
The Effect of Low Density Lipoproteins, Cholesterol, and 25
10
THE JOURNAL 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc. OF BIOLOGICAL CHEMISTRY Vol. 267, No. 10, hue of April 5, pp. 7160-7169,1992 Printed in U.S.A. The Effect of Low Density Lipoproteins, Cholesterol, and ” 25-Hydroxycholesterol on Apolipoprotein B Gene Expression in HepG2 Cells* (Received for publication, September 23, 1991) Nassrin Dashti$ From the Lipoprotein and Atherosclerosis Research Program, Oklahoma Medical Research Foundation, Oklahoma City, Oklahoma 73104 The purpose of the present study was to examine the effects of exogenous cholesterol on the apolipoprotein (Apo) B gene expression in HepG2 cells. Pure choles- terol had no significant effect on either the cellular content of cholesteryl esters or the net accumulation of neutral lipids and ApoB in the culture medium. By contrast, addition of 26-hydroxycholestero1 increased the net accumulation of cholesteryl esters incells and medium by 2-3-fold and decreased that of unesterified cholesterol by 60% in both compartments. A 33% re- duction in the cellular content of triglycerides was commensurate with a 40% increase in their accumu- lation in the medium. A significant %fold increase in the net accumulation of ApoB in themedium was pre- dominantly due to enhanced secretion of newly synthe- sized ApoB as established by pulse-chase studies. The stimulation in ApoB secretion was accompanied by a 66% increase in cellular ApoBmRNA. Under these experimental conditions, the low density lipoprotein receptor activity wasdecreased by only 12420%. Ad- dition of progesteron prevented the effects of 26-hy- droxycholesterol. The changes in theconcentration of neutral lipids and ApoB were reflected in the compo- sition of secreted “low-density” lipoproteins. These particles had increased percentage contents of choles- teryl esters and ApoB and a decreased percentage con- tent of unesterified cholesterol in comparison with li- poproteins produced by control cells. The rate of ApoB production was not correlated with the triglyceride mass in the cells but was positively correlated with the cellular and secreted cholesteryl esters and secreted triglycerides. With the exception of unchanged cellular unesterified cholesterol and ApoB mRNA levels, plasma low density lipoprotein had similar, although less pronounced, effects on the production of neutral lipids and ApoB. These results demonstrate that in HepG2 cells the synthesis and secretion of ApoB and cholesteryl esters are tightly coupled and that 26-hy- droxycholesterol increased the concentration of ApoB- containing lipoproteins primarily by stimulating their production rather thanreducing their catabolism. * This investigation was supported by Research Grant HR7-032 from the Oklahoma Health Research Program and, in part, by Re- search Grant HL-23181 from the National Institutes of Health and by the resources of Oklahoma Medical Research Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This articlemust therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ To whom correspondence should be addressed Lipoprotein and Atherosclerosis Research Program, Oklahoma Medical Research Foundation, 825 NE 13th St., Oklahoma City, OK 73104. It is well established that the elevated concentration of LDL’ cholesterolin circulation is one of the major risk factors for the development of coronary artery disease (1). The con- centration of lipoproteins, including LDL, in the plasma is markedly influenced by nutritional and hormonal manipula- tions (2, 3). It is generally agreed that in humans increased dietary cholesterol and saturated fat result in elevated LDL cholesterol and that this is due to enhanced production and/ or decreased catabolism of circulating LDL particles (4,5). In addition to plasma level of LDL cholesterol (l), the concen- tration of ApoB correlates directly with the incidence of coronary heart disease (6). ApoB exists in two immunologi- cally distinct forms, ApoB-100 and ApoB-48 (7). In humans, ApoB-100 is the only molecular form that is synthesized by the liver (8). ApoB-100 is an essential structural component of very low density lipoproteins (VLDL) and LDL and is required for their formation and secretion (8). ApoB-100 also serves as a ligand for the uptake of LDL by a variety of cells via the LDL receptor (9-11). Whereas the disorders in LDL concentration due to their impaired catabolism through the LDL receptor pathway is well characterized ( l l ) , the nature of the mechanism(s) involved in overproduction of LDL- ApoB remains unresolved. Therefore, due to its potentially crucial role as a positive risk factor in the development of atherosclerosis,an assessment of factors, including dietary fat and cholesterol, which regulate the synthesis and secretion of ApoB-containinglipoprotein particles, is of great clinical im- portance. The role of fatty acids as modulators of hepatic and intes- tinal synthesis and secretion of ApoB-containinglipoprotein particles have been studied in this laboratory (12-14). The results of these studies demonstrated that the addition of oleate to HepG2 cells caused a 2-4-fold increase in the net accumulation of triglycerides in the medium and only a 30% elevation in that of ApoB (12, 13). These studies thus dem- onstrated a lack of coordinate induction in the secretion of triglycerides and ApoB and suggested that fatty acid alone was not sufficient to stimulate ApoB synthesis and secretion. Furthermore, the similarity between the oleate-induced in- crease in the net accumulation of cholesterylesters (55%)and ApoB (30%) in the medium indicated that the rate of ApoB production might be regulated by cholesterylester rather than triglyceride content of the cells. The present study was de- signed to test this hypothesis and to assess the role of exoge- nous cholesterol in this process. The abbreviations used are: LDL, low density lipoproteins; Apo, apolipoprotein; VLDL, very low density lipoproteins; IDL, interme- diate density lipoproteins; MEM, minimum essential medium; FBS, fetal bovine serum; PBS, phosphate-buffered saline; SDS, sodium dodecyl sulfate; HMG-CoA, 3-hydroxy-3-methylglutaryl coenzyme A. 7160
Transcript of The Effect of Low Density Lipoproteins, Cholesterol, and 25

THE JOURNAL 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc.
OF BIOLOGICAL CHEMISTRY Vol. 267, No. 10, h u e of April 5, pp. 7160-7169,1992 Printed in U.S.A.
The Effect of Low Density Lipoproteins, Cholesterol, and ”
25-Hydroxycholesterol on Apolipoprotein B Gene Expression in HepG2 Cells*
(Received for publication, September 23, 1991)
Nassrin Dashti$ From the Lipoprotein and Atherosclerosis Research Program, Oklahoma Medical Research Foundation, Oklahoma City, Oklahoma 73104
The purpose of the present study was to examine the effects of exogenous cholesterol on the apolipoprotein (Apo) B gene expression in HepG2 cells. Pure choles- terol had no significant effect on either the cellular content of cholesteryl esters or the net accumulation of neutral lipids and ApoB in the culture medium. By contrast, addition of 26-hydroxycholestero1 increased the net accumulation of cholesteryl esters in cells and medium by 2-3-fold and decreased that of unesterified cholesterol by 60% in both compartments. A 33% re- duction in the cellular content of triglycerides was commensurate with a 40% increase in their accumu- lation in the medium. A significant %fold increase in the net accumulation of ApoB in the medium was pre- dominantly due to enhanced secretion of newly synthe- sized ApoB as established by pulse-chase studies. The stimulation in ApoB secretion was accompanied by a 66% increase in cellular ApoB mRNA. Under these experimental conditions, the low density lipoprotein receptor activity was decreased by only 12420%. Ad- dition of progesteron prevented the effects of 26-hy- droxycholesterol. The changes in the concentration of neutral lipids and ApoB were reflected in the compo- sition of secreted “low-density” lipoproteins. These particles had increased percentage contents of choles- teryl esters and ApoB and a decreased percentage con- tent of unesterified cholesterol in comparison with li- poproteins produced by control cells. The rate of ApoB production was not correlated with the triglyceride mass in the cells but was positively correlated with the cellular and secreted cholesteryl esters and secreted triglycerides. With the exception of unchanged cellular unesterified cholesterol and ApoB mRNA levels, plasma low density lipoprotein had similar, although less pronounced, effects on the production of neutral lipids and ApoB. These results demonstrate that in HepG2 cells the synthesis and secretion of ApoB and cholesteryl esters are tightly coupled and that 26-hy- droxycholesterol increased the concentration of ApoB- containing lipoproteins primarily by stimulating their production rather than reducing their catabolism.
* This investigation was supported by Research Grant HR7-032 from the Oklahoma Health Research Program and, in part, by Re- search Grant HL-23181 from the National Institutes of Health and by the resources of Oklahoma Medical Research Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed Lipoprotein and Atherosclerosis Research Program, Oklahoma Medical Research Foundation, 825 NE 13th St., Oklahoma City, OK 73104.
