
Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
62
UNIVERSITATEA DIN BUCURESTI FACULTATEA DE BIOLOGIE LUCRARE DE LICENTA ,, TEHNICI DE BIOLOGIE MOLECULARA IN ANALIZA SI EPIDEMIOLOGIA ENTEROVIRUSURILOR’’ COORDONATOR: INDRUMATOR: Conferenţiar Universitar Cercetător principal III Dr. Marina Nechifor Gabriela Oprişan ABSOLVENT : Alexandru Ana-Maria Secţia : Biochimie
-
Upload
matranrazvan -
Category
Documents
-
view
228 -
download
2
Transcript of Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 1/62
UNIVERSITATEA DIN BUCURESTI
FACULTATEA DE BIOLOGIE
LUCRARE DE LICENTA
,, TEHNICI DE BIOLOGIE MOLECULARA IN
ANALIZA SI EPIDEMIOLOGIAENTEROVIRUSURILOR’’
COORDONATOR: INDRUMATOR:
Conferenţiar Universitar Cercetător principal III
Dr. Marina Nechifor Gabriela Oprişan
ABSOLVENT :
Alexandru Ana-Maria
Secţia : Biochimie

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 2/62
Cuprins
Introducere……………………………………………………………………. pag. 1
Capitolul I : Enterovirusuri
1. Încadrare taxonomica……………………………………………………..…pag.5
2. Genomul viral………………………………………………………………pag.7
2.1.Regiunea 5’ necodata……………………………………………………..pag.8
2.2.Regiunea 3’ necodata……………………………………………………..pag.8
2.3.Regiunea codata a poliproteinei virale……………………………………pag.10
3. Proteine virale………………………………………………………………pag.11
3.1.Proteine structurale……………………………………………………….pag.11
3.2.Proteine nestructurale…………………………………….………………pag.12
4. Virionul.........................................................................................................pag.16
4.1.Arhitectura virionului…………………………………………………….pag.164.2.Proprietati fizico-chimice…………………………………………………pag.17
4.3.Determinanti antigenici……………………………………………………pag.18
5. Receptori celulari…………………………………………………………….pag.20
6.Multiplicarea virala…………………………………………………………..pag.23
6.1.Etapele precoce…………………………………………………………….pag.24
6.2.Sinteza virala………………………………………………………………pag.25
6.3.Etapele tardive……………………………………………………………..pag.28
7.Epidemiologia…………………………………………………………………pag.29
7.1.Moduri de transmitere………………………………………………………pag.29
7.2.Epidemiologia moleculara…………………………………………………pag.29
2

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 3/62
8. Patogeneza…………………………………………………………………….pag.30
Capitolul II :Tehnici de biologie moleculara
Reactia de polimerizare n lant(PCR)ȋ ………………………………………….pag.33
1. Generalitati………………………………………………………………...pag33
2. Componentele reactiei……………………………………………………..pag.34
2.1. Matrita ADN………………………………………………………….pag.34
2.2. Primeri oligonucleotidici……………………………………………..pag.35
2.3. ADN polimerazele termostabile……………………………………...pag.35
2.4. Deoxinucleotid trifosfaţi ( dNTP)…………………………………....pag.37
2.5. Tamponul de reacţie …………………………………………………pag.38
2.6. Uleiul mineral………………………………………………………..pag.38
3.Detectarea produsilor PCR …………………………………………… …..pag.38
3.1.Contaminarea si evitarea contaminarii in reactia PCR………………….pag.39
3.2. Probleme ce apar in reactiile PCR…………………………………… .pag. 40
3.3.Specificitatea reactiei……………………………………………………pag.41
3.4. Variante de PCR………………………………………………………..pag.413.5.Aplicatii ale tehnologiei PCR…………………………………………...pag.43
4.Restrictia enzimatica………………………………………………………pag. 44
PARTEA EXPERIMENTALA
1.PCR pentru enterovirusuri non-polio……………………………………….pag. 48
2.Extractie ARN viral…………………………………………………………..pag.49
3.Reverstranscriere pentru originea genomica VP1-2A………………………pag.51
4.Reactia PCR pentru regiunea genomica VP1-2A……………………………pag.52
5.Electroforeza in gel de agaroza……………………………………………….pag.54
6.Incarcarea ampliconilor in gel si migrarea…………………………………...pag.56
7.Concluzii...............................................................................................................pag.58
3

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 4/62
8.Bibliografie…………………………………………………………………… pag.59
INTRODUCERE
Enterovirusurile reprezintă un gen foarte larg aparţinând familiei Picornaviridae.
Virusurile cu ARN de polaritate pozitivă din această familie se remarcă prin faptul că
sunt cele mai mici ribovirusuri cunoscute (picos = mic) si reprezintă patogeni importanţi
pentru om şi zootehnie. Datorită importanţei economice şi medicale a picornavirusurilor
studiul lor a adus o contribuţie substanţială la dezvoltarea virusologiei moderne.
Apartenenţa la grupul virusurilor ARN de polaritate pozitivă este justificată de
faptul că genomul enterovirusurilor funcţionează ca ARN mesager şi prezintă o ARN
replicază dependentă de ARN.
Numelele generic de enterovirusuri provine de la faptul că majoritatea colonizează
tractul digestiv. Pe langă infecţii uşoare sau asimptomatice, cele 66 serotipuri deenterovirusuri umane cunoscute pana in prezent pot provoca şi infecţii foarte grave cum
sunt meningitele aseptice sau miocardita acuta.
Clasificarea iniţiala a enterovirusurilor s-a bazat pe diferenţele antigenice, pe
capacitatea de a se replica intr-o anumita linie celulară umană sau animală.
4

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 5/62
CAPITOLUL I
ENTEROVIRUSURI
1. ÎNCADRARE TAXONOMICA
ETIOLOGIE
Enterovirusurile (E.V.) fac parte din familia PICORNAVIRIDAE (pico = foarte
mic, RNA =tipul de acid nucleic), con in ARN, sunt icosaedrale, de aproximativ 30 nm
diametru i nu au anvelop . Clasificarea enterovirusurilor în mai multe subgrupe seş ă
bazeaz pe diferen e de patogenitate i gazde, iar divizarea în serotipuri se face prină ţ ş
seroneutralizare:
Virusurile Poliomielitice - (V.P.)
Serotipurile 1-3 (tipul 1- Brunhilde; tipul 2 - Lansing; tipul 3 - Leon) cultiv înă
culturi celulare de primate.
Tipul 2 Lansing a fost adaptat la roz toare. Provoac leziuni histopatologiceă ă
caracteristice prin inoculare direct în SNC la primate.ă
Virusurile Coxsackie sunt divizate în 2 grupuri:
Coxsackie grup A - produc la oricelul inoculat miozita generalizat , cu paralizieş ă
flasc i cuprind serotipurile 1-24 (în prezent 23 serotipuri);ă ş
5

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 6/62
Coxsackie grup B - produc miozita focal , cu infec ie generalizat în miocard,ă ţ ă
pancreas, SNC cu paralizie spastic i cuprind serotipurile 1-6;ă ş
Virusurile ECHO
Acestea produc efecte citopatice în culturi celulare de primate i cuprind 31ş
serotipuri recunoscute. Ini ial au fost 34 serotipuri, însa în urma studiilor virusologice s-ţ
au restrâns la 31. Enterovirusuri nou descoperite au fost numerotate 68-72, cu
excluderea virusului hepatitic A (enterovirus 72).
6

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 7/62
2. GENOMUL VIRAL
Genomul enterovirusurilor este constituit dintr-o moleculă de ARN monocatenar, de
polaritate pozitivă, având aproximativ 7400 de nucleotide. ARN-ul viral, infectat,
funcţionază ca ARN mesager în timpul translaţiei şi ca matriţă pentru formarea catenei
complementare de polaritate negativă în timpul replicării genomului viral. Extremitatea
3’ a genomului este poliadenilată iar extremitatea 5’ este legată covalent cu o proteină
de talie mică ( 22 de aminoacizi ), VPg, cu un rol important se pare în iniţierea sintezei
ARN-ului. Molecula de ARN prezintă o singură fază de citire deschisă, ORF
(,, open reading frame’’), încadrată de două regiuni necodante. (NC) în 5’ şi 3’
(Rueckert, 1996)
7

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 8/62
1.1. Regiunea 5’ necodantă ( 5’ NC)
În comparaţie cu ARN mesageri celulari, regiunea 5’NC a enterovirusurilor este
neobişnuit de lungă. Este constituită din aproximativ 750 de nucleotide ( ~ 10% din
lugimea totală a genomului) şi are un grad înalt de conservare în cadrul
enterovirusurilor. Aceste caracteristici se datoresc functiilor importante îndeplinite deregiunea 5’NC, funcţii care depind atât de structura sa secundară, cât şi de cea primară.
Structura sa secundară complexă, compusă din duble helixuri şi bucle, conţine două
domenii funcţionale diferite implicate în sinteza ARN viral şi în iniţierea translaţiei
proteinelor virale. Primele 100 de nucleotide au o structura caracterisică în formă de
8

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 9/62
treflă, implicată în replicarea catenei de polaritate pozitivă dar care este indispensabila
sintezei catenei de polaritate negativă. În plus, această structură sub forma de frunza de
trifoi ar putea conţine semnale comune atât pentru translaţie cât şi pentru replicarea
virală şi pare să intervină în supresia translaţiei eucariote în favoarea iniţierii sintezei de
ARN viral prin intermediul interacţiilor ARN-proteine. Urmeaza al doilea domeniu
funcţional, care est necesar în translaţia virală şi care conţine o strucutură numită IRES
(,, Internal Ribosome Entry Site’’, sediul intern de acces la ribosom), implicată în
interacţia cu proteine celulare. Mecanismul propus pentru enterovirusuri, în care IRES
joacă un rol important în iniţierea internă a sintezei poliproteinei virale (lipsind o
structură de tip ,,bonetă’’ - ,,cap’’ – în extremitatea 5’ a genomului ), este diferit de cel
de scanare a ribozomilor, specific pentru ARN mesageri celulari. Recent, a fost
demostrat faptul ca IRES este bivalent pentru ca ar conţine şi secvenţe cu acţiune în cis,
necesare pentru replicarea genomului viral.
9

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 10/62
1.2. Regiunea 3’ necodantă ( 3’NC)
Regiunea 3’NC este de mărime variabilă în funcţie de tipul de enterovirus, de
exemplu de 72 de nt la polivirus sau de 100 nt la coxackievirus B. Prezintă două sau
trei domenii distincte (al treila nu est prezent decât la două din cele cinci grupuri
filogenetice ale enterovirusurilor) , desemnate ,,X’’, ,,Y’’, ,,Z’’, cu structuri în ac de
păr. Această regiune pare să aibă un rol primordial în iniţierea sintezei ARN de
polaritate negativă în timpul replicării genomului viral, graţie structurii sale primare
dar şi a celei terţiare, sub formă de pseudo-nod. Proteinele virale şi celulare recunosc
specific complexul ribonucleoproteic format la originea replicării de către 3’NC în
procesul de iniţiere a sintezei catenei de polaritate negativă.
Regiunea 3’NC este urmată de o secvenţă poliadenilată codificată genetic, de
aproximativ 75 nt, a cărei funcţie nu este încă foarte bine definită. Aceasta secventa
pare să fie implicată în infectivitatea ARN şi poate interveni şi în procesul de
replicare ( Sarnow, 1989 ).
1.3. Regiunea codantă a poliproteinei virale
Cadrul de lectură deschis (ORF) codifică o poliproteină de aproximativ 250 kDa,care este clivată în etape succesive în timpul translaţiei de către proteaze virale. Se obţin
astfel 11 proteine virale (Fig. 2). Poliproteina nu a fost niciodată detectată în celulele
infectate deoarece est clivată în timpul sintezei sale. Este împărţită în 3 regiuni: P1, P2,
P3, corespunzând celor trei polipeptide precursoare ale proteinelor virale.
Regiunea P1 codifică poliproteina P1, precursoare a proteinelor structurale
VP1,VP2, VP3 şi VP4, care constituie capsida virală.
Regiunea P2 şi P3 codifică poliproteinele P2 şi respectiv P3, ca precursoare ale
proteinelor nestructurale, implicate în clivarea proteolitică, în replicarea genomului viral
precum şi în înhibarea funcţiilor celulare. Poliproteina P2 este precursoarea proteinelor
10

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 11/62
mature 2A, 2B şi 2C. Poliproteina P3 este precursoare pentru proteinele mature 3A, 3B,
3C şi 3D.
Procesarea poliproteinelor virale (dupa Allan )
3. PROTEINE VIRALE
11

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 12/62
Regiunile P1, P2, P3 ale proteinelor virale
3.1. Proteine structurale
Regiunea P1, prima din ORF, codifică polipeptidul P1 care eate precursor al celor
patru proteine mature ale capsidei VP4,VP2, VP3, VP1 (ordinea din P1). Imediat ce P1
este separat de restul poliproteinei de către proteinaza virală 2A, ea este la rândul său
clivată graţie activităţii proteazice a proteinei 3CD. Se obţin astfel polipeptidele VP0,
VP3 şi VP1.
Si proteina VP0 este clivată ulterior, în timpul asamblării finale a virionului,
pentru a produce proteinele structurale VP4 şi VP2. Proteina VP0 rămâne totusi
prezentă, în număr redus (două molecule per virion), în capsida virionului. O descrire
detaliată a acestor proteine este prezentată în paragraful D.1 din capitolul ,,Arhitectura
virionului’’.
12

