
Regulation of Light Energy Utilization and Distribution of Photosynthesis in Five Subtropical Woody...
11
Journal of Integrative Plant Biology 2007, 49 (3): 351−361 Received 14 Mar. 2006 Accepted 4 Jul. 2006 Supported by the National Natural Science Foundation of China (30470282). *Author for correspondence. Tel: +86 (0)20 3725 2995; Fax: +86 (0)20 3725 2831; E-mail: <[email protected]>. © 2007 Institute of Botany, the Chinese Academy of Sciences doi: 10.1111/j.1672-9072.2006.00369.x Regulation of Light Energy Utilization and Distribution of Photosynthesis in Five Subtropical Woody Plants Nan Liu 1, 3 , Chang-Lian Peng 1, 2* , Zhi-Fang Lin 1 , Gui-Zhu Lin 1 and Xiao-Ping Pan 1 ( 1 South China Botanical Garden, the Chinese Academy of Sciences, Guangdong Key Laboratory of Digital Botanical Garden, Guangzhou 510650, China; 2 College of Life Sciences, South China Normal University, Guangzhou 510631, China; 3 Graduate University of the Chinese Academy of Sciences, Beijing 100049, China) Abstract The adaptations and responses of photosynthesis to long- and short-term growth light gradient treatments were investigated in five subtropical forest plants, namely Pinus massoniana Lamb., Schima superba Gardn. et Champ., Castanopsis fissa (Champ. ex Benth.) Rehd. et Wils., Acmena acuminatissima (BI.) Merr et Perry, and Cryptocarya concinna Hance. With diurnal changes in sunlight and air temperature, the de-epoxidation state and lutein content in the five woody plants under three light intensities first increased and then decreased during the day. However, maximal photochemical efficiency (F v /F m ; where F m is the maximum fluorescence yield and F v is variable fluorescence) and the photochemical quantum yields of photosystem (PS) II ( Φ Φ Φ PSII ) of the species examined changed in the opposite manner, with those in plants grown under 100% natural light changing the most. After long-term treat- ment (21 months), anti-oxidant capacity (1,1-diphenyl-2-picrylhydrazyl radical (DPPH·)-scavenging capacity) and uti- lization of excitation energy showed differences in modulation by different light intensities. It was shown that A. acuminatissima and C. concinna, as dominant species in the late succession stage of a subtropical forest in Dinghu mountain, South China, were better able to adapt to different light environments. However, P. massoniana, the pioneer species of this forest, exhibited less adaptation to low light intensity and was definitely eliminated by the forest succession process. Key words: adaptation; chlorophyll fluorescence; community succession; light intensity; subtropical forest. Liu N, Peng CL, Lin ZF, Lin GZ, Pan XP (2007). Regulation of light energy utilization and distribution of photosynthesis in five subtropical woody plants. J. Integr. Plant Biol. 49(3), 351−361. Available online at www.blackwell-synergy.com/links/toc/jipb, www.jipb.net Physiologically, light has both direct and indirect effects. It affects metabolism directly through photosynthesis, but affects growth and development indirectly, both as a consequence of immediate metabolic responses and more subtly by its control of morphogenesis. Photosynthesis is the most important deter- minant of plant growth and its responses and adaptation to the environment enable plants to exist and develop under various light conditions. Growth in permanent shade is unavoidable for the lower leaves of a plant forming a multilayer canopy and it follows that both plastic (within a genotype) and genetic (between genotypes) differences must exist in the photosyn- thetic system. A leaf below the light compensation point uses more energy for respiration than it fixes and is effectively a non-functional organ (Fitter and Hay 2002). When a shade plant grows in full sunshine and excessive energy cannot be dissi- pated by the pigment protein complex of the photosystem, the plant may suffer from photoinhibition (Björkman and Holmgren 1963). Previous studies have shown that the xanthophyll cycle in the photosynthetic apparatus plays an important role in the dissipation of heat and the prevention of photoinhibition in higher plants to enable them to adapt to light and other environmental factors (Long et al. 1994; Gilmore 2001; Verhoeven et al. 2001).
Transcript of Regulation of Light Energy Utilization and Distribution of Photosynthesis in Five Subtropical Woody...

Journal of Integrative Plant Biology 2007, 49 (3): 351−361
Received 14 Mar. 2006 Accepted 4 Jul. 2006
Supported by the National Natural Science Foundation of China (30470282).
*Author for correspondence.
Tel: +86 (0)20 3725 2995;
Fax: +86 (0)20 3725 2831;
E-mail: <[email protected]>.
© 2007 Institute of Botany, the Chinese Academy of Sciences
doi: 10.1111/j.1672-9072.2006.00369.x
Regulation of Light Energy Utilization and Distribution ofPhotosynthesis in Five Subtropical Woody Plants
Nan Liu1, 3, Chang-Lian Peng1, 2*, Zhi-Fang Lin1, Gui-Zhu Lin1 and Xiao-Ping Pan1
(1South China Botanical Garden, the Chinese Academy of Sciences, Guangdong Key Laboratory of DigitalBotanical Garden, Guangzhou 510650, China;
2College of Life Sciences, South China Normal University, Guangzhou 510631, China;3Graduate University of the Chinese Academy of Sciences, Beijing 100049, China)
Abstract
The adaptations and responses of photosynthesis to long- and short-term growth light gradient treatments wereinvestigated in five subtropical forest plants, namely Pinus massoniana Lamb., Schima superba Gardn. et Champ.,Castanopsis fissa (Champ. ex Benth.) Rehd. et Wils., Acmena acuminatissima (BI.) Merr et Perry, and Cryptocaryaconcinna Hance. With diurnal changes in sunlight and air temperature, the de-epoxidation state and lutein contentin the five woody plants under three light intensities first increased and then decreased during the day. However,maximal photochemical efficiency (Fv/Fm; where Fm is the maximum fluorescence yield and Fv is variable fluorescence)and the photochemical quantum yields of photosystem (PS) II (ΦΦΦΦΦPSII) of the species examined changed in theopposite manner, with those in plants grown under 100% natural light changing the most. After long-term treat-ment (21 months), anti-oxidant capacity (1,1-diphenyl-2-picrylhydrazyl radical (DPPH·)-scavenging capacity) and uti-lization of excitation energy showed differences in modulation by different light intensities. It was shown that A.acuminatissima and C. concinna, as dominant species in the late succession stage of a subtropical forest in Dinghumountain, South China, were better able to adapt to different light environments. However, P. massoniana, thepioneer species of this forest, exhibited less adaptation to low light intensity and was definitely eliminated by theforest succession process.
Key words: adaptation; chlorophyll fluorescence; community succession; light intensity; subtropical forest.
