
Purification, Characterization, and Localization Protein ... · 1322 DUBREUIL ET AL. on the surface...
8
JOURNAL OF CLINICAL MICROBIOLOGY, June 1990, p. 1321-1328 0095-1137/90/061321-08$02.00/0 Copyright C 1990, American Society for Microbiology Purification, Characterization, and Localization of a Protein Antigen Shared by Thermophilic Campylobacters J. DANIEL DUBREUIL, MAGDALENA KOSTRZYNSKA, SUSAN M. LOGAN, LORI A. HARRIS, JOHN W. AUSTIN, AND TREVOR J. TRUST* Department ofBiochemistry and Microbiology, University of Victoria, Victoria, British Columbia, V8W 2 Y2, Canada Received 23 October 1989/Accepted 14 March 1990 A protein antigen with an apparent molecular weight (Mr) of 31,000 was isolated from 0.2 M glycine hydrochloride (pH 2.2) extracts of a typical human fecal isolate, Campylobacterjejuni VC74. The protein was purified to homogeneity on a preparative scale by immunoaffinity chromatography followed by molecular sieving with a Superose 12 column. Isoelectric focusing under nondenaturing conditions indicated a pI of 9.3, and amino acid composition analysis showed that the protein was unusually rich in lysine, containing 14.9 mol% of this basic amino acid. Cysteine and tryptophan were absent. The protein also contained approxi- mately 35% hydrophobic amino acid residues, and N-terminal amino acid analysis showed that 17 of the first 38 residues were hydrophobic. This amino-terminal sequence to residue 22 was virtually identical to that of an antigenically cross-reactive 31,000-Mr protein isolated from another C. jejuni strain belonging to a different heat-labile serogroup. Western blotting (immunoblotting) of glycine extracts of other C. jejuni, Campylobacter coli, and Campylobacter laridis strains belonging to different thermolabile and thermostable serotypes, as well as Campylobacterfetus, with a rabbit polyclonal antiserum raised against the purified C. jejuni VC74 protein showed that all C. jejuni, C. coli, and C. laridis strains tested contained a 31,000-Mr protein with epitopes which were antigenically cross-reactive with the C. jejuni VC74 protein. The antigenically cross-reactive epitopes of this protein were also readily detected by immunodot blot assay of glycine extracts of C. jejuni, C. coli, and C. taridis with monospecific polyclonal antisera to the 31,000-Mr protein, suggesting that this serological test could be a useful addition to those currently employed in the rapid identification of these important pathogens. Slide agglutination reactions, immunofluorescence assay, and immunogold electron microscopy with antisera to purified 31,000-Mr protein and trypsin treatment of whole cells indicated that the cross-reactive epitopes of the 31,000-Mr protein were not exposed on the cell surface. Cell fractionation analysis and immunogold electron microscopy located the protein on the outer surface of the cytoplasmic membrane. This finding suggests that the 31,000-Mr protein is not a good candidate for inclusion in a monovalent subunit Campylobacter vaccine. The thermophilic campylobacters, especially Campylo- bacterjejuni and Campylobacter coli, are now recognized as important gastrointestinal pathogens of humans, contribut- ing substantially to morbidity in developed countries and to high mortality in developing countries (7, 44). Because of the high incidence of Campylobacter disease worldwide, there is a need for effective vaccines to protect against infection by this group of organisms. The frequency of isolation of thermophilic campylobacters also means that they are the subject of many laboratory identification investigations and subsequent biotyping and serotyping studies. Unfortunately, despite numerous studies, campylobacters remain relatively difficult to rapidly identify, classify, and type biochemically. Members of the genus Campylobacter neither ferment nor oxidize carbohydrates and are inert in many other typical biochemical test media (15, 22, 36), so there are only a few criteria which can be simply applied in their rapid identifi- cation (14, 15, 22, 26, 42). A simple monovalent serological assay which could rapidly identify thermophilic campylobac- ters would clearly be useful. However, attempts to develop such a test have been unsuccessful because of the consider- able serological diversity of the thermophilic campylobac- ters. This antigenic heterogeneity has also hampered vaccine development. For example, the heat-stable scheme of Pen- ner and Hennessy (37) now contains >76 serotypes (Penner, personal communication), while the heat-labile scheme of * Corresponding author. Lior (27) contains >100 serotypes (Lior, personal commu- nication). Western blot (immunoblot) studies of thermophilic Cam- pylobacter species have identified a small number of proteins with antigenically cross-reactive epitopes (3, 4, 29, 32). The immunodominant protein antigen of the thermophilic cam- pylobacters is flagellin with a subunit molecular weight (Mr) of 59,000 to 61,000. However, while this protein carries cross-reactive epitopes, antibody absorption experiments employing live motile cells indicate that serospecific epitopes are predominant on the surface of the native flagella filament (30). Furthermore, there are numerous serotypes of Campylobacter flagella (13, 16, 27, 45, 46) and these flagella are also subject to both phase and antigenic variation (6, 13). This means that the Campylobacter flagella protein is neither a straightforward candidate for use in a monovalent vaccine nor as a target for an antibody-based identification test for thermophilic campylobacters. A second protein with cross- reactive epitopes is the major outer membrane (OM) protein (3, 4, 29, 32), but again, the majority of cross-reactive epitopes in this molecule likely represent conserved amino acid sequences buried in the OM (8, 19). The third protein with cross-reactive epitopes is a protein with an apparent molecular weight of 31,000, which has been identified in glycine hydrochloride (pH 2.2) extracts of cells of thermo- philic campylobacters (3, 4, 29, 32). Indeed, the ability to remove the 31,000-Mr protein from cells by this simple extraction procedure has led to the widely held notion that the antigenically cross-reactive 31,000-Mr protein is located 1321 Vol. 28, No. 6 on August 28, 2019 by guest http://jcm.asm.org/ Downloaded from
Transcript of Purification, Characterization, and Localization Protein ... · 1322 DUBREUIL ET AL. on the surface...