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 13/62
Proteine mature de capsida ( VP1,VP2,VP3, VP4)
3.2. Proteine nestructurale
Regiunea P2 codifică polipeptida P2 care est precursoare polipeptidelor mature 2A,
2B Şi 2C. Produsul precursor 2BC este stabil şi ar putea fi implicat în sinteza ARN
viral.
Proteina 2A este o protează care se eliberează intr-un proces autocatalitic de
poliproteina virala. Un prim clivaj se realizează in în cis la nivelul Tyr/Gly, pentru a
elibera extremitatea N-terminală a acestei enzime, separând astfel precursorul
proteinelor de capsidă ( P1) de precursorii proteinelor nestructurale ( P2-P3). Proteina
2A este responsabilă şi de un clivaj alternativ, neesenţial, al precursorului 3CD pentru a
13

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 14/62
genera produşii maturi 3C’ şi 3D’. Riolul acestui clivaj alternativ nu este încă cunoscut.
Proteina 2A ar putea fi un element esenţial în replicarea ARN deoarece nu a fost
observată sinteza virală în celulele transfectate cu ARN dicistronice, având această
proteină delatata. Acţiune proteazei 2A asupra proteinelor celulare are gazdei în timpul
infecţiei enterovirale a fost intens studiată. Proteinele 2Apro ale poliovirusurilor şi
coxackievirusurilor sunt responsabile de clivarea factorului de iniţiere a translaţiei
eucariote, eIF4G, prin care se blochează translatia ARN mesageri ai celulei gazdă
(,,shut-off’’). Structura ARN mesageri din celula eucariotă, numită de autorii englezi
,,cap’’., indispensabilă pentru iniţierea translaţiei şi aşezarea corectă a ARN pe
ribozomi, lipseşte la enterovirusuri. Înhibarea iniţierii sintezei proteiceeucariote ,,cap’’-
dependente de către proteaza virală conduce la stimularea translaţiei ARN enteroviral,
lipsit de ,,bonetă’’, dirijată de către IRES. O altă ţintă celulară a proteazei 2A este
,, poly( A)- binding protein’’ ( PABP). Clivarea acestei proteine, din familia proteinelor
eucariote care se leaga la ARN, ar putea constitui un alt mecanism prin care
enterovirusurile induc procesul de ,,shut-off’’ celular (blocarea sintezei proteinelor
celulare). Recent a fost demonstrat faptul că 2Apro a coxackievirusurilor clivează in
vitro distrofina umană şi de şoarece. Distrofina poate fi clivată şi in vivo încardiomiocitele şoarecilor în timpul infecţiei cu coxackievirus B3. Clivarea acestei
proteine de citoschelet, a cărei deficienţă de origine genetică determină o cardiomiopatie
dilatantă ereditară, ar putea fi la originea mecanismului molecular prin care infecţia
enterovirală contribuie la patogeneza cardiomiopatiei dilatante dobândite. De puţin timp
2Apro este suspectată de a fi implicată şi în procesul de moarte celulară prin apoptoză.
Proteina 2BC pare să inducă o proliferare membranară, la nivelul membranelor
citoplasmatice, ceea ce determină o acumulare de vezicule induse de virus pe care se
ancorează complexele de replicare. De asemenea, proteina 2BC ar bloca şi transportul
proteinelor prin reticulul endoplasmic către aparatul Golgi şi ar putea mări
permeabilitatea membranelor celulare.
14

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 15/62
Proteina 2B pare să joace un rol structural în complexele de replicare virală, dar
funcţia sa exactă rămâne incă neelucidată. Se bănuieşte că blochează procesul de
secreţie a proteinelor celulare la nivelul reticulului ensoplasmic şi perturbă integritatea
aparatului Golgi.
Proteina 2C este cea mai conservată dintre proteinele enterovirale, dacă se ia în
considerare structura primară. Conţine trei domenii bine caracterizate: o structură
helicoidală amfipatică la capătul amino, un situs de legare pentru nucleozid trifosfaţi
(,, NTP binding’’) caracteristic helicazelor. Această proteină conţine şi locusul de
rezistenţă la guanidină, substanţă care blochează sinteza ARN de poliovirus, fiind
capabilă să inhibe complet replicarea enterovirusurilor la concentraţii milimolare.
Proteina 2C ar interveni mai ales în cursul iniţierii sintezei catenei ARN de polaritate
negativă. În complexele de replicare, proteina 2C şi precursorul său 2BC, sunt strâns
asociate împreună cu ARN viral de membranele veziculelor din reticulul endoplasmic şi
joacă un rol în formarea şi organizarea spaţiala a complexelor de replicare la nivelul
acestor vezicule. De asemenea, ar putea fi implicată în eliberarea ARN viral din
complexul de replicare precum şi în etapele tardive ale replicării, în încapsidarea
genomului virel şi asamblarea virionilor.Regiunea P3 codifică polipeptidul P3, precursor al polipeptidelor mature 3A pana la
3D. Proteinele 3 AB şi 3CD sunt stabile, având funcţii distincte faţă de cele ale
produşilor de clivare 3A, 3B, 3C şi 3D.
Proteina 3AB intervine la mai multe niveluri în cursul replicarii genomului viral: este
precursorul direct al proteinei 3B care este legată la extremitatea 5’ a ARN virali de
polaritate pozitivă şi negativă, fiind şi un presupus co-factor al polimerazei 3Dpol.
Proteina 3AB se leagă nespecific de ARN şi, în asociere cu proteina 3CD, poate realiza
o legătură specifică cu structură treflată din capătul 5’NC al genomului. Date genetice şi
biochimice au demonstrat că acest complex nucleoproteic joacă un rol esenţial în
replicare ARN viral. In plus, asocierea peoteinei 3AB cu proteina 3CD ar stimula
15

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 16/62
autoclivarea precursorului stabil 3CD în proteinele mature 3C şi 3D. În acelaşi timp
proteina 3 AB prezintă o afinitate pentru polimeraza 3Dpol şi această interacţie ar avea
un rol central în recunoaşterea de către polimerază a matriţei, precum şi în activitatea sa
enzimatică.
Proteina 3A, acesteia i se raibuie un rol în inhibarea comunicării dintre reticulul
endoplasmic şi aparatul Golgi. Conţine o secvenţă de 22 aminoacizi hidrofobi,
responsabilă de interacţia polipeptidului cu membranele şi de modificarea
permeabilităţii acestora. Mai recent, analiza mutanţilor a sugerat un rol al proteinei 3A
în efectul citopatogen indus viral.
Proteina 3B ( VPg ) se leagă la extemitatea 5’ a ARN genomic. Ea participă direct la
replicarea genomului viral. Un studiu recent a demostrat ca VPg, ar putea servi ca
amorsă (,, primer’’) pentru ARN polimerază în timpul sintezei ARN viral.
Proteina 3CD este precursorul direct al proteazei virale 3Cpro şi al polimerazei
virale 3Dpol. Este şi proteaza direct implicată în procesarea proteolitica la nivelul
situsurilor Gln/Gly din regiunea P1, precursor al proteinelor capsidei. Recent, proteina3CD a fost implicată şin în replicarea virală. Ea ar forma un complex ribonucleoproteic
cu extremitatea 5’ în formă de treflă a genomului şi o proteină celulară asociată
ribozomilor. Pentru a iniţia în trans sinteza ARN de polaritate pozitivă, proteina 3CD
poate să se asocieze şi cu proteina 3AB şi să formeze un complex 3Cdpro/3AB/ARN,
implicat în sinteza ARN viral.
Proteina 3C este proteaza virală majoră responsabilă de clivajul legăturilor
dipeptidice Gln/Gly din precursorii P2 şi P3., de clivajul primar dintre proteinele 2C şi
3A şi de autoclivarea sa din precursorul 3DC. Recent, strctura proteazei 3C a
poliovirusului a fost determinată prin cristalografie cu raze X.: ea prezintă o strcutură
asemănătoare cu cea a unei serin-proteaze, de tip chemotripsină. Această proteină ar
16

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 17/62
cliva şi anumite proteine celulare printre care: factorul de transcriere TFIIIC ,,TATA-
binding protein’’ (TBP, sub-unitate a factorului de trasncriere TFIIID) şi factorul de
transcriere CREB (,, c-AMP-responsive element-binding protein’’). Consecinţa directă a
inactivării acestor factori de transcriere este inhibarea trascrierii genelor celulare
asigurată de către ARN polimerazele celulare I, II, şi III.
Proteina 3D este ARN polimeraza dependentă de ARN, responsabilă de sinteza
ARN de polaritate pozitivă şi negativă. Activitatea sa este strict legată de prezenţa unui
primer nucleotidic împreună cu matriţa. In plus, 3Dpol prezintă şi una din proprietăţile
helicazelor şi anume cea de a derula duplexul ARN. I se atribuie şi activitate de terminal
adenil transferază cu rol ipotetic în replicare. Este bine stabilit că 3Dpol se asociază cu
factori virali, cum este proteina 3AB sau VPg, dai şi cu proteine celulare pentru a forma
complexe de replicare. Structura cristalografică a polimerazei 3D a permis localizarea a
4 domenii: A, B, C şi D, înalt conservate printre ARN polimerazele virale.
4. VIRIONUL
4.1. Arhitectura virionului
Studiile de microscopie electronicã cât şi apoi cele de difracţie cu raze X au arãtat ca
virionul are un diametru de 24-30 nm şi posedã o capsidã cu simetrie icosaedricã lipsitã
de un înveliş lipoproteic. Structura tridimensionalã a diferitelor enterovirusuri umane a
fost elucidatã prin cristalografie cu raze X : poliovirusul de tip I ( Hogie et.al, 1985 ) ,
cokackievirus B3 ( Muckelbauer et.al., 1995 ) , echovirus 1 ( Filman et. al., 1998 ) .Toate aceste enterovirusuri au o arhitecturã a virionului similarã cu celelalte
enterovirusuri din familia Picornaviridae. Capsida viralã este constituitã din 60 de
protomeri identici, fiecare compus din cele patru proteine structurale ; VP1,VP2, VP3 şi
VP4. Cinci protomeri formeazã un pentamer şi asocierea cu 12 pentameri constituie
17

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 18/62
procapsida. Cãnd aceşti 12 pentameri sunt asamblati, proteinele VP 1 sunt orientate în
direcţia axelor de simetrie de ordinul 5, în timp ce pe axele de simetrie de ordinul 3apare
o protuberanţã formatã din proteinele VP2 Şi VP 3. Proteinele VP4, situate exclusiv în
interior, cãptuşesc capsida în strânsã legãturã cu ARN genomic.
In ciuda diferenţelor majore la nivelul structurii lor primare, cele trei proteine: VP1,
VP2 şi VP3 prezintã o structurã tridimensionalã relativ asemanãtoare. Aceastã structurã
este compusã din douã helixuri de tip alfa şi din opt planuri beta antiparalele, pliate şi
legate între ele prin patru bucle de conexiune scurte. Extremitãţile amino ale proteinelor
VP1, VP2 şi VP3 formeazã o reţea pe partea internã a capsidei, în asociere cu VP4.
Buclele de conexiune ale proteinelor VP1, formeazã platouri în jurul axelor de
simetrie de ordinul 5. Acestea sunt înconjurate de o depresiune destul de profundã la
poliovirus şi rhinovirus, constituitã din aminoacizii celor trei proteine: VP1, VP2 şi
VP3, numitã ,, canion’’ , aceastã structurã a fost propusã ca situs de ataşare la receptorul
celular, atât pentru grupul rhinovirus ( Colonno et.al. , 1998 ) şi pentru poliovirus.
Descifrarea structurii tridimensionale a capsidei mai multor enterovirusuri a
determinat un progres considerabil n n elegerea func iilor acestei organiz ri,ȋ ȋ ț ț ӑ protejarea genomului viral de nucleazele de mediu, recunoa terea receptorului celular ș
specific ca prim determinant n specificitatea de gazd i n tropismul celular, suportȋ ӑ ș ȋ
pentru determinantii antigenici, sediu pentru informa iile privind ncapsidareaț ȋ
genomului viral i maturarea virionilor, ca i pentru maturarea virionilor, ca i pentruș ș ș
eliberarea genomului viral n celulele int ale gazdei.ȋ ț ӑ
4.2. Propriet ile fizico-chimiceӑț
Enterovirusurile sunt, ntre altele, definite i prin propriet ile lor fizico-chimice,ȋ ș ӑț
caracteristice acestui gen. Ele sunt rezistente la alcool etilic 70%, lizol 5%, amoniu
18