It is well established that the elevated concentration of LDL’ cholesterol in circulation is one of the major risk factors for the development of coronary artery disease (1). The con- centration of lipoproteins, including LDL, in the plasma is markedly influenced by nutritional and hormonal manipula- tions (2, 3). It is generally agreed that in humans increased dietary cholesterol and saturated fat result in elevated LDL cholesterol and that this is due to enhanced production and/ or decreased catabolism of circulating LDL particles (4,5). In addition to plasma level of LDL cholesterol (l), the concen- tration of ApoB correlates directly with the incidence of coronary heart disease (6). ApoB exists in two immunologi- cally distinct forms, ApoB-100 and ApoB-48 (7). In humans, ApoB-100 is the only molecular form that is synthesized by the liver (8). ApoB-100 is an essential structural component of very low density lipoproteins (VLDL) and LDL and is required for their formation and secretion (8). ApoB-100 also serves as a ligand for the uptake of LDL by a variety of cells via the LDL receptor (9-11). Whereas the disorders in LDL concentration due to their impaired catabolism through the LDL receptor pathway is well characterized ( l l ) , the nature of the mechanism(s) involved in overproduction of LDL- ApoB remains unresolved. Therefore, due to its potentially crucial role as a positive risk factor in the development of atherosclerosis, an assessment of factors, including dietary fat and cholesterol, which regulate the synthesis and secretion of ApoB-containing lipoprotein particles, is of great clinical im- portance.
The role of fatty acids as modulators of hepatic and intes- tinal synthesis and secretion of ApoB-containing lipoprotein particles have been studied in this laboratory (12-14). The results of these studies demonstrated that the addition of oleate to HepG2 cells caused a 2-4-fold increase in the net accumulation of triglycerides in the medium and only a 30% elevation in that of ApoB (12, 13). These studies thus dem- onstrated a lack of coordinate induction in the secretion of triglycerides and ApoB and suggested that fatty acid alone was not sufficient to stimulate ApoB synthesis and secretion. Furthermore, the similarity between the oleate-induced in- crease in the net accumulation of cholesteryl esters (55%) and ApoB (30%) in the medium indicated that the rate of ApoB production might be regulated by cholesteryl ester rather than triglyceride content of the cells. The present study was de- signed to test this hypothesis and to assess the role of exoge- nous cholesterol in this process.
The abbreviations used are: LDL, low density lipoproteins; Apo, apolipoprotein; VLDL, very low density lipoproteins; IDL, interme- diate density lipoproteins; MEM, minimum essential medium; FBS, fetal bovine serum; PBS, phosphate-buffered saline; SDS, sodium dodecyl sulfate; HMG-CoA, 3-hydroxy-3-methylglutaryl coenzyme A.
7160

Regulation of ApoB by Cholesterol in HepG2 Celb 7161
Cholesterol-rich diets have been shown to result in a marked hypercholesterolemia and increased serum concentra- tion of apolipoproteins B and E in a variety of species (15). Both increased synthetic rate and decreased fractional cata- bolic rate of LDL-ApoB have been implicated as the mecha- nisms involved in this process (3). The purpose of the present study was to investigate the regulatory role of exogenous cholesterol, supplied as pure cholesterol, LDL cholesterol, and 25-hydroxycholestero1, in the synthesis and secretion of ApoB-containing lipoproteins by human liver using HepGZ cells as an experimental model. Alterations in the levels of cellular mRNA for apolipoproteins and modifications in the LDL receptor activity were followed concurrently. In addition, the relationship between the neutral lipid composition within the cells and the rate of ApoB production was assessed. Initial results showed that 25-hydroxycholestero1 was highly effec- tive in causing a simultaneous increase in the net accumula- tion of cholesteryl esters and ApoB in the culture medium. This observation provided a basis for further studies designed to elucidate the regulatory mechanism(s) involved in this process. Part of this work has been reported in abstract form (16).
EXPERIMENTAL PROCEDURES
Materials-Minimum essential medium (MEM), trypsin, sodium pyruvate, L-glutamine, and MEM vitamin solution were purchased from GIBCO. Fetal bovine serum (FBS) was obtained from Whittaker Bioproducts (Walkersville, MD). Triton X-100, benzamidine, phenyl- methylsulfonyl fluoride, leupeptin, bovine serum albumin, progester- one, cholesterol, and 25-hydroxycholestero1 were purchased from Sigma. ~-[~'S]Methionine, [32P]dCTP, and Amplify were obtained from Amersham Corp. Protein A-Sepharose CL-4B was from Phar- macia LKB Biotechnology Inc. All reagents used for the slab gel electrophoresis were purchased from Bio-Rad.
Cell Culture-The human hepatoblastoma cell line HepG2 was obtained from American Type Culture Collection (Rockville, MD). Cells were seeded into 100-mm diameter dishes in 10 ml of MEM supplemented with 2 mM glutamine, 1 mM sodium pyruvate, MEM vitamin solution, and 10% FBS and incubated at 37 "C in a 95% air, 5% C02 atmosphere as previously described (12, 13). In all experi- ments the medium was changed 48 h after plating unless otherwise stated.
Four days after seeding, the maintenance medium was removed, monolayers were washed twice with phosphate-buffered saline (PBS), and serum-free MEM containing the test substrate was added to each dish. Cholesterol and 25-hydroxycholestero1 were dissolved in ethanol; the same concentration of ethanol was added to the control cells. At the end of incubation, the conditioned medium was removed and, to prevent oxidative and proteolytic damage, preservative mix- ture (13) was added to obtain the final concentrations of 500 units/ ml penicillin-G, 50 pg/ml streptomycin sulfate, 20 pg/ml chloram- phenicol, 1.3 mg/ml €-amino caproic acid, 1 mg/ml EDTA, and 0.2 mg/ml glutathione. The medium was centrifuged at 2000 rpm for 30 min at 4 "C to remove small amounts of cells and debris. The supernatant fraction was concentrated approximately 10-15-fold with either polyvinylpyrrolidone or sucrose placed outside the dialysis bag (5000 molecular weight cut-off). The monolayers were washed three times with PBS, scraped off the plate after addition of 2 X 2 ml of PBS, and sonicated. Aliquots of cell suspension were analyzed for cellular neutral lipids and protein. Under the present experimental conditions, addition of ethanol (0.25-0.5% final concentration) and/ or exogenous cholesterol did not affect the cell growth and number, as judged by unchanged cellular protein content.
Isolation of Lipoprotein Density Classes-LDL (d 1.03-1.063 g/ml) was isolated from plasma of normolipidemic subjects after an over- night fast. The lipoproteins accumulated in the cell culture medium were separated into "low density" and "high density" lipoproteins by a single ultracentrifugation at the density of 1.063 g/ml (17). In studies on the binding and uptake of nascent lipoproteins, culture medium was fractionated into VLDL + intermediate-density lipopro- teins (IDL) (d < 1.03 g/ml) and LDL (d 1.03-1.063 g/ml). Plasma LDL was washed and dialyzed as previously described (10). Lipopro- teins isolated from the culture medium were dialyzed against PBS
and concentrated 10-15-fold as described above. Determination of Neutral Lipids and Apolipoproteim-The concen-
tration of triglycerides, unesterified cholesterol, and cholesteryl esters were determined by gas-liquid chromatography (18). The concentra- tions of apolipoproteins A-I, B, C-111, and E were measured by electroimmunoassays developed in this laboratory as previously de- scribed (19). Cell protein was measured by the method of Lowry et al. (20).
Antibodies-Monospecific polyclonal antisera to apolipoproteins A-I, B, C-111, and E have been used in this laboratory for determi- nation of the corresponding apolipoprotein concentration and for immunoprecipitation and immunoblotting analyses (14, 17, 21). The specificity of antisera was tested by double-diffusion analysis, cross- immunoelectrophoresis, and electroimmunoassays (19). On the basis of these tests, all antisera were found to be monospecific.
Pulse-Chase Studies of ApoB Synthesis and Secretion-HepG2 cells were grown in 35-mm diameter dishes in supplemented MEM for 4 days. The maintenance medium was removed and monolayers were washed twice with serum-free medium and preincubated for 3 h in the same medium containing either ethanol or 25-hydroxycholestero1 (dissolved in ethanol) added at 20 pg/ml of medium. The preincuba- tion medium was removed, cells were washed three times with serum- free, methionine-free medium, and pulse-labeled by incubation with 1 ml of methionine-free medium containing 100 pCi of [35S]methio- nine; ethanol and 25-hydroxycholesterol were present in appropriate dishes during the pulse period. After 10 min, the medium from each dish was rapidly removed, monolayers were washed twice, and chased for 10, 20, 40, 60, 90, and 120 min at 37 "C in MEM containing 10 mM unlabeled methionine and either ethanol or 25-hydroxycholes- terol as above. After each chase period, medium was removed, cells were washed three times with cold PBS, and immediately lysed according to the method of Bostrom et al. (22) using solubilization buffer described by Sparks and Sparks (23). Aliquots of medium and cell lysate were analyzed for 35S-labeled ApoB by immunoprecipita- tion followed by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and autofluorography as described later in this sec- tion.
