
Neocortex: Origins - Queen's Universitybrain.phgy.queensu.ca/pare/assets/Circuits2a.pdf ·...
8
Neocortex: Origins F Aboitiz, Pontificia Universidad Cato ´lica de Chile, Santiago, Chile ã 2009 Elsevier Ltd. All rights reserved. Introduction The origin of the mammalian neocortex is probably one of the most controversial themes in modern comparative neurobiology. There have been long- standing arguments on this issue (dating from early last century), and still there is no general consensus about it. This is partly due to the enormous anatomi- cal diversification that is observed in the cerebral hemispheres of distinct vertebrate groups. Starting from a relatively conserved embryonic plan that divides the hemispheres into a dorsal pallium and a ventral subpallium, different brain components have expanded in each vertebrate class, producing pro- found distortions of the ancestral adult condition. For example, cartilaginous fishes are characterized by the development of a central pallial nucleus located at the dorsal midline, while bony fish develop a brain in which the ventricular region everts to the outside, separating the medial hemispheres in the dorsal aspect. Amphibians possess quite a simple brain with little neuronal migration outside the in- ternal ventricular zone, and almost no expansion of cell masses. Their pallium has been subdivided into medial, dorsal, and lateral components. On the other hand, the pallium of reptiles and birds (saur- opsida) displays a cortical, laminated architecture containing a medial/dorsomedial (hippocampal) cor- tex, a lateral (olfactory) cortex, and a small dorsal cortex interposed between these two (in birds, the region corresponding to the dorsal cortex has been termed the Wulst, or more recently, the hyperpallium). These components correspond to the different pallial subdivisions in amphibians. In addition, the brains of sauropsids display a conspicuous nuclear structure that bulges into the ventricle, the dorsal ventricular ridge (DVR), which receives many thalamic sensory projec- tions (Figure 1). Mammals are characterized by the presence of a neocortex (Figure 1). This structure is bound medially by the medial cortex or hippocampus, and laterally by the lateral or olfactory cortex. Deep to some cortical areas like the insular region there are structures such as the claustral complex. These and other dorsal structures make up the pallium, while the ventral subpallium includes the basal ganglia and related nuclei. The neocortex of mammals is indeed a unique structure, consisting of six laminae that remain some- what conserved across species and cortical regions. Furthermore, it is subdivided into several distinct areas in the tangential domain, some of them being sensory, others motor, and still others receiving multi- modal input. In several mammalian groups there is a clear trend of tangential neocortical expansion that results in the increase of both surface area (leading to the formation of fissures and convolutions) and the number of cortical areas. The neocortex receives most of its extrinsic input from dorsal thalamic nuclei, many of them conveying sensory information from somatosensory, auditory, or visual modalities, and projects back to the thalamus, to the amygdala, to the entorhinal cortex (which in turn projects to the hippocampus), and to several subcortical structures, including the spinal cord. Historical Background Early in the last century, concepts on the evolution of the amniote brain (i.e., reptiles, birds, and mammals) were dominated by Elliot Smith’s interpretation that sauropsids were characterized by the expansion of subpallial structures like the basal ganglia, leading to the development of the DVR. On the other hand, mammals would have expanded the pallial re- gion, producing the neocortex. Interestingly, other authors, such as Nils Holmgren and Bengt Ka ¨lle ´n, proposed a pallial origin for the reptilian DVR, which they described as arising from a deep prolifera- tive region that overlaps with the lateral cortex. Holmgren termed this region as the hypopallium, and considered it to be related to deep-brain mamma- lian structures like the claustrum or parts of the amyg- dalar complex. However, these views were eclipsed by Elliot Smith’s claim for a subpallial origin of the DVR. Subsequent analyses performed in the 1970s, taking advantage of newly developed histochemical proce- dures, revealed that the reptilian DVR does not con- tain acetylcholinesterase activity as the basal ganglia do, and is therefore a pallial structure. Furthermore, tract-tracing experiments performed by Harvey Karten and collaborators, showed that the avian DVR (more specifically, its anterior part, ADVR, or nidopallium) received thalamic sensory projections similar to those reaching some parts of the mammalian neocortex. More precisely, collothalamic nuclei (i.e., thalamic nuclei receiving input from mesencephalic sensory centers, conveying visual and auditory information) project to the ADVR/nidopallium of birds, and to the lateral neocortex of mammals (visual extrastriate and Neocortex: Origins 43
-
Upload
nguyendung -
Category
Documents
-
view
219 -
download
0
Transcript of Neocortex: Origins - Queen's Universitybrain.phgy.queensu.ca/pare/assets/Circuits2a.pdf ·...

Neocortex: OriginsF Aboitiz, Pontificia Universidad Catolica de Chile,Santiago, Chile
ã 2009 Elsevier Ltd. All rights reserved.
