (Fig.6B) despite exhibiting a modest growth defect in LB broth
culture. The lrhA mutant was also recovered in numbers sim-ilar to those of the wild-type parent (Fig. 6D).
P. mirabilis swarms in the absence of protein synthesis.Because 541 genes were upregulated in consolidate comparedto swarm, but only nine genes were upregulated in swarmcompared to consolidate, we hypothesized that P. mirabilismight be able to swarm in the absence of protein synthesis. Totest this, the bacteriostatic antibiotic chloramphenicol, whichinterferes with protein synthesis by binding to the 50S subunitof the ribosome, was added to paper discs. P. mirabilis HI4320was added to the center of swarm agar and allowed to dry. Thechloramphenicol-infused discs were placed approximately 1 cmfrom the inoculation point, and the swarm colony was allowedto develop. As a control, P. mirabilis HI4320 was spread platedonto nonswarm LB agar to create a lawn and allowed to dry.
FIG. 4. Comparison of swarming on LB and minimal A agar sup-plemented with 1% tryptone. Top panel, agar plates with characteristicbull’s-eye P. mirabilis swarm colonies on LB agar (left) and MinA-Tagar (right), 36 h postinoculation. Middle panel, Gram-stained cellscollected during the consolidation (short form) and swarm (long form)phases from LB agar (left) and MinA-T agar (right). Consolidate wascollected at 14 h postinoculation, and swarm was collected at 17 hpostinoculation. All four Gram stain photos were recorded at �400magnification. Bottom panel, the swarm radius of colonies on LB(closed circles) and MinA-T (open circles) agar was measured hourly.Three plates were measured for each agar type. Error bars representthe standard errors of the means (SEM).
TABLE 5. Genes analyzed by qRT-PCRa
Gene Name AnnotationLog2 fold change
Array qRT-PCR
PMI0437 gltI Glutamate/aspartate ABCtransport
2.79 2.46 � 0.72
PMI0785 ompA Outer membrane protein A 3.18 6.64 � 0.59PMI0875 ndh NADH dehydrogenase �2.22 �2.11 � 0.77PMI0913 cspA Cold shock protein �1.38 �2.00 � 0.68PMI1475 oppA Oligopeptide ABC
transporter3.02 2.80 � 1.49
PMI1722 Phage protein 2.74 �1.77 � 1.64PMI1730 rcsB Capsular synthesis
regulator2.49 0.16 � 0.34
PMI1908 Putative colicin 3.05 1.27 � 1.11PMI2250 cysJ Sulfite reductase (NADPH)
flavoprotein2.09 3.08 � 0.98
PMI3037 acs Acetyl coenzyme Asynthetase
2.91 1.11 � 0.40
a Positive values are upregulated in consolidate compared to swarm; negativevalues are upregulated in swarm compared to consolidate.
2840 PEARSON ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.
Chloramphenicol-infused discs were placed on the lawn todetermine concentrations of antibiotic that led to a zone ofgrowth inhibition. In the presence of chloramphenicol, P. mira-bilis swarmed more rapidly, though in a thinner, more disor-
ganized manner (Fig. 7A, top left panel). However, P. mirabilisdid not swarm in the presence of the bactericidal antibiotickanamycin (Fig. 7A, bottom left panel). These results suggestthat limited swarming behavior can occur in the absence ofprotein synthesis. Bacteria from the edge of the advancingswarm colony were examined by Gram staining for cell mor-phology (Fig. 7B). Swarm fronts within the area affected by thechloramphenicol-infused discs consistently contained bacteriawith the elongated swarm morphology (Fig. 7B, top panel). Incomparison, regions of the plate unaffected by the chloramphen-icol discs alternated between swarm (Fig. 7B, middle panel) andconsolidation (Fig. 7B, bottom panel) morphologies.
DISCUSSION
This is the first microarray analysis of the P. mirabilis tran-scriptome. When cultured on an agar surface, P. mirabilis elon-gates, upregulates its flagellar regulon, and undergoes swarm-ing motility. This is followed by what was believed to be a
FIG. 5. Swarming radii of P. mirabilis and isogenic mutants onMinA-T agar, 16 h postinoculation. *, P 0.05; **, P 0.01. Errorbars represent SEM.
