
Resistance to a non-immunogenic tumor, induced by Corynebacterium parvum or Listeria monocytogenes,...
4
Int. J. Cancer: 46, 687-690 (1990) 0 1990 Wiley-Liss, Inc. RESISTANCE TO A NON-IMMUNOGENIC TUMOR, INDUCED BY Publication of the International Union Against Cancer Publication de I'Union lnternationale Contre le Cancer CORYNEBACTERIUM PARVUM OR LISTERIA MONOCYTOGENES, IS ABROGATED BY ANTI-INTERFERON Robert KELLER'.~, Ruth KEIST',Thomas P. LEI ST*^^ and Peter H. VAN DER MEIDE3 LImmunobiology Research Group, Institute for Immunology and Virology, University of Zurich, Zurich; 2Division of Experimental Pathology, Department of Pathology, University Hospital, Zurich, Switzerland; and 3The Primate Center TNO, Rijswijk, The Netherlands. The complex processes that determine the outcome of the interaction of tumor and host were explored in the opera- tionally simple and reproducible rat D- I2 ascites tumor model. Animals exhibit weak spontaneous resistance against this tumor that is not augmented by repeated inoculation, by various routes, of irradiated syngeneic D- I2 tumor cells, but considerably enhanced after local administration of heat- killed Corynebacteriurn parvurn (CP) or Listeria rnonocyto- genes (LM) organisms. Inoculation of conventional or mono- clonal anti-rat IFNy antibodies into the same compartment did not affect spontaneous tumor resistance, but largely ab- rogated the tumor-protective effect triggered by CP or LM. Our findings support the concept that IFNy, produced by T cells in the course of the specific immune response raised against immunogenic micro-organisms, is able to enhance and to maintain local tumor resistance and thus to strengthen the capacity of the host to cope with a non-immunogenic tumor. Cancer biotherapy is based on the thesis that the immune system, which protects the host from bacterial, viral, or fungal infection, might also be able to eliminate tumor cells in a similar way. Accordingly, live, attenuated, or killed facultative intracellular pathogens, known to augment immunity to infec- tious agents, were among the earliest factors introduced in cancer immunotherapy. Such microbial agents, particularly BCG and CP, are capable of inducing anti-tumor activity in defined animal models and in some solid human tumors (Laucius et al., 1974; Scott, 1974; Sadler and Castro, 1978; Herberman, 1980; Woodruff, 1980; Morales and Ersil, 1982). Such treatments may also be effective in autochthonous tumors without detectable immunogenicity, suggesting that non- specific mechanisms, as mediated by activated MP, NK and/or LAK cells, are dominant (Laucius et al., 1974; Scott, 1974; Sadler and Castro, 1978; Oldham, 1987). The present work is a further attempt to analyze the complex processes involved in the interaction of tumor and host in vivo, utilizing the opera- tionally simple and reproducible DA rat D-12 ascites tumor model (Keller, 1976, 1977, 1980). MATERIAL AND METHODS Animals Inbred DA (RTla) rats weighing 120-180 g, raised locally and maintained under conventional conditions, were utilized. Tumor The D-12 tumor, induced by administration of dimethyl- benz(a)anthracene to female DA rats, has been adapted to growth in ascites form (Keller, 1977, 1980). D-12 tumor cells were continuously passaged in vivo in ascites form. Tumor cells harvested in the exponential phase of growth, i.e., before the ascites fluid became hemorrhagic, were suspended in PBS and lo2 up to 5 x lo3 D-12 cells/animal were injected i.p., observing the necessary precautions to keep the number of tumor cells deposited along the injection channel as low as possible. Ascites tumor growth was assessed by the death of the animal and confirmation of progressive local tumor growth. The large majority of control animals died within 25 days of TABLE I - REPEATED LOCAL IMMUNIZATION OF ANIMALS WITH IRRADIATED D-12 TUMOR CELLS DID NOT ENHANCE RESISTANCE TO LIVE D-12 TUMOR CELLS Mean survival in days (number of animals surviving) Number of live D-12 cells inoculated i.p. on day 0 Conmls Immunized animals' 5 x 102 103 5 x 103 22.9 (1) 16.2 (0) 15.1 101 16.2 (0)2 16.0 (0) 13.6 10)z 10 animalslgroup.