
Sintesis de Carbohidratos
53
UNIVERSIDAD NACIONAL PEDRO RUIZ GALLO Facultad de Ingeniería Química e Industrias Alimentarias Escuela Profesional de Ingeniería de Industrias Alimentarias BIOENERGÉTICA DE LOS CARBOHIDRATOS CURSO: BIOQUÍMICA GENERAL INTEGRANTES: AYASTA GONZALES ERNESTO M. BERMEO PEREZ WILY CERVANTES CASAS CRISTIAN FARCEQUE CORDOVA DANTE CARRIÓN FARROÑAN JUAN JOSE VEGA ZULOETA BORIS
Transcript of Sintesis de Carbohidratos

UNIVERSIDAD NACIONAL PEDRO RUIZ GALLO
Facultad de Ingeniería Química e Industrias Alimentarias
Escuela Profesional de Ingeniería de Industrias
AlimentariasBIOENERGÉTICA DE LOS
CARBOHIDRATOS CURSO: BIOQUÍMICA
GENERAL INTEGRANTES:
AYASTA GONZALES ERNESTO M.
BERMEO PEREZ WILY
CERVANTES CASAS CRISTIAN
FARCEQUE CORDOVA DANTE
CARRIÓN FARROÑAN JUAN JOSE
VEGA ZULOETA BORIS

LA OXIDACIÓN EN LOS SISTEMAS BIOLÓGICOS
CADENA RESPIRATORIA
O
TRANSPORTE ELECTRONICO
y
FOSFORILACION OXIDATIVA

DESDE EL PUNTO DE VISTA QUÍMICO
OXIDACIÓN
• Ganancia de oxígeno
• Pérdida de electrones
• Pérdida de hidrógeno
REDUCCIÓN
• Pérdida de oxígeno
• Ganancia de electrones
• Ganancia de hidrógeno (en compuestos orgánicos)
El uso principal del OXÍGENO es en la RESPIRACIÓN
Y ESTE ES EL PROCESO POR EL CUAL LAS CÉLULAS OBTIENEN ENERGÍA EN FORMA DE ATP
Este principio de OXIDO- REDUCCIÓN se aplica a los sistemas bioquímicos y es un concepto importante para la
comprensión de la naturaleza de las oxidaciones biológicas.

EN LOS SISTEMAS REDOX
LOS CAMBIOS DE ENERGÍA LIBRE
PUEDEN EXPRESARSE EN TÉRMINOS DEL
POTENCIAL DE
OXIDACIÓN – REDUCCIÓN
LAS ENZIMAS QUE INTERVIENEN EN LOS
PROCESOS REDOX
SE DENOMINAN
OXIDORREDUCTASAS

Oxidorreductasas
Catalizan reacciones de oxido- reducción
Ared + Box Aox + Bred
A : es el reductor o dador electrónico; en el cursode la reacción se oxida (pierde electrones)
B : es el oxidante o aceptor electrónico; en el curso
de la reacción se reduce (gana electrones)
En las reacciones redox, siempre tienen que estarpresentes a la vez el aceptor y el dador electrónico

Estas reacciones son fuertemente exergónicas, en las cuales para evitar una liberación brusca de
energía, no aprovechable por la célula, se libera en forma fraccionada.

DISTINTAS FORMAS EN QUE LA CELULA PUEDE TRANSFERIR ELECTRONES
1.- Transferencia de 1 e-: Fe +++ Fe++
2.- Transferencia de un átomo de hidrógeno:
(H+ + e-): AH2 + B A + BH2
3.- Transferencia de un ion Hidruro (:H-)
AH2 + NAD+ → A + NADH + H+
4.- Transferencia de e- desde un reductor orgánico al oxígeno:
R-CH3 + ½ O2 RCH2-OH

