
Non-Vector Transmission of Dengue and Other Mosquito...
14
18 Dengue Bulletin – Vol 29, 2005 Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses ¶ Lin H. Chen* and Mary E. Wilson** *Harvard Medical School, Travel Medicine Center, Mount Auburn Hospital, Division of Infectious Diseases, Mount Auburn Hospital, 330 Mount Auburn Street, Cambridge, MA 02238, USA **Harvard Medical School, Division of Infectious Diseases, Mount Auburn Hospital, 330 Mount Auburn Street, Cambridge, MA 02238, USA Abstract A number of mosquito-borne viruses in the family Flaviviridae, genus Flavivirus, cause significant illnesses in humans. These diseases include dengue, yellow fever, West Nile fever, Japanese encephalitis, St. Louis encephalitis and Murray Valley encephalitis. The viruses cause syndromes that can be generally classified as one of three types: haemorrhagic fever, fever with rash and arthralgia, and encephalitis. Transmission of dengue virus following mucocutaneous exposure has recently been documented. Transmission of West Nile virus (WNV) without a mosquito vector has also been reported. We review the literature and summarize reported cases and routes of non-vector transmission of the six medically most important mosquito-borne flaviviruses. We also compare the biological characteristics of the viruses in humans and discuss how this could influence the probability of non-mosquito transmission. Keywords: Dengue, flavivirus, West Nile virus, virus transmission. ¶ Although the non-mosquito route of transmission of dengue and other mosquito-borne viral infections is rare and accidental but it has a lot of significance for physicians, who should keep this in mind and should aggressively look for these possibilities while making differential diagnosis of mosquito-borne viral infections, particularly in non-endemic areas. However, public health workers engaged in the prevention and control of dengue and other vector-borne viral infections should essentially treat these as mosquito-borne infections - Editor. [email protected]; 617 499 5026; Fax: 617 499 5453 Introduction: Flavivirus Transmission The genus Flavivirus, which belongs to the family Flaviviridae, contains 73 RNA viruses [1] . Among these viruses, 34 are mosquito-borne, 17 are tick-borne and 22 are zoonotic agents; 22 of the 34 mosquito-borne and 13 of the 17 tick-borne flaviviruses are associated with human disease [1] . There are three serological groups – dengue serological group, Japanese encephalitis serological group and yellow fever virus group [1,2] . Non-vectored flaviviruses are probably maintained in nature by animal-to- animal transmission via saliva and urinary shedding. Flavivirus infection in mosquito occurs when the mosquito ingests a blood meal containing the virus, which infects the midgut epithelial cells and subsequently the salivary gland [1] . Some days after the initial blood meal (extrinsic incubation period), the virus is
-
Upload
nguyenkhue -
Category
Documents
-
view
214 -
download
1
Transcript of Non-Vector Transmission of Dengue and Other Mosquito...

18 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Non-Vector Transmission of Dengue and OtherMosquito-Borne Flaviviruses¶
Lin H. Chen* and Mary E. Wilson**
*Harvard Medical School, Travel Medicine Center, Mount Auburn Hospital,Division of Infectious Diseases, Mount Auburn Hospital, 330 Mount Auburn Street, Cambridge,
MA 02238, USA**Harvard Medical School, Division of Infectious Diseases, Mount Auburn Hospital,
330 Mount Auburn Street, Cambridge, MA 02238, USA
Abstract
A number of mosquito-borne viruses in the family Flaviviridae, genus Flavivirus, cause significant illnessesin humans. These diseases include dengue, yellow fever, West Nile fever, Japanese encephalitis, St.Louis encephalitis and Murray Valley encephalitis. The viruses cause syndromes that can be generallyclassified as one of three types: haemorrhagic fever, fever with rash and arthralgia, and encephalitis.Transmission of dengue virus following mucocutaneous exposure has recently been documented.Transmission of West Nile virus (WNV) without a mosquito vector has also been reported. We review theliterature and summarize reported cases and routes of non-vector transmission of the six medically mostimportant mosquito-borne flaviviruses. We also compare the biological characteristics of the viruses inhumans and discuss how this could influence the probability of non-mosquito transmission.
Keywords: Dengue, flavivirus, West Nile virus, virus transmission.
¶Although the non-mosquito route of transmission of dengue and other mosquito-borne viral infections is rareand accidental but it has a lot of significance for physicians, who should keep this in mind and should aggressivelylook for these possibilities while making differential diagnosis of mosquito-borne viral infections, particularly innon-endemic areas. However, public health workers engaged in the prevention and control of dengue and othervector-borne viral infections should essentially treat these as mosquito-borne infections - Editor.
[email protected]; 617 499 5026; Fax: 617 499 5453
Introduction:Flavivirus Transmission
The genus Flavivirus, which belongs to thefamily Flaviviridae, contains 73 RNA viruses[1].Among these viruses, 34 are mosquito-borne,17 are tick-borne and 22 are zoonotic agents;22 of the 34 mosquito-borne and 13 of the 17tick-borne flaviviruses are associated withhuman disease[1]. There are three serologicalgroups – dengue serological group, Japanese
encephalitis serological group and yellow fevervirus group[1,2]. Non-vectored flaviviruses areprobably maintained in nature by animal-to-animal transmission via saliva and urinaryshedding.
Flavivirus infection in mosquito occurswhen the mosquito ingests a blood mealcontaining the virus, which infects the midgutepithelial cells and subsequently the salivarygland[1]. Some days after the initial blood meal(extrinsic incubation period), the virus is

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 19
secreted in the saliva and reaches a new hostwhen the mosquito takes another blood meal.Infection of salivary gland leads to lifelonginfection in the mosquito[3]. Some enzymeprocessing may occur in the mosquito’s midgut,for example by trypsin, and may exert animpact on the infectivity of the virus[4]. Followinga mosquito bite, which inoculates dengue virusinto skin, the virus appears to target skinLangerhans cells, and replicates in local tissuesand regional lymph nodes, then disseminatesvia lymphatics to the blood stream[1,5,6].Similarly, after inoculation from an infectedmosquito, the West Nile virus (WNV) mayinfect fibroblasts, vascular endothelial cells orthe reticuloendothelial system, leading toviraemia, and then reach the central nervoussystem to infect the host cells[2,3]. The recentstudy that documents the infection ofuninfected mosquitoes co-feeding with WNV-infected mosquitoes on the same host (mouse)shows that some local diffusion/dispersal of thevirus from bite site must occur as the mosquitois feeding[7].
