
Nephrin and CD2AP Associate with Phosphoinositide 3-OH Kinase
Upload
vuongtuongCategory
view
229download
2
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1987 by The American Society of Biological Chemists, Inc.
Vol. 262, No. 3, Issue of January 25, pp. 1105-1110 1987 Printed in c?, S. A.
Regulation of Phosphoinositide Phosphorylation in Swiss 3T3 Cells Stimulated by Platelet-derived Growth Factor*
(Received for publication, July 17, 1986)
Marnie Lynn MacDonald, Katherine F. Mack, and John A. Glomset From the Howard Hughes Medical Institute Research Laboratories and the Department of Biochemistry, University of Washington, Seattle, Washington 98195
The regulation of phosphoinositide phosphorylation was studied in Swiss 3T3 cells that were stimulated by platelet-derived growth factor (PDGF). Studies with intact cells showed that the mitogen increased the in- corporation of “P into phosphatidylinositol (PtdIns), phosphatidylinositol 4-phosphate (PtdIns-P), and phosphatidylinositol 4,5-bisphosphate (PtdIns-Pa) during the cell cycle, with distinct peaks of incorpo- ration for all three phosphoinositides after 1 h, and for PtdIns and PtdIns-P2 after 20 h. Direct measurements of the activities of PtdIns kinase and PtdIns-P kinase in freeze-thawed cells revealed that the activity of PtdIns kinase was rate-limiting for the synthesis of PtdIns-P2. Maximal activities of PtdIns kinase and PtdIns-P kinase, with exogenous substrates, were un- changed during the 1st h of PDGF treatment, but dou- bled during the next 24 h. The increase in PtdIns kinase activity began within 2-4 h, exceeded the in- crease in cell protein, and was abolished by cyclohex- imide, which suggests that the enzyme was induced specifically in response to PDGF. The increase in ac- tivity of PtdIns-P kinase paralleled the increase in cell protein. Dose-response curves for PDGF showed that the activities of PtdIns kinase and PtdIns-P kinase at 24 h increased in proportion to the extent of mitogenic stimulation of the cells. Our results support the conclu- sion that the activities of PtdIns kinase and PtdIns-P kinase increase in response to PDGF, but only after several hours of cell cycle traverse.
PDGF’ is a mitogen for 3T3 cells and other cells of mes- enchymal origin (see Ref. 1 for review). When PDGF is added to quiescent 3T3 cells, the cells are stimulated to traverse the cell cycle. DNA synthesis begins within 20 h, and mitosis occurs within 30-40 h after the addition of PDGF (2).
One of the first effects of PDGF in 3T3 cells is to stimulate the turnover of phosphatidylinositol and the formation of diacylglycerol (3) and inositol trisphosphate (4, 5), two com- pounds that are thought to act as second messengers for signal
* This work was supported by the Howard Hughes Medical Insti- tute and by Grant HL18645 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviations used are: PDGF, platelet-derived growth factor; PtdIns, phosphatidylinositol; PtdIns-P, phosphatidylinositol 4-phos- phate; PtdIns-P2, phosphatidylinositol 4,5-bisphosphate; Ins-P, ino- sitol monophosphate; Ins-Pp, inositol bisphosphate, Ins-P3, inositol trisphosphate; PDS, plasma-derived serum; MOPS, 3-(N-morpho- 1ino)propanesulfonic acid. TCA, trichloroacetic acid, DTT, dithio- threitol; the term “phosphoinositide” is used to refer in general to PtdIns, PtdIns-P, and PtdIns-P,.
transduction (see Ref. 6). Studies from our laboratory showed that both the breakdown and resynthesis of PtdIns were enhanced as early as 2 min after the addition of PDGF to quiescent 3T3 cells (3). Net breakdown of PtdIns continued for at least 60 min, while levels of diacylglycerol peaked within 10-20 min after the addition of PDGF. Berridge et al. (4) and Hasegawa-Sasaki (5) showed that, in cells prelabeled with radioactive inositol or phosphorus, the levels of PtdIns, PtdIns-P, and PtdIns-P, were decreased, and levels of Ins-P, Ins-P,, and Ins-Pa were increased at 5 min (4) or 10 min (5) after PDGF was added. The effect of PDGF on incorporation of radioactive precursors into PtdIns-P or PtdIns-Pz has not been studied.
Little is known about the mechanism by which PDGF stimulates the turnover of PtdIns. Although Berridge et al. (4) observed apparent breakdown of PtdIns, PtdIns-P, and PtdIns-P, in response to PDGF, they found that the propor- tion of label in the polyphosphoinositides was increased rel- ative to PtdIns; they suggested that this shift might be due to enhanced phosphorylation of PtdIns to PtdIns-P and PtdIns- P,. According to this scheme, PtdIns kinase and PtdIns-P kinase would be activated, PtdIns would be converted to PtdIns-P,, and the latter compound would be hydrolyzed by phospholipase C to produce Ins-Pa and diacylglycerol. The increased production of Ins-P, and Ins-P would then be explained by phosphomonoesteratic cleavage of Ins-Pa to the less phosphorylated forms.
Activation of PtdIns kinase or PtdIns-P kinase during the PtdIns response could occur by several mechanisms, including release of product inhibition (cf. Refs. 7 and 8) or covalent modification such as phosphorylation or dephosphorylation. Interest in the phosphoinositide kinases as possible targets for regulation by protein phosphorylation has been fueled by reports that the incorporation of phosphorus into PtdIns-P and PtdIns-Pz is enhanced in cells transformed by viruses that code for protein tyrosine kinases (9, 10) and that the activities of PtdIns kinase and PtdIns-P kinase are increased in these cells (11, 12). Moreover, Sugimoto et al. (9) and Macara et al. (10) reported that purified protein tyrosine kinases of viral origin could directly phosphorylate PtdIns and/or PtdIns-P. These authors concluded that the virally encoded protein tyrosine kinases might be responsible, either directly or indirectly, for the increased phosphorylation of phosphoinositides in transformed cells. Furthermore, several studies have shown that levels of inositol trisphosphate and diacylglycerol are elevated in a variety of transformed cell lines (13-15). Taken together, these reports led to the specu- lation that viral transformation might be initiated through an increase in phosphorylation of PtdIns and PtdIns-P, resulting in an increase in the supply of Ins-Pa and diacylglycerol which then would act as second messengers to stimulate cell growth. By extension, some authors have proposed (10) that growth
1105

1106 Regulation of Phosphoinositide Phosphorylation in 3T3 Cells
factor receptors that are protein tyrosine kinases, such as the PDGF receptor (161, might directly phosphorylate phosphoi- nositides and thereby initiate the PtdIns response.