• Gran parte de los sustratos oxidados en el organismo sufren deshidrogenación.
• Las reacciones de deshidrogenación son catalizadas por las ENZIMAS DESHIDROGENASAS.
• En estas reacciones el hidrógeno es captado por una coenzima.
• Las coenzimas pueden ser:
- Nicotinamida (NAD o NADP) - Flavina (FAD).
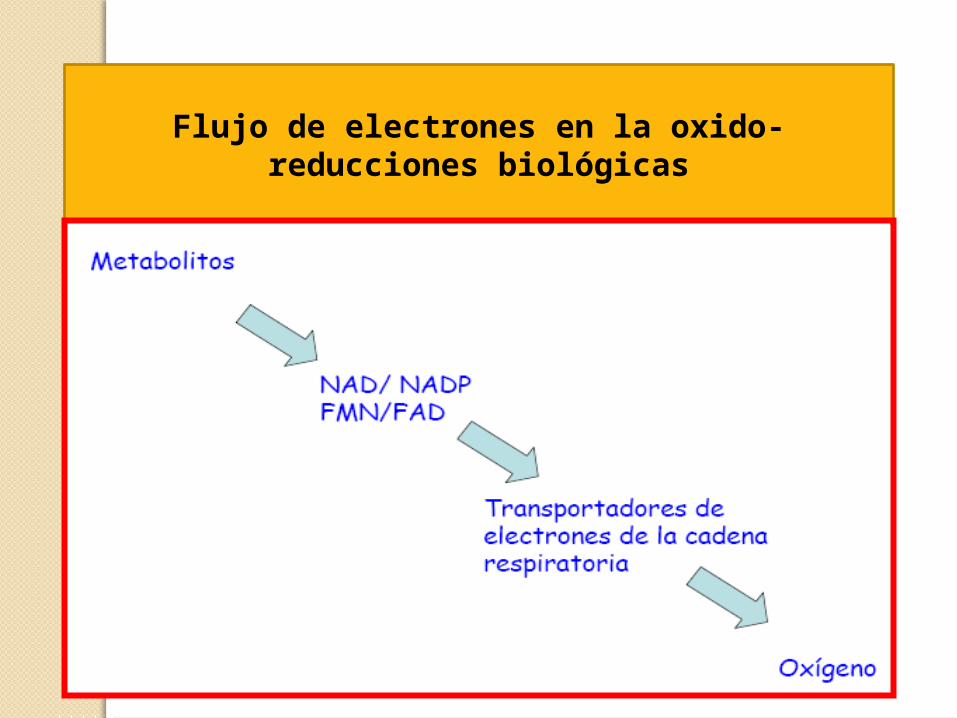
Flujo de electrones en la oxido-reducciones biológicas

Respiración Celular
Es el conjunto de reacciones en las cuales el ác. pirúvico producido por la glucólisis se desdobla a CO2 y H2O y se producen 30 ATP.
En las células eucariontes la respiración se realiza en la mitocondria.


LA MITOCONDRIA
FÁBRICA DE ENERGÍA CELULAR
ES EL SITIO DONDE TIENEN LUGAR
EL TRANSPORTE ELECTRÓNICO Y
LA FOSFORILACIÓN OXIDATIVA


Localización
30ATP
32ATP

La cadena transportadora de electrones
El NADH+H y el FADH2, obtenidos en el ciclo de Krebs, van a entrar en una cadena transportadora de electrones o cadena respiratoria, donde pasan los electrones, de una molécula reducida a otra oxidada, hasta el aceptor final que será el oxígeno molecular, que al reducirse formará agua.

CADENA DE TRANSPORTE
ELECTRONICO
• Los componentes de la cadena se encuentran en la membrana mitocondrial interna.
• Reciben equivalentes de reducción de NADH Y FADH2 producidos en la matriz.
• Los componentes actúan secuencialmente en orden creciente según sus potenciales de reducción.
• La energía que se libera durante la transferencia electrónica está acoplada a varios procesos endergónicos entre los que se destaca la síntesis de ATP.


Reacciones de la cadena de transporte de electrones
Con excepción de la coenzima Q, todos los miembros de esta cadena son
proteínas.
Pueden funcionar como enzimas como en el caso de varias deshidrogenasas.
Pueden contener hierro como parte de su centro hierro-azufre .
Los citocromos a y a3 contienen cobre.

