The Duct Cell in Cystic Fibrosis
14
17 The Duct Cell in Cystic Fibrosis ANN HARRIS a Paediatric Molecular Genetics, Institute of Molecular Medicine, Oxford University, John Radcliffe Hospital, Oxford OX3 9DS, UK ABSTRACT: The pancreatic duct cell is central to the etiology of cystic fibrosis (CF) and is the site where pathology commences in utero. We have evaluated expression of the cystic fibrosis transmembrane conductance regulator gene (CFTR) through human development and shown it to be expressed from the early mid-trimester, with highest levels in the most distal portion of the devel- oping duct system and in centroacinar cells. The precise cause of pancreatic de- struction in CF is thought to be the obstruction of pancreatic ducts with inspissated secretions. We have shown that the MUC6 mucin is a significant component of the material that obstructs the ducts and that the MUC6 gene shows a very similar pattern of expression to that of CFTR in the developing pancreas. These observations provide a starting point for investigating how mutations in CFTR lead to obstruction of the pancreatic ducts in CF. INTRODUCTION Cystic fibrosis (CF) is the result of mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) protein, a small conductance cAMP-activated chlo- ride ion channel. The protein is expressed in epithelial cells within the serous portion of the submucosal glands of the lung, intestinal crypts, sweat gland ducts, male gen- ital ducts, bile and pancreatic ducts, among other sites. 1–4 A primary function of CFTR is chloride secretion and failure of a cAMP-mediated chloride secretion is a feature of CF epithelial cells. The involvement of CFTR in the pancreatic duct is more complex as the primary role of this epithelium is in bicarbonate secretion. 5 The bicarbonate concentration in pancreatic duct secretions from CF patients is markedly reduced. 6,7 The exocrine portion of the pancreas is comprised of enzyme-secreting acini and ducts lined by epithelial cells that secrete a bicarbonate-rich fluid. The ductal tree extends from centroacinar cells within the acini, through intralobular and interlobu- lar ducts into the main pancreatic duct. Pancreatic pathology is one of the earliest features of CF, and deposition of material within the pancreatic ducts can be ob- served by the mid-trimester of human gestation. 8 85% of CF patients are pancreatic insufficient from birth and all show some pancreatic pathology. Obstruction of pan- creatic ducts in the mid-trimester of development is followed by acinar autolysis as the acini differentiate and produce pancreatic enzymes by about 24 weeks of gesta- tion. We have been particularly interested in dissecting the molecular basis of the early pathology of CF pancreatic disease. a Phone, +44 1865 222341; fax, +44 1865 222626; e-mail, [email protected]
-
Upload
ann-harris -
Category
Documents
-
view
213 -
download
1
Transcript of The Duct Cell in Cystic Fibrosis

17
The Duct Cell in Cystic Fibrosis
ANN HARRISa
Paediatric Molecular Genetics, Institute of Molecular Medicine, Oxford University, John Radcliffe Hospital, Oxford OX3 9DS, UK
ABSTRACT: The pancreatic duct cell is central to the etiology of cystic fibrosis(CF) and is the site where pathology commences in utero. We have evaluatedexpression of the cystic fibrosis transmembrane conductance regulator gene(CFTR) through human development and shown it to be expressed from theearly mid-trimester, with highest levels in the most distal portion of the devel-oping duct system and in centroacinar cells. The precise cause of pancreatic de-struction in CF is thought to be the obstruction of pancreatic ducts withinspissated secretions. We have shown that the MUC6 mucin is a significantcomponent of the material that obstructs the ducts and that the MUC6 geneshows a very similar pattern of expression to that of CFTR in the developingpancreas. These observations provide a starting point for investigating howmutations in CFTR lead to obstruction of the pancreatic ducts in CF.
INTRODUCTION
Cystic fibrosis (CF) is the result of mutations in the cystic fibrosis transmembraneconductance regulator (CFTR) protein, a small conductance cAMP-activated chlo-ride ion channel. The protein is expressed in epithelial cells within the serous portionof the submucosal glands of the lung, intestinal crypts, sweat gland ducts, male gen-ital ducts, bile and pancreatic ducts, among other sites.1–4 A primary function ofCFTR is chloride secretion and failure of a cAMP-mediated chloride secretion is afeature of CF epithelial cells. The involvement of CFTR in the pancreatic duct ismore complex as the primary role of this epithelium is in bicarbonate secretion.5 Thebicarbonate concentration in pancreatic duct secretions from CF patients is markedlyreduced.6,7
The exocrine portion of the pancreas is comprised of enzyme-secreting acini andducts lined by epithelial cells that secrete a bicarbonate-rich fluid. The ductal treeextends from centroacinar cells within the acini, through intralobular and interlobu-lar ducts into the main pancreatic duct. Pancreatic pathology is one of the earliestfeatures of CF, and deposition of material within the pancreatic ducts can be ob-served by the mid-trimester of human gestation.8 85% of CF patients are pancreaticinsufficient from birth and all show some pancreatic pathology. Obstruction of pan-creatic ducts in the mid-trimester of development is followed by acinar autolysis asthe acini differentiate and produce pancreatic enzymes by about 24 weeks of gesta-tion. We have been particularly interested in dissecting the molecular basis of theearly pathology of CF pancreatic disease.
