






on March 7, 2019 by guest - iai.asm.org fileAbstract The cyclic-di-GMP binding protein PilZ has been...
51
Analysis of the Borrelia burgdorferi cyclic-di-GMP binding protein PlzA reveals a role in motility and virulence Section: Molecular pathogenesis. Running title: A role of B. burgdorferi PilZ in motility and virulence. Key words: Borrelia burgdorferi/Lyme disease/PilZ/motility/virulence. Joshua E. Pitzer 1# , Syed Z. Sultan 1# , Yoshihiro Hayakawa 2 , Gerry Hobbs 3 , Michael R. Miller 4 and Md. A. Motaleb 1* 1 Department of Microbiology and Immunology, Brody School of Medicine, East Carolina University, Greenville, NC 27834. 2 Department of Applied Chemistry, Faculty of Engineering, Aichi Institute of Technology, 1247 Yachigusa, Yakusa-cho, Toyota 470-0392, Japan. 3 Department of Statistics, West Virginia University, Morgantown, WV 26506. 4 Department of Biochemistry, Health Sciences Center, West Virginia University, Morgantown, WV 26506. *Corresponding author: TEL: 252-744-3129; FAX: 252-744-3535; E-mail: [email protected] # these authors contributed equally to this manuscript. Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Infect. Immun. doi:10.1128/IAI.00075-11 IAI Accepts, published online ahead of print on 28 February 2011 on April 3, 2019 by guest http://iai.asm.org/ Downloaded from on April 3, 2019 by guest http://iai.asm.org/ Downloaded from on April 3, 2019 by guest http://iai.asm.org/ Downloaded from
Transcript of on March 7, 2019 by guest - iai.asm.org fileAbstract The cyclic-di-GMP binding protein PilZ has been...

1"
"
Analysis of the Borrelia burgdorferi cyclic-di-GMP binding protein PlzA reveals a 1
"
role in motility and virulence 2
"
3
"
Section: Molecular pathogenesis. 4
"
Running title: A role of B. burgdorferi PilZ in motility and virulence. 5
"
6
"
Key words: Borrelia burgdorferi/Lyme disease/PilZ/motility/virulence. 7
"
8
"
Joshua E. Pitzer1#, Syed Z. Sultan1#, Yoshihiro Hayakawa2, Gerry Hobbs3, Michael R. 9
"
Miller4 and Md. A. Motaleb1* 1 0
"
1 1
"
1Department of Microbiology and Immunology, Brody School of Medicine, East Carolina 1 2
"
University, Greenville, NC 27834. 1 3
"
2Department of Applied Chemistry, Faculty of Engineering, Aichi Institute of Technology, 1 4
"
1247 Yachigusa, Yakusa-cho, Toyota 470-0392, Japan. 1 5
"
3Department of Statistics, West Virginia University, Morgantown, WV 26506. 1 6
"
4Department of Biochemistry, Health Sciences Center, West Virginia University, 1 7
"
Morgantown, WV 26506. 1 8
"
1 9
"
*Corresponding author: TEL: 252-744-3129; FAX: 252-744-3535; E-mail: 2 0
"
"
2 2
"
#these authors contributed equally to this manuscript. 2 3
"
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.00075-11 IAI Accepts, published online ahead of print on 28 February 2011
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2"
"
Abstract 2 4
"
The cyclic-di-GMP binding protein PilZ has been implicated in bacterial motility and 2 5
"
pathogenesis. Although BB0733 (PlzA), the only PilZ domain-containing protein in B. 2 6
"
burgdorferi was reported to bind c-di-GMP, neither its role in motility or virulence nor it’s 2 7
"
affinity for c-di-GMP have been reported. We determined that PlzA specifically binds c-2 8
"
di-GMP with high affinity (Kd 1.25 µM), consistent with Kd values reported for c-di-GMP 2 9
"
binding proteins from other bacteria. Inactivation of the monocistronically transcribed 3 0
"
plzA resulted in a translucent colony morphology, whereas the wild-type colonies were 3 1
"
opaque/solid. While the swimming pattern of mutant cells appeared normal, on swarm 3 2
"
plates mutant cells exhibited a significantly reduced swarm diameter, demonstrating a 3 3
"
role of plzA in motility. Furthermore, the plzA mutant cells were significantly less 3 4
"
infectious in experimental mice (as determined by ID50) relative to wild-type spirochetes. 3 5
"
The mutant also had lowered survival rates in fed ticks than wild-type. Consequently, 3 6
"
plzA mutant cells failed to complete the mouse-tick-mouse infection cycle, indicating 3 7
"
plzA is essential for the enzootic life cycle of B. burgdorferi. All of these defects were 3 8
"
corrected when the mutant was complemented in cis. We propose that failure of plzA 3 9
"
mutant cells to infect mice was due to altered motility; however, the possibility that an 4 0
"
unidentified factor(s) contributed to interruption of the B. burgdorferi enzootic life cycle 4 1
"
cannot yet be excluded. 4 2
"
4 3
"
4 4
"
4 5
"
4 6
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3"
"
Introduction 4 7
"
4 8
"
Cyclic-dimeric-guanosine monophosphate (c-di-GMP) [bis(3',5')-cyclic diguanylic acid] is 4 9
"
a ubiquitous bacterial second messenger. C-di-GMP is produced from 2 molecules of 5 0
"
GTP by the activity of diguanylate cyclase and is degraded by phosphodiesterase 5 1
"
enzymes. The activities of these cyclase and phosphodiesterase enzymes are often 5 2
"
regulated by changing physiological conditions. C-di-GMP is implicated in the regulation 5 3
"
of diverse bacterial properties, including biosynthesis of adhesins and 5 4
"
exopolysaccharide matrix for biofilm formation, different forms of motility, long term 5 5
"
survival and response to environmental stresses, in addition to production of virulence 5 6
"
factors and to virulence (14,29,39,76,88,96). These cellular functions regulated by c-di-5 7
"
GMP often utilize multiple receptors and signaling mechanisms. Several types of c-di-5 8
"
GMP receptors have been reported, including PilZ, PelD, I-site effectors, FleQ and Clp 5 9
"
(29,30,40,76). Although many of the downstream mechanisms remain to be identified, 6 0
"
studies indicate that c-di-GMP can impact cellular signaling at multiple levels, including 6 1
"
transcription, translation, posttranslation, protein activity, protein secretion and protein 6 2
"
stability (7,15,17,30,41,48,50,57,58,70,86,93). 6 3
"
6 4
"
One c-di-GMP receptor, the PilZ protein, was originally identified in silico as a putative 6 5
"
conserved sequence based on the binding of c-di-GMP to the BcsA1 protein, a 6 6
"
component of the cellulose synthase complex of Acetobacter xylinum (2,69,94). The 6 7
"
‘PilZ domain’, named after the PilZ protein from Pseudomonas aeruginosa, is required 6 8
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4"
"
for the assembly of functional Type 4 pili (T4P) (1). Recently, proteins containing PilZ 6 9
"
domains have been shown to bind c-di-GMP in several species of bacteria ((29,76) and 7 0
"
ref. therein). Strong affinity of c-di-GMP for PilZ domains has been observed, with 7 1
"
dissociation constants (Kd) in the range of 1 nM to 2 たM, and this strong affinity is 7 2
"
consistent with the low concentration of c-di-GMP in cells (29,37,76,78). Furthermore, 7 3
"
PilZ-proteins from a number of bacteria have been implicated in the regulation of 7 4
"
specific cellular processes such as motility, polysaccharide synthesis, biofilm formation 7 5
"
and virulence of animal and plant pathogens (5,13,15,30,41,48,60,70). The role of PilZ 7 6
"
appears different in different bacterial species. For example, as mentioned above, PilZ 7 7
"
of P. aeruginosa (PA2960) is necessary for T4P biogenesis and twitching motility. A pilZ 7 8
"
knockout is defective in the secretion of PilA polymers to the bacterial surface, though 7 9
"
intracellular PilA pools are not affected (1). On the other hand, a knockout of the PilZ 8 0
"
ortholog in Neisseria meningitides is piliated but is not able to produce bacterial 8 1
"
aggregates (11); a PilZ knockout in Xanthomonas campestris pv. campestris has 8 2
"
slightly reduced T4P-dependent motility on semi-solid Eiken agar (47); and a pilZ 8 3
"
mutant in Xanthomonas axonopodis pv. citri resulted in increased swarming motility 8 4
"
relative to its parental wild-type cells (27). A PilZ domain-containing protein, Alg44, 8 5
"
was shown to up-regulate alginate production in P. aeruginosa (48). Furthermore, in E. 8 6
"
coli (and Salmonella enterica), the reduced motility of a phosphodiesterase mutant is 8 7
"
alleviated by a pilZ mutation (58,70), indicating a role of PilZ in bacterial motility. 8 8
"
Specifically, the E. coli PilZ protein, YcgR, was found to control motility by directly 8 9
"
interacting with and altering flagellar switch proteins FliG, FliM or motor protein MotA 9 0
"
function in response to c-di-GMP (3,7,17,57). In Vibrio cholerae, a mutant in genes 9 1
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

5"
"
encoding PilZ proteins also resulted in altered motility (60). These observations suggest 9 2
"
that signaling by PilZ could be modulated differently in different species. 9 3
"
9 4
"
Although c-di-GMP may have different roles in different species of bacteria, little is 9 5
"
known about this signaling system in Borrelia burgdorferi, the Lyme disease spirochete. 9 6
"
The spirochete resides in disparate host environments, including Ixodes ticks and 9 7
"
mammalian hosts. During transmission between hosts, B. burgdorferi modulates gene 9 8
"
expression in response to environmental factors such as nutrient availability, pH, CO2, 9 9
"
temperature, host factors and immune response (10,55,65,74,90,91). C-di-GMP levels, 1 0 0
"
which are controlled by a two-component system, are also believed to be involved in 1 0 1
"
bacterial adaptation to different host environments (84,88,95). While many species of 1 0 2
"
bacteria were reported to contain multiple enzymes responsible for controlling the levels 1 0 3
"
of c-di-GMP (22), bioinformatics, as well as experimental analysis, indicate that B. 1 0 4
"
burgdorferi utilizes only a limited number of enzymes to modulate c-di-GMP: a single 1 0 5
"
diguanylate cyclase; two phosphodiesterases; and a single c-di-GMP binding PilZ 1 0 6
"
homolog designated as PlzA (2,21,22,66,71,87). B. burgdorferi diguanylate cyclase 1 0 7
"
(BB0419; Rrp1) was shown to regulate a diverse set of genes, many of which were 1 0 8
"
reported to be associated with B. burgdorferi virulence, implying a role of c-di-GMP in 1 0 9
"
pathogenesis (66,71). We recently demonstrated that BB0363 is a phosphodiesterase 1 1 0
"
which specifically hydrolyzed c-di-GMP into pGpG (87). Furthermore, a bb0363/pdeA 1 1 1
"
mutant exhibited impaired motility, and those mutant cells were unable to infect 1 1 2
"
experimental mice, irrespective of the size or route of inoculum (needle or tick), 1 1 3
"
demonstrating a role of c-di-GMP in B. burgdorferi motility and virulence (87). 1 1 4
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

6"
"
1 1 5
"
Recently, PlzA was shown to specifically bind c-di-GMP, and the expression of plzA 1 1 6
"
was shown to be modulated in different environmental conditions (21). While PilZ has 1 1 7
"
different functions in different bacteria, a role for the lone c-di-GMP binding protein PlzA 1 1 8
"
has not been demonstrated in B. burgdorferi. Here we demonstrate that PlzA binds c-di-1 1 9
"
GMP in vitro with strong affinity. The deletion of plzA reduces motility and infectivity in 1 2 0
"
experimental mice. The mutant cells exhibit lowered viability in ticks. Consequently, the 1 2 1
"
plzA cells are unable to complete the mouse-tick-mouse infection cycle, consistent with 1 2 2
"
a role for PilZ in bacterial pathogenesis. Furthermore, by constructing a plzA mutant in 1 2 3
"
the pdeA mutant background where the cellular c-di-GMP level is 2x higher than the 1 2 4
"
wild-type cells we demonstrate that PlzA regulates B. burgdorferi motility in a different 1 2 5
