
Scanning transmission electron microscopic study of molluscan hemocyanins in various aggregation...
8
JOURNAL OF ULTRASTRUCTURE AND MOLECULAR STRUCTURE RESEARCH 102, 221-228 (1989) Scanning Transmission Electron Microscopic Study of Moiluscan Hemocyanins in Various Aggregation States: Comparison with Light Scattering Molecular Weights’ MARY G. HAMILTON,* THEODORE T. HERSKOVITS,~ PAUL S. FURCINITTI$.~ AND JOSEPH S. WALLS *Division of Science and Matkematics, Fordkam University, New York, New York 10023; fDepartment of Chemistry, Fordkam University, Bronx, New York 10458; and #Biology Department, Brookhaven National Laboratory, Upton, New York 11973 ReceivedAugust 14, 1989 The masses of individual particles of the hemocyanins of six members of two molluscan classes, Polyplacophora and Gastropoda, have been determined by scanning transmission electron microscopy (STEM) of unstained specimens dried from the frozen state. The decameric hemocyanins of two chitons, Mop&o muscosa and Steno- ptax conspicuu, had masses of 4.20 -C 0.18 and 4.47 f 0.56 MDa, respectively; the didecameric hemocyanins of two gaatropods, Faacidkria tulipa and PleuropCoca gigantea, had masses of 8.67 it 0.44 and 9.96 f 0.39 MDa, respec- tively; and the trid~americ hemocyanin of Lunuti hew had a mass of 13.50 * 0.44 MDa. The STEM values were in close agreement with those obtained by light scatter- ing measurements of the same samples in solution. For Bus&con cednrrium, a gastropod with a multidecameric hemocyanin, nine size classes from didecamers to deca- deeamem with masses that corresponded to multiples of a basic decanter (4.4 MDa) were detected. The appearance of unstained specimens of the cylindrical particles differs from negatively stained specimens. Viewed end-on the cylinders show no internal structure, but in well- preserved specimens cavities are apparent in the side views of the cylinders that resemble those seen in nega- tively stained specimens. Although they lack the charac- teristic “tiered” appearance, the number of decameric units can be counted and their arrangement within the particle seen. 0 1999 Academic Press, Inc. INTRODUCTION The hemocyanins of molluscs are very large pro- tein assemblies whose cylindrical architecture was revealed by electron microscopy; they look like lin- ear arrays of a repeating cylindrical unit (diameter ca. 35 nm, length ca. 19 nm) that has fivefold sym- ’ This work was supported in part by a Faculty Research Grant from Fordham University. Support for the Brookhaven STEM is provided by the N.I.H., Grant No. RR 01777, and the U.S.D.O.E. 2 Present address:BiophysicsResearchDivision, University of Michigan, Ann Arbor, MI 48109. metry (van Bruggen et al., 1981). Their large sixes, reflected in high sedimentation coeficients (50 to >200 S), made them the subject of ultracentrifugal analyses in the pioneering studies of Svedberg and Hedenius (1934). While some are monodisperse in the ultracentrifuge, others exhibit pauci&spersity. Their molecular weights (from 4 to >20 million Da) have been measured by a variety of classical physi- cal-chemical techniques including light scattering (Herskovits, 1988). The state of ation of the cylindrical units is a&&d by the ionic conditions of the solution. The basic unit is identified as a decamer because it can be further dissociated to 10 monomeric polypeptide subunits at alkaline pH or in urea or guanidine HCl (see review of Herskovits, 1988). Each monomeric subunit, paired as five dimers within the decameric cylinder, consists of eight functional units, two of which form a collar that can be seen in electron micrographs at one end of the cylinder with the other six forming the wall of the cylinder (van Breemen et al., 1977; reviewed by van Holde and Miller, 1982). So far hemocyanins have been found in the he- molymphs of animals in four of the six classes of Phylum Mollusca (Ellerton et al., 1983; Morse et al., 1986). The hemocyanins range from a single decameric unit in chitons and cephalopods to di-, tri-, and higher multiples of the decamer in gastro- pods. Since the cephalopdan hemocyanins differ from the pattern of organization described above in that they have collars at both ends of smaller cylin- ders (Lamy et al., 1986; Wichjertes et al., 19861, we have not included them in this comparative study of hemocyanins that, in size and morphology, appear to be polymers of a single kind of decameric unit. The single decamer type of hemocyanin of the present study is found in the chitona, Class Polypla- cophora , ca. 60 S, M, = 4.2-4.6 million (Werskovits and Hamilton, 1987a,b). In contrast, the gastropo- dan hemocyanins exhibit a spectrum of sizes. For 221 08&x9-1606/89 $3.00 Copyright 8 1989 by Academic Prees, Inc. All rights of reproduction in anp form reserved.
Transcript of Scanning transmission electron microscopic study of molluscan hemocyanins in various aggregation...

