
Overexpression of a Novel Activator of PAK4, the CDK5...
11
Molecular and Cellular Pathobiology Overexpression of a Novel Activator of PAK4, the CDK5 Kinase–Associated Protein CDK5RAP3, Promotes Hepatocellular Carcinoma Metastasis Grace Wing-Yan Mak 1 , Mandy Man-Lok Chan 1 , Veronica Yee-Law Leong 1 , Joyce Man-Fong Lee 2,3 , Tai-On Yau 2 , Irene Oi-Lin Ng 2,3 , and Yick-Pang Ching 1,3 Abstract The CDK5 kinase regulatory subunit-associated protein 3 (CDK5RAP3 or C53/LZAP) regulates apoptosis induced by genotoxic stress. Although CDK5RAP3 has been implicated in cancer progression, its exact role in carcinogenesis is not well established. In this article, we report that CDK5RAP3 has an important prometastatic function in hepatocarcinogenesis. An examination of human hepatocellular carcinoma (HCC) samples revealed at least twofold overexpression of CDK5RAP3 transcripts in 58% (39/67) of HCC specimens when compared with corresponding nontumorous livers. CDK5RAP3 overexpression was associated with more aggressive biological behavior. In HCC cell lines, stable overexpression of CDK5RAP3 promoted, and small interfering RNA–mediated knockdown inhibited, tumorigenic activity and metastatic potential. We found that over- expression of CDK5RAP3 and p21-activated protein kinase 4 (PAK4) correlated in human HCCs, and that CDK5RAP3 was a novel binding partner of PAK4, and this binding enhanced PAK4 activity. siRNA-mediated knockdown of PAK4 in CDK5RAP3-expressing HCC cells reversed the enhanced cell invasiveness mediated by CDK5RAP3 overexpression, implying that PAK4 is essential for CDK5RAP3 function. Taken together, our findings reveal that CDK5RAP3 is widely overexpressed in HCC and that overexpression of CDK5RAP3 promotes HCC metastasis through PAK4 activation. Cancer Res; 71(8); 2949–58. Ó2011 AACR. Introduction The CDK5 kinase regulatory subunit-associated protein 3 (CDK5RAP3, also called C53/LZAP) was first identified as a binding partner of cyclin-dependent kinase 5 activator, p35 nck5a , in yeast 2-hybrid screening (1). Northern analysis indicated that CDK5RAP3 is widely expressed in human tissues, and the expression level is relatively constant in the heart, brain, skeletal muscle, placenta, lung, liver, kidney, and pancreas (2). Overexpression of CDK5RAP3 has been shown to sensitize cells to apoptosis induced by genotoxic stress (3). CDK5RAP3 can interact with a well-known tumor suppressor, namely, the alternate reading frame (ARF; p14 ARF ), by which it stabilizes and promotes the transcription activity of p53 (4). More recently, CDK5RAP3 has been found to be underex- pressed in head and neck cancers, and forced expression of CDK5RAP3 can negatively regulate NF-kB activity (5), which suggests that CDK5RAP3 may function as a tumor suppressor. On the contrary, stable overexpression of the CDK5RAP3 isoform has been shown to promote hepatocellular carcinoma (HCC) and cardiac cell proliferation (6, 7), which indicates, furthermore, that CDK5RAP3 may enhance cell growth. CDK5RAP3 is located at chromosome region 17q21.32, which has been reported to be amplified in HCC; however, the role of CDK5RAP3 in HCC has not been explored so far (8). In this study, we found that the expression of CDK5RAP3 was frequently upregulated in human HCCs at both transcript and protein levels. More importantly, we detected a remarkable enhancement of CDK5RAP3 expression in metastatic HCC. Although little information is available on how CDK5RAP3 regulates cancer metastasis, we found that CDK5RAP3 is a novel activator of p21-activated protein kinase 4 (PAK4) and activation of PAK4 can promote HCC cell migration. There- fore, we provided, in this study, a novel mechanism by which CDK5RAP3 contributes to the metastasis of HCC by activation of PAK4. Materials and Methods Cell culture Human hepatoma cell lines PLC/PRF/5 and HepG2 were purchased from the American Type Culture Collection. The authentication of these cell lines was ensured by the provider through cytogenetic analysis. No additional test was conducted specifically for this study. The human HCC cell line Authors' Affiliations: Departments of 1 Anatomy and 2 Pathology, Li Ka Shing Faculty of Medicine, and 3 State Key Laboratory for Liver Research, The University of Hong Kong, Hong Kong, China Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Corresponding Author: Yick-Pang Ching, Department of Anatomy, The University of Hong Kong, Room L1-43, Laboratory Block, Faculty of Medicine Building, Hong Kong. Phone: 852-28199434; Fax: 852- 28170857; E-mail: [email protected] doi: 10.1158/0008-5472.CAN-10-4046 Ó2011 American Association for Cancer Research. Cancer Research www.aacrjournals.org 2949 Research. on January 2, 2020. © 2011 American Association for Cancer cancerres.aacrjournals.org Downloaded from Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046
Transcript of Overexpression of a Novel Activator of PAK4, the CDK5...

Molecular and Cellular Pathobiology
Overexpression of a Novel Activator of PAK4, the CDK5Kinase–Associated Protein CDK5RAP3, PromotesHepatocellular Carcinoma Metastasis
Grace Wing-Yan Mak1, Mandy Man-Lok Chan1, Veronica Yee-Law Leong1, Joyce Man-Fong Lee2,3,Tai-On Yau2, Irene Oi-Lin Ng2,3, and Yick-Pang Ching1,3
AbstractThe CDK5 kinase regulatory subunit-associated protein 3 (CDK5RAP3 or C53/LZAP) regulates apoptosis
induced by genotoxic stress. Although CDK5RAP3 has been implicated in cancer progression, its exact role incarcinogenesis is not well established. In this article, we report that CDK5RAP3 has an important prometastaticfunction in hepatocarcinogenesis. An examination of human hepatocellular carcinoma (HCC) samples revealedat least twofold overexpression of CDK5RAP3 transcripts in 58% (39/67) of HCC specimens when comparedwith corresponding nontumorous livers. CDK5RAP3 overexpression was associated with more aggressivebiological behavior. In HCC cell lines, stable overexpression of CDK5RAP3 promoted, and small interferingRNA–mediated knockdown inhibited, tumorigenic activity and metastatic potential. We found that over-expression of CDK5RAP3 and p21-activated protein kinase 4 (PAK4) correlated in human HCCs, and thatCDK5RAP3 was a novel binding partner of PAK4, and this binding enhanced PAK4 activity. siRNA-mediatedknockdown of PAK4 in CDK5RAP3-expressing HCC cells reversed the enhanced cell invasiveness mediated byCDK5RAP3 overexpression, implying that PAK4 is essential for CDK5RAP3 function. Taken together, ourfindings reveal that CDK5RAP3 is widely overexpressed in HCC and that overexpression of CDK5RAP3 promotesHCC metastasis through PAK4 activation. Cancer Res; 71(8); 2949–58. �2011 AACR.
