Regeneration of the eye margin in Periplaneta americana...
18
/. Embryol. exp. Morph. 76, 9-25 (1983) Printed in Great Britain © The Company of Biologists Limited Regeneration of the eye margin in Periplaneta americana (Insecta, Blattodea) By P. M. J. SHELTON 1 , H.-D. PFANNENSTIEL 2 AND E. WACHMANN 2 From the Zoology Department, University of Leicester and The Institutfiir Allgemeine Zoologie der Freien Universitdt, Berlin SUMMARY The regulative ability of the proliferative zone of the insect eye margin has been investigated in larval Periplaneta americana. After sections of the eye margin are removed the eye nevertheless recovers to form a normal shape. Using chimaeras of lavender and wild-type animals we were able to show that the margin can regenerate from the differentiated parts of the eye. When differentiated eye tissue is confronted with epidermis from the head capsule adjacent to the proliferative zone (the vertex), the regenerated margin always forms from the eye. There is no evidence that intervening levels can be intercalated between host and graft tissues when sections of the eye margin are moved to new circumferential levels. However, in that situation differences between tissues from non-adjacent circumferential positions lead to the rounding up of the graft and it fails to develop normally. INTRODUCTION The compound eyes of hemimetabolous insects grow throughout larval development by increase in the size of existing ommatidia and by the addition of more ommatidia to the perimeter of the eye (Meinertzhagen, 1973; Shelton, 1976). In Periplaneta americana eye growth is due mainly to the addition of ommatidia along the dorsal, anterior and ventral parts of the eye margin. There is no addition of ommatidia to the posterior margin (Nowel, 1981; Stark & Mote, 1981). We have shown that cells forming new ommatidia are derived from a proliferation zone within the eye margin (Nowel & Shelton, 1980; Nowel, 1981). This zone has been regarded as a persistent primitive portion of the original eye anlage in the embryo (Bodenstein, 1953). According to that view, cells within the proliferation zone are determined by their ancestry as eye-forming cells and removal of this anlage material should result in cessation of eye growth. There is some evidence to support this idea. If the whole eye including the proliferation zone is removed from a cockroach nymph, the wound becomes covered with 1 Author's address: Department of Zoology, Adrian Building, University of Leicester, Leicester LEI 7RH, U.K. 2 Authors' address: Institut fur Allgemeine Zoologie der Freien Universitat, Konigin-Luise- Str. 1-3, D-1000 Berlin 33.
Transcript of Regeneration of the eye margin in Periplaneta americana...

/. Embryol. exp. Morph. 76, 9-25 (1983)Printed in Great Britain © The Company of Biologists Limited
Regeneration of the eye margin in Periplaneta
americana (Insecta, Blattodea)
By P. M. J. SHELTON1, H.-D. PFANNENSTIEL2 ANDE. WACHMANN2
From the Zoology Department, University of Leicester and The InstitutfiirAllgemeine Zoologie der Freien Universitdt, Berlin
SUMMARY
The regulative ability of the proliferative zone of the insect eye margin has been investigatedin larval Periplaneta americana. After sections of the eye margin are removed the eyenevertheless recovers to form a normal shape. Using chimaeras of lavender and wild-typeanimals we were able to show that the margin can regenerate from the differentiated parts ofthe eye. When differentiated eye tissue is confronted with epidermis from the head capsuleadjacent to the proliferative zone (the vertex), the regenerated margin always forms from theeye. There is no evidence that intervening levels can be intercalated between host and grafttissues when sections of the eye margin are moved to new circumferential levels. However,in that situation differences between tissues from non-adjacent circumferential positions leadto the rounding up of the graft and it fails to develop normally.
INTRODUCTION
The compound eyes of hemimetabolous insects grow throughout larvaldevelopment by increase in the size of existing ommatidia and by the addition ofmore ommatidia to the perimeter of the eye (Meinertzhagen, 1973; Shelton,1976). In Periplaneta americana eye growth is due mainly to the addition ofommatidia along the dorsal, anterior and ventral parts of the eye margin. Thereis no addition of ommatidia to the posterior margin (Nowel, 1981; Stark & Mote,1981). We have shown that cells forming new ommatidia are derived from aproliferation zone within the eye margin (Nowel & Shelton, 1980; Nowel, 1981).This zone has been regarded as a persistent primitive portion of the original eyeanlage in the embryo (Bodenstein, 1953). According to that view, cells within theproliferation zone are determined by their ancestry as eye-forming cells andremoval of this anlage material should result in cessation of eye growth. Thereis some evidence to support this idea. If the whole eye including the proliferationzone is removed from a cockroach nymph, the wound becomes covered with
1 Author's address: Department of Zoology, Adrian Building, University of Leicester,Leicester LEI 7RH, U.K.
2 Authors' address: Institut fur Allgemeine Zoologie der Freien Universitat, Konigin-Luise-Str. 1-3, D-1000 Berlin 33.