"
manner than it does in E. coli and that elevated c-di-GMP in the B. burgdorferi pdeA 1 2 6
"
mutant regulates motility by a mechanism independent of PlzA. A mechanism of altered 1 2 7
"
motility and infectivity is proposed. 1 2 8
"
1 2 9
"
1 3 0
"
Materials and Methods 1 3 1
"
1 3 2
"
Bacterial strains and growth conditions 1 3 3
"
Low-passage, virulent B. burgdorferi strain B31-A3-68〉bbe002 is used as the wild type 1 3 4
"
throughout the study (a kind gift from R. Rego and P. Rosa, RML, NIH) (63). This strain 1 3 5
"
is a derivative of A3-68 (63), lacking circular plasmid 9 (cp9) and linear plasmid 56 1 3 6
"
(lp56), with the bbe002 gene inactivated using a PflgB-aadA cassette to increase 1 3 7
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

7"
"
transformation frequency (36,63). B31-A3-68〉bbe002 is a derivative of strain B31 (16). 1 3 8
"
The genome of the virulent B31 strain has been sequenced and was found to contain a 1 3 9
"
total of 21 plasmids with 12 linear and 9 circular plasmids, in addition to the 960 kb 1 4 0
"
linear chromosome (12,20). B. burgdorferi was cultured in liquid Barbour-Stoenner-Kelly 1 4 1
"
(BSK-II) medium as described; plating BSK was prepared using 0.6% agarose (52). 1 4 2
"
Swarm plate assays were carried out using 0.35% agarose in plating BSK; 1 4 3
"
approximately 1 x 106 cells in 5 たl volume were spotted onto plates containing plating 1 4 4
"
BSK medium diluted 1:10 in Dulbecco’s phosphate buffered saline without divalent 1 4 5
"
cations. Because B. burgdorferi is a slow growing organism with 8–12 h generation 1 4 6
"
time, swarm plates were incubated for 5-7 days (52,53). The paired Student’s T-Test 1 4 7
"
(Minitab Inc.) was used to compare wild-type and plzA mutant cell swarm diameters. 1 4 8
"
1 4 9
"
BB0733 recombinant protein expression 1 5 0
"
1 5 1
"
Recombinant BB0733 protein (PlzA) was expressed with an N-terminal 6x His fusion 1 5 2
"
tag. To express BB0733 protein in E. coli, the coding sequence was amplified by PCR 1 5 3
"
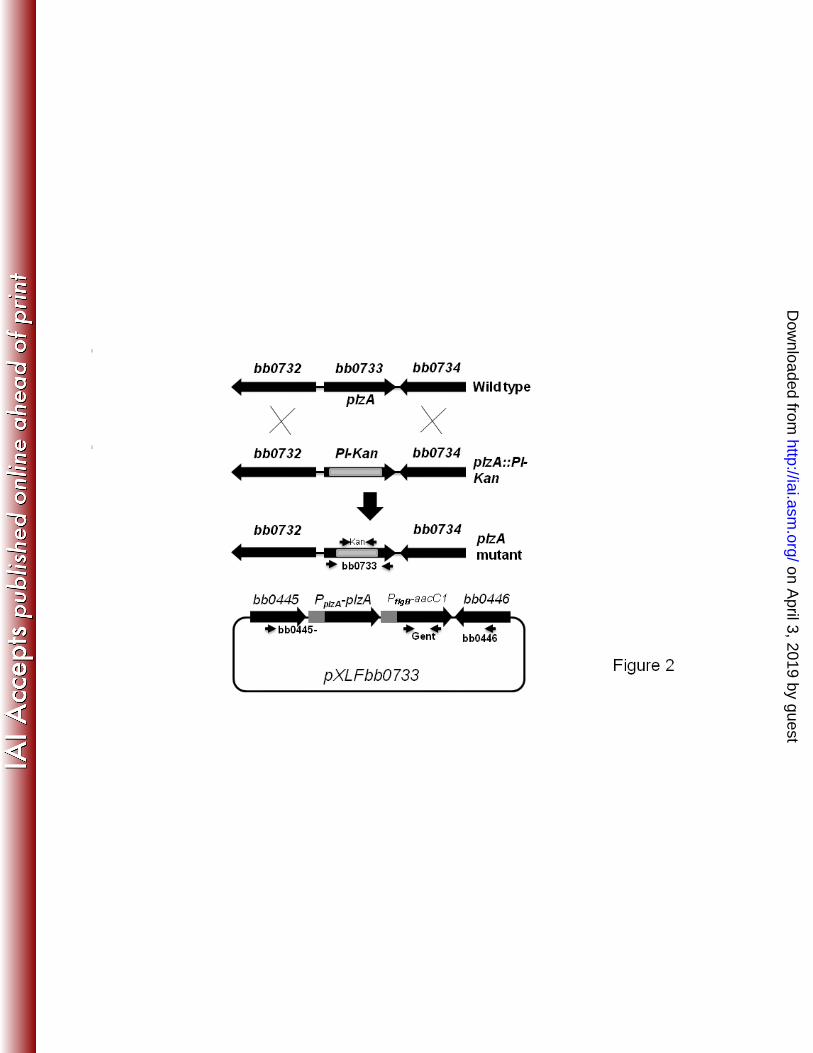
using primers (5’-3’) R-BB0733-F (CTTTTATCTAGAAAAATAAGAG) and T-BB0733-R 1 5 4
"
(CTTTGATTAATTGAAATAATC). The amplified DNA fragment was ligated into an E. 1 5 5
"
coli expression vector pTrc His TOPO (Invitrogen Inc.) using TA cloning. The vector was 1 5 6
"
then digested and sequenced to confirm its integrity. E. coli cells containing pbb0733-1 5 7
"
TRC were induced with 0.25 mM Isopropyl く-D-1-thiogalactopyranoside at 30°C for 4 h 1 5 8
"
and Ni2+-affinity purified under native conditions. Purified recombinant BB0733 protein 1 5 9
"
was resolved by SDS-PAGE to determine protein concentration and purity. 1 6 0
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

8"
"
1 6 1
"
C-di-GMP binding assays by equilibrium dialysis. 1 6 2
"
1 6 3
"
Equilibrium dialysis was performed as described (70). Briefly, DispoBiodialyzer 1 6 4
"
cassettes (The Nest Group, Southborough, MA) contain 2 chambers separated by a 5 1 6 5
"
kDa cut off membrane; 60 µl c-di-GMP (2 – 50 µM) was injected into one chamber and 1 6 6
"
60 µl test protein (10 µM) BB0733 or positive control E. coli c-di-GMP binding PilZ 1 6 7
"
receptor YcgR (70) was injected into the other chamber; c-di-GMP and test proteins 1 6 8
"
were in 300 mM NaCl, 0.5 mM EDTA, 10% glycerol, 50 mM sodium phosphate (pH7.4). 1 6 9
"
Following gentle rocking at room temperature for 24 hours to reach equilibrium, samples 1 7 0
"
from both sides of cassettes were removed, boiled for 4 min, centrifuged and 0.22 µm 1 7 1
"
microfiltered (Millipore Ultrafree MC). C-di-GMP concentrations in both samples were 1 7 2
"
determined via reverse phase HPLC on a Supelcosil LC-18T column using the same 1 7 3
"
buffer and gradient program as described in (71) and c-di-GMP was quantified at 254 1 7 4
"
nm. The binding constant was determined using Graph Pad Prism 5 software. 1 7 5
"
1 7 6
"
Reverse transcription-polymerase chain reaction (RT-PCR) 1 7 7
"
1 7 8
"
Exponentially growing B. burgdorferi wild-type cells (2 x 107 cells ml-1) were treated with 1 7 9
"
RNAprotect™ followed by total RNA isolation using the RNeasy mini kit (Qiagen Inc.). 1 8 0
"
Contaminating DNA in the RNA samples was removed by RNase-free Turbo® DNase I 1 8 1
"
(Ambion Inc.) digestion for 3 h at 37°C followed by RNeasy mini purification. For RT-1 8 2
"
PCR, cDNA was prepared from 1 たg RNA using Superscript III reverse transcriptase 1 8 3
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

9"
"
according to the manufacturer’s protocol (Invitrogen Inc.). RT-PCR primer sequences 1 8 4
"
(5’-3’) were a1 (TGTTAAATATATAAGCAGCTTATGCG); a2 1 8 5
"
(GCGGGCAATACTGTATCTTCTTGA); b1 (TTGAGATGAGAATAGGCTCTTATACC); 1 8 6
"
b2 (GCGGGCAATACTGTATCTTCTTGA); c1 (TTGAGTGCTTCTGCTTATTCTTCT) 1 8 7
"
and c2 (GATACGAAAATACTTATCACAAAAGC). A Bio-Rad iScriptTM cDNA synthesis 1 8 8
"
kit and an iCycler detection system were used to measure bb0733 transcript levels 1 8 9
"
according to the manufacturer’s instructions. B. burgdorferi enolase was used a 1 9 0
"
reference gene (54,87). The gene specific primers (5’-3’) were RT-enolase-F 1 9 1
"
(TGGAGCGTACAAAGCCAACATT); RT-enolase-R 1 9 2
"
(TGAAAAACCTCTGCTGCCATTC); Bb0733-rt-F 1 9 3
"
(TTGAGATGAGAATAGGCTCTTATACC); and Bb0733-rt-R 1 9 4
"
(GCGGGCAATACTGTATCTTCTTGA). The relative level of expression was calculated 1 9 5
"
using the 2-〉〉Ct method (46,79,87). 1 9 6
"
1 9 7
"
Construction of a plzA (bb0733) mutant 1 9 8
"
1 9 9
"
Targeted inactivation of bb0733 (a 786 bp gene) was achieved by homologous 2 0 0
"
recombination using a promoterless Pl-Kan cassette (87). PCR amplification of bb0733, 2 0 1
"
construction of the inactivation plasmids and electroporation of linear bb0733-Pl-Kan 2 0 2
"
DNA into competent cells were carried out as described previously (52,53,73,87). 2 0 3
"
Briefly, bb0733 DNA was PCR-amplified using the GoTaq PCR system (Promega Inc.) 2 0 4
"
and primers (5’-3’) BB0733-KO-F (AGTTTTGCATTTGGAAGCTGGA) and BB0733-KO-2 0 5
"
R (ATTCCCAAAAGTGCCCTCATG). The amplified DNA was ligated into the pGEM-T 2 0 6
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 0"
"
Easy vector (Promega Inc.), yielding pBB0733-Easy. Pl-Kan was similarly PCR-2 0 7
"
amplified from PflgB-Kan (8) using primers (5’-3’) PrLs-Kan AflII-F 2 0 8
"
(CTTAAGTAGTTAAAAGCAAT) and PrLs-Kan AflII-R (CTTAAGTTAGAAAAACTCAT). 2 0 9
"
The AflII-restricted Pl-Kan DNA was then inserted into the AflII sites within bb0733 2 1 0
"
(located 167 and 676 base pairs from the ATG start codon) of the pbb0733-Easy vector, 2 1 1
"
yielding pbb0733-Pl-Kan. This deletion–insertion was in-frame. Restriction mapping 2 1 2
"
indicated that the direction of transcription of Pl-Kan was the same as that of bb0733. 2 1 3
"
bb0733-Pl-Kan was digested and separated from pbb0733-Pl-Kan vector by NotI 2 1 4
"
digestion, precipitated and electroporated into the wild-type competent cells (52). 2 1 5
"
Mutants were selected on solid growth medium containing 200 µg ml-1 kanamycin plus 2 1 6
"
100 µg ml-1 streptomycin. Resistant colonies were analyzed by PCR for the presence of 2 1 7
"
the Pl-Kan cassette using primers PrLs-Kan AflII-F and -R as well as using primers 2 1 8
"
BB0733-KO-F and -R. PCR-positive mutants were further examined for their plasmid 2 1 9
"
contents using 21 sets of primers to detect 21 linear and circular plasmids (100). 2 2 0
"
2 2 1
"
Complementation of bb0733 (plzA) mutant 2 2 2
"
2 2 3
"
To complement bb0733::Pl-Kan, the promoter of bb0733 (PplzA; 400 bp upstream from 2 2 4
"
the ATG start codon of bb0733/plzA gene) and the coding sequence was PCR-amplified 2 2 5
"
using PCR with primers (5’-3’) 733 comp-F-SpeI 2 2 6
"
(GCGCCACTAGTTGTGATAATAGAAAAGCTAACTGCGAAATA) and 733-comp-R-NotI 2 2 7
"
(GCGCGCGGCCGCCAAATACAACCAAGAAGACTTAAAC). Restriction sites are in 2 2 8
"
bold font. The PCR product was ligated into pGEM-T Easy. The bb0733 promoter-2 2 9
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 1"
"
bb0733 (Pbb0733-bb0733/PplzA-plzA) DNA was then inserted into suicide vector 2 3 0
"
pXLF14301 (99) using SpeI-NotI restriction digestion, yielding pXLFBB0733. Fifty 2 3 1
"
micrograms of the resultant plasmid was electroporated into the bb0733:Pl-Kan 2 3 2
"
competent cells. Transformants were selected in plating BSK medium containing 100 µg 2 3 3
"
ml-1 streptomycin, 200 µg ml-1 kanamycin plus 40 µg ml-1 gentamicin. Resistant 2 3 4
"
transformants were analyzed by PCR for the presence of kan, gent and an intact 2 3 5
"
bb0733 gene. Positive clones were further examined for their plasmid content by PCR. 2 3 6
"
2 3 7
"
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and western 2 3 8
"
blot 2 3 9
"
SDS-PAGE and western blotting with an enhanced chemiluminescent detection method 2 4 0
"
(GE Healthcare) were carried out as previously reported (23). B. burgdorferi strains B31-2 4 1
"
A3-68〉bbe002 (wild-type), plzA or plzA-pdeA cells were cultured to 5 x 107 cells ml-1. 2 4 2
"
Cells were pelleted, resuspended in PBS and equalized to protein content using 2 4 3
"
Bradford assay reagent (Bio-Rad Inc.). Samples were boiled and approximately 5 たg of 2 4 4
"
cell lysate was loaded per lane. Proteins were transferred to Polyvinylidene Fluoride 2 4 5
"
(PVDF) membrane and western blotted using monoclonal FlaB (A. Barbour), polyclonal 2 4 6
"
MotB (J. Carroll), FliG1 and FliG2 (C. Li), FliM (D. Blair), CheY3 and FliL antisera. 2 4 7
"
Specific monoclonal or polyclonal reactivity to B. burgdorferi FlaB, MotB, FliG1, FliG2, 2 4 8
"
FliM, FliL and CheY3 has been reported (42,43,51,53,54,72); Motaleb et al. 2 4 9
"
unpublished). 2 5 0
"
2 5 1
"
Experimental mouse-tick-mouse infection model of B. burgdorferi 2 5 2
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 2"
"
2 5 3
"
Six week old female C3H/HeN mice were purchased from Charles River Laboratories, 2 5 4
"
North Carolina and housed in the East Carolina University animal facility at the Brody 2 5 5
"
School of Medicine according to the institutional guidelines for the care and use of 2 5 6
"
laboratory animals. For infection via needle, 5 x 102 to 5 x 106 in vitro-grown spirochetes 2 5 7
"
were injected subcutaneously as described (16,35,64). The number of spirochetes was 2 5 8
"
determined using a Petroff- Hausser chamber and each clone was verified for their 2 5 9
"
retention of lp25, lp28-1 and lp36 plasmids. Mice were bled 2 weeks post infection for 2 6 0
"
immunoblot analysis of mouse sera against B. burgdorferi antigens to determine 2 6 1
"
infectivity, as described (35,80,87). Reisolation of B. burgdorferi from mouse skin, 2 6 2
"
bladder and joint tissues was performed 6 weeks post infection to assess the ability of 2 6 3
"
spirochetes to infect mice as described (16,26,83,87). Mouse tissues were placed in 2 6 4
"
BSK-II growth medium and incubated for up to 35 days; the presence of spirochetes 2 6 5
"
was determined by dark-field microscopy. The ID50 was calculated as described (see 2 6 6
"
below) (35,62). 2 6 7
"
2 6 8
"
For tick infection studies, naïve Ixodes scapularis larvae were purchased from 2 6 9
"
Oklahoma State University. Two independent experiments were performed with ticks. 2 7 0
"
Ticks were kept at 23-24°C under a 14:10 light: dark photoperiod in a humidified 2 7 1
"
chamber with 85–90% relative humidity. Approximately 100 larval ticks were fed to 2 7 2
"
repletion on spirochete-infected mice for 5–7 days, allowed to fall off and collected. One 2 7 3
"
subset of larvae was dissected 7 days after repletion, and the isolated midguts were 2 7 4