Immunoprecipitation and SDS-Polyacrylamide Gel Electrophore- sis-The [35S]methionine-labeled ApoB in cell lysate and secreted into the chase medium were immunoprecipitated using monospecific antibodies to human ApoB as described in the following example. Protein A-Sepharose CL-4B was washed according to the manufac- turer's instructions. Twenty-five mg of washed protein A-Sepharose was placed in 1.5-ml microcentrifuge tubes and resuspended in 0.5 ml of buffer A (0.01 M Tris-HC1, pH 7.5,0.15 M NaCl, 0.1% (w/v) bovine serum albumin, 0.5% (w/v) Triton X-100, 0.1% (w/v) SDS, and 0.2% (w/v) sodium azide), and 10 pl of goat antiserum to human ApoB was added. The tubes were rotated at room temperature for 1 h. The unbound antibody was removed by centrifugation at 1000 rpm for 2- 3 min. The supernatant was discarded and the protein A-Sepharose- antibody complex was washed six times with buffer A. Aliquots (0.5 ml) of medium or cell lysate were added to the pelleted gels and rotated at 4 "C overnight. The mixtures were centrifuged as above and the supernatants were discarded. The gels were washed three times with buffer A and three times with buffer B (same as buffer A but without bovine serum albumin), and immunoprecipitates were collected by centrifugation as above. Labeled ApoB was eluted in 100 pl of elution buffer (0.125 M Tris-HC1, pH 6.8, 4% (w/v) SDS, 20% (v/v) glycerol, 10% (v/v) 2-mercaptoethanol, and 0.02% (w/v) brom- phenol blue) by heating twice at 100 "C for 5 min. The eluted 35S- labeled ApoB was applied to a 2.5-15% polyacrylamide gradient slab gel containing SDS using the Laemmli buffer system (24). After electrophoresis, the gels were stained, destained (24), and enhanced by agitation in Amplify. The enhanced gels were dried and fluoro- graphed at -85 "C using Kodak X-OMAT film.
Lipoprotein Binding, Uptake, and Degradation Assays-In this series of experiments, cells were seeded into 16-mm diameter multi- well dishes in complete MEM as described above. Plasma LDL and nascent VLDL + IDL and LDL were iodinated with Na'251 using Iodo-Beads according to the method of Markwell (25), and free iodide was removed by dialysis as described (10). The properties of Iz6I- labeled VLDL and LDL were similar to those reported previously (10,261. The binding at 4 'C and uptake and degradation at 37 "C of '251-labeled VLDL and LDL were performed under conditions de- scribed in detail elsewhere (10,26). Control dishes without cells were incubated under identical conditions. In all experiments the binding was expressed in terms of radioactivity associated with the cells at 4 "C. The uptake was expressed in terms of '''1 associated with the

7162 Regulation of ApoB by Cholesterol in HepG2 Cells cells at 37 "C while degradation was measured in the medium after precipitation of protein with trichloroacetic acid and removal of free iodide (10, 26).
Isolation and Characterization of Cellular RNA, Preparation of Probes, and DNA-excess Solution Hybridization Assays-RNA was isolated from HepG2 cells by the guanidine thiocyanate method (27). The integrity of all RNA samples was verified by electrophoresis in 1.2% agarose gels containing 2.2 M formaldehyde (28). Hybridization probes were prepared from human apolipoproteins A-I, ApoB-100 3' and 5', and LDL receptor cDNA fragments subcloned in bacterio- phage M13 vectors. The human apolipoprotein A-I and ApoB-100 3' cDNA clones were provided by Dr. J. Breslow (29, 30). The prepara- tion of single-stranded cDNA probes for ApoA-I (31) and ApoB-100 3' (13) and hybridization assays for determination of the correspond- ing mRNAs were carried out exactly as described (13, 31, 32). The human ApoB-100 5' cDNA probe was prepared from a fragment of pB25 (33) subcloned in M13 mpl0 as described by Sorci-Thomas et al. (34). Human LDL receptor cDNA was prepared from a fragment of pLDLR2 (35) and inserted into M13 mp8 as described (34). Single- stranded probes for ApoB-100 5' and LDL receptor were isolated by digestion with BclI and BamHI, respectively, after DNA synthesis and purified as described (34). Hybridization assays for mRNA de- terminations were carried out as described (32, 34) using the respec- tive M13-apo cDNA template as hybridization standards to calibrate absolute mRNA concentration (32). All assays were carried out using RNA from triplicate dishes, and four mRNA measurements were made on RNA prepared from each dish. ApoB mRNA was character- ized by Northern blot analysis (36) using a uniformly labeled 5' end ApoB cDNA probe.
Statistical Analysis-Statistical analysis was performed using the Student's t test and simple correlation analysis.
RESULTS
Effects of Free Cholesterol on the Net Accumulation of Neutral Lipids and Apolipoproteins in HepG2 Cells and Culture Medium-To determine the effects of cholesterol on cellular and secreted lipoproteins, HepG2 cell were incubated with increasing concentration of pure cholesterol for 17 h. As shown in Fig. l A , cholesterol caused a moderate decrease in the cellular content of both triglycerides and cholesteryl es- ters. On the other hand, addition of pure cholesterol resulted in a significant, dose-dependent increase in the concentration of'unesterified cholesterol (Fig. LA). At the highest concen- tration of cholesterol tested (100 pg/ml of medium), the cellular content of unesterified cholesterol was increased by 226% ( p < 0.001), while the levels of triglycerides and choles- teryl esters were reduced by 25 and 16%, respectively (Fig. 1A). In contrast to marked changes in cellular neutral lipids, no significant alterations were observed in their net accumu- lation in the culture medium (Fig. 1B). There was only a moderate, although not significant, increase of 20% in the concentration of both unesterified cholesterol and cholesteryl esters in the culture medium of HepG2 cells incubated with 100 pg of cholesterol/ml of medium (Fig. 1B). The net accu- mulation of apolipoproteins A-I and B in the culture medium was not affected by cholesterol (Fig. 2 A ) . The lack of effect of pure cholesterol on ApoB production corroborated the results obtained on the net accumulation of neutral lipids in the medium (Fig. 1B) and was further verified by unchanged ApoB mRNA abundance (data not shown).
Effects of Plasma LDL on the Net Accumulation of Neutral Lipids and Apolipoproteins in HepG2 Cells and Culture Me- dium-Since pure cholesterol did not affect the net accumu- lation of ApoB-containing lipoproteins in the medium, the next series of experiments were designed to examine the effects of LDL cholesterol on this process. Preincubation with LDL at 10 pg of protein/ml of medium caused a 9% increase in the cellular content of both triglycerides and cholesteryl esters without affecting that of unesterified cholesterol (Fig. I C ) . The net accumulation of triglycerides and cholesteryl esters in the culture medium was increased by 20 and 12%,
Cholesterol bglml) Cholesterol (rrs/al)
C 0
1
LDL k g protein/nll FIG. 1. Effects of pure cholesterol and LDL cholesterol on
the net accumulation of neutral lipids in HepGZ cells and culture medium. HepG2 cells were grown in MEM containing 10% FBS for 4 days. The maintenance medium was removed, cells were washed twice with PBS, and incubated for 17 h with serum-free MEM containing increasing concentration of pure cholesterol dissolved in ethanol; control cells received the same amount of ethanol. The net accumulation of triglycerides (open bars), unesterified cholesterol (dotted bars), and cholesteryl esters (diagonal bars) in the cells (panel A) and medium (panel B ) was determined by gas-liquid chromatog- raphy. In studies on the effect of LDL cholesterol, the maintenance medium was removed, cells were washed twice with PBS, and prein- cubated for 24 h in serum-free MEM containing the indicated con- centration of LDL. After removing the preincubation medium, cells were washed three times with PBS and incubated in serum-free MEM for 17 h. The net accumulation of neutral lipids in the cells (panel C) and culture medium (panel D) was determined. Values are means .t S.E. of six dishes from two separate experiments. The control values for cellular neutral lipids, expressed in terms of pg/g cell protein/h, were: triglycerides = 1897 f 80; unesterified cholesterol = 953 f 18; cholesteryl esters = 893 f 21. The control values for medium neutral lipids, expressed in terms of Fg/g cell protein/h, were: triglycerides = 132 f 18; unesterified cholesterol = 95 f 6; cholesteryl esters = 41 f 4. *p < 0.02; **p < 0.005; ***p < 0.05; ****p < 0.001; *****p < 0.01.
respectively, (Fig. 1D). When HepG2 cells were preincubated with high concentration of LDL (100 pg of protein or 96 pg of total cholesterol/ml of medium), there was a significant 2- fold increase ( p < 0.005) in the cellular content of cholesteryl esters and a 23% decrease ( p < 0.02) in that of triglycerides; there were no changes in the cellular unesterified cholesterol (Fig. I C ) . Under these experimental conditions, significant increases in the accumulation of triglycerides (29%,p < 0.01), unesterified cholesterol (53%,p < 0.02), and cholesteryl esters (70%, p < 0.005) in the culture medium were observed (Fig. 1D).
