
Nematode resistance faisal
42
Achievements and recent developments of genetic engineering in agriculture for nematode resistance Course Teacher Dr. V. Balasubramanian Student Mohammed Faisal P
-
Upload
mohammed-faisal-peeran -
Category
Education
-
view
23 -
download
3
Transcript of Nematode resistance faisal

Achievements and recent developments of genetic engineering in agriculture for
nematode resistance
Course TeacherDr. V. Balasubramanian
StudentMohammed Faisal P

Nematodes cause annual losses of over US $lOO billion to world agriculture.
Control measures are inadequate, and are too heavily dependent on environmentally hazardous nematicides.
Transgenic-plant technology offers a ‘green’ solution for controlling nematodes
(Atkinson et al., 1995)
Introduction


Mutation
Protoplast fusion
Genetic engineering
Genetic improvement of plants.
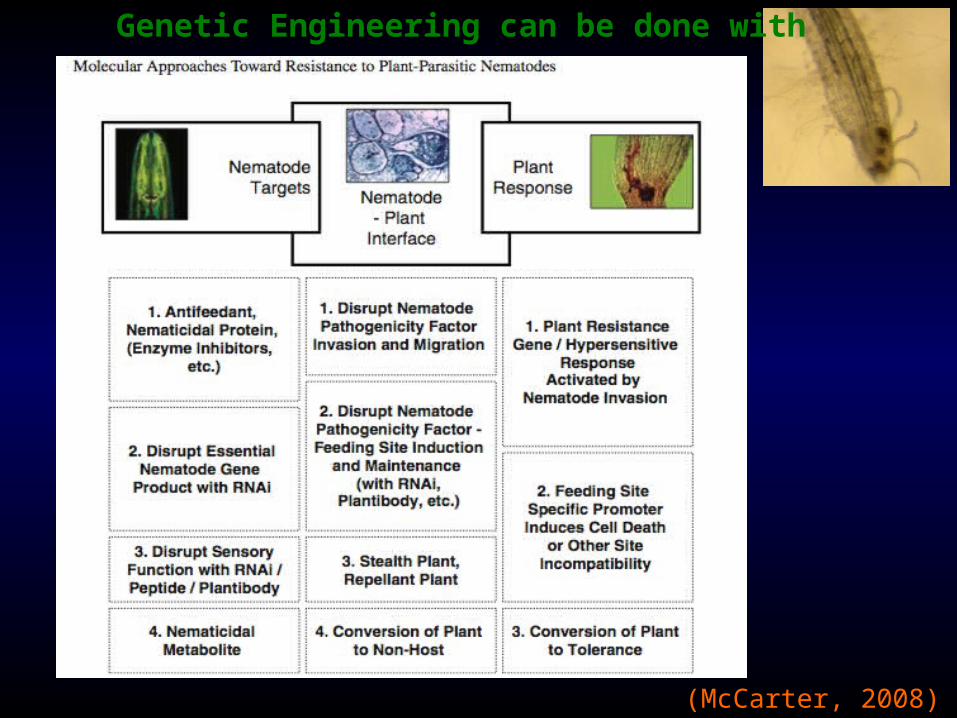
Genetic Engineering can be done with
(McCarter, 2008)
1. Antifeedant/Nematicidal Proteins
The expression of rice oryzacystatin protein, an inhibitor of cysteine proteases is most studied transgenic trait.
Nematode intestinal proteases are attractive targets for disruption for several reasons
(Lilley et al. 1999)
Function of digestion - essential to growth and reproduction is disturbed
Transgenic expression of a modified version of oryzacystatin, Oc-1∆D86, can interfere with nematode reproduction
(Urwin et al. 1995)

Beyond cysteine protease inhibitors, a serine protease inhibitor has been examined for control of the cereal cyst nematode Heterodera avenae in wheat
(Vishnudasan et al. 2005)
Cry5B, Cry6A, Cry14A, and Cry21A- Cry proteins also showed activity against nematode.
(Wei et al. 2003)
Cry5B interacts with the luminal surface of the C. elegans intestine via an invertebrate-specific glycolipid, loss of which conveys resistance.
(Griffitts et al. 2001, 2005)
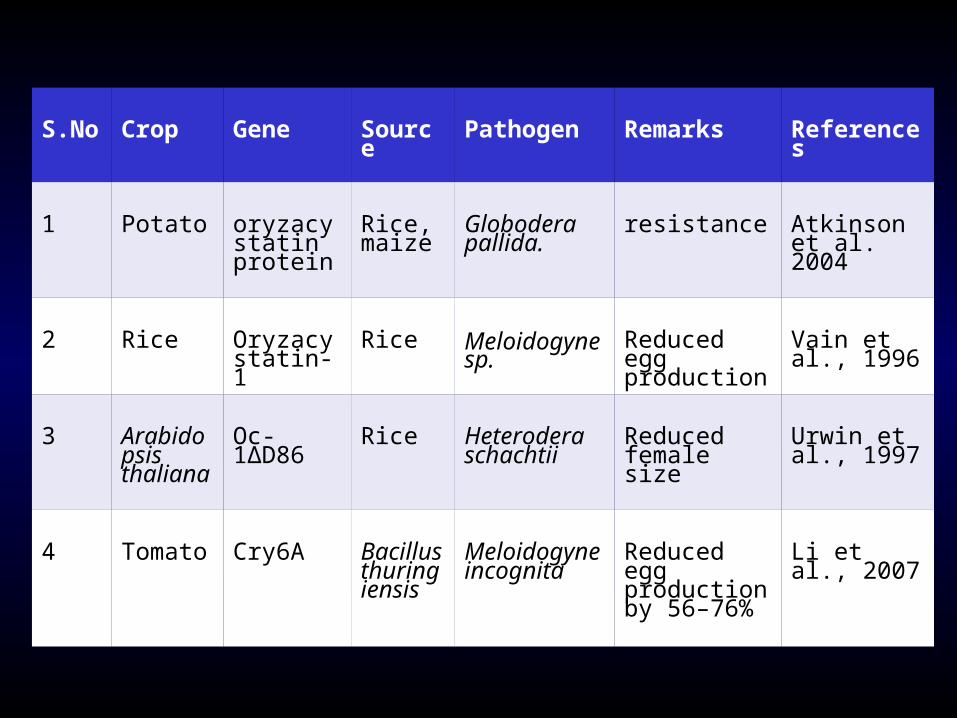
S.No Crop Gene Source Pathogen Remarks References
1 Potato oryzacystatin protein
Rice, maize
Globodera pallida.
resistance Atkinson et al. 2004
2 Rice Oryzacystatin- 1
Rice Meloidogyne sp.
Reduced egg production
Vain et al., 1996
3 Arabidopsis thaliana
Oc-1∆D86
Rice Heterodera schachtii
Reduced female size
Urwin et al., 1997
4 Tomato Cry6A Bacillus thuringiensis
Meloidogyne incognita
Reduced egg production by 56–76%
Li et al., 2007