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 19/62
cuaternar 1% i fat de al i compu i similari cu rol dezinfectant. De asemenea, suntș ț șӑ
rezistente la tratamentul cu eter, dezoxicolat i al i solven i organici sau detergen i.ș ț ț ț În
schimb, enterovirusurile sunt termolabile : expunerea la temperaturi de 500C le distruge
rapid, dar inactivarea este par ial n prezen a clorurii de magneziu 1 M. Sunt virusuriț ӑ ȋ ț
foarte stabile la ph-uri acide ( ncep nd cu 2,5 ) ceea ce ilustraz o perfect adaptare laȋ ȃ ӑ ӑ
mediu : enterovirusurile trebuie s treac prin mediul acid din stomac nainte de aӑ ӑ ȋ
ajunge n zonele primare de replicare din intestin.ȋ
4.3. Determinan i antigeniciț
Propriet ile antigenice ale virusurilor au permis dezvolatarea principalelor metodeӑț
de diferen iere a acestora, implicit de identificare i clasificare. Trebuie s se tinț ș ӑ ӑ
seama de sistemul de identificare serologic , bazat pe antigenitatea virusurilor. In plus,ӑ
n cadrul profilaxiei maladiilor provocate de infec ii cu enterovirusuri, va trebui s seȋ ț ӑ
in seama ntotdeauna de antigenitatea acestora pentru prepararea vaccinurilor.ț ӑ ȋ
Una din caracteristicile familiei Picornaviridae este un num r impresionant deӑ
serotipuri ( peste 200 ) . Serotipul este definit cu ajutorul unui antiser monospecific care
este capabil s neutralizeze infectivitatea viral . Antiserul este produs prin imunizareaӑ ӑ unui animal care nu a fost expus anterior infec iei cu virusul care este investigat.ț
Statusul antigenic al unui virion integru poate fi conservat i n cazul unei particuleș ȋ
virale, a c rei infectivitate a fost abolit n urma tratamentului cu formaldehid . Acestaӑ ӑ ȋ ӑ
este principiul de preparare al vaccinului polio inactivat ( VP1 ) . Diverse tratamente
fizice sau chimice determin o denaturare a virusului, incapabil de a se mai adsorbi peӑ
celula gazd . El prezint un status antigenic modificat care este putin specific dec t celӑ ӑ ȃ
al virusului n stare nativ deoarece proteinele de la suprafa a virusului au oȋ ӑ ț
conforma ie modificat . ( Minor et.al. , 2000) de aceea, metodele care se bazeaz peț ӑ ӑ
identificarea
19

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 20/62
virusurilor dupa astfel de tratamente ( ex: tehnicile imunochimice ca: Elisa sub
imunoblot) trebuie completate cu alte tehnici mai fiabile.
Localizarea situsurilor antigenice de la suprafa a virionului este de importanț țӑ
major pentru n elegerea mecanismelor de neutralizare viral precum i a bazelor ӑ ȋ ț ӑ ș
moleculare ale diversit ii serotipuilor. Una dintre principalele caracteristici aleӑț
enterovirusurilor este num rul im presionant de serotipuri ( 64 pana n prezent ) .ӑ ȋ
Situsurile antigenice sunt reflectate de secven a proteinelor structurale recunoscute deț
c tre sistemul imun alӑ
unui animal injectat cu antigenele virale i sunt capabile s induc anticorpi careș ӑ ӑ
neutralizeaz infectivitatea virionului nativ. Caracterizarea mutatiilor de poliovirus careӑ
au rezistat la neutralizarea cu anticorpi monoclonali precum i a structuriiș
tridimensionale a capsidei au permis localizarea situsurilor antigenice de neutralizare nȋ
proteinele VP1, VP2 i VP3, la nivelul buclelor hidrofile expuse la suprafa a particuleiș ț
virale. Secven ele de aminoacizi implicatii n eptopi de neutralizare sunt grupate n treiț ȋ ȋ
situsuri antigenice de neutralizare : situsurile 1,2,3 , ultimul fiind divizat n trei domeniiȋ
3a, 3b, 3c ( Hogle & Filman, 1989 ; Mateu, 1995; Minor et.al., 1986 ; Page et.al., 1988)Cele mai multe situsuri antigenice de neutralizare sunt de fapt epitopi discontinui.
Ace tia sunt compu i fie din secven e peptidice scurte i cel mai adesea necontinue, fieș ș ț ș
din secven e situate pe proteine diferite de capsid . Diferen ele de conforma ie celeț ӑ ț ț
mai semnificative ntre serotipurile de polivirus sunt localizate la nivelul buclelor.ȋ
Astfel, exist o diferen important de conforma ie la nivelul buclei B-C din VP1.ӑ țӑ ӑ ț
Aceasta este mult mai expus n cazul poliovirusului de tip 3 Sabin. O mai bunӑ ȋ ӑ
accesibilitate a sa ar putea fi responsabil de imunodominan a situsului antigenic 1.ӑ ț
Aceast bucl B-C este supus unei varia ii intense de secven ( modific ri deӑ ӑ ӑ ț țӑ ӑ
lungime prin inser ii sau dele ii) , ntre enterovirusuri apar in nd unor grupuriț ț ȋ ț ȃ
20

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 21/62
taxonomice diferite i constituie , probabil, o structur important n diversificareaș ӑ ӑ ȋ
enterovirusurilor.
De i situsurile antigenice au fost intens studiate pentru poliovirusuri, cele ale altor ș
enterovirusuri sunt nca pu in cunoscute. Probabil c structura antigenic a acestoraȋ ț ӑ ӑ
este asem n toare cu cea a poliovirusului. Construc ia de virusuri himericeӑ ӑ ț
coxsackieivirus B3/B4 a permis identificarea unui situs antigenic de neutralizare
specific de serotip la nivelul buclei B-C din VP1 la coxackie B4 ( Reimann et.al.,
1991 ) totu i , aceast caracteristic nu se poate extinde la totatlitateaș ӑ ӑ
enterovirusurilor. Analiza antigenic a virusului coxackie A 9, a revelat faptul cӑ ӑ
anumite situsuri antigenice corespund cu
localizarea situsurilor 2 si 3 de la poliovrus , dar nici un epitop de neutralizare nu a fost
similar situsului antigenic 1 ( Pulli et.al., 1998 ) .
Patogenitatea, capacitatea virusurilor de a induce manifest ri clinice n organismeleӑ ȋ
infectate, este rezultatul unor interac iuni complexe la care particip factori implicaț ӑ
manifest ri clinice n organismele infectate, este rezultatul unor interac iuni complexeӑ ȋ ț la care particip factori implica i n carcteristicile biologice, biochimice i genetice aleӑ ț ȋ ș
virusului, pe de o parte, i reactivitatea gazdei, pe de alt parte. Boala se manifestș ӑ ӑ
clinic dac func ii importante sau vitale ale unor grupe de celule sunt afectate deӑ ț
replicarea viral . In general, nu toate celulele i esuturile din organismul infectat suntӑ ș ț
sensibile fa de infec ia viral . Tropismul unui virus pentru o celul gazdțӑ ț ӑ ӑ ӑ
reprezint , n primul r nd, o interac iune ntre componentele de suprafa ale virusuluiӑ ȋ ȃ ț ȋ țӑ
i receptorii de la nivelul membranei celulelor infectate. De aaemenea , n acest procesș ȋ
intervin i molecule intracelulare necesare multiplic rii virale,ș ӑ
5. Receptorii celulari
21

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 22/62
Evenimentul ini ial din ciclul infec ios al virusurilor este adsorbtia pe membranaț ț
celular unde se g sesc receptorii specifici. Enterovirusurile, care induc o patologieӑ ӑ
foarte divers , recunosc diferi i receptori celulari, n ciuda unei anumite unit iӑ ț ȋ ӑț strcturale
i genetice. Unele variante mutante se multiplic ns n celule care nu sunt permisiveș ӑ ȋ ӑ ȋ
pentru tulpinile parentale, ceea ce reflect o anumit flexibilitate a interac iunii dintreӑ ӑ ț
virus i gazd . Probabil c aceasta reprezint o strategie viral pentru a permite o maiș ӑ ӑ ӑ ӑ
mare variabilitate a tropismului celular i o palet mai larg de gazde, d nd sanse maiș ӑ ӑ ȃ
mari supravie uirii genului viral ( Evans & Almond, 1998 ). In plus receptorii nu ar fiț
responsabili numai de fixarea virusului la suprafa a celulei gazda ci ar putea interveni iț ș
n etapele precoce ale ciclului viral ( decapsidare i eliberarea ARN viral n celul ).ȋ ș ȋ ӑ
Receptorul poliovirusului ( PVR ) este prezent numai pe suprafa a anumitor celuleț
de primate. Gena care codific pentru PVR uman se g se te pe cromozomul 19.ӑ ӑ ș
( Koike, et.al., 1990 ) . Acest receptor specific este o glicoprotein din familiaӑ
imunoglobulinelor ( Mendelsohn, et.al., 1989 ) , al c rei rol fiziologic nu este ncӑ ȋ ӑ
cunoscut. Exist patru isoformeare PVR uman ( dou forme membranare( α i δ), careӑ ӑ ș
func ioneaz ca receptori i dou forme secretat ( β i γ) care nu sunt func ionaleț ӑ ș ӑ ș ț
pentru infec ia viral . Aceast molecul , puternic glicozilat , comport trei domeniiț ӑ ӑ ӑ ӑ ӑ
extracelulare cu o configura ie de tip V2-C2-C2, o regiune transmembranar i unț ӑ ș
domeniu intracelular. A fost demonstrat faptul c cel care interac ioneaz cu particulaӑ ț ӑ
viral este domeniul V2 ( Kolike, et.al., 1991 ) . Cu ajutorul anticorpilor ӑ monoclonali, care au blocat infec ia viral a unor celule, au putut fi identifica iț ӑ ț
receptori poten iali, ai altor enterovirusuri, astfel receptorul celular pentru echovirusț
( acum recunoscu i ca fiind un singur serotip ) a fost identificat ca fiindț integrina VLA-2
, ai c rei ligan i naturali sunt colagenul iӑ ț ș
22

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 23/62
laminina ( Bergelson et.al., 1992 ). Aceeasi autori au demostrat ca infec ia celulelor deț
c tre acest echovirus depinde de subunitatea αӑ 2 din VLA-2. Integrinele, glicoproteinele
membranare heterodimerice, sunt moleculele responsabile de adeziunea intercelular iӑ ș
de interac iunile celulelor cu matricea extracelular , dar i de ata area viral la celuleț ӑ ș ș ӑ
i infectarea acestora. Un prim motiv identificat din proteinele matricei extracelulareș
care interac ioneaz cu integrinele este tripeptidul RGD. Nu ntampl tor,ț ӑ ȋ ӑ
coxackievirus A9
prezint o inser ie de 17 aminoacizi la extremitatea C-terminal a proteinei de capsidӑ ț ӑ ӑ
VP1 care include i motivul RGD. ( Rovainen, et.al., 1991 ), au demontrat ca peptideleș
sintetice care con in motivul RGD blocheaz ata area virusului coxsackie A9 pe celule,ț ӑ ș
ceea ce sugereaz faptul c o integrin serve te ca receptor pentru acest virus. Esteӑ ӑ ӑ ș
vorba de integrina αv β3 ( pentru care ligandul natural este vitronectina ), receptorul
celular pentru Coxackievirus A9 ( Rovainen , et.al., 1994 ). A fost demostrat existen aӑ ț
unor receptori suplimentari pentru CV-A9 ( β2 microglobulina i o protein de 70 KDaș ӑ
asociat CMH de tip 1 ), necesari n evenimentele de internalizare ( Triantafilou , et.al.,ӑ ȋ
2000 ). A fost sugerat i faptul c βș ӑ 2 microglobulina reprezint un cofactor n etapeleӑ ȋ precoce ale ciclului viral pentru echovirus 7 ( Ward, et.al, 1998 ), integrina αv β3 este
molecul candidat i pentru receptorul celular al echovirusului 9 Barty. Aceastӑ ӑ ș ӑ
secven RGD ar putea explica caracterul patogen al acestei tulpini pentru oriceii nou-țӑ ș
n scu i, motiv care lipse te din capsida tulpinii nepatogene de Echovirus 9.ӑ ț ș
Pentru numeroase tulpini de echovirus ( E 3, 6,7,11-13, 19-21, 24, 25, 29, 30, 33 ) ca
i pentru anumite coxackievirusuri B ( CV-B1, B3 i, B5 ), pentru coxackie virus A 21ș ș
sau pentru enterovirus 70, unul din elementele complexului de receptori ar fi format din
molecula DAF ( ,, Decay Accelranting Factor sau CD 55) . DAF intervine n reglareaȋ
activit ii complementului i n semnalizarea intercelular . Exprimarea sa esteӑț ș ȋ ӑ
ubicvitara, inclusiv pe eritrocite, ceea ce explic fenotipul hemaglutinant alӑ
23