FIG. 6. In vivo cochallenge of wild-type P. mirabilis HI4320 with mutant strains. (A) dppA mutant cochallenge. (B) oppB mutant cochallenge. (C) cysJmutant cochallenge. (D) lrhA mutant cochallenge. (E) hexA mutant cochallenge. (F) lrhA hexA mutant cochallenge. In all panels, closed circles representwild-type CFU, and open circles represent mutant CFU. Numbers of actual input CFU/ml for each experiment were as follows: (A) HI4320, 1.51 � 108,and dppA mutant, 4.31 � 107; (B) HI4320, 1.79 � 108, and oppB mutant, 1.44 � 108; (C) HI4320, 1.57 � 108, and cysJ mutant, 2.31 � 108; (D) HI4320,1.38 � 108, and lrhA mutant, 2.05 � 108; (E) HI4320, 1.36 � 108, and hexA mutant, 2.37 � 108; (F) HI4320, 1.60 � 108, and lrhA hexA mutant, 2.27 �108. Bars indicate median values. The limit of detection is 102 CFU per ml or g tissue. *, P 0.05; **, P 0.01.
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2841
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

resting stage, consolidation. However, this phase appears torepresent a time of preparation for the next phase of swarming,marked by the upregulation of nutrient uptake systems, centralmetabolism (TCA cycle, gluconeogenesis, and glycerol metab-olism), respiration, and cell wall synthetic enzymes that favorfilamentation but discourage septation and division.
We first compared broth-cultured P. mirabilis directly to swarmor consolidate. Most genes that were differentially regulated inconsolidate were similarly regulated in swarm, suggesting that thisexperimental approach was amenable for distinguishing the brothenvironment from the swarm agar environment, but is not asuseful for pinpointing the changes that occur as P. mirabilis cyclesfrom swarm to consolidate and back again. Thus, a second seriesof microarray experiments was conducted to compare swarm di-rectly to consolidate.
The swarming phase of P. mirabilis is dynamic, with writhingmotion easily detectable on the agar surface using light mi-croscopy. Thus, it was initially surprising to find that 541 geneswere upregulated during consolidation, yet only nine geneswere upregulated during swarming in this direct comparison.Additionally, genes expected to be upregulated during swarm-ing, especially those involved in flagellar synthesis, were in-stead either highly expressed during both swarm and consoli-dation phases or upregulated during the consolidation phaseonly. It should be noted, however, that P. mirabilis convertsbetween the swarm and consolidation phases approximatelyevery 2 h under the conditions used in this study. We hypoth-esize that the consolidation phase is a time where the bacteriaare preparing for the next swarm cycle. That is, genes whoseproducts are required during the swarming phase (i.e., flagella)should actually be upregulated during the previous consolida-tion phase, allowing time for proteins to be assembled andfunctional during the correct phase. Likewise, the activity of
thousands of coordinated flagella during the swarming phaseconsumes considerable energy, and the consolidation phasemay require the upregulation of genes involved in nutrientuptake (e.g., amino acid and peptide uptake systems) to re-plenish energy reserves. These observations are consistent witha previous report that indicated that swarm cells are metabol-ically less active than consolidate cells (7). However, our re-sults, which revealed very high levels of flagellar gene expres-sion through both swarming and consolidation phases, withhighest expression during consolidation, contrast with previouswork in which flhDC and flaA (called fliC in that report) variedin expression 30-fold by Northern blot analysis and, at lowestexpression levels, had almost undetectable transcript (23). Themajor difference between these two studies is that the currentstudy used bacteria taken from the edge of a swarming colony,while the earlier report used P. mirabilis spread evenly on anagar surface and followed transcript levels from bacteriawashed from the agar surface for 5.5 h after seeding. That is, a5.5-h-old seeded bacterium and a newly divided consolidationcell at the edge of a swarm colony may both have a shortmorphotype, but they are unlikely to be physiologically equiv-alent.