Liu N, Peng CL, Lin ZF, Lin GZ, Pan XP (2007). Regulation of light energy utilization and distribution of photosynthesis in fivesubtropical woody plants. J. Integr. Plant Biol. 49(3), 351−361.
Available online at www.blackwell-synergy.com/links/toc/jipb, www.jipb.net
Physiologically, light has both direct and indirect effects. Itaffects metabolism directly through photosynthesis, but affectsgrowth and development indirectly, both as a consequence ofimmediate metabolic responses and more subtly by its controlof morphogenesis. Photosynthesis is the most important deter-minant of plant growth and its responses and adaptation to the
environment enable plants to exist and develop under variouslight conditions. Growth in permanent shade is unavoidable forthe lower leaves of a plant forming a multilayer canopy and itfollows that both plastic (within a genotype) and genetic(between genotypes) differences must exist in the photosyn-thetic system. A leaf below the light compensation point usesmore energy for respiration than it fixes and is effectively anon-functional organ (Fitter and Hay 2002). When a shade plantgrows in full sunshine and excessive energy cannot be dissi-pated by the pigment protein complex of the photosystem, theplant may suffer from photoinhibition (Björkman and Holmgren1963). Previous studies have shown that the xanthophyll cyclein the photosynthetic apparatus plays an important role in thedissipation of heat and the prevention of photoinhibition in higherplants to enable them to adapt to light and other environmentalfactors (Long et al. 1994; Gilmore 2001; Verhoeven et al. 2001).

352 Journal of Integrative Plant Biology Vol. 49 No. 3 2007
The xanthophyll cycle, comprising the carotenoids V(violaxanthin), A (antheraxanthin), and Z (zeaxanthin), is in-volved in the adaptation of the energy conversion efficiency inphotosynthetic membranes. Carotenoid V, which contains twoepoxide groups, can be reversibly and quickly (within minutes)converted into carotenoid Z with two de-epoxidized epoxidegroups through the intermediate carotenoid A to dissipate ex-cessive exciting energy under high light intensity. Carotenoid Zcontributes to the heat dissipation of light energy, whereascarotenoid V does not. A higher de-epoxidation state (DES) ofthis cycle is related to stronger protection of the photosyn-thetic apparatus and a lower photochemical efficiency.
The chlorophyll (Chl) in thylakoid membrane is photosensitiveand can transform into 1Chl, which can be changed to 3Chlwithout de-excitation through photochemical processes. Theexcited energy can be transferred to O2 to produce 1O2 atphotosystem (PS) II and O2
-. at PSI, under oxygen, which can befurther transformed into H2O2 and ·OH (Demmig-Adams andAdams 2000). As the byproduct of photosynthesis, active oxy-gen species may damage the photosynthetic apparatus.Carotenoids, such as lutein, may also contribute to energy dis-sipation or others, such as β-carotene, carotenoid V and lowlevels of carotenoids A and Z, may quench 3Chl and 1O2 andscavenge O2
-. . In addition, with long-term adaptation to theenvironment, plants develop a highly efficient anti-oxidant de-fense system, comprising defense enzymes and low molecu-lar weight anti-oxidants, such as glutathione (GSH) and ascor-bate (Noctor and Foyer 1998). Measurements of anti-oxidantenzyme activities and anti-oxidant molecule content can be usedto evaluate the anti-oxidant capacities of plants. In recentstudies, the capacity of a biological reagent to scavenge 1,1-diphenyl-2-picrylhydrazyl (DPPH·) has been used as an indexof its anti-oxidant capacity and reflects approximately the re-sistance of the plant anti-oxidant system to extrinsic free radi-cals (Larrauri et al. 1998; Peng et al. 2000a).
A balance of absorbed light energy (or excitation energy)between the two photosystems is necessary for maximumphotochemical energy conversion efficiency. The regulation ofenergy distribution between PSI and PSII involves physicalmovement of the light-harvesting Chl a/b complex (Horton 1983).The possibility of a redistribution of excitation in favor of PSIcannot be excluded, taking into consideration the possibledestacking of grana thylakoids and rearrangement of pigment-protein complexes as a result of light treatment (Hundal et al.1990). Photoinhibitory treatment leads not only to a transforma-tion of some of the of PSII centers in quenchers, but also to arelative increase of excitation of PSI and an increase of excita-tion energy transfer from PSII to PSI, which is known as statetransition (Somersalo and Krause 1989). Measurement of Chlfluorescence is a useful technique to investigate the conditionof the photosynthetic apparatus of plants from which we canobtain considerable information regarding the structure and
funct ion of the photosystems, inc luding exci tat iontransformation, electron transportation, and PSII characteristics.The ratios of 77-K Chl fluorescence emission of PSII and PSIcan be used to investigate the excitation energy transfer be-tween the photosystems and state transitions (Krause andWeis 1991).
The responses of forest plants to sunlight vary amongspecies. As an impeller, light can promote forest successionthrough changes in the degree of shade within a community. Inthe present study, we focused on differences in the utilizationof light energy and excessive energy dissipation and distribu-tion among dominant species of subtropical forest plants inSouth China in order to expound the mechanism of differentadaptation to growth light intensity among these species in thesuccession of the forest community.
Results
Diurnal changes in the pigment content in plant leaves
Lutein is the most abundant component of carotenoids, partici-pating in the construction of chloroplast pigment antennae, lightharvesting and photoprotection (Siefermann-Harms 1987;Kühlbrandt et al. 1994). From Figure 1, it can be seen that thelutein content (based on Chl content) increased in the begin-ning and then decreased in the five woody plants examined inthe present study, namely Pinus massoniana, Schima superba,Castanopsis fissa, Acmena acuminatissima, and Cryptocaryaconcinna, with diurnal changes in light intensity. The diurnalchanges in lutein content were obvious under 100% naturallight and showed a maximum at 12:00, which indicates thatstronger sunlight with a higher temperature induced the incre-ment of lutein content. Lutein content in C. fissa, A.acuminatissima, and C. concinna was much higher under 100%natural light intensity than at lower light intensity (P<0.05). Along period of high-light treatment changed the carotenoidcomponents. An elevated lutein content is favorable for thestability of photosystem structure and function under high lightintensity. The lutein content of P. massoniana was similar un-der different light intensities, but was lower under 100% natu-ral light, which may be attributed to its adaptation to strongerlight and higher temperature, as well as its smaller antenna inphotosystem as a heliophyte species.