aPhone, +44 1865 222341; fax, +44 1865 222626; e-mail, [email protected]

18 ANNALS NEW YORK ACADEMY OF SCIENCES
CFTR EXPRESSION IN THE DEVELOPMENTOF THE PANCREATIC DUCT
The ductal and acinar epithelial cells within the pancreas share a common devel-opmental precursor. In the mid-trimester human pancreas, CFTR mRNA and proteinare present in the ductal epithelium from the 12th week of gestation.9–12 There is aclear gradient of expression of CFTR with increasing levels of mRNA moving dis-tally through the duct system, with lowest levels in large interlobular ducts and high-est levels in the distal intralobular ducts. Throughout this paper we refer to theproximal part of the pancreatic duct system as being closest to the main duct and thedistal part being in the centroacinar cells. The pattern of distribution of CFTR mRNAduring gestation led us to examine whether CFTR might be a useful marker of pan-creatic duct cell development and differentiation. A number of studies with differentanti-CFTR antibodies have shown the expression of CFTR protein in the epitheliumof intralobular ducts in adult human pancreas and in centroacinar cells.1,2,13 Howev-er, most anti-CFTR antibodies have limited usefulness in immunocytochemistrysince they cross-react with other cellular proteins. In addition one study has exam-ined CFTR expression in the adult human gastrointestinal tract by mRNA in situ hy-bridization.4 We have examined CFTR expression by mRNA in situ hybridization inhuman pancreas samples from 13 weeks gestation through to 33 years of age.12
In situ hybridization was carried out as described previously.11 Antisense (HCF-3) and sense (HCF-4) 35S-labeled cRNA probes were generated from an EcoR1/Xba1 fragment (exons 1–5, bases 62–645) of the 10-1 CFTR cDNA clone.14 Thecell-specific expression of CFTR mRNA in the pancreas was examined by in situ lo-calization in mid-trimester fetal tissue at 13, 15, 16.5, 18, 18.5, 19, 23, 24.5, and 32weeks and postnatal tissue at 2, 7, 17, and 26 years. Data from the 18.5-week pan-creas are shown in FIGURE 1, and results on postnatal tissues are shown in FIGURE 2.The first panel in each figure shows a brightfield image of the section, stained withhematoxylin and eosin, and hybridized with a single-stranded, 35S-labeled RNAprobe complementary to the CFTR mRNA (the antisense probe). The second panelshows a darkfield image of the same section. The silver grains, that appear as blackdots on the brightfield image and white dots on the darkfield image, indicate expres-sion of CFTR. In FIGURE 2, panels C and D show a section from the same 2-year-oldpancreas that is shown in panels A and B, hybridized with the negative control senseprobe. The 18.5-week-old fetus in FIGURE 1 illustrates well the pattern of CFTRmRNA expression throughout the mid- trimester of gestation, high levels of CFTRmRNA were detected in the epithelium lining the small inter- and intralobular ducts(indicated by the solid arrows) and in the developing acini (indicated by the squares).Higher magnification (×20 obj.) panels (FIG. 1C,D) illustrate the specific epitheliallocation of CFTR mRNA and tissue morphology in greater detail. Analysis of CFTRmRNA expression in postnatal tissue of all the ages examined (2, 7, 17, and 26 years)show high levels not only in interlobular and intralobular ducts (FIG. 2E,F, indicatedby the solid arrows) and in the centroacinar cells (FIG. 2G,H, indicated by thesquares) but also within the acini themselves (FIG. 2I,J, indicated by the squares).The islets of Langerhans shown by circles in FIGURE 2, panels A,B and I,J, showedno detectable CFTR mRNA expression. We have examined the expression of CFTR

19HARRIS: THE DUCT CELL IN CYSTIC FIBROSIS
in a ∆F508 CF homozygote15 at 32 weeks by in situ hybridization and have shownit to be similar to that of a normal pancreas.
CFTR protein expression was evaluated by immunocytochemistry with the M3A7anti-CFTR (kindly donated by N. Kartner16), and a control for acinar tissue humanpancreatic amylase was detected by monoclonal antibody 099(M) (Biogenex). Im-munocytochemical studies of fetal (14.5-week) and adult pancreas tissue (FIG. 3)show the localization of CFTR to the apical membrane of cells within the ductal ep-ithelium. A gradient of expression of CFTR protein is observed within the ducts withthe highest levels seen in centroacinar cells and small intralobular ducts (FIG. 3A)and decreasing levels through interlobular ducts. The identity of the acinar cells wasconfirmed with a monoclonal antibody to human pancreatic amylase (FIG. 3B). Atthis level of resolution it is impossible to determine whether there is CFTR proteinin other acinar cells, though other groups have now provided data to support this.17,18
FIGURE 1. Expression of CFTR mRNA in 18.5-week fetal pancreas. Sections showexpression of CFTR mRNA as detected by the HCF-3 (antisense) probe. Panels A and Cshow brightfield sections; and panels B and D show darkfield images of the same sections.Specific localization of CFTR mRNA is indicated in the epithelial lining of interlobularducts (solid arrows), intralobular ducts (open arrows, panels A and B) and developing acini(squares). Size bar: panels A,B = 200 µm; panels C,D = 100 µm. (From Hyde et al.12 Re-printed by permission from Gastroenterology.)