JOURNAL OF CLINICAL MICROBIOLOGY, June 1990, p. 1321-13280095-1137/90/061321-08$02.00/0Copyright C 1990, American Society for Microbiology
Purification, Characterization, and Localization of a Protein AntigenShared by Thermophilic Campylobacters
J. DANIEL DUBREUIL, MAGDALENA KOSTRZYNSKA, SUSAN M. LOGAN, LORI A. HARRIS,JOHN W. AUSTIN, AND TREVOR J. TRUST*
Department ofBiochemistry and Microbiology, University of Victoria, Victoria, British Columbia, V8W 2 Y2, Canada
Received 23 October 1989/Accepted 14 March 1990
A protein antigen with an apparent molecular weight (Mr) of 31,000 was isolated from 0.2 M glycinehydrochloride (pH 2.2) extracts of a typical human fecal isolate, Campylobacterjejuni VC74. The protein was
purified to homogeneity on a preparative scale by immunoaffinity chromatography followed by molecularsieving with a Superose 12 column. Isoelectric focusing under nondenaturing conditions indicated a pI of 9.3,and amino acid composition analysis showed that the protein was unusually rich in lysine, containing 14.9mol% of this basic amino acid. Cysteine and tryptophan were absent. The protein also contained approxi-mately 35% hydrophobic amino acid residues, and N-terminal amino acid analysis showed that 17 of the first38 residues were hydrophobic. This amino-terminal sequence to residue 22 was virtually identical to that of anantigenically cross-reactive 31,000-Mr protein isolated from another C. jejuni strain belonging to a differentheat-labile serogroup. Western blotting (immunoblotting) of glycine extracts of other C. jejuni, Campylobactercoli, and Campylobacter laridis strains belonging to different thermolabile and thermostable serotypes, as wellas Campylobacterfetus, with a rabbit polyclonal antiserum raised against the purified C. jejuni VC74 proteinshowed that all C. jejuni, C. coli, and C. laridis strains tested contained a 31,000-Mr protein with epitopes whichwere antigenically cross-reactive with the C. jejuni VC74 protein. The antigenically cross-reactive epitopes ofthis protein were also readily detected by immunodot blot assay of glycine extracts of C. jejuni, C. coli, and C.taridis with monospecific polyclonal antisera to the 31,000-Mr protein, suggesting that this serological test couldbe a useful addition to those currently employed in the rapid identification of these important pathogens. Slideagglutination reactions, immunofluorescence assay, and immunogold electron microscopy with antisera topurified 31,000-Mr protein and trypsin treatment of whole cells indicated that the cross-reactive epitopes of the31,000-Mr protein were not exposed on the cell surface. Cell fractionation analysis and immunogold electronmicroscopy located the protein on the outer surface of the cytoplasmic membrane. This finding suggests thatthe 31,000-Mr protein is not a good candidate for inclusion in a monovalent subunit Campylobacter vaccine.
The thermophilic campylobacters, especially Campylo-bacterjejuni and Campylobacter coli, are now recognized as
important gastrointestinal pathogens of humans, contribut-ing substantially to morbidity in developed countries and tohigh mortality in developing countries (7, 44). Because of thehigh incidence of Campylobacter disease worldwide, there isa need for effective vaccines to protect against infection bythis group of organisms. The frequency of isolation ofthermophilic campylobacters also means that they are thesubject of many laboratory identification investigations andsubsequent biotyping and serotyping studies. Unfortunately,despite numerous studies, campylobacters remain relativelydifficult to rapidly identify, classify, and type biochemically.Members of the genus Campylobacter neither ferment noroxidize carbohydrates and are inert in many other typicalbiochemical test media (15, 22, 36), so there are only a fewcriteria which can be simply applied in their rapid identifi-cation (14, 15, 22, 26, 42). A simple monovalent serologicalassay which could rapidly identify thermophilic campylobac-ters would clearly be useful. However, attempts to developsuch a test have been unsuccessful because of the consider-able serological diversity of the thermophilic campylobac-ters. This antigenic heterogeneity has also hampered vaccinedevelopment. For example, the heat-stable scheme of Pen-ner and Hennessy (37) now contains >76 serotypes (Penner,personal communication), while the heat-labile scheme of
* Corresponding author.
Lior (27) contains >100 serotypes (Lior, personal commu-
nication).Western blot (immunoblot) studies of thermophilic Cam-
pylobacter species have identified a small number of proteinswith antigenically cross-reactive epitopes (3, 4, 29, 32). Theimmunodominant protein antigen of the thermophilic cam-
pylobacters is flagellin with a subunit molecular weight (Mr)of 59,000 to 61,000. However, while this protein carriescross-reactive epitopes, antibody absorption experimentsemploying live motile cells indicate that serospecificepitopes are predominant on the surface of the native flagellafilament (30). Furthermore, there are numerous serotypes ofCampylobacter flagella (13, 16, 27, 45, 46) and these flagellaare also subject to both phase and antigenic variation (6, 13).This means that the Campylobacter flagella protein is neithera straightforward candidate for use in a monovalent vaccinenor as a target for an antibody-based identification test forthermophilic campylobacters. A second protein with cross-
reactive epitopes is the major outer membrane (OM) protein(3, 4, 29, 32), but again, the majority of cross-reactiveepitopes in this molecule likely represent conserved aminoacid sequences buried in the OM (8, 19). The third proteinwith cross-reactive epitopes is a protein with an apparentmolecular weight of 31,000, which has been identified inglycine hydrochloride (pH 2.2) extracts of cells of thermo-philic campylobacters (3, 4, 29, 32). Indeed, the ability toremove the 31,000-Mr protein from cells by this simpleextraction procedure has led to the widely held notion thatthe antigenically cross-reactive 31,000-Mr protein is located
1321
Vol. 28, No. 6
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

1322 DUBREUIL ET AL.
on the surface of the Campylobacter cell and is a primecandidate for inclusion in a monovalent subunit Campylo-bacter vaccine.Because of the potential importance of such a cross-
reactive antigen in the prophylaxis and serological identifi-cation of thermophilic enteropathogenic Campylobacter spe-cies, we have isolated and purified a subunit 31,000-Mracid-extractable cross-reactive protein antigen from a humanfecal isolate of C. jejuni. In this study, we report on thebiochemical characteristics of the protein and provide evi-dence that the protein may be unique to campylobacters.Further, we show that the protein is a good immunogen andthat antibodies to the protein can be used to serologicallyidentify thermophilic Campylobacter species. We also pro-vide evidence that the protein is not located on the cellsurface but is associated with the outer surface of thecytoplasmic membrane and so appears to be a poor candi-date for inclusion in a Campylobacter vaccine.
MATERIALS AND METHODSBacterial strains and growth conditions. All strains were
from the culture collection in this laboratory, unless other-wise noted. C. jejuni strains were VC74 (LIO serogroup 11),VC79 (ex-National Type Culture Collection [NCTC; Col-lindale, London, United Kingdom] strain 11392), VC82(ex-NCTC 113510), VC83 (ex-NCTC 11168, LIO4, Lior C.jejuni biotype 1 reference strain), VC88 (LIO 5), VC90 (LIO9), VC91 (LIO 11), VC94 (LIO 15), VC99 (LIO 18), VC101(LI016), VC102 (LIO 10), VC103 (LIO 17), VC104 (LIO 19),VC105, VC118, VC206, VC208 (LI036, Lior C. jejuni bio-type 2 reference strain), VC209 (ex-NCTC 11392, L106,Lior C. jejuni biotype 23 reference strain), VC210 (LI074,Lior C. jejuni biotype 4 reference strain). C. coli strains wereVC50, VC51 (seawater isolate), VC80 (ex-NCTC 11366),VC84 (LIO 6), VC85, VC93 (LIO 14), VC96 (LIO 13), VC97(LIO 20), VC113, VC114, and VC117. Campylobacter laridisstrains were VC81 (ex-NCTC 11352, LIO34, Lior C. laridisbiotype 2 reference strain), VC186 (LI035, Lior C. laridisbiotype 1 reference strain), and VC214. Campylobactercryaerophila 10685, Campylobacter cinaedi 5519, and"Campylobacter upsaliensis" 5424 were provided by H.Lior, Laboratory Center for Disease Control (Ottawa, On-tario, Canada). Bovine Campylobacter fetus strains wereVC1, VC78 (ex-NCTC 11842), VC119, VC202, and VC203.Aeromonas hydrophila TF7 (9), Escherichia coli HB101,K127, 438, and 1593, Salmonella enteritidis 3b, Salmonellaflorida, Salmonella newport, and Shigellaflexneri were alsoused. Stock cultures were maintained at -70°C in tryptic soybroth (GIBCO Laboratories, Grand Island, N.Y.) supple-mented with 15% (vol/vol) glycerol. Campylobacter cultureswere grown in a humid C02 incubator (10% C02) at 37°C onMueller-Hinton agar (Difco Laboratories, Detroit, Mich.),while Aeromonas, Escherichia, Salmonella, and Shigellastrains were grown for 18 h at 37°C on tryptone soy agar(Difco Laboratories, Detroit, Mich.).