The DES can reflect the activity of the xanthophyll cyclerelated to photoprotection. Similar to variations in lutein content,DES was elevated in the morning and then decreased duringthe day. Except for A. acuminatissim under 100% natural light,all other species showed maximum DES at 12:00 (Figure 1),which indicates that higher air temperature and sunlight at mid-day stimulated DES. The DES of P. massoniana did not changeobviously between the various light treatments, but was higher
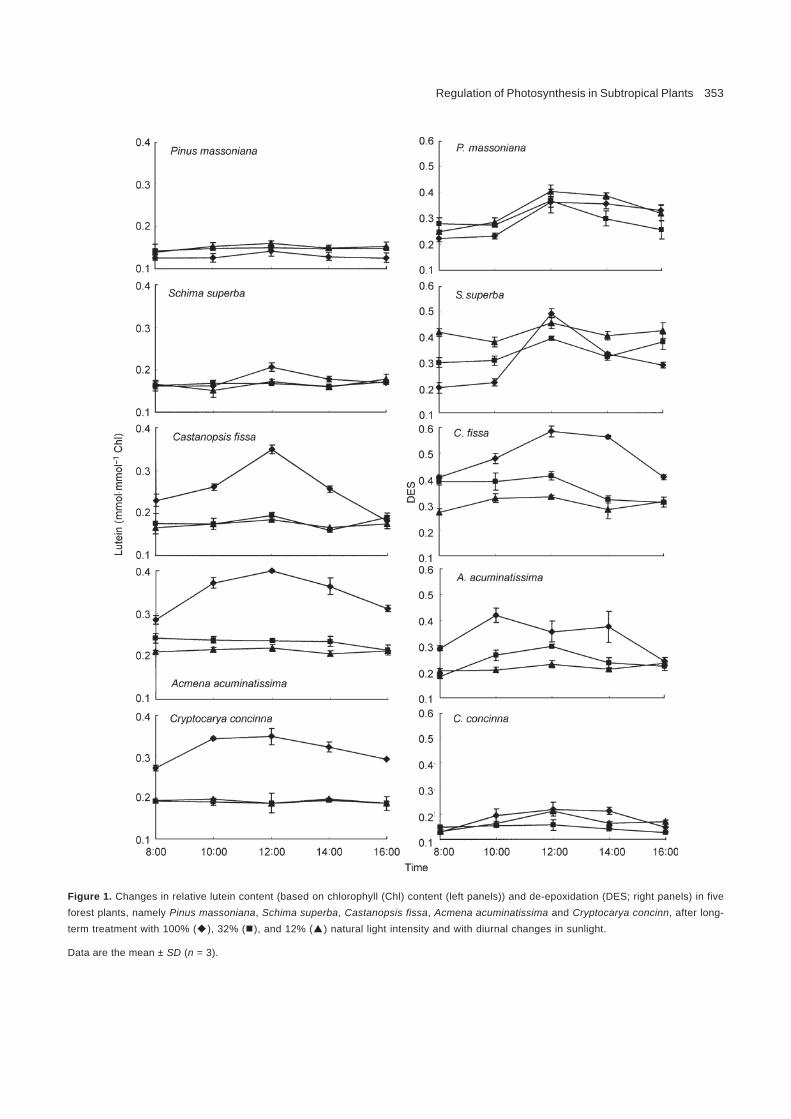
Regulation of Photosynthesis in Subtropical Plants 353
Figure 1. Changes in relative lutein content (based on chlorophyll (Chl) content (left panels)) and de-epoxidation (DES; right panels) in fiveforest plants, namely Pinus massoniana, Schima superba, Castanopsis fissa, Acmena acuminatissima and Cryptocarya concinn, after long-term treatment with 100% ( ), 32% ( ), and 12% ( ) natural light intensity and with diurnal changes in sunlight.
Data are the mean ± SD (n = 3).
354 Journal of Integrative Plant Biology Vol. 49 No. 3 2007
at 100% natural light in C. fissa, A. acuminatissima, and C.concinna. The extent of the change in DES in C. concinna wassmaller than that observed in the other four species, reflectingthe low-sensitivity of C. concinna to short-term strong sunlightand high temperature. The DES of S. superba under 100% ofnatural light was the highest among three light treatments (100%,32%, and 12% natural light), but only at 12:00.
Anti-oxidant activity (DPPH·-scavenging capacity)
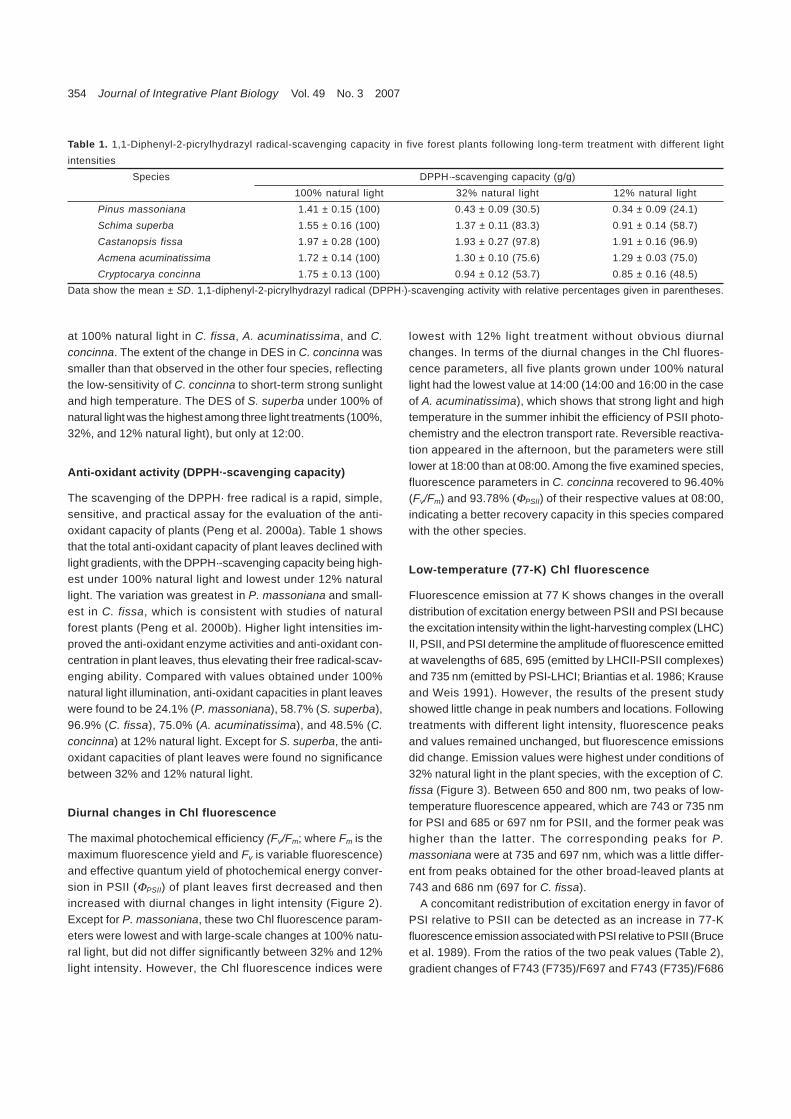
The scavenging of the DPPH· free radical is a rapid, simple,sensitive, and practical assay for the evaluation of the anti-oxidant capacity of plants (Peng et al. 2000a). Table 1 showsthat the total anti-oxidant capacity of plant leaves declined withlight gradients, with the DPPH·-scavenging capacity being high-est under 100% natural light and lowest under 12% naturallight. The variation was greatest in P. massoniana and small-est in C. fissa, which is consistent with studies of naturalforest plants (Peng et al. 2000b). Higher light intensities im-proved the anti-oxidant enzyme activities and anti-oxidant con-centration in plant leaves, thus elevating their free radical-scav-enging ability. Compared with values obtained under 100%natural light illumination, anti-oxidant capacities in plant leaveswere found to be 24.1% (P. massoniana), 58.7% (S. superba),96.9% (C. fissa), 75.0% (A. acuminatissima), and 48.5% (C.concinna) at 12% natural light. Except for S. superba, the anti-oxidant capacities of plant leaves were found no significancebetween 32% and 12% natural light.