-'Immunization schedule: animals were inoculated i.p. with 3 x lo7 irradiated D-12 cells each, 4 times at intervals of 4 weeks. Challen e with live D-12 cells i.p. was carried out 3 weeks after the last immunization.- These values were statistically significantly lower than those of the corresponding controls 5 (p < 0.001). tumor-cell challenge; animals surviving 50 days were killed. For statistical evaluation, survival of 60 days was considered for animals exhibiting no macroscopic evidence of tumor growth. For immunization, D- 12 ascites tumor cells suspended in PBS were irradiated with 10,000 rad from a Cobalt60 source. Statistical evaluation was made utilizing the Mann-Whitney U-test. Assay of IFN activity in peritoneal fluid Control rats and animals that had been inoculated with CP (mostly on day -7), were injected with 5 ml warm PBS, and peritoneal fluid was harvested after brief, careful massage. After 10-fold concentration of the samples, IFN activity was assayed in mouse L929 cells challenged with vesicular stoma- titis virus (Keller et al., 1987). Reagents Microorganisms. CP (ATCC 69 16) and LM organisms were selected from a single colony on solid brain-heart infusion agar (Difco, Detroit, MI) and cultured for 3-4 days in liquid brain- heart infusion (Difco) before harvesting by centrifugation; after washing twice in PBS, the organisms were heat-killed (240 min at 60°C), stored at 4"C, and used within 2-3 weeks. Usu- ally, 3 mg heat-killed organisms per animal were injected i.p. 4T0 whom correspondence and reprint requests should be sent, at the Immunobiology Research Group, Institute for Immunology and Virology, University of Zurich, Birchstrasse 95, CH-8050 Zurich, Switzerland. 'Present address: Department of Microbiology and Immunology, UCLA School of Medicine, Los Angeles, CA, USA. Abbreviations: BCG, bacille Calmette-Gutrin; BMMP, bone- marrow-derived mononuclear phagocytes; CP, Corynebacterium purvurn; IFN, interferon; LM, Listeriu monocytogenes; MAb, monoclonal anti- body; MP, macrophages; NU, neutralizing units; PBS, phosphate-buffered saline. Received May 18, 1990 and in revised form June 20, 1990.
-
Upload
robert-keller -
Category
Documents
-
view
215 -
download
3
Transcript of Resistance to a non-immunogenic tumor, induced by Corynebacterium parvum or Listeria monocytogenes,...

Int. J . Cancer: 46, 687-690 (1990) 0 1990 Wiley-Liss, Inc.
RESISTANCE TO A NON-IMMUNOGENIC TUMOR, INDUCED BY
Publication of the International Union Against Cancer Publication de I'Union lnternationale Contre le Cancer
CORYNEBACTERIUM PARVUM OR LISTERIA MONOCYTOGENES, IS ABROGATED BY ANTI-INTERFERON Robert KELLER'.~, Ruth KEIST', Thomas P. LEI ST*^^ and Peter H. VAN DER MEIDE3 LImmunobiology Research Group, Institute for Immunology and Virology, University of Zurich, Zurich; 2Division of Experimental Pathology, Department of Pathology, University Hospital, Zurich, Switzerland; and 3The Primate Center TNO, Rijswijk, The Netherlands.
The complex processes that determine the outcome of the interaction of tumor and host were explored in the opera- tionally simple and reproducible rat D- I 2 ascites tumor model. Animals exhibit weak spontaneous resistance against this tumor that is not augmented by repeated inoculation, by various routes, of irradiated syngeneic D- I 2 tumor cells, but considerably enhanced after local administration of heat- killed Corynebacteriurn parvurn (CP) or Listeria rnonocyto- genes (LM) organisms. Inoculation of conventional or mono- clonal anti-rat IFNy antibodies into the same compartment did not affect spontaneous tumor resistance, but largely ab- rogated the tumor-protective effect triggered by CP or LM. Our findings support the concept that IFNy, produced by T cells in the course of the specific immune response raised against immunogenic micro-organisms, is able to enhance and to maintain local tumor resistance and thus to strengthen the capacity of the host to cope with a non-immunogenic tumor.