The objective of the present study was to determine if PtdIns kinase or PtdIns-P kinase were changed in response to PDGF. We studied the rates of phosphorylation of phos- phoinositides at early and late times throughout the cell cycle. The activities of PtdIns kinase and PtdIns-P kinase were measured directly with exogenous and endogenous substrates. The results show that PDGF causes a dose-dependent in- crease in the activities of PtdIns kinase and PtdIns-P kinase, but that these changes only occur after several hours of cell cycle traverse.
EXPERIMENTAL PROCEDURES AND RESULTS~
DISCUSSION
The regulation of the phosphoinositide kinases has been a subject of considerable debate in the literature. Based on the results of labeling studies, some authors have proposed that the kinases are activated, resulting in depletion of phosphati- dylinositol by conversion to polyphosphoinositides (24-26). In this scheme, the apparent breakdown of PtdIns involves phosphorylation of PtdIns to PtdIns-Pz, hydrolysis of PtdIns- Pz by phospholipase C, and breakdown of Ins-P3 to Ins-P. The breakdown of PtdIns-P2 might itself activate PtdIns-P kinase by releasing the enzyme from product inhibition, or, activation could occur by a direct mechanism such as protein phosphorylation. Regardless of mechanism, it has been sug- gested that activation of PtdIns kinase and PtdIns-P kinase would increase the supply of PtdIns-P2 for phospholipase C and thereby increase the production of Ins-P3 (10, 27).
Our results do not support this hypothesis. We found that the activities of PtdIns kinase and PtdIns-P kinase were unchanged during the first 60 min after PDGF addition, a time when there was extensive breakdown and resynthesis of PtdIns and PtdIns-P. In addition, the rates of phosphoryla- tion of membrane PtdIns and PtdIns-P declined during this period, probably as a result of depletion of the endogenous substrates of these enzymes. Although it remains possible that enzyme activation occurred but could not be measured under the conditions we used, we did not find any activation in other studies in which inhibitors were used to guard against the action of proteinases or phosphatases. Therefore, it seems unlikely that the kinases play a role in the enhanced produc- tion of inositol phosphates and diacylglycerol in response to PDGF. In addition, the extensive depletion of PtdIns and PtdIns-P that occurs during the early phase of hormone stimulation is not due to increased conversion to PtdIns-Pz and must be explained by another mechanism. The most likely explanation is that these compounds are hydrolyzed directly by phospholipase C. Direct action of phospholipase C on PtdIns would be consistent with the rapid and sustained formation of large amounts of Ins-P (data not shown) and would imply that the diacylglycerol that is generated4 is derived primarily from PtdIns. A similar conclusion was reached by Wilson and co-workers (28,29) who used a differ-
* Portions of this paper (including “Experimental Procedures,” “Results,” Table I, Figs. 1-6, and Footnote 3) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Be- thesda, MD 20814. Request Document No. 86M-2399, cite the au- thors, and include a check or money order for $7.60 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press. ‘ See Ref. 3 for the kinetics of diacylglycerol accumulation in Swiss
3T3 cells stimulated by PDGF.
ent approach to study other cell response systems. Although there was no activation of PtdIns kinase or
PtdIns-P kinase during the earliest phase of cell stimulation, the activities of both enzymes increased progressively as cells traversed the cell cycle. The increase in activity of PtdIns-P kinase paralleled the cell cycle-dependent rise in total cellular protein, whereas the activity of PtdIns kinase rose dispropor- tionately during GI and S phases of the cell cycle. The increase in PtdIns kinase activity during GI may be due to induction of this enzyme. The increase occurs at a time when dramatic changes are occurring in cell shape, microvillus formation, and cell attachment (18). While the mechanism of these changes remains to be determined, the enhanced synthesis of polyphosphoinositides via PtdIns kinase may be important for increased formation of plasma membrane at this time.
The observation that mitogenic stimulation of untrans- formed cells causes an increase in activity of enzymes of PtdIns metabolism may help to explain why these enzymes are elevated in transformed cells. Sugimoto and Erikson (12) studied cells transformed by a temperature-sensitive mutant of Rous sarcoma virus and found that shifting the cells from a nonpermissive to a permissive temperature resulted in a small but reproducible increase in the activities of PtdIns kinase and PtdIns-P kinase. These authors suggested that the protein tyrosine kinase encoded by Rous sarcoma virus might be responsible, either directly or indirectly, for the increased phosphorylation of phosphoinositides. We also found that the activities of PtdIns kinase and PtdIns-P kinase were elevated in extracts of cells that had been transformed by Rous sar- coma virus or by Moloney leukemia virus, but there was no evidence for direct phosphorylation of PtdIns or PtdIns-P by protein tyrosine kinases (11). Jackowski and co-workers re- cently showed that the activity of a GTP-dependent phospho- lipase C was enhanced in cells transformed by the v-fms or v- fes oncogenes (13).
Our studies show that the activities not only of PtdIns kinase and PtdIns-P kinase, but also of diacylglycerol kinase: phosphatidylinositol synthase? and phospholipase C,6 are in- creased in “growing” (PDGF-stimulated) cells as opposed to quiescent cells. Thus, the increased kinase and phospholipase activities in transformed cells may be a consequence of the altered growth parameters of transformation (see Ref. 30 for review) and may not be an initial event in the transformation process. Nevertheless, the established role of these enzymes in the production of second messengers suggests that changes in these enzymes during S phase might be important for cell signalling during the period of active synthesis of DNA.
REFERENCES
1. Ross, R., Raines, E. W., and Bowen-Pope, D. F. (1986) Cell 4 6 ,
2. Habenicht, A. J. R., Glomset, J. A., and Ross, R. (1980) J. Biol.
3. Habenicht, A. J. R., Glomset, J. A., King, W. C., Nist, C., Mitchell,
4. Berridge, M. J., Heslop, J. P., Imine, R. F., and Brown, K. D.
5. Hasegawa-Sasaki, H. (1985) Biochem. J. 232,99-109 6. Nishizuka, Y. (1984) Science 225 , 1365-1370 7. Van Rooijen, L. A. A., Rossowska, M., and Bazan, N. G. (1985)