"
analyzed by indirect immunofluorescence assays (IFA) for the presence of spirochetes 2 7 5
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 3"
"
(83,87). A second subset of fed larvae were surface sterilized using 3% H2O2 followed 2 7 6
"
by 70% ethanol, crushed in BSK-II medium and plated to determine the colony-forming 2 7 7
"
units per tick. A third subset of larvae was crushed and genomic DNA was extracted 2 7 8
"
using DNeasy Blood and Tissue Kit (Qiagen Inc.). qPCR was performed to quantitate 2 7 9
"
the level of tick actin gene using primers (5’-3’) ActinT-F 2 8 0
"
(GATCATGTTCGAGACCTTCA) and ActinT-R (CGATACCCGTGGTACGA) versus the 2 8 1
"
level of B. burgdorferi flaB using primers FlaB qRT-F 2 8 2
"
(TTGCTGATCAAGCTCAATATAACCA ) and FlaB qRT-R 2 8 3
"
(TTGAGACCCTGAAAGTGATGC), as described (97,99) . The remaining fed larvae 2 8 4
"
were allowed to molt to nymphs. The infected nymphs were then fed to repletion on 2 8 5
"
naïve mice (six week old female C3H/HeN), allowed to fall off and collected. Subsets of 2 8 6
"
nymphs were processed for IFA or plated to determine the number of colony-forming 2 8 7
"
units per tick as described above. Six weeks post infection, mice sera were collected 2 8 8
"
for immunoblot analysis, and tissues were collected for reisolation of B. burgdorferi. 2 8 9
"
2 9 0
"
Determination of Infectious dose 50 (ID50) and statistical analysis 2 9 1
"
2 9 2
"
The dose required to infect 50% of the mice inoculated was experimentally determined 2 9 3
"
for wild-type, the plzA mutant and the complemented plzA+ strains as described 2 9 4
"
(35,62). The data from the ID50 infection experiment and the single dose infection 2 9 5
"
experiment for each strain were combined for the estimations of the 50% infectious 2 9 6
"
dose. Comparison between strain ID50 values was made using a generalized linear 2 9 7
"
model with a probit link function. This method is also known as probit regression (35) 2 9 8
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 4"
"
and in it we assume identical slopes in the response/log-dose relationship but different 2 9 9
"
intercepts for each strain. Graphically those assumptions manifest themselves as dose 3 0 0
"
response curves with lateral shifts corresponding to the changes in intercept. 3 0 1
"
Additionally, an overdispersion parameter was fit in order to accommodate greater 3 0 2
"
homogeneity in infection rates than would otherwise be permitted by the model. All 3 0 3
"
calculations were carried out using JMP V9 Software (SAS Institute Inc, Cary, NC). 3 0 4
"
3 0 5
"
Artificial inoculation of ticks by immersion studies 3 0 6
"
3 0 7
"
Tick immersion studies were performed as described (87). Briefly, approximately 150 3 0 8
"
tick larvae were artificially infected by immersion (in duplicate) in equal density 3 0 9
"
exponential phase B. burgdorferi cultures (5x107 cells/ml) as described (4,59,83,87). 3 1 0
"
Ticks were fed to repletion on separate naïve mice for 5-7 days, allowed to fall off and 3 1 1
"
collected. Subsets of larvae were dissected 7-10 days after repletion, and the crushed 3 1 2
"
ticks were analyzed by immunofluorescence assay (IFA) for the presence of spirochetes 3 1 3
"
(83,87). Fed larvae were treated as described above to determine the colony forming 3 1 4
"
units per tick. Reisolation of spirochetes from mice ears, joints and bladders was 3 1 5
"
performed as described above. 3 1 6
"
3 1 7
"
Indirect immunofluorescence assays (IFA) 3 1 8
"
3 1 9
"
Ticks were dissected in 25 たl PBS, 5 mM MgCl2 in Teflon coated microscopic slides, 3 2 0
"
mixed by pipetting, and then air dried (4,87). To avoid quenching by hemin in the blood, 3 2 1
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 5"
"
dissected tick contents were 10-fold serially diluted. Slides were blocked with 0.75% 3 2 2
"
BSA in PBS, 5 mM MgCl2 and washed with PBS-5 mM MgCl2. Spirochetes were 3 2 3
"
detected using a 1:100 dilution of goat anti-B. burgdorferi antisera labeled with 3 2 4
"
fluorescein isothiocyanate (Kirkegaard & Perry Laboratories). Images were captured 3 2 5
"
using a Zeiss Axio Imager M1 microscope coupled with an AxioCam MRc digital camera 3 2 6
"
(Carl Zeiss Inc.). 3 2 7
"
3 2 8
"
3 2 9
"
Results 3 3 0
"
3 3 1
"
BB0733 binds c-di-GMP with high affinity. Bioinformatics BLAST analysis suggests 3 3 2
"
that BB0733 is the only PilZ domain-containing protein in B. burgdorferi (PilZ domain; 3 3 3
"
Pfam PF07238) (1,2,20). BB0733, which has been named as PlzA, shares significant 3 3 4
"
amino acid sequence similarity with orthologs in other Borrelia strains (21). Although 3 3 5
"
PlzA shares poor homology with PilZ proteins of other species of bacteria, it contains 3 3 6
"
the conserved residues of the ‘PilZ domain’ (RxxxR; D/NzSxxG where x is any amino 3 3 7
"
acid and z is a hydrophobic residue) but lacks the so-called ‘PilZ-N’ terminal domain (N-3 3 8
"
terminal domain of E. coli PilZ protein YcgR [PF07317]) (2,70) and therefore is 3 3 9
"
considered as a ‘stand-alone’ c-di-GMP effector protein (21). Recently, using X-ray 3 4 0
"
crosslinking studies, Freedman et al. confirmed that PlzA specifically binds c-di-GMP 3 4 1
"
and that the conserved Arginine (R) residues of 160RxxxR (number is based on amino 3 4 2
"
acid residues of BB0733 protein; (20) is critical for c-di-GMP binding (21). However, a 3 4 3
"
dissociation constant (Kd) of PlzA for c-di-GMP was not determined (21). To determine 3 4 4
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 6"
"
the Kd of PlzA, synthetic c-di-GMP (38,41) and recombinant, purified His6-PlzA (Fig. 1) 3 4 5
"
were used in equilibrium dialysis, as described (70). An E. coli c-di-GMP binding PilZ 3 4 6
"
protein YcgR, was used as a positive control (70). Fig. 1, shows that His6-PlzA is highly 3 4 7
"
purified and binds c-di-GMP; His6-PlzA did not bind other nucleotides tested (cAMP, 3 4 8
"
GTP, cGMP; not shown) confirming the findings reported by Freedman et al. (21). The 3 4 9
"
dissociation constant of PlzA was determined to be 1.25 ± 0.17 µM, indicating high 3 5 0
"
affinity for c-di-GMP, consistent with Kd values (1 nM to 2 たM) reported for c-di-GMP 3 5 1
"
binding proteins from other bacteria (29,37). 3 5 2
"
3 5 3
"
Inactivation and complementation of plzA in B. burgdorferi. B. burgdorferi genome 3 5 4
"
sequence analysis predicted that the genes upstream (bb0732) and downstream 3 5 5
"
(bb0734) of bb0733 are transcribed in the opposite direction of bb0733 (Fig. 2). Using 3 5 6
"
RT-PCR we confirmed that bb0733 is a monocistronic mRNA that is expressed in 3 5 7
"
growing B. burgdorferi cells and that bb0732 and bb0734 genes are not co-transcribed 3 5 8
"
(not shown). bb0732 is annotated as a penicillin-binding protein that is predicted to be 3 5 9
"
responsible for biosynthesis and degradation of murein sacculus and peptidoglycan of 3 6 0
"
cell envelope, whereas bb0734 encodes a Sua5/YciO/YrdC/YwlC family protein of 3 6 1
"
unknown function. In Shewanella oneidensis and the psychrophile Colwellia 3 6 2
"
psychrerythraea 34H, the Sua5/YciO/YrdC/YwlC family protein is a putative translation 3 6 3
"
factor involved in translation, as well as ribosomal structure and biogenesis (28,49). 3 6 4
"
3 6 5
"
The expression of B. burgdorferi PlzA was reported to be maintained throughout the 3 6 6
"
enzootic cycle; however, its expression was significantly increased in ticks after a 3 6 7
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 7"
"
bloodmeal (21). A function of plzA has not been demonstrated. To investigate the 3 6 8
"
function of PlzA during the enzootic life cycle of B. burgdorferi, we inactivated plzA with 3 6 9
"
a promoterless-kanamycin cassette (Pl-Kan) by allelic exchange, as described (87). 3 7 0
"
Electroporation of the plzA::Pl-Kan linear DNA (Fig. 2) into the wild-type competent cells 3 7 1
"
yielded ~10 kanamycin resistant clones. PCR analysis confirmed that a 509 bp internal 3 7 2
"
fragment of plzA is replaced by the 872 bp Pl-Kan cassette in all clones, confirming 3 7 3
"
inactivation of plzA (Fig. 3 left panel). Characterization of several positive clones 3 7 4
"
confirmed identical genotype and phenotypes; one randomly chosen clone was used for 3 7 5
"
subsequent analysis. 3 7 6
"
3 7 7
"
To restore the synthesis (and phenotype; see below) of plzA at the wild-type level, we 3 7 8
"
complemented the plzA mutant by in cis, genomic integration where genes bb0445 and 3 7 9
"
bb0446 converge (see Fig. 2-bottom). The insertion of PplzA-plzA was downstream of 3 8 0
"
both bb0445-bb0446 genes and less likely to affect their expression as described 3 8 1
"
previously (44,99). We chose to complement the mutant in cis since shuttle vectors 3 8 2
"
developed so far to complement B. burgdorferi mutants are multi-copy plasmids (89) 3 8 3
"
that result in overexpression of the target gene (43,53,77,89). Accordingly, we 3 8 4
"
introduced suicide vector pXLFbb0733 (Fig. 2-bottom) containing an intact plzA gene 3 8 5
"
under its own promoter (PplzA-plzA) into the mutant cells. Positive clones were 3 8 6
"
characterized by PCR using kanamycin, gentamicin and bb0445-bb0446 specific 3 8 7
"
primers (Fig. 3 middle panel). PCR analysis confirmed that PplzA-plzA integrated 3 8 8
"
between the bb0445-bb0446 region, as expected (see Figures 2 and 3). To detect the 3 8 9
"
expression of plzA in wild-type, the plzA mutant and the complemented (plzA+) strains, 3 9 0
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 8"
"
we employed qPCR using gene specific primers. Real-time PCR detected the plzA 3 9 1
"
transcripts in wild-type and the plzA+ but not in the plzA mutant cells (Fig. 3 right panel). 3 9 2
"
Furthermore, the level of plzA transcripts synthesized in the complemented plzA+ cells 3 9 3
"
were approximately 70% the level of wild-type cells, indicating the restoration of plzA 3 9 4
"
synthesis in the complemented cells. 3 9 5
"
3 9 6
"
To confirm retention of B. burgdorferi endogenous plasmids required for infectivity 3 9 7
"
(56,61) in the plzA mutant and complemented strains, PCR-based plasmid profiling 3 9 8
"
using each of the plasmid specific primers was employed (61,100). As shown in Fig. 4 3 9 9
"
the plasmid profile of wild-type, the plzA mutant and the plzA+ strains were the same, 4 0 0
"
confirming retention of all plasmids required for infectivity (see below). 4 0 1
"
4 0 2
"
plzA mutant is deficient in swarming motility. Mutating a gene encoding PilZ has 4 0 3
"
been reported to generate various phenotypes: rough, rugose, opaque or translucent 4 0 4
"
bacterial colony morphology and altered motility (2,13,27,58,60,98). To determine if a 4 0 5
"
mutation in plzA altered B. burgdorferi swimming pattern or swarming motility (42,53), 4 0 6
"
the mutant cells were analyzed by microscopy and swarm plate motility assays in 0.35% 4 0 7
"
agarose (52). Although cellular morphology or the swimming pattern (run, pause/flex, 4 0 8
"
reverse) (42,53) of the plzA mutant cells were indistinguishable from the wild-type cells, 4 0 9
"
the swarm diameter of plzA mutant cells was significantly reduced (p value 0.001) 4 1 0
"
compared to the wild-type cells when the plates were incubated for 7 days (Fig. 5). 4 1 1
"
Additionally, while the colony morphology of the wild-type cells was solid/opaque, the 4 1 2
"
mutant cells colonies were translucent (figure was not clear to show). Both the 4 1 3
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

1 9"
"
swarming motility and colony morphology phenotypes were restored to wild-type level 4 1 4