As one would predict, when P. mirabilis undergoes the rad-ical transformation from an elongated, multinucleated cell tothe short individual bacterium, there must be a significantremodeling of peptidoglycan and the cell wall. Indeed, genesinvolved in such remodeling are significantly upregulated inthe consolidate compared to the swarmer. Genes upregulatedduring consolidation appear to prepare P. mirabilis for fila-mentation. For example, upregulation of minC (4.61-fold) andminD (2.36-fold) would produce an excess of MinCD, whichprevents cell division and results in filamentation (18). Genesinvolved in chromosomal partitioning in the multinuclear
FIG. 7. P. mirabilis swarming in the presence of antibiotics. (A) In the top panels, discs were infused with chloramphenicol (chl). In the bottompanels, discs were infused with kanamycin (kan). The panels on the left show swarm agar, while the panels on the right are nonswarm agar. Onall plates the discs contain, clockwise from left, 2, 10, 20, or 50 �g antibiotic. (B) Gram stains from a representative chloramphenicol disc swarmplate (50 �g chl/disc). Top, swarm front next to the disc; middle, swarm front from a region of the plate unaffected by chloramphenicol; bottom,consolidation front from a region of the plate unaffected by chloramphenicol. Bar, 50 �m.
2842 PEARSON ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

swarmer cell, mukB, mukE, and mukF (46, 66), are all upregu-lated (2.31-, 3.26-, and 2.88-fold, respectively). Upregulation ofmreC (2.75-fold) is consistent with maintenance of a rod-shaped swarmer cell (48). Finally, upregulation of mtgA, en-coding a monofunctional biosynthetic peptidoglycan transgly-cosylase (19), would ensure glycan chain elongation in thedeveloping swarmer cell. On the other hand, genes responsiblefor routine synthesis of peptidoglycan (ftsZ, ftsI, ftsL, murD,murE, murF, and mraY) are not altered in expression betweenswarmer and consolidate cells.
Genes differentially regulated during P. mirabilis swarminghave interesting parallels and contrasts with similar studiesconducted with swarming Salmonella (37, 63) and E. coli (32).For purposes of comparison, it should be noted that while P.mirabilis swarms on surfaces containing up to 2% agar, Salmo-nella and E. coli require a lower agar concentration (0.5 to0.8%) and richer media (i.e., LB supplemented with glucose)to swarm. Swarming Salmonella and E. coli also lack the cyclicpattern of swarming and consolidation seen in P. mirabilis andinstead swarm continuously to the agar edge (29, 37). Kim andSurette (37) examined protein expression in actively swarmingSalmonella and found that amino acid biosynthetic pathwaysand central metabolism, particularly the TCA cycle, were in-creased compared to that in swimming samples; however, ma-jor outer membrane porins, including OmpA, were decreasedduring swarming. Interestingly, cysteine biosynthesis in partic-ular was necessary for full swarm cell differentiation (60). Incontrast, we found genes involved in all of these systems(amino acid biosynthesis, TCA cycle, as well as peptide andnonspecific uptake systems) were upregulated during consoli-dation. A microarray study by Wang et al. (63) analyzed geneexpression in Salmonella on swarm agar with Salmonella onhard agar or in broth culture and found that gene expressionpatterns were more similar under the two agar-cultured con-ditions compared to broth culture; this is comparable to thefinding that P. mirabilis has more genes expressed in commonduring swarm and consolidation than during exponential-phasebroth culture. The same study also reported that genes in-volved in basic metabolism and cysteine biosynthesis were up-regulated during swarming by Salmonella (63). A saturatingknockout mutant screen of E. coli K-12 (32) also indicated theimportance of the TCA cycle in swarming yet also identifiedglucose metabolism as a requirement for swarming in thatspecies. In contrast, we did not find glucose metabolic genesupregulated during swarming by P. mirabilis, which does notrequire exogenous glucose for swarming.
Previous swarming reports also indicated that P. mirabilis ismetabolically less active during swarming and that amino aciduptake increases during consolidation (7, 36). The microarrayresults from the current study support these findings. Numer-ous markers of metabolic activity were upregulated duringconsolidation, including peptide uptake, amino and nucleicacid biosynthesis, central metabolism, and aerobic respiration.Combining the microarray results with the ability of P. mirabilisto swarm in the absence of protein synthesis suggests that theswarming state is almost wholly consumed with flagellum-me-diated motility. We propose that after this energy has beenexpended, P. mirabilis enters the consolidation phase, which isfocused upon replenishing supplies. Perhaps the replenishing
consolidation stage allows P. mirabilis to swarm under condi-tions that are not permissive for other species.
