Phytoplankton composition under resource limitations MSc research project 1
41
Phytoplankton Composition Under Resource Limitations 42 EC (20 th Feb 2013 – 30 th September 2014) Emma Greenwell : 10407995 MSc Biological sciences: Limnology and oceanography Supervisor: Amanda Burson Examiner: Maayke Stomp IBED, UvA March 28 th 2014
-
Upload
emma-greenwell -
Category
Documents
-
view
52 -
download
1
Transcript of Phytoplankton composition under resource limitations MSc research project 1

Phytoplankton Composition
Under Resource Limitations
42 EC
(20th Feb 2013 – 30th September 2014)
Emma Greenwell : 10407995 MSc Biological sciences:
Limnology and oceanography Supervisor: Amanda Burson Examiner: Maayke Stomp
IBED, UvA March 28th 2014

2
Table of Contents
I. Abstract .............................................................................................................. 3
II. Introduction ....................................................................................................... 4 2.1 Nutrient limitations ................................................................................................................................... 4 2.1.1 previous research .................................................................................................................................... 5 2.1.2 Management .............................................................................................................................................. 5
2.2 Competition ................................................................................................................................................... 7 2.2.1 nutrient limitations ................................................................................................................................ 7 2.2.2 light limitation .......................................................................................................................................... 8
2.3 Aims .................................................................................................................................................................. 9
III. Materials and methods ................................................................................... 10 3.1 Analysis ........................................................................................................................................................ 12 3.2 Dissolved inorganic nutrients ............................................................................................................ 14 3.2.1 DIN .............................................................................................................................................................. 14 3.2.2 DIP ............................................................................................................................................................... 14 3.2.3 DISi .............................................................................................................................................................. 14
IV. Results ............................................................................................................ 15 4. 1 Low Nitrogen: Low Phosphate Chemostat .................................................................................. 17 4.1.1 Phytoplankton population ................................................................................................................ 17 4.1.2 Abiotic parameters .............................................................................................................................. 17
4. 2 Mid-‐Nitrate: Mid-‐Phosphate Chemostat ....................................................................................... 17 4.2.1 Phytoplankton population ................................................................................................................ 17 4.2.2 Abiotic parameters .............................................................................................................................. 17
4.3 High Nitrate: High Phosphate Chemostat ..................................................................................... 18 4.3.1 Phytoplankton population ................................................................................................................ 18 4.3.2 Abiotic parameters .............................................................................................................................. 18
4.4 Low Nitrate: High Phosphate Chemostat ...................................................................................... 20 4.4.1 Phytoplankton population ................................................................................................................ 20 4.4.2 Abiotic parameters .............................................................................................................................. 21
4.5 High Nitrate: Low Phosphate Chemostat ...................................................................................... 21 4.5.1 Phytoplankton population ................................................................................................................ 21 4.5.2 Abiotic parameters .............................................................................................................................. 21
4.6 Mid-‐Nitrate: High Phosphate Chemostat ....................................................................................... 23 4.6.1 Phytoplankton population ................................................................................................................ 23 4.6.2 Abiotic factors ........................................................................................................................................ 24
4.7 High Nitrate: Mid-‐Phosphate chemostat ....................................................................................... 24 4.7.1 Phytoplankton population ................................................................................................................ 24 4.7.2 Abiotic parameters .............................................................................................................................. 24
4.9 All chemostats: Summary ..................................................................................................................... 25 4.9.1 Beginning ................................................................................................................................................. 25 4.9.2 final states ................................................................................................................................................ 26
V. Discussion ........................................................................................................ 28 5.1 Species and their limiting nutrients ................................................................................................ 28 5.1.1 ‘pushing’ nutrient environments .................................................................................................... 29
5.2 Species dominance (beginning and end state) ........................................................................... 30 5.3 Summary of end state species ............................................................................................................ 33 5.3 Limitations .................................................................................................................................................. 34 5.4 conclusion ................................................................................................................................................... 35
VI. Reference list .................................................................................................. 35

3
I. Abstract Phytoplankton are the basis of the pelagic food web, therefore any alterations in their composition can affect higher trophic levels and the economic value of marine products. Marine waters have traditionally been known to be nitrogen (N) limited while freshwater was phosphorous (P) limited. De-‐eutrophication measures have begun to alter this standard of thought as the reduction of phosphorus entering coastal seas has been much more successful when compared to nitrogen. Thus, the CHARLET project is looking at the recent change from nitrogen to phosphorous limitation in coastal seas. Does this change in nutrient limitation alter phytoplankton composition (through competition) in a positive or negative way, or not at all? Competition for resources can occur when the ratio of availability of N to P is skewed, or due to light limitation when nutrient loads are replete. To test this nutrient-‐ratio hypothesis, North Sea inoculum was gathered and placed into chemostats with 7 different nutrient ratios and varying loads (LowN:LowP, LowN:HighP, HighN:HighP, HighN:MidP, HighN:LowP, MidN:MidP and MidN:HighP). Lugols and flow cytometry samples were taken consistently for 3 months for further species identification and species counts. Results showed that diatoms (Nitschia agnita and Nitschia pusilla) were the best competitors in low to mid nutrient loads of 16:1 N:P , the diatom N. pusilla was best suited to phosphorous limited conditions (LowN:LowP and HighN:LowP), Chlorella sp. was the best competitor for low light situations while potentially nitrogen fixing cyanobacteria “2” was able to utilize the extremely low nitrogen levels. In conclusion changes in nutrient ratio and load do allow predictions of phytoplankton composition however only at group level (green algae, cyanobacteria, diatom etc.).

4
II. Introduction Autotrophic phytoplankton are the base of the entire pelagic food web. Therefore changes in phytoplankton dominance can disrupt the functions interacting within the aquatic ecosystem (Henriksen et al. 2002). Shifts in phytoplankton community composition can produce changes in higher trophic species (Tilman et al. 1986, Phillipart et al. 2007 and Navarro and Lobbam 2009). Further shown by Beaugrand et al. in 2003 where long term changes in Atlantic cod (Gadus morhua L.) were shown to be not only a result of over fishing, but also by changes and fluctuations of phytoplankton species and abundance. The economic value of plankton/marine products can also suffer, such as the seaweed (Nori) aquaculture in Japan which has been negatively impacted by undesired phytoplankton blooms (Imai et al. 2006). Understanding the key factors responsible for changes in phytoplankton communities is of interest to many different stake holders.
2.1 Nutrient limitations As a driving factor in phytoplankton productivity nutrients, such as nitrogen and phosphorus, are extensively investigated, especially with respect to which is limiting growth. Nutrient limitation continuously shapes phytoplankton communities as shown by Philippart et al. (2000) in a strong longitudinal high-‐resolution time series study of 20 years (1974-‐1994). A causal relationship was found between phytoplankton and nutrient concentrations on the Wadden Sea. It showed drastic changes in phytoplankton communities that coincided with changes in nutrient concentrations. Weekly sampling (1962-‐1984) in the German Bight showed increases in N concentration reducing the Si:N ratio to <0.1 resulting in a shift from a high diatom abundance to high flagellates (Radach et al., 1990). Nutrient limitation has been the focus of many marine and freshwater research over the years, and rightly so. The importance of understanding phytoplankton composition in relation to nutrient uptake and availability is paramount as this may allow for prediction

5
of toxic algal blooms, among other management related issues (Brauer et al. 2012).
2.1.1 previous research Research has repeatedly shown marine environments to be nitrogen limited (Hecky 1988; Elser et al 2007 and Phillipart et al. 2007) due to strong de-‐nitrification, low amounts of N-‐fixing cyanobacteria and high rates of SO4 reduction (through microbes) that consequently makes it harder for Fe ions to isolate phosphorous which is bound to the sea floor (Ekholm 2008). This makes phosphorous unattainable in the water column rendering it the limiting nutrient. Phosphorous, on the other hand, is the most common limiting nutrient in freshwater environments (Hecky 1988 and Elser et al 2007). However, recent studies have shown phosphorous to be the main limiting nutrient in regards to phytoplankton primary production in the western Wadden Sea (Philippart et al., 2007 and Ly et al., in prep.). While this is contradictory to the previous theories as the Wadden Sea is a highly dynamic water body (Van Raaphorst and De Jonge, 2004) with high input of freshwater from surface runoff and nearby estuaries (Graneli and Sundback 1985). This, as with some other coastal waters in close proximity with large rivers and estuaries, hint that the high freshwater input in coastal areas can result in areas of phosphorous limitation, even in marine environments. In some cases, such as the southern area of the North Sea, nutrient input from freshwater can account for around 75% of the water body (Skogen et al., 2004).
2.1.2 Management This decrease in phosphorus is not strictly due to naturally occurring P-‐limitations in the freshwater inputs, but rather is in part a result of de-‐eutrophication efforts. Due to increased occurrences of toxic algal blooms pre-‐1987 (Cadee 1986), during a international conference in early 1980’s European countries developed a target to reduce amounts of nutrients in their major rivers (mainly nitrogen and phosphorous) and therefor entering the coastal zones. Phosphorous, due to municipal improvements, was decreased by 50% (target
6
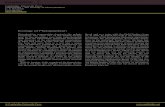
achieved) while nitrogen was decreased by the lesser 30% (Skogen et al. 2004). Figure 1 shows the phosphorous decrease This uneven decrease in the N and P loads in freshwater inputs have also brought a decrease in the retention of Si, which then also flushes to coastal zones, increasing the coastal Si load. The increase in Si is key to producing increased diatom blooms in Dutch coastal waters within the North Sea (Prins et al. 2012). Some might say this could be beneficially, as it is known that diatoms carry a greater capacity to take up carbon (Prins et al. 2012 and Smetacek 1999) providing a greater energy transfer towards higher trophic levels (Litchman et al. 2009 and Prins et al. 2012) or alternatively transporting carbon towards the ocean floor when sinking (Smetacek 1999). On the other hand there is a choice to be made whether this increased energy transfer outweighs the occurrences of harmful diatom blooms (Prins et al. 2012). Different outcomes have been recorded, some showing a decrease in biomass within already low phosphorous water environments (Skogen et al. 2004), while others have documented that even with the decreases in phosphorous and nitrogen, net increases in both concentrations can be seen over the last century (Cloern, 2001).
Still there have been recent cases of harmful algal blooms (Philippart et al. 2007; Schindler 2006; Anderson 2002 and Cloern 2001), which have researchers even more invested in the ‘nutrient load hypothesis/nutrient resource ratio’ put forward by Tilman in 1986. This theory states that total nutrient load may not be the only factor
Figure 1: Decreasing phosphorous concentrations from sea’s around the Netherlands. Decreases in phosphorus can be seen in 1985 (implementation of phosphorus reduction (Ecomare, undated)