Componentes de la cadena respiratoria
Transportadores de electrones-Coenzimas hidrosolubles:
NAD+ coenzimas de las deshidrogenasasNADP+
FMN se unen covalentemente a flavoproteínas FAD (grupo prostético), transportan 2 e- y 2 H+
- Quinonas: Coenzima Q – Ubiquinona, transportadores en medio no acuoso (membrana), transporta 1 e- y libera 2 H+ a la matriz
- Citocromos b, c, c1, a y a3 : proteínas con grupo prostético hemo, transportan 1 e-
- Proteínas ferro-sulfuradas: proteínas con Fe asociado a átomos de S, transfieren 1 e- por oxidación o reducción del Fe
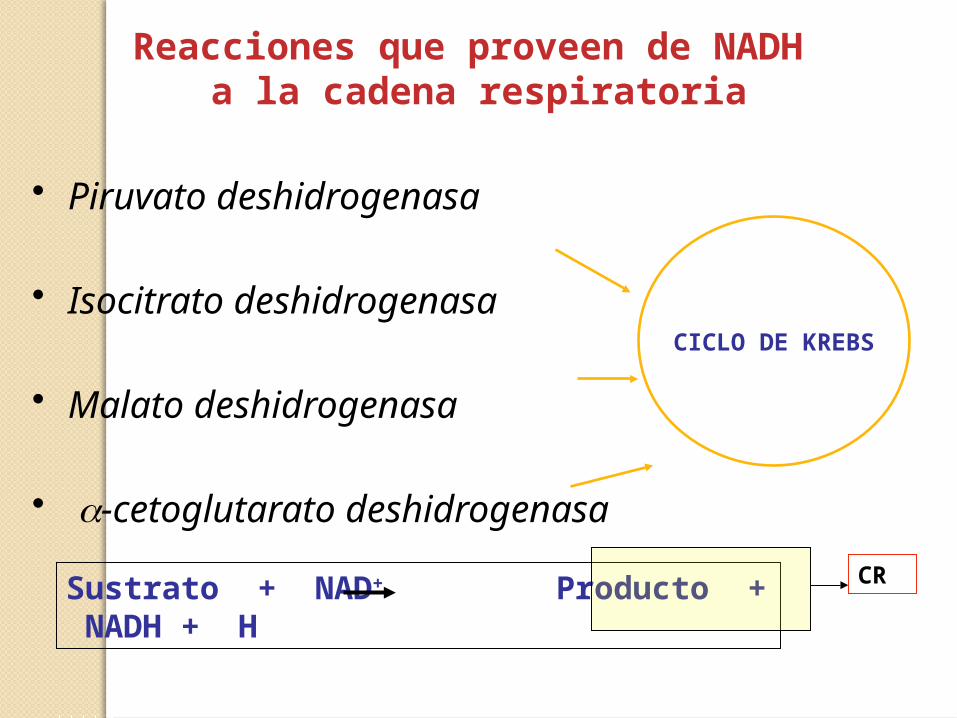
Reacciones que proveen de NADH a la cadena respiratoria
• Piruvato deshidrogenasa
• Isocitrato deshidrogenasa
• Malato deshidrogenasa
• a-cetoglutarato deshidrogenasa
CICLO DE KREBS
Sustrato + NAD+ Producto + NADH + H CR

Componentes de la Cadena de transporte electrónico
Complejo enzimático Grupos prostéticos
Complejo I (NADH deshidrogenasa) FMN, FeS
Complejo II ( succinato deshidrogenasa) FAD, FeS
Complejo III (citocromo bc1) Hemo, FeS
Citocromo c Hemo
Complejo IV (citocromo oxidasa) Hemo, Cu
Complejo V (ATP sintasa)

