
The mammalian target of rapamycin a cell key modulator of ageing
8
Xavier Viñals González Degree in Biomedical Sciencies Autonomous University of Barcelona ABSTRACT | The target of rapamycin (TOR) is a key modulator of ageing in different organisms, ranging from yeast to rodents, and it is likely that this function has been conserved also in humans (mTOR, mammalian target of rapamycin). The signaling pathway of mTOR senses and integrates a variety of environmental inputs to regulate not only the organismal growth but also its homeostasis. This pathway regulates many important cellular processes that are implicated in the appearance of age-related diseases including cancer, obesity, type 2 diabetes, and neurodegeneration in late life. Here, are been reviewed the recent knowledge in the understanding of the mTOR pathway and its role in ageing and disease. Special emphasis in mTORC1 will be done due to this particular complex can be modulated by some compounds such as rapamycin. Moreover, are going to be discussed pharmacological approaches to treat human pathologies related with mTORC1 deregulation. KEYWORDS TOR, mTOR, mTORC1, rapamycin, ageing, modulation, diseases, life, span, network, signaling. Table of Contents INTRODUCTION 1 RESULTS AND STATE OF THE ART 2 The mTORC1 signaling network 2 Ageing modulation 4 Rapamycin 7 CONCLUSIONS 7 FUTURE PERSPECTIVES 7 ACKNOWLEDGEMENTS 8 REFERENCES 8 INTRODUCTION For a long time, ageing has been conceived as a causal and random process but in the recent years, thanks to research in the field of ageing, it has been shown that this phenomenon can be controlled or at least modulated. Many hypotheses have attempted to explain the evolutionary origin of ageing in order to understand variations in life expectancy of different species. Trying to find an answer to the question of why we age has been one of the main objectives of many scientists working in the field of evolution, but of course it is the answer to the question how we age which can give all of us more information. The Mammalian Target of Rapamycin: A Cell Key Modulator of Ageing Among the many hypotheses and theories that have been formulated in order to understand ageing from an evolutionary standpoint, the most important ones are the mutation accumulation theory and antagonistic pleiotropy theory. The first of these conceives ageing as the result of an accumulation of mutations, that are the main cause of the deposition of cell products leading to deterioration of the body. The second explains that the action of a protein encoded by a particular gene not only participates in its metabolic pathway (main effect), but can also participate in other pathways (pleiotropic effect). According to this theory this gene has been selected by the advantages in the early stages of life but in later stages may cause side effects. Noting life expectancy of different species, which was seen to be significantly different between them, it was thought that ageing could be the result of different factors, which act as regulators and / or modulators that are encoded in the genetic material. The thought there might be genes responsible for ageing, did change the established dogma and since then research has focused on identifying these genes in order to delay ageing and prolong life expectancy. It was in the nineties, when the plasticity of ageing began to be seen as a tangible fact and was discovered that nutrient deprived cells delayed their growth by inhibiting certain proteins. Since then, it has been demonstrated that caloric restriction (defined as the reduction in nutrient intake without causing malnutrition) extends the maximum life span |1
-
Upload
xavier-vinals-gonzalez -
Category
Health & Medicine
-
view
214 -
download
0
Transcript of The mammalian target of rapamycin a cell key modulator of ageing

Xavier Viñals GonzálezDegree in Biomedical ScienciesAutonomous University of Barcelona
ABSTRACT | The target of rapamycin (TOR) is a key modulator of ageing in different organisms, ranging from yeast to rodents, and it is likely that this function has been conserved also in humans (mTOR, mammalian target of rapamycin). The signaling pathway of mTOR senses and integrates a variety of environmental inputs to regulate not only the organismal growth but also its homeostasis. This pathway regulates many important cellular processes that are implicated in the appearance of age-related diseases including cancer, obesity, type 2 diabetes, and neurodegeneration in late life. Here, are been reviewed the recent knowledge in the understanding of the mTOR pathway and its role in ageing and disease. Special emphasis in mTORC1 will be done due to this particular complex can be modulated by some compounds such as rapamycin. Moreover, are going to be discussed pharmacological approaches to treat human pathologies related with mTORC1 deregulation.KEYWORDS TOR, mTOR, mTORC1, rapamycin, ageing, modulation, diseases, life, span, network, signaling.
Table of Contents
INTRODUCTION 1
RESULTS AND STATE OF THE ART 2The mTORC1 signaling network 2Ageing modulation 4Rapamycin 7
CONCLUSIONS 7FUTURE PERSPECTIVES 7ACKNOWLEDGEMENTS 8REFERENCES 8
INTRODUCTION
For a long time, ageing has been conceived as a causal and random process but in the recent years, thanks to research in the field of ageing, it has been shown that this phenomenon can be controlled or at least modulated. Many hypotheses have attempted to explain the evolutionary origin of ageing in order to understand variations in life expectancy of different species. Trying to find an answer to the question of why we age has been one of the main objectives of many scientists working in the field of evolution, but of course it is the answer to the question how we age which can give all of us more information.