A particular concern when analyzing the transcriptome of acyclical event is ensuring sample uniformity. Great effort wastaken to confirm that the bacterial inputs were in the desired(swarm or consolidation) phase by observing the appearanceand motion at the colony edge and by examining a portion ofthe input sample by Gram staining. The desired morphotypepredominated in both consolidate (�99% of bacteria were10 �m) and swarm (�88% bacteria were �10 �m) cells,respectively. One strength of this study is that only the edge ofthe bacterial colony was collected for RNA isolation; thus, onlythe youngest cells in the swarm colony were analyzed for bothswarming and consolidation phases. Difficulties in obtainingpure samples of swarm or consolidate cells have been noted byothers (7, 64), and older vegetative cells from the interior ofthe swarm colony have been used in the past for comparisonswith swarm cells (1, 21). It is important to emphasize that inour study many genes previously known to be swarming regu-lated were also identified by microarray (e.g., flagella, fimbriae,and Lon and Zap proteases). Furthermore, our P. mirabilisHI4320 microarray has also been validated by comparingHI4320 transcripts from iron-restricted with iron-replete broth(S. D. Himpsl, M. M. Pearson, and H. L. T. Mobley, submittedfor publication).
In this report, we found that a hexA mutant had a significantdefect in swarming. In contrast, an earlier study (15) concludedthat hexA (called lrhA in that report) had no effect on swarm-ing. There are several possible explanations for the differentoutcomes. The first study examined swarming on LB agar in-cubated at 37°C, while the current report examined swarmingon a minimal medium supplemented with tryptone, MinA-Tagar, incubated at 30°C. Indeed, although we found that thehexA mutant tended to have reduced swarming motility on LBagar, the difference was not statistically significant (data notshown). It is also possible that, in the earlier study, the lrhAgene was able to complement the hexA mutant in the P. mira-bilis strain used. The different results may also be a conse-quence of mutant construction. The hexA mutant in our studywas constructed by inserting an intron between nucleotides 463and 464 of hexA, resulting in the first 154 amino acids of the305-amino-acid protein potentially being produced. Clemmerand Rather (15) used an internal cloned fragment of hexA ona suicide plasmid, which would result in the first 206 aminoacids potentially being produced (i.e., the C-terminal 33% ofthe protein would be missing). Although this seems sufficientto ablate HexA function, we have deleted as much as theC-terminal third of another regulatory protein, MrpJ, andfound that functional protein was still produced (50). BothhexA mutants could result in translated HexA protein with anintact helix-turn-helix domain; in addition, our mutant bears 57of 151 amino acids of the predicted substrate-binding domain,while the other hexA mutant could produce 105/151 aminoacids of this domain. As we cannot rule out the possibility thateither hexA mutant is able to produce an active, truncatedHexA protein, ideally, a complete hexA deletion would betested for swarming ability in a future study.
P. mirabilis (4, 62) and other species (24, 49, 63) upregulatevirulence factors during swarming, and in this report, morevirulence factors in ascending UTI were identified. Thus, fur-
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2843
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

ther analysis of the swarming transcriptome microarray datawill likely be useful for identifying additional virulence factorsfor this and other pathogens capable of swarming. The asso-ciation of swarming with virulence may seem to conflict withearlier work, in which P. mirabilis was found in the swarmmorphotype only rarely in a mouse model of UTI (33). How-ever, P. mirabilis swarms on urinary catheters (35, 54), whichwere not used in the previous study; this may represent amechanism by which the bacterium gains access to the cathe-terized urinary tract, the bacterium’s typical niche. It is notknown how quickly P. mirabilis switches to the vegetative formonce inside the urinary tract, and it is possible that swarming P.mirabilis cells are primed to be virulent as soon as they gainaccess to the bladder. The ability to prime genetic expressionunder a particular growth condition in preparation for enteringa new environment has already been demonstrated for Vibriocholerae, which expresses genes needed for survival in anaquatic environment during the later stages of intestinal infec-tion (55). Urinary catheters may serve as a constant source ofswarming P. mirabilis cells in the urinary tract. Further exper-imentation is needed to address this possibility.