For a mosquito-borne infection, theamount of virus in the mosquito saliva, thefrequency of bites and the duration of feedingby infected mosquitoes would determine theamount of virus inoculated. The amount ofWNV secreted in mosquito saliva has beenmeasured by real-time reverse transcriptase-polymerase chain reaction (RT-PCR) in Culexpipiens quinquefasciatus, a vector of WNV;mosquitoes infected with WNV could transmitabout 104.3 plaque-forming units of virus[8].Furthermore, mosquitoes inoculate about 1%of their total virus content when they take ablood meal, thus each mosquito could transmitat least 100 infective doses of virus[8].
The level of viraemia in humans dependson the rate of clearance by macrophages, andviraemia ceases with the production of humoralantibodies in the host[1]. In vertebrate hosts,dengue viraemia titers usually are >105/ml,
although virus can be below detectable levelsby conventional laboratory techniques[1]. Thetime of onset, duration and level of viraemiavaries with different flaviviral infections andmay vary from one subtype or strain of a specificvirus to the other.
Analogous to mosquito-borne infection,a non-vector transmission depends on theamount of virus in the inoculum and volumeof material that reaches a receptive site.Additional factors that may influence thelikelihood of direct transmission includestability of shed virus in the environment, theimmune status of those potentially exposed,the size of the inoculum and the route of viruscontact and entry[9]. Virus shedding may varyamong the flaviviruses, may be different inhumans as compared to experimental animals,and may occur in irregular patterns. Manyreports of infection follow blood exposure,but some flaviviral infections are associatedwith virus in cerebrospinal fluid (CSF), urine,or other fluids that clinicians and caretakerscould potentially be exposed to. A patientwho is critically ill and who has haemorrhageis more likely to be a source of bloodexposures in the hospital setting or at home.An important variable in the level of risk isthe time of viraemia relative to the presenceof severe illness and haemorrhage.
The viability of viruses in the environmentalso may differ. For example, WN and yellowfever viruses appeared to have prolongedsurvival at room temperature in an experimentwhere the viruses dried on filter paper weretested for infectivity by culture, titration in Verocells and assessed by RT-PCR for viral RNA[10].WNV was recovered up to 60 days after theprocedure and yellow fever up to 90 days[10].The survival of Japanese encephalitis virus (JEV)appeared to be inversely related to relativehumidity whereas the yellow fever virusinfectivity remained longer at higher relativehumidity[9].
Dengue Bulletin – Vol 29, 2005 19

20 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Virus
Dengue viruses can be associated withhaemorrhage, which can increase health careworkers’ exposure to blood, and presumablylead to a greater risk for direct or nosocomialtransmission. Dengue virus transmissionwithout mosquito vector has been reported tooccur via different routes, includingneedlestick, intrapartum, bone marrowtransplant and mucocutaneous exposure[11-21]
(Table 1).
Among the reported cases of dengue virusinfection from non-mosquito transmission,most source patients did not have haemorrhagicmanifestations. Specifically, the reported healthcare workers who acquired dengue infectionwere not exposed to haemorrhagic sourcepatients. One case of intrapartum/verticaltransmission occurred in an infant born to amother who was diagnosed with denguehaemorrhagic fever just before delivery[18].
There are no known animal models ofdengue virus transmission. Although serologicalsurveys have demonstrated dengue antibodiesin macaques in Sri Lanka, non-human primatesinfected with dengue virus do not exhibitsymptoms of illness[22]. It is not clear whetherasymptomatic infections in non-humanprimates contribute to maintenance of denguevirus in nature.
Yellow Fever
Yellow fever is the prototype virus amongstthe flaviviruses. Because yellow fever virus isassociated with haemorrhage, there may bea greater potential for transmission throughblood exposure in health care workers,compared to flaviviruses that do not causehaemorrhage. A review of literature identifiednumerous cases of non-vector transmission inthe pre-vaccine era, where scientists and
laboratorians became infected with yellowfever virus after contact with blood or tissuesof infected laboratory animals or handlingexperimentally-infected animals[23,24]. Onecase that occurred in 1930 in a hospitaltechnician who analysed the blood of a yellowfever patient in London was described againrecently; the route of transmission wasunclear[25] (Table 1).
Experimentally, monkeys have becomeinfected with yellow fever virus that wasintroduced through a gastric catheter[26].Intranasal transmission and mucocutaneoustransmission via conjunctival sac has also beendocumented in monkeys in the laboratory[27-29].Furthermore, yellow fever virus rubbed onintact abdominal skin of monkeys has led toinfection[30] (Table 2).
West Nile Virus
Human infections with WNV have beenreported to occur from needlestick,breastfeeding, blood transfusion, organtransplants, haemodialysis and intrauterine ortransplacental routes[31-38]. Two cases of WestNile fever occurred in turkey breeders wherethe route of transmission was unknown, butwas speculated to have been by aerosol, fecal-oral or percutaneous exposure[37]. Aseroprevalence survey conducted on theworkers from six turkey farms showed that 18%had recent infection with WNV; seroprevalencewas highest (55%) from the farm where thetwo cases worked[37] (Table 1).
Among the reported cases of non-mosquito transmission, many source patientswere asymptomatic. In fact, asymptomaticviraemic donors of blood products and organswere the most likely contributors to non-mosquito transmission. This shows thatasymptomatically-infected individuals areviraemic.