Genotype-independent transformation system involving particle gun bombardment of immature embryos.
Elite African varieties (‘ITA212’, ‘IDSA6’, ‘LAC23’, ‘WAB56-104’).
Twenty-five transformed clones containing genes coding for an engineered cysteine proteinase inhibitor (oryzacystatin-IΔD86, OC-IΔD86), hygromycin resistance (aphIV) and β-glucuronidase (gusA) were recovered from the four varieties.

Tranformed plants were regenerated from all clones and analysed by PCR, Southern and western blot.
Up to 0.2% total soluble protein in plant roots were measured in 12 out of 25 transformed rice lines.
This level of expression resulted in a significant 55% reduction in egg production by Meloidogyne incognita.
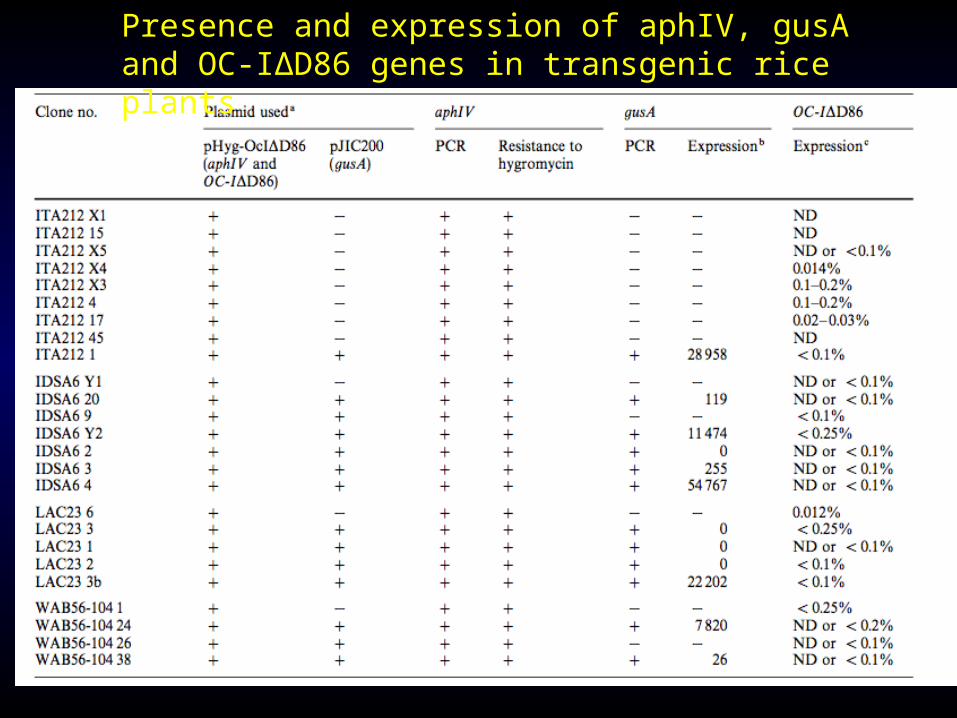
Presence and expression of aphIV, gusA and OC-IΔD86 genes in transgenic rice plants

