
Simvastatin interferes with cancer ‘stem-cell’ plasticity ...
16
https://doi.org/10.1530/ERC-18-0132 http://erc.endocrinology-journals.org © 2018 Society for Endocrinology Printed in Great Britain Published by Bioscientifica Ltd. 25:10 Endocrine-Related Cancer S Kato et al. Statins reduce metastasis in ovarian cancer 821–836 RESEARCH Simvastatin interferes with cancer ‘stem-cell’ plasticity reducing metastasis in ovarian cancer S Kato 1 , M F Liberona 1 , J Cerda-Infante 2,3 , M Sánchez 2 , J Henríquez 2 , C Bizama 4 , M L Bravo 5,6 , P Gonzalez 5 , R Gejman 4 , J Brañes 1 , K García 1 , C Ibañez 2,6 , G I Owen 5,6 , J C Roa 4,6 , V Montecinos 2 and M A Cuello 1 1 Division of Obstetrics and Gynecology, Faculty of Medicine, Pontificia Universidad Católica de Chile (PUC), Santiago, Chile 2 Department of Hematology and Oncology, Faculty of Medicine, PUC, Santiago, Chile 3 Department of Cellular and Molecular, Faculty of Biological Sciences, PUC, Santiago, Chile 4 Department of Pathology, Faculty of Medicine, PUC, Santiago, Chile 5 Department of Physiological Sciences, Faculty of Biological Sciences, PUC, Santiago, Chile 6 Millennium Institute on Immunology and Immunotherapy, PUC, Santiago, Chile Correspondence should be addressed to M A Cuello: [email protected] Abstract Cell plasticity of ‘stem-like’ cancer-initiating cells (CICs) is a hallmark of cancer, allowing metastasis and cancer progression. Here, we studied whether simvastatin, a lipophilic statin, could impair the metastatic potential of CICs in high-grade serous ovarian cancer (HGS-ovC), the most lethal among the gynecologic malignancies. qPCR, immunoblotting and immunohistochemistry were used to assess simvastatin effects on proteins involved in stemness and epithelial-mesenchymal cell plasticity (EMT). Its effects on tumor growth and metastasis were evaluated using different models (e.g., spheroid formation and migration assays, matrigel invasion assays, 3D-mesomimetic models and cancer xenografts). We explored also the clinical benefit of statins by comparing survival outcomes among statin users vs non-users. Herein, we demonstrated that simvastatin modifies the stemness and EMT marker expression patterns (both in mRNA and protein levels) and severely impairs the spheroid assembly of CICs. Consequently, CICs become less metastatic in 3D-mesomimetic models and show fewer ascites/tumor burden in HGS-ovC xenografts. The principal mechanism behind statin-mediated effects involves the inactivation of the Hippo/YAP/RhoA pathway in a mevalonate synthesis-dependent manner. From a clinical perspective, statin users seem to experience better survival and quality of life when compared with non-users. Considering the high cost and the low response rates obtained with many of the current therapies, the use of orally or intraperitoneally administered simvastatin offers a cost/effective and safe alternative to treat and potentially prevent recurrent HGS-ovCs. Introduction High-grade serous ovarian cancer (HGS-ovC) remains the deadliest gynecological cancer worldwide. Despite achieving complete response, most patients will recur and die from their disease. Unfortunately, less than 40% of patients will exceed 10-year survival (Baldwin et al. 2012). Such a poor outcome is mainly explained by the late diagnosis when the disease has already disseminated throughout the whole abdominal cavity. Accordingly, most patients will first present with ascites and carcinomatosis. For surgeons, reducing the disease to its Endocrine-Related Cancer (2018) 25, 821–836 Key Words f ovarian cancer f metabolism f stemness f therapy f statins Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PM via free access
Transcript of Simvastatin interferes with cancer ‘stem-cell’ plasticity ...

https://doi.org/10.1530/ERC-18-0132http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.
25:10Endocrine-Related Cancer
S Kato et al. Statins reduce metastasis in ovarian cancer
821–836
-18-0132
RESEARCH
Simvastatin interferes with cancer ‘stem-cell’ plasticity reducing metastasis in ovarian cancer
S Kato1, M F Liberona1, J Cerda-Infante2,3, M Sánchez2, J Henríquez2, C Bizama4, M L Bravo5,6, P Gonzalez5, R Gejman4, J Brañes1, K García1, C Ibañez2,6, G I Owen5,6, J C Roa4,6, V Montecinos2 and M A Cuello1
1Division of Obstetrics and Gynecology, Faculty of Medicine, Pontificia Universidad Católica de Chile (PUC), Santiago, Chile2Department of Hematology and Oncology, Faculty of Medicine, PUC, Santiago, Chile3Department of Cellular and Molecular, Faculty of Biological Sciences, PUC, Santiago, Chile4Department of Pathology, Faculty of Medicine, PUC, Santiago, Chile5Department of Physiological Sciences, Faculty of Biological Sciences, PUC, Santiago, Chile6Millennium Institute on Immunology and Immunotherapy, PUC, Santiago, Chile
Correspondence should be addressed to M A Cuello: [email protected]
Abstract
Cell plasticity of ‘stem-like’ cancer-initiating cells (CICs) is a hallmark of cancer, allowing
metastasis and cancer progression. Here, we studied whether simvastatin, a lipophilic
statin, could impair the metastatic potential of CICs in high-grade serous ovarian cancer
(HGS-ovC), the most lethal among the gynecologic malignancies. qPCR, immunoblotting
and immunohistochemistry were used to assess simvastatin effects on proteins involved
in stemness and epithelial-mesenchymal cell plasticity (EMT). Its effects on tumor
growth and metastasis were evaluated using different models (e.g., spheroid formation
and migration assays, matrigel invasion assays, 3D-mesomimetic models and cancer
xenografts). We explored also the clinical benefit of statins by comparing survival
outcomes among statin users vs non-users. Herein, we demonstrated that simvastatin
modifies the stemness and EMT marker expression patterns (both in mRNA and protein
levels) and severely impairs the spheroid assembly of CICs. Consequently, CICs become
less metastatic in 3D-mesomimetic models and show fewer ascites/tumor burden in
HGS-ovC xenografts. The principal mechanism behind statin-mediated effects involves
the inactivation of the Hippo/YAP/RhoA pathway in a mevalonate synthesis-dependent
manner. From a clinical perspective, statin users seem to experience better survival
and quality of life when compared with non-users. Considering the high cost and the
low response rates obtained with many of the current therapies, the use of orally or
intraperitoneally administered simvastatin offers a cost/effective and safe alternative to
treat and potentially prevent recurrent HGS-ovCs.
Introduction
High-grade serous ovarian cancer (HGS-ovC) remains the deadliest gynecological cancer worldwide. Despite achieving complete response, most patients will recur and die from their disease. Unfortunately, less than 40% of patients will exceed 10-year survival (Baldwin et al. 2012).
Such a poor outcome is mainly explained by the late diagnosis when the disease has already disseminated throughout the whole abdominal cavity. Accordingly, most patients will first present with ascites and carcinomatosis. For surgeons, reducing the disease to its
Endocrine-Related Cancer (2018) 25, 821–836
10
Key Words
f ovarian cancer
f metabolism
f stemness
f therapy
f statins
25
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
822S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
microscopic burden is considered a success. However, such a load still presents numerous single or aggregated cancer cells. By then, some cancer cells have already adapted, established cooperative metabolic cell relationships and exhibit properties that confer a survival advantage (Blatter & Rottenberg 2015).
A key cellular phenotype during the metastatic wave, responsible for drug resistance and later recurrence, is the so-called cancer-initiating cell (CIC) subpopulation (Blatter & Rottenberg 2015). These cells exhibit stem-cell-like properties and have experienced epithelial-mesenchymal transition or epithelial-mesenchymal cell plasticity (EMT). They grow slowly, are more resistant to apoptotic and cytotoxic stimuli and can reformulate a complete tumor in xenograft models (Yan et al. 2014). These cells are part of floating aggregates present in ascites (Yeung et al. 2015). Cell aggregates offer enhanced survival while in transit to their metastatic niche, better resistance to environmental menaces and increased attachment and faster formation of metastatic foci compared to single floating cells (Yeung et al. 2015). Interestingly, in vitro, the expression of CIC stemness and EMT markers tend to increase when CICs are exposed to chemotherapy pulses (Chisholm et al. 2015). They also tend to adopt a spheroid conformation and enhanced resistance, mimicking the aggregates observed in ascites (Shield et al. 2009). At the time of recurrence, many cancer cells present the CIC phenotype because of the chemotherapy-selecting effect. This phenomenon explains the lower response rates (less than 30%) to most of the consecutive treatments (Fuh et al. 2015).
Changes in TP53 function are critical events for HGS-ovC progression (Iwanicki et al. 2016). In fact, TP53 is mutated (mut-TP53) in nearly all HGS-ovCs (96%), predominantly by missense mutations (Network CGAR 2011). Besides its classical tumor-suppressive functions, TP53 regulates metabolic pathways. Additionally, new evidence suggests that mut-TP53 is associated with cancer metastasis (Hu et al. 2013). Along with changes in TP53 functions, HGS-ovCs seem to acquire a mevalonate pathway gene signature that prompts tumor growth and metastasis (Hu et al. 2013, Greenaway et al. 2016). This signature is a consequence of metabolic re-programming, an evolutionary adaptation to satisfy the demand of lipid arising from fast dividing and moving HGS-ovC cells (Baenke et al. 2013).