8. Michell, R. H., Harwood, J. L., Coleman, R., and Hawthorne, J.
9. Sugimoto, Y., Whitman, M., Cantley, C., and Erikson, R. L.
155-169
Chem. 265,5134-5140
C. D., and Ross, €2. (1981) J. Biol. Chem. 256,12329-12335
(1984) Biochem. J. 222,195-201
Bwchem. Bwphys. Res. Commun. 126 , 150-155
N. (1967) Biochim. Biophys. Acta 144,649-658
(1984) Proc. Natl. Acad. Sci. U. S. A. 81,2117-2121
M. L. MacDonald, K. F. Mack, and J. A. Glomset, unpublished
R. Kouchalakos, personal communication. results.

10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
Regulation of Phosphoinositide Phosphorylation in 3T3 Cells 1107
Macara, I. G., Marinetti, G. V., and Balduzzi, P. C. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,2728-2732
MacDonald, M. L., Kuenzel, E. A., Glomset, J. A., and Krebs, E. G. (1985) Proc. Natl. Acad. Sci. U. S. A. 82,3993-3997
Sugimoto, Y., and Erikson, R. L. (1985) Mol. Cell. Bwl. 6,3194- 3198
Jackowski, S., Rettenmier, C. W., Sherr, C. J., and Rock, C. 0. (1986) J. Bwl. Chem. 261,4978-4985
Fleischman, L. F., Chahwala, S. B., and Cantley, L. (1986) Science 231,407-410
Kaplan, D. R., Whitman, M., Schaffhausen, B., Raptis, L., Gar- cea, R. L., Pallas, P., Roberts, T. M., and Cantley, L. (1986) Pmc. Natl. Acad. Sci. U. S. A. 83,3624-3628
Pike. L. J.. Bowen-Pow. D. F.. Ross. R.. and Krebs. E. G. (1983) . , J. Bwl. Chem. 258;9383-9390
. .
b i n e s . E. W.. and Ross, R. (1982) J. Biol. Chem. 257. 5154- . . . 5160'
and Ross, R. (1982) J. Cell BwL 95, 144-153
phys. Acta 710,332-340
Schmidt, R. A., Glomset, J. A., Wight, T. N., Habenicht, A. J. R.,
Marche, P., Koutouzov, S., and Meyer, P. (1982) Biochim. Bw-
MINIPRINT SECTION
Regulation of phosphoinositide phosphorylation in Swirr 3T3 cells stimulated by platelet-derived growth factor
M a r n ~ e Lynn MacDonald, Katherine F. Mack and John A. GlOmset
cilmol) and LnP1phosphorus (Hcl-free) wece fcom New England Nuclear, Materials- Iy-'*PlAlP ( 3 0 0 0 C i l m l ) , Imethyl-'Hlthymidine (6.7
ATP were from Sigma Chemical Co., St. Louis, BW. Bovine llver Ptdlns was from Boston, HA. PtdInS-P, PtdIns-P2, cycioheximide, dithiothreitol and sodium
Avanti Polac-Lipids 1 ° C . . Birmingham, UI. Pre-Coated CelluloSe plates (0.1 m thickness) f o r thin-layer Chromatoglaphy were from Ell Science. Cherry Hill. NJ. Chromatographically pure PDGF, prepared as described (17). was a gift
described 12). Other cell culture reaaentr were from Grand Island Biolo~ical from Bonnie Ashleman and O r . Russell Ross. Human PDS was prepared as
Co., Grand.Ibland, NY.
Cell Culture and Labelling- Swiss 3T3-Dl cells were maintained in culture and were made gulescent essentially a s described ( 3 ) . Cells were plated in 1.5 ml of medium in 35 m plastic plates (for biosynthetic
Dulbecc0'9 modification of Eagle's minimum essential medium that had been labelling) or in 7 ml Of medium in 100 m plates ( f o r enzyme measurements) in
supplomented with PDS at 51. Cells were fed after two day3 with fresh radium containing PDS and were used after 4-6 days. by which tlme they were quiescent a s assessed by a lack Of mitotic figures and by low incorporation Of
acetic acid; a final concentration Of 10 nglml in cell medium was used I'Hlthymidine into DNA. Stock solutions of PDGF were 1 mq/ml in 10 nv(
routinely to StimYlate quiescent cell*.
described previou~ly (10) by incubating cell5 in the presence Of H-thymidine for 3 h0Y.S during S phase of the cell cycle (21 to 24 hoYcs
Cells Stimulated the incorporation Of thymidine lnto DNA by a factor Of at after PDGF additlon). I n a l l experiments, the addition Of PDGF to quiescent
least forty-fold.
[Nethyl-'Hlthymidine incorporation into DNA w a s determined as
TO measure the incorporation of "P into inositol phospholipids in cells stimulated by PDCF, the cell medium was first replaced vlth low (0.05 Mt-phosphorus medium containing PDS. "P (50 "Cildish) either v a s added five minutes before the additlon Of PDGF (for studies of the first hoUc of the cell cycle) or was added 1 hour before the plater were harvested (for studies throughout 24 hours of the cell cycle). For cell harvesting, the medium was
on dry ice. Dishes were stored at -70 for later analysis. Cells were then removed. 1 ml of 1 0 1 triChlc.rOaCetiC :=id was added and each dish was Placed
thawed and were scraped from the dish w ~ t h a rubbec po1icem~D. The dish was rinsed with 1 ml of 10% trichloroacetic acid and the combined TCA extracts were centrlfuged. The pellet was washed twice with 5 1 TCA. LipIdr wece extracted from TCA pellet3 by sequential addition of 1.5 ml Of 0.1 N HCI and 5.6 ml of CH,OH/CHCL, (37:19). After 30 min, 1 . 9 ml each of CHC13 and 0.1 N ncl were added to form two phases. The upper phase was removed and the
48:47:3). Carrier Ptdlns, PtdIna-P and Ptdlnr-P, ( 4 0 pg each) were added lower phase was washed twice with upper phase (CH10H/H~O/CHC13.
wece essentldl for the migration Of ladioactive PtdIns-P and PtdInS-PI an to the washed lower phases (separate experiments Showed that carrier lipids
cellulo~e plates). Samples were dried under nitrogen and were then dissolved in a small volume Of CHC1,/CH,OHIH1O (75:25:2) and spotted on cellulo~e TLC plates. . Ascending chromatography vas carried Out in butmollacetic acidlwater (75:10:25) as described (19). Plates Were stained with Nile Blue dye. bands were excised and radioactivity was determined by liquid
enzyme activities were determined, cells were plated in 100 m dishes. Because it was not possible to quantztate a11 enzyme activities in one experiment. separate experiments were performed for each enzyme Studied. At least 3 olater of cells were harvested at each time DOint. and e n l m
Assay of Ptdlns kinase and Ptdlns-P kinase- In experiments in whlch
activitlis uere~irsayed i i duplicate f o r each cell &tract. At vacious tines after the addition O f PDCF. dishes were rinsed once with 4 ml of ice-cold phosphate-buffered saline and cells were scraped with a rubber policeman into 2 ml of ice-cold 30 RUr MOPS. DH 7.25. containin. 150 nv( NaCl and 2 nll dithiothreitol. The cell susbension was sedirnanted in a Beckman miCrOfuge for 5 mi" and the pellet was suspended in 100 "1 of MOPS buffer containing 10 I glycerol ( v l v ) and 2 iM dithiothreitol and was frozen at -70' for assay the next day. For Some studies. two separate experiments were performed and
20. 21.
