Overview of Metabolic Pathways
14
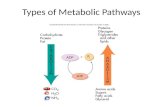
Overview of Metabolic Pathways: Catabolism Biological cells have a daunting task. They must carry out 1000s of different chemical reactions required to carry out cell function. These reactions can include opposing goals such as energy production and energy storage, macromolecule degradation and synthesis, and breakdown and synthesis of small molecules. All of these reactions are catalyzed by proteins and RNAs enzymes whose activities must be regulated, again through chemical reactions, to avoid a futile and energy wasting scenario of having opposing pathways functioning simultaneously in a cell. Metabolism can be divided into two main parts, catabolism, the degradation of molecules, usually to produce energy or small molecules useful for cell function, and anabolism, the synthesis of larger biomolecules from small precursors. CATABOLISM Catabolic reactions involve the breakdown of carbohydrates, lipids, proteins, and nucleic acids to produce smaller molecules and biological energy in the form of heat or small thermodynamically reactive molecules like ATP whose further degradation can drive endergonic process such as biosynthesis. Our whole world is reliant on the oxidation of organic hydrocarbons to water and carbon dioxide to produce energy (at the expense of releasing a potent greenhouse gas, CO 2 ). In the biological world, reduced molecules like fatty acids and partially oxidized molecules such as glucose polymers (glycogen, starch), as well as simple sugars, can be partially or fully oxidized to ultimately produce CO 2 as well. Energy released from oxidative reactions is used to produce molecules like ATP as well as heat. Oxidative pathways include glycolysis, the tricarboxylic acid cycle (aka Kreb's cycle) and mitochondrial oxidative phosphorylation/electron transport. To fully oxidize carbon in glucose and fatty acids to carbon dioxide requires splitting C-C bonds and the availability of series of oxidizing agents that can perform controlled, step-wise oxidation reactions, analogous to the sequential oxidation of methane, CH 4 to methanol (CH 3 OH), formaldehyde (CH 2 O) and carbon dixoxide. Glycolysis: This most primitive of metabolic pathways is found in perhaps all organisms. In glycolysis, glucose (C 6 H 12 O 6 ), a 6C
description
Metabolism has two processes, catabolism and anabolism. RNA plays a vital role in Anabolism.
Transcript of Overview of Metabolic Pathways
Overview of Metabolic Pathways: CatabolismBiological cells have a daunting task. They must carry out 1000s of different chemical reactions required to carry out cell function. These reactions can include opposing goals such as energy production and energy storage, macromolecule degradation and synthesis, and breakdown and synthesis of small molecules. All of these reactions are catalyzed by proteins and RNAs enzymes whose activities must be regulated, again through chemical reactions, to avoid a futile and energy wasting scenario of having opposing pathways functioning simultaneously in a cell.Metabolism can be divided into two main parts, catabolism, the degradation of molecules, usually to produce energy or small molecules useful for cell function, and anabolism, the synthesis of larger biomolecules from small precursors.
CATABOLISMCatabolic reactions involve the breakdown of carbohydrates, lipids, proteins, and nucleic acids to produce smaller molecules and biological energy in the form of heat or small thermodynamically reactive molecules like ATP whose further degradation can drive endergonic process such as biosynthesis. Our whole world is reliant on the oxidation of organic hydrocarbons to water and carbon dioxide to produce energy (at the expense of releasing a potent greenhouse gas, CO2). In the biological world, reduced molecules like fatty acids and partially oxidized molecules such as glucose polymers (glycogen, starch), as well as simple sugars, can be partially or fully oxidized to ultimately produce CO2as well. Energy released from oxidative reactions is used to produce molecules like ATP as well as heat. Oxidative pathways include glycolysis, the tricarboxylic acid cycle (aka Kreb's cycle) and mitochondrial oxidative phosphorylation/electron transport. To fully oxidize carbon in glucose and fatty acids to carbon dioxide requires splitting C-C bonds and the availability of series of oxidizing agents that can perform controlled, step-wise oxidation reactions, analogous to the sequential oxidation of methane, CH4to methanol (CH3OH), formaldehyde (CH2O) and carbon dixoxide. Glycolysis: This most primitive of metabolic pathways is found in perhaps all organisms. In glycolysis, glucose (C6H12O6), a 6C molecule, is split (or lysed) into two, 3C carbon molecules, glyceraldehyde-3-phosphate, which are then partially oxidized underanaerobicconditions (without O2) to form two molecules of pyruvate (CH3COCO2-). Instead of the very strong oxidizing agent, O2, a weaker one, NAD+is used, which is reduced in the process to form NADH. Since none of the carbon atoms is oxidized to the state of CO2, little energy is released compared to the complete oxidation to CO2. This pathway comes to a screeching halt if all cellular NAD+is converted to NADH as NAD+is not replenished by the simple act of breathing as is the case with O2in aerobic oxidation. To prevent the depletion of NAD+from inhibiting the cycle and to allow the cycle to continue under anaerobic conditions, excess NADH is reconverted to NAD+when the other product of glycolysis, pyruvate is converted to lactate by the enzyme lactate dehydrogenase. Glycolysis occurs in the cytoplasm of the cell.Figure: Summary of Glycolysis