The mevalonate pathway facilitates multiple metabolic cell functions (Kobayashi et al. 2015). Intermediate products of this pathway include sterol isoprenoids such as farnesyl pyrophosphate (FPP) and geranylgeranyl
pyrophosphate (GPP), responsible for prenylation, membrane localization and activation of different small GTPases that regulate important cell functions, and many are also well-established oncogenes (Cheng et al. 2010). The rate-limiting enzyme in the mevalonate pathway is the hydroxymethyl-glutaryl coenzyme A reductase (HMGCR) that converts HMG-CoA into mevalonic acid (Kato et al. 2010). Interestingly, some mut-TP53 forms interact with SREBP transcription factors and upregulate mevalonate genes, including HMGCR (Freed-Pastor et al. 2012). Consistently, we showed that HGS-ovCs exhibit higher HMGCR levels compared to benign tissues (Kato et al. 2010).
Simvastatin is a lipophilic statin that inhibits HMGCR activity, thereby preventing mevalonate and cholesterol synthesis and blocking protein prenylation by downstream depletion of FPP and GGP (Berndt et al. 2011). Treating mut-TP53 HGS-ovC cell lines with increasing concentrations of statins induces autophagy, cell death and drug sensitization in a mevalonate-dependent manner (Kato et al. 2010). Supporting a potential therapeutic role of statins in ovarian cancer, two recent studies showed a decrease in hazards ratio of disease-specific death and longer survival among statin users compared to non-users (Khan et al. 2015, Couttenier et al. 2017).
Statin research in ovarian cancer has been limited to test its effects in vitro or in vivo models. To our knowledge, no publication addressed its effect on the CIC-derived HGS-ovC subpopulation responsible for metastasis and recurrence. Thus, we investigated the effects of simvastatin on this subpopulation using different models. Herein, we identified new mechanisms by which statins can reduce or prevent CIC-mediated progression and thus disease recurrence.
Materials and methods
Reagents
Simvastatin was from EMD Millipore Corp. Carboplatin, etoposide, docetaxel, cisplatin, leptin, mevalonate, and geranylgeranyl pyrophosphate (GGPP) were from Sigma-Aldrich, mafosfamide from Santa Cruz Biotechnology and paclitaxel from Kampar Laboratories (Santiago, Chile).
Cell lines and spheroids
Human ovarian cancer cell lines, HeyA8 (harboring TP53 P72R polymorphism, RRID: CVCL_8878), UCI101 (TP53 WT, high p-glycoprotein and high EGFR, RRID: CVCL_5380) and
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
823S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
SKOV3 (TP53 null, RRID: CVCL_0532) were maintained in RPMI 1640 medium. In particular, HeyA8 and UCI101 cell lines were used for most of the experiments based on their capabilities of giving origin to tumors resembling HGS-ovC behavior in mouse models (Buick et al. 1985, Fuchtner et al. 1993, Domcke et al. 2013, Mitra et al. 2015, Hernandez et al. 2016, Tanenbaum et al. 2017). Cell lines, once acquired, were periodically authenticated using the Genemarker 10 kit (Promega), routinely screened for mycoplasma infection by PCR and discard before achieving 30 passages. CIC-enriched spheroids were isolated as previously described (Kato et al. 2015). To grant purity and to confirm that effects were effectively on stem-like cells, we also established CIC-only derived spheroids by combining limit dilution (single cell starting point) and stem-selecting media. To characterize CICs and assess statin effects, we studied two of the most probable stem-like cell markers (CD44 and ALDH1) described for ovarian cancer (Klemba et al. 2018).
Tissue culture, patient data and systemic immune-inflammatory index
This study was reviewed and approved by the institutional review board of PUC. All patients enrolled in this study signed an informed consent before the beginning of this study. Ascites were collected, and clinical records from patients who signed informed consent (IRB-approved protocol) were reviewed. Ascites-derived cultures were established as previously described (Kato et al. 2010). Mesothelial cells (calretinin(+)/claudin-4(−)) were isolated by differential trypsinization. Systemic immune-inflammatory index (SII) was calculated at the time of recurrence as described elsewhere (Hu et al. 2014). Briefly, the formula to calculate the score was SII = platelet * neutrophil/lymphocyte.
Xenograft mouse model
Pathogen-free NOD-Cg-Prkdcscid-IL2rgtm1Wjl/Szj (NSG) mice were obtained from Jackson Laboratories. Six- to eight-week-old mice were used for experiments. All IRB-approved experiments were conducted under Conicyt-Chile and ARRIVE/AMVA guidelines. For xenograft implantation, non-selected or CIC-isolated HeyA8 cells were injected intraperitoneally (IP). Tumor formation was monitored daily by palpation and measurement of the abdominal diameter. After 7 days of tumor growth, we treated the mice (n = 4 per group, three consecutive cohorts) with placebo or simvastatin (2 mg/kg/day via IP) for 4 weeks.
Quantitative PCR
Total RNA was extracted using TRIzol (Life Technologies). cDNA was generated from 1 µg RNA using iScript Reverse Transcription Supermix for RT-qPCR (Bio-Rad). HMGCR, CD44, POU5F1, NANOG, SOX2, ZEB2, CDH2 and CDH1 mRNA levels were assessed by qPCR and normalized to that of the porphobilinogen deaminase gene. Primers used in this study are listed in Supplementary Table 1 (see section on supplementary data given at the end of this article). The qPCR reactions were performed using SYBR Green Master Mix (Applied Biosystems) in an ECO qPCR System (Illumina San Diego, CA, USA). Relative amounts of mRNAs were calculated using the 2−∆∆ct method.
Immunoblotting (WB) and RhoA pull-down assay
Protein pellets and electrophoresis were carried out as previously described (Kato et al. 2015). Overnight primary antibody exposure was performed by using antibodies against PARP and CD44, TWIST (Santa Cruz Biotechnology), ZEB2, SOX2, E-cadherin, TAZ, YAP, phospho-YAP (Cell Signaling), OCT4, NANOG, ALDH1A1, and N-cadherin (Abcam), ACTB (β-actin) (Sigma-Aldrich) or TUBB (β-tubulin) (Thermo-Fisher). Peroxidase-conjugated goat anti-mouse/rabbit/goat IgGs were used as secondary antibodies (Bio-Rad). The reaction was developed with chemiluminescence using Western Lightning ECL Pro (Perkin-Elmer) and the signal was detected by Imagequant LAS 500 system (GE Healthcare Bio-Science AB). Densitometry analysis was made using Adobe Photoshop Software Tools and protein levels were normalized against β-actin protein levels.
RhoA pull-down assays were performed using the Rho activation assay kit according to manufacturer’s instructions (Cell Biolabs, San Diego, CA, USA).
Immunocytochemistry and immunofluorescence
Immunocytochemistry (IC) was carried out as previously described (Kato et al. 2010). Primary antibodies, CD44 (Santa Cruz Biotechnology), ALDH1A1 (Abcam), YAP and phospho-YAP (Cell Signaling) and TP53 (Dako) were diluted following the manufacturer’s instructions. For three-dimensional IC (3D-IC), spheroids were collected and spread on Superfrost Plus microscope slides (Thermo-Scientific), fixed with methanol and treated as described for 2D-IC (P-YAP, CD44, ALDH1A1). The immunofluorescence (IF) protocol has been described elsewhere (Kato et al. 2015). Each picture consisted of z-series of 1024-pixel images. Merge analysis was performed using ImageJ.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
824S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
Immunohistochemistry
Paraffin tissue-sections (5 µm thickness) from xenografts and HGS-ovC specimens were prepared for immunohistochemistry (IHC) as described previously (Ohyagi-Hara et al. 2013). PAX8 (Roche), WT1 (Dako), TP53, KI67, CD44, ALDH1A1, YAP and p-YAP staining were quantified by using the Aperio Imaging Analysis Toolbox, under Aperio nuclear algorithm. Spheroids were embedded in HistoGel and included in paraffin blocks. Five micrometer paraffin sections were prepared to carry out the IHC for TP53. Nuclei staining was quantified using Adobe Photoshop counting tools and Aperio Imaging Analysis Toolbox, under Aperio nuclear algorithm. To detect the vasculature, a double staining was performed by using huCD34, huCD31 and msCd31 antibodies and the EnVision G/2 Doublestain System (Dako), following the manufacturer’s instructions. The quantification of the vascular area was performed by using image color deconvolution and particle analysis with ImageJ.
MTS-based cell proliferation assay
Cancer cells from primary cultures were exposed for 48 h to different chemotherapeutic reagents. Cell viability was assessed as described previously (Kato et al. 2010).
Matrigel invasion assay
Transwell inserts containing 8 μm isopore membranes (Nunc) were coated with Corning Matrigel Matrix Growth Factor Reduced (GFR) diluted 1:10, as described elsewhere (Kato et al. 2015). Equal numbers of CICs suspended in the stem-selecting medium were spread over and treated as indicated for 48 h. The number of invading cells was determined by IC for pan-cytokeratin (Sigma-Aldrich). To quantify the number of cells trans-passing the isopores, the inserts were examined under a microscope (100×, Nikon) and 15 fields were counted per experiment.
Spheroid migration assay
Flat bottom 96-well plates were coated with 0.3% Corning Matrigel Matrix GFR for 30 min at 37°C. Spheroids established from a single ‘stem-like’ cell (confirmed by stemness and EMT marker expression) were treated with simvastatin 1 µM for 24 h and then transferred to the coated flat bottom plates. Effects of simvastatin were measured at 24 h, and the images obtained by optical microscopy were analyzed by ImageJ. A migration score
was calculated individually for each sphere using the following formula = ((the area at 24 h − area at time 0)/area at time 0) × 100. The graphs presented reflect the average score of at least ten replicates for each condition in three different experiments.