Preincubation of HepG2 cells with LDL had no significant effect on the production of ApoA-I (Fig. 2B). By contrast, the net accumulation of ApoB in the medium was significantly stimulated as a function of increasing concentration of LDL in the preincubation medium (Fig. 2B). At the highest con- centration of LDL tested (100 pg of protein/ml medium), the observed 56% increase in ApoB (Fig. 2B) correlated with the 70% elevation in the net accumulation of cholesteryl esters in the culture medium under the same experimental condi-

Regulation of ApoB by Cholesterol in HepG2 Cells 7163
Cholesterol (Whll LoL (M p r o t e i n h l )
FIG. 2. Effects of pure cholesterol and plasma LDL choles- terol on the net accumulation of apolipoproteins A-I and B in the culture medium. Cells were grown and incubated with the source of cholesterol as described in the legend to Fig. 1. The net accumulation of ApoA-I (open bars) and ApoB (diagonal bars) in the culture medium of cells incubated with pure cholesterol ( A ) or prein- cubated with LDL cholesterol ( B ) was measured by electroimmu- noassay. Values are means f S.E. of six dishes from two separate experiments. The control values for apolipoproteins, expressed in terms of pg/g cell protein/h, were: ApoA-I = 65 f 4; ApoB = 120 f 19. *Significant at p C 0.025; **significant at p < 0.05; ***significant at p < 0.01.
tions (Fig. 1D). The increase in ApoB accumulation in the medium was largely due to enhanced secretion as determined by [36S]methionine incorporation into ApoB (data not shown). However, LDL cholesterol had no effect on the cellular con- centration of ApoB mRNA (data not shown).
Effects of 25-Hydroxychlestero1 on the Net Accumulation of Neutral Lipids and Apolipoproteins in HepG2 Cells and Culture Medium-The above results do not totally exclude the pos- sibility that the increased concentrations of cholesteryl esters and ApoB in the culture medium of cells preincubated with LDL might have been due to residual exogenous lipoproteins. Therefore, an alternative method was used to modify the cellular cholesterol content and to examine the subsequent effect on the production of ApoB-containing lipoproteins. A number of oxygenated sterols, in particular 25-hydroxycho- lesterol, have been shown to regulate cholesterol homeostasis in cultured fetal mouse liver cells (37), in hepatoma cells (38), in perfused liver, and freshly isolated hepatocytes (39), in monolayer cultures of adult rat hepatocytes (40) and rabbit intestinal absorptive cells (41). Therefore, the effects of this sterol on cholesterol esterification and ApoB production in HepG2 cells were examined. A time study on the effect of 25- hydroxycholesterol demonstrated that changes in cellular neutral lipids were detectable after 30 min of incubation, the time of the first measurement. However, a minimum of 4- hour incubation time was necessary to allow the accumulation of sufficient amounts of neutral lipids and apolipoproteins in the culture medium that could be measured accurately by gas- liquid chromatography and electroimmunoassay methods, re- spectively, used in these experiments. The observed altera- tions in the cellular and secreted neutral lipids were main- tained for 18 h of incubation, therefore, all experiments were carried out for 14-18 h unless otherwise stated.
As shown in Table I, addition of 25-hydroxycholesterol was highly effective in altering the concentration and composition of cellular neutral lipids with the maximum effect occurring at 20 pg/ml of medium. The most predominant effect of 25- hydroxycholesterol was on the concentration of cholesteryl esters which in a series of four experiments was increased by 2-3-fold (Table I). The observed 2-fold increase in cellular cholesteryl ester concentration at 20 pg of sterol/ml of me- dium coincided with a 43% decrease in that of unesterified cholesterol (Table I), indicating marked stimulation in ester-
ification of endogenous cholesterol. Whereas the total choles- terol concentration in the cells was not significantly altered, there was a significant 33% decrease in that of triglycerides (Table I). In the control system (ethanol alone), triglycerides, unesterified cholesterol, and cholesteryl esters accounted for 49,27, and 24%, respectively, of the total neutral lipid content of the cells (Table I). In the presence of 25-hydroxycholes- terol, the contribution of triglycerides, unesterified choles- terol, and cholesteryl esters to the total neutral lipids were 34, 16, and 50%, respectively (Table I).
Addition of 25-hydroxycholesterol caused a concentration- dependent increase in the net accumulation of cholesteryl esters and a concurrent decrease in that of unesterified cho- lesterol in the medium with the maximum effect occurring at 20 pg of sterol/ml of medium (Table 11). The significant 3- fold increase in the net accumulation of cholesteryl esters was paralleled by a 50% decrease in that of unesterified cholesterol and the consequent unchanged level of total cholesterol (Table 11). There was also a moderate, although not signifi- cant, increase of 40% in the net accumulation of triglycerides in the medium (Table 11). The above changes in the absolute concentration of secreted neutral lipids brought about signif- icant alterations in their composition. Thus, in the control system, and in weight percentage, triglycerides, unesterified cholesterol, and cholesteryl esters accounted for 45, 40, and 15%, respectively, of the total neutral lipids (Table 11). In the presence of 25-hydroxycholesterol, the contribution of triglyc- erides, unesterified cholesterol, and cholesteryl esters to the total neutral lipids were 47, 15, and 38%, respectively, (Table 11). The above results clearly demonstrated that, the 25- hydroxycholesterol-mediated changes in the cellular neutral lipids were reflected in the concentration and composition of secreted lipoproteins. The observed decrease in triglycerides and cholesteryl esters in the presence of 40 pg of 25-hydrox- ycholesterol/ml of medium might be due to the possible cy- totoxic effect of high concentration of this sterol.
The net accumulation of ApoA-I in the culture medium was not significantly affected by 25-hydroxycholesterol at all con- centrations tested (Table 111). By contrast, 25-hydroxycholes- terol caused a dose-dependent increase in the concentration of ApoB in the medium (Table 111). In a series of seven experiments, the rate of ApoB accumulation in the medium was significantly increased by 70-220% upon incubation with 25-hydroxycholesterol added at 10-40 pg/ml of medium; the maximum 2.5-fold enhancement was observed at 20 pg of sterol/ml of medium (Table 111).
Effects of 25-Hydroxycholesterol on the de Novo Synthesis and Secretion of ApoB-To establish that the increase in ApoB accumulation in the culture medium (Table 111) was due to the enhanced secretion, the incorporation of [35S] methionine into newly synthesized ApoB was determined. Confluent 4-day-old HepG2 cells were incubated with serum- free MEM containing [35S]methionine (100 pCi/ml) and either ethanol (control) or 25-hydroxycholestero1 at 20 pg/ml of medium. After incubation for 2-22 h, medium was removed and 35S-labeled ApoB was analyzed by immunoprecipitation followed by SDS-gel electrophoresis and autofluorography. Results demonstrated that 25-hydroxycholesterol increased the secretion of newly synthesized ApoB and that this was apparent at the earliest time point measured (2 h) and per- sisted up to 22 h of incubation time tested (data not shown),
To examine the effect of 25-hydroxycholesterol on early stages of synthesis and secretion of ApoB, pulse-chase exper- iments were performed. After the 10-min pulse, the concen- tration of immunoprecipitable 35S-labeled ApoB was higher in cells incubated with 25-hydroxycholesterol (Fig. 3A, lane

7164 Regulation of ApoB by Cholesterol in HepG2 Cells
TABLE I Effect of 25-hydroxycholesterol on the net accumulation of neutral lipids in HepGZ cells
Cells were grown in MEM containing 10% FBS for 4 days. The maintenance medium was removed, cells were washed with PBS, and serum-free MEM containing either ethanol (control) or 25-hydroxycholestero1 dissolved in ethanol was added. After a 16-20-h incubation, the neutral lipid content of cells was determined by gas-liquid chromatography. Values are means f S.E. of 12 dishes from four different experiments.