7
in species competition and therefore composition, but that nutrient ratio also is important in determining species composition (Tilman 1986, Miller et al. 2005). There is much and equal evidence for both ratio (e.g. Vrede et al., 2009) and absolute amount (e.g. Downing et al., 2001) being the predecessor of composition with absolute amount being more important in eutrophic conditions (Brauer et al., 2012). A few have proposed the theory that nutrient enrichment causes the competition for nutrients to shift to light (Brauer et al., 2012; Passarge et al., 2006; Kilham 1986 and Huisman et al., 2002).
2.2 Competition In order to understand why certain species are dominant over others, competition experiments and models are implemented. Best described by Brauer et al. (2012) when looking at competition between two species: If both species are limited by the same nutrient then the species with the lowest R* value for said nutrient will outcompete the other (seen in figure 2A). R* value refers to the minimum amount of the resource needed to produce positive growth (Miller et al. 2005). Limiting nutrients are taken up faster than others allowing species to push their environment to favor their limiting conditions. However, when two species are competing for different limiting nutrients, a trade off can exist where stable coexistence and alternative stables states may arise (seen in figure 2B) (Litchman et al. 2007).
2.2.1 nutrient limitations Here species A has the lowest R* for phosphorous while species B has the lowest R* for nitrogen. From this we can say that species A is a good competitor for phosphorous and species B for nitrogen. However the nutrient supply point and consumption vectors determine the species composition. When the initial nutrient supply point is within the consumption vectors stable coexistence between both species is possible. When the nutrient supply point is outside of these vectors, both species may be present although consumption of their nutrients will soon push the initial conditions towards a

8
limitation (by quick consumption of limiting nutrient) (represented by bold arrows in figure 2B) that ultimately eliminates one of the species. Alternative stables states, although difficult to prove in the field, is still a possible outcome of competition. Seen in figure 2C, alternative stable states are determined by the initial abundance of species A and B and the time of the initial nutrient supply (Brauer et al. 2012). If it is able to monopolize the limiting resource, the more abundant species will exclude others.
2.2.2 light limitation When nutrients are plentiful, the composition of phytoplankton will not be dictated by competition for nutrients but rather a third resource becomes limiting; light. At a certain point species will start
A B
C D
Figure 2: competition models for competitive exclusion (A), co-‐existence (B), alternative stable states (C) and addition of light (D). Green dotted line represents consumption vectors, blue and red isocline represents species A and B, orange circles represent initial nutrient supply points and bold arrows represent ‘pushing the environment’ (made by author, based off of Tilmans work).

9
to self-‐shade each other and therefor cause light limitation (Anderson et al. 2002). When light limitation is added into competition models (figure 2D) we can see that a third species (C) is present. This is because for species C, light is the limiting resource for which it is the best competitor. Alternatively it could be that species A for example also happened to be the best competitor for light and therefore would outcompete in both environments, this is a situation where there is no tradeoff.
2.3 Aims This study was carried out alongside the CHARLET project (CHanges in Resource Limitation and Energy Transfer). The project started in January 2011 and is focusing their research on three main criteria: 1. The recent changes from nitrogen to phosphorous limitation in the coastal North Sea. 2. Whether these changes can affect the phytoplankton composition? 3.Whether these changes create positive or negative impacts in the aquatic environment. Positive referring to an increase of the nutritional value of phytoplankton thus increasing the energy to higher trophic levels, or negative leading to unhealthy cells prone to viral lysis and therefore energy remaining trapped within the microbial loop (NWO). This study focuses on criterion number 2; will changes in N:P ratio impact the phytoplankton community of the North Sea? To address this, we worked with chemostats, which are used as a model of a marine ecosystem (Groove 2000) to investigate phytoplankton composition changes. Using an inoculum taken from the North Sea during an early spring cruise and growing under different N:P ratios and loads we can see how these impact community composition. Applying the theory of Brauer et al. (2012), Tilman (1985), and Huisman & Weissing (1994, 1995) to the observed results we can begin to understand the potential impact the increasing N:P ratio of the North Sea will have on its phytoplankton composition.

10
III. Materials and methods The research cruise was carried out during 15-‐22nd March 2013 on the North Sea. Niskin bottles were attached to a conductivity, temperature and depth device and 2.5 L of seawater was collected from a depth of 7 meters. The water was then used to rinse and fill a container. This was repeated for seven different stations along the transect (figure 3). At each station, water collected was filtered through a 200-‐micrometer sieve to eliminate zooplankton. After which, CO2 gas was bubbled through the filtered seawater for 30 minutes to eliminate any leftover zooplankton. All seawater samples were then mixed together in equal proportions, to account for preconditioning, and transported back in one 10 L carboy to the lab and stored at 4oc. In the laboratory, seven chemostats were constructed as in Huisman et al. 2002 (Figure 4) using full-‐spectra white fluorescent bulbs as the light source. Magnetic stirrers were placed in each chemostat to avoid accumulation of sticky and heavy species. Chemostats were
positioned for an average Iin (light entering the chemostat) of 40μmol photons m-‐2 s-‐1 and flow-‐through rate was adjusted to allow for a 0.20 day-‐1 dilution period. Seawater inoculum was added to fill half the volume of the chemostats to which nutrient medium (Table 1A) with seven different nitrogen
(NaNO3):Phosphorous (K2HPO4•3H2O) ratios (Figure 1B) was added through peristaltic pumps set to a steady flow rate. Overflow was passed into sterile waste tanks. Required volume of sample per day
9
The CHARLET-2 cruise 2012 We studied during late spring the biology of plankton (with a major focus on phytoplankton) in the North Sea, along a transect from coastal waters to central North Sea water. This cruise was undertaken as part of larger integrated study with the main merit of assessing the production, composition and losses of phytoplankton in order to determine the limiting factors for phytoplankton growth in the North Sea and study how these limiting factors affect the food quality and species composition of the phytoplankton. This is the first of four cruises, during spring and summer. The cruise track is shown in Figure 2. Station details in Table 1 and the participant and crew list in Table 2.
Fig. 2. Cruise track CHARLET-1, summer 2011. This cruise was longer in duration. During the present CHARLET-2 cruise, May 2012, we have sampled from south to north stations 13, 11, 10, 8, 7, 4, 2, and 1.
Figure 3: satellite image of stations (transect). Water samples taken from station 1, 2, 4, 7, 8, 10, 11 and 13 (REF).
Figure 4: skematic side view diagram of the chemostat arrangement Huisman et al. (2002)

11
were calculated so that minimum +10 ml were taken from the vessel to insure minimal disruption to chemostat and phytoplankton conditions.
Table 1: A) list of solutions used for the nutrient medium. B) Chemostat, N : P ratio and corresponding concentrations of nitrogen and phosphorous solutions.
B ! !
Concentration!(µM)!Chemostat! Ratio! NaNO3! K2HPO4•3H2O!LowN:LowP' 16:1' 64' 6'MidN:MidP' 16:1' 160' 10'HighN:HighP' 16:1' 2000' 125'LowN:HighP' 0.521:1' 64' 125'HighN:LowP' 500:1' 2000' 4'MidN:HighP' 1.28:1' 160' 125'HighN:MidP' 200:1' 2000' 10'
!
A Compound:
Concentration (µmol/L)
Mol. Wgt.of Compound
Salts/Buffers: MgSO4•7H20 2.0x104 246
KCl 8.0x103 74 CaCl2•2H2O 2.5x103 146
NaCl 4.3x105 58 NaHCO3 500 84
Macro nutrients: NaNO3 2000; 160; 64 85
K2HPO4•3H2O 125; 10; 4 229 Na2SiO3•5H2O 160 261
H3BO3 550 62 Micro nutrients:
FeSO4•7H20 14 278 Na2EDTA 35 338
MnCl2•4H2O 22 197 ZnCl2 2.4 135
Na2MoO4•2H2O 5.4 242 CuSO4•5H2O 0.2 249 CoCl2•4H2O 0.5 201 Vitamins:
Thiamine•HCl (B1) 0.6 337 Biotin 4.0x10-3 244
Cyanocobalamin (B12) 7.4x10-3 1355

12
The Iout (light exiting the chemostat) and pH were measured daily, making sure pH was measured straight after the sample was taken to allow for the most accurate reading. -‐ Iout using a light meter and a template with 10 wholes an average was calculated. -‐ pH using a pH meter (SCHOTT instruments lab 860) recorded only after a constant reading for 5 seconds. When necessary, adjustments to the CO2 quantity inputted to the bubbling system were performed to maintain a pH of approximately 8. Three times per week flow cytometery and Lugol iodine samples were taken for species composition analysis. Flow cytometery samples were taken by pipetting 4 ml of each samples into two 5 ml cryogenic vials labeled accordingly and fixed with 0.5 ml formaldehyde hexamine, placed in the refrigerator for half an hour then flash frozen and stored at -‐800C. Lugol iodine samples were taken by pouring 14 ml of each sample into a 15 ml centrifuge vial labeled according. Then 0.5 ml of Lugol’s iodine solution were added, mixed thoroughly and stored in the dark. Once per week dissolved inorganic nutrients were taken (nitrogen-‐ DIN, phosphate-‐ DIP and silicate-‐ DISi). -‐ Dissolved inorganic nutrients: 20 ml (of each sample was filtered through a 25 mm Whatman GF/F filter(s) (pore size 0.45 µm) into 2 acid washed and labeled scintillation vials using the first 5 ml to double rinse the vials and stored at -‐200C.
3.1 Analysis Microscopy was used to identify and count cells larger than 10μm (magnification limitations). For this the Lugol iodine samples collected were transferred to 10 ml Utermöhl settling chambers that were left over night in the dark to settle. The entire chamber was studied under an inverted microscope (Leica DMIRB) where species and total counts were recorded. When samples became too dense, dilutions were carried out and then read on a Sedgwick – rafter