10 P. M. J. SHELTON AND OTHERS
undifferentiated cuticle and eye regeneration never takes place (Nowel & Shel-ton, 1980). On the other hand, if the eye is only partially removed we havenoticed that the eye continues to grow and that local damage to the eye marginin P. americana nymphs has very little effect on the size and shape of the adulteye (Shelton, unpublished observations). This implies that damaged sections ofthe proliferative zone can reform by some regulative mechanism. The purposeof the present investigation was to provide conclusive evidence that this is so.Having established that the eye margin can regenerate, our next objective wasto establish the source of the cells forming the regenerated eye margin. Such cellscould be derived from any of three separate sources. First, they could come fromthe head epidermis adjacent to the proliferation zone (the vertex). Second, theycould be derived from the mature eye. Finally, the ablated section of the eyemargin could reform from the remaining eye margin to either side of the wound.The present experiments show that a new eye margin including the proliferativezone can reform from mature regions of the eye. There is some evidence that thenew eye margin forms partly by migration of cells into the wound from remainingparts of the eye margin. We cannot exclude the possibility that in certain circum-stances cells derived from the vertex can form eye margin but in our experimentswe obtained no evidence that this occurs.
MATERIALS AND METHODS
The culture conditions and operation procedure have been describedpreviously (Nowel & Shelton, 1980). Chimaeric eyes were generated by ex-changing sections of eye margin between wild-type and lavender (Ross, Cochran& Smyth, 1964) P. americana at stages 5 to 8 using recently moulted animals.In some cases a second operation was performed on the eye one or two moultstages later. Details of other types of operation are given in the results section.Most animals were photographed at each moult following the operation andthey were fixed as adults in a paraformaldehyde/glutaraldehyde mixture (Kar-novsky, 1965) and embedded in Araldite. Some specimens were fixed at larvalstages to examine the anatomy of the regenerated eye margin. Experimentalanimals were photographed using a Zeiss Tessovar Photomacrographic Zoomsystem. For this they were held under water following anaesthesis by cooling.This method eliminates unwanted reflections from the eye surface. Embeddedeyes were sectioned for light microscopy at 1-0 fjm and stained with toluidineblue in the usual way. To demonstrate the proliferation zone some animals wereinjected with a 1 % colchicine solution (0-5 jul/0-1 g live weight of animal) madeup in saline (Hoyle, 1953). For the SEM, specimens were mounted onaluminium stubs after critical-point drying. They were coated with a 1-3 nmlayer of gold using an ISI sputter-coating unit and examined with an ISI-60SEM.

Regeneration of the eye margin in Periplaneta americana 11
RESULTS
Location of the proliferation zone within the eye margin
The anatomy of the nymphal cockroach eye margin has already been described(Nowel, 1981). At the extreme edge of the eye there is a region of undifferen-tiated and dividing cells; this is the proliferation zone. It is restricted to thedorsal, anterior and ventral parts of the circumference; new ommatidia are notadded to the posterior margin. Separating the proliferation zone from the matureommatidia of the differentiated eye is the maturation zone. This region containspreommatidia in various states of differentiation with occasional dividing cellsbetween them (Nowel, 1981). The least differentiated ommatidia are closest tothe proliferating zone and the most differentiated ones are closest to the fullydifferentiated part of the eye. In the present investigation the regions of the eyecontaining the proliferation zone were identified by injecting animals (3 daysafter moulting) with colchicine and fixing them for sectioning after 12 h. Insubsequent experiments to remove the eye margin this entire region was cut out(Fig. 1A).
Regeneration of the eye after removal of a section of dorsal eye margin
To test the regenerative powers of the eye margin, strips of tissue containingabout one third of the dorsal eye margin were removed from nymphs at moultstages 5 to 8. In different animals the tissue was removed from the anterior,middle or the posterior regions of the dorsal eye margin (Fig. 2A, B, C). Thecontralateral eye was used as an unoperated control. The results were similar inall cases. The animals were allowed to develop to the adult stage, 5 or 8 moultslater. If no eye margin regeneration had taken place, that part of the eye norm-ally derived from the ablated margin should have failed to form. So, for instance,where the posterior third of the dorsal eye margin had been removed the pos-terior third of the eye should have ceased to grow, resulting in a large notch inthe normally smooth outline of the eye. Regular inspection of operated animalsshowed that eye growth in the damaged region was temporarily interrupted forone or two moults after the operation. The damage was visible in all animals asa zone of undifferentiated cuticle within and at the edge of the eye. However,after two moults a new margin had formed in the damaged area and eye growthin that sector had resumed. Thus inspection of animals at the adult stage revealedoperated eyes of more or less normal shape and the dorsal regions of the eyeswere similar in size and shape to the contralateral controls (Figs 3A, B). Theseobservations provide conclusive proof that the eye margin can reform followingablation.
Regeneration of the eye margin in \avender/wild-type chimaeric eyes
Although the previous experiment shows that the eye margin can regenerate,