Triglycerides Unesterified Cholesteryl Total cholesterol esters cholesterol
Fg/g cell proteinlh Control 1719.33 f 138.57 961.07 f 33.10 824.45 f 87.74 1457.28 f 78.14 25-Hydroxycholestero1 (10 pg/ml of medium) 1345.07 f 219.89 486.18 f 28.43" 1102.78 f 59.34b 1149.35 f 63.04b 25-Hydroxycholester01(20 pg/ml of medium) 1159.20 f 75.11' 547.74 f 9.18" 1680.90 f 58.31" 1560.22 f 39.31
Significant a t p < 0.001. Significant at p C 0.05. Significant a t p < 0.01.
TABLE I1 Effect of 25-hydroxycholesterol on the net accumuhtion of neutral lipids in HepG2 cell culture medium
Cells were grown in MEM containing 10% FBS for 4 days. The maintenance medium was removed and cells were washed twice with PBS and incubated with serum-free MEM containing either ethanol (control) or 25- hydroxycholesterol dissolved in ethanol. After a 16-20-h incubation, medium was removed, concentrated, and analyzed for neutral lipids by gas-liquid chromatography. Values are means f S.E. of 12 dishes from four different experiments.
Additions Triglycerides Unesterified Cholesteryl Total cholesterol esters cholesterol
~
pg/g cell proteinlh ~~
Control 112.53 f 14.91 99.96 f 9.43 39.94 f 4.41 124.12 f 11.27 25-Hydroxycholestero1 (10 pg/ml of medium) 144.35 f 12.01 68.20 f 2.76" 86.77 k 10.55b 120.65 f 8.94 25-Hydroxycholester01(20 pg/ml of medium) 157.59 f 37.06 49.55 f 4.70b 127.20 f 12.01b 126.16 f 10.46 25-Hydroxycholestero1(40 pg/mI of medium) 99.59 f 16.36 38.21 f 2.05' 113.50 -C 7.45b 106.51 f 3.37
Significant at p < 0.005. Significant at p c 0.001.
TABLE 111 Net accumulation in HepGZ culture medium and celluhr mRNA contents of apolipoproteins A-I and B in the
presence of 25-hydroxycholestero1 Cells were grown under the experimental conditions described in the legend to Table 11. Values for accumulation
of apolipoproteins A-I and B are means f S.E. of 12 dishes from four different experiments. Values for cellular mRNA are means f S.E. of triplicate dishes. The mean for each dish represents four determinations.
Accumulation in medium Cellular mRNA content
ApoA-I ApoB APoA-I ApoB
g l g cell proteinlh Pg1l.g RNA Control 97.7 f 7.4 88.2 f 9.6 59.5 * 2.5 412.2 f 18.4 25-Hydroxycholestero1(10 pg/ml of medium) 115.3 f 7.1 152.2 f 11.3" 61.0 f 1.4 496.3 f 4.8b 25-HydroxychoIestero1(20 pg/mI of medium) 103.9 f 5.4 219.3 -C 35.5" 59.7 3.2 636.9 k 28.3' 25-Hydroxycholestero1(40 pg/ml of medium) 110.3 f 11.0 220.2 f 23.1" NDd 541.3 k 19.8"
a Significant at p < 0.001. Significant a t p < 0.025.
e Significant a t p < 0.005. ND, not determined. Significant a t p < 0.02.
2) when compared to ethanol alone (Fig. 3A, lane I). The mobility of %-labeled ApoB was identical to that of ApoB present in human plasma LDL (Fig. 3A, lane 3) which corre- sponded to molecular weight of ApoB-100, based on calibra- tion protein standards (Fig. 3A, lane 4). SDS-polyacrylamide gel electrophoresis analysis of the chase medium (Fig. 3B) demonstrated that in both control and 25-hydroxycholesterol- treated cells, ApoB-100 appeared in the medium between 20- 40 min after the start of the chase. These results indicated that the secretion of newly synthesized ApoB into the medium required longer than 20 min and that, under the experimental conditions employed, the transit time of ApoB in the secretory pathway was similar in both the control and 25-hydroxycho- lesterol-treated cells. As shown in Fig. 3B, the stimulatory effect of 25-hydroxycholesterol on the secretion of newly synthesized ApoB-100 was evident as early as 40 min after
the start of chase and was maintained throughout the 120 min of the chase period tested.
Effect of 25-Hydroxydwlesterol on the Cellular mRNA Con- tent of Apolipoproteins A-I and B-To determine if the 25- hydroxycholesterol-mediated stimulation in the secretion of ApoB was due to increase in mRNA level, cellular ApoB mRNA content was measured by DNA excess solution hy- bridization with the 3' end cDNA probe. These studies were repeated three times and showed a high degree of reproduci- bility. In accordance with the unimpaired accumulation of ApoA-I in the medium, there was no change in the correspond- ing mRNA level (Table 111). Consistent with the enhanced rate of ApoB secretion, the concentration of ApoB mRNA was significantly increased in HepG2 cells incubated with 25- hydroxycholesterol (Table 111). In agreement with results obtained for neutral lipids and ApoB production, the maxi-


7166 Regulation of ApoB by Cholesterol in HepG2 Cells
tion (r = -0.56, p < 0.02, n = 44) (Fig. 50). These results suggest that in HepG2 cells the cellular concentration of cholesteryl esters might dictate the secretion rate of ApoB- containing lipoprotein particles.
To establish that the enhanced rate of ApoB synthesis and secretion by 25-hydroxycholesterol was due to increased in- tracellular cholesterol esterification, the effects of progester- one, a known inhibitor of acyl-coenzyme Axholesterol acyl- transferase (41) on these processes were studied. As shown in Table IV, there was a 26 and 46% decrease in cellular triglyc- erides and unesterified cholesterol, respectively, and a 75% increase in that of cholesteryl esters when cells were incubated with 25-hydroxycholestero1. The net accumulation of triglyc- erides, cholesteryl esters, and ApoB in the medium were also increased by 65, 244, and 92%, respectively, while that of unesterified cholesterol decreased by 43% (Table IV). Simul- taneous addition of progesterone at 5 pg/ml of medium pre- vented the stimulatory effect of 25-hydroxycholesterol on the
ApoB Accumulation in Medium b g f g cell protein/h)
FIG. 5. Relationships between the rate of ApoB production and the net accumulation of neutral lipids in HepG2 cells and culture medium. Panel A shows the correlation between cellular cholesteryl esters content and ApoB net accumulation in the medium obtained in five separate experiments ( r = +0.66, p < 0.001). The data from seven experiments were used to establish the relationship of the rate of ApoB accumulation in the medium with those of cholesteryl esters (panel B ) , triglycerides (panel C), and unesterified cholesterol (panel D). Analysis of the data showed significant positive correlation for both cholesteryl esters ( r = +0.79, p < 0.001) and triglycerides (+0.78, p < 0.001) and a negative correlation for unes- terified cholesterol ( r = -0.56, p < 0.02) and ApoB production.
secretion of cholesteryl ester-rich ApoB-containing lipopro- tein particles (Table IV). At higher concentrations of proges- terone tested, there was a significant increase in the accu- mulation of all neutral lipids, in particular triglycerides, in the cells and a concurrent inhibition in the secretion of ApoB- containing lipoproteins (Table IV). The reduced rate of ApoB accumulation in the medium by progesterone was confirmed by a marked decrease in the incorporation of [35S]methionine into immunoprecipitable ApoB in the culture medium (data not shown).
Role of LDL Receptor in the Regulation of ApoB Produc- tion-The above studies demonstrated that the increased ApoB accumulation in the culture medium of HepG2 cells incubated with 25-hydroxycholesterol was primarily due to increased secretion of the newly synthesized ApoB. However, down-regulation of LDL receptor and subsequent decrease in the catabolism of secreted lipoproteins under the present experimental conditions may also play a role in this process. Therefore, experiments were carried out to investigate this latter possibility. As shown in Fig. 6, at all concentrations of lZ5I-labeled LDL tested, both binding at 4 "C (Fig. 6A) and uptake at 37 "C (Fig. 6B) were inhibited by 20-25% in HepG2 cells preincubated with 25-hydroxycholesterol when com- pared to ethanol alone. The degradation of lZ5I-labeled LDL was inhibited by 12-20% in 25-hydroxycholesterol-treated cells (data not shown). In addition, there was only a 10% decrease in LDL receptor mRNA abundance in HepG2 cells incubated with 25-hydroxycholesterol in comparison to ethanol alone (data not shown). This weak down-regulation of LDL receptor was also observed in HepG2 cells incubated with plasma LDL or pure cholesterol. Incubation of HepG2 cells with plasma LDL (65 pg of protein/ml of medium) for 19 h caused a moderate 13, 20, and 14% decrease in the binding, uptake, and degradation, respectively, of 1251-labeled plasma LDL (data not shown). There was even a smaller reduction of 7-13% in LDL receptor activity in HepG2 cells incubated with 50 pg of pure cholesterol/ml of medium (data not shown).