"
upon complementation (Fig. 5 and not shown), indicating that the phenotypes 4 1 5
"
associated with plzA cells are solely due to the mutation and not a secondary alteration 4 1 6
"
elsewhere. Noticeable growth defects were not observed in plzA mutant cells, indicating 4 1 7
"
the decreased swarming phenotype was not due to a growth defect (not shown). These 4 1 8
"
results indicate that plzA is essential for normal B. burgdorferi motility and colony 4 1 9
"
morphology. 4 2 0
"
4 2 1
"
To determine if the deficient motility exhibited by plzA cells resulted from alteration of 4 2 2
"
motility/chemotaxis protein synthesis, we performed western blotting using specific 4 2 3
"
antisera against the periplasmic flagella FlaB; motor switch proteins FliM, FliG1, FliG2 4 2 4
"
and MotB; flagellar protein FliL or the chemotaxis protein CheY3. As shown in Fig. 6, 4 2 5
"
the levels of proteins detected using specific antisera in the plzA mutant cells were not 4 2 6
"
altered when compared with the wild-type cells. These results indicate that the reduced 4 2 7
"
swarming observed in the plzA mutant cells were not due to a reduction in levels of 4 2 8
"
FlaB, FliL, FliG2, FliM, MotB or CheY3 proteins. 4 2 9
"
4 3 0
"
plzA mutant cells are significantly attenuated in infecting mice. 4 3 1
"
In some bacteria, pilZ was shown to be required for the host colonization/disease 4 3 2
"
processes (47,60). In order to evaluate the infection potential of the plzA mutant, groups 4 3 3
"
of C3H/HeN mice were challenged with 10-fold increasing doses of wild-type, plzA::Pl-4 3 4
"
Kan or the isogenic complemented plzA+ cells to determine the 50% infectious dose 4 3 5
"
(ID50). Two weeks post inoculation, the mice were bled and their sera were assessed for 4 3 6
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 0"
"
reactivity with B. burgdorferi antigen membrane protein A, also known as P39 (35,80). 4 3 7
"
Furthermore, to confirm the serology results, mice were sacrificed 6 weeks 4 3 8
"
postinoculation and spirochetes were reisolated from ear, bladder and joint tissues. 4 3 9
"
Serology results indicated that plzA mutant cells were attenuated in establishing an 4 4 0
"
infection in mice (not shown) and correlated well with reisolation of spirochetes from 4 4 1
"
tissues examined (Table 1). Whereas the plzA+ cells did not show an increase in the 4 4 2
"
ID50 compared to the parental wild-type cells, the ID50 for the plzA mutant was 4 4 3
"
reproducibly more than one logarithm higher than that of wild-type (or plzA+), indicating 4 4 4
"
a significant attenuation in virulence (p = 0.0031) (Table 1). 4 4 5
"
4 4 6
"
The plzA mutant B. burgdorferi is unable to complete the mouse tick mouse 4 4 7
"
infection cycle. 4 4 8
"
4 4 9
"
Because infection of and survival within the mammalian host represents only one facet 4 5 0
"
of the B. burgdorferi enzootic life cycle, a more comprehensive evaluation of the 4 5 1
"
behavior of these mutants in the tick vector was warranted. Naïve Ixodes scapularis 4 5 2
"
larval ticks were allowed to feed on mice that were infected with mutant plzA (dose, 5x 4 5 3
"
106/mouse), the wild-type or the complemented strains (dose, 5x104/mouse). These 4 5 4
"
doses were chosen to infect 100% of mice with wild-type, plzA or plzA+ cells (Table 1). 4 5 5
"
Nine days after feeding on infected mice, a sub-sample of ticks was analyzed by 4 5 6
"
immunofluorescence assays (IFA) to determine the percentage of ticks infected with the 4 5 7
"
different cells (not shown; see Fig. 7). Spirochete loads in those fed-ticks were 4 5 8
"
determined using qPCR as well as by crushing ticks and plating on B. burgdorferi 4 5 9
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 1"
"
growth plates to determine viable colony forming units per tick (Table 2). The majority of 4 6 0
"
the fed larvae were allowed to molt to nymphs. As shown in Table 2, while almost 100% 4 6 1
"
of naïve ticks that fed on wild-type or the complemented plzA+ infected mice became 4 6 2
"
infected, only 55% of the ticks that fed on plzA-infected mice were positive. 4 6 3
"
Furthermore, the spirochete burden in the plzA mutant-infected ticks was >150-fold 4 6 4
"
lower than the wild-type or the isogenic complemented strains (Table 2). These results 4 6 5
"
indicate that plzA mutant cells are deficient in ability to survive in ticks. Moreover, when 4 6 6
"
the plzA-infected-nymphs were fed on naïve mice, none of the mice became infected, 4 6 7
"
whereas all naïve mice were infected with nymphs containing wild-type or the 4 6 8
"
complemented plzA+ cells (Table 2). The failure of the plzA-ticks to transmit the 4 6 9
"
infection to naïve mice was not due to the number of nymphal ticks allowed to feed on 4 7 0
"
mice. Since only 55% of plzA mutant ticks were positive (vs. ~100% for wild-type or the 4 7 1
"
plzA+), we allowed 30 plzA-nymphs to feed on a naïve mouse vs. only 10 nymphs for 4 7 2
"
wild-type or plzA+ (Table 2). Together, these results clearly demonstrate that B. 4 7 3
"
burgdorferi PilZ protein PlzA appears essential to complete the mouse-tick-mouse 4 7 4
"
infection cycle. 4 7 5
"
4 7 6
"
plzA mutant cells are deficient in surviving in artificially infected ticks. To better 4 7 7
"
determine why plzA cells in infected mice are deficient in the ability to transmit the 4 7 8
"
infection to naïve ticks, we artificially infected naïve larval ticks with wild-type, the plzA 4 7 9
"
mutant and the complemented plzA+ cells by immersion (Table 3). Tick immersion 4 8 0
"
studies (83,85,87) allows for direct artificial tick infection and serves two purposes: (a) to 4 8 1
"
optimally infect naïve ticks with the wild-type and mutant spirochetes and determine 4 8 2
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 2"
"
their colonization and survivability within the tick vector; and (b) to examine the 4 8 3
"
spirochete’s potential to migrate from arthropod host vector to the mammalian host. 4 8 4
"
While ticks inoculated with wild-type or plzA+ cells were readily transmitted to and 4 8 5
"
infected naïve mice, the plzA infected ticks failed to transmit the infection to naïve mice 4 8 6
"
(n=2) (Table 3). We found that the plzA cells were severely defective in surviving in ticks 4 8 7
"
that fed on naïve mouse although the tick’s infection rates were similar (80%) between 4 8 8
"
the wild-type and the mutant cells (Table 3). The number of viable plzA spirochetes was 4 8 9
"
very low compared to the number of viable wild-type or the complemented plzA+ (Table 4 9 0
"
3). These results indicated that a mutation in plzA resulted in cells that are deficient in 4 9 1
"
surviving in fed ticks and consequently, failed to transmit the infection from infected ticks 4 9 2
"
to naïve mice (Tables 2-3). 4 9 3
"
4 9 4
"
4 9 5
"
Discussion 4 9 6
"
4 9 7
"
C-di-GMP controls diverse cellular processes including the motility and virulence of 4 9 8
"
some bacteria (47,60,96). B. burgdorferi is an ideal organism in which to study c-di-4 9 9
"
GMP effects, in that there is no repetition of the genes that synthesize or hydrolyze c-di-5 0 0
"
GMP. The B. burgdorferi genome encodes a single protein for the GGDEF, EAL, or 5 0 1
"
HD-GYP domains (22,66,71,87), however, relatively little is known about how these 5 0 2
"
proteins function mechanistically in different cellular process. Recently, we have shown 5 0 3
"
that the EAL domain-containing phosphodiesterase BB0363 plays a role in motility and 5 0 4
"
virulence of B. burgdorferi (87). 5 0 5
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 3"
"
5 0 6
"
The c-di-GMP-binding PilZ domain proteins are important for mediating c-di-GMP 5 0 7
"
signaling that controls bacterial motility, colony morphology and effects virulence. 5 0 8
"
PlzA/BB0733 is the only recognizable PilZ domain-containing protein in B. burgdorferi 5 0 9
"
(2), and recently it has been shown to specifically bind to c-di-GMP (21). The Kd value 5 1 0
"
we determined for B. burgdorferi PilZ was 1.25 µM which is consistent with the Kd 5 1 1
"
values of other c-di-GMP binding PilZ proteins (29,37) and indicates strong affinity for c-5 1 2
"
di-GMP (Fig. 1). Cellular c-di-GMP concentrations, together with the effector 5 1 3
"
components’ affinities for c-di-GMP, are crucial for triggering many c-di-GMP-dependent 5 1 4
"
outputs (29). 5 1 5
"
5 1 6
"
Analysis of the B. burgdorferi genome as well as our RT-PCR results indicates that plzA 5 1 7
"
is transcribed monocistronically and is separated from divergently transcribed upstream 5 1 8
"
and downstream genes by 309 and 205 bp, respectively. Although genetic 5 1 9
"
manipulations in B. burgdorferi virulent, low passage strains is challenging and often 5 2 0
"
result in failure (33,66,68), we constructed a plzA mutant in the virulent wild-type strain 5 2 1
"
B31-A3-68〉bbe002 that retained all of the endogenous plasmids (Fig. 4) required for 5 2 2
"
mouse and tick infections (Tables 1-3) (63). pilZ mutants in different bacteria exhibit 5 2 3
"
different and diverse phenotypes, including but not limited to translucent, “rdar” 5 2 4
"
morphotype (rough, dry and rugose colony morphology) and altered motility 5 2 5
"
(60,67,70,98). We discovered for the first time that the B. burgdorferi plzA mutant 5 2 6
"
exhibited a translucent colony morphology consistent with other bacteria. The colony 5 2 7
"
morphology is specific to the plzA mutation since complementation in cis resulted in 5 2 8
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 4"
"
restoration of the wild-type solid/opaque colony morphology. Phase variation in other 5 2 9
"
bacteria has been described to arise from programmed genetic rearrangements or 5 3 0
"
variable expression patterns in an isogenic population (92). Molecular mechanisms 5 3 1
"
underlying the translucent colony morphology exhibited by B. burgdorferi plzA mutant 5 3 2
"
are currently unknown and require further investigation. The plzA mutant cells run-5 3 3
"
pause-reverse swimming pattern was indistinguishable from the wild-type cells (42,53). 5 3 4
"
However the swarming motility on 0.35% agarose (B. burgdorferi do not swarm on 0.5% 5 3 5
"
agarose; our unpublished results) was markedly reduced compared to the isogenic 5 3 6
"
complemented or the parental wild-type cells when plates were incubated for 7 days 5 3 7
"
(Fig. 5). We note that when swarm plates were incubated for 5 days, the swarm 5 3 8
"
diameters of the mutant strain were only slightly smaller than the wild-type cells, these 5 3 9
"
results suggest that PilZ has a positive effect on B. burgdorferi motility. 5 4 0
"
5 4 1
"
In other species of bacteria, PilZ was reported to control motility in response to changes 5 4 2
"
in the level of c-di-GMP (13,58,70). Specifically, in Caulobacter crescentus, the c-di-5 4 3
"
GMP-binding PilZ protein, DgrA, when complexed with c-di-GMP, diminished FliL 5 4 4
"
synthesis, altering motility (13). However, this is likely not to be the case in B. 5 4 5
"
burgdorferi: the levels of motility and chemotaxis proteins FliL, FliG1, FliG2, FliM, MotB, 5 4 6
"
FlaB or CheY3 were not altered in the plzA, pdeA or plzA-pdeA double mutant (see 5 4 7
"
below) (Fig. 6 and data not shown). Furthermore, in E. coli (and S. enterica) a mutation 5 4 8
"
in the phosphodiesterase yhjH reduces motility while an ycgR (encoding a PilZ protein) 5 4 9
"
mutant exhibits normal motility. Furthermore, the motility defect of the yhjH mutant was 5 5 0
"