Lanes 1—3 were loaded with 2 μg total protein from T0 transformed rice plants regenerated from three independent lines (ITA212 nos. 45, 4 and 15, respectively).Lanes 4—8 were loaded with 2 μg total protein from an untransformed rice plant to which 0, 0.25, 0.5, 0.75, or 1% OC-IΔD86
(IDB21) and ITA212 regenerated fertile
transgenic rice plants containing the
engineered OC-IΔD86 oryzacystatin gene
Western blot with a polyclonal antibody raised against
Oryzacystatin OC-IΔD86
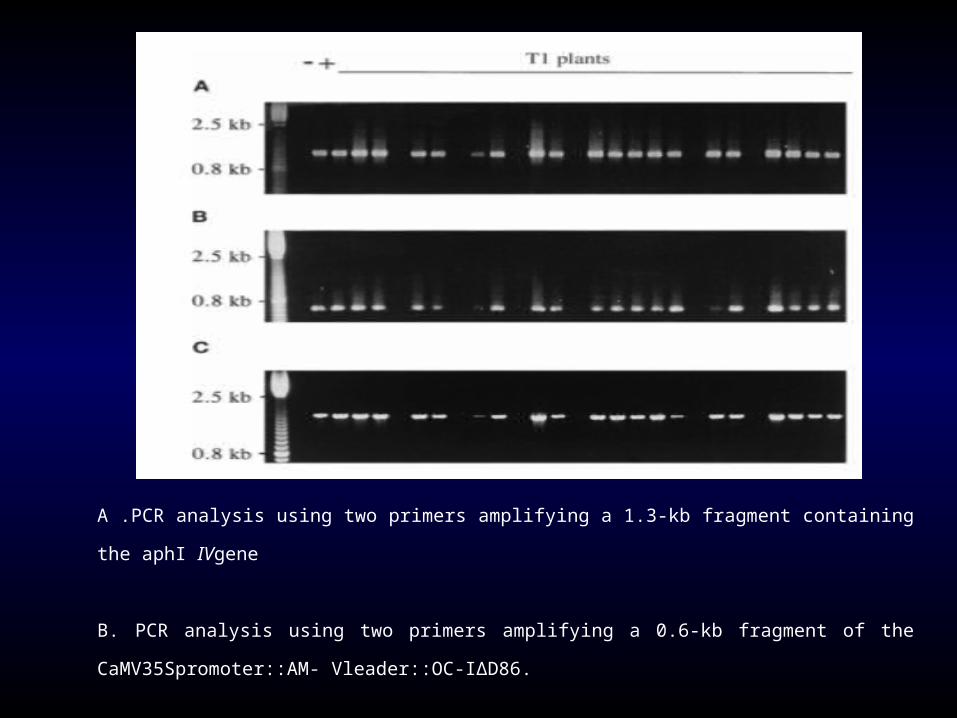
A .PCR analysis using two primers amplifying a 1.3-kb fragment containing the aphI IVgene
B. PCR analysis using two primers amplifying a 0.6-kb fragment of the CaMV35Spromoter::AM-
Vleader::OC-IΔD86.
C. PCR analysis using two primers amplifying a 1.6-kb fragment of the ubi5” region : : gusA gene.
Non- transformed plants (-) and plasmid DNA (+) were used as negative and positive controls, respectively

After a 42 day exposure of the rice plants to nematode- infested soil, established females of M. incognita
Western-negative plants had produced an overall mean for the three harvests of 3631±1381 eggs/g root mass.
The corresponding overall mean for plants expressing a low level of OC-IΔD86 was 1995±596 eggs/g root mass.
The reduction in eggs on the latter plants to 55% of the controls was statistically significant


Serine proteinase inhibitors (IP’s) are proteins found naturally in a wide range of plants with a significant role in the natural defense system of plants against herbivores.
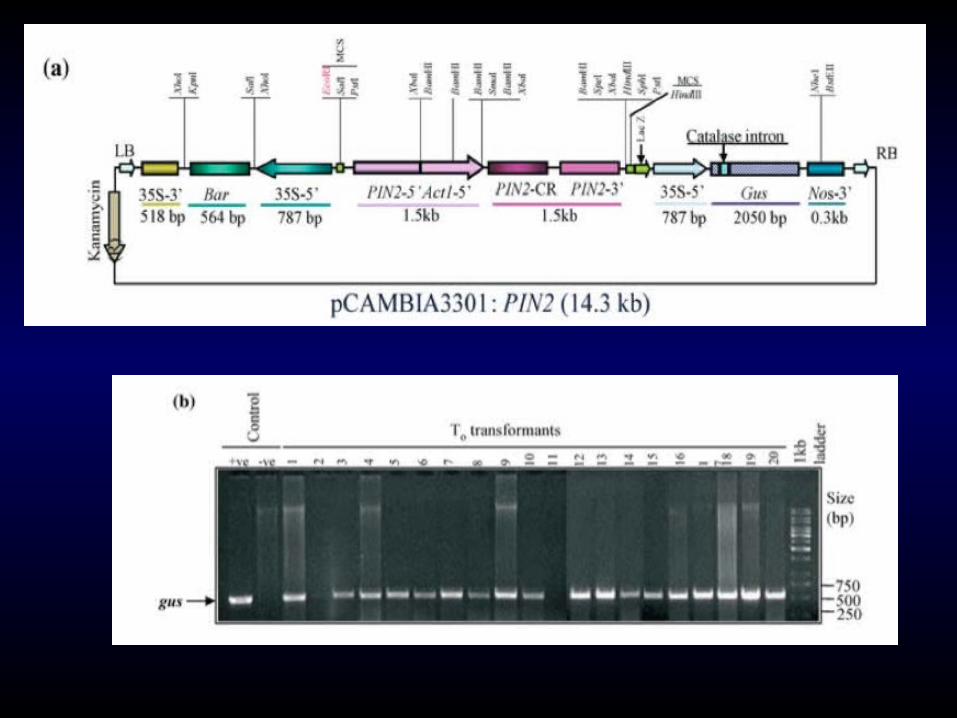
A plant serine proteinase inhibitor (pin2) gene into T. durum PDW215 by Agrobacterium-mediated transformation to combat cereal cyst nematode (Heterodera avenae) infestation
The plant proteinase inhibitors (PI’s) bind with the proteinases present in the gut and thereby hinder its proteolytic activity that further impedes the development of feeding pests.
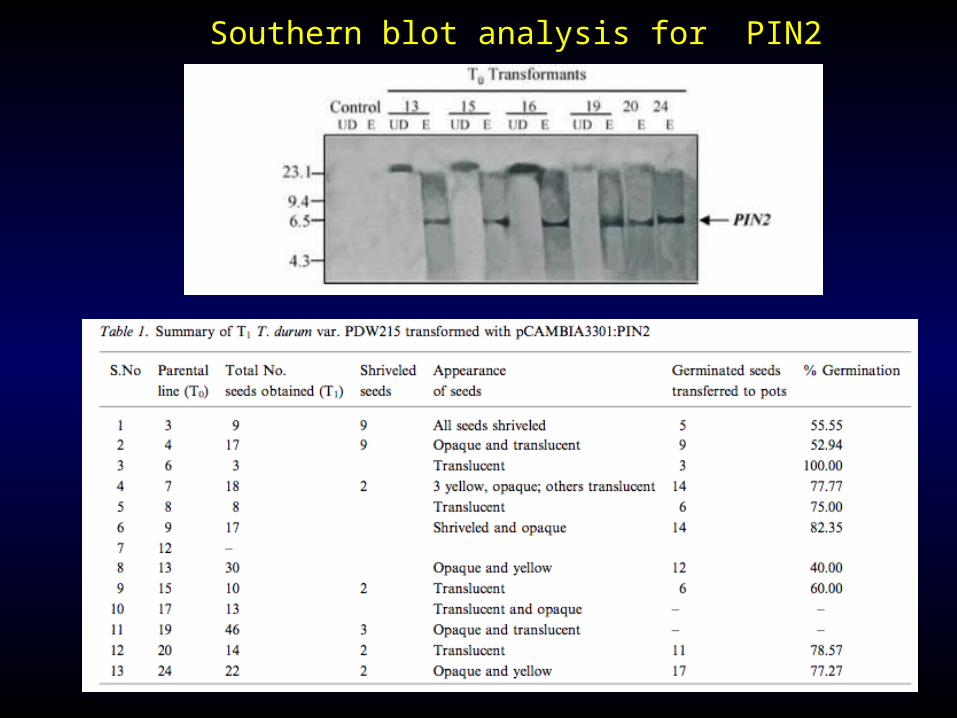
Southern blot analysis for PIN2

