
The effect of exercise and captivity on energy ...faculty.forestry.ubc.ca/hinch/Patterson et al.,...
21
The effect of exercise and captivity on energy partitioning, reproductive maturation and fertilization success in adult sockeye salmon D. A. P ATTERSON *†, J. S. M ACDONALD *, S. G. H INCH ‡, M. C. H EALEY § AND A. P. F ARRELL { *Fisheries and Oceans Canada, Science Branch, Pacific Region, Co-operative Resource Management Institute, School of Resource and Environmental Management, Simon Fraser University, Burnaby, B.C. V5A 1S6, Canada, ‡Department of Forest Sciences University of British Columbia, Vancouver, B.C. V6T 1Z2, Canada, §Institute for Resources and Envir- onment, University of British Columbia, Vancouver, B.C. V6T 1Z2, Canada and {Biological Sciences, Simon Fraser University Burnaby, B.C. V5A 1S6, Canada (Received 24 June 2003, Accepted 20 January 2004) Releasing a population of adult Fraser River sockeye salmon Oncorhynchus nerka from an energetically more expensive non-feeding natural migration (1089 km, natal groups) and allow- ing them to sexually mature in either a moderate flow (captive exercise group) or a no flow (captive non-exercise group) environment resulted in significant differences in body energy densities (MJ kg 1 ) among groups (ranked: initial natural migrator > non-exercise > exercise > natal arrivals). Similarly, per cent body lipids were significantly lower in exercise females compared with non-exercise females, experimentally confirming the central role lipid catabolism has in powering swimming. Although restricting exercise did result in greater body energy reserves at spawning, this did not result in a reallocation of energy to reproduction, as fecundity, egg size and gonad composition were unchanged among the groups. Furthermore, non-exercise females had delayed maturity, lower egg deposition rates, and were more likely to die prior to egg ovulation than exercise females and natal spawners. Eggs from captive exercise adult females were more likely to survive to the eyed stage than eggs from captive non-exercise females, but both captive groups had significantly lower egg fertilization success than natal spawners suggesting that confinement stress played a role in some of results presented. There were no significant differences in plasma levels of glucose, lactate, cortisol and reproductive hormones among the groups, but correlations among acute and chronic indicators of stress were significant among individuals. These results indicate that exercise during the late stages of maturation, as might occur normally, may be required for optimizing reproductive maturation and maximizing reproductive success. # 2004 The Fisheries Society of the British Isles Key words: maturation timing; Oncorhynchus; reproduction; spawning migration; stress. INTRODUCTION Many Pacific salmon Oncorhynchus spp. must perform an arduous upstream migration before spawning. Considerable research has gone into elucidating the †Author to whom correspondence should be addressed. Tel.: þ1 604 666 5671; fax: þ1 604 666 1995; email: [email protected] Journal of Fish Biology (2004) 64, 1039–1059 doi:10.1111/j.1095-8649.2004.00370.x, available online at http://www.blackwell-synergy.com 1039 # 2004 The Fisheries Society of the British Isles
Transcript of The effect of exercise and captivity on energy ...faculty.forestry.ubc.ca/hinch/Patterson et al.,...

The effect of exercise and captivity on energy
partitioning, reproductive maturation and fertilization
success in adult sockeye salmon
D. A. PATTERSON*†, J. S. MACDONALD*, S. G. HINCH‡,M. C. HEALEY§ AND A. P. FARRELL{
*Fisheries and Oceans Canada, Science Branch, Pacific Region, Co-operative ResourceManagement Institute, School of Resource and EnvironmentalManagement, Simon FraserUniversity, Burnaby, B.C. V5A 1S6, Canada, ‡Department of Forest Sciences University ofBritish Columbia, Vancouver, B.C. V6T 1Z2, Canada, §Institute for Resources and Envir-
onment, University of British Columbia, Vancouver, B.C. V6T 1Z2, Canada and{Biological Sciences, Simon Fraser University Burnaby, B.C. V5A 1S6, Canada
(Received 24 June 2003, Accepted 20 January 2004)
Releasing a population of adult Fraser River sockeye salmon Oncorhynchus nerka from an
energetically more expensive non-feeding natural migration (1089km, natal groups) and allow-
ing them to sexually mature in either a moderate flow (captive exercise group) or a no flow
(captive non-exercise group) environment resulted in significant differences in body energy
densities (MJkg�1) among groups (ranked: initial natural migrator>non-exercise>
exercise>natal arrivals). Similarly, per cent body lipids were significantly lower in exercise
females compared with non-exercise females, experimentally confirming the central role lipid
catabolism has in powering swimming. Although restricting exercise did result in greater body
energy reserves at spawning, this did not result in a reallocation of energy to reproduction, as
fecundity, egg size and gonad composition were unchanged among the groups. Furthermore,
non-exercise females had delayed maturity, lower egg deposition rates, and were more likely to
die prior to egg ovulation than exercise females and natal spawners. Eggs from captive exercise
adult females were more likely to survive to the eyed stage than eggs from captive non-exercise
females, but both captive groups had significantly lower egg fertilization success than natal
spawners suggesting that confinement stress played a role in some of results presented. There
were no significant differences in plasma levels of glucose, lactate, cortisol and reproductive
hormones among the groups, but correlations among acute and chronic indicators of stress
were significant among individuals. These results indicate that exercise during the late stages of
maturation, as might occur normally, may be required for optimizing reproductive maturation
and maximizing reproductive success. # 2004 The Fisheries Society of the British Isles
Key words: maturation timing; Oncorhynchus; reproduction; spawning migration; stress.
INTRODUCTION
Many Pacific salmon Oncorhynchus spp. must perform an arduous upstreammigration before spawning. Considerable research has gone into elucidating the
†Author to whom correspondence should be addressed. Tel.: þ1 604 666 5671; fax: þ1 604 666 1995;
email: [email protected]
Journal of Fish Biology (2004) 64, 1039–1059
doi:10.1111/j.1095-8649.2004.00370.x,availableonlineathttp://www.blackwell-synergy.com
1039# 2004TheFisheries Society of theBritish Isles

specific adaptations that salmonids have evolved to overcome this obstacle(Brett, 1995; Webb, 1995; Høgasen, 1998). These include optimizing swimmingbehaviour, adjusting run timing to coincide with favourable river conditions,elevating swimming performance capability and increasing lipid reserves to fuellong and difficult migrations (Gilhousen, 1980; Brett, 1995; Hinch & Rand,1998; Hodgson & Quinn, 2002; Lee et al., 2003).Early Stuart sockeye salmon Oncorhynchus nerka (Walbaum) undertake one
of the longest and most difficult freshwater migrations of any salmonid popula-tion. Migration takes place in the Fraser River, British Columbia (BC) everyJuly, spanning 3–5 weeks, during which period the average river discharge rateis decreasing from peak spring freshet level and water temperature is increasingtowards the summer high. Without feeding, they embark on a 1089 km migra-tion with 12% body lipids and then gain an elevation of 701m before reachingthe natal spawning streams with body lipids reduced to 2% (G.T. Crossin,S.G. Hinch, A.P. Farrell, D.A. Higgs, A.G. Lotto, J.D. Oakes &M.C. Healey, pers.comm.). Egg size increases three-fold from freshwater entry to final ovulation(Patterson & Farrell, 2003). Therefore, energy resources are reallocated frommother to offspring to a significant degree while the mother is expendingconsiderable energy during migration and is not feeding. Furthermore, becauseriver discharge rate and water temperature during July vary greatly from year toyear, extreme conditions in some years may further tax the finite energy reservesof Early Stuart sockeye salmon (Macdonald, 2000).Given the limited energy budget of migrating salmonids, a change in the
energy requirements of migration could result in some trade-off in reproductivedevelopment between fecundity, egg size and quality (Smith & Fretwell, 1974;Kinnison et al., 2001). It is generally believed, although not directly tested insockeye salmon, that fecundity is set months prior to freshwater re-entry (Tyleret al., 1994). Therefore, any change in migration conditions would probablyeffect egg size and quality rather than fecundity. Several mechanisms couldpotentially alter egg size. One is that the blood flow requirement of locomotorymuscles during both sustained and stressful exercise (Thorarensen et al., 1993)diverts blood flow away from the ovary, reducing nutrient depositional rates tothe eggs. Another is that release of stress hormones, associated with exercise,interferes with production of reproductive hormones necessary for egg devel-opment (Pottinger & Pickering, 1990; Kubokawa et al., 1999; Schreck et al., 2001).With respect to egg quality, previous work has shown that chronic and acutestress can reduce progeny survival at different embryonic stages in salmonids(Campbell et al., 1992, 1994).Currently, exercise associated with migration is presented as a potential
obstacle to successful reproduction (Macdonald, 2000). There has been noattempt, however, to reverse this paradigm and examine exercise as an integralpart of normal reproductive development for long distance migrators. Indeed,the unknown effect of exercise on different reproductive variables within anovary is a seemingly remarkable data gap given that a large egg size is believedto be the major maternal effect that confers a survival advantage to the off-spring (West & Larkin, 1987). No studies could be found on either the effects ofexercise on the maternal provisioning of lipids and protein constituents,although these constituents may have a profound effect on either overall egg
1040 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