12 P. M. J. SHELTON AND OTHERS
Fig. 1

Regeneration of the eye margin in Periplaneta americanaB c
13
Fig. 2. Effects of removing a third of the dorsal eye margin from a nymph (left,central third; centre, posterior third; right, anterior third). In all cases the adult eyehas a normal appearance. A region of scar tissue (dotted) is the only detectableconsequence of the operation.
it says nothing about the origin of the cells which form the reconstitutedproliferation zone. We used chimaeras in an attempt to answer this question.Chimaeric eyes consisting of patches of eye tissue carrying a distinctiveautonomous pigment marker were generated by exchanging dorsal eye materialbetween lavender and wild-type individuals. The grafts were oblong and ofvariable size being one quarter to one half the length of the dorsal eye margin.The grafts consisted of eye margin with narrow regions of adjacent vertex epider-mis and mature eye to either side of it (Fig. 3C, 4A, B). In 19 cases the graft wasaccepted and continued to grow. After one or two moults, a second operationwas performed to surgically remove the eye margin from the graft. The tissueremoved extended at least three rows of facets into the eye and contained a largearea of vertex material. This ensured that any vertex material implanted with theoriginal graft was removed and that none of the proliferation zone was left
Fig. 1. Micrographs showing sections through the eye margins in control and experi-mental nymphs of P. americana.
(A) Normal eye, showing the organization of the eye margin and the parts thatwere removed or transplanted in the various experimental procedures. Theproliferation zone (pz) is located at the extreme edge of the eye and contains amitotic figure (arrow) arrested with colchicine. Dividing cells are also found in thematuration zone (mz) which contains partly differentiated ommatidia. A well dif-ferentiated cone (c) indicates the limit of the maturation zone. In removing a marginduring ablation or transplant experiments, a region containing the zones between thedashed lines was removed together with equal areas of mature eye and head epider-mis (e) to either side.
(B) The regenerated eye margin reformed from the grafted tissue in a chimaerafrom which the graft's eye margin had been removed. The major eye margin zoneshave reformed and the proliferation zone (pz) contains a dividing cell (arrow). Otherlabels as above.
(C) An eye margin of normal appearance which has reformed from the grafted eyetissue after confrontation of mature eye with head epidermis. Mitotic figure arrowed.Other labels as before. Bars represent 25jum.

P. M. J. SHELTON AND OTHERS
Fig. 3

Regeneration of the eye margin in Periplaneta americana 15behind. The wound was repaired with an implant of vertex material from anotheranimal (Figs 3D, 4A, B). In one half of the animals the vertex was taken froman animal of host phenotype, in the rest it was taken from an animal of thephenotype of the original eye graft. Subsequent eye development was followedover successive moults. The results for both combinations of graft were similar.In the majority of cases (12 animals) the marked eye implant continued to growshowing that a new eye margin had formed (Figs 3E, F; 4A, B). Regeneratedmargins of selected individuals were fixed and sectioned at larval stages beforethe cessation of eye growth. They had the appearance of normal eye margins witha characteristic proliferation and maturation zone (Figs 1A, B). Ommatidiaformed by the regenerated margin were normal with the usual complement ofreceptor, cone and pigment cells. Most animals were allowed to develop to theadult stage before being fixed. Each animal was photographed at each moultstage following the operation. Comparisons of the same animal at differentstages showed that there was a regular increase in numbers of ommatidia withthe pigmentation of the original eye graft (Figs 3E, F). These combined observa-tions show that the regenerated margin is providing a continual supply of newcells to the eye and that the patterns observed are not due to the redistributionof ommatidia from the original graft. Where the implant of vertex was of hosttype and the regenerated eye had the phenotype of the original eye implant thisshowed that the eye itself can form a margin. In all cases where the regenerated
Fig. 3. A series of macrophotographs showing the effects of various types of opera-tion procedure. After removal of the posterior third of the dorsal eye margin froma nymph the resulting adult eye has an approximately normal shape (A). Comparisonof the operated (right) eye with the control shows that the eyes are both the same size(B). The only indication of the operation is a region of scar tissue (s). Figs 3C-Fillustrate the effects of removing the eye margin from the grafted eye in a chimaera.Chimaeras were constructed by implanting a section of donor eye margin (C). At alater nymphal stage the eye margin in the vicinity of the graft was removed andreplaced with an implant of head epidermis (D). Two moults later the original eyegraft continues to grow indicating the regeneration of the margin (E). The resultingadult eye is shown in (F). The continual addition of new ommatidia shows that thereis true growth of the original graft and not the redistribution of ommatidia fromwithin it. In some of these experimental animals the graft failed to reform a new eyemargin (G). In another type of experiment the implanted eye tissue came from thecentre of a donor eye (H). In some cases such grafts formed a margin between headcapsule and eye. Fig. 31 shows the same eye at a later stage. In a number of cases theeye margin apparently reformed but later petered out (J). In many cases the graftdid not reform a margin but was surrounded by host-type ommatidia (K). Removalof the eye margin to either side of the implanted mature eye leads to the formationof a graft-type margin in all cases and a relatively much wider region of regeneratedeye margin (L). The bottom row of figures (3M-P) shows the effect of implanting arectangular section of vertex epidermis across the eye margin. One moult after thegraft the rectangular graft has become rounded (M). After two moults it has with-drawn from the eye (N). Later, ommatidia have reformed in the previously damagedregion (O). In the adult, the eye has a more or less normal shape and the onlyevidence of the operation is scar tissue in the eye (P). Figs 3A, B, bar represents1-0 mm. Figs 3C-P, bar represents 0-5 mm.