13
gridded counting S50 microliter slide. Complete rows were analyzed until 200 counts were recorded per species. After all dates had been viewed under the microscope a second look was taken (at a higher concentrations) to dates where certain species were expected to be seen (ie. were present before and after missing dates). For cells that were less than 10μm, abundance was determined using a flow cytometer (FCM). All FCM samples were pre-‐filtered using 10μm Whatman Nuclepore membrane filters to make sure that nothing counted on the microscope was recounted. These were then placed into the FCM machine and set for a maximum total count of 20000 to reduce noise. For samples that had low abundance a flow through time of 5 minutes was set. When all samples were run, using fluorescence of chlorophyll and phycocyanin and size of cells as guides, three counting gates (Chlorella sp., cyanobacteria 1 and cyanobacteria 2) were determined and cells within those gates were recorded. All species counts were converted to counts/ml and then further to biovolume using the approximate average geometric shape of each species. The penultimate five days of counts were averaged for stable state species composition. For graphical purposes all diatoms were placed in shades of brown, dinoflagellates shades of red, green algae in shades of green and cyanobacteria in shades of blue and transformed to a logarithmic scale. Stable state species composition was tested for species similarity (presence/absence) using the Sorensen similarity index (below).
Sorenson presence/absence equation 2 (# of species in common) S = # of species in x + # of species in y

14
3.2 Dissolved inorganic nutrients
3.2.1 DIN For nitrate + nitrite analysis (the only added nitrogen source) 2.5 ml of sample were added to 20 ml scintillation vials with 8 vials used for standards. To all vials 0.5 ml ammonium chloride buffer were added. Cadmium pellets (previously activated by 6M HCL) were then added to each, capped and placed on a shaker (150 rpm) for 2.5 hours. One ml of each vial was then pipetted into 2 ml eppendorf tubes along with 50μl of Reagent 2 and 3 (appendix) (in consecutive order), capped, inverted and left to develop for 10 minutes. Once developed, 310μl were then pipetted into wells on a 96 well plate and read at 540 nm on a micro plate reader (Molecular devices Versa max tunable micro plate reader). Calculations of concentrations were determined using the linear regression of the standard curve to absorbtion.
3.2.2 DIP For phosphate analysis, 300μl of each sample and standard were pipetted into wells in a 96 well plate along with 30μl of mixed reagent (Ammonium molybdate, sulfuric acid, ascorbic acid and antimonyl potassium tartare). Well plates were left for 1 hour to develop color and then read at 850 nm (Molecular devices Versa max tunable micro plate reader). Calculations of concentrations were determined using the linear regression of the standard curve to absorbtion.
3.2.3 DISi Dissolved inorganic silicate was analyses to make sure silicate wasn’t limiting.

15
IV. Results
Figure 5: species composition in biovolume (μm3) of increased load scenario chemostats. LN:LP (a), MN:MP (b) and HN:HP (c). Text at bottom right of each graph refers to chemostat ID. Brown lines – diatoms, orange line – dinoflagellate, green line – green algae, blue lines– cyanobacteria. Dark colours represent the highest biovolume within that group.
C
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
1E+09"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
Days)Pseudonitzschia.delica0ssima. Thalassiosira.angulata. Asterionellopsis.glacialis. Cylindrotheca.closterium. Pseudonitzschia.seriata.Skeletonema.costatum. Chaetoceros.affinis. Navicula.delicatula. Navicula.transitrans. Dinoflagellete.sp..Tryblionella.sp.. Thallassorias.longissima. Navicula.directa. Nitzschia)agnita) Coscinodiscus.sp..(s).Coscinodiscus.sp..(l). Ditylum.sp.. Melosira.nummoloides. Delphinies.minu0ssima. Chaetoceros.atlan0cus.Cocconies.sp.. Nitzschia)pusilla) Chlorella)sp.) Cyanobacteria)"1") Cyanobateriac)"2")
A
LowN : LowP
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm3))
Days)Navicula(directa( Skeletonema(costatum( Pseudonitzchia(delica6ssima( Cylindrotheca(closterium( Pseudonitzchia(seriata( Thallassitrix(longissima(Dinoflagellate(sp.( Coscinodiscus(sp.((l)( Coscinodiscus(sp.((s)( Asterionellopsis(glacialis( Thallassiosira(angulata( Bellerochea(malleus(Triblionella(sp.( Chaetoceros(laciniosus( Atheya(sp.( Nitzschia)agnita) Nitzschia)pusilla) Cocconies(sp.(Chlorella)sp.) Cyanobacteria)"1") Cyanobacteria)"2")
MidN : MidP
B
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
1E+09"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
Days)Thallassiotrix+longissima+ Skeletonema+costatum+ Cylindrotheca+closterium+ Pseudonitzchia+delica9ssima+ Asterionellopsis+glacialis+ Pseudonitzchia+seriata+Thallassiosira+angulata+ Navicula+delicutula+ Coscinodiscus+sp.+(s)+ Coscinodiscus+sp.+(l)+ Chaetoceros+affinis+ Thallassiosira+sp.+Bellerochea+malleus+ Tryblionella+sp.+ Dinoflagellete+sp.+ Melosirus+nummoloides+ Guinardia+sp.+ Atheya+sp+Nitzschia)agnita) Delphinies+minu9ssima+ Cocconies+sp.+ Pleurosigma+directa+ Nitzschia+pusilla+ Chlorella)sp.)Cyanobacteria)"1") Cyanbacoteria)"2")
HighN : HighP
C

16
Figure 6: Light and dissolved inorganic nutrients (DINuts) nitrogen (DIN) and phosphorous (DIP) for increased load scenario chemostats. LowN:LowP light –A and DINuts –B, MidN:MidP light –C and DINuts –D and HighN:HighP light –E and DINuts –F. Text in middle of each graph group represents chemostat ID. X axis for DINuts apply to corresponding light graphs. Take note of secondary axis in B. Legend applies to all graphs. Red line – Phosphorous, blue line – nitrogen and green line – Iout.
0"
1"
2"
3"
4"
5"
6"
7"
8"
9"
10"
0"
20"
40"
60"
80"
100"
120"
140"
160"
180"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
DIP$concen
tra,
on$(μ
g/l)$$
DIN$con
centra,o
n$(μg/l)$
Days$
0"5"10"15"20"25"30"35"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)& A
LN : LP B
0"
50"
100"
150"
200"
250"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
DINut.'con
centra.o
n'(μg/l)'
Days'
0"5"
10"15"20"25"30"35"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)& E
HN : HP F
0"
10"
20"
30"
40"
50"
60"
70"
80"
90"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
DINut.'con
centra.o
n'(μ'g/l)'
Days'
0"5"10"15"20"25"30"35"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)&
C
MN : MP D

17
4. 1 Low Nitrogen: Low Phosphate Chemostat
4.1.1 Phytoplankton population 17 species identified in the inoculum of chemostat LowN:LowP (Figure 6A) declined to five (N. agnita, N. pusilla, Chlorella sp., Cyanobacteria 1 and 2) in the final days of the experiment. Day 81, four species increased in biovolume (N. pusilla, Chlorella sp., Cyanobacteria 1 and 2) while N. agnita showed signs of a lowering biovolume. During days 51-‐63 Cocconies sp. was recorded. This particular species increased from 6.97x104 to 4.86x109μm3/l in seven days (51-‐58 days) and subsequently decreased. At the same time N. pusilla, N. agnita and D. minutissima decreased. N. agnita showed a rapid increase (day 49) and decrease (day 51) from 7.31x105 to 8.32x107 and back to 3.61x106μm3/l. Triblionella sp. (yellow line) increased steadily until day 23 where it decreased to 6.64x105μm3/l by day 44 to where numbers were to low to be detected.
4.1.2 Abiotic parameters In the LowN:LowP chemostat (figure 6A) the Iout (light) remained level with a slight decrease. Both DIN and DIP (figure 6B) increased in the first two weeks followed by a decrease at week three. Week four (day 29) onwards DIN remained constant at <10 μg/l. After day 29 DIP stayed on average at around 3.8μg/l with a 1.4μg/l increase between days 43 and 50. Both nutrients remained steady with final DIN and DIP concentrations at 5.6 and 3.0µg/l respectively.
4. 2 Mid-‐Nitrate: Mid-‐Phosphate Chemostat
4.2.1 Phytoplankton population In the MidN: MidP chemostat (Figure 5B), species richness was the highest at day seven (13 species) which then rapidly dropped to five by week two. By day 39 the five species that remained were N. agnita, N. pusilla, Chlorella sp. and Cyanobacteria 1 and 2. From day 25 Cyanobacteria 1 had the lowest biovolume until day 81 where it outcompeted Cyanobacteria 2, N. pusilla and Chlorella sp. in the last 10 days.
4.2.2 Abiotic parameters Looking at the MidN:MidP chemostat (figure 6C) Iout shows fluctuations with an overall decrease from 29 to 18.6μE’s. Both DIN

18
and DIP showed the same trend throughout the experiment (figure 6D), a large increase during the first week (23.3 to 74.4μg/l and 7.6 to 18.9μg/l respectively) and a subsequent decrease during week two. Both remained level ending at concentrations of 2.8μg/l (DIP) and 8.2μg/l (DIN).
4.3 High Nitrate: High Phosphate Chemostat
4.3.1 Phytoplankton population Only nine species were recorded in the HighN:HighP with a maximum of 19 by day seven. At the end of the experiment only four species were recorded (N. agnita, Chlorella sp. and Cyanobacteria 1 and 2. N. pusilla remained the dominant species from day 28. At days 44–72 N. pusilla decreased (1.33x108 to 5.70x105μm3/l) while both Chlorella sp. and Cyanobacteria 1 increased until numbers of N. pusilla suddenly were below detection (Figure 5C).
4.3.2 Abiotic parameters Chemostat HighN:HighP showed a steady decrease in Iout from 29.6 to 0.4 µE’s (figure 6E) with a brisk increase day 16-‐23. At day 70, Iout remained constant between 0.3 -‐0.7µE’s. DIN remained (with slight fluctuations) at 233.5μg/L until day 57 where it decreased (209.2 to 85.5μg/l) ending on a concentration of 81.3 (Figure 6F). DIP increased (38.1 to 62.2μg/l) in the first week and then remained leveled at around 60μg/l (+/-‐ 10μg/l). At day 50 DIP decreased steadily ending on a concentration of 7.9μg/l.