3D organotypic mesomimetic assay
The assay was performed as described previously, with minimal modifications (Kenny et al. 2009). For invasion assays, six-well plates containing 8 µm pore size inserts were coated with a mix of Hs832 fibroblasts (ATCC), collagen I, and medium and incubated at 37°C for 4 h. Mesothelial cells isolated from primary cultures were cultured on top of the fibroblast layer and incubated at 37°C for 18 h. Spheroids were then seeded on top of the mesothelial layer. DMEM/F12 + 10% FBS was used as the chemoattractant stimulus.
Statistical analyses
JMP13 (SAS) was used for data analyses. Statistical significance (P values <0.05) was calculated by Student’s t-test, Chi-squared test or Mann–Whitney. Survival curves were generated using the Kaplan–Meier method and analyzed by Wilcoxon test. Cox proportional hazard regression model was carried out to assess the effect of different risk factors on survival.
Results
Cancer cell line-derived spheroids exhibit a morphology and stemness/EMT expression pattern resembling that in ascites aggregates from HGS-ovCs
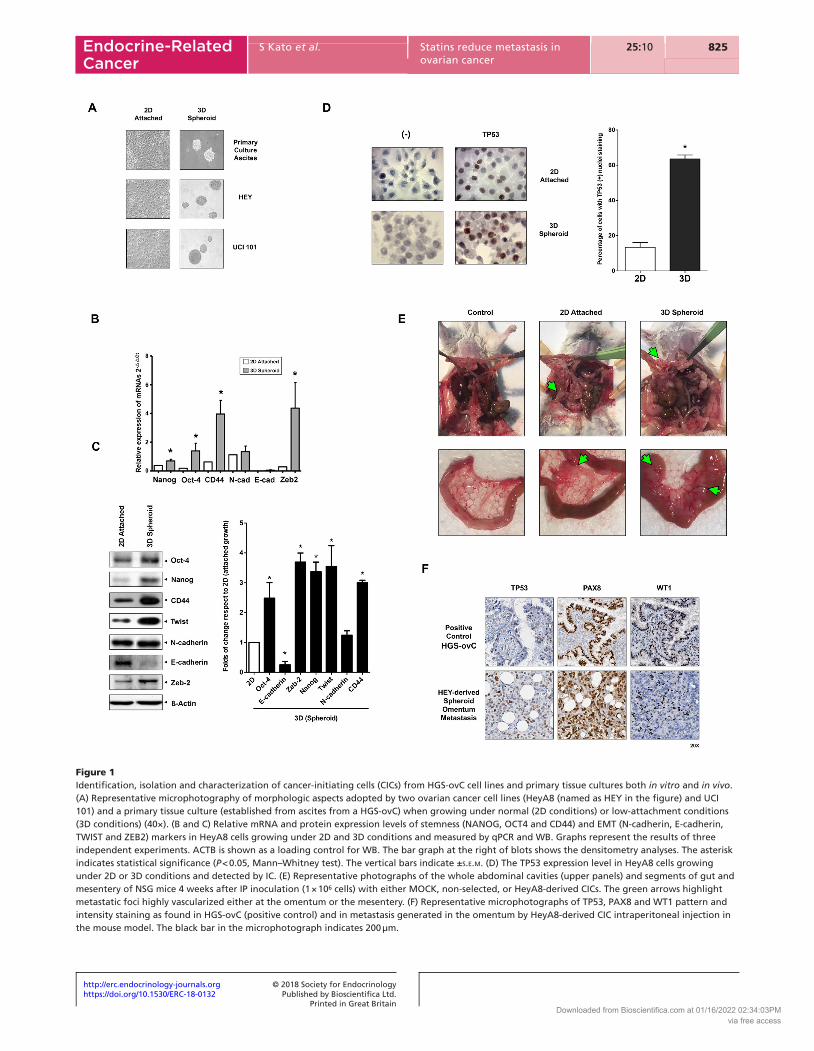
As observed in Fig. 1A, ovarian cancer aggregates collected from ascites from HGS-ovC cases exhibit a similar morphology compared with established ovarian cancer cell lines (HeyA8 and UCI 101) when cultured under normal (in 2D) or low-attachment conditions (in 3D). Under 2D conditions, all of them grew as attached cell groups until reaching full confluency. In contrast, under low-attachment, all formed 3D structures or spheroids similar in size, but different in cell density (as confirmed when visualized with magnification, identifying single cell limits and measuring cell membrane overlapping), mimicking the aggregates found in ascites.
Then, we tested whether spheroids from cancer cell lines represented an enriched CIC or
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
825S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
Figure 1Identification, isolation and characterization of cancer-initiating cells (CICs) from HGS-ovC cell lines and primary tissue cultures both in vitro and in vivo. (A) Representative microphotography of morphologic aspects adopted by two ovarian cancer cell lines (HeyA8 (named as HEY in the figure) and UCI 101) and a primary tissue culture (established from ascites from a HGS-ovC) when growing under normal (2D conditions) or low-attachment conditions (3D conditions) (40×). (B and C) Relative mRNA and protein expression levels of stemness (NANOG, OCT4 and CD44) and EMT (N-cadherin, E-cadherin, TWIST and ZEB2) markers in HeyA8 cells growing under 2D and 3D conditions and measured by qPCR and WB. Graphs represent the results of three independent experiments. ACTB is shown as a loading control for WB. The bar graph at the right of blots shows the densitometry analyses. The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). The vertical bars indicate ±s.e.m. (D) The TP53 expression level in HeyA8 cells growing under 2D or 3D conditions and detected by IC. (E) Representative photographs of the whole abdominal cavities (upper panels) and segments of gut and mesentery of NSG mice 4 weeks after IP inoculation (1 × 106 cells) with either MOCK, non-selected, or HeyA8-derived CICs. The green arrows highlight metastatic foci highly vascularized either at the omentum or the mesentery. (F) Representative microphotographs of TP53, PAX8 and WT1 pattern and intensity staining as found in HGS-ovC (positive control) and in metastasis generated in the omentum by HeyA8-derived CIC intraperitoneal injection in the mouse model. The black bar in the microphotograph indicates 200 µm.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
826S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
‘stem-like’ subpopulation. In Fig. 1B and C, we show the findings in one of the cell lines tested, the HeyA8 cell line. As shown there, cancer cells significantly modified their stemness (e.g., CD44) and EMT (e.g., TWIST) expression patterns when forming spheroids, as measured by qPCR and WB. This pattern resembled that described by others as characteristic of CICs isolated from HGS-ovCs (Shield et al. 2009). Additionally, we assessed TP53 expression. We hypothesized that spheroids also possessed a TP53(+) enriched population, favoring a pro-survival and metabolic adaptation within ascites (Hu et al. 2013). Accordingly, TP53(+) cells were significantly enriched in spheroids (Fig. 1D, left panel) as confirmed by quantification using Adobe Photoshop counting tools and the Aperio Imaging Analysis Toolbox, under Aperio nuclear algorithm (P < 0.008, Fig. 1D, right panel).
Cells forming spheroids generate tumor xenografts more rapidly
As shown in Fig. 1E, both non-selected (cells growing in 2D conditions) and the CIC-enriched population (3D-spheroids) generated xenografts upon IP injection into NSG mice. However, the number of cells required to generate a tumor and the growth curve slope were significantly different. A lower number of CICs was required to generate a tumor compared with non-selected cells (5 × 104 vs 1 × 106 cells). Once tumors were identifiable, the growth curve slope was significantly steeper for xenografts arising from CICs. Additionally, metastatic foci were more vascularized in tumors arising from CICs (as indicated by green arrows in Fig. 1E). Supporting the resemblance of the model with HGSOCs in humans, HeyA8-derived CICs, injected intraperitoneally, disseminated in the abdominal cavity in a similar fashion in the mouse model (e.g. omentum, peritoneum, diaphragm, mesentery root, so on) and expressed markers commonly used to confirm the serous histology and the tube-ovarian epithelial origin (e.g. PAX8 and WT1, Fig. 1F) (Liliac et al. 2013, Di Palma et al. 2014, Mitra et al. 2015, Xiang et al. 2018).
Simvastatin decreases CIC invasion and migration, induces spheroid disassembly and CIC cell death
Statins can induce cell death in vitro (Kato et al. 2010). Additionally, CICs from this malignancy are more invasive and resistant to commonly used chemotherapies (Kato et al. 2015). Therefore, we tested whether one lipophilic statin, simvastatin, used at different concentrations (including those clinically achievable),
affected ovarian cancer spheroids. Varying concentrations of simvastatin were added to the cultures either before or after spheroid formation in single or repeated pulses.
To mimic clinical settings, we used different administration schemes, always considering the simvastatin achievable plasma concentrations (Björkhem-Bergman et al. 2011, Lee et al. 2014). As shown in Fig. 2A and Supplementary Fig. 1A, a single dose of simvastatin (up to 5 µM) did not affect morphology or cell death in spheroids established from HeyA8 and UCI101 cells or primary cultures (the chemo-naïve patient UC172). Although not significant, the number of spheroids obtained from ascites tended to decrease upon simvastatin exposure and at higher concentrations. A significant decrease in the number of migrating or invading cells was detected upon a single pulse of simvastatin (1 µM) in matrigel invasion assays, spheroid migration assays and mesomimetic 3D cultures in both HeyA8 (Fig. 2B, C and D) and UCI101 cells (Supplementary Fig. 1B and C) either using isolated spheroids (CIC-enriched populations) or spheroids established from single stem-like cell (CIC-only derived spheroids). In contrast, upon repeated pulses, a significant spheroid disassembly was observed in a concentration-dependent manner in both cell lines and primary tissue cultures (Fig. 2E, F and Supplementary Fig. 1D.1). Meanwhile, cancer cell death significantly increased in a concentration-dependent manner as measured by detection of PARP cleavage in W-B (Fig. 2G and Supplementary Fig. 1D.2). We also assessed the effect of starting the pulses before spheroid formation. Regardless of the starting point, a decrease in spheroid cell density and a reduction in spheroid numbers were found (Supplementary Figs 1D.3, 2A and B).