The Mammalian Target of Rapamycin: A Cell Key Modulator of Ageing
Among the many hypotheses and theories that have been formulated in order to understand ageing from an evolutionary standpoint, the most important ones are the mutation accumulation theory and antagonistic pleiotropy theory. The first of these conceives ageing as the result of an accumulation of mutations, that are the main cause of the deposition of cell products leading to deterioration of the body. The second explains that the action of a protein encoded by a particular gene not only participates in its metabolic pathway (main effect), but can also participate in other pathways (pleiotropic effect). According to this theory this gene has been selected by the advantages in the early stages of life but in later stages may cause side effects. Noting life expectancy of different species, which was seen to be significantly different between them, it was thought that ageing could be the result of different factors, which act as regulators and / or modulators that are encoded in the genetic material. The thought there might be genes responsible for ageing, did change the established dogma and since then research has focused on identifying these genes in order to delay ageing and prolong life expectancy.
It was in the nineties, when the plasticity of ageing began to be seen as a tangible fact and was discovered that nutrient deprived cells delayed their growth by inhibiting certain proteins. Since then, it has been demonstrated that caloric restriction (defined as the reduction in nutrient intake without causing malnutrition) extends the maximum life span
|1

in some species such as yeast and mice (Martin et al., 2007). In these conditions it can be seen that the metabolic energy investment, which is normally used in anabolic processes (involving growth), is then used in somatic maintenance and catabolic processes (which removes the excess mass) to ensure cell survival (Holliday, 1989). If this is true, the cell must have some kind of sensor that mediates this change, sensitive to the nutritional status of the cell as well as to different growth factors that may be around the cell. One candidate which can perform this function is the target of rapamycin (TOR), since it has proven to be a conserved protein sensitive cellular nutritional status and conserved in many species.
TOR protein was discovered thanks to the previous finding of rapamycin, a fungal inhibitor detected in the soil of Easter Island in 1972. Then several experiments that demonstrated that rapamycin increases the maximum life span of laboratory mice were conducted, but it was not until 2009 when it was reported this compound not only extended the maximum life span a 12% but also increased a third the survival when administered to mice of advanced age (BOX 1). The comprehensive study of rapamycin led to different centers to be set in an ancient mechanism that regulates ageing in different species in which the protein TOR is involved. Today, many studies indicate that inhibition of TOR by rapamycin in late life proves to be beneficial for some species, in which ageing is delayed and their tissues remain longer healthy (Sharp, 2011). The main interest in the field of gerontology is to mimic these effects in humans and studies so far seem to claim that the inhibition of this protein in mammals (mTOR,
BOX1Terms in the literature on ageing
When talking about ageing we ought to differentiate different concepts often used interchangeably. The first is the term life span, which according to the World Health Organization (WHO) refers to the average number of years a person can expect to live, if in the future they experience the current age-specific mortality rates in the population; different from the maximum life span, which is the average from the greatest age reached by the 10% of the population which lives longer. Both terms described differ significantly from the concept of longevity, which usually means long life but should be used in strict sense to describe the length of life of an individual who is still alive. When referring to the average period that an individual may expect to live we must use the term life expectancy at birth, which is an abstract parameter based on demographic data.
mammalian target of rapamycin) mitigates some of the diseases associated with ageing such as type 2 diabetes, osteoporosis, myocardial degeneration and some cancers (Cornu et al., 2013; Gouras et al., 2012), to name a few. Despite all the positive effects it has, rapamycin has been excluded for human trials due to its side effects.
In the early nineties, two TOR genes (named TOR1 and TOR2) were identified and both seemed to be important in regulating growth in yeast. Subsequently the TOR gene was isolated in mammalian and nowadays it has been demonstrated that many species have these genes. TOR genes encode for a cytoplasmatic enzyme which interacts with other proteins to form, in mammals, two complexes named mTORC1 and mTORC2, being only the first one sensitive to the action of rapamycin. The study of these two complexes showed that they not only differ in their structure but also that participate in different pathways. mTORC1 comprises mTOR1 or mTOR2, raptor and mLST8 whereas mTORC2 is more complex and is composed of mTOR2, rictor, mSIN1, PRR5 and mLST8. On the different subunits that form both complexes, raptor has been perhaps the most studied one and it has been suggested that its role is to present various substrates to the mTORC1 catalytic subunit. The role of the other subunits is not known with precision. If only focusing on mTOR, it can be seen that it is a large protein with HEAT domains (huntingtin, elongation factor 3, a subunit of protein phosphatase 2A and TOR1) and FRB domains, both highlighted by several studies since most of the subunits forming mTORC1/C2 interact with them. mTOR complexes develop their functions through their carboxyl terminus portion where a protein kinase domain similar to the phosphatidyl inositol kinase can be found, placing mTOR in a group of protein kinases referred to as the phosphatidylinositol kinase-related kinases (PIKK).