19
Figure 7: Species composition in boivolume (μm3) of extreme limitation scenario chemostats. LN:HP (a) and HN:LP (b). Text at bottom right of each graph refers to chemostat ID. Brown lines – diatoms, orange line – dinoflagelatte, green line – green algae, blue lines– cyanobacteria. Dark colours represent the highest biovolume within that group.
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
1E+09"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
Days)Pseudonitzschia.delica0ssima. Pseudonitzschia.seriata. Asterionellopsis.glacialis. Thalassiosira.angulata. Skeletonema.costatum. Dinoflagellete.sp..Cylindrotheca.closterium. Chaetoceros.affinis. Coscinodiscus.sp..(s). Pleurosigma.normani. Bellerochea.malleus. Thallassiothrix.longisima.Meliosira.nummoloides. Delphinies.minu0ssima. Coscinodiscus.sp..(l). Tryblionella.sp.. Atheya.sp.. Nitzchia(agnita(Cocconies.sp.. Nitzschia(pusilla( Chlorella(sp.( Cyanobacteria("1"( Cyanobacteria("2"(
HighN : LowP
B
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
Days)Thallassiotrix+longissima+ Navicula+transitrans+ Navicula+delicutala+ Pseudonitzchia+delica7ssima+ Cylindrotheca+closterium+ Chaetoceros+tor7ssimus+Asterionellopsis+glacialis+ Pseudonitzchia+seriata+ Melosira+nummoloides+ Thallassiotrix+angulata+ Skeletonema+costatum+ Dinflagellate+sp.+Chaetoceros+affinis+ Coscinodiscus+sp.+(s)+ Coscinodiscus+sp.+(l)+ Bellerochea+malleus+ Tryblionella+sp.+ Navicula+directa+Nitzschia)agnita) Thalossiosira+delicatula+ Delphinies+minu7ssima+ Chaetoceros+lacinosus+ Atheya+sp.+ Cocconies+sp.+Nitzschia)pusilla) Chlorella)sp.) Cyanobacteria)"1") Cyanobacteria)"2")
LowN : HighP
A
20
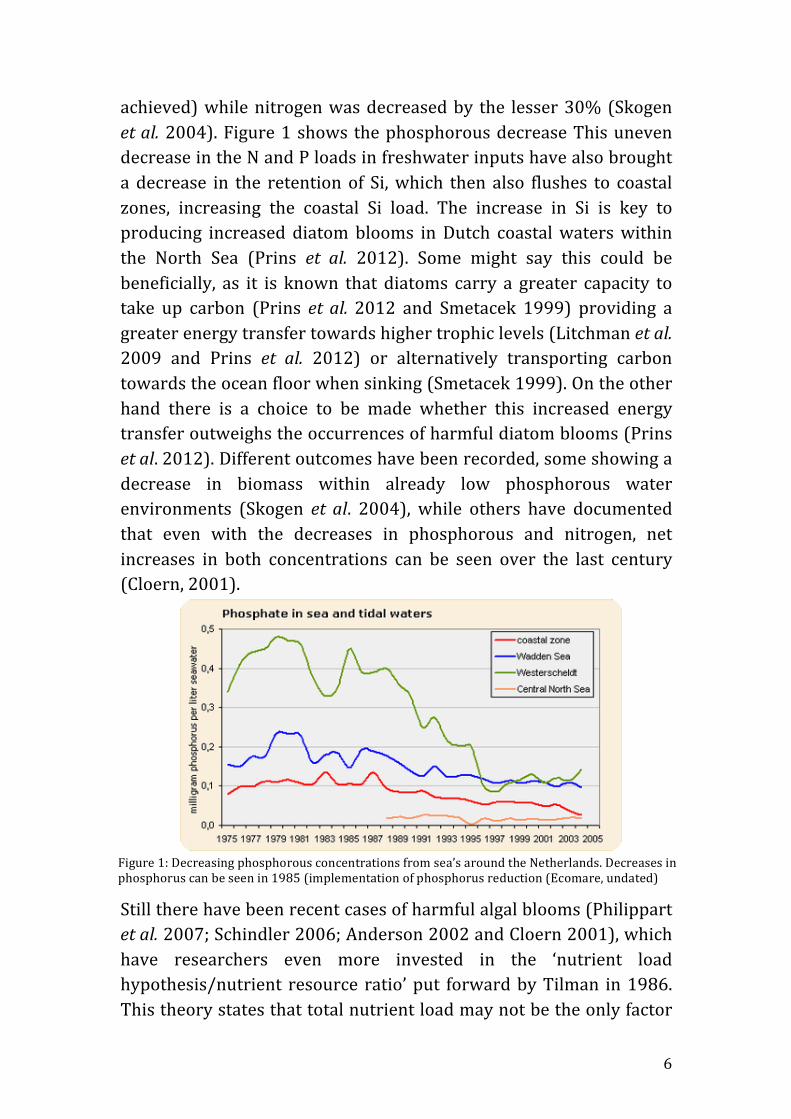
Figure 8: Light and dissolved inorganic nitrogen and phosphorous for extreme limitation scenario chemostats LowN:HighP light –A and DINuts –B and HighN:LowP light –C and DINuts –D. Text in middle of each graph group represents chemostat ID. X axis in DINuts apply to corresponding light graphs. Take note of secondary axis in D. Legend applies to all graphs. Red line – Phosphorous, blue line – nitrogen and green line – Iout.
4.4 Low Nitrate: High Phosphate Chemostat
4.4.1 Phytoplankton population Chemostat LowN:HighP (Figure 7A) started with 20 species and decreased towards 11 by the start of the third week. At this time a rapid increase in biovolume for species D. minutissima, N. agnita, N. pusilla, Chlorella sp. and both cyanbacteria species can be seen. With the exclusion of D. minutissima these five species constituted the
0"2"4"6"8"10"12"14"16"18"20"
0"
100"
200"
300"
400"
500"
600"
700"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"DIP$concen
tra,
on$(μ
g/l)$
DIN$con
centra,o
n$(μg/l)$
Days$
0"
5"
10"
15"
20"
25"
30"
35"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)&
D
C
HN : LP
0"
5"
10"
15"
20"
25"
30"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)&
0"
20"
40"
60"
80"
100"
120"
140"
160"
180"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
DINu
t.'concen
tra.o
n'(μg/l)'
Days'
B
A
LN : HP

21
final species composition. Species D. minutissima increased at day 45 where N. pusilla, Cyanobacteria 2 and Chlorella sp. showed opposite trends.
4.4.2 Abiotic parameters In chemostat LowN:HighP (Figure 8A) Iout decrease throughout (26.8 to 23.9μE’s). In regards to nutrients (Figure 8B) DIP remained at a higher concentration then DIN from the second week. DIN had a high concentration of 112.9μg/l and decreased to 85.7μg/l within the first week. It remained constant until the fourth week where DIN decrease suddenly from 30.4 to 4.1μg/l where it then remained under 10μg/l with the exception of day 64 (tenth week). DIP and DIN had final concentration of 55.6 and 3.9μg/l, respectively.
4.5 High Nitrate: Low Phosphate Chemostat
4.5.1 Phytoplankton population The widest range of species were present at the start (18) of HighN:LowP (Figure 7B), which fell to five towards the end (N. agnita, N. pusilla, Chlorella sp. and Cyanobacteria 1 and 2). Cocconies sp. were detected during days 53-‐60 with the highest overall biovolume at day 60 of 3.53x108μm3/l after which abundance rapidly dropped to below detection. Cyanobacteria 2 had many fluctuations throughout and from day 45 onwards had the lowest biovolume of all five species until day 70 where an increased to around 1x106μm3/l, where it remained constant until the end.
4.5.2 Abiotic parameters Chemostat HighN:LowP Iout stayed steady until day 23 where numbers fell to 14μE’s at day 37 reaching the minimum value (figure 8C). It then remained constant until day 58 where it increased and leveled out at around 22μE’s. Looking at figure 8D, DIN remained between 270.8 and 344.4μg/l until the seventh week (day 35-‐45) where the concentration rose to 627.9 (DIN maximum). Two weeks after, it had decreased to 280μg/l where it undulated +/-‐100μg/l ending on a concentration of 605.2μg/l. DIP started at a concentration of 8μg/l where it increased to 12.4μg/l by day 15. After its decrease by day 22 DIP fluctuated between 5.6 and 3μg/l until the end where it reached a final concentration of 2.5μg/l.