The discovery that peptide and amino acid uptake are crit-ical for P. mirabilis during UTI meshes with metabolic studiesof uropathogenic E. coli (UPEC). P. mirabilis and other uri-nary tract pathogens are capable of growth in human urine,where the main carbon sources are amino acids and peptides(6). While P. mirabilis dppA and cysJ mutants are attenuated invivo, it was surprising that an oppB mutant had no defect invivo. This result contrasts with a similar study of UPEC strainCFT073, in which both dppA and oppA mutants could notsuccessfully compete with the wild-type parent during experi-mental infection (5). A possible answer may lie in the organi-zation of the opp operons in these two species. The UPECoperon consists of the genes oppABCDF, while the P. mirabilisopp locus has two copies of oppA, designated oppA and oppA2(which are 57% identical to each other, with 77% similaramino acid sequences). Perhaps the P. mirabilis OppA2 pro-tein is able to import oligopeptides through another channel.Alternatively, other peptide import systems in P. mirabilis thatare not present in UPEC may compensate for the oppB mu-tation.
The use of microarrays to study swarming has raised newquestions about this long-studied phenomenon. Why are nu-trient uptake pathways upregulated during consolidation?What is the role played by genes that were unexpected to varyin expression during swarming, such as the tellurite resistance(ter) genes or phage? Can any of the genes in this study beharnessed to control or prevent swarming on urinary cathe-ters? How many swarming-regulated genes are also differen-tially regulated during infection? Perhaps this aptly namedpathogen will now be less able to avoid questioning even as itchanges forms.
ACKNOWLEDGMENTS
This work was funded by Public Health Service grant AI059722 fromthe National Institutes of Health (H.L.T.M.). Analysis in the labora-tory of D.A.R. is supported by University of Maryland School ofMedicine internal funds. M.M.P. was supported in part by NationalResearch Service Award F32 AI068324.
REFERENCES
1. Allison, C., N. Coleman, P. L. Jones, and C. Hughes. 1992. Ability of Proteusmirabilis to invade human urothelial cells is coupled to motility and swarmingdifferentiation. Infect. Immun. 60:4740–4746.
2. Allison, C., and C. Hughes. 1991. Closely linked genetic loci required forswarm cell differentiation and multicellular migration by Proteus mirabilis.Mol. Microbiol. 5:1975–1982.
3. Allison, C., H. C. Lai, D. Gygi, and C. Hughes. 1993. Cell differentiation ofProteus mirabilis is initiated by glutamine, a specific chemoattractant forswarming cells. Mol. Microbiol. 8:53–60.
4. Allison, C., H. C. Lai, and C. Hughes. 1992. Co-ordinate expression ofvirulence genes during swarm-cell differentiation and population migrationof Proteus mirabilis. Mol. Microbiol. 6:1583–1591.
5. Alteri, C. J., S. N. Smith, and H. L. Mobley. 2009. Fitness of Escherichia coliduring urinary tract infection requires gluconeogenesis and the TCA cycle.PLoS Pathog. 5:e1000448.
6. Altman, P. L. 1961. Physical properties and chemical composition of urine:mammals. Part 1: man, p. 363–369. In D. L. Dittmer (ed.), Blood and otherbodily fluids. Federation of American Societies for Experimental Biology,Washington, DC.
7. Armitage, J. P. 1981. Changes in metabolic activity of Proteus mirabilis duringswarming. J. Gen. Microbiol. 125:445–450.
8. Belas, R., D. Erskine, and D. Flaherty. 1991. Proteus mirabilis mutantsdefective in swarmer cell differentiation and multicellular behavior. J. Bac-teriol. 173:6279–6288.
9. Belas, R., D. Erskine, and D. Flaherty. 1991. Transposon mutagenesis inProteus mirabilis. J. Bacteriol. 173:6289–6293.
10. Belas, R., M. Goldman, and K. Ashliman. 1995. Genetic analysis of Proteusmirabilis mutants defective in swarmer cell elongation. J. Bacteriol. 177:823–828.
11. Belas, R., R. Schneider, and M. Melch. 1998. Characterization of Proteusmirabilis precocious swarming mutants: identification of rsbA, encoding aregulator of swarming behavior. J. Bacteriol. 180:6126–6139.
12. Burall, L. S., J. M. Harro, X. Li, C. V. Lockatell, S. D. Himpsl, J. R. Hebel,D. E. Johnson, and H. L. Mobley. 2004. Proteus mirabilis genes that contrib-ute to pathogenesis of urinary tract infection: identification of 25 signature-tagged mutants attenuated at least 100-fold. Infect. Immun. 72:2922–2938.