16 P. M. J. SHELTON AND OTHERS
eye margin had the phenotype of the original eye implant, the shape of the graft-derived eye had a common pattern with a constriction at the point where the eyemargin of the implant had been removed (Figs 3E, F; 4A, B). Our interpretation
Fig. 4

Regeneration of the eye margin in Periplaneta americana 17of this observation is that the wound is repaired in part by the inward migrationof host cells from either side of the graft. The same pattern of constriction is seenwhether the vertex implant is of host or donor type. Where the vertex implantis of the donor eye type, the inwardly moving cells can only have come from hosteye. In some cases (seven animals) implanted eye tissue failed to form a newmargin and consequently ended up isolated from the edge of the eye byommatidia of host phenotype (Figs 3G, 4A, B). Once again identical results wereobtained for both types of vertex implant. Certainly where the vertex was froman animal of the same eye colour as the implanted eye (four cases), the cellswhich form the regenerated margin (of host phenotype) must have come fromthe host eye. Here we must assume that the inward migration of cells is thedominant process and occurs before the implanted eye has time to regenerate itsown margin. These combined observations show that when the eye margin isremoved it can regenerate from the remaining eye tissue of the eye graft but thatit may be partly restored by the invasion of eye cells from the host to either side ofthe graft. We consider the most likely source of such host cells is the eye margin.The experiments do not exclude the possibility that in certain circumstances theregenerated eye margin can form from vertex. We have tested the ability of thevertex to contribute cells to the regenerated margin in later experiments.
Regeneration of the eye margin from implants of mature eye
Although we were careful to remove all of the eye margin from the graft-derived eye in the previous experiment (the sections of cuticle removed werealways inspected to confirm that the underlying eye margin was attached), wewished to eliminate the possible criticism that we had left a few cells behind. Thefollowing experiments show that our previous conclusions were justified. Onceagain we used wild-type and lavender stocks to identify graft- and host-derivedtissues. In this case we made chimaeras at nymphal stages by implanting squaregrafts of mature eye tissue from the centre of donor eyes into similar shaped sitesat the dorsal margins of host eyes (Figs 3H; 4C). This type of graft confrontedmature eye with the vertex. Since the implant came from the centre of the eyeit could not possibly include eye margin cells. If our previous results representedtrue regeneration of the eye margin, we predicted that a new margin should
Fig. 4. Chimaeric eyes were generated by exchanging sections of eye margin be-tween wild-type and lavender nymphs (A, B). One or two moults later a square oftissue containing the eye margin and adjacent head epidermis was removed andreplaced with a square of head epidermis. Some eye grafts failed to grow and becamesurrounded by host ommitidia. In the majority of cases a new eye margin formed andthe graft continued to grow. In a second type of operation (C) a square of tissue fromthe centre of a donor eye was implanted into a suitable site in the eye of a host so thatthe graft confronted the vertex along one edge. Some of these grafts did regeneratea narrow section of eye margin although the majority did not. The success rate wasimproved to 100 % by removing host eye margin to either side of the implant at thefollowing moult (D).