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 24/62
enterovirusurilor care utilizeaz acest receptor. DAF este o glicoprotein de 70 KDa iӑ ӑ ș
este definit prin ase domenii: patru regiuni SCR ( ,, Short Consensus Repeat’’) deӑ ș
aproximativ 60 aminoacizi, o regiune bogat n serin /treonin i o regiune C-ӑ ȋ ӑ ӑ ș
terminal membranar , fixat covalent pe un acid gras. Regiunea SCR-1 este necesar ӑ ӑ ӑ ӑ
pentru interac iunile cu CV-A21, EV-70 i mai general cuț ș Human Enterovirus C si D,
pe c nd regiunea SCR-4 este necesar pentruȃ ӑ Human Enterovirus B . Dar exprimarea sa
la suprafa a celulei nu este suficient pentru ca infec ia s aib loc.ț ӑ ț ӑ ӑ
In ultimii ani, numeroase lucr ri au ar tat c virusurile ( inclusiv enterovirusurile nuӑ ӑ ӑ
utilizeaz un receptor celular unic ci , mai degrab , un complex receptor compus dinӑ ӑ
mai multe elemente cuprinz nd proteine de ata are i molecule accesorii ( Evans&ӑ ș ș
Almond, 1998) . In lumea virusurile, enterovirusurile manifest cea mai mareӑ
diversitate privind tipul de receptor. Acest lucru este ilustrat foarte bine de
coxackievirusurile B,
capabile s utilizeze cel pu in trei receptori diferi i ( CD55, CAR i o proteinӑ ț ț ș ӑ
mambranar din familia nucleolinelor ). Pentru receptorii ale caror fun ii r m n a fiӑ ț ӑ ȃ determinate ( de ex: CAR i PVR uman ), remarcabil este faptul c ace tia reprezintș șӑ ӑ
molecule din familia imunoglobulinelor i deci, susceptibile s joace un rol nș ӑ ȋ
imunitate. Alegerea unei familii de proteine comun pentru mai multe virusuri ar puteaӑ
fi consecin a unei presiuni selective n timpul evolutiei enterovirusurilor.ț ȋ
6. MULTIPLICAREA VIRALA
Ciclul complet de multiplicare a enterovirusurilor este unul dintre cele mai scurte si
durează in vitro aproximativ 8 ore. Având o localizare exclusiv citoplasmatică, ciclul
24

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 25/62
viral este iniţiat prin fixarea virusurului pe receptorul celular şi se termină prin liza
celulei infectate şi eliberarea virionilor nou formaţi. Studiul poliovirusului – prototipul
enterovirsurilor – a furnizat ansamblul elementelor care au permis înţelegerea ciclului
de multiplicare enterovirală.
6.1. Etapele precoce: adsorbţia, pătrunderea şi decapsidarea
25

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 26/62
Pentru iniţierea ciclului de relicare, virusul trebuie sa interacţioneze cu receptorul
celular şi să elibereze acidul nucleic în compatimentul celular adecvat. Dar mecanismul
prin care enterovirusurile introduc ARN în celula ţintă ramâne controversat. In general
este admis faptul că interacţiunea enterovirusurilor cu receptorul lor specific antrenează
modificări conformaţionale importante ale particulei virale. La temperatură fiziologică,
aceste intercţiuni determină aparitia unor particule stabile numite A ( 135S), mult mai
hidrofobe decât particulele native şi capabile de internalizare celulară. Această structură
intermediară de 135S este obiect al unor controverse: Dove şi Racaniello au sugerat ca
formarea nu pare absolut necesară infecţiei virale productive, iar Curry et al. au arătat că
aceste particule, spre deosebire de particulele native, pot infecta celule care nu exprimă
receptorul. Totuşi date recente pledează în favoarea rolului de intermediar al particulei
135S pentru intrarea virusului în celulă. Aceşti autori propun chiar o etapă suplimentară
in procesul de intrare, care ar avea loc după formarea particulei 135S şi ar fi asociată cu
eliberarea ARN. Schimburile conformaţionale ale capsidei virale sunt, în principal,
caracterizate prin pierderea proteinei VP4 şi prin externalizarea extremităţii N-terminale
hidrofobe a proteinei VP1, antrenând astfel reducerea coeficientului de sedimentare de
la 160S al virionului intact la 135S pentru particula modificată. Această particulămodificată, care prezintă o mai mare afinitate pentru membrane, a fost propusă ca
intermediar obligatoriu în mecanismul internalizării virusului. Într-adevăr, aceste
schimbări conformaţionale par a fi indispensabile decapsidării, care începe imediat ce
virusul este adsorbit pe membrana celulară.
6.2. Multiplicarea la enterovirusuri
Dintre particulele adsorbite, 50% sunt internalizate, restul sunt eliberate în mediul
extracelular prin procesul numit eluţie.
26

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 27/62
Particulele neeluate intră în celulă fie pătrunzând direct prin membrana celulară dupa
legarea de receptor, fie printr-un mecanism de endocitoză mediată de receptorul celular.
În acest ultim caz, acidifierea endozomului nu este necesară pentru decapsidare şi pentru
trecerea genomului viral prin membrana endozomală. S-a sugerat ca translocarea ARN
viral in citoplasma celulei infectate ar putea să se realizeze prin formarea unor pori
trans-membranari apăruţi prin inserţia extremităţii N-terminale a VP1 şi, eventual, a
proteinei VP4 miristilate. Acest model a fost susţinut prin detectarea unui canal ionic,
după punerea în contact a particulelor virale cu membrane lipidice.
6.3. Sinteza virală: translaţia si replicarea genomului viral
După eliberarea ARN în citoplasmă, acesta este tradus direct de către aparatul
translaţional al celulei gazdă. Proces anticipat de clivarea proteinei VPg din extremitatea
5’a genomului de către o protează de origine celulară. Spre deosebire de ARN de la
eucariote, genomul enterovirusurilor nu prezintă o structură ,,cap’’ sau ,,bonetă’’ ,etilată
la extremitatea 5’. Iniţierea translaţiei se face printr-un mecanism numit ,,iniţiere
internă’’: subunitatea 40S a ribozomului recunoaşteun segment cu o structură secundară
particulară al regiunii 5’NC, IRES, care este localizat cu 150 de nucleotide situate în
amonte de codonul start. Subunitatea 40S migrează apoi până la primul AUG care se
găseşte intr-un context favorabil iniţierii translaţiei. Se pare că buna funcţionare acodonului start necesită prezenţa în amonte a unui tandem constituit dintr-o secvenţă
bogată în pirimidină şi intr-un AUG criptic. Este interesant de notat că regiunea 5’NC
conţine 6-8 codoni AUG care nu sunt utilizaţi pentru iniţierea translaţiei. În plus fată de
27

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 28/62
factorii ,,clasici’’ ai inţierii, activare translaţiei virale necesită o serie de factori celulari,
care se fixează specific pe structuri secundare din regiunea 5’NC, cum sunt
autoantigenul LA( p52), ,,polypyrimidibe-tract-binding protein’’ (PTB), factorul
eucariot de iniţiere a translaţiei eIF- 2α şi ,, poly (rC)-binding protein 2’’ ( PCBP2).
Aceste proteine care se leagă de ARN ar acţionaca molecule ,, chaperones’’ care ar
menţine IRES în conformaţia necesară pentru interacţiunea cu sistemul translaţional
celular, în special cu eIF- 2α . Aşa cum a fost deja descris, genomul enterovirusurilor
este în final translat într-o singură poliproteină, care este clivată cotranslaţional de către
proteazele virale neosintetizate – 2Apro, 3Apro şi 3Cdpro, pentru a da naştere
proteinelor de capsidă şi celor nestructurale.
În momentul în care suficiente proteine virale au fost sintetizate, se produce o
basculare de la translaţie la replicarea genomului viral, în principiu aceste două
fenomene neputând avea loc simultan. Gamarnik şi Andino au propus existenţa la
nivelul structurii în treflă din 5’NC a unor semnale care controlează atât translatia cât şi
replicarea virală, prin formarea unor complexe ribonucleoproteice. In acest model,
interacţiunea dintre PCBP celulară şi această structură ar favoriza traslaţia. După sinteza
precursorului viral 3CD, aceasta se leagă de structura treflată şi înhibă astfel translatia înfavoarea sintezei actenei de ARN de polaritate negativă.
Mecanismul replicării genomului enterovirusurilor este încă puţin înteles. In celulele
normale nu este cunoscută o activitate de tip ARN polimerază dependentă de ARN.
ARN polimeraza virală 3Dpol este strict necesară oentru virus. Totuşi, această enzimă
nu este suficientă şi replicarea necesită nu numai proteine virale nestructurale dar şi
anumite proteine celulare.
Procesele replicative au loc în citoplasmă, la suprafaţa externă a veziculelor formate
din organite celulare cum sunt: reticulul endoplasmic, aparatul Golgi şi lizozomii.
Aceste vezicule sunt probabil induse de proteinele virale 2BC şi/sau 2C şi se
organizează în rozete în jurul complexelor de replicare. Aceste complexe sunt
28

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 29/62
constituite din membrane celulare netede, molecule matrice de ARN, molecule în curs
de a fi sintetizate, moleculele de polimerază virală 3Dpol, alte proteine virale şi din alţi
factori celulari. Replicarea începe prin sinteza unei catene de ARN negativ,
complementar genomului viral; acesta va servi apoi la rândul său ca matrice pentru
sinteza numeroaselor molecule de ARN de polaritate genomică pozitivă. În citoplasma
celulei infectate se mai găsesc forme replicative (RF), constituite din actene pozitive şi
negative ca hibrizi în dublu helix precum şi forme intermediare de replicare ( RI),
constituite dintr-o matrice asociată cu mai multe catene de polaritate opusă în curs de
sinteză.
Este în general admis că numai regiunile 5’NC şi 3’NC ale genomului sunt implicate
în mecanismele de iniţiere a replicării, graţie structurilor secundare şi a prezenţei
semnalelor de repliacare care acţionează in cis, numite cre. Extremitatea 3’ ( oriR) ar
interveni în iniţierea în cis a sintezei catenei negative, iar extremitatea 5’ ( oriL) ar fi
necesară pentru iniţierea în trans a replicării catenei pozitive. Totuşi, o regiune de tip
cre a fost recent identificată şi în regiunea care codifică pentru proteina 2C .
6.4. Etapele tardive: morfogeneza şi eliberarea vironilor.
Etapa iniţială a morfogenezei virale este eliberarea procursorului miristilat P1 al
poliproteinei in urma activităţii autocatalitice a proteinei 2Apro. Precursorul P1 suferă
apoi o clivare de către proteaza 3CD, pentru a da naştere proteinelor de capsidă VP0,
VP1 şi VP3. Procesul de asamblare virală poate astfel să înceapă: proteinele structurale
se asociază pentru a forma un protomer care sedimentează la 5S şi care este cea maimică sub-unitate a capsidei. Cinci protomere se combina apoi pentru a forma sructura
pentamerică de 14S. Urmează asamblarea a 12 pentamere care formează procapsida de
75S. Etapa următoare este unirea ARN viral cu capsida imatură pentru a constitui o
particulă instabilă, provirionul de 125S.
29

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 30/62
O ultimă etapă de maturare este necesară pentru formarea virionului infectant, în
care precursorul VP0 este clivat în VP4 şi VP2 printr-un mecanism probabil
autocatalitic. Această clivare şi încapsidarea ARN declanşează un rearanjament
structural care este un proces specific, în sensul că nici un alt tip de ARN viral sau
celular nu poate fi încapsidat. Specificitatea s-ar putea explica prin cuplarea funcţională
dintre morfogeneză şi replicarea virală.
Asamblarea odată realizată, particulele mature se acumulează în citoplasma celulelor
infectate sub forma unor incluziuni cristaline, după care sunt eliberate prin spargerea
vacuolelor la suprafata celulelor. Eliberarea masivă de virioni nou sintetizaţi este
concomitentă cu liza celulei, în care proteina virală 2B ar putea juca un rol important.
7. EPIDEMIOLOGIA
Transmiterea este in general de tip fecal-oral, difuzarea virusurilor fiind
sporadică, endemică, epidemică şi chiar pandemică, cu predilecţie pentru senzorul vară-
toamnă în regiunile temperate. Anumite serotipuri domină şi circulă simultan timp de
mai mulţi ani, apoi scad ca frecvenţă, revenind după un timp pentru initierea unor noi
epidemii. Serotipurile dominante sunt insă imprevizibile de la un an la altul.
7.1. Moduri de transmitere
Sunt cunoscute moduri de transmitere a enterovirusurilor, fiecare influentând în mod
diferit caracteristicile epidemiologice ale infecţiei provocate. Schema principală de
difuzare a enterovirusurilor este contaminarea fecalo-orală, fiind confirmată în primul
rând în ţările în curs de dezvlatare unde condiţiile sanitare sunt precare dar şi în anumite
comunităţi din ţarile dezvoltate cum sunt creşele şi şcolile. De fapt, enterovirusurile de
30