Introduction
The CDK5 kinase regulatory subunit-associated protein 3(CDK5RAP3, also called C53/LZAP) was first identified as abinding partner of cyclin-dependent kinase 5 activator,p35nck5a, in yeast 2-hybrid screening (1). Northern analysisindicated that CDK5RAP3 is widely expressed in humantissues, and the expression level is relatively constant in theheart, brain, skeletal muscle, placenta, lung, liver, kidney, andpancreas (2). Overexpression of CDK5RAP3 has been shown tosensitize cells to apoptosis induced by genotoxic stress (3).CDK5RAP3 can interact with a well-known tumor suppressor,namely, the alternate reading frame (ARF; p14ARF), by which itstabilizes and promotes the transcription activity of p53 (4).More recently, CDK5RAP3 has been found to be underex-pressed in head and neck cancers, and forced expression ofCDK5RAP3 can negatively regulate NF-kB activity (5), which
suggests that CDK5RAP3 may function as a tumor suppressor.On the contrary, stable overexpression of the CDK5RAP3isoform has been shown to promote hepatocellular carcinoma(HCC) and cardiac cell proliferation (6, 7), which indicates,furthermore, that CDK5RAP3 may enhance cell growth.
CDK5RAP3 is located at chromosome region 17q21.32,which has been reported to be amplified in HCC; however,the role of CDK5RAP3 in HCC has not been explored so far (8).In this study, we found that the expression of CDK5RAP3 wasfrequently upregulated in human HCCs at both transcript andprotein levels. More importantly, we detected a remarkableenhancement of CDK5RAP3 expression in metastatic HCC.Although little information is available on how CDK5RAP3regulates cancer metastasis, we found that CDK5RAP3 is anovel activator of p21-activated protein kinase 4 (PAK4) andactivation of PAK4 can promote HCC cell migration. There-fore, we provided, in this study, a novel mechanism by whichCDK5RAP3 contributes to the metastasis of HCC by activationof PAK4.
Materials and Methods
Cell cultureHuman hepatoma cell lines PLC/PRF/5 and HepG2 were
purchased from the American Type Culture Collection. Theauthentication of these cell lines was ensured by the providerthrough cytogenetic analysis. No additional testwas conductedspecifically for this study. The human HCC cell line
Authors' Affiliations: Departments of 1Anatomy and 2Pathology, Li KaShing Faculty of Medicine, and 3State Key Laboratory for Liver Research,The University of Hong Kong, Hong Kong, China
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Author: Yick-Pang Ching, Department of Anatomy, TheUniversity of Hong Kong, Room L1-43, Laboratory Block, Faculty ofMedicine Building, Hong Kong. Phone: 852-28199434; Fax: 852-28170857; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-10-4046
�2011 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 2949
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

SMMC-7721 was a gift from the Shanghai Institute of Biochem-istry and Cell Biology, Chinese Academy of Sciences. Cells weremaintained in Dulbecco's modified Eagle medium (DMEM)-high glucose (Life Technologies) supplementedwith 1 mmol/Lsodium pyruvate and 10% heat-inactivated FBS (JRH Bios-ciences). Cells were transfected with DNA constructs, usingLipofectamine 2000 (Invitrogen) according to the man-ufacturer's protocol. For constructing the stable clones, cellswere transfected with expression constructs (Myc-CDK5RAP3or shCDK5RAP3) and were selected with the correspond-ing antibiotic puromycin at 0.4 mg/mL (Sigma) or G418 at0.8 mg/mL (Calbiochem), 48 hours after transfection.
CDK5RAP3 antibodyRabbit anti-CDK5RAP3 polyclonal antibody was generated
using purified glutathione S-transferase (GST)-CDK5RAP3fusion protein as antigen (Genscript Company). Then, theantibody was affinity purified by (CNBr)-GST-CDK5RAP3 andGST column.
ImmunohistochemistryImmunohistochemical staining for CDK5RAP3 and phos-
pho-PAK4 (p-PAK4) was done as described previously (9).Purified anti-CDK5RAP3 antibody and anti-p-PAK4 (Ser474)antibody (Cell Signaling Technology) were used at 1:100 and1:50 dilutions, respectively. Scoring of the protein expressionwas assessed by an experienced pathologist.
Real-time quantitative reverse transcriptase PCRQuantitative PCR (qPCR) was done as described previously
(9). The sequence of TaqMan probes (Applied Biosystems) forCDK5RAP3 and PAK4 are 50-AGGAAAGATGGAGGACCAT-CAGCAC-30 and 50-GCGGCGCCGAGCCGATGAGTAACCC-30,respectively; cellular 18S rRNA was used as an internal control.
Colony formation assayCells were transiently transfected with plasmids expressing
CDK5RAP3 and shCDK5RAP3 and were selected with corre-sponding antibiotics for 2 weeks. The experiments were done3 times independently.
Cell proliferation, soft agar growth, cell migration, andinvasion assays
The procedure described by Leung and colleagues (10) andWong and colleagues (11) was adopted. For soft agar assay,colonies with diameter greater than 50 mm in 10 fields werecountedanddatawere shownas average numberof colonies perfield. The experiments were carried out 3 times independently.
Nude mouse xenograft assayCDK5RAP3 stable knockdown PLC/PRF/5 cells and control
cells (5 � 106) were administered by s.c. injection into the rightand left flanks of 6-week-old male nude mice, respectively. Thelarger (a) and smaller (b) diameters of a tumor were measuredweekly. Tumor volumewas estimated according to the formula:volume ¼ 1/2 � a � b2; and it was measured from week 2 toweek 5. After 5 weeks, the mice were sacrificed in accordancewith institutional regulations for animal experiments.
Purification of proteinGST fusion proteins were purified using reduced glu-
tathione (GSH)-sepharose beads by a procedure describedpreviously (1). His-tagged proteins were purified using Nickel-NTA agarose beads (Qiagen). Bacterial cells were lysed in alysis buffer containing 20 mmol/L Tris (pH 7.5), 500 mmol/LNaCl, 5 mmol/L imidazole, 1 mmol/L dithiothreitol (DTT),1 mmol/L phenylmethylsulfonyl fluoride, 1 mmol/L leupep-tin, and 10 mg/mL lysozyme. The protein was eluted with100 mmol/L imidazole in Tris buffer.
PAK4 kinase assayAn in vitro gel kinase assay was conducted as described
previously (12). GST-CDK5RAP3 and its mutants were incu-bated with His-PAK4 and GST-PAK4 peptide substrate [aminoacid (aa) sequence AARRLSVASAK, named PAK4tide, designedon the basis of the published result (13)] in PAK4 kinase buffer[50 mmol/L HEPES (pH 7.5), 5 mmol/L MgCl2, 100 mmol/LNaCl, and 1 mmol/L DTT] containing 10 mCi [g-32P]ATP. Forpeptide-kinase assay, GST and GST-CDK5RAP3 were incu-bated with His-PAK4 in the presence of 1 mmol/L PAK4tideand 10 mCi [g-32P]ATP at 30�C for 10 minutes (14). Peptideswith single mutation (AARRLAVASAK) were included as anegative control.
GST affinity pull-down assayGST and GST-CDK5RAP3 immobilized by GSH-sepharose
beads (GE Healthcare) were incubated with His-PAK4 for 2hours at 4�C, followed by washing 3 times with NETN buffer[50 mmol/L NaCl, 5 mmol/L EDTA, 50 mmol/L Tris (pH 8.0),1% NP40]. The bound proteins were then visualized byWestern blotting.
CoimmunoprecipitationHEK293T cells, which were ectopically expressed with GFP-
PAK4 and Myc-CDK5RAP3, were lysed with NETN buffer sup-plemented with freshly prepared protease inhibitors (1 mmol/Lphenylmethylsulfonyl fluoride, 1 mg/mL leupeptin, 2 mg/mLaprotinin, and 1 mmol/L DTT). Antibodies used for immuno-precipitation include anti-Myc and rabbit anti-PAK4, and anti-bodies used for immunoblotting are rabbit anti-Myc, rabbit anti-GFP, and rabbit anti-CDK5RAP3.