The PIN2 systemic expression confers satisfactory nematode resistance.

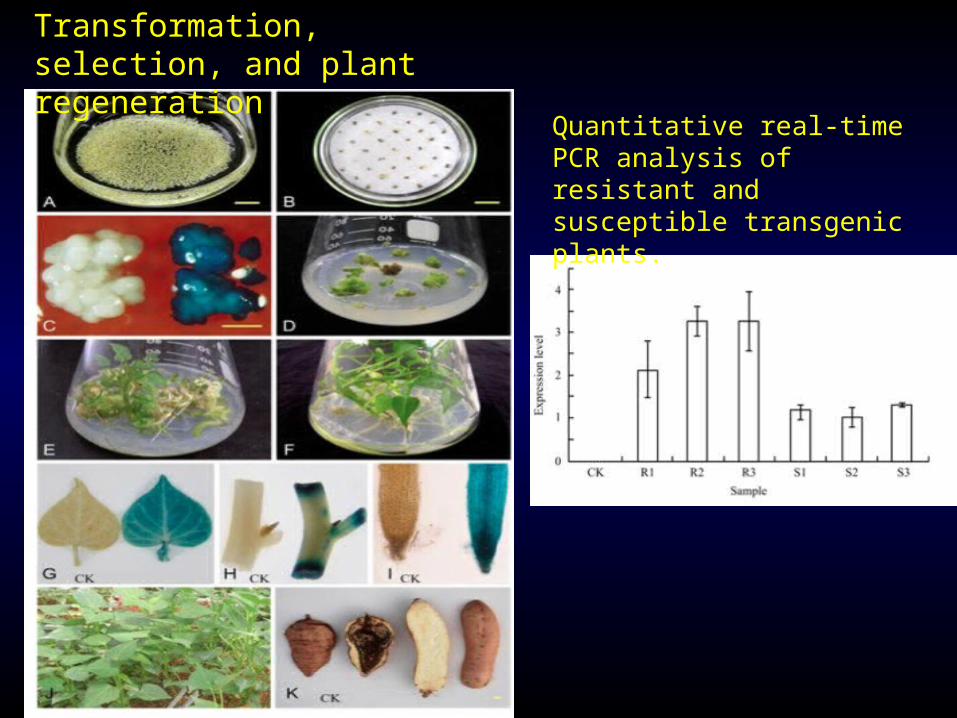
Transformation, selection, and plant regeneration
Quantitative real-time PCR analysis of resistant and susceptible transgenic plants.

2. Disruption of Essential Nematode Gene Products with RNAiThe critical role of dsRNA in silencing was first elucidated in
C. elegans
(Fire et al. 1998)
RNAi occurs by an endogenous cellular pathway, which includes the dicer protein that processes long dsRNAs into 21mers (siRNAs) and the RISC protein complex that guides siRNA-mRNA base pairing and degradation
(Mello and Conte 2004)