20 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 2. Expression of CFTR mRNA in postnatal pancreas. Sections through a 2-year (A–D); 7-year (E,F); 17-year (G,H) and 26-year (I,J) pancreas. Panels A, E, G, and Ishow brightfield sections hybridized with the HCF-3 (antisense) probe; and panels B, F, H,and J show darkfield images of the same sections. Panels C and D (2-year) show a brightfieldview (of a consecutive section to panels A and B) and a darkfield image of the same sectionhybridized with the HCF-4 (sense) negative control probe. Specific localization of CFTRmRNA is indicated in the epithelium lining of interlobular ducts (A,B and E,F, solid ar-rows), intralobular ducts (E,F, open arrows) in centroacinar cells (G,H, squares) and in aci-nar cells (I-J, squares). In pancreatic islets there is no expression of CFTR mRNA (A,B andI,J,circles). Size bar: panels A,B and E–J = 100 µm; panels C,D = 200 µm. (From Hyde etal.12 Reprinted by permission from Gastroenterology.)

21HARRIS: THE DUCT CELL IN CYSTIC FIBROSIS
The functional significance of this evolution of distribution of CFTR expressionis of interest. CFTR is known to be a cAMP-activated chloride ion channel; our datasuggest that the most active sites of secretin-activated chloride secretion in the pan-creas are in the most distal portion of the ducts and within centroacinar cells. Thedistribution of CFTR in the fetal pancreas is important, since it has been suggestedthat as well as being a cAMP-activated chloride ion channel CFTR may have addi-tional roles in epithelial development.11
The evolution in the pattern of expression of CFTR in the pancreatic duct systemthrough human development into adult life may be of relevance to the pathology ofCF. About 15% of CF patients are born pancreas sufficient, this usually being asso-ciated with “mild mutations” in the CFTR gene. It is possible that age-related chang-es in the pattern of expression of CFTR in the pancreas may be important in thedisease progression. As pancreatic sufficiency in CF is often maintained for many
FIGURE 3. Expression of CFTR pro-tein and amylase in adult pancreas. PanelA, CFTR protein detected by the anti-CFTR antibody M3A7; panel B, amylasedetected by the 099M antibody; panel Csecondary antibody alone. In panelA,CFTR is seen in interlobular and intral-obular duct cells and centroacinar cells; inpanel B, amylase is seen in acinar cells.Size bar: all panels = 100 µm. (From Hydeet al.12 Reprinted by permission fromGastroenterology.)
22 ANNALS NEW YORK ACADEMY OF SCIENCES
decades of postnatal life, it is probable that the extent of in utero tissue damage isfundamental to pancreatic function in this disease.
MUCINS IN THE PANCREATIC DUCT IN CF
The precise cause of the pancreatic destruction characteristic of CF is thought tobe obstruction of the pancreatic ducts by inspissated secretions. These secretions
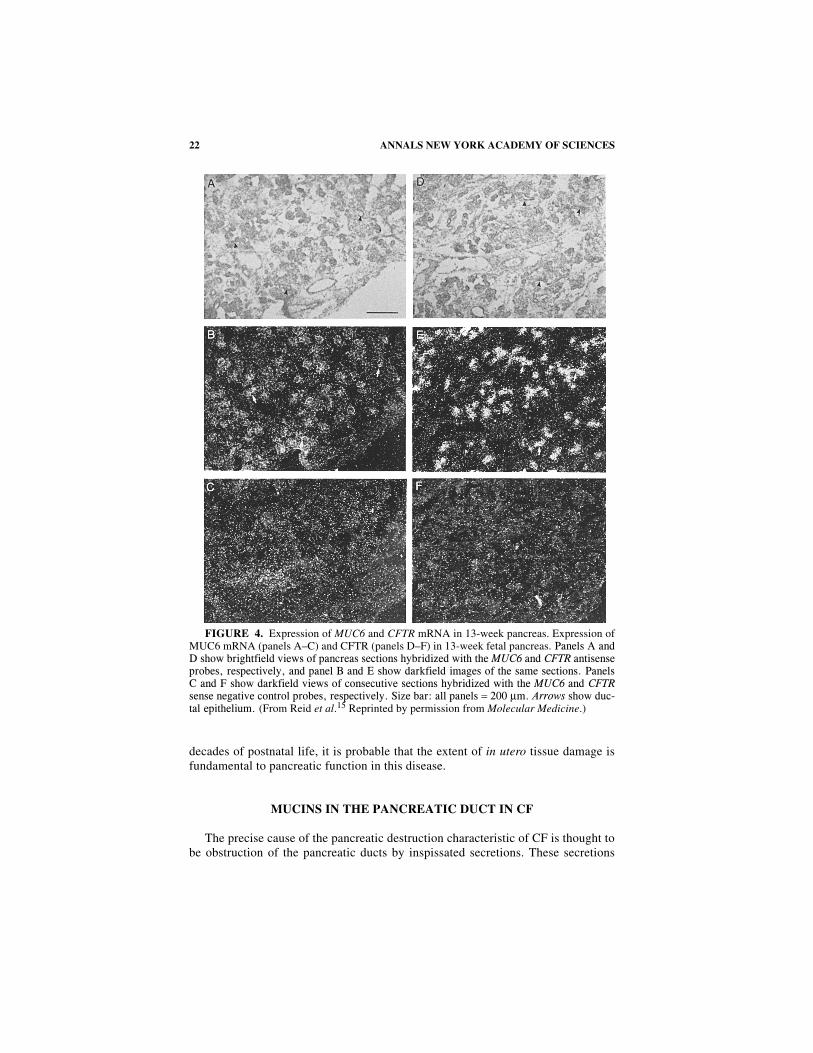
FIGURE 4. Expression of MUC6 and CFTR mRNA in 13-week pancreas. Expression ofMUC6 mRNA (panels A–C) and CFTR (panels D–F) in 13-week fetal pancreas.���������������� ��������������� �������������������������������� ����������������������������������������������������������������������� ������������������������������������������������� ���� ������������ �����������!����������������������� �����������������������������������������������������������������"�������#������������=�$%%�µ���������� �!�&������������!���(From Reid et al.15 Reprinted by permission from Molecular Medicine.)