Introduction
The origin of the mammalian neocortex is probablyone of the most controversial themes in moderncomparative neurobiology. There have been long-standing arguments on this issue (dating from earlylast century), and still there is no general consensusabout it. This is partly due to the enormous anatomi-cal diversification that is observed in the cerebralhemispheres of distinct vertebrate groups. Startingfrom a relatively conserved embryonic plan thatdivides the hemispheres into a dorsal pallium and aventral subpallium, different brain components haveexpanded in each vertebrate class, producing pro-found distortions of the ancestral adult condition.For example, cartilaginous fishes are characterizedby the development of a central pallial nucleus locatedat the dorsal midline, while bony fish develop abrain in which the ventricular region everts to theoutside, separating the medial hemispheres inthe dorsal aspect. Amphibians possess quite a simplebrain with little neuronal migration outside the in-ternal ventricular zone, and almost no expansion ofcell masses. Their pallium has been subdivided intomedial, dorsal, and lateral components. On theother hand, the pallium of reptiles and birds (saur-opsida) displays a cortical, laminated architecturecontaining a medial/dorsomedial (hippocampal) cor-tex, a lateral (olfactory) cortex, and a small dorsalcortex interposed between these two (in birds, theregion corresponding to the dorsal cortex has beentermed the Wulst, or more recently, the hyperpallium).These components correspond to the different pallialsubdivisions in amphibians. In addition, the brains ofsauropsids display a conspicuous nuclear structure thatbulges into the ventricle, the dorsal ventricular ridge(DVR), which receives many thalamic sensory projec-tions (Figure 1).Mammals are characterized by the presence of a
neocortex (Figure 1). This structure is bound mediallyby the medial cortex or hippocampus, and laterally bythe lateral or olfactory cortex. Deep to some corticalareas like the insular region there are structuressuch as the claustral complex. These and other dorsalstructures make up the pallium, while the ventralsubpallium includes the basal ganglia and relatednuclei. The neocortex of mammals is indeed a unique
structure, consisting of six laminae that remain some-what conserved across species and cortical regions.Furthermore, it is subdivided into several distinctareas in the tangential domain, some of them beingsensory, others motor, and still others receiving multi-modal input. In several mammalian groups there is aclear trend of tangential neocortical expansion thatresults in the increase of both surface area (leading tothe formation of fissures and convolutions) and thenumber of cortical areas. The neocortex receives mostof its extrinsic input from dorsal thalamic nuclei,many of them conveying sensory information fromsomatosensory, auditory, or visual modalities, andprojects back to the thalamus, to the amygdala, tothe entorhinal cortex (which in turn projects to thehippocampus), and to several subcortical structures,including the spinal cord.
Historical Background
Early in the last century, concepts on the evolution ofthe amniote brain (i.e., reptiles, birds, and mammals)were dominated by Elliot Smith’s interpretation thatsauropsids were characterized by the expansion ofsubpallial structures like the basal ganglia, leadingto the development of the DVR. On the otherhand, mammals would have expanded the pallial re-gion, producing the neocortex. Interestingly, otherauthors, such as Nils Holmgren and Bengt Kallen,proposed a pallial origin for the reptilian DVR,which they described as arising from a deep prolifera-tive region that overlaps with the lateral cortex.Holmgren termed this region as the hypopallium,and considered it to be related to deep-brain mamma-lian structures like the claustrum or parts of the amyg-dalar complex. However, these views were eclipsed byElliot Smith’s claim for a subpallial origin of the DVR.
Subsequent analyses performed in the 1970s, takingadvantage of newly developed histochemical proce-dures, revealed that the reptilian DVR does not con-tain acetylcholinesterase activity as the basal gangliado, and is therefore a pallial structure. Furthermore,tract-tracing experiments performed byHarvey Kartenand collaborators, showed that the avian DVR (morespecifically, its anterior part, ADVR, or nidopallium)received thalamic sensory projections similar to thosereaching some parts of the mammalian neocortex.More precisely, collothalamic nuclei (i.e., thalamicnuclei receiving input from mesencephalic sensorycenters, conveying visual and auditory information)project to the ADVR/nidopallium of birds, and to thelateral neocortex of mammals (visual extrastriate and