The function of these complexes depends on different signaling that can be both internal (energy state of the cell) or external, as the one it could be given by growth factors and/or nutrients.
RESULTS AND STATE OF THE ART
1. The mTORC1 signaling network
Upstream of mTORC1Cell growth is a complex process that requires coordination not only in time but also in space. In the presence of resources, macromolecules such as proteins and fats are synthesized allowing growth
2|
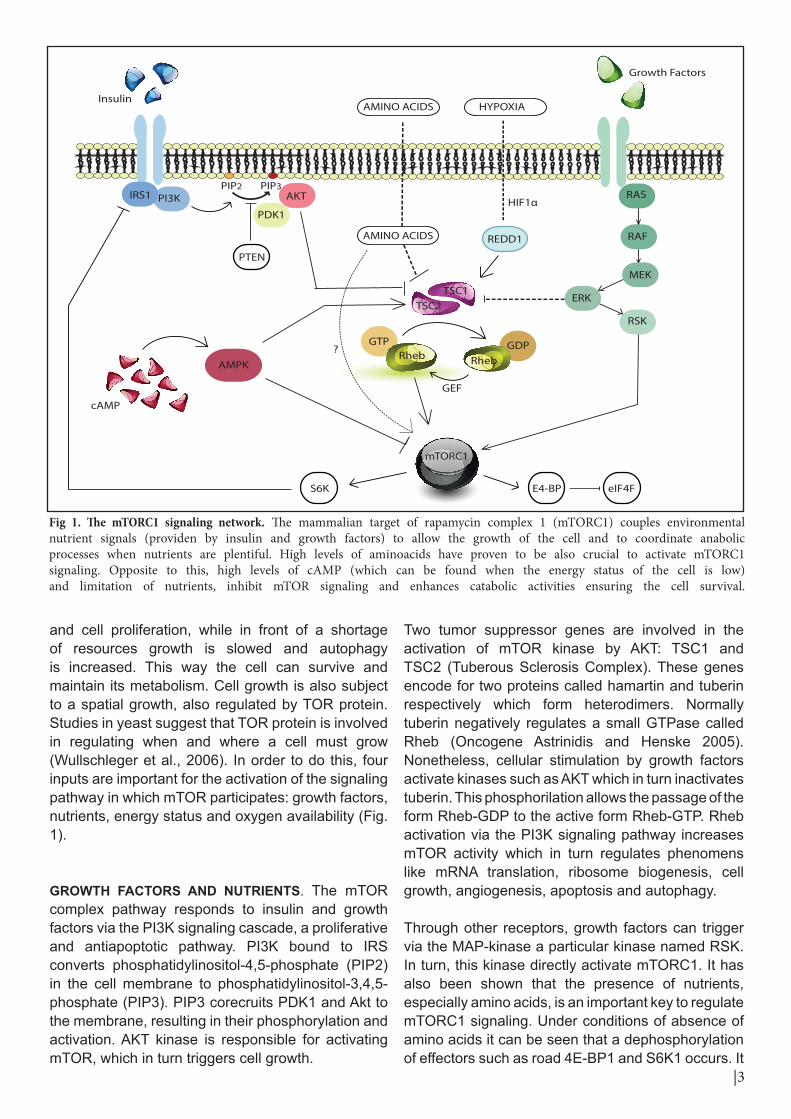
and cell proliferation, while in front of a shortage of resources growth is slowed and autophagy is increased. This way the cell can survive and maintain its metabolism. Cell growth is also subject to a spatial growth, also regulated by TOR protein. Studies in yeast suggest that TOR protein is involved in regulating when and where a cell must grow (Wullschleger et al., 2006). In order to do this, four inputs are important for the activation of the signaling pathway in which mTOR participates: growth factors, nutrients, energy status and oxygen availability (Fig. 1).
GROWTH FACTORS AND NUTRIENTS. The mTOR complex pathway responds to insulin and growth factors via the PI3K signaling cascade, a proliferative and antiapoptotic pathway. PI3K bound to IRS converts phosphatidylinositol-4,5-phosphate (PIP2) in the cell membrane to phosphatidylinositol-3,4,5-phosphate (PIP3). PIP3 corecruits PDK1 and Akt to the membrane, resulting in their phosphorylation and activation. AKT kinase is responsible for activating mTOR, which in turn triggers cell growth.
Two tumor suppressor genes are involved in the activation of mTOR kinase by AKT: TSC1 and TSC2 (Tuberous Sclerosis Complex). These genes encode for two proteins called hamartin and tuberin respectively which form heterodimers. Normally tuberin negatively regulates a small GTPase called Rheb (Oncogene Astrinidis and Henske 2005). Nonetheless, cellular stimulation by growth factors activate kinases such as AKT which in turn inactivates tuberin. This phosphorilation allows the passage of the form Rheb-GDP to the active form Rheb-GTP. Rheb activation via the PI3K signaling pathway increases mTOR activity which in turn regulates phenomens like mRNA translation, ribosome biogenesis, cell growth, angiogenesis, apoptosis and autophagy.