23
FIG
UR
E5.
Exp
ress
ion
of M
UC
6 an
d C
FT
R m
RN
A in
23-
wee
k pa
ncre
as a
nd in
28
+ 3-
wee
k C
F p
ancr
eas.
Pan
els
A–F
, exp
ress
ion
of M
UC
6m
RN
A (
pane
ls A
–C)
and
CF
TR
(pa
nels
D–F
) in
23-
wee
k fe
tal
panc
reas
. Pan
els
A a
nd D
sho
w b
righ
tfie
ld v
iew
s of
pan
crea
s se
ctio
ns h
ybri
dize
dw
ith
the
MU
C6
and
CF
TR
ant
isen
se p
robe
s, r
espe
ctiv
ely,
and
pan
el B
and
E s
how
dar
kfie
ld i
mag
es o
f th
e sa
me
sect
ions
. P
anel
s C
and
F s
how
dark
fiel
d vi
ews
of c
onse
cuti
ve s
ecti
ons
hybr
idiz
ed w
ith
the
MU
C6
and
CF
TR
sen
se n
egat
ive
cont
rol
prob
es, r
espe
ctiv
ely.
Pan
els
G–L
sho
w e
x-pr
essi
on o
f M
UC
6 m
RN
A (
pane
ls G
–I)
and
CF
TR
(pa
nels
J–L
) in
28
+ 3-
wee
k C
F f
etal
pan
crea
s. P
anel
s G
and
J s
how
a b
righ
tfie
ld v
iew
s of
panc
reas
sec
tion
s hy
brid
ized
wit
h th
e M
UC
6 an
d C
FT
R a
ntis
ense
pro
bes,
res
pect
ivel
y, a
nd p
anel
H a
nd K
sho
w d
arkf
ield
im
ages
of
the
sam
ese
ctio
ns. P
anel
s I
and
L s
how
dar
kfie
ld v
iew
s of
con
secu
tive
sec
tion
s hy
brid
ized
wit
h th
e M
UC
6 an
d C
FT
R s
ense
neg
ativ
e co
ntro
l pro
bes
resp
ec-
tive
ly. S
ize
bar:
all
pan
els
= 20
0 µ m
. Arr
ows
show
duc
tal
epit
heli
um. (
Fro
m R
eid
et a
l .15 R
epri
nted
by
perm
issi
on f
rom
Mol
ecul
ar M
edic
ine .
)

24 ANNALS NEW YORK ACADEMY OF SCIENCES
may include mucous glycoproteins and other proteins; however, the composition ofthe secretions has not been defined. As already mentioned, one of the earliest patho-logical manifestations of CF in utero is the deposition of periodic-acid Schiff-posi-tive material in the fetal pancreatic ducts at about 12 weeks of gestation,8,19
suggesting a contribution of mucins to these deposits. We have examined the devel-opmental expression of 9 human mucin genes in the pancreas by mRNA in situ hy-bridization.20,21 MUC 6 was identified as a major pancreatic mucin that is expressedby 13 weeks of gestation and shows a pattern of distribution that is very similar tothat of CFTR from this age through to adult life.15 MUC1 was the only other mucinmRNA shown to be expressed in the mid-trimester pancreatic duct,22,23 and this wasevident later in gestation in the epithelium of larger pancreatic ducts.20 However, theMUC1 mucin-like core protein is not gel-forming, and it is in part cell surface-asso-ciated via an integral transmembrane domain. MUC1 is not known to be a majorcomponent of the inspissated secretions that are found in CF pancreas, and its tem-poral and spatial patterns of expression are distinct from CFTR. Hence, MUC1 is un-likely to be involved in the initial pathobiochemical events that lead to ductalobstruction in CF. The MUC6 cDNA was isolated by expression cloning from a hu-man stomach cDNA library.24,25 The cDNA has not been fully characterized but ap-pears to be at least 15–16 kb long.26 Similar to other mucins, MUC6 contains aserine and threonine rich tandem repeat sequence with an individual repeat unit of507 bp and 169 amino acids. The probe used for MUC6 in situ hybridization was a74 bp double-stranded oligonucleotide GGTCCACACACACAGCCCCACCAGT-GACGCCGACCACCAGTGGGACGAGCCAAG CCGCGAGCTCATTCAGCACA(bases 308–381 EMBL Accession number L07517) cloned into the Bam HI andHind III sites of pBluescript. Antisense and sense MUC6 35S-labeled riboprobeswere generated from the T7 and T3 promoters, respectively. Abundant MUC6mRNA expression was detected throughout the pancreatic duct epithelia and the de-veloping acini at or by 13 weeks of gestation, the earliest fetal age examined (FIG.4A–C). A similar pattern of expression was seen at 23 weeks (FIG. 5A–C). Two CFpancreases were analyzed. One was from a male ∆F508 homozygote who was bornprematurely at 28 weeks gestation and died 21 days later from respiratory failure, atan age equivalent to 31 weeks gestation (28 + 3 weeks). The second was a 29-weekintrauterine death due to intestinal rupture. This fetus was diagnosed on the basis ofpathology consistent with CF and was heterozygous for ∆F508 and an undefinedmutation. MUC6 mRNA expression patterns in both CF pancreases appeared normal(FIG. 5G–I shows the 28 + 3-week pancreas). In postnatal pancreas (FIG. 6A–D)MUC6 expression was seen in interlobular and intralobular duct epithelial cells, cen-troacinar cells and possibly acinar cells (the in situ technique might not enable dis-crimination between centroacinar cells and acini). There was substantially lessMUC6 mRNA in larger pancreatic duct structures and no detectable expression inislets. A comparison between the MUC6 and CFTR expression patterns in the samepancreatic sections is provided by the data presented in FIGURE 4D–F and FIGURE