Simvastatin impairs stem-cell plasticity by regulating the Hippo/YAP/TAZ pathway and the RhoA activity
Since repeated simvastatin pulses affected CICs, we assessed the underlying mechanisms. Our laboratory and others showed that statins reduce cancer cell migration and induce cell death by inhibiting the mevalonate pathway and prenylation of small GTPase family members (Horiuchi et al. 2008, Kato et al. 2010). TP53 mutation is involved in controlling EMT, stemness, and metabolic adaptations of cancer cells destined to form spheroids (Vousden & Ryan 2009, Freed-Pastor et al. 2012). Interestingly, TP53 mutations correlate with the acquisition of a mevalonate gene signature in HGS-ovC (Greenaway et al. 2016, Cuello et al. 2018). Thus, we
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
827S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
postulated that the mevalonate pathway might be critical for stemness, EMT and spheroid formation. Thus, we first assessed HMGCR mRNA levels in cancer cells growing under 2D or 3D conditions. As shown in Fig. 3A, HMGCR levels were higher in cells (HeyA8, UCI 101, SKOV3
(upper panel) and primary cultures (lower panel)) forming spheroids, supporting the mevalonate dependence. We then assessed the effect of simvastatin on the expression of EMT and stemness markers. As shown in Fig. 3B, C, and Supplementary Fig. 1E, a single simvastatin pulse
Figure 2Effects of single and repeated doses of simvastatin on CIC-derived spheroid assembly, spheroid count, invasion and cell death. (A) Upper panels, HPF microphotograph (40×, contrast microscopy) showing the effect of a single pulse of vehicle (DMSO) or simvastatin (5 µM for 24 h) on HeyA8-derived and chemo-naïve primary culture-derived (patient UC172) spheroid integrity. Middle panels M, immunoblotting for PARP levels, as a cell death marker, upon the single pulse of vehicle or simvastatin (1–5 µM for 24 h). Lower panels, the graph represents the results of three separate and consistent experiments using a single pulse of vehicle or simvastatin on the total spheroid count. The vertical bars indicate ± s.e.m.. (B) Left panel, the effect of single simvastatin or vehicle pulse on HEY-derived CIC migration measured by Spheroid Migration Assay. The graph summarizes the results of three separate experiments (vertical bars indicate ± s.e.m.). The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). Right panel, representative microphotographs of the spheroid appearance at 0 h and 24 h of incubation upon MOCK or simvastatin treatment. (C) Effect of single simvastatin or vehicle pulse on HEY-derived CIC invasion measured by Matrigel Assays. The graph in the left summarizes the results of three separated experiments (vertical bars indicate ± s.e.m.). The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). At the right, a representative microphotography of cells trespassing to the reverse of the insert. (D) Effects on spheroid adherence and invasion in 3D-mesomimetic inserts. Red flags point spheroids on the obverse of the insert. Green flags indicate CICs trans-passing to the obverse of the insert. (E) The effect of repeated simvastatin pulses (four pulses, added after spheroid formation) on the number of spheroids formed from HeyA8 cells and primary tissue cultured cells (chemo-naïve patient UC172) after a week of incubation. Graphs represent the results of three separate experiments using repeated simvastatin pulses on the total spheroid count. The vertical bars indicate ± s.e.m. The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). (F) HPF microphotograph (40×, contrast microscopy) showing the effect of repeated pulses (up to four) of simvastatin (2–5 µM for 24 h) on HeyA8-derived and chemo-naïve primary culture-derived (patient UC172) spheroid integrity. (G) Detection of PARP cleavage by WB upon repeated pulses of simvastatin in HeyA8-derived and chemo-naïve primary culture-derived (patient UC172) spheroids. ACTB is shown as loading control of respective blots.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
828S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
(1 µM for 24 h) induced significant changes in EMT (e.g., decrease in N-cadherin) and stemness (e.g., reduction in CD44) marker expression as measured by qPCR and WB in HeyA8 and UCI 101 cells. Different pathway interplay/crosstalks modulate EMT and stemness during the metastatic process, among them the Hippo/YAP pathway and the activity of small GTPases (Chen et al. 2015, Sebio & Lenz 2015, Vieira & Paredes 2015). Interestingly, CD44 levels are involved in this crosstalk and the expression of EMT genes controlled by these pathways (Zhang et al. 2014, Orian-Rousseau 2015). As shown in Fig. 3D, a single pulse of simvastatin significantly decreased CD44 levels in HeyA8 cells. This decrease correlated with the accumulation of the inactive/un-prenylated form of RHOA (confirmed by pull-down, Supplementary Fig. 2C). Both CD44 levels and RHOA activity were re-established
by adding either mevalonate or GGPP (Fig. 3D upper and lower panels, respectively). Since CD44 regulates the Hippo/YAP pathway via RHOA (Zhang et al. 2014), we also assessed the effect of simvastatin pulses in this pathway. As shown in Fig. 3E, F and Supplementary Fig. 1F, a single simvastatin pulse resulted in the accumulation of the phosphorylated/inactive cytoplasmic form of YAP both in cell lines and chemo-naïve primary cultures. YAP activity was recovered by adding either mevalonate or GGPP (Fig. 3G). Finally, we studied the effect of mevalonate supplementation in spheroid disassembly and cell death induced by repeated simvastatin pulses. As seen in Fig. 3H and Supplementary Fig. 1G, adding mevalonate results in the rescue of spheroids from disassembly and cell death (Fig. 3I and Supplementary Fig. 1H) induced by the repeated pulses of simvastatin.
Figure 3Simvastatin effects on stemness and EMT marker expression and the activity of RhoA, Hippo/YAP/TAZ in HGS-ovC CICs. (A) Relative mRNA expression levels of HMGCR in ovarian cancer cell lines (UCI101, SKOV3, and HeyA8) and chemo-naïve and recurrent HGS-ovC primary cultures growing under 2D- or 3D-conditions as measured by qPCR (the figure summarizes three different experiments and vertical bars indicate ±s.e.m.). The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). (B) Changes in relative mRNA levels of stemness markers (CD44, OCT4, NANOG and SOX2) induced by a single pulse of simvastatin (1 µM per 24 h) compared to control in HeyA8-derived CICs as measured by qPCR. The graph summarizes the results of three separate experiments (vertical bars indicate ±s.e.m.). The asterisk indicates statistical significance (P < 0.05, Mann-Whitney test). (C) Representative gels of changes in protein levels of stemness (CD44, OCT4, NANOG, SOX2 and ALDH1A1) and EMT (N-cadherin) markers induced by the simvastatin pulse compared to control in CICs as measured by WB. (D) Representative gels of effects of a single pulse of simvastatin in the absence or presence of mevalonate (100 µM, upper panels) GGPP (10 µM, lower panels) on CD44 and inactive form of RhoA protein levels in HeyA8-derived CICs as measured by WB. (E) Representative gels of effects of simvastatin pulse on phospho-YAP, YAP and TAZ protein levels in HeyA8- and chemo-naïve primary culture-derived CICs (patient UC172, left and right panels, respectively) as measured by WB. ACTB and TUBB are shown as loading controls. (F) Representative microphotography of the changes in YAP cellular localization (The red-fluorescence staining corresponds to phospho-YAP; nuclei are stained in blue) in chemoresistant primary culture-derived CICs upon a single pulse as detected by IF (patient UC02). (G) Representative gels of effects of mevalonate and GGPP supplementation in simvastatin-induced changes in phospho-YAP levels in HeyA8-derived CICs. β-Actin is shown as a loading control. (H) Representative microphotographs of mevalonate supplementation in simvastatin-induced disassembly of HeyA8-derived spheroids. (I) Representative gels of effects of mevalonate supplementation in simvastatin-induced cell death in HeyA8-derived CICs (measured by detection of PARP cleavage). ACTB is shown as a loading control.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
829S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
Simvastatin reduces tumourigenesis and metastasis in xenografts
The above-described results were then confirmed in vivo.A valuable finding to mention before proceeding to
describe the simvastatin effects in cancer xenografts, the aggregates identified in the ascites of the control mice expressed CD44 and ALDH1A1 (Supplementary Fig. 2D) as seen in aggregates isolated from ascites in HGS-ovC patients.