Parasitism proteins expressed in esophageal gland cells mediate infection and parasitism of plants by root-knot nematodes (RKN).
Parasitism gene 16 D10 encodes a conserved RKN secretory peptide that stimulates root growth and functions as a ligand for a putative plant transcription factor.
in vitro and in vivo RNA interference approaches to silence
this parasitism gene in RKN and validate that the parasitism gene has an essential function in RKN parasitism of plants.
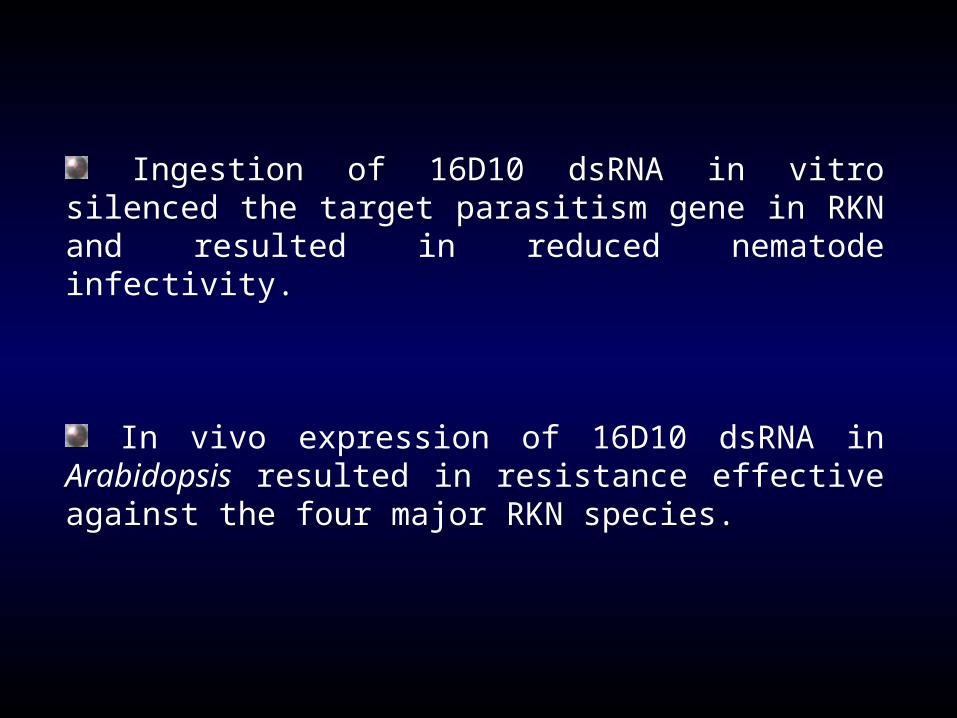
Ingestion of 16D10 dsRNA in vitro silenced the target parasitism gene in RKN and resulted in reduced nematode infectivity.
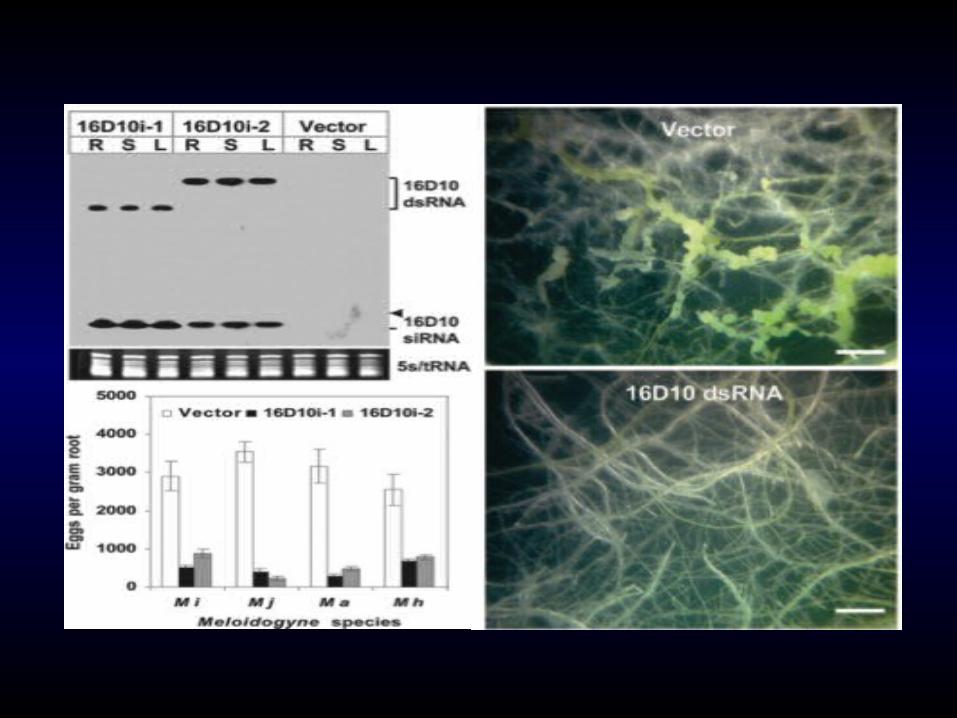
In vivo expression of 16D10 dsRNA in Arabidopsis
resulted in resistance effective against the four major RKN species.