Through other receptors, growth factors can trigger via the MAP-kinase a particular kinase named RSK. In turn, this kinase directly activate mTORC1. It has also been shown that the presence of nutrients, especially amino acids, is an important key to regulate mTORC1 signaling. Under conditions of absence of amino acids it can be seen that a dephosphorylation of effectors such as road 4E-BP1 and S6K1 occurs. It
Fig 1. The mTORC1 signaling network. The mammalian target of rapamycin complex 1 (mTORC1) couples environmental nutrient signals (providen by insulin and growth factors) to allow the growth of the cell and to coordinate anabolic processes when nutrients are plentiful. High levels of aminoacids have proven to be also crucial to activate mTORC1 signaling. Opposite to this, high levels of cAMP (which can be found when the energy status of the cell is low) and limitation of nutrients, inhibit mTOR signaling and enhances catabolic activities ensuring the cell survival.
|3
Insulin
Growth Factors
mTORC1
TSC1
PIP2 PIP3AKT
PDK1
TSC2
PI3K
PTEN
Rheb Rheb
GTP GDP
GEF
cAMP
AMPK
AMINO ACIDS HYPOXIA
AMINO ACIDS
?
REDD1
RAS
RAF
MEK
ERK
RSK
IRS1
S6K E4-BP eIF4F

may be beneficial during ageing, as corroborated by studies in mice with mutations in S6K1 (Selman et al., 2009). The phosphorylation of 4E-BP1 avoids its binding to the cap-binding protein eIF4E, required for the initiation of cap-dependent translation.
The inhibition of autophagy is surely one of the key functions of mTORC1 in promoting longevity. Autophagy is a major degradation pathway in eukaryotic cells that is essential for removing damaged organelles and macromolecules from the cytoplasm and recycling amino acids during periods of starvation (Mizushima et al., 2008). Evidence suggests a decrease in autophagy during ageing, and it has been proposed that this leads to an accumulation of damage (such as aggregated proteins and organelles degeneration). It has been postulated that the active mTORC1 complex is capable of inhibiting autophagy through a serine/threonine kinase known as ULK1. In conditions of resource abundance, ULK1 is inactivated by mTORC1 and autophagy pathway is inhibited, allowing cell growth (Alvers et al., 2009).
The decrease in the function of stem cells in mammals can be a major cause of age-related diseases (reviewed in Sharpless et al., 2007). Again, the mTORC1 appears to have a role in this event and the inhibition of the complex could restore the function of stem cells. It has been shown that inhibition of mTORC1 through the administration of rapamycin to protects old mice from an immune challenge with influenza virus (attributed to a rejuvenation of haematopoietic stem-cell function).
2. Ageing Modulation
Rapamycin increases the life span of yeast and other animals by inhibiting mTORC1 protein (caloric restriction also slows down ageing). Research on the protein function in cells and how its inhibition lengthens life indicated that mTORC1 has two stages along life, one positive and one negative (Fig. 2).
In the early life it is a sensor of nutritional status, vital for the growth and development of the organism. In the presence of abundant food, more insulin and growth factors can be found. The complex reacts to the presence of nutrients and growth factors not only by increasing the synthesis of cellular components but also promoting the cell growth and division. At the same time, mTORC1 gives to the cell the order to decrease autophagy. If the food or energy resources diminish, the complex subsides and the cells focus on keeping itself, not growing or dividing. Furthermore, autophagy increases providing elements and
is hypothesized that high levels of amino acids inhibit complex hamartin / tuberin, allowing the protein Rheb to be active. Other studies suggest that the effects induced via amino acids, is unrelated to these proteins as it would be the mTORC1 complex itself who would act as a sensor.
ENERGY STATUS. Accumulation of cell mass for cell growth requires the synthesis of many molecules. Therefore, mTORC1 must be able to know the energy status of the cell. This enzyme complex is activated in response to an increase in the ratio AMP / ATP. In other words, it is activated in low energy situations through AMPK (AMP-activated protein kinase). When this happens, AMPK performs two functions: it inhibits the phosphorylation of mTORC1 effectors and it phosphorylates TSC1-TSC2 complex, preventing mTORC1 activation.
OXYGEN AVAILABILITY. There are environmental stress situations, such as low availability of oxygen (hypoxia), in which cells are capable of developing a response to survive. When the oxygen concentration in the environment is low, the cell must optimize its resources and stop those synthetic routes that require lots of energy. The hypoxia-inducible factor or HIF1α is a key transcription factor involved in cellular adaptation in hypoxia. In these situations it has been observed an increase in the expression of REDD1 a protein which stimulates the phosphorylation of TSC1/TSC2 complex. When this happens, Rheb remains bound to GDP and does not activate mTORC1.