Confocal microscopyCells were fixed with 4% paraformaldehyde and permeabi-
lized with 0.2% Triton X-100 (9). Images were captured by aconfocal laser scanning microscope LSM510 (Carl Zeiss).
Statistical analysisThe Student t test and the Mann–Whitney test were used
for statistical analysis of data. Tests were considered signifi-cant with P < 0.05.
Results
CDK5RAP3 was overexpressed in human HCCsTo elucidate the role of CDK5RAP3 in human HCCs, we
examined CDK5RAP3 transcripts in human HCCs by using
Mak et al.
Cancer Res; 71(8) April 15, 2011 Cancer Research2950
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

qPCR. We found that CDK5RAP3 transcripts were frequently(58%, 39/67) overexpressed (�2-fold) in the HCCs as com-pared with their corresponding nontumorous livers (Fig. 1A).Moreover, the transcript levels of CDK5RAP3 in the tumorsamples were significantly higher than those of the nontu-morous livers (P < 0.001, Mann–Whitney test). To understandthe clinicopathologic significance of CDK5RAP3 in HCC, wecorrelated the overexpression of CDK5RAP3 mRNA with thepatients’ clinicopathologic features (Table 1). Overexpressionof CDK5RAP3 was found to be significantly associated with amore aggressive phenotype, namely, the presence of tumormicrosatellite formation (P ¼ 0.024) and poorer cellular dif-ferentiation (P ¼ 0.023). However, for the survival analysis,overexpression of CDK5RAP3 was not associated with theoverall and disease-free survival rates among patients(Table 1).To examine the expression level of the protein CDK5RAP3
in human HCCs, we generated an antibody that specificallyrecognized the CDK5RAP3 protein (Supplementary Fig. S1A).Using the purified antibody, we showed that CDK5RAP3 wasubiquitously expressed in a panel of HCC cell lines (Supple-mentary Fig. S1B). To confirm that CDK5RAP3 was upregu-lated in human HCCs, immunohistochemical staining wascarried out. As shown in Fig. 1B, there was a strong cytoplas-mic staining of CDK5RAP3 in the tumor cells but not in thecorresponding nontumorous hepatocytes, confirming thatCDK5RAP3 was overexpressed in HCCs. Because overexpres-sion of CDK5RAP3mRNA in HCC samples was associated witha more metastatic phenotype, we examined the expression ofCDK5RAP3 in patients with extrahepatic metastatic HCC. Atissue microarray consisting of 25 cases each, containing theprimary HCC, extrahepatic metastasis, and nontumorous liverfrom the same patient, was examined for CDK5RAP3 expres-sion. Quantification of CDK5RAP3 staining was scored by anexperienced pathologist (I. Oi-Lin Ng). Eleven of the 25 (44%)cases had higher expression levels of CDK5RAP3 in primary
HCCs than in the corresponding nontumorous livers. Amongthe 11 cases with overexpression of CDK5RAP3, 55% (6/11)had higher levels of CDK5RAP3 in tumor metastases than inthe corresponding primary HCCs (Fig. 1C), indicating upre-gulation of CDK5RAP3 during HCC progression.
CDK5RAP3 enhanced cell proliferation andtumorigenicity of HCC cells
The expression of the human CDK5RAP3 isoform (IC53-2)has previously been shown to promote the proliferation ofHCC cells (7). To study the effect of the full-length (FL) formof CDK5RAP3 on growth properties of HCC cells, we con-ducted a colony formation assay in human HCC cell lineswith transient overexpression and knockdown ofCDK5RAP3. PLC/PRF/5 cells transiently transfected withCDK5RAP3 expression plasmid formed more colonies thanthe vector control (P ¼ 0.005; Fig. 2A), whereas transientknockdown of CDK5RAP3 using short hairpin RNA (shRNA)specific to CDK5RAP3 suppressed colony formation in PLC/PRF/5 cells (P ¼ 0.005; Fig. 2B), indicating that CDK5RAP3promotes HCC cell growth. To further examine the effect ofCDK5RAP3 in HCC tumorigenesis, we established 2CDK5RAP3 stably overexpressing HepG2 cells. TheCDK5RAP3 stable overexpression was confirmed by immu-noblotting (Fig. 2C). A cell proliferation assay using thestable clones showed that the doubling time of bothCDK5RAP3 stably expressing clones 1 and 2 (CDK5RAP3#1,29.28 hours; CDK5RAP3#2, 28.56 hours) was remarkablyshorter than the vector control (31.6 hours), indicating thatstable clones grew faster than the vector (Fig. 2C). Becausethere is little information about the tumorigenic activity ofCDK5RAP3, a soft agar growth assay was conducted toevaluate the change in the anchorage-independent growthproperty of the CDK5RAP3 stably expressing HCC cells. Theresults showed that 3-fold more colonies were formed instable clones CDK5RAP3#1 (P ¼ 0.027) and CDK5RAP3#2
Figure 1. Overexpression ofCDK5RAP3 in HCC. A,CDK5RAP3 transcripts weremeasured in 67 paired humanHCCs and correspondingnontumorous liver tissues byqPCR. The horizontal linesindicate the median of mRNAexpression; P < 0.001, Mann–Whitney test; T, tumor; NT,nontumor. B, representativeimmunohistochemical staining forCDK5RAP3 in a case of HCC andits corresponding nontumorousliver tissues. C, representativeimmunohistochemical staining(magnification � 20) ofCDK5RAP3 and p-PAK4 (Ser474)in nontumorous liver, primaryHCC, and extrahepaticmetastasis.
Nontumor liver
Nontumor liver
HCC
Nontumor liver
Case250
CD
K5R
AP
3
NT0.01
0.1
1
10A B
C
Rel
ativ
e co
py n
fum
ber/
106
T
p-PA
K4
(Ser
474)C
ase
9
HCC
HCC metastasis
HCC HCC metastasis
CDK5RAP3 Activates PAK4 in HCC
www.aacrjournals.org Cancer Res; 71(8) April 15, 2011 2951
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046
(P ¼ 0.002) than in the vector control (Fig. 2D). To addressthe loss-of-function effect of CDK5RAP3 in HCC cells, weestablished CDK5RAP3 stable knockdown clones in PLC/PRF/5 and the highly motile SMMC-7721 HCC cells by usingshRNA (3). Thus, 2 different cell lines were used for estab-lishing stable clones to eliminate cell-line–specific effect.Two CDK5RAP3 stable knockdown clones were selected(shCDK5RAP3#1 and shCDK5RAP3#2) in each cell line,and the knockdown of CDK5RAP3 was confirmed by Wes-tern blotting (Fig. 2E and Supplementary Fig. S2A). Ascompared with vector control cells, the loss of CDK5RAP3significantly reduced both the cell-proliferation rate (Fig. 2E)and anchorage-independent growth of PLC/PRF/5 cells(Fig. 2F, P < 0.005). Furthermore, a similar result wasobserved in CDK5RAP3 stable knockdown SMMC-7721clones (Supplementary Fig. S2). Together with the data in
CDK5RAP3 stably overexpressing HepG2 cells, these resultsconsistently suggest that CDK5RAP3 enhanced the tumor-igenicity of HCC cells and the effect was not cell-line specific.
To further show the tumorigenic properties of CDK5RAP3in vivo, a nude mouse xenograft assay was conducted usingCDK5RAP3 stable knockdown PLC/PRF/5 cells. Although thetumor incidences for the PLC/PRF/5-shCDK5RAP3#2 (4/5)and vector control (5/5) were similar, tumors generated fromPLC/PRF/5-shCDK5RAP3#2 had remarkably lower weightsand slower growth rates than the control (Fig. 2G), suggestingthat loss of CDK5RAP3 inhibited tumor growth in vivo.