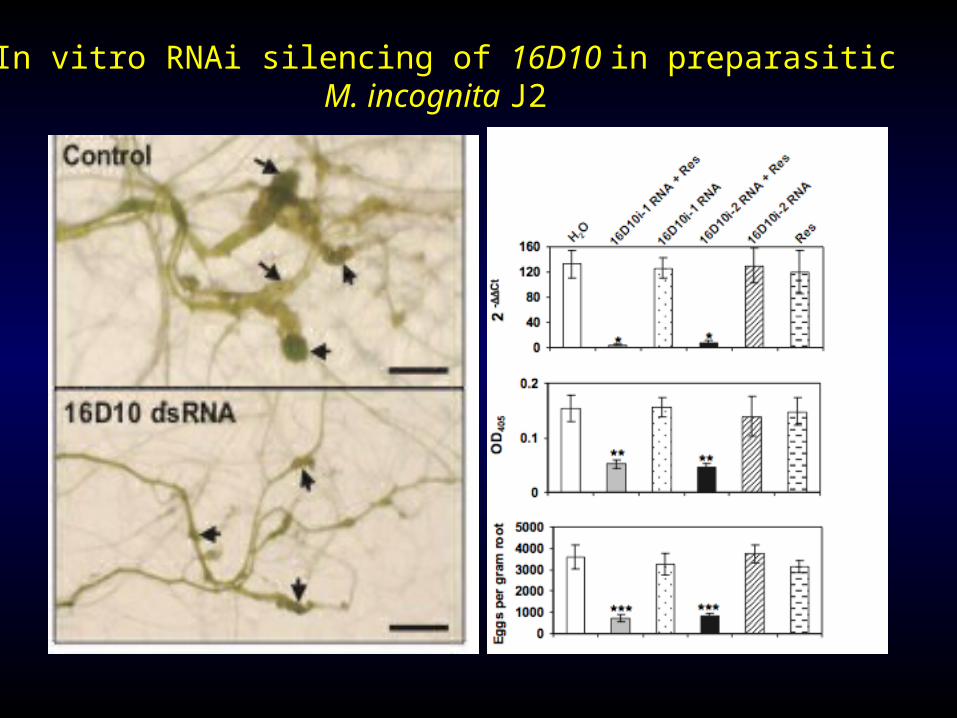
In vitro RNAi silencing of 16D10 in preparasitic M. incognita J2
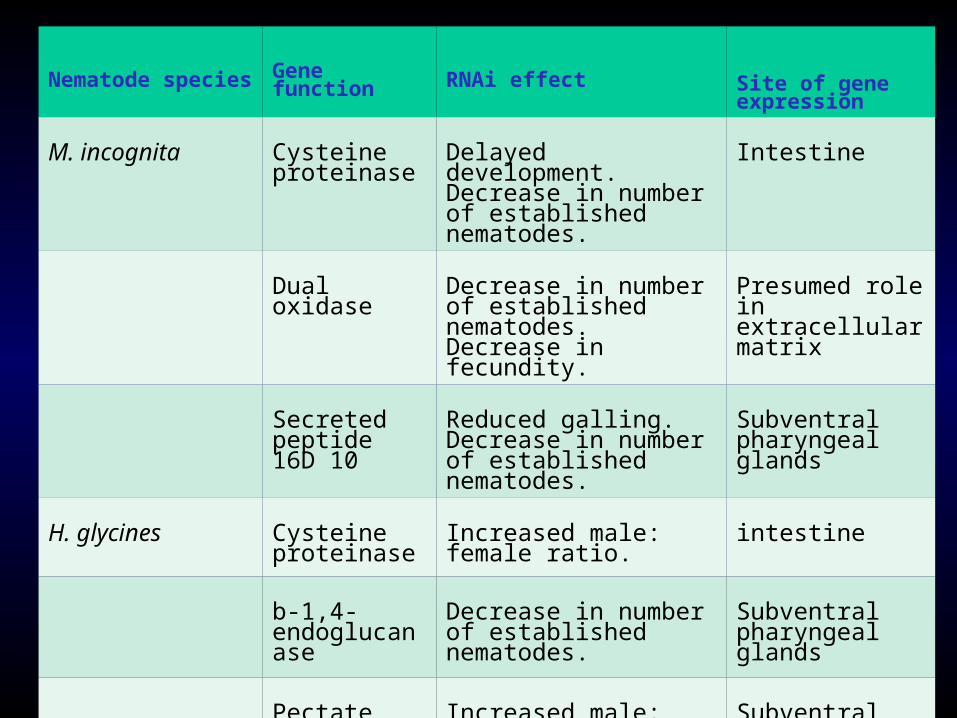
Nematode species Gene function RNAi effect Site of gene expression
M. incognita Cysteine proteinase
Delayed development.Decrease in number of established nematodes.
Intestine
Dual oxidase Decrease in number of established nematodes. Decrease in fecundity.
Presumed role in extracellular matrix
Secreted peptide 16D 10
Reduced galling.Decrease in number of established nematodes.
Subventral pharyngeal glands
H. glycines Cysteine proteinase
Increased male: female ratio.
intestine
b-1,4-endoglucanase
Decrease in number of established nematodes.
Subventral pharyngeal glands
Pectate lyase Increased male: female ratio.
Subventral pharyngeal glands
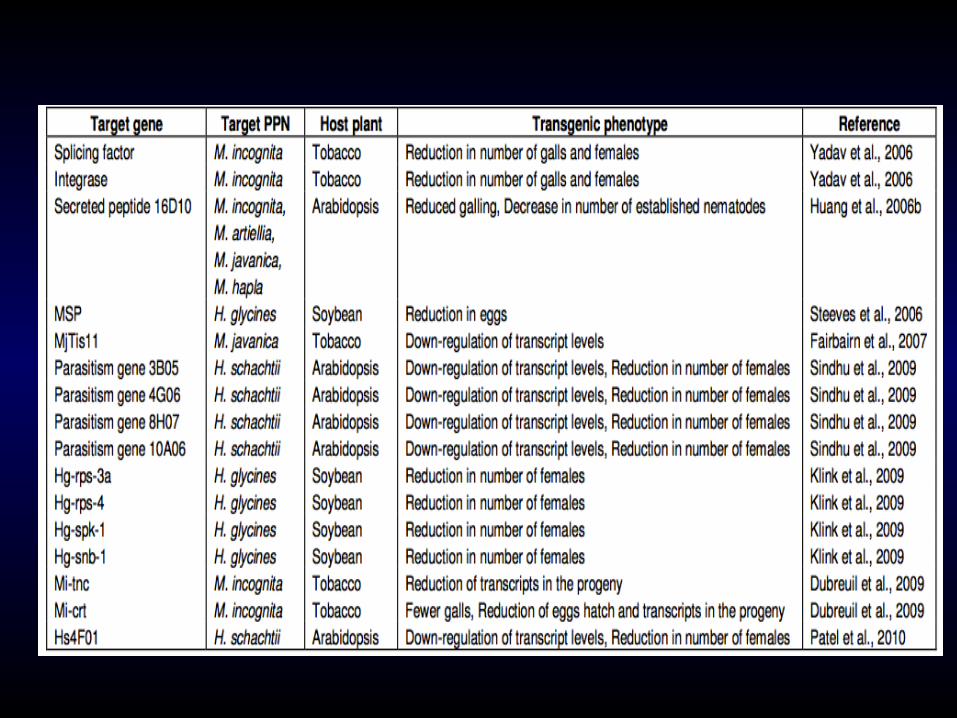