Cell fractionation. Glycine extraction of 48-h-old Cam-pylobacter cells was performed by the method of McCoy etal. (31), modified as previously described (10). The periplas-mic fraction was prepared by the osmotic shock procedureof Willis et al. (47); cell envelopes and released OM vesicleswere prepared by differential centrifugation as previouslydescribed (29). The OM fraction was isolated from cellenvelopes by the sodium lauryl sarcosinate procedure ofFilip et al. (12). Cells cultured for 24 and 48 h were alsotreated with trypsin (100 ,ug/ml for 1 h) and trypsin inhibitoras previously described (29).
Electrophoresis. Sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) was performed by themethod of Laemmli (24) in a mini-slab apparatus (Bio-RadLaboratories, Richmond, Calif.). Protein solubilized in sam-ple buffer was stacked in 4.5% acrylamide (100 V, constantvoltage) and separated with 12.5% acrylamide (200 V).Protein was stained with Coomassie blue R-250.When required, separated proteins or peptides were trans-
ferred from the slab to nitrocellulose paper (NCP) by themethanol-Tris-glycine system described by Towbin et al.(43). Electroblotting was carried out in a Bio-Rad transblotapparatus (Bio-Rad Laboratories) for 18 h at 60 V. Nonde-naturing isoelectric focusing gels were run in a mini-slab gelapparatus by the method of Robertson et al. (39) and werestained with Coomassie blue R-250 according to the Bio-Radisoelectric focusing standards instruction sheet.
Antibodies. Polyclonal antiserum was raised in adult NewZealand White rabbits by intramuscular injections. To pre-pare antiserum DD1, a rabbit was immunized with approxi-mately 30 ,ug of linearized 31,000-Mr protein in acrylamide,obtained by SDS-PAGE separation of a glycine extract,cutting the protein band out of the gel, and then homogeniz-ing the protein-acrylamide preparation in isotonic saline. Toprepare antiserum DD2, a rabbit was immunized with 50 ,ugof fast protein liquid chromatography (FPLC)-purified pro-tein in Freund complete adjuvant. To prepare antiserumLAH31, a third rabbit was immunized with 30 ,ug of high-pressure liquid chromotography (HPLC)- and SDS-PAGE-purified 31,000-Mr protein which had been electroblotted tonitrocellulose and stained by buffalo black. To prepareantiserum SML1, a fourth rabbit was immunized with 109Formalin-killed cells of a 24-h culture of C. jejuni VC74emulsified in Freund complete adjuvant. Booster doses weregiven in Freund incomplete adjuvant on days 14 and 28. Onday 42, the rabbits were exsanguinated and the serum wascollected and stored at -20°C. Control nonimmune serumwas obtained before the first injection.To prepare an immunoaffinity gel to the 31,000-Mr protein,
immunoglobulin G class antibodies in antiserum DD1 wereseparated by chromatography on a DEAE Affi-Gel Blueimmunoglobulin G purification gel (Bio-Rad Laboratories)following the instructions of the manufacturer and coupledon CNBr-activated Sepharose 4B (Pharmacia, Uppsala,Sweden) following the instructions of the manufacturer.Remaining active groups were blocked with 0.1 M Trishydrochloride (pH 8.0).
Protein purification. Two methods were used to purify the31,000-Mr protein to homogeneity. By the first method,glycine extract was subjected to 70 to 80% ammoniumsulfate precipitation and the pellet obtained by centrifugationat 16,000 x g for 30 min at 4°C was suspended in HPLC-grade distilled water and dialyzed overnight against thissolvent. This material was subjected to HPLC. A BeckmanHPLC system with a Brownlee RP8-Spheri 10 analyticalcolumn (4.6 by 250 mm) was used to separate the 31,000-Mrprotein from minor contaminants. A 20 to 50% acetonitrilegradient in 0.1% trifluoroacetic acid was applied over 60 min.The flow rate was 1 ml/min, and chromatography wasperformed at room temperature. Fractions were concen-trated and monitored by SDS-PAGE. The fraction contain-ing a single band of an Mr of 31,000 was electroblotted tonitrocellulose paper and used to prepare polyclonal mono-specific antiserum LAH31, as described above.For preparative scale isolations, lyophilized glycine ex-
tract was rehydrated in 50 mM Tris hydrochloride (pH 8.0)and ultracentrifuged at 100,000 x g for 30 min at 4°C and the
J. CLIN. MICROBIOL.
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

PROTEIN ANTIGEN SHARED BY THERMOPHILIC CAMPYLOBACTERS
supernatant was dialyzed for 16 h against 50 mM Trishydrochloride (pH 8.0). The sample was then applied to theSepharose 4B immunoaffinity column. After sample applica-tion, the column was washed with 10 volumes of 50 mM Trishydrochloride (pH 8.0) and then developed with 0.1 Mglycine hydrochloride (pH 2.5) containing 0.5 M NaCI and10% dioxane (Mallinckrodt, Inc., Paris, Ky.). The peakeluted from the column was concentrated by evaporationand dialyzed for 16 h against 20 mM Tris hydrochloride (pH7.4) containing 0.1 M NaCl. Purification to homogeneity wasachieved by molecular sieving on an FPLC system with aSuperose 12 column (Pharmacia). The system was run in 20mM Tris hydrochloride (pH 7.4) containing 0.1 M NaCl(Tris-NaCl) at a flow rate of 0.5 ml/min. The peaks elutedwere monitored at 230 nm. After chromatography, the peaksof interest were collected, pooled, and concentrated byevaporation. After addition of0.2% (wt/vol) 3-[(3-cholamido-propyl)dimethylammonio]-1-propanesulfonate (CHAPS) (Sig-ma Chemical Co., St. Louis, Mo.), the sample was rechro-matographed in 20 mM Tris-NaCl buffer containing 0.2%(wt/vol) CHAPS.Amino acid composition analysis. The purified protein was
dialyzed extensively against distilled water, lyophilized, andthen hydrolyzed in 6 N HCl at 100°C for 18 h. Amino acidcomposition was determined on a 119CL Beckman aminoacid analyzer. The method used was as described by themanufacturer for a 90-min single column procedure. Cyste-ine was analyzed as cysteic acid after performic acid oxida-tion (17). Tryptophan was determined by the method ofPenke et al. (35), which involved hydrolysis of the proteinwith mercaptoethanesulfonic acid.