18 P. M. J. SHELTON AND OTHERS
regenerate at the border between the graft eye tissue and the head capsuleepidermis. A number of these animals did regenerate an eye margin derivedfrom the graft although the success rate was rather low (3/20 animals) (Figs 31,J; 4C). In two cases the implant became separated from the margin only after twoor three moults. In these cases it appeared that the graft had initially reformeda small section of eye margin but that after several moults it petered out (Fig. 3J).A similar result can occur after a control eye margin graft where the length of theimplanted margin is short (Shelton, unpublished observation). This would beexpected if cell division within the eye margin is under some sort of probabilisticcontrol. It argues against a population of stem cells. While this type of graft hada low success rate, we were able to increase the success rate to 100% (nineanimals) by removing substantial sections of the host eye margin to either sideof the graft together with the dorsal edge of the implant, one moult after theinitial operation (Figs 3L; 4D). This finding provides support for the hypothesisthat cells from the host eye margin can migrate into the wounded area at the edgeof the eye. When these cells are removed, the eye margin regenerates mainlyfrom the implanted mature eye. Once again we sectioned eyes in the region ofthe regenerated margin and found that there was a normal appearance (Fig. 1C)and that the ommatidia derived from it had the normal complement of cells.
Implantation of marked head capsule (vertex) epidermis into the larval eye
The previous experiments show that the eye can reform a new proliferationzone from mature eye. The following experiments were designed to test theability of vertex tissue to form eye margin. Once again we used exchanges ofgrafts between lavender and wild-type animals so that the source of anyregenerated eye cells could be identified. In 50 % of the cases we used a lavenderhost and wild-type donor; in the rest we used the opposite combination. Theresults were the same for both combinations. Two types of experiment wereperformed, the object of both being the same. That was to confront epidermisfrom the vertex with mature eye and see if a new margin would form from vertex-derived cells at the junction between the two kinds of tissue. In the first type ofexperiment, a 250/im square of vertex epidermis from close to the eye marginwas implanted into the centre of the host eye at a site prepared by removing anappropriately sized piece of mature eye tissue (Fig. 5A). In all cases (25 animals)the epidermis was sloughed off after the first postoperative moult. We assumethat this failure of grafts to 'take' is due to the large differences in cell surfaceproperties of the cuboidal epidermal cells and the spindle-shaped eye cells.
In order to overcome this difficulty, a second series of experiments was devisedusing lavender and wild-type tissues as before but in a configuration where thevertex graft made contact with both the head capsule and the eye. The graftconsisted of a rectangular piece of vertex tissue. It was implanted into a rectan-gular site whose long axis crossed the eye/vertex border at right angles (Fig. 5B).These grafts had a low success rate (7/30 animals) but a significant number could

Regeneration of the eye margin in Periplaneta americana 19
B
Fig. 5. The grafts illustrated here confront vertex epidermis with eye tissue. Squaresof vertex implanted into the centre of the eye are lost by the following moult (A)leaving only a small area of scar tissue. Rectangular grafts of vertex crossing the eyeborder are retained longer (B). However, the vertex material rounds up and with-draws from the eye, the eye margin regenerates from host tissue and the adult eyehas a more or less normal shape.
be followed over a number of moults. We assume the improved success rate wasdue to the fact that one end of the graft was in continuity with the head capsule.Where the grafts were successful the part of the implant within the eye wasrounded and had lost its original rectangular shape. It often formed a bumpabove the smooth contour of the adjacent eye (Fig. 3M). By the second post-operative moult the graft had withdrawn completely from the eye and formed arounded protrusion just outside the eye in a typical case (Fig. 3N). This left alarge area of scar tissue at the edge of the eye. By the third postoperative moultthe two sides of the eye margin that had been separated previously by the grafthad become confluent (Fig. 3O). Subsequent growth resulted in an adult eye inwhich the dorsal eye region had normal proportions even though a significantpart of the original eye margin had been removed and replaced with vertex (Fig.3P). From direct observations it was clear that, in this typical case, the eyemargin reformed in the damaged area once the vertex implant had withdrawn.In all cases ommatidia in the vicinity of the regenerated amrgin were of hostphenotype. The implant never contributed cells to the regenerated eye margin,it must have formed from the eye tissue. These results do not prove that thevertex cannot in other circumstances redifferentiate to form eye margin but in