LA CADENA DE TRANSPORTE DE ELECTRONES COMPRENDE DOS PROCESOS:
1.- Los electrones son transportados a lo largo de la membrana, de un complejo de proteínas transportadoras a otro.
2. Los protones son translocados a través de la membrana, desde el interior o matriz hacia el espacio intermembrana de la mitocondria.
Esto constituye un gradiente de protones
El oxígeno es el aceptor terminal del electrón, combinándose con electrones e
iones H+ para producir agua.
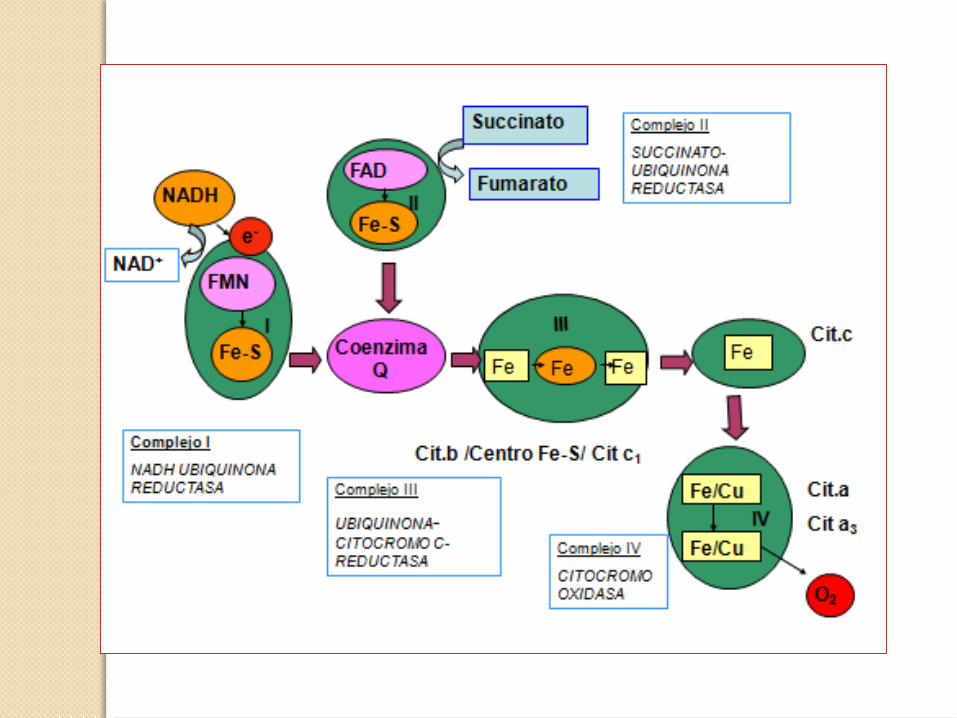

• La transferencia de electrones desde el NADH a través de la cadena respiratoria hasta el O2 es un proceso altamente exergónico.
• La mayor parte de esa energía se emplea para bombear protones fuera de la matriz.
• Por cada par de electrones transferidos al O2 los complejos I y III bombean 4 H+ y 2 el complejo IV.
• El complejo II no transfiere H+ ya que no atraviesa la membrana interna como los demás.
• Así esta energía electroquímica generada por el gradiente protónico impulsa la síntesis de ATP.

Flujo de electrones y protones a través de los cuatro complejos que forman la cadena respiratoria
EL OXÍGENO ES EL ACEPTOR TERMINAL DEL ELECTRÓN, COMBINÁNDOSE CON ELECTRONES E IONES H+ PARA
PRODUCIR AGUA.
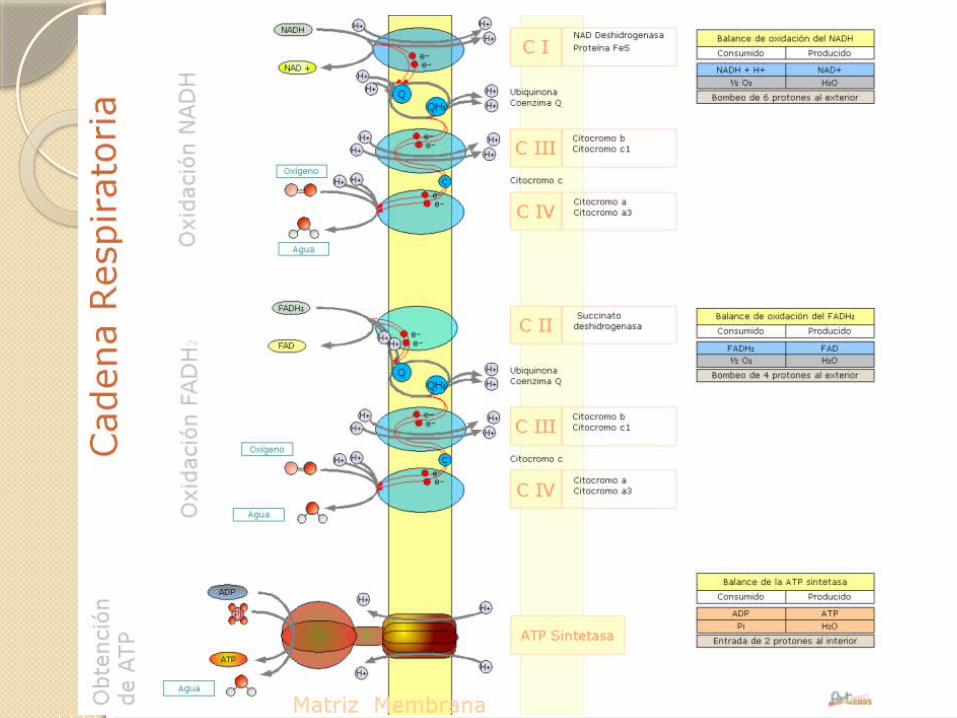