quality (Berg et al., 2001), or the viability of gametes, the ultimate test of eggquality. The broad objective of this study was to examine the effect of exerciseon maturing sockeye salmon in fresh water with respect to selected aspects ofbody morphology, blood physiology and reproduction. By releasing migratingsockeye salmon from their energetically demanding upstream migration, thisexperiment tested the hypothesis that restricting exercise would increase energyallocation toward reproductive performance in female sockeye salmon. Com-parisons were made under controlled conditions and with naturally migratingand spawning fish.
MATERIALS AND METHODS
FISH CAPTURE
Three groups of sockeye salmon termed the natural migrator (Lady Franklin Rock),natal arrival and natal spawner, were terminally sampled and used as a natural referencefor the experiments with captive fish. Twenty natural migrators, 10 females and 10 males,were captured and killed on 15 and 16 July 1999, at Lady Franklin Rock, 175 kmupstream of the Fraser River mouth. On 12 August 1999, 20 natal arrival sockeyesalmon, 10 females and 10 males, were killed upon arrival at Kynoch Creek, an EarlyStuart natal spawning stream located 1089 km upstream from the Fraser River mouth.An additional 16 females were captured on the 14 and 15 August 1999 during activespawning on Kynoch Creek. All fish in this experiment were killed by a single blow to thehead in accordance with Fisheries and Oceans Canada (DFO) animal care guidelines.For the confinement experiments, 40 adult sockeye salmon were captured on 20 and 21July 1999 from Lady Franklin Rock. They were inferred to be 4 years old Early Stuartsockeye salmon based on fish size, historic run timing, and from the analysis of scalesamples taken from 67 other sockeye salmon captured from the same portion of lowerFraser Canyon during that time period (M. Lapointe, pers. comm.). Each fish wasindividually dip-netted and placed in a 40 l container filled with 30 l of 14� C FraserRiver water (10�0mg l�1 oxygen) and 30mg of Marinil# anaesthetic (1 ppt; metomidatehydrochloride; Syndel International Laboratories Ltd., Vancouver, B.C., Canada). Thefish were then transferred to a 1000 l transport tank containing 10� C Cultus Lake water,with a Marinil# anaesthetic concentration of 0�5mg l�1. The length of time from captureof the first fish to arrival at the Fisheries and Oceans Canada Cultus Lake SalmonResearch Laboratory, Cultus Lake, B.C. was <5 h, during which time water oxygen levelswere maintained between 8�0 and 10�0mg l�1. Upon arrival, the fish were dip-nettedout of the transport tank, sexed and then randomly assigned to the two large holdingtanks. Twenty captive fish, six males and 14 females, were placed in each of the holdingswim tanks.
SWIM TANK DESIGN
Two identical ‘doughnut’ shaped concrete holding tanks were used in this experiment.The outer diameter was 5�30m and an inverted round fibreglass aquarium (1�00m deepand 1�80m diameter) was centred in the tank and raised 0�15m off the bottom. The waterdepth was maintained at 1�05m using stand pipes (1�00m height� 0�15m diameter)located in the centre of both tanks. The raised outlet increased water circulation andallowed water from the bottom of the tank to be drawn up to improve the overall waterquality. The total water volume in the tank was 22�0m3, of which 20�5m3 was availableto the fish. Each tank was serviced by two 8�75 cm internal diameter (ID) pipes, eachdischarging a regulated amount of either cool or warm water withdrawn from CultusLake at depths of 15 and 1m, respectively. The water quality of Cultus Lake is sufficientto sustain viable population of lake spawning sockeye salmon. Valves were adjusted to
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1041
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

maintain water temperature in both tanks between 12 and 14� C during the entire 37 daysof the experiment. The holding temperature was just below the optimal swimmingtemperature of 15� C for adult sockeye salmon (Brett, 1995; Tierney, 2000) and withinthe lower range of temperatures Early Stuart sockeye salmon can experience during theirFraser River migration (Macdonald et al., 2000). Salmonid mortality is known to occurat higher temperatures (Taranger & Hansen, 1993; Macdonald et al., 2000). The waterdischarge into the tank was 0�30m from the outside wall and below the water surface.Exercise was encouraged on the fish in one tank by orienting both pipes in the samedirection to create a counter-clockwise flow. In a second tank, the two water dischargepipes opposed each other thereby preventing directional flow. The fish in this tank weredesignated non-exercised.
SWIMMING PROTOCOL
There was minimal directional flow (mean velocity¼ 0�10m s�1) in the exercise tankwhen the fish were first introduced, but water discharge rate was gradually increased overa 2 day period to 0�55m s�1 (measured at mid-depth) on 23 July 1999. This velocity wasmaintained for 25 days. The water discharge rate was reduced on 17 August 1999 (meanvelocity¼ 0�35m s�1) in anticipation of fish sampling and this level was maintained untilthe experiment ended on 26 August 1999. Any changes in water discharge rate to theexercise tank were duplicated in the non-exercise tank to maintain similar water replace-ment rates and temperatures. The approximate distance swum by an exercise fish wasestimated as 1288 km, estimated by multiplying the average encounter velocity (assumingthe fish maintained position in the tank) by the average experimental duration (30 days).It was not possible to quantify the distance non-exercise fish swam due to the slow,erratic and non-directional swimming behaviour they exhibited. In comparison, thenatural spawning group swam 1014 km over c. 21 days, gaining 621m in elevationfrom Lady Franklin Rock.
SAMPLING AND SPAWNING
Early Stuart sockeye salmon normally spawn between 4 and 8 August, but in 1999 anunusually high Fraser River water discharge delayed the arrival onto the natal spawninggrounds and subsequently peak spawning, which occurred between 10 and 15 August. By19 August, all of the estimated 6630 sockeye salmon returning to Kynoch Creek in 1999had migrated through the enumeration fence and the vast majority had spawned and died(Department of Fisheries and Oceans, unpubl. data). Consequently, captive fish were allexpected to be sexually mature by 19 August but not all of them were. Ovary conditionwas recorded as either ripe (ovulated eggs free within body cavity or ‘loose’) or gravid(still attached or ‘tight’) for all females. Male condition was not recorded. Fourteen fishhad died prior to blood and gamete sampling on 19 August: five exercise females (fourloose, one tight); seven non-exercise females (seven tight) and two non-exercise males.Thirteen of 14 had died after 10 August, past the historical date for peak spawning, andas such they were all expected to be ripe. The decision to sample fish on 19 Augusttherefore represented a compromise between increasing mortality and sexual maturity.All fish were externally examined for sexually maturity and 19 were sacrificed: five ripeand two tight exercise females; three ripe and two tight non-exercise females; four exercisemales; three non-exercise males. The remaining seven fish that were not sexually maturewere held for an additional week, however, two exercise females (one tight; one loose)and one exercise male died that week. The surviving four fish were successfully spawnedon 26 August 1999 [two non-exercise females (loose), one non-exercise male, one exercisemale]. Because of the unbalanced statistical design, this second group of spawned fishwere not used in fertilization comparisons, and were used only for assessment of maturity,morphological measurements and physiological condition.
1042 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