rescued by deleting ycgR (24,58,70), indicating that YcgR mediated the reduction in 5 5 1
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 5"
"
motility due to elevated c-di-GMP (yhjH mutant). In this case, the PilZ protein YcgR 5 5 2
"
controls motility posttranslationally: in response to elevated c-di-GMP, YcgR was 5 5 3
"
reported to interact with motor switch proteins FliM, FliG or MotA, interfering with the 5 5 4
"
electrostatic interaction between the motor and switch protein controlling motility 5 5 5
"
(3,7,17,57). To determine if PlzA can function in response to c-di-GMP to regulate B. 5 5 6
"
burgdorferi motility, we inactivated plzA in a phosphodiesterase (pdeA; bb0363) mutant 5 5 7
"
background (87) where the cellular c-di-GMP level is 2x higher than the wild-type cells 5 5 8
"
(our unpublished observation). We have demonstrated that bb0363 (pdeA) mutants 5 5 9
"
exhibit reduced motility and that those mutant cells failed to reverse, whereas the wild-5 6 0
"
type cells ran-paused-reversed (87). B. burgdorferi cells do not swim well in their BSK-5 6 1
"
II growth medium, and analyzing their swimming pattern requires high-viscous medium, 5 6 2
"
such as methylcellulose (25,43,52,87). The plzA mutant constructed in the pdeA mutant 5 6 3
"
background (elevated c-di-GMP) did not rescue the pdeA cells swimming behavior: the 5 6 4
"
plzA-pdeA double mutant cells swimming pattern was indistinguishable from the pdeA 5 6 5
"
cells constant run-pause motility phenotype (87). These results suggest that PlzA 5 6 6
"
regulates B. burgdorferi motility in a different manner than it does in E. coli and that 5 6 7
"
elevated c-di-GMP in the B. burgdorferi pdeA mutant alters motility by a mechanism 5 6 8
"
independent of PlzA. It is tempting to speculate that B. burgdorferi may possess 5 6 9
"
additional c-di-GMP-binding protein(s) that responds to elevated c-di-GMP in a manner 5 7 0
"
similar to that reported in E. coli or C. crescentus to control motility (7,13,57,87). 5 7 1
"
Furthermore, our findings are consistent with the observations that PilZ proteins lacking 5 7 2
"
the N-terminal YcgR-domain are likely unable to interact with the flagellar switch FliG 5 7 3
"
protein to control bacterial motility (17). 5 7 4
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 6"
"
5 7 5
"
On the other hand, in some bacteria PilZ can act independent of c-di-GMP. For 5 7 6
"
example, V. cholerae contains five PilZ domain proteins (60), but only PlzC and PlzD 5 7 7
"
bind c-di-GMP (60). However, both plzB and plzC mutant cells were reported to exhibit 5 7 8
"
decreased motility, and a plzD mutant displayed an increased swarming motility, relative 5 7 9
"
to wild-type V. cholerae (45,60). In addition, a V. cholerae strain that produced elevated 5 8 0
"
levels of c-di-GMP exhibited reduced motility due to diminished transcription of motility 5 8 1
"
and chemotaxis genes (6,60). However, as described above, the expression of B. 5 8 2
"
burgdorferi motility or chemotaxis genes were not affected in pdeA, plzA or pdeA-plzA 5 8 3
"
mutant cells. 5 8 4
"
5 8 5
"
While the motility (Fig. 5) of plzA mutant cells was attenuated, relative to wild-type cells, 5 8 6
"
the swimming pattern of the plzA cells was indistinguishable from the wild-type cells and 5 8 7
"
expression of motility and chemotaxis genes was also unaltered. These findings may 5 8 8
"
suggest that assembly of the periplasmic flagella was not affected due to a mutation in 5 8 9
"
plzA (96). We speculate that PlzA or a PlzA-regulated factor(s) may modulate the 5 9 0
"
function of a motor protein, perhaps by modulating the MotB proton channel. A recent 5 9 1
"
study in E. coli reported that amino acid residues K53 to T64 of MotB act as a plug to 5 9 2
"
prevent proton flow before the MotA-MotB complex associates with the flagellar 5 9 3
"
structure (31,32). Because in plzA cells the swarming motility is attenuated but the 5 9 4
"
swimming pattern was not altered, it appears likely that PlzA acts as a positive regulator 5 9 5
"
in B. burgdorferi motility. 5 9 6
"
5 9 7
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 7"
"
Several studies have shown that a mutation in the gene encoding PilZ-protein affects 5 9 8
"
virulence of some bacteria. For example, a mutant in plzD of V. cholerae results in cells 5 9 9
"
that are 10-fold less infectious than the parental cells in a murine model (60). A mutation 6 0 0
"
in plant pathogen Xanthomonas campestris pv. campestris pilZ (xc3221) also resulted 6 0 1
"
in cells that are significantly less virulent in Chinese radish than their parental wild-type 6 0 2
"
cells (47). Therefore, the more than 1 log increased ID50 observed with the B. 6 0 3
"
burgdorferi plzA mutant cells during mammalian infections is consistent with these other 6 0 4
"
reports (Table 1). Although it is unknown why plzA mutant cells are significantly (p = 6 0 5
"
0.0031) less infectious than the wild-type cells, we propose that motility is likely to be a 6 0 6
"
factor and that wild-type motility is critical for B. burgdorferi infectivity ((9,43,87); our 6 0 7
"
unpublished observation). Future studies will test this hypothesis. However, we were 6 0 8
"
surprised that the plzA cells are defective in transmitting the infection from infected 6 0 9
"
mouseånaïve tickå naïve mouse (Table 2). The defect in the transmission between 6 1 0
"
tick and mammalian hosts was not due to the number of ticks allowed to feed on naïve 6 1 1
"
mice (30 plzA mutant ticks vs. 10 ticks per mouse for wild-type/plzA+). Using mouse-6 1 2
"
tick-mouse infection cycle and tick-immersion studies (Tables 2-3) we determined that 6 1 3
"
plzA cells were defective in surviving in fed ticks although the mechanism is currently 6 1 4
"
unknown (see below). Reduced motility is likely not responsible for the reduced survival 6 1 5
"
of spirochetes in ticks since motility or chemotaxis-defective mutants are able to survive 6 1 6
"
and multiply before or after a blood meal from mice ((87); our unpublished observation). 6 1 7
"
We speculate that PlzA itself is required for survivability within ticks or PlzA is a 6 1 8
"
regulator of a virulence determinant(s) impacting survivability in ticks and/or attenuation 6 1 9
"
in mice. It is also possible that the translucence colony morphology of the plzA mutant 6 2 0
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 8"
"
cells may reflect an altered cell surface membrane structure, which may be related to 6 2 1
"
the impaired ability of plzA cells to infect mice and/or decreased survivability within 6 2 2
"
ticks. Tick saliva contains bioactive molecules and a plethora of proteolytic enzymes 6 2 3
"
(19,75). Whether tick-derived elements, host-derived products acquired from tick’s 6 2 4
"
bloodmeal, or salivary glands factors (18,19,34,75,81,82) were responsible for the 6 2 5
"
reduced survival of the plzA mutant in the ticks remains to be determined. We and 6 2 6
"
others have shown that B. burgdorferi uses different mechanism to survive in different 6 2 7
"
hosts (ticks/mammalian). Future studies are warranted to decipher the wide-spectrum of 6 2 8
"
PilZ effects on B. burgdorferi. 6 2 9
"
6 3 0
"
6 3 1
"
Acknowledgement 6 3 2
"
6 3 3
"
We thank Rita Tamayo and Philip Stewart for comments on the manuscript. We would 6 3 4
"
like to thank R. Rego, P. Rosa, C. Li, J. Benach, A. Barbour, J. Carroll, M. Gomelsky, D. 6 3 5
"
Blair, U. Pal, E. Fikrig and S. Samuels for providing reagents. We appreciate technical 6 3 6
"
discussions from P. Policastro and A. Bestor, RML, NIH. The research is sponsored by 6 3 7
"
a National Institute of Arthritis, Musculoskeletal, and Skin diseases, NIH Grant 6 3 8
"
R03AR054582 and an East Carolina University Research and Development ‘start-up’ 6 3 9
"
fund to M.A.M. 6 4 0
"
6 4 1
"
6 4 2
"
"6 4 3
"6 4 4"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

2 9"
"
Reference List 6 4 5
"
6 4 6
"
1. Alm, R. A., A. J. Bodero, P. D. Free, and J. S. Mattick. 1996. Identification of a 6 4 7
"novel gene, pilZ, essential for type 4 fimbrial biogenesis in Pseudomonas
6 4 8"
aeruginosa. J. Bacteriol. 178:46-53. 6 4 9
"
2. Amikam, D. and M. Y. Galperin. 2006. PilZ domain is part of the bacterial c-di-6 5 0
"GMP binding protein. Bioinformatics. 22:3-6.
6 5 1"
3. Armitage, J. P. and R. M. Berry. 2010. Time for bacteria to slow down. Cell 6 5 2
"141:24-26.
6 5 3"
4. Battisti, J. M., J. L. Bono, P. A. Rosa, M. E. Schrumpf, T. G. Schwan, and P. 6 5 4
"F. Policastro. 2008. Outer surface protein A protects Lyme disease spirochetes
6 5 5"
from acquired host immunity in the tick vector. Infect. Immun. 76:5228-5237. 6 5 6
"
5. Benach, J., S. S. Swaminathan, R. Tamayo, S. K. Handelman, E. Folta-6 5 7
"Stogniew, J. E. Ramos, F. Forouhar, H. Neely, J. Seetharaman, A. Camilli,
6 5 8"
and J. F. Hunt. 2007. The structural basis of cyclic diguanylate signal 6 5 9
"transduction by PilZ domains. EMBO J. 26:5153-5166.
6 6 0"
6. Beyhan, S., A. D. Tischler, A. Camilli, and F. H. Yildiz. 2006. Transcriptome 6 6 1
"and phenotypic responses of Vibrio cholerae to increased cyclic di-GMP level. J.
6 6 2"
Bacteriol. 188:3600-3613. 6 6 3
"
7. Boehm, A., M. Kaiser, H. Li, C. Spangler, C. A. Kasper, M. Ackermann, V. 6 6 4
"Kaever, V. Sourjik, V. Roth, and U. Jenal. 2010. Second Messenger-Mediated
6 6 5"
Adjustment of Bacterial Swimming Velocity. Cell 141:107-116. 6 6 6
"
8. Bono, J. L., A. F. Elias, J. J. Kupko III, B. Stevenson, K. Tilly, and P. Rosa. 6 6 7
"2000. Efficient targeted mutagenesis in Borrelia burgdorferi. J. Bacteriol.
6 6 8"
182:2445-2452. 6 6 9
"
9. Botkin, D. J., A. N. Abbott, P. E. Stewart, P. A. Rosa, H. Kawabata, H. 6 7 0
"Watanabe, and S. J. Norris. 2006. Identification of potential virulence
6 7 1"
determinants by Himar1 transposition of infectious Borrelia burgdorferi B31. 6 7 2
"Infect. Immun. 74:6690-6699.
6 7 3"
10. Brooks, C. S., P. S. Hefty, S. E. Jolliff, and D. R. Akins. 2003. Global analysis 6 7 4
"of Borrelia burgdorferi genes regulated by mammalian host-specific signals.
6 7 5"
Infect. Immun. 71:3371-3383. 6 7 6
"
11. Carbonnelle, E., S. Helaine, L. Prouvensier, X. Nassif, and V. Pelicic. 2005. 6 7 7
"Type IV pilus biogenesis in Neisseria meningitidis: PilW is involved in a step
6 7 8"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 0"
"
occurring after pilus assembly, essential for fibre stability and function. Mol. 6 7 9
"Microbiol. 55:54-64.
6 8 0"
12. Casjens, S., N. Palmer, R. van Vugt, W. M. Huang, B. Stevenson, P. Rosa, R. 6 8 1
"Lathigra, G. Sutton, J. Peterson, R. J. Dodson, D. Haft, E. Hickey, M. Gwinn,
6 8 2"
O. White, and C. M. Fraser. 2000. A bacterial genome in flux: the twelve linear 6 8 3
"and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme
6 8 4"
disease spirochete Borrelia burgdorferi. Mol. Microbiol. 35:490-516. 6 8 5
"
13. Christen, M., B. Christen, M. G. Allan, M. Folcher, P. Jeno, S. Grzesiek, and 6 8 6
"U. Jenal. 2007. DgrA is a member of a new family of cyclic diguanosine
6 8 7"
monophosphate receptors and controls flagellar motor function in Caulobacter 6 8 8
"crescentus. Proc. Natl. Acad. Sci. U. S. A 104:4112-4117.
6 8 9"
14. Cotter, P. A. and S. Stibitz. 2007. c-di-GMP-mediated regulation of virulence 6 9 0
"and biofilm formation. Curr. Opin. Microbiol. 10:17-23.
6 9 1"
15. Duerig, A., S. Abel, M. Folcher, M. Nicollier, T. Schwede, N. Amiot, B. Giese, 6 9 2
"and U. Jenal. 2009. Second messenger-mediated spatiotemporal control of
6 9 3"
protein degradation regulates bacterial cell cycle progression. Genes Dev. 23:93-6 9 4
"104.