Knockdown of CDK5RAP3 inhibited cell migration andinvasiveness of HCC cells
Clinical data for HCCs in this study reveal thatCDK5RAP3 protein levels increased with HCC progression
Table 1. Association of overexpression of CDK5RAP3 mRNA with clinicopathologic features in humanHCCs
Parameters Category CDK5RAP3 notoverexpressed
CDK5RAP3overexpressed
P value
Overall survival 23 37 0.264Disease-free survival 23 37 0.701Sex Male 16 27 0.026a
Female 10 4Age, y �40 2 4 0.678
>40 24 27HBV surface antigen Absent 9 8 0.267
Present 18 30HBV core antigen Absent 14 18 0.582
Present 2 1Liver cirrhosis Absent 8 16 0.139
Present 18 16Resection margin Absent 25 37 0.642
Present 3 2Venous invasion Absent 17 18 0.239
Present 11 21Tumor microsatellite Absent 20 17 0.024a
Present 8 22Liver invasion Absent 16 20 0.192
Present 7 18Tumor encapsulation Absent 16 27 0.322
Present 11 11Cellular differentiation
(Edmondson's grading)I–III 19 15 0.023a
IV–VI 9 23Tumor size (cm) �5 9 17 0.159
>5 17 15Tumor stage I–II 12 13 0.672
III–V 14 19Number of tumor nodules Single 22 27 1.000
Multiple 4 5
aStatistically significant.
Mak et al.
Cancer Res; 71(8) April 15, 2011 Cancer Research2952
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

and that overexpression of CDK5RAP3 was statistically cor-related with clinicopathologic features of a more metastaticphenotype. To examine whether the loss of CDK5RAP3 inhi-bits cell motility, Transwell migration assay was conductedusing SMMC-7721 cells treated with small interfering RNA(siRNA) targeting CDK5RAP3 (siCDK5RAP3). The number ofmigrated SMMC-7721 cells transfected with siCDK5RAP3 wasreduced by 36% as compared with the cells transfected withsiRNA control (Fig. 3A). Furthermore, we then queriedwhether loss of CDK5RAP3 suppressed the invasiveness ofHCC cells. Compared with parental and siRNA-control trans-fected cells, the number of invaded cells was reduced by 43% insiCDK5RAP3 transfected SMMC-7721 cells (Fig. 3B). In con-trast, the CDK5RAP3 stably overexpressing HepG2 cellsshowed an increase in migration rate by 37% in the Transwellassay (Fig. 3C). Thus, both loss- and gain-of-function assaysindicated that CDK5RAP3 promoted the cell migration andinvasiveness of HCC cells.
Association analysis and physical correlation betweenCDK5RAP3 and PAK4 in HCCs
From our functional assays and clinicopathologic correla-tion data, we inferred that CDK5RAP3 was closely associatedwith HCC cell invasiveness. We then analyzed the correlationof the overexpression of CDK5RAP3 transcripts with theexpression levels of metastasis-related genes (e.g., PAK4,PAK1, NF-kB, and p53) in our database (9). Interestingly, wefound that CDK5RAP3 overexpression was significantly corre-lated with PAK4 overexpression in 63 cases of HCC (P < 0.001,Pearson correlation; Fig. 4A).
To delineate the physiologic implication of this correla-tion, we first examined the interaction of PAK4 andCDK5RAP3 by coimmunoprecipitation assay. GFP-PAK4,but not GFP-PAK1, was coprecipitated with Myc-CDK5RAP3in transfected HEK293T cells, indicating that CDK5RAP3 isspecifically associated with PAK4 (Fig. 4B). Furthermore, theendogenous CDK5RAP3 and PAK4 proteins were shown, by
Mock
Vector
CDK5RAP3
A C E
B D F
G
4540353025201510
50
MockMyc-CD5RAP3
β-Actin
β-Actinβ-Actin
Myc-CDK5RAP3HepG2
paren
tal
vecto
r
CDK5RAP3#
1
CDK5RAP3#
2
Paren
tal
vecto
r#1
vecto
r#2
shCDK5R
AP3#1
shCDK5R
AP3#2
Endogenous CDK5RAP3
β-Actin
2580
00
10
20
30
40
50C
ell n
umbe
r (×
104 )
Cel
l num
ber
(×10
4 )
Tum
or v
olum
e (m
m )3
60CDK5RAP3#2CDK5RAP3#1VectorHepG2 parental 250 Vector#1
Vector
Vector
Vector
5/5 4/5
Vector#2shCDK5RAP3#1
CDK5RAP3PLC/PRF/5
shCDK5RAP3#2 shCDK5RAP3#2
shCDK5RAP3#2
shCDK5RAP3#2
1,250
Tumor incidence
1,000
750
500
250
00 1 2 3
Week after injectionTime (h)4 5 6
200
150
100
50
00 24 48 72 96 120 144
70
80
24 48 72 96Time (h)
120 140 168
70
6050
40
30
20
10
0
20
15
10
Num
ber
of c
olon
ies
Num
ber
of c
olon
ies
Num
ber
of c
olon
ies
>50
μm
45
50 1.00
0.75
0.50
0.25
0.00
4035
30
2520
15
105
0
Num
ber
of c
olon
ies
>50
μm
Tum
or w
eigh
t (g)
5
0Vector
Vecto
r
CDK5RAP3#
1
CDK5RAP3#
2
Vecto
r#2
Vecto
r#1
CDK5RAP3#
1
CDK5RAP3#
2
shCDK5RAP3
Vector shCDK5RAP3#2CDK5RAP3
Vector CDK5RAP3
Figure 2. CDK5RAP3-enhanced tumorigenicity of HCC cells. A, the construct expressing Myc-tagged CDK5RAP3 was transfected into PLC/PRF/5cells for colony formation assay. The expression of CDK5RAP3 was confirmed by immunoblotting. Error bars, mean � SD; *, P < 0.05 compared withvector control (Student t test). Empty vector (vector) and untransfected cells (mock) were used as controls. B, same as (A), but endogenous CDK5RAP3expression was silenced by shCDK5RAP3. C, CDK5RAP3 stably expressing HepG2 cells (CDK5RAP3#1 and CDK5RAP3#2), vector control, and parental cellswere used for proliferation assay. Top, immunoblotting; bottom, proliferation curve. D, cells in (C) were used for soft agar growth assay. *, P < 0.05; **,P < 0.005, compared with HepG2–vector control cells (Student t test). Error bars, mean � SD. E, CDK5RAP3 stable knockdown PLC/PRF/5 cloneswere used for proliferation assay. Top, immunoblotting; bottom, proliferation curve. F, 2 CDK5RAP3 stable knockdown PLC/PRF/5 clones were used for softagar growth assay; *, P < 0.005 compared with PLC-vector 1 (Student t test). Error bars, mean � SD. G, CDK5RAP3 stable knockdown PLC/PRF/5clones and vector control were used for the nude mouse xenograft model. Tumor volume (error bars, mean � SEM); tumor weight (error bars, mean � SD).
CDK5RAP3 Activates PAK4 in HCC
www.aacrjournals.org Cancer Res; 71(8) April 15, 2011 2953
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

coimmunoprecipitation assay, to interact with each other inHepG2 cells (Fig. 4C). Moreover, a GST affinity pull-down assayshowed a direct interaction between CDK5RAP3 and PAK4(Fig. 5A). By confocal microscopy, CDK5RAP3 was found to besignificantly colocalized with PAK4 (Fig. 4D). The subcellularlocalization of CDK5RAP3, which was mainly nuclear, wasaltered by coexpression with PAK4 and became more concen-trated at peripheral regions (Fig. 4D).