CADENA RESPIRATORIA
En la cadena respiratoria los transportadores electrónicos que se ven involucrados son:
Flavo proteínas, estas pueden captar 2H+ y 2e-.Citocromos, que pueden captar solo 1e-.Ubiquinona, esta puede captar 2H+ y 2e-.Proteínas Hierro-Azufre, que solo captan 1e-.
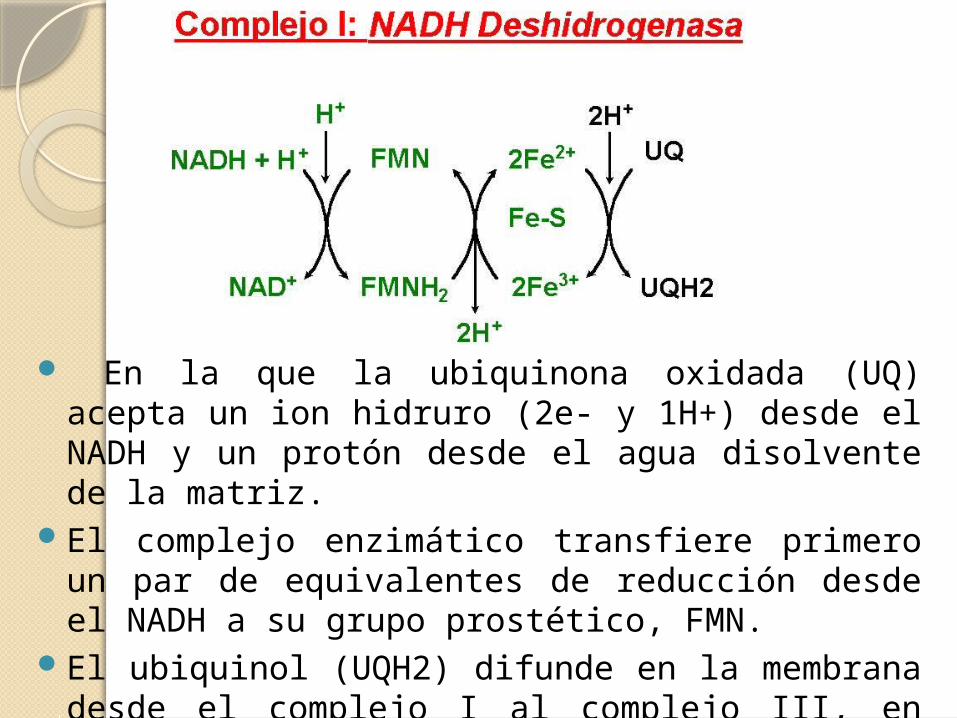
.
En la que la ubiquinona oxidada (UQ) acepta un ion hidruro (2e- y 1H+) desde el NADH y un protón desde el agua disolvente de la matriz.
El complejo enzimático transfiere primero un par de equivalentes de reducción desde el NADH a su grupo prostético, FMN.
El ubiquinol (UQH2) difunde en la membrana desde el complejo I al complejo III, en donde se oxida a UQ.