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 31/62
transmit de la individ la individ prin intermediul mâinilor contaminate, a obiectelor, a
alimentelor lichide sau solide stropite cu salivă sau prin materii fecale. În contextul
conjunctivitelor hemoragice acute provocate de enterovirus 70 sau de coxackievirus
A24, secreţiile înalt infecţioase se transferă prin intermediul mâinilor şi chiar prin
instrumentar oftamologic insuficinet sterilizat. Calea aeriană poate să joace şi ea un rol
în transmiterea unor serotipuri cu tropism pentru tractul respirator. Nu este de neglijat
nici transmiterea oral-orală la populaţii cu nivel socio-sanitar ridicat.
7.2. Epidemiologia moleculară
Epidemiologia moleculară permite recunoaşterea tulpinilor diferite din aceeaşi
specie, provenind din circumstanţe epidemiologice cariate, pe baza polimorfismului
genetic. Dintre mecanismele care determină polimorfismul genetic, mai importante sunt
variabilitatea naturală determinată de polimeraze (în special ARN polimeraze) şi
modificările genetice (deleţii, inserţii, recombinări, trasnlocaţii).De cativa ani, epidemiologia virusurilor s-a imbogăţit cu o serie de tehnici
moleculare noi care permit caracterizarea diferenţelor genetice şi antigenice între
diferite izolate. Aceste tehnici recente, fac posibile investigaţii privind originea şi
evoluţia tulpinilor, avănd astfel implicaţii în prevenirea anumitor boli enterovirale cu
importanţă deosebită pentru sănătatea publică.
In acelaşi timp, epidemiologia moleculară permite ameliorarea metodelor de
diagnostic şi terapeutice precum şi identificarea unor markeri genomici (de exemplu
secvenţele din genom care pot fi asociate cu o creştere a virulenţei sau cele implicate în
modificările către fenotipuri patogene).
31

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 32/62
8. PATOGENEZA
Poarta de intrare a enterovirusurilor este calea orală (cu excepţia enterivirusurilor
asociate cu conjuctivitele). Situsurile primare ale replicării virale sunt mucoaselefaringiană şi intestinală, virusurile ajungănd în intestin după ce au trecut de bariera
stomacală în virtutea rezistenţei lor la pH acid.
Enterovirusurile traversează bariera intestinală prin celulele M din structurile limfoide
de la nivelul plăcilor Peyer. Această fază digestivă din intestinul subţire ar fi urmată de
o fază limfatică prin care virusul ajunge în ganglionii cervicali şî mezenterici. Faza
viremică ce urmează ar putea fi întreţinută prin invadarea acestor ganglioni, precum şi a
altor ţesuturi. Viremia tranzitorie este, din punct de vedere clinic, silenţionasă sau este
însoţită de semne clinice nespecifice cum sunt vomă, febră, dureri în gât, etc. După ce
virusul trece în sânge, acesta se multiplică în celulele sistemului reticulo-endotelial apoi
ajunge în organul ţintă. Perioada de incubaţie, acoperind toate aceste faze, este în
general de 7 la 14 zile, dar poate fi şi de 2 la 35 de zile. După apariţia semneleor clinice,
care reflectă leziuni la nivelul organelor ţintă, virusul dispare destul de repede din
orofaringe, dar multiplicarea intestinală poate persista cu o excreţie virală prin materiile
fecale, timp de mai multe săptămâni. Fiecare tip de enterovirus are propriul său organ
ţintă: creier, inimă, muşchi, piele, etc. Izolarea enterovirusurilor se face din diferite
tipuri de probe clinice: fecale, secreţii faringiene, lichid cefalo-rahidian ( LCR), maduva
spinării, creier, inimă, sânge, secreţii oculare (conjunctivite) sau din leziuni ale pielii sau
ale mucoaselor. Poliovirusul infectează sistemul nervos central (SNC) şi astfel trebuie
să traverseze bariera hemato-encefalică, fie direct, fie prin infectarea celulelor endoteliale . Deşi calea sangvină rămâne ipoteza privilegiată pentru a se explica pasajul
poliovirusului către SNC, s-a propus şi ipoteza migrării virusului pe cale nervoasă prin
nervii periferici. Ţinta preferenţială a poliovirusului este neuronul motor periferic din
coarnele anterioare ale maduvei spinării sau din echivalentele lor de la nivelul
32

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 33/62
trunchiului cerebral. Distrugerea neuronului motor periferic determină paralizia
musculară flascăa zonelor dependente topografic de neuronii distruşi. Infecţiile
simultane de către mai multe serotipuri de enterovirusuri sunt posibile, dar pot apărea
competiţii la nivelul intestinului între diferitele virusuri.
Patogeneza enterovirală descrisă astfel, cu referire la o infecţie acută, este cea
adoptată în mod clasic, dar se cunosc şî câteva excepţii. Enterovirozele cronice şi
conjunctivita hemoragică acută nu corespund schemei prezentate. În conjuctivita
hemoragică acută, contaminarea este directă, pe cale conjunctivală şi incubarea este de
24 la 48 de ore. Virusul are o temperatură optimă de replicare mai scăzută, de 33-35 °C
şi se izolează din secreţiile conjunctivale şi din oro-faringe, dau nu şî din materii fecale.
Enterovirusurile sunt o cauza destul de frecventa de encefalita acuta virala,
diagnosticul etiologie este dificil, dozarea de IgM care stabileste diagnosticul de infectie
acuta nefiind efectuata intotdeauna. Un rol important il are si deficitul de IgA, precum si
valoarea scazuta a complementului seric care sunt factori esentiali in mecanismele de
aparare. De altfel, copiii cu diferite deficiente ale sistemului imunitar sunt predispusi la
infectii cu enterovirusuri, difera atat de serotipul implicat dar mai ales de statul imun al
gazdei. Este de remarcat aparitia ambelor episoade in luna august, perioada de incidentamaxima a infectiei cu enterovirusuri.
Particularitatea cazului Encefalita acuta cu enterovirusuri ( ECHO si Cocsackie),
manifestata prin ataxie cerebeloasa, repetata la un interval de un an, la un copil cu
deficit imunologic IgA pe fond de valori scazute ale complementului seric, cu raspuns
favorabil la tratamentul patogenic si simptomatic aplicat.
33

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 34/62
CAPITOLUL II
TEHNICI DE BIOLOGIE MOLECULARA
I. Reacţia de polimerizare în lanţ ( PCR)
1. Generalitãţi:Reacţia de polimerizare în lanţ ( polymerase chian reaction, PCR), este o metodã de
sintezã ADN in vitro, prin care o anumitã secvenţã din aceasta este amplificatã în mod
specific. Principalii reactanţi care intervin în procesul de amplificare sunt:
- primerii /amorse sunt douã oligonucleotide cu secvenţa complementarã capetelor
3’ ale fragmentului de ADN ţintit, orientarea lor permiţând sinteza secvenţei din
regiunea pe care o delimiteaza.- Deoxinucleozid trifosfaţii aceştia fiind unitãţile constitutive ale acizilor
deoxiribonucleici, respectiv dATP, dCTP, dGTP, dTTP
- ADN polimeraza termorezistentã , aceasta fiind enzima care catalizeaza
construirea catenelor complementare prin extensia primerilor
34

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 35/62
Reacţia se desfaşoarã într-un tampon care contine Tris-HCl, KCl, MgCl2
( concentraţia optimã de Mg2+ variind între 1-10 mM) şi alţi aditivi ( gelatina , tween 20,
Nonidet P-40, Triton X-100) .
Reacţia de PCR se desfaşoarã pe parcursul a mai multor cicluri ( 25-45), fiecare
constituit din trei etape:
1. denaturarea ADN se realizeazã prin expunerea la temperaturi de 94-95oC
2. hibridizarea primerilor la monocatenele ADN , se realizeazã la temperaturi de
45-68oC
3. extensia primerilor la capãtul 3’, respectiv sinteza de ADN la temperatura de
72oC,temperatura optima de activitate a Taq-polimerazei.
Rezultatul acestui proces ciclic repetat este o crestere exponenţialã a numãrului total
de fragmente ADN care includ secvenţeledintre primeri PCR utilizaţi. Teoretic
amplificarea finalã a matriţei ADN ţintã este de 2n, unde n reprezintã numãrul de cicluri
PCR.
Produsul de reacţie ( amplicon) cumuleazã dimensiunea celor doi primeri şi a
secvenţei dintre ei. El poate fi identificat, respectiv verificat ca specificitate, în maimulte moduri. Cel mai frecvent se recurge la aprecierea lungimii sale precis definitã
( vizualizare in UV, dupã migrare în gel de agarozã şi colorare cu bromurã de etidiu),
exprimatã în perechi de baze azotate ( bp) şi comparatã cu un martor de greutate
molecularã. Acestei analize i se poate adãuga evidenţierea perezenţei anumitor situsuri
de restricţie, prin digestia ampliconului cu endonucleaze de restricţie . Identificarea
prodului PCR se poate face şi prin hibridizarea lui cu o sondã specificã sau prin
secvenţiere.
Optimizarea parametrilor reacţiei PCR este de cea mai mare importanţã pentru
sensibilitatea si specificitatea reacţiei.
35

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 36/62
2.Componentele reacţiei:
2.1. Matriţa ADN
Proba PCR trebuie sã fie un ADN monocatenar sau dublucatenar, liniar sau
circular. Puritatea ADN folosit ca matriţã reprezintã un factor deosebit de important ( o probã infectatã cu cantitãţi mari de ARN va dispune de o activitate scãzutã a ADN
polimerazei în reacţie , ARN fiind rãspunzãtor de chelatarea ionilor de Mg2+ ).
Cantitãţile recomandate pentru o reacţie PCR sunt: maximum 500 ng pentru ADN
genomic uman, 1-10 ng pentru ADN bacterian şi 0,1-1 ng ADN plasmidial.
2.2. Primeri oligonucleotidici
In general dimensiunea primerilor utilizaţi în reacţiile PCR este cuprinsã între 15-30
nucleotide ( folosirea unor primeri mai lungi de 30 nucleotide duce la creşterea
specificitãţii). De regulãprimerii sunt cinstituiti astfel încât sã aibã complementaritate
perfectã sau aproape perfectã cu capetele 5’ şi repspectiv 3’ a matriţei de ADN. Este
recomandat ca primerii sã aibã un procent molar de citozinã ( C) + guaninã ( G) cuprins
între 40-60% şi sã aibã o distribuţie echilibratã a celor patru baze. Primerii nu trebuie sa
fie complementari între ei ( pentru a nu dimeriza), sã nu prezinte secvenţe cu
complementaritate intracatenarã ( sã nu formeze structuri secundare interne) şi sã aibã
valori ale Tm ( temperatura de topire) identice sau foarte apropiate. Se recomandã ca cei
doi primeri sã aibã Tm cuprins între 55-65 oC ( pentru creşterea specificitãţii se impun
temperaturi de ataşare ridicate: 62-65o C)
Concentraţia optimã de primer e cuprinsã între 0,1 şi 0,6 μM. Concentraţii prea
mari de primer duc la obţinerea produşilor nedoriţi.
2.3. ADN polimerazele termostabile
Iniţial, pentru reacţia PCR s-a utilizat fragmentul mare al ADN polimerazei I de la
Escherichia coli – fragmentul Klenow. Marele avantaj al acestei enzime ers faptul ca se
36

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 37/62
inactiva la temperaturile ridicate ale etapei de denaturare. Ca urmare, la fiecare ciclu de
replicare trebuia adãugatã o nouã cantitate de enzimã în tubul de reacţie.
In prezent toate reacţiile PCR folosesc ADN polimerazele termostabile derivate ( în
urma manipulãrii genetice) de la variantele naturale, izolate din microorganisme
termofile. Cantitatea optimã de polimerazã utilizatã este cuprinsã între 0,5-2,5 unitãţi /
50 μl volum de reacţie. ADN polimerazele ADN dependente termostabile se împart în
mai multe clase:
1. Clasa Taq
a. Taq polimeraza -izolatã din microorganismul Thermus aquaticus. Temperatura
optima de activitate este de 75-78 oC, iar la temperaturi de peste 90oC, activitatea sa
scade semnificativ. Polimerizeazã cu o vitezã de aproximativ 150 nc/ secundã. Are o
activitate de polimerizare ( în direcţia 5’→ 3’ ) şi exonucleazicã în aceeaşi direcţie. Este
însã lipsitã de activitate exonucleazicã în direcţia 3’→5’, responsabilã de citirea şi
încorporarea corectã a dNTP ( nu are activate proofreading ). Reacţia PCR mediatã de
aceastã enzimã are loc în trei trepte ( ataşarea primerilor şi polimerizarea au loc la treptediferite de temperaturã).
b. Ampli-Taq – ADN polimeraza recombinantã obţinutã prin exprimarea unei forme
modificate a genei de Taq pot din E.coli. Se activeazã la temperatura de 55-75 0C
( proprietate ce determinã simplificarea reacţiei PCR de la trei trepte de temperaturã la
douã trepte ). Are o stabilitate mult mai mare la temperaturi de peste 90 oC şi prezintã în
plus activitate exonucleazicã 3’→5’ ( creşte specificitatea de încorporare a
nucleotidelor)
c. Fragmentul Stoffel – este reprezentat de un Ampli-Taq trunchiat la capãtul
N-terminal. Fragmentul excizat find responsabil de activitatea exonucleazicã 3’→5’,
absenţa lui mãreşte mult viteza de polimerizare, Temperaturi de polimerizare: 55-60oC.
37