Cancer biotherapy is based on the thesis that the immune system, which protects the host from bacterial, viral, or fungal infection, might also be able to eliminate tumor cells in a similar way. Accordingly, live, attenuated, or killed facultative intracellular pathogens, known to augment immunity to infec- tious agents, were among the earliest factors introduced in cancer immunotherapy. Such microbial agents, particularly BCG and CP, are capable of inducing anti-tumor activity in defined animal models and in some solid human tumors (Laucius et al . , 1974; Scott, 1974; Sadler and Castro, 1978; Herberman, 1980; Woodruff, 1980; Morales and Ersil, 1982). Such treatments may also be effective in autochthonous tumors without detectable immunogenicity, suggesting that non- specific mechanisms, as mediated by activated MP, NK and/or LAK cells, are dominant (Laucius et al . , 1974; Scott, 1974; Sadler and Castro, 1978; Oldham, 1987). The present work is a further attempt to analyze the complex processes involved in the interaction of tumor and host in vivo, utilizing the opera- tionally simple and reproducible DA rat D-12 ascites tumor model (Keller, 1976, 1977, 1980).
MATERIAL AND METHODS
Animals Inbred DA (RTla) rats weighing 120-180 g, raised locally
and maintained under conventional conditions, were utilized.
Tumor The D-12 tumor, induced by administration of dimethyl-
benz(a)anthracene to female DA rats, has been adapted to growth in ascites form (Keller, 1977, 1980). D-12 tumor cells were continuously passaged in vivo in ascites form. Tumor cells harvested in the exponential phase of growth, i . e . , before the ascites fluid became hemorrhagic, were suspended in PBS and lo2 up to 5 x lo3 D-12 cells/animal were injected i.p., observing the necessary precautions to keep the number of tumor cells deposited along the injection channel as low as possible. Ascites tumor growth was assessed by the death of the animal and confirmation of progressive local tumor growth. The large majority of control animals died within 25 days of
TABLE I - REPEATED LOCAL IMMUNIZATION OF ANIMALS WITH IRRADIATED D-12 TUMOR CELLS DID NOT ENHANCE RESISTANCE TO LIVE
D-12 TUMOR CELLS
Mean survival in days (number of animals surviving) Number of live D-12
cells inoculated i.p. on day 0 Conmls Immunized animals'
5 x 102 1 0 3
5 x 1 0 3
22.9 ( 1 ) 16.2 (0) 15.1 101
16.2 (0)2 16.0 (0) 13.6 10)z
10 animalslgroup.-'Immunization schedule: animals were inoculated i.p. with 3 x lo7 irradiated D-12 cells each, 4 times at intervals of 4 weeks. Challen e with live D-12 cells i.p. was carried out 3 weeks after the last immunization.- These values were statistically significantly lower than those of the corresponding controls
5 (p < 0.001).
tumor-cell challenge; animals surviving 50 days were killed. For statistical evaluation, survival of 60 days was considered for animals exhibiting no macroscopic evidence of tumor growth. For immunization, D- 12 ascites tumor cells suspended in PBS were irradiated with 10,000 rad from a Cobalt60 source. Statistical evaluation was made utilizing the Mann-Whitney U-test. Assay of IFN activity in peritoneal fluid
Control rats and animals that had been inoculated with CP (mostly on day -7), were injected with 5 ml warm PBS, and peritoneal fluid was harvested after brief, careful massage. After 10-fold concentration of the samples, IFN activity was assayed in mouse L929 cells challenged with vesicular stoma- titis virus (Keller et al., 1987).
Reagents Microorganisms. CP (ATCC 69 16) and LM organisms were
selected from a single colony on solid brain-heart infusion agar (Difco, Detroit, MI) and cultured for 3-4 days in liquid brain- heart infusion (Difco) before harvesting by centrifugation; after washing twice in PBS, the organisms were heat-killed (240 min at 60°C), stored at 4"C, and used within 2-3 weeks. Usu- ally, 3 mg heat-killed organisms per animal were injected i.p.