El primer paso en la ð-oxidación de los acil graso-CoA, por la flavoproteína acil-CoA deshidrogenasa, es la transferencia de electrones desde el sustrato al FAD de la deshidrogenasa, después a su flavoproteína transferidora de electrones (ETFP) que pasa sus electrones a la ETFP-ubiquinona deshidrogenasa. Esta reductasa, proteína ferro-sulfurada también tiene unido un nucleótido de flavina, pasa electrones a la cadena respiratoria al reducir la ubiquinona en la membrana mitocondrial interna.
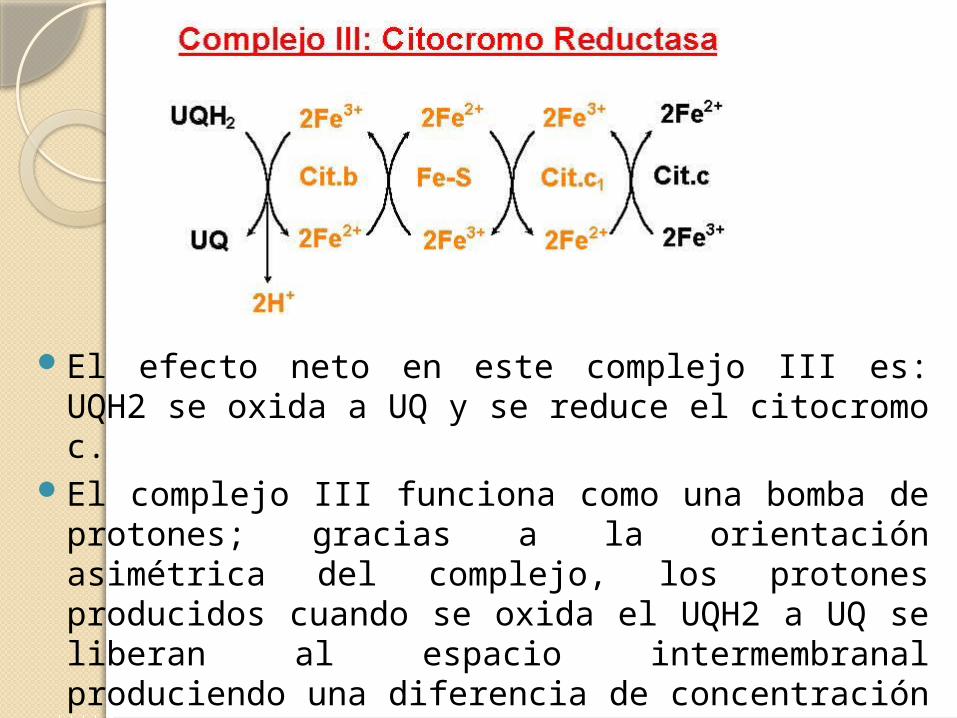
El efecto neto en este complejo III es: UQH2 se oxida a UQ y se reduce el citocromo c.
El complejo III funciona como una bomba de protones; gracias a la orientación asimétrica del complejo, los protones producidos cuando se oxida el UQH2 a UQ se liberan al espacio intermembranal produciendo una diferencia de concentración de protones transmembrana, un gradiente de protones.

COMPLEJO IV: REDUCCIÓN DE O2
La citocromo oxidasa también contiene dos iones cobre, CuA y CuB, que son de importancia crucial para la transferencia de electrones al O2.
El paso de los electrones a través del complejo IV: CuA (Cu2+) y cit. a (Fe2+) forman un centro redox bimetalito que puede aceptar dos electrones.
Aparentemente los electrones se trasladan primero desde el cit. c a CuA o al cit. a, que están en rápido equilibrio redox entre si.
Este centro bimetálico dona electrones a CuB y a cit. a3 también en equilibrio redox.
Estos, a su vez, ceden los electrones que reducen el O2 a H2O.

RESUMEN ( cadena respiratoria )
Los electrones llegan a UQ vía complejos I y II.UQH2 actúa como un transportador móvil de electrones
y protones; pasa los electrones al complejo III, el cual los pasa a otro eslabón móvil, el citocromo c.
El complejo IV transfiere electrones desde el citocromo c reducido al O2.
El flujo electrónico a través de los complejos I, III y IV va acompañado de flujo de protones desde la matriz al espacio intermembranal.
También se puede señalar que los electrones producidos en la ð-oxidación de ácidos grasos también se pueden incorporar en la cadena respiratoria a través de UQ.

INHIBICION DEL TRANSPORTE ELECTRÓNICO
• Inhibidores del transporte electrónico Inhiben solamente el transporte de e-
• Inhibidores de la fosforilación Inhiben la síntesis de ATP, indirectamente el transporte
de e-
• Desacoplantes Impiden la síntesis de ATP pero no inhiben el transporte
de electrones. Actúan como ionóforos eliminando el gradiente de protones.
• Inhibidores de la translocasa Inhiben la entrada de ADP y la salida de ATP desde la
mitocondria

REGULACION DE LACADENA
RESPIRATORIA

REGULACION: La degradación de los nutrientes y la síntesis
de ATP deben adaptarse de manera continua a los cambiantes requerimientos de energía dela cuerpo. La necesidad de coordinar el consumo y la producción de ATP se debe a que las cantidades totales de coenzimas son escasas en el organismo. El cuerpo humano forma alrededor de 65 Kg de ATP por día, pero en total contiene solo unos 3 a 4g de nucleótidos de adenina libres (AMP, ADP, ATP). Por tanto, cada molecula de ATP debe ser fosforilada hasta ADP y desfosforilada miles de veces por día.