13. Claret, L., and C. Hughes. 2000. Rapid turnover of FlhD and FlhC, theflagellar regulon transcriptional activator proteins, during Proteus swarming.J. Bacteriol. 182:833–836.
14. Clemmer, K. M., and P. N. Rather. 2008. The Lon protease regulates swarm-ing motility and virulence gene expression in Proteus mirabilis. J. Med.Microbiol. 57:931–937.
15. Clemmer, K. M., and P. N. Rather. 2007. Regulation of flhDC expression inProteus mirabilis. Res. Microbiol. 158:295–302.
16. Coker, C., C. A. Poore, X. Li, and H. L. Mobley. 2000. Pathogenesis ofProteus mirabilis urinary tract infection. Microbes Infect. 2:1497–1505.
17. D’Agata, E. M. 2004. Rapidly rising prevalence of nosocomial multidrug-resistant, Gram-negative bacilli: a 9-year surveillance study. Infect. ControlHosp. Epidemiol. 25:842–846.
18. de Boer, P. A., R. E. Crossley, and L. I. Rothfield. 1989. A division inhibitorand a topological specificity factor coded for by the minicell locus determineproper placement of the division septum in E. coli. Cell 56:641–649.
19. Derouaux, A., B. Wolf, C. Fraipont, E. Breukink, M. Nguyen-Disteche, andM. Terrak. 2008. The monofunctional glycosyltransferase of Escherichia colilocalizes to the cell division site and interacts with penicillin-binding protein3, FtsW, and FtsN. J. Bacteriol. 190:1831–1834.
20. Dufour, A., R. B. Furness, and C. Hughes. 1998. Novel genes that upregulatethe Proteus mirabilis flhDC master operon controlling flagellar biogenesisand swarming. Mol. Microbiol. 29:741–751.
21. Falkinham, J. O., III, and P. S. Hoffman. 1984. Unique developmentalcharacteristics of the swarm and short cells of Proteus vulgaris and Proteusmirabilis. J. Bacteriol. 158:1037–1040.
22. Fraser, G. M., and C. Hughes. 1999. Swarming motility. Curr. Opin. Micro-biol. 2:630–635.
23. Furness, R. B., G. M. Fraser, N. A. Hay, and C. Hughes. 1997. Negativefeedback from a Proteus class II flagellum export defect to the flhDC masteroperon controlling cell division and flagellum assembly. J. Bacteriol. 179:5585–5588.
24. Ghelardi, E., F. Celandroni, S. Salvetti, M. Ceragioli, D. J. Beecher, S.Senesi, and A. C. Wong. 2007. Swarming behavior of and hemolysin BLsecretion by Bacillus cereus. Appl. Environ. Microbiol. 73:4089–4093.
25. Griffith, D. P., D. M. Musher, and C. Itin. 1976. Urease. The primary causeof infection-induced urinary stones. Invest. Urol. 13:346–350.
26. Gygi, D., M. M. Rahman, H. C. Lai, R. Carlson, J. Guard-Petter, and C.Hughes. 1995. A cell-surface polysaccharide that facilitates rapid populationmigration by differentiated swarm cells of Proteus mirabilis. Mol. Microbiol.17:1167–1175.
27. Hagberg, L., I. Engberg, R. Freter, J. Lam, S. Olling, and C. Svanborg Eden.1983. Ascending, unobstructed urinary tract infection in mice caused by
2844 PEARSON ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.

pyelonephritogenic Escherichia coli of human origin. Infect. Immun. 40:273–283.
28. Harshey, R. M. 1994. Bees aren’t the only ones: swarming in gram-negativebacteria. Mol. Microbiol. 13:389–394.
29. Harshey, R. M., and T. Matsuyama. 1994. Dimorphic transition in Esche-richia coli and Salmonella typhimurium: surface-induced differentiation intohyperflagellate swarmer cells. Proc. Natl. Acad. Sci. U. S. A. 91:8631–8635.
30. Hatt, J. K., and P. N. Rather. 2008. Characterization of a novel gene, wosA,regulating FlhDC expression in Proteus mirabilis. J. Bacteriol. 190:1946–1955.
31. Hay, N. A., D. J. Tipper, D. Gygi, and C. Hughes. 1999. A novel membraneprotein influencing cell shape and multicellular swarming of Proteus mirabi-lis. J. Bacteriol. 181:2008–2016.