MORPHOLOGY AND PHYSIOLOGY
Measurements collected from all captive fish included the following: sex, standardlength (LS) (tip of snout to hypural plate), wet mass (M) and gonad mass (MG). Thegonado-somatic index (IG) was calculated from: 100MGM�1. For females, individualmeasurements of egg wet mass, dry mass and diameter were made. The Fulton conditionfactor (K) was calculated for each individual adult, from K¼ 100ML�3 (Anderson &Neumann, 1996), where M was in g and LS in cm.Eggs were removed from females by cutting the abdominal cavity open from the anal
vent towards the head. Total ovary mass was weighed to the nearest 0�1 g and thendivided into three sub-sets to assess: i) egg morphometrics, ii) proximate constituentanalysis and iii) cross-fertilizations. Egg measurements were made on three sets of 10eggs. Egg diameter was averaged from 10 eggs lined up in a row and measured to thenearest 0�05mm using callipers. Average wet mass was assessed to the nearest 0�0001 g.Fecundity was estimated by dividing the total egg mass by the average wet mass of an eggfrom that female. Eggs were placed in a drying oven for 48 h at 60� C to determine drymass.Body constituent analysis was used to assess fish energy reserves. Constituent analysis
was also performed on portions of all gonads to compare gonad proximate compositionamong non-exercise, exercise and natal arrival fish. Constituent data for natal spawnerswere not used because the fish had already expended energy spawning and lost anunknown amount of gonadal material prior to capture. Fish carcasses and gonads wereseparated and kept frozen at �20� C prior to proximate analysis. They were thawed at4� C before being completely homogenized in a food processor. Duplicate homogenatesof carcass and gonads from each fish were analysed for per cent moisture, dry matter,ash, protein and lipid content according to the procedures of Higgs et al. (2000). Caloricequivalents were used to convert from per cent composition of lipid and protein to energydensity (MJkg�1). The energy equivalents used for protein and lipid were 23�64 kJ g�1
and 36�4 kJ g�1, respectively (Higgs et al., 1995).Blood plasma analysis can provide an integrated measure of the physiological state of
individual fish, with respect to acute stress, osmoregulatory ability, gametogenesis andproximity to final maturation at the time of sampling (Donaldson, 1990; Pickering, 1992;Clarke & Hirano, 1995). Blood samples were taken within 5min of capture from thecaudal vein using a needle attached to heparinized vacutainer. The plasma was partitionedusing a refrigerated centrifuge and stored at �80� C. Plasma samples were subse-quently analysed for cortisol, lactate, glucose, osmolality, 17-a 20-b dihydroxy-4-pregen-3-one (17,20P), testosterone and 11-keto testosterone (11-KT; males only) according tothe methods described by Donaldson et al. (2000). Interrenal nuclear diameters were usedas an index of chronic stress levels in these fish (Fagerlund et al., 1981; Macdonald et al.,1988). Head kidney samples were preserved in Davidson’s solution prior to paraffinembedding, histological sectioning (5mm) and staining. Micrometer measurements ofnuclear diameters were made at �1200 on the major axis of interrenal cells that werespherical in appearance. Mean interrenal nuclear diameter (IRND) was based on 20 cellsper fish (Donaldson et al., 2000). Blood plasma was not taken from natal spawners andcaptive fish that died naturally, and IRNDs were not taken from the natal spawners.
REPRODUCTION
Milt samples were obtained by manually stripping each male. Every effort was made tostop blood and water contamination of the egg and milt samples since this reducesfertility. All milt and egg samples were kept cool at 4� C until fertilization took place.Individual full crosses were made among the gametes from all surviving captive malesand females using a dry fertilization technique (Herunter et al., 2000). At Kynoch Creekon 15 August 1999, 24 unique pairings were made between eight natal females and threenatal males. For each cross, c. 15 g of eggs were weighed and placed in a 120ml glass jar.Approximately 0�1ml of milt was added and both were gently swirled. The sperm werethen activated by adding 15ml of water. The mixture was gently agitated, and an
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1043
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

additional 30ml of water was added. After 2min, the excess sperm were rinsed off byreplacing the original water with a fresh 60ml aliquot of water. The eggs were kept in theglass jars and allowed to water harden for 45min before being placed into incubationcontainers. Captive eggs crosses were placed in baskets (7�0� 7�5� 5�0 cm) inside Heathtrays supplied with Cultus Lake deep water (range 10–8� C). Light was occluded duringthe incubation period to reduce mortality associated with light (Novales Flamarique &Harrower, 1999) and the eggs were not disturbed until they had reached the less sensitiveeyed stage (Jensen & Alderdice, 1989). The eggs from crosses of natal spawners wereplaced in incubation capsules (Herunter et al., 2000) with spawning size gravel and buried15 cm into spawning gravel at Kynoch Creek. The intergravel incubation temperaturewas 12� C at fertilization and declined to 9� C by capsule sampling. At the eyed stage, allof the eggs from each basket or capsule were counted and survival was calculated basedon the number of eyed eggs compared with total egg number per basket or capsule. Thesurviving eggs were monitored weekly until yolk absorption was completed. Survival totwo key development stages, hatch and maximum attainable wet mass (MAWW), wasdetermined.
STATISTICAL ANALYSIS
Individuals within all five groups were considered replicates thus restricting the infer-ences that can be made (Hurlbert, 1984). The overall influence of exercise treatment onthe response variables within a section ( i.e. morphology, plasma analysis, body consti-tuent, gonad constituents and reproductive variables) was examined by MANOVA(Sokal & Rohlf, 2000; Minitab Vers. 13). Response variables were reduced within asection if the variables were highly correlated, r> 0�8. Once an overall treatment effectwas established, one-way ANOVAs were used to test the treatment effect on individualresponse variables. In those cases involving a significant treatment effect for an individ-ual response variable, a Tukey-Kramer multiple means test was applied. Because manyof these variables had a gender specific expression, males and females were analysedseparately except for the analysis of the stress variables. To satisfy the statistical require-ments of normality and homoscedasticity, the plasma hormone data were log10-trans-formed when necessary. To examine the individual relationship among the differentvariables, Pearson’s correlation coefficients were used for all pair-wise comparisons(Minitab ver. 13). Similarly, pair-wise comparisons were used to test whether any ofmorphological, physiological or reproductive maternal traits were correlated with eggsurvival. To assess parental effects on egg survival to eyed stage, a General Linear Model(GLM) was applied, using mothers and fathers as separate factors. Tukey-Kramer multi-ple means comparison tests were then employed if an overall paternal or maternal effectwas found. w2 tests were employed to determine statistical differences in binomial datasuch as survival and full maturity. All tests were deemed statistically significant at analpha value<0�05.
RESULTS
SWIMMING BEHAVIOUR
All exercise fish voluntarily swam against the imposed water current. Theexercise fish oriented against the current and held position within the tank,swimming c. 30 cm off the bottom, maintaining swim speed of 0�55m s�1 orapproximately 1 body length s�1 (BL s�1). In contrast, the non-exercise fish weredisorientated and moved randomly throughout the tank for at least 2 days post-capture. Afterwards they became rather inactive and did not orient in anyparticular direction, remaining within 30 cm of the bottom moving very slowly(<0�05m s�1), with the occasional fish swimming on the surface. The behaviour
1044 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