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 21
Table 1. Reported routes of transmission in humans without vector

22 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Infected blood has, however, been themost frequently documented source of non-mosquito transmission. WNV RNA has beenidentified by reverse transcriptase-polymerasechain reaction in the urine of a patient withencephalitis on the 8th day of illness[39]. It isnot yet clear whether the WNV RNA in urineis infectious, and whether viruria plays asignificant role in human WNV transmission.
Similarly, an experiment in hamsters foundWNV to persist in the brain and kidneys,leading to chronic renal infection with persistentviruria[40]. Although transmission from urineexposure has not been documented inliterature, infectious WNV was cultured fromhamster urine for up to eight months and couldpossibly spread to other animals by aerosol orby ingestion[41]. WNV can also infect mice bythe oral route[42]. In hamsters, investigatorscompared the pathogenesis of WNV followinginfection by mosquito bite, needle inoculationand ingestion; infection occurred by all threeroutes, resulting in similar levels of viraemia,duration of viraemia, clinical manifestations,pathology and antibody response[43]. The onsetof viraemia was delayed following oral infectionand mortality was lower, but the importanceof oral route in the transmission of WNV innature remains unclear[43] (Table 2).
Flaviviruses are sensitive to acid pH and bile,but their transmission via ingestion of infectedmilk has been demonstrated with tick-borneencephalitis, another flavivirus infection[1,44]. It isconceivable that virus entry could occur at otherparts of oropharynx rather than in the stomachor duodenum. With breastfeeding humans, onecould postulate that virus potentially could entervia mucosa of oropharynx.
Japanese Encephalitis
No nosocomial cases in humans have beenreported. However, intrapartum, transplacental
or congenital infection has beendocumented[45,46]. A study of five pregnantwomen infected during an epidemic in UttarPradesh, India, found that two womendelivered normal infants, two womenmiscarried in the first or second trimesters, andthe virus was isolated from one aborted fetus[46]
(Table 1).
Experiments in animals havedemonstrated direct transmission of JEV. Onestudy showed that a bat that ingested infectedmosquitoes became infected[47]. Twoexperiments showed possible aerosoltransmission[48,49]. Macaques inoculatedintranasally with JEV developed symptoms 11-14 days later; viral antigen was identified byimmunofluorescent staining and histo-pathological changes were found in thenervous system[48]. Investigators speculatedthat JEV probably entered the blood streamfollowing aerosol infection and seeded thecentral nervous system. A study in miceinfected with aerosolized JEV showed lesionsin olfactory bulb, frontal lobe and olfactoryportions of cerebrum initially, followed bynecrotic lesions in olfactory bulb, cerebrum,brain stem and spinal cord; the suggestedspread was from nasopharynx across thecribriform plate to the olfactory bulb[49]. Oralinfection in mice of JEV led to antibodyproduction and protection against intracerebralchallenge of JEV[50]. JEV has also been isolatedfrom mouse urine after infection; in addition,JE viruria was shown to persist longer thanviraemia (days 5–9 versus days 1–2 afterinfection)[51] (Table 2).
St. Louis Encephalitis
Direct, non-mosquito transmission of this virusin humans has not been reported. However, astudy on mice exposed to aerosolized St. Louisencephalitis virus in a closed chamberdemonstrated intranasal/aerosol transmission[52].

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 23
Table 2. Potential routes of transmission in animals*
*WNV and St. Louis encephalitis virus have been isolated from urine of experimentally infected hamsters[40,41]
and JEV from urine of mice[51], but no reports documenting transmission from urine were found in thepublished literature.
Patients with St. Louis encephalitis were foundto have viral antigen in urine by indirectimmunofluorescence, electron microscopy andimmune electron microscopy, although virus
was not isolated from urine[53]. Recent studiesshow that hamsters experimentally infected bysubcutaneous injection of St. Louis virus canshed the virus in urine for at least four months[41].

24 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Murray Valley Encephalitis
Direct, non-mosquito transmission of this virusin humans has not been reported, which mayreflect the small number of human infectionsas compared to the other flaviviruses.
Viraemia
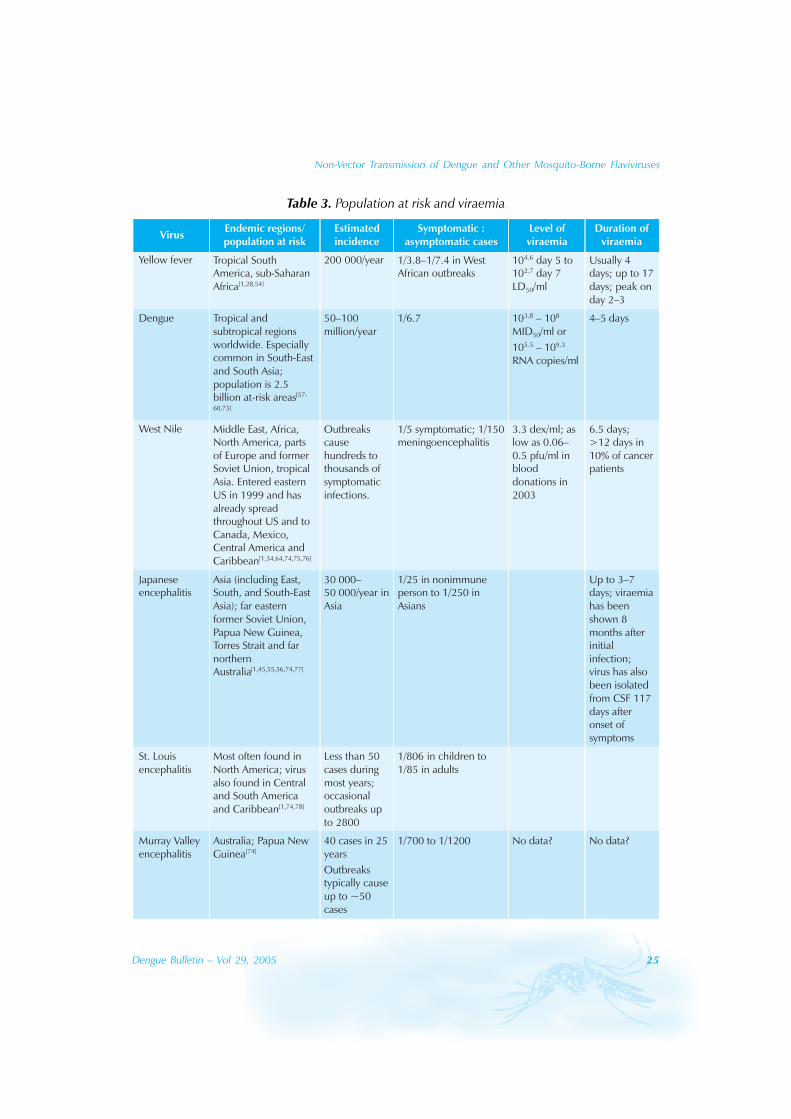
The levels of viraemia among the flavivirusesare difficult to compare because measurementsare described in different terms. Thesemeasurements include median mosquitoinfectious doses per millilitre (MID50/ml), copiesof RNA per millilitre, plaque-forming units permillilitre (pfu/ml), and intracerebral medianlethal dose per millilitre (LD50/ml) (Table 3).