4T0 whom correspondence and reprint requests should be sent, at the Immunobiology Research Group, Institute for Immunology and Virology, University of Zurich, Birchstrasse 95, CH-8050 Zurich, Switzerland.
'Present address: Department of Microbiology and Immunology, UCLA School of Medicine, Los Angeles, CA, USA.
Abbreviations: BCG, bacille Calmette-Gutrin; BMMP, bone- marrow-derived mononuclear phagocytes; CP, Corynebacterium purvurn; IFN, interferon; LM, Listeriu monocytogenes; MAb, monoclonal anti- body; MP, macrophages; NU, neutralizing units; PBS, phosphate-buffered saline.
Received May 18, 1990 and in revised form June 20, 1990.
688 KELLER ET AL.
TABLE 11 - ANTI-IFNy MAB DOES NOT AFFTCT SPONTANEOUS TUMOR RESISTANCE MANIFESTED AGAINST THE D-12 ASCITES TUMOR
Size of tumor-cell inoculum (d 0) Treatment of animals
102 5 x 102 10'
None (controls) 23 ( k5 ) 15 (k2) 13 (52 )
Anti-rat IFNy MAb 10,OOO NU i.p. d - 1 18 ( + 5 ) 16 ( 5 3 ) 13 (k2) Carrageenan 5 mg i.p. d - 1 10 (21)' ND ND Values are derived from a single experiment including 10 rats per group and represent the mean survival in days ( 2 s ~ ) . 'This value is statistically significantly different
Sheep anti-rat IFNy 10,000 NU i.p. d - 1 18 (+4) 15 (k2) 14 (&2)
from the corresponding control value (p < 0.001).
Antibodies to rat IFNy Polyclonal sheep antiserum. Repeated intramuscular inocu-
lation of concentrated purified rat recombinant IFNy (rrIFNy; Dijkema et a l . , 1985) with incomplete Freund's adjuvant into sheep resulted in polyclonal antiserum with neutralizing activ- ity of approximately 100,000 NU/ml (sheep 219; Keller et al., 1987). The antibody was purified by ammonium sulphate pre- cipitation. Normal preimmune serum derived from the same sheep was heat-inactivated (30 min at 56°C) before use.
Monoclonal DB-1 antibody against rat recombinant IFNy (rrIFNy) was obtained as described (Dijkema et a l . , 1985); the affinity-purified IgG antibody (neutralizing activity approx. 6,000 NU/mg protein) was free of IL-1 or TNFu activity.
Carrageenan (Seakem 21; Marine Colloids, Springfield, NJ) was dissolved in PBS by heating to 100°C for 15 min.
RESULTS
Repeated inoculation with irradiated D-12 cells (3-5 times lo7 D-12 cells each at intervals of 3 to 4 weeks either s.c., i.m., or i.p.) had no positive effect on resistance to i.p. chal- lenge with live D-12 cells. Results of a typical experiment, in which ascites tumor growth was slightly enhanced after immu- nization, are presented in Table I.