of the exercise and non-exercise groups differed because the imposed current inthe former allowed them to orient.
MORPHOLOGY
Standard length, body mass or overall body condition of females or maleswas not different among the groups (MANOVA, P> 0�05) (Table I). Energydensity declined in all three groups from initial collection (MANOVA, P< 0�05;females mean� S.E. 8�02� 0�20MJkg�1; males 8�52� 0�23MJkg�1). Femalenatal arrivals had significantly lower somatic energy density(4�30� 0�70MJkg�1) [Fig. 1(a)] than exercise females (5�19� 0�23MJkg�1),who in turn had significantly lower overall somatic energy density than non-exercise females (6�22� 0�21MJkg�1) after 4 weeks of continuous exercise(ANOVA, P< 0�05; Tukey’s test, P< 0�05). This decline was primarily due tolower lipid reserves in natal arrivals and exercise fish (ANOVA, P< 0�05)[Fig. 1(b)]. While energy density and lipid content was significantly lower innatal arrival v. captive spawner and in captive spawners compared to the initialnatural migrator group, there was no significant difference between exercise andnon-exercise males (ANOVA, P> 0�05).There was no difference in gonads constituents among natal arrival, exercise
and non-exercise males (MANOVA, P> 0�05) or females (MANOVA, P> 0�05)[Fig. 1(c)]. Maternal and paternal investment in gonad mass was similar amongthe three groups (MANOVA, P> 0�05). Individual egg size increased two-foldfrom Franklin Rock for all groups during the course of the experiment[Fig. 1(d)], but there was no difference in either the amount of dry egg matterdeposited or in the number of eggs among the natal and captive groups(ANOVA, P> 0�05).
PHYSIOLOGY
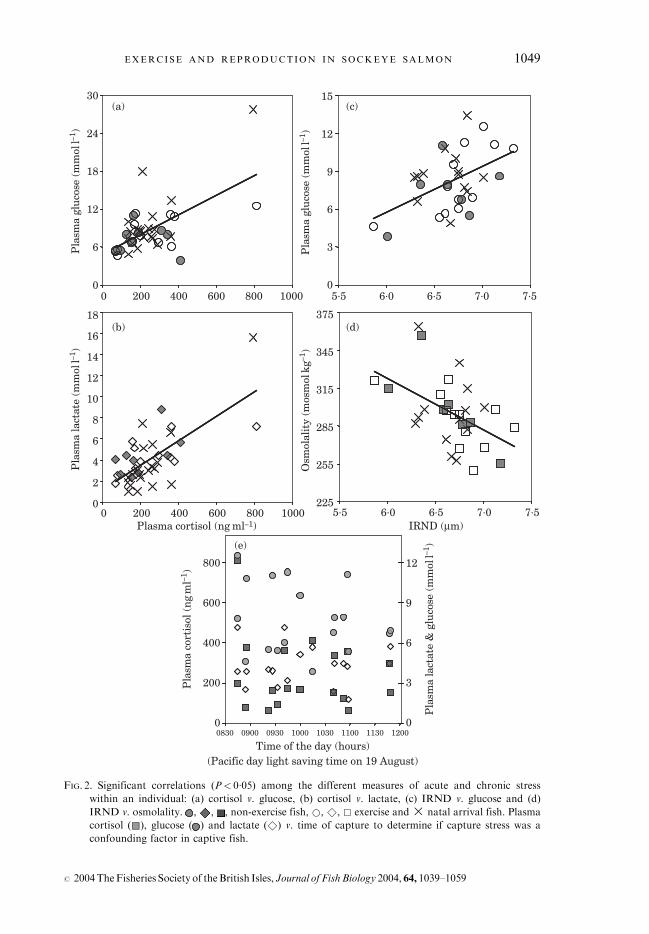
The overall levels for all of the stress variables analysed from both captivegroups and natal arrivals were high (Table II), but there was no evidence of anexercise effect (MANOVA, P¼ 0�96). Among individual fish, significant posi-tive correlations (P< 0�05) existed between cortisol and glucose [Fig. 2(a)],cortisol and lactate [Fig. 2(b)], and IRND and glucose [Fig. 2(c)]. A significantnegative relationship (P< 0�05) existed between IRND and osmolality[Fig. 2(d)]. There was no significant exercise effect in either females [Fig. 3(a)]or males [Fig. 3(b)] with respect to any of the reproductive hormones analysed(MANOVA, P> 0�05). Females from both captive groups with non-ovulatedeggs had low levels of 17–20 P [Fig. 3(a); F3, F5, F7, F8].
REPRODUCTION
On 19 August, pass the date where natal females had already matured andspawned, only 12 of 28 captive females had reached sexually maturity. Exercisefemales (nine of 14) were more likely (w2-test, P< 0�05) to be mature than non-exercise females (three of 14) at this date (Table III). Also, twice as manyexercise females (10) produced ovulated eggs, irrespective of date of death(w2-test, P< 0�05). There was no difference in longevity between non-exercise
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1045
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

(28 days) and exercise females (29 days) (w2-test, P> 0�05). Therefore, given theorder of maturating was natal, exercise and non-exercise, and there were noegg size differences, egg growth rates were probably fastest in natal females,followed by exercise females and slowest in non-exercise females.
TABLE I. Mean� S.E. values for the morphology variables of body metrics, body andgonad constituents for non-exercise, exercise and natal arrival sockeye salmon. TheF-value for each group of variables represents the overall MANOVA test result (bold).The ANOVA F-value associated with each individual variable is also included. Gonadenergy density is not included as response variables in the MANOVA because of the
high pair-wise correlations with other variables in the section
Females
Morphology parameters Non-exercise Exercise Natal arrival
Metrics 0�88 0�515Standard length (LS) (cm) 52�1� 0�8 53�0� 0�6 52�0� 0�7 0�56 0�577Whole body mass (g) 2134� 99 2234� 94 1987� 99 1�40 0�259Condition factor (K) 1�50� 0�04 1�49� 0�03 1�40� 0�03 1�93 0�161
Body constituents 6�09 0�001% Moisture 74�5� 0�5 77�2� 0�8 79�2� 0�4 15�08 0�001% Protein 15�4� 0�4 14�4� 0�3 15�5� 0�3 3�07 0�059% Lipid 6�5� 0�5 4�5� 0�5 1�8� 0�3 26�17 0�001Energy density (MJkg�1) 6�2� 0�2 5�2� 0�2 4�4� 0�1 18�80 0�001
Gonad constituents 0�53 0�781% Moisture 63�7� 1�6 63�3� 1�1 62�2� 0�6 0�79 0�466% Protein 22�8� 1�2 23�2� 0�9 23�8� 0�4 0�50 0�614% Lipid 11�2� 0�5 10�9� 0�4 11�5� 0�2 1�13 0�338Energy density (MJkg�1) 9�8� 0�4 9�8� 0�3 10�2� 0�2Sample size 14 14 10
Males
Metrics 1�08 0�395Standard Length (LS) (cm) 55�5� 1�4 55�5� 1�4 54�5� 0�6 0�32 0�731Whole body mass (g) 2568� 219 2781� 187 2444� 126 1�00 0�388Condition factor (K) 1�48� 0�03 1�62� 0�04 1�50� 0�03 3�57 0�049
Body constituents 7�39 0�001% Moisture 76�3� 1�2 77�6� 1�4 79�7� 0�4 3�70 0�044% Protein 14�5� 0�3 13�9� 0�7 15�1� 0�3 1�95 0�170% Lipid 6�4� 0�6 5�8� 0�6 1�6� 0�2 44�05 0�001Energy density (MJkg�1) 5�9� 0�2 5�6� 0�4 4�2� 0�1 18�82 0�001
Gonad constituents 1�73 0�145% Moisture 75�6� 1�0 76�0� 1�1 78�2� 0�7 2�68 0�094% Protein 22�6� 1�2 22�2� 1�2 19�5� 0�6 3�46 0�052% Lipid 1�7� 0�1 2�0� 0�1 1�8� 0�1 1�77 0�197Energy density (MJkg�1) 6�0� 0�3 6�1� 0�3 5�3� 0�2Sample size 6 6 10
1046 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

Based on survival of eggs to the eyed staged for each unique pairing, thegamete viability from captive individual male and female parents differed sig-nificantly (GLM, P< 0�05). The majority of egg mortality occurred betweenfertilization and the eyed stage, and overall survival declined marginally from57�6 to 52�4% from the eyed stage to MAWW. The three highest egg survivalrates all came from exercise females [Fig. 4(a)]. There was no overall significanttreatment effect, however, for either females (P> 0�05) or males (P> 0�05)[Fig. 4(b)] from cross-fertilizations made on 19 August. Moreover, none of theparental morphological, physiological or reproductive variables were signifi-cantly correlated with gamete survival (all r-values P> 0�05; values not given).Overall egg survival to eyed stage for natal spawners was 88�2%, significantlyhigher than captive fish (ANOVA, P< 0�05), but there were no differences ingamete survival based on parental origin (GLM, P> 0�05).
10
8
6
4
2
0
30
25
20
15
10
5
0
10
8
6
4
2
0
30
35
25
20
15
10
5
0
a
b
c
d
x x x
a a a
a
b
c
d
a
b bb
(a)
(c)
(b)
(d)
FranklinRock
Non-exercise
Exercise Natalarrival
Non-excercise Exercise Natal arrival
FranklinRock
Non-exercise
Exercise Natalarrival
FranklinRock
Non-exercise
Exercise Natalarrival
Natalspawner
Ova
ry %
pro
tein
an
d %
lipi
dB
ody
ener
gy d
ensi
ty (
MJ
kg–1
)
Dry
egg
mas
s (m
g)B
ody
lipid
(%
Com
posi
tion
)b
FIG. 1. (a) Female body energy density MJkg�1 and (b) per cent body lipid composition density by
sample group. (c) Female ovary composition of % lipid ( ) and % protein ( ) and (d) dry egg
mass by location and sample group. Means� S.E. Common lower case letters indicate no significant
difference among group means (ANOVA; Tukey’s test).
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1047
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