Neocortex: Origins 43
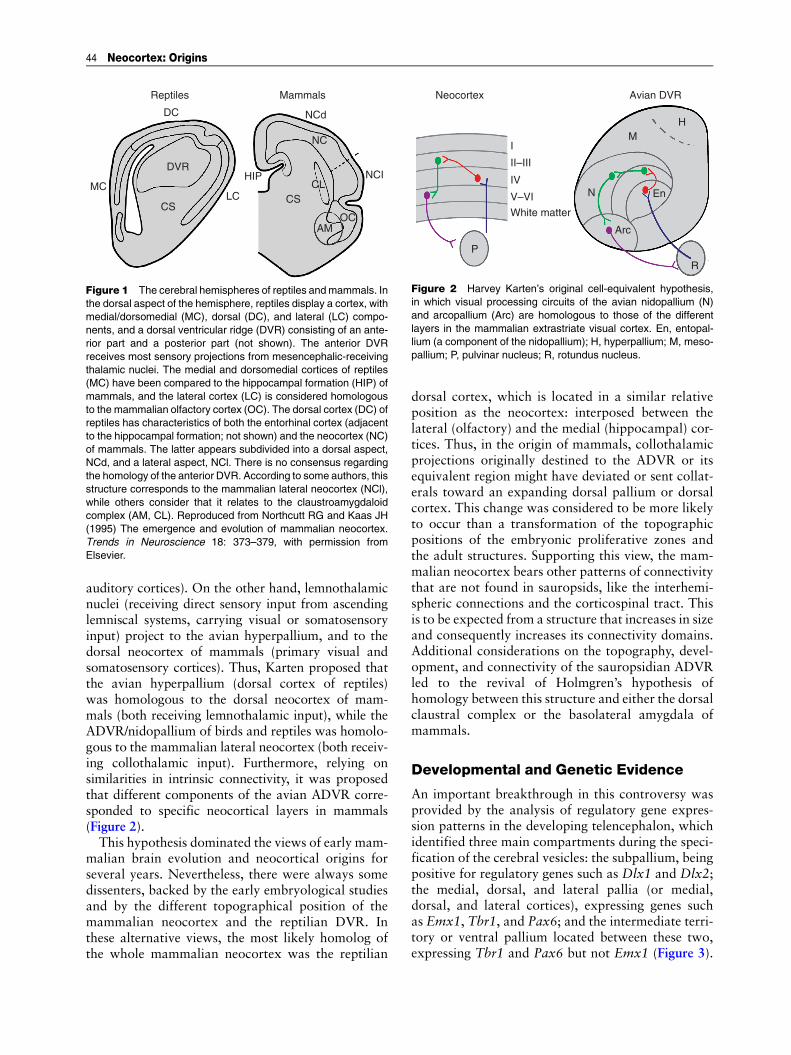
auditory cortices). On the other hand, lemnothalamicnuclei (receiving direct sensory input from ascendinglemniscal systems, carrying visual or somatosensoryinput) project to the avian hyperpallium, and to thedorsal neocortex of mammals (primary visual andsomatosensory cortices). Thus, Karten proposed thatthe avian hyperpallium (dorsal cortex of reptiles)was homologous to the dorsal neocortex of mam-mals (both receiving lemnothalamic input), while theADVR/nidopallium of birds and reptiles was homolo-gous to the mammalian lateral neocortex (both receiv-ing collothalamic input). Furthermore, relying onsimilarities in intrinsic connectivity, it was proposedthat different components of the avian ADVR corre-sponded to specific neocortical layers in mammals(Figure 2).This hypothesis dominated the views of early mam-
malian brain evolution and neocortical origins forseveral years. Nevertheless, there were always somedissenters, backed by the early embryological studiesand by the different topographical position of themammalian neocortex and the reptilian DVR. Inthese alternative views, the most likely homolog ofthe whole mammalian neocortex was the reptilian
dorsal cortex, which is located in a similar relativeposition as the neocortex: interposed between thelateral (olfactory) and the medial (hippocampal) cor-tices. Thus, in the origin of mammals, collothalamicprojections originally destined to the ADVR or itsequivalent region might have deviated or sent collat-erals toward an expanding dorsal pallium or dorsalcortex. This change was considered to be more likelyto occur than a transformation of the topographicpositions of the embryonic proliferative zones andthe adult structures. Supporting this view, the mam-malian neocortex bears other patterns of connectivitythat are not found in sauropsids, like the interhemi-spheric connections and the corticospinal tract. Thisis to be expected from a structure that increases in sizeand consequently increases its connectivity domains.Additional considerations on the topography, devel-opment, and connectivity of the sauropsidian ADVRled to the revival of Holmgren’s hypothesis ofhomology between this structure and either the dorsalclaustral complex or the basolateral amygdala ofmammals.
Developmental and Genetic Evidence
An important breakthrough in this controversy wasprovided by the analysis of regulatory gene expres-sion patterns in the developing telencephalon, whichidentified three main compartments during the speci-fication of the cerebral vesicles: the subpallium, beingpositive for regulatory genes such as Dlx1 and Dlx2;the medial, dorsal, and lateral pallia (or medial,dorsal, and lateral cortices), expressing genes suchas Emx1, Tbr1, and Pax6; and the intermediate terri-tory or ventral pallium located between these two,expressing Tbr1 and Pax6 but not Emx1 (Figure 3).
I
II–III
IV
V–VIWhite matter
Neocortex Avian DVR
P
R
Arc
N
MH
En
Figure 2 Harvey Karten’s original cell-equivalent hypothesis,in which visual processing circuits of the avian nidopallium (N)and arcopallium (Arc) are homologous to those of the differentlayers in the mammalian extrastriate visual cortex. En, entopal-lium (a component of the nidopallium); H, hyperpallium; M, meso-pallium; P, pulvinar nucleus; R, rotundus nucleus.
DC
DVR
MC
CSLC
HIP
NCd
NCI
NC
Reptiles Mammals
CLCS
AMOC
Figure 1 The cerebral hemispheres of reptiles andmammals. Inthe dorsal aspect of the hemisphere, reptiles display a cortex, withmedial/dorsomedial (MC), dorsal (DC), and lateral (LC) compo-nents, and a dorsal ventricular ridge (DVR) consisting of an ante-rior part and a posterior part (not shown). The anterior DVRreceives most sensory projections from mesencephalic-receivingthalamic nuclei. The medial and dorsomedial cortices of reptiles(MC) have been compared to the hippocampal formation (HIP) ofmammals, and the lateral cortex (LC) is considered homologousto the mammalian olfactory cortex (OC). The dorsal cortex (DC) ofreptiles has characteristics of both the entorhinal cortex (adjacentto the hippocampal formation; not shown) and the neocortex (NC)of mammals. The latter appears subdivided into a dorsal aspect,NCd, and a lateral aspect, NCl. There is no consensus regardingthe homology of the anterior DVR. According to some authors, thisstructure corresponds to the mammalian lateral neocortex (NCl),while others consider that it relates to the claustroamygdaloidcomplex (AM, CL). Reproduced from Northcutt RG and Kaas JH(1995) The emergence and evolution of mammalian neocortex.Trends in Neuroscience 18: 373–379, with permission fromElsevier.