Yellow fever virus is readily isolated duringthe first four days of illness, but may berecovered from serum up to 17 days[54]. Yellowfever viraemia in a human case showed thatvirus titres were 104.6 LD50 at 5 days after theonset of symptoms and 102.7 LD50 at 7 daysafter the onset of illness[53]. Experience withJapanese encephalitis also shows that viraemiacan persist following resolution of symptomsand in recurrently symptomatic patients[55,56].
In natural infection with dengue virus, theduration and level of viraemia varied amongdifferent strains and serotypes[57]. The durationof viraemia ranged from 1–7 days (2–12 days asper Gubler, 1981), with a mean of 4.5 days,median of 5 days[58]. The duration of viraemia isunderestimated in the study because some ofthe children were still viraemic on the last blooddraw. The duration is longer in primary infection(5.1 days), compared to secondary infection (4.4days), and the peak virus titre was 108.2 MID50/ml (median mosquito infectious doses permillilitre)[58]. Patients with primary infections hadpersistent viraemia until 1.6 days afterdefervescence whereas those with secondaryinfections cleared viraemia by 0.6 days after
defervescence[59]. Higher levels of viraemiacorrelated with the severity of disease, and therate of virus clearance in the two days beforedefervescence was greater in denguehaemorrhagic fever than in dengue fever[59].
The measurement of plasma RNA byquantitative competitor (RT-PCR) derived arange of 105.5 to 109.3 copies/ml, whichcorrelated with MID50 by a geometric meannumber of 0.15 RNA copies per MID50
[60].These RNA levels peaked two days beforedefervescence and declined to belowdetection in two thirds of patients bydefervescence[60]. Therefore, one third ofpatients had detectable dengue RNA atdefervescence and they were possiblyinfectious after defervescence[60]. A study ofadult patients infected with DENV-3 insouthern Taiwan in 1998 using the quantitativeRT-PCR method showed that duringdefervescence, viral RNA remained high inDHF patients but was undetectable in DFpatients; viraemia in DHF patients persistedfor up to six days after defervescence[61].
Since the level and duration of viraemiaappear to correlate with the severity of thedisease, it is likely that asymptomatic dengueinfection would have lower levels and shorterduration of viraemia than DF or DHF. Likewise,other flavivirus infections are expected to havelower levels of viraemia in asymptomaticinfections, compared to that in symptomaticinfections. Cases of WNV acquired throughtransfusion demonstrate viraemia inasymptomatic donors. Estimated viral loads inblood donations that led to transfusion-associated transmission of WNV were 0.8–75pfu/ml for 2002 and 0.06–0.5 pfu/ml in 2003[34].Because RNA titres are higher than plaque-forming units by about 400[62], the RNA levelswould range from 24–30 000 copies/ml.
In West Nile infections, viraemia is presentduring the two days before until about fourdays after the onset of illness[63]. Rarely the
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 25
Table 3. Population at risk and viraemia

26 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
virus is isolated from the CSF in patients withmeningoencephalitis. The virus has also beenisolated from organs such as liver, spleen, lungand pancreas, with a high concentration ofreticuloendothelial cells[64]. In immuno-compromised patients intentionally infectedwith West Nile virus as part of a study toexamine novel approaches to cancertreatment, the virus could be recovered fromblood up to 28 days after inoculation.
In the case of percutaneous ormucocutaneous exposure, the viral load woulddepend on the amount of blood in contact.Assuming a mean volume of blood inoculatedvia needlestick to be 1 ul[65], the viral load aftera needlestick exposure would be in the rangeof 103 to 107 RNA copies/ml for dengue virusand up to 300 RNA copies/ml for West Nilevirus. Despite the minuscule amount of bloodtransferred, needlestick transmission has beendocumented for both dengue and West Nile.
Quantification of Risk
The risk of transfusion-associated transmissionof West Nile virus in Queens, New York, in2002, was estimated to be 1.8/10 000donations (mean) up to 2.7/10 000 donations(peak) (or 1 in 3700 to 1 in 5555 donations)[66].These estimates were based on the findingsthat 80% of West Nile virus infections areasymptomatic, that the incubation period lasts2–6 days, and that the virus is detected in blood1–2 days after mosquito inoculation; thereforean asymptomatic donor may have a mean ofthree viraemic days before the onset ofsymptoms[66]. In comparison, the estimatedfrequency of transfusion-associated hepatitis Bvirus (HBV) infection in the United States withcurrently available testing technologies is 1/30 000–250 000 units, hepatitis C virus (HCV)infection is 1/30 000–150 000 units, andhuman immunodeficiency virus (HIV) infectionis 1/200 000 to 2 000 000 units[67].