Plants are defined as resistant to nematodes when they support reduced levels of reproduction. Nematode resistance genes are present in several crop species and are an important component of many breeding programs including those for tomato, potato, soybeans and cereals.
Several resistance genes have been mapped to chro- mosomal locations or linkage groups.
A few have been cloned and a number of additional genes are likely to be cloned in the near future.
3. Nematode resistance genes
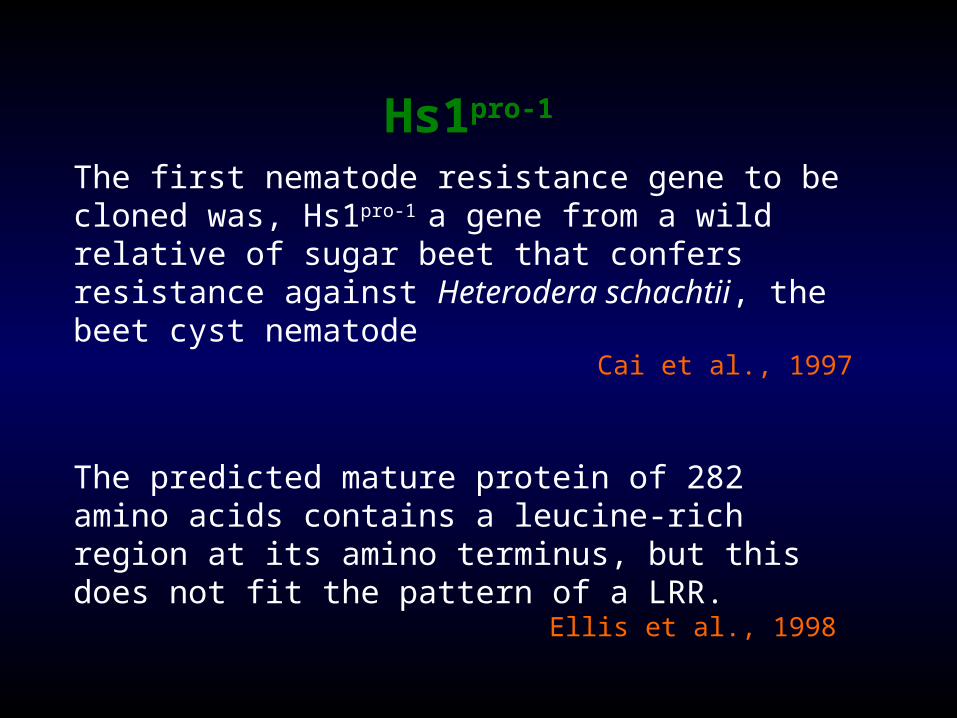
Hs1pro-1
The first nematode resistance gene to be cloned was, Hs1pro-1 a gene from a wild relative of sugar beet that confers resistance against Heterodera schachtii, the beet cyst nematode
Cai et al., 1997
The predicted mature protein of 282 amino acids contains a leucine-rich region at its amino terminus, but this does not fit the pattern of a LRR.
Ellis et al., 1998