44 Neocortex: Origins

Interestingly, the ventral pallium maps into parts ofthe claustrum and the amygdala of mammals and intothe DVR of sauropsids, indicating a common devel-opmental origin for these structures, and supportingembryonic homology between the territories pro-ducing the reptilian ADVR and parts of the claus-troamygdalar complex of mammals. Nevertheless,it is not clear that in the adult state there is anyprecise homology between the different ventral pallialderivatives in sauropsids and mammals, as both
structures are likely to have undergone distinct evolu-tionary histories. In fact, fossil evidence indicates thatthe lineage of mammal-like reptiles diverged quiteearly from the reptilian line, and that their brainswere extremely simple in their gross morphology,resembling more the brains of modern amphibiansthan those of modern reptiles.
In an attempt to accommodate these data toKarten’s original hypothesis, it has been proposedthat the lateral neocortex of mammals arose by virtueof a transformation of a ventral pallial structure intoa dorsal pallial phenotype. This may have resultedeither from the acquisition of dorsal characteristicsof a yet unspecified territory, or from duplication ofthe ventral pallium and subsequent transformationof its dorsal moiety. Nevertheless, these hypothesesmight be difficult to refute, as any dorsal pallial fea-ture in the lateral neocortex could be consideredto result from the transformation from a ventral toa dorsal pallial structure. Instead, these proposalsrequire positive evidence, such as a direct sign ofventral pallial origin in the lateral neocortex.
Reptilian Dorsal Cortex: Neocortical andEntorhinal/Subicular Features
Furthermore, the presumed homology – betweenvisual and somatosensory areas in the reptilian dorsalcortex, and primary visual and somatosensory areasof the mammalian neocortex – has been questionedby some authors. For example, somatosensory pro-jections to the dorsal cortex have not been observedin either lizards or turtles, and thus may not representan ancestral character of sauropsids. In addition,visual projections to the dorsal pallium reach a regionthat in turtles includes the dorsal cortex and thepallial thickening, and in lizards, are restricted tothe pallial thickening. The pallial thickening is locat-ed in the lateral aspect of the pallium of some reptilesand its homologies and embryonic origins are unclear.Moreover, the multimodal dorsomedial pallium ofamphibians and reptiles has several features reminis-cent of the mammalian entorhinal cortex. Like thelatter, the reptilian dorsal cortex has important con-nections with the medial/dorsomedial or hippocam-pal cortices. In addition, the reptilian dorsal cortexdoes not participate in vision but rather in spatiallearning and memory. Thus, the dorsal cortex ofreptiles shares features with the entorhinal/subicularcortex of mammals, but also contains a direct visual(and sometimes somatosensory) sensory input that,if perhaps not being strictly equivalent to the mam-malian visual (or somatosensory) cortex, may serve tofeed sensory information to the hippocampus forestablishing sensory associations during learning.
DPvI
vI
vI
vc
vc
vc
ADVRa
a
a
SP
AM
SP
SP
N
LP Reptiles
Birds
Mammals
W
M
Cx
Lateral, dorsal, andmedial pallium
Intermediate territoryand ventral pallium
Subpallium
Emx1+/Tbr1+/Pax6+
Tbr1+/Pax6+
DIx-2+
Figure 3 Patterns of regulatory gene expression in the devel-oping telencephalon of sauropsids and mammals. All pallial com-ponents (medial, dorsal, and lateral; blue) express the markersEmx1, Tbr1, and Pax6, with the exception of the ventral pallium(yellow), which is positive for Tbr1 and Pax6 but not for Emx1. Thesubpallium (SP, red) expresses Dlx genes. According to thisevidence, the ventral pallium gives rise to the reptilian anteriordorsal ventricular ridge (ADVR), to the avian nidopallium (N), andto the claustroamygdalar complex (AM) of mammals. a,vc, termination of auditory and visual collothalamic pathways,respectively; vl, termination of visual lemniscal pathways; DP,dorsal pallium; LP, lateral pallium; Cx, cortex; M, mesopallium;W, Wulst or hyperpallium. Adapted from Aboitiz F, Morales D, andMontiel J (2003) The evolutionary origin of the mammalian iso-cortex: Towards an integrated developmental and functionalapproach. Behavioral and Brain Sciences 26: 535–586, with per-mission from Cambridge University press.