JOURNAL OF ULTRASTRUCTURE AND MOLECULAR STRUCTURE RESEARCH 102, 221-228 (1989)
Scanning Transmission Electron Microscopic Study of Moiluscan Hemocyanins in Various Aggregation States: Comparison with
Light Scattering Molecular Weights’
MARY G. HAMILTON,* THEODORE T. HERSKOVITS,~ PAUL S. FURCINITTI$.~ AND JOSEPH S. WALLS
*Division of Science and Matkematics, Fordkam University, New York, New York 10023; fDepartment of Chemistry, Fordkam University, Bronx, New York 10458; and #Biology Department, Brookhaven National Laboratory, Upton, New York 11973
Received August 14, 1989
The masses of individual particles of the hemocyanins of six members of two molluscan classes, Polyplacophora and Gastropoda, have been determined by scanning transmission electron microscopy (STEM) of unstained specimens dried from the frozen state. The decameric hemocyanins of two chitons, Mop&o muscosa and Steno- ptax conspicuu, had masses of 4.20 -C 0.18 and 4.47 f 0.56 MDa, respectively; the didecameric hemocyanins of two gaatropods, Faacidkria tulipa and PleuropCoca gigantea, had masses of 8.67 it 0.44 and 9.96 f 0.39 MDa, respec- tively; and the trid~americ hemocyanin of Lunuti hew had a mass of 13.50 * 0.44 MDa. The STEM values were in close agreement with those obtained by light scatter- ing measurements of the same samples in solution. For Bus&con cednrrium, a gastropod with a multidecameric hemocyanin, nine size classes from didecamers to deca- deeamem with masses that corresponded to multiples of a basic decanter (4.4 MDa) were detected. The appearance of unstained specimens of the cylindrical particles differs from negatively stained specimens. Viewed end-on the cylinders show no internal structure, but in well- preserved specimens cavities are apparent in the side views of the cylinders that resemble those seen in nega- tively stained specimens. Although they lack the charac- teristic “tiered” appearance, the number of decameric units can be counted and their arrangement within the particle seen. 0 1999 Academic Press, Inc.
INTRODUCTION
The hemocyanins of molluscs are very large pro- tein assemblies whose cylindrical architecture was revealed by electron microscopy; they look like lin- ear arrays of a repeating cylindrical unit (diameter ca. 35 nm, length ca. 19 nm) that has fivefold sym-
’ This work was supported in part by a Faculty Research Grant from Fordham University. Support for the Brookhaven STEM is provided by the N.I.H., Grant No. RR 01777, and the U.S.D.O.E.
2 Present address: Biophysics Research Division, University of Michigan, Ann Arbor, MI 48109.
metry (van Bruggen et al., 1981). Their large sixes, reflected in high sedimentation coeficients (50 to >200 S), made them the subject of ultracentrifugal analyses in the pioneering studies of Svedberg and Hedenius (1934). While some are monodisperse in the ultracentrifuge, others exhibit pauci&spersity. Their molecular weights (from 4 to >20 million Da) have been measured by a variety of classical physi- cal-chemical techniques including light scattering (Herskovits, 1988). The state of ation of the cylindrical units is a&&d by the ionic conditions of the solution. The basic unit is identified as a decamer because it can be further dissociated to 10 monomeric polypeptide subunits at alkaline pH or in urea or guanidine HCl (see review of Herskovits, 1988). Each monomeric subunit, paired as five dimers within the decameric cylinder, consists of eight functional units, two of which form a collar that can be seen in electron micrographs at one end of the cylinder with the other six forming the wall of the cylinder (van Breemen et al., 1977; reviewed by van Holde and Miller, 1982).
So far hemocyanins have been found in the he- molymphs of animals in four of the six classes of Phylum Mollusca (Ellerton et al., 1983; Morse et al., 1986). The hemocyanins range from a single decameric unit in chitons and cephalopods to di-, tri-, and higher multiples of the decamer in gastro- pods. Since the cephalopdan hemocyanins differ from the pattern of organization described above in that they have collars at both ends of smaller cylin- ders (Lamy et al., 1986; Wichjertes et al., 19861, we have not included them in this comparative study of hemocyanins that, in size and morphology, appear to be polymers of a single kind of decameric unit.
The single decamer type of hemocyanin of the present study is found in the chitona, Class Polypla- cophora , ca. 60 S, M, = 4.2-4.6 million (Werskovits and Hamilton, 1987a,b). In contrast, the gastropo- dan hemocyanins exhibit a spectrum of sizes. For
221 08&x9-1606/89 $3.00
Copyright 8 1989 by Academic Prees, Inc. All rights of reproduction in anp form reserved.