22
Figure 9: Species composition in boivolume (μm3) of middle ratio scenario chemostats. MidN:HighP (a) and HighN:MidP (b). Text at bottom right of each graph refers to chemostat ID. Brown lines – diatoms, orange line – dinoflagelatte, green line – Chlorella sp., blue lines– cyanobacteria. Dark colours represent the highest biovolume within that group.
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
1E+09"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
)
Days)
Skeletonema*costatum* Pseudonitzschia*delica3ssima* Cylindeotheca*closterium* Thallassiosira*angulata* Thallassiothrix*longissima* Navicula*directa*Pseudonitzschia*seriata* Coscinodiscus*sp.*(s)* Coscinodiscus*sp.*(l)* Tryblionella*sp.* Atheya*sp.* Bellerochea*malleus*Dinoflagellate*sp.* Thalassiosira*delicatula* Asterionellopsis*glacialis* Chaetoceros*affinis* Nitzschia)agnita) Pleurosigma*directa*Melosira*nummoloides* Nitschia)pusilla) Cocconies*sp.* Chlorella)sp.) Cyanobacteria)"1") Cyanobacteria)"2")
MidN : HighP
A
1"
10"
100"
1000"
10000"
100000"
1000000"
10000000"
100000000"
1E+09"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Biovolum
e)(μm
3))
Days)Navicula(directa( Skeletonema(costatum( Cylindrotheca(closterium( Thallassiotrix(angulata( Dinoflagellate(sp.( Pseudonitzschia(seriata(Asterionellopsis(glacialis( Pseudonitzschia(delica@ssima( Atheya(sp.( Coscinodiscus(sp.((s)( Coscinodiscus(sp.((l)( Bellerochea(malleus(Cheatoceros(affinis( Dictyocha(sp.( Tryblionella(sp.( Chaetoceros(tor@ssimus( Nitzschia)agnita) Cocconies(sp.(Delphinies(minu@ssima( Navicula)pusilla) Chlorella)sp.) Cyanobacteria)"1") Cyanobacteria)"2")
B
HighN : MidP
23
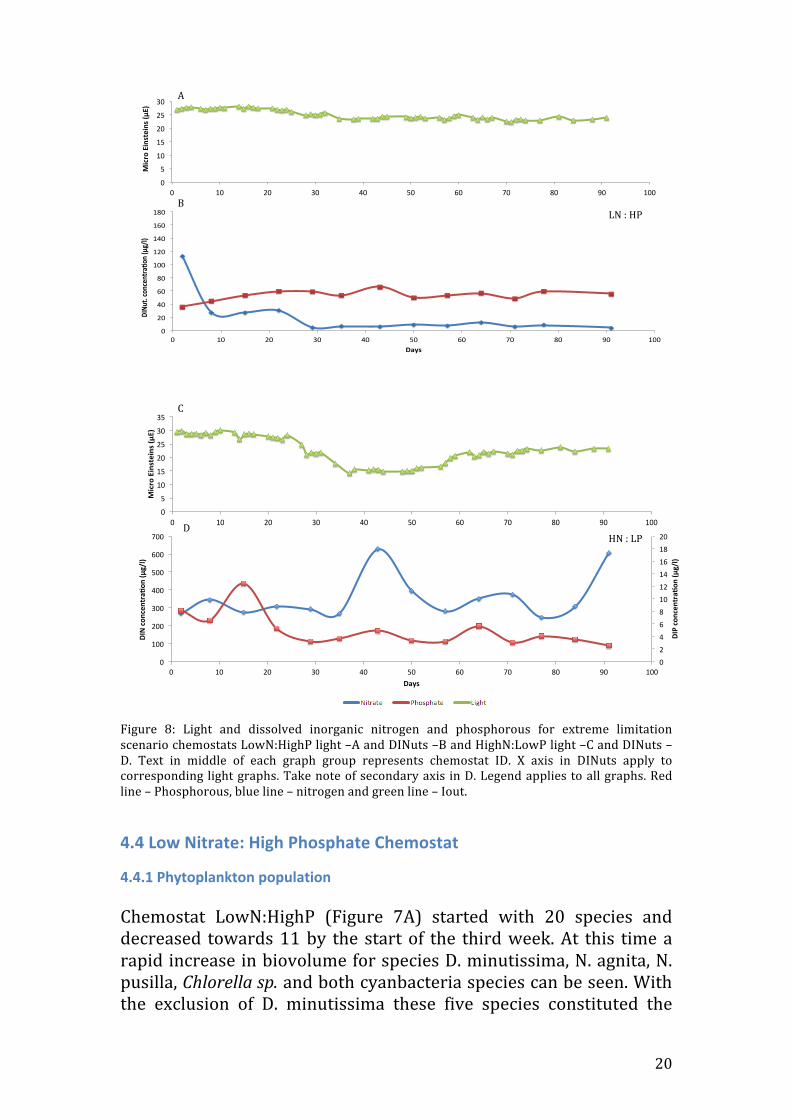
Figure 10: Light and dissolved inorganic nitrogen and phosphorous for middle raio scenario chemostats. MidN:HighP light –A and DINuts –B and HighN:MidP light –C and DINuts –D. Text in middle of each graph group represents chemostat ID. X axis in DINuts apply to corresponding light graphs. Take note of secondary axis in D. Legend applies to all graphs. Red line – Phosphorous, blue line – nitrogen and green line – Iout.
4.6 Mid-‐Nitrate: High Phosphate Chemostat
4.6.1 Phytoplankton population MidN:HighP (figure 9A) started with 12 species and dropped to five (N. agnita, N. pusilla, Chlorella sp. and Cyanobacteria 1 and 2) towards the end of the time series. N. pusilla had the largest biovolume of all species present from days 28 to 63 until Chlorella sp. overtook between days 65 -‐74. Cyanobacteria 2 had a fluctuating
0"
10"
20"
30"
40"
50"
60"
70"
80"
90"
100"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
DINut&con
centra-o
n&(μg/l)&
Days&
0"
5"
10"
15"
20"
25"
30"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)&
A
MN : HP
0"5"10"15"20"25"30"35"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"
Micro&Einsteins&(μ
E)&
0"2"4"6"8"10"12"14"16"18"20"
0"
100"
200"
300"
400"
500"
600"
700"
0" 10" 20" 30" 40" 50" 60" 70" 80" 90" 100"DIP$concen
tr+o
n$(μg/l)$
DIN$con
centra+o
n$(μg/l)$
Days$
D
C
HN: MP

24
biovolume throughout, and at day 18 became one of the lowest three biovolumes. It then drastically increased from 4114μm3/l to 1x108μm3/l outcompeting all species for the largest biovolume for the last two days.
4.6.2 Abiotic factors In MidN:HighP Iout started at 27.9μE’s and remained steady until day 10 where it decreased and reached 21.4μE’s by day 16 (figure 10A). It then proceeded to increase again to 25.5μE’s by day 24 to which it decreased steadily to day 56 (14.6μE’s). From here on Iout fluctuated until the end ranging between 14.6 and 18.1μE’s. From weeks one to two (days 2 to 8) both DIN and DIP increased, DIN from 26.1 to 34.7μg/l and DIP from 48.9 to almost double at 82.5μg/l. DIN and DIP stayed at a constant level further on the time series (with a slight fluctuation in DIP during day 60-‐91) ending on concentration of 2.3 and 57.4μg/l respectively (Figure 10B).
4.7 High Nitrate: Mid-‐Phosphate chemostat
4.7.1 Phytoplankton population In the HighN:MidP (figure 9B) nine species were recorded at day three which increased to 16 by day nine to which species diversity declined until 5 species left (day 30) (N. agnita, N. pusilla, Chlorella sp. and Cyanobacteria 1 and 2), with the exception of Cocconies sp appearing at day 58. Chlorella sp. dominated in biovolume from day 42 until the end with Cyanobacteria 1 and N. agnita switching between second and third. Cyanobacteria 2 had the lowest biovolume.
4.7.2 Abiotic parameters The HighN:MidP chemostat (figure 10C) showed a steady decrease throughout with the exception of day 71 where the alternative chemostat was measured (due to breakage). Overall Iout decreased from a value of 31.4μE’s to 8.5μE’s. Both DIN and DIP showed a similar trend throughout (figure 10D). DIN started at a concentration of 341μg/l and increased to 442.5 in the first week then decreased to 277.2μg/l where it stayed constant throughout and decreased to 158.8μg/l in the last 2 weeks. DIP started at 9.6μg/l where it increased 2-‐fold in the first week (18.4μg/l). During the third week it decreased to 4.5μg/l and remained constant between 5.5 and 3μg/l.

25
Figure 11: Comparing diversity (A) and dominance (B) between chemostats at day one. Diversity calculated by quarter root of day one counts.
Table 2: Sorensen index comparing similarity (presence or absence of species) of the start composition between each chemostat (equation in method section). A Sorensen index of 1 indicates 100% similarity, 0 means 100% dissimilar
4.9 All chemostats: Summary
4.9.1 Beginning The beginning composition of all chemostats was varied. The extreme conditions and low load (LowN:HighP, HighN:LowP and LowN:LowP) contained the largest diversity with 18, 17 and 14 species respectively (Figure 11A). The remaining chemostats contained eight species while MidN:HighP contained nine. The chemostats LowN:LowP, MidN:HighP and MidN:MidP showed a dominance by Chlorella sp. (Figure 11B). In HighN:HighP Thallasiothrix angulata seems to be the most dominant at 70%. The
!! LH! LL! HH! HM! HL! MH! MM!LH! !! 0.81! 0.62! 0.64! 0.80! 0.52! 0.54!LL! 0.81! !! 0.64! 0.55! 0.77! 0.52! 0.64!HH! 0.62! 0.64! !! 0.38! 0.56! 0.71! 0.50!HM! 0.64! 0.55! 0.38! !! 0.56! 0.47! 0.75!HL! 0.80! 0.77! 0.56! 0.56! !! 0.54! 0.56!MH! 0.52! 0.52! 0.71! 0.47! 0.54! !! 0.71!MM! 0.54! 0.64! 0.50! 0.75! 0.56! 0.71! !!!
26
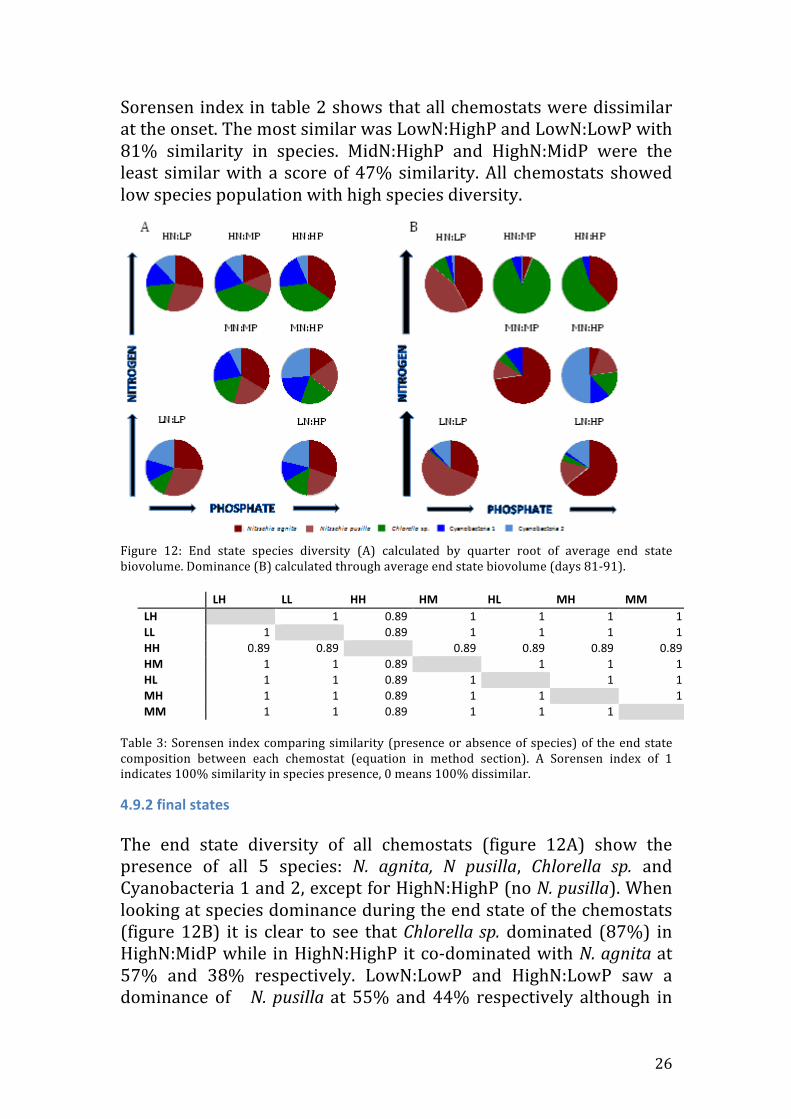
Sorensen index in table 2 shows that all chemostats were dissimilar at the onset. The most similar was LowN:HighP and LowN:LowP with 81% similarity in species. MidN:HighP and HighN:MidP were the least similar with a score of 47% similarity. All chemostats showed low species population with high species diversity.
Figure 12: End state species diversity (A) calculated by quarter root of average end state biovolume. Dominance (B) calculated through average end state biovolume (days 81-‐91).
Table 3: Sorensen index comparing similarity (presence or absence of species) of the end state composition between each chemostat (equation in method section). A Sorensen index of 1 indicates 100% similarity in species presence, 0 means 100% dissimilar.
4.9.2 final states The end state diversity of all chemostats (figure 12A) show the presence of all 5 species: N. agnita, N pusilla, Chlorella sp. and Cyanobacteria 1 and 2, except for HighN:HighP (no N. pusilla). When looking at species dominance during the end state of the chemostats (figure 12B) it is clear to see that Chlorella sp. dominated (87%) in HighN:MidP while in HighN:HighP it co-‐dominated with N. agnita at 57% and 38% respectively. LowN:LowP and HighN:LowP saw a dominance of N. pusilla at 55% and 44% respectively although in
!! LH# LL# HH# HM# HL# MH# MM#LH# !! 1! 0.89! 1! 1! 1! 1!LL# 1! !! 0.89! 1! 1! 1! 1!HH# 0.89! 0.89! !! 0.89! 0.89! 0.89! 0.89!HM# 1! 1! 0.89! !! 1! 1! 1!HL# 1! 1! 0.89! 1! !! 1! 1!MH# 1! 1! 0.89! 1! 1! !! 1!MM# 1! 1! 0.89! 1! 1! 1! !!!