5D–F, which show CFTR expression using the same probes described above at therelevant developmental stages. CFTR and MUC6 transcripts have nearly identicalexpression patterns in the 13- and 23-week fetal pancreas. A similar pattern of co-localization of MUC6 (panels A–D) and CFTR mRNA (panels E–H) is seen in post-natal pancreas (FIG. 6).

25HARRIS: THE DUCT CELL IN CYSTIC FIBROSIS
The localization of MUC6 expression by mRNA in situ hybridization was con-firmed by immunocytochemistry. The M6P antibody, a chicken polyclonal antibody
FIGURE 6. Expression of MUC6 and CFTR mRNA in 2-year-old pancreas. Expressionof MUC6 mRNA (panels A–D) and CFTR (panels E–H) in 2-year-old pancreas. Panels Aand D show brightfield views of pancreas sections hybridized with the MUC6 antisenseprobe; panels E and F show brightfield views of pancreas sections hybridized with the CFTRantisense probe. Panels B and G show darkfield images of the same sections shown in panelsA and F. Panels C and H show darkfield views of consecutive sections hybridized with theMUC6 and CFTR sense negative control probes, respectively. Size bar: panels A–C and F–H = 200 µm; panels D,E = 100 µm. Arrows show ductal epithelium. (From Reid et al.15 Re-printed by permission from Molecular Medicine.)

26 ANNALS NEW YORK ACADEMY OF SCIENCES
that was raised against a MUC6 tandem repeat peptide (kindly donated by Sam Ho),was used to localize the MUC6 glycoprotein in normal and CF pancreas (FIG. 7).Panels A–D show hematoxylin and eosin-stained sections of normal 32-week-gesta-tion fetal pancreas (panels A,B) and the 28 + 3 week CF pancreas (panels C,D). Anabnormal histology is apparent in the CF pancreas with material obstructing dilatedducts (see arrow in panel D). MUC6 protein expression is seen in 23-week pancreasin developing pancreatic ducts and acini (panels E–G: E, preimmune serum; F andG, M6P). The arrow in panel G identifies MUC6 expression in developing acini/cen-troacinar cells. MUC6 protein expression in a 2-year-old normal pancreas is illus-trated in panels K–M (K, preimmune serum; L and M, M6P). The arrow in panel Mshows acinar expression of MUC6, which is also seen in the epithelial cells lining
FIGURE 7. Expression of MUC6 protein in 32-week and 2-year-old normal pancreasand 28 + 3-week CF pancreas. Immunoperoxidase detection of MUC6 protein in the pancreas.Panels A-D, hematoxylin and eosin-stained pancreatic tissue, A and B, normal 32-week pan-creas; C and D, 28 + 3-week CF fetal pancreas. Panels E–G show MUC6 protein expression,detected with the M6P antibody, in 23 week pancreas, E, preimmune serum, F and G, M6P.Panels H–J show MUC6 protein expression in 28 + 3-week CF fetal pancreas. Panels K–Mshow MUC6 protein expression in 2-year normal pancreas, K, preimmune serum, L and M,M6P. Size bars: panels A,C,E,F,I,H,K and L = 200 µm; panels B,D,G,I,J and M = 100 µm.(From Reid et al.15 Reprinted by permission from Molecular Medicine.)

27HARRIS: THE DUCT CELL IN CYSTIC FIBROSIS
small ducts and in centroacinar cells. Panels H–J show MUC6 protein expression in28 + 3-week CF fetal pancreas detected by the M6P antibody. The arrows in panelsI and J show MUC6 to be a significant component of the material obstructing thesmall intralobular ducts in this CF pancreas. Similar results were obtained on a 29-week CF fetal pancreas, in which the histology was less abnormal, but there was alsoevidence of MUC6 in inspissated secretions within the intralobular ducts (notshown).