To sum up, there are different inputs such as growth factors and nutrients, energy status and availability of oxygen which signaling converges in the TSC1/TSC2 complex, which regulates mTORC1 through Rheb.
Downstream of mTORC1It has been well demonstrated that signaling via TOR plays a key role in various processes related to growth in yeast and higher eukaryotes. The best studied effectors of mTOR in mammalian cells are the translational regulators 4E-BP1 (eukaryote initiation factor 4E-binding protein 1) and S6K1 (ribosomal S6 kinase).
Of all the functions mTORC1 has, the most important one is related to the promotion of the mRNA translation and protein synthesis in favourable conditions (Kaeberlein et al., 2011) through one of its effectors, S6K1. As discussed below, the continued function of mTORC1 after the maturity of the individual may be one of the mechanisms of ageing. For this reason, it is proposed that a global reduction of mRNA translation 4|
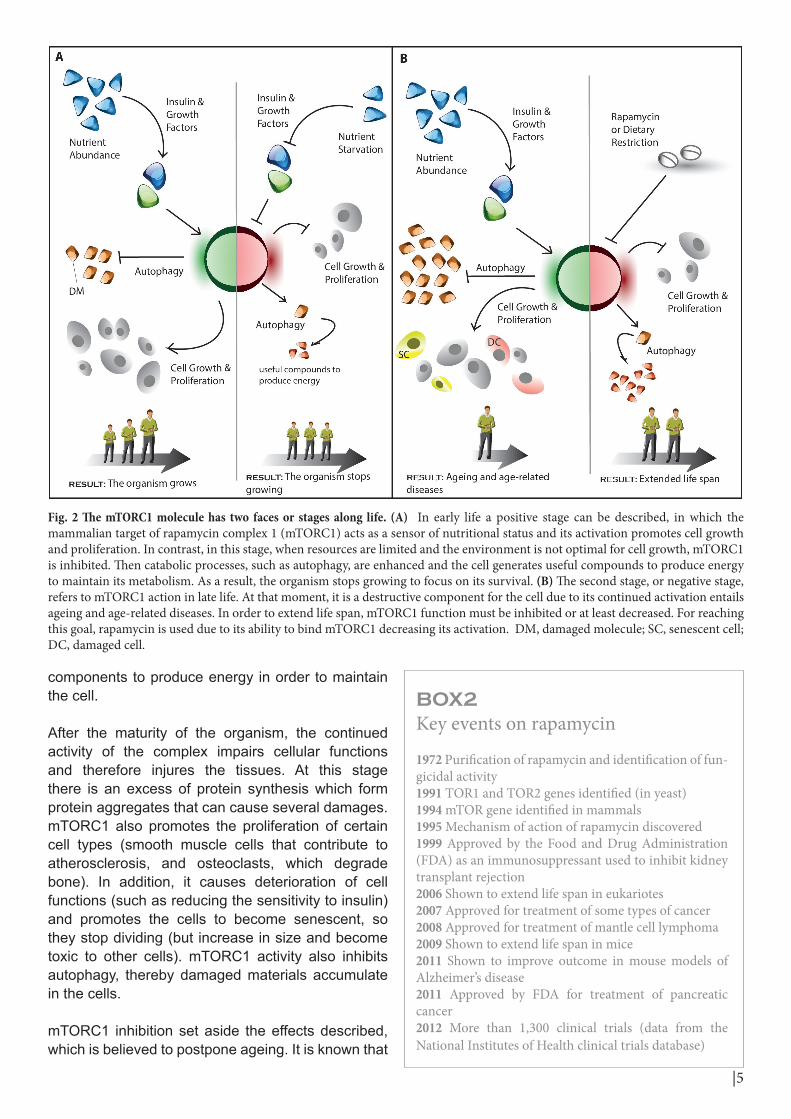
components to produce energy in order to maintain the cell.
After the maturity of the organism, the continued activity of the complex impairs cellular functions and therefore injures the tissues. At this stage there is an excess of protein synthesis which form protein aggregates that can cause several damages. mTORC1 also promotes the proliferation of certain cell types (smooth muscle cells that contribute to atherosclerosis, and osteoclasts, which degrade bone). In addition, it causes deterioration of cell functions (such as reducing the sensitivity to insulin) and promotes the cells to become senescent, so they stop dividing (but increase in size and become toxic to other cells). mTORC1 activity also inhibits autophagy, thereby damaged materials accumulate in the cells.