N-terminal sequence analysis. Amino acid sequence analy-sis was performed on an Applied Biosystems model 470Agas phase sequenator (Applied Biosystems, Foster City,Calif.) running a standard operating program. Phenylthiohy-dantoin derivatives were separated on an IBM cyanocolumn(4.6 by 250 mm) as described by Hunkapiller and Hood (20)fitted to a Beckman model 332 high-pressure liquid chro-matograph equipped with a Hewlett-Packard 3390A integra-tor (Hewlett-Packard Co., Palo Alto, Calif.). The amino-terminal sequence of the 31,000-Mr protein of C. jejuni VC96was determined after electrophoretic separation of a glycineextract on a 12.5% acrylamide gel and electroblotting to anImmobilon transfer membrane (Millipore Corp., Bedford,Mass.) by the method of LeGendre and Matsudaira (25).Western blotting. After electroblotting, unreacted sites on
the NCP were blocked with a 1% (wt/vol) solution of gelatinin 10 mM Tris hydrochloride-0.9% NaCl (pH 7.4) (GTS) for1 h at room temperature. The NCP was then incubated withan appropriate dilution of antiserum in the same buffer for 2h. The NCP was washed five times with 10 mM Trishydrochloride-O.9% NaCl (pH 7.4). Goat anti-rabbit immu-noglobulin conjugated to alkaline phosphatase (CaltagLabs., South San Francisco, Calif.) was then added in GTSbuffer and incubated for 1 h at room temperature. Afterincubation, the NCP was washed five times in 10 mM Trishydrochloride-0.9% NaCl (pH 7.4). The reactive bands werevisualized as described by Blake et al. (2) with 5-bromo-4-chloro-3-indolyl phosphate (Boehringer GmbH, Mann-heim, Federal Republic of Germany) as the alkaline phos-phatase substrate and Nitro Blue Tetrazolium (Sigma) as thecolor development reagent.Immunodot blotting. Dot blotting was performed by trans-
ferring 10 ,ug of glycine-extractable protein of each strain tobe tested to NCP. After drying at 37°C for 1 h, the NCP wasblocked with GTS, reacted with antiserum LAH31, washed,
and developed by the procedures described for Westernblotting.
Slide agglutination. The ability of antisera to agglutinatelive cells of Campylobacter species was tested by themethod of Lior et al. (27). Presence or absence of agglutina-tion was determined after 1 min.
Electron microscopy. Samples were routinely negativelystained with a solution of 1% (wtl/vol) ammonium molybdateand 0.1% (vol/vol) glycerol on Formvar-coated grids. Forimmunogold electron microscopy, a grid covered with aFormvar film was floated on a 50-,ul drop of bacterial cellssuspended in 100 mM Tris (pH 7.5) containing 150 mMNaCl, 0.05% Tween 20 (TW), and 0.5% bovine serumalbumin (BSA) (Tris-NaCl-TW-BSA) for 5 min. The grid wasthen removed from the drop and floated on a drop ofTris-NaCl-TW-BSA containing 10% (vol/vol) antiserumLAH31. After a 1-h incubation, the grid was removed andnonspecifically bound immunoglobulin G was removed byfloating the grid on two drops of Tris-NaCl-TW-BSA. Thegrid was then floated on a drop of Tris-NaCl-TW-BSAcontaining a 1:10 dilution of 15-nm colloidal gold particlescoated with protein A (Jannsen Biotech, Olen, Belgium).After incubation for 1 h, the nonspecifically bound colloidalgold particles were removed by floating the grid on twodrops ofTris-NaCl-TW-BSA and one drop of distilled water.The grids were stained by floating on a drop of 1% (wtl/vol)ammonium molybdate containing 0.1% (vol/vol) glycerol(pH 7.5) and were examined in a Philips EM-300 electronmicroscope (Philips Electronic Instruments, Inc., Mahwah,N.J.) operated at an accelerating voltage of 60 kV. Imageswere recorded on 70-mm Fine Grain Release Film (EastmanKodak, Rochester, N.Y.).
Indirect fluorescent-antibody testing. Indirect fluorescent-antibody testing was performed as previously described (30).
RESULTS AND DISCUSSION
The most efficient recovery of the 31,000Mr protein wasobtained by the glycine buffer (pH 2.2) extraction procedureof McCoy et al. (31) with cells cultured for 48 h. Thistechnique was used by McCoy and co-workers to preferen-tially isolate the surface layer protein from C. fetus. Themethod has also been successfully employed to isolatesurface layer proteins from Aeromonas species (9) andflagella from Campylobacter species (29, 30). SDS-PAGEanalysis showed that the 31,000-Mr protein and the 61,000-Mr flagellin protein (3, 4, 29, 30, 33) were the principalcomponents in the glycine extract. The 45,000-Mr majorouter membrane protein was also present (3, 4, 29, 32),indicating contamination by OM material, presumably asvesicles.To purify sufficient protein to allow antibodies to be
prepared for subsequent immunoaffinity purification, the31,000-Mr protein was cut directly from an SDS-PAGE gel ofthe glycine-extracted fraction and used to immunize a rabbit(antiserum DD1). In an alternative procedure, a 70 to 80%ammonium sulfate fraction was subjected to HPLC by usinga reverse-phase C8 Spheri-10 column and a 20 to 50%gradient of acetonitrile in a 0.1% trifluoroacetic acid-aceto-nitrile system. The 31,000-Mr protein was eluted at approx-imately 39% acetonitrile. This fraction contained a singleband, which was electroblotted to NCP, and this was used toprepare polyclonal monospecific antiserum LAH31.For preparative scale isolation of the 31,000-Mr protein,
immunoaffinity chromatography was employed. Contami-nating flagella and OM material were removed from a
VOL. 28, 1990 1323
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
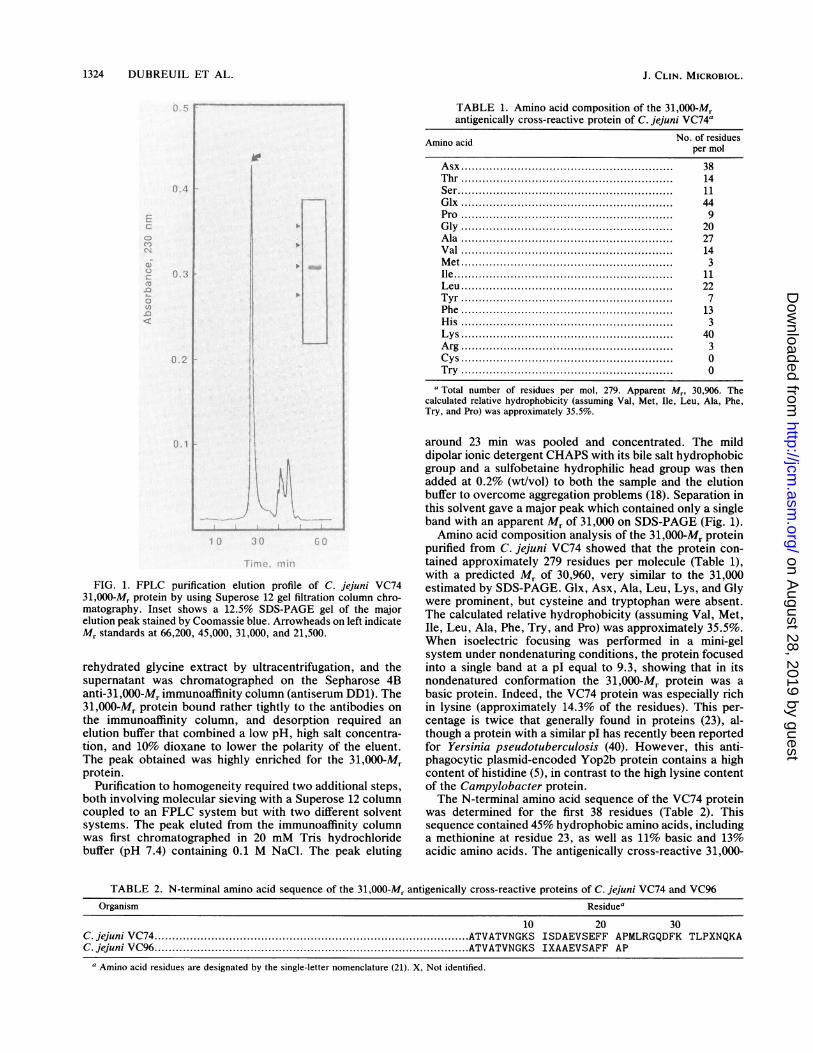
1324 DUBREUIL ET AL.
FIG. 1. FPLC purification elution profile of C. jejuni VC7431,000-Mr protein by using Superose 12 gel filtration column chro-matography. Inset shows a 12.5% SDS-PAGE gel of the majorelution peak stained by Coomassie blue. Arrowheads on left indicateMr standards at 66,200, 45,000, 31,000, and 21,500.