6 9 5"
16. Elias, A. F., P. E. Stewart, D. Grimm, M. J. Caimano, C. H. Eggers, K. Tilly, J. 6 9 6
"L. Bono, D. R. Akins, J. D. Radolf, T. G. Schwan, and P. Rosa. 2002. Clonal
6 9 7"
polymorphism of Borrelia burgdorferi strain B31 MI: Implications for mutagenesis 6 9 8
"in an infectious strain background. Infect. Immun. 70:2139-2150.
6 9 9"
17. Fang, X. and M. Gomelsky. 2010. A posttranslational, c-di-GMP-dependent 7 0 0
"mechanism regulating flagellar motility. Mol. Microbiol. 76:1295-1305.
7 0 1"
18. Fogaca, A. C., S. P. da, Jr., M. T. Miranda, A. G. Bianchi, A. Miranda, P. E. 7 0 2
"Ribolla, and S. Daffre. 1999. Antimicrobial activity of a bovine hemoglobin
7 0 3"
fragment in the tick Boophilus microplus. J. Biol. Chem. 274:25330-25334. 7 0 4
"
19. Francischetti, I. M., A. Sa-Nunes, B. J. Mans, I. M. Santos, and J. M. Ribeiro. 7 0 5
"2009. The role of saliva in tick feeding. Front Biosci. 14:2051-2088.
7 0 6"
20. Fraser, C. M., S. Casjens, W. M. Huang, G. G. Sutton, R. Clayton, R. 7 0 7
"Lathigra, O. White, K. A. Ketchum, R. Dodson, E. K. Hickey, M. Gwinn, B.
7 0 8"
Dougherty, J. F. Tomb, R. D. Fleischmann, D. Richardson, J. Peterson, A. R. 7 0 9
"Kerlavage, J. Quackenbush, S. Salzberg, M. Hanson, R. van Vugt, N.
7 1 0"
Palmer, M. D. Adams, and J. Gocayne. 1997. Genomic sequence of a Lyme 7 1 1
"disease spirochaete, Borrelia burgdorferi. Nature 390:580-586.
7 1 2"
21. Freedman, J. C., E. A. Rogers, J. L. Kostick, H. Zhang, R. Iyer, I. Schwartz, 7 1 3
"and R. T. Marconi. 2009. Identification and molecular characterization of a
7 1 4"
cyclic-di-GMP effector protein, PlzA (BB0733): additional evidence for the 7 1 5
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 1"
"
existence of a functional cyclic-di-GMP regulatory network in the Lyme disease 7 1 6
"spirochete, Borrelia burgdorferi. FEMS Immunol. Med. Microbiol. 58:285-294.
7 1 7"
22. Galperin, M. Y., A. N. Nikolskaya, and E. V. Koonin. 2001. Novel domains of 7 1 8
"the prokaryotic two-component signal transduction systems. FEMS Microbiol.
7 1 9"
Lett. 203:11-21. 7 2 0
"
23. Ge, Y., C. Li, L. Corum, C. A. Slaughter, and N. W. Charon. 1998. Structure 7 2 1
"and expression of the FlaA periplasmic flagellar protein of Borrelia burgdorferi. J.
7 2 2"
Bacteriol. 180:2418-2425. 7 2 3
"
24. Girgis, H. S., Y. Liu, W. S. Ryu, and S. Tavazoie. 2007. A comprehensive 7 2 4
"genetic characterization of bacterial motility. PLoS. Genet. 3:1644-1660.
7 2 5"
25. Goldstein, S. F., N. W. Charon, and J. A. Kreiling. 1994. Borrelia burgdorferi 7 2 6
"swims with a planar waveform similar to that of eukaryotic flagella. Proc. Natl.
7 2 7"
Acad. Sci. U. S. A. 91:3433-3437. 7 2 8
"
26. Grimm, D., A. F. Elias, K. Tilly, and P. A. Rosa. 2003. Plasmid stability during 7 2 9
"in vitro propagation of Borrelia burgdorferi assessed at a clonal level. Infect.
7 3 0"
Immun. 71:3138-3145. 7 3 1
"
27. Guzzo, C. R., R. K. Salinas, M. O. Andrade, and C. S. Farah. 2009. PILZ 7 3 2
"protein structure and interactions with PILB and the FIMX EAL domain:
7 3 3"
implications for control of type IV pilus biogenesis. J. Mol. Biol. 393:848-866. 7 3 4
"
28. Heidelberg, J. F., I. T. Paulsen, K. E. Nelson, E. J. Gaidos, W. C. Nelson, T. 7 3 5
"D. Read, J. A. Eisen, R. Seshadri, N. Ward, B. Methe, R. A. Clayton, T.
7 3 6"
Meyer, A. Tsapin, J. Scott, M. Beanan, L. Brinkac, S. Daugherty, R. T. 7 3 7
"DeBoy, R. J. Dodson, A. S. Durkin, D. H. Haft, J. F. Kolonay, R. Madupu, J.
7 3 8"
D. Peterson, L. A. Umayam, O. White, A. M. Wolf, J. Vamathevan, J. 7 3 9
"Weidman, M. Impraim, K. Lee, K. Berry, C. Lee, J. Mueller, H. Khouri, J. Gill,
7 4 0"
T. R. Utterback, L. A. McDonald, T. V. Feldblyum, H. O. Smith, J. C. Venter, 7 4 1
"K. H. Nealson, and C. M. Fraser. 2002. Genome sequence of the dissimilatory
7 4 2"
metal ion-reducing bacterium Shewanella oneidensis. Nat. Biotechnol. 20:1118-7 4 3
"1123.
7 4 4"
29. Hengge, R. 2009. Principles of c-di-GMP signalling in bacteria. Nat. Rev. 7 4 5
"Microbiol. 7:263-273.
7 4 6"
30. Hickman, J. W. and C. S. Harwood. 2008. Identification of FleQ from 7 4 7
"Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol.
7 4 8"
Microbiol. 69:376-389. 7 4 9
"
31. Hosking, E. R. and M. D. Manson. 2008. Clusters of charged residues at the C 7 5 0
"terminus of MotA and N terminus of MotB are important for function of the
7 5 1"
Escherichia coli flagellar motor. J. Bacteriol. 190:5517-5521. 7 5 2
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 2"
"
32. Hosking, E. R., C. Vogt, E. P. Bakker, and M. D. Manson. 2006. The 7 5 3
"Escherichia coli MotAB proton channel unplugged. J. Mol. Biol. 364:921-937.
7 5 4"
33. Hyde, J. A., D. K. Shaw, R. Smith, III, J. P. Trzeciakowski, and J. T. Skare. 7 5 5
"2009. Characterization of a Conditional bosR Mutant in Borrelia burgdorferi.
7 5 6"
Infect. Immun. 78:265-274. 7 5 7
"
34. Hynes, W. L., S. M. Ceraul, S. M. Todd, K. C. Seguin, and D. E. Sonenshine. 7 5 8
"2005. A defensin-like gene expressed in the black-legged tick, Ixodes scapularis.
7 5 9"
Med. Vet. Entomol. 19:339-344. 7 6 0
"
35. Jewett, M. W., K. Lawrence, A. C. Bestor, K. Tilly, D. Grimm, P. Shaw, M. 7 6 1
"VanRaden, F. Gherardini, and P. A. Rosa. 2007. The critical role of the linear
7 6 2"
plasmid lp36 in the infectious cycle of Borrelia burgdorferi. Mol. Microbiol. 7 6 3
"64:1358-1374.
7 6 4"
36. Kawabata, H., S. J. Norris, and H. Watanabe. 2004. BBE02 disruption mutants 7 6 5
"of Borrelia burgdorferi B31 have a highly transformable, infectious phenotype.
7 6 6"
Infect. Immun. 72:7147-7154. 7 6 7
"
37. Ko, J., K. S. Ryu, H. Kim, J. S. Shin, J. O. Lee, C. Cheong, and B. S. Choi. 7 6 8
"2010. Structure of PP4397 reveals the molecular basis for different c-di-GMP
7 6 9"
binding modes by Pilz domain proteins. J. Mol. Biol. 398:97-110. 7 7 0
"
38. Kulasakara, H., V. Lee, A. Brencic, N. Liberati, J. Urbach, S. Miyata, D. G. 7 7 1
"Lee, A. N. Neely, M. Hyodo, Y. Hayakawa, F. M. Ausubel, and S. Lory. 2006.
7 7 2"
Analysis of Pseudomonas aeruginosa diguanylate cyclases and 7 7 3
"phosphodiesterases reveals a role for bis-(3'-5')-cyclic-GMP in virulence. Proc.
7 7 4"
Natl. Acad. Sci. U. S. A 103:2839-2844. 7 7 5
"
39. Kumar, M. and D. Chatterji. 2008. Cyclic di-GMP: a second messenger required 7 7 6
"for long-term survival, but not for biofilm formation, in Mycobacterium smegmatis.
7 7 7"
Microbiol. 154:2942-2955. 7 7 8
"
40. Leduc, J. L. and G. P. Roberts. 2009. Cyclic di-GMP allosterically inhibits the 7 7 9
"CRP-like protein (Clp) of Xanthomonas axonopodis pv. citri. J. Bacteriol.
7 8 0"
191:7121-7122. 7 8 1
"
41. Lee, V. T., J. M. Matewish, J. L. Kessler, M. Hyodo, Y. Hayakawa, and S. 7 8 2
"Lory. 2007. A cyclic-di-GMP receptor required for bacterial exopolysaccharide
7 8 3"
production. Mol. Microbiol. 65:1474-1484. 7 8 4
"
42. Li, C., R. G. Bakker, M. A. Motaleb, M. L. Sartakova, F. C. Cabello, and N. W. 7 8 5
"Charon. 2002. Asymmetrical flagellar rotation in Borrelia burgdorferi
7 8 6"
nonchemotactic mutants. Proc. Natl. Acad. Sci. U. S. A. 99:6169-6174. 7 8 7
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 3"
"
43. Li, C., H. Xu, K. Zhang, and F. T. Liang. 2010. Inactivation of a putative flagellar 7 8 8
"motor switch protein FliG1 prevents Borrelia burgdorferi from swimming in highly
7 8 9"
viscous media and blocks its infectivity. Mol. Microbiol. 75:1563-1576. 7 9 0
"
44. Li, X., U. Pal, N. Ramamoorthi, X. Liu, D. C. Desrosiers, C. H. Eggers, J. F. 7 9 1
"Anderson, J. D. Radolf, and E. Fikrig. 2007. The Lyme disease agent Borrelia
7 9 2"
burgdorferi requires BB0690, a Dps homologue, to persist within ticks. Mol. 7 9 3
"Microbiol. 63:694-710.
7 9 4"
45. Liu, X., S. Beyhan, B. Lim, R. G. Linington, and F. H. Yildiz. 2010. 7 9 5
"Identification and Characterization of a Phosphodiesterase that Inversely
7 9 6"
Regulates Motility and Biofilm Formation in Vibrio cholerae. J. Bacteriol. 7 9 7
"192:4541-4552.
7 9 8"
46. Livak, K. J. and T. D. Schmittgen. 2001. Analysis of relative gene expression 7 9 9
"data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method.
8 0 0"
Methods 25:402-408. 8 0 1
"
47. McCarthy, Y., R. P. Ryan, K. O'Donovan, Y. Q. He, B. L. Jiang, J. X. Feng, J. 8 0 2
"L. Tang, and J. M. Dow. 2008. The role of PilZ domain proteins in the virulence
8 0 3"
of Xanthomonas campestris pv. campestris. Mol. Plant Pathol. 9:819-824. 8 0 4
"
48. Merighi, M., V. T. Lee, M. Hyodo, Y. Hayakawa, and S. Lory. 2007. The 8 0 5
"second messenger bis-(3'-5')-cyclic-GMP and its PilZ domain-containing receptor
8 0 6"
Alg44 are required for alginate biosynthesis in Pseudomonas aeruginosa. Mol. 8 0 7
"Microbiol. 65:876-895.
8 0 8"
49. Methe, B. A., K. E. Nelson, J. W. Deming, B. Momen, E. Melamud, X. Zhang, 8 0 9
"J. Moult, R. Madupu, W. C. Nelson, R. J. Dodson, L. M. Brinkac, S. C.
8 1 0"
Daugherty, A. S. Durkin, R. T. DeBoy, J. F. Kolonay, S. A. Sullivan, L. Zhou, 8 1 1
"T. M. Davidsen, M. Wu, A. L. Huston, M. Lewis, B. Weaver, J. F. Weidman, H.
8 1 2"
Khouri, T. R. Utterback, T. V. Feldblyum, and C. M. Fraser. 2005. The 8 1 3
"psychrophilic lifestyle as revealed by the genome sequence of Colwellia
8 1 4"
psychrerythraea 34H through genomic and proteomic analyses. Proc. Natl. Acad. 8 1 5
"Sci. U. S. A 102:10913-10918.
8 1 6"
50. Monds, R. D., P. D. Newell, R. H. Gross, and G. A. O'Toole. 2007. Phosphate-8 1 7
"dependent modulation of c-di-GMP levels regulates Pseudomonas fluorescens
8 1 8"
Pf0-1 biofilm formation by controlling secretion of the adhesin LapA. Mol. 8 1 9
"Microbiol. 63:656-679.