222 HAMILTON ET AL.
example, while the hemocyanin of the marine snail Fasciolaria tulipa sediments at 106 S (M, = 8.6 mil- lion) (Herskovits et al., 1988) and exists entirely in the didecameric form, the major component of the hemocyanin of a moon snail, Lunatia heros, sedi- ments at 136 S and is a tridecamer (Herskovits et al., 1985). In contrast, ultracentrifugal analysis of the hemocyanin of Busycon contrarium, a member of the Melongenidae family, reveals a multicomponent mixture of species with sedimentation coefficients of 105, 132,155,170, 185, and ca. 200 S (Herskovits et al., 1989b). A similar complexity has been observed in the hemocyanins of two other gastropods, Dola- bella auricularia (Makino, 1971) and Megathura crenulata (Senozan et al., 1981).
Scanning transmission electron microscopy (STEM) uses the electron scattering at wide angles to determine the masses of individual, unstained particles (Wall and Hainfeld, 1986). In this paper we first compare the STEM masses with the weight- average molecular weights obtained by light scat- tering for homogeneous populations of hemocyanin molecules: those of two chitons, Mopalia muscosa and Stenoplax conspicuu, those of two members of the tulip family of gastropods, F. tulipa and Pleu- roploca gigantea, and that of the moon snail, L. heros. We have also measured the masses of individ- ual particles in the pauci-disperse hemocyanin of B. contrarium, information unattainable by any solu- tion method without prior fractionation of the mix- ture.
The basis for the wide diversity of sizes among proteins with the same function is not understood. One aim of the mass measurements was to deter- mine whether the mass increment in the assembly of a typical population of multidecameric particles is uniform.
Preliminary reports of this work have been pre- sented (Furcinitti et al., 1988; Hamilton et al., 1989). The particle masses for the B. contrarium hemocy- anin have been quoted with additional STEM mass measurements for four other members of the Melon- genidae family in a comparative study of their sub- unit structure and higher order assembly (Hersko- vits et al., 1989a).
MATERIALS AND METHODS
Live specimens of chitons and gastropods were purchased from Pacific Bio-Marine Laboratory (Venice, CA), Gulf Specimen Co. (Panacea, FL), and the Marine Biology Laboratory (Woods Hole, MA). Hemolymphs were collected in the presence of proteolytic inhibitors and purified by gel filtration column chromatography on BioGel A-5m in 0.1 A4 Tris3-HCl, pH 8.0,0.05 M MgCl,, 0.01 M CaCle as previously described (Herskovits et al., 1985). Sam- ples of I?. contrarium hemocyanin were dialyzed against other buffers: 0.1 M sodium acetate, pH 5.0,O.Ol A4 MgCl,; 0.1 M Tris- HCl, pH 8.0, 0.01 M MgCl,. Samples were analyzed by velocity centrifugation in a Beckman Model E instrument equipped with schlieren and ultraviolet-photoelectric scanner optics.
For STEM analysis samples were either dialyzed against 0.1 M Hepes, pH 7.4,0.05 M MgCl,, or diluted at least lo-fold into that buffer because Tris-HCI is difficult to wash from the specimen. The best preservation of the B. contmrium hemocyanin was ob- tained in 0.01 M MgCl,. Samples were either freeze-dried or neg- atively stained with uranyl acetate. Samples were deposited by diffusion. Tobacco mosaic virus, used as an internal standard, was deposited first by injection of ca. 2 ~1 of a 60 pg/ml solution into a drop of buffer already on the grid. The grid was washed twice with buffer and ca. 2 p,l of the hemocyanin solution (ca. 100 kg/ml) was injected into the remaining buffer. After 1 min the grid was washed eight times with 20 mM ammonium acetate (pH 71, 1 mM MgCl,. The remaining liquid was wicked to less than l-pm thickness and the grid was plunged immediately into liquid nitrogen slush. The grid was transferred under liquid nitrogen to the freeze-drier where six grids were freeze-dried overnight and transferred under vacuum to the STEM as previously described (Wall and Hainfeld, 1986). Negatively stained specimens were prepared in the same way up to the last two washes which were done with 2% uranyl acetate; the grid was air-dried. The Brookhaven STEM is operated at 40 kV using a - 140°C cold stage. The elastically scattered electron signal from the STEM large-angle annular detector was used to form the images. Freeze-dried specimens were imaged using a 512 x 512 raster with 10-A pixels at a dose of 6-10 electrons/A’ or 20 A pixels at a dose of l-5 electrons/A’. Negatively stained specimens were imaged with 5 A pixels at a dose of 25 electrons/A’. Molecular weights of individual unstained, freeze-dried hemocyanin mole- cules were measured as previously described (Wall and Hainfeld, 1986). Particles for measurement were selected for clean sur- rounding background and circular (or 5-fold) symmetry for end- views and rectangular shape for side-views. A correction for the decreased scattering intensity caused by the thickness of long cylindrical particles viewed end on was applied to the mass mea- surements (Steven et al., 1983; Wall, 1988).
3 Abbreviations used: Tris, tris(hydroxymethyl)amino- methane; Hepes, 4-(2-hydroxyethyll-1-piperaxineethanesulfonic acid.