Mi, a single, dominant locus conferring resistance to three rootknot nematode species (M. incognita, M. javanica, and M. arenaria)
Mi – 1 gene was introduced into cultivated tomato, Lycoperisicon esculentum, from the wild species L. peruvianum by embryo rescue of the interspecific cross.
(Roberts and Thomason, 1986)
Mi is one of the NBS/LRR class of resistance genes. Genes in this class gave nucleotide binding sites (NBS) and leucine rich repeats (LRR) as part of the protein architecture.
Mi-1

Hypersensitive response on the part of the host.
Necrotic plant cells are visible around the head of the invading nematode within six hours of inoculation of roots with nematode juveniles
(Dropkin et al., 1969; Dropkin, 1969)
Necrosis of host cells precludes larval growth, and the larvae die within 96 hours
(Riggs and Winstead, 1959; Dropkin et al., 1969).
Resistance mediated by Mi is lost at elevated temperatures.
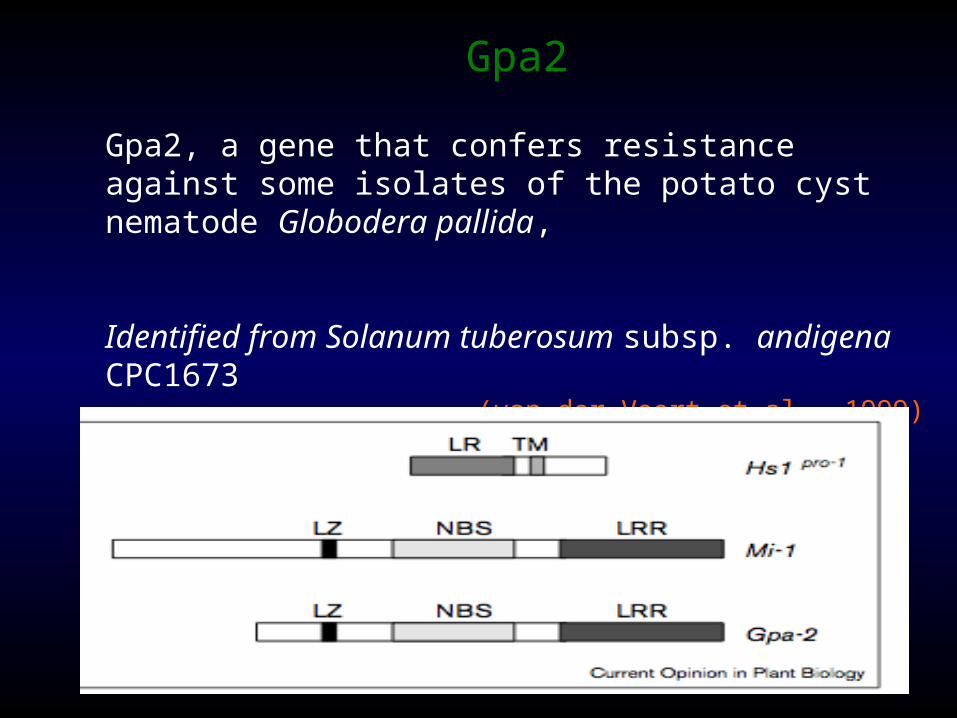
Gpa2
Gpa2, a gene that confers resistance against some isolates of the potato cyst nematode Globodera pallida,
Identified from Solanum tuberosum subsp. andigena CPC1673
(van der Voort et al., 1999)
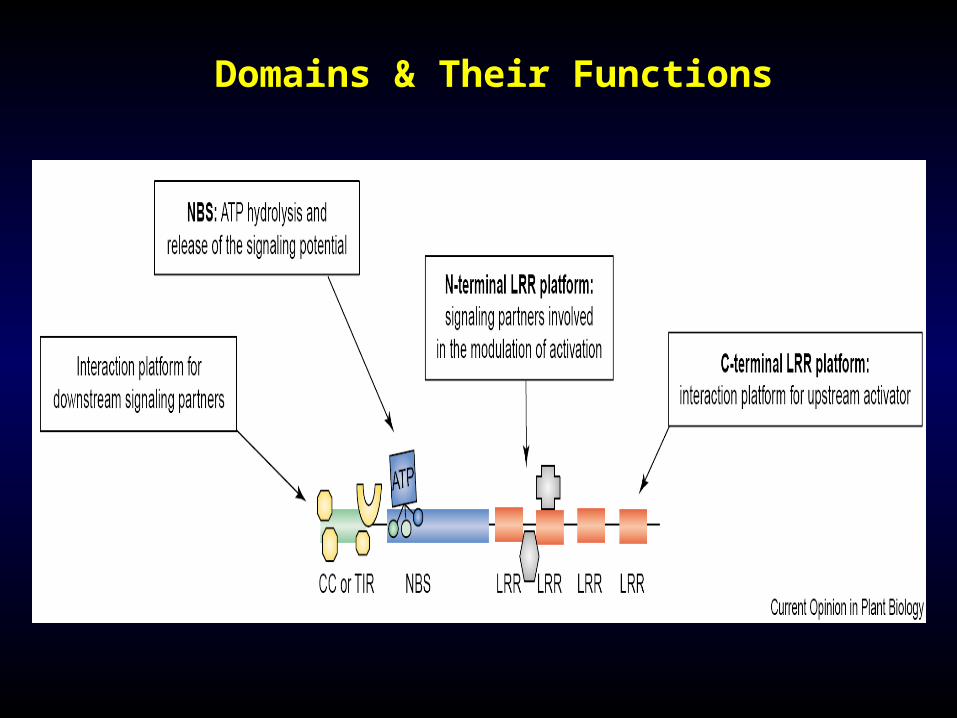
Domains & Their Functions


Results

Mi-1.2 gene, which encodes a leucine-rich repeat protein and confers resistance to three Meloidogyne species as well as aphids and white flies.
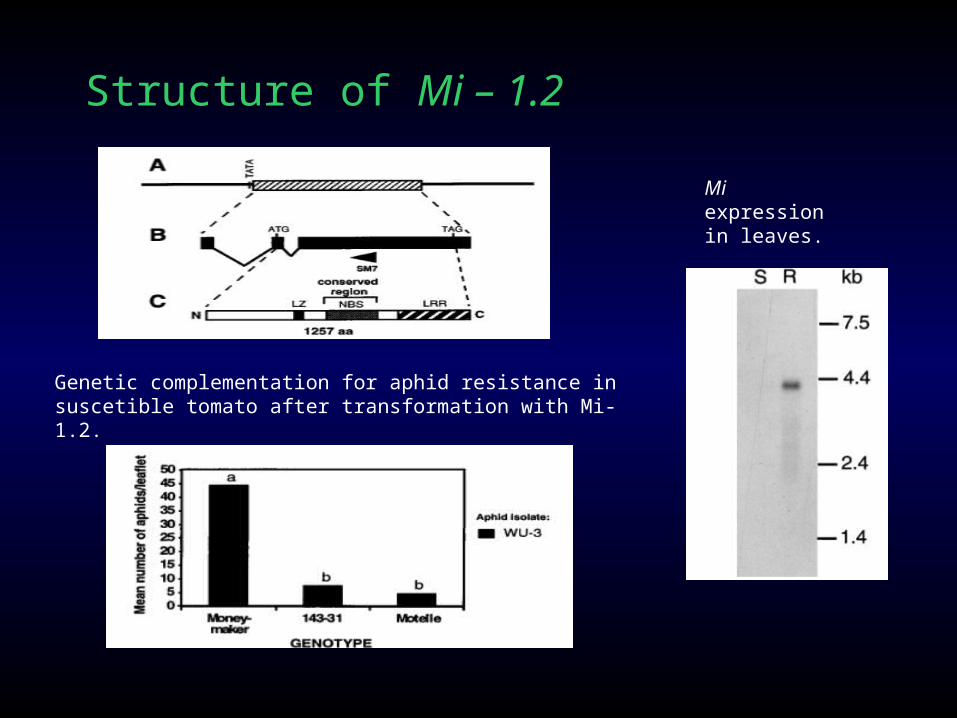
Structure of Mi – 1.2
Genetic complementation for aphid resistance in suscetible tomato after transformation with Mi-1.2.
Mi expression in leaves.

4. Nematicidal Metabolites
Naturally occurring nematicidal metabolites from plants have been characterized including certain polythienyls, alkaloids, lipids, isoflavonoids, and diterpenoids
Major limiting factor in transgenic production of nematicidal metabolites is a lack of knowledge of the biosynthetic pathways responsible for the production of many of these molecules.



