32. Inoue, T., R. Shingaki, S. Hirose, K. Waki, H. Mori, and K. Fukui. 2007.Genome-wide screening of genes required for swarming motility in Esche-richia coli K-12. J. Bacteriol. 189:950–957.
33. Jansen, A. M., C. V. Lockatell, D. E. Johnson, and H. L. Mobley. 2003.Visualization of Proteus mirabilis morphotypes in the urinary tract: the elon-gated swarmer cell is rarely observed in ascending urinary tract infection.Infect. Immun. 71:3607–3613.
34. Johnson, D. E., C. V. Lockatell, M. Hall-Craigs, H. L. Mobley, and J. W.Warren. 1987. Uropathogenicity in rats and mice of Providencia stuartii fromlong-term catheterized patients. J. Urol. 138:632–635.
35. Jones, B. V., R. Young, E. Mahenthiralingam, and D. J. Stickler. 2004.Ultrastructure of Proteus mirabilis swarmer cell rafts and role of swarming incatheter-associated urinary tract infection. Infect. Immun. 72:3941–3950.
36. Jones, H. E., and R. W. Park. 1967. The short forms and long forms ofProteus. J. Gen. Microbiol. 47:359–367.
37. Kim, W., and M. G. Surette. 2004. Metabolic differentiation in activelyswarming Salmonella. Mol. Microbiol. 54:702–714.
38. Li, X., D. E. Johnson, and H. L. Mobley. 1999. Requirement of MrpH formannose-resistant Proteus-like fimbria-mediated hemagglutination by Pro-teus mirabilis. Infect. Immun. 67:2822–2833.
39. Li, X., H. Zhao, C. V. Lockatell, C. B. Drachenberg, D. E. Johnson, and H. L.Mobley. 2002. Visualization of Proteus mirabilis within the matrix of urease-induced bladder stones during experimental urinary tract infection. Infect.Immun. 70:389–394.
40. Liaw, S. J., H. C. Lai, and W. B. Wang. 2004. Modulation of swarming andvirulence by fatty acids through the RsbA protein in Proteus mirabilis. Infect.Immun. 72:6836–6845.
41. Liu, Q., M. Z. Li, D. Leibham, D. Cortez, and S. J. Elledge. 1998. Theunivector plasmid-fusion system, a method for rapid construction of recom-binant DNA without restriction enzymes. Curr. Biol. 8:1300–1309.
42. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(�delta delta C(T)) method.Methods 25:402.
43. Marr, A. K., J. Overhage, M. Bains, and R. E. Hancock. 2007. The Lonprotease of Pseudomonas aeruginosa is induced by aminoglycosides and isinvolved in biofilm formation and motility. Microbiology 153:474–482.
44. Matsuyama, T., Y. Takagi, Y. Nakagawa, H. Itoh, J. Wakita, and M. Mat-sushita. 2000. Dynamic aspects of the structured cell population in a swarm-ing colony of Proteus mirabilis. J. Bacteriol. 182:385–393.
45. Mobley, H. L., and J. W. Warren. 1987. Urease-positive bacteriuria andobstruction of long-term urinary catheters. J. Clin. Microbiol. 25:2216–2217.
46. Niki, H., A. Jaffe, R. Imamura, T. Ogura, and S. Hiraga. 1991. The new genemukB codes for a 177 kD protein with coiled-coil domains involved inchromosome partitioning of E. coli. EMBO J. 10:183–193.
47. O’Fallon, E., S. Gautam, and E. M. D’Agata. 2009. Colonization with mul-tidrug-resistant gram-negative bacteria: prolonged duration and frequentcocolonization. Clin. Infect. Dis. 48:1375–1381.
48. Osborn, M. J., and L. Rothfield. 2007. Cell shape determination in Esche-richia coli. Curr. Opin. Microbiol. 10:606–610.
49. Overhage, J., M. Bains, M. D. Brazas, and R. E. Hancock. 2008. Swarmingof Pseudomonas aeruginosa is a complex adaptation leading to increasedproduction of virulence factors and antibiotic resistance. J. Bacteriol. 190:2671–2679.
50. Pearson, M. M., and H. L. Mobley. 2008. Repression of motility duringfimbrial expression: identification of 14 mrpJ gene paralogues in Proteusmirabilis. Mol. Microbiol. 69:548–558.