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 38/62
Este de doua ori mai stabilã decât Ampli-Taq şi işi pãstreazã activitatea optimã pe o
plaja mare de concentraţii de Mg2+. Se utilizeazã în PCR cu douã trepte.
2. Clasa Vent şi Deep Vent
a. Vent pol – izolatã din Thermococcus litoralis, bacterie ce trãieşte în izvoarele
marine calde. Temperatura optimã : 55-60oC şi desfãşoarã ambele tipuri de activitate
exonucleazicã ( 3’→ 5’ şi respectiv 5’→3’ )
b. Vent pol ( exo)- similarã cu Vent pol, dar nu are activitate exonucleazicã 5’→3’
c. Deep Vent pol – izolatã dintr-o tulpinã de Pyrococcus, din izvoarele marine calde
de mare adâncime. Temperatura optimã: 55-60oC şi desfãşoarã ambele tipuri de
activitate exonucleazicã, dar este mai rezinstentã decât Vent pol şi Ampli Taq pol la
temperaturi mai mari de 100oC.
d. Deep Vent Pol ( exo) – similarã cu DEP Vent pol, dar nu prezintã activitate
exonucleazicã 5’→3’.
3. Clasa ADN polimerazelor ambivalente termostabilerTth pol – ADN polimerazã recombinantã. Caracterul ambivalent constã în faptul cã
poate fi folositã ca ADN polimerazã ARN dependentã în prezenţa MnCl2, dupã care,
prin chelaterea ionilor de Mn2+ şi adãugarea MgCl2, poate acţiona ca ADN polimerazã
ADN dependentã. Cantitatea optimã de enzimã utilizatã într-o reacţie PCR este de 0,5-
2,5 unitãţi/ 50μl volum de reacţie. Cantitãţi crescute de enzimã sau timpi prelungiţi de
polimerizare pot genera artefacte, datoritã activitãţii exonucleazice 5’→
3’ a Taq pol.
2.4. Deoxinucleotid trifosfaţi ( dNTP)
DNTP ( dATP, dGTP, dCTP, dTTP ) sunt folosiţi de polimerazã în etapa de elongare
a PCR. În amestecul de reacţie de introduc cantitãţi echimoleculare din cele patru tipuri
38

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 39/62
de dNTP ( în caz contar scade fidelitatea ADN polimerazei ). Cantitatea optimã utilizatã
în PCR este de aproximativ 200 μm. Concentraţii mai mari conduc la reducerea
concentraiei de Mg2+ liber, influenţând activitatea polimerazei şi scade ataşarea
primerilor. Din acestã cauzã concentraţia lor trebuie corelatã cu concentraţia de Mg2+.
2.5. Tamponul de reacţie
Tamponul de reacţie conţine : TRIS-HCl, pH=8,3, KCl, MgCl2, în soluţie
concentratã 10X care trebuie diluatã 1:10 ( v/v). Concentraţia MgCl2 are o importanţã
crucialã în reacţie, ionii de Mg2+ fiind necesari pentru buna funcţionare a polimerazei.
Ionii de Mg2+ formeazã complexe solubile cu moleculele dNTP pentru a produce
substratul propriu-zis recunoscut de adn apolimerazã. Un exces de Mg2+ va determina
însã reducerea fidelitãţii enzimei, creşterea valorii Tm şi creşterea amplificãrilor
nespecifice. Concentraţia optimã intr-o reacţie sã poatã fi optimizatã, existã şi varianteîn care tamponul este livrat fãrã Mg2+, acesta fiind livrat separat.
2.6. Uleiul mineral
In cazul aparatelor fãrã capac încãlzit , un strat de ulei mineral care sã acopere
amestecul de reacţie este necesar, în vederea prevenirii evaporãrii volumului de reacţie.
Orice evaporare determinã o cocentrare a componentelor PCR şi o descreştere a
temperaturii.
3. Detectarea produşilor PCR
39

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 40/62
Cea mai utilizatã metodã de detecţie este electroforeza în gel de agarozã sau
poliacrilamidã. Se mai poate utiliza şi Southern Blotting ( detectarea şi identificarea
produşilor PCR se realizeazã prin hibridizare cu sinde specifice marcate radioactiv sau
neradioacvtiv).
Gel electroforeza- este ceam mai simplã metodã. Detectarea Produşilor PCR se
realizeazã pe un gel de agarozã, de concentraţie diferitã în funcţie de lungimea
fragmentului de amplificat. Gelul va fi marcat cu bromurã de etidiu, un colorant
fluoreşcent ce se intercaleazã între perechile de baze ale ADN şi favorizeazã
vizualizarea benzilor carecateristice ampliconilor ( sub acţiunea UV). De asemenea, pe
gel se incarcã şi un marker de greutate molecularã ( pentru identificarea fragmentului de
interes), cât şi martorii pozitivi şi negativi.
De obicei, pentru vizulaizarea produşilor de amplificare este suficientã agaroza cu
rezoluţie standard . Pentru evidenţierea produşilor mici se folosesc însã geluri de
agarozã de înaltã rezoluţie.
3.1. Contaminarea şi evitarea contaminãrii în reaţia PCR Potenţiale surse de contaminare : probe biologice ( alt ADN ), metoda de colectare a
probei, manipularea probei de mai multe persoane, utilizarea de reactivi şi
instrumentarul de laborator ( tuburi de reacţie, pipete, tipsuri, termocycler, autoclav,
centrifugi, gheaţã, etc) contaminat, mediu de laborator sau aer condiţionat.
Este important sã se realizeze controlul pozitiv şi negativ pentru fiecare reacţie PCR
efectuatã. Prin martori negativi se verificã dacã reactivi sunt contaminaţi cu ADN , iar
prin martorii pozitivi se verificã eficientã şi specificitatea reacţiei PCR.
Evitarea contaminãrii este un lucru foarte important. Se realizeazã prin utilizareade
hote de flux laminar şi lampã de UV, utilizarea de mãnuşi, tipsuri cu filtru, utilizarea de
40

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 41/62
pipete diferite pentru mix şi probã, reactivii PCR trebuie sã fie alicotaţi şî pãstraţi la
-20oC, orice material ce poate fi autoclavat trebuie sterilizat corespunzãtor.
Alte procedee de decontaminare : iradierea UV, tratamentul enzimatic,uracil N-
glicolaza.
Tratamentul cu UV – ADN din tuburile PCR contaminate este degradat cu formarea
dimerilor de timinã ( formã neamplificabilã ). Susceptibilitatea ADN de a fi distrus de
radiaţii depinde şi de: conţinutul în pirimidine ( în special timinã ), lungimea ţintei ADN
, timpul de iradiere, lungimea de undã, energia.
Tratamentul enzimatic – se realizeazã cu DNA-ze şi enzime de restrsicţie care au
situsuri de recunoaştere în ţinta ADN. Acest lucru are ca dezavantaj introducerea de noi
paşi în reacţia PCR, ceea ce coduce la risculo crescut de contaminare şi scãderea
eficienţei reacţiei PCR.
Uracil N-glicozilaza (UNG) - utilizatã pentru decontaminarea reacţiilor PCR
înaintea realizãrii ciclurilor termice. dUTP se îndepãrteazã prin digestia enzimaticã cu
uracil N-glicozilazã, ce se adaugã la mix ( fãrã Taq ). Atunci când reacţiile sut realizate
în prezenţa UNG, se eliminã eventualiicontaminanţi din reacţiile anterioare, ce conţin
uracil in moleculã. Metoda nu e utilizatã pentru toate polimerazele, deoarece nu toateîncorporeazã dUTP atât de eficient ca Taq polimeraza.
3.2. Probleme ce apar în reacţiile PCR
a. Nici un produs nu este identificat sau este slab amplificat
Cauze posibile:
- reactie dezechilibratã → se recomandã recalcularea concentraţiilor finale a
componentelor PCR
- ciclurile de amplificare→ se recomandã revederea programului PCR sau creşterea
numãrului de cicluri
- concentraţia micã de matriţã→ se recomandã creşterea concentraţiei şi verificarea
41

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 42/62
ei din punct de vedere calitativ
- concentraţia primerilor sau secvenţa lor → se recomandã optimizarea concentraţiei.
Verificarea secvenţei iar dacã e necesar reconstruirea lui
- mix de reacţie degradat sau incomplet → se reacomandã realizarea de controale
pozitive pentru verificarea diecãrui component
- temperatura de hibridizare prea mare → se recomandã reacalcularea Tm a
primerilor şi descreşterea temperaturii de hibridizare
- temperatura de denaturare subotimalã → se recomandã optimizarea temperaturii de
denaturare şi durata acestei etape
- timp de extensie suboptimal → se recomandã creşterea cu un minut a timpului de
extensie.
b. Trene ( ‘’ Smiruri ‘’)
Cauze posibile şi recomandãri :
- amplificãri nespecifice → se recomandã verificarea concentraţiilor şi a cilurilor de
amplificare, optimizarea magneziului şi a primerilor, descreşterea numãrului de
cicluri, verificarea matriţei, utilizarea procedeului hot startc.Produşi multimpli
Cauze posibile şi recomandãri :
- numãr mare de cicluri→ se recomandã scãderea numãrului de ciluri
- temperaturã de hibridizare prea joasã→ se recomandã creşterea temperaturii de
hibridizare
3.3. Specificitatea reacţiei
Condiţii ce favorizeazã creşterea specificitãţii : scãderea cantitãţii de Mg2+, dNTP,
Taq ADN polimeraza, primeri; scãderea numãrului de cicluri şi scurtarea duratei
42

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 43/62
etapelor ciclurilor; cresterea temperaturii de specificitate ; creşterea vitezei de rampã ;
utilizarea de hot-star PCR , nested-PCR şi touchdown PCR.
Specificitatea orocãrei reacţii PCR depinde de specificitatea de legare a primerilor la
matriţã, iar reacţiile se standardizeazã astefel încât sã se evite pãstrarea mixului mai
mult decât este necesar, pânã la introducerea în Temal Cycler.
3.4. Variante de PCR
3.4.1.Hot start PCR – este o metodã prin care se separã unul sau mai multe
componenete ale reacţiei, astfel încât toţi compuşii ce iau parte la reacţie sa ia
contact abia dupã denaturarea matriţei. Se realizeazã separarea fizicã a
componentelor, cu
ajutorul unui material ce funcţioneazã ca o barierã iniţialã, dar care se topeşte odata cu
creşterea temperaturii, ceea ce duce la amestecul tuturor componentelor reacţiei la
punctul START. Avantaj : se minimizeazã hibridizarea primerilor la ţinte nespecifice şi
se reduce obţinerea oligomerilor de primeri.
3.4.2. Touchdown PCR – temperatura de hibridizare este scãzutã progresiv în timpulciclurilor de reacţie, de la o valoare iniţialã mai mare decât a TM, pânã la o
valoare inferioarã acestuia. Acest lucru încurajeazã hibridizarea optimã a
primerilor la secvenţa ţintã, iar ampliconii urmãriţi se vor acumula înaintea
altor produşi nespecifici.
3.4.3. Invers PCR – utilizat pentru amplificareaunor segmente de ADN cu secvenţã
necunoscutã ( ex: situsurile în care se inserã traspozomii sau genomul unor
virusuri) . Secvenţa de ADN cunoscutã ( încadratã de secvenţe necunoscute )
se selecţioneazã cu endonucleaze. Molecula se circularizeazã şi se introduce în
PCR cu primeri complementari secvenţei cunoscute. Se obţine secvenţa
necunoscutã.
43

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 44/62
3.4.4. Multiplex PCR- în reacţie se introduce mai mult de o pereche de primeri, cu
scopul de a detecta mai multe ţinte
3.4.5. Real time PCR – sinteza ADN este detectatã pe mãsurã ce se produce. Cea mai
simplã metodã foloseşte un colorant care , prin legare la ADN dublu catenar,
emite fluorenşcenţã proporţionalã cu cantitatea de PCR acumulat. Avantaje :
nu necesitã metode de detecţie suplimentarã, aceasta având loc direct în tubul
de reacţie; se reduc şansele de contaminare ; se reduce durata experimentului.
3.4.6. RT-PCR – permite amplificarea ARN cu o etapã suplimentarã de revers
transcriere de la ARN la ADN complementar ( ADNc). Pentru sinteza primei
catene de
3.4.7. ADNc se pot folosi: primeri ,, random’’ , un primer complementar cu secvenţa
ARN , primeri oligo(dT) care hibridizeazã pe secvenţa poli (A) de la capãtul
3’. Polimerizarea se realizeazã cu ajutorul unei reverstranscriptaze : rTth –
ADN polimerazã ambivalentã ; AMV ( Avian Myeloblastosis Virus ) – are
activitate endonucleazicã şi se
3.4.8. utilizeazã în scopul obţinerii de fragmente mici de ADNc ; MMLV ( MaloneyMurine Leukemia Virus ) - este lipsitã de activitate endonucleazicã şi se
utilizeazã pentru sinteza moleculelor mari de ADNc. Reacţia are loc la
temperatura ce poate fi ajustatã la o valoare între 37 oC şi 55 oC ( în funcţie de
revers-trabscriptaza utilizatã ), în timp de 30 minute şi este urmatã de o reacţie
obişnuitã de PCR. Aplicaţii : se analizeazã ARN mesager şi virusuri ARN.
4. Aplicaţii ale tehnologiei PCR
Multe tehnici biochimice şi genetice se bazeazã pe PCR, pentru cã de cele mai multe
ori probele de ADN de analizat sunt în cantitãţi insuficiente. Aplicaţiile se încadreazã în
douã categorii :
44