rehydrated glycine extract by ultracentrifugation, and thesupernatant was chromatographed on the Sepharose 4Banti-31,000-Mr immunoaffinity column (antiserum DD1). The31,000-Mr protein bound rather tightly to the antibodies onthe immunoaffinity column, and desorption required anelution buffer that combined a low pH, high salt concentra-tion, and 10% dioxane to lower the polarity of the eluent.The peak obtained was highly enriched for the 31,000-Mrprotein.
Purification to homogeneity required two additional steps,both involving molecular sieving with a Superose 12 columncoupled to an FPLC system but with two different solventsystems. The peak eluted from the immunoaffinity columnwas first chromatographed in 20 mM Tris hydrochloridebuffer (pH 7.4) containing 0.1 M NaCl. The peak eluting
TABLE 1. Amino acid composition of the 31,000-Mrantigenically cross-reactive protein of C. jejuni VC74'
Amino acid No. of residuesper mol
Asx ..................................... 38Thr ..................................... 14Ser..................................... ilGlx ..................................... 44Pro ..................................... 9Gly ..................................... 20Ala ..................................... 27Val ..................................... 14Met ..................................... 3Ile..................................... ilLeu ..................................... 22Tyr ..................................... 7Phe ..................................... 13His ..................................... 3Lys ..................................... 40Arg ..................................... 3
Cys..........................................OTry .....................................O0
a Total number of residues per mol, 279. Apparent Mr, 30,906. Thecalculated relative hydrophobicity (assuming Val, Met, Ile, Leu, Ala, Phe,Try, and Pro) was approximately 35.5%.
around 23 min was pooled and concentrated. The milddipolar ionic detergent CHAPS with its bile salt hydrophobicgroup and a sulfobetaine hydrophilic head group was thenadded at 0.2% (wt/vol) to both the sample and the elutionbuffer to overcome aggregation problems (18). Separation inthis solvent gave a major peak which contained only a singleband with an apparent Mr of 31,000 on SDS-PAGE (Fig. 1).Amino acid composition analysis of the 31,000-Mr protein
purified from C. jejuni VC74 showed that the protein con-tained approximately 279 residues per molecule (Table 1),with a predicted Mr of 30,960, very similar to the 31,000estimated by SDS-PAGE. Glx, Asx, Ala, Leu, Lys, and Glywere prominent, but cysteine and tryptophan were absent.The calculated relative hydrophobicity (assuming Val, Met,Ile, Leu, Ala, Phe, Try, and Pro) was approximately 35.5%.When isoelectric focusing was performed in a mini-gelsystem under nondenaturing conditions, the protein focusedinto a single band at a pI equal to 9.3, showing that in itsnondenatured conformation the 31,000-Mr protein was abasic protein. Indeed, the VC74 protein was especially richin lysine approximatelyy 14.3% of the residues). This per-centage is twice that generally found in proteins (23), al-though a protein with a similar pI has recently been reportedfor Yersinia pseudotuberculosis (40). However, this anti-phagocytic plasmid-encoded Yop2b protein contains a highcontent of histidine (5), in contrast to the high lysine contentof the Campylobacter protein.The N-terminal amino acid sequence of the VC74 protein
was determined for the first 38 residues (Table 2). Thissequence contained 45% hydrophobic amino acids, includinga methionine at residue 23, as well as 11% basic and 13%acidic amino acids. The antigenically cross-reactive 31,000-
TABLE 2. N-terminal amino acid sequence of the 31,000-Mr antigenically cross-reactive proteins of C. jejuni VC74 and VC96Organism Residue
10 20 30C.jejuniVC74.......... ATVATVNGKS ISDAEVSEFF APMLRGQDFK TLPXNQKAC.jejuniVC96.......... ATVATVNGKS IXAAEVSAFF AP
a Amino acid residues are designated by the single-letter nomenclature (21). X, Not identified.
J. CLIN. MICROBIOL.
ll
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

PROTEIN ANTIGEN SHARED BY THERMOPHILIC CAMPYLOBACTERS
--- --M-4 N---
1 2 3 4 5 6 7 8 9 10 1 1 1 2
v -» -M - -
13 14 15 16 17 18 19 20 21 22 23
FIG. 2. Western blot of glycine extracts of C. jejuni strains VC74(lane 1), VC79 (lane 2), VC82 (lane 3), VC83 (lane 4), VC88 (lane 5),VC90 (lane 6), VC91 (lane 7), VC94 (lane 8), VC105 (lane 9), VC118(lane 10), and VC206 (lane 11); C. coli strains VC50 (lane 12), VC51(lane 13), VC80 (lane 14), VC84 (lane 15), VC93 (lane 16), VC96(lane 17), VC97 (lane 18), and VC113 (lane 19); C. laridis VC81 (lane20); C. fetus VC78 (lane 21); C. fetus VC119 (lane 22); and A.hydrophila TF7 (lane 23) reacted with a 1:5,000 dilution of rabbitpolyclonal anti-31,000-Mr protein antiserum DD2.