TABLE II. Mean� S.E. values for the physiological variables of stress and reproductivehormones for non-exercise, exercise and natal arrival sockeye salmon. The F-value inbold for each group of variables represents the overall MANOVA test result and theANOVA F-value associated with each individual variable is also given. Cortisol and
reproductive hormone data were log10-transformed for statistical analysis
Females
Non-exercise Exercise Natal arrival F P
StressCortisol (ngml�1) 256�8� 62�7 244�3� 34�4 292�3� 59�8Lactate (mmol l�1) 5�4� 0�9 4�8� 0�5 4�4� 1�4Glucose (mmol l�1) 7�4� 1�2 8�5� 0�8 11�6� 2�1Osmolality (mosmol kg�1) 300�4� 12�8 286�6� 8�8 286� 4�8IRND (mm) 6�61� 0�19 6�86� 0�06 6�52� 0�11
Reproductive 0�74 0�54917, 20 P (ngml�1)* 335�9� 327�5 418�4� 201�9 110�5� 51�3 1�62 0�223Testosterone (ngml�1) 136�8� 49�1 128�8� 26�7 128�3� 35�8 0�14 0�868Sample size 5 7 10
Males
StressCortisol (ngml�1) 139�6� 20�0 280�7� 144�4 206�6� 17�2Lactate (mmol l�1) 3�1� 0�5 3�6� 1�0 3�2� 0�4Glucose (mmol l�1) 7�1� 0�6 7�8� 1�6 7�8� 0�5Osmolality (mosmol kg�1) 298�5� 20�3 296�0� 9�4 301�0� 9�4IRND (mm) 6�67� 0�14 6�59� 0�27 6�7� 0�1
Reproductive 0�52 0�78917, 20 P (ngml�1)* 68�4� 26�3 45�9� 19�2 32�0� 8�5 0�46 0�640Testosterone (ngml�1) 45�3� 10�9 68�4� 44�3 55�5� 8�6 0�46 0�64111 keto-testosterone (ngml�1) 96�6� 41�2 114�3� 63�3 100�7� 9�0 1�01 0�389
5 4 10
Males and females combined
Stress 1�29 0�259Cortisol (ngml�1) 204�7� 68�6 259�5� 59�7 249�5� 63�5 0�62 0�544Lactate (mmol l�1) 4�4� 0�7 4�3� 0�5 3�8� 1�4 0�23 0�796Glucose (mmol l�1) 7�3� 0�9 8�2� 0�8 9�7� 2�2 1�24 0�301Osmolality (mosmol kg�1) 299�6� 9�3 290�5� 6�3 293�5� 10�8 0�33 0�723IRND (mm)** 6�6� 0�1 6�8� 0�1 6�6� 0�1 0�50 0�610Sample size 9 12 20
*, 17-alpha 20-beta dihydroxy-4-pregen-3-one.
**, sample size for IRND natal fish is 13.
1048 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059
30(a) (c)
(d)
(e)
(b)
24
18
12
6
00 200 400 600 800 1000
Pla
sma
gluc
ose
(mm
ol l–1
)
15
12
9
6
3
05.5 6.0 6.5 7.0 7.5
Pla
sma
gluc
ose
(mm
ol l–1
)375
345
315
285
255
2255.5 6.0 6.5 7.0 7.5
Osm
olal
ity
(mos
mol
kg–1
)18
16
14
12
10
6
4
2
8
00 200 400 600 800 1000
Pla
sma
lact
ate
(mm
ol l–1
)
Pla
sma
cort
isol
(n
g m
l–1)
Pla
sma
lact
ate
& g
luco
se (
mm
ol l–1
)
800
600
400
200
00830 0900 0930 1000 1030 1100 1130 1200
12
9
6
3
0
Time of the day (hours)
Plasma cortisol (ng ml–1) IRND (µm)
(Pacific day light saving time on 19 August)
FIG. 2. Significant correlations (P< 0�05) among the different measures of acute and chronic stress
within an individual: (a) cortisol v. glucose, (b) cortisol v. lactate, (c) IRND v. glucose and (d)
IRND v. osmolality. , , , non-exercise fish, *, &, & exercise and� natal arrival fish. Plasma
cortisol (&&), glucose ( ) and lactate ( &) v. time of capture to determine if capture stress was a
confounding factor in captive fish.
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1049
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

DISCUSSION
The voluntary swimming of fish under moderate flow conditions during thefinal 4 weeks of maturation had a significant effect on selected morphologicaland reproductive variables compared with non-exercise fish. Importantly,energy density for non-exercise fish was 50% higher than that of natal fishand 20% higher than that for exercise fish. None of the physiological stressmeasurements taken at the end of the experiment, however, were different forexercise, non-exercise or natal arrival sockeye salmon. In the absence of a water
10000
1000
100
10
1
1000
100
10
1
F2 F3 F8 F9 F13 F1 F5 F6 F7 F10 F11 F12 F16 F17 F18 F19 F20 F21 F22 F23 F24 F25
M4 M5 M6 M8 M1 M2 M3 M7 M9 M10 M11 M12 F13 M14 M15 M16 M17 M18 M19
Pla
sma
leve
ls o
f re
prod
ucti
ve h
orm
ones
(n
g m
l–1)
Non-exercise Exercise Natal arrival
Non-exercise Exercise Natal arrival
(a)
(b)
Fish number
FIG. 3. Reproductive hormone values for 17-a 20-b dihydroxy-4-pregen-3-one (17,20P) ( ), testosterone
( &), and 11-keto-testosterone (�) are given for each individual fish sampled to demonstrate large
differences among individuals group by (a) females and (b) males. The fish are organized by treatment
group to show that no observed pattern exists with respect to treatment (MANOVA, P> 0�05).
1050 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059
current, salmonids are reported to swim continuously near 1BL s�1 (Petrellet al., 1997), but this response was not observed in the non-exercise sockeyesalmon used in this experiment. The exercise fish in this experiment averaged0�55m s�1 (1 BL s�1), which approximates the lower range of swim speedsattained over the natural migration of Early Stuart sockeye salmon in theFraser River (Hinch & Rand, 1998). The natural migrators average swimspeed increases from 0�6m s�1 in the lower river stages of migration to1�2m s�1 during the upper river portions and is characterized by periods of
TABLE III. Mean� S.E. values for the reproductive variables of gonads, egg metrics andreproductive success for non-exercise, exercise, natal arrival and natal spawner females.The F-value in bold for each group of variables represents the overall MANOVA testresult and the ANOVA F-value associated with each individual variable is also given.No overall MANOVA was performed on egg variables because of the high correlationamong the egg size variables (r> 0�9). w2 analysis was performed on binomial data
Variables Non-exercise ExerciseNatalarrivals
Natalspawners F P
Maternal 1�66 0�143Gonad mass (g) 282�8� 18�7 339�7� 16�2 295�5� 23�3 2�62 0�087IG 13�3� 0�7 15�3� 0�6 14�7� 0�6 3�16 0�055Fecundity 3896� 240 4044� 130 3998� 253 0�11 0�895
Egg variablesEgg dry mass (mg) 29�66� 1�23 32�25� 0�92 29�26� 0�57 30�98� 0�79 1�92 0�138Egg wet mass (mg) 73�71� 4�16 84�03� 2�82 73�41� 1�51 85�60� 2�30Egg diameter (mm) 4�56� 0�09 4�79� 0�05 4�53� 0�03 4�75� 0�06
Female reproductivestatusNumber ovulated by 19
August3 9 5�25 0�022
Number non-ovulatedat natural death
7 2 7�78 0�005
Number ovulated beforedeath*
5 10 4�44 0�035
Mean time tomaturity
32 29
Sacrificed 7 7Died naturally 7 7
Sample size 14 14 10 16
Paternal variables 2�55 0�056Gonad mass (g) 49�4� 5�6 53�6� 5�3 60�9� 5�1 1�19 0�326IG 1�9� 0�2 1�9� 0�1 2�5� 0�1 4�45 0�026
Male reproductivestatusSacrificed 4 5Died naturally 2 1Sample size 6 6 10
*, w2 excluded fish killed with tight skeins.
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1051
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