CDK5RAP3-regulated PAK4 kinase activityPAK4 has been shown to induce tumorigenesis and cell
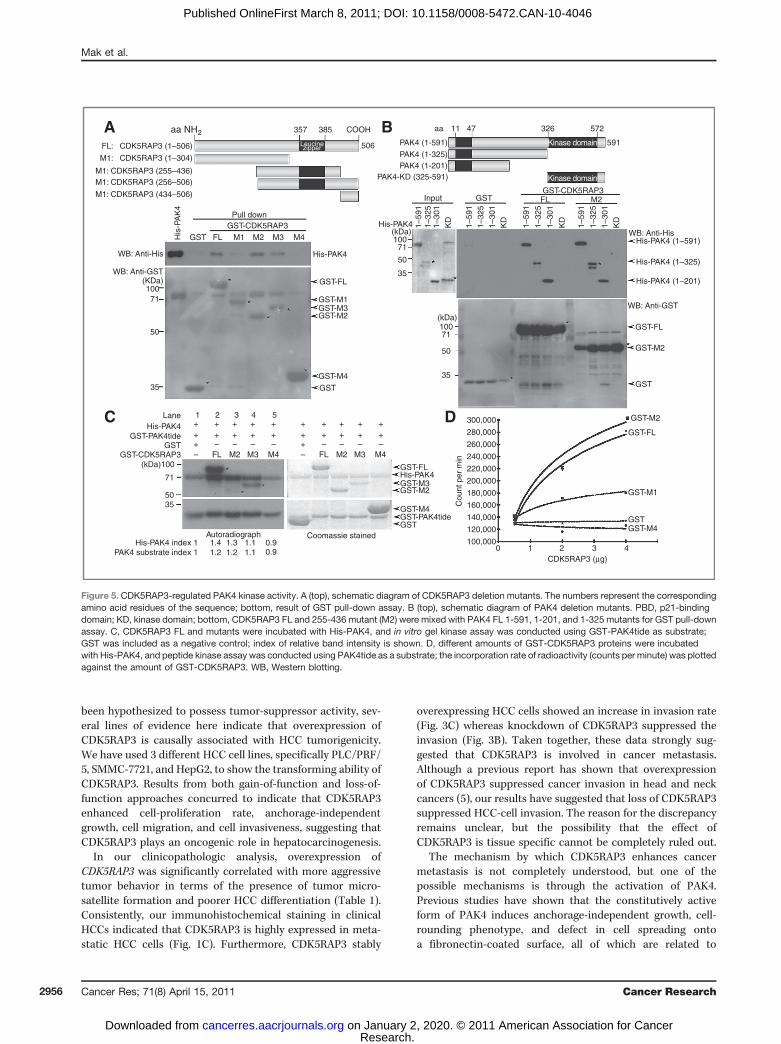
adhesion by phosphorylating its substrates (15), and, therefore,wewonderedwhetherCDK5RAP3promotesHCC formation byregulating PAK4 activity. To evaluate this, we constructed apanel of CDK5RAP3 truncated mutants (Fig. 5A) and assessedtheir interaction with PAK4 as well as their effect on PAK4activity. The results of the GST affinity pull-down assayrevealed that the GST control, 434-506 and 1-304 mutants ofCDK5RAP3, did not bind to PAK4. However, FL and 255-436mutant (M2) showed strong binding affinity to PAK4, whereasthe 256-506mutant (M3) displayed a weaker binding affinity toPAK4 (Fig. 5A). It is possible that the additional aa 437-506mayhave attenuated the binding of M3 to PAK4. To further inves-tigate how CDK5RAP3 interacted with PAK4, we constructed 3deletion mutants of PAK4: 2 of them (aa 1–201 and 1–325)contained the N-terminal p21-binding domain (PBD) and onecontained the C-terminal kinase domain (aa 325–591). Using aGST affinity pull-downassay, we found that both the FL andM2
of CDK5RAP3were associatedwith N-terminal region of PAK4,which contained the PBD domain but not the C-terminalkinase domain (Fig. 5B). To further examine whetherCDK5RAP3 regulated PAK4 activity, a PAK4 kinase assaywas conducted. The result revealed that incubation of FL,M2, and M3 of CDK5RAP3 with PAK4 not only promotedthe PAK4 activity measured by a specific GST-PAK4tide sub-strate but also promoted the autophosphorylation of PAK4(Fig. 5C, lanes 2–4). In contrast, the 434-506 mutant (M4) ofCDK5RAP3 showed no PAK4 activation, similar to the GSTcontrol (Fig. 5C, lane 5). This result indicated that the aasequence 255–436 of CDK5RAP3 contains an activationdomain of PAK4. Moreover, as revealed by kinase assay,CDK5RAP3 was a good substrate for PAK4 (Fig. 5C). Using aPAK4 peptide kinase assay, we further illustrated that the FLand M2 of CDK5RAP3 significantly enhanced the activity ofPAK4, but not the 1-304mutant (M1),M4, andGST control, in adose-dependent manner (Fig. 5D). Together with the interac-tion data, our results implied that the direct binding ofCDK5RAP3 to PAK4 elicited the activation of PAK4. Further-more, the result from Fig. 5B indicated that CDK5RAP3 mayinteract with the N-terminal domain, possibly the PBD, thusleading to the activation of PAK4.
Knockdown of PAK4 abolished the promotion of cellinvasiveness in CDK5RAP3 overexpression cells
To investigate whether the overexpression of CDK5RAP3activated PAK4 in HCCs, the correlation between CDK5RAP3
*
600CDK5RAP3
β-Actin500
400
300
Num
ber
of in
vade
d ce
llsN
umbe
r of
imm
igra
ted
cells
Num
ber
of im
mig
rate
d ce
lls
200
100
0
500
350
300
250
200
150
100
50
0
C
BA
400
300
200
100
0
Mock
Vecto
r
CDK5RAP3#
1
CDK5RAP3#
2
siControl siCDK5RAP3
Mock siControl siCDK5RAP3
Mock siControl siCDK5RAP3
*Moc
ksiC
ontro
l
siCDK5R
AP3
Mock siControl siCDK5RAP3
Figure 3. Knockdown ofCDK5RAP3 suppressed HCC cellmigration and invasiveness.SMMC-7721 cells weretransfected with siCDK5RAP3 andsiRNA control, andnontransfected cells weresubjected to (A) the Transwellmigration assay and (B) theinvasion assay. *, P ¼ 0.05compared with siControl (Studentt test). Immunoblotting showedthe CDK5RAP3 expression.Representative photomicrographsare shown. C, CDK5RAP3 stablyoverexpressing HepG2 cells wereused for the Transwell migrationassay. *, P < 0.05 compared withHepG2–vector control cells(Student t test). Error bars, mean� SD.
Mak et al.