Neocortex: Origins 45

The Dorsalization Hypothesis
Considering the evidence discussed herein, Aboitizand collaborators proposed that the mammalian neo-cortex arose as an expansion of an ancestral pallialregion with dorsal characteristics, which acquired amassive lemnothalamic and collothalamic input. Thismay be conceived as a dorsalization effect, in whichthe activity of genes (FGFs,Wnts, Emxs, Lhx2, Pax6,and others) or signaling pathways involved in dorsalpallial specification was enhanced, thus producingan increase in proliferative activity leading to theexpansion of a primitive dorsal cortex into a neocor-tex. More precisely, the embryonic dorsal palliumrepresents a transitional zone that receives influencefrom several signaling sources, including the anteriorforebrain expressing FGF8; the medially locatedcortical hem expressing Wnts and BMPs, and regulat-ing other genes like Emxs (these signals specify boththe medial pallium and the neocortex); and the Pax6-dependent antihem that specifies the lateral and ven-tral pallium. The expansion of this transitional tissuemay have somehow resulted from the interactionbetween these three morphogenetic fields.This process was not only the result of a develop-
mental transformation; probably it was also favoredfor functional reasons. As nocturnal animals, earlymammals probably had refined senses of smell andhearing, which was reflected in the development of alarge olfactory bulb, a middle ear, and external outerears. Strong associative networks involving olfactionand other senses developed in the dorsal pallium andthe hippocampus, which participated in the establish-ment of learned maps of space. These networks bene-fited from the expansion of the dorsal pallium, whichin turn facilitated the projection of collothalamicpathways that were originally directed to the ventralpallium or to the subpallium into this structure.
Lamination of the Neocortex: ThePreplate
The neocortex is not only the result of the tangentialexpansion of the dorsal pallium. There is also a dra-matic increase in radial thickness, as the reptiliancortex is only a few cells deep while the mammalianneocortex is organized in vertical columns about100 cells in depth. As mentioned, the adult neocortexconsists of six laminae, the most superficial (layer I)being a cell-poor layer in which tangential axonsfrom Martinotti cells and other axons run alongvarious distances. The development of the neocortexis quite a conspicuous one, in which the first cellsto arrive make up a transient embryonic structure
termed the preplate, consisting of Cajal–Retzius cellsand pioneer neurons. The former derive in large partfrom the medially located cortical hem and migratelaterally into the neocortex, while pioneer neuronsderive mainly from the subpallium. Reelin-secretingCajal–Retzius cells are involved in radial glia mainte-nance and in controlling the latest steps of neuronalmigration to the cortex, and pioneer neurons displayan axon directed subcortically that acts as a guide forgrowing thalamocortical axons. Subsequently, radi-ally migrating neurons that make up the true neocor-tical plate arrive from the cortical ventricular zoneand split the preplate into two layers: a superficialmarginal zone, containing the Cajal–Retzius cells andother cells, and a deep subplate below the emergingcortical plate, containing most pioneer neurons andother cells such as excitatory neurons (Figure 4).The latter receive transient contacts from arrivingthalamic axons before they enter into the maturecortex. Most of the cells in the marginal zone andthe subplate die during development or dilute withcortical expansion, while cells of the cortical plateremain in adulthood. Interestingly, the developmentof the cortical plate occurs in an inside-out sequencein which the earliest-produced cells end up in thedeepest cortical layers, and the latest-produced cells
Figure 4 Lamination of the mammalian neocortex. The earliestcells to arrive at the embryonic cortex make up a preplate (notshown). The preplate becomes divided into two layers: a superfi-cial marginal zone (MZ, future layer I), containing Cajal–Retziuscells (diamond-shaped cells above, brown) that secrete reelin(yellow stars), and a deep subplate (SP), containing pioneer neu-rons (diamond-shaped cells below, red) whose axon serves as aguide for growing thalamocortical axons. Cells that locate be-tween these layers make up the cortical plate (green ovals) andarrange in five laminae (VI–II in the mature neocortex). Theselaminae are generated in an inside-out sequence in which early-produced neurons locate above late-produced ones (arrow repre-sents progressively later neurogenetic times).
46 Neocortex: Origins

end up in the more superficial layers. This implies thatlate-produced neurons have to cross the layers ofearly-produced neurons and locate above them.Additionally, inhibitory neurons originating in thesubpallium populate the developing cortical plate,also following an inside-out gradient in whichearly-produced cells arrive at deeper layers and late-produced cells arrive at superficial layers. This wholeprocess contrasts with the situation of the much thin-ner reptilian cortex, where cells migrate in an outside-in sequence, early layers locating more superficiallythan late-produced layers.Is there any correspondence between the reptilian
cortical cells and the different layers making upthe mammalian neocortex? Miguel Marın-Padillaoriginally proposed that neurons of the mammalianpreplate (more specifically, those making up the sub-plate) were homologous to the reptilian cortex, thecortical plate being a developmental addition to themammalian brain. However, other authors pro-posed that the mammalian preplate and the subplateemerged as devices to support the development ofcorticocortical connectivity in mammals. Insteadof corresponding to the mammalian preplate, thereptilian cortex was claimed to be homologous tothe deep neocortical layers V and (especially) VI ofthe mammalian cortical plate.There are reports of a subplate-like structure in rep-
tiles consisting of tangentially migrating interneurons,but no excitatory neurons have yet been described in it.On the other hand, in a marsupial, no subplate wasdetected and corticothalamic axons were seen directlypenetrating the cortical plate, instead of establishingtransient contacts below it as they do in placentalmammals. If this observation is correct and true forall marsupials, it would imply that at least part of thesubplate (its excitatory component, which receivesarriving thalamic axons) is an acquisition of placentalmammals. Moreover, in some mammals, includingsome marsupials, there is an adult cortical layer inferi-or to layer VI, termed layer VIb or VII, which containspersisting subplate cells. These cells have been consid-ered to be closely related to the deepest neurons in thecortical plate. Furthermore, there is molecular evi-dence for an intimate relation between the inferiorlayers of the cortical plate and the subplate. The geneTbr1 marks both subplate neurons and the earliestcortical plate neurons, and mutations in this genecause defects in both structures. Thus, in the origin ofmammals (or of placental mammals), a subplate-likestructure containing tangentially migrating neuronsmight have incorporated excitatory cells that originallybelonged to the cortical plate, that aided in theguidance of thalamocortical axons.