A recent report of HCV transmissionassociated with saline flushes (re-use ofdisposable syringes and contamination ofshared saline bags) showed an attack rate of27%, and the dose leading to infection of 50%of exposed population for patients was threeflushes (30–60 ml of saline)[68,69]. Given therelative rates, one could estimate that WNVcould be transmitted from up to 5% of viraemiccontacts if similar breaches in sterile techniquesoccurred with saline flushes.
Health care workers (HCWs) regularlyencounter occupational exposure to blood andbody fluids, and the risk of injury is related toprecautions taken. A study in a communityhospital found that one-quarter of HCWs hadmucocutaneous blood exposure in the threemonths prior to the survey, whereas one thirdof HCWs had percutaneous injury in the sameperiod.[70] Another study that assessed the riskof sharps injuries in nurses caring for diabeticpatients found nearly 80% of nursesexperienced at least one needlestick; theAmerican Nursing Association reported 600 000to 1 000 000 sharps injuries in HCWs eachyear[71]. The frequency of exposure in HCWsillustrates the vulnerability to blood-bornetransmission of infections, includingflaviviruses. Health care workers in developingcountries face a substantially riskierenvironment because of inadequate suppliesof sterile needles, masks, gloves and gogglesthat could protect them from blood exposures,though few studies are available to documentthe magnitude of these risks[72].
Conclusion
Multiple routes of non-mosquito transmissionof flaviviruses in humans have been describedrecently. Infections with dengue and West Nileviruses have been the most commonlydocumented. These two viruses can beassociated with asymptomatic infection, febrile

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 27
illness with rash and arthralgia, or more severeillnesses such as dengue haemorrhagic fever/dengue shock syndrome and West Nilemeningoencephalitis. The asymptomaticallyinfected persons appear to be particularlyimportant as sources in the transfusion-associated and transplant-associatedtransmission of West Nile virus.
Syndromes with haemorrhage may increasethe risk of exposure to blood-borne pathogens,possibly due to higher level of viraemia, greaterdifficulty in avoiding accidental contact and moreintense contact needed to care for the severelyill patients. In spite of the fulminant course andhaemorrhagic sequelae of yellow feverinfection, the only documented and publishedcases of possible direct transmission of yellowfever virus were reported before the availabilityof the yellow fever vaccines. Vaccinationprobably has provided significant protection tohealth care workers and laboratorians. It is alsopossible that health care workers take greaterpersonal protective measures in caring forseverely ill patients (for example, those withhaemorrhagic symptoms) and thereforeminimize their exposure.
Risks of non-mosquito transmission offlaviviruses may differ among the viruses formany reasons. These include the size of thepopulation exposed to the infection (which mayvary from year to year and change over time),the availability and state of medical resources(e.g. adequacy of infection control, testing ofdonated blood, availability of organtransplantation), biological characteristics of thevirus and immunity of the human populationbecause of prior infection or vaccination. Asdiscussed, the level of viraemia, the durationof viraemia and peak viraemia differ amongthe flaviviruses. The infective dose of virus mayalso differ among the flaviviruses. Finally, theperiod of clinical illness and the period ofviraemia may have different patterns ofoverlap. Persistent shedders of virus would posea greater risk of transmission to their contacts.
Infection may not be detected in anendemic population where the majority ofindividuals have previously been infected orvaccinated. For example, non-immuneyoungsters are more likely to be infected bydengue virus, but an association with non-mosquito transmission may not be apparent tothem. Furthermore, WNV is the only flavivirusdocumented in the literature to be transmittedby transfusion of blood products. One possibleexplanation is that youngsters, in whomflavivirus infections such as dengue occur at ahigher incidence, are less likely to donate blood.Another explanation is that non-mosquitotransmission is unlikely to be recognized in anarea where an infection is endemic. In addition,many flavivirus infections result in mild orasymptomatic illness and may not prompt anywork-up, so could not be recognized astransfusion-acquired infections.
Adequacy of infection control educationand materials will influence risks in the healthcare setting. In many countries endemic forJE, dengue and yellow fever, resources forinfection control are lacking and adherence toprecautions may be difficult or impossible.Diagnostic laboratories and technologies todocument infections and their sources may beunavailable in many endemic countries. It ismost difficult to differentiate between non-mosquito transmission and mosquito-borneinfection in endemic areas where the vector iswidespread. Hence, most cases of non-mosquito flavivirus transmission in humanshave been documented in developed countries.Our review would suggest that nosocomialtransmission of these infections probably doesoccur in endemic areas, and health careworkers should be aware of the these risks.
The authors searched Medline/Pubmed forviraemia, nosocomial transmission andmucocutaneous transmission of flaviviruses aswell as yellow fever, dengue, West Nile,Japanese encephalitis, St. Louis encephalitisand Murray Valley encephalitis for pertinentinformation on the topic.

28 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
References
[1] Burke DS and Monath TP. Flaviviruses. In: KnipeDM, Howley, Griffin DE, Lamb RA, Martin MA,Roizman B, Straus SE, editors. Fields virology. 4th
ed. Vol.1 .Philadelphia: Lippincott Williams &Wilkins, 2001: pp.1043-1125.
[2] Mackenzie JS, Gubler DJ and Petersen LR.Emerging flaviviruses: the spread and resurgenceof Japanese encephalitis, West Nile and dengueviruses. Nat Med. 2004; 10(12 Suppl): S98-109.
[3] Gea-Banacloche J, Johnson RT, Bagic A, ButmanJA, Murray PR and Agrawal AG. West Nile virus:pathogenesis and therapeutic options. Ann InternMed. 2004; 140(7): 545-553.