20 P. M. J. SHELTON AND OTHERS
this situation it is unable to do so. They do indicate that there are considerabledifferences between head capsule and eye and that these differences cause animplant to withdraw from the eye.
Effects of local reversal of the eye margin
So far we have been concerned with regulative behaviour mainly along theradial axis of the eye. Next we wished to establish whether there is any evidencefor regulation around the circumferential axis. Where pattern regulation occursin postembryonic development of insects, missing intervening zones can be inter-calated when non-congruent levels are combined (French, 1981; Wright &Lawrence, 1981). In this series of experiments non-congruent parts of the eyemargin were confronted by exchanging sections of nymphal eye margin betweenleft and right eyes (Fig. 6C). In a parallel set of control operations, sections ofeye margin were exchanged between eyes of the same side (either left or right)(Fig. 6A). Once again we used lavender and wild-type animals for each ex-change. Exchanging grafts between left and right sides produces a local reversalof the eye margin and consequently brings different regions of the circumferenceinto juxtaposition. The grafts were of a standard size and included a 250jumlength of dorsal eye margin and several rows of ommatidia. The animals werephotographed at each moult stage following the operation and they were fixedas adults. The eyes with a section of the eye margin reversed differed from thecontrols in only one detectable way. After one or two moults, tissues derivedfrom the reversed eye margin protruded beyond the normally smooth surfacecontours of the eye to form a slight bump. As development continued thisunevenness gradually disappeared so that in the adult the eye surface was assmooth after reversals as it always was after controls. The other feature of thegrafts that we considered was their overall shape (Figs 7C, D). For each success-ful reversal (8 animals) we made a direct comparison with a control operation tothe same part of the dorsal eye margin (Fig. 7A). We had access to at least 40control operations from a previous study (Nowel, 1981) in addition to 10 of ourown. We found that the shape of the graft-derived eye after a reversal was notdetectably different from the appropriate control (Fig. 7D). If intercalation hadtaken place after reversals two consequences should follow. First, the overall sizeof the eye should be larger compared with the contralateral unoperated eye orwith the relevant control graft. This is because intercalation should increase thelength of the eye margin. Second, the graft-derived retina after reversals shouldbe larger than in controls for the same reason. Neither of these predictions wasborne out (figs 6C; 7A, C, D). We concluded that no intercalation had takenplace.
Exchanges between dorsal and anterior eye margin
As a second test for intercalation we exchanged similar 250 jum sections of eyemargin between the dorsal and anterior parts of the eye (Fig. 6D). The results
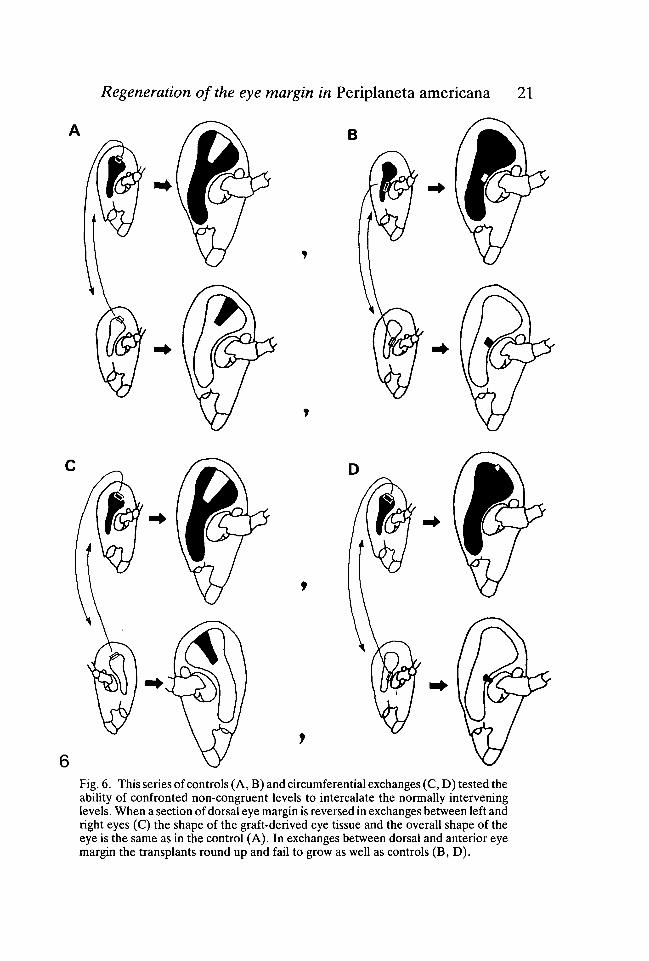
Regeneration of the eye margin in Periplaneta americana 21
Fig. 6. This series of controls (A, B) and circumferential exchanges (C, D) tested theability of confronted non-congruent levels to intercalate the normally interveninglevels. When a section of dorsal eye margin is reversed in exchanges between left andright eyes (C) the shape of the graft-derived eye tissue and the overall shape of theeye is the same as in the control (A). In exchanges between dorsal and anterior eyemargin the transplants round up and fail to grow as well as controls (B, D).

22 P. M. J. SHELTON AND OTHERS
/
\ • f
Fig. 7. Control grafts (A, B) show the normal pattern of eye growth. Figs 7C & Dshow the same experimental animal at two different stages. When sections of the eyemargin are reversed as in (C) the shape of the resulting adult graft-derived eye (D)is indistinguishable from a control graft to the same part of the eye margin. In Fig.7C eye tissue dorsal to the arrows has formed after grafting operation. Dotted linesin Fig. 7D show the outlines of graft-derived eye in a comparable control. When eyemargin is grafted from anterior to dorsal regions (E) and vice versa (F, G. H) the graftrounds up shortly afterwards (E) and forms a bumpy projection. Figs 7F, G & H showthe resulting adult eye of an animal that received dorsal eye margin in an anteriorlocation. A side view shows the rounding of the graft (F); seen from a dorsal positionthe typical projection is visible (G). An SEM preparation (H) viewed from theanterior shows how the graft has become isolated from the anterior edge of the eye(lower left). Figs 7A-G, bar represents 0-5 mm. Fig. 7H, bar represents 100/an.
of these experiments were compared with control grafts at each of the two sites(Figs 6A, B; 7A, B). The notable feature of the exchanges was the relatively lowsuccess rate. Often there was no sign of the graft at the following moult (21animals). Where the graft was detectable at the new site subsequent growth wasextremely poor (10 animals). In most cases the implanted graft rounded up; inextreme cases it produced a raised projection (Figs 7E, F, G, H). It seems thatnormally the host eye margin to either side of the graft outcompetes the graftedmargin, the graft becomes isolated from the edge of the eye and it ceases to grow.The control grafts show that the eye grows much more at the dorsal than at theanterior region (Figs 6A, B; 7A, B). The failure of the anterior grafts to growsatisfactorily in a dorsal location could be attributed to differential growth rates.