As shown in upper panel Fig. 4A, CICs widely spread into the abdomen, mimicking a massive peritoneal carcinomatosis in placebo mice. Interestingly, a reduction in the number and distribution of metastatic implants was observed after 3 weeks of IP administration of simvastatin compared to placebo (lower panel Fig. 4A). Additionally, no implant was identified on the diaphragm, liver or gastric surfaces in statin-treated mice. This reduction in tumor burden, upon statin treatment, was associated
Figure 4Effects of 4-week simvastatin treatment (2 mg/kg/day IP) on tumor growth, ascites, weight, metastasis distribution and expression of CD44, ALDH1A1 in HeyA8-derived CIC inoculated NSG mice. (A) Whole animal view (closed and opened abdomen) of a representative mouse for each condition (n = 3–4 per group, three experiments). The small circle under each right panel shows the amount of disease collected after debulking procedure. The green arrows highlight main disease foci. (B) Bar graphs showing the average effect of simvastatin on ascites volume (left panel) and body weight (right panel) compared with MOCK treatment. Vertical bars indicate ±s.e.m. The asterisk indicates statistical significance (P < 0.05, Mann–Whitney test). (C) Representative microphotographs of changes in the expression of CD44 (membrane), ALDH1A1 (cytoplasm), YAP (nucleus and cytoplasm) and p-YAP (cytoplasm) staining upon simvastatin treatment (panels in third and fourth rows) compared to MOCK treatment (panels in first and second rows) at the injection site (implantation site). The small black and blue frames included in the microphotographs (first and third rows, respectively) indicate the areas shown magnified under the corresponding one (second and fourth rows, respectively). The black bar in each microphotograph indicates 200 µm. The green arrows highlight the brown staining of the nucleus and reflect the nuclear localization of YAP.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
830S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
with a significant decrease in ascites volume and lower increase in body weight (Fig. 4B, left and right panels, respectively). Given that simvastatin significantly reduced ascites and the number of metastatic foci found in the treated xenografts, we decided then to measure by immunohistochemistry the expression of CD44, ALDH1A1, YAP and p-YAP at the site of peritoneal injection, the place from where carcinomatosis was originated. Additionally, we analyzed the expression of markers associated with the cell proliferation or tumor activity (KI67), the metabolic adaptation (TP53) and the neo-angiogenesis in the most common metastatic site, the omentum. As shown in the magnified lower blue frame panelslower blue frame panels of Fig. 4C, upon IP simvastatin treatment, a significant decrease in CD44 and ALDH1A1 staining and changes in localization and phosphorylation status of YAP were seen compared to untreated conditions (four magnified upper black frame panelsupp in Fig. 4C). KI67 and TP53 expression were
significantly reduced in simvastatin-treated mice (lower panels in Fig. 5A). A decrease in the recruited vasculature (mouse origin, msCD31) was detected in simvastatin-treated mice (lower panels in Fig. 5B). Finally, no CIC-derived neo-angiogenesis was observed in any conditions since no huCD34 or huCD31 staining was detectable using the double staining technique and species-specific antibodies.
Simvastatin use improves survival outcomes among patients with HGS-ovC and the quality of life in those recurrent cases
Patients with recurrent HGS-ovC who received any statins, during their disease, were identified from our clinical database and the disease evolution among statin user and non-users was evaluated. In total, we reviewed charts in 128 cases of stage III/IV HGS-ovC treated at our institution, 32 of them received statins.
Figure 5Effects of 4-week simvastatin treatment (2 mg/kg/day IP) on KI67 (cell proliferation), TP53 expression (a reflex of metabolic adaptation), and vascular recruitment at metastatic foci (omentum) in HeyA8-derived CIC inoculated NSG mice. (A) Representative microphotographs of omental metastasis as captured under LPF (10×, upper panels in each condition) and HPF (40×, respective underlying panels). Left upper panels, shown hematoxylin-eosin staining (H&E); middle and right upper panels show TP53 and KI67 staining in IHC. Immediately underlying panels show magnified fields of equivalent areas included in the upper panels and using the same staining technique. (B) Mouse CD31 staining of any vascular structure detected at the injection or metastasis site (panels at the left). No staining suggestive of human CIC-derived neo-angiogenesis was detected (using huCD31 or huCD34 antibody). The table, at the right, summarizes the quantification of the vascular area under each condition. The *, #, and ^ signs indicate statistical significance for specific comparisons (P < 0.05, t-test).
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
831S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
When comparing clinical variables related to adverse outcomes, no significant differences were found between groups, including age, stage, primary debulking surgery and optimal debulking rates and complete primary response rate upon completion of planned chemotherapy (more details on demographics and clinical variables are shown in Supplementary Table 2). Among this cohort, there were 81 recurrent cases, of which 15 patients received statins. As shown in Fig. 6A and B, statin users exhibited longer progression-free interval (median 26 vs 14 months), and overall survival (OS; median 93 vs 43 months) compared with non-users. Interestingly, the clinical picture of recurrent cases among statin users was characterized by the absence or lower volume of ascites and less tumor burden (confirmed by CT scan), similar to the phenomenon found in xenografts. Besides, statin users exhibited a significantly lower SII compared with non-users (Fig. 6C), a recently identified powerful prognostic indicator of poorer outcomes in patients with solid tumors (Zhong et al. 2017). In fact, we found out that none of statin users exhibited a SII higher than 1000 while 34.4% of non-users did it (P value <0.002). Finally, we confirmed that a SII lower than 1000 (the group in which all statin users were included) constituted an independent variable associated with better OS in our cohort of recurrent cases (Fig. 6D).
In this cohort, we identified a case of recurrent HGS-ovC (stage IIIC, optimally debulked) and heavily treated with chemotherapy. This patient had been diagnosed with hypercholesterolemia and treated intermittently in previous years. At her last visit, she was in progression (after a third relapse and
failing to respond to consecutive rescue schemes), in palliative care and was using alternative medicine. Simultaneously, she underwent paracentesis, twice a week, to alleviate symptoms from recurrent massive ascites. Due to shortening in symptomatic relief after each paracentesis, we proposed a compassionate measure to try simvastatin (10 mg/day PO, as prescribed for hypercholesterolemia). We obtained informed consent to collect ascites samples before and after finishing a 4-week, close follow-up, single course. Supplementary Figure 3A shows the original histology and the pleomorphic aggregates observed in the ascites before treatment. Most of the cells forming aggregates were CD44+. To assess chemosensitivity, we carried out an in vitro MTS assay using different options. As shown in Supplementary Fig. 3B, the ascites-isolated cancer cells were mostly resistant to all chemotherapeutics, except taxanes (a drug previously discontinued for this patient due to neurotoxicity). Simultaneously, we exposed the aggregates to repeated pulses of simvastatin. As shown in Supplementary Fig. 3C, simvastatin reduced the number of spheroids but did not induce cell death in vitro. Encouragingly from a clinical perspective, a significant decrease and stabilization in CA125 levels were observed upon 4 weeks of treatment (Supplementary Fig. 3D, upper panel) that correlated with normalization of cholesterol levels (Supplementary Fig. 3D, lower panel).Importantly, an evident symptomatic relief and a decrease in the number of paracentesis were observed. Finally, we measured CD44, Oct-4 and Nanog levels in aggregates collected before and after treatment. As shown in Supplementary Fig. 3E, after 2 weeks of
Figure 6Clinical impact of using statins among patients with HGS-ovC. (A and B) Comparison of progression-free survival (PFS) and overall survival (OS) curves among statin users (green line, n = 28) and non-users (violet line, n = 91). (C) Calculated systemic immune-inflammatory index (SII) among statin users and non-users. Box, line, bars and the asterisk in the graph indicate 25–75 interquartile, median, standard deviation, and statistical significance, respectively. (D) Cox proportional-hazards regression model including significant variables known for influencing OS in HGS-ovC.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
832S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
treatment, a decrease in CD44, Oct-4 and Nanog levels was observed.
Discussion
A better understanding of ovarian cancer unveils key facts that will affect prognosis. First, optimal treatment, which implies a surgical reduction of the disease to its minimal expression (at the microscopic level) and hopefully the obliteration of the remaining disease with chemotherapy are needed (Bookman 2016). Secondly, the microenvironment should be targeted to prevent the conditions favoring the establishment of harmful and sustainable relationships between cancer cells and the surrounding non-cancerous cells, part of the microenvironment (Hansen et al. 2016). We recently showed that high leptin levels, observed among obese patients, contribute to a more favorable microenvironment for cancer cells to easily transit and metastasize (Kato et al. 2015). Even more recently, we have shown that HGS-ovCs exhibiting gene signatures related to abnormal lipid metabolism and obesity resulted in poorer outcomes, independently of the expression profile of well-known gene drivers for this disease (e.g., WT or mutant TP53) (Cuello et al. 2018). Under these conditions, cancer cells can easily aggregate, adhere to the mesothelium and remain dormant and protected within the metastatic foci (Liu et al. 2015). If such conditions remain unmodified, the recurrence will be inevitable. After establishing a metastatic niche, a dormant cancer cell can enter a period of proliferative growth, demonstrating all characteristics of cells resistant to chemotherapy (Blatter & Rottenberg 2015).
Herein, we demonstrate that at any time during disease progression, including after chemotherapy, a subset of cancer cells form spheroids or aggregates (Figs 1, 2 and 3) as a mechanism of self-defense, survival and resistance. To do so, cancer cells adopt a 3D conformation and exhibit several adaptive changes. Many cells within the spheroids experience a cadherin switch that allows them to aggregate and provides rescue from anoikis when floating in ascites (Vieira & Paredes 2015, Plutoni et al. 2016). They also accumulate TP53 (Yang-Hartwich et al. 2015) and express stemness markers such as CD44, which facilitate drug resistance and metastasis (Gao et al. 2015, Zhao et al. 2016). Also, they take advantage of other metabolic pathways (e.g., mevalonate pathway) to obtain energy and regulate gene transcription, particularly of genes involved in metastasis and drug resistance (Kobayashi et al. 2015, Sato et al. 2016).