27
the HighN:LowP N. agnita co-‐dominated contributing 42% of the total biomass. In the cases of MidN:MidP and LowN:HighP N. agnita dominated in both cases with 73% and 65% respectively. Cyanobacteria 2 dominated 50% of the species biovolume in MidN:HighP. The Sorensen index in table 3 shows that all chemostats are 100% similar (values of 1) while that of the HighN:HighP was different from the rest with 89% similarity.
Figure 13: Changes in ratio from start to end and final light (ave) value. Chemostat, beginning ratio -‐> end ratio, beginning -‐> end concentration and end Iout. From the figure above (figure 13) it is clear that none of the chemostats became ‘more’ phosphorous limited than had been intended. Instead the five chemostats LowN:LowP, LowN:HighP, MidN:MidP, MidN:HighP and HighN:HighP were pushed more towards nitrogen limited environments. For the other two chemostats HighN:LowP and HighN:MidP phosphorous was still the limiting nutrient but nitrogen was the one nutrient being consumed at a higher rate. For the middle ratio scenarios it is clear to see that HighN:MidP pushed the environment towards a light limitation as both nutrients were not limiting (40N:1P), and that of MidN:HighP pushed the environment towards an extreme nitrogen limitation (even more than the LowN:LowP at 0.431N:1P).
Phosphorous
Nitrogen
HN:LP HN:MP HN:HP500:1*+>*29:1 200:1*+>*40:1 16:1*+>*4.5:12000:4*+>*88:3*μM 2000:10*+>*161:4*μM 2000:125*+>*82:18*μMIout*23*μ'Es Iout*9.5*μE's Iout*0.4*μE's
MN:MP MN:HP16:1*+>*3.:1 1.28:1*+>*0.034:1160:10*+>*9:3*μM 160:125*+>*2:59*μMIout*19*μE's Iout*17*μE's
LN:LP LN:HP16:1*+>*2:1 0.521:1*+>*0.103:164:6*+>*6:3*μM 64:125*+>*6:58*μMIout*26*μE's Iout*24*μE's

28
V. Discussion Looking at both the biovolume and dissolved nutrient/light graphs correlations between the two can be seen. The beginning of all seven chemostats shows a high diversity but relatively low biovolume. Due to the low abundance (low biovolume) but relatively high diversity of species, the flow-‐through rate of nutrients was quicker than the consumption rates, especially by small, low storage capacity cells such as Chlorella sp. and cyanobacteria 2. After a couple of weeks competition was strong which can be seen by the species diversity decreased and the subsequent shifted in composition towards one dominated with diatoms and dinoflagellates such as in the case of LowN:LowP. This increase in size (diatoms/dinoflagellates) and abundance cause nutrient consumption to increase, which is reflected in the prominent decreases (around day 20) in all DIN and DIP graphs except HighN:HighP due to its extremely high load meeting no match of consumption until around day 50.
5.1 Species and their limiting nutrients When looking at species and their effect on nutrients (DIN/DIN) more correlations can be found. For example in LowN:LowP where the slight increase in DIP during days 43-‐49 show a correlation with species N. pusilla, Triblionella sp. and Cocconies sp. When comparing graphs (5A and 8A) at day 49 N. pusilla biovolume decreased where DIP increased. This could be due to N. pusilla previously consuming more phosphorous than other species present, thus when biovolume (abundance) decreases the phosphorous within the chemostat increases. Triblionella sp. was not recorded after day 44, which, if like N. pusilla, also had a high affinity for phosphorous could contribute to the increase in phosphorous availability. At day 77, N. agnita decreased while all other four species increased causing a slight increase in the DIP availability within the system. This gives evidence of a low R* value for diatoms in regards to phosphorous leading to diatoms being the best competitors for phosphorous which corresponds to previous research (Tilman et al., 1986) where they

29
too found diatoms to be superior competitors in phosphorous limitation which may be due to the eradication of predation from dinoflagellates (Hu et al., 2011). HighN:HighP, due to the extreme high load of nutrient input, showed no change in DIN or DIP when the biovolume increased, even at a biovolume of approximately 1.0 X 107 which can be seen by the large decrease in light (high abundance of species) in figures 4C, 8E and 8F). During days 50-‐72 a large decrease in DIN occurred (142μg/l) which coincided with Chlorella sp. and cyanobacteria 2 increase, giving evidence to the notion that both species are nitrogen competitors (Wang et al., 2013). This may also be due to the small size cyanobacteria and Chlorella sp. share, compared to diatoms, as these tend to require more nitrogen than phosphorous (Maranon et al., 2013) therefore through the increased consumption of nitrogen, reduced the nitrogen within the chemostat. Within all three load scenarios, the onset of final state species diversity appeared earlier as load increased. In HighN:HighP the onset appeared at day 29, MidN:MidP on day 35 and in LowN:LowP on day 43. This shows that increased load invoke a more intense competition therefore producing dominance earlier. Tubay et al., (2013) reported that low levels of nutrients produce a lower level of competition. This coheres with other studies that have shown that eutrophication (increase in nutrient load) can lead to an increased biomass of phytoplankton (blooms) which lead to diversity loss (Tubay et al., 2013 and Anderson et al., 2002). These blooms can be toxic/harmful although not all HABs are linked with nutrient enrichment (Anderson et al., 2002).
5.1.1 ‘pushing’ nutrient environments When looking at how the species composition push the environments in MidN:HighP and HighN:MidP, figure 13 showed that limitations were pushed to extremes. MidN:HighP was pushed towards an extreme nitrogen limitation (0.034:1) even more extreme than LowN:HighP (0.103:1). Similarly HighN:MidP was pushed towards a light limitation, as both nutrients were not limiting enough to cause

30
nutrient limitation. HighN:HighP although having a lower Iout value then HighN:MidP also produced an end ratio of 4.5N:1P showing the chemostat to be co-‐limited (Nitrogen and light). This particularly shows that species composition is important as the species most competitive will reduce their limiting resources to their lowest R* (Tilman, 1982) present in the environment as said in the introduction, therefor unavailable to other species (Litchman et al., 2012). In the cases of the other five chemostats, all appeared to be ‘more’ nitrogen limited then phosphorous limited which correlates to Anderson et al., 2002 which showed a relationship between marine phytoplankton production and nitrogen. HighN:LowP and HighN:MidP were the only two chemostats to show signs of phosphorous limitation (29N:1P and 40N:1P respectively). This fits previous consensus showing nitrogen to be the limiting nutrient (Howarth and Marino, 2006).
5.2 Species dominance (beginning and end state) To understand why certain species dominated or why environments were pushed figure 11B, 12B and 13 must be taken into consideration. N. pusilla clearly is favorable towards low phosphate environment which would grant N. pusilla the lowest R* value for phosphorous therefore outcompeting all other species (competitive exclusion) (Tilman et al., 1986). In HighN:LowP however N. agnita is co-‐dominating proposing that is has broader N:P tolerance rage. Due to both having similar biovolume it can be assume N. agnita has the lowest R* for nitrogen therefor both species are able to co-‐exist together (trade offs). To further the statement that N. agnita has the lowest R* for nitrogen we can look at HighN:MidP and HighN:HighP. Here the light limitation predicted turned out to be a co-‐limitation of nitrogen and light while chemostat HighN:MidP was clearly light limiting as both nutrients were not limiting. The N:P ratio shifted towards N limitation rather than exasperating P limitation, which would have been the predicted nutrient to become limiting. If Chlorella sp. was dominant in low light conditions it must have the lowest R* for light resources. This implies successful competition in