Regeneration of the eye margin in Periplaneta americana 23However, in the converse situation, implanted dorsal margin should be an ad-vantage over the anterior host margin. Nevertheless, even here the implantsfailed to grow properly and were outcompeted. If intercalation occurs, a graft ofthe sort performed here should produce a grossly abnormally shaped eye, and thegraft-derived eye should expand with successive moults. The interpretation ofthese results is that intercalation does not occur but that different parts of the eyemargin are non-equivalent in some way. Thus grafts from one region to anotherfail to develop properly in the new site.
DISCUSSION
These experiments provide the first conclusive proof that the eye margin hasregulative properties. Although probably there is normally an unbroken lineagerelating the postembryonic proliferation zone of the eye to the original opticanlage (Bodenstein, 1953), that lineage can be broken and a regenerated marginwill give rise to perfectly normal eye tissue. The results are consistent with thoseof Mouze (1972) using dragonflies. However, our results are more conclusivebecause we were able to use grafts carrying autonomous cellular markers. In thecase of the dragonfly experiments no markers were available and cell migrationfrom the remaining eye margin could have restored the ablated section.
The clearest result was that the eye itself can regenerate a proliferation zone.Although one has to be cautious about the possible failure to completely removethe ablated section, we took great trouble to ensure that this always occurred.The most persuasive argument is that fully differentiated eye tissue can reforma dividing margin when confronted with vertex epidermis. This result satisfies usthat true regeneration does occur.
The shape of the graft-derived eye tissue following removal of an implantedeye margin tells us something of the processes of wound repair and regeneration.A consistent feature is the constriction which occurs in the outline of the graft-derived tissue at the point where the grafted eye margin was removed. In somecases this constriction is total showing that the graft has not regenerated a mar-gin. Where the eye and vertex implants both carry the same marker to distinguishthem from the host, the constriction can only be caused by inward movementsof host cells from either side of the eye implant. The most likely source of cellsis the host eye margin but without direct evidence we cannot exclude the possibil-ity that the invading cells come from slightly more mature parts of the eye.
Our conclusions rest upon the assumption that the trails of graft cells extendingto the edge of the eye are the result of true growth and not the redistribution ofcells from within the graft. This conclusion is justified because sections throughthe regenerated region reveal a normal eye margin with proliferation andmaturation zones. Also the number of ommatidia with the graft phenotypeincreases in a regular way from moult to moult and they have the normal comple-ment of receptor, pigment and cone cells.

24 P. M. J. SHELTON AND OTHERS
At present we do not know which class or classes of cells are respecified to formeye margin. However, we are well aware that dividing cells are found in thematuration zone and in mature parts of the eye (Anderson, 1978; Nowel, 1981).In the maturation zone ommatidia already have the full complement of receptorand cone cells so it is likely that the dividing cell population normally gives riseto pigment cells. This is supported by the fact that such dividing cells are oftenfilled with pigment grains characteristic of secondary pigment cells (Shelton,unpublished observations). During normal development, cell division within themature region of the eye is probably associated with the need for more secondarypigment cells as the ommatidia increase in size (Nowel, 1981). It seems unlikelythat highly differentiated receptor cells could revert to a primitive undifferen-tiated state where they are capable of cell division. That is also the case for thecone cells. However, since secondary pigment cells seem capable of cell divisionit is possible that they could revert to a more primitive condition. The questionof whether the new proliferation zone is formed from the dividing cell populationor from dedifferentiation of specialized cells, requires further investigation.
The pattern-forming mechanism that allows the regeneration of a proliferationzone also remains obscure but we see no reason why it should be the same onethat allows the regeneration of cuticular ridges (Stumpf, 1968) or even segmentalboundaries (Wright & Lawrence, 1981). This could alter the way we think of themechanisms for setting up patterns within the eye. While we thought of the eyemargin as a persistent primitive part of the eye anlage it was logical to assumethat it was the eye margin which acted as a progress zone and laid down thepattern of positional values within the eye. If the eye margin reforms by the samesort of mechanism that causes predictable cuticular features to form at particularsegment levels, then it is logical to assume that the eye margin does not generatea sequence of positional values but that it has fixed positional value and inconsequence cells at that level form a proliferation zone.
While we could establish regulative ability in the radial axis of the eye that wasnot the case for the circumferential axis. It is known that cells in different regionsof the eye margin divide at different rates (Nowel, 1981). The controlling factorcould be related to differences in positional value around the circumference. Ourexperiments to shift particular circumferential levels to new locations on theperimeter of the eye were designed with the idea that intercalation of interveninglevels would occur wherever differences in positional information are present.Since intercalation does not occur, we concluded that the circumferential axis ofthe eye may be organized in the same way as the mediolateral axis of the insectsegment. Here with only one known exception (the ecdysial line; Shelton, 1979),transplants along this axis do not lead to intercalation. However, in the case ofthe eye, transplanted sections of eye margin do not grow normally. Characteris-tically they round up and the neighbouring host margin cells seem to outcompetethe transplant. This tendency of grafts to round up would be consistent with theoccurrence of adhesive differences at different points on the circumference. Such