These results indicated that 25-hydroxycholesterol caused a small to moderate decrease in LDL receptor activity and subsequent reduction in the catabolism of plasma LDL. How- ever, these results do not rule out the possible differential reuptake of the newly synthesized lipoproteins. Therefore, the effect of 25-hydroxycholesterol on the reuptake of secreted nascent ApoB-containing lipoproteins was assessed. Nascent
TABLE IV Effect of progesterone on the 25-hydroxycholesterol-mediated accumulation of neutral lipids and apolipoprotein B in
HepG2 cells and culture medium Cells were grown under the experimental conditions described in the legends to Tables 1-111. Progesterone, at
the indicated concentration, was added simultaneously with 25-hydroxycholestero1(25-OH) at 20 pg/ml of medium. Control cells were incubated with ethanol alone. Values are mean f S.E. of triplicate dishes. TG, triglycerides; FC, free cholesterol; CE, cholesteryl esters.
Neutral lipids in cells Neutral lipids in medium Progesterone
pglrnl medium pg/g cell proteinlh
ApoB in medium TG FC CE TG FC CE
Control 0 1818 f 46 1057 f 3 1038 f 13 6 5 f 11 92 f 8 2 7 f 2 74 f 6 25-OH 0 1344 f 55" 568 f 19' 1816 f 32' 107 f 6 52 f 1' 9 3 f 8 " 142 f lod 25-OH 5 1731 f 126 915 f 14' 1505 f 36' 73 f 3 39 f 2" 46 f 1" 85 f 2 25-OH 10 2066 f 58' 1095 f 15 1383 -+ 44" 33 f 4 24 f 1" 19 f lb 49 f 3' 25-OH 20 2491 f 50' 1290 f 6' 1342 f 16' 6 f Id 7 f 1' 4 f 1" Trace
a Significant at p < 0.005. Significant at p < 0.05.
e Significant at p < 0.001. Significant at p < 0.01.
e Significant at p < 0.025.
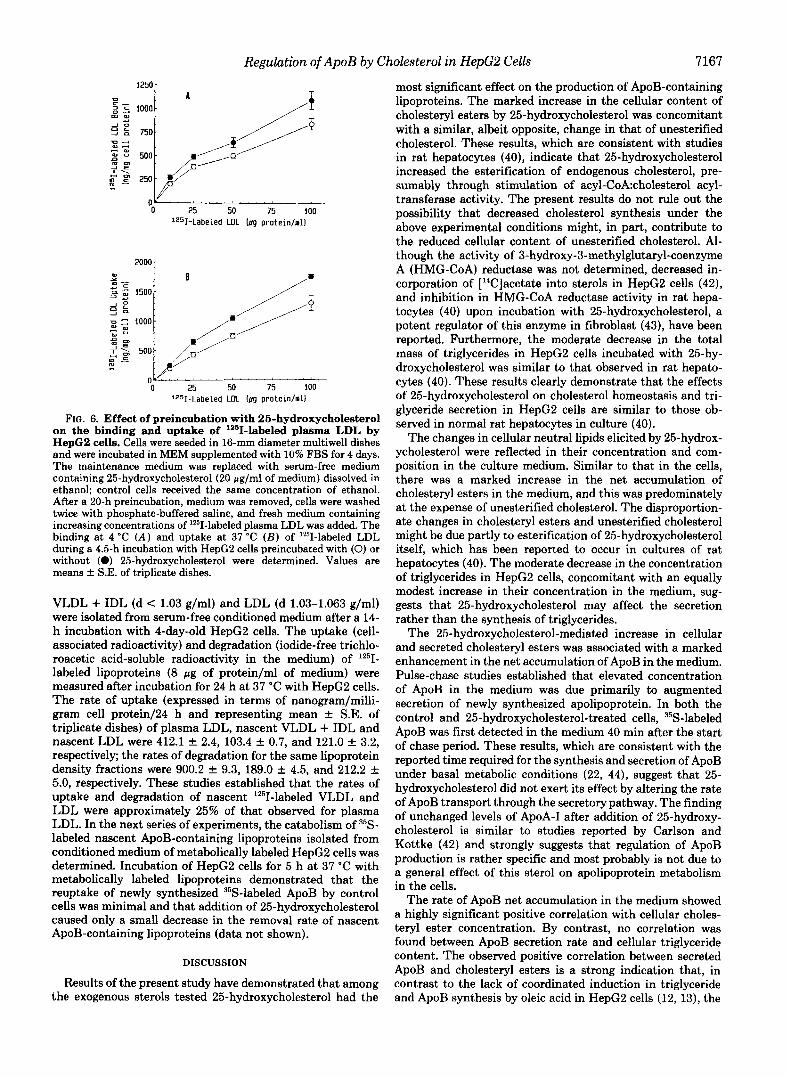
Regulation of ApoB by Cholesterol in HepG2 Cells 7167
0 25 50 75 100 1251-Labeled LOL (ag prote in /n l l
2000 r
0 25 50 75 100 1251-Labeled LOL lug protein/mll
FIG. 6. Effect of preincubation with 26-hydroxycholestero1 on the binding and uptake of '261-labeled plasma LDL by HepG2 cells. Cells were seeded in 16-mm diameter multiwell dishes and were incubated in MEM supplemented with 10% FBS for 4 days. The maintenance medium was replaced with serum-free medium containing 25-hydroxy~holestero1(20 .ug/ml of medium) dissolved in ethanol; control cells received the same concentration of ethanol. After a 20-h preincubation, medium was removed, cells were washed twice with phosphate-buffered saline, and fresh medium containing increasing concentrations of lZ5I-labeled plasma LDL was added. The binding at 4 "C ( A ) and uptake at 37 "C ( B ) of '251-labeled LDL during a 4.5-h incubation with HepG2 cells preincubated with (0) or without (0) 25-hydroxycholestero1 were determined. Values are means f S.E. of triplicate dishes.
VLDL + IDL (d < 1.03 g/ml) and LDL (d 1.03-1.063 g/ml) were isolated from serum-free conditioned medium after a 14- h incubation with 4-day-old HepG2 cells. The uptake (cell- associated radioactivity) and degradation (iodide-free trichlo- roacetic acid-soluble radioactivity in the medium) of lZ5I- labeled lipoproteins (8 pg of protein/ml of medium) were measured after incubation for 24 h at 37 "C with HepG2 cells. The rate of uptake (expressed in terms of nanogram/milli- gram cell protein/24 h and representing mean k S.E. of triplicate dishes) of plasma LDL, nascent VLDL + IDL and nascent LDL were 412.1 f 2.4, 103.4 f 0.7, and 121.0 f 3.2, respectively; the rates of degradation for the same lipoprotein density fractions were 900.2 k 9.3, 189.0 k 4.5, and 212.2 f 5.0, respectively. These studies established that the rates of uptake and degradation of nascent lZ5I-labeled VLDL and LDL were approximately 25% of that observed for plasma LDL. In the next series of experiments, the catabolism of 35S- labeled nascent ApoB-containing lipoproteins isolated from conditioned medium of metabolically labeled HepG2 cells was determined. Incubation of HepG2 cells for 5 h at 37 "C with metabolically labeled lipoproteins demonstrated that the reuptake of newly synthesized 35S-labeled ApoB by control cells was minimal and that addition of 25-hydroxycholesterol caused only a small decrease in the removal rate of nascent ApoB-containing lipoproteins (data not shown).
DISCUSSION
Results of the present study have demonstrated that among the exogenous sterols tested 25-hydroxycholesterol had the
most significant effect on the production of ApoB-containing lipoproteins. The marked increase in the cellular content of cholesteryl esters by 25-hydroxycholesterol was concomitant with a similar, albeit opposite, change in that of unesterified cholesterol. These results, which are consistent with studies in rat hepatocytes (40), indicate that 25-hydroxycholesterol increased the esterification of endogenous cholesterol, pre- sumably through stimulation of acyl-CoAcholesterol acyl- transferase activity. The present results do not rule out the possibility that decreased cholesterol synthesis under the above experimental conditions might, in part, contribute to the reduced cellular content of unesterified cholesterol. Al- though the activity of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase was not determined, decreased in- corporation of ['*C]acetate into sterols in HepG2 cells (42), and inhibition in HMG-CoA reductase activity in rat hepa- tocytes (40) upon incubation with 25-hydroxycholesterol, a potent regulator of this enzyme in fibroblast (43), have been reported. Furthermore, the moderate decrease in the total mass of triglycerides in HepG2 cells incubated with 25-hy- droxycholesterol was similar to that observed in rat hepato- cytes (40). These results clearly demonstrate that the effects of 25-hydroxycholesterol on cholesterol homeostasis and tri- glyceride secretion in HepG2 cells are similar to those ob- served in normal rat hepatocytes in culture (40).