Origin of the Superficial NeocorticalLayers and the Inside-OutNeurogenetic Gradient
Additional molecular evidence supports the concept ofthe superficial neocortical layers IV–II as developmen-tally distinct from the inferior layers VI and V. Amongother features, mutants for the gene Pax6 exhibitcortical migration defects, consisting of the inabilityto migrate and differentiate supragranular neocorticalneurons belonging to layers IV to II. Furthermore,early-produced cortical plate neurons destined to neo-cortical layers VI and V are produced in the embryon-ic ventricular zone and express the marker geneOtx1.On the other hand, late-produced neurons, which de-pend onPax6 for their development and are destined tosuperficial layers IV–II, are produced in the subventri-cular zone surrounding the ventricular zone, and ex-press the marker gene Svet1. Thus, Pax6 may beinvolved in the origin of the superficial neocorticallayers by recruiting the subventricular zone in the neu-rogenetic process. Further findings indicate that theproneural genes Ngn1 and Ngn2 specify the laminarcharacteristics of early-born cortical neurons, butupper-layer cortical neurons are specified in an Ngn-independent manner, requiring instead the synergisticactivities of Pax6 and Tlx. There is also comparativeevidence indicating a progressive development of thesubventricular zone in large-brainedmammals, its nearabsence in the reptilian embryonic dorsal cortex, andits independent development in the avian nidopallium.It will be of interest to determine what, if any, is thecontribution of the subventricular zone and of thegenes Pax6, Svet1, and Tlx to dorsal cortical develop-ment in reptiles.
In this context, the effect of mutations in somegenes involved in the regulation of cortical cell migra-tion has provided further clues to the development ofthe laminar organization of the neocortex. For exam-ple, the reeler mutant mouse, deficient in the extra-cellular protein reelin, shows a striking inversion ofthe developmental gradient in the cortical plate, inwhich early-produced neurons become located super-ficially and late-produced neurons arrange in theinferior layers. Furthermore, in this mutant the pre-plate is not split and is compressed above the corticalplate. In the adult, no cell-poor layer I is detectable, asthe cortical plate cells are observed immediatelybelow the subpial layer. Electron microscopy analysesindicate abnormal attachments between the neuronsand the radial glia, suggesting that these neurons didnot detach correctly from the latter. There have beenseveral hypotheses for the participation of reelin inthe control of neuronal migration, including a role as
Neocortex: Origins 47

a stop signal, as a glial detachment signal, or as anattractor for migrating neurons. Overall, the evidenceindicates that reelin has a role in the final stagesof neuronal migration and, more important to ourpurposes, in detaching migrating neurons from theradial glia, and in the generation of a cell-free super-ficial marginal zone. Reelin is present in several otherbrain regions besides the cerebral cortex, and its pat-terns of expression reflect an evolutionarily stableframework. However, reelin expression is stronglyamplified in the mammalian cortical marginal zonein relation to the reptilian marginal zone, indicatingthat this molecule has been one key element in theorigin of the neocortex. Furthermore, the level ofreelin expression correlates with the degree of colum-narity present in the dorsal cortex of different reptiles,suggesting that it may have contributed to the radialorganization of the mammalian neocortex.Another molecule of interest is the cyclin-
dependant kinase cdk5; cdk5 and its activator, p35,are fundamental for the adequate lamination of theneocortex. Recently, two modes of radial migrationhave been characterized: nuclear translocation, inwhich the leading process of the neuron is attachedto the subpial layer and the cell body is draggedtoward the periphery as a consequence of the shorten-ing of this process; and cellular locomotion, in whichcells migrate along the radial glia in their pathwayto the cortex. Interestingly, early-produced, deep-layer cortical neurons migrate mainly by transloca-tion, while late-produced, superficial neurons useglial-guided locomotion to reach the cortical plate.Mutants for the enzyme cdk5 show strong deficits inlocomotion, and therefore the superficial layers of thecortex fail to form. In these animals, neurons are notable to reach the cortical plate by birth and remain inthe subcortical intermediate zone. However, early-produced layer VI cells which migrate by transloca-tion do so correctly and are able to split the preplatenormally. Moreover, mutants for the cdk5 activatorp35 display milder defects in locomotion, and neu-rons from all layers are able to reach the cortical plateby birth. However, within the cortical plate, cellsarrange in an inverted outside-in gradient reminiscentof the reeler mouse, but with the difference that thepreplate has been partially split and there is a normalmarginal zone. N-Cadherin-mediated cell aggrega-tion in migrating neurons is inhibited by cdk5/p35,which provides a possible mechanism by whichnormally migrating cells may use this cascade to by-pass a migration-suppressing signal (N-cadherin or arelated molecule) present in postmigratory neurons,tomove above them. Considering that early-producedcortical plate cells are more likely to correspondto reptilian cortical cells, it is conceivable that the
cdk5-independent mechanism of translocation repre-sents an ancestral condition, while the activationof the cdk5 pathway represents an evolutionary inno-vation of the mammalian brain, associated with newmodes of neuronal migration and, by virtue of theactivator p35, contributing to the generation ofthe inside-out neurogenetic gradient that is character-istic of the neocortex.