It is of particular interest in the context of CF that the localization of expressionof the CFTR gene coincides with that of MUC6 in the pancreas. The temporal andspatial distribution of MUC6 and CFTR transcripts shows complete concordance inthe pancreas (FIGS. 4–6). These results suggest that MUC6 and CFTR are expressedin the same cell types within the pancreas. These observations may be of consider-able importance to our understanding of the disease process in the CF pancreas. Asalready stated, the CFTR gene encodes a small conductance cAMP-activated�chlo-ride ion channel$' that is expressed at high levels in the pancreatic duct epitheliumin utero and postnatally.$( A principal physiological defect in the CF pancreas is thefailure to secrete bicarbonate ions that are essential for the normal flow of pancreaticduct secretions. One model that would explain the pathophysiology of CF in the pan-creas, for which there is some physiological evidence, proposes that chloride ion ef-flux through CFTR is coupled to bicarbonate ion secretion via a chloride/bicarbonate exchanger in the apical membrane of the pancreatic duct cell.) The CFpancreatic duct fluid has reduced water and bicarbonate content' and this is believedto contribute to the deposition of material in the small intralobular ducts. The com-position of these deposits has not been characterized, though it has been suggestedthat they may contain secreted proteins and/or mucus glycoproteins. We have nowidentified MUC6 as a significant constituent of these ductal deposits. Future exper-iments will determine the relationship between altered CFTR functioning and thesolubility of MUC6.
HOW DO MUTATIONS IN CFTR LEAD TO OBSTRUCTIONOF PANCREATIC DUCTS IN CF?
The cause of the mucus clearance problems in CF remains poorly understood.Several hypotheses to explain the mucus abnormalities have been put forward, in-cluding: mucin hypersecretion; dehydration of mucins due to ion transport defectsthat result from mutations in the CFTR cAMP-activated chloride ion channel; andbiochemical abnormalities in the glycosylation of mucins in CF epithelial cells in-cluding increased sulphation and fucosylation, and reduced sialylation. However,there is currently no conclusive evidence to provide support for any one of thesehypotheses.
Since the biochemical and biophysical properties of a mucin are dependent on O-glycosylation, this aspect of mucin biology has been investigated in CF. Reports ofincreased glycosylation and sulphation of mucins from CF airway tissue29–31 andintestine32 are complicated by the potential for secondary modifications in mucinssecreted by diseased epithelia. Increased sulphation of glycoconjugates secreted byCF nasal epithelial cells in culture33 or CF bronchial epithelial cells in a xenograft

28 ANNALS NEW YORK ACADEMY OF SCIENCES
model34 have also been observed. The cause of mucin abnormalities in the pancreashas not been studied extensively.
All the experiments in which CF mucins have been analyzed to date, regardlessof the experimental model, have analyzed a mixture of glycoproteins secreted fromepithelial cells, rather than evaluated the biochemistry of individual mucins. Our ob-servation that MUC6 is a significant component of the material that obstructs thepancreatic ducts in CF provides a handle on investigating the molecular basis of mu-cin abnormalities in the CF pancreas. Our aim is to evaluate the O-glycosylation ofMUC6 alone in matched pairs of pancreatic duct cells that differ with respect toCFTR expression status, for example, one cell line carrying mutations in both CFTRgenes, and a derivative line in which normal CFTR expression has been restored bythe introduction of a wild-type transgene. Examples of these cell lines exist, for ex-ample, the CFPAC line and its corrected pair that carries a retroviral vector express-ing wild-type CFTR.35 We will exploit an epitope-tagged MUC1 mucin cDNA36
that, when stably expressed in cell lines, can be purified with an antibody specificfor the epitope. This approach detected variation in mucin glycosylation (expressionof blood group antigens) in different cell lines36 including pancreatic and colon car-cinoma cell lines. We have extended these studies to examine the expression of bloodgroup antigens on MUC1F mucin in matched pairs of Caco2 cell lines that either ex-press wild-type CFTR or have spontaneously switched off CFTR expression and somay be considered equivalent to CFTR null mutants.37 Further, metabolic labelingexperiments were then carried out to estimate the gross levels of glycosylation andsulphation of MUC1F mucin in these matched pairs of cell lines. Expression ofCFTR in this experimental system did not affect the gross levels of glycosylation orsulphation of the MUC1F mucin or the antigenicity of the carbohydrates structuresattached to the MUC1F protein. Similar experiments with MUC6 need to be per-formed in pancreatic duct cells. Given the size of the MUC6 cDNA it will be difficultto generate a full length epitope-tagged MUC6F cDNA construct. Hence we are us-ing the MUC1F backbone in a tandem repeat-deleted form (Burdick et al., submit-ted) as a vehicle for the tandem repeats of other mucins. The tandem repeats ofmucin are rich in serine and threonine and they provide most of the sites for the O-glycosylation that is characteristic of these glycoproteins. We have generated a hy-brid mucin carrying tandem repeat sequences from MUC5AC in a tandem repeat-de-leted construct of the epitope-tagged MUC1 (MUC1/5ACF) and shown it to carrydifferent carbohydrate structures from the native MUC1F construct when transfectedin to the same cell line (unpublished data). This suggests that a hybrid MUC1/6TRFmucin will provide a powerful tool for investigating the glycosylation of the MUC6mucin tandem repeats in differentiated pancreatic duct cells. These experimentsshould establish whether a primary defect in mucin processing is responsible for theobstruction of CF pancreatic ducts or whether the cause is dehydration of the epithe-lium, mucin hypersecretion or some other mechanism.