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 45/62
4.1. preparative –sintezã de ADN ( RT-PCR, clonare, sinteza şi marcarea sondelor )
4.2. analitice – diagnosticul unor infecţii umane sau a unor boli monogenice ; studii
privind mecanisme genetice ce acompaniazã procese maligne sau chiar
diagnosticul cancerului; biologia populaţiilor; studii de taxonomie, sistematicã
şi filogenie molecularã; aplicatii în medicina legalã; amplificarea moleculelor de
ADN preistoric.
4. Restricţia enzimaticӑ
Enzimele de restricţie , cunoscute şi sub numele de endonucleaze de restricţie, sunt
responsabile de fenomenul restricţie-modificare. Sistemele de restricţie şi modificare
( metilare) au fost descoperite în urma studiilor interactiilor bacteriofag-bacterie, în care
s-a constatat cã o tulpinã bacterianã prezintã metilaze si endonucleaze care au aceeaşi
specificitate de secvenţã ( metilaza va adãuga o grupare metil la aceeaşi secvenţã care
este recunoscutã de enzimele de restricţie , astfelîncât în urm metilãrii un situs ţintã
devine rezistent la restricţie ) ( T.Vass, 2001 ). Metilarea reprezintã un mecanism de
apãrare a celulei baxteriene faţã de propriile endonucleaze.
Endonucleazele de restricţie sunt enzime care recunosc secvenţe de ADN specifice,
scurte, legarea fiind urmatã de clivare, fie la situsul de recunoaştere, fie la o distanţã
oarecare de acesta, în funcţie de tipul de enzimã.
45

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 46/62
PARTEA EXPERIMENTALA
46

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 47/62
Materiale necesare
- amestec dNTP, 10 mM fiecare- Taq ADN polimeraza 5U/μl
- Tampon pentru PCR 10X
- primer fimH1 20pmoli/μl
- primer fimH2 20pmoli/μl
47

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 48/62
- apã ultrapurã sterilã
- lizat termic ( ADN)
- vârfuri albe şi galbene cu/fãrã filtru pentru pipete ( sterile)
- tuburi Eppendorf de 0,2 , 1,5ml ( sterile)
- gheaţã şi cutii pentru gheaţã
- stative pentru tuburi Eppendorf
- hârtie prosop
Aparaturã:
- pipete automate P10, P20, P200 ( dedicate celor douã arii de lucru)
- aparat termocycler cu capac incãlzit
- microcentrifugã
- vortex
- hota cu flux laminar
- frigider
- congelator
Mod de lucru :
A. Prepararea master mixului:Se lucreazã în spaţiu special ADN free, cu echipamente dedicate camerei
respective
Se schimbã vârfurile de pipetã dupã adãugarea fiecãrui reactiv
1. se decongeleazã urmãtorii reactivi, pãstrându-i în gheaţã:
- tamponul pentru PCR
- amestecul de dNTP
- perechea de primeri fimH1/fimH2
2. se pregãteşte pe un stativ 1 tub Eppendorf de 1,5 ml şi 4 tuburi de 0,2 ml.; se noteazã
pe capac cu M
3. pentru a efectua în paralel cele n reacţii PCR, se preparã în tubul de 1,5ml
48

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 49/62
(notat M) un amestec ( master mix) adaugând volumele calculate de apã, tampon,
dNTP, primeri şi Taq ADN polimeraza. Volumele corespunzãtoare se calculeazã
pentru n+1 reacţii. Pentru o reacţie este nevoie de volumele indicate in tabelul de
mai jos:
Tabelul 1: Cantitãţi de reactanţi pentru PCR într-un volum final de 50 μl
Reactiv (concentraţie iniţialã) Volum ( μl) Concentraţie finalã
Apã ultrapurã 30,7 -
tampon pentru pcr 10x 5 1xamestec dNTP ( 10mM) 1 0,2 mM
primer fimH1 ( 20pmoli/μl) 1,5 30 pmoli
primer fimH2 ( 20pmoli/μl) 1,5 30 pmoli
Taq ADN polimeraza ( 5U/μl) 0,3 1,5 U
4. amestecul se vortexeazã scurt, apoi se centrifugheazã scurt5. folosind un singur vârf de pipetã se distribuie câte 40μl din acest amestec în cele
4 tuburi de 0,2.
B. Adãugarea ADN
Se transferã tuburile în spaţiul seprat de cel în care s-a preparat master mixul.
1. Folosind pipeta dedicatã acestei arii şi vârfuri de pipetã diferite se adaugã în
Tuburi.
2. Se centrifugheazã scurt toate tuburile şi se pun în gheaţã pânã când se
introduc în maşina PCR
C. Amplificarea
49

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 50/62
1. Se porneşte maşina de PCR. Se verificã programul de amplificare:
Programul de amplificare:
1 ciclu: 94oC - minute
25 cicluri: 94oC- 30 secunde
60oC- 30secunde
72oC- 1 minut
2. Dupã ce se verificã ca sunt bine închise, tuburile se pun în maşina şi se porneşte
programul de amplificare .
3. Dupã terminarea programului, tuburile se scot pe gheaţã şi se pãstreazã la +4oC
pânã la verificarea produşilor de amplificare.
1. PCR pentru enterovirusuri non-polio
Descriere: Metoda RT-PCR ( Reverstranscription-Polymerization Chain Reaction)
Constã în conversia ARN în ADN complementar ( ADNc) în prezenţa unei
reverstranscriptaze şi amplificare enzimaticã în regiunea genomicã VP1-2A.
Probe: Suspensie materii fecale, LCr ( lichid cefalorahidian), alte produse
patologice specifice, supernatant culturã celularã)
Condiţii de transport: + 4oC, condiţii de securitate conform normelor în viguare.
Se lucreazã la cãteva ore de la recoltare sau dupã stocare la -20oC.
Controlul reacţiei PCR: se face introducând în sistem doi martori : un martor
negativ şi un martor pozitiv. Ca martor negativ se introduce apa în loc de ARN. Scopul
acestui martor este de a detecta contaminãrile din timpul lucrului sau potenţialelecontaminãri ale reactivilor. Martorul pozitiv conţine ARN extras din enterovirusuri; prin
compararea probelor cu martorul pozitiv se confirmã sau infirmã prezenţa virusului în
proba analizatã.
50

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 51/62
2. Extractie ARN viral ( protocol adaptat dupa kit-ul de extracţie Qiagen)
Se lucreazã la hota cu flux laminar destinatã acizilor nucleici pentru a evita .
contaminarea probei şi a pentru a proteja operatorul de o potenţialã infectare cu agenţii
patogeni din probã.
Principiul metodei:
Extracţia ARN are la bazã o serie de proceduri care vizeazã dezorganizarea
virionului şi separarea materialului genetic de celelalte componente ale virionului.
Etapele extracţiei:
a. Dezorganizarea structurii virionilor
În acest scop se utilizeazã tamponul de lizã AVL. Acest tampon conţine agenţi
caotropici care denatureazã proteinele virale de înveliş şi deorganizeazã capsida.
Deasemenea tamponul conţine un inhibitor de ribonucleazã ( previne degradarea
enzimaticã a ARN) şi un carrier care leagã ARN.
b. Precipitarea ARN
Se realizeazã cu etanol 96%
c. Fixarea ARN pe coloana de separare
Fixarea se realizeazã prin intermediul carrier-ului conţinut în AVL
d. spãlarea coloanei
Are ca scop îndepãrtarea celorlalte componenete din amestec provenite din
materialul biologic. Se efectueazã cu tampoanele AWI şi AW2
e. Eluarea ARN
Eluarea ARN este etapa în care ARN este desprins din coloanã şi reluat sub
formã purã în tamponul AVE.
Materiale necesare:
51

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 52/62
- kit(Qiagen) destinat exracţiei ARN ( QlAamp viral RNA mini kit cat nr. 52906)
- vârfuri: 200-1000 μl
- tuburi Eppendorf: 1,5 ml
- mãnuşi de unicã folosinţã
- dezinfectant
- stativ pentru tuburi Eppendorf
- pipete automate
- frizetã
- vas pentru dezinfectat
Aparatura :
- hotã cu flux laminar
- vortex
- minicentrifugã
Mod de lucru:
a. se pipeteazã 560μl tampon AVL într-un tub Eppendorf 1,5 ml
b. se pipeteazã în tub 140μl supernatant culturã celularã şi se vortexeazã
c. se incubeazã la temperatura camerei timp de 10 minuted. se pipeteazã 560μl etanol 96-100% şi se vortexeazã timp de 15 secunde
e. din tub se reiau 630μl amestec şi se introduc în coloana de extracţie ( care a
fost fixatã intr-un tub de colectare de 2 ml); se centrifugheazã la 6000g ( 8000rpm). Se
fixeazã coloana intr0un tub nou de colectare
f. se repeta punctul e
g. coloana se spalã cu 500 μl tampon AW2 şi se centrifugheazã la 20.000g
( 14.000rpm) timp de trei minute şi se reia intr-un tub de colecţie nou şi se
centrifugheazã 1 minut la 20.000rpm , pentru eliminarea completa a tamponului AW20
52

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 53/62
h. coloana este trasferatã intr-un tub steril de 1,5 ml şi arn este eluat cu 50 μl
tampon AVE. Se incubeazã la temperatura camerei timp de un minut. Se centrifigheazã
la 6000g
3. Reverstranscriere pentru originea genomica VP1-2A
Principiul metodei:
Reverstranscrierea reprezintã transcrierea unei matriţe de ARN sub forma ADN.
Procesul se întâlneşte în natura la retrovirusuri şi stã la baza tehnicii RT-PCR.
Pentru reverstranscriere se folosesc enzime ( reverstranscriptaze) produse de
microorganisme recombinate ( în genomul microorganismului a fost introdusã gena
viralã care codificã pentru reverstranscriptazã).
Reverstranscrierea necesitã: o reverstranscriptazã ( exemplu: M-MLV: Moloney-
Murine Leukemia Virus Reverse Transcriptase) şi un primer ( primer =amorsa,
oligodeoxinucleoid; fragment sintetic de ADN de la care începe reverstranscrierea).
In tehnica RT-PCR, reverstranscrierea se face în douã etape succesive:
• RT-1: în acestã etapã primer-ul se leagã la matriţa ARN prin complementaritate.Procesul se numeşte,, anneling’’ ( hibridizare)
• RT-2: reverstranscriptaza sintetizeazã o catenã de ARN complementarã matriţei
ARN. Procesul se numeşte ,, elongation’’ ( elongare)
Materiale necesare:
- inhibitor de RN-aza( Promega RNAsin 40U/μl); 20 U per probã ( 0,5 μl)- primeri EUC2a, EUC2b; 20 pmoli per probã din fiecare primer ( 2μl)
- dNTP mix ( dGTP, dCTP, dATP, dTTP) ; 10 milimoli per probã ( 1μl mix)
- reverstranscriptaza ( M-MLV Promega) ; 200 U per probã ( 1μl)
- tampon reverstranscriptaza – concentrat 5X ( 4μl)
53

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 54/62
- apã distilatã sterilã
- vârfuri cu filtru 0,5-10μl
- vârfuri cu filtru 2-20μl
- vârfuri cu filtru 20-200 μl
- tuburi Eppendorf 0,2 ml
- tuburi Eppendorf 0,5 ml
- mãnuşi de unicã utilizare
- stativ pentru Eppendorf
- frizetã
- pahar Berzelius pentru deşeuri
- dezinfectant
- gheaţã
Aparatura:
-hotã cu flux laminar, vortex, minicentrifugã de masã, maşinã PCR, pipetã automatã,
maşinã de gheatã, congelator, frigider.
4. Reacţia PCR pentru regiunea genomicã VP1-2A Principiul metodei:
Reacţia PCR constã in replicarea unei matriţe ADN cu ajutorul ADN polimerazelor
izolate din microorganisme termofile este necesarã datoritã faptului cã reacţia PCR
implicã utilizare unor valori înalte de temperaturi care ar denatura polimerazele
obişnuite.
Etapele reacţiei:
• Denaturarea
În acestã etapã se realizeazã desfacerea duplexului ADN ( trecerea din forma
bicatenarã în forma monocatenarã ) . Procesul se realizeazã la temperaturi
apropiate de valoarea 90oC.
54