Tricarboxylic Acid (Kreb's) Cycle: The TCA cycle is an aerobic pathway which takes place in an intracellular organelle called the mitochondria. It takes pyruvate, the incompletely oxidized product from glycolysis, and finishes the job of oxidizing the 3C atoms all the way to CO2. First the pyruvate moves into the mitochondria where is is oxidized to the 2C molecule acetylCoA with the release of one CO2by the enzyme pyruvate dehydrogenase. The acetyl-CoA then enters the TCA cycle where two more CO2are released. As in glycolysis, C-C bonds are cleaved and C is oxidized by NAD+and another related oxidizing agent, FAD. What is very different about this pathway is that instead of being a series of linear, sequential reactions with one reactant (glucose) and one product (two pryuvates), it is a cyclic pathway. This has significant consequences since if any of the reactants within the pathways becomes depleted, the whole cyclic pathway can slow down and stop. To see how this happens consider the molecule oxaloacetate (OAA) which condenses with acetyl-CoA to form citrate (see diagram below). In this reaction, one OAA is consumed. However, when the cycle returns, one malate is converted to OAA so there is no net loss of OAA, unless OAA is pulled out of the TCA cycle for other reactions, which happens.Figure: Pyruvate Dehydrogenase (mitochondrial) and the TCA Cycle
Mitochondrial Oxidative Phosphorylation/Electron Transport: The TCA cycle accomplishes what glycolysis didn't, that is the cleavage of all C-C bonds in glucose (in the form of pyruvate and acetyl-CoA, and the complete oxidation of all C atoms to CO2. Yet two problem remains. The pool of oxidizing molecules, NAD+and FAD get converted to their reduced forms, NADH and FADH2. Unless NAD+and FAD are regenerated, as was the case in anaerobic conditions when pyruvate gets converted to lacate, the pathway would again come to a grinding halt. In addition, not much ATP is made in the cycle (in the form of a related molecule GTP). Both these problems are resolved as the resulting NADH and FADH2formed are reoxidized by mitochondrial membrane enzyme complexes which pass electrons from the oxidized NADH and FADH2to increasingly potent oxidizing agents until they are accepted by the powerful oxidant O2,which is converted reduced to water. The net oxidation of NADH and FADH2by dioxygen is greatly exergonic, and the energy released by the process drives the synthesis of ATP from ADP and Pi by an mitochondrial enzyme complex, the F0F1ATPase.Figure: Mitochondrial Electron Transport/Oxidative Phosphorylation
Feeder Pathways: Other catabolic pathways produce products that can enter glycolysis or the TCA cycle. Two examples are given below. Complex carbohydrates: In mammals, the major carbohydrate storage molecule is glycogen, a polymer of glucose linkeda1-4 witha1-6 branches. The terminal acetal linkages in this highly branched polymer is cleaved sequentially at the ends not through hydrolysis but through phosphorolysis to produce lots of glucose-1-phosphate which can enter glycolysis.
Lipids: Lipids are stored mostly as triacylglycerides in fat cells (adipocytes). When needed for energy, fatty acids are hydrolyzed from the glycerol backbone of the triacylglyceride, and send into cells where they broken down in an oxidative process to form acetyl-CoA with the concomitant production of lots of NADH and FADH2. These can then enter the mitochondrial oxidative phosphorylation/electrons transport system, which produces, under aerobic conditions, lots of ATP.
Proteins: When intracellular proteins get degraded, they from individual amino acids. The amine N is lost as it enters the urea cycle. The rest of some amino acid structures can be ultimately converted to acetyl-CoA or keto acids (like alpha-ketoglutarate-a-KG) that are TCA intermediate. These amino acids are called ketogenic. Alternatively, some amino acids, after deamination, are coveted to pyruvate which can either enter the TCA cycle or in the liver be used to synthesize glucose in an anabolic process.These amino acids are called glucogenic. Chemical reactions such as these can be used to replenish intermediates in the TCA cycle which can become depleted as they are withdraw for other reactions.
Now on to anabolic reactions.