ACKNOWLEDGMENTS
This work was supported by Grant DK46589 from the National Institutes ofHealth and by the Cystic Fibrosis Research Trust UK. Karen Hyde, Colm Reid, ScottTebbutt and Deborah Harrison contributed data described in this paper. I am grateful

29HARRIS: THE DUCT CELL IN CYSTIC FIBROSIS
to N. Kartner for the M3A7 antibody, and to S. Ho for the M6P antibody. We thankW. B. Saunders Co. for permission to reproduce FIGURES 1–3 and Molecular Medi-cine, a joint publication of the Picower Institute Press and Springer-Verlag, NY, Inc.for permission to reproduce FIGURES 4–7.
REFERENCES
1. CRAWFORD, I.C., P.C. MALONEY, P.L. ZEITLIN, W.B. GUGGINO, S.C. HYDE, H. TUR-LEY, K.C. GATTER, A. HARRIS & C.F. HIGGINS. 1991. Immuno-cytochemical local-ization of the cystic fibrosis gene product CFTR. Proc. Natl. Acad. Sci. USA 88:9262–9266.
2. DENNING, G.M., L.S. OSTEDGAARD, S.H. CHENG, A.E. SMITH & M.J. WELSH. 1992.Localization of the cystic fibrosis transmembrane conductance regulator in chloridesecretory epithelia. J. Clin. Invest. 89: 339–349.
3. ENGELHARDT, J.F., J.R. YANKASKAS, S.A. ERNST, Y. YANG, C.R. MARINO, R.C.BOUCHER, J.A. COHN & J.M. WILSON. 1993. Submucosal glands are the predominantsite of CFTR expression in the human bronchus. Nature Genet. 2: 240–247.
4. STRONG, T.V., K. BOEHM & F.S. COLLINS. 1994. Localization of cystic fibrosis trans-membrane conductance regulator mRNA in the human gastrointestinal tract by insitu hybridization. J. Clin. Invest. 93: 347–354.
5. GRAY, M.A., C.E. POLLARD, A. HARRIS, L. COLEMAN, J.R. GREENWELL & B.E.ARGENT. 1990. Anion selectivity and block of the small-conductance chloride chan-nel on pancreatic duct cells. Am. J. Physiol. 259: C752–761.
6. GASKIN, K.J., P.R. DURIE, M. COREY, P. WEI & G.G. FORSTNER. 1982. Evidence for aprimary defect of pancreatic HCO3
− secretion in cystic fibrosis. Pediatr. Res. 16:554–557.
7. KOPELMAN, H., P.R. DURIE, K. GASKIN, Z. WEISMAN & G. FORSTNER. 1985. Pancre-atic fluid secretion and protein hyperconcentration in cystic fibrosis. N. Engl. J. Med.312: 329–334.
8. HARRIS, A.& L. COLEMAN. 1987. Establishment of a tissue culture system for epithe-lial cells derived from human pancreas: a model for the study of cystic fibrosis. J.Cell Sci. 87: 695–703.
9. FOULKES, A.G. & A. HARRIS. 1993. Localization of expression of the cystic fibrosisgene in human pancreatic development. Pancreas 8: 3–6.
10. HARRIS, A., G. CHALKLEY, S. GOODMAN & L. COLEMAN. 1991. Expression of the cys-tic fibrosis gene in human development. Development 113: 305–310.
11. TREZISE, A.E.O., J.A. CHAMBERS, C.J. WARDLE, S. GOULD & A. HARRIS. 1993.Expression of the cystic fibrosis gene in human fetal tissues. Hum. Mol. Genet. 2:213–218.
12. HYDE, K., C.J. REID, S.J. TEBBUTT, L. WEIDE, M.A. HOLLINGSWORTH & A. HARRIS.*++'�� ,��� ������� ��������� �������������� ���!������� ���!������ ��� ��������� ���!���� ����������� !��� ����������� ��� ���������������� -���������������� 113:+*.–+*+�
13. MARINO, C.R., L.M. MATOVCIK, F.S. GORELICK & J.A. COHN. 1991. Localization ofthe cystic fibrosis transmembrane conductance regulator in pancreas. J. Clin.Invest. 88: 712–716.
14. RIORDAN, J.R., J.M. ROMMENS, B.-S. KEREM, N. ALON, R. ROZMAHEL, G. GRZELC-ZAK, S. LOK, N. PLAVSIC, J.-L. CHOU, M.L. DRUMM, M.C. IANNUZZI, F.S. COLLINS
& L.-C. TSUI. 1989. Identification of the cystic fibrosis gene: cloning and charac-terisation of complementary DNA. Science 245: 1066–1073.
15. REID, C.J., K. HYDE, S.B. HO & A, HARRIS��*++'��������������������� ������������#������������ ���/0�1��!���� ��� �����!������ ��� ����������� !�����/����/��� 3:.%2–.**�
16. KARTNER, N., O. AUGUSTINAS, T.J. JENSEN, A.L. NAISMITH & J.R. RIORDAN. 1992.Mislocalization of ∆F508 CFTR in cystic fibrosis sweat gland. Nature Genet. 1:321–327.

30 ANNALS NEW YORK ACADEMY OF SCIENCES
17. KOPELMAN, H., E. FERRETTI, C. GAUTHIER & P.R. GOODYER. 1995. Rabbit pancreaticacini express CFTR as a cAMP-activated chloride efflux pathway. Am. J. Physiol.269: C626–C631.