Regeneration of the eye margin in Periplaneta americana 25adhesive differences are thought to occur in other insect systems (Nardi &Kafatos, 1976; Nardi, 1977; Niibler-Jung, 1977) and they cannot be excludedhere. The tendency of vertex grafts to withdraw from the eye could also beinterpreted in terms of differences in adhesive properties.
Our results lead us to conclude that it is only the eye itself that can regeneratea new margin. This would explain why it is that the Periplaneta eye never reformswhen the whole eye is removed (Nowel & Shelton, 1980) but that the eye iscapable of considerable regulation when it is only partially ablated.
REFERENCESANDERSON, H. (1978). Postembryonic development of the visual system of the locust, Schisto-
cerca gregaria. 1. Pattern of growth and developmental interactions in the retina and opticlobe. J. Embryol. exp. Morph. 45, 55-83.
BODENSTEIN, D. (1953). Postembryonic development. In Insect Physiology (ed. K. D.Roeder), pp. 822-865. New York: Wiley.
FRENCH, V. (1981). Pattern regulation and regeneration. Phil. Trans. R. Soc. Lond. B 295,601-617.
HOYLE, G. (1953). Potassium ions and insect nerve muscle. J. exp. Biol. 30, 121-135.KARNOVSKY, M. J. (1965). A formaldehyde-glutaraldehyde fixative of high osmolarity for use
in electron microscopy. J. Cell Biol. 27, 137A.MEINERTZHAGEN, I. A. (1973). Development of the compound eye and optic lobe of insects.
In Developmental Neurobiology of Arthropods (ed. D. Young), pp. 51-104. London: Cam-bridge University Press.
MOUZE, M. (1972). Croissance et metamorphose de l'appareil visuel des Aeschnidae(Odonata). Int. J. Insect Morphol. & Embryol. 1,181-200.
NARDI, J. B. (1977). The construction of the insect compound eye: the involvement of celldisplacement and cell surface properties in the positioning of cells. Devi Biol. 61,287-298.
NARDI, J. B. & KAFATOS, F. C. (1976). Polarity and gradients on lepidopteran wing epidermisII. The differential adhesiveness model: gradient of a non-diffusible cell surface parameter./. Embryol. exp. Morph. 36, 489-512.
NOWEL, M. S. (1981). Postembryonic growth of the compound eye of the cockroach. J.Embryol. exp. Morph. 62, 259-275.
NOWEL, M. S. & SHELTON, P. M. J. (1980). The eye margin and compound eye developmentin the cockroach: evidence against recruitment. /. Embryol. exp. Morph. 60, 329-343.
NUBLER-JUNG, K. (1977). Pattern stability in the insect segment. 1. Pattern reconstitution byintercalary regeneration and cell sorting in Dysderous intermedius Dist. Wilhelm Roux'sArch, devl Biol. 183, 17-40.
Ross, M. H., COCHRAN, D. G. & SMYTH, T. (1964). Eye-color mutations in the Americancockroach, Periplaneta americana. Ann. ent. Soc. Am. 57, 790-792.
SHELTON, P. M. J. (1976). The development of the insect compound eye. In Insect Develop-ment - Symposia of the Royal Entomological Society, no. 8 (ed. P. A. Lawrence), pp.152-169. Blackwell Scientific Publications.
SHELTON, P. M. J. (1979). Post-embryonic determination of the ecdysial line in the cockroach:evidence for pattern regulation in the medio-lateral axis./. Embryol. exp. Morph. 49,27-46.
STARK, R. J. & MOTE, M. I. (1981). Post-embryonic development of the visual system ofPeriplaneta americana. 1. Patterns of growth and differentiation. J. Embryol. exp. Morph.66, 235-255.
STUMPF, H. (1968). Further studies on gradient-dependent diversification in the pupal cuticleof Galleria mellonella. J. exp. Biol. 49, 49-60.
WRIGHT, D. A. & LAWRENCE, P. A. (1981). Regeneration of the segment boundary in Onco-peltus. Devi Biol. 85, 317-327.
(Accepted 22 April 1983)