8 2 0"
51. Motaleb, M. A., L. Corum, J. L. Bono, A. F. Elias, P. Rosa, D. S. Samuels, 8 2 1
"and N. W. Charon. 2000. Borrelia burgdorferi periplasmic flagella have both
8 2 2"
skeletal and motility functions. Proc. Natl. Acad. Sci. U. S. A 97:10899-10904. 8 2 3
"
52. Motaleb, M. A., M. R. Miller, R. G. Bakker, C. Li, and N. W. Charon. 2007. 8 2 4
"Isolation and characterization of chemotaxis mutants of the Lyme disease
8 2 5"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 4"
"
Spirochete Borrelia burgdorferi using allelic exchange mutagenesis, flow 8 2 6
"cytometry, and cell tracking. Methods Enzymol. 422:421-437.
8 2 7"
53. Motaleb, M. A., M. R. Miller, C. Li, R. G. Bakker, S. F. Goldstein, R. E. 8 2 8
"Silversmith, R. B. Bourret, and N. W. Charon. 2005. CheX is a phosphorylated
8 2 9"
CheY phosphatase essential for Borrelia burgdorferi chemotaxis. J. Bacteriol. 8 3 0
"187:7963-7969.
8 3 1"
54. Motaleb, M. A., M. S. Sal, and N. W. Charon. 2004. The decrease in FlaA 8 3 2
"observed in a flaB mutant of Borrelia burgdorferi occurs posttranscriptionally. J.
8 3 3"
Bacteriol. 186:3703-3711. 8 3 4
"
55. Ojaimi, C., C. Brooks, S. Casjens, P. Rosa, A. Elias, A. Barbour, A. 8 3 5
"Jasinskas, J. Benach, L. Katona, J. Radolf, M. Caimano, J. Skare, K.
8 3 6"
Swingle, D. Akins, and I. Schwartz. 2003. Profiling of temperature-induced 8 3 7
"changes in Borrelia burgdorferi gene expression by using whole genome arrays.
8 3 8"
Infect. Immun. 71:1689-1705. 8 3 9
"
56. Ouyang, Z., M. Kumar, T. Kariu, S. Haq, M. Goldberg, U. Pal, and M. V. 8 4 0
"Norgard. 2009. BosR (BB0647) governs virulence expression in Borrelia
8 4 1"
burgdorferi. Mol. Microbiol. 74:1331-1343. 8 4 2
"
57. Paul, K., V. Nieto, W. C. Carlquist, D. F. Blair, and R. M. Harshey. 2010. The 8 4 3
"c-di-GMP Binding Protein YcgR Controls Flagellar Motor Direction and Speed to
8 4 4"
Affect Chemotaxis by a "Backstop Brake" Mechanism. Mol. Cell 38:128-139. 8 4 5
"
58. Pesavento, C., G. Becker, N. Sommerfeldt, A. Possling, N. Tschowri, A. 8 4 6
"Mehlis, and R. Hengge. 2008. Inverse regulatory coordination of motility and
8 4 7"
curli-mediated adhesion in Escherichia coli. Genes Dev. 22:2434-2446. 8 4 8
"
59. Policastro, P. F. and T. G. Schwan. 2003. Experimental infection of Ixodes 8 4 9
"scapularis larvae (Acari: Ixodidae) by immersion in low passage cultures of
8 5 0"
Borrelia burgdorferi. J. Med. Entomol. 40:364-370. 8 5 1
"
60. Pratt, J. T., R. Tamayo, A. D. Tischler, and A. Camilli. 2007. PilZ domain 8 5 2
"proteins bind cyclic diguanylate and regulate diverse processes in Vibrio
8 5 3"
cholerae. J. Biol. Chem. 282:12860-12870. 8 5 4
"
61. Purser, J. E. and S. J. Norris. 2000. Correlation between plasmid content and 8 5 5
"infectivity in Borrelia burgdorferi. Proc. Natl. Acad. Sci. U. S. A 97:13865-13870.
8 5 6"
62. Reed, L. J. and H. Muench. 1938. A simple method of estimating fifty percent 8 5 7
"endpoint. Am. J. Hyg. 27: 493-497.
8 5 8"
63. Rego, R. O., A. Bestor, and P. A. Rosa. 2011. Defining the plasmid-encoded 8 5 9
"restriction-modification systems of the Lyme disease spirochete Borrelia
8 6 0"
burgdorferi. J. Bacteriol. 193: 1161-71. 8 6 1
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 5"
"
64. Revel, A. T., J. S. Blevins, C. Almazan, L. Neil, K. M. Kocan, F. J. de la, K. E. 8 6 2
"Hagman, and M. V. Norgard. 2005. bptA (bbe16) is essential for the persistence
8 6 3"
of the Lyme disease spirochete, Borrelia burgdorferi, in its natural tick vector. 8 6 4
"Proc. Natl. Acad. Sci. U. S. A 102:6972-6977.
8 6 5"
65. Revel, A. T., A. M. Talaat, and M. V. Norgard. 2002. DNA microarray analysis 8 6 6
"of differential gene expression in Borrelia burgdorferi, the Lyme disease
8 6 7"
spirochete. Proc. Natl. Acad. Sci. U. S. A 99:1562-1567. 8 6 8
"
66. Rogers, E. A., D. Terekhova, H. M. Zhang, K. M. Hovis, I. Schwartz, and R. T. 8 6 9
"Marconi. 2009. Rrp1, a cyclic-di-GMP-producing response regulator, is an
8 7 0"
important regulator of Borrelia burgdorferi core cellular functions. Mol. Microbiol. 8 7 1
"71:1551-1573.
8 7 2"
67. Romling, U. 2005. Characterization of the rdar morphotype, a multicellular 8 7 3
"behaviour in Enterobacteriaceae. Cell Mol. Life Sci. 62:1234-1246.
8 7 4"
68. Rosa, P. A., K. Tilly, and P. E. Stewart. 2005. The burgeoning molecular 8 7 5
"genetics of the Lyme disease spirochaete. Nat. Rev. Microbiol. 3:129-143.
8 7 6"
69. Ross, P., H. Weinhouse, Y. Aloni, D. Michaeli, P. Weinberger-Ohana, R. 8 7 7
"Mayer, S. Braun, V. E. de, G. A. van der Marel, J. H. van Boom, and M.
8 7 8"
Benziman. 1987. Regulation of cellulose synthesis in Acetobacter xylinum by 8 7 9
"cyclic diguanylic acid. Nature 325:279-281.
8 8 0"
70. Ryjenkov, D. A., R. Simm, U. Romling, and M. Gomelsky. 2006. The PilZ 8 8 1
"domain is a receptor for the second messenger c-di-GMP: the PilZ domain
8 8 2"
protein YcgR controls motility in enterobacteria. J. Biol. Chem. 281:30310-30314. 8 8 3
"
71. Ryjenkov, D. A., M. Tarutina, O. V. Moskvin, and M. Gomelsky. 2005. Cyclic 8 8 4
"diguanylate is a ubiquitous signaling molecule in bacteria: insights into
8 8 5"
biochemistry of the GGDEF protein domain. J. Bacteriol. 187:1792-1798. 8 8 6
"
72. Sal, M. S., C. Li, M. A. Motalab, S. Shibata, S. Aizawa, and N. W. Charon. 8 8 7
"2008. Borrelia burgdorferi uniquely regulates its motility genes and has an
8 8 8"
intricate flagellar hook-basal body structure. J. Bacteriol. 190:1912-1921. 8 8 9
"
73. Samuels, D. S., K. E. Mach, and C. F. Garon. 1994. Genetic transformation of 8 9 0
"the Lyme disease agent Borrelia burgdorferi with coumarin-resistant gyrB. J.
8 9 1"
Bacteriol. 176:6045-6049. 8 9 2
"
74. Samuels, D. S. and J. D. Radolf. 2009. Who is the BosR around here anyway? 8 9 3
"Mol. Microbiol. 74:1295-1299.
8 9 4"
75. Sauer, J. R., J. L. McSwain, A. S. Bowman, and R. C. Essenberg. 1995. Tick 8 9 5
"salivary gland physiology. Annu. Rev. Entomol. 40:245-267.
8 9 6"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 6"
"
76. Schirmer, T. and U. Jenal. 2009. Structural and mechanistic determinants of c-8 9 7
"di-GMP signalling. Nat. Rev. Microbiol. 7:724-735.
8 9 8"
77. Seshu, J., M. D. Esteve-Gassent, M. Labandeira-Rey, J. H. Kim, J. P. 8 9 9
"Trzeciakowski, M. Hook, and J. T. Skare. 2006. Inactivation of the fibronectin-
9 0 0"
binding adhesin gene bbk32 significantly attenuates the infectivity potential of 9 0 1
"Borrelia burgdorferi. Mol. Microbiol. 59:1591-1601.
9 0 2"
78. Simm, R., M. Morr, U. Remminghorst, M. Andersson, and U. Romling. 2009. 9 0 3
"Quantitative determination of cyclic diguanosine monophosphate concentrations
9 0 4"
in nucleotide extracts of bacteria by matrix-assisted laser desorption/ionization-9 0 5
"time-of-flight mass spectrometry. Anal. Biochem. 386:53-58.
9 0 6"
79. Simm, R., U. Remminghorst, I. Ahmad, K. Zakikhany, and U. Romling. 2009. 9 0 7
"A role for the EAL-like protein STM1344 in regulation of CsgD expression and
9 0 8"
motility in Salmonella enterica serovar Typhimurium. J. Bacteriol. 191:3928-3937. 9 0 9
"
80. Simpson, W. J., W. Burgdorfer, M. E. Schrumpf, R. H. Karstens, and T. G. 9 1 0
"Schwan. 1991. Antibody to a 39-kilodalton Borrelia burgdorferi antigen (P39) as
9 1 1"
a marker for infection in experimentally and naturally inoculated animals. J. Clin. 9 1 2
"Microbiol. 29:236-243.
9 1 3"
81. Sonenshine, D. E., S. M. Ceraul, W. E. Hynes, K. R. Macaluso, and A. F. 9 1 4
"Azad. 2002. Expression of defensin-like peptides in tick hemolymph and midgut
9 1 5"
in response to challenge with Borrelia burgdorferi, Escherichia coli and Bacillus 9 1 6
"subtilis. Exp. Appl. Acarol. 28:127-134.
9 1 7"
82. Sonenshine, D. E. and W. L. Hynes. 2008. Molecular characterization and 9 1 8
"related aspects of the innate immune response in ticks. Front Biosci. 13:7046-
9 1 9"
7063. 9 2 0
"
83. Stewart, P. E., A. Bestor, J. N. Cullen, and P. A. Rosa. 2008. A tightly 9 2 1
"regulated surface protein of Borrelia burgdorferi is not essential to the mouse-tick
9 2 2"
infectious cycle. Infect. Immun. 76:1970-1978. 9 2 3
"
84. Stock, A. M., V. L. Robinson, and P. N. Goudreau. 2000. Two-component 9 2 4
"signal transduction. Annu. Rev. Biochem. 69:183-215.
9 2 5"
85. Strother, K. O. and S. A. de. 2005. Role of Borrelia burgdorferi linear plasmid 25 9 2 6
"in infection of Ixodes scapularis ticks. J. Bacteriol. 187:5776-5781.
9 2 7"
86. Sudarsan, N., E. R. Lee, Z. Weinberg, R. H. Moy, J. N. Kim, K. H. Link, and R. 9 2 8
"R. Breaker. 2008. Riboswitches in eubacteria sense the second messenger
9 2 9"
cyclic di-GMP. Science 321:411-413. 9 3 0
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 7"
"
87. Sultan, S. Z., J. E. Pitzer, M. R. Miller, and M. A. Motaleb. 2010. Analysis of a 9 3 1
"Borrelia burgdorferi phosphodiesterase demonstrates a role for cyclic-di-
9 3 2"
guanosine monophosphate in motility and virulence. Mol. Microbiol. 77:128-142. 9 3 3
"
88. Tamayo, R., J. T. Pratt, and A. Camilli. 2007. Roles of cyclic diguanylate in the 9 3 4
"regulation of bacterial pathogenesis. Annu. Rev. Microbiol. 61:131-148.
9 3 5"
89. Tilly, K., J. G. Krum, A. Bestor, M. W. Jewett, D. Grimm, D. Bueschel, R. 9 3 6
"Byram, D. Dorward, M. J. Vanraden, P. Stewart, and P. Rosa. 2006. Borrelia
9 3 7"
burgdorferi OspC protein required exclusively in a crucial early stage of 9 3 8
"mammalian infection. Infect. Immun. 74:3554-3564.
9 3 9"
90. Tilly, K., P. A. Rosa, and P. E. Stewart. 2008. Biology of infection with Borrelia 9 4 0
"burgdorferi. Infect. Dis. Clin. North Am. 22:217-234.
9 4 1"
91. Tokarz, R., J. M. Anderton, L. I. Katona, and J. L. Benach. 2004. Combined 9 4 2
"effects of blood and temperature shift on Borrelia burgdorferi gene expression as
9 4 3"
determined by whole genome DNA array. Infect. Immun. 72:5419-5432. 9 4 4
"
92. van der Woude, M. W. and A. J. Baumler. 2004. Phase and antigenic variation 9 4 5
"in bacteria. Clin. Microbiol. Rev. 17:581-611.
9 4 6"
93. Weber, H., C. Pesavento, A. Possling, G. Tischendorf, and R. Hengge. 2006. 9 4 7
"Cyclic-di-GMP-mediated signalling within the sigma network of Escherichia coli.