FIGS. l-6. STEM electron micrographs of unstained, freese-dried, and negatively stained specimens of representative molluscan hemocyanins. The specimens also contained tobacco mosaic virus (diameter 18 nm). In the unstained specimens, the scan width is 0.512 pm and in the negatively stained specimens it is 0.256 pm. The scale bars represent 0.1 pm.
FIG. 1. Pbumploca gigantea hemocyanin: unstained, freeze-dried. FIG. 2. P. gigantea hemocyanin: negatively stained with uranyl acetate. FIG. 3. Lunutiu Aeros hemocyanin unstained, freeze-dried. Reproduced with permission from Hamilton et al. (1989); copyright by San
Francisco Press, Inc., San Francisco, CA 94101-6800. FIG. 4. L. Aeros hemocyanin: negatively stained with uranyl acetate. FIG. 5. Busycon contmrium hemocyanin: unstained, freeze-dried. The semi-detached circles in the lower left corner resemble the
chiton decamers in mass and appearance. FIG. 6. B. contmrium hemocyanin: negatively stained with uranyl acetate.

ti66

224 HAMILTON ET AL.
TABLE I Comparison of STEM Particle Masses with the Light-Scattering Molecular Weights of Several
Molluscan Decameric, Didecamer and Tridecameric Hemocyanins
Light- STEM scattering
Number particle molecular of
Species decamers CElzi weight (MDa)*
Mopalia muscosac 1 4.20 + 0.18 (6) 4.5 + 0.3 stmwplnz conspicua= 1 4.47 f 0.56 (17) 4.2 + 0.3 Fasciolaria tulipa 2 8.67 2 0.44 (142) 8.6 2 0.6 Pleumploca gigantea 2 8.96 + 0.39 (79) 8.8 + 0.6 Lumtia hems 3 13.50 f 0.44 (38) (14)
a Mean, standard deviation, number of measurements. s Light-scattering molecular weights for M. muscosa, S. conspicua, F.
tulipa, and P. gigantea hemocyanins are from Herskovits and Hamilton (1987a,b) and Herskovite et al. (1988); the L. hems value was taken as 30 times the average monomer value obtained at pH 10 and in 8 M urea (Herskovits et al., 1985).
c The names of the two chitons were inadvertently interchanged in Pur- cinitti et al. (1988).
RESULTS
Unstained Images of Molluscan Hemocyanins
Electron micrographs of hemocyanins typically show both circular and rectangular profiles. Figures 1,3, and 5 show electron micrographs of freeze-dried specimens of the didecameric hemocyanin of P. gi- gantea (Fig. 11, the predominantly tridecameric hemocyanin of L. heros (Fig. 31, and the multide- cameric hemocyanin of B. contrarium (Fig. 51, for which mass measurements were made by STEM. For comparison, specimens that were negatively stained with uranyl acetate are shown in Figs. 2,4, and 6. The STEM pictures of unstained specimens show less internal structure than is evident with negative staining. The characteristic tiers are not visible in the side-views, but recognizable cavities are seen. Computer enhancement (not shown) of end-on views of a freeze-dried specimen of the de- tamer of the chiton, S. conspicua, showed five-fold symmetry similar to that found by Mellema and Klug (1972) in image reconstruction of negatively stained electron micrographs of half molecules of di- decameric Kelletia kelletia and B. canaliculatum hemocyanins. One detached unit in the lower left corner of Fig. 5, indistinguishable from the authen- tic decameric particle of the chiton hemocyanin, shows the five-point “star” shape of the central cav- ity.
STEM Mass Measurements of Various Molluscan Hemocyanins Compared with Light-Scattering Molecular Weights
For four hemocyanins that are monodisperse in sedimentation analysis, the STEM particle masses
TABLE II Summary of STEM Measurements on Busycon
contrurium Hemocyanin
Mass/ Designation Mass (MDa) Length (nm) No. decamers
A. Summary of particle masses and lengths for side-views Didecamer 8.69 2 0.33 (3) 38.3 “- 2.2 (3) 4.34 Tridecamer 13.34 ” 0.26 (16) 50.6 -c 2.3 (16) 4.45 Tetradecamer 17.88 2 0.61(33) 68.1 * 3.5 (33) 4.47 Pentadecamer 22.13 2 0.58 (21) 84.5 ” 2.9 (21) 4.43 Hexadecamer 26.65 + 0.39 (5) 97.6 ” 3.7 (5) 4.44 Heptadecamer 31.01 f 0.72 (8) 117.2 r 4.3 (8) 4.43 Octadecamer 35.09 k 0.65 (5) 130.2 * 2.4 (5) 4.39 Nonadecamer 38.87 (1) 153.3 (1) 4.32 Decadecamer 43.40 (1) 169.4 (1) 4.34
4.40 2 0.06
B. Summary of particle masses for end-on views Didecamer 8.91 2 0.38 (34) 4.46 Tridecamer 13.15 2 0.38 (87) 4.38 Tetradecamer 17.12 -t 0.37 (28) 4.28 Pentadecamer 21.55 -t 0.66 (4) 4.31
4.36 ” 0.08
C. Summary of mass measurements of both side- and end-views Didecamer 8.89 2 0.37 4.45 Tridecamer 13.20 + 0.35 AA0
. . “ ”
Tetradecamer 17.35 2 0.46 4.34 Pentadecamer 22.06 2 0.59 4.41
4.40 2 05
can be compared directly with the light-scattering molecular weights shown in Table I. The hemocya- nins of the two chitons, M. muscosa and S. con- spicuu, that have sedimentation coefficients of ca. 60 S, are decamers with masses ca. 4.4 x 106; the hemo- cyanins of the two gastropods of the tulip family, F. tulipa and P. gigantea, that have sedimentation co- efficients close to 100 S, are didecamers with masses ca. 8.8 x 106. The STEM particle masses are very close to the light-scattering values for all four hemo- cyanins.