31
HighN:HighP for the light which fits theory that Chlorella sp. are the best competitors for light (Tilman et al., 1986). Stole et al., (1994) stated that in eutrophic conditions phytoplankton of larger sizes would dominate due to their increased storage availability, but when compared to the light limiting – eutrophic (HighN:MidP) conditions it is clear that Chlorella sp. dominated even though much smaller than all diatoms. While N. agnita is not present in low light conditions (HighN:MidP) it is present in low light and nitrogen limited conditions (HighN:HighP). This gives evidence that N. agnita may be the source of the nitrogen limitation in HighN:HighP due to its high affinity for nitrogen. This also connects to MidN:MidP and LowN:HighP where N. agnita is a stronger competitor (strong grower) in condition that are initially 16N:1P, but is able to skew the availability of nutrient towards N limitation and still thrive, thus outgrowing its competitors.
However in extremely low nitrate conditions cyanobacteria 2 were able to survive (LowN:LowP and LowN:HighP). Although not dominating like N. pusilla or N. agnita most likely due to species biovolume (Maranon et al., 2013 and Irwin et al., 2006) as diatoms are larger and therefore able to take up more nutrients than required (as well as have a larger storage capacity) than smaller phytoplankton. As a trade-‐off larger phytoplankton growth is limited by biomass. When nitrogen limitation is extreme (MidN:HighP -‐
Figure 14: Pictures of final state species A) Cyanobacteria 1, B) Cyanobacteria 2, C) Chlorella sp., D) Nitzschia pusilla, E) Nitzschia agnita. All photos at 50X using water immersion lense. Scale bar in E applies to all.
A B C
D E

32
0.034N:1P) the diatom’s need for nitrogen isn’t met therefore allowing cyanobacteria 2 to over-‐take and out compete. In low nutrient oceanic waters mutualistic relationships have been found between diatoms and nitrogen fixing bacteria (heterocystous cyanobacteria) where heterocystous cyanobacteria provide nitrogen to the diatoms in close proximity sometimes even 30 minutes after fixation (Foster et al. 2011). In some cases as with Richelia and Calothrix (heterocystic cyanobacteria) an overload of nitrogen fixation was encountered and more than 90% of the total fixed nitrogen is transferred to the diatom partner. After biochemical analysis performed by a colleague on the CHARLET cruise (Julia Grosse) the low nitrogen chemostats showed evidence of nitrogen fixation. This suggests that cyanobacteria 2 is a nitrogen fixating cyanobacteria capable of withstanding low nitrogen environments, which explains the 51% dominance of cyanobacteria 2 within the MidN:HighP chemostat. This finding also coincides with Michard et al. (1996) where he too concluded through water observations of the French reservoir that cyanobacteria blooms (in this case Microcystis aeruginosa) were forming when the ratio of N:P was less than 5 (such as our MidN:HighP) therefore concluding that cyanobacteria would dominant in low nitrogen environments. This study however looked at a reservoir where water stratification would be imminent due to low or no turbulence making conditions perfect for cyanobacteria to float to the top of the water column through gaseous vesicles within their cells (Huisman et al., 2004). Using Huiman et al 2004 findings of low turbulence favoring cyanobacteria and turbulent environments favoring diatoms and Chlorella sp., perhaps if this study was repeated with no turbulence (air mixing) a different composition would arise favoring cyanobacteria.

33
5.3 Summary of end state species In summary it can be said that N. pusilla is the best competitor in low phosphorous conditions, Chlorella sp. for low light conditions, N. agnita and cyanobacteria 2 for low and very low nitrogen conditions, respectively and cyanobacteria 1 is able to withstand exclusion. Overall diatoms are better competitors in low nutrient loads and low phosphorous environments while Chlorella sp. thrive in high nutrient and low light conditions (Potapova and Charles 2002; Kilham 1986) while nitrogen fixing cyanobacteria are superior in extreme nitrogen limitations (Vrede et al., 2009; Ekhom, 2008 and Tilman et al., 1986). Species diversity may shift (through competition) by changing nutrient ratios in low nutrient conditions (Anderson et al., 2002). Nutrient ratio (N:P) can also be a good predictor for the presence of nitrogen fixing cyanobacteria in ratio’s less than 16N:1P (Vrede et al., 2009). On the other hand nutrient load (LowN:LowP, MidN:MidP and HighN:MidP) becomes a better predictor for species dominance especially in high nutrient conditions producing a stronger competition (resulting diversity loss). From the results we can say that the North Sea shows signs of nitrogen limitation due to the increased nitrogen consumption in all chemostats. Both ratio and load are important in predicting species composition where load is more important in light limiting conditions and in phytoplankton dominance. If nutrient ratio and or load change in controlled environments then phytoplankton composition changes. The change, as to whether the composition diversity or dominance will change all depends on what the nutrient change will alter, ratio or load. For the North Sea this still remains uncertain due to its dynamic system but perhaps small changes in either direction (dominance or diversity) may appear with nutrient changes in the North Sea. From the study one could accept an increased load (during pre-‐spring bloom) to increase the likely hood of green algae blooms.

34
Care must be taken when dealing with management of nutrients as nitrogen and phosphorous are linked meaning that changes in one may effect the other such as in the Baltic Sea where depletions of nitrogen could stimulate nitrogen fixing cyanobacteria causing an increased risk of blooms (Ekholm 2008). Alternatively this reduced nitrogen load could decrease phytoplankton biomass therefore contributing towards the release of phosphorous from sediments therefore lowering the chance of cyanobacteria blooms (Ekholm, 2008) therefore shifting toward diatom dominated waters.
5.3 Limitations With controlled environments however come disadvantages when applying to the field, as stable states are rare to come across due to intermittent nutrient fluxes favoring larger diatoms than constant supply (Litchman et al. 2009) and constant changing physical environments such as temperature dependence which has been shown to have an effect on species dominance (Tilman et al 1986). Perhaps the temperature of the chemostats during this experiment altered the original composition as different seasons would (Anderson et al., 2002 and Potapova 2002). Predation is also an important factor where in this experiment dinoflagellates were eliminated by week two allowing other species to compete where otherwise not (Anderson et al., 2002 and Elser et al. 1988). Research conducted on the role of grazers (zooplankton) also showed interesting findings. It has been suggested that grazers alter the nutrient limitation patters and therefore communities receiving their nutrients through grazers are either to become nitrogen or phosphorous limited but not both (Sterner 1990). The study used pre spring bloom inoculum where abundance was very low at the start giving no prior nutrient limitation or adaptive species a head start. In the future comparing mid/post spring bloom inoculum with the results of the present study would prove beneficial. Would the diversity of mid/post end states prove a similar diversity (in respect to number and similarity of diversity) and would they show similar dominance in regards to species?

35
5.4 conclusion To answer the questions of this paper ‘will changes in N:P ratio impact the phytoplankton community of the North Sea?’ from the results it can be said that changes in nutrient ratio and load have the potential to change phytoplankton composition. The maximum degree to which these change in composition can be predicted only lies within the different groups of phytoplankton (diatom, dinoflagellate, green algae, cyanobacteria etc.).
VI. Reference list Anderson D. M, Glibert P. M ad Burkholder J. M (2002) ‘Harmful alfal blooms and eutrophication: nutrient sources, composition, and consequences’ Estuaties. 25 (4b) : 704-‐726 Beaugrand G, Brander K.M, Lindley J. A, Souissi S and Reid P. C (2003) ‘Plankton effect on cod recruitment in the North Sea’ Nature. 426 : 661-‐664 Brauer V. S, Stomp M, Huisman J (2012) ‘The nutrient-‐load hypothesis: Patterns of resource limitation and community structure driven by competition for nutrients and light’ The American naturalist. 127 (6) : 179-‐740 Bulgakov N. G and Levich A. (undated) ‘The nitrogen:phosphorus ratio as a factor regulating phytoplankton community structure’ [online] http://www.chronos.msu.ru/EREPORTS/levich_the_nitrogen/levich_the_nitrogen.htm Cadee G. C (1986) ‘Recurrent and changing seasonal patterns in phytoplankton of the westernmost inlet of the Dutch Wadden Sea from 1969 to 1985’ Marine biology. 93 : 281-‐289

36
Cloern J. E (2001) ‘Our evolving conceptual model of the coastal eutrophication problem’ Marine ecology progress series. 210 : 223-‐253 Dow A. R, Gilliand J. L, Raymond R. A and Shiring J. J (2006) ‘Competition between Chlorella sp. and Chlorococcum sp.: a study in nutrient limitation’ Journal of ecological research. 8 : 20-‐25 Downing J. A, Watson S. B and McCauley E (2001) ‘Predicting cyanobacteria dominance in lakes’ Candaian journal of fisheries and aquatic sciences. 58 (10) : 1905-‐1908 Ecomare (undated) http://www.ecomare.nl/en/encyclopedia/natural-‐environment/matter-‐and-‐materials/nutrients/phosphoric-‐compounds/ [accessed on 3/3/2014] Ekholm (2008) [online] http://www.cost869.alterra.nl/FS/FS_NPratio.pdf [accessed on 19/12/2013] Elser J. J, Bracken M. E. S, Cleland E. E, Gruner D. S, Harpole W. S, Hillebrand H, Ngai J. T, Seabloom E. W, Shurin J. B and Smith J. E (2007) ‘Global analysis of nitrogen and phosphorous limitation of primary producers in freshwater, marine and terrestrial ecostystems’ Ecology letters. 10 : 1135-‐1142 Elser J. J, Elser M. M, sMackay N and Carpenter S. R (1988) ‘Zooplankton-‐mediated transitions between N-‐ and P-‐limited algal growth’ Limnology and oceanography. 33 (1) : 1-‐14 Fisher T. R, Carlson P. R and Barber R. T (1982) ‘Carbon and nitrogen primary productivity in three North Carolina estuaries’ Estuarine, coastal and shelf science. 15 (6) : 621-‐644 Foster R. A, Kuypers M. M. M, Vagner T, Paerl R. W, Musat N and Zehr J. P (2011) ‘Nitrogen fixation and transfer I nopen ocean diatom-‐cyanobacterial symbioses’ Multidisciplinary journal of microbial ecology. 5 (9) : 1484-‐1493