Mr protein of C. jejuni VC96 was also selectively isolatedfrom glycine extracts by the Immobilon transfer technique(25) and, when subjected to N-terminal amino acid sequenceanalysis, exhibited a sequence virtually identical to the VC74protein to residue 22. The single difference of an Ala for anAsp at residue 13 could result from a single nucleotidechange at the codon level. A search of the National Biomed-ical Research Foundation protein sequence library failed toreveal any sequences with significant homology to the N-terminal of the 31,000-Mr protein (28), which suggests that itmay be a protein unique to Campylobacter species.The 31,000-Mr protein was a good immunogen in rabbits,
both when rabbits were immunized with the purified C.jejuni VC74 31,000-Mr protein or with intact formalinized C.jejuni VC74 cells. Western blotting of glycine extracts of arange of Campylobacter species with antisera DD2 orLAH31 to the VC74 31,000-Mr protein showed that anantigenically cross-reactive protein with an Mr of 31,000 waspresent in all C. jejuni and C. coli strains tested, regardlessof heat-labile serotype. The protein was also present in C.laridis, which can also cause enteritis in humans (36), butwas not present in glycine extracts of C. fetus. Identicalresults were also obtained with antiserum SML1 preparedagainst intact cells of VC74. Figure 2 shows the resultsobtained when antiserum DD2 was reacted with C. jejuniVC74, VC79, VC82, VC83, VC88, VC90, VC91, VC94,VC105, VC118, and VC206 (lanes 1 to 11); C. coii VC5O,VC51, VC80, VC84, VC93, VC96, VC97, and VC113 (lanes12 to 19); C. laridis VC81 (lane 20); and C. fetus VC78 andVC119 (lanes 21 and 22). The peptide was not present inglycine extracts of control strains A. hydrophila TF7 (lane23) and E. coli HB101 (data not shown). A dot blot assay ofglycine extracts with the monospecific polyclonal antiserafurther revealed that the cross-reactive epitopes could bereadily used to detect C. jejuni, C. coli, and C. Iaridis.Weaker, but still positive, signals were also obtained withglycine extracts of "C. upsaliensis," and C. cinaedi. "C.upsaliensis" is a newly described catalase-negative orweakly positive species of the genus Campylobacter and hasbeen isolated with gastroenteritis in humans and dogs (34,41), while C. cinaedi has been associated with proctitis andenteritis in homosexual males (11). This latter Campylobac-ter species does not grow at 42°C, however, and shouldneither be coisolated nor have its identity confused with themuch more common thermophilic enteropathogenic speciesof the genus Campylobacter. No reaction was seen withglycine extracts of C. fetus, the catalase-positive C. cryaero-
'i,., , a#
:e* * e e e* ea
FIG. 3. Immunodot blot of glycine extracts (10 ,ug of protein)reacted with a 1:500 dilution of rabbit polyclonal anti-31,000-M,protein antiserum LAH31. (al) C. jejuni VC74; (a2) C. jejuni VC79;(a3) C. jejuni VC82; (a4) C. jejuni VC83; (aS) C. jejuni VC88; (a6) C.jejuni VC90; (a7) C. jejuni VC99; (a8) C. jejuni VC101; (a9) C. jejuniVC102; (alO) C. jejuni VC103; (bl) C. jejuni VC104; (b2) C. jejuniVC208; (b3) C. jejuni VC209; (b4) C. jejuni VC210; (b5) C. coliVC85; (b6) C. coli VC97; (b7) C. coli VC114; (b8) C. coli VC117;(b9) C. laridis VC81; (blO) C. laridis VC186; (ci) C. laridis VC214;(c2) C. cryaerophila 10685; (c3) C. cinaedi 5519; (c4) "C. upsalien-sis" 5424; (c5) C. fetus VCi; (c6) C. fetus VC78; (c7) C. fetusVC119; (c8) C. fetus VC202; (c9) C. fetus VC203; (c1O) E. coliHB101; (dl) E. coli K127; (d2) E. coli 438; (d3) E. coli 1593; (d4) S.enteritidis 3b; (d5) S. florida; (d6) S. newport; (d7) S. flexneri; (d8)FPLC-purified VC74-31K protein (5 ,ug).
phila, or the nonspiral enteric pathogens tested, including E.coli, Salmonella species, and S. flexneri (Fig. 3).
Initial reports of the 31,000-Mr antigenically cross-reactiveprotein of Campylobacter species had suggested that theantigen was positioned on the cell surface by virtue of itssensitivity to trypsin and by its ability to be iodinated by thelactoperoxidase surface radiolabeling technique (3, 29, 32).These studies were performed on cells grown for 48 h. Whenwe treated 24-h-old cells with trypsin, there was no loss ofepitope reactivity although the protein was susceptible totrypsin cleavage in 48-h cultures and when purified (Fig. 4,lanes 2 to 5). Moreover, monospecific antisera DD2 andLAH31 both failed to agglutinate cells of homologous strainC. jejuni VC74 and indirect fluorescent-antibody testing andimmunogold electronmicroscopy with these antisera failedto reveal antibody binding to the cell surface or to releasedOM (Fig. SA). Cell fractionation studies further showed thatthe protein was not present in the released OM vesiclefraction (Fig. 4, lane 6), and although the protein was presentin the cell envelope fraction (Fig. 4, lane 7), it did notcopurify with OM prepared by sarcosyl solubilization (Fig.4, lane 8). Unfortunately, because of extensive fusionsbetween the two membranes, physical separation proce-dures, such as sucrose gradients, do not allow separation ofthe outer and inner membranes of Campylobacter species(33); however, the protein was sarcosyl soluble (Fig. 4, lane
FIG. 4. Western blot of cellular fractions of C. jejuni VC74reacted with a 1:500 dilution of rabbit polyclonal anti-31,000-Mrprotein antiserum LAH31. Lane 1, glycine extract; lane 2, whole-cell lysate; lane 3, whole-cell lysate of trypsin-treated 24-h-old cells;lane 4, whole-cell lysate of trypsin-treated 48-h-old cells; lane 5,trypsin-treated purified 31,000-Mr protein; lane 6, released OMvesicles; lane 7, cell envelope; lane 8, sarcosyl-insoluble OMfraction; lane 9, sarcosyl-soluble envelope fraction.
VOL. 28, 1990 1325
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
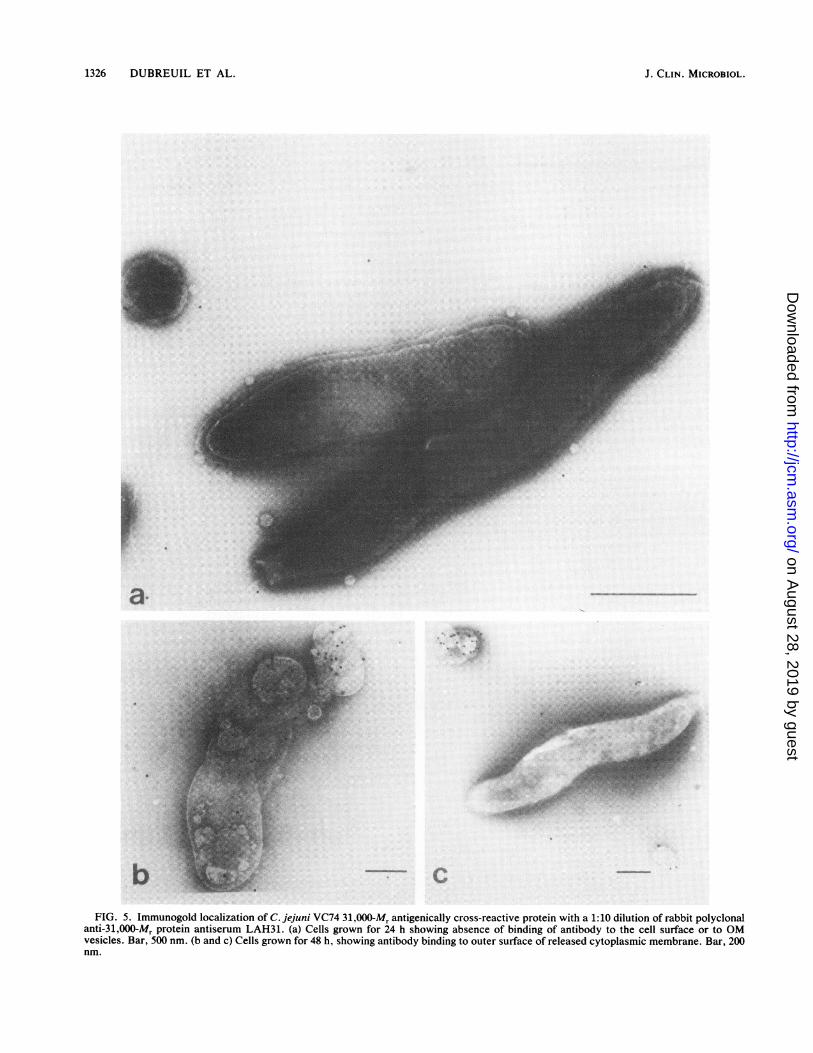
1326 DUBREUIL ET AL.
b c-l%FIG. 5. Immunogold localization of C. jejuni VC74 31,000-Mr antigenically cross-reactive protein with a 1:10 dilution of rabbit polyclonal
anti-31,000-Mr protein antiserum LAH31. (a) Cells grown for 24 h showing absence of binding of antibody to the cell surface or to OMvesicles. Bar, 500 nm. (b and c) Cells grown for 48 h, showing antibody binding to outer surface of released cytoplasmic membrane. Bar, 200nm.