Earlier findings have established that one single i.p. injec- tion into the normal rat of so-called anti-macrophage agents (e .g . silica particles, carrageenan) shortly before tumor-cell
' I "1.- 15 20 25 30 35
DAYS AFTER CHALLENGE
4.1;> r 8 . .%.,i.. , 1 ' 7 -5.. I r
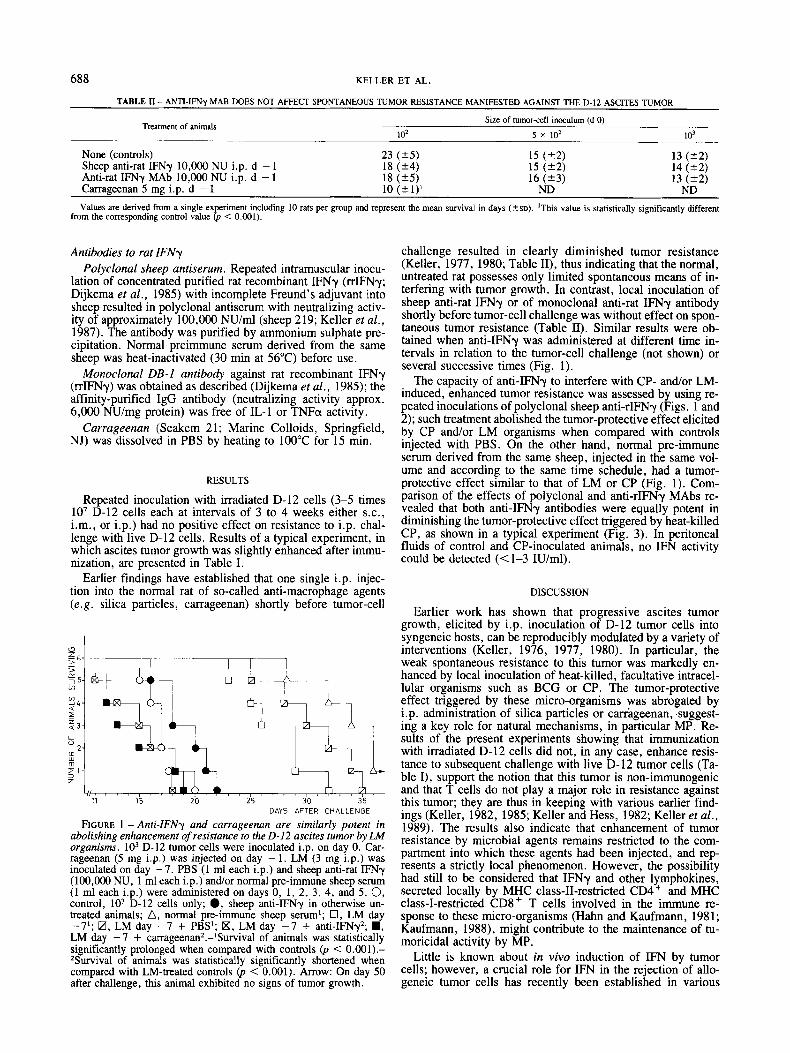
FIGURE 1 - Anti-IFNy and carrageenan are similarly potent in abolishing enhancement of resistance to the 0 - 1 2 ascites tumor by L,M organisms. lo3 D-12 tumor cells were inoculated i.p. on day 0. Car- rageenan (5 mg i.p.) was injected on day - l . LM (3 mg i.p.) was inoculated on day - 7. PBS (1 ml each i.p.) and sheep anti-rat IFNy (100,000 NU, 1 ml each i.p.) and/or normal pre-immune sheep serum (1 ml each i.p.) were administered on days 0, 1 , 2, 3, 4, and 5. 0, control, lo3 D-12 cells only; 0, sheep anti-IFNy in otherwise un- treated animals; A , normal pre-immune sheep serum1; 17, LM day -7l; m, LM day -7 + PBS'; IXI, LM day -7 + anti-IFNy2; ., LM day -7 + carrageenan2.-lSurvival of animals was statistically significantly prolonged when compared with controls (p < 0.001).- *Survival of animals was statistically significantly shortened when compared with LM-treated controls (p < 0.001). Arrow: On day 50 after challenge, this animal exhibited no signs of tumor growth.
challenge resulted in clearly diminished tumor resistance (Keller, 1977, 1980; Table 11), thus indicating that the normal, untreated rat possesses only limited spontaneous means of in- terfering with tumor growth. In contrast, local inoculation of sheep anti-rat IFNy or of monoclonal anti-rat IFNy antibody shortly before tumor-cell challenge was without effect on spon- taneous tumor resistance (Table 11). Similar results were ob- tained when anti-IFNy was administered at different time in- tervals in relation to the tumor-cell challenge (not shown) or several successive times (Fig. 1).