22. 23.
24. 25.
26.
27.
28.
29.
30.
Bradford, M. (1976) Anal. Bwchem. 72,248-254 Huang, E. M., and Detwiler, T. C. (1986) Biochem. J. 236,895-
Hokin, L. E. (1985) Annu. Reu. Biochem. 64, 205-235 Habenicht, A. J. R., Glomset, J. A., Goerig, M., Gronwald, R.,
Grulich, J., Loth, U., and Schettler, G. (1985) J. Bwl. Chem.
901
260,1370-1373 Berridge, M. J. (1983) Biochem. J. 212,849-858 Downes, C. P., and Wusteman, M. M. (1983) Biochem. J. 216,
Agranoff, B. W., Murthy, P., and Seguin, E. B. (1983) J. Biol.
Taylor, M. V., Metcalfe, J. C., Hesketh, T. R., Smith, G. A., and
Wilson, D. B., Bross, T. E., Hofmann, S. L., and Majerus, P. W.
Wilson, D. B., Neufeld, E. J., and Majerus, P. W. (1985) J. Biol.
Baserga, R. (1976) Multiplication and Division in Mammalian
633-640
Chem. 258,2076-2078
Moore, J. P. (1984) Nature 312,462-465
(1984) J. Bwl. Chern. 259, 11718-11724
Chem. 260,1046-1051
Cells, Marcel Dekker, Inc., New York
measuring the incorporation of "P from Iy-"PlATP into PtdIna-P and PtdIns-P,, respectively. Standard phosphorylation a s s a y s Were carried Out for 2 min at I4 in 35-40 V I . A 1 0 - y 1 aliquot of the cell Preparation containing 3-20 p9 Of protein w a s the source of enzyme. Other assay
dithiothreitol. 18 mn NgC1, and 450 rn Iy-"PlATP (600-1500 cprn/pmol). Separate ezperiments showed that these Concentrations Of Mg" and ATP allowed maximal rates of phosphorylation. Maximum activities (V.) were determined by adding miled micelles Of Ptdlns or Ptdlnr-P in Triton X-100 to give f i n a l concentrations of 0.171 Triton and 3.5 mglrnl ( 4 nll) Ptdlns o r
of Ptdlns and Ptdlns-P in CHCI, yere dried under nitrogen. resuspended in 0.5 mplml (0.5 M) PtdInr-P. To prepare the lipid Substrates, stock Solutions
the appropriate volume Of 1.21 Triton. and vortexed YigorousIY until the su~pension *as clear. When activity was measured with endogenous substrates, cell preparations were pipetted into a buffer that contained the MOPS. MgCl,, dithiothreitol and 1 M sodium deoxycholate. Assays Were started by the addition of ("PIATP. After 2 min. reactions were stopped by pipetting 3 0 y l of the reaction mixture into 0 . 9 ml Of CHCl,/CHIOH/i2 N HC1 (66:13:lI. After 30 minutes. phases were separated by the addition Of 0.5 ml of pure upper phase (CH,OH/H~0lCHCl,. 4 8 : 4 1 : 3 ) . The upper phase was
Lipids in the lower phase wece separated by thin-layer chromatography a s removed and the lower phase was washed t w x e with additional upper phase.
described above. Residual ATP that was not washed completely Out Of the lower phase remained at the origin of the ~ e l i u i o ~ e piate. in COntIOi incvbations
the lover phase waz ["PIATP. in which no cell preparation was added. the only "P-Labelled material in
The activities of Ptdlns kinase and Ptdlns-P kinase were determined by
Constituents were (in final c m ~ e n t ~ a r i o n ~ ~ 50 nv( MOPS a t PH 7 . 2 5 . I M
Standard values fo1 V. Of Ptdlns kinase and PtdIns-P kInaae in freshly-harvested quiescent cells were 78 pmollminlmg protein and 292 pmollminlmg protein, celpectively. Activities with endogenous Substrates were 29 pmllmin/nq protein and 28 pmol/nin/mg protein. respectively. In Order to array Simultaneously all of the samples from one elperiwnt it wa3 necewary to freeze the cells. Freeze-thawing did not affect the activity Of Ptdlns-P kinase but resulted in a tWOfold or oreater increase in V- for Ptdlns kinase
Standard values fo1 V. Of Ptdlns kinase and PtdIns-P kInaae in freshly-harvested quiescent cells were 78 pmollminlmg protein and 292 pmollminlmg protein, celpectively. Activities with endogenous Substrates were 29 pmllmin/nq protein and 28 pmol/nin/mg protein. respectively. In Order to array Simultaneously all of the samples from one elperiwnt it wa3 necewary to freeze the cells. Freeze-thawing did not affect the activity Of Ptdlns-P kinase but resulted in a tWOfold or oreater increase in V- for Ptdlns kinase and a three- to four-fold increase i n the rates of phosphorylation Of endogenous PtdInS and PtdIna-P, suggesting that access Of substrate to PtdInS
magnitude Of effects Of PDCF on enzyme aetlvitiel, nor were any changes in kinase was improved by freeze-thawing. However, freezing did not change the
enzyme activity seen with fresh extracts that were not observed when extracts had been frozen. Also. the results obtained were identical whether 01 not proteinase inhibitors or phosphatase inhibitors Were ueed in harvesting the 5811s (NaF inhibited kinase activities).
activity for each culture (expressed as pmol of PhoSphorOUP incorporated into Specific enzyme activities were Calculated by dividing the total
subftcate pec minute) by the awunt of proteln in each culture. Protein concentrations were measured by the method of Bradford (20).