very rapid swimming through difficult high velocity stretches of the river (Hinch& Rand, 1998). It is also important to note that the minimum cost of transportfor several stocks of Fraser River salmonids, including sockeye salmon, wasrecently found to be c. 1 BL s�1 (Lee et al., 2003). Therefore, captive exercisefish may have inadvertently been challenged with a swim speed that optimizedenergy utilization.
MORPHOLOGY
Although exercise fish did not face an equivalent challenge to the morerigorous natural migration environment, as confirmed by the lower energy
F9 F12 F2 F11 F4 F10 F1
M7Fish number
M4 M3 M5 M2 M1 M6
F6
100
75
50
25
0
100
75
50
25
0
Em
bryo
sur
viva
l to
eyed
sta
ge (
%)
(a)
(b)
A
A
B
B
A
A,B
A,B
A,BA,B
A,B
A,B,CB,C
B,C
B,CC
FIG. 4. Mean� S.E. survival to eyed stage for eachuniquepairing completedon19August 1999among exercise
(&) and non-exercise fish (&), organized by (a) captive dams (crossed with seven males) and (b) captive
sires (crossed with eight females). Common upper case letters indicate no significant difference among
means (Tukey’s test; n¼ 7 crosses for captive dams and n¼ 8 crosses for captive sires).
1052 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

densities of natal arrivals, conditions were sufficient to modulate energy andproximate body constituents of the sockeye salmon compared with the initialnatural migrator sample collection. The significant decrease in lipid reserves ofexercise females v. non-exercise females is consistent with previous studies demon-strating that freshwater migration is primarily powered by lipid catabolism(Idler & Clemens, 1959; Brett, 1973; G.T. Crossin, S.G. Hinch, A.P. Farrell,D.A. Higgs, A.G. Lotto, J.D. Oakes & M.C. Healey, pers. comm.).Despite the differences in whole body composition, the per cent composition
of lipids and protein in ovaries was not significantly different for natal arrivals,non-exercise and exercise females. The implication is that relative egg composi-tion was not compromised unduly by either the exercise or stress encounteredby the fish in the present study. Therefore, in contrast to the original hypothesis,releasing both the non-exercise and exercise captive fish from the more energetic-ally demanding in river migration did not result in a reallocation of energy intoreproduction because eggs were not larger and fecundity was not increased.Instead, sockeye egg growth rate appeared to be slowed in non-exercise fish,similar to an earlier finding for maturing rainbow trout Oncorhynchus mykiss(Walbaum) which found egg growth rates can be altered (Tyler et al., 1996).Whether a slower egg growth rate contributed to delay in reproductive matura-tion is unclear from the present study. Proximate egg composition, however, wasnot a factor in the observed effects of captivity on reproduction since there wereno differences with natal fish. Further experiments are needed to resolve whethereither gonad composition, fecundity or egg size decrease when migrationdifficulty exceeds the natural range experienced in-river (Kinnison et al., 2001).
PHYSIOLOGY
Surveys on Atlantic salmon Salmo salar L. at aquaculture facilities havefound that continuous swimming reduced stress (Petersen et al., 2001). Incontrast, no detected differences were found in stress levels for exercise andnon-exercise fish. In fact, all plasma variables were consistent with the fish beingstressed at the time of or as a result of blood sampling since all plasma lactatevalues were >1mmol l�1 (Jain et al., 1998), most plasma glucose values were>5�5mmol l�1(Donaldson et al., 2000), and all plasma cortisol values were>10 ngml�1 (Fagerlund et al., 1995). Nevertheless, these values for captivefish were comparable with those for natal arrival fish, all of which fall withinbaseline measures previously recorded for Early Stuart and Horsefly sockeyesalmon sampled at their natal spawning grounds (Macdonald, 2000; Macdonaldet al., 2000). Clearly the difficultly here is an inability to partition stress associatedwith the blood sampling event from the stress of holding wild salmonids incaptivity, not to mention the high stress associated with final maturation andnatural spawning (Kubokawa et al., 1999; Carruth et al., 2000). Sampling bloodfrom large adult salmonids is difficult and various approaches have been used tominimize stress prior to sampling (Booth et al., 1995; Farrell et al., 2001). Inaddition, sampling fish from a large tank throughout the day may have led tocumulative stress. There was no relationship, however, between sampling orderwith either plasma cortisol or plasma glucose [Fig. 2(e)]. Therefore, even if samplingstress was a confounding factor, there was no systematic effect of sampling order.
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1053
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

The positive correlations among individual values for cortisol and lactate,cortisol and glucose, and IRND and glucose strongly suggest that these acuteand chronic stress variables can collectively provide a reliable indicator ofoverall stress within an individual. The pair-wise comparisons of these stressorsamong individuals have not always been explicitly identified in the literature.Donaldson et al. (2000) noted that sockeye salmon in poor condition had higherlevels of lactate, glucose and cortisol, and large IRND’s than sockeye salmon ingood condition. An inverse correlation between osmolality and IRND providesfurther evidence that hypertrophy of interrenal cells and the resultant increase inplasma corticosteroids leads to tissue degeneration and whole body edema(Clarke & Hirano, 1995).
REPRODUCTION
The novel finding of the present work is the discovery of an exercise effect onthe reproductive development of wild female sockeye salmon in captivity, withexercise females being more likely to become sexually mature and at an earlierstage than non-exercise females. The present work indicates that exercise can beadded to the list of previous manipulations that have altered salmonid matura-tion timing. These include force-feeding, gonadectomies, chronic stress, accel-erating growth rates and hormone injections (McBride et al., 1963, 1965;Donaldson, 1990; Campbell et al., 1992).The mechanisms by which exercise improved reproductive development are
unclear. No studies have investigated the role of exercise in mobilizing lipidsand proteins as a means to alter to egg depositional rates. It is well recognizedthat exercise training does improve the catabolism of fatty acids in feeding fishes(Johnston & Moon, 1980a, b). Exercise also increases the deposition of lipids towhite muscles during the growth phase (East & Magnan, 1987). Sockeye salmonused in the present study, however, had stopped feeding and were depleting lipidstores (Table I). It is also possible that exercise stimulated vitellogenesis andfinal egg maturation, and so future studies may need to monitor hormonalcontrol of these events more closely than the present one. Although a mechan-ism cannot be assigned the present study supports the idea that the normalphysiological changes associated with the exercise during freshwater migrationin sockeye salmon may play a key role in regulating the timing of sexualmaturation.The overall confinement stress on both captive groups probably contributed
to delayed maturation, higher egg retention rates, and especially depressedoverall fertilization success compared with natal fish. The mean egg survivalfrom captive fish (56%) were lower than eggs from natal spawners (88%) andpublished values for wild sockeye salmon (>90%) (Hoysak & Liley, 2001;Galbraith, 2003). Campbell et al. (1994) reported that confinement stressreduced progeny survival from 96 to 54% in rainbow trout and 96 to 68% inbrown trout Salmo trutta L.. Similarly, the mortality of 38% captive femalesbefore producing ovulated eggs is well above the historical estimate of 13% forEarly Stuart sockeye salmon (Department of Fisheries and Oceans, unpubl.data). Exercise may act to partially ameliorate the effect of confinement stress(Jørgensen & Jobling, 1993; Petersen et al., 2001). None of the standard
1054 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

physiological stress responses measured, however, were different betweenexercise and non-exercise fish. Therefore, confinement stress alone is unlikelyto account for the significant differences observed here for reproductivedevelopment between exercise and non-exercise fish.Consistent with other research, there were larger differences in viability
among eggs from different females than among milt from different males(Nagler et al., 2000; Hoysak & Liley, 2001). This individual variability wasnot correlated with any of the morphological, physiological or reproductivevariables recorded from each parent. The possible physiological mechanismsconnecting stress variables, reproductive endocrinology and ultimately repro-ductive success have been discussed by others (Donaldson, 1990; Pottinger &Pickering, 1990; Campbell et al., 1992; Pankhurst & Van Der Kraak, 1997;Schreck et al., 2001). These studies found a consistent association betweenhigh plasma cortisol levels and suppressed levels of androgens and oestrogens.Pottinger & Pickering (1990) went further by using cortisol implants to increaseplasma cortisol levels, which changed oestradiol-binding capacity. This implieda possible mechanism whereby stress can suppress vitellogenesis and therebydelaying maturation timing and ovulation (Campbell et al., 1992). No correl-ation was found in the present study, however, between either high cortisolvalues or large IRNDs (chronic stress) and reproductive endocrine suppressionduring this one-time plasma sampling design. Similarly, Campbell et al. (1994)did not always find a connection between stress and gonadal steroid suppres-sion, but did find a reduction in fertilization success. Sustained high levels ofcirculating cortisol, prior to sampling, may have suppressed 17-a 20-b dihy-droxy-4-pregen-3-one production in non-ovulated captive females, all of whichhad low levels of this hormone (17, 20 P< 13 ngml�1). Temporal non-lethalplasma sampling maybe required to provide empirical evidence of a rate changein vitellogenesis associated with particular stressors. Future work should con-centrate on separating confinement stress from exercise manipulation. Thiscould be accomplished by capturing fish in the river, tagging and re-releasingthe fish above and below the capture point to both decrease and increase energyexpenditure without introducing a confinement stress.In summary, wild sockeye salmon can be intercepted during upstream migra-
tion and then swum in captivity under different water conditions to maturity.This type of manipulation is appropriate for a controlled assessment of energyallocations and to monitor the timing of different physiological changes asso-ciated migration, maturation and senescence. The main finding was that reduc-ing the high energy expenditure associated with a natural migration did notresult in a reallocation of energy into greater reproductive investment. Femalesockeye salmon, however, invested lipids and proteins into gonads at a differentrates (natal> exercise> non-exercise) without compromising the relative percent compositional makeup. A possible consequence of this rate differencewas the inability of some exercise females and half of the non-exercise femalesto reach full sexual maturity before dying. While stress seemed unlikely to havecontributed to the difference in reproductive development between exercise andnon-exercise fish, confinement stress probably played a role in the reducedoverall reproductive success of captive v. natal fish. Ultimately, a lack ofexercise impaired the reproductive performance of sockeye salmon, suggesting
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1055
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