Cancer Res; 71(8) April 15, 2011 Cancer Research2954
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

and p-PAK4 expression levels in the same set of tissuemicroarray was assessed by immunohistochemical staining.Phosphorylated PAK4 showed strong nuclear staining, and thestaining was higher in the primary HCCs than in the corre-sponding nontumorous liver tissues (10/20 cases); however,the expression levels of CDK5RAP3 and p-PAK4 positivelycorrelated with one another (Fig. 1C; P ¼ 0.024, Pearsoncorrelation). In addition, 3 of 5 cases with higher CDK5RAP3expression in the metastases than in their primary HCCsshowed a higher level of p-PAK4 in metastases than in primaryHCCs (Fig. 1C), suggesting a positive correlation of CDK5RAP3and PAK4 activity during HCC progression. In addition, PAK4activity, as revealed by p-PAK4 staining, was quantitativelyreduced in the tumor tissues obtained from the CDK5RAP3-knockdown, nude mouse xenograft (Supplementary Fig. S3).To further confirm the specific activation of PAK4 byCDK5RAP3 within cells, PAK4, in combination with increasingdoses of CDK5RAP3, was cotransfected into HepG2 cells andp-PAK4 was determined by immunoblotting. As shown inFig. 6A, increased expression of CDK5RAP3 promoted the totalprotein and phosphorylation levels of PAK4 in HepG2 cells in adose-dependent manner. The upregulation of phosphorylatedPAK4 was functionally active, as indicated by the increase inPAK4 activity in CDK5RAP3 cotransfected cells (Fig. 6A).Consistently, the phosphorylation of PAK4 by CDK5RAP3was observed in CDK5RAP3 stably overexpressing HepG2 cells(Fig. 6B). To examine whether CDK5RAP3 promoted HCC-cellinvasiveness through activation of PAK4, we used siRNA tospecifically knock down PAK4 in CDK5RAP3 stably overex-pressing HepG2 cells and conducted the cell-invasion assay.
The result showed that loss of PAK4, as compared with thevector control cells, significantly reduced the invasiveness ofboth CDK5RAP3 stably expressing clones (Fig. 6C) and, thus,strongly suggests that CDK5RAP3 enhanced HCC-cell inva-siveness through activation by PAK4.
Discussion
Several studies have investigated the roles of CDK5RAP3 incarcinogenesis by using various cancer models; however, sofar, its definite roles remain contradictory. CDK5RAP3 hasbeen found to promote apoptosis induced by genotoxic stressin HeLa cells by triggering G2/M arrest (3). In addition,CDK5RAP3 has been proposed to be a tumor suppressorbecause CDK5RAP3 inhibits the NF-kB cell-survival pathwayand its protein level is significantly reduced in head and necksquamous cell carcinomas (5). However, a similar NF-kB–suppressive effect of CDK5RAP3 was not observed in our studyusing HCC cells, and this could probably be attributed to sometissue-specific effects. In contrast, overexpression of theCDK5RAP3 isoform has been reported to promote cell pro-liferation of HCC cells (7). Recently, CDK5RAP3 has beenshown to be overexpressed in lung adenocarcinoma (16). Inthe present study, using qPCR and immunohistochemicalstaining assays, we showed that both transcripts and proteinof CDK5RAP3 were frequently and significantly overexpressedin human HCCs (Fig. 1). The mechanism of such overexpres-sion is currently unclear, but it has been reported that thechromosomal region 17q, which contains the CDK5RAP3, isfrequently amplified in HCCs (8). Although CDK5RAP3 has
Figure 4. CDK5RAP3 interactedwith PAK4. A, CDK5RAP3 andPAK4 transcripts were measuredin 63 paired HCC samples byqPCR. The line from the originindicates the positive correlation;P < 0.001 (Pearson correlation). B,HEK293T cells werecotransfected with GFP-PAK4and Myc-CDK5RAP3 in thecoimmunoprecipitation assay.GFP-PAK1 was included as acontrol for specific interaction. C,endogenous CDK5RAP3 andPAK4 in HepG2 cells werecoimmunoprecipitated with theindicated antibodies. D, confocalmicroscopy with Myc-CDK5RAP3and GFP-PAK4 cotransfected inHepG2 cells. Myc-CDK5RAP3was visualized by anti-Mycantibody (Ab) and secondaryantibody conjugated with TexasRed. IP, immunoprecipitation;WB, Western blotting.
kDa85
50C
otra
nsfe
ctio
nS
ingl
e tr
ansf
ectio
n
50WB: lgG
OverlayMyc -CDK5RAP3GFP-PAK4
Myc -CDK5RAP3GFP-PAK4
WB: α-CDK5RAP3
WB: α-PAK4IP: α-Myc
α-GFP
α-Myc
α-GFP
CDK5RAP3 (log10)0.1
0.1
1
10
100
A
D
B C
PAK
4 (lo
g 10)
1 10 100
IP
α-PA
K4
No
Ab
cont
rol
Inpu
t
Moc
k
Vec
tor
GF
P-P
AK
1
GF
P-P
AK
4
Input
85
CDK5RAP3 Activates PAK4 in HCC
www.aacrjournals.org Cancer Res; 71(8) April 15, 2011 2955
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046
been hypothesized to possess tumor-suppressor activity, sev-eral lines of evidence here indicate that overexpression ofCDK5RAP3 is causally associated with HCC tumorigenicity.We have used 3 different HCC cell lines, specifically PLC/PRF/5, SMMC-7721, and HepG2, to show the transforming ability ofCDK5RAP3. Results from both gain-of-function and loss-of-function approaches concurred to indicate that CDK5RAP3enhanced cell-proliferation rate, anchorage-independentgrowth, cell migration, and cell invasiveness, suggesting thatCDK5RAP3 plays an oncogenic role in hepatocarcinogenesis.
In our clinicopathologic analysis, overexpression ofCDK5RAP3 was significantly correlated with more aggressivetumor behavior in terms of the presence of tumor micro-satellite formation and poorer HCC differentiation (Table 1).Consistently, our immunohistochemical staining in clinicalHCCs indicated that CDK5RAP3 is highly expressed in meta-static HCC cells (Fig. 1C). Furthermore, CDK5RAP3 stably
overexpressing HCC cells showed an increase in invasion rate(Fig. 3C) whereas knockdown of CDK5RAP3 suppressed theinvasion (Fig. 3B). Taken together, these data strongly sug-gested that CDK5RAP3 is involved in cancer metastasis.Although a previous report has shown that overexpressionof CDK5RAP3 suppressed cancer invasion in head and neckcancers (5), our results have suggested that loss of CDK5RAP3suppressed HCC-cell invasion. The reason for the discrepancyremains unclear, but the possibility that the effect ofCDK5RAP3 is tissue specific cannot be completely ruled out.