Diurnal changes in Chl fluorescence
The maximal photochemical efficiency (Fv/Fm; where Fm is themaximum fluorescence yield and Fv is variable fluorescence)and effective quantum yield of photochemical energy conver-sion in PSII (ΦPSII) of plant leaves first decreased and thenincreased with diurnal changes in light intensity (Figure 2).Except for P. massoniana, these two Chl fluorescence param-eters were lowest and with large-scale changes at 100% natu-ral light, but did not differ significantly between 32% and 12%light intensity. However, the Chl fluorescence indices were
lowest with 12% light treatment without obvious diurnalchanges. In terms of the diurnal changes in the Chl fluores-cence parameters, all five plants grown under 100% naturallight had the lowest value at 14:00 (14:00 and 16:00 in the caseof A. acuminatissima), which shows that strong light and hightemperature in the summer inhibit the efficiency of PSII photo-chemistry and the electron transport rate. Reversible reactiva-tion appeared in the afternoon, but the parameters were stilllower at 18:00 than at 08:00. Among the five examined species,fluorescence parameters in C. concinna recovered to 96.40%(Fv/Fm) and 93.78% (ΦPSII) of their respective values at 08:00,indicating a better recovery capacity in this species comparedwith the other species.
Low-temperature (77-K) Chl fluorescence
Fluorescence emission at 77 K shows changes in the overalldistribution of excitation energy between PSII and PSI becausethe excitation intensity within the light-harvesting complex (LHC)II, PSII, and PSI determine the amplitude of fluorescence emittedat wavelengths of 685, 695 (emitted by LHCII-PSII complexes)and 735 nm (emitted by PSI-LHCI; Briantias et al. 1986; Krauseand Weis 1991). However, the results of the present studyshowed little change in peak numbers and locations. Followingtreatments with different light intensity, fluorescence peaksand values remained unchanged, but fluorescence emissionsdid change. Emission values were highest under conditions of32% natural light in the plant species, with the exception of C.fissa (Figure 3). Between 650 and 800 nm, two peaks of low-temperature fluorescence appeared, which are 743 or 735 nmfor PSI and 685 or 697 nm for PSII, and the former peak washigher than the latter. The corresponding peaks for P.massoniana were at 735 and 697 nm, which was a little differ-ent from peaks obtained for the other broad-leaved plants at743 and 686 nm (697 for C. fissa).
A concomitant redistribution of excitation energy in favor ofPSI relative to PSII can be detected as an increase in 77-Kfluorescence emission associated with PSI relative to PSII (Bruceet al. 1989). From the ratios of the two peak values (Table 2),gradient changes of F743 (F735)/F697 and F743 (F735)/F686
Table 1. 1,1-Diphenyl-2-picrylhydrazyl radical-scavenging capacity in five forest plants following long-term treatment with different lightintensities
Species DPPH·-scavenging capacity (g/g)100% natural light 32% natural light 12% natural light
Pinus massoniana 1.41 ± 0.15 (100) 0.43 ± 0.09 (30.5) 0.34 ± 0.09 (24.1)Schima superba 1.55 ± 0.16 (100) 1.37 ± 0.11 (83.3) 0.91 ± 0.14 (58.7)Castanopsis fissa 1.97 ± 0.28 (100) 1.93 ± 0.27 (97.8) 1.91 ± 0.16 (96.9)Acmena acuminatissima 1.72 ± 0.14 (100) 1.30 ± 0.10 (75.6) 1.29 ± 0.03 (75.0)Cryptocarya concinna 1.75 ± 0.13 (100) 0.94 ± 0.12 (53.7) 0.85 ± 0.16 (48.5)
Data show the mean ± SD. 1,1-diphenyl-2-picrylhydrazyl radical (DPPH·)-scavenging activity with relative percentages given in parentheses.

Regulation of Photosynthesis in Subtropical Plants 355
Figure 2. Maximal photochemical efficiency (Fv/Fm; where Fm is the maximum fluorescence yield and Fv is variable fluorescence; left panels)and the photochemical quantum yields of photosystem (PS) II (ΦPSII; right panels) in five forest plants, namely Pinus massoniana, Schimasuperba, Castanopsis fissa, Acmena acuminatissima and Cryptocarya concinna, after long-term treatment with 100% ( ), 32% ( ), and 12%( ) natural light intensity and with diurnal changes in sunlight.
Data are the mean ± SD (n = 5). Measurements were obtained in plant leaves after 30 min dark adaptation.

356 Journal of Integrative Plant Biology Vol. 49 No. 3 2007
were found according to different natural light treatments. TheF735/F686 ratio of P. massoniana did not differ between 100%and 32% natural light treatment, but increased 24.55% under12% sunlight, lower than any other species except for C. fissa(21.00%). Compared with 100% natural light, the F743 (F735)/F697 increments of C. fissa and P. massoniana were 20.2%and 39.1% with weak light (12%) treatments, whereas theincrements of F743/F686 for A. acuminatissima, S. superba,and C. concinna were 40.2%, 34.5%, and 29.7%, respectively.The results demonstrate that weak light results in a tendencyfor more excitation energery to be distributed from PSII to PSI;however, strong light may increase the distribution of excita-tion light to PSII rather than PSI.