J. CLIN. MICROBIOL.
-. i -;-:4,'..,
b'i...r-W
èr
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

PROTEIN ANTIGEN SHARED BY THERMOPHILIC CAMPYLOBACTERS
9) and only small amounts were released from the cell byosmotic shock (data not shown). Taken together, thesefindings suggested that the 31,000-Mr antigenically cross-reactive protein of the enteropathogenic campylobacterswas associated with the inner membrane. Immunogold elec-tron microscopy confirmed that this was the case. Noantibody labeling was observed on the surface of intact cells(Fig. 5A), but, as the Campylobacter culture aged, anincreasing number of cells lysed and either released orexposed their cytoplasmic membrane and antibodies to the31,000-Mr cross-reactive protein were seen to bind to thesurface of this membrane (Fig. 5B and C). However, whileobviously associated with the inner membrane, it seemsunlikely that the 31,000-Mr protein is an integral membraneprotein because it displays none of the major solubilityproblems associated with integral membrane proteins. Onthe basis of amino acid composition, the structure-functionRatio 3 value of 1.8 calculated by the algorithm of Barrantes(1) also suggests a peripherally located protein rather than anintegral membrane protein, as does the pI value.
Normally, it is expected that for a cellular immunogen toconfer protective immunity, the antigen needs to be exposedon the surface of live infecting bacteria, where it will beaccessible to the host immune defense mechanisms. Thisfinding that the 31,000-Mr antigenically cross-reactive pro-tein is not exposed on the cell surface indicates that thisprotein is unlikely to be a strong candidate for inclusion in avaccine against thermophilic Campylobacter infections.Even though as many as 61% of individuals seroconvert toan acid-extractable protein of an apparent Mr of 30,000 overthe course of a Campylobacter infection (Z. Pei, R. T.Ellison III, C. D. Deal, and M. J. Blaser, Program Abstr.29th Intersci. Conf. Antimicrob. Agents Chemother., abstr.no. 46, 1989), this seroconversion must result from antigenexposure during cell lysis or during macrophage processing.The fact that the epitopes are not exposed on the cell surfacedoes not rule out their potential diagnostic value, however,and indeed may be fortuitous when it comes to developmentof a specific serological test for the thermophilic entero-pathogenic campylobacters, given the serologic diversity ofthe surface of the Campylobacter cell. Crude glycine ex-tracts are already used to detect anti-Campylobacter anti-bodies (38). However, these crude extracts contain antigen-ically variable components such as flagellin andlipopolysaccharide, so use of the purified antigenically cross-reactive 31,000-Mr protein should improve the specificity ofdetection of anti-Campylobacter antibodies in sera frompatients. Antisera to the 31,000-Mr protein can also be usedfor rapid laboratory identification of such isolates as thosefrom Campylobacter species. Glycine extraction to releasethe antigen is easy to perform and can be done on a smallscale, such as in microcentrifuge tubes, and the immunodotblot assay is within the capabilities of most laboratories andis simple to perform on a routine basis. The nucleotidesequence encoding the thermophilic campylobacter-specificepitope could also provide for a DNA probe. Efforts aretherefore underway to clone the gene coding for the 31,000-Mr cross-reactive protein antigen of the thermophilic entero-pathogenic campylobacters.
ACKNOWLEDGMENTS
This work was supported in part by grants to T.J.T. from theMedical Research Council of Canada. J.D.D. was the recipient of apostdoctoral award from the Fonds de la Recherche en Santé duQuébec.
LITERATURE CITED1. Barrantes, F. J. 1975. The nicotinic cholinergic receptor: dif-
ferent compositions evidenced by statistical analysis. Biochem.Biophys. Res. Commun. 62:407-414.
2. Blake, M. S., K. H. Johnston, G. J. Russell-Jones, and E. C.Gotschlich. 1984. A rapid, sensitive method for detection ofalkaline phosphatase-conjugated anti-antibody on Westernblots. Anal. Biochem. 136:175-179.
3. Blaser, M. J., J. A. Hopkins, R. M. Berka, M. L. Vasil, andW.-L. L. Wang. 1983. Identification and characterization ofCampylobacterjejuni outer membrane proteins. Infect. Immun.42:276-284.
4. Blaser, M. J., J. A. Hopkins, and M. L. Vasil. 1984. Campylo-bacterjejuni outer membrane proteins are antigenic for humans.Infect. Immun. 43:986-993.
5. Bolin, I., and H. Wolf-Watz. 1988. The plasmid-coded Yop2protein of Yersinia pseudotuberculosis is a virulence determi-nant regulated by calcium and temperature at the level oftranscription. Mol. Microbiol. 2:237-245.
6. Caldwell, M. B., P. Guerry, E. C. Lee, J. P. Burans, and R. I.Walker. 1985. Reversible expression of flagella in Campylobac-terjejuni. Infect. Immun. 50:941-943.
7. Chowdhury, M. N. H. 1984. Campylobacter jejuni enteritis: areview. Trop. Geogr. Med. 36:215-222.
8. DiRenzio, J. M., K. Nakamura, and M. Inouye. 1978. The outermembrane proteins of gram-negative bacteria: biosynthesis,assembly, and functions. Annu. Rev. Biochem. 47:481-532.
9. Dooley, J. S. G., W. D. McCubbin, C. M. Kay, and T. J. Trust.1988. Isolation and biochemical characterization of the S-layerprotein from a pathogenic Aeromonas hydrophila strain. J.Bacteriol. 170:2631-2638.
10. Dubreuil, J. D., S. M. Logan, S. C. Cubbage, D. Ni Eidhin,W. D. McCubbin, C. M. Kay, T. J. Beveridge, F. G. Ferris, andT. J. Trust. 1988. Structural and biochemical analysis of asurface array protein of Campylobacter fetus. J. Bacteriol.170:4165-4173.
11. Fennell, C. L., A. P. Totten, T. C. Quinn, D. L. Patton, K. K.Holmes, and W. E. Stamm. 1984. Characterisation of Campylo-bacter-like organisms isolated from homosexual men. J. Infect.Dis. 149:58-66.
12. Filip, C., G. Fletcher, J. L. Wulff, and C. F. Earhart. 1973.Solubilization of the cytoplasmic membrane of Escherichia coliby the ionic detergent sodium-lauryl sarcosinate. J. Bacteriol.115:717-722.