The mechanism by which CDK5RAP3 enhances cancermetastasis is not completely understood, but one of thepossible mechanisms is through the activation of PAK4.Previous studies have shown that the constitutively activeform of PAK4 induces anchorage-independent growth, cell-rounding phenotype, and defect in cell spreading ontoa fibronectin-coated surface, all of which are related to
aa NH2
FL: CDK5RAP3 (1–506)
A B
C D
Leucinezipper
M1: CDK5RAP3 (1–304)
M1: CDK5RAP3 (255–436)M1: CDK5RAP3 (256–506)
M1: CDK5RAP3 (434–506)
WB: Anti-His
His
-PA
K4
GST FL M1 M2 M3 M4
His-PAK4
His-PAK4
PAK4 (1-591)
aa 11 47 326 572
591
PAK4 (1-325)PAK4 (1-201)
PAK4-KD (325-591)
Input GSTGST-CDK5RAP3
Kinase domain
Kinase domain
FL
WB: Anti-HisHis-PAK4 (1–591)
His-PAK4 (1–325)
His-PAK4 (1–201)
GST-FL
WB: Anti-GST
GST-M2
GST
GST-M2
GST-FL
GST-M1
GST-M4
43CDK5RAP3 (μg)
210100,000
120,000
140,000
160,000Cou
nt p
er m
in
180,000
200,000
220,000
240,000
260,000
280,000
300,000
GST
*
M2
1–59
11–
325
1–30
1K
D
1–59
11–
325
1–30
1K
D
1–59
1
1–32
5
1–30
1
KD
1–59
1
1–32
5
1–30
1
KD
GST-FL
GST-M1
35
(kDa)10071
50
35
50
71100(kDa)
GST-M3GST-M2
GST-M4
GST
GST-FLHis-PAK4GST-M3GST-M2
GST-M4GST-PAK4tideGST
Coomassie stainedAutoradiograph
3550
71
(kDa)100GST-CDK5RAP3
GSTGST-PAK4tide
His-PAK4Lane 1
+ + + + + + + + + ++ + + + + + + + + ++– FL M2 M3 M4 – FL M2 M3 M4
– – – – + – – – –
2 3 4 5
His-PAK4 index 1 1.4 1.3 1.11.2 1.2 1.1
0.90.9PAK4 substrate index 1
Pull downGST-CDK5RAP3
WB: Anti-GST(KDa)100
71
50
35
357 385 COOH
506
Figure 5. CDK5RAP3-regulated PAK4 kinase activity. A (top), schematic diagram of CDK5RAP3 deletion mutants. The numbers represent the correspondingamino acid residues of the sequence; bottom, result of GST pull-down assay. B (top), schematic diagram of PAK4 deletion mutants. PBD, p21-bindingdomain; KD, kinase domain; bottom, CDK5RAP3 FL and 255-436 mutant (M2) were mixed with PAK4 FL 1-591, 1-201, and 1-325 mutants for GST pull-downassay. C, CDK5RAP3 FL and mutants were incubated with His-PAK4, and in vitro gel kinase assay was conducted using GST-PAK4tide as substrate;GST was included as a negative control; index of relative band intensity is shown. D, different amounts of GST-CDK5RAP3 proteins were incubatedwith His-PAK4, and peptide kinase assay was conducted using PAK4tide as a substrate; the incorporation rate of radioactivity (counts per minute) was plottedagainst the amount of GST-CDK5RAP3. WB, Western blotting.
Mak et al.
Cancer Res; 71(8) April 15, 2011 Cancer Research2956
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

cancer-cell migration (17, 18). With regard to the upstreamsignaling, PAK4 is found to be activated by hepatocyte andkeratinocyte growth factors during cell migration (19, 20). Ourdata indicate that overexpression of CDK5RAP3 in HCC cellswas significantly associated with enhanced expression andactivity of PAK4 through Ser474 phosphorylation (Figs. 1C, 4A,6A and B). With regard to the mechanism by which theoverexpression of CDK5RAP3 could increase the expressionof PAK4 in HCCs, we hypothesized that CDK5RAP3 mightstabilize PAK4 through inhibition of PAK4 ubiquitination.Furthermore, recent reports have shown that CDK5RAP3mutually stabilizes its binding partner, KIAA0776, by inhibit-ing the ubiquitination of KIAA0776 (21). Thus, it is possiblethat CDK5RAP3 not only activates PAK4 but also stabilizesPAK4, making it a potent regulator of PAK4 activity.We have shown that CDK5RAP3 is a PAK4 cellular binding
partner, using coimmunoprecipitation, coimmunostaining,and GST affinity pull-down assays. Although we have notexamined the interaction of CDK5RAP3 with all other mem-bers of the PAK family, the coimmunoprecipitation dataclearly show that CDK5RAP3 only binds to PAK4 but notto PAK1, indicating that the binding of CDK5RAP3 is relativelyspecific for PAK4. In addition, the analysis in human HCCsshowed no significant correlation of CDK5RAP3 and PAK1overexpression. Interestingly, our confocal immunofluores-cence staining data revealed that ectopic expression ofCDK5RAP3 with PAK4 altered the localization of CDK5RAP3,which was mainly nuclear and cytoplasmic, to be concen-trated at the membrane peripheral (Fig. 4D). Whether theperipheral localization of CDK5RAP3 plays a role in theactivation of PAK4 and the regulation of cell migration
requires further investigation. However, it is worth notingthat CDK5RAP3 is a good substrate of PAK4 (Fig. 5C); thepossibility that PAK4 can modulate the phosphorylation, and,therefore, the translocation and function of CDK5RAP3, iscurrently under investigation.
With our in vitro kinase assay, we showed that CDK5RAP3significantly and directly promoted the autophosphorylationand activity of PAK4 (Fig. 5C). In addition, we showed thatincreased expression of CDK5RAP3 remarkably enhanced thephosphorylation and activity of PAK4, both in CDK5RAP3transiently transfected cells and in stably expressing HCC cells(Fig. 6A and B), thus strongly indicating that CDK5RAP3 is acellular activator of PAK4. With regard to the mechanism bywhich CDK5RAP3 activates PAK4, we mapped the centralregion of CDK5RAP3, which contains a leucine zipper domain,required for the activation of PAK4. In our GST affinity pull-down experiment, we found that CDK5RAP3 interacted withthe N-terminal region of PAK4, which contains the PBD, andFigure 5A shows that the CDK5RAP3 mutant (aa 255–436) is,indeed, the activator for PAK4, as compared withmutants thatshowed lower or absent binding ability. This result led us tospeculate that, similar to small Rho GTPase, Cdc42, and Rac1,the CDK5RAP3 activates PAK4 through direct protein–proteininteraction for which PBD is a requisite factor.
PAK4 has been shown to play important roles in cellmigration and adhesion by regulating the actin-cytoskeletonorganization and integrin signaling (22). Furthermore, studieshave shown that PAK4 is an important mediator for hepato-cyte growth factor–stimulated cell migration in epithelial(20) and cancer cells (23). Although the role of PAK4 inHCC has not been characterized, a recent report has shown
Figure 6. CDK5RAP3 promotedHCC-cell invasiveness mediatedthrough activation by PAK4 kinaseactivity. A, increasing amounts ofCDK5RAP3 were transientlycotransfected with PAK4 inHepG2 cells; expression of GFP-PAK4 and Myc-CDK5RAP3 andphosphorylation of PAK4 weredetermined by immunoblotting;bottom, graph showing PAK4kinase activity in cell lysates. B,cell lysates from vector andCDK5RAP3 stably expressingHepG2 cells (CDK5RAP3#1 andCDK5RAP3#2) were analyzed byWestern blotting. C, CDK5RAP3stably overexpressing HepG2cells were transfected withsiPAK4, as indicated for invasionassay; top, Western blotting;bottom, graph showing numberof invaded cells after transfectionof PAK4 and control siRNA.*P ¼ 0.005 (Student t test);representative photomicrographsare shown.
A C
B
GFP-PAK4
Vector Myc-CDK5RAP3
α-CDK5RAP3
α-p-PAK4 (ser474)
α-GFPα-β-Actin
Vector
Moc
k
siC
ontr
ol
siControlsiPAK4
Negative substratePAK4tide
30,000
25,000
20,000C
ount
s pe
r m
in
Vect
orC
DK5
RAP
3#1
CDK5RAP
3#2
15,000
10,000
5,000
0
GFP-PAK4 – +
0
Num
ber
of in
vade
d ce
lls
20406080
100120140160180
––
+ +
Myc-CDK5RAP3
CDK5RAP3
Vecto
r
CDK5RAP
3#1
CDK5RAP3#
2
p-PAK4 (ser474)PAK4β-Actin
PAK4myc-CDK5RAP3Endogenous CDK5RAP3
β-Actin
siP
AK
4
Moc
k
siC
ontr
ol
siP
AK
4
Moc
k
siC
ontr
ol
siP
AK
4
CDK5RAP3#1 CDK5RAP3#1
1μg 2μg
1 μg 2 μg
CDK5RAP3 Activates PAK4 in HCC
www.aacrjournals.org Cancer Res; 71(8) April 15, 2011 2957
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

that overexpression of PAK4 in prostate cancer promotes cellmigration and invasion (15). Thus, we hypothesized that theactivation of PAK4 by CDK5RAP3 may promote HCC-cellinvasion. For this purpose, we used siRNA to specificallyknock down PAK4 in CDK5RAP3 stably overexpressing HepG2cells and showed that the loss of PAK4 attenuated the inva-siveness of stable clones back to similar level as the vectorcontrol, indicating that PAK4 plays a key role in CDK5RAP3-mediated HCC cancer metastasis (Fig. 6C). Our findings haveestablished that upregulation of CDK5RAP3may occur in HCCprogression and metastasis through the regulation of PAK4.