Perults
Turnover Of Ptdlns, PtdIns-P and PtdIns-P. in Swis3 3T3 cells stimulated by PDGp- I n initial erperimnta. in which the breakdown Of Polyphosphoinolitides was studied in intact cells that had been labelled to equilibrium with I'HIinonitol, it was c lear that the reswnsa of 3T3 cells to POGF was Y IY I IU~~ in that the production Of inositol phosphates was sustained for at least an hour after g w w t h factor addition (data not shown). In contrast, in platelets and Other secretoTy cells. the response lasts only a
been no studies Of the effect Of PDGF an the incorporation of labelled few minutes (21.22). Because Of these differences, and becauae there hawe
precursors into PtdInr-P and PtdInS-PI. we conducted studies to determine if PDCF increased the incorporation of '*P into the phosphoinoaitides durinq the period Of phorphoinositide breakdown. Tholle studies *ere designed to asseas the kinetics Of labelling Of Ptdlns-P and Ptdlns-PI in intact cells
Of Ptdlns kinase and Ptdlna-P kinase night be expected to occur. in order to determine at what times PDGF-dependent Changes in the activities
Ptdlns. PtdIn8-P. PtdIns-P1 and phosphatidic acid within 2-S minutes after growth factor addition. The radioactivity in a11 4 compounds increased progressively throughout the LO-minute period. The relative awunts of
phOJphOlipids LID determined by equilibrium labelling with I'Hlinositol (data not shown). The PDCF-dependent stimulation Of lsbellinq was gceatez for PtdIns and phosphatidic acid than for Ptdlnr-P or PtdIns-Pz.
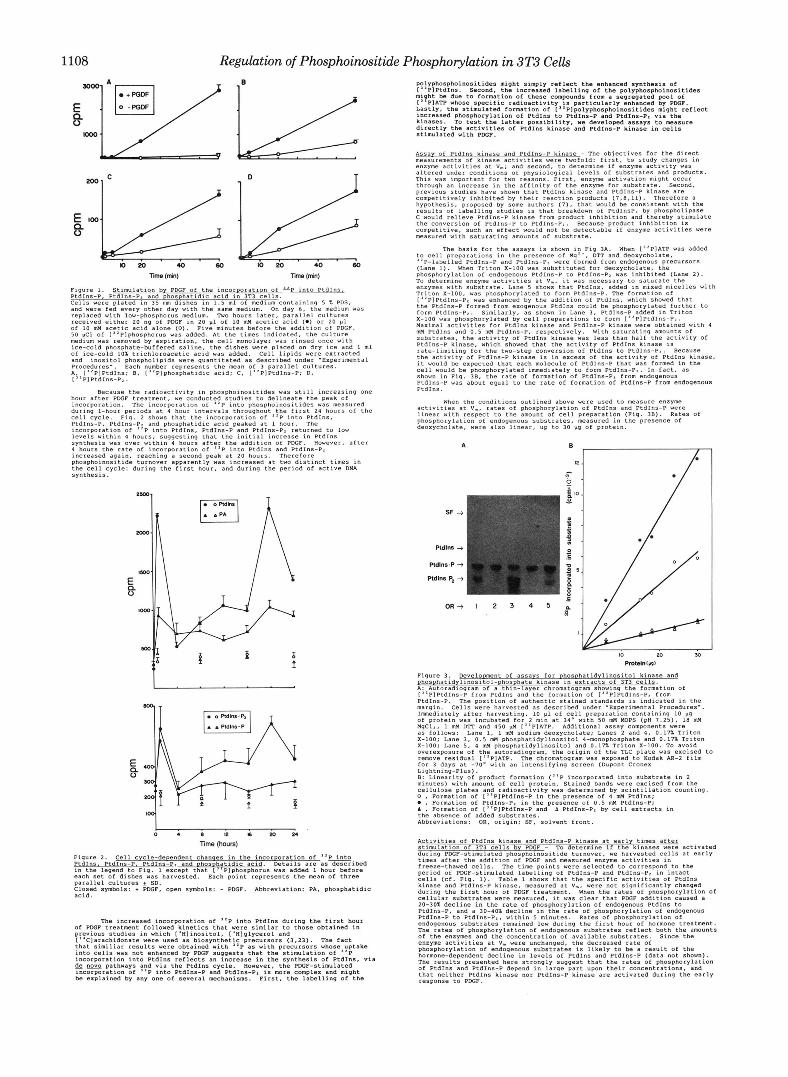
Fig. 1 shows that PDGF stimulated the incorporstion Of label into
'P incorporated were proportional to the relative amounts of the
1108
SQW
E 8
KKK)
Regulation of Phosphoimsitide Phosphorylation in 3T3 Cells
Figure 1. Stimulation by PDGF Of the incorporation Of into PtdIn?lL PtdInS-P, PtdIns-P, and phos hatidic acid in 3T3 cells. Cells were plated in 35 m dfshes in 1.5 ml Of medium containing 5 % PDS, and were fed every other day with the same medium. On day 6, the medium Was replaced With 1ow"phosphocus medium. Two hours later, parallel cultures
Of 10 mM acetic acid alone ( 0 ) . Five minutes before the addition Of PDGF. received either 20 ng of PDGF ~n 20 pl of 10 mM acetic acid 1.) or 20 p l
50 pci Of [''Plphosphorus was added. At the times indicated. the Culture medium was removed by aspiration, the cell monolayer was rinsed once with
Of ice-cold 10% trichlomacetic acid was added. Cell lipids were extracted ice-cold phosphate-buffered saline, the dishes were placed on dry I C ~ and 1 ml
and inositol phosphollpids were quantitated as described under -Experimental
A;21"P1PtdIns: 8, ["Plphosphatidic acid; C, ["PlPtdIns-P; D, Procedures-. Each number represents the mean Of 3 parallel Cultures.
I PIPtdIns-PI.
hour after PDGF treatment, we conducted studies to delineate the peak of
during I-hour periods at 4 hour intervals throughout the first 24 hours Of the incorporation. The xncorporation of "P into PhoSPhoinOsitideS was measured
cell cycle. Fig. 2 Shows that the tncorporation of "e into PtdIns. PtdIns-P, PtdIns-P. and phosphatidic acid peaked at 1 hour. The incorporation of "P into PtdInS, PtdIns-P and PtdIns-P. returned to low levels within 4 hours, suggesting that the initial increase in PtdInS
4 hours the Iate Of incorporation of "P into PtdIns and PtdInS-PI Increased again. reaching a second peak at 20 hours. Therefore phosphoinoaitide turnover apparently was increased at two distinct times in the cell cycle: during the first hour. and during the period of active DNA synthesis.
Because the radioactivity in phosphoinositides was still increasing one
synthesis was over within 4 houcs after the additla" Of PDGF. However, after
E, 0
Figure 2. Cell cycle-dependent chanqes in the incOrDOration Of ''P, PtdInh, PtdIns-P, PtdInr-P. and Ohosohatidic acid. Details a r e as descrlbed in the legend to Fig. 1 except that ["Plphosphorus was added 1 hour before each set of dishes vas harvested. Each poxnt represents the m a n of three parallel Cultures 5 SD. Closed symbols: + PDGF, open symbols: - PDGF. Abbreviation: PA, phosphatidic acid.