mTORC1 inhibition set aside the effects described, which is believed to postpone ageing. It is known that
BOX2Key events on rapamycin
1972 Purification of rapamycin and identification of fun-gicidal activity 1991 TOR1 and TOR2 genes identified (in yeast)1994 mTOR gene identified in mammals 1995 Mechanism of action of rapamycin discovered 1999 Approved by the Food and Drug Administration (FDA) as an immunosuppressant used to inhibit kidney transplant rejection 2006 Shown to extend life span in eukariotes2007 Approved for treatment of some types of cancer 2008 Approved for treatment of mantle cell lymphoma2009 Shown to extend life span in mice2011 Shown to improve outcome in mouse models of Alzheimer’s disease2011 Approved by FDA for treatment of pancreatic cancer2012 More than 1,300 clinical trials (data from the National Institutes of Health clinical trials database)
|5
Fig. 2 The mTORC1 molecule has two faces or stages along life. (A) In early life a positive stage can be described, in which the mammalian target of rapamycin complex 1 (mTORC1) acts as a sensor of nutritional status and its activation promotes cell growth and proliferation. In contrast, in this stage, when resources are limited and the environment is not optimal for cell growth, mTORC1 is inhibited. Then catabolic processes, such as autophagy, are enhanced and the cell generates useful compounds to produce energy to maintain its metabolism. As a result, the organism stops growing to focus on its survival. (B) The second stage, or negative stage, refers to mTORC1 action in late life. At that moment, it is a destructive component for the cell due to its continued activation entails ageing and age-related diseases. In order to extend life span, mTORC1 function must be inhibited or at least decreased. For reaching this goal, rapamycin is used due to its ability to bind mTORC1 decreasing its activation. DM, damaged molecule; SC, senescent cell; DC, damaged cell.

BOX3Relationship between mTORC1 and age-related diseasesStudies in animals and in humans suggest that some disorders, such as Alzheimer’s disease, cancer and heart disease are improved by administration of rapamycin. However, in other disorders, the evidence suggests that there may be both beneficial and adverse consequences to the inhibition of mTORC1.
•Neurodegenerative diseases: These diseases, such as Parkinson disease, Alzheimer disease, Huntington disease or amyotrophic lateral sclerosis, are related with permanent loss of both neuronal structure and neuronal functions. One of the key processes in many neurodegenerative diseases is the accumulation of elements in neurons, which normally are degraded by the process of autophagy. mTORC1 is an important regulator of autophagy and its continuous activation result in the deposition of protein aggregates in cells. mTORC1 inhibition by rapamycin activates the autophagy of these aggregates in vitro, reducing the severity of the disease (reviewed in Sarkar and Rubinsztein).
•Type 2 diabetes: This type of diabetes is characterized by insulin resistance by tissues, coursing with hyperglycemia and hyperlipidemia. One of the effectors of mTORC1, S6K, has a substrate-inactivating function of the insulin receptor (IRS1), responsible for activating the PI3K pathway in the presence of insulin and insulin-like growth factors (IGF-1). The continuous mTORC1 signaling pathway involves a long-term sensibilization to insulin by the cells (Um et al., 2004). Although insulin resistance is reduced by rapamycin administration to treat type 2 diabetes, rapamycin induces a diabetes like syndrome by impairing the function of the muscles, liver, adipose tissue, and pancreatic β-cells.
•Heart disease: cardiac hypertrophy, is a risk factor for cardiac morbidity and mortality. It is hypothesized that the mTORC1 complex plays a key role due to it has been seen that inhibition by rapamycin, has shown to attenuate cardiac hypertrophy (Shioi et al., 2003). Rapamycin administration has also been linked to high levels of triglycerides in the blood, which can lead to heart disease.
•Metabolic disease: chronic administration of rapamycin impairs glucose tolerance and insulin action. It has been demonstrated that rapamycin disrupt (not directly) a second mTOR complex, mTORC2, in vivo and that mTORC2 was required for the insulin-mediated suppression of hepatic gluconeogenesis (Ricoult and Manning, 2013).
•Cancer: The mTORC1 complex is active in many types of cancer (reviewed in Dazert, 2011). Many components of the PI3K pathway, which we have seen that regulates the activity of mTORC1, are mutated in human cancers. Mutations in the tumor supressor genes (green) encoding TSC1, TSC2, PTEN and Nf1 (neurofibromatosis type 1) are some examples and all of them result in activation of mTORC1. Furthermore, loss of p53 (common in neoplastic processes) promotes the activation of mTORC1 complex (Guertin and Sabatini, 2007). Mutations in proteins that act as oncogens (red) such as Akt and PI3K can stimulate cell growth and proligeration through mTORC1 activation.
•Kidney disease: mTOR plays an important role in a number of renal diseases, including acute kidney injury, diabetic nephropathy, and polycystic kidney diseases. The activity of mTORC1 is necessary for renal regeneration and repair and inhibition of mTORC1 by rapamycin has been shown to prolong delayed graft rejection in humans who have received a kidney transplant (Liu, 2006).