Overview of Metabolic Pathways: Anabolism
Anabolic reactions are those that lead to the synthesis of biomolecules. In contrast to the catabolic reactions just discussed (glycolysis, TCA cycle and electron transport/oxidative phosphorylation) which lead to the oxidative degradation of carbohydrates and fatty acids and energy release, anabolic reactions lead to the synthesis of more complex biomolecules including biopolymers (glycogen, proteins, nucleic acids) and complex lipids. Many biosynthetic reactions, including those for fatty acid synthesis, are reductive and hence require reducing agents. Reductive biosynthesis and complex polymer formation require energy input, usually in the form of ATP whose exergonic cleavage is coupled to endergonic biosynthesis.Cells have evolved interesting mechanism so as not to have oxidative degradation reactions (which release energy) proceed at the same time and in the same cell as reductive biosynthesis (which requires energy input). Consider this scenario. You dive into a liver cell and find palmitic acid, a 16C fatty acid. From where did it come? Was it just synthesized by the liver cell or did it just enter the cell from a distant location such as adipocytes (fat cells). Should it be oxidized, which should happen if there is a demand for energy production by the cell, or should the liver cell export it, perhaps to adipocytes, which might happen if there is an excess of energy storage molecules? Cells have devised many ways to distinguish these opposing needs. One is by using a slightly different pool of redox reagents for anabolic and catabolic reactions. Oxidative degradation reactions typically use the redox pair NAD+/NADH (or FAD/FADH2) while reductive biosynthesis often uses phosphorylated variants of NAD+, NADP+/NADPH. In addition, cells often carry out competing reactions in different cellular compartments. Fatty acid oxidation of our example molecule (palmitic acid) occurs in the mitochondrial matrix, while reductive fatty acid synthesis occurs in the cytoplasm of the cell. Fatty acids entering the cell destined for oxidative degradation are transported into the mitochondria by the carnitine transport system. This transport system is inhibited under conditions when fatty acid synthesis is favored. We will discuss the regulation of metabolic pathways in a subsequent section. One of the main methods, as we will see, is to activate or inhibit key enzymes in the pathways under a given set of cellular conditions. The key enzyme in fatty acid synthesis, acetyl-CoA carboxylase, is inhibited when cellular conditions require fatty acid oxidation.The following examples give short descriptions of anabolic pathways. Compare them to the catabolic pathways from the previous section. Glucose synthesis, better known as Gluconeogenesis:In glycolysis, glucose (C6H12O6), a 6C molecule, is converted to two, 3C molecules (pyruvate) in an oxidative process that requires NAD+and makes two net ATP molecules. In a few organs, most predominately in the liver, the reverse pathway can take place. The liver does this to provide glucose to the brain when the body is deficient in circulating glucose, for example, under fasting and starving conditions. (The liver under these conditions can get its energy from oxidation of fatty acids). The reactions in gluconeogenesis are the same reactions in glycolysis but run in reverse, with the exception of three glycolytic steps which are essentially irreversible. These three steps have bypass enzymes in the gluconeogenesis pathway. Although the synthesis of glucose is a reductive pathway, it uses NADH instead of NADPH as the redundant as the same enzyme used in glycolysis is simply run in reverse. Gluconeogenesis, which also occurs in the cortex of the kidney, is more than just a simple reversal of glycolysis, however. It can be thought of as the net synthesis of glucose from non-carbohydrate precursors. Pyruvate, as seen in the section on catabolism, can be formed from protein degradation to glucogenic amino acids which can be converted to pyruvate. It can also be formed from triacylglycerides from the 3C molecule glycerol formed and released from adipocytes after hydrolysis of three fatty acids from triacylglycerides. However, in humans, glucose can not be made innetfashion from fatty acids. Fatty acids can be converted to acetyl-CoA by fatty acid oxidation. The resulting acetyl-CoA cannotform pyruvate since the enzyme that catalyzes the formation for acetyl-CoA from pyruvate, pyruvate dehydrogenase, is irreversible and there is no bypass reaction known. The acetyl-CoA can enter the TCA cycle but since the pathway is cyclic and proceeds in one direction, it can not form in net fashion oxaloacetate. Although oxaloacetate can be remove from the TCA cycle and be use to form phosphoenolpyuvate, a glycolytic intermediate, one acetyl-CoA condenses with one oxaloacetate to form citrate which leads back to one oxaloacetate. Hence fatty acids can not be converted to glucose and other sugars in a net fashion.Figure: Gluconeogenesis
Pentose Phosphate Shunt: This two-part pathway doesn't appear to start as a reductive biosynthetic pathway as the first part is the oxidative conversion of a glycolytic intermediate, glucose-6-phosphate, to ribulose-5-phosphate. The next, nonoxidative branch leads to the formation of ribose-5-phosphate, a key biosynthetic intermediate in nucleic acid synthesis as well as erthyrose-4-phosphate used for biosynthesis of aromatic amino acids . The oxidative branch is important in reductive biosynthesis as it is a major source of the reductant NADPH used in biosynthetic reactions.