For the tridecameric hemocyanin of a moon snail, L. heros (last entry in Table I), STEM measurements gave a particle mass of 13.5 x 106. The weight- average molecular weight from light-scattering can- not be compared directly because the hemocyanin was not monodisperse: 55% sedimented at 130 S and 45% as components with higher sedimentation coef- ficients. However, the mass for the tridecamer can be estimated to be ca. 14 x 106, i.e., 30 times the average monomer mass of 4.7 x lo6 obtained by light-scattering measurements at pH 10 or in 8 M urea (Herskovits et al., 1985).
Table I shows that good agreement between the STEM masses and the light-scattering values was obtained for those specimens which best survived the preparative procedures as indicated by a large number of particles suitable for measurement. The hemocyanins of the chitons were more sensitive than the gastropodan hemocyanins and there were fewer intact particles.

STEM INVESTIGATION OF MOLLUSCAN HEMOCYANINS 225
P a r t i c I e
M a s S
i n
ii 0 0
I I I I I I I I
20 40 60 80 100 120 140 160 180 Particle Length (nm)
FIG. 7. Scatter diagram for side-view particle masses vs. length of Busycon contmrium hemocyanin at pH 8.0. The particle identifi- cations are indicated on the ordinate.
Mass Analysis of the Multidecameric Hemocyanin of B. Contrarium
Table II presents a detailed analysis for B. con- trarium hemocyanin at pH 5.0 and pH 8.0 in the presence of 0.01 M MgCl,. Other specimens at pH 7.4, 0.05 M MgCls had given similar masses (Fur- cinitti et al., 19681, but there was more fragmenta- tion and flattening (central cavity collapsed) of the particles in those specimens.
For B. contrarium hemocyanin, the relative num- bers of end-on views and side-views varied from 48 to 86% even for different specimens of the same sam- ple. Fragmentation of the cylinders was apparent in some side-views. In some cases one end had a frayed appearance. Occasionally, a circle (end-view) close to or attached to a rectangle (see Fig. 5) may have originated from attachment of the cylinder to the grid followed by its breaking at a decamer-decamer interface and falling over on its side. This explana- tion seems supported by the approximate masses of the attached circles which were usually about 4 MDa. No decamers were observed in side-views and would not be expected since decamers are observed in sed- imentation analysis of B. contrarium hemocyanin only at pH values above 9.0.
The particle masses of side-views and end-views have been evaluated separately because the masses of the particles that attached to the grid end-on re- quired a correction for the attenuation of the inten- sity of scattering by their thickness (Steven et al., 1983; Wall, 1988). The masses of the particles that
attached on their sides were not significantly af- fected by application of such a correction.
For those side-views where the width of the rect- angle was commensurate with the diam&er of the end-on view, i.e., those which showed little flatten- ing, fraying of the ends, or other signs of Eragmen- tation, length as well as mass measurements were made. The data for measurements of the specimen at pH 8.0 are shown in Fig. 7 as a scatter &gram of the mass vs length where each point rapresents one side-view particle. That the mass varies directly as the length indicates that classification by either mass or length is unambiguous. The data fall into clusters that conform with the incremental in- creases in the mass. The various particles are iden- tified by labels on the ordinate. M axaauaments at pH 5.0 gave an identical distribution excel& that a few didecamers were also observed. The data for both pHs have been combined in the means given in Ta- ble II, part A.
While the number of decamers in a particle on its side can be counted in the best cases, or deduced from the particle length, the oligomeric state of a particle viewed on end cannot be mxl by in- spection. It can, however, be determined by mass measurement. Only the masses of those views that were directly end-on as judged by a clerrr central hole with baseline scattering values and an outer profile independent of the height (or number of units) were selected as likely to be in&&. The cor- rection factors ranged from 2.5% for &decamers to 10% for the pentadecamers. None of the particles
226 HAMILTON ET AL.
r
N 0 .