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 55/62
• Alinierea primerilor ( hibridizare)
Taq polimeraza necesitã fragmente scurte de ADN ( oligonucleotide ).
complementare cu ADN ţintã, de la care sã se inceapã sinteza. Alinierea primeri-
lor la catena matriţã se face prin complementaritate. Hibridizarea are loc latemperaturi in jurul valorii de 50oC.
• Extensia
Extensia reprezintã polimerizarea deoxinucleotideloraflate în amestecul de reacţie
prin acţiunea ADN polimerazei. Pentru sinteza catenei noi, polimeraza ,, aplicã’’
principiul complemetaritãţii utilizând catena matriţã. Extensia are loc la
aproximativ 70oC.
Materiale necesare:
- apa distilatã sterilã
- tampon Taq polimeraza – concentrat 10X; 5 μl /probã
- dNTP mix ( dGTP, dCTP, dATP, dTTP ) ; 10 mM per reacţie pentru fiecare
deoxinucleotidã ( 1μl )
- Taq polimeraza ( Q-BIOgene ); 1,25 U/ probã
- Primeri: EUG2, EUG3a, EUG3b, EUG3c ; 5 picomoli/probã din fiecare primer
( 0,5 μl)
- MgCl2 25 mM ; 2 mM/ probã ( 4 μl)
- Vârfuri cu filtru
5. Electroforeza în gel de agarozã
Principiul metodei:
Electroforeza reprezintã mişcarea dirijatã a moleculelor încãrcate electric sub
acţiunea unui câmp electric continuu.
55

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 56/62
Direcţia de deplasare a moleculelor este disctatã de sarcina electricã totalã a acestora;
moleculele cu sarcina electricã pozitivã vor migra cãtre catod ( polul negativ), iar
moleculele cu sarcinã electricã negativã totalã vor migra spre anod ( polul pozitiv).
Viteza de migrare a particulelor este determinatã atât de sarcina electricã cãt si de
masa lor molecularã, moleculele cu masa molecularã mai micã vor migra mai repede
decât moleculele care au masa molecularã mai mare dar sunt încârcate cu aceeaşi
sarcinã electricã.
Acizii nucleici ( ADN şi ARN ) sunt încãrcaţi cu sarcini electrice negative datoritã
grupelor fosfat ionizate, în consecinţã ei vor migra de la polul negativ catre polul
pozitiv. Mediul de migrare este asigurat de gelul de electroforezã; gelul este separat prin
hidratarea unui polimer natural extras din alge marine ( agarozã). Agaroza este un
polimer cu masã molecularã mare, cu lanţuri liniare şi ramificate, alcãtuite din retsuri de
glucozã şi derivaţi ai acestuia.
În uram hidratãrii agarozei, rezultã un gel care prezintã o microretea tridimensionalã
prin porii cãreia se deplaseazã acizii nucleici sub acţiunea curentului electric cintinuu.
Dimenisunea porilor este direct proporţionalã cu concentraţia gelului de agarozã;
creştera concentraţiei de agarozã favorizeazã deplasarea fragmentelor mai mici de acizinucleici şi reţinerea pentru un timp mai îndelungat a fragmentelor mari; acest lucru duce
la o mai bunã separare a fragmentelor de acizi nucleici.
Tamponul de migrare are rolul de a menţine sarcina electricã negativã a ADN
( datoritã pH-ului alcalin) dar şi de a închide circuitul electric.
In funcţie de mãrimea ampliconului se stabileşte concentraţia gelului de agarozã.
Concentraţia gelului este invers proporţionalã cu dimensiunea ampliconului.
Materiale necesare electroforezei :
- agarozã ( polimer natural extras din alege)
- tampon TAE ( tris-etilaminãhidroximetan, acid acetic, EDTA ) 1 X
56

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 57/62
- bromurã de etidiu soluţie stoc 10 mg/ml
- spatulã
- vas Erlenmeyer
- cilindru gradat
- piepten
- bandã adezivã sau dispozitiv pentru fixarea tãviţei
Aparatura: pipete automate, balanţa de laborator, sistem de electroforezã, tãviţã
specialã pentru formarea godeurilor, cuptor cu microunde
Mod de lucru:
• se fizeazã cu bandã adezivã capetele libere ale tãviţei
• se cântãreşte agaroza în funcţie de concentraţia doritã ( 0,8-2%) . Pentru
migrarea ampliconilor obţinuţi prin tehnica prezentatã, este necesarã o
cencentraţie de agarozã de 1,5 %
• se mãsoarã volum de tampon necesar preparãrii gelului
• se încãlzeşte amestecul de agarozã –TAE în cuptorul cu microunde pânã la
obţinerea unei soluţii transparente
• se rãceşte treptat soluţia de agarozã ( soluţia trebuie sã fie la aproximativ 600C)
• se adaugã bromurã de etidiu ( 1,25 la 25 ml soluţie agarozã ) , se omogenizeazã
Si se toarnã în tãviţa în care s-a fixat pieptenul
• gelul se solidificã la temperatura camerei
Incãrcarea ampliconilor în gel şi migrarea
Materiale necesare:
- gel pregãtit anterior
- tanc de electroforezã
57

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 58/62
- sursã de curent continuu
- pipete
- vârfuri sterile fãrã filtru
- marker de masã molecularã ( Promega , 100 pb )
- colorant albastru de bromfenol
- parafilm
Mod de lucru:
• se aşeazã gelul în tancul de electroforezã şi se acoperã cu TAE 1X
• pe parafilm se repatizeazã un numãr de ,,n’’ picãturi albastru de
abromfenol; n fiind numarul de ampliconi plus markerul de greutate
molecularã
• în fiecare picãturã de colorant se adaugã 5 μl amplicon şi se omogenizeazã
cu pipeta prin aspirare
• se încarcã în godeuri ( se noteazã ordinea încãrcãrii probelor)
• se seteazã parametrii de migrare
• gelul se vizualizeazã la lumina UV şi se forografiazã
Profil electroforectic al regiunii VP1-2A
MK 278 279 280 292 295 370 371
58
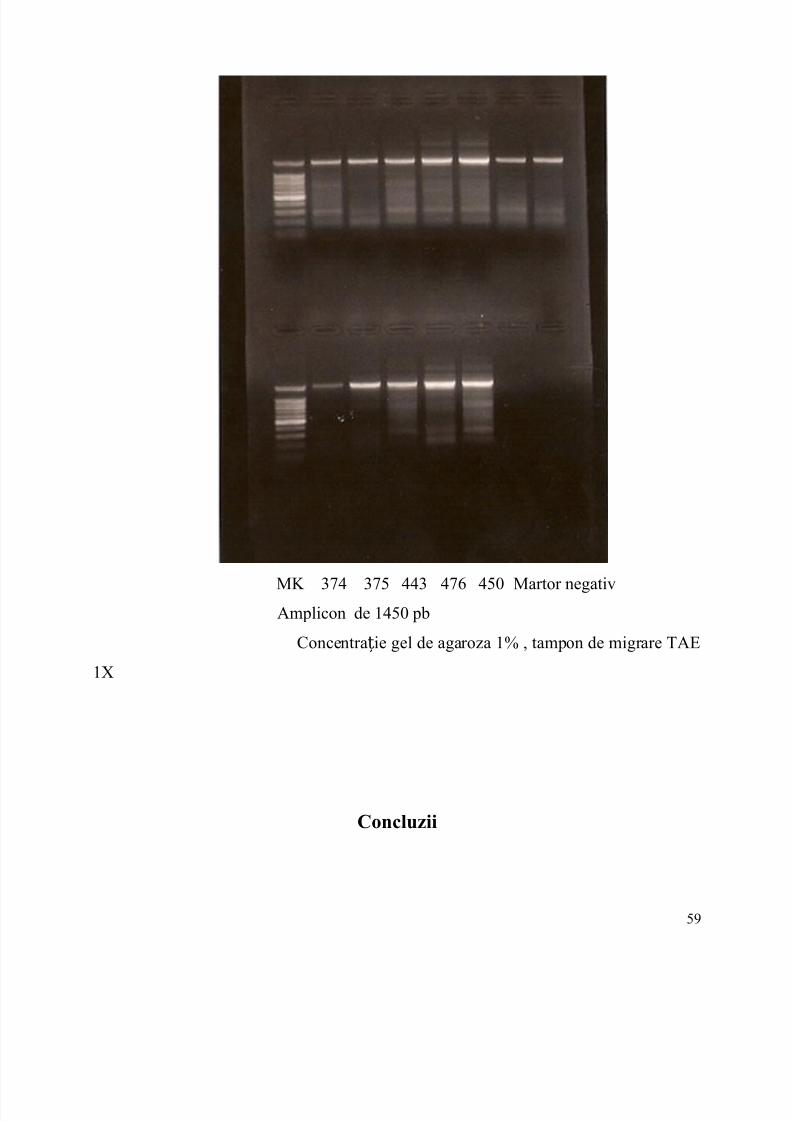
7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 59/62
MK 374 375 443 476 450 Martor negativ
Amplicon de 1450 pb
Concentra ie gel de agaroza 1% , tampon de migrare TAEț
1X
Concluzii
59

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 60/62
Scopul acestei lucr ri a fost eviden ierea eficien ei sistemului PCR-RFLP, n tipareaӑ ț ț ȋ
celor mai r sp ndite genotipuri la noi n tar . Probele au fost furnizate de Insitutul deӑ ȃ ȋ ӑ
Boli Infec ioase ,, Matei Bal ’’ au fost supuse restric ei enzimatice. Majoritateaț ș ț
probelor aveau profile electroforetice u or de interpretat prin PCR-RFLP.ș
Epidemiologia moleculară permite recunoaşterea tulpinilor diferite din aceeaşi
specie, provenind din circumstanţe epidemiologice cariate, pe baza polimorfismului
genetic. Dintre mecanismele care determină polimorfismul genetic, mai importante sunt
variabilitatea naturală determinată de polimeraze (în special ARN polimeraze) şi
modificările genetice (deleţii, inserţii, recombinări, trasnlocaţii). Epidemiologia
virusurilor s-a imbogăţit cu o serie de tehnici moleculare noi care permit caracterizarea
diferenţelor genetice şi antigenice între diferite izolate. Aceste tehnici recente, fac
posibile investigaţii privind originea şi evoluţia tulpinilor, avănd astfel implicaţii în
prevenirea anumitor boli enterovirale cu importanţă deosebită pentru sănătatea publică.
Epidemiologia studiat prin tehnici de biologie molecular permite ameliorareaӑ ӑ
metodelor de diagnostic şi terapeutice precum şi identificarea unor markeri genomici
(de exemplu secvenţele din genom care pot fi asociate cu o creştere a virulenţei sau cele
implicate în modificările către fenotipuri patogene).
BIBLIOGRAFIE
60

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 61/62
1. Colllier L. , Balows A., Sussman. M‘’ Microbiology and microbial
infections”
Edit.Topley and Wilson’s, 1998
2. Fields B., Knipe M.D, Howlley P. ‘’ Virology’’ fourth edition, Lippincott
Williams and Wilkins vol.1, ( 2001)
3. Oprisan G. Teza de doctorat : ‘’ Markeri moleculari utilizabili n ncadrareaȋ ȋ
taxonomic i caracterizarea enterovirusurilor ‘’ , 2002ӑ ș
4. Rolfs A., Schuller I., Finckh U., Weber-Rolfs I., : ‘’ PCR Clinical diagnostics and
reseaarch ‘’ Edit. Springer Laboratory, 1992
5.Rebedea I ,, Boli infec ioase’’ edit Medical , Bucure ti, 2000ț ӑ ș
6.SarnbrookJ., Fritsch E.F, Maniatis T.‘’ Molecular cloning-a Laboratory manual ‘’
CSH ( 2000)
7.Vassu T., Stoica L., Csutak O.,Mu at Fș . ‘’ Genetica microorgansimelor i inginerieș
genetic microbian ‘’ , Edit. Petrion, Bucure ti 2001ӑ ӑ ș
8.Blomqvist S., Bruu A-L.,Stenvik M.,Hovi T. ‘’Characterization of a recombinant
type 3/type 2 poliovirus isolated from a healthy vaccinee and containing chimeric
capsid protein VP1‘’ Journal of General Virology 2003,vol.84,573-5809.Brown B.,Oberste S.,Maher K.,Pallansch M. ‘’ Complete genomic sequencing
Shows that polioviruses and members of human enterovirus species C are closely
Related in the noncapsid codin region‘’Journal of Virology,vol.77,2003,8973-8984
10.Dove A.W Racaniello V.R. ‘’The Polio Eradication Effort:Should Vaccine
Eradication Be Next? ‘’ Science,1997,vol.277,779-780
11.Guillot Sophie. ‘’ Application a l’etude de la variabilite’ genetique des suoshes du
VPO.Digestion enzimatique‘’ ,2004
12.Hull h. F.,Aylward R. ‘’ Invited commentary:The Scientific Basis for Stopping
Polio Imunization‘’ American Journal of Epidemiology,1999,vol.150,1022-1025
13.Lukashev N. A ‘’ Role of recombination in evolution of enteroviruses‘’ Reviews
61

7/28/2019 Tehnici de Biologie Moleculara in Analiza Si Epidemiologia Enterovirusurilor
http://slidepdf.com/reader/full/tehnici-de-biologie-moleculara-in-analiza-si-epidemiologia-enterovirusurilor 62/62
In Medical Virology,2004
14.Masatoshi N.,Sudhir K. ‘’ Molecular evolution and phylogenetics‘’ Edit.Oxford
University ress,2000,187-207
15.Sloan K.,Eustace B.,Steward J.,Zehetmeier C.,Torella C., Simeone H., Roy J.,
Unger C., Louis D., Ilag L., Jay D. ‘’ CD155/PVR plays a key role in cell motility
During tumor cell invasion and migration‘’ BMC Cancer,2004,vol.4,73


