37
Fukuyo Y, Imai I, Kodama M and Tamai K (undated) [online] http://www.pices.int/publications/scientific_reports/report23/HAB_Japan.pdf Groove J. P (2000) ‘Resource competition and community structure in aquatic micro-‐organisms: experimental studies of algae and bacteria along a gradient of organic carbon to inorganic phosphorous supply’ Journal of plankton research. 22 98) : 1591-‐1610 Graneli E and Sundback K (1985) ‘The response of planktonic and microbenthi algal assemblages to nutrient enrichment in shallow coastal waters, southwest Sweden’ Journal of experimental marine biology and ecology. 85 (3) : 253-‐268 Hecky R. E and Kilham P (1988) ‘Nutrient limitation on phytoplankton freshwater and marine environments: a review of recent evidence on the effects of enrichment’ Limnology and oceangrapthy. 33 : 786-‐822 Henriksen P, Riemann B, Kaas H, Sorensen H. M and Sorensen H. L (2002) ‘Effects of nutreitn-‐limitation and irradiance on marine phytoplankton pigments’ Journal of plankton research. 24 (9) : 835-‐858 Howarth R. W and Marino R (2006) ‘Nitrogen as the limiting nutrient for eutrophication in coastal marine ecosystems: evolving views over three decades’ Limnology and oceanography. 51 (1 pt.2) : 364-‐376 Hu H, Zhang J and Chen W (2011) ‘Competition of bloom-‐forming marine phytoplankton at low nutrient concentrations’ Journal of environmental science. 23 (4) : 656-‐663 Huisman J, Sharples J, Stroom J. M, Visser P. M, Kardinaal W. E. A, Verspagen J. M. H and Sommeijer B (2004) ‘Changes in turbulent mixing shift competition for light between phytoplankton species’ ecological society of America. 85 (11) : 2960-‐2970 Huisman J, Matthijs H. C. P, Visser P. M, Balke H, Signon C. A. M, Passarge J, Weissing F. J and Mur L. R (2002) ‘Principles of the light-‐limited chemostat: theory and ecological application’ Antonie can leeuwenhoek. 81 : 117-‐13

38
Huisman J and Weissing F. J (1995) ‘Competition for nutrients and ight in a mixed water column: a theoretical analysis’ The ecological society of america. 72 (2) : 507-‐520 Huisman J and Weissing F. J (1994) ‘Light-‐limited growth and competition for light in well-‐mixed aquatic environments: an elementary model’ American naturalist. 146 (4) : 536-‐564 Imai I, Yamagughi M and Hori Y (2006) ‘Eutrophication and occurrences of harmful algal blooms in the Seto Inland Sea, Japan’ Plankton benthos research. 1 (2) : 71-‐84 Irwin A. J, Finkel Z. V, Schofield O. M and Falkowski P. G (2006) ‘Scaling-‐up from nutrient physiology to the size-‐structre of phytiplaknton cimmunties’ Journal of plankton research. 28 (5) : 459-‐471 Kilham S. S (1986) ‘Dynamics of lake Michigan natural phytoplankton communities in continuous cultures along a Si:P loading gradtient’ Candaian journal of fisheries and aquatic science. 43 (2) : 351-‐360 Litchman E, Edwards K. F, Klausmeier C. A and Thomas M. K (2012) ‘Phytoplankton niches, traits and eco-‐evolutionary responses to global environmental changes’ Marnie evology progress series. 470 : 235-‐248 Litchman E, Klausmeier C. A and Yoshiyama K (2009) ‘Contrasting size evolution in marine and freshwater diatoms’ Proceedings of the national academy of science. 106 (8) : 2665-‐2670 Litchman E, Klausmeier C. A, Schofield O. M and Falkowski P. G (2007) ‘The role of functional traits and trade-‐offs in structuring phytoplankton communities: scaling from cellular to ecosystem level’ Ecology letters. 10 (2) : 1170-‐1181M Maranon E, Cermeno P, Lopez-‐Sandoval D. C, Todriguesz-‐Ramos T, Sobrino C, Huete-‐Ortega M, Blanco J. M and Rodriguez J (2013) ‘Unimodal size scaling of phytoplankton growth and the size dependence of nutrient uptake and use’ Ecology letters. 16 (3) : 371-‐379

39
Menzel D. W and Corwin N (1965) ‘The measurement of total phosphorous in seawater based on the liberation of organically bound fraction by persulfate oxidation’ Limnology and oceanography. 10 : 280 -‐ 282 Michard M, Aleya L and Verneaux J (1996) ‘Mass occurrence of cyanobacteria Microcystis aeruginosa in the hypertrophic Villerest reservoir (Roanne, France): usefulness of biyearly examination of N/P and P/C couplings’ Archieve for hydrobiology. 135 (3) : 337-‐359 Miller T. E, Burns J. H, Munguia P, Walters E. L, Kneitel J.M, Richards P.M, Mouquet N and Buckley H. L (2005) ‘A critical review of twenty years’ use of the resource-‐ratio theory’ The American naturalist. 126 (4) : 439-‐448 Navarro J. N and Lobbam C. S (2009) ‘Freshwater and marine diatoms from the western pacific islands of Yap and Guan, with notes on some diatoms in damselfish territories’ Diatom research. 24 (1) : 123-‐157 NWO (undated) [online] http://www.nwo.nl/en/research-‐and-‐results/research-‐projects/82/2300158382.html [accessed on 19/12/2013] Passarge J, Hol S, Escher M and Huisman J (2006) ‘Competition for nutrients and light: stable coexcistene, alternative stable states or competitive exclusion?’ Ecologial monographs. 76 (1) : 57-‐72 Philippart C. J. M, Beukema J. J, Cadee G C, Dekker.R, Goedhart P. W, Van Iperen J. M, Leopold M. F and Herman P. M. J (2007) ‘Impacts of nutrient reduction on coastal communities’ Ecosystems. 10 : 95-‐118 Philippart C. J. M, Cadee G. C, Van Raaphorst W and Riegman R (2000) ‘Long term phytoplankton nutrient interactions in shallow coastal sea: algal community structure, nutrient budgets and denitrification potential’ Limnology and oceanography. 45 : 131-‐144 Potapova M. G and Charles D. F (2002) ‘Benthic diatoms in USA rivers: distribution along spatial and environemtnal gradients’ Jounal of biogeography. 29 (2) : 167-‐187

40
Prins T. C, Desmit X and Baretta-‐Bekker J. G (2012) ‘Phytoplankton composition in Dutch coastal waters responds to changes in riverine nutrient loads’ Journal of sea research. 73 : 49-‐62 Radach G, Berg J, Hagmeier E (1990) ‘Long term changes of the annual cycles of meterologial, hydrigraphc, nutrient and phytoplankton time series at Helgoland and at LV eLEBE 1 in the German Bight’ CONT SHELF RESS. 10 : 305-‐328 Roelke D. L, Eldridge P. M and Cifuenter L. A (1999) ‘A model of phytoplankton completion for limiting and nonlimiting mutrients: Implications for development of estuarine and nearshore management schemes’ Estuaries. 22 : 92-‐104 Ryther H. J and Dunstan W. M (1971) ‘Nitrogen, phosphorous and eutrophication in the coastal marine environment’ science. 171 (3975) : 1008-‐1113 Schindler D. W (2006) ‘Recent advances in understanding and management of europhication’ limnology and oceanography. 51 : 356-‐363 Skogen M. D, Soiland H, Svendsen E (2004) ‘Effects of changing nutrient loads to the North Sea’ Journal of marine ecosystems. 46 : 23-‐38 Smetacek V (1999) ‘Diatoms and the coean carbon cycle’ Protist. 150 (1) : 25-‐32 Stephen-‐Bishop S, Emmanuele K. A and Joder J. A (1984) ‘Nutrient limitation on phytoplankton growth in Georgia nearshore waters’ Estuaries 7 (4) : 506-‐512 Sterner R. W (1990) ‘The ratio of nitrogen to phosphorous resupplied by herbivores: zooplankton and the algal competitive arena’ The American naturalist. 136 (2) : 209-‐229 Stolte W, McCollin T, Noordeloos A.A.M and Riegman R (1994) ‘Effect of nitrogen source on the size distribution within marine phytoplankton populations’ Experimental marine biology and ecology 184 (1) : 83-‐97

41
Tilman D, Kiesling R, Sterner R, Kilham S. S and Johnson F. A (1986) ‘Green, bluegreen and diatom algae: taxonomic differences in competitive ability for phosphorous, silicon and nitrogen’ archieve of hydrobiology. 106 (4) : 473-‐485 Tilman D (1985) ‘the resource-‐ratio hypothesis of plant succession’ The American naturalist. 125 (6) : 827-‐852 Tilman D (1982) ‘Resource competition and community structure’ Princeton, New Jersey. Princeton university press Trommer G, Leynaert A, Klein C, Naegelen A and Beker B (2013) ‘Phytiplankton phosphorous limitation in North Atlantic coastal ecosystem not predicted by nutrient load’ Journal of plankton research. 35 (6) : 1207-‐1219 Tubay J. M, Ito H, Uehara T, Kakishima S, Morita S, Togashi T, Tainaka K, Niraula M. P, Casareto B. E, Suzuki Y and Yoshimura J (2013) ‘The paradox of enrichment in phytoplankton by incudced competitive interactions’ Scientific reports. 3 no. 2835 Van Raaphorst W and De Jonge V. N (2004) ‘Reconstruction of the total N and P inputs from the Ijsselmeer into the western Wadden Sea between 1935-‐1998’ Journal of sea research. 51 : 109-‐131 Van Ruth P. D, Qin J. G and Branford A. J (2012) ‘Size dependant competition in centric diatoms as a function of nitrogen and silicon availablility’ Journal of marine science. 2 : 33-‐42 Vrede T, Ballantyne A, Mille-‐Lindblom C, Algesten Gm Gudasz C, Lindahl A and Brunberg A. K (2009) ‘Effects of N:P loading ratios on phytoplankton community composition, primary production and N fixation in eutrophic lake’ .Freshwater biology. 54 (2) : 331-‐344 Wang C, Yu X, Lv H and Yan g J (2013) ‘Nitrogen and phosphorous removal from municipal wastewater by the green algae Chlorella sp.’ Journal of environmental biology. 34 : 421-‐425 Wang P, Shen H and Zie P (2012) ‘Can hydrodynamics change phosphorous strategies of diatoms? – Nitrate levels and diatom blooms in lotic and lentic ecosystems’ Microbial ecology. 63 : 369-‐382