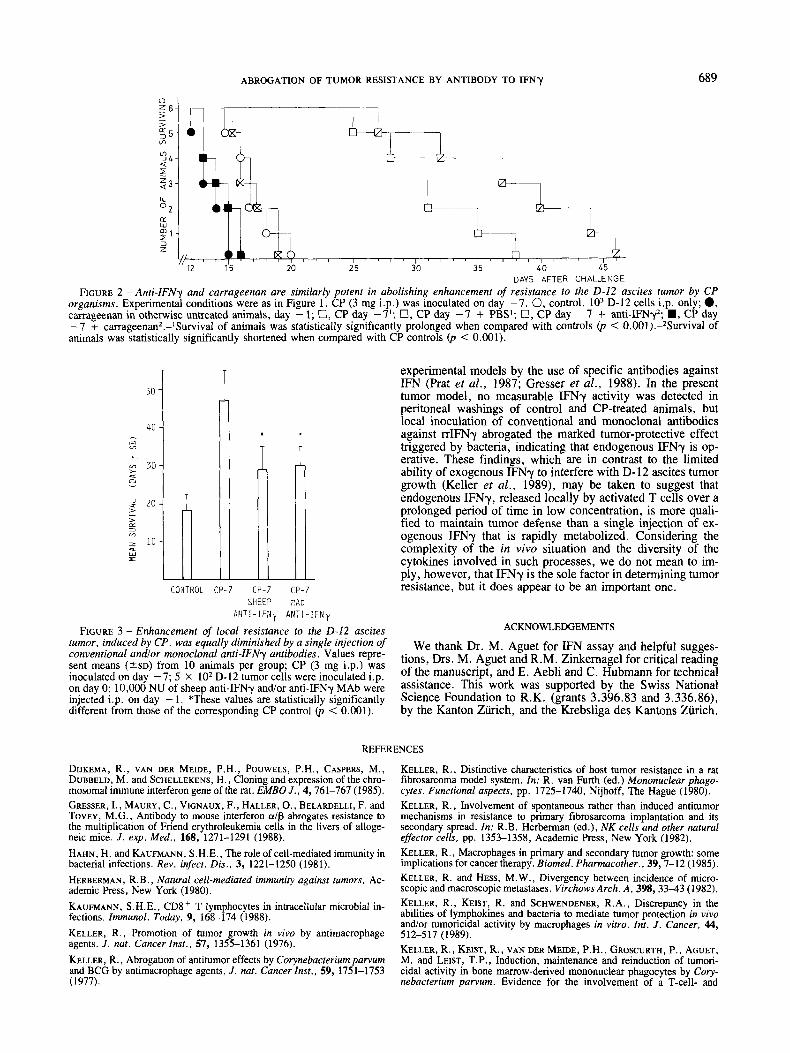
The capacity of anti-IFNy to interfere with CP- and/or LM- induced, enhanced tumor resistance was assessed by using re- peated inoculations of polyclonal sheep anti-rIFNy (Figs. 1 and 2); such treatment abolished the tumor-protective effect elicited by CP and/or LM organisms when compared with controls injected with PBS. On the other hand, normal pre-immune serum derived from the same sheep, injected in the same vol- ume and according to the same lime schedule, had a tumor- protective effect similar to that of LM or CP (Fig. 1). Com- parison of the effects of polyclonal and anti-rIFNy MAbs re- vealed that both anti-IFNy antibodies were equally potent in diminishing the tumor-protective effect triggered by heat-killed CP, as shown in a typical experiment (Fig. 3). In peritoneal fluids of control and CP-inoculated animals, no IFN activity could be detected (<1-3 IU/ml).
DISCUSSION
Earlier work has shown that progressive ascites tumor growth, elicited by i.p. inoculation of D-12 tumor cells into syngeneic hosts, can be reproducibly modulated by a variety of interventions (Keller, 1976, 1977, 1980). In particular, the weak spontaneous resistance to this tumor was markedly en- hanced by local inoculation of heat-killed, facultative intracel- lular organisms such as BCG or CP. The tumor-protective effect triggered by these micro-organisms was abrogated by i.p. administration of silica particles or carrageenan, suggest- ing a key role for natural mechanisms, in particular MP. Re- sults of the present experiments showing that immunization with irradiated D-12 cells did not, in any case, enhance resis- tance to subsequent challenge with live D-12 tumor cells (Ta- ble I), support the notion that this tumor is non-immunogenic and that T cells do not play a ma.jor role in resistance against this tumor; they are thus in keeping with various earlier find- ings (Keller, 1982, 1985; Keller and Hess, 1982; Keller et al . , 1989). The results also indicate that enhancement of tumor resistance by microbial agents remains restricted to the com- partment into which these agents had been injected, and rep- resents a strictly local phenomenon. However, the possibility had still to be considered that IFNy and other lymphokines, secreted locally by MHC class-11-restricted CD4+ and MHC class-I-restricted CD8+ T cells involved in the immune re- sponse to these micro-organisms (Hahn and Kaufmann, 1981; Kaufmann, 1988), might contribute to the maintenance of tu- moricidal activity by MP.