A Scenario for the Origin of NeocorticalLamination
Based on this evidence, a possible scenario for theevolution of cortical lamination is that initially therewas a radial expansion of the primitive mammaliancortex, producing late-born, phylogenetically newneurons which participated in local processing cir-cuits, as opposed to the older neurons that functionas an output layer. These new elements originated inan expanded subventricular zone, by virtue of theaction of the gene Pax6, among others. The acquisi-tion of an inside-out neurogenetic gradient was possi-bly related to the ancestral organization of afferentsto the primitive cortex. The neocortex differs fromother structures, including the hippocampus, theolfactory cortex, and the reptilian cortex, in thestrong radial organization of its inputs, in whichafferents penetrate the cortical plate from the under-lying white matter (aided by subplate cells and pio-neer neurons); in these other structures the afferentsarrange tangentially in the superficial marginal zone(Figure 5). Radial expansion of the primitive cortex inthe context of a primitive outside-in developmentalpattern would have left the new, local processingneurons located below the early-produced outputneuronal layer, separating them from the superficiallylocated afferents. In such context, there would be abenefit to those individuals in which late-producedneurons were able to cross the layer of early-producedcells, generating an intermediate layer between theinput axons and the output cells. In other words, theinside-out neurogenetic gradient was produced inorder to maximize synaptic contacts between thenew, late-born neurons, and the superficially locatedcortical afferents. In this process, the cdk5 signalingpathway facilitated new mechanisms of radial migra-tion, and especially its activator p35 was important inpermitting late-born neurons to cross the barrierformed by previously migrated cells. Finally, animportant increase in the expression of reelin byCajal–Retzius cells permitted the preservation of aneuron-free marginal zone. Reelin also contributedto stop radial migration by detaching migrating neu-rons from the radial glia, thus facilitating the migra-tion of young neurons past old ones.
48 Neocortex: Origins

As the neocortex expanded tangentially, however,the distance traveled by tangential afferent axonsbecame increasingly longer, with a consequent im-pairment in transmission time. A solution to this prob-lem was provided by the rerouting of these axonsvia the subcortical white matter, making a shortcutto their final targets. This was possible by virtueof the displacement of the pioneer neurons attractingthe thalamic afferents, from a superficial positionin the marginal zone of reptiles, to a deeper posi-tion in the subplate of mammals. Thus, there wereseveral steps in the acquisition of a neocortex, includ-ing the increase in cortical thickness, the origin of theinside-out gradient, and the displacement of axonaltrajectories from a superficial to a subcortical posi-tion. Interestingly, intermediate stages may be foundbetween the reptilian cortex, displaying superficial,tangential inputs, and the advanced mammalian neo-cortex, with radially organized inputs. For example,like the neocortex, the hippocampus has an inside-outdevelopmental gradient, but also a superficial localiza-tion of afferents, indicating a step above the reptiliancortex. A subsequent stage may be represented bythe neocortex of the primitive insectivore Erinaceus,in which axons penetrate the cortex radially fromthe underlying white matter, cross obliquely to themarginal zone, and run tangentially there for somedistance. Finally, in the sensory neocortex of ad-vanced mammals, afferents enter the cortex radially
and synapse in spiny stellate neurons of layer IV(Figure 5).
Final Comments
Summarizing the views presented here, the mammali-an neocortex displays several unique characteristicsand therefore the term ‘neocortex’ is indeed appropri-ate. One main dicussion in relation to its origins con-cerns the ancestral structure that gave rise to it. Thereare two main proposals in this regard, one that claimsthat the neocortex partly arose from the DVR ofreptiles, and the other that prescribes that it origi-nated as an expansion of the dorsal pallium. Currentdevelopmental evidence tends to favor the latter, asthe reptilian DVR has embryonic origins similar tothose of the mammalian claustroamygdalar complexin the ventral pallium. The neocortex is also unique inits laminar structure. Recent genetic and developmen-tal analyses have provided important insights as tothe mechanisms and possible sequence of events thatgave rise to the neocortex, producing its characteristicinside-out neurogenetic gradient and the reroutingof thalamic afferents from a superficial to a deep,subcortical position. All this evidence strengthensthe view that development may not only give cluesto evolutionary processes, but also that variations inthe developmental processes are the main events inthe generation of new structures in evolution.