9 4 8"
Mol. Microbiol. 62:1014-1034. 9 4 9
"
94. Weinhouse, H., S. Sapir, D. Amikam, Y. Shilo, G. Volman, P. Ohana, and M. 9 5 0
"Benziman. 1997. c-di-GMP-binding protein, a new factor regulating cellulose
9 5 1"
synthesis in Acetobacter xylinum. FEBS Lett. 416:207-211. 9 5 2
"
95. West, A. H. and A. M. Stock. 2001. Histidine kinases and response regulator 9 5 3
"proteins in two-component signaling systems. Trends Biochem. Sci. 26:369-376.
9 5 4"
96. Wolfe, A. J. and K. L. Visick. 2008. Get the message out: cyclic-Di-GMP 9 5 5
"regulates multiple levels of flagellum-based motility. J. Bacteriol. 190:463-475.
9 5 6"
97. Yang, X. F., U. Pal, S. M. Alani, E. Fikrig, and M. V. Norgard. 2004. Essential 9 5 7
"role for OspA/B in the life cycle of the Lyme disease spirochete. J. Exp. Med.
9 5 8"
199:641-648. 9 5 9
"
98. Yildiz, F. H. and K. L. Visick. 2009. Vibrio biofilms: so much the same yet so 9 6 0
"different. Trends Microbiol. 17:109-118.
9 6 1"
99. Zhang, X., X. Yang, M. Kumar, and U. Pal. 2009. BB0323 function is essential 9 6 2
"for Borrelia burgdorferi virulence and persistence through tick-rodent
9 6 3"
transmission cycle. J. Infect. Dis. 200:1318-1330. 9 6 4
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 8"
"
100. Zhou, X., M. R. Miller, M. Motaleb, N. W. Charon, and P. He. 2008. Spent 9 6 5
"culture medium from virulent Borrelia burgdorferi increases permeability of
9 6 6"
individually perfused microvessels of rat mesentery. PLoS. ONE. 3:e4101. 9 6 7
"
9 6 8"
9 6 9
"9 7 0"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

3 9"
"
Figure legend 9 7 1
"
9 7 2
"
Fig. 1. Left panel, SDS-PAGE analysis of expressed His6-BB0733/PlzA recombinant 9 7 3
"
protein from E. coli. Lane 1, protein size marker with their sizes in kDa. Lanes 2 and 3, 9 7 4
"
uninduced and induced E. coli lysates, respectively. Lane 4, His6-PlzA protein after 9 7 5
"
purification in a Ni2+-agarose column. Arrowhead indicates the His6-PlzA (~32 kDa) 9 7 6
"
protein band. Right panel, plot of equilibrium binding between PlzA and synthetic c-di-9 7 7
"
GMP (cdG). Equilibrium dialysis was performed as described above, and the binding 9 7 8
"
constant was determined using GraphPad Prism 5 (one site, specific binding). Results 9 7 9
"
with GTP, cAMP or cGMP were not shown since no binding with His6-PlzA was 9 8 0
"
detected. 9 8 1
"
9 8 2
"
Fig. 2. Inactivation and complementation of bb0733 (plzA). (Top) In-frame insertion of 9 8 3
"
the Pl-Kan cassette within bb0733 gene by homologous recombination resulted in the 9 8 4
"
creation of plzA mutant. Approximate locations of kan and bb0733 primer sets are 9 8 5
"
shown by thin arrows that were used to confirm the bb0733 inactivation by PCR (see 9 8 6
"
Fig. 3). (Bottom) Complementation plasmid pXLFbb0733 containing the bb0733 9 8 7
"
promoter (PplzA) that drives the expression of bb0733 (plzA) gene was used to 9 8 8
"
complement the plzA mutant in cis by genomic integration in a region where genes 9 8 9
"
bb0445 and bb0446 converge. Approximate locations of primer sets (bb0445-bb0446 9 9 0
"
and Gent) used to confirm the integration of PplzA-PlzA-PflgB-aacC1 was shown using 9 9 1
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4 0"
"
thin horizontal arrows. PCR primer products of each of these primer set is shown in Fig. 9 9 2
"
3. 9 9 3
"
9 9 4
"
Fig. 3. Confirmation of plzA inactivation, complementation in cis, and restoration of plzA 9 9 5
"
transcript synthesis. Left panel - PCR was used to confirm the integration of Pl-Kan 9 9 6
"
within the bb0733 (plzA) gene. Location of PCR primers were shown in Fig. 2. An 892 9 9 7
"
bp DNA containing the Pl-Kan cassette was inserted after deleting a 509 bp fragment 9 9 8
"
from the plzA gene. Sizes of PCR products for lanes plzA and positive ‘control’ DNA are 9 9 9
"
1132 bp; Lane, wild-type is 783 bp. A 100 bp ladder was used (Fermentas Inc.). Middle 1 0 0 0
"
panel-confirmation of the integration of PplzA-PlzA-PflgB-aacC1 between genes bb0445 1 0 0 1
"
and bb0446. The PCR primers used to amplify a specific region are indicated at the top 1 0 0 2
"
of the agarose gels. Sizes of all PCR products were amplified as expected. Controls in 1 0 0 3
"
both gels were the plasmid DNAs that were used to inactivate plzA or complement the 1 0 0 4
"
plzA mutant. 1 kb DNA ladder (Fermentas Inc.) was used in middle panel. Right panel- 1 0 0 5
"
real-time PCR detecting bb0733 mRNA from the indicated strains. The Bar represents 1 0 0 6
"
the mean + SEM from 2 independent experiments. WT is wild-type. Average relative 1 0 0 7
"
expression of bb0733 from each strain was calculated using 2〉〉-CT method. Enolase 1 0 0 8
"
was used as an internal control (54,87). 1 0 0 9
"
1 0 1 0
"
Fig. 4. PCR analysis confirmed that the wild-type, plzA mutant and the plzA+ 1 0 1 1
"
complemented cells retained endogenous plasmids required for infection of mice and 1 0 1 2
"
ticks. 21 sets of PCR primers were used to detect 12 linear and 9 circular B. burgdorferi 1 0 1 3
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4 1"
"
plasmids (12,100). lp indicates linear plasmids and cp indicates circular plasmids. 1 0 1 4
"
Arrows indicate cp9 and lp56 plasmids were lost from all strains. 1 0 1 5
"
1 0 1 6
"
Fig. 5. Swarm plate motility assays confirmed that the plzA mutant was defective in 1 0 1 7
"
motility. A representative swarm assay with plzA mutant cells incubated for 7 days is 1 0 1 8
"
shown. Since the swarm diameters were not clearly visible, a black bar on each swarm 1 0 1 9
"
was placed which represents the size of their swarm diameters (left panel). Swarm 1 0 2 0
"
diameters from 4 assays were measured in millimeter ± standard deviation (right panel). 1 0 2 1
"
A flagellin-less, non-motile flaB mutant was used as a control (51). *"indicates significant 1 0 2 2
"
difference (p = 0.001) between wild-type and the plzA mutant cells.""1 0 2 3
"
1 0 2 4
"
Fig. 6. The levels of prominent motility or chemotaxis proteins were not altered in the 1 0 2 5
"
plzA mutant. Western blotting using 5 たg cell lysates from indicated strains were probed 1 0 2 6
"
with antisera against indicated motility and chemotaxis proteins. Specific antisera 1 0 2 7
"
against proteins used are indicated on the right side of each blot. The values on the left 1 0 2 8
"
are molecular sizes in kilodaltons. 1 0 2 9
"
1 0 3 0
"
Fig. 7. Immunofluorescence assays (IFA) of immersed larval ticks 7 days post feeding. 1 0 3 1
"
Spirochetes were detected in wild-type, plzA mutant and the complemented plzA+ 1 0 3 2
"
strains; however, this is the only tick where several plzA B. burgdorferi cells were 1 0 3 3
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4 2"
"
detected. In nearly all ticks infected with plzA mutant spirochetes by immersion, no 1 0 3 4
"
spirochetes were detected by IFA 7 days post feeding (n=15). 1 0 3 5
"
"1 0 3 6
"Table 1: plzA mutant cells are significantly attenuated in C3H/HeN mice infection
1 0 3 7"
Strain 5x102 5x103 5x104 5x105 5x106 ID50
Wild-type 0/8 3/8 10/10 8x103
plzA 0/8 1/8 6/12 6/10 10/10 9x104
plzA+ 1/8 4/8 10/10 4x103
1 0 3 8
"
1 0 3 9
"
1 0 4 0
"
Table 2: plzA mutant cells are unable to complete mouse-tick-mouse infection cycle 1 0 4 1
"
Strain Dose % mice infected
% ticks positive
# Bb/ticka # nymphs
per mouse
# mice infected
Wild-type 5x104 100 95 3947 ± 725 10 3/3
plzA 5x106 100 55 20b 30 0/5
plzA+ 5x104 100 100 3006 ± 537 10 5/5
aBb/tick, viable B. burgdorferi per tick. 1 0 4 2
"
bplzA-infected ticks had 20 or less spirochetes/tick (n=10). 1 0 4 3
"
1 0 4 4
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
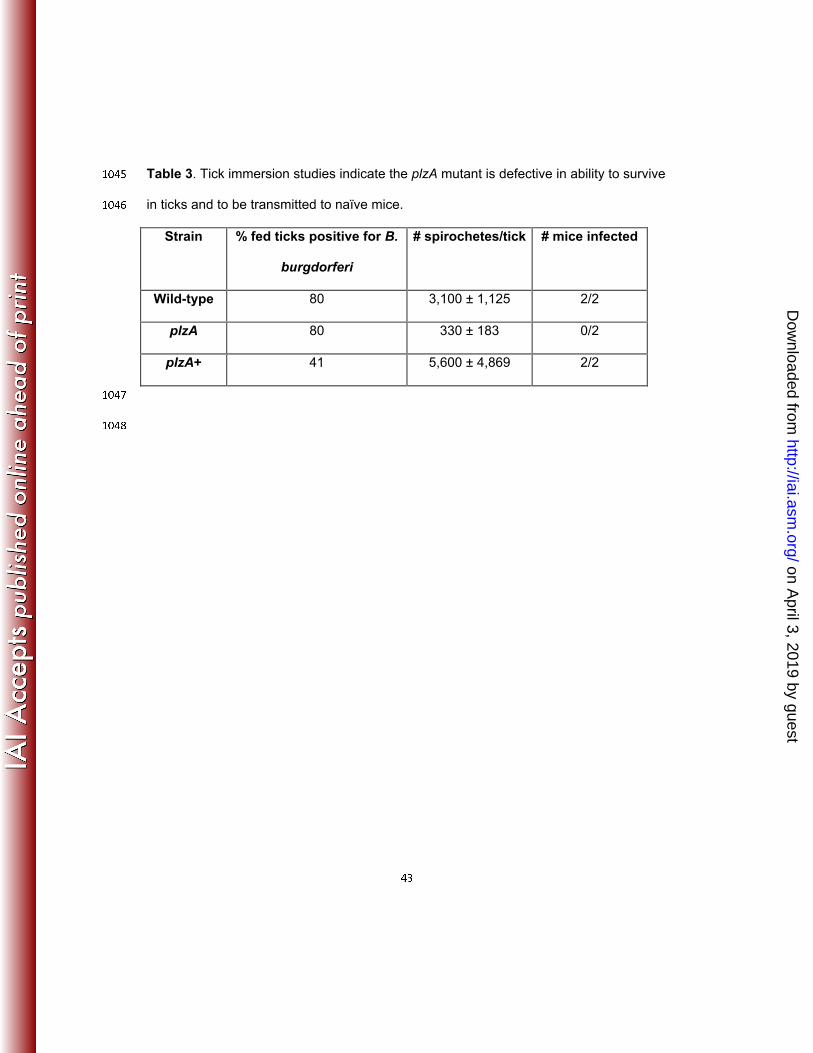
4 3"
"
Table 3. Tick immersion studies indicate the plzA mutant is defective in ability to survive 1 0 4 5
"
in ticks and to be transmitted to naïve mice. 1 0 4 6
"
Strain % fed ticks positive for B.
burgdorferi
# spirochetes/tick # mice infected
Wild-type 80 3,100 ± 1,125 2/2
plzA 80 330 ± 183 0/2
plzA+ 41 5,600 ± 4,869 2/2
1 0 4 7
"
1 0 4 8
"
on April 3, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, June 2011, p. 2499 Vol. 79, No. 60019-9567/11/$12.00 doi:10.1128/IAI.05273-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
ERRATUM
Analysis of the Borrelia burgdorferi Cyclic-di-GMP-Binding Protein PlzA Revealsa Role in Motility and Virulence
Joshua E. Pitzer, Syed Z. Sultan, Yoshihiro Hayakawa, Gerry Hobbs,Michael R. Miller, and Md A. Motaleb
Department of Microbiology and Immunology, Brody School of Medicine, East Carolina University, Greenville, North Carolina 27834;Department of Applied Chemistry, Faculty of Engineering, Aichi Institute of Technology, 1247 Yachigusa, Yakusa-cho, Toyota 470-0392,
Japan; Department of Statistics, West Virginia University, Morgantown, West Virginia 26506; and Department ofBiochemistry, Health Sciences Center, West Virginia University, Morgantown, West Virginia 26506
Volume 79, no. 5, p. 1815–1825, 2011. Page 1819, column 2, line 4 from bottom: “FligG1” should read “FliG1.”Page 1822: column 1, lines 15 and 16: “translucent colony morphology” should read “opaque colony morphology.”
2499


