In CICs collected from ascites of advanced HGS-ovCs, we demonstrated that the this cell subset grows forming spheroids or aggregates that express higher TP53 and HMGCR levels compared with the non-selected population. CICs are highly tumorigenic and more resistant to apoptotic-inducing agents, including statins, a consequence of intratumoral heterogeneity; a concept that has been confirmed through the exploitation of high-throughput genomic analysis of multiple biopsies from individual tumors or by isolation and analysis of single cells (Gerlinger et al. 2012). Thus, cancer cell heterogeneity within a tumor can manifest itself in two ways: (1) by major genetic events (e.g., somatic copy number aberrations, mutations) or (2) by phenotypic variations in transcript and protein expression levels and metabolic rewiring (Eason & Sadanandam 2016). Providing evidence for the second, we observed that the same cancer cells behave differentially when grown under 2D or 3D conditions. 3D spheroids not only contain a higher TP53 population or express higher HMGCR levels, but they also upregulate the expression of factors needed to form spheroids (e.g., CD44, ALDH1A1). CICs manifesting higher TP53, CD44 and HMGCR levels seem to arise from context-specific metabolic re-programming, a cellular adaptation to microenvironmental conditions. Confirming this re-programming hypothesis, Greenaway and colleagues recently showed in an orthotopic, syngeneic model of epithelial ovarian cancer that ascites-derived cancer cells upregulate the mevalonate pathway compared with cancer cells originally used for tumor induction (in the ovarian bursa). and this upregulation requires changes in TP53 function (Greenaway et al. 2016).
In recent years, worldwide opinion-leaders in ovarian cancer have reached a consensus regarding recommendations to improve current clinical outcomes (Vaughan et al. 2011), including the need to identify novel therapeutic strategies directly targeting CICs and the recruited microenvironment (a cytokine milieu in which CICs can thrive, becoming more resistant and aggressive). Additionally, a single targeted agent is not sufficient to inhibit adaptive tumor changes in cancer cells. Based on the tumor adaptability and the epigenetic influence of the tumor microenvironment, future treatment selection should consider cancer profiling of primary tumor, metastatic foci and patient-conditioned microenvironment (e.g., obese vs thin women). The same principle could be considered at the time of recurrence when cancer cells have already evolved to provide further resistance to therapies. We are currently far from identifying specific profiles in a cost/effective manner to
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
833S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
successfully offer more effective personalized treatments to patients with HGS-ovCs (Wallbillich et al. 2016). Meanwhile, educated choices and combining multi-targeted therapies with those that have already been proven effective (e.g., chemotherapy) seem to constitute the best option. In this current scenario, statins arise as an attractive alternative to prevent or treat recurrent disease.
Statins possess pleiotropic properties, mainly due to the inhibition of the mevalonate pathway and prenylation of small GTPases (e.g., RAS and RHO), rather than through reduction of cholesterol synthesis (Mullen et al. 2016). From an oncological perspective, lipophilic statins, such as simvastatin, seem to severely impair the metabolic adaptability of ovarian cancer cells to the microenvironmental conditions. We previously showed that statins induce autophagy, diminish migration and trigger cell death either alone or in combination with chemotherapy in a mevalonate-dependent manner (Kato et al. 2010). Herein, we demonstrate that simvastatin, at concentrations clinically achievable and used in repeated doses (daily administered IP), impairs spheroid formation and induces spheroid disassembly, major adaptive changes pertinent to metastasis and resistance. Simvastatin modifies and reverts stemness (e.g., CD44) and EMT. As an immediate consequence of simvastatin action, the migration, adherence and invasion of CICs decrease in mesomimetic 3D cultures. This phenomenon may be explained by the decrease in CD44 levels (Cieply et al. 2015). As the spheroid structure offers a conformational barricade, by hampering spheroid assembly, simvastatin also jeopardizes one of the major self-defense mechanisms against hostile environmental conditions. Thus, CICs may become more vulnerable and accessible to any pro-apoptotic agents. Encouragingly, almost all these effects are completely abrogated by mevalonate or GGPP supplementation in vitro, demonstrating the mechanism by which simvastatin may offer its beneficial effects to patients with HGS-ovC.
By inhibiting the mevalonate pathway, CICs cannot use different metabolic substrates (e.g., glucose, acetate and glutamine), orchestrate oncogenic upstream and downstream signaling pathways (e.g., AKT, mTORC1 and AMPK) and activate the dysfunctional TP53-SREBP-YAP-TAZ axis (Mullen et al. 2016). Thus, statins may inhibit CIC redundant mechanisms that protect them against specific targeted therapies. Concurrently, simvastatin inhibits GGPP-mediated prenylation of RhoA, leading to cytoplasmic accumulation of its inactive form and decrease of its GTPase activity. Besides these effects, simvastatin also decreases YAP/TAZ activity by inducing
YAP phosphorylation and its cytoplasmic accumulation. Thus, simvastatin may disrupt the crosstalk among pathways, modulating the transcription of genes involved in stemness/EMT and drug resistance.
Residual metastatic disease post-surgery, even at the microscopic level, represents the principal threat of recurrence. An important rate-limiting step resides in tumor dormancy, where CICs remain in a quiescent state in the metastatic foci. When CICs reach a target site (e.g., the omentum), a tight regulation imposed by the hosting microenvironment will dictate the fate of these cells, which implies a balance in the secretion of soluble factors, modulation of the extracellular matrix and the angiogenic switch (Bleau et al. 2014). If the microenvironment becomes favorable, CICs will re-enter into the cell cycle, divide and eventually manifest as recurrent disease. The traditional approach for the recurrent disease has been the sequential selection and prescription of different chemotherapies, ideally not sharing cross-resistance mechanisms (Bookman 2016). Unfortunately, many of the alternatives cannot be used chronically due to their high cost or toxicity. Thus, an alternative is to develop new therapies capable of maintaining CICs in long-term tumor dormancy (Yeh & Ramaswamy 2015). Dormancy involves stagnation of overall tumor growth due to equilibrium of proliferation and cell death. One of the mechanisms controlling the dormancy state is the so-called ‘angiogenic switch’ that is regulated by a balance between pro- and anti-angiogenic factors produced by the tumor and its microenvironment (Yeh & Ramaswamy 2015). Metastatic cancer cells need to recruit nearby vessels or induce angiogenesis to grow beyond 1 or 2 mm (Yeh & Ramaswamy 2015). In our xenograft model, tumors arising from CICs were more vascularized compared with those emerging from non-CICs. The long-term use of metronomic chemotherapy, bevacizumab or its combination is among the strategies used to avoid the angiogenic switch in HGS-ovC (Barber et al. 2013). Simvastatin administered at conventional concentrations (0.2 µM for 24 h; equivalent to 40–80 mg/day PO) inhibits angiogenesis in vitro in colon cancer cell lines and synergizes with bevacizumab by suppressing angiogenic mediators such as angiopoietin 2, binding immunoglobulin protein (BiP) and Hsp90α (Lee et al. 2014). Thus, statins could constitute an alternative to avoid the angiogenic switch. In this study, upon initiation of simvastatin treatment in our xenograft model, a significant decrease in vascular recruitment occurs and these
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
834S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
vessels are already smaller and collapsed. Our findings reaffirm the potential of statins as anti-angiogenic agents as observed in other cancers (Lee et al. 2014, Sounni et al. 2014).
We demonstrated the good tolerance and anti-tumor effect of simvastatin when administered intraperitoneally to mice harboring HGS-ovC xenografts. Simvastatin administered orally is well tolerated and induced a clinical response, at least regarding quality of life, when used in an adverse scenario such as recurrence.
However, our study presents some limitations. Our numbers are still small to draw definitive conclusions, despite clinical reports and our data showing that statin users exhibit better outcomes regarding progression-free survival (PFS) and OS (Khan et al. 2015). These outcomes must be validated in prospective cohorts. Besides, statin use offers benefits, reducing systemic inflammation (as reflected by the lower SII found among those recurrent cases who were users), ascites accumulation and delaying cancer progression. Thus, its use should not be prohibited or suspended when already prescribed.
In summary, simvastatin, used in a daily-based scheme, can revert stemness and EMT and impair spheroid assembly of HGS-ovC-derived CICs. Consequently, CICs become less metastatic and potentially more vulnerable to other therapies (work in progress by our group). The principal mechanism behind statin-mediated effects involves the inhibition of the mevalonate pathway, a critical pathway for CIC adaptation to microenvironmental conditions.
Clinically, our findings on better PFS and OS in a cohort of patients with recurrent HGS-ovC are promising. Considering the high cost, low tolerability and the little response rates of many of the current therapies, the use of oral or intraperitoneal simvastatin arise as a cost/effective and safe alternative to prevent or treat recurrent HGS-ovCs.
Supplementary dataThis is linked to the online version of the paper at https://doi.org/10.1530/ERC-18-0132.
Declaration of interestThe authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
FundingThis work was supported by FONDECYT 1120292, 1160800 (M A C), 1140970 (G I O), 11140657 (M L B), 1171463 (C B), BMRC 13CTI-21526-P6
(M L B, G I O), IMII P09/016-F (M L B, G I O, C I, J C R) and CONICYT-FONDAP 15130011 (G I O, J C R).
Authors’ contribution statementConception and design: Kato S, Liberona F, Cuello M. Development of methodology: Kato S, Liberona F, Cerda-Infante J, Henríquez J, Bizama C, Roa J C, Montecinos V, Cuello M. Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): Kato S, Liberona F, Cerda-Infante J, Henríquez J, Sánchez M, Bizama C, Bravo L, Gonzalez P, Gejman R, Brañes J, García K, Ibañez C, Owen G, Roa J C, Montecinos V, Cuello M. Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): Kato S, Liberona F, Cerda-Infante J, Henríquez J, Ibañez C, Montecinos V, Roa J C, Cuello M. Writing, review, and/or revision of the manuscript: Kato S, Liberona F, Cerda-Infante J, Sánchez M, Henríquez J, Bravo L, Gonzalez P, Gejman R, Brañes J, Ibañez C, García K, Owen G, Roa J C, Montecinos V, Cuello M. Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): Kato S, Liberona F, Bravo L, Gonzalez P, Gejman R, Brañes J, García K, Ibañez C, Owen G, Cuello M. Study supervision: Kato S, Montecinos V, Roa J C, Cuello M.