The changes in cellular neutral lipids elicited by 25-hydrox- ycholesterol were reflected in their concentration and com- position in the culture medium. Similar to that in the cells, there was a marked increase in the net accumulation of cholesteryl esters in the medium, and this was predominately at the expense of unesterified cholesterol. The disproportion- ate changes in cholesteryl esters and unesterified cholesterol might be due partly to esterification of 25-hydroxycholesterol itself, which has been reported to occur in cultures of rat hepatocytes (40). The moderate decrease in the concentration of triglycerides in HepG2 cells, concomitant with an equally modest increase in their concentration in the medium, sug- gests that 25-hydroxycholesterol may affect the secretion rather than the synthesis of triglycerides.
The 25-hydroxycholesterol-mediated increase in cellular and secreted cholesteryl esters was associated with a marked enhancement in the net accumulation of ApoB in the medium. Pulse-chase studies established that elevated concentration of ApoB in the medium was due primarily to augmented secretion of newly synthesized apolipoprotein. In both the control and 25-hydroxycholesterol-treated cells, 35S-labeled ApoB was first detected in the medium 40 min after the start of chase period. These results, which are consistent with the reported time required for the synthesis and secretion of ApoB under basal metabolic conditions (22, 44), suggest that 25- hydroxycholesterol did not exert its effect by altering the rate of ApoB transport through the secretory pathway. The finding of unchanged levels of ApoA-I after addition of 25-hydroxy- cholesterol is similar to studies reported by Carlson and Kottke (42) and strongly suggests that regulation of ApoB production is rather specific and most probably is not due to a general effect of this sterol on apolipoprotein metabolism in the cells.
The rate of ApoB net accumulation in the medium showed a highly significant positive correlation with cellular choles- teryl ester concentration. By contrast, no correlation was found between ApoB secretion rate and cellular triglyceride content. The observed positive correlation between secreted ApoB and cholesteryl esters is a strong indication that, in contrast to the lack of coordinated induction in triglyceride and ApoB synthesis by oleic acid in HepG2 cells (12,13), the

7168 Regulation of ApoB by Cholesterol in HepG2 Cells
secretion of ApoB appears to be tightly coupled to cellular cholesterol esterification. This conclusion was supported by studies on the effect of progesterone, an inhibitor of acyl- CoA:cholesterol acyltransferase (41), on the sterol-mediated changes in the secretion of ApoB-containing lipoproteins. Progesterone prevented the stimulatory effect of 25-hydrox- ycholesterol on cellular cholesterol esterification and abol- ished the marked enhancement in the secretion of cholesteryl esters and ApoB. Consistent with studies by Cianflone et al. (45), the effect of progesterone was not specific, i.e. it also decreased the net accumulation of triglycerides in the me- dium. This decrease was due to diminished rate of secretion since triglyceride accumulation in the cells was increased in the presence of progesterone. These results indicated that inhibition of the intracellular cholesterol esterification coin- cided with a decreased rate of ApoB synthesis and consequent reduction in the pool of ApoB that is needed for the assembly of lipoprotein particles. This aberration resulted in retention of triglycerides in the cells and decreased the secretion of cholesteryl ester-, triglyceride-rich ApoB-containing lipopro- tein particles.
Further support for the essential role of cholesteryl ester synthesis in regulation of ApoB production was provided by studies on the effects of pure cholesterol and LDL cholesterol on these processes. Pure cholesterol, which had no regulatory effect on the cellular cholesteryl esters, also failed to modulate the rate of ApoB production. The lack of effect of cholesterol- rich liposomes on ApoB secretion has also been reported by Craig et al. (46). Furthermore, plasma LDL cholesterol caused a marked increase in the net accumulation of cholesteryl esters in the cells and medium, and this was paralleled by similar changes in the net concentration of ApoB in the medium. The LDL-dependent increase in cellular cholesteryl esters was previously shown to be mediated by stimulation in the activity of acyl-CoA:cholesterol acytransferase (10). Stud- ies by other investigators (46) have demonstrated that chy- lomicron remnants and @-VLDL, which also deliver choles- terol to the cells, enhanced the rate of ApoB production by HepG2 cells. It should be pointed out that the inability of pure cholesterol to regulate cellular cholesterol esterification in HepG2 cells, which is consistent with that observed in rat cultured hepatocytes (40), might be due to the low solubility of this sterol impairing its access into the cells. This possibil- ity is supported by the recovery of large amounts of unester- ified cholesterol in cells (Fig. lA), presumably caused by adherence of the added sterol to plasma membrane. Although a part of ApoB in culture medium of LDL-treated cells might be due to a trace amount of exogenous LDL-ApoB, the fact remains that with either pure cholesterol or LDL cholesterol, the net accumulation of ApoB in the medium was directly related to the cellular cholesteryl ester concentration.
The 25-hydroxycholesterol-induced changes in the neutral lipids and ApoB were reflected primarily in the concentration and composition of low density lipoproteins. The main com- positional changes in lipoproteins with d < 1.063 g/ml were a significant increase in the relative contents of cholesteryl esters and ApoB and equally significant decrease in those of unesterified cholesterol and ApoA-I. The ratio of core lipids to ApoB in LDL produced in the presence of 25-hydroxycho- lesterol(l.85 f 0.24) was similar to that in the control system (2.08 f 0.43). This, together with similar ratios in the unfrac- tionated culture medium of control (1.57 f 0.13) and treated cells (1.34 f 0.09) obtained in eight experiments, suggests that 25-hydroxycholesterol increased the number of ApoB- containing lipoprotein particles produced by HepG2 cells. This observation is consistent with studies in both choles-
terol-fed human subjects (47) and nonhuman primates (48). The increase in the net accumulation of ApoB-containing
lipoproteins in the culture medium reflects the balance be- tween the rates of secretion and catabolism. Since HepG2 cells express LDL receptor (10, 49), reduced LDL receptor activity and subsequent decrease in the removal of newly synthesized lipoproteins under these experimental conditions is potentially an important regulatory mechanism. However, results of the present study demonstrated that, consistent with similar studies by Craig et al. (46), the catabolism of both plasma LDL and nascent ApoB-containing lipoproteins was only minimally decreased in HepGZ cells incubated with either LDL or 25-hydroxycholesterol. These results, together with the small decrease in LDL receptor mRNA abundance in 25-hydroxycholesterol-treated cells, clearly and reproduci- bly demonstrated that reduced catabolism contributed only minimally to the marked increase in ApoB accumulation in the medium. This conclusion is at variance with that reported for cholesterol-fed African green monkeys (34) in which de- creased catabolism of ApoB-containing lipoproteins was pro- posed to be the major mechanism responsible for increased LDL concentration. Whereas the variations in the two studies may possibly be explained by differences in the experimental conditions, e.g. the composition of diet used in the in vivo study (34) and/or species differences, the exact factors for the contrasting results require further investigation. The weak down-regulation of LDL receptor by cholesterol observed in the present study supports the hypothesis (49) that exogenous LDL-derived cholesterol is preferentially channeled to the acyl-CoAcholesterol acyltransferase-substrate pool rather than to the free cholesterol pool involved in the regulation of LDL receptor activity.
The increase in ApoB mRNA concentration observed in the present study indicates that 25-hydroxycholestero1 also regulates ApoB synthesis at the molecular level. The marked stimulation of 150% in the secretion of ApoB was accom- panied by a 55% increase in ApoB mRNA concentration. Northern blot analysis of ApoB mRNA showed the presence of a single 14-kilobase mRNA species under all experimental conditions, confirming that 25-hydroxycholesterol increased the secretion of ApoB-100 which is the only molecular form of this protein secreted by HepG2 cells (50). The dispropor- tionate increase in ApoB secretion and its mRNA concentra- tion suggest that in HepG2 cells regulation of ApoB gene expression by 25-hydroxycholesterol occurs both at the mRNA and post-translational levels. The present results do not permit discrimination between transcriptional and post- transcriptional mRNA stability as the mechanism responsible for increased ApoB mRNA abundance. The molecular mech- anism responsible for the regulation of ApoB gene expression at the DNA or mRNA level remains to be elucidated. A potential mechanism for post-translational regulation of ApoB by 25-hydroxycholesterol and LDL cholesterol (which did not change ApoB mRNA concentration) is decreased intracellular degradation of the newly synthesized ApoB which has been shown to occur in HepG2 cells (44, 51). Further studies are needed to elucidate the above regulatory mechanisms.