18. ZENG, W., M.G. LEE, M. YAN, J. DIAZ, I. BENJAMIN, C.R. MARINO, R. KOPITO, S.FREEDMAN, C. COTTON, S. MUALLEM & P. THOMAS. 1997. Immuno and functionalcharacterization of CFTR in submandibular and pancreatic acinar and duct cells.Am. J. Physiol. 273: C442-C455.
19. BOUE, A., F. MULLER, C. NEZELOF et al. 1986. Prenatal diagnosis in 200 pregnancieswith a 1-in-4 risk of cystic fibrosis. Hum. Genet. 74: 288–297.
20. CHAMBERS, J.A., M.A. HOLLINGSWORTH, A. TREZISE & A. HARRIS. 1994. Develop-mental expression of mucin genes MUC1 and MUC2. J. Cell Sci. 107: 413–424.
21. REID, C.J. & A. HARRIS.� *++(�� ������������� �3��������� ��� �!���� ������ ��� ����!���������������������-!��42: $$%–$$1�
22. LAN, M.S., S.K. BATRA, W.-N. QI, R.S. METZGAR & M.A. HOLLINGSWORTH. 1990.Cloning and sequencing of a human pancreatic tumor cDNA. J. Biol. Chem. 265:15294–15299.
23. BATRA, S., R.S. METZGAR & M.A. HOLLINGSWORTH. 1992. Human MUC1 mucingene expression in fetal pancreas. Pancreas 7: 391–393.
24. HO, S., A.M. ROBERTSON, L.L. SHEKELS, C.T. LYFTOGT, G.A. NIEHANS & N.W. TOR-IBARA. 1995. Expression cloning of gastric mucin complementary cDNA and local-ization of mucin gene expression. Gastroenterology 109: 735–747.
25. TORIBARA, N.W., A.M. ROBERTSON, S. HO et al. 1993. Human gastric mucin. J. Biol.Chem. 268: 5879–5885.
26. TORIBARA, N.W., S.B. HO, E. GUM, J.R. GUM, P. LAU & Y.S. KIM. 1997. The car-boxyl-terminal sequence of the human secretory mucin, MUC6. Analysis of theprimary amino acid sequence. J. Biol. Chem. 272: 16398-16403
27. ANDERSON, M.P., D.P. RICH, R.J. GREGORY, A.E. SMITH & M.J. WELSH. 1991. Gen-eration of cAMP-activated chloride currents by expression of CFTR. Science 251:679–682.
28. GRAY, M.A., C.E. POLLARD, A. HARRIS, L. COLEMAN, J.R. GREENWELL & B.E.ARGENT. *++%�� ������ ������������ ��� ������ ��� ���� �����&���!������� �����������������������������!��������������4�����������259:��')$–'1*�
29. BOAT, T. F., P.W. CHENG, R.N. LYER, D.M. CARLSON & I. POLONY. 1976. Humanrespiratory tract secretions: mucous glycoproteins of nonpurulent tracheobronchialsecretions, and sputum of patients with bronchitis and cystic fibrosis. Arch. Bio-chem. Biophys. 177: 95–104.
30. FRATES, R.C., T.T. KAIZU & J.A. LAST. 1983. Mucus glycoproteins secreted by res-piratory epithelial tissue from cystic fibrosis patients. Paediatr. Res. 17: 30–34.
31. LO-GUIDICE, J., J. WIERUSZESKI, J. LEMOINE, A. VERBERT, P. ROUSSEL & G. LAMBLIN.1994. Sialylation and sulfation of the carbohydrate chains in respiratory mucinsfrom a patient with cystic fibrosis. J. Biol. Chem. 269: 18794–18813.
32. WESLEY, A., J. FORSTNER, R. QURESHI, M. MANTLE & G. FORSTNER. 1983. Humanintestinal mucin in cystic fibrosis. Paediatr. Res. 17: 65–69.
33. CHENG, P.W., T.F. BOAT, K. CRANFILL, J.R. YANKASKAS & R.C. BOUCHER. 1989.Increased sulphation of glyconjugates by cultured nasal epithelial cells frompatients with cystic fibrosis. J. Clin. Invest. 84: 68–72.
34. ZHANG, Y., B. DORANZ, J.R. YANKASKAS & J.F. ENGELHARDT. 1995. Genotypic anal-ysis of respiratory mucus sulfation defects in cystic fibrosis. J. Clin. Invest. 96:2997–3004.
35. CLIFF, W.H., R.A. SCHOUMACHER & R.A. FRIZZELL. 1992. cAMP-activated Cl chan-nels in CFTR-transfected cystic fibrosis pancreatic epithelial cells. Am. J. Physiol.262: C1154 -C1160.
36. BURDICK, M.D., A. HARRIS, C.J. REID, T. IWAMURA & M.A. HOLLINGSWORTH��*++'�5��������������� �3������� ���/0�*� ���!��� ��� ����������� ��� ������ �!���������������4��������������272:$.*+(–$.$%$��
37. REID, C.J., M.D. BURDICK, M.A. HOLLINGSWORTH & A. HARRIS. 1998. CFTR expres-sion does not influence glycosylation of an epitope-tagged MUC1 mucin in coloncarcinoma cell lines. Glycobiology. In press.