0 f
P a r t i C I e S
25
20
15
10
7.5 10 12.5 15 17.5
Particle Mass (Md) 20
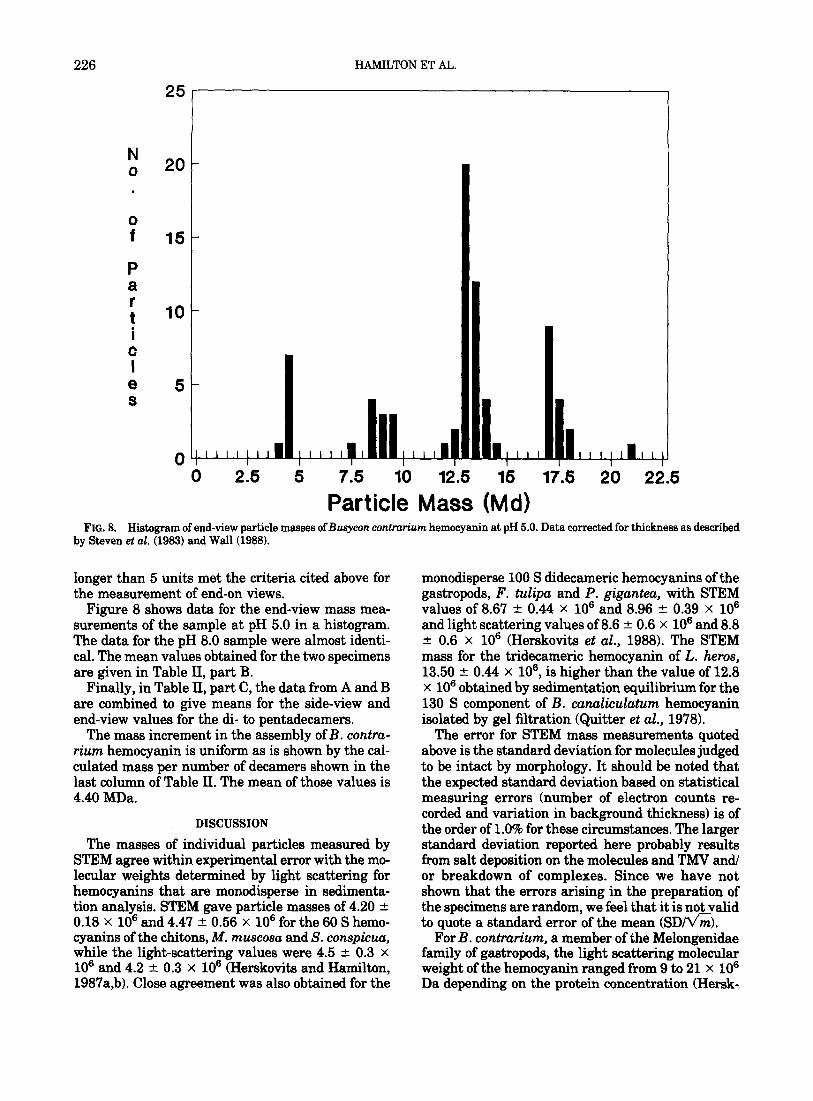
FIG. 8. Histmram of end-view oarticle masse ofBusvcon contrarium hemocyanin at pH 5.0. Data corrected for thickness as described by Steven et al. 5983) and Wall (i988).
longer than 5 units met the criteria cited above for the measurement of end-on views.
Figure 8 shows data for the end-view mass mea- surements of the sample at pH 5.0 in a histogram. The data for the pH 8.0 sample were almost identi- cal. The mean values obtained for the two specimens are given in Table II, part B.
Finally, in Table II, part C, the data from A and B are combined to give means for the side-view and end-view values for the di- to pentadecamers.
The mass increment in the assembly of B. contru- rium hemocyanin is uniform as is shown by the cal- culated mass per number of decamers shown in the last column of Table II. The mean of those values is 4.40 MDa.
DISCUSSION
The masses of individual particles measured by STEM agree within experimental error with the mo- lecular weights determined by light scattering for hemocyanins that are monodisperse in sedimenta- tion analysis. STEM gave particle masses of 4.20 f 0.18 X lo6 and 4.47 +- 0.56 X lo6 for the 60 S hemo- cyanins of the &tons, M. muscosa and S. conspicuu, while the light-scattering values were 4.5 r 0.3 x lo6 and 4.2 +- 0.3 x 10’ (Herskovits and Hamilton, 1987a,b). Close agreement was also obtained for the
monodisperse 100 S didecameric hemocyanins of the gastropods, F. tulipa and P. gigunteu, with STEM values of 8.67 + 0.44 x lo6 and 8.96 + 0.39 x lo6 and light scattering values of 8.6 + 0.6 X lo6 and 8.8 f 0.6 x lo6 (Herskovits et al., 1988). The STEM mass for the tridecameric hemocyanin of L. heros, 13.50 lr: 0.44 x 106, is higher than the value of 12.8 x lo6 obtained by sedimentation equilibrium for the 130 S component of B. cundicdatum hemocyanin isolated by gel filtration (Quitter et al., 1978).