Of PDGF treatment followed kinetics that were similar to those obtained in previous Studies in which ['H1inositol, I'HIglycerol and I'*Clarachidonate were used an biosynthetic precursors (3.23). The fact that similiar results were obtained with "P (IS with precurso~s whose uptake into cells was not enhanced by PDGF suggests that the stimulation Of "P
de "yo pathways and via the PtdIns cycle. However, the PDGF-stimulated incorporation into PtdIns reflects an increase in the synthesis Of PtdIns, via
incorporation Of "P into PtdInS-P and PtdIns-P, is m r e complex and might be explained by any one Of several mechanisms. First, the labelling Of the
The increased incorporation Of "P into PtdIns during the first hour
polyphobphoino*itide~ might simply reflect the enhanced synthesis Of ["PlPtdIns. Second. the increased labelling of the polyphosphoinositides might be due to formation Of these compounds from a segregated pool of I' PIATP whose specific radioactivity is particularly enhanced by PDOF. Lastly, the stimulated formation of I''P1polyphosphoinositides might reflect increased DhoSDhorvlation Of PtdIns to PtdIns-P and PtdInS-P1 via the kinases. io test the latter possibility, we developed assay; to Masure directlv the activities Of PtdInS kinase and PtdInr-P kinase in cells stimulated with PDGF
Assay Of PtdInS kinase and PtdIns-P kinase - The objectives for the direct measurements Of kinase activities were twofold: flrst, to study Changes in enzyme actlvitles at v.; and second, to determine if enzyme activity was altered under conditions of physiological levels Of Substrates and products. This was important for two reasons. First, enzyme activation might OCCYC through an increase in the affinity of the enzyme for Substrate. Second. previous studies have Shown that PtdIns kinase and PtdIns-P kinase are competitively inhibited by thelr reaction products (7,8,11). Therefore a hypothesis. proposed by some authors 17). that would be Consistent with the TeSults Of labelling studies 1s that breakdown Of PtdInSP, by phospholipase C would relieve PtdIns-P kinase from product inhibition and thereby Stimulate
comoetitive. Such a n effect would not be detectable if enzyme activities were the conversion of PtdIns-P to PtdIns-Pz. Because prOdUCt inhibition is
to cell preparations in the presence Of Mg", DTP and deoxycholate, "P-labelled PtdIns-P and PtdIns-Pz were formed from endogenous PreCUCSOrS
phosphorylation Of endogenous PtdIns-P to Ptdlns-P. was inhibited (Lane 2). (Lane I). When Triton x-100 was Substituted for deoxycholate. the
TO deterrnlne enzyme activities at V., it was necessa~y to saturate the enzymes Wlth Substrate. Lane 5 Shows that PtdInS, added in mixed micelles with Triton X-100. was phosphorylated to form PtdIns-P. The formation Of
the Ptdlns-P formed from exogenous Ptdlnr could be phosphorylated further to ["PIPtdIns-P, was enhanced by the addition Of PtdIns, Which showed that
X-100 was phosphorylated by cell preparations to form ["PIPtdIns-Pz. form PtdlnS-P2. Smxlarly, a s shown in Lane 3, PtdInS-P added 1" Triton
Maximal activities for PtdIns kinase and PtdIns-P kxnase were obtained With 4 mM PtdIns and 0 . 5 mM PtdIns-P lezpectively. With SatUrating amounts Of substrates, the activity of Ptdlns kinase was less than half the activity of PtdIns-P kinase, Which Showed that the activity Of PtdIns kinase is rate-llmitlng for the two-step conversion of Ptdlns to PtdInS-Pz. Because the activity of PtdIns-P kinase in in excess of the activity of PtdIns kinase, it would be expected that each molecule of PtdInn-P that was formed in the cell would be phosphorylated immediately to form PtdInS-P.. In fact, as shown in Fig. 36, the rate of formation of PtdIns-Pr from endogenous PtdIns-P was about equal to the rate of formation Of PtdIDs-P from endogenous
The basis for the assays is shown in Fig 3A. When 1"PlATP was added
PtdIna.
When the conditions outlined above were used to measure enzyme activities at V., cates Of phosphorylation Of PtdIns and PtdIns-P W e C e
phosphorylation of endogenous substrates, measured in the presence of linear with respect to the amount of cell preparation (Fig. 31). Rates of
deoxycholate, were a150 linear, up to 30 pg Of protein.
Figure 3. Development Of assays for phosphatidylin0SitOl kinase and e h o s p h a t i d v l i n o s i t ~ l - p h ~ ~ p h ~ t ~ kinase in extracts of 3T3 cells. A: Autoradiogram Of a thin-layer chromatogram Showing the formation of ["PlPtdIns-P from PtdInS and the formation of ["PIPtdIns-Pz from PtdIns-P. The position of authentic stained standacds 1s Indicated in the margin. Cells were harvested as dercrxbed under .Experimental PrOcedUres-.
of protein was incubated for 2 min at 34' with 50 mM MOPS (pH 7.25). 18 M Immediately after harvesting, 10 p l Of cell preparation Containing 10 P9
MgClz, 1 mM DTI and 450 pI4 ["PIATP. Additional assay components were
X-100: Lane 3, 0.5 mM phosphatidylinositol 4-monophosphate and 0.17% Triton a s follows: Lane 1. 1 mM sodium deoxycholate; Lanes 2 and 4, 0.17% Triton
x-loo' Lane 5, 4 mH phosphatidylinositol and 0.17% Triton X-100. TO avoid 0Ye~e;POZUTe Of the autoradiogram, the orlgln Of the TLC plate was excised to remove residual ["PIATP. The chromatogram vas exposed to Kodak AR-2 film for 3 days at -70- with an intensifying screen IDupont Crone= Lightning-Plus). 8: Linearity of product formation ("P incorporated into Substrate in 2 minutes) with amount of cell protein. Stained bands were excised from the cellulose plates and radioactivity was determined by scintillation counting. 0 , Formation Of 1"PIPtdIns-P in the presence Of 4 mM PtdInr; 0 , Formation Of PtdInr-Pz in the presence Of 0.5 M PtdIns-P: A , Formation Of ["PlPtdIns-P and A PtdIns-Pz by cell extracts in
Abbrevxations: OR, origin; SF, solvent front. the absence Of added substrates.