Discussion
Light as the only energy source for photosynthesis, is funda-mental in plant metabolism. The chemical energy transformedfrom light by the photosynthetic apparatus can be used in CO2
fixation and other assimilation. However, when the energyabsorbed is in excess of the amount that can be used forcarbon fixation, the photosystems become overexcited andthe electron transport chains may become over-reduced. Thiseffect is known as photoinhibition, but it is a complex phenom-enon and the more precise terms “photoprotection” and“photoinactivation” are preferred (Osmond et al. 1999). Theratio Fv/Fm is an important parameter in measuring PSII photo-chemical potential and its reduction is one of the significantcharacteristics of photoinhibition (Björkman and Demmig 1987;Long et al. 1994). The term ΦPSII is directly related to the non-cyclic electron transport rate of PSII and reflects the activity ofthe PSII reaction center. The value of ΦPSII depends on theefficiency of the PSII opening center in capturing excitationenergy, as well as the number of PSII opening centers (Kralland Edward 1992; Valentini et al. 1995). These two Chl fluo-rescence parameters first decreased and then increased withdiurnal changes in sunlight intensity (Figure 2). The lowestvalues appeared at 14:00 or 16:00, which showed that tempo-rary inactivation of PSII was brought about by the strong sun-light and high temperature at midday and that the short-termphotoinhibition was significant. Compared with the weak lighttreatments, the fluorescence parameters of broad-leaved spe-cies showed obvious changes under 100% natural light andwere different from those in the conifer species P. massoniana.The latter only exhibited little variation in these two parametersduring the day and the values of Fv/Fm and ΦPSII were lowerunder 12% natural light. This may be due to the fact that P.massoniana, as a pioneer heliophyte species, has a higherlight saturation point and shows less photoinhibition than otherspecies. In contrast, weak light may become a problem for P.massoniana. Without enough sunlight, the growing condition
Figure 3. Fluorescence emission spectra at 77 K of five forest plants,namely Pinus massoniana, Schima superba, Castanopsis fissa,Acmena acuminatissima and Cryptocarya concinna, after long-termtreatment at different light intensities.
Chlorophyll fluorescence was exited at 436 nm. Spectra represent anaverage of three scans. Each experiment was repeated four times.
Regulation of Photosynthesis in Subtropical Plants 357
of P. massoniana was poor, with withered and yellow needlesand low-level fluorescence. The Fv/Fm of A. acuminatissimaand C. concinna in the dark recovered to nearly 96% of that atdawn, showing the best recovery from photoinhibition amongthe five species examined.
Despite effective long-term acclimation, only a part of theabsorbed energy is utilized for photosynthesis. The remaining
excited energy captured by the LHC must be dissipated harm-lessly or too much excessive excited energy could lead to theproduction of damaging reactive oxygen species as byproductsof photosynthesis, which can damage the photosynthetic ap-paratus (Osmond 1994; Müller et al. 2001). Xanthophyll cycle-dependent thermal dissipation and detoxification of reactiveoxygen species by the anti-oxidant system are two
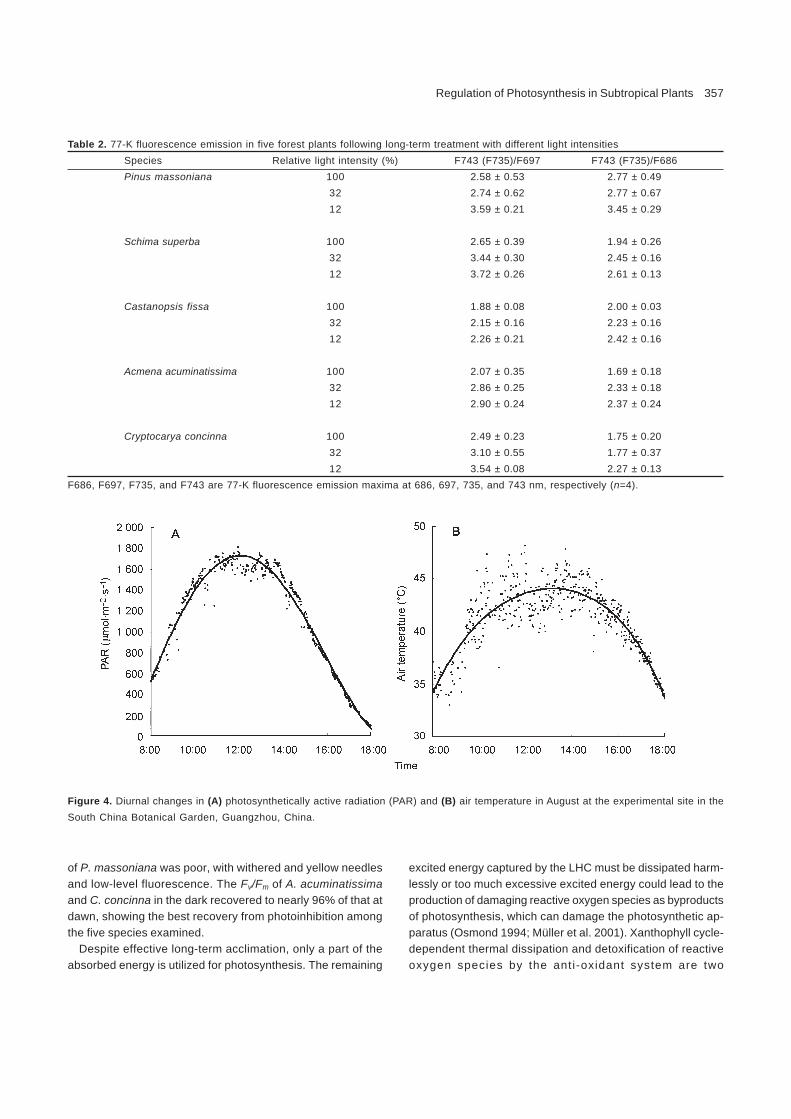
Table 2. 77-K fluorescence emission in five forest plants following long-term treatment with different light intensitiesSpecies Relative light intensity (%) F743 (F735)/F697 F743 (F735)/F686Pinus massoniana 100 2.58 ± 0.53 2.77 ± 0.49
32 2.74 ± 0.62 2.77 ± 0.6712 3.59 ± 0.21 3.45 ± 0.29
Schima superba 100 2.65 ± 0.39 1.94 ± 0.2632 3.44 ± 0.30 2.45 ± 0.1612 3.72 ± 0.26 2.61 ± 0.13
Castanopsis fissa 100 1.88 ± 0.08 2.00 ± 0.0332 2.15 ± 0.16 2.23 ± 0.1612 2.26 ± 0.21 2.42 ± 0.16
Acmena acuminatissima 100 2.07 ± 0.35 1.69 ± 0.1832 2.86 ± 0.25 2.33 ± 0.1812 2.90 ± 0.24 2.37 ± 0.24
Cryptocarya concinna 100 2.49 ± 0.23 1.75 ± 0.2032 3.10 ± 0.55 1.77 ± 0.3712 3.54 ± 0.08 2.27 ± 0.13
F686, F697, F735, and F743 are 77-K fluorescence emission maxima at 686, 697, 735, and 743 nm, respectively (n=4).
Figure 4. Diurnal changes in (A) photosynthetically active radiation (PAR) and (B) air temperature in August at the experimental site in theSouth China Botanical Garden, Guangzhou, China.