6|
mTORC1
Autophagy is decreased
Protein aggrega-tes and inclusion body formation
DISEASE
mTORC1 activa-tion in late life
mTORC1
Cell Growth isstimulated
CARDIAC HIPERTROPHY DISEASE
+ high levels oftriglycerides
mTORC1
mTORC1 conti-nuous activation
S6K
PI3KIRS1
DISEASE
sensibilization to insulin
mTORC1
mTORC1 conti-nuous activation
CELL GROWTH AND PROLIFERATION
PTENTSC1/2 p53
AKT PI3K
Ras Raf
Nf1
mTORC1
mTORC1 activation
Renal regenera-tion in injured
kidney
Rapamycin
Proliferation and cell growth stops
No graftrejection

ageing is the cause of the onset and/or progression of multiple disorders, which in turn are modulated by mTORC1 (Box 3). Note than in early human trials, rapamycin showed to have serious side effects. Due to it is an immunosuppressant, it can make patients susceptible to opportunistic infections.
3. Rapamycin
After having discovered the mechanism of action of rapamycin, it was approved its use for renal transplant patients to prevent rejection (BOX 2). In these patients rapamycin was proved to be a protective element for certain types of cancer (Liu, 2006) and an attenuator of the effects of some types of existing tumors. As the body mass index in these patients years after transplantation was significantly lower, it can be thought that rapamycin may prevent obesity. Moreover, improvement of cognitive function in adult mice under the administration of rapamycin is shown in some studies (Halloran et al., 2012).
As seen in this study, rapamycin antagonizes and/or modulates the mTORC1 effects on the organism. But why is mTORC1 sensitive to rapamycin whereas mTORC2 is not? The answer lies in the structure of the complexes and the FRB domain, which has previously been cited. This domain acts as a docking site for FKBP12-rapamycin complex. FKBP12 is a proline isomerase that in the absence of rapamycin is free within the cell. In the presence of rapamycin, this isomerase is kidnapped by this compound and the union of these two elements form a complex capable of inhibiting mTORC1. In the mTORC2 complex the FRB domain is masked by one of the subunits of the same complex, making it resistant to the action of rapamycin.Side effects of rapamycin are the reason why this compound is not used in humans due to it increases blood cholesterol levels, cause anemia and delayed wound healing, within other alterations.
Metformin, the most prescribed treatment for diabetes which is useful for reducing blood glucose concentration, might be an alternative (Gallagher and LeRoith, 2011). Its mechanism of action is not fully understood, but it is known to inhibit the TOR pathway in mice and it activates another enzyme that may be related to ageing, AMPK, which is also stimulated by caloric restriction. It has also been shown that metformin emulates the effect of caloric restriction on gene activity in mice, and some studies suggest that it could increase the maximum life span in rodents. Many studies need to be done in order to know if metformin has the same effects that an
hypocaloric diet in humans.
CONCLUSIONS
Achieving an increase to human longevity like the one rapamycin induces to mice would add an average of five to ten years to a person’s life. As mortality has declined during the early stages of life, further extend of the life span implies we must fight against age-related diseases. The development of drugs able to slow ageing could be used as preventive medicine to postpone common diseases of ageing (such as dementia, osteoporosis, cancer or diabetes), just as nowadays medications that reduce blood pressure and lower cholesterol reduce the number of heart attacks in middle-aged people. They would provide us a time of life-quality and prolong the period of vitality before the body starts to weak and die.
Due to all this, mTOR is becoming one of the candidates for mediating the effects of life span extension just as like dietary restriction does. All data emphasize the importance of mTORC1 signaling in mediating life span extension through species. As mentioned before, mTORC1 is essential in early life for the development in many species. Opposite to this, inhibition of this pathway during late life extends life span. This behaviour is consistent with the predictions of the theory of antagonistic pleiotropy, which suggests that some genes which are important for the development early in life limit life span during adulthood. Knowing this, there are many challenges in the field and many groups have now been able to understand and link different processes downstream of this molecule. It is far unknown the possible cross talk of mTORC1 with other ageing pathways. Another challenge is to describe the contribution of each of the effectors of mTORC1 and the different inputs that have a great influence in determining life span across species.
FUTURE PERSPECTIVES
In order to understand the role of mTORC1 signaling in coordinating overall body growth, further studies of the role in individual tissues are required. Precisely, it will be not only interesting but also important to examine its role in tissues that are most sensitive to insulin and nutrients such as adipose tissue, muscle and liver.
When talking about drugs for treating diseases nothing is more important that their dose. Very little information exists on the most effective dose of rapamycin for longevity or healthspan in mice.
|7

To avoid the side effects of rapamycin, newer ATP-competitive inhibitors of mTOR kinase activity have been developed and these can block the phosphorylation of all known substrates of both mTORC1 and mTORC2.
The search for a way to delay human ageing has proven to be long and difficult to reach. Although still far from reality, there is reason to be optimistic that inhibitors of mTOR may be helpful to accomplish this goal. mTORC1 inhibition has proven to slow ageing in yeast and invertebrates, extends lifespan in mice, and has an impact on several age-related diseases. Only time will tell, but research in mTOR is going to be very promising and interesting in the next years.