Activities Of PtdIns kinase and PtdInS-P kinase at early tines after
during PDGF-Stimulated phosphoinositide tucnovec, we harvested cells at early Stimulation Of 3T3 cells by PDGF - TO determine if the kinases were activated
times after the addition of PDGF and measured enzyme activitles in freeze-thawed cells. The time points were selected to correspond t0 the period Of PDGF-stmulated labelling of PtdInS-P and PtdIns-P. in intact cells (cf. Fig. 1). Table 1 Shows that the specific activities of PtdInS
during the first hour Of PDGF treatment. When the rates Of phosphorylation Of kinase and PtdIns-P kinase, measured at v., yere not significantly changed
cellular substrates were measured, it was clear that PDGF addition caused a 20-301 decline in the rate of phosphorylation of endogenous PtdIns to PtdIns-P, and a 30-101 decline in the rate of phosphorylation Of endogenous PtdIns-P to PtdIns-Pz, within 5 minutes. Rates of phosphorylation Of endogenous Substrates remained low during the firzt hour of hormone treatment. The rates of phosphorylation of endogenous substrates reflect both the amOYnt8 of the enzymes and the concentration of available substrates. Since the enzyme activities at V. were unchanged, the decreased rate Of phosphorylation of endogenous substrates is lLkely to be a result of the hormone-dependent decline in levels Of PtdInS and PtdIns-P (data not shown). The results presented here strongly Suggest that the rates Of phosphorylation Of PtdIns and PtdIns-P depend in large part upon their Concentrations, and that neither PtdIns klnase nor PtdInS-P kinase are activated during the early response to PDGF.
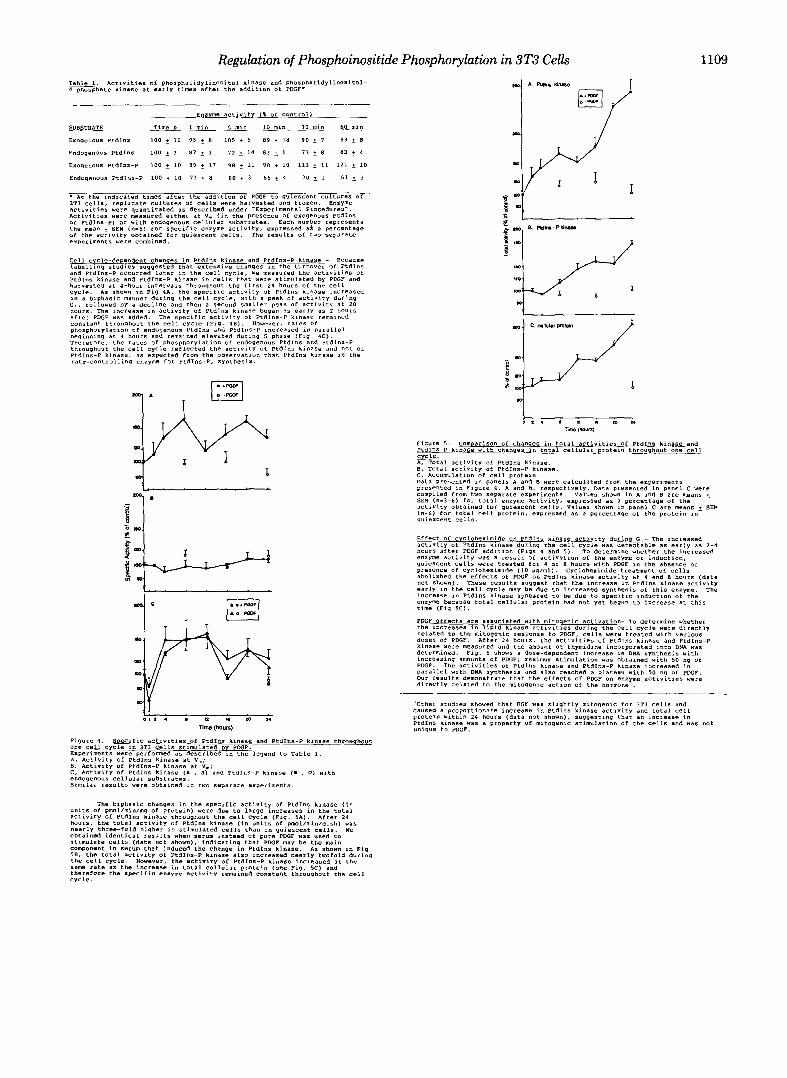
Regulation of Phosphoinositide Phosphorylation in 3T3 Cells 1109 T W . Activities of PhosphatidylinOsitOI kinase and phoSph~tidylinOSitD1- 4-phosphate kinase at early tiles after the additlon of PDGF*
* At the indicated times after the addition of PDGF to quiescent Cultures Of 313 cells. replacate cultures Of cells were haryented and frozen. Enzyme
Activities yere measured either at v. (in the presence of exogenous PtdIns activities were quantitated as described under Experimental FCOcedurBJ".
or PtdIns-P) or with endogenous cellular substrates. Each number represents the mean + SEM ("-6) for rpeclfic enzyme activity, expressed a?. a percentage Of the actlviry obtained for qviescene cell% T h e x e ~ ~ l t h Of t w o separate experiments were combined.
Cell c cle-deeendent chanqes in PtdIns kinase and Ptdlns-P kinase - Because
and PtdIns-P occurred later in the cell cycle, Ye measured the activities Of labellrng rtudles suggested that extenrlve changes ~n the turnover Of PtdIns
Ptdlns kinase and PtdIns-P kinase in Cells that were stimulated by PDGF and harvested at 4-hour intervals throughout the first 24 hours of the cell cycle. As shown in Fig 4A, the SpecifLC activity of PtdIns klnase increased
G I , followed by a decline and then a Second smaller peak of activity at 20 in a biphasic manner during the cell cycle. with a peak of activity during
houcs. The increase in activity of PtdIns klnase began a s early as 2 hours after PDGF w a s added. The specific actlvity of PtdInS-P kinase remained constant throughout the cell cycle (F>g. l B ) . Howeve,, rater of phosphorylation Of endogenous PtdInS and PtdInr-P ,"creased in p a r a l l e l
Therefore. the rates of phoephorylation of endogenous PtdInS and PtdIns-P heglnning at 4 hours and remained elevated during S phase (F14. 4C).
throughout the cell cycle reflected the activity of PtdIns kinase and not of PtdIns-P kinase. as expected from the observation that PtdInS kinase is the rate-controlling enzyme for Ptdlns-PI synthesis.