AcknowledgementsThe authors would like to thank Editage (www.editage.com) and Grammarly for English language and grammar editing.
ReferencesBaenke F, Peck B, Miess H & Schulze A 2013 Hooked on fat: the role of
lipid synthesis in cancer metabolism and tumour development. Disease Models and Mechanisms 6 1353–1363. (https://doi.org/10.1242/dmm.011338)
Baldwin LA, Huang B, Miller RW, Tucker T, Goodrich ST, Podzielinski I, DeSimone CP, Ueland FR, van Nagell JR & Seamon LG 2012 Ten-year relative survival for epithelial ovarian cancer. Obstetrics and Gynecology 120 612–618. (https://doi.org/10.1097/AOG.0b013e318264f794)
Barber EL, Zsiros E, Lurain JR, Rademaker A, Schink JC & Neubauer NL 2013 The combination of intravenous bevacizumab and metronomic oral cyclophosphamide is an effective regimen for platinum-resistant recurrent ovarian cancer. Journal of Gynecologic Oncology 24 258–264. (https://doi.org/10.3802/jgo.2013.24.3.258)
Berndt N, Hamilton AD & Sebti SM 2011 Targeting protein prenylation for cancer therapy. Nature Reviews Cancer 11 775–791. (https://doi.org/10.1038/nrc3151)
Björkhem-Bergman L, Lindh JD & Bergman P 2011 What is a relevant statin concentration in cell experiments claiming pleiotropic effects? British Journal of Clinical Pharmacology 72 164–165. (https://doi.org/10.1111/j.1365-2125.2011.03907.x)
Blatter S & Rottenberg S 2015 Minimal residual disease in cancer therapy – small things make all the difference. Drug Resistance Updates 21–22 1–10. (https://doi.org/10.1016/j.drup.2015.08.003)
Bleau AM, Agliano A, Larzabal L, de Aberasturi AL & Calvo A 2014 Metastatic dormancy: a complex network between cancer stem cells and their microenvironment. Histology and Histopathology 29 1499–1510. (https://doi.org/10.14670/HH-29.1499)
Bookman MA 2016 Optimal primary therapy of ovarian cancer. Annals of Oncology 27 (Supplement 1) i58–i62. (https://doi.org/10.1093/annonc/mdw088)
Buick RN, Pullano R & Trent JM 1985 Comparative properties of five human ovarian adenocarcinoma cell lines. Cancer Research 45 3668–3676.
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
835S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
Chen X, Chen S, Xiu YL, Sun KX, Zong ZH & Zhao Y 2015 RhoC is a major target of microRNA-93-5P in epithelial ovarian carcinoma tumorigenesis and progression. Molecular Cancer 14 31. (https://doi.org/10.1186/s12943-015-0304-6)
Cheng KW, Agarwal R & Mills G 2010 Ras-superfamily GTP-ases in ovarian cancer. In Ovarian Cancer, 2nd ed., pp 229–240. Eds MS Stack & D Fishman. New York, NY, USA: Springer.
Chisholm RH, Lorenzi T, Lorz A, Larsen AK, de Almeida LN, Escargueil A & Clairambault J 2015 Emergence of drug tolerance in cancer cell populations: an evolutionary outcome of selection, nongenetic instability, and stress-induced adaptation. Cancer Research 75 930–939. (https://doi.org/10.1158/0008-5472.CAN-14-2103)
Cieply B, Koontz C & Frisch SM 2015 CD44S-hyaluronan interactions protect cells resulting from EMT against anoikis. Matrix Biology 48 55–65. (https://doi.org/10.1016/j.matbio.2015.04.010)
Couttenier A, Lacroix O, Vaes E, Cardwell CR, De Schutter H & Robert A 2017 Statin use is associated with improved survival in ovarian cancer: a retrospective population-based study. PLoS ONE 12 e0189233. (https://doi.org/10.1371/journal.pone.0189233)
Cuello M, Kato S & Liberona MF 2018 The impact on high-grade serous ovarian cancer of obesity and lipid metabolism-related gene expression patterns: the underestimated driving force affecting prognosis. Journal of Cellular and Molecular Medicine 22 1805–1815. (https://doi.org/10.1111/jcmm.13463)
Di Palma T, Lucci V, de Cristofaro T, Filippone MG & Zannini M 2014 A role for PAX8 in the tumorigenic phenotype of ovarian cancer cells. BMC Cancer 14 292. (https://doi.org/10.1186/1471-2407-14-292)
Domcke S, Sinha R, Levine DA, Sander C & Schultz N 2013 Evaluating cell lines as tumour models by comparison of genomic profiles. Nature Communications 4 2126. (https://doi.org/10.1038/ncomms3126)
Eason K & Sadanandam A 2016 Molecular or metabolic reprograming: what triggers tumor subtypes? Cancer Research 76 5195–5200. (https://doi.org/10.1158/0008-5472.CAN-16-0141)
Freed-Pastor WA, Mizuno H, Zhao X, Langerød A, Moon SH, Rodriguez-Barrueco R, Barsotti A, Chicas A, Li W, Polotskaia A, et al. 2012 Mutant p53 disrupts mammary tissue architecture via the mevalonate pathway. Cell 148 244–258. (https://doi.org/10.1016/j.cell.2011.12.017)
Fuchtner C, Emma DA, Manetta A, Gamboa G, Bernstein R & Liao SY 1993 Characterization of a human ovarian carcinoma cell line: UCI 101. Gynecologic Oncology 48 203–209. (https://doi.org/10.1006/gyno.1993.1034)
Fuh KC, Secord AA, Bevis KS, Huh W, ElNaggar A, Blansit K, Previs R, Tillmanns T, Kapp DS & Chan JK 2015 Comparison of bevacizumab alone or with chemotherapy in recurrent ovarian cancer patients. Gynecologic Oncology 139 413–418. (https://doi.org/10.1016/j.ygyno.2015.06.041)
Gao Y, Foster R, Yang X, Feng Y, Shen JK, Mankin HJ, Hornicek FJ, Amiji MM & Duan Z 2015 Up-regulation of CD44 in the development of metastasis, recurrence and drug resistance of ovarian cancer. Oncotarget 6 9313–9326. (https://doi.org/10.18632/oncotarget.3220)
Gerlinger M, Rowan AJ, Horswell S, Larkin J, Endesfelder D, Gronroos E, Martinez P, Matthews N, Stewart A, Tarpey P, et al. 2012 Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. New England Journal of Medicine 366 883–892. (https://doi.org/10.1056/NEJMoa1113205)
Greenaway JB, Virtanen C, Osz K, Revay T, Hardy D, Shepherd T, DiMattia G & Petrik J 2016 Ovarian tumour growth is characterized by mevalonate pathway gene signature in an orthotopic, syngeneic model of epithelial ovarian cancer. Oncotarget 7 47343–47365. (https://doi.org/10.18632/oncotarget.10121)
Hansen JM, Coleman RL & Sood AK 2016 Targeting the tumour microenvironment in ovarian cancer. European Journal of Cancer 56 131–143. (https://doi.org/10.1016/j.ejca.2015.12.016)
Hernandez L, Kim MK, Lyle LT, Bunch KP, House CD, Ning F, Noonan AM & Annunziata CM 2016 Characterization of ovarian cancer cell lines as in vivo models for preclinical studies. Gynecologic Oncology 142 332–340. (https://doi.org/10.1016/j.ygyno.2016.05.028)
Horiuchi A, Kikuchi N, Osada R, Wang C, Hayashi A, Nikaido T & Konishi I 2008 Overexpression of RhoA enhances peritoneal dissemination: RhoA suppression with Lovastatin may be useful for ovarian cancer. Cancer Science 99 2532–2539. (https://doi.org/10.1111/j.1349-7006.2008.00977.x)
Hu J, Liu Z & Wang X 2013 Does TP53 mutation promote ovarian cancer metastasis to omentum by regulating lipid metabolism? Medical Hypotheses 81 515–520. (https://doi.org/10.1016/j.mehy.2013.06.009)
Hu B, Yang XR, Xu Y, Sun YF, Sun C, Guo W, Zhang X, Wang WM, Qiu SJ, Zhou J, et al. 2014 Systemic immune-inflammation index predicts prognosis of patients after curative resection for hepatocellular carcinoma. Clinical Cancer Research 20 6212–6222. (https://doi.org/10.1158/1078-0432.CCR-14-0442)
Iwanicki MP, Chen HY, Iavarone C, Zervantonakis IK, Muranen T, Novak M, Ince TA, Drapkin R & Brugge JS 2016 Mutant p53 regulates ovarian cancer transformed phenotypes through autocrine matrix deposition. JCI Insight 1 1–20. (https://doi.org/10.1172/jci.insight.86829)
Kato S, Smalley S, Sadarangani A, Chen-Lin K, Oliva B, Branes J, Carvajal J, Gejman R, Owen GI & Cuello M 2010 Lipophilic but not hydrophilic statins selectively induce cell death in gynaecological cancers expressing high levels of HMGCoA reductase. Journal of Cellular and Molecular Medicine 14 1180–1193. (https://doi.org/10.1111/j.1582-4934.2009.00771.x)
Kato S, Abarzua-Catalan L, Trigo C, Delpiano A, Sanhueza C, Garcia K, Ibanez C, Hormazabal K, Diaz D, Branes J, et al. 2015 Leptin stimulates migration and invasion and maintains cancer stem-like properties in ovarian cancer cells: an explanation for poor outcomes in obese women. Oncotarget 6 21100–21119. (https://doi.org/10.18632/oncotarget.4228)
Kenny HA, Dogan S, Zillhardt M, K Mitra A, Yamada SD, Krausz T & Lengyel E 2009 Organotypic models of metastasis: a three-dimensional culture mimicking the human peritoneum and omentum for the study of the early steps of ovarian cancer metastasis. Cancer Treatment and Research 149 335–351. (https://doi.org/10.1007/978-0-387-98094-2_16)
Khan MS, Fatima K & Rameez 2015 Impact of statins on risk and survival of ovarian cancer. Journal of Gynecologic Oncology 26 240–241. (https://doi.org/10.3802/jgo.2015.26.3.240)
Klemba A, Purzycka-Olewiecka JK, Wcislo G, Czarnecka AM, Lewicki S, Lesyng B, Szczylik C & Kieda C 2018 Surface markers of cancer stem-like cells of ovarian cancer and their clinical relevance. Contemporary Oncology 22 48–55. (https://doi.org/10.5114/wo.2018.73885)
Kobayashi Y, Kashima H, Wu RC, Jung JG, Kuan JC, Gu J, Xuan J, Sokoll L, Visvanathan K, Shih IM, et al. 2015 Mevalonate pathway antagonist suppresses formation of serous tubal intraepithelial carcinoma and ovarian carcinoma in mouse models. Clinical Cancer Research 21 4652–4662. (https://doi.org/10.1158/1078-0432.CCR-14-3368)
Lee SJ, Lee I, Lee J, Park C & Kang WK 2014 Statins, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors, potentiate the anti-angiogenic effects of bevacizumab by suppressing angiopoietin2, BiP, and Hsp90α in human colorectal cancer. British Journal of Cancer 111 497–505. (https://doi.org/10.1038/bjc.2014.283)
Liliac L, Carcangiu ML, Canevari S, Caruntu ID, Ciobanu Apostol DG, Danciu M, Onofriescu M & Amalinei C 2013 The value of PAX8 and WT1 molecules in ovarian cancer diagnosis. Romanian Journal of Morphology and Embryology 54 17–27.