The error for STEM mass measurements quoted above is the standard deviation for molecules judged to be intact by morphology. It should be noted that the expected standard deviation based on statistical measuring errors (number of electron counts re- corded and variation in background thickness) is of the order of 1.0% for these circumstances. The larger standard deviation reported here probably results from salt deposition on the molecules and TMV and/ or breakdown of complexes. Since we have not shown that the errors arising in the preparation of the specimens are random, we feel that it is not valid to quote a standard error of the mean <SD/V’&).
For B. contrurium, a member of the Melongenidae family of gastropods, the light scattering molecular weight of the hemocyanin ranged from 9 to 21 x lo6 Da depending on the protein concentration (Hersk-

STEM INVESTIGATION OF MOLLUSCAN HEMOCYANINS 227
ovits et al., 1989b). Ultracentrifugal analysis of the hemocyanin showed a complex pattern of at least seven components with sedimentation coefficients from 105 S to 220 S. While one tentatively correlates these components with di- to octadecameric species, STEM analysis of the side-views of the particles has unambiguously established the molecular masses of the individual multimeric species (Table II). The masses of 8.89 and 13.20 MDa obtained for the di- and tridecameric forms are close to the values for the homogeneous types (Table I). The STEM data also reveal that each decamer has virtually the same molecular mass, suggesting that no protein (or possibly carbohydrate) was lost and that no addi- tional protein components were added which pro- moted the polymerization. This is apparent from the relatively narrow ranges (from 4.3 X lo6 to 4.47 X 106) of the calculated molecular masses per decamer (see last column of Table II).
The mean length per decamer for the B. contra- rium hemocyanin of 17.0 + 0.9 nm is lower than the value of 19 nm cited by van Bruggen et al. (1981) for the length of the gastropodan decameric unit in neg- atively stained specimens. However, the lengths of the unstained particles are expected to differ from those obtained with negatively stained specimens because the particles are not embedded in stain, and hence shrinkage may occur during the freeze-drying process.
While the primary advantage of STEM lies in the measurement of the masses of individual molecules, visual examination of the various assemblies shows how the decameric units are organized within them. Although the images of the freeze-dried specimens are not as detailed as those of negatively stained specimens, features of the mass distribution such as cavities are recognizable in many specimens (see Figs. 1, 3, and 5). The unique arrangement of the decamers in the longer hemocyanin particles, best seen in the images of the hemocyanin of the moon snail, L. heros (Figs. 3 and 41, appears to be the addition of half-molecules (decamers) to a “nucleating” didecamer. This arrangement is also seen in the longer assemblies of B. contrurium (Figs. 5 and 6) as well as in the hemocyanins of other mem- bers of the Melongenidae family. These results have been described elsewhere (Hamilton et al., 1989; Herskovits et al., 1989a).
As to the question of what promotes or limits the association of the basic decamers into multiunit as- semblies, we have no definitive answer. Our mass measurements of the multidecameric hemocyanin of B. contrurium indicate a uniform mass increment and set an upper limit of +lOO kDa for any piece that is cleaved or added in its assembly. Mellema and Klug (1972) suggested that there might be a cap of some kind at the collar end of the didecameric hemocyanin of K. kelletiu. van Breemen et al. (1975)
showed that limited proteolysis led to the formation of long tubular particles from We&r pom.&iu hemo- cyanin particles that do not normally form particles larger than didecamers. Differences in the chemical nature of the collar-ends of the decamers, such as strong specific binding sites for bridging divalent ions, or in the carbohydrate moieties, may account for the diversity of particle sizes.
STEM offers the ultimate tool for characterizing hemocyanins of animals whose hemocyanins have not been studied because of the animal’s small size or its unavailability in sufficient quantity for isola- tion, among other reasons. For example, STEM mea- surements showed that the particle mass of the re- cently discovered hemocyanin of the primitive bi- valve, Yoldiu Zimatula, (Morse et al., 1986) is very close to the didecameric hemocyanins of gastropods. Light-scattering measurements confirmed this re- sult when larger amounts of the purified hemocya- nin became available (Herskovits et al., 1990).
We thank Grace G. Shiue for grid and sample preparation for electron microscopy and Martha Simon and Frank Hito for STEM operation. We thank James F. Hainfeld for helpful advice.
REFERENCES
ELLXRTON, H. D., ELLERTON, N. F., AND ROBINSON, H. A. (1983) Prog. Biophys. Mol. Biol. 41, 143-248.