Little is known about in vivo induction of IFN by tumor cells; however, a crucial role for IFN in the rejection of allo- geneic tumor cells has recently been established in various
ABROGATION OF TUMOR RESISTANCE BY ANTIBODY TO IFNy 689
DAYS AFTER CHALLENGE
FIGURE 2 - Anti-IFNy and carrageenan are similarly potent in abolishing enhancement of resistance to the 0 - 1 2 ascites tumor by CP organisms. Experimental conditions were as in Figure 1. C P (3 mg i.p.) was inoculated on day -7. 0, control, lo3 D-12 cells i.p. only; 0, carrageenan in otherwise untreated animals, day - 1; 0, CP day -7l; 0, CP day - 7 + PBS; 0, CP day - 7 + anti-IFNyz; W , C P day - 7 + carrageenan2.-'Survival of animals was statistically significantly prolonged when compared with controls (p < O.OOl).-*Survival of animals was statistically significantly shortened when compared with C P controls (p < 0.001).
I I
I; - CONTROL CP-7 C P - 7 C P - 7
SHEEP mAb ANTI-IF14 y ANTI -1FNy
FIGURE 3 - Enhancement of local resistance to the D-12 ascites tumor, induced by CP, was equally diminished by a single injection of conventional andlor monoclonal anti-IFNy antibodies. Values repre- sent means ( ~ s D ) from 10 animals per group; C P (3 mg i.p.) was inoculated on day - 7; 5 X lo2 D-12 tumor cells were inoculated i.p. on day 0; 10,000 NU of sheep anti-IFNy and/or anti-IFNy MAb were injected i.p. on day - 1. *These values are statistically significantly different from those of the corresponding C P control (p < 0.001).
experimental models by the use of specific antibodies against IFN (ha t et al., 1987; Gresser er al., 1988). In the present tumor model, no measurable lFNy activity was detected in peritoneal washings of control and CP-treated animals, but local inoculation of conventional and monoclonal antibodies against rrIFNy abrogated the marked tumor-protective effect triggered by bacteria, indicating that endogenous IFNy is op- erative. These findings, which are in contrast to the limited ability of exogenous IFNy to interfere with D-12 ascites tumor growth (Keller er al., 1989), may be taken to suggest that endogenous IFNy, released locally by activated T cells over a prolonged period of time in low concentration, is more quali- fied to maintain tumor defense than a single injection of ex- ogenous IFNy that is rapidly metabolized. Considering the complexity of the in vivo situation and the diversity of the cytokines involved in such processes, we do not mean to im- ply, however, that IFNy is the sole factor in determining tumor resistance, but it does appear to be an important one.
ACKNOWLEDGEMENTS
We thank Dr. M. Aguet for IFN assay and helpful sugges- tions, Drs. M. Aguet and R.M. Zinkernagel for critical reading of the manuscript, and E. Aebli and C. Hubmann for technical assistance. This work was supported by the Swiss National Science Foundation to R.K. (grants 3.396.83 and 3.336.86), by the Kanton Ziirich, and the Krebsliga des Kantons Zurich.