358 Journal of Integrative Plant Biology Vol. 49 No. 3 2007
photoprotective mechanisms that plants have evolved to copewith excess absorbed photon flux density (PFD). The frequentlyobserved relationship between the contribution of carotenoidsZ and A to the total pool of xanthophyll-cycle pigments (VAZ)and non-photochemical fluorescence quenching has led to theconclusion that these carotenoids are involved in the short-term adjustment of the protective dissipation of excess excita-tion energy (Demmig-Adams and Adams 1996). Some studieshave reported that the most abundant carotenoid component inthylakoid, namely lutein, may participate in the dissipation ofnon-radiation energy through quenching of Chl fluorescence(Kühlbrandt et al. 1994; Niyogi et al. 1997). In addition, caro-tenoids Z, A, and lutein may act as anti-oxidants to scavengesinglet oxygen and free radicals and eliminate the damage tothe photosynthetic apparatus and cellular structure. In thepresent study, lutein content (based on Chl content) and DESlevels were all increased at first and decreased later withdiurnal changes in light intensity; the DPPH·-scavenging ca-pacity was improved under high light treatment (Figure 1; Table1). These results indicate that, following either long- or short-term treatment with strong light, protective mechanisms againstphotoinhibition and photooxidation are activated. Because oftheir different characteristics and functions in forest succes-sion processes, excessive excitation energy dissipation andanti-oxidant capacities varied among species under strongersunlight. As a pioneer heliophyte species, the Fv/Fm and ΦPSII
of P. massoniana were lower under 12% light compared with32% and 100% natural light. Diurnal changes in fluorescenceparameters, DES, and lutein content were small at various lightintensities, but obvious differences in anti-oxidant capacitieswere found among the different light treatments. So, we de-duced that the efficiencies of light harvesting, energytransformation, and photosynthetic electron transportation inthe photosynthetic apparatus were much lower at 12% and32% of natural light so that oxidative stress and photoinhibitiondid not occur in plant cells. Under 100% natural light, the incre-ments of DPPH·-scavenging capacities of P. massoniana werehigher than DES and lutein related to heat dissipation, whichindicates that an elevated anti-oxidant capacity may be thepredominant protection mechanism in the adaptation of thisspecies to strong light. The medium succession species S.superba and C. fissa were less light dependent than P.massoniana, with a lesser increment in the anti-oxidant capac-ity after stronger light treatment. The plant leaves of these twospecies were significantly inhibited at noon (decrement in Fv/Fm and ΦPSII), with a rapidly increased DES, indicating that thexanthophyll cycle plays an important role in heat dissipation.The climax species of the subtropical forest A. acuminatissimaand C. concinna exhibited little diurnal change in Fv/Fm andΦPSII under 12% and 32% natural light, but DES and lutein con-tent fluctuated not significantly with 50%–75% anti-oxidant ca-pacity compared with strong light treatments. Photoinhibition
existed at noon with the increase in DES, especially for lutein,the content of which was markedly decreased at 14:00 in theafternoon. In addition, the Fv/Fm and ΦPSII of these two speciesrecovered more quickly in the dark than did those of the otherthree woody plants. The results demonstrate that the speciesrequiring medium light adjusted their metabolism in terms of thecarotenoids (lutein, A, and Z) and PSII function in order to im-prove anti-oxidant capacity, accelerate heat dissipation, and torecover the ability to resist photoinhibition when exposured tofull sunshine over a long period. From these results, it is evi-dent that different plant species adopt distinct primary tacticsto enable them to cope with environmental variations.
Highly efficient photosynthesis requires a balanced excita-tion of the two photosystems. The balanced modulation of ex-citation energy flux to the two photosystems is achieved bythe state transitions (Bonaventura and Myers 1969). This phe-nomenon occurs via the reversible transfer of a fraction ofLCHII from PSII to PSI (state 1 to state 2 transition), as a conse-quence of its phosphorylation (Bennett 1991; Allen 1992; Gal etal. 1997). Energy interaction between the main pigment-proteincomplexes of thylakoid membranes depends on their mutualorganization and distance between their light-capturing anten-nae complexes. Changes in membrane organization induced byunstacking under conditions of low salt, state 1–state 2 transi-tion or short-duration heat stress influence the relative intensi-ties of the main fluorescent bands at 77 K of the isolatedthylakoids. The ratio of F735/F685 (F735/F695) reflects thesechanges and is commonly used to estimate changes in energydistribution between both photosystems (Ivanov and Velitchkova1990). In addition, the observed decrease in the ratio of themaximum fluorescence of PSII to the maximum fluorescence ofPSI at 77 K during photoinhibition of spinach leaves has beenregarded as a possible increase in excitation energy transferfrom PSII to PSI (Somersalo and Krause 1989). In the presentstudies, the low-temperature fluorescence curves were simi-lar in shape among species, with two peaks related to bothphotosystems, but some differences in peak locations and dis-tances were observed. The PSI fluorescence peaks of P.massoniana and C. fissa were blue shifted and their corecomplexes of PSII were reflected in fluorescence emissions at697 nm. After long-term light gradient treatments, the relativeemission of low-temperature fluorescence in plant leaves wasaltered. The values of ΦPSI/ΦPSII in P. massoniana were highest,but with least change among the different light intensities; in A.acuminatissima ΦPSI/ΦPSII varied most under different naturallight conditions (Figure 3; Table 2), which indicates differencesbetween the species in their ability to adjust to changes in thelight environment. On the basis of the ΦPSI/ΦPSII ratios, elevatedlight intensities decreased the transfer of energy from PSII toPSI. Therefore, we assume that changes in state transitionsmay occur in all five plants species in order for them to adapt tolong-term sunlight conditions.

Regulation of Photosynthesis in Subtropical Plants 359
Arid climates are found near the Tropic of Cancer. However,in South China, which is controlled by the monsoon from thePacific Ocean, it is rainy and humid throughout the year and asubtropical evergreen broadleaf forest is found (Chen et al.1996). The forest developed over a succession process: fromconifer forest to coniferous and broad-leaved mixed forestdominated by conifers, and then to coniferous and broad-leavedmixed forest dominated by heliophyte broad-leaved species,thereafter to evergreen broad-leaved forest dominated byheliophyte broad-leaved species, until the climax evergreenbroad-leaved forest with shade-tolerant species (Peng 1996).The five woody species chosen in the present study weredominant species at different times in the forest successionprocess. Taking the results of the present study together, P.massoniana, as a pioneer species in the succession process,is fond of a strong light environment, with most physiologicalindices being less sensitive to light gradient treatments anddiurnal changes in sunlight and fewer adjustment capacity interms of the redistribution of excitation energy betweenphotosystems; therefore, P. massoniana cannot develop nor-mally under low light intensity and is replaced by other plantspecies. The climax species of subtropical forest succession,namely A. acuminatissima and C. concinna, are plants depen-dent on medium light and can grow naturally under differentlight intensities because of long- and short-term adjustments inthe absorbtion, dissipation, and transfer of light energy, so thatthey can survive in different succession periods and are themost predominant subtropical forest species.