LOS INHIBIDORES DE LA CADENA RESPIRATORIA IMPIDEN EL
PASAJE DE ELECTRONESLas moléculas que actúan como inhibidores impiden el flujo de e- entre los transportadores, y por lo tanto la síntesis de ATP.
Estas moléculas pueden actuar como venenos.
El Amital (un barbitúrico) o la Rotenona (un insecticida) bloquean el flujo de e- desde el NADH a la CoQ, mientras la Antimicina (un antibiótico) lo hace entre CoQ y Cit b, y el cianuro, la azida y el monóxido de carbono actúan sobre la citocromo oxidasa.
La aplicación de estos inhibidores bloquea el pasaje de e- por la cadena respiratoria. De esta forma “por detrás” del punto de inhibición los transportadores quedan reducidos.
Como consecuencia de esto no se produce ATP porque no se genera el gradiente protónico, ni agua porque los e- no llegan a reducir al O2: la cadena está interrumpida.

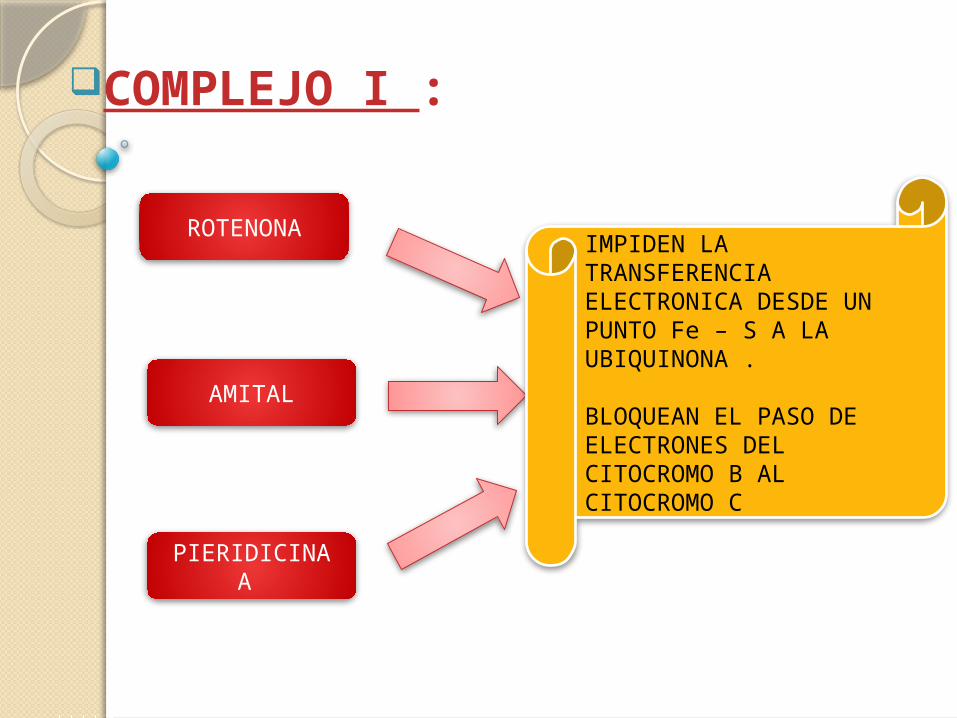
COMPLEJO I :
ROTENONA
AMITAL
PIERIDICINA A
IMPIDEN LA TRANSFERENCIA ELECTRONICA DESDE UN PUNTO Fe – S A LA UBIQUINONA .
BLOQUEAN EL PASO DE ELECTRONES DEL CITOCROMO B AL CITOCROMO C

COMPLEJO II :
MALONATO
INHIBIDOR COMPETITIVO DE LA
SUCCINATO DESHIDROGENASA

COMPLEJO III:
ANTIMICINA A BLOQUEA LA
TRANSFERENCIA ELECTRONICA
DESDE EL CITOCROMO b AL CITOCROMO c1.