[4] Molina-Cruz A, Gupta L, Richardson J, Bennett K,Black W IV and Barillas-Mury C. Effect of mosquitomidgut trypsin activity on dengue-2 virus infectionand dissemination in Aedes aegypti. Am J Trop MedHyg. 2005; 72(5): 631-637.
[5] Wu SJ, Grouard-Vogel G, Sun W, Mascola JR, BrachtelE, Putvatana R, Louder MK, Filgueira L, MarovichMA, Wong HK, Blauvelt A, Murphy GS, Robb ML,Innes BL, Birx DL, Hayes CG and Frankel SS. Humanskin Langerhans cells are targets of dengue virusinfection. Nat Med. 2000; 6(7): 816-820.
[6] Marovich M, Grouard-Vogel G, Louder M, Eller M,Sun W, Wu SJ, Putvatana R, Murphy G,Tassaneetrithep B, Burgess T, Birx D, Hayes C,Schlesinger-Frankel S and Mascola J. Humandendritic cells as targets of dengue virus infection. JInvestig Dermatol Symp Proc. 2001; 6(3): 219-224.
[7] Higgs S, Schnieder BS, Vanlandingham DL, KlingerKA and Gould EA. Nonviremic transmission ofWest Nile virus. Proc Nat Acad Sci. 2005; 102(25):8871-8874.
[8] Vanlandingham DL, Schneider BS, Klingler K, FairJ, Beasley D, Huang J, Hamilton P and Higgs S.Real-time reverse transcriptase-polymerase chainreaction quantification of West Nile virustransmitted by Culex pipiens quinquefasciatus. AmJ Trop Med Hyg. 2004; 71(1): 120-123.
[9] Kuno G. Transmission of arboviruses withoutinvolvement of arthropod vectors. Acta Virol. 2001;45(3): 139-150.
[10] Guzman H, Ding X, Xiao S-Y and Tesh RB. Durationof infectivity and RNA of Venezuelan equineencephalitis, West Nile, and yellow fever virusesdried on filter paper and maintained at roomtemperature. Am J Trop Med Hyg. 2005; 72(4):474-477.
[11] Wagner D, de With K, Huzly D, Hufert F,Weidmann M, Breisinger S, Eppinger S, Kern WVand Bauer TM. Nosocomial acquisition of dengue.Emerg Infect Dis. 2004; 10(10): 1872-1873.
[12] Nemes Z, Kiss G, Madarassi EP, Peterfi Z, FerencziE, Bakonyi T, Ternak G. Nosocomial transmissionof dengue. Emerg Infect Dis. 2004; 10(10): 1880-1881.
[13] de Wazieres B, Gil H, Vuitton DA and Dupond JL.Nosocomial transmission of dengue from aneedlestick injury. Lancet. 1998; 351(9101): 498.
[14] Hirsch JF, Deschamps C and Lhuillier M.Transmission métropolitaine d’une dengue parinoculation accidentelle hospitalière. Ann MedInterne (Paris). 1990; 141(7): 629.
[15] Langgartner J, Audebert F, Schölmerich J, Gluck T.Dengue virus infection transmitted by needle stickinjury. J Infect. 2002; 44(4): 269-270.
[16] Rigau-Perez JG, Vorndam AV and Clark GG. Thedengue and dengue hemorrhagic fever epidemicin Puerto Rico, 1994-1995. Am J Trop Med Hyg.2001; 64(1-2): 67-74.
[17] Chye JK, Lim CT, Ng KB, Lim JMH, George R andLam SK. Vertical transmission of dengue. Clin InfectDis. 1997; 25(6): 1374-1377.
[18] Kerdpanich A, Watanaveeradej V, Samakoses R,Chumnanvanakij S, Chulyamitporn T, Sumeksri P,Vuthiwong C, Kounruang C, Nisalak A and Endy T.Perinatal dengue infection. Southeast Asian J TropMed Public Health. 2001; 32(3): 488-493.
[19] Boussemart T, Babe P, Sibille G, Neyret C andBerchel C. Prenatal transmission of dengue: twonew cases. J Perinatol. 2001; 21(4): 255-257.
[20] Thaithumyanon P, Thisyakorn U, Deerojnawong Jand Innis BL. Dengue infection complicated bysevere hemorrhage and vertical transmission in aparturient woman. Clin Infect Dis. 1994; 18(2):248-249.
[21] Chen LH and Wilson ME. Transmission of denguevirus without a mosquito vector: nosocomialmucocutaneous transmission and other routes oftransmission. Clin Infect Dis. 2004; 39(6): e56-60.
[22] De Silva AM, Dittus WP, Amerasinghe PH andAmerasinghe FP. Serologic evidence for anepizootic dengue virus infecting toque macaques(Macaca sinica) at Polonnaruwa, Sri Lanka. Am JTrop Med Hyg. 1999; 60(2): 300-306.

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 29
[23] Low CG and Fairley NH. Laboratory and hospitalinfections with yellow fever in England. Br Med J.1931: 125-128.
[24] Berry GP and Kitchen SF. Yellow fever accidentallycontracted in the laboratory: a study of seven cases.Am J Trop Med Hyg. 1931; 11: 365-434.
[25] Cook GC. Fatal yellow fever contracted at theHospital for Tropical Diseases, London, UK, in1930. Trans R Soc Trop Med Hyg. 1994; 88(6):712-713.
[26] Findlay GM and MacCallum FO. Transmission ofyellow fever virus to monkeys by mouth. J PatholBacteriol. 1939; 49: 53-61.
[27] Niedrig M, Stolte N, Fuchs D, Hunsmann G andStahl-Hennig C. Intra-nasal infection of macaqueswith yellow fever vaccine 17D: a novel andeconomical approach for YF vaccination in mice.Vaccine. 1999; 17(9-10): 1206-1210.
[28] Monath TP. Yellow fever vaccine. In: Plotkin SA,Orenstein WA, editors. Vaccines. 4th ed.Pennsylvania: Saunders, 2004. pp. 1095-1176.