In conclusion, the present results add more information to theforest succession theory at Dinghu Mountain. The pine forestthat was originally planted in the 1930s is now dominated by P.massoniana in the tree layer mainly because of its unique pho-tosynthetic adaptation to strong sunlight. Thereafter, the conif-erous and broad-leaved mixed forest develops from the pineforest, with a gradual invasion of some heliophyte broadleafspecies, such as S. superba and C. fissa. In this successionperiod, P. massoniana seedlings in the shrub layer cannot ob-tain enough sunlight for ordinary photosynthesis because ofthe increased degree of shade under the tree layer and arethus eliminated from forest development. Eventually, C. concinnaand A. acuminatissima, with well-regulated function of theirphotosynthetic apparatus, inevitably become the dominant spe-cies in the evergreen broad-leaved forest, the subtropical re-gional climax of vegetation.
Materials and Methods
Field site and plant materials
Dinghu Mountain Natural Reserve (112°30'39''–112°33'41'' E,
23°09'21''–23°11'30'' N) is located in the middle of GuangdongProvince, South China. In this forest succession process,heliophyte species (e.g. S. superba and C. fissa) graduallyinvade the pine forests (e.g. P. massoniana) to form mixedforests and the mesophytes species (e.g. C. concinna and .acuminatissima) subsequently invade the mixed forests, even-tually transforming them into broadleaf forests. Therefore, fivetypical woody plants (with longevity ranging from several de-cades to centuries), namely P. massoniana Lamb, S. superbaGardn. and Champ, C. fissa R and W, A. acuminatissima Bi.merr. and Perry, and C. concinna Hance, representing threeperiods in subtropical forest succession, were chosen forinvestigation.
The experimental site was at the South China BotanicalGarden, Guangzhou, Guangdong Province, South China, be-longing to the south subtropical monsoon climate. Potted seed-lings of the five woody plant species were grown under differ-ent light intensities (100%, 32%, and 12% of natural light), withregular management of water and fertilizer for 21 months. Have-locks were used in setting light gradients and 12 potted plantswere subjected to each light intensity treatment. Diurnal changesin natural light intensity and air temperature (Figure 4) weremeasured using photosynthetically active radiation (PAR) andtemperature sensors (PAM-2100; Walz, Efeltrich, Germany).For measurements, mature leaves born in the same year of theexperiment were chosen from different individuals of each plantspecies.
Leaf pigment analysis
At 2-h intervals throughout the day, leaf discs of known area(or weight for P. massoniana needles) were first frozen imme-diately in liquid nitrogen and then ground in a mortar with theaddition of CaCO3, silica, and cold acetone. The mixture wascentrifuged at 11 300g for 10 min at 0–4 °C, with the superna-tant fluid passed through a 0.45-μm filter. Liquid samples wereanalyzed using an HPLC system with a Pinnacle II C18 chro-matographic column (2695; Waters, Milford, MA, USA) accord-ing to the method described by Gilmore and Yamamoto (1991)with slight modification. Pigments were eluted at a flow rate of1.8 mL/min with a detector wavelength for integration of peakareas at 440 nm. The pigment content was calculated using theconversion factors for peak area as well as two relatedparameters. The DES was defined as (Z+0.5×A)/(V+A+Z) andlutein content was estimated on the basis of the Chl content(Gilmore and Yamamoto 1993).
Anti-oxidant capacity (DPPH·-scavenging capacity)
Leaf samples (0.1 g) were obtained from plants grown at dif-ferent light intensities, ground in a mortar with 50% ethanol,and centrifuged for 15 min at 5 000g. The DPPH· solution has a

360 Journal of Integrative Plant Biology Vol. 49 No. 3 2007
unique absorption at 525 nm. The decline in A525 can be used asan index of the organic free radical-scavenging capacity(ORSC) of a plant extract. The total reaction volume was 2 mLand DPPH· was dissolved using 0.5 mL methanol followed bythe addition of 50% ethanol to give a final concentration of 120μmol/L DPPH·. Then, 0.1 mL plant extract (dissolved in 50%ethanol) was added to 1.9 mL DPPH· and A525 was measuredafter 20 min. The remaining DPPH· (R%) and scanvenging ra-tios (ORSC) were determined using the following formulae:
R (%) = ((A–B)/Ao) × 100ORSC (%) = (1–R) × 100
Where Ao is DPPH· absorption in the absence of leaf extract (0.1 mL of 50% ethanol + 1.9 mL DPPH·), A is DPPH· absorption afterreaction with the sample and B is the absorption of the blank (0.1mL sample + 1.9 mL of 50% ethanol). The DPPH·-scavengingcapacity was calculated as ORSC (%)×DPPH· in the reaction (g)/ sample weight (g), as described by Peng et al. (2000a).
Chlorophyll fluorescence
The maximum efficiency of PSII photochemistry (Fv/Fm) and ΦPSII
were measured using a portable pulse-modulated fluorimeterPAM-2100 (Walz, Efeltrich, Germany). Leaf disks were darkadapted with leaf clips for 30 min and measurements wereperformed at 2-h intervals on a sunny day. The Fv/Fm ratio wascalculated as (Fm–Fo)/Fm and ΦPSII was equal to 1–Fs/Fm', where Fm
and Fo are the maximum and basal fluorescence yields after darkadaptation, respectively, Fm' is the maximum fluorescence yield afterlight adaptation, and Fs is steady state fluorescence yield.
77-K Chl fluorescence emission spectra
Plant leaves under different natural light intensities were col-lected after they had been covered with a wet cloth. Beforemeasurements, leaves were cut into strips and inserted intoquartzose tubes. The 77-K Chl fluorescence emission spectrawere recorded using a fluorescence spectrophotometer withlow temperature affiliation (LS55; Perkin-Elmer, Norwalk, CT,USA). The excitation wavelength was 436 nm (slit 10 nm) andthe emission was between 600 and 800 nm (slit 10 nm).
Acknowledgements
The authors thank Jian Liu (South China Botanical Garden, theChinese Academy of Sciences) for his technical assistancewith the HPLC procedure.
References
Allen JF (1992). Protein phosphorylat ion in regulat ion of
photosynthesis. Biochim. Biophys. Acta 1098, 275–335.Bennett J (1991). Protein phosphorylation in green plant chloroplast.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 42, 281–311.Björkman O, Demmig B (1987). Photon yield of O2 evolution and
chlorophyll fluorescence characteristics at 77 K among vascularplants of diverse origins. Planta 170, 489–504.