13. Harris, L. A., S. M. Logan, P. Guerry, and T. J. Trust. 1987.Antigenic variation of Campylobacter flagella. J. Bacteriol.169:5066-5071.
14. Harvey, S. M. 1980. Hippurate hydrolysis by Campylobacterfetus. J. Clin. Microbiol. 11:435-437.
15. Hébert, G. A., D. G. Hollis, R. E. Weaver, M. A. Lambert, M. J.Blaser, and C. W. Moss. 1982. 30 years of campylobacters:biochemical characteristics and a biotyping proposal for Cam-pylobacterjejuni. J. Clin. Microbiol. 15:1065-1073.
16. Hébert, G. A., D. G. Hollis, R. E. Weaver, A. G. Steigerwalt,R. M. McKinney, and D. J. Brenner. 1983. Serogroups ofCampylobacterjejuni, Campylobacter coli, and Campylobacterfetus defined by direct immunofluorescence. J. Clin. Microbiol.17:529-538.
17. Hirs, C. H. W. 1965. The oxidation of ribonuclease withperformic acid. J. Biol. Chem. 219:611-621.
18. Hjelmeland, L. M. 1980. A nondenaturing zwitterionic detergentfor membrane biochemistry: design and synthesis. Proc. Natl.Acad. Sci. USA 77:6368-6370.
19. Hofstra, H., and J. Dankert. 1979. Antigenic cross-reactivity ofmajor outer membrane proteins in Enterobacteriaceae species.J. Gen. Microbiol. 111:293-302.
20. Hunkapiller, M. W., and L. E. Hood. 1983. Analysis of phe-nylthiohydantoins by ultrasensitive gradient high-performanceliquid chromatography. Methods Enzymol. 91:486-493.
21. IUPAC-IUB Commission on Biochemical Nomenclature. 1968. Aone-letter notation for amino acid sequences: tentative rules. J.Biol. Chem. 243:3557-3559.
22. Karmali, M. A., and M. B. Skirrow. 1984. Taxonomy of the
VOL. 28, 1990 1327
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

1328 DUBREUIL ET AL.
genus Campylobacter, p. 1-20. In J. P. Butzler (ed.), Campylo-bacter infection in man and animals. CRC Press, Boca Raton,Fla.
23. Klapper, M. H. 1977. The independent distribution of aminoacid near neighbor pairs into polypeptides. Biochem. Biophys.Res. Commun. 78:1018-1024.
24. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
25. LeGendre, N., and P. Matsudaira. 1988. Direct protein micro-sequencing from Immobilon-P transfer membrane. Biotech-niques 6:154-159.
26. Lior, H. 1984. New, extended biotyping scheme for Campylo-bacter jejuni, Campylobacter coli, and "Campylobacter lari-dis." J. Clin. Microbiol. 20:636-640.
27. Lior, H., D. L. Woodward, J. A. Edgar, and L. J. Laroche. 1981.Serotyping by slide agglutination of C. jejuni and epidemiology.Lancet ii:1103-1104.
28. Lipman, D. J., and W. R. Pearson. 1985. Rapid and sensitiveprotein similarity searches. Science 227:1435-1441.
29. Logan, S. M., and T. J. Trust. 1983. Molecular identification ofsurface protein antigens of Campylobacter jejuni. Infect. Im-mun. 42:675-682.
30. Logan, S. M., and T. J. Trust. 1986. Location of epitopes onCampylobacterjejuni flagella. J. Bacteriol. 168:739-745.
31. McCoy, E. C., D. Doyle, K. Burda, L. B. Corbeil, and A. J.Winter. 1975. Superficial antigens of Campylobacter (Vibrio)fetus: characterization of an antiphagocytic component. Infect.Immun. 11:517-525.
32. Newell, D. G., H. McBride, and A. D. Pearson. 1984. Theidentification of outer membrane proteins and flagella of Cam-pylobacterjejuni. J. Gen. Microbiol. 130:1201-1208.
33. Page, W. J., and D. E. Taylor. 1988. Comparison of methodsused to separate the inner and outer membranes of cell enve-lopes of Campylobacter species. J. Gen. Microbiol. 134:2925-2932.
34. Patton, C. M., N. Shaffer, P. Edmonds, T. J. Barrett, M. A.Lambert, C. Baker, D. M. Periman, and D. J. Brenner. 1989.Human disease associated with "Campylobacter upsaliensis"(catalase-negative or weakly positive Campylobacter species) inthe United States. J. Clin. Microbiol. 27:66-73.
35. Penke, B., R. Ternezi, and K. Kovacs. 1974. A new acid
hydrolysis method for determining tryptophan in peptides andproteins. Anal. Biochem. 60:45-50.
36. Penner, J. L. 1988. The genus Campylobacter: a decade ofprogress. Clin. Microbiol. Rev. 1:157-172.
37. Penner, J. L., and J. N. Hennessy. 1980. Passive hemagglutina-tion technique for serotyping Campylobacterfetus subsp. jejunion the basis of soluble heat-stable antigens. J. Clin. Microbiol.12:732-737.
38. Rautelin, H., and T. U. Kosunen. 1983. An acid extract as acommon antigen in Campylobacter coli and Campylobacterjejuni strains. J. Clin. Microbiol. 17:700-701.
39. Robertson, E. F., H. K. Dannelly, P. J. Malloy, and H. C.Reeves. 1987. Rapid isoelectrofocusing in a vertical polyacryl-amide minigel system. Anal. Biochem. 167:290-294.
40. Rosqvist, R., I. Bolin, and H. Wolf-Watz. 1988. Inhibition ofphagocytosis in Yersinia pseudotuberculosis: a virulence plas-mid-encoded ability involving the Yop2 protein. Infect. Immun.56:2139-2145.
41. Sandstedt, K., J. Ursing, and M. Walder. 1983. ThermotolerantCampylobacter with no or weak catalase activity isolated fromdogs. Curr. Microbiol. 8:209-213.
42. Skirrow, M. B., and J. Benjamin. 1980. Differentiation ofenteropathogenic Campylobacter. J. Clin. Pathol. 33:1122.
43. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 74:4350-4354.
44. Walker, R. I., M. B. Caldwell, E. C. Lee, P. Guerry, T. J. Trust,and G. M. Ruiz-Palacios. 1986. Pathophysiology of Campylo-bacter enteritis. Microbiol. Rev. 50:81-94.
45. Wenman, W. M., J. Chai, T. J. Louie, C. Goudreau, H. Lior,D. G. Newell, A. Pearson, and D. E. Taylor. 1985. Antigenicanalysis of Campylobacter flagellar protein and other proteins.J. Clin. Microbiol. 21:108-112.
46. Wenman, W. M., D. E. Taylor, and H. Lior. 1985. The flagellaprotein determines Campylobacter heat-labile serotype, p. 212.In A. D. Pearson, M. B. Skirrow, H. Lior, and B. Rowe (ed.),Campylobacter III. Public Health Laboratory Service, London.
47. Willis, R. C., R. G. Morris, C. Cirakoglu, G. D. Schellenberg,N. H. Gerber, and C. E. Furlong. 1974. Preparation of theperiplasmic binding proteins from Salmonella typhimurium andEscherichia coli. Arch. Biochem. Biophys. 161:64-75.
J. CLIN. MICROBIOL.
on August 28, 2019 by guest
http://jcm.asm
.org/D
ownloaded from


