COMPLEJO IV :
CIANURO
AZIDA DE SODIO
INHIBEN LA CITOCROMO
OXIDASA

ATP sintasa :
OLIGOMICINA
INHIBEN LA FO Y
CFO

LOS DESACOPLADOTES DE LA CADENA RESPIRATORIA NO IMPIDEN EL PASAJE DE ELECTRONES
El Dinitrofenol (DNP) y la termogenina, entre otras moléculas, pueden actuar como desacopladores, es decir desacoplan el transporte de e- de la síntesis de ATP.
Los desacopladores disminuyen la generación del gradiente de H+ porque permiten que los H+ difundan desde el espacio intermembrana hacia la matriz. La membrana mitocondrial interna se hace entonces permeable a los H+, lo que interfiere con la generación del gradiente.

En presencia de desacopladores se consume oxígeno, porque no está alterado el transporte de e- a través de la cadena, pero se forman menos ATP porque no se establece normalmente el gradiente protónico.
La termogenina es una proteína presente en la membrana mitocondrial interna del tejido adiposo pardo. A través de esta proteína pasan los H+, de manera que se genera un gradiente protónico mucho menor, y por lo tanto la cantidad de ATP generado también es menor. Como consecuencia de este desacople entre el transporte de e- y la fosforilación oxidativa, se libera energía como calor. El tejido adiposo pardo es abundante en animales que hibernan.

2 – 4 DINITROFENOL
TERMOGENINA

INHIBIDORES DE LA ATP- ASA
Hay moléculas que inhiben la síntesis de ATP, pero que no actúan sobre la cadena respiratoria. Por ejemplo la oligomicina, un antibiótico que interfiere en el transporte de protones. Esta molécula se une a la subunidad Fo de la ATPasa e impide de esta forma el pasaje de H+.


RENDIMIENTO DE ATP EN LA OXIDACION COMPLETA DE LA GLUCOSA

En este punto será de utilidad realizar una contabilidad para ver cuantas moléculas de ATP se producen por cada molécula de glucosa que se oxida.
Recordemos que parte del ATP se produce en la glucólisis, pero una cantidad mucho mayor de ATP es producida por el metabolismo aeróbico

En esta tabla resumiremos la producción de ATP y describiremos el reciclado de NADH y FADH2-

VIA NADH FADH
TRANSPORTADOR DE GLICEROL -
FOSFATO
TRANSPORTADOS DE MALATO - ASPARTATO
GLUCOLISIS: GLUCOSA A PIRUVATO (CITOSOL)
Glucosa de fosforilación -1 -1
Fosforilación de fructuosa-6-fosfato
-1 -1
Defosforilación de 2 moléculas de 1.3-BPG
+2 +2
Defosforilación de 2 moléculas de PEP
+2 +2
La oxidación de 2 moléculas de gliceraldehído-3-fosfato da 2 NADH
+2
CONVERSIÓN DE PIRUVATO A ACETIL-CoA (mitocondria)Produce 2 NADH +2
RENDIMIENTO DE ATP POR GLUCOSA

CICLO DEL ÁCIDO CÍTRICO (mitocondria)
2 moléculas de gtp a partir de 2 moléculas de succinil-CoA
+2 2
La oxidación de 2 moléculas de isocitrato de α-cetoglutarato y de malato, da 6 NADH
+6
La oxidación de 2 moléculas de succinato
de 2 FADH2-
+2

FOSFORILACION OXIDATIVA (mitocondria)
2 NADH de la glucólisis da 1.5 ATP cada uno si el NADH se oxida por el transportador glicerol-fosfato; 2.5 ATP por el transportador malato-aspartato.
+3 +5 -2
Decarboxilación oxidativa de 2 piruvato a 2 acetil-CoA:
2 NADH producen 2.5 ATP cada uno.
+5 +5 -2
2 FADH2 de cada ciclo del ácido cítrico producen 1.5 ATP cada uno.
+3 +3 -2
6 NADH del ciclo del ácido cítrico producen 2.5 ATP cada uno.
+15 +15 -6
RENDIMIENTO NETO +30 +32 0 0


