Björkman O, Holmgren P (1963). Adaptability of the photosyn-thetic apparatus to light intensity in ecotype from exposed andshaded habitats. Physiol. Plant. 16, 889–914.
Bonaventura C, Myers J (1969). Fluorescence and oxygen evolu-tion from Chlorella pyrenoidosa. Biochim. Biophys. Acta 189,366–383.
Briantias JM, Vernotte C, Krause GH, Weis E (1986). Chlorophylla fluorescence of higher plants: Chloroplasts and leaves. In:Govindjee JA, Fork D, eds. Light Emission by Plants and Bacteria.Academic, New York. pp. 539–583.
Bruce D, Brimble S, Bryant DA (1989). State transitions in ap h y c o b i l i s o m e - l e s s m u t a n t o f t h e c y a n o b a c t e r i u mSynechococcus sp PCC 7002. Biochim. Biophys. Acta 974, 66–73.
Chen ZH, Wang BS, Zhang HD (1996). Productivity of EvergreenBroad-leaved Forest in South Subtropics. Guangdong HigherEducation Press, Guangzhou (in Chinese).
Demmig-Adams B, Adams WWIII (1996). The role of xanthophyllcycle carotenoids in the protection of photosynthesis. TrendsPlant Sci. 1, 21–26.
Demmig-Adams B, Adams WWIII (2000). Harvesting sunlight safely.Nature 403, 371–374.
Fitter A, Hay R (2002). Environmental Physiology of Plants, 3rdedn. Academic Press, London.
Gal A, Zer H, Ohad I (1997). Redox-controlled thylakoid proteinphosphorylation. News and views. Physiol. Plant 100, 869–885.
G i l m o r e A M ( 2 0 0 1 ) . X a n t h o p h y l l c y c l e - d e p e n d e n tnonphotochemical quenching in photosystem II: Mechanisticinsight gained form Arabidopsis thaliana L. mutants that lackviolaxanthin deepoxidase activity and or lutein. Photosynth.Res. 67, 87–99.
Gilmore AM, Yamamoto HY (1991). Resolution of lutein and zeax-anthin using a non-endcapped lightly carbon-loaded C18 high-performance liquid chromatographic column. J. Chromatogr.543, 137–145.
Gilmore AM, Yamamoto HY (1993). Linear models relatingxanthophylts and lumen acidity to non-photochemical fluores-cence quenching: Evidence that antheraxanthin explains zeax-anthin-independent quenching. Photosynth. Res. 35, 67–78.
Horton P (1983). Control of chloroplast electron transports by phos-phorylation thylakoid proteins. FEBS Lett. 118, 1–10.
Hundal T, Virgin I, Stryng S, Andersson B (1990). Changes of theorganization of photosystem II following light-induced D1-pro-tein degradation. Biochim. Biophys. Acta 1017, 235–241.
Ivanov AG, Velitchkova MY (1990). Heat induced changes of theefficiency of P700 photooxidation in pea chloroplast membranes.

Regulation of Photosynthesis in Subtropical Plants 361
J. Photochem. Photobiol. B: Biol. 4, 307–320.Krall JP, Edward GE (1992). Relationship between photosystem II
activities and CO2 fixation in leaves. Physiol. Plant. 86, 180–197.
Krause GH, Weis E (1991). Chlorophyll f luorescence andphotosynthesis. The basis. Annu. Rev. Plant Physiol. Plant Mol.Biol. 42, 313–349.
Kühlbrandt W, Wang DN, Fujiyoshi Y (1994). Atomic model ofplant light-harvesting complex by electron crystallography. Na-ture 367, 614–621.
Larrauri JA, Sanchez-Moreno C, Saura-Calixto F (1998). Effect oftemperature on the free radical scavenging capacity of extractsfrom red and white grape pomace peels. J. Agr. Food Chem. 46,2694–2697.
Long SP, Humphries S, Falkowski PG (1994). Photoinhibition ofphotosynthesis in nature. Annu. Rev. Plant Physiol. Plant Mol.Biol. 45, 633–662.
Müller P, Li XP, Niyogi KK (2001). Non-photochemical quenching.A response to excess light energy. Plant Physiol. 125, 1558–1566.
Niyogi KK, Bjorkman O, Grossman AR (1997). Chlamydomonasxanthophylls cycle mutants identified by video imaging of chlo-rophyll fluorescence quenching. Plant Cell 9, 1369–1380.
Noctor G, Foyer C (1998). Ascorbate and glutanthione: Keepingactive oxygen under control. Annu. Rev. Plant Physiol. PlantMol. Biol. 49, 249–279.
Osmond CB (1994). What is photoinhibition? Some insights fromcomparisons of shade and sun plants. In: Baker NR, Bowyer JR,eds. Photoinhibition of Photosynthesis. From Molecular Mecha-
nisms to the Field. BIOS Scientific, Oxford. pp. 1–24.Osmond CB, Anderson, JM, Ball MC, Egerton JJG (1999). Com-
promising efficiency: The molecular ecology of light-resourceutilization in plants. In: Press MC, Scholes JD, Barker MG, eds.Plant Physiology Ecology. Blackwell Science, Oxford. pp. 1–24.
Peng CL, Chen SW, Lin ZF, Lin GZ (2000a). Detection ofantioxidative capacity in plants by scavenging organic free radi-cal DPPH. Prog. Biochem. Biophys. 27, 658–661 (in Chinesewith an English abstract).
Peng CL, Lin ZF, Lin GZ (2000b). Effect of light on scavengingcapacity for organic free radical in leaves of four woody plants.Acta Bot. Sin. 42, 393–398 (in Chinese with an English abstract).
Peng SL (1996). Dynamics of Forest Community in South Subtropics.Science China Press, Beijing (in Chinese).
Siefermann-Harms D (1987). The light-harvesting and protectivefunctions of carotenoids in photosynthetic membranes. Physiol.Plant. 69, 561–568.
Somersalo S, Krause GH (1989). Photoinhibition at chillingtemperature. Fluorescence characteristics of unhardened andcold acclimated leaves. Planta 177, 409–416.
Valentini R, Epron D, De Angelis P, Matteucci G, Dreyer E (1995).In situ estimation of net CO2 assimilation, photosynthetic elec-tron flow and photorespiration in Turkey oak (Quercus cerris L.)leaves: Diurnal cycles under different levels of water supply.Plant Cell Environ. 18, 631–640.
Verhoeven AS, Bugos RC, Yamamoto HY (2001). Transgenic to-bacco with suppressed zeaxanthin formation is susceptible tostress-induced photoinhibition. Photosynth. Res. 67, 27–39.
(Handling editor: Cong-Ming Lu)


