51. Pearson, M. M., and H. L. Mobley. 2007. The type III secretion system ofProteus mirabilis HI4320 does not contribute to virulence in the mouse modelof ascending urinary tract infection. J. Med. Microbiol. 56:1277–1283.
52. Pearson, M. M., M. Sebaihia, C. Churcher, M. A. Quail, A. S. Seshasayee,N. M. Luscombe, Z. Abdellah, C. Arrosmith, B. Atkin, T. Chillingworth, H.Hauser, K. Jagels, S. Moule, K. Mungall, H. Norbertczak, E. Rabbinowitsch,D. Walker, S. Whithead, N. R. Thomson, P. N. Rather, J. Parkhill, and H. L.Mobley. 2008. Complete genome sequence of uropathogenic Proteus mira-bilis, a master of both adherence and motility. J. Bacteriol. 190:4027–4037.
53. Rauprich, O., M. Matsushita, C. J. Weijer, F. Siegert, S. E. Esipov, and J. A.Shapiro. 1996. Periodic phenomena in Proteus mirabilis swarm colony de-velopment. J. Bacteriol. 178:6525–6538.
54. Sabbuba, N., G. Hughes, and D. J. Stickler. 2002. The migration of Proteusmirabilis and other urinary tract pathogens over Foley catheters. BJU Int.89:55–60.
55. Schild, S., R. Tamayo, E. J. Nelson, F. Qadri, S. B. Calderwood, and A.Camilli. 2007. Genes induced late in infection increase fitness of Vibriocholerae after release into the environment. Cell Host Microbe 2:264–277.
56. Schneider, R., C. V. Lockatell, D. Johnson, and R. Belas. 2002. Detectionand mutation of a luxS-encoded autoinducer in Proteus mirabilis. Microbi-ology 148:773–782.
57. Stahl, S. J., K. R. Stewart, and F. D. Williams. 1983. Extracellular slimeassociated with Proteus mirabilis during swarming. J. Bacteriol. 154:930–937.
58. Stevenson, L. G., and P. N. Rather. 2006. A novel gene involved in regulatingthe flagellar gene cascade in Proteus mirabilis. J. Bacteriol. 188:7830–7839.
59. Sturgill, G., and P. N. Rather. 2004. Evidence that putrescine acts as anextracellular signal required for swarming in Proteus mirabilis. Mol. Micro-biol. 51:437–446.
60. Turnbull, A. L., and M. G. Surette. 2008. D-Cysteine is required for inducedantibiotic resistance in actively swarming Salmonella enterica serovar Typhi-murium. Microbiology 154:3410–3419.
61. Verstraeten, N., K. Braeken, B. Debkumari, M. Fauvart, J. Fransaer, J.Vermant, and J. Michiels. 2008. Living on a surface: swarming and biofilmformation. Trends Microbiol. 16:496–506.
62. Walker, K. E., S. Moghaddame-Jafari, C. V. Lockatell, D. Johnson, and R.Belas. 1999. ZapA, the IgA-degrading metalloprotease of Proteus mirabilis, isa virulence factor expressed specifically in swarmer cells. Mol. Microbiol.32:825–836.
63. Wang, Q., J. G. Frye, M. McClelland, and R. M. Harshey. 2004. Geneexpression patterns during swarming in Salmonella typhimurium: genes spe-cific to surface growth and putative new motility and pathogenicity genes.Mol. Microbiol. 52:169–187.
64. Williams, F. D., D. M. Anderson, P. S. Hoffman, R. H. Schwarzhoff, and S.Leonard. 1976. Evidence against the involvement of chemotaxis in swarmingof Proteus mirabilis. J. Bacteriol. 127:237–248.
65. Williams, F. D., and R. H. Schwarzhoff. 1978. Nature of the swarmingphenomenon in Proteus. Annu. Rev. Microbiol. 32:101–122.
66. Yamanaka, K., T. Ogura, H. Niki, and S. Hiraga. 1996. Identification of twonew genes, mukE and mukF, involved in chromosome partitioning in Esch-erichia coli. Mol. Gen. Genet. 250:241–251.
Editor: S. M. Payne
VOL. 78, 2010 TRANSCRIPTOME OF SWARMING PROTEUS MIRABILIS 2845
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 08
Feb
ruar
y 20
22 b
y 20
8.10
7.36
.47.


