In summary, we provide the first evidence that CDK5RAP3is overexpressed in human HCCs and that overexpression ofCDK5RAP3 promotes metastasis of HCC. We also provideevidence that supports a single mechanism whereinCDK5RAP3 enhances HCC metastasis through the activationof PAK4. Therefore, inhibition of CDK5RAP3 can potentially beused to suppress HCC formation, providing a new moleculartarget for therapeutic intervention in HCC.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Dr. Honglin Li for providing the plasmids and Dr. Abel Chun forcritical reading of manuscript. Irene O.L. Ng is the Loke Yew Professor inPathology.
Grant Support
The Hong Kong Research Grant Council (N_HKU715/08, HKU 1/06Cand 7/CRF/09) and The University of Hong Kong, Seed Funding Pro-gramme (200711159100; to Y.P. Ching), supported this research study.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must, therefore, be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
Received November 11, 2010; revised February 1, 2011; accepted February 22,2011; published OnlineFirst March 8, 2011.
References1. Ching YP, Qi Z, Wang JH. Cloning of three novel neuronal Cdk5
activator binding proteins. Gene 2000;242:285–94.2. Yin X, Warner DR, Roberts EA, Pisano MM, Greene RM. Novel
interaction between nuclear co-activator CBP and the CDK5 activatorbinding protein—C53. Int J Mol Med 2005;16:251–6.
3. Jiang H, Luo S, Li H. Cdk5 activator-binding protein C53 regulatesapoptosis induced by genotoxic stress via modulating the G2/M DNAdamage checkpoint. J Biol Chem 2005;280:20651–9.
4. Wang J, He X, Luo Y, Yarbrough WG. A novel ARF-binding protein(LZAP) alters ARF regulation of HDM2. Biochem J 2006;393:489–501.
5. Wang J, An H, Mayo MW, Baldwin AS, Yarbrough WG. LZAP, aputative tumor suppressor, selectively inhibits NF-kappaB. CancerCell 2007;12:239–51.
6. Chen J, Liu B, Liu Y, Han Y, Yu H, Zhang Y, et al. A novel gene IC53stimulates ECV304 cell proliferation and is upregulated in failing heart.Biochem Biophys Res Commun 2002;294:161–6.
7. Xie YH, He XH, Tang YT, Li JJ, Pan ZM, Qin WX, et al. Cloning andcharacterization of human IC53-2, a novel CDK5 activator bindingprotein. Cell Res 2003;13:83–91.
8. Raidl M, Pirker C, Schulte-Hermann R, Aubele M, Kandioler-Eckers-berger D, Wrba F, et al. Multiple chromosomal abnormalities in humanliver (pre)neoplasia. J Hepatol 2004;40:660–8.
9. Ching YP, Leong VY, Lee MF, Xu HT, Jin DY, Ng IO. p21-activatedprotein kinase is overexpressed in hepatocellular carcinoma andenhances cancer metastasis involving c-Jun NH2-terminal kinaseactivation and paxillin phosphorylation. Cancer Res 2007;67:3601–8.
10. Leung TH, Ching YP, Yam JW, Wong CM, Yau TO, Jin DY, et al.Deleted in liver cancer 2 (DLC2) suppresses cell transformation bymeans of inhibition of RhoA activity. Proc Natl Acad Sci U S A2005;102:15207–12.
11. Wong CC, Wong CM, Tung EK, Man K, Ng IO. Rho-kinase 2 isfrequently overexpressed in hepatocellular carcinoma and involvedin tumor invasion. Hepatology 2009;49:1583–94.
12. Ching YP, Leong VYL, Wong CM, Kung HF. Identification of anautoinhibitory domain of p21-activated protein kinase 5. J BiolChem2003;278:33621–4.
13. Rennefahrt UE, Deacon SW, Parker SA, Devarajan K, Beeser A,Chernoff J, et al. Specificity profiling of Pak kinases allows identifica-tion of novel phosphorylation sites. J Biol Chem 2007;282:15667–78.
14. Lew J, Beaudette K, Litwin CM, Wang JH. Purification and character-ization of a novel proline-directed protein kinase from bovine brain.J Biol Chem 1992;267:13383–90.
15. Wells CM, Whale AD, Parsons M, Masters JR, Jones GE. PAK4: apluripotent kinase that regulates prostate cancer cell adhesion. J CellSci 2010;123:1663–73.
16. Stav D, Bar I, Sandbank J. Usefulness of CDK5RAP3, CCNB2, andRAGE genes for the diagnosis of lung adenocarcinoma. Int J BiolMarkers 2007;22:108–13.
17. Callow MG, Clairvoyant F, Zhu S, Schryver B, Whyte DB, Bischoff JR,et al. Requirement for PAK4 in the anchorage-independent growth ofhuman cancer cell lines. J Biol Chem 2002;277:550–8.
18. Dan C, Kelly A, Bernard O, Minden A. Cytoskeletal changes regulatedby the PAK4 serine/threonine kinase aremediated by LIM kinase 1 andcofilin. J Biol Chem 2001;276:32115–21.
19. Lu Y, Pan ZZ, Devaux Y, Ray P. p21-activated protein kinase 4 (PAK4)interacts with the keratinocyte growth factor receptor and participatesin keratinocyte growth factor-mediated inhibition of oxidant-inducedcell death. J Biol Chem 2003;278:10374–80.
20. Wells CM, Abo A, Ridley AJ. PAK4 is activated via PI3K in HGF-stimulated epithelial cells. J Cell Sci 2002;115:3947–56.
21. Kwon J, Cho HJ, Han SH, No JG, Kwon JY, Kim H. A novel LZAP-binding protein, NLBP, inhibits cell invasion. J Biol Chem 2010;285:12232–40.
22. Jaffer ZM, Chernoff J. p21-activated kinases: three more join the Pak.Int J Biochem Cell Biol 2002;34:713–7.
23. Paliouras GN, Naujokas MA, Park M. Pak4, a novel Gab1 bindingpartner, modulates cell migration and invasion by the Met receptor.Mol Cell Biol 2009;29:3018–32.
Mak et al.
Cancer Res; 71(8) April 15, 2011 Cancer Research2958
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046

Published OnlineFirst March 8, 2011.Cancer Res Grace Wing-Yan Mak, Mandy Man-Lok Chan, Veronica Yee-Law Leong, et al. Hepatocellular Carcinoma Metastasis
Associated Protein CDK5RAP3, Promotes−Kinase Overexpression of a Novel Activator of PAK4, the CDK5
Updated version
10.1158/0008-5472.CAN-10-4046doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2011/03/07/0008-5472.CAN-10-4046.DC1
Access the most recent supplemental material at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/early/2011/04/08/0008-5472.CAN-10-4046To request permission to re-use all or part of this article, use this link
Research. on January 2, 2020. © 2011 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst March 8, 2011; DOI: 10.1158/0008-5472.CAN-10-4046


