REFERENCES
DUKEMA, R., VAN DER MEIDE, P.H., POUWELS, P.H., CASPERS, M., DUBBELD, M. and SCHELLEKENS, H., Cloning and expression of the chro- mosomal immune interferon gene of the rat. EMBO J . , 4,761-767 (1985). GRESSER, I., MAURY, C., VIGNAUX, F., HALLER, O., BELARDELLI, F. and TOVEY, M.G., Antibody to mouse interferon a/$ abrogates resistance to the multiplication of Friend erythroleukemia cells in the livers of alloge- neic mice. J . exp. Med., 168, 1271-1291 (1988). HAHN, H. and KAUFMANN, S.H.E., The role of cell-mediated immunity in bacterial infections. Rev. infect. Dis., 3, 1221-1250 (1981). HERBERMAN, R.B., Natural cell-mediated immunity against tumors, Ac- ademic Press, New York (1980). KAUFMANN, S.H.E., CD8+ T lymphocytes in intracellular microbial in- fections. fmmunol. Today, 9, 168-174 (1988). KELLER, R . , Promotion of tumor growth in vivo by antimacrophage agents. J. nut. Cancer Inst., 57, 1355-1361 (1976). KELLER, R., Abrogation of antitumor effects by Corynebacterium parvum and BCG by antimacrophage agents. J. nut. Cancer Inst., 59, 1751-1753 (1977).
KELLER, R., Distinctive characteristics of host tumor resistance in a rat fibrosarcoma model system. In: R. van Furth (ed.) Mononuclear phago- cytes. Functional aspects, pp. 1725-1740, Nijhoff, The Hague (1980). KELLER, R., Involvement of spontaneous rather than induced antitumor mechanisms in resistance to primary fibrosarcoma implantation and its secondary spread. In: R.B. Herberman (ed.), NK cells and other natural effector cells, pp. 1353-1358, Academic Press, New York (1982). KELLER, R., Macrophages in primary and secondary tumor growth: some implications for cancer therapy. Biomed. Pharmacother., 39,7-12 (1985). KELLER, R. and HESS, M.W., Divergency between incidence of micro- scopic and macroscopic metastases. Virchows Arch. A, 398 ,3343 (1982). KELLER, R., KEIST, R. and SCHWENDENER, R.A., Discrepancy in the abilities of lymphokines and bacteria to mediate tumor protection in vivo and/or tumoricidal activity by macrophages in vitro. Int. J . Cancer, 44,
KELLER, R., KEIST, R., VAN DER MEIDE, P.H., GROSCURTH, P., AGUET, M. and LEIST, T.P., Induction, maintenance and reinduction of tumori- cidal activity in bone marrow-derived mononuclear phagocytes by Cory- nebacterium parvum. Evidence for the involvement of a T-cell- and
512-517 (1989).

690 KELLER ET AL.
interferon-y-independent pathway of macrophage activation. J. Immunol., 138, 2366-2371 (1987). LAUCIUS, J.F., BODURTHA, A.J., MASTRANGELO, M.J. and CREECH, R.M., Bacillus Calmette-Guerin in the treatment of neoplastic diseases. J. Reticuloendothel. SOC., 16, 347-373 (1974). MORALES, A. and ERSIL, A., Adjuvant BCG immunotherapy in the pro- phylaxis and treatment of noninvasive bladder cancer. In: W.D. Teny and S.A. Rosenberg (eds.), lmmunotherapy of human cancer, p. 301, Elsevier/ North Holland, New York (1982). OLDHAM, R.K., Principles of cancer biotherapy. Raven Press, New York (1987).
F’RAT, M., BRETTI, S., AMEDEO, M., LANDOLFO, S. and COMOGLIO, P.M., Monoclonal antibodies against murine IFN-y abrogate in vivo tumor immunity against RSV-induced murine sarcomas. J. Zmmunol., 138, 453W533 (1987). SADLER, T.E. and CASTRO, J.E., Experimental non-specific immunother- apy. In: J.E. Castro (ed.), Immunological aspects of cancer, p. 357, MTP Press, Lancaster (1978). SCOTT, M.T., Corynebacterium parvum as an immunotherapeutic anti- cancer agent. Sen. Oncol., 1, 367-378 (1974). WOODRUFF, M.F.A., The interaction of cancer and host. Grune and Strat- ton, New York (1980).


