exercise is an important component in ensuring successful sexual maturationand maximizing reproductive success in wild female sockeye salmon.
The authors would like to acknowledge the following people for their invaluablecontributions: Yale First Nations and R. Galbraith for their Herculean efforts in catch-ing and transporting fish in Fraser Canyon; J. Beckett, H. Herunter, K. Mullen,B. Jensen for their assistance in broodline generation; J. Oakes and D. Higgs for proximateanalysis; E. Donaldson and J. Smith for blood chemistry and reproductive hormoneanalysis. Special thanks go to D. Barnes for his assistance in sockeye salmon sampling,plasma collection and IRND reading; and to E. MacIsaac for his continuous support ofthis project. This study was funded by contributions from an NSERC Strategic grant toM.C. Healey, S.G. Hinch, A.P. Farrell, Forest Renewal British Columbia grant toJ.S. Macdonald, and DFO Freshwater Habitat Science Environmental Watch Program.
References
Anderson, R. O. & Neumann, R. M. (1996). Length, weight, and associated structuralindices. In Fisheries Techniques, 2nd edn. (Murphy, B. R. & Willis, D. W., eds),pp. 447–482. Bethesda, MD: American Fisheries Society.
Berg, O. K., Hendry, A. P., Svendsen, B., Bech, C., Arnekleiv, J. V. & Lohrmann, A. (2001).Maternal provisioning of offspring and the use of those resources during ontogeny:variation within and between Atlantic salmon families. Functional Ecology 15, 13–23.
Booth, R. K., Kieffer, J. D., Davidson, K., Bielak, A. T. & Tufts, B. L. (1995). Effects oflate-season catch and release angling on anaerobic metabolism, acid–base status,survival, and gamete viability in wild Atlantic salmon (Salmon salar). CanadianJournal of Fisheries and Aquatic Sciences 52, 283–290.
Brett, J. R. (1973). Energy expenditure of sockeye salmon, Oncorhynchus nerka, duringsustainedperformance. Journal of theFisheriesResearchBoard ofCanada 30,1799–1809.
Brett, J. R. (1995). Energetics. In Physiological Ecology of Pacific Salmon (Groot, C.,Margolis, L. & Clarke, W. C., eds), pp. 3–68. Vancouver: University of BritishColumbia Press.
Campbell, P. M., Pottinger, T. G. & Sumpter, J. P. (1992). Stress reduces the quality ofgametes produced by rainbow trout. Biological Reproduction 47, 1140–1150.
Campbell, P. M., Pottinger, T. G. & Sumpter, J. P. (1994). Preliminary evidence thatchronic confinement stress reduces the quality of gametes produced by brown andrainbow trout. Aquaculture 120, 151–169.
Carruth, L. L., Dores, R. M., Maldonado, T. A., Norris, D. O., Ruth, T. & Jones, R. E.(2000). Elevation of plasma cortisol during the spawning migration of landlockedkokanee salmon (Oncorhynchus nerka kennerlyi). Comparative Biochemistry andPhysiology C 127, 123–131.
Clarke, W. C. & Hirano, T. (1995). Osmoregulation. In Physiological Ecology of PacificSalmon (Groot, C., Margolis, L. & Clarke, W. C., eds), pp. 317–377. Vancouver:University of British Columbia Press.
Donaldson, E. M. (1990). Reproductive indices as measures of the effects ofenvironmental stressors in fish. American Fisheries Society Symposium 8, 109–122.
Donaldson, E. M., Smith, J., Barnes, D., Clarke, W. C., Gordon, R. & Martens, D.(2000). Physiological and endocrine changes during the anadromous migration ofEarly Stuart sockeye salmon (Oncorhynchus nerka) in 1997. InMortality during theMigration of Fraser River Sockeye Salmon (Oncorhynchus nerka): a Study of theEffect of Ocean and River Environmental Conditions in 1997 (Macdonald, J. S., ed.).Canadian Technical Report of Fisheries and Aquatic Sciences 2315, 67–90.
East, P. & Magnan, P. (1987). The effect of locomotor activity on the growth of brookcharr, Salvelinus fontinalis Mitchell. Canadian Journal of Zoology 65, 843–846.
Fagerlund, U. H. M., McBride, J. R. & Stone, E. T. (1981). Stress-related effects ofhatchery-rearing density in coho salmon. Transactions of the American FisheriesSociety 110, 644–649.
1056 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

Fagerlund, U. H. M., McBride, J. R. & Williams, I. V. (1995). Stress and tolerance.In Physiological Ecology of Pacific Salmon (Groot, C., Margolis, L. & Clarke,W. C.,eds), pp. 459–503. Vancouver: University of British Columbia Press.
Farrell, A. P., Gallaugher, P. E. & Routledge, R. (2001). Rapid recovery of exhaustedadult coho salmon after commercial capture by troll fishing. Canadian Journal ofFisheries and Aquatic Sciences 58, 2319–2324.
Galbraith, R. V. (2003). Suspended sediment effects on fertilization success in PacificSalmon (Oncorhynchus spp.). MSc Thesis. Simon Fraser University, Canada.
Gilhousen, P. (1980). Energy sources and expenditures in Fraser River sockeyesalmon during their spawning migration. International Pacific Salmon FisheriesCommission, Bulletin XXII.
Herunter, H. E., Pedersen, L. F., Patterson, D. A. & Galbraith, R. V. (2000). Early Stuartsockeye salmon (Oncorhynchus nerka) egg quality and egg-to-alevin survival for the1997 escapement. In Mortality during the Migration of Fraser River SockeyeSalmon (Oncorhynchus nerka): a Study of the Effect of Ocean and RiverEnvironmental Conditions in 1997 (Macdonald, J. S., ed.). Canadian TechnicalReport of Fisheries and Aquatic Sciences 2315, 105–116.
Higgs, D. A., Macdonald, J. S., Levings, C. D. & Dosanjh, B. S. (1995). Nutrition andfeeding habits in relation to life-history stage. In Physiological Ecology of PacificSalmon (Groot, C., Margolis, L. & Clarke, W. C., eds), pp. 159–315. Vancouver:University of British Columbia Press.
Higgs, D. A., Rowshandeli, M., Greig, M., Barnes, D. P. & Macdonald, J. S. (2000).Changes in the proximate constituents and gross energy in the muscle (fillet)of 4 year-old Early Stuart sockeye salmon (Oncorhynchus nerka) during the1997 upstream Fraser River spawning migration. In Mortality during the Migrationof Fraser River Sockeye Salmon (Oncorhynchus nerka): a Study of the Effect ofOcean and River Environmental Conditions in 1997 (Macdonald, J. S., ed.).Canadian Technical Report of Fisheries and Aquatic Sciences 2315, 91–104.
Hinch, S. G. & Rand, P. S. (1998). Swim speeds and energy use of upriver-migratingsockeye salmon (Oncorhynchus nerka): role of local environment and fishcharacteristics. Canadian Journal of Fisheries and Aquatic Sciences 55, 1821–1831.
Hodgson, S. & Quinn, T. P. (2002). The timing of adult sockeye salmon migrations intofresh water: adaptations by populations to prevailing thermal regimes. CanadianJournal of Zoology 80, 542–555.
Høgasen, R. H. (1998). Physiological changes associated with the diadromous migrationof salmonids. Canadian Special Publication of Fisheries and Aquatic Sciences 127.
Hoysak, D. J. & Liley, R. N. (2001). Fertilization dynamics in sockeye salmon and acomparison of sperm from alternative male phenotypes. Journal of Fish Biology 58,1286–1300. doi: 10.1006/jfbi.2000.1536.
Hurlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments.Ecological Monographs 54, 187–211.
Idler, D. R. & Clemens, W. A. (1959). The energy expenditures of Fraser River sockeyesalmon during the spawning migration to Chilko and Stuart Lakes. InternationalPacific Salmon Fisheries Commission, Progress Report 6.
Jain, K. E., Birtwell, I. K. & Farrell, A. P. (1998). Repeat swimming performance ofmature sockeye salmon following a brief recovery period: a proposed measure offish health and water quality. Canadian Journal of Zoology 76, 1488–1496.
Jensen, J. O. T. & Alderdice, D. F. (1989). Comparison of mechanical shock sensitivity ofeggs of five Pacific salmon (Oncorhynchus) species and steelhead trout (Salmogairdneri). Aquaculture 78, 163–181.
Johnston, I.A.&Moon,T.W. (1980a). Endurance exercise training in the fast and slowmusclesof a teleost fish (Pollachius virens). Journal of Comparative Physiology, B 135, 147–156.
Johnston, I. A. & Moon, T. W. (1980b). Exercise training in skeletal muscle of brooktrout (Salvelinus fontalis). Journal of Experimental Biology 87, 177–194.
Jørgensen, E. H. & Jobling, M. (1993). The effects of exercise on growth, food utilizationand osmoregulatory capacity of juvenile Atlantic salmon, Salmo salar. Aquaculture116, 233–246.
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1057
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059