Thus, the present study has demonstrated that the rate of ApoB secretion was directly correlated with the extent of intracellular cholesterol esterification. The increase in ApoB accumulation in the medium was due primarily to increased secretion rather than reduced catabolism of ApoB-containing lipoproteins. These data indicate that the increase in ApoB secretion by 25-hydroxycholesterol was mediated through both an increased mRNA concentration and post-transla-

Regulation of ApoB by Cholesterol in HepG2 Cells 7169
tional processes. The inhibitory effect of progesterone on both cholesteryl ester formation and ApoB secretion implicates the regulatory role of acyl-CoA:cholesterol acyltransferase in the synthesis and secretion of ApoB-containing lipoprotein par- ticles through mechanisms that remain to be determined.
Acknowledgments-I thank Dr. David L. Williams and Dr. Mary Sorci-Thomas for providing ApoA-I, 3' and 5' end ApoB, and LDL receptor cDNA with the kind permission of Dr. J. Breslow, Dr. A. Protter, and Dr. D. Russell. Polyclonal antibodies to apolipoproteins were kindly provided by Dr. P. Alaupovic. The assistance of Randall Whitmer in performing lipid analyses and Margo French in preparing this manuscript is acknowledged.
REFERENCES 1. Gofman, J. W., Young, W., and Tandy, R. (1966) Circulation 3 4 ,
679-697 2. Goodnight, S. H., Jr., Harris, W. S., Connor, W. E., and Illing-
3. Grundy, S. M., and Denke, M. A. (1990) J. Lipid Res. 31 , 1149-
4. Mistry, P., Miller, N. E., Laker, M., Hazzard, W. R., and Lewis,
5. Schonfeld, G., Patsch, W., Rudel, L. L., Nelson, C., Epstein, M.,
6. Brunzell, J. D., Sniderman, D. A., Albers, J. J., and Kwiterovich,
7. Kane, J. P. (1983) Annu. Rev. Physiol. 45,637-650 8. Dolphin, P. J. (1985) Can. J. Biochem. Cell Biol. 6 3 , 850-869 9. Mahley, R. W., Innerarity, T. L., Rall, S. C., Jr., and Weisgraber,
K. H. (1984) J. Lipid Res. 2 5 , 1277-1294 10. Dashti, N., Wolfbauer, G., Koren, E., Knowles, B. B., and Alau-
povic, P. (1984) Biochim. Biophys. Acta 794,373-384 11. Brown, M. S., and Goldstein, J. L. (1986) Science 2 3 2 , 34-47 12. Dashti, N., and Wolfbauer, G. (1987) J. Lipid Res. 28,423-436 13. Dashti, N., Williams, D. L., and Alaupovic, P. (1989) J. Lipid
14. Dashti, N., Smith, E. A., and Alaupovic, P. (1990) J. Lipid Res.
15. Mahley, R. W. (1978) in Disturbances in Lipid and Lipoprotein Metabolism. (Dietschy, J . M., Gotto, A. M., Jr., and Ontko, J. A., eds) pp. 181-197, American Physiological Society, Bethesda, Maryland
worth, D. R. (1982) Arteriosclerosis 2 , 87-113
1172
B. (1981) J. Clin, Invest. 67,493-502
and Olson., R. E. (1982) J. Clirz. Inuest. 6 9 , 1072-1080
P. O., Jr. (1984) Arteriosclerosis 4 , 79-83
Res. 30,1365-1373
31,113-123
16. Dashti, N. (1990) Arteriosclerosis 10 , 777 (abst.) 17. Dashti, N., Alaupovic, P., Knight-Gibson, C., and Koren, E.
(1987) Biochemistry 26, 4837-4846 18. Kuksis, A., Myher, J. J., Marai, L., and Geher, L. (1975) J.
Chromatogr. Sci. 13 , 423-430 19. McConathy, W. J., Koren, E., Wieland, H., Campos, E. M., Lee,
D. M., Kloer H. U., and Alaupovic, P. (1985) J. Chromutogr.
20. Lowry, 0. H., Rosebrough, N. J., Farr A. L., and Randall, R. J .
21. Dashti, N., Koren E., and Alaupovic, P. (1989) Biochem. Biophys,
22. Bostrom, K., Wettesten, M., Boren, J., Bondjers, J., Wiklund O.,
342,47-66
(1951) J. Biol. Chem. 193,265-275
Res. Commun. 163,574-580
and Olofsson, S. 0. (1986) J. Biol. Chem. 2 6 1 , 13800-13806
23. Sparks, J. D., and Sparks, C. E. (1990) J. Biol. Chem. 265,8854-
24. Laemmli, U. K. (1970) Nature 227,680-685 25. Markwell, M. A. K. (1982) Anal. Biochem. 125,427-432 26. Dashti, N., and Wolfbauer, G. (1986) Biochim. Biophys. Acta
27. Chirgwin, J. M., Przylbyla, A. E., MacDonald R. J., and Rutter, W. J. (1979) Biochemistry 18,5294-5299
28. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) in Molecular Cloning. DD. 202-203. Cold SDrine Harbor Laboratorv. Cold
8862
876,473-486
SpriniHaibor, NY '
"
29. Breslow. J. L.. Ross. D.. McPherson. J.. Williams. H.. Kurnit. K.. Nussbaum, A. L.,'Karathanasis, S . K., and Zannis,'V. I. (19823 Proc. Natl. Acud. Sci. U. S.A. 7 9 , 6861-6865
30. Huang, L.-S., Clark Bock, S., Feinstein, S. I., and Breslow, J. L. (1985) Proc. Natl. Acad. Sci. U. S. A. 82,6825-6829
31. Sorci-Thomas, M., Prack, M. M., Dashti, N., Johnson, F., Rudel, L. L., and Williams, D. L. (1988) J. Biol. Chem. 263 , 5183- 5189
32. Williams, D. L., Newman, T. C., Shelness, G. S., and Gordon, D. A. (1985) Methods Enzymol. 128,671-689
33. Protter, A. A., Hardman, D. A., Schilling, J. W., Miller, J., Appleby, V., Chen, G. C., Krischer, S. W., McEnroe, G., and Kane, J. P. (1986) Proc. Natl. Acud. Sci. U. S. A . 8 3 , 1467- 1471
34. Sorci-Thomas, M., Wilson, M. D., Johnson, F. L., Williams, D. L., and Rudel, L. L. (1989) J. Biol. Chem. 264,9039-9045
35. Yamamoto, T., Davis, C. G., Brown, M. S., Schneider, W. J., Casey, M. L., Goldstein, J. L., and Russell, D. W. (1984) Cell 39,27-38
36. Thomas, P. S. (1980) Proc. Natl. Acud. Sci. U. S.A. 7 7 , 5201- 5205
37. Kandutsch, A. A., and Chen, H. W. (1974) J. Biol. Chem. 2 4 9 ,
38. Bell, J. J., Sargeant, T. E., and Watson, J. A. (1976) J. Biol.
39. Erickson, S. K., Matsui, S. M., Shrewsbury, M. A., Cooper, A. D.,
40. Drevon, C. A., Weinstein, D. B., and Steinberg, D. (1980) J. Biol.
41. Field, F. J., and Mathur, S. N. (1983) J. Lipid Res. 24 , 1049-
42. Carlson, T. L., and Kottke, B. A. (1989) Biochem. J. 264 , 241-
43. Brown, M. S., and Goldstein, J. L. (1974) J. Biol. Chem. 249,
44. Dixon, J . L., Furukawa, S., and Ginsberg, H. N. (1991) J. Biol.
45. Cianflone, K. M., Yasruel, Z., Rodriguez, M. A., Vas, D., and
46. Craig, W. Y., Nutik, R., and Cooper, A. D. (1988) J. Biol. Chem.
47. Packard, C. J., McKinney, L., Carr, K., and Shepherd, J. (1983)
48. Johnson, F. L., St. Clair, R. W., and Rudel, L. L. (1983) J. Clin.
49. Havekes, L. M., De Wit, E. C. M., and Princen, H. M. G. (1987)
50. Dashti, N., Lee, D. M., and Mok, T. (1986) Biochem. Biophys.
51. Sato, R., Imanaka, T., Takatsuki, A., and Takano, T. (1990) J.
6057-6061
Chem. 251,1745-1758
and Gould, G. (1978) J. Biol. Chem. 253,4159-4164
Chem. 255,9128-9137
1059
247
7306-7314
Chem. 266,5080-5086
Sniderman, A. D. (1990) J. Lipid Res. 31 , 2045-2055
263,13880-13890
J. Clin. Invest. 7 2 , 45-51
Invest. 7 2 , 221-236
Biochem. J. 247 , 739-746
Res. Commun. 137,493-499
Biol. Chem. 265, 11880-11884


