Fatty acid and isoprenoid/sterol biosynthesis: Acetyl-CoA is the source of carbon atoms for the synthesis of more complex lipids such as fatty acids, isoprenoids, and sterols. When energy needs in a cell are not high, citrate, the condensation product of oxaloacetate and acetyl-CoA in the TCA cycle, builds up in the mitochondrial matrix. It is then transported by the citrate transporter (an inner mitochondrial membrane protein) to the cytoplasm, where it is cleaved back to oxaloacetate and acetyl-CoA by the cytoplasmic enzyme citrate lyase. The oxaloacetate is returned to the mitochondria by conversion first to malate (reduction reaction using NADH), which can move back into the mitochondria through the malate transporter, or further conversion to pyruate, using the cytosolic malic enzyme, which uses NADP+to oxidize malate to pyruvate which then enters the mitochondria. The acetyl-CoA formed in the cytoplasm can then be used in reductive biosynthesis using NADPH as the reductant to form fatty acids, isoprenoids, and sterols. The NADPH for the reduction comes from the oxidative branch of the pentose phosphate pathway and from the reaction catalyzed by malic enzyme. The liver cells can still run the glycolytic pathway as the NADH/NAD+ratio is low in the cytoplasm while NADPH/NADP+ratio is high.
Now its time to see how the various pathways fit together to form an integrated set of pathways. There is a very large number of metabolic pathways. In humans, the most important metabolic pathways are: glycolysis- glucose oxidation in order to obtain ATP citric acid cycle (Krebs' cycle)- acetyl-CoA oxidation in order to obtain GTP and valuable intermediates. oxidative phosphorylation- disposal of the electrons released by glycolysis and citric acid cycle. Much of the energy released in this process can be stored as ATP. pentose phosphate pathway- synthesis of pentoses and release of the reducing power needed for anabolic reactions. urea cycle- disposal of NH4+in less toxic forms fatty acid -oxidation- fatty acids breakdown into acetyl-CoA, to be used by the Krebs' cycle. gluconeogenesis- glucose synthesis from smaller percursors, to be used by the brain.Metabolic profiles of key tissuesBrainUsually neurons use only glucose as energy source. Since the brain stores only a very small amount of glycogen, it needs a steady supply of glucose. During long fasts, it becomes able to oxidize ketone bodies.LiverThe maintenance of a fairly steady concentration of glucose in the blood is one of the liver's main functions. This is accomplished throughgluconeogenesisand glycogensynthesisanddegradation. It synthesizes ketone bodies when acetyl-CoA is plenty. It is also the site of urea synthesis.Adipose tissueIt synthesizes fatty acids and stores them as triacylglycerols.Glucagonactivates a hormone-sensitive lipase, which hydrolizes triacylglycerols yielding glycerol and fatty acids. These are then released into the bloodstream in lipoproteins.MuscleMuscles use glucose, fatty acids, ketone bodies and aminoacids as energy source. It also contains a reserve of creatine-phosphate, a compound with a high phosphate-transfer potential that is able to phosphorilate ADP to ATP, thereby producing energy without using glucose. The amount of creatine in the muscle is enough to sustain about 3-4 s of exertion. After this period, the muscle uses glycolysis, first anaerobically (since it is much faster than the citric acid cycle), and later (when the increased acidity slows phosphofrutokinase enough for the citric acid cycle to become non-rate-limiting) in aerobic conditions.KidneyIt can perform gluconeogenesis and release glucose into the bloodstream. It is also responsible for the excretion of urea, electrolytes, etc.Metabolic acidosismay be increased by the action of the urea cycle, since urea synthesis (which takes place in the liver) uses HCO3-, thereby further lowering blood pH. Under these circunstances, nitrogen may be eliminated by the joint action of kidney and liver: excess nitrogen is first incorporated in glutamine byglutamine synthetase. Kidney glutaminase then cleaves glutamine in glutamate e NH3, which the kidney immediately excretes. This process allows nitrogen excretion without affecting blood bicarbonate levels.Hormone controlHormone control is mainly effected through the action of two hormones synthesized by the pancreas:insulinandglucagon. Insulin is released by the pancreas when blood glucose levels are high, i.e., after a meal. Insulin stimulates glucose uptake by the muscle, glycogen synthesis, and triacylglyceride synthesis by the adipose tissue. It inhibits gluconeogenesis and glycogen degradation. Glucagon is released by pancreas when blood glucose levels drop too much. Its effects are opposite those of insulin: in liver, glucagon stimulates glycogen degradation and the absorption of gluconeogenic aminoacids. It inhibits glycogen synthesis and promotes the release of fatty acids by adipose tissue.
R.A #9Overview of Metabolism
Submitted by: Nealeth B. NanquilSubmitted to: Professor Cyda O. Meimban