Figure 4. Specific activities Of PtdIns kinase and PtdIns-P kinase throuqhout One cell cycle in IT3 cells Stimulated by PDGF. Experiments Were performed a s described in the legend to Table 1. A, Activity of PtdInS kinase at v.; B, Activity O f PtdIns-P kinase at V.; C, Activity Of PtdIns kinase (A , B ) and PtdInS-P kinase (a , 0 ) with endogenous cellular substrates. Similar resYlt5 were obtained in two separate experiments.
figure 5 . ComDarison of changes in total activities Of PtdIns kinase and PtdIns-P kinase with chanqes in total cellular protein throuqhout one cell
A, Total activity of PtdIns kinase.
c, ACCUmUlation of cell Droteln. 8 , Total activity of PtdIns-P kinase.
qYieICenC cells.
Effect Of cycloheximide on PtdIn. kinase activity during G I - The increased activity Of PtdIns kinase during the Cell cycle w a s detectable as early as 2-4
enzyme activity was 3 result of activation of the enzyme or induction. hours after PDGF addition (Figs 4 and 5 ) . TO determine whether the Increased
quiescent cells were treated for I or 8 hours with PDGF l n the absence or presence Of Cyclohexlrnide (10 pg/ml). Cycloheximide treatment of cells abolished the effects of PDGF on PtdInS kinase activity at 4 and 8 hours (data not shown). These results w w e s t that the increase in PtdlnS kinase activity e a r l y in the cell cycle may be-due to increased syntherls of this enzyme. The inareare ln PtdInS klnase aoleared to be due to SoecifIc lndwtion of the e n r W because total cellule; protein had not yet'begun to increase at this time (Fig S C ) .
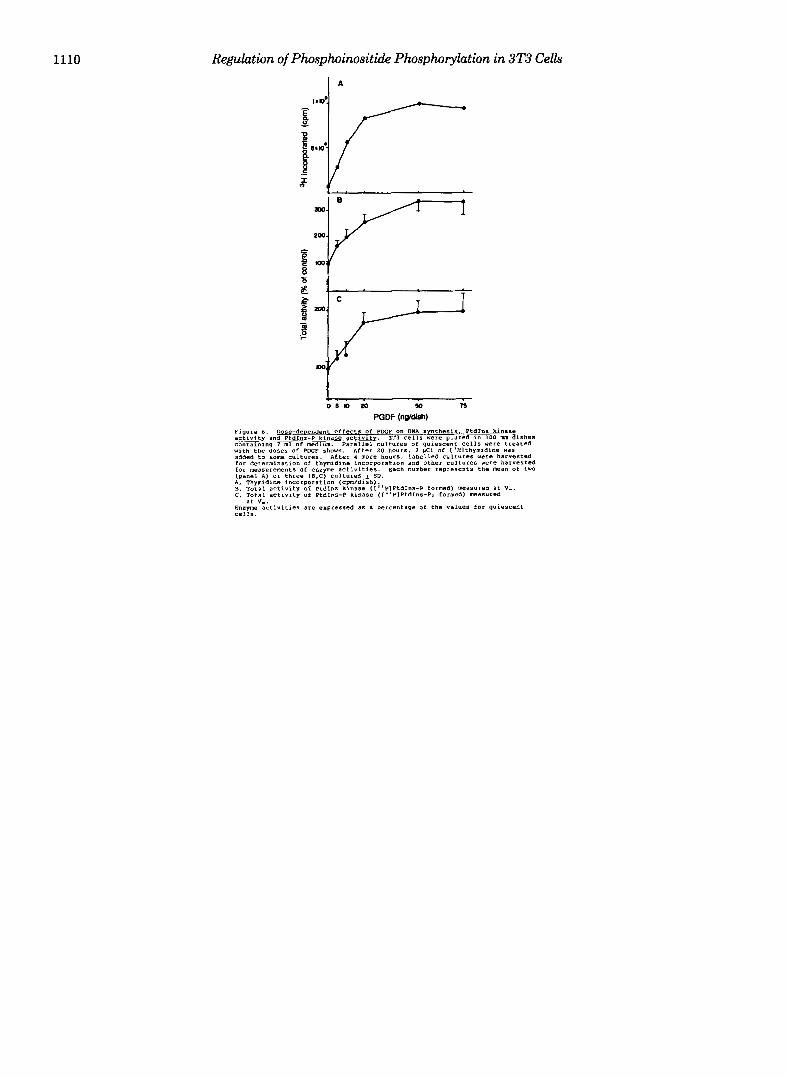
PDGF effects are associated with mitoaenic activation- To determine whether the increases in lipid kinase activities during the cell cycle were directly
dose5 Of PDGF. After 24 hours. the activitles of Ptdlns kinase and PtdIns-P related to the mitogenic response to PDGF, cells were treated with various
kinase were measured and the amount of thmidine incormrated into DNA was determined. Fig. 6 shows a dose-dependent increase in -DNA 6ynthesis with increasing amounts of PDGF; maximum stimulation was Obtained vith 50 ng of PDGF. The activitieS of PtdIns kinase and PtdIns-P kinase increased in
Our results demonstrate that the effects of PDGF on e n z y e activities were parallel with DNA Synthesis and also reached a plateau with 50 "9 Of PDGF.
directly related to the mitogemc action of the hormone .
caused a proportionate increase in PtdIns kinase activity and total cell 'Other studies showed that EGF was slightly mitogenic foc 3T3 cells and
protein within 24 hours (data not shown), suggesting that an increase in PtdInJ kinase was a property Of mitogenic stimulation Of the cells and was not unique to PDGF.
units of pmol/min/mg of protein) were due to large increases in the total The biphasic changes in the specific activity of PtdIns kinase (in
aCtlvify Of PtdIns kinase throughout the Cell cycle (Fig. 5A). After 24 hours, the total activity of PtdInS klnase (in units of pmol/min/dish) was nearly three-fold higher in stimulated cells than in quiescent cells. we Obtained identical results when serum instead of pure PCGP was used to stimulate cell5 (data not shown), indicating that PCGF may be the main component in serum that induced the change in Ptdlns kinase. As shown in Fig 58. the t o t a l activity O f PtdIns-P kinase a 1 9 0 increased nearly twofold during
sane rate as the increase I" total cellular protein (see Fig. 5C) and the cell cycle. However, the activity of PtdIns-P kinase increased at the
therefere the specific enzyme activity remained Constant throvqhout the cell Cycle.
1110 Regulation of Phosphoinositide Phosphorylation in 3T3 Cells


