ACKNOWLEDGEMENTS
I would like to thank Anna Genescà not only for her task in being my tutor but also for her task on giving me the basic guidelines for developing the present review.
REFERENCES
Alvers, A.L., Wood, M.S., Hu, D., Kaywell, A.C., Dunn, W.A.,Jr & Aris, J.P. 2009, “Autophagy is required for extension of yeast chronological life span by rapamycin”, Autophagy, vol. 5, no. 6, pp. 847-849.
Cornu, M., Albert, V. & Hall, M.N. 2013, “mTOR in aging, metabolism, and cancer”, Current opinion in genetics & development,.
Dazert, E. & Hall, M.N. 2011, “mTOR signaling in disease”, Current opinion in cell biology, vol. 23, no. 6, pp. 744-755.Gallagher, E.J. & LeRoith, D. 2011, “Diabetes, cancer, and metformin: connections of metabolism and cell proliferation”, Annals of the New York Academy of Sciences, vol. 1243, pp. 54-68.
Gouras, G.K. 2012, “mTOR: at the crossroads of aging, chaperones, and Alzheimer’s disease”, Journal of neurochemistry, .
Guertin, D.A. & Sabatini, D.M. 2007, “Defining the role of mTOR in cancer”, Cancer cell, vol. 12, no. 1, pp. 9-22.
Halloran, J., Hussong, S.A., Burbank, R., Podlutskaya, N., Fischer, K.E., Sloane, L.B., Austad, S.N., Strong, R., Richardson, A., Hart, M.J. & Galvan, V. 2012, “Chronic inhibition of mammalian target of rapamycin by rapamycin modulates cognitive and non-cognitive components of behavior throughout lifespan in mice”, Neuroscience, vol. 223, pp. 102-113.
Holliday, R. 1989, “Food, reproduction and longevity: is the extended lifespan of calorie-restricted animals an evolutionary adaptation?”, BioEssays : news and reviews in molecular,
cellular and developmental biology, vol. 10, no. 4, pp. 125-127.
Kaeberlein, M. & Kennedy, B.K. 2011, “Hot topics in aging research: protein translation and TOR signaling, 2010”, Aging cell, vol. 10, no. 2, pp. 185-190.
Liu, Y. 2006, “Rapamycin and chronic kidney disease: beyond the inhibition of inflammation”, Kidney international, vol. 69, no. 11, pp. 1925-1927.
Martin, B., Golden, E., Egan, J.M., Mattson, M.P. & Maudsley, S. 2007, “Reduced energy intake: the secret to a long and healthy life?”, IBS journal of science, vol. 2, no. 2, pp. 35-39.
Mizushima, N., Levine, B., Cuervo, A.M. & Klionsky, D.J. 2008, “Autophagy fights disease through cellular self-digestion”, Nature, vol. 451, no. 7182, pp. 1069-1075.Ricoult, S.J. & Manning, B.D. 2013, “The multifaceted role of mTORC1 in the control of lipid metabolism”, EMBO reports, .
Sarkar, S. & Rubinsztein, D.C. 2008, “Small molecule enhancers of autophagy for neurodegenerative diseases”, Molecular bioSystems, vol. 4, no. 9, pp. 895-901.
Selman, C., Tullet, J.M., Wieser, D., Irvine, E., Lingard, S.J., Choudhury, A.I., Claret, M., Al-Qassab, H., Carmignac, D., Ramadani, F., Woods, A., Robinson, I.C., Schuster, E., Batterham, R.L., Kozma, S.C., Thomas, G., Carling, D., Okkenhaug, K., Thornton, J.M., Partridge, L., Gems, D. & Withers, D.J. 2009, “Ribosomal protein S6 kinase 1 signaling regulates mammalian life span”, Science (New York, N.Y.), vol. 326, no. 5949, pp. 140-144.
Sharp, Z.D. 2011, “Aging and TOR: interwoven in the fabric of life”, Cellular and molecular life sciences : CMLS, vol. 68, no. 4, pp. 587-597.
Sharpless, N.E. & DePinho, R.A. 2007, “How stem cells age and why this makes us grow old”, Nature reviews.Molecular cell biology, vol. 8, no. 9, pp. 703-713.
Shioi, T., McMullen, J.R., Tarnavski, O., Converso, K., Sherwood, M.C., Manning, W.J. & Izumo, S. 2003, “Rapamycin attenuates load-induced cardiac hypertrophy in mice”, Circulation, vol. 107, no. 12, pp. 1664-1670.
Um, S. H. et al. 2004, “Absence of S6K1 protects against age- and diet-induced obesity while enhancing insulin sensitivity”. Nature 431, 200–205.
Wullschleger, S., Loewith, R. & Hall, M.N. 2006, “TOR signaling in growth and metabolism”, Cell, vol. 124, no. 3, pp. 471-484.
8|


