Liu Y, Metzinger MN, Lewellen KA, Cripps SN, Carey KD, Harper EI, Shi Z, Tarwater L, Grisoli A, Lee E, et al. 2015 Obesity contributes to ovarian cancer metastatic success through increased lipogenesis,
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access

Printed in Great BritainPublished by Bioscientifica Ltd.https://doi.org/10.1530/ERC-18-0132
http://erc.endocrinology-journals.org © 2018 Society for Endocrinology
836S Kato et al. Statins reduce metastasis in ovarian cancer
25:10Endocrine-Related Cancer
enhanced vascularity, and decreased infiltration of M1 macrophages. Cancer Research 75 5046–5057. (https://doi.org/10.1158/0008-5472.CAN-15-0706)
Mitra AK, Davis DA, Tomar S, Roy L, Gurler H, Xie J, Lantvit DD, Cardenas H, Fang F, Liu Y, et al. 2015 In vivo tumor growth of high-grade serous ovarian cancer cell lines. Gynecologic Oncology 138 372–377. (https://doi.org/10.1016/j.ygyno.2015.05.040)
Mullen PJ, Yu R, Longo J, Archer MC & Penn LZ 2016 The interplay between cell signalling and the mevalonate pathway in cancer. Nature Reviews Cancer 16 718–731. (https://doi.org/10.1038/nrc.2016.76)
Network CGAR 2011 Integrated genomic analyses of ovarian carcinoma. Nature 474 609–615. (https://doi.org/10.1038/nature10166)
Ohyagi-Hara C, Sawada K, Kamiura S, Tomita Y, Isobe A, Hashimoto K, Kinose Y, Mabuchi S, Hisamatsu T, Takahashi T, et al. 2013 miR-92a inhibits peritoneal dissemination of ovarian cancer cells by inhibiting integrin alpha5 expression. American Journal of Pathology 182 1876–1889. (https://doi.org/10.1016/j.ajpath.2013.01.039)
Orian-Rousseau V 2015 CD44 acts as a signaling platform controlling tumor progression and metastasis. Frontiers in Immunology 6 154. (https://doi.org/10.3389/fimmu.2015.00154)
Plutoni C, Bazellieres E, Le Borgne-Rochet M, Comunale F, Brugues A, Séveno M, Planchon D, Thuault S, Morin N, Bodin S, et al. 2016 P-cadherin promotes collective cell migration via a Cdc42-mediated increase in mechanical forces. Journal of Cell Biology 212 199–217. (https://doi.org/10.1083/jcb.201505105)
Sato M, Kawana K, Adachi K, Fujimoto A, Yoshida M, Nakamura H, Nishida H, Inoue T, Taguchi A, Takahashi J, et al. 2016 Spheroid cancer stem cells display reprogrammed metabolism and obtain energy by actively running the tricarboxylic acid (TCA) cycle. Oncotarget 7 33297–33305. (https://doi.org/10.18632/oncotarget.8947)
Sebio A & Lenz HJ 2015 Molecular pathways: hippo signaling, a critical tumor suppressor. Clinical Cancer Research 21 5002–5007. (https://doi.org/10.1158/1078-0432.CCR-15-0411)
Shield K, Ackland ML, Ahmed N & Rice GE 2009 Multicellular spheroids in ovarian cancer metastases: biology and pathology. Gynecologic Oncology 113 143–148. (https://doi.org/10.1016/j.ygyno.2008.11.032)
Sounni NE, Cimino J, Blacher S, Primac I, Truong A, Mazzucchelli G, Paye A, Calligaris D, Debois D, De Tullio P, et al. 2014 Blocking lipid synthesis overcomes tumor regrowth and metastasis after antiangiogenic therapy withdrawal. Cell Metabolism 20 280–294. (https://doi.org/10.1016/j.cmet.2014.05.022)
Tanenbaum LM, Mantzavinou A, Subramanyam KS, Del Carmen MG & Cima MJ 2017 Ovarian cancer spheroid shrinkage following continuous exposure to cisplatin is a function of spheroid diameter. Gynecologic Oncology 146 161–169. (https://doi.org/10.1016/j.ygyno.2017.04.014)
Vaughan S, Coward JI, Bast RC Jr, Berchuck A, Berek JS, Brenton JD, Coukos G, Crum CC, Drapkin R, Etemadmoghadam D, et al. 2011 Rethinking ovarian cancer: recommendations for improving outcomes. Nature Reviews Cancer 11 719–725. (https://doi.org/10.1038/nrc3144)
Vieira AF & Paredes J 2015 P-cadherin and the journey to cancer metastasis. Molecular Cancer 14 178. (https://doi.org/10.1186/s12943-015-0448-4)
Vousden KH & Ryan KM 2009 p53 and metabolism. Nature Reviews Cancer 9 691–700. (https://doi.org/10.1038/nrc2715)
Wallbillich JJ, Forde B, Havrilesky LJ & Cohn DE 2016 A personalized paradigm in the treatment of platinum-resistant ovarian cancer – a cost utility analysis of genomic-based versus cytotoxic therapy. Gynecologic Oncology 142 144–149. (https://doi.org/10.1016/j.ygyno.2016.04.024)
Xiang L, Rong G, Zhao J, Wang Z & Shi F 2018 Identification of candidate genes associated with tubal origin of high-grade serous ovarian cancer. Oncology Letters 15 7769–7775. (htpps://doi.org/10.3892/ol.2018.8346)
Yan HC, Fang LS, Xu J, Qiu YY, Lin XM, Huang HX & Han QY 2014 The identification of the biological characteristics of human ovarian cancer stem cells. European Review for Medical and Pharmacological Sciences 18 3497–3503.
Yang-Hartwich Y, Soteras MG, Lin ZP, Holmberg J, Sumi N, Craveiro V, Liang M, Romanoff E, Bingham J, Garofalo F, et al. 2015 p53 protein aggregation promotes platinum resistance in ovarian cancer. Oncogene 34 3605–3616. (https://doi.org/10.1038/onc.2014.296)
Yeh AC & Ramaswamy S 2015 Mechanisms of cancer cell dormancy – another hallmark of cancer? Cancer Research 75 5014–5022. (https://doi.org/10.1158/0008-5472.CAN-15-1370)
Yeung TL, Leung CS, Yip KP, Au Yeung CL, Wong ST & Mok SC 2015 Cellular and molecular processes in ovarian cancer metastasis. A review in the theme: cell and molecular processes in cancer metastasis. American Journal of Physiology: Cell Physiology 309 C444–C456. (https://doi.org/10.1152/ajpcell.00188.2015)
Zhang Y, Xia H, Ge X, Chen Q, Yuan D, Leng W, Chen L, Tang Q & Bi F 2014 CD44 acts through RhoA to regulate YAP signaling. Cell Signaling 26 2504–2513. (https://doi.org/10.1016/j.cellsig.2014.07.031)
Zhao S, Chen C, Chang K, Karnad A, Jagirdar J, Kumar AP & Freeman JW 2016 CD44 expression level and isoform contributes to pancreatic cancer cell plasticity, invasiveness, and response to therapy. Clinical Cancer Research 22 5592–5604. (https://doi.org/10.1158/1078-0432.CCR-15-3115)
Zhong JH, Huang DH & Chen ZY 2017 Prognostic role of systemic immune-inflammation index in solid tumors: a systematic review and meta-analysis. Oncotarget 8 75381–75388. (https://doi.org/10.18632/oncotarget.18856)
Received in final form 13 May 2018Accepted 30 May 2018Accepted Preprint published online 30 May 2018
Downloaded from Bioscientifica.com at 01/16/2022 02:34:03PMvia free access


