Figure 5 Postulated sequence in the evolution of neocortical lamination. In a primitive state (I) like that of reptilian cortex, with anoutside-in neurogenetic gradient (arrow downwards), pioneer neurons (pointed ovals, red) locate in the superficial marginal zone andguide thalamocortical axons (Y-shaped lines) tangentially above the cortex (ovals, blue, green, and yellow). A second stage (II) may beexemplified by the hippocampus, which already displays an inside-out gradient (arrow upwards) while its afferents are located superficially.The neocortex of the hedgehog (Insectivora) represents a third stage (III), where axons enter the cortical plate from below but many runobliquely and end tangentially in the marginal zone. Finally (IV), the primary sensory cortex of advanced mammals displays axons radiallyentering the cortex and synapsing with neurons in layer IV, and a large proportion of pioneer neurons locate in the subplate below the cortex.
Neocortex: Origins 49

See also: Cerebral Cortex; Evolution of Vertebrate Brains;Homology and Homoplasy.
Further Reading
Aboitiz F, Montiel J, and Garcıa RR (2005) Ancestry of the mam-malian preplate and its derivatives: Relicts or embryonic adap-tations. Reviews in the Neurosciences 16: 359–376.
Aboitiz F, Morales D, andMontiel J (2003) The evolutionary originof the mammalian isocortex: Towards an integrated develop-mental and functional approach. Behavioral and Brain Sciences26: 535–586.
Bar I, Lambert de Rouvroit C, and Goffinet A (2000) The evolutionof cortical development. An hypothesis based on the reelinsignaling pathway. Trends in Neuroscience 23: 633–638.
Bruce LL and Neary TJ (1995) The limbic system of tetrapods:A comparative analysis of cortical and amygdalar populations.Brain, Behavior and Evolution 46: 224–234.
Butler AB and Hodos W (1996) Comparative Vertebrate Neuro-anatomy. Evolution and Adaptation. New York: Wiley-Liss.
Caviness VS, Crandall JE, and Edwards MA (1988) The reelermalformation. Implications for neocortical histogenesis. In:Peters A and Jones EG (eds.) Cerebral Cortex, Vol. 7. Develop-ment and Maturation of the Cerebral Cortex, pp. 59–89. NewYork: Plenum.
Doron NN and LeDoux JE (1999) Organization of projections tothe lateral amygdala from auditory and visual areas of thethalamus in the rat. Journal of Comparative Neurology 412:383–409.
Goffinet AM, Daumerie C, Langerwerf B, and Pieau C (1986)Neurogenesis in reptilian cortical structures: 3H-thymidineautoradiographic analysis. Journal of Comparative Neurology243: 106–116.
Karten HJ (1997) Evolutionary developmental biology meets thebrain: The origins of mammalian neocortex. Proceedings of theNational Academy of Sciences of the United States of America94: 2800–2804.
KriegsteinARandNoctor SC (2004) Patterns of neuronalmigrationin the embryonic cortex. Trends in Neuroscience 27: 392–399.
Marın-Padilla M (1978) Dual origin of the mammalian neocortexand evolution of the cortical plate. Anatomy and Embryology152: 109–126.
Molnar Z, Metin C, Stoykova A, et al. (2006) Comparative aspectsof cerebral cortical development. European Journal of Neuro-science 23: 921–934.
Nadarajah B and Parnavelas JG (2002) Modes of neuronal migra-tion in the developing cerebral cortex. Nature Reviews Neuro-science 3: 423–432.
Northcutt RG and Kaas JH (1995) The emergence and evolution ofmammalian neocortex. Trends in Neuroscience 18: 373–379.
Powers AS (1990) Brain mechanisms of learning in reptiles. In:Kesner RP and Olton DS (eds.) Neurobiology of ComparativeCognition, pp. 75–87. New York: Erlbaum Press.
Puelles L, Kuwana E, Puelles E, et al. (2000) Pallial and subpallialderivatives in the embryonic chick and mouse telencephalon,traced by the expression of the genes Dlx-2, Emx-1, Nkx-2.1,Pax-6 and Tbr-1. Journal of Comparative Neurology 424:409–438.
Schuurmans C, Armant O, Nieto M, et al. (2004) Sequentialphases of cortical specification involve neurogenin-dependentand -independent pathways. EMBO Journal 23: 2892–2902.
SmithFernandez A, Pieau C, Reperant J, et al. (1998) Expression ofthe Emx-1 and Dlx-1 homeobox genes defines three molecularlydistinct domains in the telencephalon of mouse, chick, turtleand frog embryos: Implications for the evolution of telencephalicsubdivisions in amniotes. Development 125: 2099–2111.
Striedter GF (2005) Principles of Brain Evolution. Sunderland,MA: Sinauer Associates.
Super H and Uylings HB (2001) The early differentiation of theneocortex: A hypothesis on neocortical evolution. CerebralCortex 11: 1101–1109.
Sur M and Rubenstein JL (2005) Patterning and plasticity of thecerebral cortex. Science 310: 805–810.
Voogd J, Nieuwenhuys R, and VanDongen PAM (1998) Mammals.In: Nieuwenhuys R, Ten Donkelaar HJ, and Nicholson C (eds.)The Central Nervous System of Vertebrates, vol. 3, pp. 1637–2098. Berlin: Springer.
50 Neocortex: Origins


