F~R~INFITI, P. S., WALL, J. S., HAMILMN, M. G., AND HER@KO- VITS, T. T. (1988) in BAILEY, G. W. (Ed.), Proceedings of the 46th Annual Meeting of the Electron Microscopy Society of America, pp. 162163, San Francisco Press Inc., San Francisco.
HAMILTON, M. G., RODRIGUEZ, R. R., HERSKOVITS, T. T., AND WALL, J. S. (1989) in BAILEY, G. W. (Ed.), Prowedings of the 47th Annual Meeting of the Electron Microscopy Satiety of America, pp. 248-249, San Francisco Press Inc., San Francisco.
HER~KO~ITS T. T. (1988) Comp. B&hem. Physiol. 918,597-611. HER~KOV~ T. T., BLAKE, P. A., GONZALEZ, J. A., HAMILTON,
M. G., AND WALL, J. S. (1989a) Comp. B&x&m. Physiol., 94B, 415-421.
HERSKOV~ T. T., BLAKE, P. A., AND HAMILTON, M. G. (1988) Comp. B&hem. Physiol. 99B, 869-874.
HERSKOVITS T. T., COUSINS, C. J., HAMILTON, M. G., AM) WALL, J. S., (1990) in PREAUX, G., and LONT& R. @Is.), Invertebrate Dioxygen Carriers, in press, The Leuven University Press, Leuven, Belgium.
HERSKOVITS T. T., GUZMAN, A. E., AND HAMILTON, M. G. (1989b) Comp. Biochem. Physiol. 92B, 181-187.
HERSKOVIT~ T. T., AND HAMILTON, M. G. (1987a) Arch. Biochem. Biophys. 258,607-614.
HERSKOVITS T. T., AND HAMILTON, M. G. 11987h) Camp. Bio- chem. Physid. 88B, 127-132.
HERSKOVITS T. T., MAZZELLA, L. J., AND VILLAN~JEVA, G. B. (1985) Biochemistry 24, 38623870.
LAMY, J., LAMY, J. N., LECLERC, M., CoNcpIN, S., MILLER, K. I., AND VAN HOLDE, K. E. (1986) in LINZEN, B. (Ed.). Invertebrate Oxygen Carriers, pp. 231-234, Springer-Verlag, Berlin.
MAKINO, N. (1971) J. Biochem. Uupun) 70, 149-155.
MELLEMA, J. E., AND KLUG, A. (1972) Nature rlondon) 239, 14&-150.

228 HAMILTON ET AL.
MORSE, M. P., MEYHOFER, E., Om, J. J., ANLI KUZIRIAN, A. M. (1986) Science 231, 1302-1304.
Q~I’ITER, S., WA’ITS, L. A., CROSBY, C., AND ROXBY, R. (1978) J. Bid. Chem. 253, 525430.
SENOZAN, N. M., LANDRUM, J., BONAVENTURA, J., AND BON- AVENTURA, C. (1981) in LAMY J., AND LAMY, J. (Eds.), Inver- tebrate Oxygen-Binding Proteins: Structure, Active Sites and Function, pp. 703-716, Dekker, New York/BaseI.
STEVEN, A, C., HAINFELD, J. F., WALL, J. S., AND STEER, C. J. (1983) J. Cell Biol. 97, 1714-1723.
SVEDBERG, T., AND HEDENIUS, A. (1934) Biol. Bull. 66,191-223. VAN BREEMEN, J. F. L., SCHUURHUIS, G. J., AND VAN BRUGGEN,
E. F. J. (1977) in BANNISTER, J. V. (Ed.), Structure and Func- tion of Hemocyanin, pp. 122-127, Springer-Verlag, Berlin.
VAN BREEMEN, J. F. L., WICHJZRTJES, T., MUUER, M. F. J., VAN
DRIEL, R., AND VAN BRUGGEN, E. F. J. (1975) Eur. J. Biochem. 60, 129-135.
VAN BRUGGEN, E. F. J., SCHIJ?TER, W. G., VAN BREEMEN, J. F. L., BIJLHOLT, M. M. C., AND WICHERTJES, T. (1981) in HARRIS, J. R. (Ed.), Electron Microscopy of F’roteins, Vol. 1, pp. 2-38, Academic Press, New York.
VAN HOLDE, K. E., AND MIUER, K. I. (1982) Q. Reu. Biophys. 15, 1-129.
WALL, J. S. (1988) in BAILEY, G. W. (Ed.), Proceedings of the 46th Annual Meeting of the Electron Microscopy Society of America, pp. 236-237, San Francisco Press Inc., San Francisco.
WALL, J. S., ANLI HAINFELD, J. F. (1986) Annu. Reu. Biophys. Biophys. Chem. 15, 355-376.
WICHERTJES, T., GIELENS, C., SCHUTTER, W. G., PREAUX, G., LONTIE, R., AND VAN BRUGGEN, E. F. J. (1986) Biochim. Bio- phys. Acta 872, 183-194.


