[29] Findlay GM and Clarke LP. Infection withneurotropic yellow fever virus following instillationinto the nares and conjunctival sac. J PatholBacteriol. 1935; xl: 55.
[30] Bauer JH, Hudson NP. Passage of virus of yellowfever through skin. Am J Trop Med. 1928; 8: 371-378.
[31] Centers for Disease Control and Prevention.Possible West Nile virus transmission to an infantthrough breast-feeding – Michigan, 2002. MorbMortal Wkly Rep. 2002; 51(39): 877-878.
[32] Centers for Disease Control and Prevention.Intrauterine West Nile Virus infection – UnitedStates, 2002. Morb Mortal Wkly Rep. 2002; 51(50):1135-1136.
[33] Centers for Disease Control and Prevention.Laboratory-acquired West Nile Virus infections –United States, 2002. Morb Mortal Wkly Rep. 2002;51(50): 1133-1135.
[34] Centers for Disease Control and Prevention.Update: West Nile virus screening of blooddonations and transfusion-associatedtransmission—United States, 2003. Morb MortalWkly Rep. 2004; 53(13): 281-284.
[35] Pealer LN, Marfin AA, Petersen LR, Lanciotti RS,Page PL, Stramer SL, Stobierski MG, Signs K,Newman B, Kapoor H, Goodman JL and
Chamberland ME; West Nile virus transmissioninvestigation team. Transmission of West Nile virusthrough blood transfusion in the United States in2002. N Engl J Med. 2003; 349(13): 1236-1245.
[36] Iwamoto M, Jernigan DB, Guasch A, Trepka MJ,Blackmore CG, Hellinger WC, Pham SM, Zaki S,Lanciotti RS, Lance-Parker SE, DiazGranados CA,Winquist AG, Perlino CA, Wiersma S, Hillyer KL,Goodman JL, Marfin AA, Chamberland ME andPetersen LR; West Nile Virus in TransplantRecipients Investigation Team. Transmission of WestNile virus from an organ donor to four transplantrecipients. N Engl J Med. 2003; 348(22): 2196-2203.
[37] Centers for Disease Control and Prevention. WestNile virus infection among turkey breeder farmworkers—Wisconsin, 2002. Morb Mortal Wkly Rep.2003; 52(42): 1017-1019.
[38] Centers for Disease Control and Prevention.Possible dialysis-related West Nile virustransmission – Georgia, 2003. Morb Mortal WklyRep. 2004; 53(32): 738-739.
[39] Tonry JH, Brown CB, Cropp CB, Co JK, BennettSN, Nerurkar VR, Kuberski T and Gubler DJ. WestNile virus detection in urine. Emerg Infect Dis.2005; 11(8):1294-1296.
[40] Tonry JH, Xiao S-Y, Siirin M, Chen H, Travassos DaRosa APA and Tesh RB. Persistent shedding ofWest Nile virus in urine of experimentally infectedhamsters. Am J Trop Med Hyg. 2005; 72(3): 320-324.
[41] Tesh RB, Siirin M, Guzman H, Travassos da RosaAP, Wu X, Duan T, Lei H, Nunes MR and Xiao SY.Persistent West Nile virus infection in the goldenhamster: studies on its mechanism and possibleimplications for other flavivirus infections. J InfectDis. 2005; 192(2): 287-295.
[42] Odelola HA and Oduye O. West Nile virusinfection of adult mice by oral route. Arch Virol.1977; 54(3): 251-253.
[43] Sbrana E, Tonry JH, Xiao S-Y, Travassos Da RosaAPA, Higgs S and Tesh RB. Oral transmission ofWest Nile Virus in a hamster model. Am J TropMed Hyg. 2005; 72(3): 325-329.
[44] Kerbo N, Donchenko I, Kutsar K, Vasilenko V.Tickborne encephalitis outbreak in Estonia linkedto raw goat milk, May-June 2005. Eurosurveillance.2005; 10(6). (http://www.eurosurveillance. /ew/2005/050623. asp#2, accessed 30 December2005).

30 Dengue Bulletin – Vol 29, 2005
Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
[45] Halstead SB and Tsai TF. Japanese encephalitisvaccines. In: Plotkin SA, Orenstein WA, editors.Vaccines. 4th ed. Pennsylvania: Saunders, 2004.pp. 919-958.
[46] Chaturvedi UC, Mathur A, Chandra A, Das SK,Tandon HO and Singh UK. Transplacental infectionwith Japanese encephalitits virus. J Infect Dis. 1980;141(6): 712-715.
[47] La Motte LC. Japanese encephalitis in bats duringsimulated hibernation. Am J Hyg. 1958; 67: 101-108.
[48] Myint KS, Raengsakulrach B, Young GD,Gettayacamin M, Ferguson LM, Innis BL, Hoke CHJr and Vaughn DW. Production of lethal infectionthat resembles fatal human disease by intranasalinoculation of macaques with Japanese encephalitisvirus. Am J Trop Med Hyg. 1999; 60(3): 338-342.
[49] Larson EW, Dominik JW and Slone TW. Aerosolstability and respiratory infectivity of Japanese Bencephalitis virus. Infect Immun. 1980; 30(2): 397-401.
[50] Ramakrishna C, Desai A, Shankar SK, ChandramukiA and Ravi V. Oral immunization of mice with liveJapanese encephalitis virus induces a protectiveimmune response. Vaccine. 1999; 17(23-24):3102-3108.
[51] Mathur A, Khanna N, Kulshreshtha R, Maitra SCand Chaturvedi UC. Viruria during acute Japaneseencephalitis virus infection. Int J Exp Pathol. 1995;76(2): 103-109.
[52] Phillpotts RJ, Brooks TJ and Cox CS. A simple devicefor the exposure of animals to infectiousmicroorganisms by the airborne route. EpidemiolInfect. 1997; 118(1): 71-75.