Kinnison, M. T., Unwin, M. J., Hendry, A. P. & Quinn, T. P. (2001). Migratory costsand the evolution of egg size and number in introduced and indigenous salmonpopulations. Evolution 55, 1656–1667.
Kubokawa, K., Watanabe, T., Yoshioka, M. & Iwata, M. (1999). Effects of acutestress on plasma cortisol, sex steroid hormone and glucose levels in male andfemale sockeye salmon during the breeding season. Aquaculture 172, 335–349.
Lee, C. G., Farrell, A. P., Lotto, A., MacNutt, M. J., Hinch, S. G. & Healey, M. C. (2003).The effect of temperature on swimming performance and oxygen consumption inadult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon stocks. Journal ofExperimental Biology 203, 3239–3251.
Macdonald, J. S. (2000). Mortality during the migration of Fraser River sockeye salmon(Oncorhynchus nerka): a study of the effect of ocean and river environmentalconditions in 1997. Canadian Technical Report of Fisheries and Aquatic Sciences 2315.
Macdonald, J. S., Levings, C. D., McAllister, C. D. S., Fagerlund, U. H.M., McBride, J. R.(1988). A field experiment to test the importance of estuaries for chinook salmon(Oncorhynchus tshawytscha) survival: short-term results. Canadian Journal ofFisheries and Aquatic Sciences 45, 1366–1377.
Macdonald, J. S., Foreman, M. G. G., Farrell, T., Williams, I. V., Grout, J., Cass, A.,Woodey, J. C., Enzenhofer, H., Clarke, W. C., Houtman, R., Donaldson, E. M. &Barnes, D. (2000). The influence of extreme water temperatures on migratingFraser River sockeye salmon (Oncorhynchus nerka) during the 1998 spawningseason. Canadian Technical Report of Fisheries and Aquatic Sciences 2326.
McBride, J. R., Fagerlund, U. H. M., Smith, M. & Tomlinson, N. (1963). Resumption offeeding by and survival of adult sockeye salmon (Oncorhynchus nerka) and theeffect of gonadectomy. Journal of the Fisheries Research Board of Canada 20,95–100.
McBride, J. R., Fagerlund, U. H. M., Smith, M. & Tomlinson, N. (1965). Post-spawningdeath of Pacific salmon. Sockeye salmon (Oncorhynchus nerka) maturing andspawning in captivity. Journal of the Fisheries Research Board of Canada 22,775–785.
Nagler, J. J., Parsons, J. E. & Cloud, J. G. (2000). Single pair mating indicates maternaleffects on embryo survival in rainbow trout, Oncorhynchus mykiss. Aquaculture184, 177–183.
Novales Flamarique, I. & Harrower, W. L. (1999). Mortality of sockeye salmon raisedunder light backgrounds of different spectral composition. Environmental Biologyof Fishes 55, 279–293.
Pankhurst, N. W. & Van Der Kraak, G. (1997). Effects of stress on reproduction andgrowthof fish. InFishStress andHealth inAquaculture (Iwama,G.K., Pickering,A.D.,Sumpter, J. P. & Schreck, C. B., eds), pp. 73–93. Cambridge: CambridgeUniversity Press.
Patterson, D. A. & Farrell, A. P. (2003). Intergenerational effects: the relationshipof adult migration conditions land-use impacts and offspring survival. In ACompendium of Research from the Stuart-Takla Fish-Forestry Interaction Study,(Year 2000) (MacIsaac, E. A., ed.). Canadian Technical Report of Fisheries andAquatic Sciences 2509.
Petersen, R. H., Page, F., Steeves, G. D., Wildfish, D. J., Harmon, P. & Loiser, R. (2001).A survey of 20 Atlantic salmon farms in the Bay of Fundy: influence ofenvironmental and husbandry variables on performance. Canadian TechnicalReport of Fisheries and Aquatic Sciences 2337.
Petrell, R. J., Shi, X., Ward, R. K., Naiberg, A. & Savage, C. R. (1997). Determining fishsize and swimming speed in cages and tanks using simple video techniques.Aquacultural Engineering 16, 45–62.
Pickering, A. D. (1992). Rainbow trout husbandry: management of the stress response.Aquaculture 100, 125–139.
Pottinger, T. G. & Pickering, A. D. (1990). The effect of cortisol administration onhepatic and plasma estradiol-binding capacity in immature female rainbow trout(Oncorhynchus mykiss). General and Comparative Endocrinology 80, 264–273.
1058 D. A . PATTERSON ET AL .
# 2004TheFisheries Society of the British Isles, Journal of FishBiology 2004, 64, 1039–1059

Schreck, C. B., Contreras-Sanchez, W. & Fitzpatrick, M. S. (2001). Effects of stress onfish reproduction, gamete quality, and progeny. Aquaculture 197, 3–24.
Smith, C. C. & Fretwell, S. D. (1974). The optimal balance between size and number ofoffspring. American Naturalist 108, 499–506.
Sokal, R. R. & Rohlf, F. J. (2000). Biometry, 3rd edn. New York: W. H. Freeman andCompany.
Taranger, G. L. & Hansen, T. (1993). Ovulation and egg survival following exposure ofAtlantic salmon, Salmo salar L., broodstock to different temperatures. Aquacultureand Fisheries Management 24, 151–156.
Thorarensen, H., Gallaugher, P. E., Kiessling, A. K. & Farrell, A. P. (1993). Intestinalblood flow in swimming chinook salmon Oncorhynchus tshawytscha and the effectsof haematocrit on blood flow distribution. Journal of Experimental Biology 179,115–129.
Tierney, K. (2000). Repeated swimming performance of sockeye, coho, and rainbow troutin varying conditions. MSc Thesis, Simon Fraser University, Canada.
Tyler, C. R., Nagler, J. J., Pottinger, T. G. & Turner, M. A. (1994). Effects of unilateralovariectomy on recruitment and growth of follicles in the rainbow trout,Oncorhynchus mykiss. Fish Physiology and Biochemistry 13, 309–316.
Tyler, C. R., Pottinger, T. G., Santos, E., Sumpter, J. P., Price, S.-A., Brooks, S. &Nagler, J. J. (1996). Mechanisms controlling egg size and number in rainbowtrout, Oncorhynchus mykiss. Biology of Reproduction 54, 8–15.
Webb, P. W. (1995). Locomotion. In Physiological Ecology of Pacific Salmon (Groot, C.,Margolis, L. & Clarke, W. C., eds), pp. 69–99. Vancouver: University of BritishColumbia Press.
West, C. J. & Larkin, P. A. (1987). Evidence for size-selective mortality of juvenilesockeye salmon (Oncorhynchus nerka) in Babine Lake, British Columbia. CanadianJournal of Fisheries and Aquatic Sciences 44, 712–721.
EXERCISE AND REPRODUCTION IN SOCKEYE SALMON 1059
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 1039–1059


