[53] Luby JP, Murphy FK, Gilliam JN, Kang CY and FrankR. Antigenuria in St. Louis encephalitis. Am J TropMed Hyg. 1980; 29(2): 265-268.
[54] Nassar Eda S, Chamelet EL, Coimbra TL, de SouzaLT, Suzuki A, Ferreira IB, da Silva MV, Rocco IMand Travassos da Rosa AP. Jungle yellow fever:clinical and laboratorial studies emphasizingviremia on a human case. Rev Inst Med Trop SaoPaulo. 1995; 37(4): 337-341.
[55] Sharma S, Mathur A, Prakash V, Kulshreshtha R,Kumar R, Chaturvedi UC. Japanese encephalitisvirus latency in peripheral blood lymphocytes andrecurrence of infection in children. Clin ExpImmunol. 1991; 85(1): 85-89.
[56] Ravi V, Desai AS, Shenoy PK, Satishchandra P,Chandramuki A, Gourie-Devi M. Persistence ofJapanese encephalitis virus in the human nervoussystem. J Med Virol. 1993; 40(4): 326-329.
[57] Gubler DJ, Suharyono W, Tan R, Abidin M and SieA. Viremia in patients with naturally acquireddengue infection. Bull World Health Organ. 1981;59(4): 623-630.
[58] Vaughn DW, Green S, Kalayanarooj S, Innis BL,Nimmannitya S, Suntayakorn S, Endy TP,Raengsakulrach B, Rothman AL, Ennis FA andNisalak A. Dengue viremia titer, antibody responsepattern, and virus serotype correlate with diseaseseverity. J Infect Dis. 2000; 181(1): 2-9.
[59] Vaughn DW, Green S, Kalayanarooj S, Innis BL,Nimmannitya S, Suntayakorn S, Rothman AL, EnnisFA and Nisalak A. Dengue in the early febrilephase: viremia and antibody responses. J InfectDis. 1997; 176(2): 322-330.
[60] Sudiro TM, Zivny J, Ishiko H, Green S, VaughnDW, Kalayanarooj S, Nisalak A, Norman JE, EnnisFA and Rothman AL. Analysis of plasma viral RNAlevels during acute dengue virus infection usingquantitative competitor reverse transcription-polymerase chain reaction. J Med Virol. 2001; 63:29-34.
[61] Wang WK, Chao DY, Kao CL, Wu HC, Liu YC, LiCM, Lin SC, Ho ST, Huang JH and King CC. Highlevels of plasma dengue viral load duringdefervescence in patients with denguehemorrhagic fever: implications for pathogenesis.Virology. 2003; 305(2): 330-338.
[62] Dodd RY. Emerging infections, transfusion safety,and epidemiology. N Engl J Med. 2003; 349(13):1205-1206.
[63] Campbell GL, Marfin AA, Lanciotti RS and GublerDJ. West Nile virus. Lancet Infect Dis. 2002; 2(9):519-529.
[64] Southam CM and Moore AE. Induced virusinfections in man by the Egypt isolates of WestNile virus. Am J Trop Med Hyg. 1954; 3(1): 19-50.
[65] Napoli VM and McGowan JE Jr. How much bloodis in a needlestick? J Infect Dis. 1987; 155(4): 828.
[66] Biggerstaff BJ and Petersen LR. Estimated risk ofWest Nile virus transmission through bloodtransfusion during an epidemic in Queens, NewYork City. Transfusion. 2002; 42(8): 1019-1026.

Non-Vector Transmission of Dengue and Other Mosquito-Borne Flaviviruses
Dengue Bulletin – Vol 29, 2005 31
[67] Goodnough LT, Brecher ME, Kanter MH andAuBuchon JP. Transfusion medicine. First of twoparts - blood transfusion. N Eng J Med. 1999;340(6): 438-447.
[68] Macedo de Oliveira A, White KL, Leschinsky DP,Beecham BD, Vogt TM, Moolenaar RL, Perz JFand Safranek TJ. An outbreak of hepatitis C virusinfections among outpatients at a hematology/oncology clinic. Ann Intern Med. 2005; 142(11):898-902.
[69] Wenzel RP and Edmond MB. Patient-to-patienttransmission of hepatitis C virus. Ann Intern Med.2005; 142(11): 940-941.
[70] Doebbeling BN, Vaughn TE, McCoy KD, BeekmannSE, Woolson RF, Ferguson KJ and Torner JC.Percutaneous injury, blood exposure, andadherence to standard precautions: are hospital-based health care providers still at risk? Clin InfectDis. 2003; 37(8): 1006-1013.
[71] Lee JM, Botteman MV, Nicklasson L, Cobden Dand Pashos CL. Needlestick injury in acute carenurses caring for patients with diabetes mellitus: aretrospective study. Curr Med Res and Op. 2005;21(5): 741-747.
[72] Sagoe-Moses C, Pearson RD, Perry J and Jagger J.Risks to health care workers in developingcountries. N Engl J Med. 2001; 354(7): 538-541.
[73] Burke DS, Nisalak A, Johnson DE and Scott RM. Aprospective study of dengue infections in Bangkok.Am J Trop Med Hyg. 1988; 38(1): 172-180.
[74] Solomon T. Flavivirus encephalitis. N Engl J Med.2004; 351(4): 370-378.
[75] Petersen LR and Marfin AA. West Nile virus: aprimer for the clinician. Ann Intern Med. 2002;137(3): 173-179.
[76] Goldblum N. West Nile fever in the Middle East.Proc 6th Int Cong. Trop Med Parasitol. 1959; 5:112-125.
[77] Halstead SB and Grosz CR. Subclinical Japaneseencephalitis. I. Infection of Americans with limitedresidence in Korea. Am J Hyg. 1962; 75: 190-201.
[78] Monath TP and Tsai TF. St. Louis encephalitis:lessons from the last decade. Am J Trop Med Hyg.1987; 37(3 Suppl): 40S-59S.


















